
Submitted:
25 December 2025
Posted:
26 December 2025
You are already at the latest version
Abstract
Cadherin-5 (CDH5), also known as vascular endothelial cadherin (VE-cadherin), plays essential roles in endothelial cell adhesion, vascular barrier function, and signaling. CDH5 coordinates endothelial cell–cell junction during vascular remodeling, which is indispensable for both vascular homeostasis and adaptive responses to pathological stimuli. Although anti-CDH5 monoclonal antibodies (mAbs) can be used for individual applications including flow cytometry, western blotting, and immunohistochemistry (IHC), highly sensitive and versatile anti-CDH5 mAbs for all applications remain limited. Here, novel anti-human CDH5 mAbs, designated Ca5Mabs, were developed using a flow cytometry-based high-throughput screening. Among them, a clone Ca5Mab-8 (IgG2a, κ) recognized CDH5-overexpressed Chinese hamster ovary-K1 (CHO/CDH5) cells in flow cytometry. Furthermore, Ca5Mab-8 also recognized endogenous CDH5-expressing human endothelial cell lines (HUVEC/TERT2 and HDMVEC/TERT164-B) and a cervical cancer cell line (Hela). These reactivities were superior to a commercially available anti-CDH5 mAb (clone BV9). The dissociation constant value of Ca5Mab-8 for CHO/CDH5 was determined as 6.1 × 10⁻9 M. Ca5Mab-8 can detect endogenous CDH5 in Western blotting. Moreover, Ca5Mab-8, but not BV9, is available for IHC to detect endothelial cells in formalin-fixed paraffin-embedded tissues. These results indicate that Ca5Mab-8 is versatile for research and are expected to contribute to clinical applications, such as tumor diagnosis and therapy.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Plasmid Construction and Establishment of Stable Transfectants
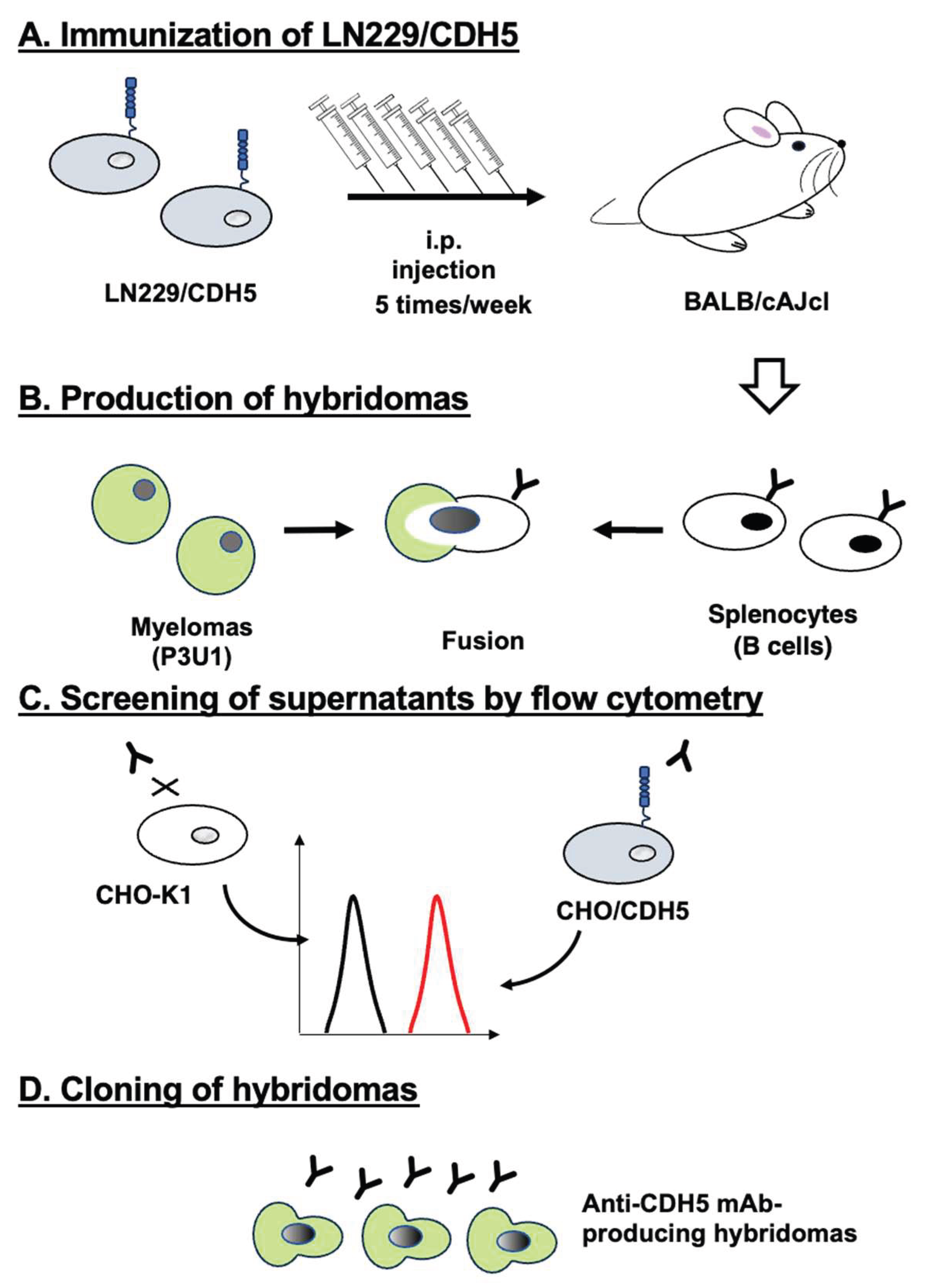
2.3. Production of Hybridomas
2.4. Flow Cytometry
2.5. Determination of Dissociation Constant Values Using Flow Cytometry
2.6. Western Blotting
2.7. IHC Using Cell Blocks
2.8. IHC Using a Tissue Array
3. Results
3.1. Development of anti-CDH5 mAbs
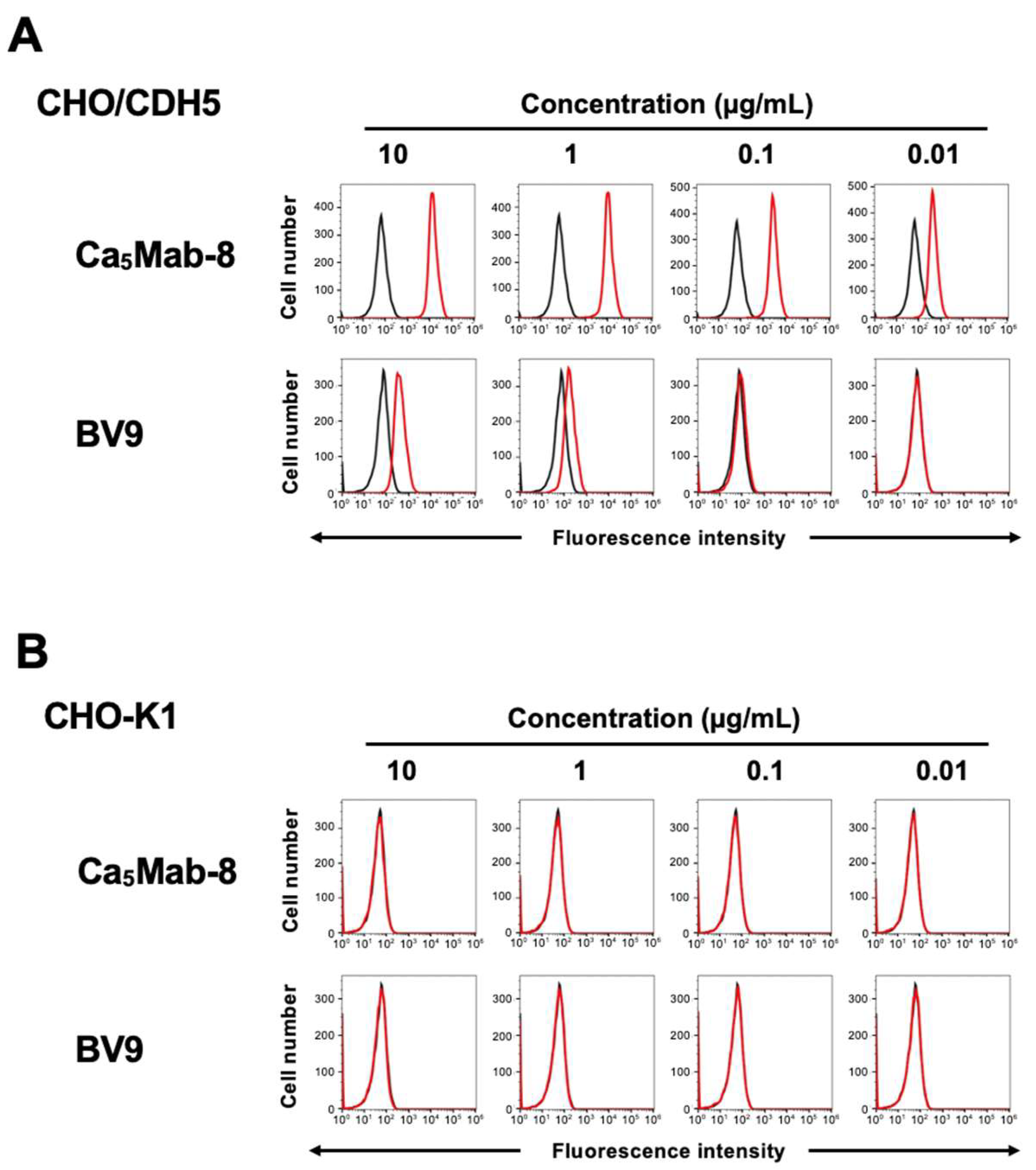
3.2. Flow Cytometry Using anti-CDH5 mAb Against CDH5-Overexpressed CHO-K1
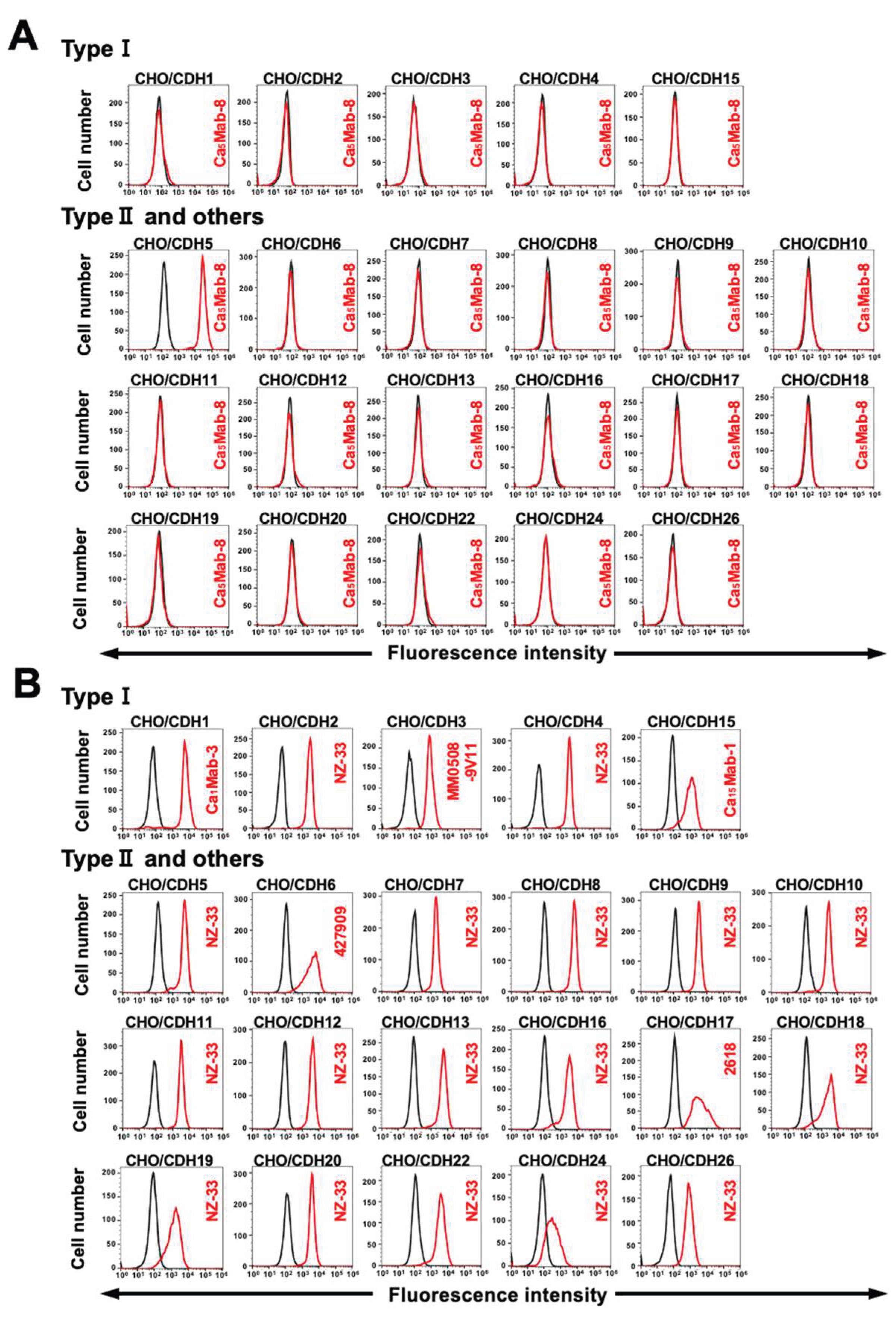
3.3. Specificity of Ca5Mab-8 to CDHs-Overexpressed CHO-K1
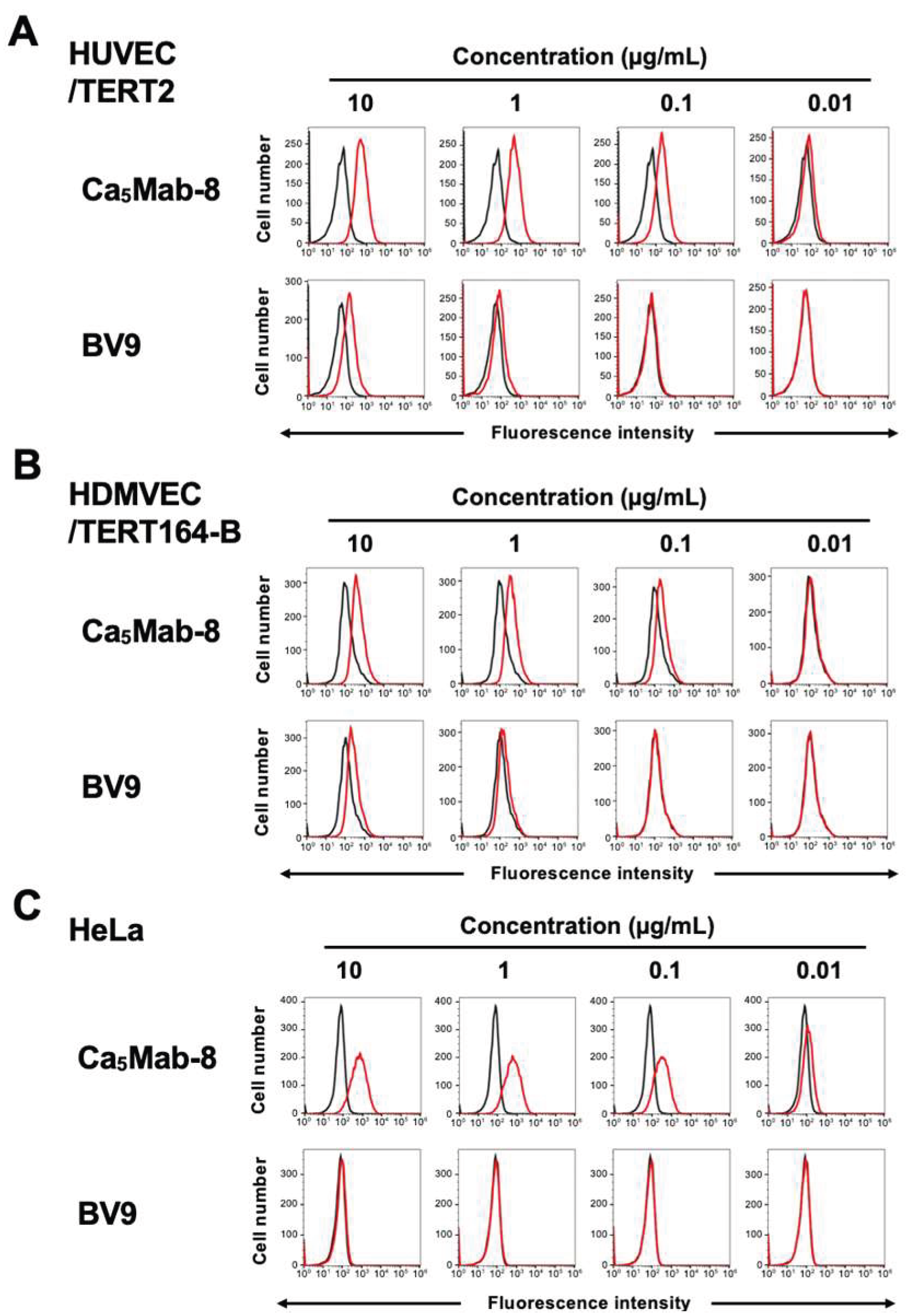
3.4. Flow Cytometry Using anti-CDH5 mAb Against Endogenous CDH5-Expressing Cells
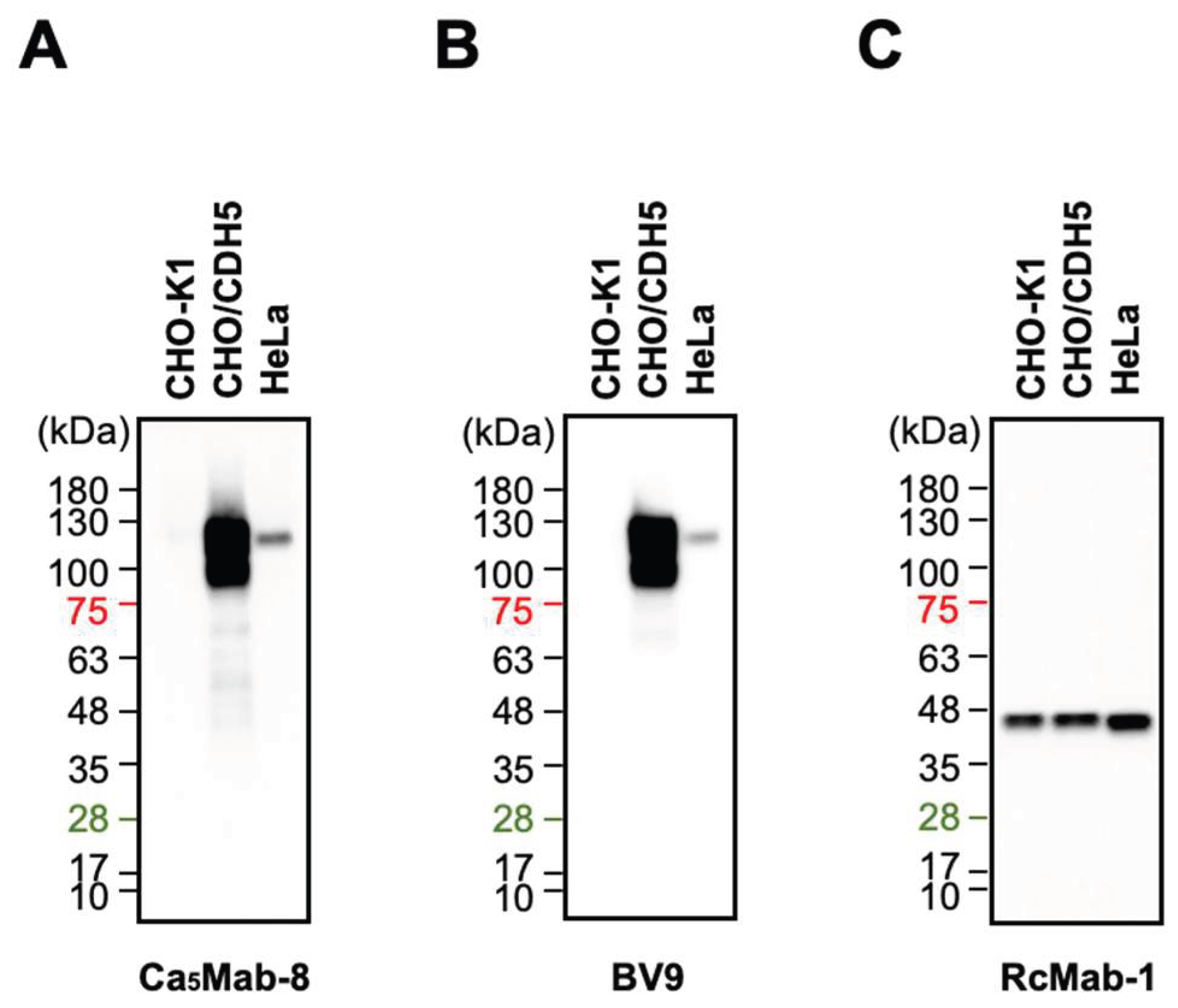
3.5. Western Blotting Using Ca5Mab-8 and BV9
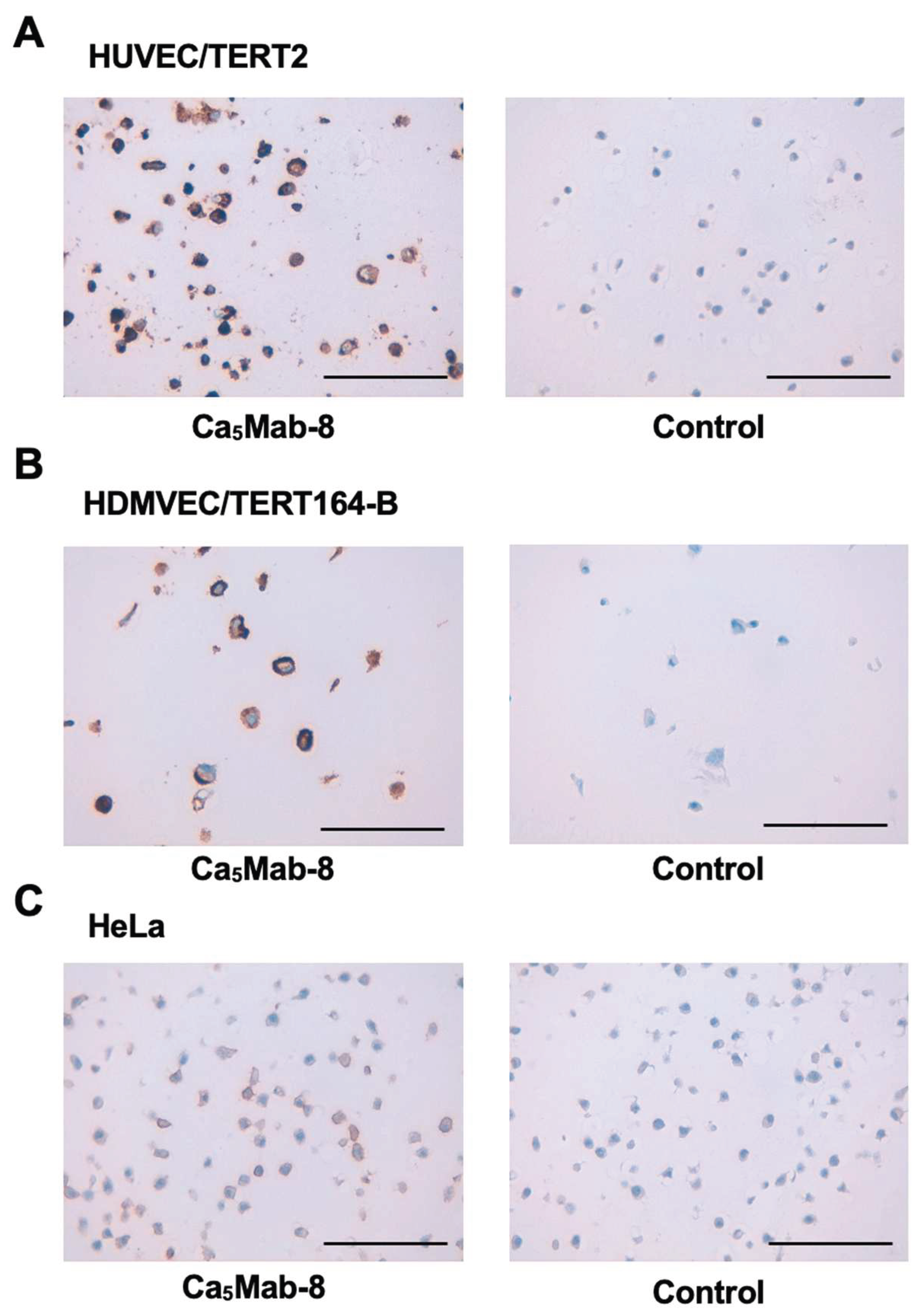
3.6. IHC Using Ca5Mab-8 in Formalin-Fixed Paraffin-Embedded Cell Blocks
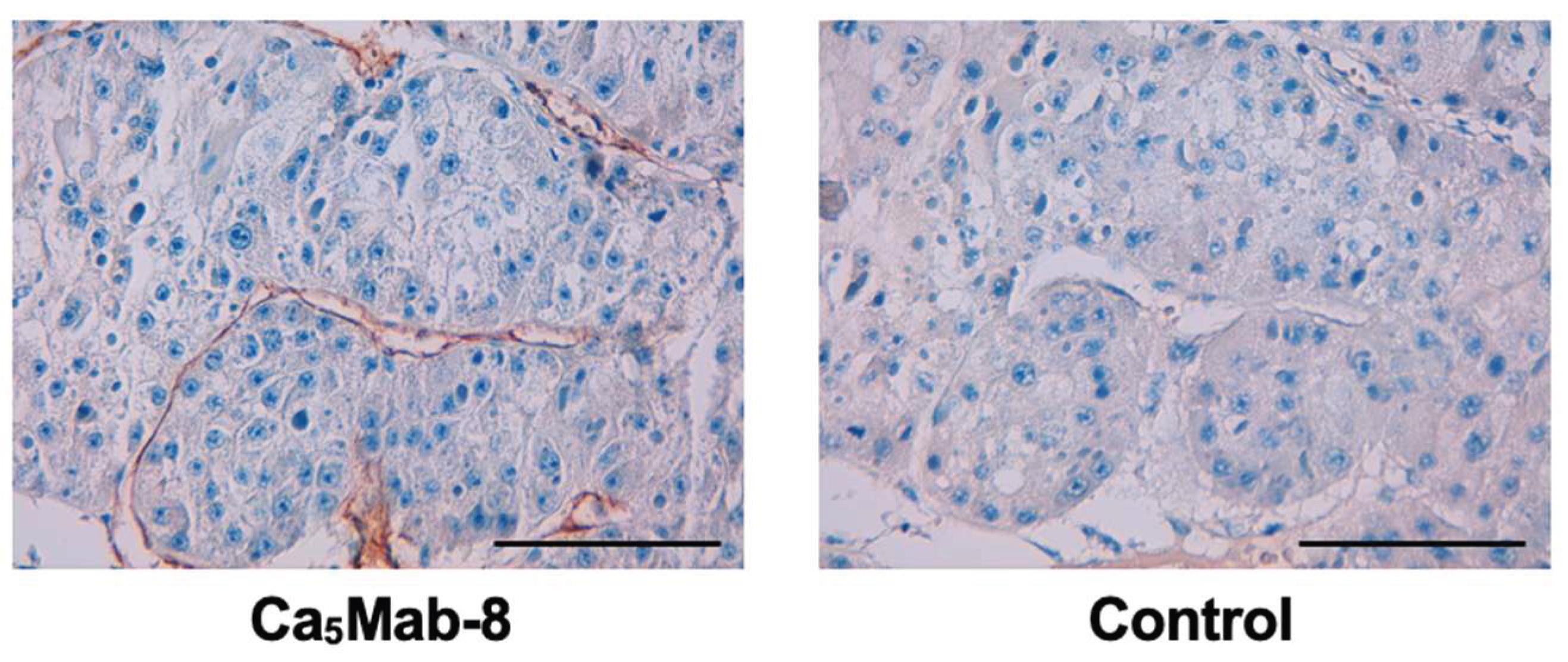
3.7. IHC Using Ca5Mab-8 in Formalin-Fixed Paraffin-Embedded Tissues
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nan, W.; He, Y.; Wang, S.; Zhang, Y. Molecular mechanism of VE-cadherin in regulating endothelial cell behaviour during angiogenesis. Front Physiol 2023, 14, 1234104. [CrossRef]
- van Roy, F. Beyond E-cadherin: roles of other cadherin superfamily members in cancer. Nat Rev Cancer 2014, 14, 121-134. [CrossRef]
- Lin, W.H.; Cooper, L.M.; Anastasiadis, P.Z. Cadherins and catenins in cancer: connecting cancer pathways and tumor microenvironment. Front Cell Dev Biol 2023, 11, 1137013. [CrossRef]
- Ratheesh, A.; Yap, A.S. A bigger picture: classical cadherins and the dynamic actin cytoskeleton. Nat Rev Mol Cell Biol 2012, 13, 673-679. [CrossRef]
- Dejana, E.; Orsenigo, F. Endothelial adherens junctions at a glance. J Cell Sci 2013, 126, 2545-2549. [CrossRef]
- Bazzoni, G.; Dejana, E. Endothelial cell-to-cell junctions: molecular organization and role in vascular homeostasis. Physiol Rev 2004, 84, 869-901. [CrossRef]
- Dejana, E.; Giampietro, C. Vascular endothelial-cadherin and vascular stability. Curr Opin Hematol 2012, 19, 218-223. [CrossRef]
- Rahimi, N. Defenders and Challengers of Endothelial Barrier Function. Front Immunol 2017, 8, 1847. [CrossRef]
- Marcos-Ramiro, B.; García-Weber, D.; Millán, J. TNF-induced endothelial barrier disruption: beyond actin and Rho. Thromb Haemost 2014, 112, 1088-1102. [CrossRef]
- Parnell, E.; Smith, B.O.; Palmer, T.M.; Terrin, A.; Zaccolo, M.; Yarwood, S.J. Regulation of the inflammatory response of vascular endothelial cells by EPAC1. Br J Pharmacol 2012, 166, 434-446. [CrossRef]
- Weis, S.; Cui, J.; Barnes, L.; Cheresh, D. Endothelial barrier disruption by VEGF-mediated Src activity potentiates tumor cell extravasation and metastasis. J Cell Biol 2004, 167, 223-229. [CrossRef]
- Delgado-Bellido, D.; Oliver, F.J.; Vargas Padilla, M.V.; Lobo-Selma, L.; Chacón-Barrado, A.; Díaz-Martin, J.; de Álava, E. VE-Cadherin in Cancer-Associated Angiogenesis: A Deceptive Strategy of Blood Vessel Formation. Int J Mol Sci 2023, 24. [CrossRef]
- Vittet, D.; Buchou, T.; Schweitzer, A.; Dejana, E.; Huber, P. Targeted null-mutation in the vascular endothelial-cadherin gene impairs the organization of vascular-like structures in embryoid bodies. Proc Natl Acad Sci U S A 1997, 94, 6273-6278. [CrossRef]
- Wen, L.; Yan, W.; Zhu, L.; Tang, C.; Wang, G. The role of blood flow in vessel remodeling and its regulatory mechanism during developmental angiogenesis. Cell Mol Life Sci 2023, 80, 162. [CrossRef]
- Wallez, Y.; Vilgrain, I.; Huber, P. Angiogenesis: the VE-cadherin switch. Trends Cardiovasc Med 2006, 16, 55-59. [CrossRef]
- Coon, B.G.; Baeyens, N.; Han, J.; Budatha, M.; Ross, T.D.; Fang, J.S.; Yun, S.; Thomas, J.L.; Schwartz, M.A. Intramembrane binding of VE-cadherin to VEGFR2 and VEGFR3 assembles the endothelial mechanosensory complex. J Cell Biol 2015, 208, 975-986. [CrossRef]
- Tzima, E.; Irani-Tehrani, M.; Kiosses, W.B.; Dejana, E.; Schultz, D.A.; Engelhardt, B.; Cao, G.; DeLisser, H.; Schwartz, M.A. A mechanosensory complex that mediates the endothelial cell response to fluid shear stress. Nature 2005, 437, 426-431. [CrossRef]
- Dabravolski, S.A.; Kashtalap, V.V.; Rozhkova, U.V.; Maksaeva, A.O.; Sukhorukov, V.N.; Orekhov, A.N. The role of cell junctions in atherosclerosis: implications for inflammation, endothelial dysfunction, and plaque stability. J Physiol Biochem 2025, 81, 573-587. [CrossRef]
- Brandon, K.D.; Frank, W.E.; Stroka, K.M. Junctions at the crossroads: the impact of mechanical cues on endothelial cell-cell junction conformations and vascular permeability. Am J Physiol Cell Physiol 2024, 327, C1073-c1086. [CrossRef]
- Berx, G.; van Roy, F. Involvement of members of the cadherin superfamily in cancer. Cold Spring Harb Perspect Biol 2009, 1, a003129. [CrossRef]
- Blaschuk, O.W.; Devemy, E. Cadherins as novel targets for anti-cancer therapy. Eur J Pharmacol 2009, 625, 195-198. [CrossRef]
- Corada, M.; Liao, F.; Lindgren, M.; Lampugnani, M.G.; Breviario, F.; Frank, R.; Muller, W.A.; Hicklin, D.J.; Bohlen, P.; Dejana, E. Monoclonal antibodies directed to different regions of vascular endothelial cadherin extracellular domain affect adhesion and clustering of the protein and modulate endothelial permeability. Blood 2001, 97, 1679-1684. [CrossRef]
- Corada, M.; Zanetta, L.; Orsenigo, F.; Breviario, F.; Lampugnani, M.G.; Bernasconi, S.; Liao, F.; Hicklin, D.J.; Bohlen, P.; Dejana, E. A monoclonal antibody to vascular endothelial-cadherin inhibits tumor angiogenesis without side effects on endothelial permeability. Blood 2002, 100, 905-911. [CrossRef]
- Liao, F.; Li, Y.; O'Connor, W.; Zanetta, L.; Bassi, R.; Santiago, A.; Overholser, J.; Hooper, A.; Mignatti, P.; Dejana, E.; et al. Monoclonal antibody to vascular endothelial-cadherin is a potent inhibitor of angiogenesis, tumor growth, and metastasis. Cancer Res 2000, 60, 6805-6810.
- Ubukata, R.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Development of novel anti-CDH1/E-cadherin monoclonal antibodies for versatile applications. Biochemistry and Biophysics Reports 2026, 45, 102401. [CrossRef]
- Ubukata, R.; Suzuki, H.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Development of an anti-CDH15/M-cadherin monoclonal antibody Ca(15)Mab-1 for flow cytometry, immunoblotting, and immunohistochemistry. Biochem Biophys Rep 2025, 43, 102138. [CrossRef]
- Fujii, Y.; Kaneko, M.; Neyazaki, M.; Nogi, T.; Kato, Y.; Takagi, J. PA tag: a versatile protein tagging system using a super high affinity antibody against a dodecapeptide derived from human podoplanin. Protein Expr Purif 2014, 95, 240-247. [CrossRef]
- Fujii, Y.; Kaneko, M.K.; Kato, Y. MAP Tag: A Novel Tagging System for Protein Purification and Detection. Monoclon Antib Immunodiagn Immunother 2016, 35, 293-299. [CrossRef]
- Satofuka, H.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Development of Anti-Human Cadherin-26 Monoclonal Antibody, Ca26Mab-6, for Flow Cytometry. Preprints 2025. [CrossRef]
- Fujisawa, S.; Yamamoto, H.; Tanaka, T.; Kaneko, M.K.; Suzuki, H.; Kato, Y. Development and characterization of Ea7Mab-10: A novel monoclonal antibody targeting ephrin type-A receptor 7. MI 2025. [CrossRef]
- Zhou, L.; Yang, Y.; Ye, Y.; Qiao, Q.; Mi, Y.; Liu, H.; Zheng, Y.; Wang, Y.; Liu, M.; Zhou, Y. Notch1 signaling pathway promotes growth and metastasis of gastric cancer via modulating CDH5. Aging (Albany NY) 2024, 16, 11893-11903. [CrossRef]
- Hung, M.S.; Chen, I.C.; Lung, J.H.; Lin, P.Y.; Li, Y.C.; Tsai, Y.H. Epidermal Growth Factor Receptor Mutation Enhances Expression of Cadherin-5 in Lung Cancer Cells. PLoS One 2016, 11, e0158395. [CrossRef]
- de Visser, K.E.; Joyce, J.A. The evolving tumor microenvironment: From cancer initiation to metastatic outgrowth. Cancer Cell 2023, 41, 374-403. [CrossRef]
- Leone, P.; Malerba, E.; Susca, N.; Favoino, E.; Perosa, F.; Brunori, G.; Prete, M.; Racanelli, V. Endothelial cells in tumor microenvironment: insights and perspectives. Front Immunol 2024, 15, 1367875. [CrossRef]
- Watabe, T.; Takahashi, K.; Pietras, K.; Yoshimatsu, Y. Roles of TGF-β signals in tumor microenvironment via regulation of the formation and plasticity of vascular system. Semin Cancer Biol 2023, 92, 130-138. [CrossRef]
- Huang, M.; Liu, T.; Ma, P.; Mitteer, R.A., Jr.; Zhang, Z.; Kim, H.J.; Yeo, E.; Zhang, D.; Cai, P.; Li, C.; et al. c-Met-mediated endothelial plasticity drives aberrant vascularization and chemoresistance in glioblastoma. J Clin Invest 2016, 126, 1801-1814. [CrossRef]
- Corada, M.; Mariotti, M.; Thurston, G.; Smith, K.; Kunkel, R.; Brockhaus, M.; Lampugnani, M.G.; Martin-Padura, I.; Stoppacciaro, A.; Ruco, L.; et al. Vascular endothelial-cadherin is an important determinant of microvascular integrity in vivo. Proc Natl Acad Sci U S A 1999, 96, 9815-9820. [CrossRef]
- Liao, F.; Doody, J.F.; Overholser, J.; Finnerty, B.; Bassi, R.; Wu, Y.; Dejana, E.; Kussie, P.; Bohlen, P.; Hicklin, D.J. Selective targeting of angiogenic tumor vasculature by vascular endothelial-cadherin antibody inhibits tumor growth without affecting vascular permeability. Cancer Res 2002, 62, 2567-2575.
- Singh, S.; Dey, D.; Barik, D.; Mohapatra, I.; Kim, S.; Sharma, M.; Prasad, S.; Wang, P.; Singh, A.; Singh, G. Glioblastoma at the crossroads: current understanding and future therapeutic horizons. Signal Transduct Target Ther 2025, 10, 213. [CrossRef]
- Nowacka, A.; Śniegocki, M.; Smuczyński, W.; Bożiłow, D.; Ziółkowska, E. Angiogenesis in Glioblastoma-Treatment Approaches. Cells 2025, 14. [CrossRef]
- Wang, R.; Chadalavada, K.; Wilshire, J.; Kowalik, U.; Hovinga, K.E.; Geber, A.; Fligelman, B.; Leversha, M.; Brennan, C.; Tabar, V. Glioblastoma stem-like cells give rise to tumour endothelium. Nature 2010, 468, 829-833. [CrossRef]
- Buruiana, A.; Florian, S.I.; Florian, A.I.; Soritau, O.; Susman, S. Endothelial transdifferentiation of glioma stem cells: a literature review. Acta Neuropathol Commun 2025, 13, 181. [CrossRef]
- Ricci-Vitiani, L.; Pallini, R.; Biffoni, M.; Todaro, M.; Invernici, G.; Cenci, T.; Maira, G.; Parati, E.A.; Stassi, G.; Larocca, L.M.; et al. Tumour vascularization via endothelial differentiation of glioblastoma stem-like cells. Nature 2010, 468, 824-828. [CrossRef]
- Kaneko, M.K.; Suzuki, H.; Kato, Y. Establishment of a Novel Cancer-Specific Anti-HER2 Monoclonal Antibody H(2)Mab-250/H(2)CasMab-2 for Breast Cancers. Monoclon Antib Immunodiagn Immunother 2024, 43, 35-43. [CrossRef]
- Arimori, T.; Mihara, E.; Suzuki, H.; Ohishi, T.; Tanaka, T.; Kaneko, M.K.; Takagi, J.; Kato, Y. Locally misfolded HER2 expressed on cancer cells is a promising target for development of cancer-specific antibodies. Structure 2024, 32, 536-549.e535. [CrossRef]
- Hosking, M.P.; Shirinbak, S.; Omilusik, K.; Chandra, S.; Kaneko, M.K.; Gentile, A.; Yamamoto, S.; Shrestha, B.; Grant, J.; Boyett, M.; et al. Preferential tumor targeting of HER2 by iPSC-derived CAR T cells engineered to overcome multiple barriers to solid tumor efficacy. Cell Stem Cell 2025, 32, 1087-1101.e1084. [CrossRef]
- Ubukata, R.; Ohishi, T.; Kaneko, M.K.; Suzuki, H.; Kato, Y. EphB2-Targeting Monoclonal Antibodies Exerted Antitumor Activities in Triple-Negative Breast Cancer and Lung Mesothelioma Xenograft Models. Int J Mol Sci 2025, 26. [CrossRef]
- Kaneko, M.K.; Suzuki, H.; Ohishi, T.; Nakamura, T.; Tanaka, T.; Kato, Y. A Cancer-Specific Monoclonal Antibody against HER2 Exerts Antitumor Activities in Human Breast Cancer Xenograft Models. Int J Mol Sci 2024, 25. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.



