
Submitted:
23 December 2025
Posted:
24 December 2025
You are already at the latest version
Abstract
Antimicrobial resistance (AMR) is a global threat to human, animal and environmental health, underscoring the need for integrated surveillance to understand its dynamics and ecosystem interactions. This study investigated the potential of swifts (Apus spp.), long-distance migratory birds, as valuable bioindicator of environmental AMR dissemination. Four sampling sessions were conducted over two years (2023-24) at a wildlife rehabilitation center in Trieste, Italy. Buccal and cloacal swabs were collected from 47 swifts: 10 sampled at arrival and 37 before autumn migration. Swabs were streaked on selective media for targeted isolation of Enterobacterales, Bacillales and Lactobacillales, yielding 168 bacterial isolates. Bacteria were identified using MALDI-TOF and antimicrobial susceptibility was assessed through disk diffusion method. Of the 168 bacterial isolates, 51 (30.36%) were non-wild type (NWT), with highest per-centages of NWT isolates for clarithromycin (33.33%), erythromycin (31.50%), clindamycin (21.88%), and tetracycline (14.29%). Methicillin-resistant staphylococci (45.83%) and carbapenem NWT isolates (9.38%) were also detected. Bacillales isolates showed significantly higher NWT proportion (58.33%; p < 0.0001), compared to Enter-obacterales and Lactobacillales. These findings, in clinically healthy non-antimicrobial treated swifts, suggest environmental exposure to resistant bacteria, and support a possible role of swifts as bioindicators of environmental AMR contamination, high-lighting the need to strengthen environmental AMR surveillance within a One Health perspective.
Keywords:
antimicrobialresistance
; wildlife
; swifts
; wildbirds
; migratorybirds
; AMR
; resistantbacteria
; Bacillales
; Enterobacterales
; Africa
; environment
; surveillance
1. Introduction
Antimicrobial resistance (AMR) constitutes a natural adaptive mechanism that allows microorganisms, including bacteria, fungi, parasites and viruses, to survive in the presence of antimicrobial agents [1]. Nowadays, AMR represents a critical global threat to both human, animal and environmental health. It is estimated that by 2050, AMR could potentially become the primary cause of death worldwide, since the number of deaths directly linked to AMR has risen to more than 1.2 million in 2019 and are forecast to increase to approximately 10 million per year by 2050 [2]. Major drivers of AMR include the overuse or misuse of antimicrobials, mainly in human and veterinary medicine [3]. Moreover, external pressures such as intensification of industrial activities and rapid urbanization can create selective pressures in the environment and facilitate the spread of resistant strains, contributing to the dissemination of resistance genes in bacterial populations and ecosystems [4]. In response to AMR, mitigation strategies recognizing the interdependence of human, animal and environmental health are needed, as highlighted by the global “One Health” approach [5]. Environmental monitoring of AMR has been recognized as crucial to understand the resistance diffusion mechanisms, the factors that influence the phenomenon and the interactions between all the compartments of the ecosystem [6]. Numerous studies have emphasized the importance of AMR surveillance also in wildlife, as wild animals are not conventionally exposed to antimicrobials, but the presence of resistant bacterial strains in their microbiota could be closely related to the presence of AMR in their surrounding ecosystems. Wild animals are increasingly studied, considered sentinels, reservoirs and vectors of AMR at the human–animal–environment interface. Wild migratory birds are of particular interest in this context, since they could exhibit long-range movements, potentially acting as silent vectors between different geographical regions, and easily being exposed to resistant bacteria and antimicrobial resistance genes (ARGs), making them possible good bioindicators of AMR environmental contamination [7]. Among migratory species, the swift (order Apodiformes, family Apodidae, genus Apus) is considered a long-distance migratory bird, moving from sub-Saharan Africa during wintering period over to Europe for breeding [8]. Specifically, in Europe, its arrival is in early spring, spending the breeding period in colonies in urban and peri-urban areas and raising their young before departing in late summer [8,9]. Despite their unique potential resulting from their migration routes and ecological behaviors to the best of our knowledge, no studies have yet investigated their role as indicators of the spread of AMR. Hence, this research aimed at two main objectives: (i) to investigate the antimicrobial resistance patterns of selected commensal bacterial orders (Enterobacterales, Bacillales and Lactobacillales) isolated from swifts; and (ii) to assess the potential role of swifts as bioindicator of environmental AMR.
2. Materials and Methods
2.1. Population Sampling and Data Collection
A total of four sampling sessions were conducted over two consecutive years (2023-2024) during two distinct moments corresponding to the swift’s migratory cycle: (i) at the arrival of swifts in Italy (July 2023; June 2024), and (ii) in late summer (September 2023; August 2024), after the breeding season and before autumn migration. According to the sampling period, swifts were classified in two groups to evaluate possible differences in AMR carriage before and after the breeding season. Moreover, age distinction was also assigned: adult swifts at the arrival in Italy, adult swifts after the breeding season, and juveniles swifts born in Italy. Samples were collected at the wildlife rehabilitation center “Liberi di Volare” in Trieste (Friuli-Venezia Giulia, Italy), where the swifts were brought for their recovery and sampled at least few days after. Only clinically healthy birds that did not receive any antibiotic treatment in the previous 90 days were included in the study. A buccal and cloacal swab was performed on each bird using Copan ESwab® with Amies liquid transport medium (Copan Italia S.p.A., Brescia, Italy). For each individual, a comprehensive data form was filled out recording essential data relating to the bird’s signalment, such as bird’s identifier name, species classification (common swift/pallid swift/alpine swift), age (adult, nestling or fledgling), sex (male/female), as well as sampling date. After sampling, the individuals were released back into the wild; therefore, it was not possible to re-sample the same birds at both sampling periods. The swabs were sent under refrigeration conditions within 24 h to the Veterinary Bacteriology Laboratory (VeLaBac) of the Department of Veterinary Medical Sciences (University of Bologna), and processed within 24-48 h.
2.2. Bacterial Isolation and Identification
Ten microliters (10 μL) of the Amies liquid transport medium from cloacal swabs were streaked onto MacConkey agar (Oxoid, Basington, UK) for selective isolation of Enterobacterales, while 10 μL from oral swabs were streaked onto Mannitol Salt Agar (Oxoid, Basington, UK) for selective growth of Bacillales and Lactobacillales. All plates were incubated aerobically at 40 ±1 ◦C overnight, a temperature selected to approximate the birds’ physiological body temperature and to favor the growth of their commensal bacteria. Following 24–48 h of incubation, the colonies isolated on the different agar plates were evaluated based on their morphology and lactose/mannitol fermentation, cloned on Tryptone Soy Agar (Oxoid, Basington, UK) and incubated aerobically at 37 ±1 °C overnight. After 24 h, the identification at species level of each isolate was assessed using the matrix-assisted laser desorption–ionization time- of-flight mass spectrometry method (MALDI-TOF MS) (Biotyper, Bruker Daltonics, Billerica, MA), following manufacturer’s instructions (Bruker Daltonik, GmbH, Bremen, Germany) and considered a species-level identification when the ID score was > 1.80 using the MALDI Biotyper version 3.0 software. Correctly identified isolates were stored in cryovials containing 1 mL of Tryptic Soy Broth with 20% glycerol at -80 ◦C until further analysis.
2.3. Antibiotics Susceptibility Testing (AST)
All isolates were tested for antibiotic susceptibility using the agar disc diffusion method (Kirby-Bauer), following EUCAST guidelines [10]. Enterobacterals and Lactobacillales were tested against 7 different antimicrobials belonging to 6 different antimicrobials classes; Bacillales were tested against 10 antimicrobials from 9 classes. For Staphylococcus spp., cefoxitin was also tested to screen for methicillin resistance [11]. All the discs were purchased from Oxoid (Oxoid S.p.A., Milan, Italy). Antimicrobials tested are listed in Table 1.
For the interpretation of inhibition zone diameters, each isolate was classified into wild-type (WT) or non-wildtype (NWT) using the standardized epidemiological cut-off values (ECOFFs) from EUCAST as reference values. This evaluation was carried out for the antimicrobials with published ECOFF value for the investigated bacterial genera. Isolates were considered WT when susceptible to all tested antimicrobial, and NWT when indicating the presence of phenotypically detectable acquired resistance mechanisms to at least one tested antimicrobial. Moreover, isolates showing no inhibition zone (diameter < 6 mm, indicating bacterial growth extending close to the edge of the disk without a discernible inhibition zone [12]) for the tested antimicrobials were classified as NWT, given clear evidence of phenotypic resistance, even in absence of ECOFF values. For Staphylococcus spp. isolates, interpretation of cefoxitin inhibition diameters followed the 2023 EUCAST clinical breakpoints [13] and isolates were subsequently classified as cefoxitin susceptible (S) or resistant (R) [14]. Intrinsic resistances for each bacterial species were excluded from AST evaluation, in accordance with EUCAST (2023) guidelines on expected resistance phenotypes [15].
2.4. Molecular Analysis
Staphylococcus spp. isolates exhibiting phenotypically cefoxitin resistance were subjected to molecular analysis by a multiplex PCR to detect methicillin resistance genetic determinants. Bacterial DNA was extracted using a standard boiling method. The isolates were screened for the presence of mecA and mecC genes, according to the protocol described by Stegger et al. (2012) [16]. Staphylococcus aureus ATCC 43300 (mecA-positive) and Staphylococcus aureus BAA-2313 (mecC-positive) were used as positive control, while a Staphylococcus spp. strain, previously confirmed to be negative for both investigated genes, and sterile water, were used as negative controls. Primers and expected amplicon sizes for the investigated genes are detailed in Table 2.
2.5. Quality Control
To ensure the reliability of incubation conditions, culture media performance, as well as the quality and accuracy of AST procedures, American Type Culture Collection (ATCC) standard reference E. coli ATCC 25922, Staphylococcus aureus ATCC 25923 and Staphylococcus aureus ATCC 43300 strains were used to verify test performances.
2.6. Statistical Analysis
Descriptive statistical analysis was performed to describe the occurrence of antimicrobial resistance (AMR), including overall percentages of NWT distribution and resistance to single antimicrobials, within the sampled population and their bacterial isolates. Specifically, swifts were grouped by age (adults at the arrival/adults at the end of breeding/juveniles) and sampling period (arrival/end of breeding period), and this classification was also applied to their corresponding bacterial isolates. Associations between the outcome variables (single antimicrobial tested and NWT proportions) and the included variables (birds’ age and sampling period) were assessed using Fisher’s test or Pearson’s chi-squared (χ2) test. Differences in AMR prevalence between bacterial orders were also statistically evaluated using the Fisher’s exact test. A p-value of ≤ 0.05 was considered statistically significant. Statistical analyses were conducted with EasyMedStat® (version 3.29).
3. Results
3.1. Population Sampling and Data Collection
A total of 47 swifts were sampled across the four sampling sessions. Of these, 20 were identified as common swifts (Apus apus), 2 as pallid swifts (Apus pallidus), and 1 as alpine swift (Tachymarptis melba), while species identification could not be determined for the remaining 24 birds. Sex determination revealed 15 female, 29 male, and 3 individuals of undetermined sex. Age classification showed a total of 13 adult individuals and 34 juveniles (including 3 fledging and 31 nestlings).
Ten adult swifts were sampled shortly after their arrival in Italy: 6 sampled in July 2023 and 4 in June 2024. Thirty-seven swifts were sampled at the end of the breeding season, before leaving for autumn migration. Specifically, 3 adult swifts were sampled in September 2023 and 34 juveniles’ swifts sampled as follows: 11 in July 2023 (1 fledgling and 10 nestlings), 5 nestlings in September 2023, 6 nestlings in June 2024, and 12 in August 2024 (2 fledglings and 10 nestlings).
3.2. Bacterial Isolation and Identification
From the 47 sampled birds, a total of 168 bacterial isolates were obtained. These comprised 71/168 (42.26%) Enterobacterales, 49/168 (29.17%) Lactobacillales and 48/168 (28.57%) Bacillales. A total of 10 different bacterial genera and 24 distinct bacterial species were identified (Figure 1). The most frequently identified species were Enterococcus faecalis (n = 45; 26.79%), Escherichia coli (n = 28; 16.67%), Mammaliicoccus sciuri (n = 14; 10.12%), and Staphylococcus gallinarum (n = 13; 7.74%). The distribution of isolates by bacterial order among swifts at the two sampling points is presented in Table 3.
3.3. Antibiotics Susceptibility Testing (AST)
Of the 168 bacterial isolates recovered from all sampled swifts, 117 (69.64%) were classified as WT, whereas 51 (30.36%) were NWT. The prevalence of NWT isolates emerged particularly among Bacillales isolates (28/48; 58.33%), followed by Lactobacillales (9/49; 18.37%) and Enterobacterales (13/71; 18.31%). Non-wild type isolates were detected towards all antimicrobial tested drugs, with highest percentages towards clarithromycin (33.33%), erythromycin (31.50%), clindamycin (21.88%), tetracycline (14.29%) and ertapenem (9.38%). Moreover, 55.6% of isolates were classified as resistant to cefoxitin. A summary of NWT proportion for each tested antimicrobial drug from swifts at both sampling points is provided in Figure 2.
3.3.1. Comparison of NWT Proportions Across Sampling Periods and Age Classes
A comparative analysis of NWT profiles associated with both sampling periods (at arrival/after breeding season) and the age class (adults/juveniles) revealed notable differences, summarized in Table 4.
When all isolates were considered, those from swifts sampled upon their arrival exhibited NWT proportion of 32.22%, while those from swifts sampled before leaving for autumn migration showed NWT of 26.92% (Figure 2). At the arrival, higher NWT percentages were observed for tetracycline (15.56%), rifampicin (8.47%), and ertapenem (9.36%); whereas 58.1% of isolates were resistant to cefoxitin. Conversely, isolates obtained from swifts before migration showed a higher number of NWT isolates to erythromycin and clarithromycin (44.44%), clindamycin (43.75%) and enrofloxacin (11.54%).
When considering only adult swift isolates, those sampled at their arrival showed 33.33% of NWT prevalence, with higher percentages to rifampicin (5.26%), compared with adults sampled before the autumn migration which showed similar NWT prevalence (38.46%), but with an higher percentage towards enrofloxacin (23.08%).
Age-related comparisons highlighted similar overall NWT proportions between adults and juveniles. Nonetheless, juvenile isolates exhibited higher NWT percentages to several antibiotics, particularly rifampicin (5.63%), ertapenem (8%), and 54.3% were resistant to cefoxitin, mirroring the pattern observed in adults sampled at their arrival. Likewise, percentages of NWT isolates to clarithromycin (34.21%) and erythromycin (31.60%) resemble that of adults sampled before migration. Moreover, low percentages for ampicillin (4.85%) and amoxicillin-clavulanic acid (3.45%) were observed only in juveniles.
3.4. Molecular Analysis
Among the 48 Staphylococcus spp. isolates, 25 (52.08%) were phenotypically resistant to cefoxitin. From them, 22 (88%) carried the mecA gene, as confirmed by multiplex PCR, whereas the mecC gene was not detected in any of them. Notably, most mecA positive isolates were more frequently from swifts at their arrival (60%), compared to those sampled before migration (16%).
3.5. Statistical Analysis
Univariable statistical analysis revealed a significant association between the prevalence of NWT isolates and bacterial order. Bacillales isolates showed a significantly higher proportion of NWT isolates compared with Enterobacterals and Lactobacillales, both from adults and juveniles (p < 0.0001) and at both sampling points (p < 0.0001).
4. Discussion
In the present study, we investigated the presence of NWT bacteria, indicative of acquired phenotypical resistance mechanisms, within selected bacterial orders (Enterobacterales, Bacillales and Lactobacillales) of the commensal microbiota of swifts (Apus spp.), a long-distance migratory species covering migration areas across Africa and Europe, to evaluate their role as bioindicator of environmental AMR. Despite growing attention to the role of wild migratory birds as bioindicator of environmental AMR contamination within the One Health framework, studies specifically exploring the role of swifts as valuable bioindicators of the spread of AMR in natural ecosystems are still lacking. Swifts have several ecological traits that already make them sensitive indicators: they are long-lived species (up to 12 years), breeding in colonies in urban areas, and feeding exclusively on aerial insects and airborne particles [17]. Previous studies have already demonstrated their role in biomonitoring environmental quality, including the accumulation of persistent organic contaminants and airborne particulates [18].
From our findings, the commensal microbiota of sampled swifts, within the bacterial orders targeted in this study, resulted predominantly composed by Enterococcus faecalis and Escherichia coli, the most frequently recovered bacteria species. This is consistent with previous studies reporting these taxa as ubiquitous bacteria inhabiting various environmental habitats and commonly found as commensals in mammals, birds, reptiles and invertebrates [19,20]. However, our result is mainly attributable to the choice of using selective media, such as MacConkey agar for Enterobacterales, that contributed to the predominant isolation of E. coli, a dominant commensal in the intestinal microbiota of wild birds; and the use of Mannitol Salt Agar which favors the growth of Bacillales and Lactobacillales, like Enterococcus spp. and staphylococci. Among these, coagulase-negative staphylococci (CoNS), particularly Mammaliicoccus sciuri and Staphylococcus gallinarum, were the most frequently recovered, aligning with previous observations reporting that birds are more frequently colonized by CoNS than mammals [21]. M. sciuri (formerly known as Staphylococcus sciuri) has been described as one of the most prevalent CoNS in avian species and widely distributed in nature [22], while S. gallinarum, although less frequently reported, has also been isolated from various animal species, including birds [21].
Overall, 30.36% of isolates revealed a non-wild type (NWT) phenotype, indicative of the occurrence of phenotypically detectable acquired resistance. The levels of antimicrobial resistance reported in wild birds vary widely across studies, reflecting heterogeneous available data. Similar prevalence has been documented, such as in a research from Spain reporting AMR rate of 47.4% in white storks and seagulls [23]. In comparison with our study, also higher AMR prevalence has been described in other migratory species, such as in the study by Akhil Prakash et al. (2022), which reported AMR rates of 94% in the commensal microbiota of Arctic terns [24]. However, in our study the prevalence of an acquired resistance may have been underestimated. This may be partly due to the interpretative criteria applied, which were based primarily on ECOFFs, but for several bacterial species and antimicrobials tested specific ECOFF values are currently unavailable, leading to the inclusion of no inhibition zone isolates within NWT category. This adoption should be considered for future environmental and non-clinical bacterial isolates to significantly enhance the AMR assessment and the detection of acquired resistance circulation in the context of environmental monitoring.
Specifically, in our study, a higher NWT prevalence was observed among Bacillales (58.33%), compared to Lactobacillales (18.37%) and Enterobacterales (18.31%). This finding was supported by statistical analysis, which confirmed a significant association between NWT proportion and Bacillales isolates from swifts sampled at both two sampling periods and both adults and juveniles (p < 0.0001). Notably, it has been widely reported that Staphylococcus spp. possesses a broad host range and strong environmental adaptability, allowing them to persist long-term in diverse habitats where antibiotic pressure may drive the selection and spread of resistant strains, potentially making them indicators of environmental contamination [7,21].
Among all bacterial isolates, the higher percentages of NWT phenotypes were observed for clarithromycin (33.33%), erythromycin (31.50%), clindamycin (21.90%), and tetracycline (14.29%); whereas 55.6% isolates were resistant to cefoxitin. Moreover, particularly concerning was the detection of NWT isolates even to last-resort antimicrobials, such as ertapenem (5.04%), and the presence of critical resistance determinants like mecA gene (76%). Carbapenem resistance represents a global critical issue, as these antibiotics represent one of the last therapeutic options against multidrug-resistant bacteria. Although typically associated with Enterobacterales [25], in our study carbapenem NWT phenotypes emerged largely among Staphylococcus isolates, possibly reflecting selective pressures and the circulation of resistant strains even within Gram-positive bacteria Indeed, it has been reported carbapenem resistance among multi-drug-resistant Staphylococcus spp., particularly methicillin-resistant staphylococci [26]. In our study, the high prevalence of cefoxitin resistance, combined with the genotypic detection of the mecA gene, confirmed the presence of methicillin-resistant staphylococci (MRS) among our isolates, raising questions about the possible acquisition of such critical resistance genes. Similar research of Abdullahi et al. (2023) reported methicillin resistance in 100% of S. aureus from migratory white storks, attributing it to nest exposure to human residues, considered a risk factor for AMR acquisition [27]. This finding supports a possible explanation to our result, considering the swifts’ habit of nesting in strictly urban environments. However, the detection of carbapenem resistance and MRS, both representing major concern in human and veterinary medicine, underscores the need for continued environmental surveillance of AMR and a deeper investigation of the role of migratory birds as potential sentinels or reservoirs of critical resistance determinants at human-wildlife-environment interface.
Our findings primarily aim to highlight the prevalence of AMR within the targeted commensal bacterial orders of swifts, but beyond the overall AMR prevalence comparative insights emerged regarding acquired antimicrobial resistance patterns among isolates from swifts sampled shortly after the arrival in Italy, swifts that had possibly spent the breeding season in Italy, and juveniles’ swifts born in Italy. Swifts, as long-distance migratory birds spending most of their active time flying and moving between different areas of the world as part of their annual life cycle [28], may be able to provide significant insights on AMR contamination even across distinct geographical areas. Understanding the swift’s migratory cycle is fundamental to evaluating their exposure to different ecosystems. So far, migration routes and exact wintering areas of swifts remain poorly defined due to limited ringing recoveries; but, it has been recently reported that during winter, the swift migrates to sub-Saharan Africa stopover sites, spanning regions such as the Sahel, the Congo Basin, and parts of East and Southern Africa, conducting their aerial lifestyle and continuous mobility during the non-breeding season [29]. In Europe, and in Italy, the swift’s typical arrival is in early spring for the breeding period, departing in late summer [29]. Their unique ecological cycle exposes swifts to different environmental contexts, even across continents, making them interesting candidates as bioindicators of environmental AMR.
Isolates from swifts sampled shortly after their arrival revealed a NWT proportion of 32.22%, compared to 26.92% in isolates from swifts sampled before the autumn migration. Moreover, when considering only adults swifts, the NWT percentage was 34.88%, compared to juveniles (28.8%). These results may indicate a potential exposure to antimicrobial resistant bacteria during migration routes. Some of the wintering sites across several African regions may be characterized by widespread and often unregulated use of antimicrobials in human and veterinary medicine, combined with a lack of surveillance AMR programs, which strongly create a substantial selective pressure for resistant bacterial strains [30]. Nevertheless, the presence of NWT isolates in juveniles, that had never migrated, may suggest rapid acquisition from the environment or from parents’ transmission.
Isolates from swifts at the arrival showed elevated resistance rate against cefoxitin (58.06%) and NWT phenotypes to ertapenem (9.38%), consistent with the circulation of MRS detected in this study, as previously described. High NWT percentage to tetracycline (15.56%) was also observed, possibly reflecting its extensive use in veterinary contexts. Tetracycline is considered the main antimicrobial drug used in poultry farming, especially in African areas [31], considered a dominant livestock sector in the continent [32]. Finally, NWT isolates toward rifampicin (8.47%) were also observed, a widely used antibiotic drug for the treatment of tuberculosis across the African continent [33]. Interestingly, in a previous study from our research group, we highlighted a correlation between rifampicin phenotypical resistance and isolates from migratory wild birds [7], suggesting a possible trend in migratory birds’ species and epidemiological significance of rifampicin acquired resistance from African areas, but further studies are needed to clarify this association.
On the other hand, isolates from swifts sampled before autumn migration, after spending the breeding season in Europe, possibly in Italy, showed a marked increase in NWT percentages toward macrolides, specifically clarithromycin (44.44%) and erythromycin (44.44%), clindamycin (43.75%) and enrofloxacin (11.58%). The results emerged may suggest the possible acquisition of locally originated acquired resistance during the breeding season. In Europe, swifts predominantly nest in urban areas where contact with contaminated surface waters, urban waste, or their diet based on insects from polluted environments may enhance exposure to resistant strains. According to the Italian National Antibiotic Use Report from 2023, macrolides are among most frequently used in the Italian population [34]. It is important to note that macrolides belong to the Watch group of antibiotics in the AWaRe classification introduced by the World Health Organization in 2022 [35]. Hence, these antibiotics are broad-spectrum molecules, recommended only as first-choice treatment towards pathogens more likely to be resistant to narrow-spectrum options. Furthermore, macrolides environmental persistence, especially for clarithromycin, has been well documented [36], emphasizing the importance to pay attention to this antimicrobial class both for its clinical relevance and impact at environmental level. Clindamycin is an antibiotic widely used in birds, especially for the treatment of Staphylococcus spp. infections, and resistance is frequent in staphylococci isolated from wildlife [37]. For what concern fluoroquinolones, they also have been frequently used in veterinary medicine [38], and according to European Medicines Agency (EMA), Italy still records high antimicrobial consumption rates in veterinary medicine across Europe [39]. A possible explanation for the higher prevalence of NWT isolates across sampled swifts after the breeding season may be linked to the conditions of the wildlife rehabilitation center where sampling occurred. In such settings, the aggregation between individuals, possible environmental contamination, and occasional exposure to antimicrobial treatments could facilitate the selection and circulation of resistant bacteria [40]. Specifically, enrofloxacin is one of the most administrated antibiotics in birds [41], which implies a plausible direct or indirect exposure in individuals in rehabilitation center.
Age-stratified analysis, comparing adult and juveniles swift isolates, revealed further details. Indeed, juveniles isolates showed higher NWT proportion to clarithromycin (34.21%), erythromycin (31.60%), and enrofloxacin (9.91%), similar to isolates from swifts sampled before the autumn migration, strengthening the hypothesis of a local acquisition during the breeding season. Whereas NWT percentage to rifampicin (5.63%) was comparable between juveniles and isolates from swifts sampled at their arrival, supporting the possible acquisition of certain resistance during the wintering grounds. These findings may be related to an early-life exposure to resistant bacteria, possibly acquired through feeding behaviors from parents; a hypothesis supported by literature reporting that birds’ gut microbiota is largely shaped by maternal transfer diet [42]. Moreover, an immature immune system of nestlings, which may facilitate the colonization and the persistence of resistant strains, but also the nest microenvironment, characterized by high bacterial density, could represent a hotspot for the resistance genes exchange [43]. Considering the swift’s ecological behaviors, its tendency to nest in urban settings make it a highly exposed species to anthropogenic contaminants, including AMR bacteria and resistance determinants. The low NWT phenotypes to ampicillin (4.85%) and amoxicillin–clavulanic acid (3.45%) observed exclusively in juveniles may further highlight potential antimicrobial pressure in urban setting or at the recovery center, since these antimicrobials are used as therapeutic agents in both human and veterinary medicine [44]. Nonetheless, specific contributions of all these potential transmission routes and the long-term consequences of early-life microbial exposure in birds remain to be clarified.
Despite the relevance of these findings in advancing environmental monitoring of AMR through the evaluation of NWT profiles from the investigated commensal bacterial microbiota of swifts, they should be interpreted with caution due to certain limitations. First, NWT profiles emerged from swifts sampled at their arrival have been related to a possible exposure during wintering periods in African regions, however the possibility of early AMR acquisition within the rehabilitation center cannot be excluded. Similarly, NWT profiles from swifts sampled at the end of the breeding period could have acquired resistance phenotypes locally or be chronic AMR carriers. This highlights an intrinsic bias introduced by sampling individuals recovered, even temporarily, in a captive environment where exposure to human or environmental related bacterial communities may occur. Additionally, individuals were not sampled longitudinally across both time points, limiting the ability to track within-individual AMR acquisition. Another limitation is a relatively small sample size and the consequently underrepresentation of adult swifts at the arrival in Italy. Furthermore, genomic analyses such as Whole Genome Sequencing (WGS) could provide insights into the genetic basis of AMR determinants. Therefore, future research should aim to larger and more balanced samples, longitudinal monitoring of the same individuals, sampling in natural habitats rather than rehabilitation centers, and inclusion of advanced genomic analyses, to allow a more accurate evaluation of the potential role of swifts as bioindicators of environmental AMR dissemination.
5. Conclusions
This study provides novel insights supporting the unexplored role of the swift (Apus spp.), a long-distance migratory bird moving from sub-Saharan Africa to Europe, as bioindicator for environmental AMR monitoring. Our findings demonstrated the presence of non-wild type (NWT) bacteria within the commensal microbiota of swifts, specifically among selected bacterial orders (Enterobacterales, Bacillales and Lactobacillales), highlighting the presence of acquired phenotypical resistance. Comparative analysis revealed partially overlapping NWT prevalences, but also the emerging of some differences across sampling periods and age classes, suggesting that swifts may acquire resistant bacteria possibly both in wintering grounds as well as in breeding areas. High percentages of NWT isolates observed in swifts at the arrival may reflect the selective pressure exerted during their migratory routes (tetracycline, rifampicin), whereas those observed in swifts sampled after the breeding season (macrolides, clindamycin, enrofloxacin) may be indicative of local selective pressures or those present within recovery centers. Moreover, the detection of methicillin-resistant staphylococci and NWT isolates to last-resort antimicrobials emphasize the circulation of resistant bacteria and ARGs across humans, wildlife and environment interface. In conclusion, our findings suggest the role of swifts (Apus spp.) as promising bioindicators of AMR contamination and reinforce the need for continuous AMR environmental surveillance within One Health strategies.
Author Contributions
Writing – review & editing, Writing – original draft, Supervision, Investigation, Data curation, E. E.; Writing – review & editing, Supervision, Formal analysis, Data curation, R. S.; Investigation, Conceptualization, T. D. L.; Visualization, Validation, Supervision, Project administration, Conceptualization, A. Z.; Conceptualization, Validation, G. M.; Visualization, Methodology, E. M.; Resources, Methodology, S. P.; Writing – review & editing, Validation, Supervision, Resources, Project administration, Conceptualization, S. P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The research project object of this study obtained a favourable ethical-scientific opinion from the Animal Welfare Committee (Prot. N. 0016546 in 09/15/2023).
Conflicts of Interest
The authors declare no conflicts of interest.:
Abbreviations
The following abbreviations are used in this manuscript:
| AMR | Antimicrobial resistance |
| MALDI-TOF MS | Matrix-assisted laser desorption–ionization time- of-flight mass spectrometry |
| WT | Wild type |
| NWT | Non-wild type |
| CN | Gentamicin |
| AMP | Ampicillin |
| AMC | Amoxicillin + clavulanic acid |
| FOX | Cefoxitin |
| RD | Rifampicin |
| DA | Clindamycin |
| TE | Tetracycline |
| E | Erythromicyn |
| CLR | Clarithromycin |
| ENR | Enrofloxacin |
| SXT | Trimethoprim-sulfamethoxazole |
| ETP | Ertapenem |
| PCR | Polymerase chain reaction |
| AST | Antimicrobial susceptibility test |
| CoNS | Coagulase-negative staphylococci |
| MRS | Methicillin-resistant staphylococci |
| ARGs | Antimicrobial resistance genes |
References
- World Health Organization. (2023). Antimicrobial resistance. World Health Organization. https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance.
- O’Neill, J. (2016). Tackling drug-resistant infections globally: Final report and recommendations. Review on Antimicrobial Resistance. Wellcome Trust & HM Government. Retrieved from https://amr-review.org/sites/default/files/160525_Final%20paper_with%20cover.pdf.
- Llor, C., & Bjerrum, L. (2014). Antimicrobial resistance: risk associated with antibiotic overuse and initiatives to reduce the problem. Therapeutic advances in drug safety, 5(6), 229–241. [CrossRef]
- Abbassi, M. S., Badi, S., Lengliz, S., Mansouri, R., Salah, H., & Hynds, P. (2022). Hiding in plain sight—Wildlife as a neglected reservoir and pathway for the spread of antimicrobial resistance: A narrative review. FEMS Microbiology Ecology, 98(6), fiac045. [CrossRef]
- Food and Agriculture Organization (FAO), United Nations Environment Programme (UNEP), World Health Organization (WHO), & World Organisation for Animal Health (WOAH) (Founded as OIE). (2022). One Health Joint Plan of Action 2022–2026: Working together for the health of humans, animals, plants and the environment. Rome: FAO. [CrossRef]
- Bengtsson-Palme, J., Abramova, A., Berendonk, T. U., Coelho, L. P., Forslund, S. K., Gschwind, R., Heikinheimo, A., Jarquín-Díaz, V. H., Khan, A. A., Klümper, U., Löber, U., Nekoro, M., Osińska, A. D., Ugarcina Perovic, S., Pitkänen, T., Rødland, E. K., Ruppé, E., Wasteson, Y., Wester, A. L., & Zahra, R. (2023). Towards monitoring of antimicrobial resistance in the environment: For what reasons, how to implement it, and what are the data needs?. Environment international, 178, 108089. [CrossRef]
- Esposito, E., Scarpellini, R., Celli, G., Marliani, G., Zaghini, A., Mondo, E., Rossi, G., & Piva, S. (2024). Wild birds as potential bioindicators of environmental antimicrobial resistance: A preliminary investigation. Research in veterinary science, 180, 105424. [CrossRef]
- Borrelli, L., Fioretti, A., Russo, T. P., Barco, L., Raia, P., De Luca Bossa, L. M., Sensale, M., Menna, L. F., & Dipineto, L. (2013). First report of Salmonella enterica serovar Infantis in common swifts (Apus apus). Avian Pathology, 42(4), 323–326. [CrossRef]
- Miniero, R., Carere, C., De Felip, E., Iacovella, N., Rodriguez, F., Alleva, E., & Di Domenico, A. (2008). The use of common swift (Apus apus), an aerial feeder bird, as a bioindicator of persistent organic microcontaminants. Annali dell’Istituto Superiore di Sanità, 44(2), 187–194. https://pubmed.ncbi.nlm.nih.gov/18660568/.
- EUCAST (European Committee on Antimicrobial Susceptibility Testing) (2025). EUCAST disk diffusion method for antimicrobial susceptibility testing (Version 13.0). https://www.eucast.org/fileadmin/eucast/pdf/disk_test_documents/2025/Manual_v_13.0_EUCAST_Disk_Test_2025.pdf.
- Fernandes, C. J., Fernandes, L. A., & Collignon, P., on behalf of the Australian Group on Antimicrobial Resistance (AGAR). (2005). Cefoxitin resistance as a surrogate marker for the detection of methicillin-resistant Staphylococcus aureus. Journal of Antimicrobial Chemotherapy, 55(4), 506–510. [CrossRef]
- Jensen, A. B., Lau, E. F., Greve, T., & Nørskov-Lauritsen, N. (2025). The EUCAST disk diffusion method for antimicrobial susceptibility testing of oral anaerobes. APMIS, 133(2), Article e70002. [CrossRef]
- EUCAST (European Committee on Antimicrobial Susceptibility Testing) (2025). EUCAST breakpoint tables for interpretation of MICs and zone diameters (Version 15.0). https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_15.0_Breakpoint_Tables.pdf.
- EUCAST (European Committee on Antimicrobial Susceptibility Testing) (2019). Definitions of S, I and R. https://www.eucast.org/newsiandr.
- EUCAST (European Committee on Antimicrobial Susceptibility Testing) (2023). Expected resistant phenotypes Version 1.2. Retrived from https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Expert_Rules/2023/Expected_Resistant_Phenotypes_v1.2_20230113.pdf.
- Stegger, M., Andersen, P. S., Kearns, A., Pichon, B., Holmes, M. A., Edwards, G., Laurent, F., Teale, C., Skov, R., & Larsen, A. R. (2012). Rapid detection, differentiation and typing of methicillin-resistant Staphylococcus aureus harbouring either mecA or the new mecA homologue mecA(LGA251). Clinical Microbiology and Infection, 18(4), 395–400. [CrossRef]
- Lack, D. (1955). The food of the swift. Journal of Animal Ecology, 24(1), 120–136. https://www.jstor.org/stable/1882.
- Wayman, C., Fernández-Piñas, F., Fernández-Valeriano, R., García-Baquero, G. A., López-Márquez, I., González-González, F., Rosal, R., & González-Pleiter, M. (2024). The potential use of birds as bioindicators of suspended atmospheric microplastics and artificial fibers. Ecotoxicology and Environmental Safety, 282, 116744. [CrossRef]
- Smith, H. G., Bean, D. C., Clarke, R. H., Loyn, R., Larkins, J. A., Hassell, C., & Greenhill, A. R. (2022). Presence and antimicrobial resistance profiles of Escherichia coli, Enterococcus spp. and Salmonella sp. in 12 species of Australian shorebirds and terns. Zoonoses and Public Health, 69(6), 615–624. [CrossRef]
- Musa, L., Stefanetti, V., Casagrande Proietti, P., Grilli, G., Gobbi, M., Toppi, V., Brustenga, L., Magistrali, C. F., & Franciosini, M. P. (2023). Antimicrobial Susceptibility of Commensal E. coli Isolated from Wild Birds in Umbria (Central Italy). Animals, 13(11), 1776. [CrossRef]
- Sousa, M., Silva, N., Igrejas, G., Silva, F., Sargo, R., Alegria, N., Benito, D., Gómez, P., Lozano, C., Gómez-Sanz, E., Torres, C., Caniça, M., & Poeta, P. (2014). Antimicrobial resistance determinants in Staphylococcus spp. recovered from birds of prey in Portugal. Veterinary Microbiology, 171(3-4), 436–440. [CrossRef]
- Ruiz-Ripa, L., Gómez, P., Alonso, C. A., Camacho, M. C., Ramiro, Y., de la Puente, J., Fernández-Fernández, R., Quevedo, M. Á., Blanco, J. M., Báguena, G., Zarazaga, M., Höfle, U., & Torres, C. (2020). Frequency and characterization of antimicrobial resistance and virulence genes of coagulase-negative Staphylococci from wild birds in Spain: Detection of tst-carrying S. sciuri isolates. Microorganisms, 8(9), 1317. [CrossRef]
- Martín-Maldonado, B., Rodríguez-Alcázar, P., Fernández-Novo, A., González, F., Pastor, N., López, I., Suárez, L., Moraleda, V., & Aranaz, A. (2022). Urban birds as antimicrobial resistance sentinels: White storks showed higher multidrug-resistant Escherichia coli levels than seagulls in central Spain. Animals, 12(19), 2714. [CrossRef]
- Prakash, A. E., Hromádková, T., Jabir, T., Vipindas, P. V., Krishnan, K. P., Mohamed Hatha, A. A., & Briedis, M. (2022). Dissemination of multidrug resistant bacteria to the polar environment: Role of the longest migratory bird Arctic tern (Sterna paradisaea). Science of the Total Environment, 815, 152727. [CrossRef]
- Colín-Castro, C. A., López-Jácome, L. E., Rodríguez-García, M. J., Garibaldi-Rojas, M., Rojas-Larios, F., Vázquez-Larios, M. del R., … Garza-González, E. (2025). The ongoing antibiotic resistance and carbapenemase-encoding genotypes surveillance: The first quarter report of the INVIFAR network for 2024. PLOS ONE, 20(4), e0319441. [CrossRef]
- Jubeh, B., Breijyeh, Z., & Karaman, R. (2020). Resistance of Gram-Positive Bacteria to Current Antibacterial Agents and Overcoming Approaches. Molecules, 25(12), 2888. [CrossRef]
- Abdullahi, I.N., Juárez-Fernández, G., Höfle, U. et al. Staphylococcus aureus Carriage in the Nasotracheal Cavities of White Stork Nestlings (Ciconia ciconia) in Spain: Genetic Diversity, Resistomes and Virulence Factors. Microb Ecol 86, 1993–2002 (2023). [CrossRef]
- Wellbrock, A. H. J., Bauch, C., Rozman, J., & Witte, K. (2017). ‘Same procedure as last year?’ Repeatedly tracked swifts show individual consistency in migration pattern in successive years. Journal of Avian Biology, 48(6), 797–805. [CrossRef]
- Åkesson, S., Klaassen, R., Holmgren, J., Fox, J. W., & Hedenström, A. (2012). Migration routes and strategies in a highly aerial migrant, the common swift Apus apus, revealed by light-level geolocators. PLoS ONE, 7(7), e41195. [CrossRef]
- Gulumbe, B. H., Haruna, U. A., Almazan, J., Ibrahim, I. H., Faggo, A. A., & Bazata, A. Y. (2022). Combating the menace of antimicrobial resistance in Africa: A review on stewardship, surveillance and diagnostic strategies. Biological Procedures Online, 24(1), 19. [CrossRef]
- Kimera, Z. I., Mshana, S. E., Rweyemamu, M. M., Mboera, L. E. G., & Matee, M. I. N. (2020). Antimicrobial use and resistance in food-producing animals and the environment: An African perspective. Antimicrobial Resistance & Infection Control, 9(1), 37. [CrossRef]
- Guèye, E. F. (2024). Trends and prospects of poultry value chains in Africa. Journal of Agriculture, Science and Technology, 23(4), 19–46. [CrossRef]
- Said, B., Pétermann, Y., Howlett, P., & Guidi, M. (2025). Rifampicin exposure in tuberculosis patients with comorbidities in Sub-Saharan Africa: Prioritising populations for treatment — a systematic review and meta-analysis. Clinical Pharmacokinetics, 64(8), 1149–1163. [CrossRef]
- Agenzia Italiana del Farmaco (AIFA). (2024). L’uso degli antibiotici in Italia. Rapporto Nazionale 2023. Roma: AIFA. https://www.aifa.gov.it/documents/20142/2766777/Rapporto_Antibiotici_2023.pdf.
- World Health Organization. (2022). The WHO AWaRe (Access, Watch, Reserve) antibiotic book. World Health Organization. https://iris.who.int/handle/10665/365135.
- Baranauskaite-Fedorova, I., & Dvarioniene, J. (2023). Management of Macrolide Antibiotics (Erythromycin, Clarithromycin and Azithromycin) in the Environment: A Case Study of Environmental Pollution in Lithuania. Water, 15(1), 10. [CrossRef]
- Sánchez-Ortiz, E., Blanco Gutiérrez, M. D. M., Calvo-Fernandez, C., Mencía-Gutiérrez, A., Pastor Tiburón, N., Alvarado Piqueras, A., Pablos-Tanarro, A., & Martín-Maldonado, B. (2024). Addressing challenges in wildlife rehabilitation: Antimicrobial-resistant bacteria from wounds and fractures in wild birds. Animals, 14(8), 1151. [CrossRef]
- Grabowski, Ł., Gaffke, L., Pierzynowska, K., Cyske, Z., Choszcz, M., Węgrzyn, G., & Węgrzyn, A. (2022). Enrofloxacin—the ruthless killer of eukaryotic cells or the last hope in the fight against bacterial infections? International Journal of Molecular Sciences, 23(7), 3648. [CrossRef]
- EMA (European Medicines Agency) (2025). European sales and use of antimicrobials for veterinary medicine: Annual surveillance report for 2023. European Medicines Agency. https://www.ema.europa.eu/en/documents/report/european-sales-use-antimicrobials-veterinary-medicine-annual-surveillance-report-2023_en.pdf.
- Russo, T. P., Minichino, A., Gargiulo, A., Varriale, L., Borrelli, L., Pace, A., Santaniello, A., Pompameo, M., Fioretti, A., & Dipineto, L. (2022). Prevalence and phenotypic antimicrobial resistance among ESKAPE bacteria and Enterobacterales strains in wild birds. Antibiotics, 11(12), 1825. [CrossRef]
- Soh, H. Y., Tan, P. X. Y., Ng, T. T. M., Chng, H. T., & Xie, S. (2022). A Critical Review of the Pharmacokinetics, Pharmacodynamics, and Safety Data of Antibiotics in Avian Species. Antibiotics, 11(6), 741. [CrossRef]
- Chen, C. Y., Chen, C. K., Chen, Y. Y., Fang, A., Shaw, G. T., Hung, C. M., & Wang, D. (2020). Maternal gut microbes shape the early-life assembly of gut microbiota in passerine chicks via nests. Microbiome, 8(1), 129. [CrossRef]
- Teyssier, A., Lens, L., Matthysen, E., & White, J. (2018). Dynamics of gut microbiota diversity during the early development of an avian host: Evidence from a cross-foster experiment. Frontiers in Microbiology, 9, 1524. [CrossRef]
- Pawłowska, B., Sysa, M., Godela, A., & Biczak, R. (2024). Antibiotics amoxicillin, ampicillin and their mixture—Impact on bacteria, fungi, ostracods and plants. Molecules, 29(18), 4301. [CrossRef]
Figure 1.
Percentage distribution of bacterial species isolated from all sampled swifts in the present study.
Figure 1.
Percentage distribution of bacterial species isolated from all sampled swifts in the present study.
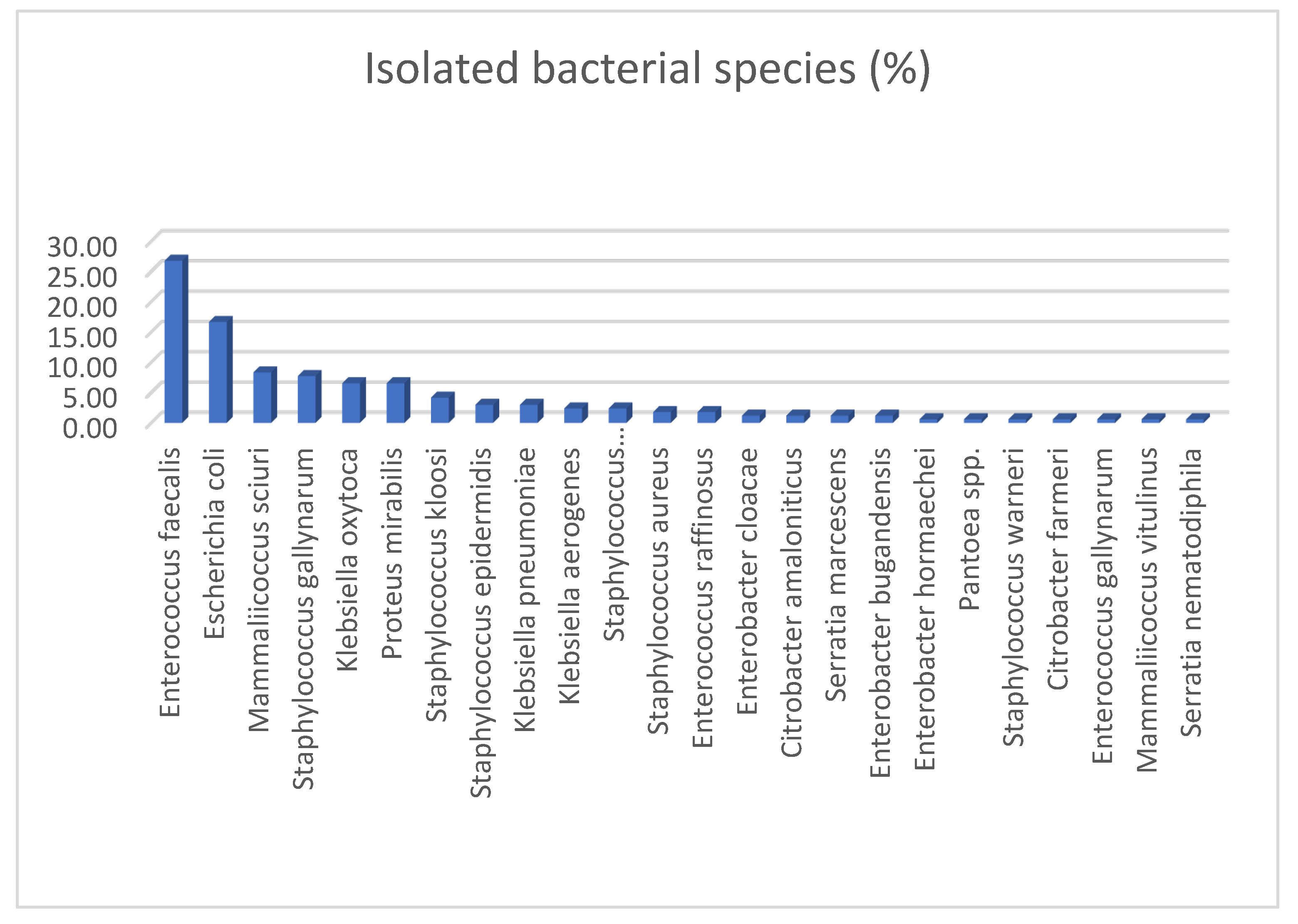
Figure 2.
Non-wild type proportions among bacterial isolates from sampled swifts. This figure summarizes the percentage of NWT isolates for each tested antimicrobial drug, except cefoxitin, for which the percentage of resistant isolates is reported, highlighting differences in resistance prevalence between the two sampling periods. The last two bars show the total percentage of NWT isolates across all antimicrobials at the two sampling periods.
Figure 2.
Non-wild type proportions among bacterial isolates from sampled swifts. This figure summarizes the percentage of NWT isolates for each tested antimicrobial drug, except cefoxitin, for which the percentage of resistant isolates is reported, highlighting differences in resistance prevalence between the two sampling periods. The last two bars show the total percentage of NWT isolates across all antimicrobials at the two sampling periods.
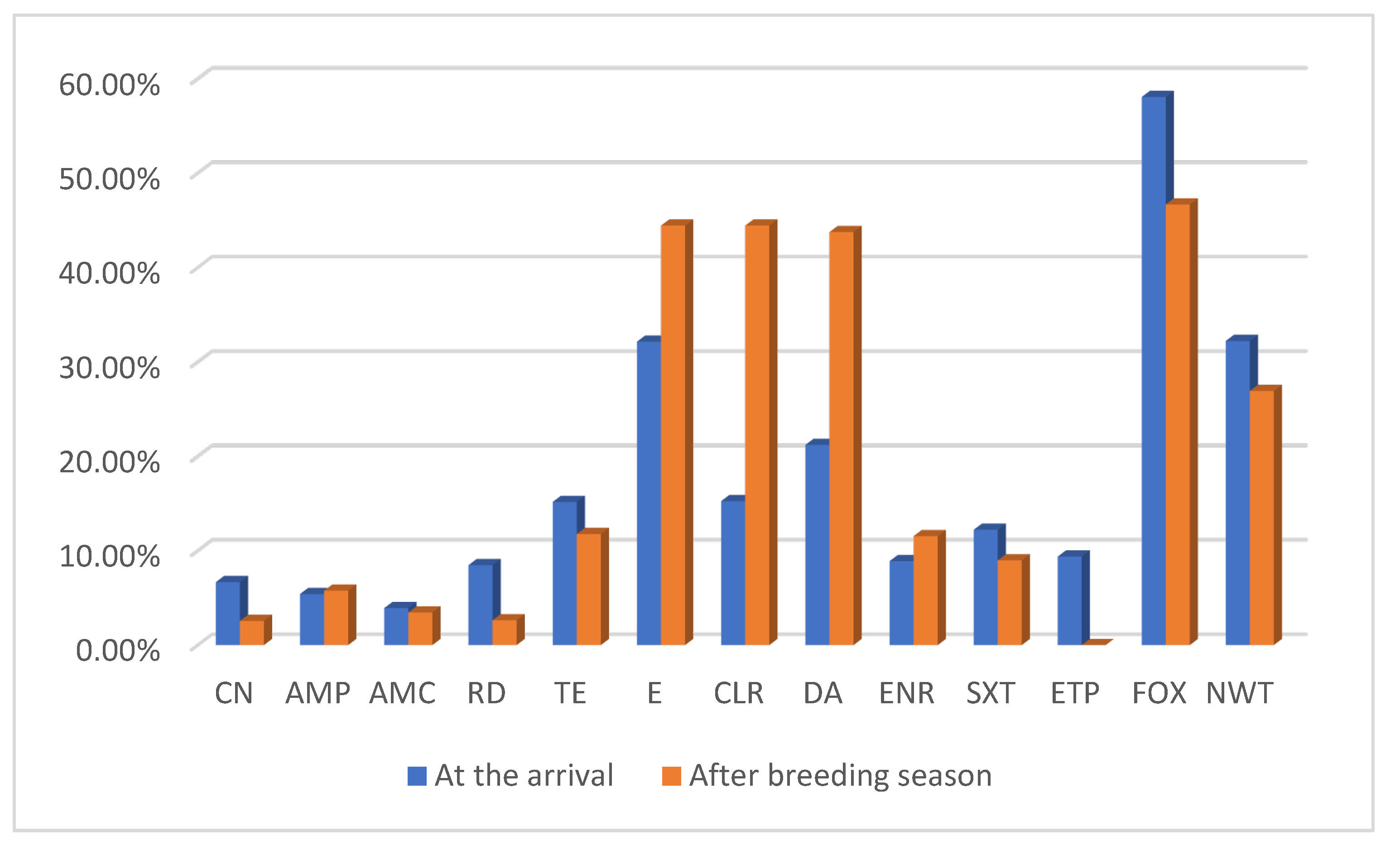

Table 1.
List of tested antimicrobials grouped by antimicrobial class with corresponding code abbreviation/disk concentration (µg), bacterial order tested, availability of ECOFFs value for specific bacterial species, and EUCAST clinical zone diameter breakpoints applied.
Table 1.
List of tested antimicrobials grouped by antimicrobial class with corresponding code abbreviation/disk concentration (µg), bacterial order tested, availability of ECOFFs value for specific bacterial species, and EUCAST clinical zone diameter breakpoints applied.
| Antimicrobial class | Antimicrobial | Code (μg) | Bacterial order tested | ECOFFs (NWT<mm) | EUCAST clinical zone diameter breakpoints (R<mm) |
| Aminoglycosides | Gentamicin | CN (10) | Bacillales, | S. aureus (18); S. epidermidis (20) | |
| Enterobacterals | E. coli, P. mirabilis, Serratia marcescens (16); K. oxytoca, C. freundii, E. cloacae (15); K. pneumoniae, K. aerogenes (14); | ||||
| Penicillins +/- beta-lactamase inhibitors | Ampicillin | AMP (10) | Enterobacterals, | E. coli (13); P. mirabilis (19) | |
| Lactobacillales | E. faecalis (12) | ||||
| Amoxicillin + clavulanic acid | AMC (30) | Enterobacterals | E. coli (14); K. pneumoniae (22); P. mirabilis (22) | ||
| Lactobacillales | N. A. | ||||
| Cephalosporins | Cefoxitin | FOX (30) | Bacillales | - | S. aureus, CoNs (22); S. epidermidis (27) |
| Rifamycins | Rifampicin | RD (5) | Bacillales | S. aureus (24); S. epidermidis (30) | |
| Lactobacillales | N. A. | ||||
| Lincosamides | Clindamycin | DA (2) | Bacillales | S. aureus, S. epidermidis (21) | |
| Tetracyclines | Tetracycline | TE (30) | Bacillales, | S. aureus (20) | |
| Enterobacterals | E. coli (21) | ||||
| Lactobacillales | N. A. | ||||
| Macrolides | Erythromycin | E (15) | Bacillales | S. aureus (22) | |
| Clarithromycin | CLR (15) | Bacillales | N. A. | ||
| Fluoroquinolones | Enrofloxacin | ENR (5) | Bacillales, | N. A. | |
| Lactobacillales, | N. A. | ||||
| Enterobacterals | N. A. | ||||
| Sulfonamides + dihydrofolate reductase inhibitors | Trimethoprim-sulfamethoxazole | SXT (25) | Bacillales, | S. aureus (23) | |
| Lactobacillales, | E. faecalis (26) | ||||
| Enterobacterals | E. coli (22); K. oxytoca (21); K. pneumoniae (18); E. cloacae, P. mirabilis (20) | ||||
| Carbapenems | Ertapenem | ETP (10) | Enterobacterals, | E. coli (24); K. pneumoniae (22) | |
| Bacillales | N. A. |
Table 2.
Primers sequences and expected product size used for the detection of methicillin-resistance genes in the present study.
Table 2.
Primers sequences and expected product size used for the detection of methicillin-resistance genes in the present study.
| Gene | Sequences (5′–3′) | Product size (bp) | References |
| mecA | Fw: TCCAGATTACAACTTCACCAGG | 162 | Stegger et al. (2012) |
| Rev: CCACTTCATATCTTGTAACG | |||
| mecC | Fw: AAAAAAAGGCTTAGAACGCCTC | 138 | Stegger et al. (2012) |
| Rev: GAAGATCTTTTCCGTTTTCAGC |
Table 3.
Distribution of bacterial isolates recovered from grouped swifts, according to bacterial order. Percentages for each category are calculated relative to the total number of isolates per group.
Table 3.
Distribution of bacterial isolates recovered from grouped swifts, according to bacterial order. Percentages for each category are calculated relative to the total number of isolates per group.
| Grouped swifts | Tot isolates (n) | Enterobacterales (n); (%) | Bacillales (n); (%) | Lactobacillales (n); (%) |
| TOTAL | 168 | 71; (42.26%) | 48; (28.57%) | 49; (29.17%) |
| AT THE ARRIVAL | 90 | 31; (34.44%) | 33; (36.67%) | 26; (28.89%) |
| AFTER BREEDING SEASON | 78 | 40; (51.28%) | 15; (19.23%) | 23; (29.49%) |
Table 4.
NWT percentages, and in brackets the number of NWT isolates, for each antimicrobial tested in the total of bacteria isolates from swifts grouped according sampling period and age. For cefoxitin, the percentage of resistant isolates is reported. The percentage value of the total NWT isolates for each category is given in the last row.
Table 4.
NWT percentages, and in brackets the number of NWT isolates, for each antimicrobial tested in the total of bacteria isolates from swifts grouped according sampling period and age. For cefoxitin, the percentage of resistant isolates is reported. The percentage value of the total NWT isolates for each category is given in the last row.
| Adults at the arrival | Adults after breeding season | Juveniles | |
| CN | 3.33% (1/30) | 0% (0/13) | 5.60% (7/125) |
| AMP | 0% (0/24) | 0% (0/10) | 4.85% (5/103) |
| AMC | 0% (0/29) | 0% (0/13) | 3.45% (4/116) |
| RD | 5.26% (1/19) | 0% (0/7) | 5.63% (4/71) |
| TE | 16.67% (5/30) | 15.38% (2/13) | 13.51% (15/111) |
| E | 22.22% (2/9) | 20% (2/10) | 31.60% (12.38) |
| CLR | 22.22% (2/9) | 20% (2/10) | 34.21% (13/38) |
| DA | 22.22% (2/9) | 20% (2/10) | 26.31% (10/38) |
| ENR | 10% (3/30) | 23.08% (3/13) | 9.91% (11/111) |
| SXT | 3.33% (1/30) | 15.38% (2/13) | 9.91% (11/111) |
| ETP | 0% (0/20) | 0% (0/10) | 8% (6/75) |
| FOX | 44.44% (4/9) | 50% (2/4) | 54.29% (19/35) |
| NWT | 33.33% (10/30) | 38.46% (5/13) | 28.8% (36/125) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



