
Submitted:
23 December 2025
Posted:
24 December 2025
You are already at the latest version
Abstract
Zucchini plants are prone to powdery mildew, a fungal disease that can hamper plant growth and yield. This study investigated whether silicon supplementation, applied via fertigation or foliar sprays, could enhance plant resilience and reduce powdery mildew disease severity. Two independent greenhouse experiments were conducted, testing six treatments that varied in both silicon (Si) concentration and application method. The most pronounced effect was observed when fertigation with 1.5 mmol/L Si was combined with weekly foliar spraying (12.5–25 g/L Fertigro Sil), resulting in the lowest disease severity. Microscopic analysis further revealed significantly thicker cell walls in silicon-treated plants, suggesting a mechanical defense mechanism that may hinder fungal penetration. These findings highlight the potential of silicon as a sus-tainable and effective component in integrated crop management strategies for zuc-chini cultivation.
Keywords:
powdery mildew
; silicon
; induced plant resilience
; zucchini
; biological cultivation systems
; integrated crop management
1. Introduction
Powdery mildew, caused by various fungal species, is a globally widespread plant disease that affects a broad range of crops, including zucchini (Cucurbita pepo subsp. pepo). The pathogen species is usually highly specialized on 1 host species. The disease is characterized by the appearance of white, powder-like fungal spots on leaf surfaces, stems, and occasionally fruits. Infection leads to reduced photosynthetic capacity, premature leaf senescence and yield loss [1]. The pathogen thrives in environments with high humidity, moderate temperatures, and limited air circulation, making disease management a significant challenge in greenhouse and field production.
Developing sustainable and effective disease management strategies that minimize negative impact on human health and the environment is increasingly important. Silicon (Si) supplementation – applied via fertigation and/or foliar sprays – is one promising approach to become less dependent on pesticides. Silicon uptake and accumulation in plant tissue varies considerably between plant species: while some species absorb very little silicon, others accumulate large amounts in the above-ground plant parts [2]. However, the high uptake by some plants are not in themselves proof of the essentiality of silicon, since plants also grow well without silicon in the root environment [2,3,4]. Some authors suggest that silicon accumulation in plants is only due to non-selective passive transport in the transpiration stream [5]. Nevertheless, substantial evidence indicates that silicon can have a beneficial effect on plant growth and development, or plant resistance to diseases and pests. Therefore, silicon is referred to as a beneficial and not an essential plant nutrient [2]. In addition to its role in disease resistance, silicon has been shown to enhance plant tolerance to various abiotic stresses, such as drought and salinity, improve nutrient use efficiency, and alleviate heavy metal toxicity [6,7].
The role of silicon in disease suppression has been demonstrated in several crops. Both root and foliar applications of silicon can reduce powdery mildew severity in Cucurbitaceae [8,9,10], and similar protective effects have been observed against other pathogens, including Alternaria sp., Fusarium sp., Pythium sp., or Phytophthora sp. [11,12,13]. The modes of action of silicon include structural reinforcement of epidermal tissues through silicon deposition and activation of systemic induced resistance. This activation involves upregulation of defense-related enzymes and phenolic compounds, enhancing the plant’s overall defense response [14,15,16,17,18].
Plants absorb silicon from soil or nutrient solutions in the form of monosilicic acid (H4SiO4) [19]. The amount of silicon uptake varies greatly between species, depending on the presence of specific transport mechanisms. It appears in nature that there are large differences in silicon uptake capacity between plant species [2]. Not only with a clear distinction between mono- and dicots, though among the dicots only specific plant genera show a significant silicon uptake [2]. The Cucurbitaceae belong to the genera that absorb a relatively large amount of silicon [3]. Plants with a silicon content of approximately 0.5% to over 1% (dry weight) and a silicon-to-calcium ratio below 1 are classified as intermediate Si accumulators [20]. Cucurbitaceae, including zucchini, fall into this category, possessing NIP-III (Nod26-like intrinsic protein III) channels that facilitate silicon transport [21]. While zucchini does not accumulate silicon as extensively as hyperaccumulators, it is hypothesized that it can still benefit substantially from silicon supplementation.
Liang et al. (2005) demonstrated that both root and foliar applications of silicon can control infection of powdery mildew in cucumber (Cucumis sativus) [9], where foliar application lead to a strengthened physical barrier and root application can enhance defence resistance upon infection via enhanced activities of pathogenesis-related proteins (peroxidase, polyphenol oxidase and chitinase). For zucchini (Cucurbita pepo), studies have mainly focused on foliar applications, with evidence by Tesfagiorgis et al. (2011) that runoff reaching the root zone can increase efficacy [22]. Although studies such as Savvas et al. (2009) have also demonstrated the beneficial effects of silicon supplied via the nutrient solution on mildew suppression in zucchini [23], the comparative effects of fertigation, foliar application, and their combination have yet to be systematically evaluated in this crop.
This study therefore aims to determine whether silicon supplementation via fertigation and foliar sprays—applied individually or in combination—can mitigate powdery mildew infection in zucchini under greenhouse conditions. By evaluating different silicon concentrations and application methods (fertigation and/or foliar sprays), this research aims to determine the effectiveness of silicon in enhancing plant resilience of zucchini. By linking physiological responses (such as silicon uptake and cell wall thickening) to disease suppression, this research contributes to understanding how silicon can be used effectively in integrated, sustainable crop production systems for zucchini cultivation. This work contributes to bridging the gap between controlled experimental findings and practical applicability in greenhouse and open-field systems used by growers.
2. Materials and Methods
2.1. Location and Experimental Setup
Two independent experiments were conducted in an east-west oriented greenhouse at Wageningen University & Research, Business Unit Greenhouse Horticulture in Bleiswijk, the Netherlands. The total greenhouse area was 144 m². The first experiment was conducted from September to October 2024, and the second from April to May 2025.
2.2. Plant Material and Cultivation Conditions
Zucchini seeds of the cultivar Milos (Syngenta) with intermediate susceptibility to powdery mildew were sown in Ø15 cm pots filled with GO PP4 substrate (Jiffy). After sowing, the pots were covered with fiberfill (1mm, Royal Brinkman) for two days to restrict evaporation and ensure sufficient moisture for seed germination. Plants were cultivated under controlled greenhouse conditions for six weeks. The climate conditions were managed using an ISII climate computer (Hoogendoorn). After sowing, the greenhouse was maintained at a daytime temperature of 22.3 ± 1.9 °C and a nighttime temperature of 20.2 ± 0.4 °C. Relative humidity (RH) averaged 76.5 ± 9.5% during the day and 82.9 ± 1.3% at night. Corresponding vapor pressure deficits (VPD) were 4.9 ± 2.6 g/m³ during the day and 3.0 ± 0.3 g/m³ at night. After two weeks, temperature and humidity setpoints were lowered, resulting in adjusted conditions averaged 20.0 ± 2.2 °C during the day and 17.9 ± 0.8 °C at night, with RH values of 66.7 ± 11.1% and 77.5 ± 5.5% for day and night, respectively. VPDs during this period were 6.0 ± 2.8 g/m³ in the daytime and 3.5 ± 0.9 g/m³ at night.
Irrigation was provided using an ebb-and-flood system, with 7-minute irrigation cycles. During the first two weeks, plants were irrigated once per day. In the following two weeks, irrigation was increased to twice per day, and during the final two weeks, plants received water three to four times daily to meet increasing water demands by the growing plants.
2.3. Treatments
The greenhouse was arranged in four rows of six tables, in total 24 tables. Each table measured 1.25x2.00m and was capable of receiving individual nutrient solutions, allowing for precise fertigation treatments. Six treatments were tested in the experiments, varying in both silicon concentration and method of application (fertigation and/or foliar sprays). The treatments were randomly distributed across the tables within each row, providing four independent replicates per treatment. The treatments included a control (nutrient solution without added silicon); two fertigation treatments with silicon supplied at 0.75 mmol/L and 1.5 mmol/L concentrations respectively; and two foliar spray treatments with silicon applied weekly at concentrations of 12.5 g/L and 25 g/L (corresponding to 40.4 mmol Si/L and 80.7 mmol Si/L, respectively). A full nutrient solution was used for fertigation, the composition of the solution is provided in Appendix A Table A1, and was prepared using liquid fertilisers (YARA SUBSTRAFEED). Potassium metasilicate (K₂SiO₃) from YaraTera SUBSTRAFEED SIKAL, with 6.5, 3.25 and 6.5 mol/kg for K+, Si and OH- respectively, was used as the silicon source in the fertigation solution. To neutralize the OH-, equal molar rates of HNO3 were supplied, and equally the additional inputs of K+ and NO3- were compensated by equally reducing the input of K and NO3 from fertilisers in the basic fertigation solution for the two Si treatments.
Fertigro Sil (FS), with the same composition as SIKAL, was used for the foliar application. Foliar-Si treatments were fertigated with a Si-free nutrient solution identical to the control. The sixth treatment combined 1.5 mmol/L silicon via fertigation with a weekly 25 g/L FS foliar spray. For all treatments, the targets for the electrical conductivity (EC) was 2.3 dS/m and for the pH between 5 and 6. Silicon treatments via fertigation were applied from the start and the foliar spraying with silicon started two weeks after sowing and were repeated weekly.
2.4. Experimental Design and Powdery Mildew Experiment
Initially, each table accommodated 10 plants, resulting in 40 plants per treatment. Four weeks after sowing, half of the plants on each table were removed to prevent overcrowding and allow sufficient space for further development, resulting in a total of 20 plants per treatment for the remainder of the experiment. Two leaves per remaining plant were inoculated with 1-1.5·104 spores/mL powdery mildew (Podosphaera xanthii): the topmost mature leaf (minimum hand-size, horizontal orientation) and the leaf directly below. The suspension was applied with a fine mist atomizer to ensure even spore distribution on the leaf surface, avoiding excessive run-off. Inoculation was performed one day after the foliar spray treatment, and weekly applications continued thereafter. Two weeks after inoculation, powdery mildew severity was evaluated using the Spencer index [24]. Additionally, silicon content in plant sap was analyzed by Normec Groen Agro Control. A composite (pooled) sample was made for the following treatments: control, 1.5 mmol/L fertigation, and the combined treatment with 1.5 mmol/L fertigation and 25 g/L FS foliar sprays. For each treatment, a separate pooled sample was prepared from young leaves and from fully-grown leaves. Per table, two young and two fully-grown leaves were randomly collected.
2.5. Modifications in the Second Experiment
The study was conducted in two independent experimental runs. To minimize spatial bias, treatment allocation was re-randomized in the second trial. The second trial followed the same methodology as the first, with three modifications. First, due to phytotoxicity observed in the first trial from the 25 g/L FS foliar sprays (evidenced by necrotic leaf lesions), this treatment was replaced with a commercial silicon product M Vita Forte using 23 mg Si/L (0,8mmol Si/L) (Mertens B.V.). The combined treatment was therefore also adjusted to 1.5 mmol/L Si via fertigation and 12.5 g/L FS foliar sprays. Second, mildew severity scoring was refined based on insights from the first experiment to improve accuracy. Instead of using the Spencer index, disease severity was assessed by visually estimating both the percentage of leaf area covered with mildew and counting the number of newly infected leaves besides those initially inoculated. Third, leaves for plant sap analysis of all treatments were taken and assessed by Eurofins Scientific in the second experiment. For each treatment, one young and one fully-grown leaf were collected per table and pooled together, resulting in a composite sample combining young and old leaves for each treatment.
2.6. Microscopic Analysis of Cell Wall Thickness
Lastly, cell wall thickness was measured via microscopy after the second experiment. From each of the five plants per table, two fully developed leaves were sampled. Leaf sections (2 × 2 cm) were excised, and thin cross-sections (<1 mm) were manually prepared and mounted on microscope slides. Microscopic analysis was performed using a Nikon Eclipse Ni-L Upright Microscope with a 20× objective lens. Images were captured using NIS-Elements imaging software connected to the microscope. Cell wall thickness was quantified using the software’s manual measurement tool. Three measurements were taken per leaf. For presentation in the figures, original images were cropped to facilitate visualization. All images were minimally processed, and full, uncropped images are available in the Supplementary Material.
2.7. Statistical Analysis
The experiments were set-up in a randomized block design. Statistical analyses were performed in R (version 4.5.1). Depending on the response variable, different mixed models were applied: a cumulative link mixed model (clmm) for ordinal data, a beta mixed model (glmmTMB, logit link) for proportional data, a Poisson generalized linear mixed model (glmer, log link) for count data, and a linear mixed model (lmer) for continuous data. All models included the treatment as fixed effect and row allocation in the greenhouse as random effect. Model assumptions were verified by residual diagnostics. Post-hoc pairwise comparisons were conducted using estimated marginal means (emmeans) with Tukey adjustment. A significance threshold of p ≤ 0.05 was applied.
3. Results
3.1. Effect of Silicon Application on Powdery Mildew Severity
To evaluate whether silicon application - administered either through fertigation or foliar sprays - can reduce the severity of powdery mildew infection in zucchini plants, two independent greenhouse experiments were conducted. In both trials, silicon application via either method significantly reduced severity of infection with powdery mildew compared to the control without silicon.
In the first experiment, all silicon treatments – except for the 12.5 g/L FS foliar sprays treatment, led to a significant reduction in powdery mildew severity (Figure 1). The most pronounced effect was observed with the combined treatment of 1.5 mmol/L Si via fertigation and the 25 g/L FS foliar sprays. For both fertigation and the foliar sprays treatment, a significant dose response effect was observed.
The 25 g/L FS foliar sprays caused mild phytotoxicity, shown by necrotic lesions on treated leaves. This effect may have been caused by the high concentration, or by the high alkalinity of the monosilic acid solution, which can damage leaf tissue. Because of this phytotoxicity, the 25 g/L FS treatment was replaced in the second experiment with a commercially available silicon-based product (M Vita Forte). The combination treatment was adjusted to 1.5 mmol/L silicon via fertigation with 12.5 g/L FS foliar sprays, as this foliar sprays concentration did not show any phytotoxicity symptoms in the first experiment.
In the second experiment, the extent of mildew infection was assessed as percentage coverage on inoculated leaves, because in the first experiment Spencer index resulted in the score 4 or 5 for 65% of the plants. This time, fertigation at a dose rate of 0.75 mmol/L did not reduce disease severity. Both 1.5 mmol/L Si via fertigation and 12.5 g/L FS via foliar sprays significantly reduced powdery mildew severity, each reducing average leaf coverage to approximately half of that observed in the untreated control (30% ± 2% and 29% ± 4% vs. 60% ± 4%, respectively). The combined application yielded the greatest effect, reducing mildew coverage to just 11% ± 1%, an approximately 5.5-fold decrease compared to the control (Figure 2). In contrast, treatments with 0.75 mmol/L Si via fertigation or the M Vita Forte product alone did not lead to a reduction in disease severity. Visual differences between the control treatment and the combined application of 1.5 mmol/L Si via fertigation and 12.5g/L FS foliar sprays are illustrated in Figure 3, which shows severe infection on a plant cultivated without silicon compared to visibly fewer symptoms on a plant receiving the combined silicon treatment.
In addition to scoring inoculated leaves, the number of leaves that became naturally infected during the trial was also recorded. The 12.5 g/L FS foliar sprays treatment significantly reduced the number of additional infected leaves compared to the control treatment without silicon. Similarly, the greatest reduction was observed with the combined application of 1.5 mmol/L Si via fertigation and 12.5 g/L FS via foliar sprays (Figure 4).
3.2. Microscopic Analysis of Cell Wall Thickness
To explore a possible physiological mechanism underlying the observed reduction in disease severity, leaf samples were collected at the end of the second experiment from both the control treatment without silicon and the combined application of 1.5 mmol/L Si via fertigation and 12.5g/L FS foliar sprays, as these results were most pronounced, for microscopic analysis of cell wall thickness. This because silicon is known to be involved in structural reinforcement of the epidermal tissue. Leaves from the combined application treatment exhibited significantly thicker cell walls (3.3 ± 0.1 µm) compared to the control treatment without silicon (2.2 ± 0.1 µm), suggesting a structural strengthening of leaf tissues as mode of action of silicon that may contribute to enhanced plant resilience against powdery mildew on zucchini (Figure 5 and Figure 6).
3.3. Silicon Content in Plant Sap
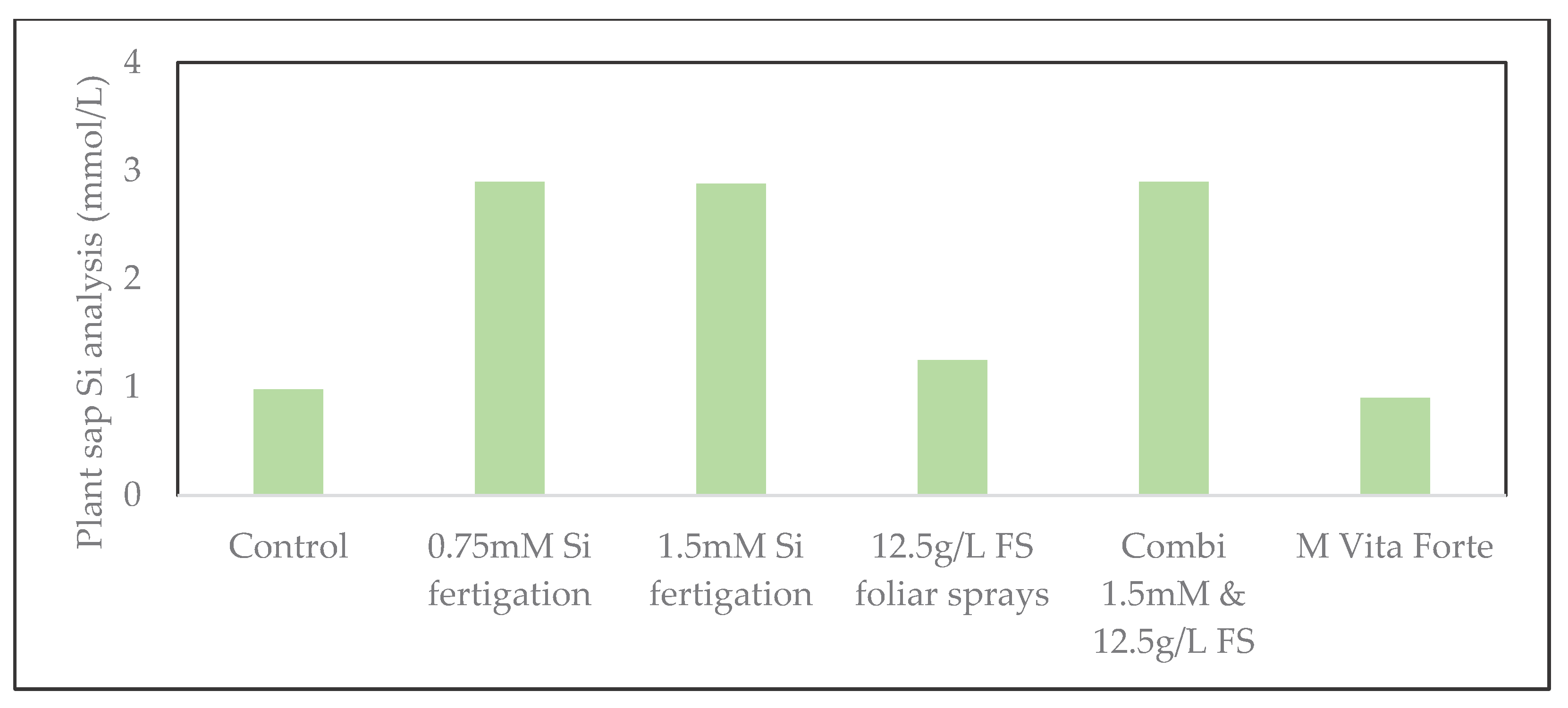
In both 2024 and 2025, plant sap analyses, based on composite (pooled) samples from multiple plants per treatment, revealed a consistent increase in silicon concentration following silicon treatments compared to the control. In 2024, fertigation with silicon already led to higher silicon levels in both young and fully-grown leaves, and the combined application of fertigation and foliar sprays resulted in an slightly higher Si accumulation for fully-grown leaves (Figure 7). The increase was more pronounced in fully developed leaves than in young leaves, suggesting active Si translocation within the plant. In 2025, the positive effect of Si supplementation was confirmed. Both fertigation and the combined application substantially enhanced Si concentrations in the plant sap compared to the control treatment (Figure 8). Here, 12.5g/L FS foliar spraying alone produced a moderate response, while the M Vita Forte treatment showed no measurable improvement. Overall Si accumulation was higher in 2025 than in 2024, most likely due to the higher plant transpiration as affected by the higher radiation in the 2025 trial, resulting in a higher Si uptake. Overall, these results demonstrate that Si fertigation, is an effective strategy to enhance Si uptake and distribution in zucchini. Note that these measurements represent pooled samples per treatment; therefore, variability among individual plants within treatments cannot be reported.
4. Discussion
Most commercial grown zucchini cultivars are susceptible to powdery mildew. This study demonstrates that silicon supplementation, applied either via fertigation or foliar sprays, significantly reduces disease severity under greenhouse conditions. In both independent trials, silicon-treated plants consistently exhibited lower disease severity compared to control plants that did not receive silicon. Notably, in the first experiment, the combination of 1.5 mmol/L Si via fertigation and 25 g/L FS foliar sprays resulted in the strongest reduction in disease, corresponding to an estimated ~3.5-fold decrease in affected leaf area compared to the control. However, this estimate should be interpreted with caution due to the non-linear nature of the Spencer index used to assess disease severity. In the second experiment, the combination of 1.5 mmol/L Si via fertigation and 12.5 g/L FS foliar sprays also had the most pronounced effect, reducing the percentage of leaf area affected by approximately 5.5-fold and limiting the spread of the pathogen to non-inoculated leaves. This treatment also resulted in a marked increase in silicon concentration in plant sap, confirming effective uptake and translocation, and indicating that zucchini actively absorbs and redistributes silicon to strengthen plant tissues and potentially enhancing defense mechanisms.
These results are consistent with previous reports in Cucurbitaceae, including pumpkin, cucumber, and zucchini, where increased silicon uptake reduced powdery mildew severity [8,9,10,25,26]. Liang et al. (2005) showed in cucumber that combined root and foliar silicon applications enhance induced resistance upon infection via enhanced activities of PR proteins [9]. Our study confirms this observation in zucchini and demonstrates that the combined treatment enhances resilience. The similarity across species indicates that zucchini, as an intermediate Si accumulator [20,21], can indeed respond strongly to silicon addition, with combined application further enhancing efficacy.
The magnitude of the response may vary considerably among species and experimental conditions. Factors such as intrinsic silicon accumulation capacity, chemical form and solubility of the silicon source, and the intensity of pathogen pressure can influence the degree of protection achieved. Additionally, root-absorbed silicon is transported to shoots via the xylem along transpiration streams [27], so variations in transpiration rate and soil moisture can affect overall silicon uptake. The concentrations used in this study are comparable to or slightly lower than those effective in Cucurbitaceae studies [8,9,10,28], suggesting that moderate silicon levels can already confer substantial protection.
Foliar application at higher concentrations (25 g/L FS in the first experiment) caused mild phytotoxicity, highlighting the importance of optimizing both concentration and pH of the formulation. The M Vita Forte treatment (10 % SiO₂, ≈ 4.7 % Si) supplied only about 0.8 mmol Si/L. In contrast, application of 25 g/L FS (19.4% water-soluble SiO₂) corresponds to 80.7 mmol Si/L, around 100 times higher. Silicon supplied via fertigation is continuously available for uptake through the roots, depending on transpiration rate, whereas foliar-applied silicon is taken up only shortly after each application. These results underscore that both product formulation and application strategy are critical for maximizing silicon uptake while minimizing phytotoxicity.
The protective effects of silicon can be explained by both structural and biochemical mechanisms. Structurally, silicon reinforces the epidermal cell wall through deposition and enhanced callose and lignin accumulation [29]. In our study, microscopy data showed that the combination of 1.5 mmol/L Si via fertigation and 12.5 g/L FS foliar sprays significantly increased cell wall thickness (3.3 ± 0.1 µm) compared to the control (2.2 ± 0.1 µm). While cell wall thickness was not measured for individual applications, both fertigation and foliar sprays alone reduced disease severity, suggesting that they too may contribute to structural reinforcement. Thicker cell walls form a physical barrier limiting fungal penetration and spread. Biochemically, silicon modulates both salicylic acid- and jasmonic acid–mediated defense pathways, activating defense-related enzymes such as polyphenoloxidase (PPO) and phenylalanine ammonia-lyase (PAL), stimulating the accumulation of secondary metabolites such as phenolics, flavonoids, and phytoalexins, and enhancing antioxidant defenses to reactive oxygen species (ROS) [15,16,30,31,32,33,34,35]. Although we did not measure PPO and PAL, it can be assumed that this mechanism also occurred in our experiments.
From a practical perspective, these findings are highly relevant for zucchini growers. Dutch zucchini is irrigated by using driplines, which provides an opportunity to apply silicon through fertigation. Foliar sprays can be applied with conventional spraying equipment, and in practice, foliar fertilization is often combined with pesticide applications. The combined use of fertigation and foliar spraying resulted in the greatest reduction in powdery mildew severity, indicating that silicon supplementation strategies can be optimized according to the irrigation system in use. Additionally, substrate and soil conditions may influence the efficacy of silicon supplementation. In open-field zucchini cultivation, crops are typically grown on sandy soils, where silicon is largely present in forms that are poorly available to plants [36]. This limited availability highlights the potential benefit of external silicon applications, particularly under conditions where root uptake from the soil is restricted.
Although this study focused on the fungal disease powdery mildew, the potential role of silicon in managing insect pests, such as aphids, also deserves further attention. Several studies have reported that silicon application can reduce aphid populations, primarily in plant species with a high capacity for silicon uptake, where silicon accumulation can substantially enhance mechanical barriers and possibly affecting host plant palatability and metabolism [37,38,39]. In contrast, the relevance of these effects for plant species with lower silicon accumulation capacity remains less clear but seem to affect herbivore performance indirectly via nutritional quality [40,41]. The observed increase in cell wall thickness in this study suggests that that silicon-induced structural changes may contribute to reduced insect feeding efficiency and reproduction. Further research is needed to determine whether, and under which conditions, silicon-mediated resistance against insect pests occurs. Overall, this study confirms that silicon supplementation, particularly when applied as a combination of fertigation and foliar sprays, is a promising strategy for managing powdery mildew in zucchini. The observed increase in cell wall thickening highlights the role of silicon in improving plant resilience. Given the increasing demand for sustainable and residue-free crop protection, silicon supplementation offers a promising, environmentally friendly approach to enhance plant resilience and reduce reliance on pesticides for specific crops.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, E.F.G., A.E., and J.B.M.; methodology, E.F.G., A.E., and J.B.M.; validation, E.F.G. and J.B.M.; formal analysis, E.F.G.; writing—original draft preparation, E.F.G.; writing—review and editing, E.F.G., A.E., and J.B.M.; visualization, E.F.G.; supervision, J.B.M.; project administration, J.B.M.; funding acquisition, J.B.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded within the framework of the Sectorale interventie Groenten & Fruit (SIG&F) program, supported by RVO (Netherlands Enterprise Agency).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
The authors greatly acknowledge the contributions of Erik Doorduin, Anne Hunting, Caroline Munts, Marianne Noordam, and Floortje Tilli for the practical execution and technical support of this study and Wim Voogt for his support in design and interpretation.
Conflicts of Interest
The funder had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A

Table A1.
Composition of the standard nutrient solution.
| Nutrient | Concentration | Micronutrient | Concentration |
|---|---|---|---|
| NH₄⁺ | 1.00 mmol/L | Fe | 30.0 µmol/L |
| K⁺ | 7.84 mmol/L | Mn | 10.0 µmol/L |
| Ca²⁺ | 4.31 mmol/L | Zn | 7.0 µmol/L |
| Mg²⁺ | 2.77 mmol/L | B | 25.0 µmol/L |
| NO₃⁻ | 16.87 mmol/L | Cu | 1.0 µmol/L |
| SO₄²⁻ | 2.30 mmol/L | Mo | 0.5 µmol/L |
| H2PO₄⁻ (P) | 1.53 mmol/L |
References
- Jiang, L.; Xiao, X. Research Progress on Powdery Mildew in Cucurbitaceae Plants, A Systematic Review. Molecular Pathogens 2024, 15. [Google Scholar] [CrossRef]
- Epstein, E. The Anomaly of Silicon in Plant Biology. Proc Natl Acad Sci U S A 1994, 91, 11–17. [Google Scholar] [CrossRef]
- Voogt, W.; Sonneveld, C. Silicon in Horticultural Crops Grown in Soilless Culture. In Silicon in agriculture; Datnoff, L.E., Snyder, G.H., Korndörfer, G.H., Eds.; Elsevier, 2001; Vol. 8, pp. 115–129. [Google Scholar]
- Sonneveld, C.; Voogt, W. Plant Nutrition of Greenhouse Crops, 1st ed.; Springer Dordrecht, 2009; ISBN 9789048125326. [Google Scholar]
- Jones, L.H.P.; Handreck, K.A. Silica In Soils, Plants, and Animals. Advances in Agronomy 1967, 19, 107–149. [Google Scholar] [CrossRef]
- da Silva, M.N.; Machado, J.; Osorio, J.; Duarte, R.; Santos, C.S. Non-Essential Elements and Their Role in Sustainable Agriculture. Agronomy 2022, 12, 888. [Google Scholar] [CrossRef]
- Zargar, S.M.; Mahajan, R.; Bhat, J.A.; Nazir, M.; Deshmukh, R. Role of Silicon in Plant Stress Tolerance: Opportunities to Achieve a Sustainable Cropping System. 3 Biotech 2019, 9. [Google Scholar] [CrossRef]
- Rodrigues, F.Á.; Dallagnol, L.J. Silicon and Plant Disease Control. In Silicon Advances for Sustainable Agriculture and Human Health; de Mello Prado, R., Etesami, H., Srivastava, A.K., Eds.; Springer: Cham, 2024; pp. 381–413. ISBN 978-3-031-69876-7. [Google Scholar]
- Liang, Y.C.; Sun, W.C.; Si, J.; Römheld, V. Effects of Foliar- and Root-Applied Silicon on the Enhancement of Induced Resistance to Powdery Mildew in Cucumis Sativus. Plant Pathol 2005, 54, 678–685. [Google Scholar] [CrossRef]
- Menzies, J.; Bowen, P.; Ehret, D.; Glass, A.D.M. Foliar Applications of Potassium Silicate Reduce Severity of Powdery Mildew on Cucumber, Muskmelon, and Zucchini Squash. Journal of the American Society for Horticultural Science 1992, 117, 902–905. [Google Scholar] [CrossRef]
- Bi, Y.; Tian, S.P.; Guo, Y.R.; Ge, Y.H.; Qin, G.Z. Sodium Silicate Reduces Postharvest Decay on Hami Melons: Induced Resistance and Fungistatic Effects. Plant Dis 2006, 90, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Mohaghegh, P.; Khoshgoftarmanesh, A.H.; Shirvani, M.; Sharifnabi, B.; Nili, N. Effect of Silicon Nutrition on Oxidative Stress Induced by Phytophthora Melonis Infection in Cucumber. Plant Dis 2011, 95, 455–460. [Google Scholar] [CrossRef]
- Chérif, M.; Benhamou, N.; Menzies, J.G.; Bélanger, R.R. Silicon Induced Resistance in Cucumber Plants against Pythium Ultimum. Physiol Mol Plant Pathol 1992, 41, 411–425. [Google Scholar] [CrossRef]
- Kim, S.G.; Kim, K.W.; Park, E.W.; Choi, D. Silicon-Induced Cell Wall Fortification of Rice Leaves: A Possible Cellular Mechanism of Enhanced Host Resistance to Blast. Phytopathology 2007, 92, 1095–1103. [Google Scholar] [CrossRef]
- Rodrigues, F.A.; Resende, R.S.; Dallagnol, L.J.; Datnoff, L.E. Silicon Potentiates Host Defense Mechanisms Against Infection by Plant Pathogens. Silicon and Plant Diseases 2015, 109–138. [Google Scholar] [CrossRef]
- Fauteux, F.; Rémus-Borel, W.; Menzies, J.G.; Bélanger, R.R. Silicon and Plant Disease Resistance against Pathogenic Fungi. FEMS Microbiol Lett 2005, 249, 1–6. [Google Scholar] [CrossRef]
- Vivancos, J.; Labbé, C.; Menzies, J.G.; Bélanger, R.R. Silicon-Mediated Resistance of Arabidopsis against Powdery Mildew Involves Mechanisms Other than the Salicylic Acid (SA)-Dependent Defence Pathway. Mol Plant Pathol 2015, 16, 572–582. [Google Scholar] [CrossRef]
- Islam, W.; Tayyab, M.; Khalil, F.; Hua, Z.; Huang, Z.; Chen, H.Y.H. Silicon-Mediated Plant Defense against Pathogens and Insect Pests. Pestic Biochem Physiol 2020, 168, 104641. [Google Scholar] [CrossRef]
- Sahebi, M.; Hanafi, M.M.; Siti Nor Akmar, A.; Rafii, M.Y.; Azizi, P.; Tengoua, F.F.; Nurul Mayzaitul Azwa, J.; Shabanimofrad, M. Importance of Silicon and Mechanisms of Biosilica Formation in Plants. Biomed Res Int 2015, 1, 396010. [Google Scholar] [CrossRef]
- Ma, J.F.; Miyake, Y.; Takahashi, E. Silicon as a Beneficial Element for Crop Plants. In Silicon in Agriculture; Datnoff, L.E., Snyder, G.H., Korndörfer, G.H., Eds.; Elsevier, 2001; Vol. 8, pp. 17–39. [Google Scholar]
- Coskun, D.; Deshmukh, R.; Sonah, H.; Menzies, J.G.; Reynolds, O.; Ma, J.F.; Kronzucker, H.J.; Bélanger, R.R. The Controversies of Silicon’s Role in Plant Biology. New Phytologist 2019, 221, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Tesfagiorgis, H.; Laing, M.D.; Tesfagiorgis, H.B.; Laing, M.D. Effects of Concentration, Frequency of Application and Runoff of Foliar-Applied Soluble Silicon on Powdery Mildew of Zucchini. Afr J Agric Res 2011, 6, 2243–2248. [Google Scholar] [CrossRef]
- Savvas, D.; Giotis, D.; Chatzieustratiou, E.; Bakea, M.; Patakioutas, G. Silicon Supply in Soilless Cultivations of Zucchini Alleviates Stress Induced by Salinity and Powdery Mildew Infections. Environ Exp Bot 2009, 65, 11–17. [Google Scholar] [CrossRef]
- Spencer, D.M. Standardized Methods for the Evaluation of Fungicides to Control Cucumber Powdery Mildew. In Crop protection agents – their biological evaluation.; Academic Press: London, 1977; pp. 455–464. [Google Scholar]
- Lepolu Torlon, J.; Heckman, J.; Simon, J.; Wyenandt, C. Silicon Soil Amendments for Suppressing Powdery Mildew on Pumpkin. Sustainability 2016, 8, 293. [Google Scholar] [CrossRef]
- Vercelli, M.; Minuto, A.; Minuto, G.; Contartese, V.; Devecchi, M.; Larcher, F. The Effects of Innovative Silicon Applications on Growth and Powdery Mildew Control in Soilless-Grown Cucumber (Cucumis Sativus L.) and Zucchini (Cucurbita Pepo L.). Acta Physiol Plant 2017, 39, 1–6. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. A Cooperative System of Silicon Transport in Plants. Trends Plant Sci 2015, 20, 435–442. [Google Scholar] [CrossRef]
- Laane, H.M. The Effects of Foliar Sprays with Different Silicon Compounds. Plants 2018, 7, 45. [Google Scholar] [CrossRef]
- Guerriero, G.; Hausman, J.F.; Legay, S. Silicon and the Plant Extracellular Matrix. Front Plant Sci 2016, 7, 463. [Google Scholar] [CrossRef]
- Ghosh, R.; Ganguly, S. Silicon Fertilizers Inciting Plant Defence against Pathogenic Fungi. J. Mycopathol. Res. 2022, 60, 515–521. [Google Scholar] [CrossRef]
- Kumar, A.; Choudhary, A.; Kaur, H.; Singh, K.; Guha, S.; Choudhary, D.R.; Sonkar, A.; Mehta, S.; Husen, A. Exploring the Role of Silicon in Enhancing Sustainable Plant Growth, Defense System, Environmental Stress Mitigation and Management. Discover Applied Sciences 2025, 7, 406. [Google Scholar] [CrossRef]
- Khursheed, S.; Bashir Surma, S.; Rahman Mohammad Al Tawaha, A.; Shah, M.D.; A. Padder, B.; Afroza, B.; Rashid, R.; Al-Tawaha, A.R. Silicon’s Defensive Role against Biotic and Abiotic Stress: A Review. Phosphorus Sulfur Silicon Relat Elem 2025, 200, 325–344. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Yang, Y. Mechanisms of Silicon-Induced Fungal Disease Resistance in Plants. Plant Physiology and Biochemistry 2021, 165, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Harizanova, A. Silicon Application Unveiled: A Review of Insights into Plant Defense Mechanisms under Biotic Challenges. Agricultural Sciences 2024, 16, 39–56. [Google Scholar] [CrossRef]
- Sangster, A.G.; Hodson, M.J.; Tubb, H.J. Silicon Deposition in Higher Plants. In Silicon in Agriculture; Datnoff, L.E., Snyder, G.H., Korndörfer, G.H., Eds.; Elsevier, 2001; Vol. 8, pp. 85–113. [Google Scholar]
- Thakral, V.; Raturi, G.; Sudhakaran, S.; Mandlik, R.; Sharma, Y.; Shivaraj, S.M.; Tripathi, D.K.; Sonah, H.; Deshmukh, R. Silicon, a Quasi-Essential Element: Availability in Soil, Fertilizer Regime, Optimum Dosage, and Uptake in Plants. Plant Physiology and Biochemistry 2024, 208, 108459. [Google Scholar] [CrossRef]
- Reynolds, O.L.; Keeping, M.G.; Meyer, J.H. Silicon-augmented Resistance of Plants to Herbivorous Insects: A Review. Annals of Applied Biology 2009, 155, 171–186. [Google Scholar] [CrossRef]
- de Oliveira, R.S.; Sampaio, M.V.; Carvalho, F.J.; Albuquerque, C.J.B.; Korndorfer, G.H. Silicon Amendments Reduce Aphid Numbers and Improve Yield of Aphid-Resistant and Susceptible Wheat Cultivars in a Dose-Dependent Manner. Crop Protection 2023, 172, 106296. [Google Scholar] [CrossRef]
- Santos, D.G.; Dias, L.L.C.; Avellar, G.S.; Simeone, M.L.F.; Parrella, R.A.C.; Santos, N.M.; Silva, T.F.; Neto, A.A.; Mendes, S.M. Biomass Sorghum (Sorghum Bicolor) Agronomic Response to Melanaphis Sorghi (Hemiptera: Aphididae) Infestation and Silicon Application. Insects 2025, 16, 566. [Google Scholar] [CrossRef] [PubMed]
- Leroy, N.; de Tombeur, F.; Walgraffe, Y.; Cornélis, J.-T.; Verheggen, F.J.; Leroy, N.; de Tombeur, F.; Walgraffe, Y.; Cornélis, J.-T.; Verheggen, F.J. Silicon and Plant Natural Defenses against Insect Pests: Impact on Plant Volatile Organic Compounds and Cascade Effects on Multitrophic Interactions. Plants 2019, 8, 444. [Google Scholar] [CrossRef]
- Frew, A.; Weston, L.A.; Gurr, G.M. Silicon Reduces Herbivore Performance via Different Mechanisms, Depending on Host–Plant Species. Austral Ecol 2019, 44, 1092–1097. [Google Scholar] [CrossRef]
Figure 1.
Reduction in powdery mildew severity scores in zucchini following silicon treatments via fertigation and foliar sprays in 2024. Silicon concentrations are expressed in mM (mmol/L). Data are presented as means ± SE (n=20). Statistical analysis was performed using R with a cumulative link mixed model (clmm) for ordinal data. Treatments differed significantly (p < 0.001 (***)), and different letters indicate statistically significant differences between treatments.
Figure 1.
Reduction in powdery mildew severity scores in zucchini following silicon treatments via fertigation and foliar sprays in 2024. Silicon concentrations are expressed in mM (mmol/L). Data are presented as means ± SE (n=20). Statistical analysis was performed using R with a cumulative link mixed model (clmm) for ordinal data. Treatments differed significantly (p < 0.001 (***)), and different letters indicate statistically significant differences between treatments.
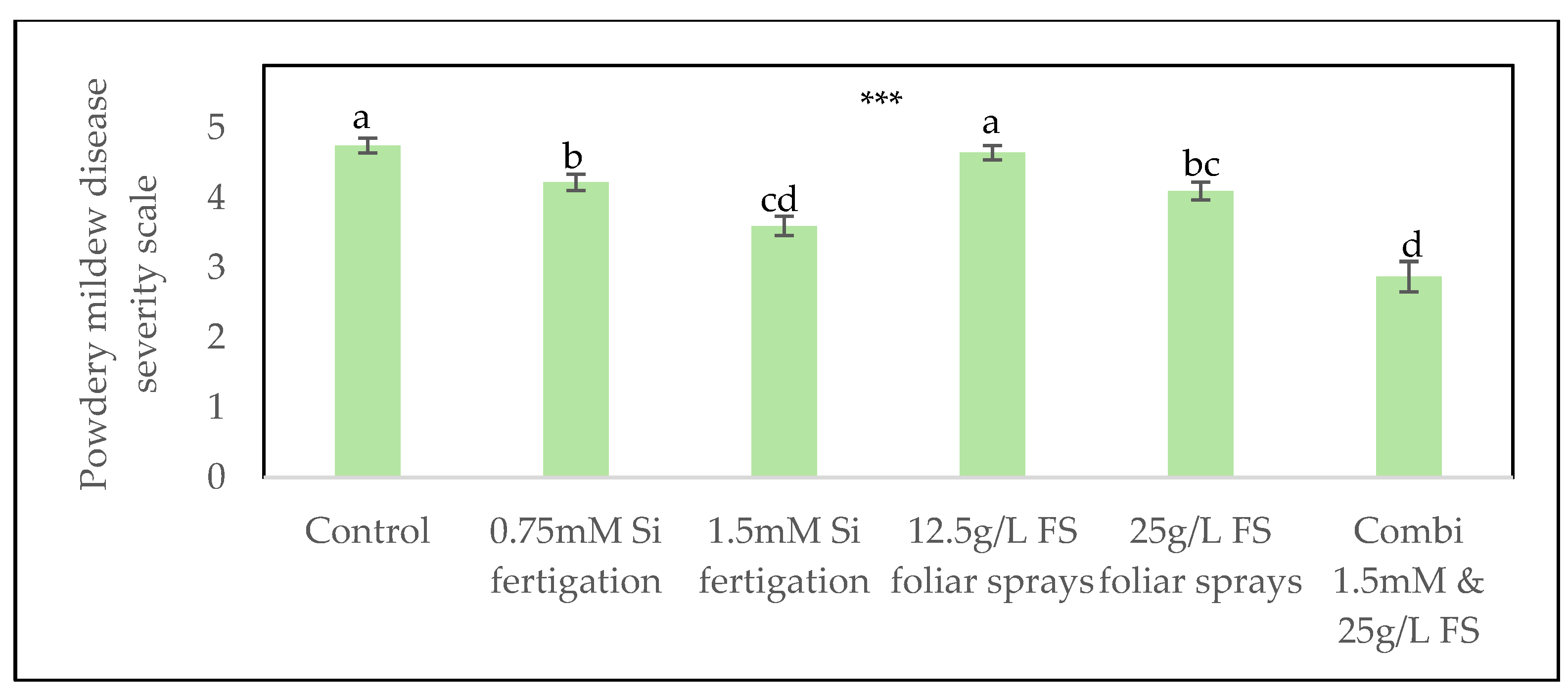
Figure 2.
Reduction in powdery mildew coverage (%) on inoculated zucchini leaves following silicon treatments via fertigation and foliar sprays in 2025. Silicon concentrations are expressed in mM (mmol/L). Data are presented as means ± SE (n=20) Statistical analysis was performed using R with a beta mixed model (glmmTMB) using a logit link. Treatments differed significantly (p < 0.001 (***)), and different letters indicate statistically significant differences between treatments.
Figure 2.
Reduction in powdery mildew coverage (%) on inoculated zucchini leaves following silicon treatments via fertigation and foliar sprays in 2025. Silicon concentrations are expressed in mM (mmol/L). Data are presented as means ± SE (n=20) Statistical analysis was performed using R with a beta mixed model (glmmTMB) using a logit link. Treatments differed significantly (p < 0.001 (***)), and different letters indicate statistically significant differences between treatments.
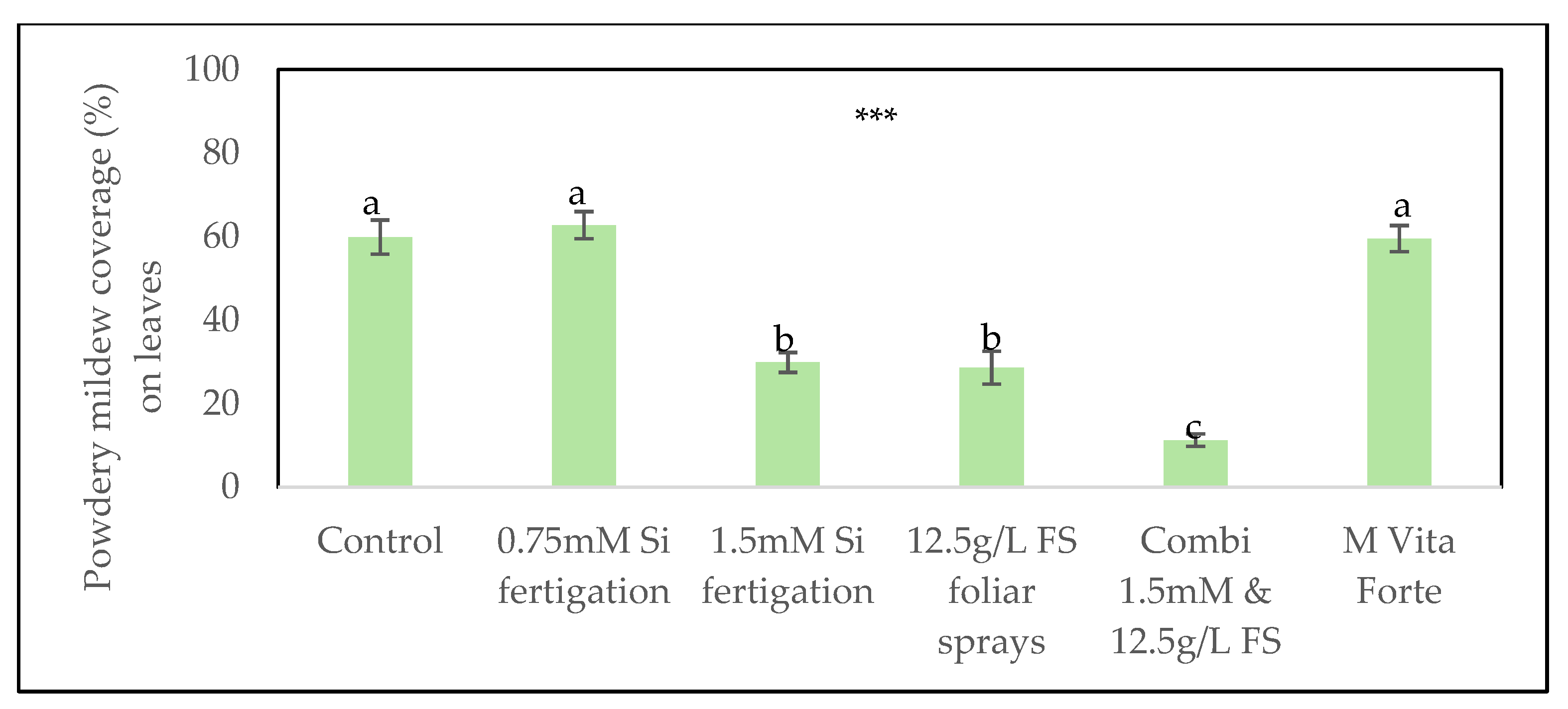
Figure 3.
Powdery mildew symptoms on zucchini plants in 2025: (a) control plant without silicon application and (b) plant treated with the combined application of 1.5 mmol/L Si via fertigation and 12.5 g/L FS foliar sprays treatment.
Figure 3.
Powdery mildew symptoms on zucchini plants in 2025: (a) control plant without silicon application and (b) plant treated with the combined application of 1.5 mmol/L Si via fertigation and 12.5 g/L FS foliar sprays treatment.

Figure 4.
Reduction in the number of naturally infected zucchini leaves following silicon treatments in 2025. Silicon concentrations are expressed in mM (mmol/L). Data are presented as means ± SE (n=20) Statistical analysis was performed in R using a Poisson generalized linear mixed model (glmer). Treatments differed significantly (p < 0.001 (***)), and different letters indicate statistically significant differences between treatments.
Figure 4.
Reduction in the number of naturally infected zucchini leaves following silicon treatments in 2025. Silicon concentrations are expressed in mM (mmol/L). Data are presented as means ± SE (n=20) Statistical analysis was performed in R using a Poisson generalized linear mixed model (glmer). Treatments differed significantly (p < 0.001 (***)), and different letters indicate statistically significant differences between treatments.
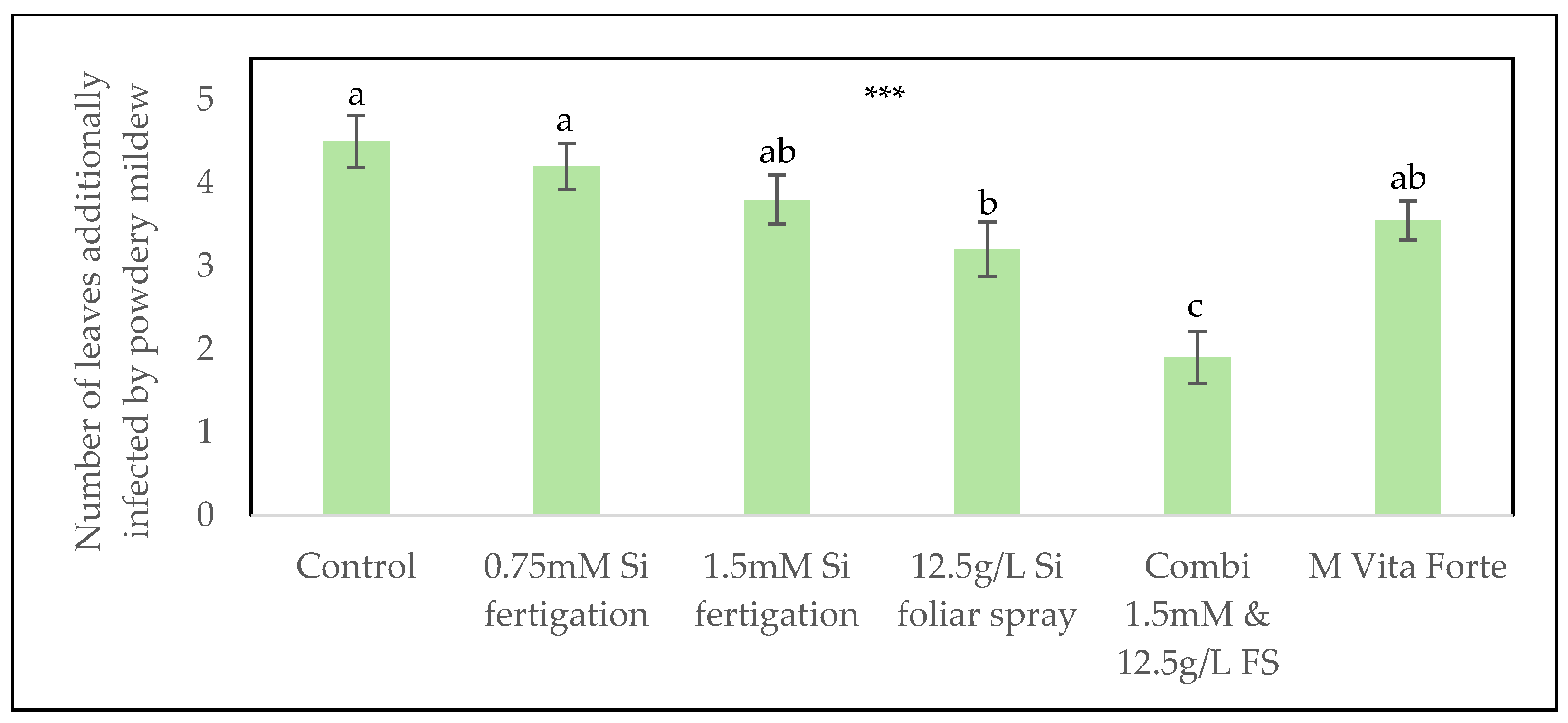
Figure 5.
Representative microscopic images showing average cell wall thickness in (a) the control treatment without silicon; (b) in the combined application of 1.5 mmol/L Si via fertigation and 12.5 g/L FS foliar sprays treatment. Panels show cropped images highlighting individual zucchini plant cells. Full, uncropped micrographs are provided in the Supplementary Material.
Figure 5.
Representative microscopic images showing average cell wall thickness in (a) the control treatment without silicon; (b) in the combined application of 1.5 mmol/L Si via fertigation and 12.5 g/L FS foliar sprays treatment. Panels show cropped images highlighting individual zucchini plant cells. Full, uncropped micrographs are provided in the Supplementary Material.
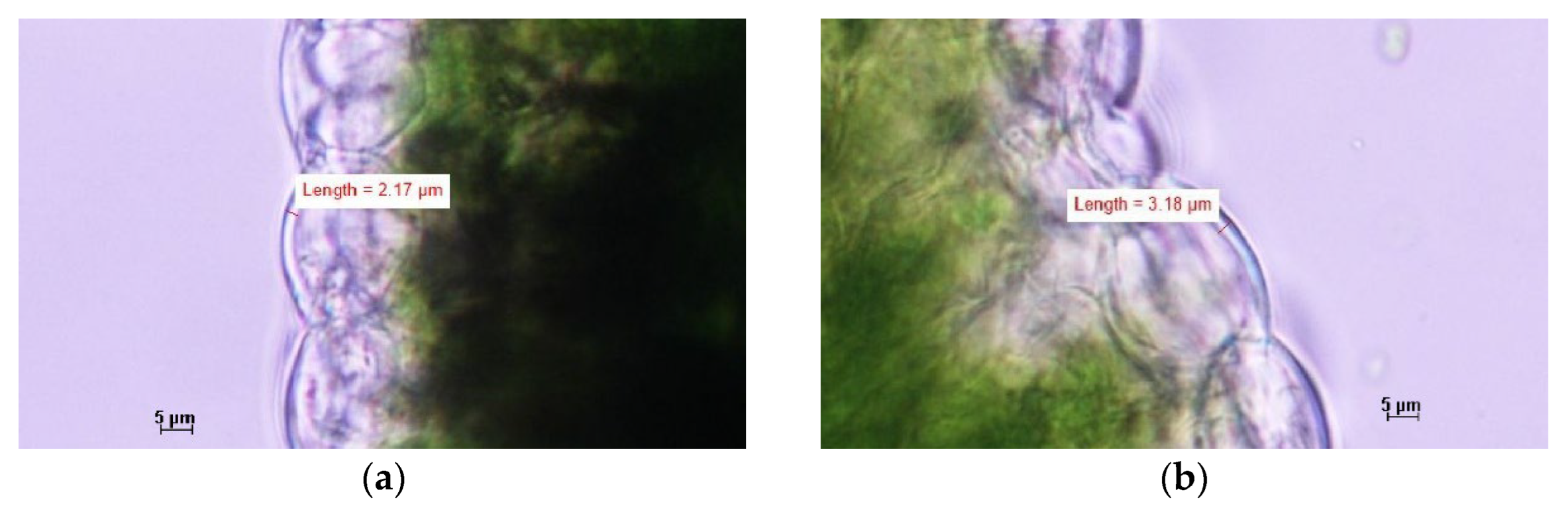
Figure 6.
Comparison of cell wall thickness in leaf samples from the control treatment without silicon and the combined application of 1.5 mmol/L Si via fertigation and 12.5 g/L FS foliar sprays treatment. Data are presented as means ± SE (n = 30). Statistical analysis was performed using a linear mixed model (lmer). Significant difference among treatments are indicated (***p < 0.001).
Figure 6.
Comparison of cell wall thickness in leaf samples from the control treatment without silicon and the combined application of 1.5 mmol/L Si via fertigation and 12.5 g/L FS foliar sprays treatment. Data are presented as means ± SE (n = 30). Statistical analysis was performed using a linear mixed model (lmer). Significant difference among treatments are indicated (***p < 0.001).
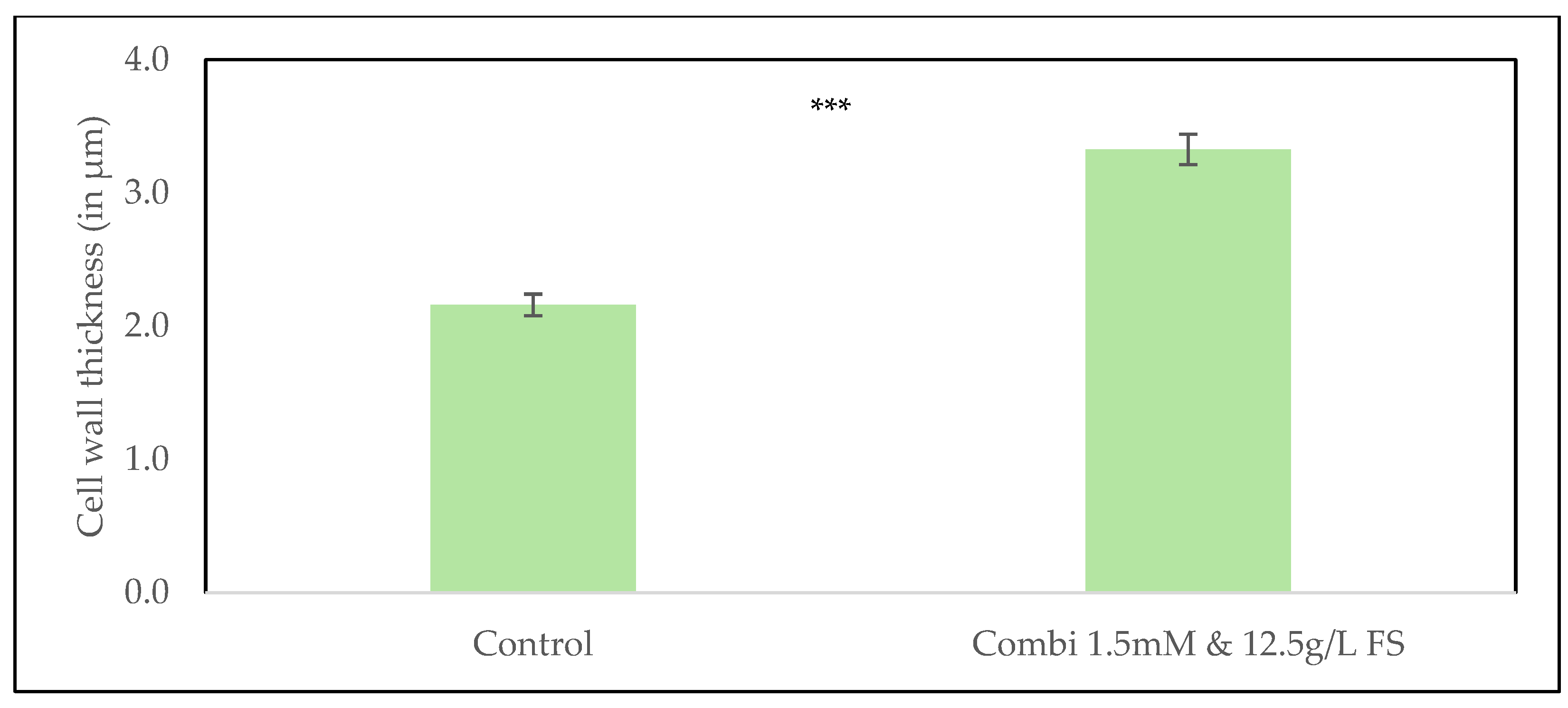
Figure 7.
Silicon concentration in zucchini plant sap in young and fully-grown leaves under various Si treatments in 2024. Both 1.5 mmol/L Si fertigation and the combined treatment of fertigation with foliar sprays resulted in increased Si accumulation compared to the control, with the highest accumulation in fully-grown leaves.
Figure 7.
Silicon concentration in zucchini plant sap in young and fully-grown leaves under various Si treatments in 2024. Both 1.5 mmol/L Si fertigation and the combined treatment of fertigation with foliar sprays resulted in increased Si accumulation compared to the control, with the highest accumulation in fully-grown leaves.
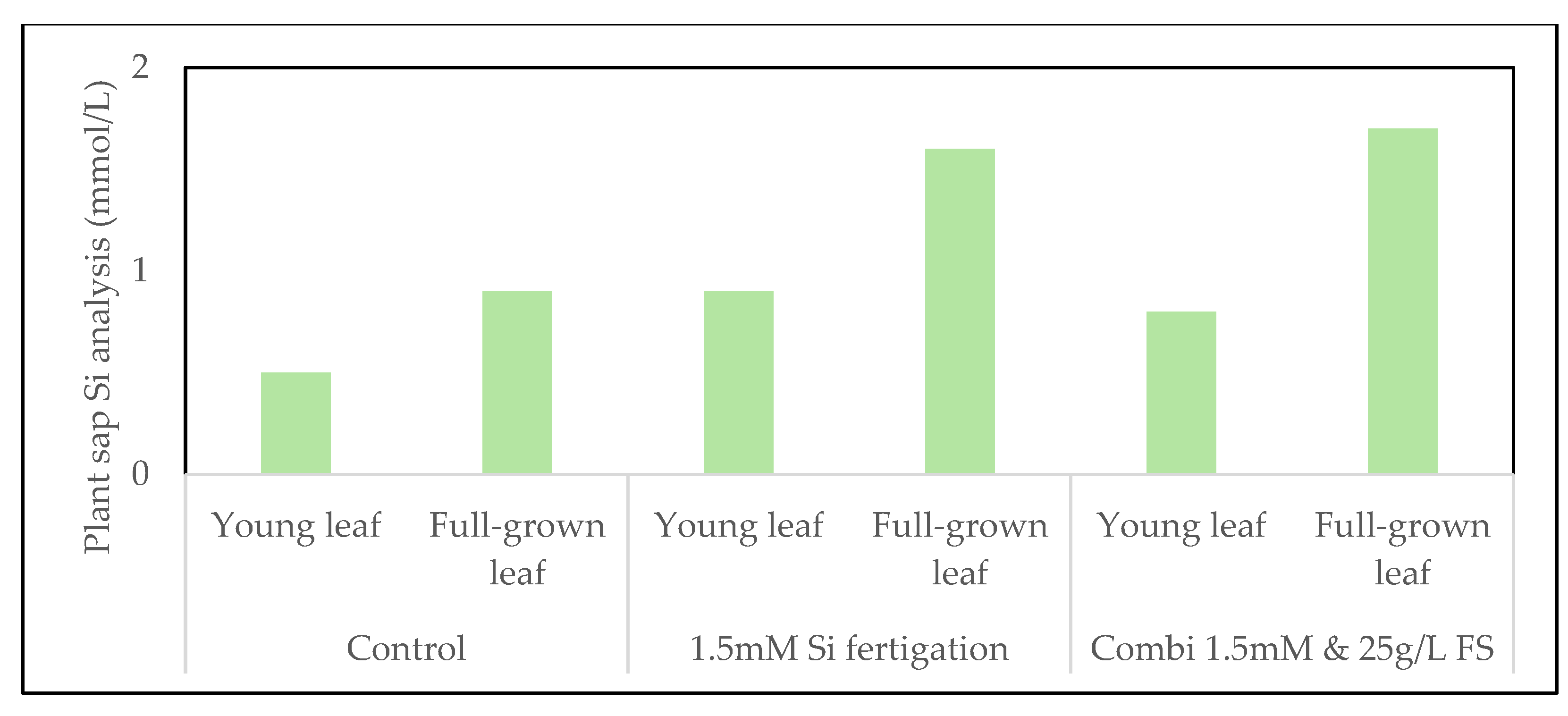
Figure 8.
Silicon concentration in zucchini plant sap, of composite (pooled) samples with young and fully-grown leaves combined, under various Si treatments in 2025. Silicon concentrations are expressed in mM (mmol/L). Fertigation treatments, as well as the combination of fertigation and foliar sprays, markedly increased Si uptake compared to the control.
Figure 8.
Silicon concentration in zucchini plant sap, of composite (pooled) samples with young and fully-grown leaves combined, under various Si treatments in 2025. Silicon concentrations are expressed in mM (mmol/L). Fertigation treatments, as well as the combination of fertigation and foliar sprays, markedly increased Si uptake compared to the control.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



