1. Introduction
Forest ecosystems are under significant pressure from anthropogenic factors, including logging, soil degradation, and climate change, leading to biodiversity loss and a reduction in their carbon sequestration capacity. The disruption of the natural carbon cycle exacerbates the climate crisis, making forest conservation and restoration one of the key challenges for sustainable development [
1,
2].
Recent studies emphasize that enhancing photosynthetic carbon uptake through increased forest cover and the restoration of degraded forests could become one of the most effective methods for climate change mitigation. According to the Intergovernmental Panel on Climate Change [
2], limiting global warming to below 1.5 °C by 2050 would require the establishment of an additional 1 billion hectares of forest plantations. A more recent analysis by Bastin et al. refined this estimate, suggesting that planting 0.9 billion hectares of forests could sequester up to 205 Gt of carbon, based on a global assessment of land available for reforestation [
3].
In the context of Russia's Low-Greenhouse-Gas-Emissions Socio-Economic Development Strategy until 2050 and the implementation of the federal "Forest Conservation" project, the country faces the task of increasing reforestation volumes. This involves creating carbon-depositing forest stands and enhancing their resilience to climatic and anthropogenic stressors. Against this background, there is growing focus on cultivating tree species that can combine high growth rates with resilience to adverse conditions [
4].
Russian forests play a key role in the global carbon balance, with their contribution to CO₂ uptake significantly exceeding that of forests in many industrialized countries, such as the states of the European Union [
5]. Pine forests are of particular importance due to their high carbon sequestration potential. Notably, pine forests – the most widespread forest type in Russia – cover an area of approximately 110 million hectares. In the north, they border the tundra, while in the south, they extend to the dry steppes [
6]. Their role is critical in restoring degraded lands, protecting soil from erosion, and enhancing biodiversity in regions that have experienced prolonged adverse environmental impacts.
This study investigates the seasonal dynamics of photosynthetic activity in Scots pine (Pinus sylvestris L.) needles in the central region of European Russia during the 2024 growing season. The research aimed to determine the dependence of photosynthesis intensity, transpiration, and stomatal conductance on meteorological conditions, such as temperature and irradiance
2. Materials and Methods
The study objects were Scots pine (P. sylvestris L.) plants. The seed material was provided by the Voronezh Forest Breeding and Seed Production Center. To assess the rate of carbon dioxide uptake by needles attached to the plant, a portable CI-340 gas exchange system (CID Bioscience, USA) was used. This system allows for the measurement of various photosynthetic parameters in real-time.
Gas exchange and transpiration measurements were performed on the needles of well-lit apical shoots in their second year of growth, which ensured reliable data on the photosynthetic characteristics of this species. Three pairs of needles were placed in the chamber without overlapping each other, following the standard methodology [
7].
Figure 1.
Measurement of gas exchange and transpiration parameters in pine using the CI-340 infrared gas analyzer.
Figure 1.
Measurement of gas exchange and transpiration parameters in pine using the CI-340 infrared gas analyzer.
The measurements were replicated 3 to 5 times for each data point. The net photosynthesis rate was calculated by the device using the built-in software according to the following formula (1):
where:
Pn: the net photosynthetic rate for open systems (µmol m⁻² s⁻¹);
W: the mass flow rate per unit leaf area for open systems (mol m⁻² s⁻¹);
C0, Ci are the CO₂ concentrations at the outlet and inlet of the gas analyzer, respectively (µmol mol⁻¹);
V: the gas flow rate through the leaf chamber (L min⁻¹);
P: the atmospheric pressure (bar);
Ta: the air temperature (K);
A: the leaf area (cm²).
The study also utilized data from a local meteorological station (Voronezh city) on air temperature and precipitation for the multi-year average period (10 years) and for the period from April to September 2024. Statistical analysis was performed using Statistica software, version 12.6 (StatSoft, Inc., USA).
3. Results
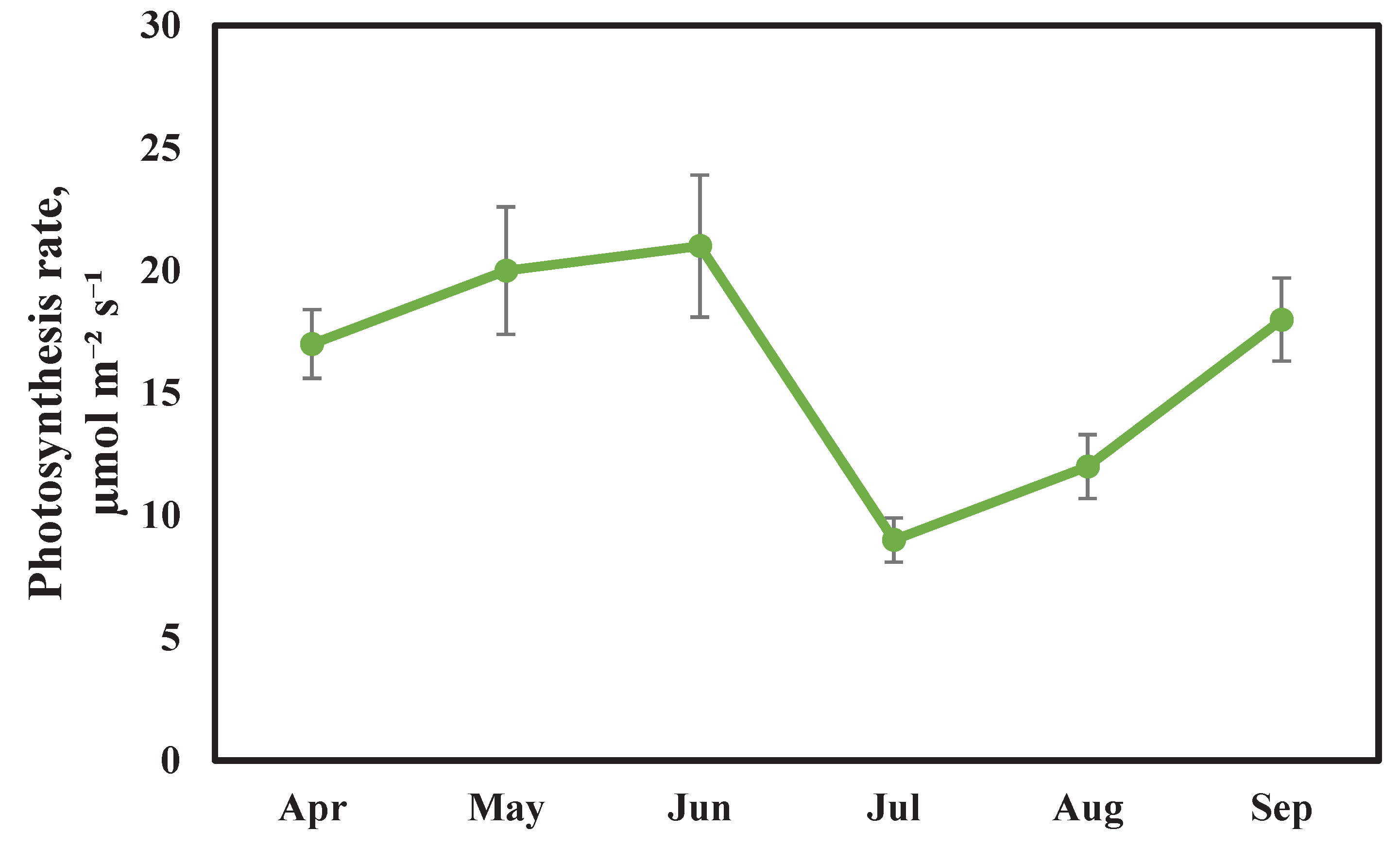
Photosynthesis is a key process that affects plant productivity and is highly dependent on climatic conditions. The dynamics of photosynthesis intensity in Scots pine plants during the 2024 growing season are presented in
Figure 2.
From April to May, the level of photosynthetic activity in the experimental Scots pine plants increased from 17 ± 1.4 to 20 ± 2.6 µmol m⁻² s⁻¹, representing a 15% growth. This is associated with changes in solar radiation intensity and photoperiod length.
During this period, the irradiance levels recorded in the experiments ranged from 1465 to 1889 µmol m⁻² s⁻¹ (
Table 1). The transition of the average daily air temperature above 5 °C was recorded as early as the last decade of March. Moreover, April was exceptionally warm, with a mean monthly temperature of 14.6 °C, which exceeded the multi-year average by 5.5 °C.
Scots pine is a pioneer species that is light-demanding and, therefore, not overly specialized to specific site conditions [
8]. Thanks to the pronounced folding of its mesophyll cells, the total assimilative surface area of pine needles is increased. Under conditions where the irradiance level is 60-90% of that in an open area, the size of the mesophyll cells, as well as the size and number of chloroplasts in the needle cells, remain constant. In contrast, when the irradiance level drops below 25%, noticeable changes occur in the size and number of cells per unit needle length, as well as in the number of stomata and chloroplasts per cell [
9].
During the specified spring period, alongside the increase in carbon dioxide assimilation rate, a rise in transpiration rate (from 3.1 ± 0.21 to 5.4 ± 0.64 mmol m⁻² s⁻¹) and stomatal conductance (from 205 ± 24 to 296 ± 36 mmol m⁻² s⁻¹) was also observed. Analyzing the influence of solar radiation intensity on stomatal activity under field conditions is complicated by the fact that the stomata of coniferous plants are affected by numerous exogenous and endogenous factors. Nevertheless, it is known that changes in stomatal conductance in woody plants, especially conifers, do not always directly correlate with fluctuations in light intensity, as stomata adapt to the latter over an extended period. For example, in Scots pine, the recovery period for high stomatal conductance after the removal of shading can be up to 4 hours [
10,
11].
In general, the stomatal conductance of Scots pine is weakly dependent on irradiance but is sensitive to low temperatures. Although the northern boundary of the pine's range often coincides with the limits of forest vegetation distribution, this species is still considered relatively thermophilic [
10,
12].
The average ambient air temperature at the experimental site of the calibration carbon polygon, where the physiological parameters of the experimental plants were measured, varied from 13.99 to 21.45 °C between May and June 2024. A particularly noticeable drop occurred during the May frosts. For instance, on May 15 (one week before our measurements), the air temperature fell to 2 °C, and the minimum soil surface temperature at night reached –1 °C. This likely explains the slight increase in the photosynthetic rate by only 4.7% compared to the previous month, given that the optimal temperature for the assimilatory apparatus in coniferous plants is 18–23 °C [
13].
The thermophilic nature of the root system is linked to the rare occurrence of pine on northern slopes and in shaded river valley headwaters, as well as on mossy bogs, where the soil warms up slowly [
12]. Due to low temperatures in the spring, Scots pine may not begin growth for an extended period. It has been repeatedly emphasized in the literature that in the north, influenced by weather factors, the onset and cessation of shoot growth in pine can be shifted [
13].
Furthermore, low temperatures adversely affect chloroplast structure, leading to the disruption of their ultrastructure and affecting the concentration of metabolites in the organelles. Cold stress also alters the structure of chloroplast membranes and enzyme activity, resulting in reduced photosynthetic activity, accumulation of excess reactive oxygen species (ROS), and abnormal thylakoid morphology. Enzymatic reactions can slow down, and high ROS levels can damage membrane structures. These factors can lead to chloroplast dysfunction and degradation, ultimately affecting the performance of the assimilatory apparatus [
14].
In July, the minimum level of photosynthetic activity in Scots pine was recorded, at 9.0 ± 0.92 µmol m⁻² s⁻¹. This decline is likely associated with the onset of a drought period in that month. The average air temperature between the measurements in June and July was 24.1 °C, with precipitation amounting to 33 mm. The high temperatures and moisture deficit observed during this period negatively affected the experimental Scots pine plants, manifesting not only in a reduced photosynthetic rate but also in a 52.8% decrease in transpiration rate and a 43.8% reduction in stomatal conductance.
In studies by R. Rehschuh and N. K. Ruehr on the impact of drought on Scots pine, it is noted that heat stress combined with drought amplifies the negative effect on the carbon and water balance of pine seedlings compared to heat stress alone. Under heat conditions, increased transpiration can provide leaf cooling if there is sufficient soil moisture and stomata remain partially open. However, during extreme heat, drought stress is exacerbated: the demand for water increases, leading to greater water loss by trees. This can slow the decline in internal water potential but simultaneously inhibits effective transpirational cooling, resulting in elevated leaf temperatures. Consequently, photosynthetic activity may decline due to reduced activity of key enzymes, such as Rubisco and components of Photosystem II [
15].
Between the measurements in July and August, a decrease in the average air temperature to 20.9 °C was recorded, along with an increase in total precipitation to 45 mm. As a result, an increase in the photosynthetic rate of Scots pine was observed, reaching 11.9 µmol m⁻² s⁻¹ in August. A similar stimulating effect of mild stress conditions has been reported in studies on other plant species [
16,
17].
Scots pine, classified as a xerophyte, possesses specific features of its assimilatory apparatus and root system that enable it to adapt to various soils with diverse hydrological regimes. In the early stages of development, pine is highly demanding of moisture. However, after the formation of a taproot and a system of lateral roots with a large number of vascular endings in 3-5-year-old saplings, the drought resistance of the plants increases [
12].
In September, under conditions of water deficit, the Scots pine plants exhibited a rise in photosynthetic activity, which reached 18 ± 1.7 µmol m⁻² s⁻¹, while transpiration activity dropped to its lowest value (2.3 ± 0.47 mmol m⁻² s⁻¹). Scots pine can maintain viability for extended periods under severe soil dryness by minimizing transpiration intensity. The active absorptive root surface area contracts, becoming covered with protective tissues that prevent water loss to the soil. In a dormant state, the roots can endure prolonged (1-2 months) and severe soil desiccation (water potential of 40-50 atm). Nevertheless, even under these conditions, portions of the absorbing roots with living primary cortex persist. The survival of pine is also favored by the xeromorphic structure of its needles [
12].
Water balance regulation at the whole-plant level could also occur through changes in stem hydraulic conductivity, which can significantly decrease due to cavitation – the formation of air-filled cavities. According to literature data, cavitation in Scots pine can occur even under drought conditions when stomata are closed. On one hand, this phenomenon can be viewed as a mechanism for limiting water flow to the crown during drought; on the other hand, it can be seen as a mechanism for rapidly redirecting water to the needles and optimizing photosynthesis. In some cases, mild embolism proves to be a beneficial adaptation, allowing species to maximize their photosynthetic rate [
9,
18].
However, there is currently a limited number of studies dedicated to the ability of Scots pine to recover from drought-induced embolism. In the work by R. Rehschuh and his co-authors, which examined the impact of xylem embolism on gas exchange in Scots pine, it was reported that no signs of recovery of stem hydraulic conductivity or reversal of xylem embolism were detected, either immediately after rewatering or one month later. These results were confirmed by both intrusive and non-intrusive methods. Despite the absence of refilling of the embolized tracheids, leaf gas exchange partially recovered, which was associated with a reduced water transport capacity [
19].
Furthermore, in September, the Scots pine plants exhibited a 39% increase in stomatal conductance compared to the minimum value recorded during the entire growing season in August (73.4 mmol m⁻² s⁻¹). This was presumably facilitated by the ability of coniferous plants to absorb atmospheric moisture. During drought, moistening by dew increases stomatal conductance, root growth, and shoot water potential. These findings have practical significance for understanding the distribution and growth patterns of conifers in regions with limited water availability and frequent dew and fog events [
9].
Thus, the seasonal dynamics of the photosynthetic rate in Scots pine during the 2024 growing season exhibited a bimodal pattern. The maximum level of photosynthetic activity, reaching 20.8 µmol m⁻² s⁻¹, was recorded in June. A sharp decline in this parameter to the minimum level of 8.9 µmol m⁻² s⁻¹ was observed in July, followed by a gradual increase to 18.10 µmol m⁻² s⁻¹ by the end of the season in September. For comparison, during the 2023 growing season, which was characterized by alternating periods of heavy rainfall and droughts, peaks of photosynthetic activity in the Scots pine samples were also recorded in June and September [
20]. These data may indicate that in 2023, the influence of arid conditions could have contributed to the development of a "drought memory" effect in the trees [
21].
Although the mechanisms contributing to the formation of the plant memory effect regarding abiotic stresses are not fully understood, a growing body of evidence points to the key role of chromatin structure changes in this process. Chromatin remodeling mechanisms, such as DNA methylation and histone modifications, can alter the transcriptional state of genes, activating or suppressing them without changing their nucleotide sequences. To cope with environmental stress conditions, plants employ extensive transcriptional reprogramming. Recent data indicate that these cis-memory mechanisms play a key role in the stress memory response in plants [
22].
This phenomenon likely facilitated more efficient carbon dioxide assimilation at the end of the 2024 growing season, highlighting the adaptive capacity of plants to unfavorable climatic conditions. Research exists confirming that plants which have previously experienced stressful conditions such as drought demonstrate higher photosynthetic rates compared to plants that have not undergone similar stress, even under comparable stomatal conductance and respiratory processes [
23,
24].