Submitted:
23 December 2025
Posted:
24 December 2025
You are already at the latest version
Abstract
Abnormal repetitive behaviours ('ARBs') in captive animals are a heterogeneous group of troubling activities (e.g. stereotypic pacing by Carnivora; feather-plucking by poultry). To assess and improve their construct validity as welfare indicators, we reviewed their responsiveness to mood-improving drugs; links with imprisonment, stress and self-reported poor well-being in humans; and in animals, the impact of welfare-compromising (e.g. aversive) treatments. Considerable evidence links ARB development with negative moods or mood disorders, and early/prolonged/recurrent negative experiences (potentially via dose-response-like effects). Findings also indicate effects of cumulative affective experience ('cumulative stress'). Furthermore, if ARBs transiently help subjects cope, such effects are only partial. Therefore, whenever husbandry or housing causes ARB-prone phenotypes, negative affect can reliably be inferred (with more severe ARBs indicating poorer welfare). However, ARBs are rather prone to false nulls as welfare indicators: prolonged negative affect does not always cause the emergence or increase of ARBs, primarily due to threshold effects, ceiling effects, and inactivity being an alternate response. Furthermore, in ARB-prone subjects, the onset/offset of bouts appears not to reliably track moment-by-moment levels of negative emotion. Additionally, because variation in activity, behavioural flexibility and stress-response style are potential confounds, ARBs are not advised for comparing welfare across individuals, strains, species or prenatal treatments. Overall, ARBs have strong construct validity as indicators of negative moods/mood disorders; and our additional rules-of-thumb should further refine their accuracy. Future research should investigate underlying mechanisms (e.g. those suggested by human and biomedical findings), especially to clarify the boundaries and biological sub-types of ARBs.
Keywords:
stereotypic behaviour
; construct validity
; affective states
; welfare assessment
; moods
; cumulative stress
1. Introduction
1.1. The Nature of the Problem: ARBs Are Heterogeneous and Seem to Have Complex Links to Welfare
If you have visited a zoo, you may have seen tigers repeatedly walking a worn-down track along their enclosure’s edge, or elephants swaying rhythmically from side to side. And if you have visited an intensive farm where sows are kept in small stalls, or a biomedical research facility during the dark phase of the circadian light cycle (when rodents are most active), you are even more likely to have witnessed strange repetitive behaviours: pigs making chewing movements until their saliva froths, or mice running in circles in their cages. Indeed, one conservative estimate is that over 70 million zoo, farm and lab animals a year perform abnormal repetitive behaviours like these (Mason and Latham, 2004). Abnormal repetitive behaviour, or ‘ARB’, is an umbrella term for a heterogeneous group of seemingly functionless, unusual or troubling activities (e.g., Mills and Luescher, 2006). They include ‘stereotypic behaviours’—defined by either their predictable appearance, or the role of frustration or brain dysfunction in their aetiology (Mason, 2006)—such as those described above. They also include more variable behaviours such as plucking fur or feathers from the self or a conspecific, and less repetitive forms such as the recurrent adoption of strange postures (e.g., holding out limbs, as sometimes seen in laboratory primates).
Collectively, such ARBs are very common in captive animals. Mechanistically, some appear to derive from ethologically normal responses to frustration or motivational conflict (e.g., Dixon et al., 2008; Keiper, 1969; Rushen and de Passillé, 1995; Wiedenmayer, 1997). However, neurological research and parallels with the ARBs seen in some human clinical populations highlight growing evidence for a role of captivity-induced, potentially dysfunctional neurological changes within the brain (e.g., Kitchenham and Mason, 2021; Muehlmann and Lewis, 2012; Wilkes and Lewis, 2018). These complementary, non-mutually exclusive explanations (see Figure 1) typically reflect research driven by different traditions and methodologies (e.g., Hall et al., 2003; Mason, 2006). And together, they identify several potential sources of heterogeneity across ARBs. One is the degree to which ARBs are best explained by motivational frustration, neurological change, or a combination of these processes (Díez-León et al., 2019; Hall et al., 2003; Latham and Mason, 2007; Mason, 2006; Pomerantz et al., 2012). A second, when motivational frustration is implicated, is the type of natural behaviour that is frustrated (e.g., interactions with the mother, grooming or being groomed, roaming, foraging, etc.); while a third is the type of response that follows, be it vacuum or intention movements mimicking the frustrated behaviour (motivationally perhaps even being a surrogate or substitute for it), escape attempts reflecting the aversiveness of frustration, or instead irrelevant-seeming displacement activities (e.g., Latham and Mason, 2007; Mason and Rushen, 2006). Where neurological changes are implicated, a fourth source of heterogeneity is the brain regions involved (typical candidates being various behavioural control pathways of the basal ganglia, or various parallel sets of corticostriatal loops that are also involved in the control of behaviour, running between basal ganglia and cortical regions like the prefrontal cortex; e.g., Garner, 2006; Lewis et al., 2006; reviewed by Kitchenham and Mason 2021); while a fifth source is the nature and lability of such changes: whether they involve readily reversible state changes (cf. e.g., Mcklveen et al., 2015; Soares et al., 2012); or longer-lasting, perhaps harder-to-reverse structural changes potentially caused by abnormal developmental environments (e.g., Lewis et al., 2000; Martin et al., 1991; Spann et al., 2012). And yet more sources of heterogeneity include the neurotransmitters suggested as most implicated (e.g., dopamine [McBride et al., 2022], endogenous opioids [Hemmings et al., 2018] and serotonin [de Haas and van der Eijk, 2018]); and whether the gut-brain axis is involved (e.g., Mindus et al., 2021; Wang et al., 2023; see also review by Homer et al., 2023). Small wonder then that ARBs vary greatly in their appearance, correlates, and (to human performers) their ‘feel’.
Ethological pathway (LH side): When performing motivated species-typical behaviours is prevented by sub-optimal environments, frustrated animals show vacuum activities (behaviours performed in the absence of the stimuli which normally elicit them), intention movements (incomplete or truncated fragments of a behavioral sequence, expressed when an animal is unable to perform the full action), or displacement activities (normal behaviours like grooming that seem irrelevant to the current context). These ARB ‘source behaviours’ are repeated, perhaps partly due to positive reinforcement from beneficial consequences of performance (‘coping’); and they may become less variable through normal processes of routine formation.
Neurological (potentially dysfunctional) pathway (RH side): Sub-optimal environments can alter brain development (e.g., by lacking necessary cognitive stimulation, social experiences and environmental complexity), and/or induce emotional state-dependent changes in function. These affect the cortico-basal ganglia circuits (or avian equivalents) that normally regulate behaviour selection. As a result, subjects may show behavioural disinhibition (inappropriate behaviours are triggered or fail to be suppressed) and inflexibility (difficulty adapting due to e.g., habit formation and the dominance of pre-potent responses): effects assessable in tests for compulsion, impulsion and perseveration.
This complexity acknowledged, the broad question we address here is, irrespective of mechanism, do ARBs in captive animals indicate poor welfare? By ‘poor welfare’, we mean poor affective well-being—for instance, experiencing psychological distress, low mood, or unpleasant symptoms of ill health. The strange appearance of ARBs, and their apparent increase when captive conditions are particularly restrictive, have long suggested yes. Some of the earliest observers described ARBs as “emotional and often neurotic symptoms” (Foley, 1934, p. 93) or “a sure sign of wrong treatment” (Hediger, 1950, p. 75). Thrope’s (1965) Appendix to the Brambell Report likewise characterized them as signs of "prolonged and intense emotional disturbances", while Broom and Johnson (1993) identified repetitive and stereotyped behaviours as “amongst the most important indicators of long-term-welfare problems” (p. 79). As a result, today ARBs are often used in welfare assessment and even policy-making. For example, for pigs, it is standard practice to use ARBs as welfare indicators in research studies, and also when setting new husbandry guidelines or laws (e.g., Broom et al., 1995; NFACC, 2012; Hoorweg et al., 2017; Wallgren et al., 2016; Zhang et al., 2017). The same holds for poultry (EFSA AHAW PANEL, 2023). It is also standard practice to use ARBs to assess laboratory animal welfare (e.g., CCAC, 2021; NC3Rs, 2025) and zoo animal welfare (e.g., Fazio et al., 2020; Hoorweg et al., 2017; Shyne and Block, 2010; Swaisgood and Shepherdson, 2005; and many others).
However, other perspectives on ARBs are more nuanced, or even skeptical. Most researchers agree that conditions which never elicit ARBs are likely to be better for animal welfare; however that ARBs are actually rather complex as welfare indicators has been recognised by statements like "It is unlikely we can use variation in the incidence of stereotypies to measure welfare in any simple fashion" (Duncan et al., 1993, p. 203); "not all stereotypies are sensitive indicators of current stress or frustration" (Mason and Latham, 2004, p. S64); "stereotypy may sometimes prevent the onset of distress" (NRC, 2008); and "there is no one to one correspondence between stereotypy and welfare" (Ashley, 2007, p. 216). Dawkins (2021) even argues that "stereotypy is not a valid welfare correlate on its own. Some stereotypies indicate poor welfare. Others show the opposite — they are part of the animal’s coping mechanism and are no more indicative of poor welfare than a chicken drinking or a dog repeatedly chasing a ball on a beach" (p. 55). Correspondingly, some animal carers discount ARBs as welfare-relevant, treating them instead as signs of successful coping, or as mere ‘habits’ that are legacies of welfare problems past (e.g., reviewed by Mason et al., 2007).
These positions reflect certain results in the by-now vast body of research on ARBs by applied ethologists and animal welfare scientists. The more study that ARBs have attracted, the more complex their relationships with well-being have appeared to be, with three main issues emerging. First, ARBs are not always elicited by poor conditions or acute stressors. For example, some pigs tethered in stalls do not develop ARB, even though their initial reactions (very strong escape attempts) and later elevated heart rates both indicate great stress and aversion (Schouten and Wiepkema, 1991). Second, ARBs often correlate with enhanced abilities to cope with the challenges of captivity. For example, in calves prematurely removed from their mothers, the performance of non-nutritive sucking and repetitive tongue-playing covaries with reduced heart rates, increased resting behaviour, and greater physiological signs of nutritional satiety (de Passillé and Rushen, 1997; et al., 1998; Veissier et al., 2002). Third, some ARBs persist even after poor conditions are improved; in particular, forms that develop in barren housing may become hard to treat by ‘enriching’ this housing, even with resources that would have successfully prevented the emergence of ARB in younger animals (reviewed by Tilly et al., 2010).
This narrative review therefore seeks to collate the evidence linking ARBs with poor welfare, and also to make sense of such complexities and counter-examples. Our primary aim is thus to rigorously assess the construct validity of ARBs as welfare indicators. Our secondary aim is to analyse and explain both cases where ARBs appear to be insensitive, unresponsive indicators (failing to track relevant changes in affective state), and cases where their emergence or increase instead appears caused by factors having little or nothing to do with welfare (suggesting poor specificity or selectivity to changes in affective state). To do this, we use a framework of five tests for the construct validation of welfare indicators (Mason and Mendl, in press). These validation tests were collated from studies of the evolution and psychology of human and animal emotion; veterinary research on the affective impacts of health problems; biomedical research aiming to validate lab animal “models” of pain, anxiety and other affective states/disorders clinically important to humans; and best practice in animal welfare research (Mason and Mendl, in press). Each is based on clearly laid out (yet differing) assumptions. We lay this validation framework out below, and also distinguish between emotions, moods and cumulative affective experience, before summarising the scope and structure of the review.
1.2. Five Tests for the Validity of ARBs as Welfare (Affect) Indicators
Validation Test 1 uses humans as models for understanding other species by assessing which measurable variables change in ways that track self-reported affective states in people. It then treats these variables as indicators of similar states in other species (Mason & Mendl in press). Here, Test 1 thus probes what ARBs reveal about negative affective states in humans (such as those in prison or affected by certain clinical conditions). Test 1’s advantages include that we can be rather certain of the valence and intensity of these subjects’ affective states, thanks to their verbal self-report and our own experiences or abilities to empathize. It also capitalizes on a vast amount of research on human affect and ARB performance. A potential disadvantage of Test 1, however, is that it assumes biological homology between humans and other species. Validation Tests 2-5 differ from Test 1 in freeing us from these assumptions of cross-species homology. These tests instead focus only on the animal species of interest (using pigs to assess whether porcine ARBs are signs of porcine negative affect, for instance), as follows.
Test 2 makes use of stimuli that animals will approach or work for (‘rewards’), and/or aversive stimuli that animals will avoid, or work to prevent or escape from (‘punishers’). Animals then exposed to such empirically identified rewards are assumed to be in relatively positive states, while animals exposed to punishers are assumed to be in relatively negative ones. Measurable variables that change in such subjects can then be assessed, with those reliably doing so (i.e., across a range of rewards or punishers) passing Test 2. Here, Test 2 thus assesses whether ARBs emerge/increase when animals are exposed to aversive stimuli, and decrease/vanish during exposure to preferred ones. Test 2’s main advantage is that it is theory-driven, building on the apparent proximate functions of affect, especially emotions, in driving motivated seeking and avoidance responses (e.g., Dawkins, 1988; Mendl and Paul, 2020; Rolls, 2014). Correspondingly, its main limitation is that it is better at validating indicators of short-term emotions than longer-term affective states (a topic we cover further below). Furthermore, the aversiveness of some challenges (such as prematurely losing maternal care) has not been evaluated in the sense of empirically ascertaining whether they are punishers. Validation Test 3 solves this problem by assuming that ancestrally, animals evolved to feel negative affect if exposed to treatments that would have harmed their ancestors’ fitness (cf.Cabanac, 1971; Dawkins, 2023; Mendl and Paul, 2020; Rolls, 2014). It thus involves assessing which variables change in animals exposed to ancestrally fitness-boosting or -harming situations. Here, Test 3 therefore assesses whether ARBs emerge/increase in animals exposed to threats to ancestral fitness, and decrease/vanish during exposure to rewards. Like Test 2, Test 3 is theory-driven, building on the presumed adaptive functions of affect. It is therefore problematic if we are unsure what would have influenced ancestral fitness, or our subjects are so domesticated that what their brains treat as rewards or punishers cannot be predicted by long-distant evolutionary selection pressures. However, Test 3 is a useful complement to Test 2, especially for wild animals or species with close extant ancestors whose natural lifestyles are well understood.
Validation Test 4 instead takes a pharmacological approach, manipulating affective states with drugs to reveal which measurable variables change in dosed subjects. Here, Test 4 thus assesses whether ARBs emerge/increase in animals given drugs that induce negative affective states, and decrease/vanish with exposure to drugs that alleviate negative states or induce positive ones (consistent with affective states not just correlating with ARBs, but actually causing them). Test 4 obviously relies on assumptions that we know how a given drug influences animals’ affective states. When such assumptions seem reasonable, however (due to homologous actions in humans, and/or their proven status as rewards or punishers for our species of interest), this approach can be very scientifically persuasive. Finally, Validation Test 5 involves assessing which variables covary with indicators of affect that have already been validated in our species of interest. Here, ARBs thus pass Test 5 if they covary with existing valid indicators of affective state (an approach obviously assuming that these other indicators are indeed valid). And overall, the more of these five tests that ARBs collectively pass, the more confident we can be that ARBs are signs of poor welfare.
1.3. The Nature of Affect: Timescale as a Crucial Consideration
While applying these tests and collating examples that pass or fail, we also recognise that the nature of affective states (and, crucially, the nature of their correlates) typically varies with the timescale under consideration, ranging on a continuum from short-term emotions to entire lives worth (or not worth) living.
Emotions are typically considered to last seconds to minutes (sometimes hours) (Oatley and Johnson-Laird, 2014), and have been operationalised by Rolls (2014) as states elicited by specific motivating stimuli that animals either seek out or actively avoid. Anticipating such stimuli can also elicit emotions (Mendl and Paul, 2020; Rolls, 2014), and they include states like hunger, thirst and pain, which some term homeostatic or primordial emotions (e.g., Craig, 2003). Moods are longer-lasting (with durations of hours to days; sometimes weeks; Oatley and Johnson-Laird 2014). From the perspective of a human experiencing them, moods may seem not to be caused by any obvious stimulus. However, research indicates that the prolonged malaise or lassitude associated with sickness reflects immune responses to infection (Bucks et al., 2008; Schrock et al., 2024); while other moods appear to integrate a subject’s emotional experiences over time, such that repeated negative emotions lead to low moods, while repeated positive emotions lead to positive ones (e.g., Mendl et al., in press.; Nettle and Bateson, 2012). If particularly prolonged, as well as severe, hard to reverse and/or seemingly disproportionate, negative moods may be deemed instead affective or mood disorders (e.g., generalised anxiety disorder, PTSD, major depression). Adversity in infancy, as well as repeated or severely negative experiences over the lifetime, are lifelong risk factors here; affective disorders can thus reflect past as well as current poor well-being (Mendl et al. in press), as can associated brain changes such as hippocampal volume loss (Poirier et al., 2019a). But these are not the only forms of disease vulnerability that reflect past poor well-being over prolonged periods: cancer, cardiovascular disease, stroke and susceptibility to viruses all become more likely and more severe in individuals exposed to chronic adversity (e.g., Cohen et al., 2019; Walker et al., 2012). This reflects the physiological ‘wear and tear’ arising from repeated or chronic negative affect, which compromises self-maintenance/-repair and immunity to infection, so accelerating biological ageing (e.g., Bateson and Poirier, 2019; Cait et al., 2022). Cumulative net negative experience over long periods thus has quite different signs or correlates than do brief negative emotions. The question "Do ARBs in captive animals indicate poor welfare?", therefore, should ideally be parsed out into sub-questions: Do ARBs in captive animals indicate negative emotions? Negative moods? Affective disorders? Or something yet longer-term still: the effects of cumulative negative affect over prolonged periods?
1.4. The Scope, Structure and Limitations of This Review
The sections below are ordered from Test 1 to 5. Within each, we begin with cases where ARBs pass the respective test. For Test 1, we first cover ARBs displayed by non-clinical populations of humans (acknowledging that the boundary between non-clinical and clinical is what arbitrary), then lasting ARBs induced in formerly neurotypical humans by environmental challenges (e.g., impoverished institutional rearing), before covering the role of affect in human clinical populations that are typified by ARB (e.g., Tourette’s syndrome, Autism Spectrum Disorder and Obsessive-Compulsive Disorder). We also cover cases where mood-enhancing drugs are effective for human ARBs. Turning to non-human animals (henceforth, ‘animals’), to limit the text for each of Tests 2 and 3 (both the focus of extensive animal welfare research), here for each we cover just three highly-studied topics: effects of aversive barren housing conditions, exposure to threatened or actual physical harm, and food deprivation for Test 2; and effects of fitness-threatening illness and infection, early maternal/parental loss, and broken or absent social bonds for Test 3. For Test 4, we primarily cover the impacts on animal ARBs of (assumed) antidepressants and anxiolytics. And for Test 5, we first review evidence that populations with elevated ARBs show signs of more negative affect (compared to differentially treated groups with lower levels), before reviewing evidence that within such populations, individuals with elevated ARB show signs of more negative affect (compared to identically-treated subjects spontaneously showing lower levels): a deliberate split by the source of variation in ARB, because this affects the patterns that emerge (Mason & Latham 2004).
Within each section, we then present counter-examples wherein ARBs fail the respective test. Thus, we review cases where human ARBs fail to increase during negative states/experiences; when animal ARBs fail to increase in aversive situations or in response to threats to fitness; when ARBs are not reduced by anti-depressants and anxiolytics; and when treatments, groups or individuals displaying relatively high ARB do not show more evidence of negative affect than comparators displaying less ARB. These all illustrate the potential poor responsiveness (or poor sensitivity) of ARBs to changes in welfare. We then review cases where ARBs even show poor selectivity (or poor specificity) to changes in welfare: thus where human ARBs are elevated during neutral or positive states compared to more negative ones; when animals’ ARBs are elevated in situations that are preferred rather than aversive, or that promote fitness rather than decrease it; when animals’ ARBs are increased by drugs that are affect-neutral or even promote positive affect; and lastly, when animals’ ARBs positively covary with signs of relatively good rather than poor welfare.
Our structure is thus logical and based on principles. However, in contrast, our literature searching process was not systematic. This is because its core is a collection of papers first started by one of us (GJM) in 1988 and steadily added to whenever particular research projects required. This means that the animal-based studies cited are skewed by the nature of these projects: towards zoo animals (especially elephants, Ungulates and Carnivora), fur animals (especially mink) and lab animals (especially mice and rhesus monkeys), while being far more superficial for farm animals (especially poultry), fish, horses, and zoo-housed birds. This is undoubtedly a weakness. We do judge this to not be a source of systematic bias in terms of the ideas emerging here, especially given the hundreds of papers collated and utilized in this review. However, we will recognise this limitation in how we present our final conclusions: not as proven, definitive truths, but instead as specific, clear hypotheses that can now be tested via experiment or meta-analysis. Note too that there is some arbitrariness in which ARBs were included or excluded, primarily driven by which behaviours were decreed to be ‘stereotypic’ or ‘abnormal’ by the authors read, and also by which involved the manipulations or data needed for Tests 2-5. In the wrap-up of this review, we therefore also reconsider terminology (especially ARB versus ‘stereotypic behaviour’) and discuss the issues arising when identifying animal ARBs.
We will then conclude with an analysis of the links between animal ARBs/stereotypic behaviours and welfare, highlighting the problems of using them to infer animal emotions, but their great value for assessing animal moods/mood disorders or cumulative negative affect over long periods. We will lay out the rules of thumb (or testable working hypotheses) that arise from such conclusions (and that should help them be used more accurately in welfare assessment), as well as highlighting several other research questions arising along the way.
2. Are ARBs Valid Indicators of Poor Welfare?
2.1. Test 1: LINKS between ARBs and Negative Affect in Humans
2.1.1. Non-Clinical Human ARBs
Nail-biting, gum-chewing, knee-joggling and other ‘fidgets’ are common everyday behaviours that may barely deserve the label ‘abnormal’; and as long as they only occur in brief bouts, take negligible time, are easily relinquished, and lack negative consequences (e.g., embarrassment or physical harm), perhaps they seem of little affective significance. However, prolonged negative experiences (including institutionalization/imprisonment) and self-reported negative moods are linked with increases in both the prevalence of such behaviours and the time devoted to them, as well as seeming to cause unusual new forms to emerge: one reason some authors call these behaviours ‘nervous habits’ (e.g., Young 1947).
Thus, ARBs in children (including potentially harmful forms like air-swallowing, self-scratching and skin-picking) emerge and/or increase during periods of stress at home or school (e.g., Hwang et al., 2007; Devanarayana and Rajindrajith, 2012; Câmara-Souza et al., 2023), after being moved into residential care (Tröster, 1994), or if children have abusive or indifferent, depressed parents (Egeland et al., 1983). Body-rocking—common in infants but normally grown out of—also persists and even increases in some children when they start going to school (Rafaeli-Mor et al., 1999). And in adults, several repetitive ‘fidgets’ are associated with anxious personalities (e.g., Mehrabian and Friedman, 1986); and one WWII study suggested that they increase in college students during wartime (Young, 1947). People who nail-bite also have a higher prevalence of anxiety and depression—or, if veterans, a greater likelihood of Post Traumatic Stress Disorder (PTSD)—than do non-nail-biters (Maraz et al., 2017; Sisman et al., 2017; Litz et al., 1992). Furthermore, drugs that reduce anxiety or depression, such as tricyclics, often reduce these ARBs (e.g., Van Moffaert, 1995; Hwang et al., 2007; Shukla and Pandey, 2020). Imprisoning adults – highly negative in being boring, frightening, and a cause of anxiety and depression (e.g., Haney, 2003; Morin, 2016)—also dramatically increases ARBs: pacing becomes prevalent (e.g., Vivar, 2014); rituals like repeatedly folding clothes may develop (Prison UK, 2014); and harmful ARBs may emerge too, such as self-cutting (e.g., Haines et al., 1995; Prison UK, 2014). Being isolated when captive has further effects (e.g., Metcalf, 2020); most prisoners-of-war report pacing, repetitively exercising and/or performing ‘ritualistic activities’ after 4 weeks of solitary confinement (Deaton et al., 1977). And as discussed below, such effects of confinement recur in other groups of people, like children raised from babyhood in institutions (with obvious relevance for caged animals).
Furthermore, some neurotypical human ARBs display graded relationships with the intensity of negative mood and/or extent of previous negative experiences, indicating potentially good responsiveness as welfare indicators. ARBs positively covaried with anxiety in one study of school-children (Keating et al., 2023). Likewise, tooth-grinding severity positively covaries with anxiety in college students (Câmara-Souza et al., 2023), while pacing increases with increasing time in jail (Vivar, 2014) and more weeks in solitary confinement (Deaton et al., 1977). Nail-biting severity is also greater in adolescents who had experienced more traumas as children (e.g., death, divorce, abuse, illness: Yasir et al., 2021); while ‘lifetime stress scores’, a tally of all past negative events (here, weighted by their aversiveness), predicts levels of tooth-grinding in ‘Type A’ personality (i.e., neurotic) adults (Pingitore et al., 1991). Furthermore, in juvenile residential homes, certain ARB forms (e.g., self-scratching) are most prevalent in children whose previous family situations were judged to involve abuse, and more kinds of ARBs also emerge the longer children are in these homes (Tröster, 1994).
Children Raised in Deprivation
After the Ceausescu regime in Romania collapsed in the 1980s, tens of thousands of children were rescued from so-called ‘orphanages’ where they experienced extreme neglect: little social contact or affection from caregivers (and sometimes abuse), nutritional restriction, and often confinement in their cribs (e.g., Fisher et al., 1997; Chugani et al., 2001; Beckett et al., 2002). These children would have been neurotypical if not institutionalized, yet instead their brain development was altered, arguably compromised (as indicated by e.g., small cranial volumes, enlarged amygdalae, altered cortico-basal ganglia connections: Chugani et al., 2001; Behen et al., 2009; Tottenham et al., 2010). There seems little doubt that welfare was very poor there; their institutionalisation caused malnourishment (Beckett et al., 2002), physical health problems (Beckett et al., 2003), anxiety (Ellis et al., 2004; Nelson et al., 2014), signs of cellular ageing (shortened telomeres: Drury et al., 2012; cf. Bateson and Seeker, in press), and sometimes even physical scars from abuse (Chugani et al., 2001). ARBs were also prevalent in these institutions (e.g., Carlson and Earls, 1997): 50-80% of children rocked back and forth on hands and knees, or stood, holding their cribs’ railings, and shifting from foot to foot (Bos and Van Balen, 2010; Fisher et al., 1997).
Some ARBs, including body-rocking, repetitive hand movements, head-banging and eye-poking, even persisted once children were adopted into normal family homes (Beckett et al., 2002; Benoit et al., 1996; Fisher et al., 1997; Maclean, 2003). Three factors, all welfare-relevant, predicted this. One was how long children had lived in the previous poor conditions, post-adoption ARBs being more prevalent if they had spent more time institutionalized (Beckett et al., 2002; Bos and Van Balen, 2010). This likely reflected greater neurological dysfunction: children showing the most persistent ARBs post-adoption had poorer cognitive abilities, and other ‘quasi-autistic’ behaviours such as circumscribed interests (Beckett et al., 2002; Bos and Van Balen, 2010; Rutter et al., 2007). Second, ARBs declined more (often even vanishing altogether), the longer children spent in normal family homes (Fisher et al., 1997; Hoksbergen et al., 2005). Having such stable home environments also allowed for developmental “catch-up” of brain growth (Sheridan et al., 2012) and reduced risks of psychopathology in early adolescence (Humphreys et al., 2015). Third, ‘quasi-autistic’ behaviour was also more marked in children who did not have normal, secure attachments with their adopted parents, although what is cause versus effect here is uncertain (Sonuga-Barke et al., 2020).
Children with Adverse Autoimmune Responses to Infections
Some children who would otherwise have been neurotypical develop ARBs after infections as part of an acute autoimmune response known as Pediatric Acute-onset Neuropsychiatric Syndrome (‘PANS’; Swedo, 2012) or Pediatric Autoimmune Neuropsychiatric Disorders Associated with Streptococcal Infections (‘PANDAS’). However, PANS/PANDAS remains a controversial diagnosis (La Bella et al., 2023) and is still considered a research topic or clinical concept rather than a formal psychiatric/medical diagnosis. PANS/PANDAS is characterized by the sudden onset of obsessions, compulsions, and/or tics and a variety of neuropsychiatric symptoms (Swedo et al., 2024). These are thought to be mediated by inflammation of the brain, specifically the basal ganglia (reviewed by Leonardi et al., 2024); and they therefore co-occur with other behavioural changes, including in eating habits, motor skills, hyperactivity and inattention.
Affected children may show suicidality and self-injury (e.g., Wang et al., 2024); anxiety, irritability, and extreme mood swings; as well as sleep disturbance and bed-wetting (National Institute of Mental Health, 2025; PANDAS Network, 2025). This suggests that poor welfare (inevitable during illness) is not just involved in the initial trigger but also co-occurs with these childrens’ new ARBs. Furthermore, chronic stress, by promoting inflammation and immune dysregulation, may act as a risk factor for symptom onset following infection and for subsequent flare-ups (reviewed by Gagliano et al., 2025). For example, children who experience repeated infections display higher rates of ARBs (Murphy et al., 2007). And stress may also exacerbate existing symptoms: during the COVID-19 lockdown, children with PANS/PANDAS who experienced heightened stress and daily disruptions showed worsening of multiple symptoms including motor and vocal tics, irritability, anxiety, and sadness, as well as both the emergence of new ARBs symptoms such as complex tics, and new signs of poor welfare such as generalized anxiety and fears related to illness or death (Guido et al., 2021). Similar interactions between past streptococcal infections, stress, and symptom severity have also been observed in certain clinical populations, as we will see below.
2.1.2. Human Clinical Populations’ ARBs
Humans with Clinical Conditions Characterised by ARBs
ARBs define several clinical conditions, including the choreas of Huntington’s disease, tics of Tourette’s syndrome, stereotypies of Rett’s syndrome, autism spectrum disorder and stereotyped movement disorders, body-focused repetitive behaviours like trichotillomania (hair plucking) and excoriation (skin-picking) disorder, and rituals of obsessive-compulsive disorder (OCD). These ARBs are very diverse: they look different to observers, feel different to performers, have different risk factors and correlates (including being male-biased for some conditions, female-biased for others), and are heterogeneous even within each condition. Nevertheless, they share commonalities: in terms of broad mechanism, they share evidence of altered fronto-striatal/basal ganglia functioning (e.g., Langen et al., 2011; Shukla and Pandey, 2020). Most are therefore viewed as movement or neurodevelopmental disorders rather than primarily affective conditions. However, despite that, "most are exacerbated by emotional stress" (Shulman et al., 1996), and as we show below, these ARBs are often associated with anxious moods or mood disorders.
As Table 1 summarizes, negative moods, especially anxious ones, are typically elevated in humans with these conditions (compared to neurotypical controls). And within each condition, negative experiences like being bullied, imprisoned, exposed to recent trauma, or relationship problems are often associated with, and likely cause, increased ARB (see Table 1). Correspondingly, increased ARB is often associated with more negative moods. Indeed, across subjects, ARB severity covaries in a graded, dose-response type way with scores for anxiety, distress or depression in populations affected by OCD, Tourette’s syndrome, autism spectrum disorder, Rett’s syndrome, and the body-focused ARBs of excoriation disorder and trichotillomania (see Table 1). Mood-enhancing drugs can also sometimes reduce such subjects’ ARBs. In particular, selective serotonin reuptake inhibitors (SSRIs) like fluoxetine (a.k.a. ‘Prozac’) are commonly used to treat OCD (e.g., Stein, 2002; Hugo et al., 2003; Singer, 2010; Lutz, 2014). They reportedly may also reduce chorea (Bhidayasiri and Truong, 2004), and additionally have been used to treat ARB and anxiety in people with autism (Ferretti et al., 2018; although recent meta-analysis indicates no overall effects of antidepressants: Maneeton et al., 2024). Clomipramine (a tricyclic antidepressant) can also be effective for stereotyped movement disorder or trichotillomania (reviewed by Shukla and Pandey, 2020), while benzodiazepines may help reduce the tics of Tourette’s, especially in anxious subjects (Singer, 2010).
These negative moods are sometimes severe enough to represent mood disorders (e.g., Grant et al., 2017b; Oakley et al., 2015; Özten et al., 2015; Spitzer et al., 2022). And given the roles that early life challenges and repeated negative experiences over the lifespan play in mood disorders (see Mendl et al., in press; Poirier, in press), it is perhaps unsurprising that such experiences can similarly help promote severe ARBs, at least in some conditions. Thus, the presence of a traumatic life event the year before onset can predict more chronic OCD, although not in all sub-groups studied (Goldberg et al., 2015). Compared to neurotypical controls, adults with excoriation disorder (Özten et al., 2015; Spitzer et al., 2022) and trichotillomania (Lochner et al., 2002; Özten et al., 2015) are more likely to have experienced early traumas (e.g., childhood abuse or neglect). They are also more likely to have experienced traumatic events in adulthood (excoriation disorder [Kłosowska et al., 2021; Spitzer et al., 2022]; trichotillomania [Ötzen et al., 2015]), as are adults with OCD (Horesh et al., 2018; Sarkhel et al., 2011; Vidal-Ribas et al., 2015; reviewed Goldberg et al., 2015).
Furthermore, within affected populations, the degree of exposure to such traumas can increase ARB severity. Thus, across adults with OCD or Tourette’s, more severe compulsions (Boger et al., 2020) or tics (Yang et al., 2022) are predicted by having had more severe or numerous adverse experiences in childhood. And such impacts are not restricted to childhood adversity. In adults with OCD or trichotillomania, more severe ARB is predicted by having previously experienced any major stressor in their lifetime (OCD [Cromer et al., 2007; Imthon et al., 2020]; trichotillomania [Houghton et al., 2016]). Furthermore—again suggesting graded dose-response effects, this time of cumulative stress—more severe OCD (reviewed Cromer et al., 2007; Sarkhel et al., 2011; Vidal-Ribas et al., 2015) and more prolonged hair-plucking disorders (Gershuny et al., 2006), as well as more severe tics in Tourette’s syndrome (Horesh et al., 2018; Steinberg et al., 2013), are predicted by more severe, numerous or diverse adverse experiences over adulthood. In parallel, Lin et al. (2010) find that prior Streptococcus infections and psychosocial stress have additive effects on OCD and Tourette’s tic severity.
Thus, in populations at risk of clinical conditions characterized by ARBs, negative moods or mood disorders commonly co-occur or even covary with these ARBs. Furthermore, this may reflect increased exposure and/or increased sensitivity to major stressors (and/or infections) experienced in infancy or cumulatively over the lifespan.
Other Neuroatypical Humans, if Institutionalized
People with developmental disabilities or psychiatric conditions that are not diagnostically characterized by ARB (e.g., Down’s syndrome), may still display such behaviours if living in institutions (e.g., Kaufman and Levitt, 1965; Francis, 1971; Phemister et al., 1978; reviewed Tröster, 1994 and Hall, 2003). For children, such institutions generally provided less social contact and fewer toys, and are more likely to restrain children in cots, chairs or playpens, compared to family homes (e.g., Francis, 1971; Phemister et al., 1978). Tröster (1994) further suggested a graded relationship between the duration of institutionalization and ARB severity, though not all cited studies support this claim clearly. Similar dose-response/graded effects have been proposed for adults with schizophrenia (Luchins et al., 1992), though this claim was not supported by evidence.
2.1.3. Counter-Examples: When Human ARBs Seem to Fail Test 1
When Human ARBs Fail to Increase During Negative States/Experiences
Despite the evidence above, if used as welfare indicators for humans, human ARBs would be very prone to false nulls. For example, children with neglectful parents are unhappy, and yet (unlike children with depressed or abusive parents) they do not perform more ARBs than well-parented controls (Egeland et al. 1983), instead showing both apathy and non-repetitive behavioural changes. Likewise, ‘lifetime stress scores’ do not predict levels of tooth-grinding in people who are not ‘Type A’ personality (Pingitore et al., 1991). Similarly, a substantial minority of children in Romanian orphanages did not develop ARBs, despite being no less anxious or physically healthier than their ARB-prone peers (Beckett et al., 2003, Bos et al., 2010); and not every prisoner-of-war reports having paced, repetitively exercised and/or performed ‘ritualistic activities’, especially at the beginning of captivity (Deaton et al., 1977). Furthermore, while Streptococcus infection is common in children (about 12% may carry the bacteria asymptomatically, and roughly 37% of sore throats are caused by it: Martin, 2022; Shaikh et al., 2010), just a tiny fraction develop PANDAS/PANS (roughly 1 in 10 000 children: Kronenberg et al., 2023; Wald et al., 2023): only individuals predisposed genetically and/or immunologically are at risk (Gagliano et al., 2025; Swedo et al., 2024). Finally, not every study of ARBs and anxiety in autism finds links (e.g., Sullivan et al., 2014); and SSRIs are also often ineffective for such people too (Maneeton et al., 2024; Williams et al., 2011).
Furthermore, many studies do not find links between exposure to stress or trauma and the onset of clinical conditions characterized by ARB. We could find no compelling evidence for this in ASD or SMD, for example. Furthermore, several studies find null results counter to those in Table 1: affected people were found not to have experienced more negative events (e.g., assaults, bereavements) over their lifetimes in some papers on OCD (Landau et al., 2011), skin-picking (Özten et al., 2015), and trichotillomania (Gershuny et al., 2016). For OCD, Goldberg et al. (2015) suggest genetic predispositions as an explanation (either increasing susceptibility to stressors or playing such a key role in disease aetiology that such environmental effects are negligible); and in their study, stressful life events only increased OCD severity (chronicity) in women with negligible familial OCD risk.
Although such patterns might be true nulls (reflecting psychological resilience to stressors in some fortunate people), we suspect they are false ones. To illustrate, roughly 70% of people worldwide experience at least one traumatic event in their lifetime (Benjet et al., 2016); and yet only a tiny subset respond by developing OCD, trichotillomania or skin picking (population prevalences ranging from just 1-4% for each of these conditions: Grant. 2019; Grant and Chamberlain, 2020; Stein et al., 2025), others instead developing different health conditions including depression (~5.7% of adults experiencing a depressive disorder globally: World Health Organization, 2025) or PTSD (lifetime prevalence ~3.9% globally: Koenen et al., 2017). Thus, while ARBs and mood disorders (i.e., depression, anxiety, PTSD) do often co-occur, the latter can often occur without the former (cf. Gershuny et al., 2006).
When Levels of Human ARB Are Higher During Positive States/Experiences than Negative Situations
In people prone to ARBs, ARB expression seems not to be a reliable indicator of current negative emotion. For one, bouts of ARB are often elicited by emotionally neutral or even positive triggers (not just negative ones). Thus, ARBs are also commonly performed during moments of excitement or intense focus that are not affectively negative (reviewed by: Muthugovindan and Singer, 2009; Srinivasan and Mink, 2012; e.g., in autism [Willemsen-Swinkels et al., 1998, Lanzarini et al., 2021] and SMD [Harris et al., 2008; Singer, 2009; Oakley et al., 2015]). In addition, they may be performed in moments of relaxation or calm (e.g., in trichotillomania: Chamberlain, 2007), or during low-arousal emotionally neutral activities (e.g., body rocking while waiting for something/someone: Rafaeli-Mor et al., 1999), as well as in the absence of any apparent emotional triggers (e.g., ARBs in Rett syndrome: Wales et al., 2004; Quest et al., 2014; Cianfaglione et al., 2016; see also Shulman et al., [1996] and Tröster, [1994] for further examples on immediate context of ARB performance). Furthermore, ARB bouts may have immediately beneficial consequences, with feedback from performance being emotionally positive. In neurotypical humans, for example, chewing gum can reduce anxious feelings and cortisol responses during acute stress exposure, as well as ease day-to-day stress (e.g., feelings of agitation or tension) in self-reported regular chewers (Zibell and Madansky, 2009; Smith, 2016), while rhythmic leg-swinging in school-children can lower heart rate (Soussignan and Koch, 1985). In trichotillomania, the act of hair plucking is sometimes described as gratifying or relieving (Chamberlain 2007), while in skin-picking disorder, bouts can also be accompanied by a sense of relief (Kłosowska et al., 2021). Likewise, in autism, ARBs can be calming, soothing and anxiety-reducing (Uljarević et al., 2017), and associated with heart rate deceleration (Heathers et al., 2019)—perhaps explaining their frequent occurrence during periods of high stress or anxiety.
One example of a “false positive” for Test 1 is more relevant to longer-term moods: cases where subjects develop ARBs (e.g., ‘punding’: Fasano and Petrovic, 2010) following the prolonged use of dopaminergic agents. These may be prescribed for symptom-reduction in Parkinson’s disease, or taken recreationally for their euphoric effects (although this subjective pleasure typically declines in the chronic users who are also prone to ARB: Robinson and Berridge, 2009). In these cases of ‘false leads’, ARBs thus arise not from poor welfare but from voluntary drug-taking that over time induces dysregulation of dopaminergic systems, causing behavioural disinhibition (something also true for non-human animals, as we will see in Test 4).
2.2. Test 2: Effects of Aversive Experiences on Animals’ ARBs
Moving away from humans, many aversive experiences promote ARB performance in animals too. Here, we cover three that are particularly well studied: barren housing conditions; exposure to threatened or actual physical harm; and food deprivation.
2.2.1. The Effects of Barren Housing
ARBs are often promoted by long-term housing conditions lacking ‘enrichments’: resources that we know (thanks to studies of motivation) that animals prefer (see Table 2). Assuming that barren environments are therefore generally aversive even to species whose housing- or resource- preferences have not been assessed, then their impact is even more ubiquitous, as shown by studies of ARBs in birds (e.g., orange-winged Amazon parrots, Amazon amazonica [Meehan et al., 2004]; Japanese quail, Corturnix japonica [Laurence et al., 2014]; and chickens, Gallus gallus domesticus [van Staaveren et al., 2021]); along with meta-analyses of ARB in multiple species, mainly mammalian, in zoos [Shyne, 2006; Swaisgood and Shepherdson, 2006, 2005]).
Furthermore, such effects of housing can be graded, ARBs declining the better resourced or more preferred are animals’ living conditions. In American mink (Neogale vision), for instance, common locomotor ARBs like pacing are not reduced by merely adding a ball and chew chain to farm cages (Meagher et al., 2014). However, they are approximately halved by providing suspended tunnels as well as chewing objects (Hansen et al., 2007), or highly valued running wheels (Malmkvist et al., 2024); reduced by three-quarters or more by climbing structures and highly motivating large cages full of diverse preferred resources (Díez-León et al., 2013; 2016); and absent altogether in mink housed in large outdoor enclosures with ponds (Rauch, pers. comm.). Similarly, the more motivated animals are to engage with particular resources, the more effectively these reduce ARB. Thus, compared to lifelong barren-housed laboratory mice (Mus musculus), mice moved to barren cages from well-resourced ones were more likely to route-trace there and more motivated to leave these sub-optimal conditions (pushing heavier weights to do so) (Latham and Mason, 2010). And within middle-aged barren-raised mice, those individuals who were most motivated to access large well-resourced cages also showed the greatest ARB reductions if rehomed there (Tilly et al., 2010). Perhaps similarly in bank voles (Clerionomys glareolus), as animals mature, their preference for cover decreases (Cooper and Nicol, 1991), and correspondingly, providing them with cover also has increasingly less ability to reduce their jumping ARB as they age (Cooper et al., 1996).
The duration of barren housing can have similar graded effects. For example, in chickens, feather pecking increases steadily over 3 months without litter, while not doing so if litter is present over that same period (Blokhuis and Arkes, 1984); and in young orange-winged Amazon parrots (Meehan et al., 2004), ARBs similarly increase over 8 months in barren housing, while barely doing so if instead provisioned with foraging, perching, climbing, swinging, chewing and exploratory opportunities. Conversely, ‘enrichment’ provision can also have greater impacts on ARB the longer the duration of provision (as suggested in a meta-analysis of zoo animals: Swaisgood and Shepherdson, 2006).
2.2.2. The Effects of Actual or Threatened Physical harm
Studies of acrimonious social groups and research animals undergoing aversive scientific procedures show that recurrent threats or experiences of physical harm can exacerbate ARBs. Across group-housed chimpanzees (Pan troglodytes), individual time budgets spent performing self-directed ARBs and nodding covaried with the proportion of life spent in research protocols (Warniment and Brent, 1996; although whether this reflected research procedures or just being housed indoors, often alone—see Test 3—could not be ascertained). An impact of invasive protocols has also been identified in rhesus monkeys (Macacca mulatta). Across dozens of single-housed monkeys, experiencing more blood draws in recent years is a risk factor for self-injurious behaviour, largely self-biting (Novak, 2003; Lutz et al., 2003), and also repetitive ‘saluting’ (Lutz et al., 2003b). In addition, the number of research projects a monkey was used in over their lifetime helped predict levels of ‘motor stereotypy’ (a broad category including pacing, twisting and bouncing: Gottlieb et al., 2013). And if young maternally-deprived rhesus monkeys were exposed to 4-8 weeks of Chronic Unpredictable Stress, CUS (a paradigm meant to induce depression-like behaviours, including restraint and ‘intimidation’ from humans brandishing capture nets), this increased the time spent pacing, rocking, bouncing, and hair pulling (Zhang et al., 2016). Finally, for a very different ARB in a very different taxon, similar effects also occur for feather-pecking in chickens: this increases if birds are experimentally exposed to CUS (via procedures such as restraint and social disruption: Mindus et al., 2021).
Moving to physical harms from conspecifics, across pig farms, tail-biting is higher in facilities where competition for food is more intense (reviewed by Boyle et al., 2022), consistent with a role for increased agonism in ARB; and levels of food deprivation that increase oral ARBs in both sows and broiler breeders, also increase intraspecific aggression too, although obviously cause and effect cannot be ascertained here (reviewed by D’Eath et al., 2009). Likewise, in mice, levels of route-tracing, jumping and similar ARBs are higher in barren cages that also elevate aggression between cage-mates (Clipperton-Allen et al., 2015; Harper et al., 2015; Nip et al., 2019); while in well-resourced cages, if ‘enrichments’ are clustered and reduced in number, levels of ARB and aggression both increase in a correlated way (Akre et al., 2011).
2.2.3. The Effects of Food Restriction
Commercially, some farm animals are often fed much less food than they would consume with free access, especially broiler chicken breeders (Savory et al., 1993; Dixon et al., 2014), sows (Lawrence and Illius, 1989), and dairy calves (De Paula Vieira et al., 2008). This promotes oral ARBs: object-pecking in chickens (Savory and Maros, 1993; Kostal et al., 1992), chain and bar manipulation in pigs (Spoolder et al., 1995), and non-nutritive sucking in calves (de Passillé and Rushen, 1997; Goeller et al., 2023; Rushen and de Passillé, 1995). In hungry chickens, pacing can occur too (Duncan and Woodgush, 1972; Savory and Mann, 1999), as can increased feather-pecking (Morrissey et al., 2014). Furthermore, if levels of feed deprivation are varied, dose-response-like effects may occur: food-thwarted hens show more pacing the less food they are given, and the more days the restriction continues (Duncan and Wood-Gush, 1972); broiler breeders show more object-pecking the more food-deprived they are (Savory et al., 1993); and the same holds for bar-biting and related ARBs in sows (Appleby and Lawrence, 1987; Appleby et al., 1989).
Food restriction similarly exacerbates ARBs in other species. Cases include oral ARBs in dairy cows (Redbo et al., 1996); rocking and swaying in chimpanzees, along with ‘regurgitation and reingestion’ (an oral ARB as disgusting-looking as it sounds: Berkson and Mason, 1964; Baker and Easley, 1996); pacing in captive Carnivora (e.g., mink: Bildsøe et al., 1991; Mason, 1993); and repetitive jumping to the cage-lid and climbing on it in DBA/2 mice (Cabib and Bonaventura, 1997). Assuming that food restriction is similarly aversive to these animals (e.g., because access to food is crucial for fitness, cf. Test 3), this supplies further cases passing Test 2.
2.2.4. Counter-Examples: When ARBs Seem to Fail Test 2
When ARBs Fail to Increase in Aversive Situations, or to Decrease in Preferred Ones
Despite the supportive evidence above, null results do occur. These include potential delays before impoverished conditions begin to induce ARBs (e.g., ARB taking 3-4 weeks to emerge in isolated, barren-housed young Amazon parrots; Meehan et al., 2004) and ceiling effects if these conditions are prolonged. For instance, in these parrots (Meehan et al., 2004), and also mice (Nip, 2018), ARBs increase over the first weeks or months of barren housing, but performance then plateaus. In addition, seemingly preferred resources may sometimes be ineffective for particular ARBs or particular populations. As examples, in male mink, large complex cages were less effective at abolishing scrabbling at cage partitions (a conspecific-directed ARB reduced by removing male neighbours) than they were at abolishing ‘whole body’ ARBs (e.g., pacing) and repetitive head movements (Polanco et al., 2018); and likewise if mink are given running wheels, scrabbling does not decline (nor an oral ARB: bar-licking), despite pacing and head-twirling/nodding doing so (Malmkvist et al., 2024). Providing swimming water for mink also does not always reduce their ARBs (reviewed by Vinke et al., 2008). Similarly, ‘enrichments’ typically fail to reduce SIB in rhesus monkeys, despite reducing rocking and pacing (Lutz et al., 2003b; Novak et al., 2006); while complex pens did not reduce tongue-rolling in dairy calves despite reducing allo-sucking (Zhang et al., 2021).
In other cases, ARBs are hard to reduce once they have appeared, even using treatments that would be effective if used pre-emptively (Novak et al., 2006). Such ‘treatment resistance’ varies between different forms of ARB and may also increase with age. To illustrate, being raised in large, well-resourced cages successfully reduces the prevalence of fur-plucking in mice (Bechard et al., 2011), but for mice in small, barren cages, once this ARB has developed, being moved to well-resourced conditions fails to reduce it (Young et al., in prep.). This is despite route-tracing, bar-mouthing and related ARBs greatly declining in such cages (Forget, 2025; Tilly et al., 2010).
Turning to age effects, ARBs can become harder to alleviate with well-resourced housing in rodents who are barren-housed into middle age or beyond (e.g., Hadley et al., 2006; Tilly et al., 2010). These persistent ARBs could thus perhaps be mere ‘scars of the past’ that fail to reflect animals’ current, presumably improved welfare (e.g., Mason, 1991). However, there are alternative explanations which do not involve a failure of Test 2, as we will consider in the Discussion. (And we will meet a somewhat similar uncertainty in the next section, concerning the lasting protective effects of well-resourced housing).
As two final null findings relating to barren versus well-resourced housing, sometimes its impact on ARB appears context-dependent. Thus, providing swimming water to mink barely reduced ARBs if their cages were large, despite greatly reducing them if their cages were small (Hansen and Jeppesen, 2000). Likewise, stereotypic pacing was no greater in barren-housed quail versus those housed with plastic grass and a cork screen (to peck and to hide behind), until birds were subject to a CUS regime (Laurence et al., 2014).
Null findings also occur in some studies of physical threat. In classic, albeit very unpleasant, work by Seligman (e.g., Maier and Seligman, 1976), dogs were subjected to repeated electric shocks in an arena. Initially, they cried, yelped and tried to escape, but if shocks were inescapable, even after several trials, the dogs never reportedly developed ARB (becoming quiet and passive instead). Turning to rhesus monkeys in biomedical research, Lutz et al. (2003b) did not find blood draws to be a risk factor for most ARBs (only self-biting and saluting); Gottlieb et al. (2013) did not find that the number of research protocols experienced predicted self-injurious behaviour, only locomotor ARBs; and in pig-tailed macaques, Macaca nemestrina (Bellanca and Crockett, 2002), no ARB was predicted by the invasiveness of the research they were involved in (although the number of past protocols was not taken into account). In their CUS work, Zhang et al. (2016) did not find significant effects in mother-reared animals; in the maternally-deprived subjects who did develop significantly more ARB, this was not seen after 1 week of CUS, only more prolonged treatment; and levels after 8 weeks of CUS were not significantly elevated over those seen at 4 weeks. Turning to agonism by conspecifics, some species respond to sustained social threat with inactivity rather than ARB, notably cynomolgus monkeys (Macaca fascicularis) (e.g., Shively et al., 2005). In two further null findings concerning acute effects in rhesus monkeys, exposure to a stranger or videotapes of fighting did not induce bouts of SIB, despite increasing aggression (Lutz et al., 2003a), and stress-inducing instances of agonism between nearby conspecifics did not induce bouts of pacing (Poirier et al., 2019b). Similarly, mink hearing the screams of alarmed conspecifics tend to hide, freeze or show vigilance rather than perform ARB (GJM, pers. obs.)
Turning to food restriction, ARBs seem not to emerge immediately after deprivation, even though animals must be hungry. Thus, in Duncan and Wood-Gush (1972)’s food-fraunted hens and Cabib and Bonaventura (1997)‘s food-restricted mice, only after 3-8 days of treatment did the chickens pace and DBA/2 mice start repetitively climbing. A similar delayed effect of daily food restriction (combined with tethering) is seen in pigs, where oral ARBs in young females take several days to develop (e.g., Cronin and Wiepkema, 1984). Cabib and Bonaventura’s (1997) mouse study also illustrates the importance of genotype: despite being food restricted for as long as the DBA/2s, and losing just as much weight, C57Bl/6 mice developed little cage-lid climbing. And in further evidence that inactivity helps promote null results, object-pecking increases if broiler breeders are food restricted in the daytime, but not if they are food restricted at night (despite similar physiological signs of deprivation), because of birds’ natural inactivity at that time (Dixon et al., 2022).
When Levels of ARB Are Higher in Preferred than Aversive Situations
Occasionally, studies of relatively preferred stimuli reveal ways in which ARBs may generate ‘false alarms’ or false leads as welfare indicators. In one subtle effect, beef cattle supplied with grooming brushes showed more tongue-rolling; however, this increase was much smaller than their concomitant decrease in bar-licking, such that overall, oral ARBs did fall (Table 2, Park et al., 2020). In mink, farmed animals given simple toy-like enrichments (e.g., balls) increased their performance of locomotor ARBs (e.g., pacing and route-tracing: Jeppesen and Falkenberg, 1990; Meagher et al., 2014). However, overall activity levels were measured in the second study and found to have increased too; statistically correcting for this then removed the apparent increase in pacing (Meagher et al., 2014). Activity was also a confound in another mink study, where animals were repeatedly restrained in a tiny catching cage (something they avoid if they can) for an hour daily for a week (Bildsøe et al., 1991). During this time, their ARBs fell to very low levels, but inactivity dramatically increased. If we assume that repeated spatial restriction is also aversive for deer mice, then similar suppressive effects on ARB were reported by Burke and colleagues (2022).
As further possible examples of ARBs being lower in more aversive situations, in deer mice (Hadley et al., 2006), African striped mice (Rhabdomys pumilio: Jones et al., 2011) and mink (Díez-León et al., 2016) raised lifelong in well-resourced housing but then moved to barren cages, the prevalence and/or individual levels of ARB remain lower than those of conspecifics housed lifelong in barren conditions. These previously ‘enriched’ animals probably have poorer current welfare than conspecifics who have never known better (Jones et al., 2011a; Latham and Mason, 2010). However, their ARBs do not reflect this (perhaps because they are too neurologically normal to perform sustained ARBs, no matter how frustrated they are in their new, sub-optimal housing). These ‘housing legacy’ effects are still a welfare puzzle, however: one we return to in the Discussion.
Finally, anticipating and consuming food can also have paradoxical effects: despite arguably being a positive moment in an animal’s day, the imminent arrival of a signalled meal often elicits bouts of ARB. Examples include pacing in broiler breeders (Kostal et al., 1992; Kostal and Savory, 1996); weaving and head-bobbling in Asian (Elephas maximus) and African (Loxodonta africana) elephants (Friend, 1999); pacing in American mink and other Carnivora (Mason and Mendl, 1997); self-directed ARBs, pacing, rocking and wall-licking in stump-tailed macaques (Macaca arctoides: Waitt and Buchanan-Smith, 2001); and weaving in stabled horses (Equus caballus: McAfee et al., 2002; Wickens, 2009). Ingesting a meal can also elicit post-prandial ARBs, typically oral, as seen in calves (de Passillé et al., 1992; 1993; Rushen and de Passillé, 1995), sows (Terlouw and Lawrence, 1993; Robert et al., 2002) and broiler chickens (Kostal et al., 1992). And in some dogs, food (along with ‘happiness’ and ‘excitement’) is a trigger for tail-chasing (Tiira et al., 2012). Furthermore, sometimes the better the meal, the more ARB is promoted. Thus in calves, ingesting milk induces non-nutritive sucking, but ingesting the same volume of water does not (de Passillé et al., 1992); in hungry sows, a ‘surprise extra meal’ triggers extra ARB performance (Terlouw et al., 1993); in chimpanzees prone to regurgitation and re-ingestion, this ARB often follows eating treats and palatable foods (e.g., Baker and Easley, 1996); and bouts of post-prandial stereotypic object-pecking pecking by broiler breeders increase in length, the larger the preceding meal (Savory and Mann, 1999). Thus, while sustained hunger can promote ARB, paradoxically, so too can meals.
2.3. Test 3: The Effects of Threats to Fitness on Animals’ ARBs
Along with food deprivation, other threats to fitness (ancestral ones that animals should have evolved to find negative promote ARB performance too. Again, we will survey three particularly well-documented examples of this type of challenge here: illness and infection; early maternal/parental loss (in species naturally reliant on parental care); and broken or absent social bonds (in naturally social species).
2.3.1. The Effects of Illness and Infection
Infection or inflammation can cause or predict elevated levels of ARB (e.g., Desmarchelier et al., 2016; Doneley, 2009; Maslak et al., 2013). Comparisons across farms reveal that in chickens, feather-pecking is more likely in flocks affected by egg peritonitis and infectious bronchitis (Green et al., 2000); and that in pigs, tail- and ear-biting is more likely in herds with high levels of respiratory disease and sometimes arthritis (Boyle et al., 2022). Comparisons across individual horses and pigs have also revealed associations between oral ARB performance and gastritis or gastric ulcers (Arias-Esquivel et al., 2024; Bergeron et al., 2006; Hemmings et al., 2007), and equine colic (Scantlebury et al., 2015). And treatment interventions in veterinary studies of dogs indicate that clinical conditions can play a causal role. In 19 dogs prone to ‘surface-licking’ (Bécuwe-Bonnet et al., 2012), 15 had gastrointestinal disorders; and within 17 treated with hypoallergenic diets and antacid/anti-nausea medication, the ARB waned or vanished in 10. Likewise, in 9 dogs showing repetitive ‘fly snapping’ (repetitive biting at the air), all had gastric or duodenal problems; and within eight treated with medication and dietary changes, the ARB declined or vanished in 7 (Frank et al., 2012).
A possible reason for such patterns, at least for farm-level associations, is that sub-optimal husbandry exacerbates both infectious disease and ARBs, without a direct causal connection between the two (Boyle et al., 2022; Green et al., 2000). Alternative, causal hypotheses include that ARBs are exacerbated by the pain and discomfort of clinical conditions (suggested by Bécuwe-Bonnet et al., 2012; Desmarchelier, 2019; Hemmings et al., 2007); for gastric issues, that oral ARBs are attempts to alleviate this by generating and swallowing saliva (Bergeron et al., 2006; Hemmings et al., 2007); and/or that ARBs reflect autoimmune-mediated neuroinflammation akin to ‘PANS/PANDAS’ (as covered under Test 1; see also Nordgreen et al., 2020; Wilson et al., 2023). Some animal models support this latter perspective: rats and mice show ARBs like head bobbing and excessive self-grooming after repeated streptococcal immunization or infection (reviewed by: Mora et al., 2020; also see Wilson et al., 2023). Whether autoimmune responses induce other animal ARBs is little explored, but Nordgreen et al. (2020) propose a similar hypothesis for farmed pigs.
2.3.2. The Effects of Early Parental Loss
The absence of parental care in infancy, in a way that would increase mortality in the wild, often increases ARB. Thus, the prevalence of feather-pecking in chickens may be increased in chicks raised without a hen rather than with (Perré et al., 2002); and data from adult zoo parrots indicate that birds who were human-reared perform more ARB than parent-raised birds (Williams et al., 2017). Likewise, in pet parrots, surveys suggest that feather-plucking is more prevalent in human-reared birds than parent-raised ones (Costa et al., 2016); and that within human-reared birds, those removed earliest from the nest develop the most ARB (Schmid et al., 2006). From other owner surveys, kittens who suck body parts emerge as being much more likely to be orphans than those who do not (Delgado et al., 2020); cats who overgroom as adults, as well as tail-chasing dogs, were reportedly removed earlier from their mothers than non-affected conspecifics (Ahola et al., 2017; Tiira et al., 2012); and equine facilities that separate mothers from foals rather than allowing natural weaning have more prevalent ARBs (Parker et al., 2008).
Experimental work on mammals reveals similar patterns (Latham and Mason, 2007), with primates being particularly well researched. One infamous early study removed rhesus monkeys from their mothers immediately after birth (isolating them in barren cages). Infants soon developed digit-sucking, self-biting, and rhythmic rocking (Cross and Harlow, 1965); and the latter two ARBs persisted into adulthood, new forms like pacing also developing (Cross and Harlow, 1965; Harlow and Harlow, 1962). In another study, when juvenile squirrel monkeys (Saimiri sciureus) were separated from their mothers for 5 days, they responded by pacing (ceasing when reunited). In today’s research primates, premature maternal separation (although not involving isolation, thanks to ‘nursery rearing’) is still a risk factor for ARBs like pacing, rocking, bouncing, flipping, hair-pulling and self-injurious behaviour, displayed years later in adulthood (e.g., Bellanca and Crockett, 2002; Lutz et al., 2003b; 2007; Rommeck et al., 2009b; Gottlieb et al., 2013; see also Zhang et al., 2016). Furthermore, the magnitude of such effects may covary in a graded way with parental deprivation: across research chimpanzees, there was an inverse linear relationship between adults’ ARB levels and the ages at which, years earlier, they had been removed from their mothers (Warniment and Brent 1996).
Turning to non-primates, ARBs in adult rodents are more time-consuming in subjects experimentally removed from their mothers at younger rather than older ages, as seen in CD-1 laboratory mice (Würbel and Stauffacher, 1997) and African striped mice (Jones et al., 2010b). Striped mice raised by mothers alone, instead of biparentally as would happen in nature, also display more ARB in adulthood (Jones et al., 2010a). Similar experiments on farms find that that tail-biting is more prevalent, and pacing more time-consuming, in adult mink removed from their mothers at younger rather than older ages (Mason, 1994; Jeppesen et al., 2000); that oral responses (object- and allo-sucking) soon appear in calves and lambs prematurely weaned from their mothers (e.g., Napolitano et al., 2008); and that in piglets, repetitive belly nosing (movements resembling udder massage) is also triggered by premature weaning (e.g., Weary et al., 1999; Widowski et al., 2008). Furthermore, in a graded way, belly-nosing effects are more prevalent and time-consuming the younger animals were when separated from their mothers (e.g., Weary et al., 1999; Widowski et al., 2008; Faccin et al., 2020).
2.3.3. The Effects of Social Isolation
In species for whom group-living is evolutionarily adaptive, being isolated or living in smaller groups than in the wild is another well-studied risk factor. In birds, such effects occur in canaries (Serinus canaria: Keiper, 1970; Sargent and Keiper, 1967) and several parrots (Meehan et al., 2003; Polverino et al., 2015; Williams et al., 2017). In mammals, they also occur in many species (albeit often confounded with space allowance), including rats, Rattus norvegicus (Hurst et al., 1998, 1997), mice (Lewis and Hurst, 2004), various primates (e.g., rhesus monkeys: Rommeck et al., 2009a; Gottlieb et al., 2013; chimpanzees: Brent et al., 1989), horses (e.g., Visser et al., 2008; Flamand et al., 2025), dholes (Canis alpinus: Maisch et al., 2017), dogs (e.g., Hubrecht et al., 1992; Corsetti et al., 2023), calves (e.g., Leruste et al., 2014) and perhaps pigs (though here social contact was confounded with straw provision; e.g., Broom et al., 1995; Karlen et al., 2007). Past isolation can have lasting effects too (just as for maternal deprivation). In Hawaiian crows (Corvus hawaiiensis), for instance, being reared without conspecifics increases rates of various ARBs like self-biting, spinning, body flipping, and feather pecking in adulthood (Harvey et al., 2002). Furthermore, graded effects of the degree of social restriction have been demonstrated in some zoo animals. For elephants (African and Asian), how much time they currently spend alone predicts the time they devote to swaying, head-bobbing and other ARBs (Greco et al., 2016); while for zoo-housed primates, across 24 species the difference between average group sizes in the wild and those in captivity (typically smaller) predicts levels of self-plucking (Pomerantz et al., 2013).
The duration of treatment can also have dose-response-like effects, suggesting cumulative impacts over time. For example, in Broom et al.’s (1995) study, single-housed sows’ levels of sham-chewing and bar-biting increased four- to fivefold between their first to fourth parity (while group-housed animals’ ARBs stayed low over the same period). In orange-winged Amazon parrots (Meehan et al., 2003), young birds who were isolated steadily developed increasing levels of ARB over the 12 months of treatment (while no such effect occurred in pair-housed conspecifics). Likewise, in deer mice from well-resourced cages, 12 hours of isolation in a barren test cage every few days steadily increased how much ARB they displayed there (something not seen in control animals: Bechard et al., 2017; Figure 2).
Lasting effects of the degree of past isolation or social disruption can even have graded effects on ARB, once current conditions are normalised or statistically controlled for. Across rescued apes in a chimpanzee sanctuary, for example, levels of abnormal behaviour (including ARB) increased with age in animals who had lived in isolation prior to rescue (but did not in previously socially-housed subjects, whose levels were always low: Chen et al., 2025). Likewise, in laboratory rhesus monkeys, having spent more of their past time in isolation steadily increases their risk of several current ARBs (Gottlieb et al., 2013; Lutz et al., 2003b; Rommeck et al., 2009a; Vandeleest et al., 2011), and the same holds for pig-tailed macaques, Macaca nemestrina (Bellanca and Crockett, 2002: Figure 3). In rhesus monkeys, being paired with multiple different cage-mates over the lifespan (thus experiencing repeated social disruption) also steadily increases the risk of current ARBs (Gottlieb et al., 2013). Likewise, in Greco et al.’s (2016) zoo elephants, current ARB levels were predicted by the number of previous transfers between institutions: something typically involving broken social bonds, as animals are generally moved on their own.
2.3.4. Counter-Examples: When ARBs Seem to Fail Test 3
When ARBs Fail to Increase in Response to Threats to Fitness
Illness and infection often do not lead to ARB (see e.g., Maslak et al., 2013; Boyle et al., 2022). Furthermore, if housing conditions are improved, ARB may wane even if painful infections persist (as reported for one bear by Maslak et al., 2013).
Likewise, the impacts of parental loss on ARB are not always strong or robust. Some studies do not find that chicks raised with a hen develop less feather-pecking (Angevaare et al., 2012; Rodenburg et al., 2009), nor that feather-plucking is more prevalent in human-reared parrots than parent-raised ones (e.g., Ebisawa et al., 2021; Schmid et al., 2006); two mouse experiments failed to find effects of maternal separation age (Bailoo et al., 2020; Bechard et al., 2012); and the impact of early maternal loss in young adult mink wanes and then vanishes with age (GJM, pers. obs.).
In other cases, maternal loss only affects some but not all ARBs, again highlighting these behaviours’ heterogeneity. Thus despite causing belly-nosing, early weaning has no obvious influence on tail- and ear-biting in growing pigs (reviewed by Boyle et al., 2022); despite clear effects on self-directed ARBs, nursery-rearing does not increase motor ARBs like pacing and twirling in pigtailed macaques (Bellanca and Crockett, 2002); and despite increasing hair-plucking and rocking/twirling, early maternal loss has no significant effects on pacing in rhesus monkeys (Polanco, 2021).
Social isolation also shows some null effects. In one, repeated social isolation did not exacerbate ARBs in deer mice, if already ARB-prone thanks to living in small lab cages (see ceiling effects in Figure 2). Genotype plays a role too: in cynomolgus monkeys again, adult females show passive responses to social isolation: depression-like forms of inactivity rather than ARB (Li et al., 2013; Willard and Shively, 2012). Social isolation or instability also did not exacerbate ARBs in quail (though this may not be a failure to pass Test 3, since it was not clear that such manipulations reduced welfare: Miller and Mench, 2005; Schweitzer and Arnould, 2010). Other ARBs may be untouched by social isolation even as other forms increase. For example, despite its effects on motor or whole body ARBs, social isolation seems not to exacerbate feather-plucking in parrots (Mellor et al., 2021; Van Zeeland et al., 2009). Likewise, in rhesus monkeys, one study found that current social isolation increased locomotor and whole-body ARBs but not self-injurious behaviour (Gottlieb et al., 2013), while another found it increased rocking/twirling, and also hair-plucking, but not pacing (Polanco, 2021); and another found that past social isolation decreased current locomotor and whole-body ARBs, despite increasing self-directed forms like hair-plucking and self-biting (Lutz et al., 2003b).
2.3.5. When Levels of ARB Increase with Decreasing Threats to Fitness
When ARBs fail Test 3, this is far more often due to null effects than to ‘false leads’. Nevertheless, we know of two instances when ARBs paradoxically increase when threats to fitness are relatively high and decrease when they are relatively low. The first is in systems where ARBs are ubiquitous in physically healthy animals. Here, their sudden absence in particular individuals can indicate sickness or injury: we have personally seen such cases in both farmed mink and conventionally housed lab mice. The second concerns a particular behaviour of some apes: coprophagia (faeces-eating). Well-powered surveys of dozens of chimpanzees in zoos and sanctuaries link this with being mother-reared rather than nursery or human-reared (Jacobson et al., 2016; Nash et al., 1999), and with being raised in larger rather than smaller social groups (Hopper et al., 2016).
2.4. Test 4: The Effects of Drugs That Reduce Negative Affect on Animal ARBs
In Test 1, we reviewed several cases where mood-improving (e.g., anxiety-reducing drugs) reduce ARB performance in humans. In other species, both therapeutic treatments of individual animals, and larger-scale experimental studies, have shown that mood-improving drugs (anti-depressants and anxiolytics) often reduce ARBs here too. That these do enhance mood, rather than merely causing sedation (cf. concerns expressed by Carroll et al., 2023, 2020) may be indicated by other signs of improved welfare (e.g., reduced startle responses and less fearfulness: Bauer et al., 2013; Gage, 2005; Meers and Ödberg, 2005; Yalcin and Aytug, 2007); that animals may self-administer them (Olsson and Sherwin, 2006); and that they can reduce ARBs without impacting normal activity (e.g., Duncan and Wood-Gush, 1974; Meers and Ödberg, 2005; though cf. Olsson and Sherwin, 2006).
In the earliest of such studies, anxiolytic benzodiazepines reduced spot-picking and route-tracing in caged canaries (Ronald R. Keiper, 1969). Duncan and Wood-Gush (1974) found that benzodiazepines reduced repetitive pacing in their food-thwarted hungry hens (even preventing it, if given before pacing emerged). Likewise, for conventionally caged mice exposed to CUS, benzodiazepines significantly reduced the time they spent backflipping and route-tracing (Olsson and Sherwin, 2006). The anxiolytic buspirone (offered in combination with more environmental complexity) also eliminated self-biting in an American badger (Taxidea taxus: Gage, 2005).
Anti-depressants like SSRIs, and less commonly tricyclics, have also had success (e.g., Mills and Luescher, 2006; D’Angelo et al., 2014; Grados et al., 2016; Poroshinska et al., 2024). SSRIs have reduced self-injurious behaviours in rhesus monkeys (Fontenot et al., 2005), somersaulting and weaving in lab-housed vervet monkeys (Chlorocebus pygerythrus: Hugo et al., 2003), route-tracing and repetitive jumping in lab-caged, single-housed deer mice (Korff et al., 2009, 2008; Wolmarans et al., 2013) ARBs in highly stereotypic bank voles (Meers and Ödberg, 2005), and fur-chewing in farmed chinchillas (Chinchilla lanigera: Galeano et al., 2013). Fluoxetine has also reduced, sometimes even eliminated, pacing in zoo-housed bears of various species (e.g., Poulsen and Cambell Teskey, 2006; Yalcin and Aytug, 2007; Jeong et al., 2019), and a tiger (Baker, 2002), as well as self-licking and tail-chasing in pet dogs and ‘compulsive sucking’ in cats (e.g., Rapoport et al., 1992; Wynchank and Berk, 1998; Tynes and Sinn, 2014; Mosallanejad et al., 2015). Similarly, feather-picking in pet cockatoos has been reduced with clomipramine, a tricyclic (Seibert et al., 2004; van Zeeland, 2018).
2.4.1. Counter-Examples: When ARBs Seem to Fail Test 4
When ARBs Are Not Reduced (or Are Even Increased) by SSRIs or Benzodiazepines
Just as for humans (Test 1), purported anxiolytics and anti-depressants do not always reduce ARBs. Thus SSRIs do not always ameliorate feather-picking in parrots (Mertens, 1997; Mills and Luescher, 2006; Seibert, 2007; van Zeeland, 2018); SSRIs did not reduce hair-plucking, pacing, saluting or head tossing in Fontenot et al. (2005)‘s lab-housed rhesus macaques; and in bank voles, they failed to be effective in males, despite working in females (Schoenecker and Heller, 2003). Occasionally, they may even exacerbate ARB performance, as they did in low-ARB individual bank voles (Meers and Ödberg, 2005). Likewise, benzodiazepines reversed the ARB-suppressing effects of regular restraint in deer mice (Burke et al., 2022).
One possible pattern here is that antidepressants and anxiolytics may be most effective on the ARBs of subjects with very poor current welfare. In mice given midazolam by Olsson and Sherwin (2006), for example, ARBs only decreased in the CUS condition, not controls; while in bank voles, SSRIs were particularly effective on the ARBs of individuals stressed by handling and injections (Schoenecker and Heller, 2001). Conversely, they seem least effective on ARBs that have been performed for longer (e.g., as shown in food-frustrated hens by Duncan and Woodgush, 1974; and reported for pet cats and dogs [Luescher, 1997 cited by Mills and Luescher, 2006] and pacing zoo-housed bears [Jeong et al., 2019]): a pattern reminiscent of such ARBs’ similar resistance to treatment with ‘enrichment’ (see Test 2).
Of the three possible explanations for why these drugs do not always reduce ARBs, two represent failures of Test 4. In the first, altered activity levels act as confounds: if negative moods cause animals to spend much of their non-stereotypic time hiding or showing depression-like inactivity (a topic developed in the Discussion), then reducing these responses by improving mood could paradoxically free up active time for ARB. In the second, the drugs’ negligible impact could indicate that current mood contributes little to the current performance of ARB—analogous to the scar of the past suggestion for ARBs that resist treatment with ‘enrichment’ (see Test 2). But in the third possible explanation, Test 4 is not failed in contrast, because the drugs do not actually lift mood. After all, human research shows that antidepressants can fail, take weeks or months to work, or even be counter-productive (e.g., Nierenberg et al., 2000; Fergusson et al., 2005; Kverno et al., 2021). For example, for people with ‘agitated depression’, SSRIs typically make restlessness and other symptoms worse (Sohel et al., 2025). Which of these explanations is likely is currently hard to say.
When ARBs Are Increased by Drugs that Are Affect-Neutral or Even Promote Positive Affect
Studies on lab rats and mice show that animals self-administer and prefer contexts associated with stimulant/dopaminergic drugs such as cocaine, apomorphine and methamphetamines (reviewed by Sanchis-Segura and Spanagel, 2006). ARBs like repetitive licking, gnawing and sniffing are also induced during self-administered doses of such drugs, suggesting they co-occur with positive drug-induced states (e.g., Fowler et al., 2007; Schlussman et al., 1998). These drug-induced ARBs can even be potentiated by other mood-enhancing drugs such as SSRIs (Lamoureux et al., 2023). Thus, as for Test 1, some drugs that induce ARBs are not doing so via inducing negative moods; indeed, they may even be inducing positive states.
2.5. Test 5: ARBs and the Co-Occurrence of Other Validated Animal Welfare Indicators
2.5.1. When Treatments That Promote ARB Cause Other Signs of Negative Affect
Experiences that render ARBs prevalent and time-consuming very often induce other signs of poor welfare. A simple meta-analysis of publications on ‘stereotypies’ (Mason and Latham, 2004) found that across 196 studies comparing animals treated in ways that either increase or reduce this subset of ARBs, those increasing it induced other signs of poor welfare in 68% of cases: significantly above chance (see Figure 1b in Mason and Latham, 2004).
Admittedly, this involved very basic ‘vote-counting’, with no calculation of effect sizes, weighting by sample size, nor parsing out effects of species or treatment types. However, the pattern it revealed seems mirrored by much research since. For example, starting with studies relating to Test 2, barren housing that promotes ARBs in mink also decreases their spleen and thymus weights, increases their fear of humans, increases skeletal asymmetry, and reduces both male testosterone levels and female fecundity (Díez-León et al., 2013, 2016; Meagher et al., 2014); while barren housing that promotes ARB in mice also increases behavioural and neurological signs of depression and anxiety, decreases resilience to disease, and shortens lifespans (Cait et al., 2022). Calves in barren pens have slower growth than conspecifics housed with ARB-reducing resources (Zhang et al., 2021). Hamsters (Mesocricetus auratus) housed without deep bedding for burrowing do not just perform more wire-gnawing: they also have poorer body condition (Hauzenberger et al., 2006). Dholes (Canis alpinus) housed in small bare enclosures are not just more prone to pacing: they also reportedly have more reproductive failure (Maisch et al., 2017). Rhesus macaques moved to a new building (and given new neighbours), do not just show more SIB: they also have more elevated cortisol and disrupted sleep (Davenport et al., 2008). And coyotes (Canis latrans) housed in small kennels rather than large enclosures do not just pace more: they also show greater heart rate and hiding responses to human proximity (Brummer et al., 2010).
Turning to studies relevant to Test 3, for social animals like parrots and primates, living alone does not just promote ARB: it can shorten telomeres (Aydinonat et al., 2014), reduce hippocampal neurogenesis (e.g., Cinini et al., 2014), and impair health and survival (e.g., Lewis et al., 2000; Capitanio et al., 2019). Furthermore, disrupting social bonds can increase both rhesus monkeys’ vulnerability to viral disease (e.g., Capitanio et al., 2019) and elephants’ mortality rates (Clubb et al., 2008). And tethering sows alone rather than group-housing them does not just elevate ARB: it seems to increase adrenal weight (van der Staay et al., 2010). Turning to parental deprivation, chicks reared without hens are not just (potentially) more prone to feather-pecking, but are also more fearful in behavioural tests (Rodenburg et al., 2009); rhesus monkeys raised without mothers grow up prone to both ARBs and shortened telomeres (Schneper et al., 2016), higher rates of physical illness (Conti et al., 2012), and curtailed lifespans (Lewis et al., 2000), as well as significant weight loss if faced with ARB-inducing CUS (Zhang et al., 2016); and similarly, nursery-reared chimpanzees with poor bonds to their human carers grow up more prone to both stereotypic rocking and respiratory infections (Clay et al., 2015).
Furthermore, studies manipulating degrees of welfare improvement may reveal graded relationships between ARB and other welfare indicator: for brown capuchins (Cebus apella) exposed to a series of enrichments, the more effectively these reduced abnormal behaviour (including ARB), the more effectively they reduced alarm responses to humans (Boinski et al., 1999); while in piglets, being weaned at progressively younger ages predicted both progressively increasing prevalences of belly-nosing and also progressively decreasing growth rates (Faccin et al., 2020).
2.5.2. When Populations with Elevated ARB Show Signs of More Negative Affect
Turning to comparisons involving combinations of treatment and genetic effects, across flocks of chickens, feather pecking severity (inferred from plumage scores) and fear of humans covaried (De Haas et al., 2014). Across stabled horses varying in background and age, more ARB-prone subjects slept less (Hausberger et al., 2007), showed more depression-like ‘withdrawn’ behaviour (Hausberger et al., 2007), were more prone to gastric ulceration (Patiño et al., 2020), and were less fertile (Benhajali et al., 2014). Keeper surveys and endocrinological analyses of 13 female white rhinoceroses (Ceratotherium simum) across six zoos revealed that animals without oestrous cycles were rated as higher in pacing (and other ‘stereotypies’: Carlstead and Brown, 2005). Further keeper surveys found that across 52 Felidae individuals spanning thirteen species and five zoos, ratings for being ‘stereotypical’ (‘fixed and oversimplified in behaviours’) were mirrored by high ratings for being fearful, tense and anxious (O’Connor and Vonk, 2022). Similarly, owner surveys indicate that across pet cats, fearfulness and over-grooming are related (Mikkola et al., 2022), as are ‘shyness’ and tail-chasing in pet dogs (Tiira et al., 2012).
Some species and genotype differences show similar patterns. Thus, in zoos, wide-ranging, semi-nomadic Carnivora species are not just more prone to stereotypic route-tracing than naturally more ‘stay-at-home’ species: they also have reduced fecundity and higher infant mortality (Bandeli et al., 2023; Miller et al., 2019). Naturally browsing Ungulate species are not just more prone to oral ARB than naturally grazing species: they also have shorter lifespans (Mellor and Mason, 2023). Furthermore, compared to hens genetically selected for low feather pecking, high-feather pecking lines are more reactive to aversive stimuli, with higher corticosterone levels and heart rates, and lower heart rate variability, after handling and restraint (Kjaer and Guémené, 2008; Kjaer and Jørgensen, 2011).
2.5.3. When Individuals with Elevated ARB Show Signs of More Negative Affect
Turning to correlates of ARB across genetically similar animals who are treated alike, here, individual differences in ARB also sometimes covary with other signs of poor welfare. Across individual laying hens, feather pecking correlates with longer latencies to move after tonic immobility (thought to indicate greater fearfulness: Grams et al., 2014; Iffland et al., 2020; see also Rodenburg et al., 2004; De Haas et al., 2014). Across orange-winged Amazons, birds with ‘neurotic’ personalities (anxious and aggressive) perform more feather-plucking (Cussen and Mench, 2015). In chinchillas, fur-chewing females have higher mortality in their offspring (Galeano et al., 2014). And in mice, greater corticosterone responses to challenge have been linked with higher levels of ARB (Latham and Mason, 2010; Engel et al., 2011). However, these particular types of studies also reveal many counter-examples, as we will cover next.
2.5.4. Counter-Examples: When ARBs Seem to Fail Test 5
When Treatments, Groups or Individuals Showing More ARB Do Not Display Other Signs of More Negative Affect
Comparisons of spontaneously high and low ARB individuals with similar genetic and experiential backgrounds rather often show that ARBs fail Test 5. In a review of 90 studies, Mason and Latham (2004) compared welfare indicators in high and low ARB individuals (or within-individual changes in welfare during ARB performance): only 20 linked high ARB to more negative affect, while 16 found no apparent relationship (and 54 suggested that high ARB individuals have less negative affect, as discussed in the next section). Findings of similar patterns since then include that tongue-playing calves and tongue-rolling heifers show no cortisol differences in response to stressful challenge (being moved from their home pens and restrained in metabolic cages (Webb et al., 2017); crib-biting horses and controls do not differ in heart-rate variability (Nagy et al., 2009); and across individual mink in barren housing, levels of locomotor ARB do not correlate with decreased spleen and thymus weights, greater fear of humans, more fluctuating asymmetry, or lower male testosterone levels (Díez-León et al., 2013, 2016).
At the treatment level, similar null effects also sometimes occur. Chronic Unpredictable Stress (CUS) causes several signs of poor welfare in mice (e.g., increased adrenal weight: Monteiro et al., 2015), yet in one study, it did not increase their ARB (Olsson and Sherwin, 2006). And in research on coyotes (Brummer et al., 2010), animals seemed to similarly engage in ARBs in small pens (8-9% observations) and versus large ones (6-7% observations), even though in the latter, animals never hid, and their heart rates recovered faster after human disturbance. Genotypes differing in ARB may also show no apparent differences in welfare. For example, although C57Bl/6 and DBA strains showed clear behavioural and physiological responses to restraint stress and food deprivation (reviewed: Cabib, 2006), only the latter respond with ARB (e.g., Cabib et al., 2021); and likewise C58 mice show far more ARB than C57Bl/6 mice (both during tests and in their home cages), yet seem no more fearful in tests for anxiety (Moy et al., 2014).
When High ARB Treatments, Groups or Individuals Show Signs of Less Negative Affect
As mentioned, across studies of welfare correlates of individual variation in ARB, 54 of 90 [60%] reported signs that relatively higher negative affect corresponds to relatively lower ARB: a pattern significantly different from chance (and from that seen when comparing different treatments). Some early examples of this seemingly common type of pattern come from pigs, e.g., across genetically similar, same age, identically stalled sows, individuals that do more oral ARBs like chain-manipulation, chomping and object licking have lower average heart rates (Schouten and Wiepkema, 1991), horses (e.g., lower heart rate during periods of cribbing: Lebelt et al., 1998) and calves (e.g., non-nutritive sucking helps lower heart rate (Seo et al., 1998). More recent research on rodents often indicates similar paradoxical effects (e.g., low ARB laboratory mice perform more floating in Porsolt tests for depressive-like helplessness: Fureix et al., 2016). In bank voles, individuals performing more ARBs had higher reproductive success and survival (Schønecker, 2009); in African striped mice, high-ARB females weaned more offspring (Jones et al., 2010b) and were also less anxious in behavioural tests (Joshi and Pillay, 2016); in deer mice, repetitive back-flipping increased alongside reproductive success over multiple generations (Lacy et al., 2013), and selecting against back-flipping had very negative effects on reproduction with low ARB mice seeming inactive and overweight (R. Lacy, pers. comm.). Farmed mink provide further evidence. Individuals selected from high-ARB lines show greater fertility, larger litters, and lower pup mortality (Jeppesen et al., 2004; Meagher et al., 2012); and individual mink who pace more than identically-treated peers have higher levels of hippocampal neurogenesis (Malmkvist et al., 2012). Furthermore, some find that fearful mink perform less ARB than do bold ones (Hansen and Jeppesen, 2005; Svendsen et al., 2007; though cf. Malmkvist et al. [2012] and Meagher et al. [2012] who find no such relationships). Some similar patterns also occur in other taxa: feather-pecking chickens are described as bolder than their low-ARB conspecifics (Jensen et al., 2005), showing lower corticosterone response to restraint stress than low-feather-pecking lines (Korte et al., 1997); crib-biting horses have been rated as less anxious in one survey (Nagy et al., 2010); and chimpanzees engaging in coprophagy (which already fails Test 3) are more likely to copulate and are described as being more affiliative and pro-social (Fritz et al., 1992; Hopper et al., 2016). Finally some studies find that ‘shy’, non-exploratory rhesus monkeys are less rather than rather than more likely to develop ARBs (Gottleib et al. 2013); that bouts of self-biting in rhesus monkeys are followed by reductions in heart rate (e.g., Novak, 2003); in another rhesus monkey study, displaying high levels of ‘self-directed’ ARB predicted significantly smaller corticosteroids responses to a frightening stimulus (Pomerantz et al., 2012); and finally, when rhesus monkeys who did or did not display self-biting were exposed to the close proximity of an unfamiliar human, all reacted as though this was aversive, but the self-biters reacted less than the ‘controls’ (Peterson et al., 2017). Collectively, these examples suggest that within populations of similar subjects all faced with equally challenging conditions, the higher-ARB individuals can sometimes have relatively better welfare than those individuals displaying negligible ARB there.
Less common are instances where treatments that clearly worsen welfare are paradoxically associated with less ARB. In the study mentioned above, where rhesus monkeys were exposed to a nearby unfamiliar human, the treatment triggered immediate threat responses as well as behaviours like retreating and freezing, and ARB performance declined (Peterson et al., 2017). In another study, construction noise increased stress markers and hiding behaviour in zoo Carnivora, but dramatically reduced pacing (Chosy et al., 2014). Similarly, CUS in rats induced signs of poor welfare—including anhedonia (Willner, 2017), chromodacryorrhea (Young et al., 2024), and pessimistic judgment biases (Harding et al., 2004)—yet ARB again actually declined (Young et al., 2024). In some species, being captured from the wild produces a comparable outcome: wild-caught voles and striped mice display little to no locomotor ARB (Cooper and Nicol, 1996; Jones et al., 2011a), despite generally being more fearful than their captive-bred counterparts.
3. Discussion
So, are ARBs clear signs of poor welfare, as early commentators believed? Or is their aetiology so complex that, as others have since argued, they reveal nothing reliable about animals’ affective states? To resolve this, we applied five tests for construct validity; distinguished two ways in which ARBs fail such tests, either not increasing in affectively negative contexts (poor responsivity) or being increased by affectively neutral or positive treatments (poor selectivity); and wherever possible, clarified timescales (for instance differentiating the slow emergence of high ARB phenotypes from the immediate triggers of ARB bouts). Along the way, several unknowns and research needs emerged (including the biological mechanisms linking prolonged negative affect with ARB, and how ARBs should be identified objectively). These we will discuss here. But first, we address the central question: how ARBs reflect affective states. We then consider the long-recognised complexities raised in the Introduction, especially ARBs’ proneness to false nulls as welfare indicators. To end, we propose some practical recommendations: six ‘rules of thumb’ for the valid, accurate use of ARBs during welfare assessments. We also suggest new research avenues, and advocate for a central role of poor welfare when deeming behaviours such as these ‘abnormal’.
3.1. ARBs Reflect Prolonged Negative Affect (But Not Transient Negative Emotions)
A substantial body of research indicates that collectively, the ARBs reviewed indicate poor welfare in both human and non-human animals. Furthermore, even though the breadth of our analysis necessarily limited depth (all five tests not being covered for all forms of ARB), some remarkably diverse types of ARB in animals passed all of Tests 2-5 (see Table 3). Thus, the Carnivora and Rodent behaviours typically labelled as ‘stereotypic’, and the body-focused, harmful ARBs of birds and primates, not typically so labelled, have remarkably similar links with poor welfare. Furthermore, to be more specific, ARBs in humans reflect negative moods or even mood disorders; and correspondingly, in both humans and animals, they reflect not only prolonged negative experiences but also often early negative experiences (e.g., during infancy), recurrent previous negative experiences, and potentially cumulative affective experience even over a whole lifespan, doing so in graded, dose-response-like ways. We review the evidence for this next; and also show how such conclusions do not apply for natural behaviours incorrectly labelled ‘abnormal’, nor for the assessment of transient negative emotions (since whether or not a subject is currently engaged in an ARB is not, in contrast, a reliable sign of current negative emotion).
In humans, being prone to ARB is thus often accompanied by anxiety or other low moods (sometimes severe enough to be deemed clinical disorders), including those arising from childhood maltreatment, the threat of war, being imprisoned, or such challenging conditions as OCD or trichotillomania. Likewise, animal ARBs are promoted by early, prolonged or repeated challenges (lasting or recurring over several days to months or years) that are known to be aversive, to threaten ancestral fitness in the wild, and/or to cause other signs of chronic stress. These include the premature loss of parental care, repeated exposure to physical harm, food deprivation, living in barren conditions or unnaturally alone, exposure to ‘CUS’ protocols, and sometimes also health conditions likely to cause discomfort or pain. Furthermore, consistent with a central role for negative mood, mood-improving drugs (anti-depressants and anxiolytics) often reduce ARBs in both humans and animals.
Our review further shows that relationships between ARBs and long-lasting negative moods, mood disorders or even cumulative negative experience are often graded. Many cases illustrated positive relationships between the extent of ARB and the intensity of subjects’ negative states, including self-reported anxiety or depression in neurotypical and clinical humans, and as inferred in animals from (as examples) their degrees of maternal loss in infancy, sustained food deprivation or social restriction. Other cases showed positive relationships between the extent of ARB and the duration of subjects’ negative states, such as how long they have lived in impoverished conditions (for children, parrots and chickens), or alone (again for parrots, and also rhesus monkeys, chimpanzees and pigs). Furthermore, sometimes such challenges act in concert, as for example in the additive effects of Streptococcus infection and stress on humans or the synergistic effects of early maternal loss and single housing on macaques. And in extensions of such patterns, yet other cases highlighted positive relationships between the extent of ARB and the number of challenges endured. Total numbers of severe life events or traumas in a lifetime can thus predict symptom severity in people at risk of tooth-grinding, OCD, trichotillomania, skin-picking and ticcing in Tourette’s, while in animals (for whom such lifetime tallies have barely been studied), similar results emerged in repeatedly isolated deer mice, repeatedly relocated elephants, and primates repeatedly subject to challenges like invasive research procedures.
In one clear instance, however, repetitive behaviour did not reflect prolonged negative affect, because it was probably not actually ‘abnormal’. This was coprophagia by chimpanzees. Data only allowed Tests 2 and 3 to be tested for this behaviour, but it failed both. Coprophagia failed Test 3 in being more common in animals raised in large groups with their mothers than in small groups without; and it failed Test 5 in that across multiple sites, it covaried with signs of relatively good welfare (greater sociability and libido). Indeed, authors studying coprophagia (e.g., Hopper et al., 2016) note that it occurs in the wild, arguing "It is a natural behaviour for chimpanzees, but one that we find distasteful, and our reaction to it may cloud our interpretation of its meaning" (p. 116). This highlights how repetitive behaviours may be judged to be normal or abnormal via rather subjective criteria (such as perceived unpleasantness): a classification challenge we return to at the end.
Furthermore, even in the many cases where ARBs seem both unnatural and linked with prolonged negative affect, the evidence that performing a bout of ARB indicates current negative emotion is highly equivocal. For one, short-term negative emotions are often not triggers. Even though negative emotions must immediately be felt when humans are first imprisoned, when mice and chickens are first food restricted, and when other species are first placed in barren, socially restrictive housing, our review did not find that ARBs emerge immediately; instead often needing days or weeks of exposure before developing. In addition, once developed, negative stimuli or events do not always trigger performance. ARBs can be notably absent when animals are alarmed by threats, as shown for monkeys and mink, while in people, the reviewed literature reports that bouts of ARB do not necessarily correspond to times of peak distress. Indeed, human ARB performance often occurs during moments of happy excitement or quiet calm; while in animals, bouts of ARB are often triggered by the arrival of meals: surely rewarding. Furthermore, ARB performance can have immediate positive consequences. One of the oldest human studies (Deaton et al., 1977) expresses certainty that ARBs are one of several "time-killing activities or adaptational strategies" that can help prisoners-of-war while captive, and a recurrent theme in the human literature was that bouts of ARB-performance are reportedly tension-relieving, stress-reducing, or calming in other ways. Physiological data even reveal that bouts can be followed by, e.g., heart rate reductions. In animals, data suggesting similar come from a few within-subject studies of heart rate reduction in horses, monkeys and calves during and after the performance of ARB bouts; there is also evidence that animals may find ARB-performance motivating or rewarding (see e.g., Houpt, 2012 on horses working to crib-bite). Whether such ‘coping’ effects are so beneficial that the welfare of ARB-prone subjects should then be considered neutral, or even good, is an important issue we cover later. But regardless, it seems clear that ARB performance should not be used to infer current negative emotion: doing so would result in false nulls in the immediate aftermath of negative events, and ‘false alarms’ when bouts are triggered by neutral or positively exciting events or have apparently positive immediate consequences.
Our overall main conclusion is therefore that when it is reasonable to deem repetitive behaviours ‘abnormal’, their presence and extent reflects the presence and severity of early, recurrent or prolonged challenges, and in humans (consistent with this), their presence and severity reflects the presence and severity of negative moods, sometimes even mood disorders. Furthermore, such relationships can reflect the degree of exposure to negative events over long periods, suggesting that at least some ARBs indicate cumulative negative affect (‘cumulative stress’).
Now, which of these affective constructs is precisely most relevant is hard to say: appropriate research is lacking; the lines between moods, mood disorders and cumulative affective impacts are anyway blurred; and since ARBs are heterogeneous, the answer would likely vary case-to-case. Nevertheless, it seems clear that challenges that induce abnormally repetitive phenotypes also typically induce prolonged negative states; and that in at least some instances, multiple causes of poor welfare can combine to promote ARB. This view supports recent suggestions that feather-pecking and other ARBs in farmed poultry, and tail-biting in farmed pigs, integrate the effects of multiple welfare challenges or ‘cumulative stressors’ (Cronin and Glatz, 2021; EFSA AHAW PANEL, 2023; Šemrov and Patt, 2024). It also suggests new ways to investigate the aetiology of animals’ ARB, by cataloguing all negative experiences from infancy onwards to assess their collective impacts (perhaps over entire lifespans). An additional conclusion is a corollary of this: that the onset and offset of ARB bouts should not, in contrast, be inferred to track immediate changes in emotional state: doing so during welfare assessment would risk both false null and ‘false alarm’ type errors.
3.2. ARBs Are Not Perfect Welfare Indicators, but Their Imperfections are Understandable
Despite the compelling picture above, our review still collated many apparent failures of Tests 1-5, even for clearly abnormal behaviours being used to infer long-term negative states. Indeed, even for the cases in Table 3, there are counter-examples (as we will see in Table 4). Some could reflect the Type 1 and Type 2 errors inevitable in research; others, that the assumptions Tests 1-5 rely on were not met (as we judged especially likely for Test 4). However, such explanations cannot plausibly account for the sheer number of counter-examples, nor the patterns evident within them. Thus, just as outlined in the Introduction, we found evidence linking ARBs with enhanced abilities to cope; cases where ARBs are not elicited or increased by prolonged poor conditions; and instances where ARBs fail to decline when welfare has apparently improved. We also found a fourth problem: occasional poor selectivity, shown in cases where ARBs were enhanced by factors other than prolonged negative affect. We review these four issues next, try to explain them, and discuss their implications for ARB as a welfare assessment tool.
3.2.1. Evidence that ARBs Are Associated with Enhanced Coping, and Implications for Their Use in Welfare Assessment
The hypothesis that ARBs help performers cope is longstanding in both human and animal literatures, as we met in Section 3.1. While our aim was not specifically to test it, many collated studies showed relevant findings: evidence that ARBs can correlate with enhanced abilities to cope with challenge both immediately (as already reviewed in Section 3.1) and for animals, in the longer term too. The evidence for this in animals largely comes from between-subject correlational data. As we reviewed for a host of species (in Test 5), data on other welfare indicators often show that across similarly poorly treated individuals, the most ARB-prone ones fare better than peers who display little or no ARB (e.g., showing fewer other signs of chronic stress). Thus, in chickens, pigs, cattle, horses, mice, rhesus monkeys and various rodents (African striped mice, deer mice, bank voles, albeit with laboratory house mouse findings less consistent), comparisons of other chronic-stress-sensitive measures between the most and least ARB-prone individuals within treatment groups, reveal that that the most ARB-prone individuals appear least impacted by their sub-optimal conditions. This could thus indicate that ARBs help those animals who develop them to cope better with challenges than do other animals.
However, as also reviewed in Test 5, such patterns are not ubiquitous. Furthermore, correlational studies like these cannot actually prove cause and effect. Indeed, an alternative explanation for such patterns is that across similarly poorly treated individuals, those reacting with the most negative affect develop the least ARB, while those reacting with the least negative affect develop the most. Two inter-twined concepts make this a reasonable suggestion. The first is that being inactive (for example, hiding) is known to be an alternate, non-ARB response to sustained severe challenge: an important theme that will recur in the sections below. The second is that whether to show active or passive responses to challenge is influenced, at least in part, by individual personality. Thus, for captive humans, Deaton et al. (1977) suggested that developing ARBs typifies active, assertive individuals, with apathetic passivity being more likely in other personality types. And decades later, Ijichi et al. (2013) proposed that for animals, ‘proactive’ personalities (responding to challenges with aggression, attempts to remove the stressor, or escape) predispose individuals to ARB; while ‘reactive’ personalities (responding to challenges with passivity) predispose individuals to inactivity instead. Furthermore, there is some empirical evidence for this in mammals, including that ‘gentle’, timid infant rhesus monkeys go on to display less ARB in adulthood (see Gottlieb et al., 2013 in Test 5; though cf. Vandeleest et al., 2011); and that within C57Bl/6 mice, individuals who spontaneously perform the least ARB, also perform the most floating in Porsolt tests for ‘helplessness’, as well as the most ‘inactive but awake’ inert sitting behaviour in their home cages (see Fureix et al., [2016] in Test 5; see also limited support for mice with respect to drug-induced ARBs from Koolhaas et al., 1999; and for African striped mice from Joshi and Pillay, 2016). Now, this particular proactive/reactive framework is not always supported (e.g., for high/low feather-pecking chickens: van der Eijk et al., 2019), but nevertheless, the broader issue holds: being endogenously prone to, or protected from, ARB does not map neatly onto respectively greater or lesser susceptibilities to negative affect (another theme developed further below). This thus provides an alternative explanation for patterns that superficially seem to support the coping hypothesis.
Overall, the possibility that ARBs help animals to cope thus remains plausible but uncertain. Yet as we saw in the Introduction, some use it to downgrade the importance of ARBs as welfare indicators. So does the possibility of coping effects render ARBs welfare neutral? We argue no, on three grounds. One is simply the lack of truly convincing evidence across multiple species. But additionally, our results from applying Test 2 and Test 5 reject this. For a start, if ARBs help animals cope with welfare challenges so effectively that performers no longer experience negative affect, then ARB-promoting housing should not be avoided over more naturalistic ‘enriched’ housing: animals should be indifferent between them (ARB performance compensating for what is lacking in the barren environments). One intriguing finding of this type did emerge in bank voles (Cooper and Nicol 1991): long barren-housed voles became indifferent to more naturalistic enclosures as their ARBs developed. However, this has never been replicated; and importantly, this is not what most data show (cf. Table 2): ARB-promoting environments are typically avoided if animals can choose better-resourced ones. Turning to Test 5, if ARB helps animals cope with welfare challenges so effectively that they no longer experience negative affect, then comparisons of treatments promoting high versus low ARB should find no other differences in terms of welfare. (For example, if ARB fully compensates for sub-optimal housing, then mice and rats in ARB-potentiating barren cages should live as long as those in larger, well-resourced cages). However, again, as we reviewed, this is not what most data show. As Tatemoto et al. (2022) nicely sum up, here for pigs, "The welfare of confined sows that show apathy and unresponsiveness may well be worse than those that show stereotypies, but neither has good welfare" (p. 5).
Now, this counter-evidence against the coping hypothesis could reflect the research being insufficiently granular. The studies summarised typically did not differentiate between high and low ARB subjects, and this is important. If ARBs enable subjects to so fully cope with sub-optimal treatment that this is no longer perceived as sub-optimal, then strictly this hypothesis predicts that ARB-promoting housing should not be avoided for more naturalistic ‘enriched’ housing by ARB-performers only; and comparisons of treatments promoting high versus low ARB should find no other differences in terms of welfare, if only ARB-performers are included in the former. Currently, studies do not reveal whether these more nuanced predictions are supported or rejected. Nevertheless, empathizing with the human subjects reviewed in Test 1 may provide further insights. Despite their ARB, people with OCD, autism spectrum disorder, and related clinical conditions still report being more anxious/depressed than controls. Furthermore, no one would ever argue that performing ARBs is so stress-relieving that it makes being imprisoned welfare-neutral. Current data support this same reasonable intuition being applied to animals. Thus, our third main conclusion is that while some ARBs may help animals to partially cope, they are not so effective as to neutralize their welfare problems.
3.2.2. Problems with Responsiveness: How and Why ARBs Can Fail to Track Long-Term Negative States, and the Implications for Their Use in Welfare Assessment
It has long been recognised that a major weakness of ARBs as welfare indicators is their insensitivity, and our review found numerous instances of ARBs failing to be promoted by long-term negative states. In humans, for instance, most people do not develop ARBs after trauma or repeated negative experiences. (Nor are ARBs so closely linked to mood disorders that they are diagnostic of such conditions). Additionally, ARBs are not ubiquitous in even the poorest human environments studied (solitary confinement in prisoner-of-war camps; Romanian ‘orphanages’): some individuals, despite undoubted poor well-being, did not display ARBs here. Turning to animals, ARBs are similarly low or even absent in several very low-welfare scenarios. Furthermore, just as in humans, even when welfare-compromising treatments do induce ARBs at the group level, some subjects so exposed will not develop them (and as we saw above, not because these low-ARB subjects have the best welfare). In addition, sometimes ARBs fail to increase when subjects are exposed to additional challenges. ARBs are thus often unresponsive to declines in welfare.
However, moving forward, understanding why could reduce the risks that such ‘false null’ results mislead us during welfare assessments. Based on the reviewed literature, we propose five explanations, two of which follow on from the Discussion so far: ARBs can be unresponsive to long-term negative states because i) neurological traits may not promote ARB; and ii) some contexts or types of negative state promote inactivity instead. In addition, sometimes the influence of negative affect on ARB is inhibited or masked by iii) threshold and priming effects, or iv) ceiling effects. Finally, v) some ARBs show motivational selectivity: they are increased or decreased by specific challenges only. These five explanations for poor responsiveness are reviewed next.
3.2.3. i) Neurological traits may not promote ARB
Just as we met for individual differences, certain endogenous traits protect some subjects from developing ARB even in sub-optimal environments. Above, we discussed the role of reactive personalities, but more broadly, this phenomenon is manifest in other familial predispositions (e.g., ones involving corticostriatal systems), along with similar population-level differences between strains or species.
For example, cognitive flexibility reduces ARB-proneness in neurotypical humans, with less routine-prone children being less likely to develop ARBs (Zohar et al., 1995; Happaney and Zelazo, 2004; see also Suñol et al., 2018): effects apparently with genetic/familial bases (e.g., ‘neurocognitive endophenotypes’: Fineberg et al., 2014; Robbins et al., 2012; Tiego et al., 2020; Yang et al., 2021). Likewise, people exposed to severe stressors but not genetically at risk of OCD, skin-picking or trichotillomania may still experience trauma (and even have other clinical consequences), yet still not develop ARBs because of their neurological predispositions (e.g., Hu et al., 2025; see also Goldberg et al., 2015). Neurological differences with genetic bases also play roles in animals. Thus, when C57Bl/6 mice are faced with recurrent restraint stress or prolonged food deprivation, and become inactive instead of developing ARB, their prefrontal cortex inhibits dopamine activity in the basal ganglia; in contrast, in the DBA mice more prone to ARB have weaker inhibition or even enhanced dopamine activity (Cabib, 2006; Cabib et al., 2021, 2020, 1998). Likewise, C58 mice compared to C57Bl/6 are neurologically predisposed to be less flexible (e.g., showing impaired reversal learning: Whitehouse et al., 2017), and in their brains, there is less activity in key inhibitory regions of the basal ganglia (e.g., Lewis et al., 2018; see also Wilkes et al., 2019). As one last possible example of species-level predispositions to not develop ARB despite poor welfare, cynomolgus monkeys show a relative lack of ARB compared to other macaques (e.g., rhesus monkeys): as we reviewed, they tend to become quiet and inactive after premature separation from their mothers in infancy, and if isolated or socially subordinate as adults. Long-recognised as "passive or ‘reserved’" (Clarke and Lindburg, 1993), monkeys of this species develop distinctive, slumped, depressed-seeming postures instead of ARB (e.g., Shively et al., 2005).
Some lasting developmental effects can have similar effects too. African striped mice born and raised in the wild before being placed in lab cages show negligible ARB there. They are also less behaviourally inflexible in maze tests (Jones et al., 2011b) than lab-raised conspecifics, indicating less behavioural disinhibition and presumably—given the barren conditions and premature parental loss typical of the lab, and the many studies showing larger brains and better learning ability of wild-caught compared to captive-raised conspecifics (e.g., Burns et al., 2009; Guay and Iwaniuk, 2008)—more normal brain development (Jones et al., 2011b). They are also less active because of anxiety: they hide more, bringing us to our next point.
3.2.2. ii) Some Prolonged Negative States Promote Inactivity Instead of ARB
That certain long-term negative states promote inactivity instead of ARB has been seen in several populations, even ones otherwise prone to ARB. In Maier and Seligman’s shocked dogs (1976), for instance, subjects gave up trying to escape after several trials, only to become passive and silent instead. When repeated restraint caused mink ARBs to decline, their hiding in the nestbox markedly increased (see Bildsøe et al., [1991] in Test 2). Likewise, when months of construction noise caused the pacing of zoo Carnivora to decline, their hiding also increased (see Chosy et al., 2014 in Test 5). Similar effects can arise when animals are in chronic pain or sick (at least anecdotally), likely due to ‘sickness behaviour’ typified by lethargy and extended rest or sleep (e.g., Dantzer, 2009). Furthermore, in Brummer et al. (2010)’s study of coyotes, individual rates of ARBs seemed similar between small and large enclosures when expressed as proportions of observations. However, overall activity declined in the former. Accounting for this reveals a housing effect: ARB as a proportion of active time was nearly doubled in smaller enclosures compared to larger (17-19% as opposed to 8-10%).
Thus, when circumstances promote negative states typified by inactivity, the result can be reduced or even absent ARB. Inactivity is then a potential confound, masking treatment effects on ARB that might be evident if this were statistically controlled for.
3.2.3. iii) The Influence of Prolonged Negative Affect on ARB Is Inhibited by Threshold/Priming Effects
Some failures of responsiveness suggest that negative experiences must accrue above certain levels before they promote ARBs. We have already described cases where it takes days or weeks of, say, solitary confinement or food restriction before ARBs emerge, presenting such delays as if reflecting the time it takes for negative emotions to integrate into negative moods. But such delays could also reflect the time required for relevant neurological changes to occur; and/or that negative affect increases in severity over the duration, eventually crossing some threshold above which ARBs emerge. Indeed, in further support of this last idea, three studies we reviewed reported distinctive interactive effects, such that a sub-optimal treatment did not promote ARB until combined with a second. In the first, on mink (see Hansen and Jeppesen, 2000 in Test 2), small cages and a lack of swimming water only potentiated ARB in combination: each on its own had negligible effects. In the other, on quail (see Laurence et al., 2014 in Test 2), a week of barren caging had little effect on ARBs unless birds were also subject to CUS, and likewise a week of CUS had little effect on ARB unless birds were also barren-caged. Finally, in rhesus monkeys, CUS had little effect on ARB unless animals were also maternally deprived (Zhang et al. 2016 in Test 2). Together, this evidence of threshold effects also suggests that affectively negative experiences may sometimes merely prime subjects to ARBs, increasing their vulnerability, yet in ways that remain latent until they are challenged further.
3.2.4. iv) The Influence of Prolonged Negative Affect on ARB Is Masked by Ceiling Effects
Other studies suggest ceiling effects on ARB performance. Reviewed examples include how housing mice and parrots in preferred versus non-preferred conditions causes animals to diverge in ARB over time, until that of the sub-optimally housed subjects reaches a plateau; and how repeatedly isolating well-housed deer mice in a test chamber causes their ARB to increase but only until reaching the same level shown by barren-housed conspecifics. Adding to this, the high ARB of quails exposed to either barren housing or CUS for 2 weeks does not seem to be elevated if the two challenges are combined (Laurence et al., 2014). In all these cases, ARBs thus level out at an apparent maximum, not increasing further with increasingly prolonged or severe negative affect: false nulls (assuming they do not indicate ceiling effects on underlying negative affect). Speculatively, such effects could reflect constraints on how much time or energy animals can devote to ARB; or perhaps even competition from increasing levels of inactivity (if poor health, fearfulness, or depression-like states have started to emerge).
3.2.5. v) Motivational Selectivity: Certain ARBs Reflect Specific Negative Experiences Only
Finally, in cases highlighting the aetiological heterogeneity of ARBs, some ARBs reflect only very specific challenges, while being insensitive to others. We suspect this particularly occurs when specific frustrated motivations play a major role in the aetiology of an ARB, and the resultant response is a relevant type of intention, vacuum or redirected movement. Thus, as reviewed in Test 3 (citing Boyle et al., 2022), for piglets, the early loss of the mother increases belly-nosing: a direct response to maternal loss seemingly derived from normal udder massage. But it seems to play little if any role in other, later developing ARBs like tail-biting. Likewise, as reviewed in Test 2 (citing Polanco et al., 2018; Malmkvist et al., 2024), scrabbling at cage partitions by male mink—a conspecific-directed ARB that, in this naturally solitary species, seems agonistic—is exacerbated by the proximity of male neighbours, yet this has little effect on other ARBs. And reciprocally, providing barren housing, versus large complex cages or running wheels, is less impactful on scrabbling than on ‘whole body’ ARBs. Such examples build on classic prior work by Keiper (1969); and arguably illustrate how welfare-relevant treatments may impact motivationally related ARBs only (such that every ARB does not respond to all possible causes of negative mood).
3.2.6. vi) Understanding the Poor Responsiveness of ARBs: A Summary
To wrap up this sub-section, ARBs are clearly not always elicited or increased by prolonged poor conditions. This makes them a somewhat blunt instrument in welfare assessment, even for assessing prolonged negative states; and it means that their absence does not necessarily demonstrate that all is well. However, understanding why ARBs are so unresponsive can help refine their use in welfare assessment. Low or absent ARBs are likely to be false nulls (i.e., not reflecting good welfare) in subjects who have negligible ARB for endogenous reasons (such as personality or strain); when levels of inactivity are high (especially if involving apparent hiding or unusual depression-like postures); and when subjects have only experienced brief or single challenges. Furthermore, once ARBs have developed, failures of further challenges to increase their levels can reflect the constraints of ceiling effects, or that they have frustration-driven aetiologies involving very specific motivational systems.
3.2.7. Evidence That ARBs May Persist Even After Poor Conditions Are Improved, and Implications for Their Use in Welfare Assessment
A puzzle, perhaps related to these last examples, is that apparent improvements in welfare can fail to reduce ARBs, once they have developed and been performed for a long period. Thus, as we saw in Test 1, ARBs may persist in children adopted from ‘orphanages’, even when now in normal family homes; and this is especially so for children who had spent many prior years in deprived conditions. As we saw in Test 2, ARBs can become hard to alleviate with well-resourced housing in rodents who have been barren-housed into middle age or beyond. And as we saw in Test 4, mood-enhancing drugs may also become less effective on ARBs that have been performed for longer periods of time. Together, such persistent ARBs could perhaps be mere ‘scars of the past’ that fail to reflect animals’ current, presumably improved welfare. And a potential explanation could be the obverse of what we saw for wild-caught rodents (where behaviourally flexible, presumed neurologically normal animals were protected from ARB): that these subjects’ corticostriatal systems have been so profoundly altered by their prolonged poor housing that normalization of function is hard, even if welfare is improved. After all, institution-reared children showing persistent ARBs post-adoption also had poor cognitive abilities and other signs of greater neurological impairment (e.g., Beckett et al., 2002; Bos and Van Balen, 2010; Rutter et al., 2007). In both such scenarios, persistent ARBs are not responding to genuine improvements in underlying welfare: a problem for their valid use in welfare assessment.
However, there are alternative explanations for such persistence. One is that the treatments we are assuming improve welfare are not actually doing so. For example in animals, ‘enrichments’ that we assume are welfare-enhancing (and indeed are preferred by animals raised lifelong with them), may be less motivating for subjects who have long done without (Chow and Hogan, 2005; Tilly et al., 2010); while for adopted ‘orphans’, perhaps the normal family homes are not quite as restorative as we hope. Likewise, drugs we hope are mood-improving, perhaps also are failing to do so for subjects with longstanding problems. If this explanation is correct, then the treatment-resistant ARBs are validly useful in welfare assessment, because they indicate treatment-resistant negative affect. The second explanation is more to do with the timescale being considered: it is that ARBs more validly reflect cumulative affective experience over a lifetime than they do just current mood. If this conception is correct, then persistent ARBs of aged or long-deprived subjects again are validly useful in welfare assessment, as long as that assessment is aiming to assess cumulative lifelong well-being.
This fascinating and potentially worrying topic thus needs much more research, but in the interim, we propose that persistent ARBs are not dismissed as mere habits but are treated as something more concerning.
3.2.8. Selectivity Problems: When ARBs Are Enhanced by Factors Other Than Prolonged Negative Affective States, and Implications for Their Use in Welfare Assessment
If using ARBs appropriately to assess long-term negative affect, then they appear to have good specificity as welfare indicators. Our review suggests just two instances where ARBs are enhanced by factors other than prolonged negative states, both related to phenomena we have met already: activity as a confound, and neurological predispositions affecting behavioural selection.
In the first instance, ARBs are elevated in affectively relative positive contexts because overall activity increases. For example, mink given simple ‘enrichments’ (balls and chew-chains in their cages) showed a surprising increase in ARB, but this simply reflected an overall increase in activity levels (see Meagher et al., 2014 in Test 2). Overall, changes in overall activity levels can thus cause ‘false alarms’ when using ARBs as welfare indicators (not just the false nulls we have met already).
In the second instance, ARBs are elevated because the behavioural selection systems of subjects’ brains predispose them to ARBs, quite independent of their affective states. Some examples are simply the obverse of those already discussed in the false nulls section (3.2.2): for example, if it would be wrong to infer relatively good welfare in reactive individual personalities, C57Bl/6 strain mice, and cynomolgus monkeys who seem resistant to developing ARBs, then the corollary is that it would also be wrong to infer poorer welfare in their high ARB comparators (proactive individual personalities, DBA and C58 strain mice, and rhesus monkeys). Further examples of similar effects, but caused by drugs instead of genotype, were met in the Test 1 and Test 4 sections. Here we learned that drugs altering dopaminergic activity in the basal ganglia—e.g., neuroleptics, potentially taken therapeutically by e.g., subjects with Parkinson’s disease; and stimulant drugs both taken recreationally by people and self-administered by rodents – can alter fronto-striatal systems of the brain to predispose subjects to ARB (e.g., Everitt and Robbins, 2005; Shukla and Pandey, 2020). Furthermore, prenatal conditions can have similar effects: rhesus monkeys exposed in utero either to radiation (Selemon and Friedman, 2013) or certain maternal antibodies (from human mothers of children with ASD (Martin et al., 2008), later go on to display more ARB as well as more behavioural inflexibility (with similar effects in mice: Camacho et al., 2014).
This all illustrates how prolonged negative affect is not the only cause of the impulsive, compulsive or perseverant behavioural selection systems that promote ARB: genetic background, prenatal effects and certain drugs can do this too. Thus, avoid these potential ‘false leads’. In welfare assessment, ARBs should only be used to compare treatments at the group level that are applied to genetically similar animals; environmental (not pharmacological), and applied post-natally (not pre-natally).
3.3. Using This Understanding to Refine the Use of ARBs in Welfare Assessment
To distill our conclusions on the relationships between ARBs and welfare, we provide six rules of thumb (or working hypotheses) for their valid use in welfare assessment. We also present counter-examples to the evidence laid out in Table 4 and show how they can now be understood. Scientifically, the rules of thumb that follow should be treated as testable working hypotheses: for certainty, each now needs testing (via experiment and/or meta-analysis), and on a case-by-case basis. This is because, as acknowledged in the Introduction, this review was narrative rather than systematic: our search process is non-replicable as well as not exhaustive, and our collation or interpretation of papers could have been skewed by unconscious biases. Furthermore, because ARBs are heterogeneous in their properties and underlying mechanisms, what seems true for most ARBs need not be true for every ARB. Nevertheless, pending such work, we propose these as reasonable rules of thumb for the valid use of ARBs in welfare assessment: ones far more likely to generate accurate, true assessments of animals’ states that either treating every bout as a sign of current distress (at one extreme) or ignoring ARBs altogether (at the other).
3.3.1. i) Sub-Optimal Environments That Promote High ARB Phenotypes Induce Negative Moods, Mood Disorders and/or Cumulative Negative Affect (Rule of Thumb 1)
Sustained negative experiences, and/or early negative experiences and/or recurrent negative experiences (with either additive or synergistic effects, perhaps accumulated over a lifespan), can be inferred when post-natal environments cause the emergence of ARBs. Such effects can include some infectious diseases, and they are accompanied by negative (e.g., anxious) moods, or even mood disorders (clinical anxiety, clinical depression), and might reflect cumulative negative affect. Furthermore, an increased severity/duration/recurrence of negative affect can be inferred from increasing extents of ARB (greater prevalence and/or time-budgets).
3.3.2. ii) Bouts of ARBs Do Not Validly Reflect Peaks of Negative Emotion (Rule of Thumb 2)
The onset/offset of ARB bouts should not be used to infer moment-to-moment changes in current negative emotion. Doing so risks false null errors (because bouts can be inhibited during negative events), and also ‘false leads’ (because ARB bouts can be expressed during calm or positively valenced moments; and some also appear to cause calming, relief and other relatively positive emotions).
3.3.3. iii) ARBs May Be Associated with Modest Improvements in Coping, but They Do Not Fully Rectify Welfare (Rule of Thumb 3)
Despite evidence linking ARBs with improved coping, humans with ARBs still report low mood, and for animals, challenges that lead to ARBs are still aversive and cause other signs of chronic stress. Thus, even where there is evidence for partial coping, treatments that induce ARB are still inducing sub-optimal welfare. However, the possibility of partial coping does mean that ARBs should not be used to compare welfare across identically-treated individuals who differ in ARB.
3.3.4. iv) As Indicators of Prolonged Negative Affect, ARBs Can Lack Responsiveness (So Risking ‘False Null’ Errors) (Rule of Thumb 4)
As welfare indicators, ARBs can be rather unresponsive to changes in negative affect: early, recurrent or prolonged negative experiences do not always promote the emergence of ARB. However, the risk of making false null errors in such cases may be reduced by, e.g., recognising inactivity as a competing response; probing with extra challenges to pass affective thresholds or reveal past priming effects; or measuring other variables that are sensitive to long-term negative affect. Prolonged or recurrent negative experiences also do not always promote increases in ARB. However, the risk of making ‘false null’ errors in such cases may be reduced by recognising the potential for ceiling effects, and for motivationally specific aetiologies, and again by measuring other variables sensitive to long-term negative affect.
3.3.5. v) Animals’ Brains Can Predispose Subjects to ARB, or Protect Them from It, via Non-Affective Processes That Are Potential Confounds (Rule of Thumb 5)
Genetic, prenatal and pharmacological effects on the neurological systems involved in behavioural selection (or on styles of responding to challenge) mean that ARBs should not be used to compare welfare between different individuals, families, strains or species; nor between animals exposed to drugs likely to affect such systems; nor between animals differing in prenatal experience. Doing so risks both false null errors (due to animals predisposed to, e.g., develop inactive responses instead), and ‘false lead’ errors (if neurological predispositions to ARB simply reflect predispositions to be active, impulsive, compulsive or perseverant/inflexible).
3.3.6. vi) General Activity Can Be a Confound That Covaries with ARBs, Potentially Generating ‘False Nulls’ and ‘False Leads’ (Rule of Thumb 6)
ARBs often covary with how active animals are overall (as affected by states as well as traits), and this can inflate or attenuate ARB performance in potentially misleading ways. Increases in general activity can promote ARBs even in relatively positive contexts (a potential ‘false lead’ or ‘false alarm’); while decreases in general activity can make ARBs seem less severe, even in aversive contexts (a potential) false null. Treating general activity as a potential confound, and correcting for it statistically, can therefore improve the accuracy of ARB in welfare assessment.
These six rules of thumb summarize both the utility and the potential limitations of ARBs as welfare indicators. To illustrate these caveats, Table 4 summarizes how different forms of ARB can misbehave as welfare indicators, even in taxa and for forms we know have construct validity. It also links each case to the relevant rule of thumb.
3.4. Some Outstanding Questions
3.4.1. What New Insights Does This Review Yield About Potential Mechanisms?
The evidence reviewed here raises the question, what mechanisms link exposure to prolonged or repeated negative experiences (including some infectious diseases) with the emergence of abnormally repetitive phenotypes? This vast topic is best served by another extensive review, but here we briefly summarise four complementary candidates, as well as outlining how cumulative negative experience could play a role (see also Figure 4).
The first is that performance of ARB ‘source behaviours’ increases during negative moods or mood disorders. After all, anxious subjects are more likely to perform both displacement activities (e.g., Norscia et al., 2021; Schino et al., 1996) and escape attempts (e.g., Jiao et al., 2011; Sege et al., 2018). Furthermore, clinically anxious people, or those with ‘agitated’ subtypes of depression, are often restless and hyperactive; and although this may be disorganised-looking in free-living humans, it can still manifest as predictable activities like pacing (e.g., Franklin et al., 2021; Lindenmayer, 2000; Sampogna et al., 2020). It is also easy to picture how such restlessness could be channelled into predictable repetition if subjects were confined in small, monotonous enclosures.
Neurophysiologically, anxiety- or depression-driven agitation has been hypothesized to reflect changes in cortico-striatal pathways, which then promote behavioural hyperactivity (Lindenmayer, 2000; Sharma and Huecker, 2024). And the broad role of such systems in ARB has long been appreciated, especially their rapid changes in function during acute stress, and their long-term impairment by abnormal developmental environments (Figure 1). However, on top, prolonged or repeated negative experiences are also now known to be impactful, acting on these systems so as to increase behavioural impulsivity (e.g., Girotti et al., 2022; Sanchez and Bangasser, 2022), compulsivity (e.g., (Adams et al., 2018; Wilson et al., 2023), and/or persistence (e.g., Hurtubise and Howland, 2017; Ko et al., 2024; Taylor et al., 2014). Thus, the sustained negative experiences that promote ARB may well do so via altering, perhaps even impairing, behavioural selection (e.g., cortico-striatal) functioning in heterogeneous ways that then cause the distinctive (yet varying) ‘oddness’ of all the ARBs: our second proposal.
Sustained negative affect may typically be the mediator here, perhaps thanks to changes in glucocorticoid levels and/or effects on neuroinflammation via microglia, the brain’s resident immune cells (e.g., Calcia et al., 2016; see also suggestions by Nordgreen et al., [2020] and Lawrence et al., [2025]; and reviews by Gagliano et al., [2025] and Wilson et al., [2023]). However, neuroinflammation can also result when negative experiences involve bacterial infections that cause the types of autoimmune effects implicated in PANDAS: cross-reactive antistreptococcal antibodies crossing the blood-brain barrier, binding to basal ganglia cells (e.g., Leonardi et al., 2024). Indeed, there seems to be an array of potential neuro-inflammatory risk factors for ARB (including gut microbiome composition, which can alter immune responses: e.g., Cheng et al., 2019; see also Mindus et al., 2021, Wang et al., 2023, and review by Homer et al., 2023). Neuroinflammation perhaps explains why stress and infection can have such similar, additive effects on ARB. This important area of research is still in its infancy, but it clearly highlights novel routes by which infectious diseases on farms, for example, could promote ARBs.
Our final candidate mechanism is a little different: it involves the adverse consequences of ARBs, and how being ARB-prone could itself reduce mood. In research on humans, especially children, much research sadly shows that ARB performance can trigger exclusion or bullying by others. Teens with Tourette’s syndrome can face social stigmatization and ridicule, for example (Cox et al., 2019), while in trichotillomania, hair loss from plucking is associated with low self-esteem and avoidance of social interactions (Christensen et al., 2023). In animals, the consequences of being ARB-prone can likewise be harmful, including bodily injuries from behavioural repetition (e.g., Baker, 2002; Kryukova et al., 2024), disruptions to maternal care (reviewed Díez-León and Mason, 2016) and reduced mating success Díez-León et al., 2013): all effects that would threaten fitness and so which a priori are likely to lower mood and reduce welfare.
A final issue concerns how cumulative negative experiences enhance ARB performance. The most basic explanation is via additive effects: thus, if one challenge is ‘bad’ (e.g., in terms of motivating functionless behaviours, or reducing behavioural disinhibition, or lowering mood), then multiple challenges are simply worse. But they can clearly also work non-additively, such as when combining synergistically to greatly increase an existing ARB, or when having latent priming effects (rendering animals more likely to develop ARBs only if challenged in the future). Mechanisms here may involve subjects being pushed over thresholds in terms of how negative their moods become; how likely they are to perform ARB source behaviours; and/or how dysfunctional their behavioural selection systems have become (predisposing them to compulsivity, impulsivity, and perseveration). But potentially, cumulative negative experiences could also enhance ARB performance by promoting the emergence of additional forms of ARB, with multiple sub-types then ‘stacking up’ as animals are progressively exposed to different types of motivational frustration and/or different types of threats to neurological function. We raise this because counting the number of forms (or ARB repertoire size) could then be a valid way to quantify the severity and welfare significance of animals’ ARB: one already common in human clinical work (e.g., García-Villamisar and Rojahn, 2015; Haas et al., 2021; Lam and Aman, 2007; Miller et al., 2006), yet not yet used in applied ethology.
Numbered arrows are explained below.
- Early/prolonged/recurrent sub-optimal environments induce not only repeated behaviour, and potential neurological dysfunction (as in Figure 1), but also negative moods (including states caused by infection), or even mood disorders.
- Negative moods (including states caused by infection) potentially promote ARB emergence by increasing motivations to escape, perform displacement activities or self-soothe; while mood disorders promote repeated behaviours by increasing ‘agitation’ / ‘restlessness’.
- Negative moods (including states caused by infection)/mood disorders potentially promote ARB emergence by leading to state-dependent neurological changes (perhaps involving neuroinflammation) in brain regions involved in behavioural control, so increasing behavioural disinhibition and inflexibility (impulsive, compulsive or perseverant behavioural selection).
- ARB performance then potentially feeds back to further lower mood, by compromising adaptive behaviour (e.g., normal social interactions, etc.)
Note that recurrent negative experiences can thus increase a given ARB via priming, synergistic effects or additive effects in terms of motivating functionless behaviours, by reducing behavioural disinhibition and/or by lowering low mood. They could also generate different forms of ARB, with animals potentially increasing their ARB repertoire sizes as they are exposed to more cumulative stress.
* Other developmental influences on neurological function of regions involved in behavioural control can be completely unrelated to affective states (e.g., reflecting genetic/prenatal effects, cf. Rule of Thumb 5)
3.4.2. How Should ARBs Be Categorised/Classified?
Throughout this review, we have alluded to knotty issues of identification and definition while avoiding tackling them. To end, we highlight the inadequacies of the deliberately loose term ‘ARB’, and discuss some alternatives, some of which rely on investigating mechanisms in the ways we suggest above.
The term ‘ARB’ was originally coined to avoid being too prescriptive and to provide a collective label for odd-looking behaviours (especially those whose mechanisms are not understood) (e.g., Mason, 2006; Mills and Luescher, 2006). However, it then potentially covers a truly vast array of heterogeneous activities, and with no direction as to what should be included versus excluded. This is one reason why the many behaviours we did not cover in this review (including hoarding and self-cutting in humans, and wheel-running and repetitive vocalisations in animals, to name but a few) are an arbitrary collection whose exclusion was more pragmatic than intellectually rigorous.
To date, alternatives to this loose ‘ARB’ term have included the traditional definition of ‘stereotypy’ as being repetitive, unvarying and having no apparent goal or function, but this too is problematic (e.g., Mason, 2006). For one, if we discover that a behaviour so-classified is sought out (like cribbing for horses) or has beneficial consequences (like self-biting in monkeys), it must lose the label ‘stereotypy’. It is also vague (e.g., how ‘repetitive’ does something have to be?). Furthermore, it assumes that being predictable and unvarying reveals something important about a behaviour. This is perhaps true in free-living humans, but it seems a flawed assumption when subjects’ very environments are predictable and unvarying. After all, as (Clubb and Vickery, 2006) put it, “there are only so many ways an animal can walk around in a small square cage”(p. 73); and note too how Table 3 finds some profound commonalities between ARBs typically termed ‘stereotypies’ and body-focused ones that are not.
Some have proposed the label of ‘compulsive disorder’, particularly in veterinary behavioural medicine (e.g., Frank, 2013; Luescher, 2004; Plato, 2022), as goal-directed behaviours that stem from conflict or frustration but become abnormal when repeated out of context (Mills and Luescher, 2006). While this is a welcome nod to human clinical perspectives, compulsiveness can be hard to operationalise or assess in animals, and furthermore, this then excludes those ARBs that are not compulsive (even if impulsive or perseverant in other ways). For example, if a dog repeatedly tail-chases (while never catching their tail), are they compulsive because the act is goal-directed (trying to catch the tail), or not, because the repetition is fruitless? And how could we know, given that we cannot access the animal’s thoughts or feelings (e.g., obsessions or irresistible ‘urges’ to perform the behaviour)? In either case, the behaviour remains worrying: a potential byproduct of poor welfare and/or underlying brain dysfunction.
To more explicitly capture this ‘wrongness’ that makes so many ARBs concerning, one of us suggested yet another definition. Mason (2006) proposed that all ARBs arising from frustration, repeated coping attempts, or neurological dysfunction be classified as ‘stereotypic behaviours’: a broad, heterogeneous category encompassing distinct mechanistic sub-groups, yet all reflecting that something is wrong with welfare, functioning or both. If this approach to a definition has merit, it could clearly be greatly refined by further research into underlying mechanism. However, it is a challenging type of definition for applied ethologists because it is reliant on fundamental research that may well not yet exist. For example, many of the ARBs linked with low mood or prolonged negative affect in this review are likely stereotypic behaviours under this definition, yet not all of them, because we currently lack a full enough understanding of their mechanistic aetiologies. More research is one obvious solution. But another could be to develop a definition based entirely on animal welfare.
We thus propose to label as ‘abnormal’ any repetitive behaviour that reliably indicates poor welfare, as demonstrated by construct validity tests. This definition thus relies on empirical evidence rather than superficial aspects of appearance or putative non-function; is flexible enough to accommodate heterogeneity in underlying mechanism; does not arbitrarily separate behaviours that share some fundamental commonalities; and is practical, providing applied ethologists with clear welfare-focused inclusion criteria. To refine this, we suggest that a behaviour should pass two or more of the four-to-five validity tests (recognising that Test 1 might be extremely relevant for non-human primates, but not for crib-biting in horses, say, due to non-homology across species), with consistent patterns within and across tests (see examples in Table 3). Furthermore, as in Table 3, evidence across species within Orders could be combined for similar-appearing ARBs. In addition, if there is counter-evidence, it should be easily explainable via our rules of thumb (cf. examples in Table 4). This welfare-/construct validity-based definition of ARB would span the potential sub-categories of ‘stereotypy’, ‘compulsive disorder’, and ‘stereotypic behaviour’, while usefully pooling the diverse ARBs that reflect poor welfare. We hope this would encourage similar research programs on a wider range of behaviours (not just ones that look highly repetitive or invariant, or which cause physical harm); and direct research attention towards affect-related mechanisms, including those recently revealed in biomedical and human clinical studies yet new to applied ethology.
As our own review has demonstrated, considering ARBs in this way offers a path toward a more rigorous, evidence-based, and biologically meaningful understanding of ARBs: one capable not only of identifying when animals are suffering, but also of perfecting their utility as welfare assessment tools, and revealing the underlying processes that give rise to these compelling, often misunderstood, behaviours.
References
- Adams, T.G.; Kelmendi, B.; Brake, C.A.; Gruner, P.; Badour, C.L.; Pittenger, C. The role of stress in the pathogenesis and maintenance of obsessive-compulsive disorder. Chronic Stress 2018, 2. [Google Scholar] [CrossRef] [PubMed]
- Ahola, L.; Mononen, J.; Mohaibes, M. Effects of access to extra cage constructions including a swimming opportunity on the development of stereotypic behaviour in singly housed juvenile farmed mink (Neovison vison). Appl. Anim. Behav. Sci. 2011, 134, 201–208. [Google Scholar] [CrossRef]
- Ahola, M.K.; Vapalahti, K.; Lohi, H. Early weaning increases aggression and stereotypic behaviour in cats. Sci. Rep. 2017, 1, 10412. [Google Scholar] [CrossRef] [PubMed]
- Akre, A.K.; Bakken, M.; Hovland, A.L.; Palme, R.; Mason, G.J. Clustered environmental enrichments induce more aggression and stereotypic behaviour than do dispersed enrichments in female mice. Appl. Anim. Behav. Sci. 2011, 131, 145–152. [Google Scholar] [CrossRef]
- Angevaare, M.J.; Prins, S.; van der Staay, F.J.; Nordquist, R.E. The effect of maternal care and infrared beak trimming on development, performance and behavior of Silver Nick hens. Appl. Anim. Behav. Sci. 2012, 140, 70–84. [Google Scholar] [CrossRef]
- Anonymous. OCD Inside [WWW Document]. Prison UK An Insid. View. 2014. Available online: https://prisonuk.blogspot.com/2014/07/ocd-inside.html.
- APA. Diagnostic and statistical manual of mental disorders: DSM-5-TR (5th edition, text revision.); American Psychiatric Association Publishing, 2022. [Google Scholar]
- Appleby, M.C.; Lawrence, A.B. Food restriction as a cause of steretypic behvaiour in tethered gilts. Am. J. Econ. Sociol. 1987, 45, 103–110. [Google Scholar] [CrossRef]
- Appleby, M.C.; Lawrence, A.B.; Illius, A.W. Influence of neighbours on stereotypic behaviour of tethered sows. Appl. Anim. Behav. Sci. 1989, 24, 137–146. [Google Scholar] [CrossRef]
- Arias-Esquivel, A.M.; Vasco, A.C.C. de M.; Lance, J.; Warren, L.K.; Rodriguez-Campos, L.A.; Lee, M.C.; Rodriguez, C.N.; Wickens, C.L. Investigating the gastrointestinal physiology of mature horses with and without a history of cribbing behavior in response to feeding a digestive support supplement. J. Equine Vet. Sci. 2024, 132, 104964. [Google Scholar] [CrossRef]
- Ashley, P.J. Fish welfare: Current issues in aquaculture. Appl. Anim. Behav. Sci. 2007, 104, 199–235. [Google Scholar] [CrossRef]
- Aydinonat, D.; Penn, D.J.; Smith, S.; Moodley, Y.; Hoelzl, F.; Knauer, F.; Schwarzenberger, F. Social isolation shortens telomeres in African Grey parrots (Psittacus erithacus erithacus). PLoS One 2014, 9, 2–6. [Google Scholar] [CrossRef]
- Bailoo, J.D.; Voelkl, B.; Varholick, J.; Novak, J.; Murphy, E.; Rosso, M.; Palme, R.; Würbel, H. Effects of weaning age and housing conditions on phenotypic differences in mice. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.G. Combination therapy for footpad lesions in a captive Bengal tiger (Panthera tigris tigris). J. Zoo Wildl. Med. 2002, 33, 389–391. [Google Scholar] [CrossRef] [PubMed]
- Baker, K.C.; Easley, S.P. An analysis of regurgitation and reingestion in captive chimpanzees. Appl. Anim. Behav. Sci. 1996, 49, 403–415. [Google Scholar] [CrossRef]
- Bandeli, M.; Mellor, E.L.; Kroshko, J.; Maherali, H.; Mason, G.J. The welfare problems of wide-ranging Carnivora reflect naturally itinerant lifestyles. R. Soc. Open Sci. 2023, 10. [Google Scholar] [CrossRef]
- Baribeau, D.A.; Vigod, S.; Pullenayegum, E.; Kerns, C.M.; Mirenda, P.; Smith, I.M.; Vaillancourt, T.; Volden, J.; Waddell, C.; Zwaigenbaum, L.; Bennett, T.; Duku, E.; Elsabbagh, M.; Georgiades, S.; Ungar, W.J.; Zaidman-Zait, A.; Szatmari, P. Repetitive behavior severity as an early indicator of risk for elevated anxiety symptoms in autism spectrum disorder. J. Am. Acad. Child Adolesc. Psychiatry 2020, 59, 890–899.e3. [Google Scholar] [CrossRef]
- Barnes, K. V.; Coughlin, F.R.; O’Leary, H.M.; Bruck, N.; Bazin, G.A.; Beinecke, E.B.; Walco, A.C.; Cantwell, N.G.; Kaufmann, W.E. Anxiety-like behavior in Rett syndrome: Characteristics and assessment by anxiety scales. J. Neurodev. Disord. 2015, 7, 1–14. [Google Scholar] [CrossRef]
- Bateson, M.; Poirier, C. Can biomarkers of biological age be used to assess cumulative lifetime experience? Anim. Welf. 2019, 28, 41–56. [Google Scholar] [CrossRef]
- Bateson, M.; Seeker, L.A. Telomere length Assessing Animal Welfare: A Guide to the Valid Use of Indicators of Affective States. In UFAW Animal Welfare Series; Mason, G.J., Nielsen, B.L., Mendl, M.T., Eds.; John Wiley & Sons Ltd.: Oxford, n.d. [Google Scholar]
- Bauer, E.; Babitz, M.; Boedeker, N.; Hellmuthh, H. Approaches to understanding and managing pacing in sloth bears in a zoological setting. Int. J. Comp. Psychol. 2013, 26. [Google Scholar] [CrossRef]
- Bechard, A.; Meagher, R.; Mason, G. Environmental enrichment reduces the likelihood of alopecia in adult C57BL/6J mice. J. Am. Assoc. Lab. Anim. Sci. 2011, 50, 171–174. [Google Scholar]
- Bechard, A.R.; Bliznyuk, N.; Lewis, M.H.; Bliznyuk, N.; Lewis, M.H. The development of repetitive motor behaviors in deer mice: Effects of environmental enrichment, repeated testing, and differential mediation by indirect basal ganglia pathway activation. Dev. Psychobiol. 2017, 59, 390–399. [Google Scholar] [CrossRef]
- Bechard, A.R.; Nicholson, A.; Mason, G.J. Litter size predicts adult stereotypic behavior in female laboratory mice. J. Am. Assoc. fr Lab. Anim. Sci. 2012, 51, 407–411. [Google Scholar]
- Beckett, C.; Bredenkamp, D.; Castle, J.; Groothues, C.; O’Connor, T.G.; Rutter, M. Behavior patterns associated with institutional deprivation: A study of children adopted from Romania. J. Dev. Behav. Pediatr. 2002, 23, 297–303. [Google Scholar] [CrossRef]
- Beckett, C.; Castle, J.; Groothues, C.; O’Connor, T.; Rutter, M. Health problems in children adopted from Romania: Association with duration of deprivation and behavioural problems. Adopt. Foster. 2003, 27, 19–29. [Google Scholar] [CrossRef]
- Bécuwe-Bonnet, V.; Bélanger, M.C.; Frank, D.; Parent, J.; Hélie, P. Gastrointestinal disorders in dogs with excessive licking of surfaces. J. Vet. Behav. Clin. Appl. Res. 2012, 7, 194–204. [Google Scholar] [CrossRef]
- Behen, M.E.; Muzik, O.; Saporta, A.S.D.; Wilson, B.J.; Pai, D.; Hua, J.; Chugani, H.T. Abnormal fronto-striatal connectivity in children with histories of early deprivation: A diffusion tensor imaging study. Brain Imaging Behav. 2009, 3, 292–297. [Google Scholar] [CrossRef]
- Bellanca, R.U.; Crockett, C.M. Factors predicting increased incidence of abnormal behavior in male pigtailed macaques. Am. J. Primatol. 2002, 58, 57–69. [Google Scholar] [CrossRef]
- Benhajali, H.; Ezzaouia, M.; Lunel, C.; Charfi, F.; Hausberger, M. Stereotypic behaviours and mating success in domestic mares. Appl. Anim. Behav. Sci. 2014, 153, 36–42. [Google Scholar] [CrossRef]
- Benjet, C.; Bromet, E.; Karam, E.G.; Kessler, R.C.; Mclaughlin, K.A.; Ruscio, A.M.; Shahly, V.; Stein, D.J.; Petukhova, M.; Hill, E.; Alonso, J.; Atwoli, L.; Bunting, B.; De Girolamo, G.; Florescu, S.; Gureje, O.; Lepine, J.P.; Kawakami, N.; Kovess-Masfety, V.; Piazza, M.; Scott, K.M.; Shalev, A.; Slade, T.; Torres, Y.; Viana, M.C.; Zarkov, Z.; Koenen, K.C. The epidemiology of traumatic event exposure worldwide: results from the World Mental Health Survey Consortium. In Psychol. Med.; The, 2016; Volume 46, pp. 327–343. [Google Scholar] [CrossRef]
- Benoit, T.C.; Jocelyn, L.J.; Moddemann, D.M.; Embree, J.E. Romanian adoption: The Manitoba experience. Arch. Pediatr. Adolesc. Med. 1996, 150, 1278–1282. [Google Scholar] [CrossRef]
- Berg, K.L.; Shiu, C.S.; Acharya, K.; Stolbach, B.C.; Msall, M.E. Disparities in adversity among children with autism spectrum disorder: a population-based study. Dev. Med. Child Neurol. 2016, 58, 1124–1131. [Google Scholar] [CrossRef]
- Bergeron, R.; Badnell-Waters, A.J.; Lambton, S.; Mason, G. Stereotypic oral behvaiour in captive ungulates: Foraging, diet and gastrointestinal function. In Stereotypic Animal Behaviour Funadamentals and Applications to Welfare. CABI; Mason, G., Rushen, J., Eds.; Wallingford, 2006; pp. 19–57. [Google Scholar]
- Berkson, G.; Mason, W.A. Stereotyped behaviors of chimpanzees: Relation to general arousal and alternative activities. Percept. Mot. Skills 1964, 19, 635–662. [Google Scholar] [CrossRef]
- Bhidayasiri, R.; Truong, D.D. Chorea and related disorders. Postgrad. Med. J. 2004, 80, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Bildsøe, M.; Heller, K.E.; Jeppesen, L.L. Effects of immobility stress and food restriction on stereotypies in low and high stereotyping female ranch mink. Behav. Processes 1991, 25, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Blokhuis, H.J.; Arkes, J.G. Some observations on the development of feather-pecking in poultry. Appl. Anim. Behav. Sci. 1984, 12, 145–157. [Google Scholar] [CrossRef]
- Boger, S.; Ehring, T.; Schwarzkopf, W.; Werner, G.G. Potential mediators of the association between childhood maltreatment and obsessive-compulsive disorder in adulthood. J. Obsessive. Compuls. Relat. Disord. 2020, 27, 100587. [Google Scholar] [CrossRef]
- Boinski, S.; Gross, T.S.; Davis, J.K. Terrestrial predator alarm vocalizations are a valid monitor of stress in captive brown capuchins (Cebus apella). Zoo Biol. 1999, 18, 295–312. [Google Scholar] [CrossRef]
- Bos, H.; Van Balen, F. Children of the new reproductive technologies: Social and genetic parenthood. Patient Educ. Couns. 2010, 81, 429–435. [Google Scholar] [CrossRef]
- Bos, K.; Zeanah, C.; Smyke, A.; Fox, N.; Nelson, C. Stereotypies in children with a history of early institutional care. Arch. Pediatr. Adolesc. Med. 2010, 164, 406–411. [Google Scholar] [CrossRef]
- Boyle, L.A.; Edwards, S.A.; Bolhuis, J.E.; Pol, F.; Šemrov, M.Z.; Schütze, S.; Nordgreen, J.; Bozakova, N.; Sossidou, E.N.; Valros, A. The evidence for a causal link between disease and damaging behavior in pigs. Front. Vet. Sci. 2022, 8. [Google Scholar] [CrossRef]
- Brent, L.; Lee, D.R.; Eichberg, J.W. The effects of single caging on chimpanzee behavior. Lab. Anim. Sci. 1989, 39, 345–346. [Google Scholar]
- Broom, D.; Mendl, M.; Zanella, A. A comparison of the welfare of sows in different housing conditions. Anim. Sci. 1995, 61, 369–385. [Google Scholar] [CrossRef]
- Broom, D.M.; Johnson, K.G. Stress and Animal Welfare; Chapman && Hall: London, 1993. [Google Scholar]
- Brummer, S.P.; Gese, E.M.; Shivik, J.A. The effect of enclosure type on the behavior and heart rate of captive coyotes. Appl. Anim. Behav. Sci. 2010, 125, 171–180. [Google Scholar] [CrossRef]
- Bucks, R.S.; Gidron, Y.; Harris, P.; Teeling, J.; Wesnes, K.A.; Perry, V.H. Selective effects of upper respiratory tract infection on cognition, mood and emotion processing: A prospective study. Brain. Behav. Immun. 2008, 22, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Buisson, N.B.; Hully, M.; Celestin, E. Stereotypies and movement disorders in Rett syndrome [WWW Document]. www.rettsyndrome.eu. 2017. Available online: https://www.rettsyndrome.eu/wp-content/uploads/2018/05/Stereotypies.pdf.
- Burke, J.T.; Mograbi, D.C.; Wolmarans, D.W. Behavioral restriction, lorazepam, and escitalopram uniquely influence the expression of naturalistic stereotypy in deer mice: perspectives on anxiety- and compulsive-like behavior. Front. Behav. Neurosci. 2022, 16. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.G.; Saravanan, A.; Helen Rodd, F. Rearing environment affects the brain size of guppies: Lab-reared guppies have smaller brains than wild-caught guppies. Ethology 2009, 115, 122–133. [Google Scholar] [CrossRef]
- Buse, J.; Kirschbaum, C.; Leckman, J.F.; Münchau, A.; Roessner, V. The modulating role of stress in the onset and course of Tourette’s syndrome: A review. Behav. Modif. 2014, 38, 184–216. [Google Scholar] [CrossRef]
- Cabanac, M. Physiological role of pleasure. Science (80-. ) 1971, 173, 1103–1107. [Google Scholar] [CrossRef]
- Cabib, S. The neurobiology of stereotypy II: The role of stress. In Stereotypic Animal Behaviour Funadamentals and Applications to Welfare; Mason, G.J., Rushen, J., Eds.; CABI: Wallingford, 2006; pp. 227–255. [Google Scholar]
- Cabib, S.; Bonaventura, N. Parallel strain-dependent susceptibility to environmentally-induced stereotypies and stress-induced behavioral sensitization in mice. Physiol. Behav. 1997, 61, 499–506. [Google Scholar] [CrossRef]
- Cabib, S.; Campus, P.; Conversi, D.; Orsini, C.; Puglisi-Allegra, S. Functional and dysfunctional neuroplasticity in learning to cope with stress. Brain Sci. 2020, 10. [Google Scholar] [CrossRef]
- Cabib, S.; Campus, P.; Latagliata, E.C.; Orsini, C.; Tarmati, V. Repetitive and inflexible active coping and addiction-like neuroplasticity in stressed mice of a helplessness – resistant inbred strain. Behav. Sci. (Basel) 2021, 11, 174. [Google Scholar] [CrossRef]
- Cabib, S.; Giardino, L.; Calza, L.; Zanni, M.; Mele, A.; Puglisi-Allegra, A. Stress promotes major changes in dopamine receptor densities within the mesoaccumbens and nigrostraital systems. Neuroscience 1998, 84, 193–200. [Google Scholar] [CrossRef]
- Cait, J.; Cait, A.; Scott, R.W.; Winder, C.B.; Mason, G.J. Conventional laboratory housing increases morbidity and mortality in research rodents: results of a meta-analysis. BMC Biol. 2022, 20, 1–22. [Google Scholar] [CrossRef]
- Calcia, M.A.; Bonsall, D.R.; Bloomfield, P.S.; Selvaraj, S.; Barichello, T.; Howes, O.D. Stress and neuroinflammation: A systematic review of the effects of stress on microglia and the implications for mental illness. Psychopharmacology (Berl) 2016, 233, 1637–1650. [Google Scholar] [CrossRef] [PubMed]
- Camacho, J.; Jones, K.; Miller, E.; Ariza, J.; Noctor, S.; Van de Water, J.; Martínez-Cerdeño, V. Embryonic intraventricular exposure to autism-specific maternal autoantibodies produces alterations in autistic-like stereotypical behaviors in offspring mice. Behav. Brain Res. 2014, 266, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Câmara-Souza, M.B.; Carvalho, A.G.; Figueredo, O.M.C.; Bracci, A.; Manfredini, D.; Rodrigues Garcia, R.C.M. Awake bruxism frequency and psychosocial factors in college preparatory students. Cranio 2023, 41, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Campbell, D.L.M.; Dallaire, J.A.; Mason, G.J. Environmentally enriched rearing environments reduce repetitive perseveration in caged mink, but increase spontaneous alternation. Behav. Brain Res. 2013, 239, 177–187. [Google Scholar] [CrossRef]
- Canadian Council on Animal Care CCAC guidelines: Animal welfare assessment. 2021.
- Capitanio, J.P.; Cacioppo, S.; Cole, S.W. Loneliness in monkeys: neuroimmune mechanisms. Curr. Opin. Behav. Sci. 2019, 28, 51–57. [Google Scholar] [CrossRef]
- Cappadocia, M.C.; Weiss, J.A.; Pepler, D. Bullying experiences among children and youth with autism spectrum disorders. J. Autism Dev. Disord. 2012, 42, 266–277. [Google Scholar] [CrossRef]
- Carlson, M.; Earls, F. Psychological and neuroendocrinological sequelae of early social deprivation in institutionalized children in Romania. Ann. N. Y. Acad. Sci. 1997, 807, 419–428. [Google Scholar] [CrossRef]
- Carlstead, K.; Brown, J.L. Relationships between patterns of fecal corticoid excretion and behavior, reproduction, and environmental factors in captive black (Diceros bicornis) and white (Ceratotherium simum) rhinoceros. Zoo Biol. 2005, 24, 215–232. [Google Scholar] [CrossRef]
- Carroll, S.L.; Sykes, B.W.; Mills, P.C. Understanding and treating equine behavioural problems. Vet. J. 2023, 296–297, 105985. [Google Scholar] [CrossRef]
- Carroll, S.L.; Sykes, B.W.; Mills, P.C. An online survey investigating perceived prevalence and treatment options for stereotypic behaviours in horses and undesirable behaviours associated with handling and riding. Equine Vet. Educ. 2020, 32, 71–81. [Google Scholar] [CrossRef]
- Castellanos, F.X.; Ritchie, G.F.; Marsh, W.L.; Rapoport, J.L. DSM-IV stereotypic movement disorder: Persistence of stereotypies of infancy in intellectually normal adolescents and adults. J. Clin. Psychiatry 1996, 57, 116–122. [Google Scholar] [PubMed]
- Cavic, E.; Valle, S.; Chamberlain, S.R.; Grant, J.E. Sleep quality and its clinical associations in trichotillomania and skin picking disorder. Compr. Psychiatry 2021, 105, 152221. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, S. Lifting the veil on trichotillomania. Am. J. Psychiatry 2007, 164, 568. [Google Scholar] [CrossRef]
- Chen, E.; Pipolo, G.; Crailsheim, D.; Morimoto, J. The lasting impact of social isolation: behavioral insights from former pet and entertainer chimpanzees in a sanctuary in Spain. Am. J. Primatol. 2025, 87. [Google Scholar] [CrossRef]
- Cheng, H.Y.; Ning, M.X.; Chen, D.K.; Ma, W.T. Interactions between the gut microbiota and the host innate immune response against pathogens. Front. Immunol. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Chosy, J.; Wilson, M.; Santymire, R. Behavioral and physiological responses in felids to exhibit construction. Zoo Biol. 2014, 33, 267–274. [Google Scholar] [CrossRef]
- Chow, A.; Hogan, J.A. The development of feather pecking in Burmese red junglefowl: the influence of early experience with exploratory-rich environments. Appl. Anim. Behav. Sci. 2005, 93, 283–294. [Google Scholar] [CrossRef]
- Christensen, R.E.; Tan, I.; Jafferany, M. Recent advances in trichotillomania: A narrative review. Acta Dermatovenerologica Alpina, Pannonica Adriat. 2023, 32, 151–157. [Google Scholar] [CrossRef]
- Chugani, H.T.; Behen, M.E.; Muzik, O.; Juhá, C.; Nagy, F.; Chugani, D.C. Local brain functional activity following early deprivation: A study of postinstitutionalized romanian orphans. Neuroimage 2001, 14, 1290–1301. [Google Scholar] [CrossRef]
- Cianfaglione, R.; Clarke, A.; Kerr, M.; Hastings, R.P.; Oliver, C.; Moss, J.; Heald, M.; Felce, D. A national survey of Rett syndrome: Behavioural characteristics. J. Neurodev. Disord. 2015, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cianfaglione, R.; Meek, A.; Clarke, A.; Kerr, M.; Hastings, R.P.; Felce, D. Direct observation of the behaviour of females with Rett syndrome. J. Dev. Phys. Disabil. 2016, 28, 425–441. [Google Scholar] [CrossRef] [PubMed]
- Cinini, S.M.; Barnabe, G.F.; Galvão-Coelho, N.; de Medeiros, M.A.; Perez-Mendes, P.; Sousa, M.B.C.; Covolan, L.; Mello, L.E. Social isolation disrupts hippocampal neurogenesis in young non-human primates. Front. Neurosci. 2014, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.S.; Lindburg, D.G. Behavioral contrasts between male cynomolgus and lion-tailed macaques. Am. J. Primatol. 1993, 29, 49–59. [Google Scholar] [CrossRef]
- Clay, A.W.; Bloomsmith, M.A.; Bard, K.A.; Maple, T.L.; Marr, M.J. Long-term effects of infant attachment organization on adult behavior and health in nursery-reared, captive chimpanzees (Pan troglodytes). J. Comp. Psychol. 2015, 129, 145–159. [Google Scholar] [CrossRef]
- Clipperton-Allen, A.E.; Ingrao, J.C.; Ruggiero, L.; Batista, L.; Ovari, J.; Hammermueller, J.; Armstrong, J.N.; Bienzle, D.; Choleris, E.; Turner, P. V. Long-term provision of environmental resources alters behavior but not physiology or neuroanatomy of male and female BALB/c and C57BL/6 Mice. J. Am. Assoc. Lab. Anim. Sci. 2015, 54, 718–30. [Google Scholar]
- Clubb, R.; Mason, G. Capitivity effects on wide-range carnivores. Nature 2003, 425, 473–474. [Google Scholar] [CrossRef]
- Clubb, R.; Rowcliffe, M.; Lee, P.; Mar, K.U.; Moss, C.; Mason, G.J. Compromised survivorship in zoo elephants. Science (80-. ) 2008, 322, 1649. [Google Scholar] [CrossRef]
- Clubb, R.; Vickery, S. Locomotory stereotypies in carnivores: Does pacing stem from hunting, ranging or frustrated escape? In Stereotypic Animal Behaviour Funadamentals and Applications to Welfare; Mason, G., Rushen, J., Eds.; CABI: Wallingford, 2006; pp. 58–85. [Google Scholar]
- Cohen, S.; Murphy, M.L.M.; Prather, A.A. Ten surprising facts about stressful life events and disease risk. Annu. Rev. Psychol. 2019, 70, 577–597. [Google Scholar] [CrossRef]
- Conti, G.; Hansman, C.; Heckman, J.J.; Novak, M.F.X.; Ruggiero, A.; Suomi, S.J. Primate evidence on the late health effects of early-life adversity. Proc. Natl. Acad. Sci. U. S. A 2012, 109, 8866–8871. [Google Scholar] [CrossRef]
- Cooper, J.J.; Appleby, M.C. Motivational aspects of individual variation in response to nestboxes by laying hens. Anim. Behav. 1997, 54, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.J.; Appleby, M.C. Nesting behaviour of hens: Effects of experience on motivation. Appl. Anim. Behav. Sci. 1995, 42, 283–295. [Google Scholar] [CrossRef]
- Cooper, J.J.; Mason, G.J. The use of operant technology to measure behavioral priorities in captive animals. Behav. Res. Methods, Instruments Comput. 2001, 33, 427–434. [Google Scholar]
- Cooper, J.J.; Mason, G.J. Increasing costs of access to resources cause re-scheduling of behaviour in American mink (Mustela vison): Implications for the assessment of behavioural priorities. Appl. Anim. Behav. Sci. 2000, 66, 135–151. [Google Scholar] [CrossRef]
- Cooper, J.J.; Nicol, C.J. Stereotypic behaviour in wild caught and laboratory bred bank voles. Anim. Welf. 1996, 5, 245–257. [Google Scholar] [CrossRef]
- Cooper, J.J.; Nicol, C.J. Stereotypic behaviour affects environmental preference in bank voles, Clethrionomys glareolus. Anim. Behav. 1991, 41, 971–977. [Google Scholar] [CrossRef]
- Cooper, J.J.; Ödberg, F.O.; Nicol, C.J. Limitations on the effectiveness of environmental improvement in reducing stereotypic behaviour in bank voles (Clethrionomys glareolus). Appl. Anim. Behav. Sci. 1996, 48, 237–248. [Google Scholar] [CrossRef]
- Corsetti, S.; Natoli, E.; Palme, R.; Viggiano, E. Intraspecific interactions decrease stress affecting welfare in shelter dogs: A comparison of four different housing conditions. Animals 2023, 13, 1–12. [Google Scholar] [CrossRef]
- Costa, P.; Macchi, E.; Tomassone, L.; Ricceri, F.; Bollo, E.; Scaglione, F.E.; Tarantola, M.; De Marco, M.; Prola, L.; Bergero, D.; Schiavone, A. Feather picking in pet parrots: Sensitive species, risk factor and ethological evidence. Ital. J. Anim. Sci. 2016, 15, 473–480. [Google Scholar] [CrossRef]
- Cox, J.H.; Nahar, A.; Termine, C.; Agosti, M.; Balottin, U.; Seri, S.; Cavanna, A.E. Social stigma and self-perception in adolescents with tourette syndrome. Adolesc. Health. Med. Ther. 2019, 10, 75–81. [Google Scholar] [CrossRef]
- Craig, A.D. A new view of pain as a homeostatic emotion. Trends Neurosci. 2003, 26, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Cromer, K.R.; Schmidt, N.B.; Murphy, D.L. An investigation of traumatic life events and obsessive-compulsive disorder. Behav. Res. Ther. 2007, 45, 1683–1691. [Google Scholar] [CrossRef] [PubMed]
- Cronin, G.M.; Glatz, P.C. Causes of feather pecking and subsequent welfare issues for the laying hen: a review. Anim. Prod. Sci. 2021, 61, 990–1005. [Google Scholar] [CrossRef]
- Cronin, G.M.; Wiepkema, P.R. An analysis of stereotyped behaviour in tethered sows. Ann. Rech. Vétérinaires 1984, 15, 263–270. [Google Scholar]
- Cross, H.A.; Harlow, H.F. Prolonged and progressive effects of partial isolation on the behaviour of macaque monkeys. J. Exp. Res. Personal. 1965, 1, 39–49. [Google Scholar]
- Cussen, V.A.; Mench, J.A. The relationship between personality dimensions and resiliency to environmental stress in orange-winged amazon parrots (Amazona amazonica), as indicated by the development of abnormal behaviors. PLoS One 2015, 10, e0126170. [Google Scholar] [CrossRef]
- D’Angelo, L.-S.C.; Eagle, D.M.; Grant, J.E.; Fineberg, N.A.; Robbins, T.W.; Chamberlain, S.R. Animal models of obsessive-compulsive spectrum disorders. CNS Spectr. 2014, 19, 28–49. [Google Scholar] [CrossRef]
- D’Eath, R.B.; Tolkamp, B.J.; Kyriazakis, I.; Lawrence, A.B. “Freedom from hunger” and preventing obesity: The animal welfare implications of reducing food quantity or quality. Anim. Behav. 2009, 77, 275–288. [Google Scholar] [CrossRef]
- Dale, M.; van Duijn, E. Anxiety in Huntington’s disease. J. Neuropsychiatry Clin. Neurosci. 2015, 27, 262–271. [Google Scholar] [CrossRef]
- Dallaire, J.A.; Meagher, R.K.; Díez-León, M.; Garner, J.P.; Mason, G.J. Recurrent perseveration correlates with abnormal repetitive locomotion in adult mink but is not reduced by environmental enrichment. Behav. Brain Res. 2011, 224, 213–222. [Google Scholar] [CrossRef]
- Dallaire, J.A.; Meagher, R.K.; Mason, G.J. Individual differences in stereotypic behaviour predict individual differences in the nature and degree of enrichment use in caged American mink. Appl. Anim. Behav. Sci. 2012, 142, 98–108. [Google Scholar] [CrossRef]
- Dantzer, R. Cytokine, sickness behavior, and depression. Immunol. Allergy Clin. North Am. 2009, 29, 247–264. [Google Scholar] [CrossRef] [PubMed]
- Davenport, M.D.; Lutz, C.K.; Tiefenbacher, S.; Novak, M.A.; Meyer, J.S. A rhesus monkey model of self-injury: Effects of relocation stress on behavior and neuroendocrine function. Biol. Psychiatry 2008, 63, 990–996. [Google Scholar] [CrossRef] [PubMed]
- Dawkins, M.S. Farm animal welfare: Beyond “natural” behavior. Science (80-. ) 2023, 379, 326–328. [Google Scholar] [CrossRef]
- Dawkins, M.S. The Science of Animal Welfare: Understanding what Animals Want; Oxford University Press, 2021. [Google Scholar]
- Dawkins, M.S. Behavioural deprivation: A central problem in animal welfare. Appl. Anim. Behav. Sci. 1988, 20, 209–225. [Google Scholar] [CrossRef]
- Dawkins, M.S.; Beardsley, T. Reinforcing properties of access to litter in hens. Appl. Anim. Behav. Sci. 1986, 15, 351–364. [Google Scholar] [CrossRef]
- De Haas, E.N.; Bolhuis, J.E.; De Jong, I.C.; Kemp, B.; Janczak, A.M.; Rodenburg, T.B. Predicting feather damage in laying hens during the laying period. Is it the past or is it the present? Appl. Anim. Behav. Sci. 2014, 160, 75–85. [Google Scholar] [CrossRef]
- de Haas, E.N.; van der Eijk, J.A.J. Where in the serotonergic system does it go wrong? Unravelling the route by which the serotonergic system affects feather pecking in chickens. Neurosci. Biobehav. Rev. 2018, 95, 170–188. [Google Scholar] [CrossRef]
- de Passillé, A.M.; Christopherson, R.; Rushen, J. Non-nutritive sucking by the calf and postprandial secretion of insulin, CCK and gastrin. Physiol. Behav. 1993, 54, 1069–1073. [Google Scholar] [CrossRef]
- de Passillé, A.M.; Metz, J.H.M.; Mekking, P.; Wiepkema, P.R. Does drinking milk stimulate sucking in young calves? Appl. Anim. Behav. Sci. 1992, 34, 23–36. [Google Scholar] [CrossRef]
- de Passillé, A.M.; Rushen, J. Motivational and physiological analysis of the causes and consequences of non-nutritive sucking by calves. Appl. Anim. Behav. Sci. 1997, 53, 15–31. [Google Scholar] [CrossRef]
- De Paula Vieira, A.; Guesdon, V.; de Passillé, A.M.; von Keyserlingk, M.A.G.; Weary, D.M. Behavioural indicators of hunger in dairy calves. Appl. Anim. Behav. Sci. 2008, 109, 180–189. [Google Scholar] [CrossRef]
- Deaton, J.E.; Berg, S.W.; Richlin, M.; Litrownik, A.J. Coping activities in solitary confinement of U.S. Navy POWs in Vietnam. J. Appl. Soc. Psychol. 1977, 7, 239–257. [Google Scholar] [CrossRef]
- Delgado, M.M.; Walcher, I.; Buffington, C.A.T. A survey-based assessment of risk factors for cross-sucking behaviors in neonatal kittens, Felis catus. Appl. Anim. Behav. Sci. 2020, 230, 105069. [Google Scholar] [CrossRef]
- Dellapiazza, F.; Michelon, C.; Picot, M.C.; Baghdadli, A. Early risk factors for anxiety disorders in children with autism spectrum disorders: results from the ELENA Cohort. Sci. Rep. 2022, 12, 1–8. [Google Scholar] [CrossRef]
- Desmarchelier, M.; Ferrell, S.T.; Frank, D. A systematic approach to behavior cases: The key to the right diagnosis - clinical cases. In Proceedings of the American Association of Zoo Veterinarians Annual Conference, 2016. [Google Scholar]
- Desmarchelier, M.R. A systematic approach in diagnosing behavior problems. In Fowler’s Zoo and Wild Animal Medicine Current Therapy; Miller, R.E., Calle, P.P., Lamberski, N., Eds.; Elsevier Inc., 2019; Volume 9, pp. 76–82. [Google Scholar] [CrossRef]
- Devanarayana, N.M.; Rajindrajith, S. Aerophagia among Sri Lankan schoolchildren: Epidemiological patterns and symptom characteristics. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 516–520. [Google Scholar] [CrossRef]
- Díez-León, M.; Bowman, J.; Bursian, S.; Filion, H.; Galicia, D.; Kanefsky, J.; Napolitano, A.; Palme, R.; Schulte-Hostedde, A.; Scribner, K.; Mason, G.J. Environmentally enriched male mink gain more copulations than stereotypic, barren-reared competitors. PLoS One 2013, 8, 1–12. [Google Scholar] [CrossRef]
- Díez-León, M.; Bursian, S.; Galicia, D.; Napolitano, A.; Palme, R.; Mason, G. Environmentally enriching American mink (Neovison vison) increases lymphoid organ weight and skeletal symmetry, and reveals differences between two sub-types of stereotypic behaviour. Appl. Anim. Behav. Sci. 2016, 177, 59–69. [Google Scholar] [CrossRef]
- Díez-León, M.; Kitchenham, L.; Duprey, R.; Bailey, C.D.C.; Choleris, E.; Lewis, M.H.; Mason, G.J. Neurophysiological correlates of stereotypic behaviour in a model carnivore species. Behav. Brain Res. 2019, 373, 112056. [Google Scholar] [CrossRef]
- Díez-León, M.; Mason, G. Effects of environmental enrichment and stereotypic behavior on maternal behavior and infant viability in a model carnivore, the American mink (Neovison vison). Zoo Biol. 2016, 35, 19–28. [Google Scholar] [CrossRef]
- Dixon, L.M.; Duncan, I.J.H. Changes in substrate access did not affect early feather-pecking behavior in two strains of laying hen chicks. J. Appl. Anim. Welf. Sci. 2010, 13, 1–14. [Google Scholar] [CrossRef]
- Dixon, L.M.; Duncan, I.J.H.; Mason, G. What’s in a peck? Using fixed action pattern morphology to identify the motivational basis of abnormal feather-pecking behaviour. Anim. Behav. 2008, 76, 1035–1042. [Google Scholar] [CrossRef]
- Dixon, L.M.; Dunn, I.C.; Brocklehurst, S.; Baker, L.; Boswell, T.; Caughey, S.D.; Reid, A.; Sandilands, V.; Wilson, P.W.; D’Eath, R.B. The effects of feed restriction, time of day, and time since feeding on behavioral and physiological indicators of hunger in broiler breeder hens. Poult. Sci. 2022, 101, 101838. [Google Scholar] [CrossRef] [PubMed]
- Dodds, R.L. An exploratory review of the associations between adverse experiences and autism. J. Aggress. Maltreatment Trauma 2021, 30, 1093–1112. [Google Scholar] [CrossRef]
- Doneley, R.J.T. Bacterial and parasitic diseases of parrots. Vet. Clin. NA Exot. Pet 2009, 12, 417–432. [Google Scholar] [CrossRef]
- Drury, S.S.; Theall, K.; Gleason, M.M.; Smyke, A.T.; De Vivo, I.; Wong, J.Y.Y.; Fox, N.A.; Zeanah, C.H.; Nelson, C.A. Telomere length and early severe social deprivation: Linking early adversity and cellular aging. Mol. Psychiatry 2012, 17, 719–727. [Google Scholar] [CrossRef]
- Duncan, I.J.H.; Rushen, J.; Lawrence, A.B. Conclusions and implications for animal welfare. In Stereotypic Animal Behaviour: Fundamentals and Applications to Welfare; Lawrence, A.B., Rushen, J., Eds.; CAB International: Wallinford, 1993; pp. 193–206. [Google Scholar]
- Duncan, I.J.H.; Wood-Gush, D.G.M. The effect of a rauwolfia tranquilizer on stereotyped movements in frustrated domestic fowl. Appl. Anim. Ethol. 1974, 1, 67–76. [Google Scholar] [CrossRef]
- Duncan, I.J.H.; Wood-Gush, D.G.M. Thwarting of feeding behaviour in the domestic fowl. Anim. Behav. 1972, 20, 444–451. [Google Scholar] [CrossRef]
- Ebisawa, K.; Nakayama, S.; Pai, C.; Kinoshita, R.; Koie, H. Prevalence and risk factors for feather-damaging behavior in psittacine birds: Analysis of a Japanese nationwide survey. PLoS One 2021, 16, 1–15. [Google Scholar] [CrossRef]
- Egeland, B.; Sroufe, L.A.; Erickson, M. The developmental consequence of different patterns of maltreatment. Child Abus. Negl. 1983, 7, 459–469. [Google Scholar] [CrossRef]
- Ellis, H.H.; Fisher, P.A.; Zaharie, S. Predictors of disruptive behavior, developmental delays, anxiety, and affective symptomatology among institutionally reared Romanian children. J. Am. Acad. Child Adolesc. Psychiatry 2004, 43, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Engel, A.K.J.; Gross, A.N.; Richter, S.H.; Rommen, J.; Touma, C.; Würbel, H. Variation in stress reactivity affects cage-induced stereotypies in female CD-1 (ICR) mice. Appl. Anim. Behav. Sci. 2011, 133, 101–108. [Google Scholar] [CrossRef]
- Everitt, B.J.; Robbins, T.W. Neural systems of reinforcement for drug addiction: From actions to habits to compulsion. Nat. Neurosci. 2005, 8, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Faccin, J.E.G.; Laskoski, F.; Hernig, L.F.; Kummer, R.; Lima, G.F.R.; Orlando, U.A.D.; Goncalves, M.A.D.; Mellagi, A.P.G.; Ulguim, R.R.; Bortolozzo, F.P. Impact of increasing weaning age on pig performance and belly nosing prevalence in a commercial multisite production system. J. Anim. Sci. 2020, 98, 1–8. [Google Scholar] [CrossRef]
- Fasano, A.; Petrovic, I. Insights into pathophysiology of punding reveal possible treatment strategies. Mol. Psychiatry 2010, 15, 560–573. [Google Scholar] [CrossRef]
- Fazio, J.M.; Barthel, T.; Freeman, E.W.; Garlick-Ott, K.; Scholle, A.; Brown, J.L. Utilizing camera traps, closed circuit cameras and behavior observation software to monitor activity budgets, habitat use, and social interactions of zoo-housed asian elephants (Elephus maximus). Animals 2020, 10, 1–21. [Google Scholar] [CrossRef]
- Ferretti, C.J.; Taylor, B.P.; Shinall, J.; Hollander, E. Psychiatric assessment and pharmacological treatment. In Autism Spectrum Disorders; Hollander, E., Hagerman, R.J., Fein, D., Eds.; American Psychiatric Publishing, 2018; pp. 111–141. [Google Scholar]
- Fineberg, N.A.; Chamberlain, S.R.; Goudriaan, A.E.; Stein, D.J.; Vanderschuren, L.J.M.J.; Gillan, C.M.; Shekar, S.; Gorwood, P.A.P.M.; Voon, V.; Morein-Zamir, S.; Denys, D.; Sahakian, B.J.; Moeller, F.G.; Robbins, T.W.; Potenza, M.N. New developments in human neurocognition: Clinical, genetic, and brain imaging correlates of impulsivity and compulsivity. CNS Spectr. 2014, 19, 69–89. [Google Scholar] [CrossRef]
- Fisher, L.; Ackland, T.; Carter, M.; Ellwood, A.-L.; Gilman, L.; Mainemer, H.; Morison, S.; Thompson, S.; Aitken, H.; Blamey, J.; Boecker, C.; Brown, K.; Carley, S.; Drew, R.; Dykman, L.; Krygsveld, S.; Lawes, P.; Mann, S.; Morum, B.; Mulloy, R.; Ogmundson, A.; Olsen, T.; Plante, K.; Rodrigues, L.; Rutherford, R.; Sayani, S.; Vigue, T.; Wang, C.; Wiebe, V.; Wong, S.; Ames, E.W.; Chisholm, K.; Savoie, L. Problems reported by parents of romanian orphans adopted to British Columbia. Int. J. Behav. Dev. 1997, 20, 67–82. [Google Scholar] [CrossRef]
- Flamand, A.; Robinet, L.; Raskin, A.; Braconnier, M.; Bouhamidi, A.; Derolez, G.; Lochin, C.; Helleu, C.; Petit, O. The social dimension of equine welfare: social contact positively affects the emotional state of stalled horses. Anim. Behav. 2025, 221, 123055. [Google Scholar] [CrossRef]
- Foley, J.P. First year development of a rhesus monkey (Macaca mulatta) reared in isolation. Pedagog. Semin. J. Genet. Psychol. 1934, 45, 39–105. [Google Scholar] [CrossRef]
- Fontenot, M.B.; Padgett, E.E.; Dupuy, A.M.; Lynch, C.R.; De Petrillo, P.B.; Higley, J.D. The effects of flouxetine and buspirone on self-injurious and stereotypic behavior in adult male rhesus macaques. Comp. Med. 2005, 55, 67–74. [Google Scholar]
- Forget, S. Treatment-resistant stereotypic behaviours in laboratory mice (Mus musculus): Investigating causes and behavioural correlates. In The University of Guelph; 2025. [Google Scholar]
- Fowler, S.C.; Covington, H.E.; Miczek, K.A. Stereotyped and complex motor routines expressed during cocaine self-administration: Results from a 24-h binge of unlimited cocaine access in rats. Psychopharmacology (Berl) 2007, 192, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Francis, S.H. The effects of own-home and institution-rearing on the behavioural development of normal and mongol children. J. Child Psychol. Psychiatry. 1971, 12, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Frank, D. Repetitive behaviors in cats and dogs: Are they really a sign of obsessive-compulsive disorders (OCD)? Can. Vet. J. 2013, 54, 129. [Google Scholar] [PubMed]
- Frank, D.; Bélanger, M.C.; Bécuwe-Bonnet, V.; Parent, J. Prospective medical evaluation of 7 dogs presented with fly biting. Can. Vet. J. 2012, 53, 1279–1284. [Google Scholar]
- Franklin, A.R.; Mathersul, D.C.; Raine, A.; Ruscio, A.M. Restlessness in generalized anxiety disorder: using actigraphy to measure physiological reactions to threat. Behav. Ther. 2021, 52, 734–744. [Google Scholar] [CrossRef]
- Friend, T.H. Behavior of picketed circus elephants. Appl. Anim. Behav. Sci. 1999, 62, 73–88. [Google Scholar] [CrossRef]
- Fritz, J.; Nash, L.T.; Alford, P.L.; Bowen, J.A. Abnormal behaviors, with a special focus on rocking, and reproductive competence in a large sample of captive chimpanzees (Pan troglodytes). Am. J. Primatol. 1992, 27, 161–176. [Google Scholar] [CrossRef]
- Fuld, S. Autism spectrum disorder: The impact of stressful and traumatic life events and implications for clinical practice. Clin. Soc. Work J. 2018, 46, 210–219. [Google Scholar] [CrossRef]
- Fureix, C.; Walker, M.; Harper, L.; Reynolds, K.; Saldivia-Woo, A.; Mason, G.J. Stereotypic behaviour in standard non-enriched cages is an alternative to depression-like responses in C57BL/6 mice. Behav. Brain Res. 2016, 305, 186–190. [Google Scholar] [CrossRef]
- Gage, L.J. Use of buspirone and enrichment to manage aberrant behavior in an American badger (Taxidea taxus). J. Zoo Wildl. Med. 2005, 36, 520–522. [Google Scholar] [CrossRef] [PubMed]
- Gagliano, A.; Cucinotta, F.; Giunta, I.; Di Modica, I.; De Domenico, C.; Costanza, C.; Germanò, E.; Frankovich, J. Immune/inflammatory underpinnings of neurodevelopmental disorders and Pediatric Acute-Onset Neuropsychiatric Syndrome: A scoping review. Int. J. Mol. Sci. 2025, 26, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Galeano, M.G.; Cantarelli, V.I.; Ruiz, R.D.; Fiol de Cuneo, M.; Ponzio, M.F. Reproductive performance and weaning success in fur-chewing chinchillas (Chinchilla lanigera). Reprod. Biol. 2014, 14, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Galeano, M.G.; Ruiz, R.D.; de Cuneo, M.F.; Ponzio, M.F. Effectiveness of fluoxetine to control fur-chewing behaviour in the chinchilla (Chinchilla lanigera). Appl. Anim. Behav. Sci. 2013, 146, 112–117. [Google Scholar] [CrossRef]
- García-Villamisar, D.; Rojahn, J. Comorbid psychopathology and stress mediate the relationship between autistic traits and repetitive behaviours in adults with autism. J. Intellect. Disabil. Res. 2015, 59, 116–124. [Google Scholar] [CrossRef]
- Garner, J.P. Perseveration and stereotypy – Systems-level insights from clinical psychology. In Stereotypic Animal Behaviour Funadamentals and Applications to Welfare; Mason, G.J., Rushen, J., Eds.; CAB International: Oxford, 2006; pp. 121–152. [Google Scholar]
- Gershuny, B.S.; Keuthen, N.J.; Gentes, E.L.; Russo, A.R.; Emmott, E.C.; Jameson, M.; Dougherty, D.D.; Loh, R.; Jenike, M.A. Current posttraumatic stress disorder and history of trauma in trichotillomania. J. Clin. Psychol. 2006, 62, 1521–1529. [Google Scholar] [CrossRef]
- Girotti, M.; Carreno, F.R.; Morilak, D.A. Role of orbitofrontal cortex and differential effects of acute and chronic stress on motor impulsivity measured with 1-choice serial reaction time test in male rats. Int. J. Neuropsychopharmacol. 2022, 25, 1026–1036. [Google Scholar] [CrossRef]
- Goeller, H.B.; Downey, B.C.; Tucker, C.B. Limit feeding total mixed rations exacerbates intersucking in year-old dairy heifers. J. Dairy Sci. 2023, 106, 9494–9506. [Google Scholar] [CrossRef]
- Goldberg, X.; Soriano-Mas, C.; Alonso, P.; Segalàs, C.; Real, E.; López-Solà, C.; Subirà, M.; Via, E.; Jiménez-Murcia, S.; Menchón, J.M.; Cardoner, N. Predictive value of familiality, stressful life events and gender on the course of obsessive-compulsive disorder. J. Affect. Disord. 2015, 185, 129–134. [Google Scholar] [CrossRef]
- Gonyou, H.W.; Develliers, N.; Faucitano, L.; Friendship, R.; Pasma, T.; Widowski, T.M.; Ringgenberg, N.; Possberg, Fl. Code of practice for the care & handling of pigs: review of scientific research on priority issues. NFACC Pig Code Pract. Sci. Comm. 2012. [Google Scholar]
- Gottlieb, D.H.; Capitanio, J.P.; Mccowan, B. Risk factors for stereotypic behavior and self-biting in rhesus macaques (Macaca mulatta): Animal’s history, current environment, and personality. Am. J. Primatol. 2013, 75, 995–1008. [Google Scholar] [CrossRef] [PubMed]
- Grados, M.; Prazak, M.; Saif, A.; Halls, A. A review of animal models of obsessive-compulsive disorder: A focus on developmental, immune, endocrine and behavioral models. Expert Opin. Drug Discov. 2016, 11, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Grams, V.; Bögelein, S.; Grashorn, M.A.; Bessei, W.; Bennewitz, J. Quantitative genetic analysis of traits related to fear and feather pecking in laying hens. Behav. Genet. 2014, 45, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.E. Trichotillomania (hair pulling disorder). Indian J. Psychiatry 2019, 61, S136–S139. [Google Scholar] [CrossRef]
- Grant, J.E.; Chamberlain, S.R. Prevalence of skin picking (excoriation) disorder. J. Psychiatr. Res. 2020, 130, 57–60. [Google Scholar] [CrossRef]
- Grant, J.E.; Leppink, E.; Chamberlain, S. Body focused repetitive behavior disorders and perceived stress: Clinical and cognitive associations. J. Obsessive. Compuls. Relat. Disord. 2015, 5, 82–86. [Google Scholar] [CrossRef]
- Grant, J.E.; Redden, S.A.; Leppink, E.W.; Chamberlain, S.R. Trichotillomania and co-occurring anxiety. Compr. Psychiatry 2017a, 72, 1–5. [Google Scholar] [CrossRef]
- Grant, J.E.; Redden, S.A.; Medeiros, G.C.; Odlaug, B.L.; Curley, E.E.; Tavares, H.; Keuthen, N.J. Trichotillomania and its clinical relationship to depression and anxiety. Int. J. Psychiatry Clin. Pract. 2017b, 21, 302–306. [Google Scholar] [CrossRef]
- Greco, B.J.; Meehan, C.L.; Hogan, J.N.; Leighty, K.A.; Mellen, J.; Mason, G.J.; Mench, J.A. The days and nights of zoo elephants: Using epidemiology to better understand stereotypic behavior of African elephants (Loxodonta africana) and Asian elephants (Elephas maximus) in North American zoos. PLoS One 2016, 11, e0144276. [Google Scholar] [CrossRef]
- Green, L.E.; Lewis, K.; Kimpton, A.; Nicol, C.J.; House, L. Cross-sectional study of the prevalence of feather pecking in laying hens in alternative systems and its associations with management and disease. Vet. Rec. 2000, 147, 233–238. [Google Scholar] [CrossRef]
- Gross, A.N.M.; Engel, A.K.J.; Würbel, H. Simply a nest? Effects of different enrichments on stereotypic and anxiety-related behaviour in mice. Appl. Anim. Behav. Sci. 2011, 134, 239–245. [Google Scholar] [CrossRef]
- Guay, P.J.; Iwaniuk, A.N. Captive breeding reduces brain volume in waterfowl (Anseriformes). Condor 2008, 110, 276–284. [Google Scholar] [CrossRef]
- Guido, C.A.; Loffredo, L.; Zicari, A.M.; Pavone, P.; Savasta, S.; Gagliano, A.; Brindisi, G.; Galardini, G.; Bertolini, A.; Spalice, A. The impact of the COVID-19 epidemic during the lockdown on children with the pediatric acute-onset neuropsychiatric syndrome (PANDAS/PANS): The importance of environmental factors on clinical conditions. Front. Neurol. 2021, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Haas, M.; Jakubovski, E.; Fremer, C.; Dietrich, A.; Hoekstra, P.J.; Jäger, B.; Müller-Vahl, K.R. Yale Global Tic Severity Scale (YGTSS): Psychometric quality of the gold standard for tic assessment based on the large-scale EMTICS study. Front. Psychiatry 2021, 12, 1–10. [Google Scholar] [CrossRef]
- Hadley, C.; Hadley, B.; Ephraim, S.; Yang, M.; Lewis, M.H. Spontaneous stereotypy and environmental enrichment in deer mice (Peromyscus maniculatus): Reversibility of experience. Appl. Anim. Behav. Sci. 2006, 97, 312–322. [Google Scholar] [CrossRef]
- Haines, J.; Williams, C.L.; Brain, K.L. The psychopathology of incarcerated self-mutilators. Can. J. Psychiatry 1995, 40, 514–522. [Google Scholar] [CrossRef]
- Hall, S.; Thorns, T.; Oliver, C. Structural and environmental characteristics of stereotyped behaviors. Am. J. Ment. Retard. 2003, 108, 391–402. [Google Scholar] [CrossRef]
- Haney, C. Mental health issues in long-term solitary and “supermax” confinement. Crime Delinq 2003, 49, 124–156. [Google Scholar] [CrossRef]
- Hansen, C.P.B.; Jeppesen, L.L. Short term behavioural consequences of denied access to environmental facilities in mink. Agric. Food Sci. Finl 2000, 9, 149–155. [Google Scholar] [CrossRef]
- Hansen, S.W.; Jensen, M.B. Quantitative evaluation of the motivation to access a running wheel or a water bath in farm mink. Appl. Anim. Behav. Sci. 2006, 98, 127–144. [Google Scholar] [CrossRef]
- Hansen, S.W.; Jeppesen, L.L. Temperament, stereotypies and anticipatory behaviour as measures of welfare in mink. Appl. Anim. Behav. Sci. 2005, 99, 172–182. [Google Scholar] [CrossRef]
- Hansen, S.W.; Malmkvist, J.; Palme, R.; Damgaard, B.M. Do double cages and access to occupational materials improve the welfare of farmed mink? Anim. Welf. 2007, 16, 63–76. [Google Scholar] [CrossRef]
- Happaney, K.; Zelazo, P.D. Resistance to extinction: A measure of orbitofrontal function suitable for children? Brain Cogn. 2004, 55, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Harding, E.J.; Paul, E.S.; Mendl, M. Cognitive bias and affective state. J. R. Soc. Med. 2004, 427, 312–312. [Google Scholar] [CrossRef]
- Harlow, H.F.; Harlow, M.K. Social deprivation in monkeys. Sci. Am. 1962, 207, 136–150. [Google Scholar] [CrossRef]
- Harper, L.; Choleris, E.; Ervin, K.; Fureix, C.; Reynolds, K.; Walker, M.; Mason, G.J. Stereotypic mice are aggressed by their cage-mates, and tend to be poor demonstrators in social learning tasks. Anim. Welf. 2015, 24, 463–473. [Google Scholar] [CrossRef]
- Harris, K.M.; Mark Mahone, E.; Singer, H.S. Nonautistic motor stereotypies: Clinical Features and longitudinal follow-up. Pediatr. Neurol. 2008, 38, 26–272. [Google Scholar] [CrossRef]
- Harvey, N.C.; Farabaugh, S.M.; Druker, B.B. Effects of early rearing experience on adult behavior and nesting in captive Hawaiian crows (Corvus hawaiiensis). Zoo Biol. 2002, 21, 59–75. [Google Scholar] [CrossRef]
- Hausberger, M.; Gautier, E.; Müller, C.; Jego, P. Lower learning abilities in stereotypic horses. Appl. Anim. Behav. Sci. 2007, 107, 299–306. [Google Scholar] [CrossRef]
- Hauzenberger, A.R.; Gebhardt-Henrich, S.G.; Steiger, A. The influence of bedding depth on behaviour in golden hamsters (Mesocricetus auratus). Appl. Anim. Behav. Sci. 2006, 100, 280–294. [Google Scholar] [CrossRef]
- Heathers, J.A.J.; Gilchrist, K.H.; Hegarty-Craver, M.; Grego, S.; Goodwin, M.S. An analysis of stereotypical motor movements and cardiovascular coupling in individuals on the autism spectrum. Biol. Psychol. 2019, 142, 90–99. [Google Scholar] [CrossRef]
- Hediger, H. Wild Animals in Captivity; Butterworth Scientific Publications: London, 1950. [Google Scholar]
- Hemmings, A.; McBride, S.D.; Hale, C.E. Perseverative responding and the aetiology of equine oral stereotypy. Appl. Anim. Behav. Sci. 2007, 104, 143–150. [Google Scholar] [CrossRef]
- Hemmings, A.J.; Parker, M.O.; Hale, C.; McBride, S.D. Causal and functional interpretation of mu-and delta-opioid receptor profiles in mesoaccumbens and nigrostriatal pathways of an oral stereotypy phenotype. Behav. Brain Res. 2018, 353, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Hoksbergen, R.; Ter Laak, J.; Rijk, K.; Van Dijkum, C.; Stoutjesdijk, F. Post-institutional autistic syndrome in Romanian adoptees. J. Autism Dev. Disord. 2005, 35, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Homer, B.; Judd, J.; Mohammadi Dehcheshmeh, M.; Ebrahimie, E.; Trott, D.J. Gut microbiota and behavioural issues in production, performance, and companion animals: A systematic review. Animals 2023, 13, 1–20. [Google Scholar] [CrossRef]
- Hoorweg, F.A.; Vermeer, H.M.; Pedersen, L.J.; Spoolder, H.A. Review on hunger induced behaviours: Aggression and stereotypies. Regul. 625 2017. [Google Scholar]
- Hopper, L.M.; Freeman, H.D.; Ross, S.R. Reconsidering coprophagy as an indicator of negative welfare for captive chimpanzees. Appl. Anim. Behav. Sci. 2016, 176, 112–119. [Google Scholar] [CrossRef]
- Horesh, N.; Shmuel-Baruch, S.; Farbstein, D.; Ruhrman, D.; Milshtein, N.B.A.; Fennig, S.; Apter, A.; Steinberg, T. Major and minor life events, personality and psychopathology in children with tourette syndrome. Psychiatry Res. 2018, 260, 1–9. [Google Scholar] [CrossRef]
- Houghton, D.C.; Mathew, A.S.; Twohig, M.P.; Saunders, S.M.; Franklin, M.E.; Compton, S.N.; Neal-Barnett, A.M.; Woods, D.W. Trauma and trichotillomania: A tenuous relationship. J. Obsessive. Compuls. Relat. Disord. 2016, 11, 91–95. [Google Scholar] [CrossRef]
- Houpt, K.A. Motivation for cribbing by horses. Anim. Welf. 2012, 21, 1–7. [Google Scholar] [CrossRef]
- Hu, S.; Wang, Y.; Wang, X.; Ji, Y.; Li, C.; Qiu, B. Transcriptomic profiles link corticostriatal microarchitecture to genetics of neurodevelopment and neuropsychiatric risks. Transl. Psychiatry 2025, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Huber-Eicher, B.; Audige, L.; Audigé, L. Analysis of risk factors for the occurrence of feather pecking in laying hen growers. Br. J. Pharmacol. 1999, 40, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Huber-Eicher, B.; Sebo, F. Reducing feather pecking when raising laying hen chicks in aviary systems. Appl. Anim. Behav. Sci. 2001, 73, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Huber-Eicher, B.; Wechsler, B. Feather pecking in domestic chicks: its relation to dustbathing and foraging. Anim. Behav. 1997, 54, 757–768. [Google Scholar] [CrossRef]
- Hubrecht, R.C.; Serpell, J.A.; Poole, T.B. Correlates of pen size and housing conditions on the behaviour of kennelled dogs. Appl. Anim. Behav. Sci. 1992, 34, 365–383. [Google Scholar] [CrossRef]
- Hugo, C.; Seier, J.; Mdhluli, C.; Daniels, W.; Harvey, B.H.; Toit, D.; Wolfe-Coote, S.; Nel, D.; Stein, D.J. Fluoxetine decreases stereotypic behavior in primates. Prog. Neuropsychopharmacol. Biol. Psychiatry 2003, 27, 639–643. [Google Scholar] [CrossRef]
- Humphreys, K.L.; Gleason, M.M.; Drury, S.S.; Miron, D.; Nelson, C.A.; Fox, N.A.; Zeanah, C.H. Effects of institutional rearing and foster care on psychopathology at age 12 years in Romania: Follow-up of an open, randomised controlled trial. Lancet Psychiatry 2015. [Google Scholar] [CrossRef]
- Hurst, J.L.; Barnard, C.J.; Nevison, C.M.; West, C.D. Housing and welfare in laboratory rats: The welfare implications of social isolation and social contact among females. Anim. Welf. 1998, 7, 121–136. [Google Scholar] [CrossRef]
- Hurst, J.L.; Barnard, C.J.; Nevison, C.M.; West, C.D. Housing and welfare in laboratory rats: welfare implications of isolation and social contact among caged males. Anim. Welf. 1997, 6, 329–347. [Google Scholar] [CrossRef]
- Hurtubise, J.L.; Howland, J.G. Effects of stress on behavioral flexibility in rodents. Neuroscience 2017, 345, 176–192. [Google Scholar] [CrossRef]
- Hwang, J.B.; Kim, J.S.; Ahn, B.H.; Jung, C.H.; Lee, Y.H.; Kam, S. Clonazepam treatment of pathologic childhood aerophagia with psychological stresses. J. Korean Med. Sci. 2007, 22, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Iffland, H.; Wellmann, R.; Bennewitz, J.; Preuß, S.; Tetens, J.; Bessei, W.; Piepho, H.-P. A novel model to explain extreme feather pecking behavior in laying hens. Behav. Genet. 2020, 50, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Ijichi, C.L.; Collins, L.M.; Elwood, R.W. Evidence for the role of personality in stereotypy predisposition. Anim. Behav. 2013, 85, 1145–1151. [Google Scholar] [CrossRef]
- Imthon, A.K.; Caldart, C.A.; Do Rosário, M.C.; Fontenelle, L.F.; Miguel, E.C.; Ferrão, Y.A. Stressful life events and the clinical expression of obsessive–compulsive disorder (Ocd): An exploratory study. J. Clin. Med. 2020, 9, 1–19. [Google Scholar] [CrossRef]
- Jacobson, S.L.; Ross, S.R.; Bloomsmith, M.A. Characterizing abnormal behavior in a large population of zoo-housed chimpanzees: Prevalence and potential influencing factors. PeerJ 2016, 2016, 1–14. [Google Scholar] [CrossRef]
- Jensen, P.; Keeling, L.; Schütz, K.; Andersson, L.; Mormède E, P.; Brändström, H.; Forkman, B.; Kerje, S.; Fredriksson, R.; Ohlsson, C.; Larsson, S.; Mallmin, H.; Kindmark, A. Feather pecking in chickens is genetically related to behavioural and developmental traits. Physiol. Behav. 2005, 86, 52–60. [Google Scholar] [CrossRef]
- Jeong, D.H.; Yang, J.J.; Yeon, S.C. Fluoxetine therapy to decrease stereotypic behavior in the asiatic black bear (ursus thibetanus). J. Zoo Wildl. Med. 2019, 50, 718–722. [Google Scholar] [CrossRef]
- Jeppesen, L.L.; Falkenberg, H. Effects of play balls on pelt-biting, behaviour, and level of stress in ranch mink. Scientifur 1990, 14, 179–186. [Google Scholar]
- Jeppesen, L.L.; Heller, K.E.; Bildsøe, M. Stereotypies in female farm mink (Mustela vison) may be genetically transmitted and associated with higher fertility due to effects on body weight. Appl. Anim. Behav. Sci. 2004, 86, 137–143. [Google Scholar] [CrossRef]
- Jeppesen, L.L.; Heller, K.E.; Dalsgaard, T. Effects of early weaning and housing conditions on the development of stereotypies in farmed mink. Appl. Anim. Behav. Sci. 2000, 68, 85–92. [Google Scholar] [CrossRef]
- Jiao, X.; Pang, K.C.H.; Beck, K.D.; Minor, T.R.; Servatius, R.J. Avoidance perseveration during extinction training in Wistar-Kyoto rats: An interaction of innate vulnerability and stressor intensity. Behav. Brain Res. 2011, 221, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Johansson, V.; Sandin, S. Repetitive behaviors and life-quality in adults with autism spectrum disorder. Eur. Psychiatry 2023, 66, S471–S472. [Google Scholar]
- Jones, M.A.; Mason, G.; Pillay, N. Early environmental enrichment protects captive-born striped mice against the later development of stereotypic behaviour. Appl. Anim. Behav. Sci. 2011a, 135, 138–145. [Google Scholar] [CrossRef]
- Jones, M.A.; Mason, G.; Pillay, N. Early social experience influences the development of stereotypic behaviour in captive-born striped mice Rhabdomys. Appl. Anim. Behav. Sci. 2010a, 123, 70–75. [Google Scholar] [CrossRef]
- Jones, M.A.; Mason, G.J.; Pillay, N. Correlates of birth origin effects on the development of stereotypic behaviour in striped mice. Rhabdomys. Anim. Behav. 2011b, 82, 149–159. [Google Scholar] [CrossRef]
- Jones, M.A.; Van Lierop, M.; Mason, G.J.; Pillay, N. Increased reproductive output in stereotypic captive Rhabdomys females: Potential implications for captive breeding. Appl. Anim. Behav. Sci. 2010b, 123, 63–69. [Google Scholar] [CrossRef]
- Joshi, S.; Pillay, N. Association between personality and stereotypic behaviours in the African striped mouse Rhabdomys dilectus. Appl. Anim. Behav. Sci. 2016, 174, 154–161. [Google Scholar] [CrossRef]
- Karlen, G.A.M.; Hemsworth, P.H.; Gonyou, H.W.; Fabrega, E.; David Strom, A.; Smits, R.J. The welfare of gestating sows in conventional stalls and large groups on deep litter. Appl. Anim. Behav. Sci. 2007, 105, 87–101. [Google Scholar] [CrossRef]
- Kaufman, M.E.; Levitt, H. Some determinants of stereotyped behaviours in institutionalized mental defectives. J. Intellect. Disabil. Res. 1965, 9, 201–209. [Google Scholar] [CrossRef]
- Keating, J.; Van Goozen, S.; Uljarevic, M.; Hay, D.; Leekam, S.R. Restricted and repetitive behaviors and their developmental and demographic correlates in 4–8-year-old children: A transdiagnostic approach. Front. Behav. Neurosci. 2023, 17, 1–15. [Google Scholar] [CrossRef]
- Keiper, R.R. Studies of stereotypy in the canary (Serinus canarius). Anim. Behav. 1970, 18, 353–357. [Google Scholar] [CrossRef]
- Keiper, Ronald R. Causal factors of stereotypies in caged birds. Anim. Behav 1969, 17, 1–4. [Google Scholar] [CrossRef]
- Keiper, Ronald R. Drug effects on canary stereotypies. Psychopharmacologia 1969, 16, 16–24. [Google Scholar] [CrossRef] [PubMed]
- King, R.A.; Scahill, L.; Vitulano, L.A.; Schwab-Stone, M.; Tercyak, K.P.; Riddle, M.A. Childhood trichotillomania: Clinical phenomenology, comorbidity, and family genetics. J. Am. Acad. Child Adolesc. Psychiatry 1995, 34, 1451–1459. [Google Scholar] [CrossRef] [PubMed]
- Kitchenham, L.; Mason, G.J. The neurobiology of environmentally induced stereotypic behaviours in captive animals: Assessing the basal ganglia pathways and cortico-striatal-thalamo-cortical circuitry hypotheses. Behaviour 2021, 158, 1251–1302. [Google Scholar] [CrossRef]
- Kjaer, J.B.; Guémené, D. Adrenal reactivity in lines of domestic fowl selected on feather pecking behavior. Physiol. Behav. 2008, 96, 370–373. [Google Scholar] [CrossRef]
- Kjaer, J.B.; Jørgensen, H. Heart rate variability in domestic chicken lines genetically selected on feather pecking behavior. Genes, Brain Behav. 2011, 10, 747–755. [Google Scholar] [CrossRef]
- Kłosowska, J.; Antosz-Rekucka, R.; Kałużna-Wielobób, A.; Prochwicz, K. Dissociative experiences mediate the relationship between traumatic life events and types of skin picking. Findings from non-clinical sample. Front. Psychiatry 2021, 12, 1–10. [Google Scholar] [CrossRef]
- Ko, C.H.; Lu, Y.C.; Lee, C.H.; Liao, Y.C. The influence of adverse childhood experiences and depression on addiction severity among methamphetamine users: exploring the role of perseveration. Front. Psychiatry 2024, 15, 1–12. [Google Scholar] [CrossRef]
- Koenen, K.C.; Ratanatharathorn, A.; Ng, L.; McLaughlin, K.A.; Bromet, E.J.; Stein, D.J.; Karam, E.G.; Meron Ruscio, A.; Benjet, C.; Scott, K.; Atwoli, L.; Petukhova, M.; Lim, C.C.W.; Aguilar-Gaxiola, S.; Al-Hamzawi, A.; Alonso, J.; Bunting, B.; Ciutan, M.; De Girolamo, G.; Degenhardt, L.; Gureje, O.; Haro, J.M.; Huang, Y.; Kawakami, N.; Lee, S.; Navarro-Mateu, F.; Pennell, B.E.; Piazza, M.; Sampson, N.; Ten Have, M.; Torres, Y.; Viana, M.C.; Williams, D.; Xavier, M.; Kessler, R.C. Posttraumatic stress disorder in the World Mental Health Surveys. Psychol. Med. 2017, 47, 2260–2274. [Google Scholar] [CrossRef]
- Koistinen, T.; Ahola, L.; Mononen, J. Blue foxes’ (Alopex lagopus) preferences between earth floor and wire mesh floor. Appl. Anim. Behav. Sci. 2008, 111, 38–53. [Google Scholar] [CrossRef]
- Koistinen, T.; Ahola, L.; Mononen, J. Blue foxes’ motivation for access to an earth floor measured by operant conditioning. Appl. Anim. Behav. Sci. 2007, 107, 328–341. [Google Scholar] [CrossRef]
- Koistinen, T.; Korhonen, H.T.; Hämäläinen, E.; Mononen, J. Blue foxes’ (Vulpes lagopus) motivation to gain access and interact with various resources. Appl. Anim. Behav. Sci. 2016, 176, 105–111. [Google Scholar] [CrossRef]
- Koolhaas, J.M.; Korte, S.M.; De Boer, S.F.; Van Der Vegt, B.J.; Van Reenen, C.G.; Hopster, H.; De Jong, I.C.; Ruis, M.A.W.; Blokhuis, H.J. Coping styles in animals: current status in behavior and stress-physiology. Neurosci. Biobehav. Rev. 1999, 23, 925–935. [Google Scholar] [CrossRef]
- Korff, S.; Stein, D.J.; Harvey, B.H. Cortico-striatal cyclic AMP-phosphodiesterase-4 signalling and stereotypy in the deer mouse: Attenuation after chronic fluoxetine treatment. Pharmacol. Biochem. Behav. 2009, 92, 514–520. [Google Scholar] [CrossRef]
- Korff, S.; Stein, D.J.; Harvey, B.H. Stereotypic behaviour in the deer mouse: Pharmacological validation and relevance for obsessive-compulsive disorder. Prog. Neuro-Psychopharmacoloy 2008, 32, 348–355. [Google Scholar] [CrossRef]
- Korte, S.M.; Beuving, G.; Ruesink, W.; Blokhuis, H.J. Plasma catecholamine and corticosterone levels during manual restraint in chicks from a high and low feather pecking line of laying hens. Physiol. Behav. 1997, 62, 437–441. [Google Scholar] [CrossRef]
- Kostal, L.; Savory, C.J.; Hughes, B.O. Diurnal and individual variation in behaviour of restricted-fed broiler breeders. Appl. Anim. Behav. Sci. 1992, 32, 361–374. [Google Scholar] [CrossRef]
- Kostal, L.; Savory, G.J. Behavioral responses of restricted-fed fowls to pharmacological manipulation of 5-HT and GABA receptor subtypes. Pharmacol. Biochem. Behav. 1996, 53, 995–1004. [Google Scholar] [CrossRef]
- Kronenberg, S.; Shouldice, M.; Bitnun, A.; Gill, P.; Levy, D.M.; Logan, W.; Pringsheim, T.; Sandor, P.; Yeh, E.A.; Wilbur, C. Frequency and impact of PANDAS/PANS diagnosis. Can. Pediatr. Surveill. Progr 2023. [Google Scholar]
- Kryukova, N. V.; Manukhova, D.A.; Slavina, M.D. Deviations in the development of captivity walrus (Odobenus rosmarus) skull. Biol. Bull. 2024, 51, 358–370. [Google Scholar] [CrossRef]
- La Bella, S.; Scorrano, G.; Rinaldi, M.; Di Ludovico, A.; Mainieri, F.; Attanasi, M.; Spalice, A.; Chiarelli, F.; Breda, L. Pediatric Autoimmune Neuropsychiatric Disorders Associated with Streptococcal Infections (PANDAS): Myth or reality? The state of the art on a controversial disease. Microorganisms 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Lacy, R.C.; Alaks, G.; Walsh, A.; Kozak, C.A. Evolution of Peromyscus leucopus mice in response to a captive environment. PLoS One 2013, 8, e72452. [Google Scholar] [CrossRef] [PubMed]
- Lam, K.S.L.; Aman, M.G. The repetitive behavior scale-revised: Independent validation in individuals with autism spectrum disorders. J. Autism Dev. Disord. 2007, 37, 855–866. [Google Scholar] [CrossRef]
- Lamoureux, L.; Beverley, J.A.; Marinelli, M.; Steiner, H. Fluoxetine potentiates methylphenidate-induced behavioral responses: Enhanced locomotion or stereotypies and facilitated acquisition of cocaine self-administration. Addict. Neurosci. 2023, 9, 100131. [Google Scholar] [CrossRef]
- Landau, D.; Iervolino, A.C.; Pertusa, A.; Santo, S.; Singh, S.; Mataix-Cols, D. Stressful life events and material deprivation in hoarding disorder. J. Anxiety Disord. 2011, 25, 192–202. [Google Scholar] [CrossRef]
- Langen, M.; Durston, S.; Kas, M.J.H.; van Engeland, H.; Staal, W.G. The neurobiology of repetitive behavior:...and men. Neurosci. Biobehav. Rev. 2011, 35, 356–365. [Google Scholar] [CrossRef]
- Lanzarini, E.; Pruccoli, J.; Grimandi, I.; Spadoni, C.; Angotti, M.; Pignataro, V.; Sacrato, L.; Franzoni, E.; Parmeggiani, A. Phonic and motor stereotypies in autism spectrum disorder: Video analysis and neurological characterization. Brain Sci. 2021, 11. [Google Scholar] [CrossRef]
- Latham, N.; Mason, G.J. Frustration and perseveration in stereotypic captive animals: Is a taste of enrichment worse than none at all? Behav. Brain Res. 2010, 211, 96–104. [Google Scholar] [CrossRef]
- Latham, N.; Mason, G.J. Maternal deprivation and the development of stereotypic behaviour. Appl. Anim. Behav. Sci. 2007, 110, 84–108. [Google Scholar] [CrossRef]
- Laurence, A.; Lumineau, S.; Calandreau, L.; Arnould, C.; Leterrier, C.; Boissy, A.; Houdelier, C. Short- and long-term effects of unpredictable repeated negative stimuli on Japanese quail’s fear of humans. PLoS One 2014, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, A.B.; Brown, S.M.; Bradford, B.M.; Mabbott, N.A.; Bombail, V.; Rutherford, K.M.D. Non-neuronal brain biology and its relevance to animal welfare. Neurosci. Biobehav. Rev. 2025, 173, 106136. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, A.B.; Illius, A.W. Methodology for measuring hunger and food needs using operant conditioning in the pig. Appl. Anim. Behav. Sci. 1989, 24, 273–285. [Google Scholar] [CrossRef]
- Lebelt, D.; Zanella, A.J.; Unshelmt, J. Physiological correlates associated with cribbing behaviour in horses: changes in thermal threshold, heart rate, plasma P-endorphin and serotonin. Equine Clin. Behavoiur 1998, 27, 21–27. [Google Scholar] [CrossRef]
- Leonardi, L.; Perna, C.; Bernabei, I.; Fiore, M.; Ma, M.; Frankovich, J.; Tarani, L.; Spalice, A. Pediatric Pediatric Acute-Onset Neuropsychiatric Syndrome (PANS) and Pediatric Autoimmune Neuropsychiatric Disorders Associated with Streptococcal Infections (PANDAS): Immunological features underpinning controversial entities. Children 2024, 11. [Google Scholar] [CrossRef]
- Leruste, H.; Brscic, M.; Cozzi, G.; Kemp, B.; Wolthuis-Fillerup, M.; Lensink, B.J.; Bokkers, E.A.M.; van Reenen, C.G. Prevalence and potential influencing factors of non-nutritive oral behaviors of veal calves on commercial farms. J. Dairy Sci. 2014, 97, 7021–7030. [Google Scholar] [CrossRef]
- Lewis, M.H. Stereotyped movement disorder. In Encycl. autism Spectr. Disord.; 2013. [Google Scholar]
- Lewis, M.H.; Gluck, J.P.; Petitto, J.M.; Hensley, L.L.; Ozer, H. Early social deprivation in nonhuman primates: Long-term effects on survival and cell-mediated immunity. Biol. Psychiatry 2000, 47, 119–126. [Google Scholar] [CrossRef]
- Lewis, M.H.; Lindenmaier, Z.; Boswell, K.; Edington, G.; King, M.A.; Muehlmann, A.M. Subthalamic nucleus pathology contributes to repetitive behavior expression and is reversed by environmental enrichment: Genes. Brain Behav. 2018, 17, e12468. [Google Scholar] [CrossRef]
- Lewis, M.H.; Presti, M.F.; Lewis, J.B.; Turner, C.A. The neurobiology of stereotypy I: Environmental Complexity. In Stereotypic Animal Behaviour Funadamentals and Applications to Welfare; Mason, G.J., Rushen, J., Eds.; CAB International: Oxford, 2006; pp. 190–226. [Google Scholar]
- Lewis, R.; Hurst, J. The assessment of bar chewing as an escape behaviour in laboratory mice. Anim. Welf. 2004, 13, 19–25. [Google Scholar] [CrossRef]
- Li, X.; Xu, F.; Xie, L.; Ji, Y.; Cheng, K.; Zhou, Q.; Wang, T.; Shively, C.; Wu, Q.; Gong, W.; Fang, L.; Zhan, Q.; Melgiri, N.D.; Xie, P. Depression-like behavioral phenotypes by social and social plus visual isolation in the adult female Macaca fascicularis. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Lin, H.; Katsovich, L.; Ghebremichael, M.; Findley, D.B.; Grantz, H.; Lombroso, P.J.; King, R.A.; Zhang, H.; Leckman, J.F. Psychosocial stress predicts future symptom severities in children and adolescents with Tourette syndrome and/or obsessive-compulsive disorder. J. Child Psychol. Psychiatry Allied Discip. 2007, 48, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Williams, K.A.; Katsovich, L.; Findley, D.B.; Grantz, H.; Lombroso, P.J.; King, R.A.; Bessen, D.E.; Johnson, D.; Kaplan, E.L.; Landeros-Weisenberger, A.; Zhang, H.; Leckman, J.F. Streptococcal upper respiratory tract infections and psychosocial stress predict future tic and obsessive-compulsive symptom severity in children and adolescents with Tourette syndrome and obsessive-compulsive disorder. Biol. Psychiatry 2010, 67, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Lindenmayer, J.P. The pathophysiology of agitation. J. Clin. Psychiatry 2000, 61, 5–10. [Google Scholar] [PubMed]
- Litz, B.T.; Keane, T.M.; Fisher, L.; Marx, B.; Monaco, V. Physical health complaints in combat-related post-traumatic stress disorder: A preliminary report. J. Trauma. Stress 1992, 5, 131–141. [Google Scholar] [CrossRef]
- Lochner, C.; Du Toit, P.L.; Zungu-Dirwayi, N.; Marais, A.; Van Kradenburg, J.; Seedat, S.; Niehaus, D.J.H.; Stein, D.J. Childhood trauma in obsessive-compulsive disorder, trichotillomania, and controls. Depress. Anxiety 2002, 15, 66–68. [Google Scholar] [CrossRef]
- Luchins, D.J.; Goldman, M.B.; Lieb, M.; Hanrahan, P. Repetitive behaviors in chronically institutionalized schizophrenic patients. Schizophr. Res. 1992, 8, 119–123. [Google Scholar] [CrossRef]
- Luescher, A.U. Diagnosis and management of compulsive disorders in dogs and cats. Clin. Tech. Small Anim. Pract. 2004, 19, 233–239. [Google Scholar] [CrossRef]
- Lumeij, J.T.; Hommers, C.J. Foraging “enrichment” as treatment for pterotillomania. Appl. Anim. Behav. Sci. 2008, 111, 85–94. [Google Scholar] [CrossRef]
- Lutz, C.; Marinus, L.; Chase, W.; Meyer, J.; Novak, M. Self-injurious behavior in male rhesus macaques does not reflect externally directed aggression. Physiol. Behav. 2003a, 78, 33–39. [Google Scholar] [CrossRef]
- Lutz, C.; Well, A.; Novak, M. Stereotypic and self-injurious behavior in rhesus macaques: A survey and retrospective analysis of environment and early experience. Am. J. Primatol. 2003b, 60, 1–15. [Google Scholar] [CrossRef]
- Lutz, C.K. Stereotypic behavior in nonhuman primates as a model for the human condition. ILAR J. 2014, 55, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Lutz, C.K.; Davis, E.B.; Ruggiero, A.M.; Suomi, S.J. Early predictors of self-biting in socially-housed rhesus macaques (Macaca mulatta). Am. J. Primatol. 2007, 69, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Maclean, K. The impact of institutionalization on child development. Dev. Psychopathol. 2003, 15, 853–884. [Google Scholar] [CrossRef] [PubMed]
- Maier, S.F.; Seligman, M.E. Learned helplessness: Theory and evidence. J. Exp. Psychol. Gen. 1976, 105, 3–46. [Google Scholar] [CrossRef]
- Maisch, H.; Vellinga, N.R.; Boersma, H. Best Practice Guideline Dhole (C. alpinus) (1. Edition). In EAZA Best Practice Guidelines; 2017. [Google Scholar]
- Malmkvist, J.; Brix, B.; Henningsen, K.; Wiborg, O. Hippocampal neurogenesis increase with stereotypic behavior in mink (Neovison vison). Behav. Brain Res. 2012, 229, 359–364. [Google Scholar] [CrossRef]
- Malmkvist, J.; Díez-León, M.; Christensen, J.W. Animals with various forms of abnormal behaviour differ in learning performance and use of enrichment: Evidence from farm mink. Appl. Anim. Behav. Sci. 2024, 271, 106167. [Google Scholar] [CrossRef]
- Maneeton, P.; Maneeton, B.; Kienngam, N.; Maneeton, N.; Winichaikul, Y.; Kawilapat, S. Updated systematic review and meta-analysis of randomized controlled trials: Antidepressants for restricted and repetitive behaviors in autism spectrum disorder. Neuropsychiatr. Dis. Treat. 2024, 20, 1711–1723. [Google Scholar] [CrossRef]
- Maraz, A.; Hende, B.; Bert Urban, R.; Demetrovics, Z. Pathological grooming: Evidence for a single factor behind trichotillomania, skin picking and nail biting. PLoS One 2017, 12, 1–13. [Google Scholar] [CrossRef]
- Martin, J. The Carrier State of Streptococcus pyogenes. In Basic Biology to Clinical Manifestations; Ferretti, J., Stevens, D., Fischetti, V., Eds.; University of Oklahoma Health Sciences Center: Oklahoma City, 2022; pp. 1–12. [Google Scholar]
- Martin, L.A.; Ashwood, P.; Braunschweig, D.; Cabanlit, M.; Van de Water, J.; Amaral, D.G. Stereotypies and hyperactivity in rhesus monkeys exposed to IgG from mothers of children with autism. Brain. Behav. Immun. 2008, 22, 806–816. [Google Scholar] [CrossRef]
- Martin, L.J.; Spicer, D.M.; Lewis, M.H.; Gluck, J.P.; Cork, L.C. Social deprivation of infant rhesus monkeys alters the chemoarchitecture of the brain: I. Subcortical regions. J. Neurosci. 1991, 11, 3344–3358. [Google Scholar] [CrossRef]
- Maslak, R.; Sergiel, A.; Hill, S.P. Some aspects of locomotory stereotypies in spectacled bears (Tremarctos ornatus) and changes in behavior after relocation and dental treatment. J. Vet. Behav. 2013, 8, 335–341. [Google Scholar] [CrossRef]
- Mason, G.; Mendl, M. Do the stereotypies of pigs, chickens and mink reflect adaptive species differences in the control of foraging? Appl. Anim. Behav. Sci. 1997, 53, 45–58. [Google Scholar] [CrossRef]
- Mason, G.J. Stereotypic behaviour in captive animals: Fundamentals and implications for welfare and beyond. In Stereotypic Animal Behaviour Fundamentals and Applications to Welfare; Mason, G.J., Rushen, J., Eds.; CABI: Wallingford, 2006; pp. 325–356. [Google Scholar]
- Mason, G.J. Tail-biting in mink (Mustela vison) is influenced by age at removal from the mother. Anim. Welf. 1994, 3, 305–311. [Google Scholar] [CrossRef]
- Mason, G.J. Age and context affect the stereotypies of caged mink. Behaviour 1993, 127, 191–229. [Google Scholar] [CrossRef]
- Mason, G.J. Stereotypies and suffering. Behav. Processes 1991, 25, 103–115. [Google Scholar] [CrossRef]
- Mason, G.J.; Clubb, R.; Latham, N.; Vickery, S. Why and how should we use environmental enrichment to tackle stereotypic behaviour? Appl. Anim. Behav. Sci. 2007, 102, 163–188. [Google Scholar] [CrossRef]
- Mason, G.J.; Cooper, J.; Clarebrough, C. Frustrations of fur-farmed mink. Nat. 2001, 410, 35–36. [Google Scholar] [CrossRef]
- Mason, G.J.; Latham, N. Can’t stop, won’t stop: Is stereotypy a reliable animal welfare indicator? Anim. Welf. 2004, 13, 57–69. [Google Scholar]
- Mason, G.J.; Mendl, M.T. Measuring the unmeasurable: The construct validation of affective state indicators Assessing Animal Welfare: A Guide to the Valid Use of Indicators of Affective States. In UFAW Animal Welfare Series; Mason, G.J., Nielsen, B.L., Mendl, M.T., Eds.; John Wiley & Sons Ltd.: Oxford, n.d. [Google Scholar]
- Mason, G.J.; Rushen, J. Stereotypic Animal Behaviour Funadamentals and Applications to Welfare, 2nd ed.; CABI: Wallingford, 2006. [Google Scholar]
- McAfee, L.M.; Mills, D.S.; Cooper, J.J. The use of mirrors for the control of stereotypic weaving behaviour in the stabled horse. Appl. Anim. Behav. Sci. 2002, 78, 159–173. [Google Scholar] [CrossRef]
- Mcbride, S.D.; Roberts, K.; Hemmings, A.J.; Ninomiya, S.; Parker, M.O. The impulsive horse: Comparing genetic, physiological and behavioral indicators to those of human addiction. Physiol. Behav. 2022, 254, 113896. [Google Scholar] [CrossRef]
- McConnachie, E.; Smid, A.M.C.; Thompson, A.J.; Weary, D.M.; Gaworski, M.A.; Von Keyserlingk, M.A.G. Cows are highly motivated to access a grooming substrate. Biol. Lett. 2018, 14, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Mcklveen, J.M.; Myers, B.; Herman, J.P. The medial prefrontal cortex: coordinator of autonomic, neuroendocrine and behavioural responses to stress. J. Neuroendocrinol. 2015, 27, 446–456. [Google Scholar] [CrossRef]
- Meagher, R.K.; Campbell, D.L.M.; Dallaire, J.A.; Díez-León, M.; Palme, R.; Mason, G.J. Sleeping tight or hiding in fright? The welfare implications of different subtypes of inactivity in mink. Appl. Anim. Behav. Sci. 2013, 144, 138–146. [Google Scholar] [CrossRef]
- Meagher, R.K.; Dallaire, A.A.; Campbell, J.L.M.; Ross, D.; Møller, M.H. Benefits of a ball and chain: Simple environmental enrichments improve welfare and reproductive success in farmed American mink (Neovison vison). PLoS One 2014, 9, 110589. [Google Scholar] [CrossRef] [PubMed]
- Meagher, Rebecca; Bechard, Allison; Palme, R.; Díez-Leó, M.; Bruce Hunter, D.; Mason, G.; Meagher, R; Bechard, A; Leo´n, D.-L. Decreased litter size in inactive female mink (Neovison vison): Mediating variables and implications for overall productivity. Can. J. Anim. Sci. 2012, 92, 131–141. [Google Scholar] [CrossRef]
- Meehan, C.L.; Garner, J.P.; Mench, J.A. Environmental enrichment and development of cage stereotypy in orange-winged amazon parrots (Amazona amazonica). Dev. Psychobiol. 2004, 44, 209–218. [Google Scholar] [CrossRef]
- Meehan, C.L.; Garner, J.P.; Mench, J.A. Isosexual pair housing improves the welfare of young Amazon parrots. Appl. Anim. Behav. Sci. 2003, 81, 73–88. [Google Scholar] [CrossRef]
- Meers, L.; Ödberg, F.O. Paradoxical rate-dependent effect of fluoxetine on captivity-induced stereotypies in bank voles. Prog. Neuropsychopharmacol. Biol. Psychiatry 2005, 29, 964–971. [Google Scholar] [CrossRef]
- Mehrabian, A.; Friedman, S.L. An analysis of fidgeting and associated individual differences. J. Pers. 1986, 54, 406–429. [Google Scholar] [CrossRef]
- Mehtar, M.; Mukaddes, N.M. Posttraumatic Stress Disorder in individuals with diagnosis of Autistic Spectrum Disorders. Res. Autism Spectr. Disord. 2011, 5, 539–546. [Google Scholar] [CrossRef]
- Mellor, E.L.; Mason, G.J. Feeding, mating and animal wellbeing: new insights from phylogenetic comparative methods. Proc. R. Soc. B Biol. Sci. 2023, 290, 20222571. [Google Scholar] [CrossRef] [PubMed]
- Mellor, E.L.; McDonald Kinkaid, H.K.; Mendl, M.T.; Cuthill, I.C.; Van Zeeland, Y.R.A.; Mason, G.J. Nature calls: Intelligence and natural foraging style predict poor welfare in captive parrots. Proc. R. Soc. B Biol. Sci. 2021, 288, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mendl, M.; Paul, E.S. Animal affect and decision-making. Neurosci. Biobehav. Rev. 2020, 112, 144–163. [Google Scholar] [CrossRef] [PubMed]
- Mendl, M.T.; Paul, E.S.; Mason, G.J. Animal welfare and affective states Assessing Animal Welfare: A Guide to the Valid Use of Indicators of Affective States. In UFAW Animal Welfare Series; Mason, G.J., Nielsen, B.L., Mendl, M.T., Eds.; John Wiley & Sons Ltd.: Oxford, n.d. [Google Scholar]
- Mertens, P.A. Pharmacological treatment of feather-picking in pet birds. In Proceedings of the First International Meeting Veterinary Behavioural Medicine; Mills, D.S., Heath, S.E., Harrington, L.J., Eds.; UFAW: Birmingham, 1997; pp. 209–211. [Google Scholar]
- Metcalf, J. A prisoner’s three tips for beating boredom [WWW Document]. Prison Writ. 2020. Available online: https://prisonwriters.com/boredom-in-prison/.
- Mikkola, S.; Salonen, M.; Hakanen, E.; Lohi, H. Fearfulness associates with problematic behaviors and poor socialization in cats. iScience 2022, 25, 105265. [Google Scholar] [CrossRef]
- Miller, J.M.; Singer, H.S.; Bridges, D.D.; Waranch, H.R. Behavioral therapy for treatment of stereotypic movements in nonautistic children. J. Child Neurol. 2006, 21, 119–125. [Google Scholar] [CrossRef]
- Miller, K.A.; Mench, J.A. The differential effects of four types of environmental enrichment on the activity budgets, fearfulness, and social proximity preference of Japanese quail. Appl. Anim. Behav. Sci. 2005, 95, 169–187. [Google Scholar] [CrossRef]
- Miller, L.J.; Ivy, J.A.; Vicino, G.A.; Schork, I.G. Impacts of natural history and exhibit factors on carnivore welfare. J. Appl. Anim. Welf. Sci. 2019, 22, 188–196. [Google Scholar] [CrossRef]
- Mills, D.S.; Luescher, A. Veterinary and pharmacological approaches to abnormal repetitive behaviour. In Stereotypic Animal Behaviour Fundamentals and Applications to Welfare; Mason, G.J., Rushen, J., Eds.; CABI: Wallingford, 2006; pp. 286–324. [Google Scholar]
- Mindus, C.; van Staaveren, N.; Bharwani, A.; Fuchs, D.; Gostner, J.M.; Kjaer, J.B.; Kunze, W.; Mian, M.F.; Shoveller, A.K.; Forsythe, P.; Harlander-Matauschek, A. Ingestion of Lactobacillus rhamnosus modulates chronic stress-induced feather pecking in chickens. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Monteiro, S.; Roque, S.; de Sá-Calçada, D.; Sousa, N.; Correia-Neves, M.; Cerqueira, J.J. An efficient chronic unpredictable stress protocol to induce stress-related responses in C57BL/6 mice. Front. Psychiatry 2015, 6, 1–11. [Google Scholar] [CrossRef]
- Moore, H.L.; Brice, S.; Powell, L.; Ingham, B.; Freeston, M.; Parr, J.R.; Rodgers, J. The mediating effects of alexithymia, intolerance of uncertainty, and anxiety on the relationship between sensory processing differences and restricted and repetitive behaviours in autistic adults. J. Autism Dev. Disord. 2022, 52, 4384–4396. [Google Scholar] [CrossRef]
- Mora, S.; Martín-González, E.; Flores, P.; Moreno, M. Neuropsychiatric consequences of childhood group A streptococcal infection: A systematic review of preclinical models. Brain. Behav. Immun. 2020, 86, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Morin, K.M. Carceral space: Prisoners and animals, First edit. ed, Antipode; Taylor and Francis, 2016. [Google Scholar] [CrossRef]
- Morrissey, K.L.H.; Widowski, T.; Leeson, S.; Sandilands, V.; Arnone, A.; Torrey, S. The effect of dietary alterations during rearing on growth, productivity, and behavior in broiler breeder females. Poult. Sci. 2014, 93, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Mosallanejad, B.; Varzi, H.N.; Avizeh, R.; Pourmahdi, M.; Khalili, F. Comparative evaluation between hypericin (hypiran) and fluoxetine in treatment of companion dogs with tail chasing. Vet. Res. Forum 2015, 6, 167–172. [Google Scholar] [PubMed]
- Mount, R.H.; Hastings, R.P.; Reilly, S.; Cass, H.; Charman, T. Behavioural and emotional features in Rett syndrome. Disabil. Rehabil. 2001, 23, 129–138. [Google Scholar] [CrossRef]
- Moy, S.S.; Riddick, N. V; Nikolova, V.D.; Teng, B.L.; Agster, K.L.; Nonneman, R.J.; Young, N.B.; Baker, L.K.; Nadler, J.J.; Bodfish, J.W. Repetitive behavior profile and supersensitivity to amphetamine in the C58/J mouse model of autism. Behav. Brain Res. 2014, 259, 200–214. [Google Scholar] [CrossRef]
- Muehlmann, A.M.; Lewis, M.H. Abnormal repetitive behaviours: Shared phenomenology and pathophysiology. J. Intellect. Disabil. Res. 2012, 56, 427–440. [Google Scholar] [CrossRef]
- Murayama, K.; Nakao, T.; Ohno, A.; Tsuruta, S.; Tomiyama, H.; Hasuzawa, S.; Mizobe, T.; Kato, K.; Kanba, S. Impacts of stressful life events and traumatic experiences on onset of obsessive-compulsive disorder. Front. Psychiatry 2020, 11, 1–6. [Google Scholar] [CrossRef]
- Murphy, T.K.; Snider, L.A.; Mutch, P.J.; Harden, E.; Zaytoun, A.; Edge, P.J.; Storch, E.A.; Yang, M.C.K.; Mann, G.; Goodman, W.K.; Swedo, S.E. Relationship of movements and behaviors to Group A Streptococcus infections in elementary school children. Biol. Psychiatry 2007, 61, 279–284. [Google Scholar] [CrossRef]
- Muthugovindan, D.; Singer, H. Motor stereotypy disorders. Curr. Opin. Neurol. 2009, 22, 131–136. [Google Scholar] [CrossRef]
- Nagy, K.; Bodó, G.; Bárdos, G.; Bánszky, N.; Kabai, P. Differences in temperament traits between crib-biting and control horses. Appl. Anim. Behav. Sci. 2010, 122, 41–47. [Google Scholar] [CrossRef]
- Nagy, K.; Bodó, G.; Bárdos, G.; Harnos, A.; Kabai, P. The effect of a feeding stress-test on the behaviour and heart rate variability of control and crib-biting horses (with or without inhibition). Appl. Anim. Behav. Sci. 2009, 121, 140–147. [Google Scholar] [CrossRef]
- Napolitano, F.; De Rosa, G.; Sevi, A. Welfare implications of artificial rearing and early weaning in sheep. Appl. Anim. Behav. Sci. 2008, 110, 58–72. [Google Scholar] [CrossRef]
- Nash, L.T.; Fritz, J.; Alford, P.A.; Brent, L. Variables influencing the origins of diverse abnormal behaviors in a large sample of captive chimpanzees (Pan troglodytes). Am. J. Primatol. 1999, 48, 15–29. [Google Scholar] [CrossRef]
- National Instititute of Mental Health. PANS and PANDAS: Questions and Answers [WWW Document]. U.S. Dep. Heal. Hum. Serv. Natl. Institutes Heal. 2025. Available online: https://www.nimh.nih.gov/health/publications/pandas.
- National Research Council (US) Committee on Recognition and Alleviation of Distress in Laboratory Animals. Recognition and alleviation of distress in laboratory animals; National Academies Press (US): Washington, DC, 2008. [Google Scholar] [CrossRef]
- NC3Rs. Behavioural indicators [WWW Document]. 2025. Available online: https://macaques.nc3rs.org.uk/welfare-assessment/behavioural-indicators.
- Nelson, C.A.; Fox, N.A.; Zeanah, C.H. Romania’s abandoned children: deprivation, brain development, and the struggle for recovery; Harvard University Press, 2014. [Google Scholar]
- Nettle, D.; Bateson, M. The evolutionary origins of mood and its disorders. Curr. Biol. 2012, 22, R712–R721. [Google Scholar] [CrossRef]
- Nielsen, S.S.; Alvarez, J.; Bicout, D.; Calistri, P.; Canali, E.; Drewe, J.; Garin-Bastuji, B.; Gonzales Rojas, J.; Schmidt, C.; Herskin, M.; Miranda Chueca, M.; Padalino, B.; Pasquali, P.; Roberts, H.; Spoolder, H.; Stahl, K.; Velarde, A.; Viltrop, A.; Winckler, C.; Tiemann, I.; de Jong, I.; Gebhardt-Henrich, S.; Keeling, L.; Riber, A.; Ashe, S.; Candiani, D.; Garc ́ıa Matas, R.; Hempen, M.; Mosbach-Schulz, O.; Rojo Gimeno, C.; Van der Stede, Y.; Vitali, M.; Bailly-Caumette, E.; Michel, V. Scientific opinion on the welfare of broilers on farm. EFSA J. 2023, 21, 7788. [Google Scholar] [CrossRef]
- Nierenberg, A.A.; Farabaugh, A.H.; Alpert, J.E.; Gordon; Worthington, Johanna; Rosenbaum, John J; Fava, Jerrold F M. Timing of onset of antidepressant response with fluoxetine treatment. Am. J. Psychiatry 2000, 157, 1423–1428. [Google Scholar] [CrossRef]
- Nimmo-Smith, V.; Heuvelman, H.; Dalman, C.; Lundberg, M.; Idring, S.; Carpenter, P.; Magnusson, C.; Rai, D. Anxiety disorders in adults with Autism Spectrum Disorder: A population-based study. J. Autism Dev. Disord. 2020, 50, 308–318. [Google Scholar] [CrossRef]
- Nip, E. The long-term effects of environmental enrichment on agonism in female C57BL/6, BALB/c, and DBA/2 mice; University of Guelph, 2018. [Google Scholar]
- Nip, E.; Adcock, A.; Nazal, B.; Maclellan, A.; Niel, L.; Choleris, E.; Levison, L.; Mason, G.J. Why are enriched mice nice? Investigating how environmental enrichment reduces agonism in female C57BL/6, DBA/2, and BALB/c mice. Appl. Anim. Behav. Sci. 2019, 217, 73–82. [Google Scholar] [CrossRef]
- Nordgreen, J.; Edwards, S.A.; Boyle, L.A.; Bolhuis, J.E.; Veit, C.; Sayyari, A.; Marin, D.E.; Dimitrov, I.; Janczak, A.M.; Valros, A. A proposed role for pro-inflammatory cytokines in damaging behavior in pigs. Front. Vet. Sci. 2020, 7, 1–15. [Google Scholar] [CrossRef]
- Norscia, I.; Collarini, E.; Cordoni, G. Anxiety behavior in pigs (Sus scrofa) decreases through affiliation and may anticipate threat. Front. Vet. Sci. 2021, 8, 1–14. [Google Scholar] [CrossRef]
- Novak, M.A. Self-injurious behavior in rhesus monkeys: New insights into its etiology, physiology, and treatment. Am. J. Primatol. 2003, 59, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Novak, M.A.; Meyer, J.S.; Lutz, C.; Gimpel, J.; Mason, G.J. The effects of enrichment in biomedical facilities: Some insights into their effects on laboratory primates’ stereotypies. In Stereotypic Animal Behaviour Funadamentals and Applications to Welfare; Mason, G., Rushen, J., Eds.; CABI: Wallingford, 2006; p. 266. [Google Scholar]
- O’Connor, V.L.; Vonk, J. Scaredy-cats don’t succeed: behavioral traits predict problem-solving success in captive felidae. PeerJ 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Oakley, C.; Mahone, E.M.; Morris-Berry, C.; Kline, T.; Singer, H.S. Primary complex motor stereotypies in older children and adolescents: Clinical features and longitudinal follow-up. Pediatr. Neurol. 2015, 52, 398–403.e1. [Google Scholar] [CrossRef] [PubMed]
- Oatley, K.; Johnson-Laird, P.N. Cognitive approaches to emotions. Trends Cogn. Sci. 2014, 18, 134–140. [Google Scholar] [CrossRef]
- Ödberg, F.O. The influence of cage size and environmental enrichment on the development of stereotypies in bank voles (Clethfionomys glareolus). Behav. Processes 1987, 14, 155–173. [Google Scholar] [CrossRef]
- Olsson, I.A.S.; Keeling, L.J. The push-door for measuring motivation in hens: Laying hens are motivated to perch at night. Anim. Welf. 2002, 11, 11–19. [Google Scholar] [CrossRef]
- Olsson, I.A.S.; Sherwin, C.M. Behaviour of laboratory mice in different housing conditions when allowed to self-administer an anxiolytic. Lab. Anim. 2006, 40, 392–399. [Google Scholar] [CrossRef]
- Özten, E.; Sayar, G.H.; Eryılmaz, G.; Kağan, G.; Işık, S.; Karamustafalıoğlu, O. The relationship of psychological trauma with trichotillomania and skin picking. Neuropsychiatr. Dis. Treat. 2015, 11, 1203–1210. [Google Scholar] [CrossRef]
- PANDAS Network Understanding PANDAS and PANS: A comprehensive resource guide [WWW Document]. 2025. Available online: https://pandasnetwork.org/understanding-pandas/.
- Park, R.M.; Schubach, K.M.; Cooke, R.F.; Herring, A.D.; Jennings, J.S.; Daigle, C.L. Impact of a cattle brush on feedlot steer behavior, productivity and stress physiology. Appl. Anim. Behav. Sci. 2020, 228, 104995. [Google Scholar] [CrossRef]
- Parker, M.; Goodwin, D.; Redhead, E.S. Survey of breeders’ management of horses in Europe, North America and Australia: Comparison of factors associated with the development of abnormal behaviour. Appl. Anim. Behav. Sci. 2008, 114, 206–215. [Google Scholar] [CrossRef]
- Patiño, J.J.; Vélez, S.A.; Martínez, J.R. Ethological, endocrinological, and gastroscopic evaluation of crib-biting Colombian creole horses. J. Vet. Behav. 2020, 40, 92–97. [Google Scholar] [CrossRef]
- Pedersen, L.J.; Jensen, M.B.; Hansen, S.W.; Munksgaard, L.; Ladewig, J.; Matthews, L. Social isolation affects the motivation to work for food and straw in pigs as measured by operant conditioning techniques. Appl. Anim. Behav. Sci. 2002, 77, 295–309. [Google Scholar] [CrossRef]
- Perré, Y.; Wauters, A.-M.; Richard-Yris, M.-A. Influence of mothering on emotional and social reactivity of domestic pullets. Appl. Anim. Behav. Sci. 2002, 75, 133–146. [Google Scholar] [CrossRef]
- Peterson, E.J.; Worlein, J.M.; Lee, G.H.; Dettmer, A.M.; Varner, E.K.; Novak, M.A. Rhesus macaques (Macaca mulatta) with self-injurious behavior show less behavioral anxiety during the human intruder test. Am. J. Primatol. 2017, 79, e22569. [Google Scholar] [CrossRef] [PubMed]
- Phemister, M.R.; Richardson, A.M.; Thomas, G. V. Observations of young normal and handicapped children. Child. Care. Health Dev. 1978, 4, 247–259. [Google Scholar] [CrossRef]
- Pineda, M. Treatment for anxiety and future treatments in rett syndrome. Biomed. J. Sci. Tech. Res. 2024, 54, 46496–46500. [Google Scholar] [CrossRef]
- Pingitore, G.; Chrobak, V.; Petrie, J. The social and psychological factors of bruxism. J. Prosthet. Dent. 1991, 65, 443–446. [Google Scholar] [CrossRef]
- Plato, S.M. Comparing the pathology of equine stereotypical behaviours to obsessive-compulsive and related disorder in humans: An exploratory Delphi study. Appl. Anim. Behav. Sci. 2022, 105571. [Google Scholar] [CrossRef]
- Poirier, C. Hippocampal structural plasticity Assessing Animal Welfare: A Guide to the Valid Use of Indicators of Affective States. In UFAW Animal Welfare Series; Mason, G.J., Nielsen, B.L., Mendl, M.T., Eds.; John Wiley & Sons Ltd.: Oxford, n.d. [Google Scholar]
- Poirier, C.; Bateson, M.; Gualtieri, F.; Armstrong, E.A.; Laws, G.C.; Boswell, T.; Smulders, T. V. Validation of hippocampal biomarkers of cumulative affective experience. Neurosci. Biobehav. Rev. 2019a, 101, 113–121. [Google Scholar] [CrossRef]
- Poirier, C.; Oliver, C.J.; Castellano Bueno, J.; Flecknell, P.; Bateson, M. Pacing behaviour in laboratory macaques is an unreliable indicator of acute stress. Sci. Reports 2019b, 2019 91 9, 1–8. [Google Scholar] [CrossRef]
- Polanco, A. Validating markers of cumulative experience in laboratory rhesus monkeys (Macaca mulatta); University of Guelph, 2021. [Google Scholar]
- Polanco, A.; Díez-León, M.; Mason, G.J. Stereotypic behaviours are heterogeneous in their triggers and treatments in the American mink, Neovison vison, a model carnivore. Anim. Behav. 2018, 141, 105–114. [Google Scholar] [CrossRef]
- Polverino, G.; Manciocco, A.; Vitale, A.; Alleva, E. Stereotypic behaviours in Melopsittacus undulatus: Behavioural consequences of social and spatial limitations. Appl. Anim. Behav. Sci. 2015, 165, 143–155. [Google Scholar] [CrossRef]
- Pomerantz, O.; Meiri, S.; Terkel, J. Socio-ecological factors correlate with levels of stereotypic behavior in zoo-housed primates. Behav. Processes 2013, 98, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Pomerantz, O.; Paukner, A.; Terkel, J. Some stereotypic behaviors in rhesus macaques (Macaca mulatta) are correlated with both perseveration and the ability to cope with acute stressors. Behav. Brain Res. 2012, 230, 274–280. [Google Scholar] [CrossRef]
- Ponzio, M.F.; Busso, J.M.; Ruiz, R.D.; de Cuneo, M.F. A survey assessment of the incidence of fur-chewing in commercial chinchilla (Chinchilla lanigera) farms. Anim. Welf. 2007, 16, 471–479. [Google Scholar] [CrossRef]
- Poroshinska, O.; Polishchuk, A.; Shmayun, S.; Kozii, N.; Shahanenko, R.; Chornozub, M.; Stovbetska, L.; Shahanenko, V.; Koziy, V. The use of anxiolytic drugs for the correction of behavioral disorders in mammals. Regul. Mech. Biosyst. 2024, 15, 42–48. [Google Scholar] [CrossRef]
- Poulsen, E.M.B.; Cambell Teskey, G. Pacing, prozac and a polar bear. In Stereotypic Animal Behaviour Fundamentals and Applications to Welfare; Mason, G.J., Rushen, J., Eds.; CABI: Wallingford, 2006; p. 309. [Google Scholar]
- Quest, K.M.; Byiers, B.J.; Payen, A.; Symons, F.J. Rett syndrome: A preliminary analysis of stereotypy, stress, and negative affect. Res. Dev. Disabil. 2014, 35, 1191–1197. [Google Scholar] [CrossRef]
- Rafaeli-Mor, N.; Foster, L.; Berkson, G. Self-reported body rocking and other habits in college students. Am. J. Ment. Retard. 1999, 104, 1–10. [Google Scholar] [CrossRef]
- Rapoport, J.L.; Ryland, D.H.; Kriete, M. Drug treatment of canine acral lick: An animal model of obsessive-compulsive disorder. Arch. Gen. Psychiatry 1992, 49, 517–521. [Google Scholar] [CrossRef]
- Redbo, I.; Emanuelson, M.; Oredsson, N.; Lundberg, K. Feeding level and oral stereotypies in dairy cows. Anim. Sci. 1996, 62, 199–206. [Google Scholar] [CrossRef]
- Redman, K.R. The effectiveness of dusting as a reinforcer for the chinchilla (Chinchilla lanigera); University of North Dakota, 1974. [Google Scholar]
- Robbins, T.W.; Gillan, C.M.; Smith, D.G.; de Wit, S.; Ersche, K.D. Neurocognitive endophenotypes of impulsivity and compulsivity: Towards dimensional psychiatry. Trends Cogn. Sci. 2012, 16, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Robert, S.; Bergeron, R.; Farmer, C.; Meunier-Salaün, M.C. Does the number of daily meals affect feeding motivation and behaviour of gilts fed high-fibre diets? Appl. Anim. Behav. Sci. 2002, 76, 105–117. [Google Scholar] [CrossRef]
- Robinson, T.E.; Berridge, K.C. The neural basis of drug craving: An incentive-sensitization theory of addiction. In Psychology of Learning; Shanks, D., Ed.; SAGE Publications Ltd, 2009. [Google Scholar]
- Rodenburg, T.B.; Buitenhuis, A.J.; Ask, B.; Uitdehaag, K.A.; Koene, P.; Van Der Poel, J.J.; Van Arendonk, J.A.M.; Bovenhuis, H. Genetic and phenotypic correlations between feather pecking and open-field response in laying hens at two different ages. Behav. Genet. 2004, 34, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Rodenburg, T.B.; Uitdehaag, K.A.; Ellen, E.D.; Komen, J. The effects of selection on low mortality and brooding by a mother hen on open-field response, feather pecking and cannibalism in laying hens. Anim. Welf. 2009, 18, 427–432. [Google Scholar] [CrossRef]
- Rodgers, J.; Glod, M.; Connolly, B.; McConachie, H. The relationship between anxiety and repetitive behaviours in autism spectrum disorder. J. Autism Dev. Disord. 2012, 42, 2404–2409. [Google Scholar] [CrossRef]
- Rolls, E.T. Emotion and Decision-Making Explained; Oxford University Press: Oxcford, 2014. [Google Scholar]
- Rommeck, I.; Anderson, K.; Heagerty, A.; Cameron, A.; McCowan, B. Risk factors and remediation of self-injurious and self-abuse behavior in rhesus macaques. J. Appl. Anim. Welf. Sci. 2009a, 12, 61–72. [Google Scholar] [CrossRef]
- Rommeck, I.; Gottlieb, D.H.; Strand, S.C.; McCowan, B. The effects of four nursery rearing strategies on infant behavioral development in rhesus macaques (Macaca mulatta). J. Am. Assoc. Lab. Anim. Sci. 2009b, 48, 395–401. [Google Scholar]
- Roper, T.J. Nest material and food as reinforcers for fixed-ratio responding in mice. Learn. Motiv. 1975, 6, 327–343. [Google Scholar] [CrossRef]
- Rozek, J.C.; Millam, J.R. Preference and motivation for different diet forms and their effect on motivation for a foraging enrichment in captive Orange-winged Amazon parrots (Amazona amazonica). Appl. Anim. Behav. Sci. 2011, 129, 153–161. [Google Scholar] [CrossRef]
- Rushen, J.; de Passillé, A.M. The motivation of non-nutritive sucking in calves, Bos taurus. Anim. Behav. 1995, 49, 1503–1510. [Google Scholar] [CrossRef]
- Rutter, M.; Kreppner, J.; Croft, C.; Murin, M.; Colvert, E.; Beckett, C.; Castle, J.; Sonuga-Barke, E. Early adolescent outcomes of institutionally deprived and non-deprived adoptees. III. Quasi-autism. J. Child Psychol. Psychiatry Allied Discip. 2007, 48, 1200–1207. [Google Scholar] [CrossRef] [PubMed]
- Sampogna, G.; Del Vecchio, V.; Giallonardo, V.; Luciano, M.; Fiorillo, A. Diagnosis, clinical features, and implications of agitated depression. Psychiatr. Clin. North Am. 2020, 43, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, E.O.; Bangasser, D.A. The effects of early life stress on impulsivity. Neurosci. Biobehav. Rev. 2022, 137. [Google Scholar] [CrossRef] [PubMed]
- Sanchis-Segura, C.; Spanagel, R. Behavioural assessment of drug reinforcement and addictive features in rodents: An overview. Addict. Biol. 2006, 11, 2–38. [Google Scholar] [CrossRef]
- Sargent, T.D.; Keiper, R.R. Stereotypies in caged canaries. Anim. Behav. 1967, 15, 62–66. [Google Scholar] [CrossRef]
- Sarkhel, S.; Praharaj, S.K.; Sinha, V.K. Role of life events in obsessive compulsive disorder. Isr. J. Psychiatry Relat. Sci. 2011, 48, 182–185. [Google Scholar] [CrossRef]
- Savory, C.J.; Mann, J.S. Stereotyped pecking after feeding by restricted-fed fowls is influenced by meal size. Appl. Anim. Behav. Sci. 1999, 62, 209–217. [Google Scholar] [CrossRef]
- Savory, C.J.; Maros, K.; Rutter, S.M. Assessment of hunger in growing broiler breeders in relation to a commercial restricted feeding programme. Anim. Welf. 1993, 2, 131–152. [Google Scholar] [CrossRef]
- Scantlebury, C.E.; Archer, D.C.; Proudman, C.J.; Pinchbeck, G.L. Management and horse-level risk factors for recurrent colic in the UK general equine practice population. Equine Vet. J. 2015, 47, 202–206. [Google Scholar] [CrossRef]
- Schino, G.; Perretta, G.; Taglioni, A.M.; Monaco, V.; Troisi, A. Primate displacement activities as an ethopharmacological model of anxiety. Anxiety 1996, 2, 186–191. [Google Scholar] [CrossRef]
- Schlussman, S.D.; Ho, A.; Zhou, Y.; Curtis, A.E.; Kreek, M.J. Effects of “binge” pattern cocaine on stereotypy and locomotor activity in C57BL/6J and 129/J mice. Pharmacol. Biochem. Behav. 1998, 60, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Schmid, R.; Doherr, M.G.; Steiger, A. The influence of the breeding method on the behaviour of adult African grey parrots (Psittacus erithacus). Appl. Anim. Behav. Sci. 2006, 98, 293–307. [Google Scholar] [CrossRef]
- Schneper, L.M.; Brooks-Gunn, J.; Notterman, D.A.; Suomi, S.J. Early-life experiences and telomere length in adult rhesus monkeys: An exploratory study. Psychosom. Med. 2016, 78, 1066–1071. [Google Scholar] [CrossRef] [PubMed]
- Schoenecker, B.; Heller, K.E. Stimulation of serotonin (5-HT) activity reduces spontaneous stereotypies in female but not in male bank voles (Clethrionomys glareolus) Stereotyping female voles as a new animal model for human anxiety and mood disorders? Appl. Anim. Behav. Sci. 2003, 80, 161–170. [Google Scholar] [CrossRef]
- Schoenecker, B.; Heller, K.E. The involvement of dopamine (DA) and serotonin (5-HT) in stress-induced stereotypies in bank voles (Clethrionomys glareolus). Appl. Anim. Behav. Sci. 2001, 73, 311–319. [Google Scholar] [CrossRef]
- Scholey, A.; Haskell, C.; Robertson, B.; Kennedy, D.; Milne, A.; Wetherell, M. Chewing gum alleviates negative mood and reduces cortisol during acute laboratory psychological stress. Physiol. Behav. 2009, 97, 304–312. [Google Scholar] [CrossRef]
- Schønecker, B. Increased survival and reproductive success associated with stereotypical behaviours in laboratory-bred bank voles (Clethrionomys glareolus). Appl. Anim. Behav. Sci. 2009, 121, 55–62. [Google Scholar] [CrossRef]
- Schouten, W.G.P.; Wiepkema, P.R. Coping styles of tethered sows. Behav. Processes 1991, 25, 125–132. [Google Scholar] [CrossRef]
- Schrock, J.M.; Sugiyama, L.S.; Snodgrass, J.J. Lassitude: A coordination system to support host immunity. Oxford Handb. Evol. Emot. 2024, 297–316. [Google Scholar] [CrossRef]
- Schweitzer, C.; Arnould, C. Emotional reactivity of Japanese quail chicks with high or low social motivation reared under unstable social conditions. Appl. Anim. Behav. Sci. 2010, 125, 143–150. [Google Scholar] [CrossRef]
- Sege, C.T.; Bradley, M.M.; Lang, P.J. Avoidance and escape: Defensive reactivity and trait anxiety. Behav. Res. Ther. 2018, 104, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Seibert, L.M. Pharmacotherapy for behavioral disorders in pet birds. J. Exot. Pet Med. 2007, 16, 30–37. [Google Scholar] [CrossRef]
- Seibert, L.M.; Crowell-Davis, S.L.; Wilson, G.H.; Ritchie, B.W. Placebo-controlled clomipramine trial for the treatment of feather picking disorder in cockatoos. J. Am. Anim. Hosp. Assoc. 2004, 40, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Selemon, L.D.; Friedman, H.R. Motor stereotypies and cognitive perseveration in non-human primates exposed to early gestational irradiation. Neuroscience 2013, 248, 213–224. [Google Scholar] [CrossRef]
- Šemrov, M.Z.; Patt, A. The tail as an iceberg indicator: interrelationships with welfare problems. In Tail Biting in Pigs: A Comprehensive Guide to Its Aetiology, Impact and Wider Significance in Pig Management; Driscoll, K., Valros, A., Eds.; Brill: Wageningen, the Netherlands, 2024; pp. 351–326. [Google Scholar]
- Seo, T.; Sato, S.; Kosaka, K.; Sakamoto, N.; Tokumoto, K. Tongue-playing and heart rate in calves. Appl. Anim. Behav. Sci. 1998, 58, 179–182. [Google Scholar] [CrossRef]
- Shaikh, N.; Leonard, E.; Martin, J.M. Prevalence of streptococcal pharyngitis and streptococcal carriage in children: a meta-analysis. Pediatrics 2010, 126, e557-564. [Google Scholar] [CrossRef]
- Sharma, N.P.; Huecker, M.R. Agitation [WWW Document]. StatPearls [Internet]. 2024. Available online: https://www.ncbi.nlm.nih.gov/books/NBK493153/.
- Sheridan, M.A.; Fox, N.A.; Zeanah, C.H.; McLaughlin, K.A.; Nelson, C.A. Variation in neural development as a result of exposure to institutionalization early in childhood. Proc. Natl. Acad. Sci. U. S. A 2012, 109, 12927–12932. [Google Scholar] [CrossRef]
- Sherwin, C.M. The use and perceived importance of three resources which provide caged laboratory mice the opportunity for extended locomotion. Appl. Anim. Behav. Sci. 1998, 55, 353–367. [Google Scholar] [CrossRef]
- Sherwin, C.M. Laboratory mice persist in gaining access to resources: A method of assessing the importance of environmental features. Appl. Anim. Behav. Sci. 1996, 48, 203–213. [Google Scholar] [CrossRef]
- Sherwin, C.M.; Nicol, C.J. Behavioural demand functions of caged laboratory mice for additional space. Anim. Behav. 1997, 53, 67–74. [Google Scholar] [CrossRef]
- Sherwin, C.M.; Nicol, C.J. Reorganization of behaviour in laboratory mice, Mus musculus, with varying cost of access to resources. Anim. Behav. 1996, 51, 1087–1093. [Google Scholar] [CrossRef]
- Shively, C.A.; Register, T.C.; Friedman, D.P.; Morgan, T.M.; Thompson, J.; Lanier, T. Social stress-associated depression in adult female cynomolgus monkeys (Macaca fascicularis). Biol. Psychol. 2005, 69, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Shukla, T.; Pandey, S. Stereotypies in adults: A systematic review. Neurol. Neurochir. Pol. 2020, 54, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Shulman, L.M.; Sanchez-Ramos, J.R.; Weiner, W.J. Defining features, clinical conditions, and theoretical constructs of stereotyped movements. In Stereotyped Movements - Brain & Behavior Relationships; Sprague, R.L., Newell, K.M., Eds.; American Psychiatric Association: Washington, DC, 1996; pp. 17–24. [Google Scholar]
- Shyne, A. Meta-analytic review of the effects of enrichment on stereotypic behavior in zoo mammals. Zoo Biol. 2006, 25, 317–337. [Google Scholar] [CrossRef]
- Shyne, A.; Block, M. The effects of husbandry training on stereotypic pacing in captive African wild dogs (Lycaon pictus). J. Appl. Anim. Welf. Sci. 2010, 13, 56–65. [Google Scholar] [CrossRef]
- Singer, H.S. Treatment of tics and Tourette syndrome. Curr. Treat. Options Neurol. 2010, 12, 539–561. [Google Scholar] [CrossRef]
- Singer, H.S. Motor stereotypies. Semin. Pediatr. Neurol. 2009, 16, 77–81. [Google Scholar] [CrossRef]
- Sisman, F.N.; Tok, O.; Ergun, A. The effect of psychological state and social support on nail-biting in adolescents: An exploratory study. Sch. Psychol. Int. 2017, 38, 304–318. [Google Scholar] [CrossRef]
- Smith, A.P. Chewing gum and stress reduction. J. Clin. Transl. Res. 2016, 2, 52–54. [Google Scholar] [CrossRef]
- Soares, J.M.; Sampaio, A.; Ferreira, L.M.; Santos, N.C.; Marques, F.; Palha, J.A.; Cerqueira, J.J.; Sousa, N. Stress-induced changes in human decision-making are reversible. Transl. Psychiatry 2012, 2. [Google Scholar] [CrossRef]
- Sohel, A.J.; Shutter, M.C.; Patel, P.; Molla, M. Fluoxetine [WWW Document]. StatPearls [Internet]. 2025. Available online: https://www.ncbi.nlm.nih.gov/books/NBK459223/.
- Sonuga-Barke, E.; Kennedy, M.; Golm, D.; Knights, N.; Kovshoff, H.; Kreppner, J.; Kumsta, R.; Maughan, B.; O’Connor, T.G.; Schlotz, W. Adoptees’ responses to separation from, and reunion with, their adoptive parent at age 4 years is associated with long-term persistence of autism symptoms following early severe institutional deprivation. Dev. Psychopathol. 2020, 32, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Soussignan, R.; Koch, P. Rhythmical stereotypies (leg-swinging) associated with reductions in heart-rate in normal school children. Biol. Psychol. 1985, 21, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Spann, M.N.; Mayes, L.C.; Kalmar, J.H.; Guiney, J.; Womer, F.Y.; Pittman, B.; Mazure, C.M.; Sinha, R.; Blumberg, H.P. Childhood abuse and neglect and cognitive flexibility in adolescents. Child Neuropsychol. 2012, 18, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, C.; Lübke, L.; Müller, S.; Lindstädt, T.; Gallinat, C. Childhood maltreatment, traumatic experiences, and posttraumatic stress disorder in pathological skin picking: An online case-control study. Gen. Hosp. Psychiatry 2022, 78, 9–13. [Google Scholar] [CrossRef]
- Spoolder, H.A.M.; Burbidge, J.A.; Edwards, S.A.; Howard Simmins, P.; Lawrence, A.B. Provision of straw as a foraging substrate reduces the development of excessive chain and bar manipulation in food restricted sows. Appl. Anim. Behav. Sci. 1995, 43, 249–262. [Google Scholar] [CrossRef]
- Srinivasan, J.; Mink, J.W. Stereotypies and other developmental hyperkinesias. In Hyperkinetic Movement Disorders: Differential Diagnosis and Treatment, 1st Ed.; Albanese, A., Jankovic, J., Eds.; Blackwell Publishing Ltd., 2012; pp. 353–362. [Google Scholar]
- Stein, D.J. Obsessive-compulsive disorder. Lancet 2002, 360, 397–405. [Google Scholar] [CrossRef]
- Stein, D.J.; Flessner, C.A.; Franklin, M.; Keuthen, N.J.; Lochner, C.; Woods, D.W. Is trichotillomania a stereotypic movement disorder? An analysis of body-focused repetitive behaviors in people with hair-pulling. Ann. Clin. Psychiatry 2008, 20, 194–198. [Google Scholar] [CrossRef]
- Stein, D.J.; Ruscio, A.M.; Altwaijri, Y.; Chiu, W.T.; Sampson, N.A.; Gaxiola, S.A.; Al Hamzawi, A.; Alonso, J.; Chardoul, S.; Gureje, O. Obsessive-compulsive disorder in the World Mental Health surveys. BMC Med. 2025, 23, 416. [Google Scholar] [CrossRef]
- Steinberg, T.; Shmuel-Baruch, S.; Horesh, N.; Apter, A. Life events and Tourette syndrome. Compr. Psychiatry 2013, 54, 467–473. [Google Scholar] [CrossRef]
- Sullivan, J.; Rodgers, J.; Lidstone, J.; Uljarevic, M.; Mcconachie, H.; Freeston, M.; Le, A.; Prior, M.; Leekam, S. Relations among restricted and repetitive behaviors, anxiety and sensory features in children with autism spectrum disorders. Res. Autism Spectr. Disord. 2014, 8, 82–92. [Google Scholar] [CrossRef]
- Suñol, M.; Contreras-Rodríguez, O.; Macià, D.; Martínez-Vilavella, G.; Martínez-Zalacaín, I.; Subirà, M.; Pujol, J.; Sunyer, J.; Soriano-Mas, C. Brain structural correlates of subclinical obsessive-compulsive symptoms in healthy children. J. Am. Acad. Child Adolesc. Psychiatry 2018, 57, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Svendsen, P.M.; Krogh Hansen, B.; Malmkvist, J.; Hansen, S.W.; Palme, R.; Lau Jeppesen, L. Selection against stereotypic behaviour may have contradictory consequences for the welfare of farm mink (Mustela vison). Appl. Anim. Behav. Sci. 2007, 107, 110–119. [Google Scholar] [CrossRef]
- Swaisgood, R.; Shepherdson, D. Environmental enrichment as a strategy for mitigating stereotypies in zoo animals: A literature review and meta-analysis. In Stereotypic Animal Behaviour Fundamentals and Applications to Welfare; Mason, G.J., Rushen, J., Eds.; CAB International: Oxford, 2006; pp. 256–285. [Google Scholar]
- Swaisgood, R.R.; Shepherdson, D.J. Scientific approaches to enrichment and stereotypies in zoo animals: What’s been done and where should we go next? Zoo Biol. 2005, 24, 499–518. [Google Scholar] [CrossRef]
- Swedo, S.; Menendez, C.M.; Cunningham, M.W. Pediatric Autoimmune Neuropsychiatric Disorder Associated with Streptococcal Infectionn (PANDAS): A case series. In Streptococcus Pyogenes: Basic Biology to Clinical Manifestations; Ferretti, J.J., Stevens, D.L., Fischetti, V.A., Eds.; University of Oklahoma Health Sciences Center: Oklahoma City, 2024; pp. 75–79. [Google Scholar] [CrossRef]
- Swedo, S.E. From research subgroup to clinical syndrome: Modifying the PANDAS criteria to describe PANS (Pediatric Acute-Onset Neuropsychiatric Syndrome). Pediatr. Ther. 02 2012. [Google Scholar] [CrossRef]
- Tan, C.Y.; Chiu, N.C.; Zeng, Y.H.; Huang, J.Y.; Tzang, R.F.; Chen, H.J.; Lin, Y.J.; Sun, F.J.; Ho, C.S. Psychosocial stress in children with Tourette syndrome and chronic tic disorder. Pediatr. Neonatol. 2024, 65, 336–340. [Google Scholar] [CrossRef]
- Tatemoto, P.; Broom, D.M.; Zanella, A.J. Changes in stereotypies: Effects over time and over generations. Anim. 2022, 12, 2504. [Google Scholar] [CrossRef]
- Taylor, S.B.; Anglin, J.M.; Paode, P.R.; Riggert, A.G.; Olive, M.F.; Conrad, C.D. Chronic stress may facilitate the recruitment of habit- and addiction-related neurocircuitries through neuronal restructuring of the striatum. Neuroscience 2014, 280, 231–242. [Google Scholar] [CrossRef]
- Terlouw, E.M.C.; Lawrence, A.B. Long-term effects of food allowance and housing on development of stereotypies in pigs. Appl. Anim. Behav. Sci. 1993, 38, 103–126. [Google Scholar] [CrossRef]
- Terlouw, E.M.C.; Wiersma, A.; Lawrence, A.B.; Macleod, H.A. Ingestion of food facilitates the performance of stereotypies in sows. Anim. Behav. 1993. [Google Scholar] [CrossRef]
- The assessment of pain and distress in animals. In Appendix III in report of the technical committee to enquire into the welfare of animals kept under intensive husbandry conditions; Thrope, W.H., Brambell, F.W.R., Eds.; London, 1965. [Google Scholar]
- Tiego, J.; Chamberlain, S.R.; Harrison, B.J.; Dawson, A.; Albertella, L.; Youssef, G.J.; Fontenelle, L.F.; Yücel, M. Heritability of overlapping impulsivity and compulsivity dimensional phenotypes. Sci. Rep. 2020, 10, 1–17. [Google Scholar] [CrossRef]
- Tiira, K.; Hakosalo, O.; Kareinen, L.; Thomas, A.; Hielm-Björkman, A.; Escriou, C.; Arnold, P.; Lohi, H. Environmental effects on compulsive tail chasing in dogs. PLoS One 2012, 7, e41684–e41684. [Google Scholar] [CrossRef] [PubMed]
- Tilly, S.L.C.; Dallaire, J.; Mason, G.J. Middle-aged mice with enrichment-resistant stereotypic behaviour show reduced motivation for enrichment. Anim. Behav. 2010, 80, 363–373. [Google Scholar] [CrossRef]
- Tottenham, N.; Hare, T.A.; Quinn, B.T.; McCarry, T.W.; Nurse, M.; Gilhooly, T.; Millner, A.; Galvan, A.; Davidson, M.C.; Eigsti, I.M.; Thomas, K.M.; Freed, P.J.; Booma, E.S.; Gunnar, M.R.; Altemus, M.; Aronson, J.; Casey, B.J. Prolonged institutional rearing is associated with atypically large amygdala volume and difficulties in emotion regulation. Dev. Sci. 2010, 13, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Tröster, H. Prevalence and functions of stereotyped behaviors in nonhandicapped children in residential care. J. Abnorm. Child Psychol. 1994, 22, 79–97. [Google Scholar] [CrossRef]
- Tynes, V. V; Sinn, L. Abnormal repetitive behaviors in dogs and cats: A guide for practitioners. Vet. Clin. North Am. Small Anim. Pract. 2014, 44, 543–564. [Google Scholar] [CrossRef]
- Uljarević, M.; Evans, D.W. Relationship between repetitive behaviour and fear across normative development, autism spectrum disorder, and down syndrome. Autism Res. 2017, 10, 502–507. [Google Scholar] [CrossRef]
- van der Eijk, J.A.J.; Lammers, A.; Kjaer, J.B.; Rodenburg, T.B. Stress response, peripheral serotonin and natural antibodies in feather pecking genotypes and phenotypes and their relation with coping style. Physiol. Behav. 2019, 199, 1–10. [Google Scholar] [CrossRef]
- van der Staay, F.J.; Schuurman, T.; Hulst, M.; Smits, M.; Prickaerts, J.; Kenis, G.; Korte, S.M. Effects of chronic stress: A comparison between tethered and loose sows. Physiol. Behav. 2010, 100, 154–164. [Google Scholar] [CrossRef]
- Van Moffaert, M. Clinical features and drug treatment of psychodermatological disorders. J. Eur. Acad. Dermatology Venereol. 1995, 5, S36. [Google Scholar] [CrossRef]
- van Staaveren, N.; Ellis, J.; Baes, C.F.; Harlander-Matauschek, A. A meta-analysis on the effect of environmental enrichment on feather pecking and feather damage in laying hens. Poult. Sci. 2021, 100, 397–411. [Google Scholar] [CrossRef]
- van Zeeland, Y. Medication for behavior modification in birds. Vet. Clin. North Am. - Exot. Anim. Pract. 2018, 21, 115–149. [Google Scholar] [CrossRef] [PubMed]
- Van Zeeland, Y.R.A.; Spruit, B.M.; Rodenburg, T.B.; Riedstra, B.; Van Hierden, Y.M.; Buitenhuis, B.; Mechiel Korte, S.; Lumeij, J.T. Feather damaging behaviour in parrots: A review with consideration of comparative aspects. Appl. Anim. Behav. Sci. 2009, 121, 75–95. [Google Scholar] [CrossRef]
- Vandeleest, J.J.; Mccowan, B.; Capitanio, J.P. Early rearing interacts with temperament and housing to influence the risk for motor stereotypy in rhesus monkeys (Macaca mulatta). Appl. Anim. Behav. Sci. 2011, 132, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Veissier, I.; De Passillé, A.M.; Després, G.; Rushen, J.; Charpentier, I.; Ramirez De La Fe, A.R.; Pradel, P. Does nutritive and non-nutritive sucking reduce other oral behaviors and stimulate rest in calves? J. Anim. Sci. 2002, 80, 2574–2587. [Google Scholar] [CrossRef]
- Vidal-Ribas, P.; Stringaris, A.; Rück, C.; Serlachius, E.; Lichtenstein, P.; Mataix-Cols, D. Are stressful life events causally related to the severity of obsessive-compulsive symptoms? A monozygotic twin difference study. Eur. Psychiatry 2015, 30, 309–316. [Google Scholar] [CrossRef]
- Vinke, C.M.; Hansen, S.W.; Mononen, J.; Korhonen, H.; Cooper, J.J.; Mohaibes, M.; Bakken, M.; Spruijt, B.M. To swim or not to swim: An interpretation of farmed mink’s motivation for a water bath. Appl. Anim. Behav. Sci. 2008, 111, 1–27. [Google Scholar] [CrossRef]
- Visser, E.K.; Ellis, A.D.; Van Reenen, C.G. The effect of two different housing conditions on the welfare of young horses stabled for the first time. Appl. Anim. Behav. Sci. 2008, 114, 521–533. [Google Scholar] [CrossRef]
- Vivar, J. Dispatches from the Canadian carceral state: The truth about provincial prisons. J. Prison. Prison. 2014, 23, 6–9. [Google Scholar] [CrossRef]
- Waiblinger, E.; König, B. Refinement of gerbil housing and husbandry in the laboratory. Altern. to Lab. Anim. 2004, 32, 163–169. [Google Scholar] [CrossRef]
- Waitt, C.; Buchanan-Smith, H.M. What time is feeding? How delays and anticipation of feeding schedules affect stump-tailed macaque behavior. Appl. Anim. Behav. Sci. 2001, 75, 75–85. [Google Scholar] [CrossRef]
- Wald, E.R.; Eickhoff, J.; Flood, G.E.; Heinz, M. V.; Liu, D.; Agrawal, A.; Morse, R.P.; Raney, V.M.; Veerapandiyan, A.; Madan, J.C. Estimate of the incidence of PANDAS and PANS in 3 primary care populations. Front. Pediatr. 2023, 11, 1–8. [Google Scholar] [CrossRef]
- Wales, L.; Charman, T.; Mount, R.H. An analogue assessment of repetitive hand behaviours in girls and young women with Rett syndrome. J. Intellect. Disabil. Res. 2004, 48, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.; Mason, G.J. A comparison of two types of running wheel in terms of mouse preference, health, and welfare. Physiol. Behav. 2018, 191, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.D.; Duggan, G.; Roulston, N.; Van Slack, A.; Mason, G.J. Negative affective states and their effects on morbidity, mortality and longevity. Anim. Welf. 2012, 21, 497–509. [Google Scholar] [CrossRef]
- Wallgren, T.; Lundeheim, N.; Wallenbeck, A.; Westin, R.; Gunnarsson, S. Rearing pigs with intact tails-experiences and practical solutions in Sweden. Animals 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Wallgren, T.; Westin, R.; Gunnarsson, S. A survey of straw use and tail biting in Swedish pig farms rearing undocked pigs. Acta Vet. Scand. 2016, 58, 1–11. [Google Scholar] [CrossRef]
- Wang, C.; Li, Y.; Wang, H.; Li, M.; Rong, J.; Liao, X.; Wu, Y.; Wang, Y. Differences in peripheral and central metabolites and gut microbiome of laying hens with different feather-pecking phenotypes. Front. Microbiol. 2023, 14, 1–14. [Google Scholar] [CrossRef]
- Wang, M.; Ricardi, R.; Ritfeld, G.J. PANDAS, a series of difficult decisions: a case report. BMC Psychiatry 2024, 24. [Google Scholar] [CrossRef]
- Warniment, A.; Brent, L. Abnormal behavior in a captive chimpanzee colony. Anim. Welf. Inst. 1996, 10–13. [Google Scholar]
- Weary, D.M.; Appleby, M.C.; Fraser, D. Responses of piglets to early separation from the sow. Appl. Anim. Behav. Sci. 1999, 63, 289–300. [Google Scholar] [CrossRef]
- Webb, L.E.; Van Reenen, C.G.; Engel, B.; Berends, H.; Gerrits, W.J.J.; Bokkers, E.A.M. Understanding oral stereotypies in calves: Alternative strategies, hypothalamic-pituitary-adrenal axis (re)activity and gene by environment interactions. Animal 2017, 11, 1054–1062. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, C.M.; Curry-Pochy, L.S.; Shafer, R.; Rudy, J.; Lewis, M.H. Reversal learning in C58 mice: Modeling higher order repetitive behavior. Behav. Brain Res. 2017, 332, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Wickens, C.L. Investigation of specific stereotypic behaviors in horses; Michigan State University, 2009. [Google Scholar]
- Widowski, T.M.; Torrey, S.; Bench, C.J.; Gonyou, H.W. Development of ingestive behaviour and the relationship to belly nosing in early-weaned piglets. Appl. Anim. Behav. Sci. 2008, 110, 109–127. [Google Scholar] [CrossRef]
- Wiedenmayer, C. Causation of the ontogenetic development of stereotypic digging in gerbils. Anim. Behav. 1997, 53, 461–470. [Google Scholar] [CrossRef]
- Wigham, S.; Rodgers, J.; South, M.; McConachie, H.; Freeston, M. The interplay between sensory processing abnormalities, intolerance of uncertainty, anxiety and restricted and repetitive behaviours in autism spectrum disorder. J. Autism Dev. Disord. 2015, 45, 943–952. [Google Scholar] [CrossRef]
- Wilkes, B.J.; Bass, C.; Korah, H.; Febo, M.; Lewis, M.H. Volumetric magnetic resonance and diffusion tensor imaging of C58/J mice: Neural correlates of repetitive behavior. Brain Imaging Behav. 2019, 14, 2084–2096. [Google Scholar] [CrossRef]
- Wilkes, B.J.; Lewis, M.H. The neural circuitry of restricted repetitive behavior: Magnetic resonance imaging in neurodevelopmental disorders and animal models. Neurosci. Biobehav. Rev. 2018, 92, 152–171. [Google Scholar] [CrossRef]
- Willard, S.L.; Shively, C.A. Modeling depression in adult female cynomolgus monkeys (Macaca fascicularis). Am. J. Primatol. 2012, 74, 528–542. [Google Scholar] [CrossRef]
- Williams, I.; Hoppitt, W.; Grant, R. The effect of auditory enrichment, rearing method and social environment on the behavior of zoo-housed psittacines (Aves: Psittaciformes); implications for welfare. Appl. Anim. Behav. Sci. 2017, 186, 85–92. [Google Scholar] [CrossRef]
- Williams, K.; Wheeler, D.M.; Silove, N.; Hazell, P. Cochrane review: Selective serotonin reuptake inhibitors (SSRIs) for autism spectrum disorders (ASD). Evidence-Based Child Heal. 2011, 6, 1044–1078. [Google Scholar] [CrossRef]
- Willner, P. The chronic mild stress (CMS) model of depression: History, evaluation and usage. Neurobiol. Stress 2017, 6, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.; Gattuso, J.J.; Hannan, A.J.; Renoir, T. Mechanisms of pathogenesis and environmental moderators in preclinical models of compulsive-like behaviours. Neurobiol. Dis. 2023, 185, 106223. [Google Scholar] [CrossRef] [PubMed]
- Wolmarans, D.W.; Brand, L.; Stein, D.J.; Harvey, B.H. Reappraisal of spontaneous stereotypy in the deer mouse as an animal model of obsessive-compulsive disorder (OCD): Response to escitalopram treatment and basal serotonin transporter (SERT) density. Behav. Brain Res. 2013, 256, 545–553. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Depressive disorder (depression) [WWW Document]. 2025. Available online: https://www.who.int/news-room/fact-sheets/detail/depression?
- Würbel, H.; Stauffacher, M. Age and weight at weaning affect corticosterone level and development of stereotypies in ICR-mice. Anim. Behav. 1997, 53, 891–900. [Google Scholar] [CrossRef]
- Wynchank, D.; Berk, M. Behavioural changes in dogs with acral lick dermatitis during a 2 month extension phase of fluoxetine treatment. Hum. Psychopharmacol. 1998, 13, 435–437. [Google Scholar] [CrossRef]
- Yalcin, E.; Aytug, N. Use of fluoxetine to treat stereotypical pacing behavior in a brown bear (Ursus arctos). J. Vet. Behav. Clin. Appl. Res. 2007, 2, 73–76. [Google Scholar] [CrossRef]
- Yang, K.; Essa, A.; Noriega, D.; Yu, D.; Osiecki, L.; Gauvin, C.A.; Illmann, C.; Bortolato, M.; Dunn, E.C.; Mathews, C.A.; Scharf, J.M. Relationship between adverse childhood experiences and symptom severity in adult men with Tourette syndrome. J. Psychiatr. Res. 2022, 155, 252–259. [Google Scholar] [CrossRef]
- Yang, Z.; Wu, H.; Lee, P.H.; Tsetsos, F.; Davis, L.K.; Yu, D.; Lee, S.H.; Dalsgaard, S.; Haavik, J.; Barta, C.; Zayats, T.; Eapen, V.; Wray, N.R.; Devlin, B.; Daly, M.; Neale, B.; Børglum, A.D.; Crowley, J.J.; Scharf, J.; Mathews, C.A.; Faraone, S. V.; Franke, B.; Mattheisen, M.; Smoller, J.W.; Paschou, P. Investigating shared genetic basis across Tourette syndrome and comorbid neurodevelopmental disorders along the impulsivity-compulsivity spectrum. Biol. Psychiatry 2021, 90, 317–327. [Google Scholar] [CrossRef]
- Yasir, W.; Sherin, M.; Kazmi, S.F. The role of childhood trauma in developing nail biting and trichotillomania among adolescents. Int. J. Innov. Creat. Chang. 2021, 15, 1314–1332. [Google Scholar]
- Yeo, S.K.; Lee, W.K. The relationship between adolescents’ academic stress, impulsivity, anxiety, and skin picking behavior. Asian J. Psychiatr. 2017, 28, 111–114. [Google Scholar] [CrossRef]
- Young, F.M. The incidence of nervous habits observed in college students. J. Pers. 1947, 15, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Young, L.E.; McCallum, R.T.; Perreault, M.L.; Mason, G.J. Opportunistically using a chronic unpredictable stress study to investigate ‘inactive-but-awake’ behaviour as a potential welfare indicator in laboratory rats. Appl. Anim. Behav. Sci. 2024, 274. [Google Scholar] [CrossRef]
- Yue, S.; Duncan, I.J.H. Frustrated nesting behaviour: Relation to extra-cuticular shell calcium and bone strength in White Leghorn hens. Br. Poult. Sci. 2003, 44, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Zadegan, S.A.; Ramirez, F.; Park, J.W.; Rocha, N.P.; Furr Stimming, E.; Teixeira, A.L. Frequency of depression in Huntington’s disease: A systematic review and meta-analysis. J. Huntingtons. Dis. 2025, 14, 43–58. [Google Scholar] [CrossRef]
- Zhang, C.; Juniper, D.T.; Meagher, R.K. Effects of physical enrichment items and social housing on calves’ growth, behaviour and response to novelty. Appl. Anim. Behav. Sci. 2021, 237, 105295. [Google Scholar] [CrossRef]
- Zhang, M. yue; Li, X.; Zhang, X. hui; Liu, H. gui; Li, J. hong; Bao, J. Effects of confinement duration and parity on stereotypic behavioral and physiological responses of pregnant sows. Physiol. Behav. 2017, 179, 369–376. [Google Scholar] [CrossRef]
- Zhang, Z.-Y.; Mao, Y.; Feng, X.-L.; Zheng, N.; Lü, L.-B.; Ma, Y.-Y.; Qin, D.-D.; Hu, X.-T. Early adversity contributes to chronic stress induced depression-like behavior in adolescent male rhesus monkeys. Behav. Brain Res. 2016, 306, 154–159. [Google Scholar] [CrossRef]
- Zibell, S.; Madansky, E. Impact of gum chewing on stress levels: Online self-perception research study. Curr. Med. Res. Opin. 2009, 25, 1491–1500. [Google Scholar] [CrossRef]
- Zimmerman, P.H.; Koene, P.; Van Hooff, J.A.R.A.M. Thwarting of behaviour in different contexts and the gakel-call in the laying hen. Appl. Anim. Behav. Sci. 2000, 69, 255–264. [Google Scholar] [CrossRef]
- Zohar, A.H.; LaBuda, M.; Moschel-Ravid, O. Obsessive-compulsive behaviors and cognitive functioning: A study of compulsivity, frame shifting and type a activity patterns in a normal population. Neuropsychiatry, Neuropsychol. Behav. Neurol. 1995, 8, 163–167. [Google Scholar]
Figure 1.
Recent conceptions of the aetiology of ARBs in captive animals. Sub-optimal treatment (e.g., barren housing, rearing in isolation, early weaning, chronic exposure to aversive stimuli, pathogen threat, etc.) promotes ARBs in two non-mutually exclusive ways.
Figure 1.
Recent conceptions of the aetiology of ARBs in captive animals. Sub-optimal treatment (e.g., barren housing, rearing in isolation, early weaning, chronic exposure to aversive stimuli, pathogen threat, etc.) promotes ARBs in two non-mutually exclusive ways.
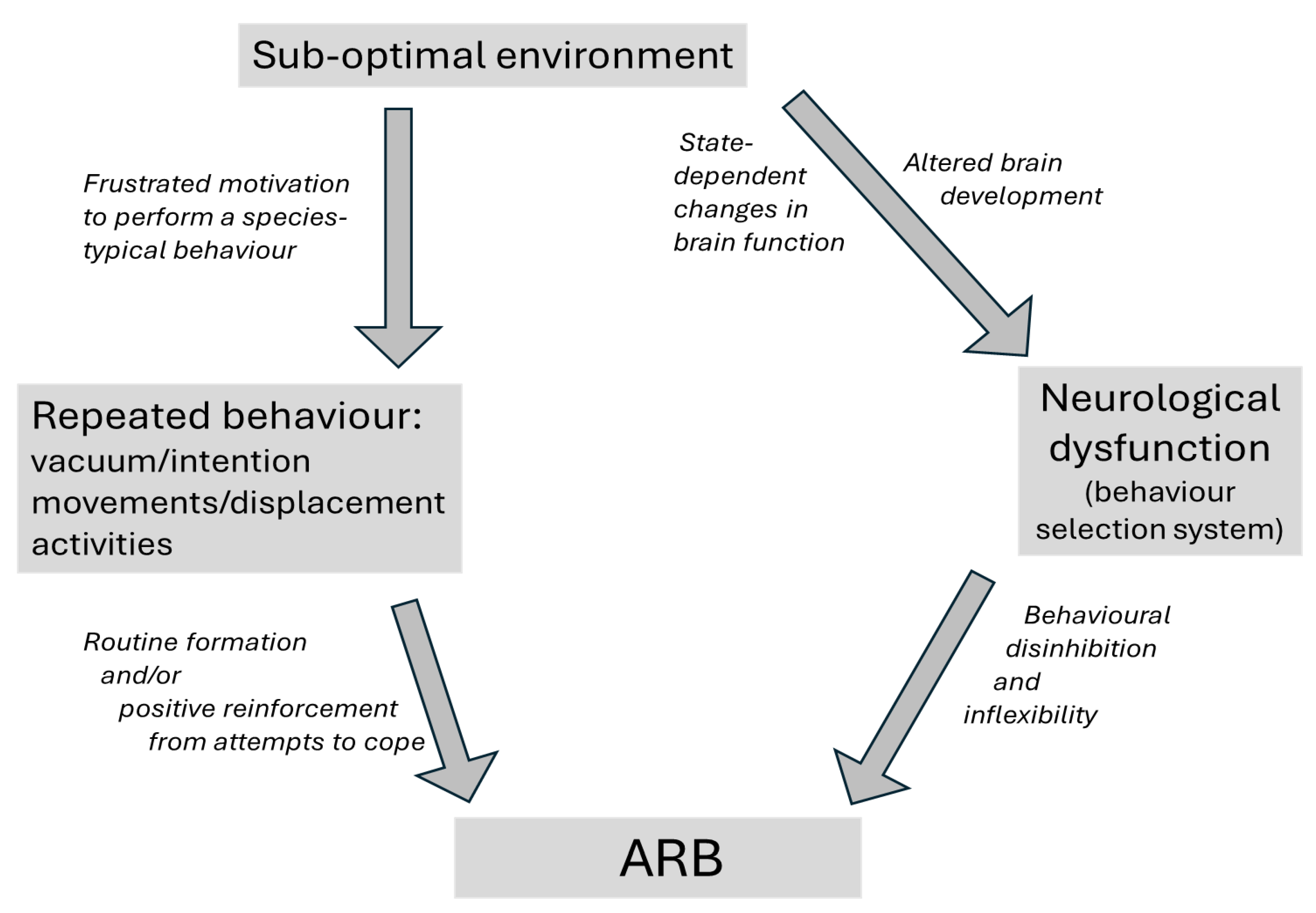
Figure 2.
Figure 2. The development of ARB in differentially housed deer mice, and the impact of regular isolation in a barren test cage. A) CONTROL GROUP: Each mouse is placed alone in a barren test cage (for automated ARB assessment) just once at one of the eight ages shown. Data are from mice reared in ‘enriched’ (n = 93) and barren housing (n = 123). B). RECURRENT ISOLATION GROUP: Each mouse is placed alone in a barren assessment cage eight times, i.e., approximately every 5 days. Data again are from mice reared in ‘enriched’ (n = 40) and barren housing (n = 19). Error bars are ± SEM. Reproduced from Bechard et al. (2017).
Figure 2.
Figure 2. The development of ARB in differentially housed deer mice, and the impact of regular isolation in a barren test cage. A) CONTROL GROUP: Each mouse is placed alone in a barren test cage (for automated ARB assessment) just once at one of the eight ages shown. Data are from mice reared in ‘enriched’ (n = 93) and barren housing (n = 123). B). RECURRENT ISOLATION GROUP: Each mouse is placed alone in a barren assessment cage eight times, i.e., approximately every 5 days. Data again are from mice reared in ‘enriched’ (n = 40) and barren housing (n = 19). Error bars are ± SEM. Reproduced from Bechard et al. (2017).
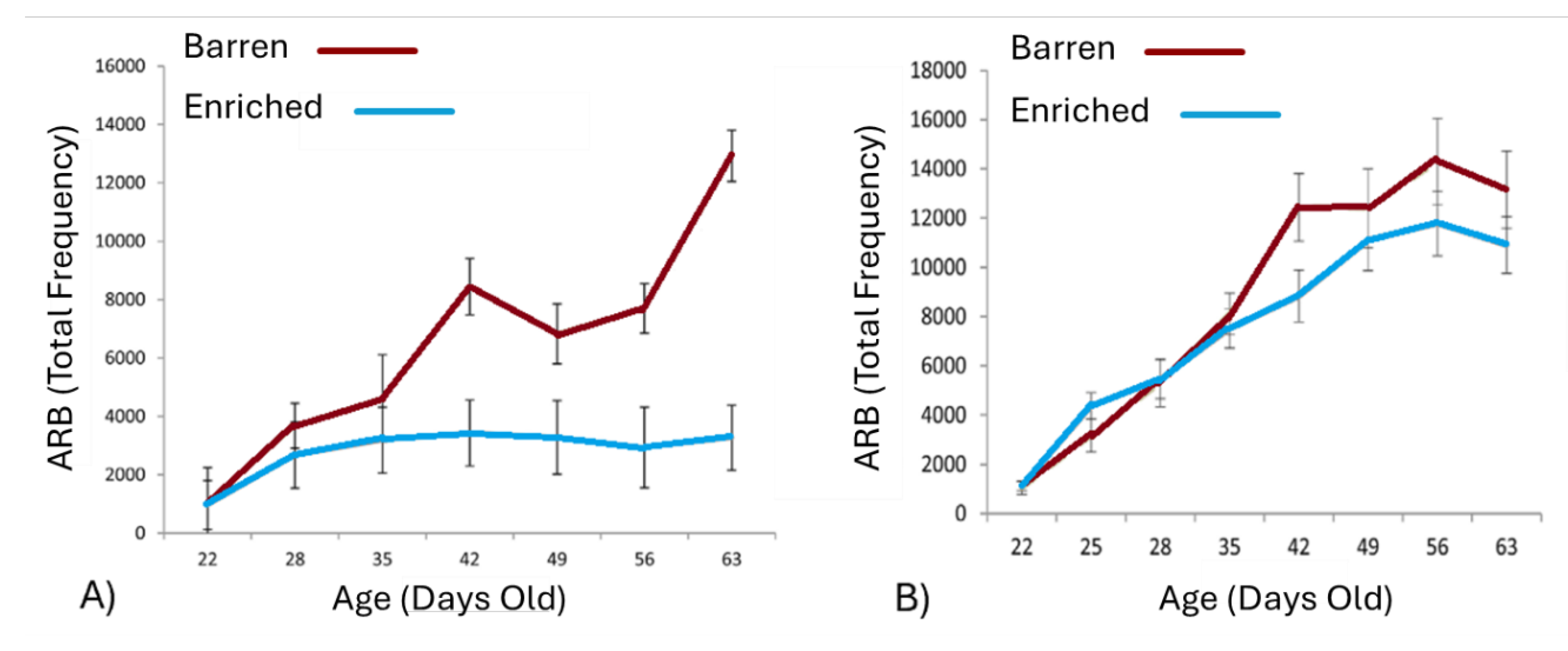
Figure 3.
In adult male pigtailed macaques (Macaca nemestrina), the current proportion of time spent in total abnormal behaviour (including ARBs) is predicted by the proportion of time spent singly housed before 48 months of age, and by whether mother-reared or nursery-reared without the mother (Bellanca and Crockett, 2002). Furthermore, re-analysis of these data reveals a significant interaction between the two, because they have synergistic effects (p < 0.05): having been nursery-reared promotes ARB more in monkeys who spent more time alone than in monkeys who spent more time socially-housed; while spending time alone also promoted ARB more in nursery-reared than in mother-reared animals. Data extracted and replotted from Bellanca and Crockett (2002).
Figure 3.
In adult male pigtailed macaques (Macaca nemestrina), the current proportion of time spent in total abnormal behaviour (including ARBs) is predicted by the proportion of time spent singly housed before 48 months of age, and by whether mother-reared or nursery-reared without the mother (Bellanca and Crockett, 2002). Furthermore, re-analysis of these data reveals a significant interaction between the two, because they have synergistic effects (p < 0.05): having been nursery-reared promotes ARB more in monkeys who spent more time alone than in monkeys who spent more time socially-housed; while spending time alone also promoted ARB more in nursery-reared than in mother-reared animals. Data extracted and replotted from Bellanca and Crockett (2002).
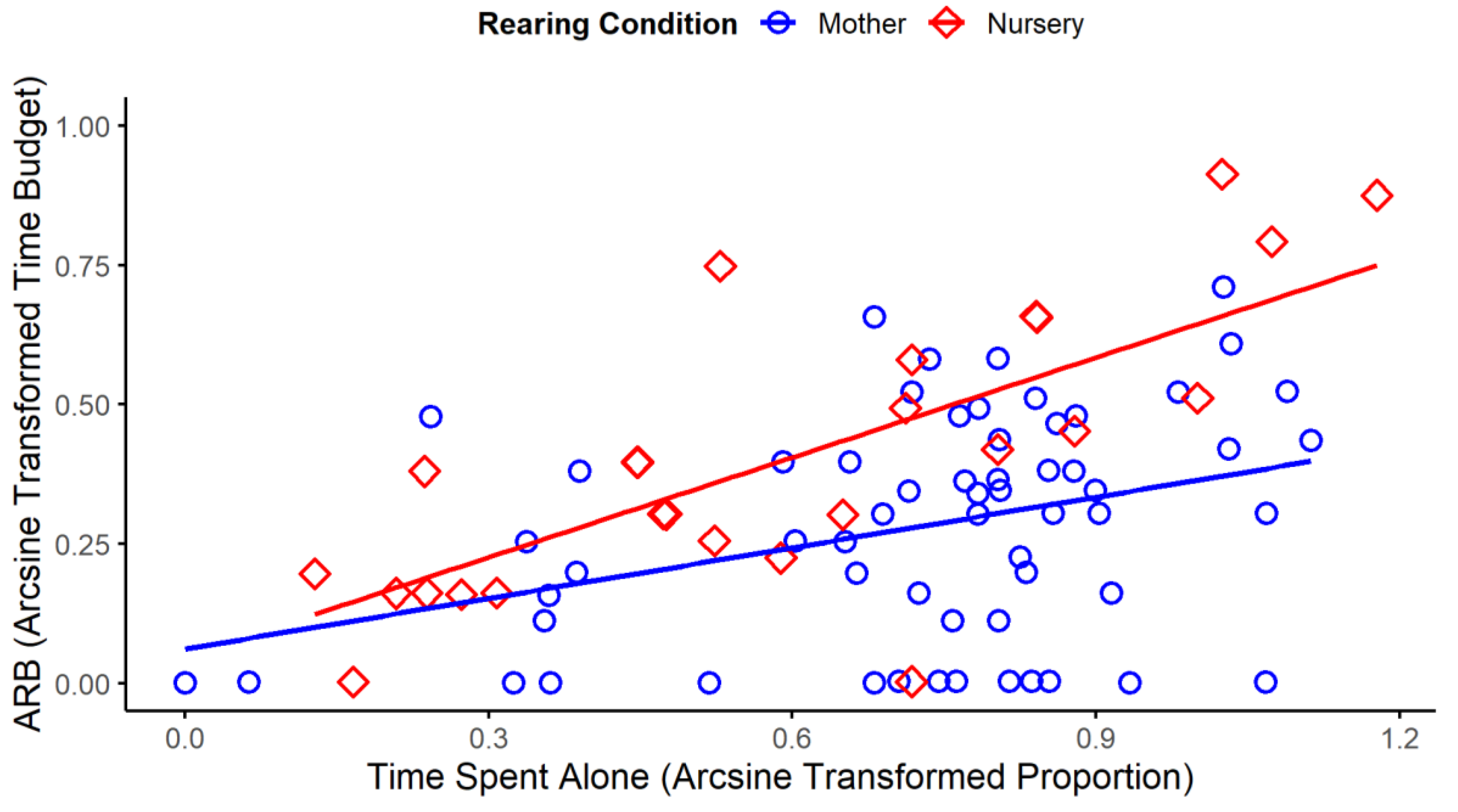
Figure 4.
Our updated conception of the aetiology of ARBs.
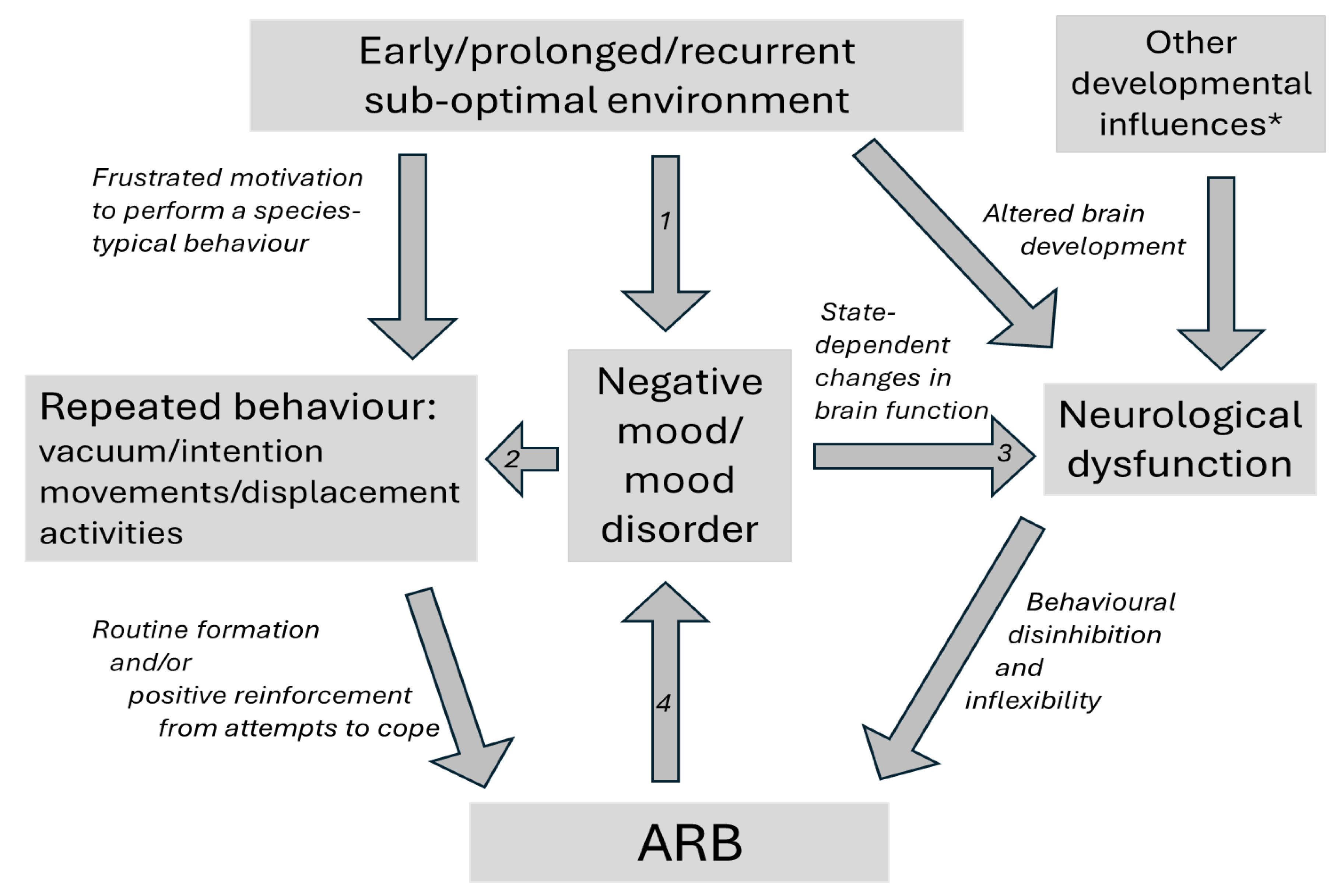

Table 1.
Associations between negative moods, mood disorders and ARBs in human clinical conditions typified by ARB.
Table 1.
Associations between negative moods, mood disorders and ARBs in human clinical conditions typified by ARB.
| Condition (ARB) | Levels of negative mood or affective disorders compared to neurotypical humans | Within condition, associations between ARB and negative experiences, negative mood or affective disorders |
| Autism Spectrum Disorder (various repetitive behaviours) | Higher prevalence of anxiety in people with autism (e.g., Fuld, 2018; Nimmo-Smith et al., 2020), and elevated levels of anxiety (e.g., Rodgers et al., 2012, rev. Baribeau et al., 2020; García-Villamisar and Rojahn, 2015) Depression is more common too (e.g., Fuld, 2018) |
Children with high anxiety perform more ARB than those without anxiety (Rodgers et al., 2012) Children who are more bullied show more anxiety and more ARB (Cappadocia et al., 2012) Childhood trauma or stress exposure is associated with more severe symptoms in children/adolescents (Berg et al., 2016; Mehtar and Mukaddes, 2011; reviewed Dodds, 2021) Individual ARB scores positively covary with scores for stress (i.e., number of perceived stressors experienced day-to-day) and anxiety (Dellapiazza et al., 2022; García-Villamisar and Rojahn, 2015; Moore et al., 2022; Uljarević and Evans, 2017; Wigham et al., 2015) Compared to individuals reporting relatively high quality of life, those who self-report low quality of life show more ARB (Johansson and Sandin, 2023) |
| Excoriation Disorder (skin-picking) | Greater depressive tendencies (Özten et al., 2015); also higher PTSD scores (Özten et al., 2015; Spitzer et al., 2022) Higher ‘psychological distress’ (Spitzer et al., 2022), and more likely to sleep poorly than controls (Cavic et al., 2021) |
Academic stress and trait anxiety scores predict skin-picking severity in adolescents (Yeo and Lee, 2017). Skin-picking severity covaries with self-reported distress in adults (Maraz et al., 2017) |
| Obsessive-Compulsive Disorder (various compulsions) | Involves persistent worry and distress (APA, 2022) In children/adolescents, higher levels of psychosocial stress (Lin et al., 2007), and more anxiety and depression (Horesh et al., 2018) Increased risks of depression (e.g., reviewed Vidal-Ribas et al., 2015) |
Levels of psychosocial stress in children/ adolescents predict future OCD symptom severity (Lin et al., 2007), especially in subjects previously exposed to Streptococcus (Lin et al., 2010) OCD is anecdotally elevated in prison (Prison UK, 2014) Experiencing stressful life events or traumas often seems to trigger OCD onset (e.g., Murayama et al., 2020) |
|
Rett’s Syndrome (various repetitive behaviours) |
High anxiety (e.g., Barnes et al., 2015) Elevated anxiety and low mood (Mount et al., 2001) |
Anxiety leads to more ARB (Pineda, 2024); see also Buisson et al. (2017) Positive relationship between ARBs and anxiety scores (Cianfaglione et al., 2015) |
| Stereotypic Movement Disorder (SMD) (various repetitive behaviours) | Among college students who body rock, anxiety disorders and ‘general distress’ are higher than in the general population (Rafaeli-Mor et al., 1999). Elevated anxiety (Lewis, 2013), including clinical levels (Oakley et al., 2015), is common in SMD; most subjects also reported challenges at school (see also Castellanos et al., 1996) |
|
| Tourette’s syndrome (tics) | In children/adolescents, elevated levels of psychosocial stress (Lin et al., 2007); anxiety and depression (Horesh et al., 2018) | Levels of psychosocial stress in children/adolescents predict future tic severity (Lin et al., 2007), especially in subjects previously exposed to Streptococcus (Lin et al., 2010) Weak but significant positive correlation between motor tics severity and the quantity of daily minor negative events (Steinberg et al., 2013); clinical experience also suggests stressful experiences (e.g., starting school, relationship challenges) increase tic severity (Buse et al., 2014) More stressful events in the preceding months were associated with increased severity of tics (Tan et al., 2024) |
| Trichotillomania (hair-pulling) | Adults are more likely to experience anxious or depressed moods (e.g., Özten et al., 2015; Houghton et al., 2016; Cavic et al., 2021); also higher PTSD scores (Ötzen et al., 2015; (though cf. Gershuny et al., [2006] who find less PTSD despite trauma exposure) More likely to sleep poorly than controls (Cavic et al., 2021) Linked with depression in children (King et al., 1995; cited by Chamberlain et al., 2007) |
Severe hair-pulling is linked with elevated depression (Houghton et al., 2016), higher distress scores (Maraz et al., 2017), and poorer sleep quality (Cavic et al., 2021) Affected individuals develop more severe forms of hair pulling if clinically depressed, especially if also have an anxiety disorder (Grant et al., 2017b, 2017a, reviewed Yasir et al., 2021) Yasir et al. (2021) find that degrees of childhood trauma predict degrees of hair-pulling in adolescents |
|
Other Hair-pulling and other self-directed ARBs pooled Chorea (e.g., in Huntington’s disease) |
Depression and anxiety are common in Huntington’s disease (Dale and van Duijn, 2015; Zadegan et al., 2025) |
The degree to which individuals find their lives unpredictable, uncontrollable, and stressful predicts time spent in self-directed ARBs (e.g., hair-pulling, skin-picking: Grant et al., 2015) Higher scores of depression, anxiety and stress linked with performing more forms of self-directed ARB, along with hair-pulling (e.g., skin-picking, nail-biting: Stein et al., 2008) Severity of hair-plucking, skin-picking and nail-biting covaries with distress (Maraz et al., 2017) Chorea is usually worsened by anxiety and stress (Bhidayasiri and Truong, 2004) |
Table 2.
Table 2. Resources that animals are motivated to interact with (i.e., spend more time with than expected by chance, or perform appetitive behaviours to reach), and their impact on ARB if absent.
Table 2.
Table 2. Resources that animals are motivated to interact with (i.e., spend more time with than expected by chance, or perform appetitive behaviours to reach), and their impact on ARB if absent.
| Species | Preferred resource | Effect on ARB in animals housed without this |
|---|---|---|
| Chickens (Gallus gallus domesticus) | Nestboxes (e.g., Cooper and Appleby, 1995; 1997) |
Increased pacing before egg-laying (Yue and Duncan, 2003; Zimmerman et al., 2000) |
| Foraging/dust-bathing substrates (e.g., Dawkins and Beardsley, 1986) | More prevalent, more time-consuming feather pecking (e.g., Blokhuis and Arkes, 1984; Nicol et al., 2001; Dixon and Duncan, 2010) | |
| Perches for sleeping on at night (Olsson and Keeling, 2002) | More prevalent feather pecking (Huber-Eicher and Audige, 1999) | |
| Parrots (assuming similarities across species: Mellor et al., 2021) | Opportunities to forage (chew and manipulate—studied in orange-winged Amazon parrots (Rozek and Millam, 2011) | Reduced feather-plucking/chewing (Lumeij and Hommers, 2008)—studied in African grey parrots (Psittacus erithacus) |
|
Chinchillas (Chinchilla lanigera) |
Dust-bathing substrates (Redman, 1974) |
More prevalent fur-chewing (trend: Ponzio et al., 2007) |
| Mice (Mus musculus) | Nesting material (Roper, 1975) |
More cage-top twirling (Gross et al., 2011) |
| Diverse resources, including more space, diverse forms of shelter and nesting material, running wheels, hammocks, and chewable cardboard (Sherwin, 1998, 1996; Sherwin and Nicol, 1997, 1996; Tilly et al., 2010; Walker and Mason, 2018) | More prevalent, time-consuming ARB including backflipping, route-tracing, bar-mouthing, cage-top twirling and fur-plucking (e.g., Olsson and Sherwin, 2006; Tilly et al., 2010; Bechard et al., 2011; Nip et al., 2019) | |
| Bank voles (Clerionomys glareolus) | Loose hay and twigs as cover (Cooper et al., 1996) |
More prevalent repetitive jumping ARB (Ödberg, 1987) |
|
Gerbils (Meriones unguiculatus) |
Opaque artificial burrows (chambers plus tunnels) (Waiblinger and König, 2004) | Increased stereotypic digging in cage corners (Waiblinger and König, 2004; Wiedenmayer, 1997) |
| Mink (Neogale vision) | Water in which animals can swim/head-dip (Cooper and Mason, 2000; Mason et al., 2001) | Locomotor ARBs (e.g., pacing) are more prevalent and time-consuming (Hansen and Jeppesen, 2000; Ahola et al., 2011; though cf. Vinke et al., 2008) |
| Running wheels (Hansen and Jensen, 2006) | Increased locomotor ARBs (e.g., pacing) and ‘stationary’ whole-body ARBs (e.g., head-twirling: Malmkvist et al., 2024) | |
| Diverse resources, including space, flowing water, and novel and/or manipulable objects (Dallaire et al., 2012; Díez-León et al., 2016) | Pacing, head-twirling and scrabbling are more prevalent and time-consuming (e.g., Dallaire et al., 2011; Campbell et al., 2013; Díez-León et al., 2016) | |
| Blue Foxes (Vulpes lagopus) | Dirt floors for digging (Koistinen et al., 2016, 2007) | More time spent in oral ARBs (Koistinen et al., 2008) |
| Pigs (Sus scrofa) | Straw to manipulate and forage in (e.g., Pedersen et al., 2002) | Increased prevalence of tail-biting (reviewed by Wallgren et al., 2019) |
| Cattle (Bos taurus) | Brushes to self-scratch and groom with (McConnachie et al., 2018) | More time spent bar-licking (Park et al., 2020) |
Table 3.
Forms of ARBs investigated across Tests 2–5: Supporting evidence for associations with poor welfare (assuming it is defensible to pool similar-looking ARBs performed by animals of the same taxonomic order or class).
Table 3.
Forms of ARBs investigated across Tests 2–5: Supporting evidence for associations with poor welfare (assuming it is defensible to pool similar-looking ARBs performed by animals of the same taxonomic order or class).
| ARB Type | Test 2: Aversive Stimuli | Test 3: Fitness Threats | Test 4: Affect Modulating Drugs | Test 5: Concordance with Other Valid Indicators |
|---|---|---|---|---|
| Self-plucking in Psittaciformes | ↑ Barren housing | ↑ Illness and infection; early parental loss; social isolation |
↓ By tricyclic antidepressant | Social isolation can ↓ telomeres Individual ↑ fearfulness and “neuroticism” |
| Route-tracing / Repetitive Jumping in Rodents | ↑ Barren housing and food deprivation | ↑ Early weaning and social isolation | ↓ By anxiolytics antidepressants | Barren housing ↓ disease resilience and lifespan Individual ↑ corticosterone response to challenge |
| Self-injurious Behaviour in Primates | ↑ Barren housing and repeated/long-term exposure to aversive stimuli (e.g., CUS) | ↑ Maternal deprivation and social isolation | ↓ By SSRIs | Social isolation ↓ hippocampal neurogenesis) and ↓ health and survival Early maternal loss ↓ telomeres ↑ illness and ↓ survival Group-level ↑ cortisol response to stressor and sleep disruption |
| Pacing in Carnivora | ↑ Barren housing and food deprivation | ↑ Illness and infection, and early loss of the mother | SSRI ↓ pacing | ↑ Heart rate in small enclosures that induce pacing Group ↓ reproductive success Individual ↑ pacing correlates with ↓ reproductive success |
Table 4.
Table 4. How the forms of ARB presented in Table 3 can misbehave as welfare indicators, with explanations. ‘RoT’ = rule of thumb.
Table 4.
Table 4. How the forms of ARB presented in Table 3 can misbehave as welfare indicators, with explanations. ‘RoT’ = rule of thumb.
| ARB Type | Test 2: Aversive Stimuli | Test 3: Fitness Threats | Test 4: Affect Modulating Drugs | Test 5: Covariation with Valid Indicators |
|---|---|---|---|---|
| Self-plucking in Psittaciformes | ARBs take time to develop, plus eventually plateau RoT 4: False nulls due to threshold and ceiling effects |
Rearing with a parent does not always ↓ feather-plucking, nor does social isolation always ↑ RoT 4: False nulls (likely threshold effects), but possible priming to respond to futurechallenges with ARB |
Not always ↓ by SSRIs Meaning uncertain: affective impact of SSRIs is highly variable. Best to follow RoT 1 |
N/A |
| Route-tracing / Repetitive Jumping in Mice | Not all genotypes respond to stressors with ARB RoT 5: False nulls:ARBs not suitable for comparing genotypes ARBs take time to develop and also eventually plateau RoT 4: False nulls due to threshold and ceiling effects With time, becomes hard to alleviate with ‘enriched’ housing Possibly indicates hard-to-treat negative affect/cumulative poor welfare over a lifetime (more research needed) Best to follow RoT 1 |
No apparent effect of maternal separation age RoT 4: False null (likely threshold effect), but possible priming to respond to futurechallenges with ARB |
↑ When benzodiazepine given to restraint-stressed mice False lead possibly caused by reduced inactivity, in which case ideally corrected with RoT 6 (though N. B. freezing, hiding and inactive-but-awake were unfortunately not recorded) |
High ARB individuals have ↑ reproductive success and are less anxious RoTs 3 & 5: ARBs are not suitable for comparing individuals |
| Self-injurious Behaviour in Primates | Not ↓ by ‘enrichment’ (not ↑ by all stressors) RoT 4: False null (perhaps caused by motivational specificity) |
Social isolation does not ↑ SIB RoT 4: False null (perhaps caused by motivational specificity) |
Self-biters are less reactive to human presence and respond with ↓ ARB—replaced with freezing RoTs 3 & 5: ARBs not suitable for comparing individuals |
|
| Pacing in Carnivora | Pacing is ↓ by aversive stimuli when animals hide instead Pacing may ↑ with general activity levels and anticipation of food arrival RoT 2: ARB bout onset/offset does not reliably indicate negative emotion RoT 6: False nulls caused by increased inactivity |
May not reflect pain if housing is improved RoT 4: False null due to possible threshold effects |
Long-term pacing may be treatment-resistant Possibly indicates hard-to-treat negative affect/cumulative poor welfare over a lifetime (more research needed); best to follow RoT 1 |
Not always consistent at the individual level RoTs 3 & 5: ARBs not suitable for comparing individuals |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



