
Submitted:
24 December 2025
Posted:
26 December 2025
You are already at the latest version
Abstract
Development of rapid, precise and fieldable detection methods for foodborne pathogen is one of the essential requirements in food safety and public health. In this research, the single nucleotide polymorphisms (SNPs) in the eae gene of Escherichia coli O157:H7 is well visually identified with the designed amplification refractory mutation system-polymerase chain reaction (ARMS-PCR) mediated lateral flow strip (LFS). Allele specific primers were designed to discriminate the wild-type eae genes from the mutant-type with the single-nucleotide resolution in the simple visual LFS format. The single nucleotide variation in eae gene could be easily differentiated by the observation of optical signal on the T line of LFS without any devices. Assay performance results show that it has a high sensitivity and specificity with the single-nucleotide differentiation ratio as low as 0.1%. This genetic polymorphisms screening performance could enumerate complex genetic variation into a simple and direct yes/no readout, highlighting the ultra-easy SNP sensing mode and the simplicity of the results output for practical applications. This ARMS-PCR mediated LFS offers a straightforward, swift, and economical strategy for SNP identification with great potential using in evolution of bacterial resistance genes and viral evolution under different environmental stresses.
Keywords:
E.coli O157:H7
; eae gene
; single nucleotide polymorphism (SNP)
; lateral flow strips (LFS)
; direct visual detection
; ARMS-PCR
; food born pathogen
1. Introduction
Bacterial infections, a major global health concern, impact millions of individuals every year [1,2].The majority of these bacterial infections are those that can be transmitted through the food chain [3]. According to the public report, every year forty-eight million cases of food-related illness are reported in the US. Additionally, due to the consumption of contaminated foods, cost of medical expenses and lost productivity in low-and middle-income countries is as high as US$110 billion annually [4]. Pathogenic E. coli is one of the most prevalent bacteria responsible for foodborne disease [5]. Although E.coli is commonly found in human as well as animal gut microbiome, this form of colonization is usually asymptomatic and some strains of E. coli have evolved into pathogenic organisms that cause clinical illness, such as diarrhea [6,7]. Human diarrheic Escherichia coli (DEC) strains are categorized into at least five pathotypes, which reflect the pathology caused during infection and, to some extent, the virulence factors they express: Enteroaggregative E. Coli (EAEC), Enterotoxigenic E. Coli (ETEC), Enteropathogenic E. Coli (EPEC), and Shiga toxic E. Coli (STEC), of which Enterohaemorrhagic E. Coli (EHEC) is a subset. Because of its low infectious dose (less than 100 organisms) and dangerous nature, EHEC has been considered as a particularly concerning zoonotic and foodborne illness, particularly in young children, the elderly, and those with impaired immune systems [8,9,10].
Among EHEC, E. Coli O157:H7 is well known for being a serious foodborne pathogen and a primary cause of foodborne illnesses including diarrhea and even death [11]. Most strains of EHEC have a plasmid that codes for a haemolysin and a chromosomally located locus of enterocyte effacement (LEE) pathogenicity island that enhance EHEC’s ability to cause intestinal injury and systemic complications by attaching bacteria tightly to host cells that are hallmarks of attaching and effacing (A/E) lesions [12].These genes that enable bacteria to create adhesion and effacing lesions in intestinal mucosa cells increase the severity of human infections. One of the most important and representative genes is the eae gene residing within the LEE pathogenicity island and encoding the intimin [13]. The eae gene is also crucial for the development of attaching and effacing lesions, promoting intimate adherence to host epithelial cells. Through a special attaching and effacing process mediated by intimin protein, E. coli O157:H7 can adheres to intestinal epithelial cells, causing illness [14]. The importance of eae gene in the pathogenesis has prompted the use of eae gene as a molecular diagnostic marker of pathogenic E. coli [15]. Nonetheless, eae gene is highly genetically polymorphic and introduces a variety of intimin subtypes that may affect virulence of the pathogen and the communication of this pathogen with tissues of the host [16]. Such variations, which are discovered by means of single nucleotide polymorphisms (SNPs), insertions, or deletions, are important in host specificity and tissue tropism, and provide a warning that close attention should be paid to these genetic alterations or evolutions in epidemiological studies and clinical diagnosis.
Conventional tools used to identify polymorphism largely center on sequencing technology including Sanger sequencing, which is costly, time intensive and advanced equipment required [17]. Next-generation sequencing (NGS) [18] has become a high-throughput substitute of traditional sequencing techniques; it also requires sophisticated infrastructure and is prohibitively costly to many diagnostic laboratories. Of great significance, the resolution of the current NGS sequencing methods cannot be as precise as the single nucleotide level. Other techniques such as the real time PCR [19,20] using allele-specific probe and high-resolution melting (HRM) [21] analysis could provide faster results, but the problems in terms of affordability and availability still exist especially in resource constrained environments. For FISH, it is routinely employed to image RNA or genes at single-cell resolution [22]. However, for one reason, it could only be used for in vivo image; for the other, the resolution of FISH is a little lower and difficult to be used to screen less gene alterations, making SNVs identification nearly impossible[23]. The sensitivity of gene detection can be effectively improved by using nucleic acid amplification while the precise primer design could be helpful for even the SNV identification. For example, Liu et al. Reported the in-situ loop-mediated isothermal amplification for detecting SNVs [24]. But the complicate multiple primer sets design and the subsequent high probability of false-positive results greatly constrain the practical applications. Consequently, there is growing need for straightforward, economical, and accessible methods that can be broadly utilized for easy and effective identification of SNP in pathogenic bacteria. Among the available molecular approaches, a promising solution to this requirement is the amplification refractory mutation system PCR (ARMS-PCR) [25]. This technique involves the design and application of allele-specific primers, which identify amplification on the basis of the consistence of the 3' terminal base of the primer with the target DNA, and leads to a high level of specificity in the identification of SNP [26]. In addition, ARMS-PCR does not require any special high-value instruments since it could be performed in regular portable thermal cyclers. That is the reason for its affordable application in low-resource labs. Although it has these benefits, traditional ARMS-PCR detection usually relies heavily on agarose gel electrophoresis in order to visualize the final amplification products, which is tedious and complex [27]. To overcome this intrinsic drawback, the lateral flow strip (LFS) assay was designed and well combined with ARMS-PCR to facilitate the process of detection for providing direct readable output without the complexity of gel electrophoresis. LFS assays have the potential to provide direct judgement results with classic visualization which is why they can be widely used for point-of-care tests with the early pragency test strip as one of the most famous and typical popular home test products[28]. Such kind approach not only simplifies the whole workflow of the detection but also significantly reduces turnaround time for the final judgements [29].
In this research, the significant eae gene of E. coli O157:H7 was taken as the target gene and the genetic polymorphism of eae gene was easily and visual screened with the designed ARMS-PCR-LFS platform in short time. Well designed allele-specific primer set combinations realized the differentiation of the wild-type and mutant-type of eae genes, and the amplicons were visualized and analyzed based on the signals on the T line of LFS. Assessment of the assay’s performance well showed that it could perform the discrimination of genetic polymorphism reliably, backed up by identifiable visual representations, thereby justifying its potential as a cost-efficient and effective method for routine dynamic pathogen surveillance. The validation of the mutated genes in real samples was also conducted with satisfied results, which formed the basis for practical application of this method in genetic alteration of pathogens under environmental stress and precise identification of genetic polymorphism in target genes.
2. Materials and Methods
2.1. Reagents and Instruments
All oligonucleotides including the functionalized forward primer 5-FITC-TTGATCAAACCAAGGCCAGC and reverse primer 5-biotin-CTAACAGTCGCTTTACCGAA (amplified length 250 bp), 2XPCR Mix with blue Dye, streptavidin (SAV), DNA Marker (25-500 bp), and 4S Red Plus nucleic acid dye (1000×), HAuCl4, sucrose, Tween-20, bovine serum albumin (BSA), trisodium citrate and agrose were purchased from Sangon Biotech Co., Ltd. (Shanghai, China). Goat-anti-mouse secondary antibody (Anti-Ab), fluorescein isothiocyanate antibody (FITC Ab), and bovine serum albumin (BSA) were purchased from Baird Bio-tech Co., Ltd. (Beijing, China). Lateral flow strip components include absorbent pads, conjugate pads, sample pads, CN 95, nitrocellulose (NC) membranes, sample pads, and adhesive backing pads were all purchased from Jie-ning Biotech, Shanghai. To confirm the specificity and sensitivity of specified primer sets, and to optimize the detection conditions of process, plasmids of eaeA gene were synthesized by Sangon Biotech (Shanghai, China).
2.2. Extraction of Genomic DNA and Sample Preparation for ARMS-PCR
Samples of milk, and orange juice were collected from the local supermarket and confirmed to be free of E. coli contamination through the standard culture method. DNA of E. coli (Mutant and Wild-type) was introduced into milk and juice samples to create combinations with varying mutated ratios (100%, 50%, 25%, 10%, 5%, 1% 0.1% and 0). To mitigate the impact of food-derived matrix inhibition effects to PCR, DNA was extracted from each mixture utilizing a commercial DNA extraction kit (Tiangen, China) in accordance with the manufacturer's guidelines. The extracted DNA was diluted in 50 μL of 1× TE buffer and and utilized as a template for further analysis.
2.3. Preparation of AuNPs and AuNP Labeled Anti-FITC Antibody Conjugates for LFS
The gold nanoparticles were prepared using the standard trisodium citrate reduction methodology with minimal alterations [30]. Overall, 2.55 mL of HAuCl4 (5 g/L) was mixed with 150 mL deionized water in a flask. After that, the resulting solution was heated to boiling while being stirred magnetically at 1000 rpm. Following that, 2.25 mL of trisodium citrate solution (1%) was added quickly to react with HAuCl4, causing the solution's color to change from black to gray, then to a stable wine-red. Ultimately, the resulting AuNPs solution was stirred and allowed to cool to room temperature and kept at 4 oC in refrigerator before using. For preparation of the AuNP/FITC-Ab conjugates, the pH of AuNPs was firstly adjusted with 10 μL K2CO3 (0.1 M), and then 4 μL FITC-Ab (1 mg/mL, dissolved by 10 mM PB buffer) was added. After incubating at room temperature for 1 hour, 100 μL 10% BSA was added to block the residual active sites on AuNPs for 60 min to prevent non-specific adsorption. After that the AuNP/FITC-Ab conjugates were concentrated by freezing (4 oC) centrifugation at 9500 r/min for 10 min. For the subsequent LFS assembly, the precipitate was re-dissolved in 100 μL resuspension E, which was then sprayed onto the conjugation pad (6×300 mm) by the Bio-Dot sprayers, which was then allowed to dry overnight at 27 oC.
2.4. Assembly of LFS
The LFS is made up of a PVC back-plastic plate (60×300 mm), an absorbent pad (18×300 mm), an NC membrane (25×300 mm), a conjugation pad (6×300 mm), and a sample pad (18×300 mm). The buffer solution (pH 8.0) containing 200 mL ddH2O, 50 mM Tris-HCl, 0.15 mM NaCl, and 0.25% Triton-100 was used to treat the sample pad while the solution (pH 8.0) containing 100 mL PB buffer, 5% sucrose, 2.5% PEG-20000, 1% alginate, 0.3% Tween-20 was used to pretreat the conjugation pad. After that, both the conjugation pad and sample pad were dried in an oven at 27 oC overnight. Using a Bio-Dot spraying device, the NC membrane was sprayed with 20 μL SAV (1 mg/mL, dissolved by 10 mM PB buffer) and 20 μL second anti-mouse Ab (1 mg/mL, dissolved by 10 mM PB buffer) at rate of 0.5 μL/cm to prepare the test line (T line) and the control line (C line), respectively. Following these pretreatments, the LFS was assembled by overlapping the sample pad, conjugation pad, NC membrane, and absorbent pad on the back-plastic plate with overlap 2 mm with the neighbor pad. Finally, the LFS was sliced into 3 mm width using an automatic strip cutter.
2.5. Primer Design for the Accurate Identification of SNP in the Target Eae Gene
The primers used in this study were designed in accordance with the ARMS-PCR amplification principle using Primer Premier 6.0 and synthesized by the Generay Biotech Co., Ltd., (Anhui, China). The detailed sequence information of primer and probe are given in (Table S1). The concept behind the design of ARMS-PCR primers is that when the target DNA is normal, a single nucleotide mismatch at the 3'-OH end of primer prevents Taq DNA polymerase from extending the primer under the proper PCR conditions, resulting in only the one-base mutated targets being amplified. To improve the ability to differentiate the normal and mutant alleles, an intentional mismatch was also introduced at the third nucleotide site of ARMS primer. NCBI Primer-BLAST (http://www.ncbi.nlm.nih.gov/tools/primer-blast/) was used to check their specificity to other potential templates. The online Oligo Evaluator program (http://www.oligoevaluator.com) was used to examine the suspected primer-dimer and hairpin structures.
2.6. ARMS-qPCR for Primer Design Evaluation
To identify the most effective primer sets for distinguishing the wild and mutant alleles, allele-specific real-time PCR (ARMS-qPCR) was performed using a Bio-Rad CFX96 system. Each 25 µL reaction system contained 12.5 µL qPCR master mix, 0.7 µL (10 µM) of forward and reverse primer, 1.2 µL (10 µM) TaqMan probe, 10.5 μL of H2O and 1 µL template DNA.
2.7. ARMS-PCR Assisted Rapid and Visual Identification Of SNP in Eae Gene of E. coli with the Lateral Flow Strip
Primers that demonstrated the highest discrimination capability between wild-type and single nucleotide mutant alleles in eae gene with ARMS-qPCR were designed and optimized for additional verification using conventional ARMS-PCR. ARMS-PCR amplification was performed in a 25 μL volume containing 0.5 μL of primer sets (10 μM) 1 μL of DNA templates , 12.5 μL of 2×sanTaq PCR Mix, and 10.5 μL H2O with the following amplification conditions: initial denaturation at 94°C for 3 min, followed by 30 cycles of denaturation at 94°C for 45 s, annealing at 60°C for 45s, and extension at 72°C for 1 min, with an additional extension step at 72°C for 5 min. For the convenient visual identification of SNP in the eae gene of E. coli, both the forward and reverse primer were modified with the FITC and biotin at the 5’ end, respectively. Subsequently, the amplicons would be dual labeled with both FITC and biotin in the amplification, which would be measured by the designed lateral flow strip.
3. Results
3.1. Detection of Mutation in Eae Gene of E. coli by the Designed ARMS-PCR Mediated Lateral Flow Strip in the Visual Mode
Rapid and easy identification of single nucleotide polymorphism (SNP) in eae gene of E. coli helps distinguishing the potential dangerous evolved or mutated strains of E. coli and provides information on genetic variants linked to different virulence factors. As demonstrated in the Figure 1, with the designed primer set combinations, the amplification efficiency of the wild-type is hampered by several mismatched nucleotides in the designed primer set. And the ARMS assay can realize the identification of SNP with the acceptable sensitivity and specificity. As shown in Figure 1B, with the help of two labeled primers, DNA amplification products are simultaneously labeled at one end with FITC and at the other end with biotin. And further integrating with the lateral flow strip can lead to the rapid visuable identification of target SNP. Typically, in the presence of the target mutated DNA in the sample, it will bind with the AuNP-modified FITC antibody and the pre-immobilized SAV on the T line of LFS to form the AuNP-amplicon-SAV structure on T line, retaining the AuNPs on the T line and producing the optical signal on T line. On the contrary, with the wild-type or the absence of the mutated genes, no dual-labeled amplicons will be formed and the AuNP-anti-FITC will not be retained on the T line and no signal could be observed. And under any conditions, the AuNP-anti-FITC could be recognized by the second anti-mouse antibody on the C line, showing optical signals on C line and indicating the validity of this detection. With this ARMS-PCR mediated LFS, single nucleotide variation in the target gene could be easily monitored in the visible observation mode, providing the direct results and information for timely response and effective treatment of the mutated strains of E. coli.
3.1. Possibility Verification of the Designed ARMS-PCR Mediated LFS Assay for Precise Visual SNP Identification
The main purpose of this assay is to realize rapid and easy identification of single nucleotide polymorphisms (SNPs) within the eae gene of Escherichia coli 0157:H7 in the visible mode. In general, it displays exceedingly challenging to differentiate alleles with solely PCR primers featuring a single nucleotide alteration at the 3′-end. Therefore, by introducing an additional mismatch at the different position from the 3′-terminus in the eae allele-specific reverse primers, the specificity of the SNP differentiation may be further improved and guaranteed especially in the low mutation abundance. To verify the SNP identification performance with the different primer sets, the wild-type and the mutated-type genes are detected with the real-time PCR. From the real time PCR results in the Figure 2, for the primer R1, R2 and R4, the wide-type genes were all not measured (with Ct>40) while the primer R0 and R3 could not with positive signals for both wild-type and mutated-type genes. In detail, among primer R1, R2 and R4, the primer R1 had the best amplification effect for the mutated-type gene. Therefore, the designed R1 was adopted for the further visuable identification research. Meanwhile, both the agarose gel electrophoresis and lateral flow strip results demonstrated that only the mutant-type gene had the characteristic band of 250 bp in the gel and the positive signal on the T line of LFS, further indicating the technical feasibility of the designed ARMS-PCR mediated LFS platform for rapid and visuable identification of SNP in eae genes of Escherichia coli 0157:H7 Figure S2.
3.3. Visual-Identification Performance of the SNP in the Mutant Genes with the Designed ARMS-PCR-LFS
After verifying the technical feasibility for rapid identification of SNP in the eae genes, some critical parameters of PCR were optimized to get the best amplification effect with the highest identification efficiency for on-site applications. 0.5 µL 10 µM primer for each reaction was adopted for the specific amplification. And the annealing temperature of 60 ℃ was determined as optimal for the balance between specificity and signal strength of target band. To get the most efficient amplification efficiency without any non-specific products, 30 cycle was adopted as the best condition to integrated with lateral flow strip. The mutant-type gene was spiked to the wide-type gene to prepare the samples with various mutation ratio (from 0 to 100%) and then the samples were all detected with the developed ARMS-PCR mediated LFS. From the results in Figure 3, it could be observed that with the increase of mutant ratio from 0 to 100%, the signal on the T line of LFS is increased accordingly. The quantitative optical intensities on T line and C line are further treated with the software as shown in Figure 3B. Then the calibration curve is also constructed based on the quantitative signals on T line of LFS and the detection limit of mutation ratio could be as low as 0.1% according to the 3σ criterion. With this designed primer set and the ARMS-PCR mediated LFS, the specificity against other common non-target bacteria including the Salmonella, Vibrio parahaemolyticus, Staphylococcus aureus, Pseudomonas aeruginosa, and Cronobacter sakazakii was also considered. Results in Figure 4A and B well demonstrate that other common non-target bacteria and the wide-type E.coli O157:H7 with the eae gene could not induce the signals on the T line while only the mutation-type E.coli O157:H7 could induce the obvious observable signal on the T line of LFS. All these results have strongly shown the satisfied detection performance and specificity for the SNP gene samples.
3.4. Practical Detection Research of SNP Mutated Eae Genes of E. coli in Spiked Wild-Type E.coli Samples
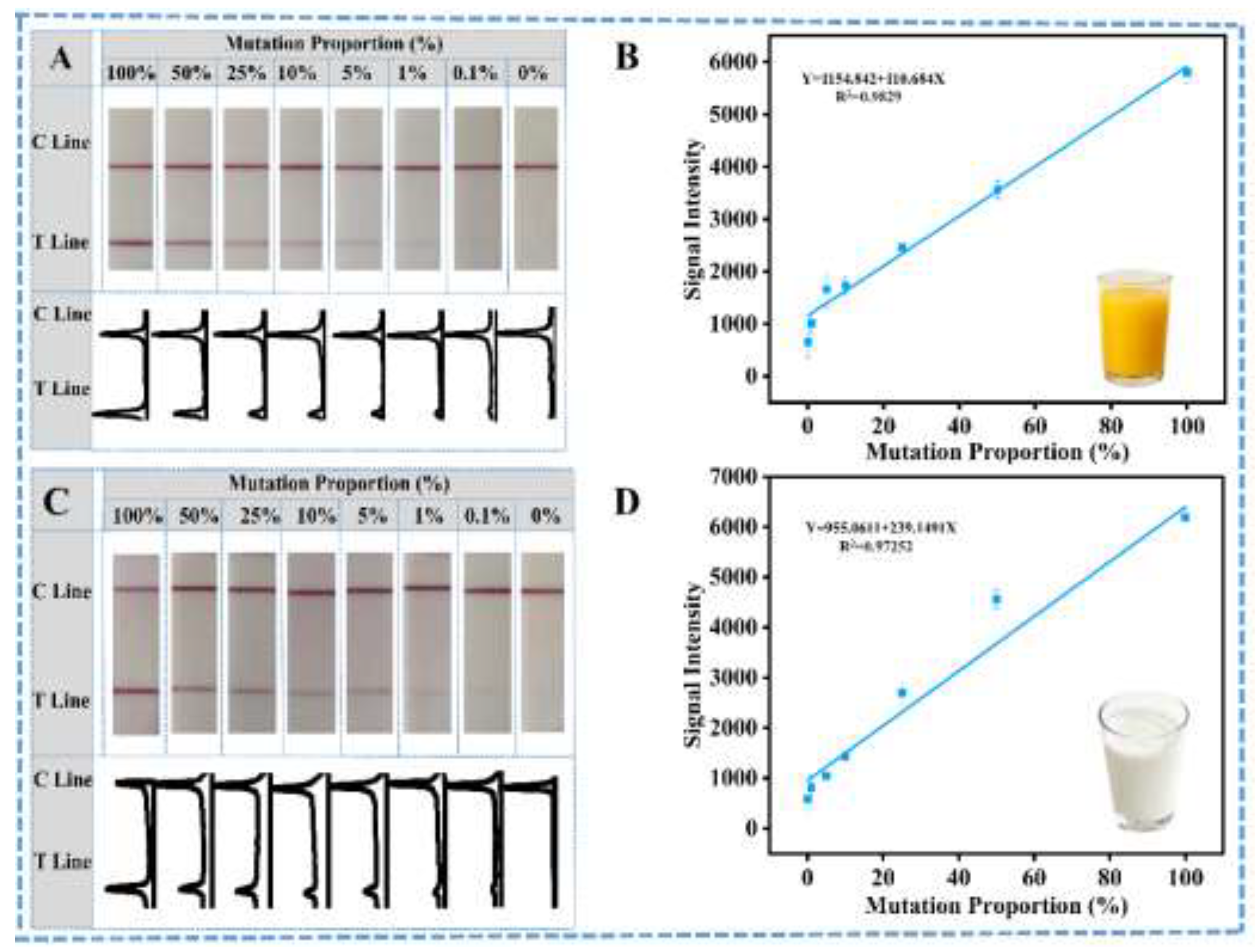
Finally, to further verify the practical application performance of this ARMS-PCR mediated LFS, the mutated eae genes of E. coli was spiked into the samples with different matrix. Then, all samples were detected with the ARMS-PCR mediated LFS and the results are shown in Figure 5. In detail, all spiked mutated eae genes could be well detected and the corresponding calibration curves could be constructed for quantitative analysis for different sample matrix. Comparatively, the detection results of the spiked samples could come to the same conclusion as the standard gene samples. All these results well demonstrate the excellent capability of this ARMS-PCR mediated LFS for rapid and easy identification of SNPs in the visible mode.
The developed ARMS-PCR-LFS has been constructed for rapid and easy identification of SNP in eae genes of E. coli O157:H7 in the visible mode without any additional high-value instruments, which is suitable for on-site or resource-limited settings. The traditional complicated SNP assay is converted into the binary presence/absence signal with the designed ARMS-PCR mediated LFS, highlighting the potential of ARMS-PCR mediated LFS as a powerful alternative for SNP assay. The whole operation process and the final results judgement are also very simple even for non-professional personnel, with the presence of T line on LFS indicating the occurrence of SNP in the target genes. And the cost of each identification is also acceptable for various scenarios with each SNP identification of less than $1 dollar. However, to realize the rapid high throughput screening of potential evolution or mutation of target pathogen for epidemic or clinical precise therapy, simultaneous multiplex screening of SNPs at different sites is of great importance. And to further simplify and avoid the dependence of the device, the isothermal amplification should be adopted for the replacement of PCR. Related research is still ongoing in our lab for practical applications.
4. Conclusions
In this study, the single nucleotide polymorphism (SNP) in the eae gene of E. coli O157:H7 has been well rapid identified in the easy visible mode with the designed ARMS-PCR mediated LFS. With the designed primer set, the difference of single nucleotide in the target genes could be precisely identified. And the identification of SNP could be converted into the simple, visible, yes/no outcome on T line of LFS without any sophisticated instrumentation. The SNPs in the target eae gene has been simply judged: the presence of T line indicates the occurrence of SNP in the target genes while only the C line of LFS indicates no SNP in the specific site of the genes of E. coli O157:H7, making it an ideal candidate for point-of-care and field-deployable SNP diagnostics. The single nucleotide mutated genes has been well identified with the mutation ratio as low as 0.1% by observation and quantitative analysis in even food samples with complicated matrix, strongly demonstrating the great potential for rapid and on-site identification of SNP in the genes of interest. Multiplex screening of different mutated sites and integration with isothermal amplifications will further widen this designed SNP identification platform for practical applications in the surveillance of pathogen evolution and related research is ongoing in our lab. .
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Optimization Conditions of ARMS-PCR-LFS; Figure S2: Detection of Mutant allele by ARMS-PCR followed by LFS; Table S1: Primers and Probe sequences used to detect SNP of eae gene.
Noor Fatima, Liangxia Jiang2, Siying Sun1, Li Yao3, Yubo Peng1, Daoli Chen2,*, Wei Chen
Author Contributions
Conceptualization, D. C. and W. C.; methodology, N.F.; L.J.; S.S.; W.C.; validation, L.J.; D.C.; Y.Y.; investigation, N.F.; S.S.; L.Y.; Y.P.; resources, D.L; W.C.; data curation, N.F.; writing—original draft preparation, N.F.; W.C.; writing—review and editing, W.C.; supervision, D.C.; W.C.; project administration, W.C.; funding acquisition, W.C.. All authors have read and agreed to the published version of the manuscript.”
Funding
This research was funded by the grants received from the National Key Research Program (2022YFF1102900, 2024YFF0618104), the NSFC program (32172295), the National Key Research Program (2023YFF0611500) and the Hainan Key Research Program (ZDYF2022XDNY248).
Data Availability Statement
The data presented in this study are available on request from the corresponding author due to the intellectual property.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
Appendix A.1

Table S1.
Primers and Probe sequences used to detect SNP of eae gene
| Primers and Probe | Primer and probe Sequence | Size(bp) | Method |
| F PCR R1 |
TTGATCAAACCAAGGCCAGC CTAACAGTCGCTTTACCGAA |
250 | ARMS-PCR |
| F LFS R1 |
FITC-TTGATCAAACCAAGGCCAGC Biotin-CTAACAGTCGCTTTACCGAA |
250 | ARMS PCR-LFS |
| F qPCR R1 |
TTGATCAAACCAAGGCCAGC CTAACAGTCGCTTTACCGAA |
250 | ARMS-qPCR |
| P- qPCR | 5’-FAMTCCCGTGGTTGCTTGCGTTTGAGACT-BHQ1 |
F Forward Primer, R Reverse Primer, P Probe
Figure S1.
Optimization Conditions of ARMS-PCR-LFS (a) primer concentration (b) amplification temperature, (c) Number of cycles (d)loading buffer (Running buffer, PB, PBS, Tris-HCL) (e) volume of K2CO3 (0.1M) (f) type of resuspension buffer (Buffer C (10 mM PB, 1% BSA, 0.25% Tween-20, 10% Sucrose), Buffer E (1 mM Tris-HCl, 1% BSA, 0.25% PEG 20000, 10% sucrose), Casein, Casein-Na) (g) Volume of FITC-Ab.
Figure S1.
Optimization Conditions of ARMS-PCR-LFS (a) primer concentration (b) amplification temperature, (c) Number of cycles (d)loading buffer (Running buffer, PB, PBS, Tris-HCL) (e) volume of K2CO3 (0.1M) (f) type of resuspension buffer (Buffer C (10 mM PB, 1% BSA, 0.25% Tween-20, 10% Sucrose), Buffer E (1 mM Tris-HCl, 1% BSA, 0.25% PEG 20000, 10% sucrose), Casein, Casein-Na) (g) Volume of FITC-Ab.
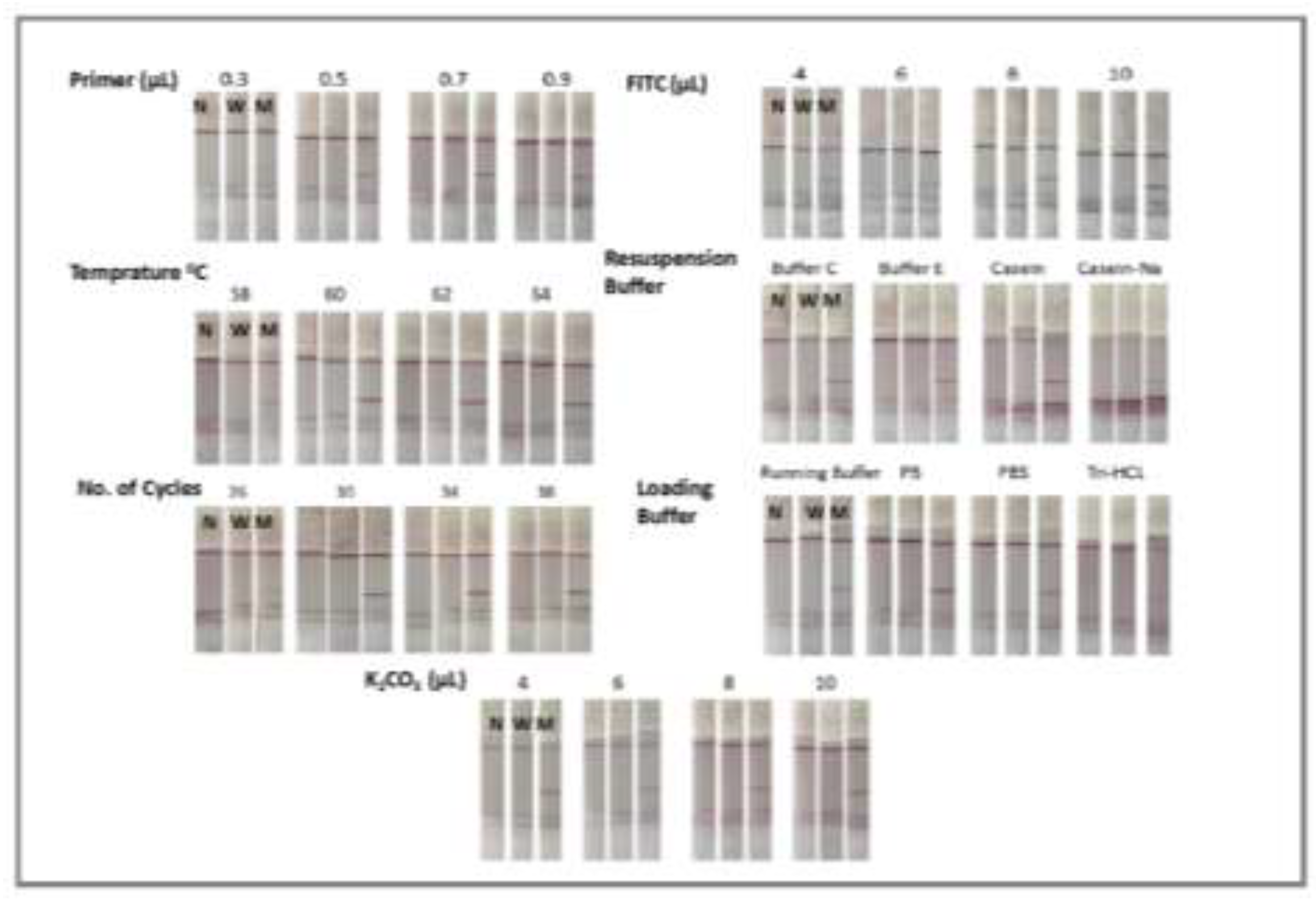
Optimization Experimental Conditions of ARMS-PCR-LFS
We optimized the experimental conditions for LFS to identify the target gene. Primer( 0.3μL ,0.5μL ,0.7μL ,0.9μL ), Temprature (58°C, 60°C, 62°C, 64°C), Cycle numbers (27,30,33,36) FITC-Ab, K2CO3, Type of loading buffer (Running buffer, PB, PBS, Tris-HCL), Type of Resuspension Buffer (Buffer C, Buffer E, Casein, Casein-Na were optimized. The optimal conditions determined for each parameter as shown in Figure S1 (primer 0.5 μL, annealing Temprature 60°C, 30 cycles, FITC 4 μL , K2CO3 10 μL (0.1M), Loading Buffer PBS, Resuspension Buffer E), were used for all subsequent experiments.
Figure S2.
Detection of Mutant allele by ARMS-PCR followed by LFS (A) LFS result, Lane N Negative, Lane Wt Wild gene, Lane M Mutant gene, T: Test line C: Control line, band in lane M shows amplification of mutant gene (B) Gel electrophoresis, mutant sample displaying 250bp band confirm successful amplification of mutant allele.
Figure S2.
Detection of Mutant allele by ARMS-PCR followed by LFS (A) LFS result, Lane N Negative, Lane Wt Wild gene, Lane M Mutant gene, T: Test line C: Control line, band in lane M shows amplification of mutant gene (B) Gel electrophoresis, mutant sample displaying 250bp band confirm successful amplification of mutant allele.

References
- Lee, H.; Yoon, Y. Etiological agents implicated in foodborne illness world wide. Food science of animal resources 2021, 41, 1. [Google Scholar] [CrossRef] [PubMed]
- Kariuki, S. Global burden of antimicrobial resistance and forecasts to 2050. The Lancet 2024, 404, 1172–1173. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Alsayeqh, A.F. Review of major meat-borne zoonotic bacterial pathogens. Frontiers in Public Health 2022, 10, 1045599. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO bacterial priority pathogens list, 2024: Bacterial pathogens of public health importance, to guide research, development, and strategies to prevent and control antimicrobial resistance. 2024: World Health Organization.
- Odo, S.E.; Uchechukwu, C.F.; Ezemadu, U.R. Foodborne diseases and intoxication in Nigeria: Prevalence of Escherichia coli 0157: H7, Salmonella, Shigella and Staphylococcus aureus. J Adv Microbiol 2021, 20, 84–94. [Google Scholar] [CrossRef]
- Bonten, M.; et al. Epidemiology of Escherichia coli bacteremia: A systematic literature review. Clinical Infectious Diseases 2021, 72, 1211–1219. [Google Scholar] [CrossRef]
- Mueller, M.; Tainter, C.R. Escherichia coli infection, in StatPearls [Internet]. 2023, StatPearls Publishing.
- Pokharel, P.; Dhakal, S.; Dozois, C.M. The diversity of Escherichia coli pathotypes and vaccination strategies against this versatile bacterial pathogen. Microorganisms 2023, 11, 344. [Google Scholar] [CrossRef]
- Hasan, J.M.; Najim, S.S. A review of the Prevalence of Enterohemorrhagic E. coli in Iraq. Journal of Biotechnology Research Center 2024, 18, 33–39. [Google Scholar] [CrossRef]
- Heo, N.; et al. The epidemiological and clinical characteristics of the largest outbreak of enterohemorrhagic Escherichia coli in Korea. J. Korean Med. Sci. 2023, 38. [Google Scholar] [CrossRef]
- Shuvra, S. Isolation of Shiga Toxin Producing Escherichia coli 0157: H7 from Environmental and Clinical Samples in Dhaka City—A Review. J Immuno Allerg 2023, 4, 40–62. [Google Scholar] [CrossRef]
- Jiang, L.; et al. Virulence-related O islands in enterohemorrhagic Escherichia coli O157: H7. Gut microbes 2021, 13, 1992237. [Google Scholar] [CrossRef]
- Liu, Q.; et al. Genetic diversity and expression of Intimin in Escherichia albertii isolated from humans, animals, and food. Microorganisms 2023, 11, 2843. [Google Scholar] [CrossRef]
- Perraud, Q.; Sperandio, V. Enterohemorrhagic E. coli (EHEC) and the microbiome. PLoS pathogens 2025, 21, e1013224. [Google Scholar] [CrossRef] [PubMed]
- Rosauer, M.L.; et al. Validation of the 3M™ Molecular Detection Assay 2-STEC Gene Screen (stx and eae) for the Detection of Shiga Toxin Gene (stx and eae) in Fresh Raw Beef Trim, Fresh Raw Ground Beef and Fresh Spinach: AOAC Performance Tested Method SM 071902. J. AOAC Int. 2022, 105, 902–914. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; et al. Molecular characteristics of eae-positive clinical Shiga toxin-producing Escherichia coli in Sweden. Emerging microbes & infections 2020, 9, 2562–2570. [Google Scholar] [CrossRef]
- Rahman, M.-M.; Lim, S.-J.; Park, Y.-C. Development of single nucleotide polymorphism (SNP)-based triplex PCR marker for serotype-specific Escherichia coli detection. Pathogens 2022, 11, 115. [Google Scholar] [CrossRef]
- Cheng, C.; Fei, Z.; Xiao, P. Methods to improve the accuracy of next-generation sequencing. Frontiers in bioengineering and biotechnology 2023, 11, 982111. [Google Scholar] [CrossRef]
- Yang, H.; et al. Sensitive detection of a single-nucleotide polymorphism in foodborne pathogens using CRISPR/Cas12a-signaling ARMS-PCR. Journal of Agricultural and Food Chemistry 2022, 70, 8451–8457. [Google Scholar] [CrossRef]
- Han, Y.; Wu, H. Fluorescent primers amplification refractory mutation system qPCR (FP ARMS-qPCR) for MTHFR C677T SNP genotyping. Molecular Biology Reports 2024, 51, 1122. [Google Scholar] [CrossRef]
- Muneeswaran, K.; et al. Genotyping SNPs and indels: A method to improve the scope and sensitivity of high-resolution melt (HRM) analysis based applications. Clinica Chimica Acta 2024, 562, 119897. [Google Scholar] [CrossRef]
- Kishi, J.Y.; et al. SABER amplifies FISH: Enhanced multiplexed imaging of RNA and DNA in cells and tissues. Nature methods 2019, 16, 533–544. [Google Scholar] [CrossRef]
- Dardani, I.; et al. ClampFISH 2.0 enables rapid, scalable amplified RNA detection in situ. Nature methods 2022, 19, 1403–1410. [Google Scholar] [CrossRef]
- Liu, X.; et al. In situ Cas12a-based allele-specific PCR for imaging single-nucleotide variations in foodborne pathogenic bacteria. Analytical Chemistry 2024, 96, 2032–2040. [Google Scholar] [CrossRef]
- Islam, M.T.; et al. A rapid and cost-effective multiplex ARMS-PCR method for the simultaneous genotyping of the circulating SARS-CoV-2 phylogenetic clades. Journal of medical virology 2021, 93, 2962–2970. [Google Scholar] [CrossRef]
- Mesrian Tanha, H.; et al. Modified tetra-primer ARMS PCR as a single-nucleotide polymorphism genotyping tool. Genetic testing and molecular biomarkers 2015, 19, 156–161. [Google Scholar] [CrossRef]
- Khehra, N.; Padda, I.S.; Swift, C.J. Polymerase chain reaction (PCR). 2023.
- Kinyua, D.M.; et al. Advancements and Applications of Lateral Flow Assays (LFAs): A Comprehensive Review. Sensors 2025, 25, 5414. [Google Scholar] [CrossRef]
- Zhao, L.; et al. A PCR-based lateral flow assay for the detection of Turkey ingredient in food products. Food Control 2020, 107, 106774. [Google Scholar] [CrossRef]
- Li, B.; et al. Filtration assisted pretreatment for rapid enrichment and accurate detection of Salmonella in vegetables. Food Science and Human Wellness 2023, 12, 1167–1173. [Google Scholar] [CrossRef]
Figure 1.
Schematic illustration of ARMS-PCR combined with LFS for distinguishing between Wild type and Mutant gene of E.coli 0157:H7 with single-nucleotide resolution.
Figure 1.
Schematic illustration of ARMS-PCR combined with LFS for distinguishing between Wild type and Mutant gene of E.coli 0157:H7 with single-nucleotide resolution.
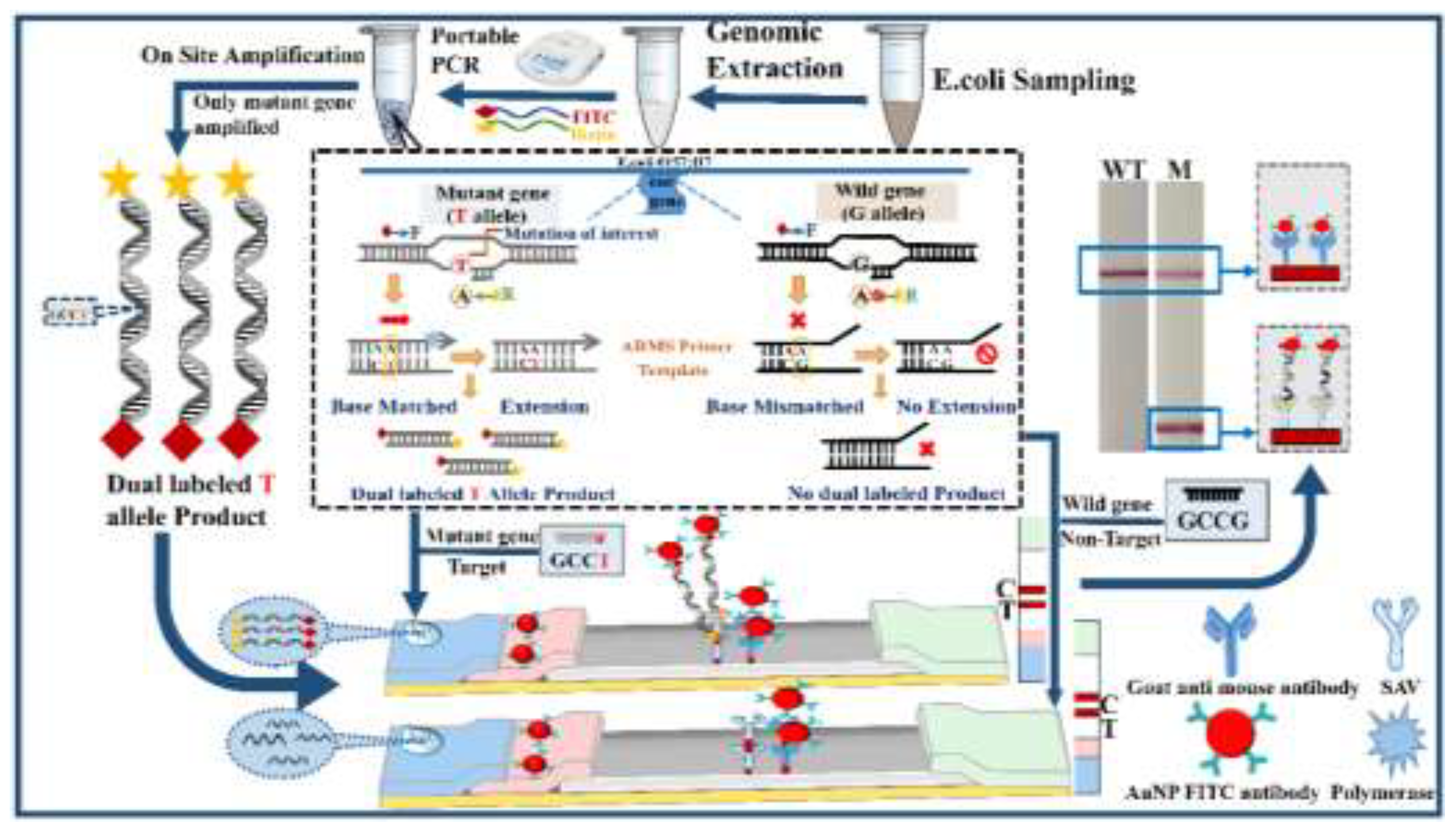
Figure 2.
ARMS-PCR primers Optimization. (A) ARMS-PCR assay methodology for identifying wild-type (WT) and mutant (MT) DNA sequences. (B) Nucleotide sequence of the R primer, Mismatched base locations are marked in red. (C) Comparison of Ct values for mutant (MT) and wild-type (WT) genes. (D) qPCR curves of WT and MT genes using different R primers.
Figure 2.
ARMS-PCR primers Optimization. (A) ARMS-PCR assay methodology for identifying wild-type (WT) and mutant (MT) DNA sequences. (B) Nucleotide sequence of the R primer, Mismatched base locations are marked in red. (C) Comparison of Ct values for mutant (MT) and wild-type (WT) genes. (D) qPCR curves of WT and MT genes using different R primers.
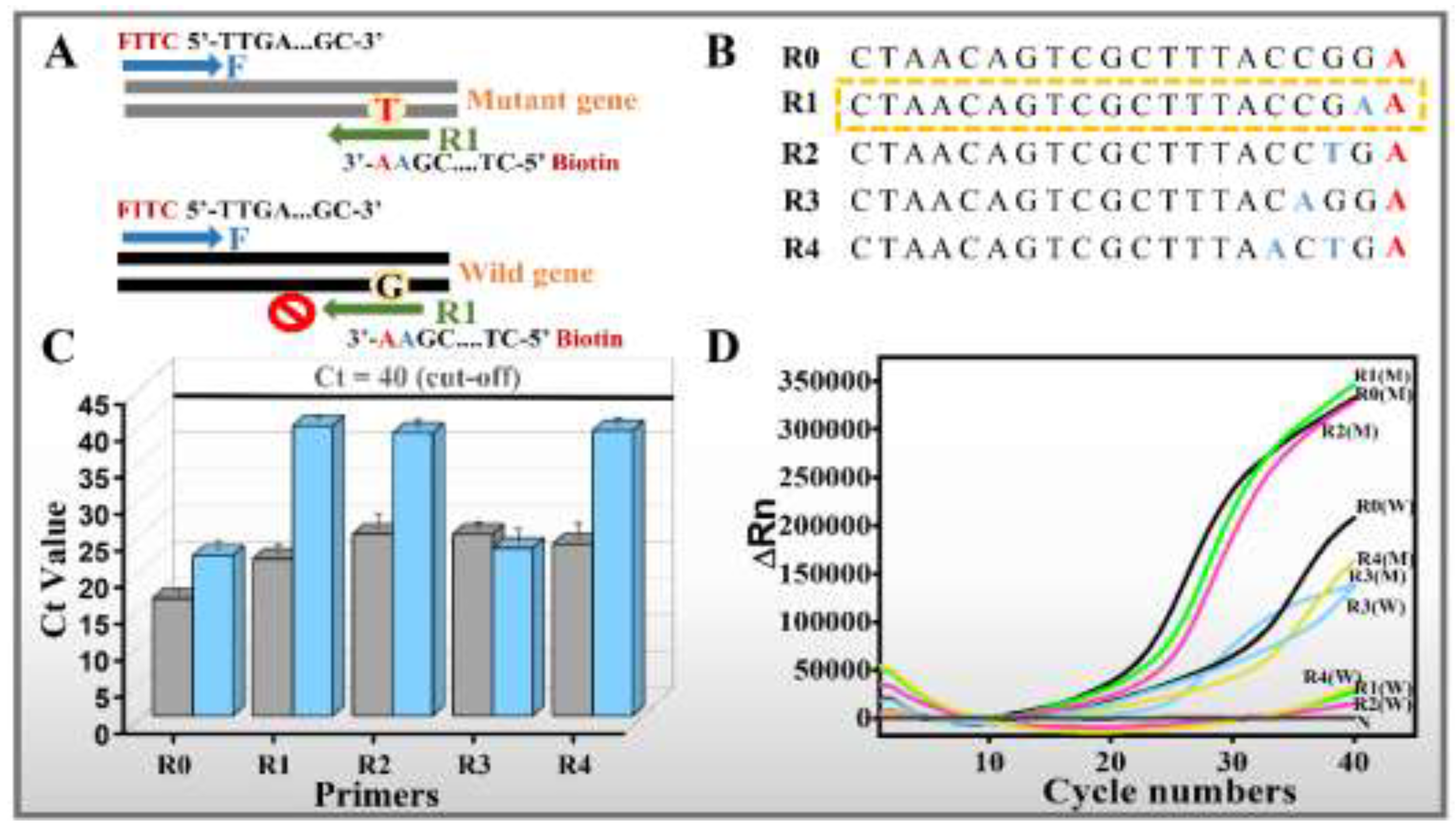
Figure 3.
Authentication performance of ARMS PCR LFS (A) Sensitivity evaluation using different concentrations of mutant DNA (B) Linear correlation between T-line signal intensity and E.coli concentration.
Figure 3.
Authentication performance of ARMS PCR LFS (A) Sensitivity evaluation using different concentrations of mutant DNA (B) Linear correlation between T-line signal intensity and E.coli concentration.
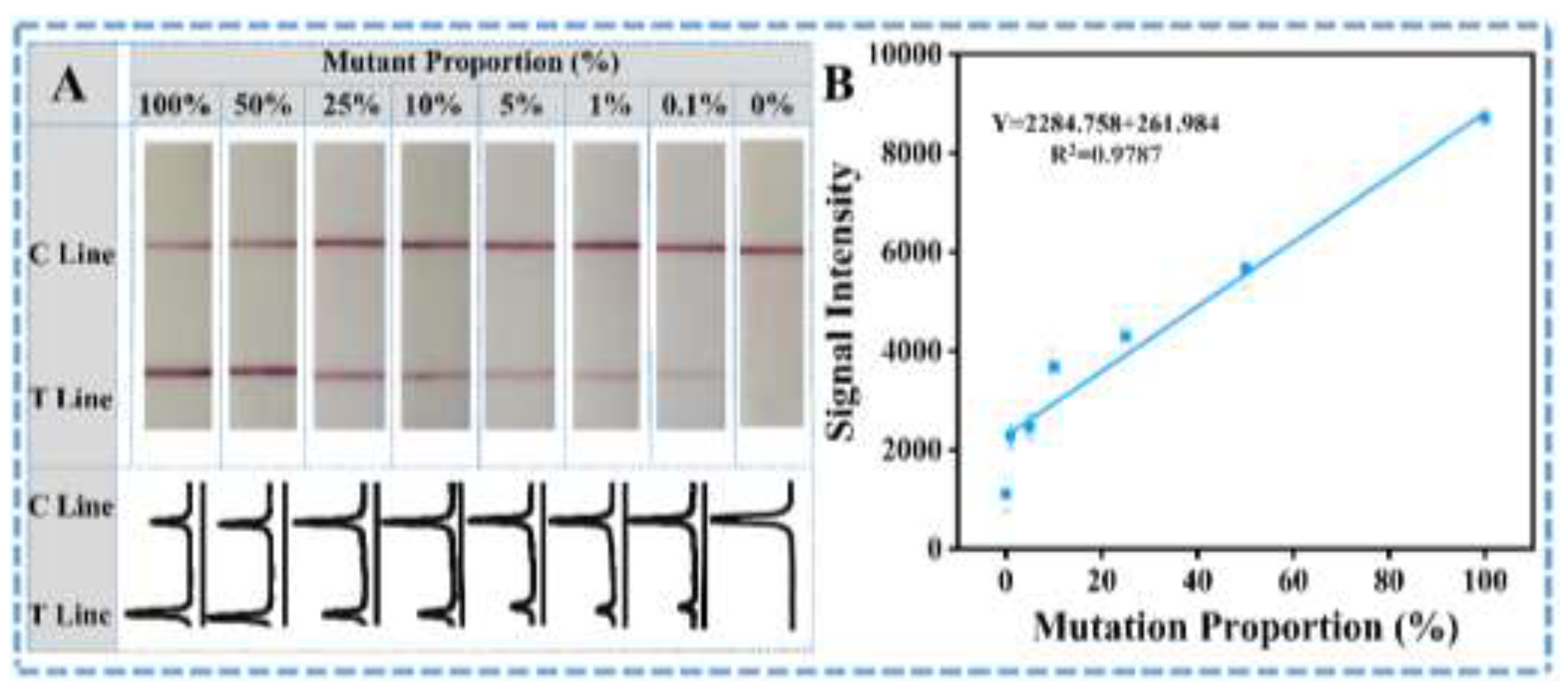
Figure 4.
Selectivity assay results of amplification assisted molecular LFS (A) Visual observation result of amplification assisted molecular LFS and corresponding curves of ImageJ treated results (B) Quantification of T line intensity induced by different analytes using software ImageJ.
Figure 4.
Selectivity assay results of amplification assisted molecular LFS (A) Visual observation result of amplification assisted molecular LFS and corresponding curves of ImageJ treated results (B) Quantification of T line intensity induced by different analytes using software ImageJ.
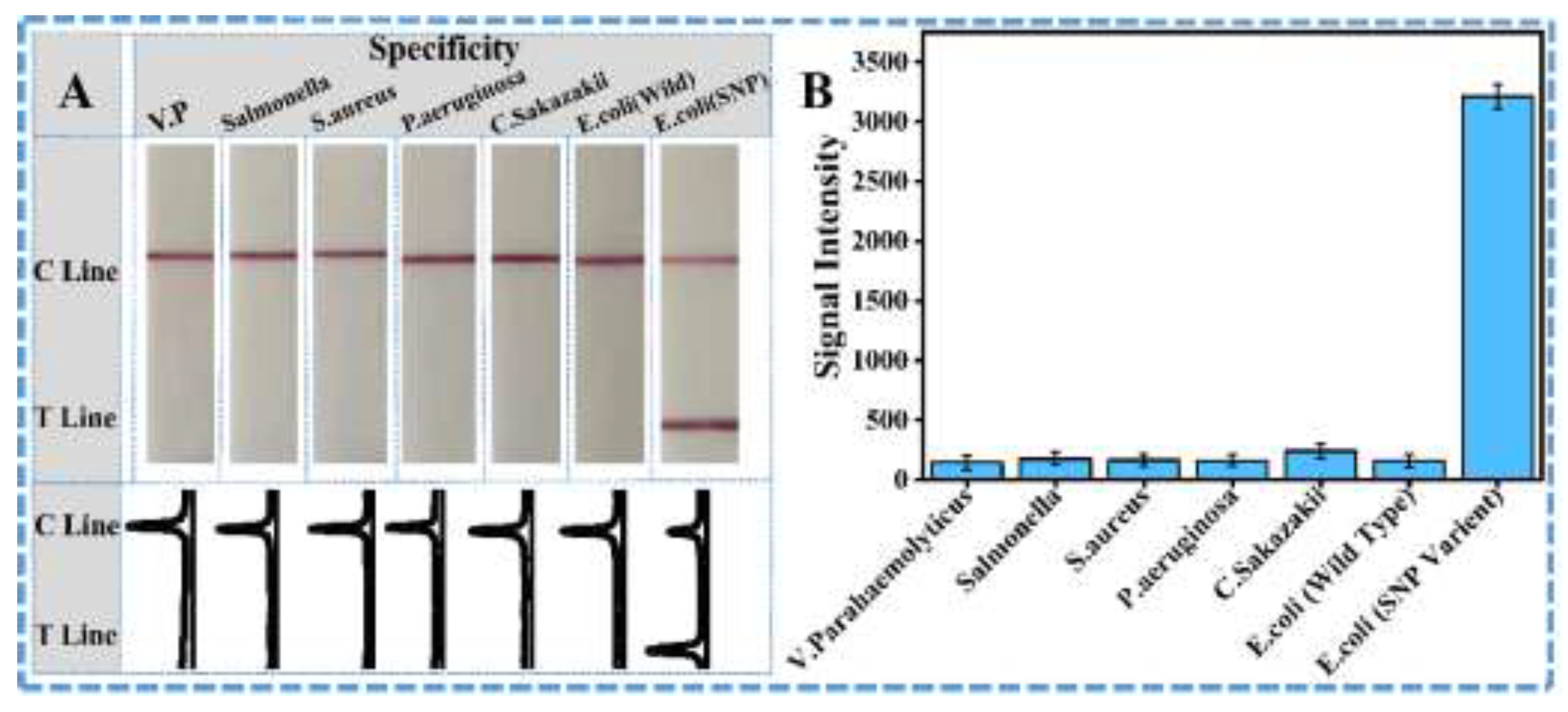
Figure 5.
Detection of E.coli in real food samples (A) Sensitivity evaluation using juice samples spiked with different ratio of mutant DNA and corresponding curves of ImageJ treated results (B) Linear correlation between T-line signal intensity and E.coli concentration in juice sample (C) Sensitivity evaluation using Milk samples spiked with different ratio of mutant DNA and corresponding curves of ImageJ treated results (D) Linear correlation between T-line signal intensity and E.coli concentration in Milk sample.
Figure 5.
Detection of E.coli in real food samples (A) Sensitivity evaluation using juice samples spiked with different ratio of mutant DNA and corresponding curves of ImageJ treated results (B) Linear correlation between T-line signal intensity and E.coli concentration in juice sample (C) Sensitivity evaluation using Milk samples spiked with different ratio of mutant DNA and corresponding curves of ImageJ treated results (D) Linear correlation between T-line signal intensity and E.coli concentration in Milk sample.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



