Submitted:
19 December 2025
Posted:
22 December 2025
You are already at the latest version
Abstract
Metachromatic leukodystrophy (MLD) results from arylsulfatase A (ARSA) deficiency and progressive demyelination. This study evaluates the safety and therapeutic potential of intravenously administered allogeneic mesenchymal stem cells transduced with AAV9 encoding human ARSA in a porcine in vivo study. While ARSA activity in plasma and cerebrospinal fluid did not significantly change, CNS tissues showed a marked increase in ARSA activity, indicating successful CNS targeting and local enzyme expression. Biochemical parameters and cytokine profiles remained within physiological ranges, demonstrating good tolerability and absence of systemic inflammation. These findings suggest that MSC-based delivery of AAV9-ARSA is a safe approach capable of enhancing ARSA activity in the CNS and may represent a promising therapeutic strategy for MLD.
Keywords:
metachromatic leukodystrophy
; gene cell therapy
; mesenchymal stem cells
1. Introduction
Metachromatic leukodystrophy (MLD) is an autosomal recessive neurodegenerative disease belonging to the group of lysosomal storage disorders (LSDs). The pathogenesis of the disease is caused by a deficiency of the lysosomal enzyme arylsulfatase A (ARSA), leading to progressive demyelination of nerve fibers in the central (CNS) and peripheral nervous system (PNS). Clinically, this manifests as progressively worsening motor and cognitive impairment [1]. MLD is one of the most common leukodystrophies, with a global incidence of 1:40,000–160,000. In LSDs, undegraded macromolecules accumulate inside lysosomes and eventually spread to other cellular compartments. Disease progression and lysosomal dysfunction lead to secondary deficiencies of other enzymes, ultimately resulting in complete disruption of lysosomal function and cell death [2].
Despite advances in gene therapy, effective treatments for MLD remain limited. In 2024, the U.S. Food and Drug Administration (FDA) approved the first therapy of this kind — Lenmeldy™ (atidarsagene autotemcel, Orchard Therapeutics) is an autologous genetically modified hematopoietic stem cell (HSC) therapy with overexpression of the ARSA gene. A key limitation of this approach is the slow migration of transplanted HSCs to the CNS and their subsequent differentiation into ARSA-expressing macrophages and microglia. As a result, the clinical efficacy of the therapy is demonstrated mainly when used at the presymptomatic stage of the disease [3].
Direct gene therapy using viral vectors represents a promising approach for MLD treatment. This strategy aims to compensate for the deficiency of functional proteins — in this case, the ARSA enzyme. Among various viral vectors, adeno-associated viruses (AAV) have gained wide use due to their relative safety and ability to infect non-dividing cells, including neurons and glial cells of the CNS [4].
For effective CNS gene delivery, AAV serotypes with strong neurotropism are commonly used, including AAV1, 5, 8, 9, and rh10 [5]. In particular, AAV9 demonstrates high efficiency in crossing the blood–brain barrier (BBB) after systemic administration [6]. Promising results were obtained in preclinical animal studies: intravenous administration of AAV9 encoding human ARSA increased enzyme activity and improved phenotype in both MLD mouse models [7] and a more relevant large-animal model — mini-pigs [8].
Despite its potential, direct intrathecal or systemic injection of recombinant AAV has several limitations, including immune responses to the viral capsid, limited specificity for target cells, small genome capacity that restricts packaging of large genes, and potentially short-lived transgene expression due to the lack of genomic integration [9]. These limitations highlight the need for new delivery strategies combining the efficiency of AAV with prolonged activity and enhanced targeting.
An alternative and promising approach is cell therapy, particularly using mesenchymal stem cells (MSCs). MSCs are multipotent progenitor cells with strong immunomodulatory properties and the ability to migrate to sites of tissue damage (the “tissue homing” phenomenon). Key advantages of MSCs include low immunogenicity, enabling the use of allogeneic transplants, and the ability to secrete a broad range of bioactive factors [10]. Upon entering the pathological region, MSCs integrate into tissue and exert therapeutic effects through paracrine secretion of neurotrophins, cytokines, and chemokines. For example, in models of neurodegenerative diseases such as Alzheimer’s and Parkinson’s, MSCs secrete brain-derived neurotrophic factor (BDNF) and vascular endothelial growth factor (VEGF), supporting neuronal survival and synaptic plasticity [11].
The combination of these two platforms — AAV-based gene delivery and the unique biological properties of MSCs — forms the basis of combined gene–cell therapeutic strategies. One such approach involves ex vivo genetic modification of MSCs followed by transplantation. These engineered cells retain their homing ability, allowing them to migrate directly to lesions and act as biological “microfactories” that continuously secrete the therapeutic protein [10]. As proof of concept, human umbilical cord MSCs transduced with self-complementary AAV carrying the gene for coagulation factor IX (FIX) demonstrated stable secretion of functional protein. The plasma FIX levels achieved were comparable to direct AAV injection but required significantly lower viral doses, indicating improved gene-delivery efficiency via cellular carriers [12].
Thus, the synergy of two technologies — long-term transgene expression provided by AAV and targeted tissue tropism combined with the immunomodulatory properties of MSCs — opens new prospects for MLD therapy. This study aims to evaluate the efficacy and safety of allogeneic MSCs transduced with AAV9 encoding the ARSA gene following intravenous administration to pigs.
2. Materials and Methods
2.1. Isolation and Genetic Modification of Mesenchymal Stem Cells
MSCs were isolated from pig adipose tissue as described previously [13] using enzymatic digestion of adipose tissue with 0.2% crab collagenase (1.4.8.1., Biolot, Russia) at 37 °C for 1 hour. Red blood cells were removed using RBC lysis buffer (420301, BioLegend, USA). The isolated cells expressed markers characteristic of pig MSCs and were capable of adipogenic, chondrogenic, and osteogenic differentiation. Detailed characterization is provided in [13].
Species-specific allogeneic porcine MSCs were genetically modified with recombinant AAV9 encoding the ARSA gene cDNA (AAV9-ARSA) at a multiplicity of infection (MOI) of 100 (100 viral particles per cell). Twelve hours after viral exposure, cells were washed with saline by centrifugation and resuspended in saline at the required volume. Functionality of AAV9-ARSA in vitro has been shown in our prior publication [8].
2.2. Laboratory Animals and Housing
The animal study was reviewed and approved by the local ethics committee of Kazan Federal University (No. 23, June 30, 2020). Healthy male pigs aged 4 months (weighing 9–12 kg) were used in the study. The experimental pigs (10 individuals) were divided into two groups: (1) intravenous administration of allogeneic MSCs genetically modified with AAV9-ARSA (MSC+AAV9-ARSA) at a dose of 2.7 million cells/kg; (2) control group of animals — without administration of MSC+AAV9-ARSA.
The pigs were kept in specialized facilities at the Kazan State Academy of Veterinary Medicine named after N.E. Bauman (KSAM) under the supervision of qualified personnel. The animals were euthanized using methods that comply with the principles set out in the European Commission’s Recommendations on the Euthanasia of Experimental Animals.
2.3. Material Collection
Before the introduction of the virus and cells, samples of cerebrospinal fluid (CSF) and blood were collected as baseline samples. At 7, 14, 21, 28, and 35 days after the introduction of MSC+AAV9-ARSA, CSF and blood samples were also collected for analysis of ARSA enzymatic activity over time. On day 35 after the introduction of MSC+AAV9-ARSA, the pigs were euthanized to determine the enzymatic activity of ARSA in homogenates of the nervous system organs. Fragments of the following organs were taken from each animal: cerebral cortex, cerebellum, spinal cord cervical (C6-7), thoracic (Th6-7), and lumbar (L2-3) sections of the spinal cord.
2.4. Confirmation of ARSA Enzymatic Activity
The enzymatic activity of ARSA was determined in plasma, CSF, and organ homogenates. Plasma was isolated from whole blood by centrifugation at 553 g for 20 min. The organs were homogenized in 500 μl of sodium acetate buffer with the addition of glass beads, and the Eppendorf tubes were placed in a homogenizer. They were treated for 20 seconds at a frequency of 5 Hz. The homogenate was then centrifuged for 5 min at 10,000 g, and the supernatant was used to determine ARSA activity.
The total protein concentration in the samples was determined using the Pierce™ BCA Protein Assay Kit (23227, ThermoFisher Scientific Inc., USA). The samples were normalized relative to the total protein concentration. To determine ARSA activity, 50 μl of the sample was incubated with a solution of the substrate nitrocatechol sulfate (0.01M p-Nitrocatechol sulfate dipotassium salt (ρ.NCS) (N7251, Sigma-Aldrich, USA), 0.5 M sodium acetate, 5 × 10-4 Na4P2O7, 10% sodium chloride, pH = 5) for 1 hour at 37 °C, after which the reaction was stopped by adding 1 N sodium hydroxide. Sulphatase dilutions (S9626, Sigma-Aldrich, USA) were used as standards. The optical density was measured at a wavelength of 515 nm. Enzymatic activity was calculated using the formula:
Where:
1 — incubation time (h);
0.25 — total reaction volume (mL);
12.6 — absorption coefficient of p.NCS at 515 nm;
0.05 — sample volume (mL).
Conversion to nmol/mL:
Conversion to nmol/mg protein:
2.5. Safety Assessment
Biochemical blood tests were performed to assess safety. For biochemical testing, whole blood was collected from animals into tubes containing gel and a blood coagulation activator. They were then centrifuged for 20 minutes at 553 g. Serum levels of aspartate aminotransferase (AST), alanine aminotransferase (ALT), total bilirubin, and creatinine were determined using a ChemWell 2900 biochemical analyzer (USA).
2.6. Cytokine Profile Analysis
The MILLIPLEX MAP Porcine Cytokine/Chemokine Magnetic Bead Panel - Immunology Multiplex Assay (Cat. No. PCYTMG-23K-13PX, Merck, Germany) was used to analyze the pig blood serum samples. The kit allows the following analytes to be measured: interleukin (IL) 1α, IL-1β, IL-1ra (IL-1 antagonist), IL-2, IL-4, IL-6, IL-8, IL-10, IL-12, IL-18, tumor necrosis factor (TNF) α. Fifty microliters of sample were used to determine the concentration of analytes, and the data were analyzed using a Luminex 200 analyzer (Immucor Transplant Diagnostics, Inc., USA) with MasterPlex CT control software and MasterPlex QT analysis software (MiraiBio, a division of Hitachi Software, USA).
2.7. Statistical Analysis
The data obtained were analyzed using GraphPad Prism 9 software (GraphPad Software) with the Shapiro-Wilk test and one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test. The Student’s t-test was used to evaluate the differences between the two groups. Statistically significant differences are indicated as * — p <0.05, ** — p <0.01, *** — p <0.001, **** — p <0.0001.
3. Results
After intravenous administration of MSC+AAV9-ARSA, the enzymatic activity of ARSA was measured dynamically in plasma and CSF. No statistically significant difference in the enzymatic activity of ARSA in plasma and CSF of animals from different groups was found (Figure 1).
Analysis of enzymatic activity in CNS homogenates showed an increase in ARSA enzymatic activity compared to the control group of animals. An increase in ARSA enzymatic activity was observed in the cortex, where activity increased by 206%, reaching a value of 99.12 ± 31.02 nM/mg/18 hours compared to the control (48.56 ± 0.79 nM/mg/18 hours). In the cerebellum, enzymatic activity increased by 205% (from 51.82 ± 0.26 nM/mg/18 hours to 105.14 ± 18.2 nM/mg/18 hours). In the cervical, thoracic, and lumbar sections of the spinal cord, a significant increase in activity was also recorded by 237% (from 106.1 ± 4.36 nM/mg/18 hours to 252.9 ± 71.49 nM/mg/18 hours), 383% (from 71.9 ± 2.9 nM/mg/18 hours to 276.3 ± 8.87 nM/mg/18 hours) and 337%, respectively (from 75.9 ± 3.5 nM/mg/18 hours to 253.2 ± 38.5 nM/mg/18 hours) (Figure 2).
Changes in biochemical parameters in the blood serum of pigs after administration of MSC transduced with AAV9-ARSA were studied (Figure 3). ALT dynamics ranged from 46 to 57 U/L, with no statistically significant increase indicating liver damage. AST values ranged from 46 to 97 U/L, with an increase observed on day 7, but no statistically significant increase. Serum creatinine concentration remained stable in the range of 0.6 to 0.73 mg/dL at all stages of the study. It should be noted that some of the creatinine values in the test animals were below the reference range (~0.70–1.85 mg/dL) [14]. The key argument in favor of this not being a consequence of therapy is the absence of both statistically significant changes and any growth dynamics of this indicator during the experiment. This indicates that the observed low level is a background characteristic of a specific group of animals. The most likely cause is age-related features. The total bilirubin level ranged from 0.4 to 1.4 mg/dL at all stages of the study. On day 35, there was a statistically significant decrease in its concentration compared to the baseline level. However, the values obtained were completely within the physiological range (0.2–1.0 mg/dL), indicating no clinically significant changes in this indicator.
A comparative analysis of cytokine and chemokine concentrations in blood serum did not reveal any statistically significant differences between the indicators recorded before administration and at various time points after it. No significant increase in the levels of key pro-inflammatory cytokines and chemokines was detected, indicating the absence of a systemic inflammatory response to the administration of MSC+AAV9-ARSA (Figure 4).
4. Discussion
Currently, promising approaches for treating MLD include gene and gene-cell therapy. Gene-cell therapy uses genetically modified cells transduced using retro- and lentiviruses. In 2024, the FDA approved a gene-cell drug called Libmeldy for transplantation in patients with presymptomatic or very early symptomatic CD34+ HSC transduced with a lentiviral vector encoding ARSA. The use of lentiviruses can potentially lead to the oncological transformation of genetically modified cells due to insertional mutagenesis, which is also a limiting factor in therapy [15].
As part of this study, we analyzed the functional activity and safety of administering allogeneic mesenchymal stem cells transduced with an adeno-associated virus encoding the cDNA of the human ARSA gene to pigs. Previous studies have demonstrated the functionality of the vector construct in vitro [8].
Our results demonstrated the effectiveness of allogeneic transduced MSCs and revealed no side effects, which is consistent with the literature data on the safety of allogeneic MSC therapy. In patients with MSD, after hematopoietic cell transplantation followed by infusion of allogeneic MSCs, rapid cell engraftment without significant toxicity and complete stabilization of neurological symptoms were observed. Clinical case data showed that combination therapy with allogeneic MSCs is safe and can halt disease progression [16].
Analysis of enzymatic activity did not reveal a statistically significant increase in ARSA enzyme activity in blood plasma or CSF after administration of MSC+AAV9-ARSA. At the same time, an increase in ARSA was shown in homogenates of the CNS. This indicates that transduced MSCs successfully penetrated the CNS and carried out local expression of the active enzyme. Such observations confirm the ability of MSCs to cross the BBB and migrate into nervous tissue [17].
Safety analysis of the therapy showed no hepatotoxicity or systemic inflammatory response. Biochemical blood analysis revealed no pathological shifts in liver enzymes—AST, ALT, and bilirubin levels remained normal in all groups. This is consistent with one of the key advantages of MSCs — their low immunogenicity [18]. The absence of statistically significant changes in the cytokine profile of blood serum indicates good tolerability of MSC+AAV9-ARSA administration and a low risk of inducing a systemic inflammatory response. This may be due to the known immunomodulatory properties of MSCs, which are capable of suppressing the activation of the immune system. Our results indicate good tolerability and efficacy of the method.
Thus, we have demonstrated the safety and efficacy of combined therapy for MLD using allogeneic MSCs transduced with AAV9-ARSA. The absence of systemic toxicity and a significant increase in ARSA enzymatic activity in CNS tissues confirm the therapeutic potential of the developed method.
Author Contributions
A.I.A. and A.A.F. contributed to the investigation and the writing of the original draft. V.V.S. and A.A.R. contributed to the reviewing and editing of the manuscript. Y.O.M., contributed to the investigation. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
This study was conducted in accordance with the local ethics committee of Kazan Federal University (protocol No. 23, 30 June 2020).
Acknowledgments
The work was carried out at the expense of the subsidy allocated to Kazan Federal University to fulfill the state task in the field of scientific activity (PROJECT No. FZSM-2023-0011).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shaimardanova, A.A.; Chulpanova, D.S.; Solovyeva, V.V.; Mullagulova, A.I.; Kitaeva, K.V.; Allegrucci, C.; Rizvanov, A.A. Metachromatic Leukodystrophy: Diagnosis, Modeling, and Treatment Approaches. Front Med (Lausanne) 2020, 7, 576221. [Google Scholar] [CrossRef] [PubMed]
- Shaimardanova, A.A.; Chulpanova, D.S.; Solovyeva; Mullagulova, A.I.; Kitaeva, K.V.; Rizvanov, A.A. New therapeutic strategies for the treatment of metachromatic leukodystrophy. Genes & Cells 2020, 15, 41–50. [Google Scholar] [CrossRef]
- Piguet, F.; Sondhi, D.; Piraud, M.; Fouquet, F.; Hackett, N.R.; Ahouansou, O.; Vanier, M.T.; Bieche, I.; Aubourg, P.; Crystal, R.G.; et al. Correction of brain oligodendrocytes by AAVrh.10 intracerebral gene therapy in metachromatic leukodystrophy mice. Hum Gene Ther 2012, 23, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Mullagulova, A.I.; Timechko, E.E.; Solovyeva, V.V.; Yakimov, A.M.; Ibrahim, A.; Dmitrenko, D.D.; Sufianov, A.A.; Sufianova, G.Z.; Rizvanov, A.A. Adeno-Associated Viral Vectors in the Treatment of Epilepsy. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Issa, S.S.; Shaimardanova, A.A.; Solovyeva, V.V.; Rizvanov, A.A. Various AAV Serotypes and Their Applications in Gene Therapy: An Overview. Cells 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yao, S.; Ding, Y.E.; Xie, M.; Feng, D.; Sha, P.; Tan, L.; Bei, F.; Yao, Y. Designing and optimizing AAV-mediated gene therapy for neurodegenerative diseases: from bench to bedside. J Transl Med 2024, 22, 866. [Google Scholar] [CrossRef] [PubMed]
- Miyake, N.; Miyake, K.; Asakawa, N.; Yamamoto, M.; Shimada, T. Long-term correction of biochemical and neurological abnormalities in MLD mice model by neonatal systemic injection of an AAV serotype 9 vector. Gene Ther 2014, 21, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Mullagulova, A.; Shaimardanova, A.; Solovyeva, V.; Mukhamedshina, Y.; Chulpanova, D.; Kostennikov, A.; Issa, S.; Rizvanov, A. Safety and Efficacy of Intravenous and Intrathecal Delivery of AAV9-Mediated ARSA in Minipigs. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Moldavskii, D.; Gilazieva, Z.; Fattakhova, A.; Solovyeva, V.; Issa, S.; Sufianov, A.; Sufianova, G.; Rizvanov, A. AAV-Based Gene Therapy: Opportunities, Risks, and Scale-Up Strategies. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef] [PubMed]
- Issa, S.S.; Shaimardanova, A.A.; Valiullin, V.V.; Rizvanov, A.A.; Solovyeva, V.V. Mesenchymal Stem Cell-Based Therapy for Lysosomal Storage Diseases and Other Neurodegenerative Disorders. Front Pharmacol 2022, 13, 859516. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Liao, R.; Li, X.; Zhang, C.; Huo, S.; Qin, L.; Xiong, Y.; He, T.; Xiao, G.; Zhang, T. Mesenchymal stem cells in treating human diseases: molecular mechanisms and clinical studies. Signal Transduct Target Ther 2025, 10, 262. [Google Scholar] [CrossRef] [PubMed]
- Bu, Z.; Lou, J.; Xu, W.; Zhang, L.; Tang, Y. Human umbilical cord mesenchymal stem cell-based gene therapy for hemophilia B using scAAV-DJ/8-LP1-hFIXco transduction. Stem Cell Res Ther 2024, 15, 210. [Google Scholar] [CrossRef] [PubMed]
- Zakirova, E.; Aimaletdinov, A.; Tambovsky, M.; Rizvanov, A. Comparative characteristics of mesenchymal stem cell lines of different animal species. Cytology 2021, 63, 139–146. [Google Scholar]
- Voitenko, N.; Makarova, M. Variability of blood biochemical parameters and establishment of reference intervals in preclinical studies. Report 3: minipigs. Laboratory animals for scientific research 2020, 7–15. [Google Scholar] [CrossRef]
- Cavazza, A.; Moiani, A.; Mavilio, F. Mechanisms of retroviral integration and mutagenesis. Hum Gene Ther 2013, 24, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Meuleman, N.; Vanhaelen, G.; Tondreau, T.; Lewalle, P.; Kwan, J.; Bennani, J.; Martiat, P.; Lagneaux, L.; Bron, D. Reduced intensity conditioning haematopoietic stem cell transplantation with mesenchymal stromal cells infusion for the treatment of metachromatic leukodystrophy: a case report. Haematologica 2008, 93, e11-13. [Google Scholar] [CrossRef] [PubMed]
- Tashima, T. Mesenchymal Stem Cell (MSC)-Based Drug Delivery into the Brain across the Blood-Brain Barrier. Pharmaceutics 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wu, Q.; Tam, P.K.H. Immunomodulatory Mechanisms of Mesenchymal Stem Cells and Their Potential Clinical Applications. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Analysis of ARSA enzymatic activity in pig plasma after administration of MSC+AAV9-ARSA. ARSA enzymatic activity was determined using ρ.NCS. The data are presented as the mean value of 5 biological replicates ± S.D.
Figure 1.
Analysis of ARSA enzymatic activity in pig plasma after administration of MSC+AAV9-ARSA. ARSA enzymatic activity was determined using ρ.NCS. The data are presented as the mean value of 5 biological replicates ± S.D.
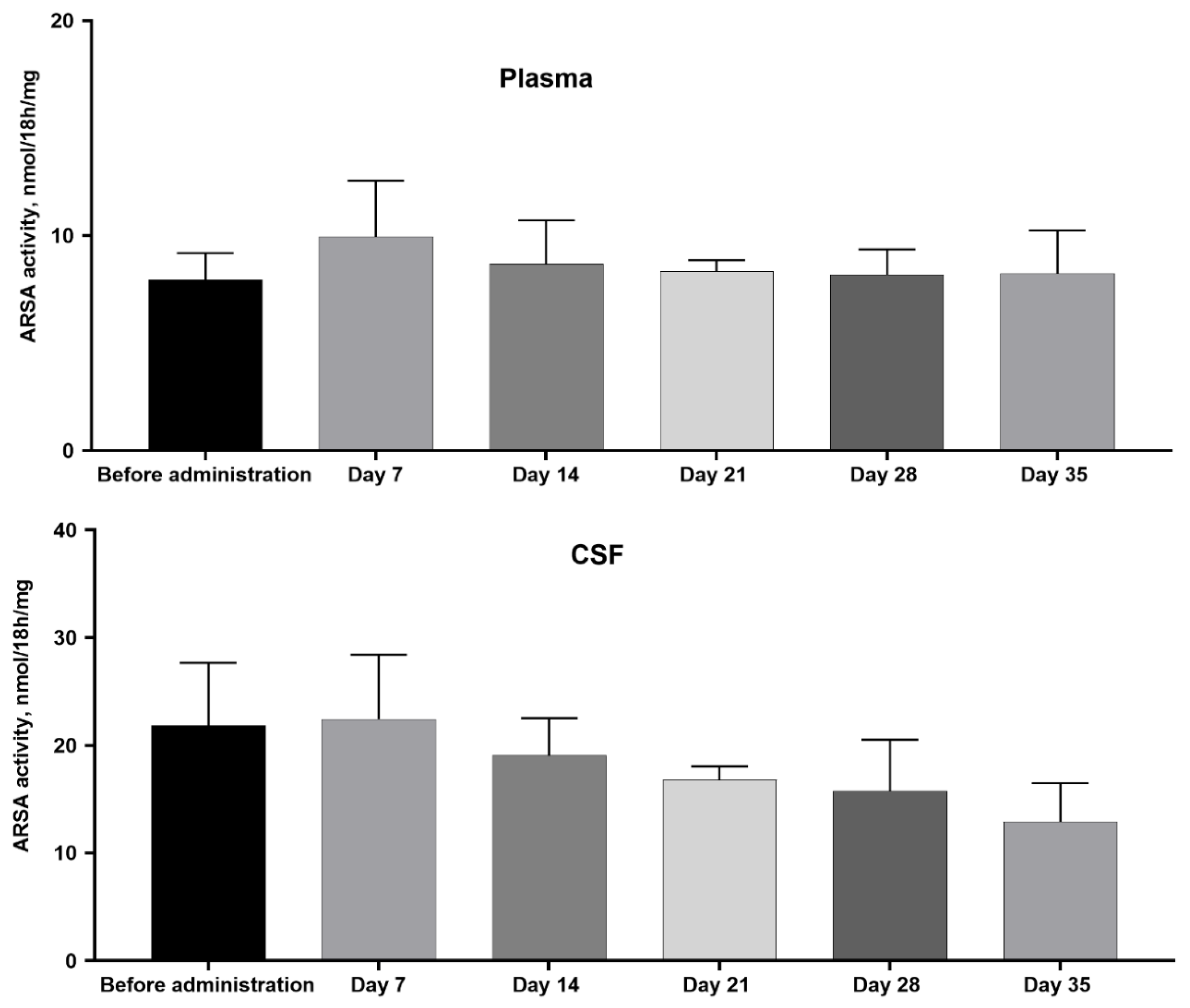
Figure 2.
Analysis of ARSA enzymatic activity in homogenates of various parts of the nervous system of pigs 35 days after administration of MSC+AAV9-ARSA. ARSA enzymatic activity was determined using ρ.NCS. Control — control group of animals; MSC+AAV9-ARSA — intravenous administration of MSC+AAV9-ARSA. Data are presented as the mean value of 5 biological replicates ± S.D. * — p <0.05, ** — p <0.01, **** — p <0.0001.
Figure 2.
Analysis of ARSA enzymatic activity in homogenates of various parts of the nervous system of pigs 35 days after administration of MSC+AAV9-ARSA. ARSA enzymatic activity was determined using ρ.NCS. Control — control group of animals; MSC+AAV9-ARSA — intravenous administration of MSC+AAV9-ARSA. Data are presented as the mean value of 5 biological replicates ± S.D. * — p <0.05, ** — p <0.01, **** — p <0.0001.
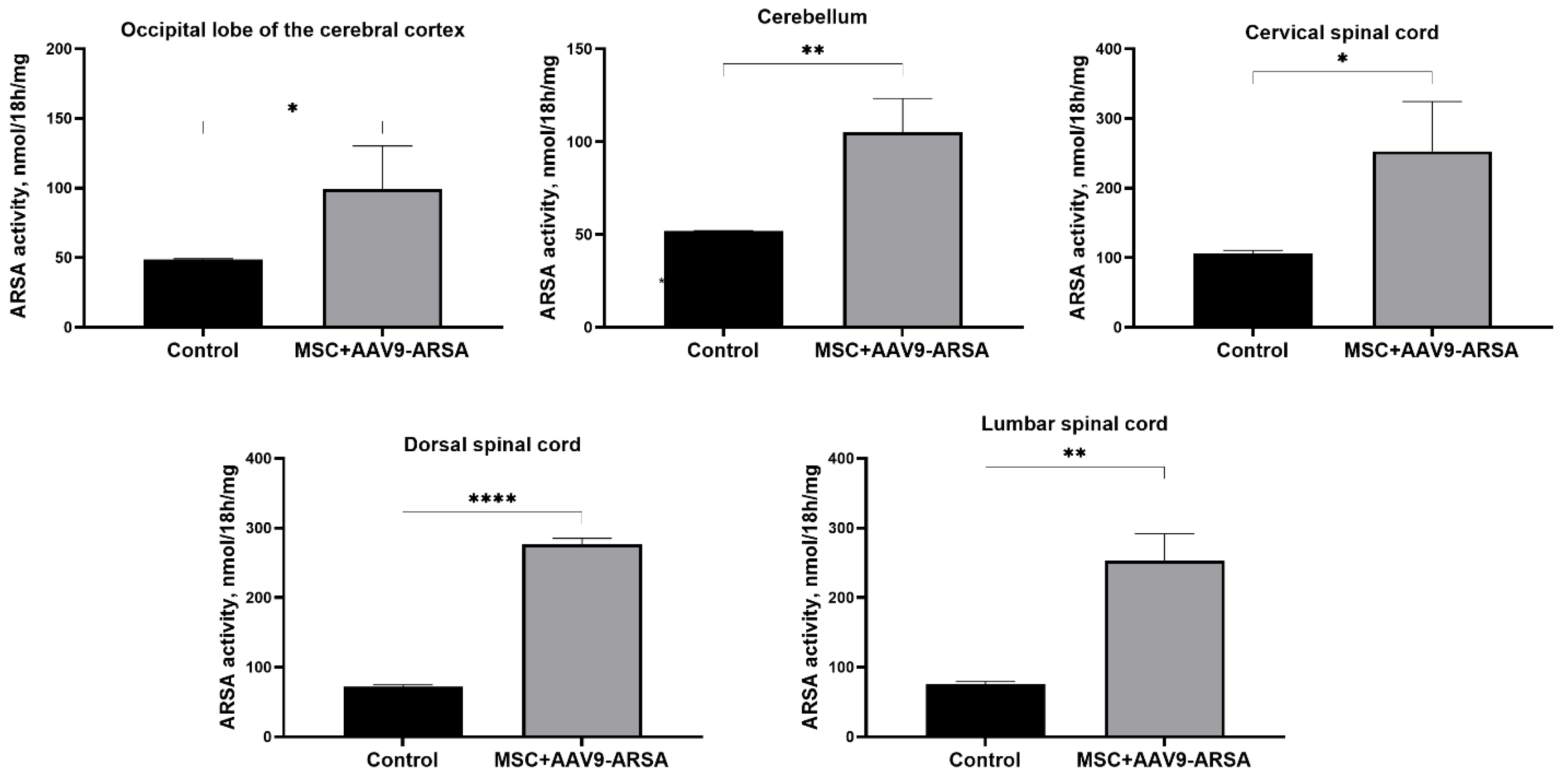
Figure 3.
Biochemical parameters of pig blood serum after administration of MSC+AAV9-ARSA. Data were obtained using a ChemWell 2900 biochemical analyzer (USA) and are presented as the mean value of 5 biological replicates ± SD. * — p < 0.05.
Figure 3.
Biochemical parameters of pig blood serum after administration of MSC+AAV9-ARSA. Data were obtained using a ChemWell 2900 biochemical analyzer (USA) and are presented as the mean value of 5 biological replicates ± SD. * — p < 0.05.
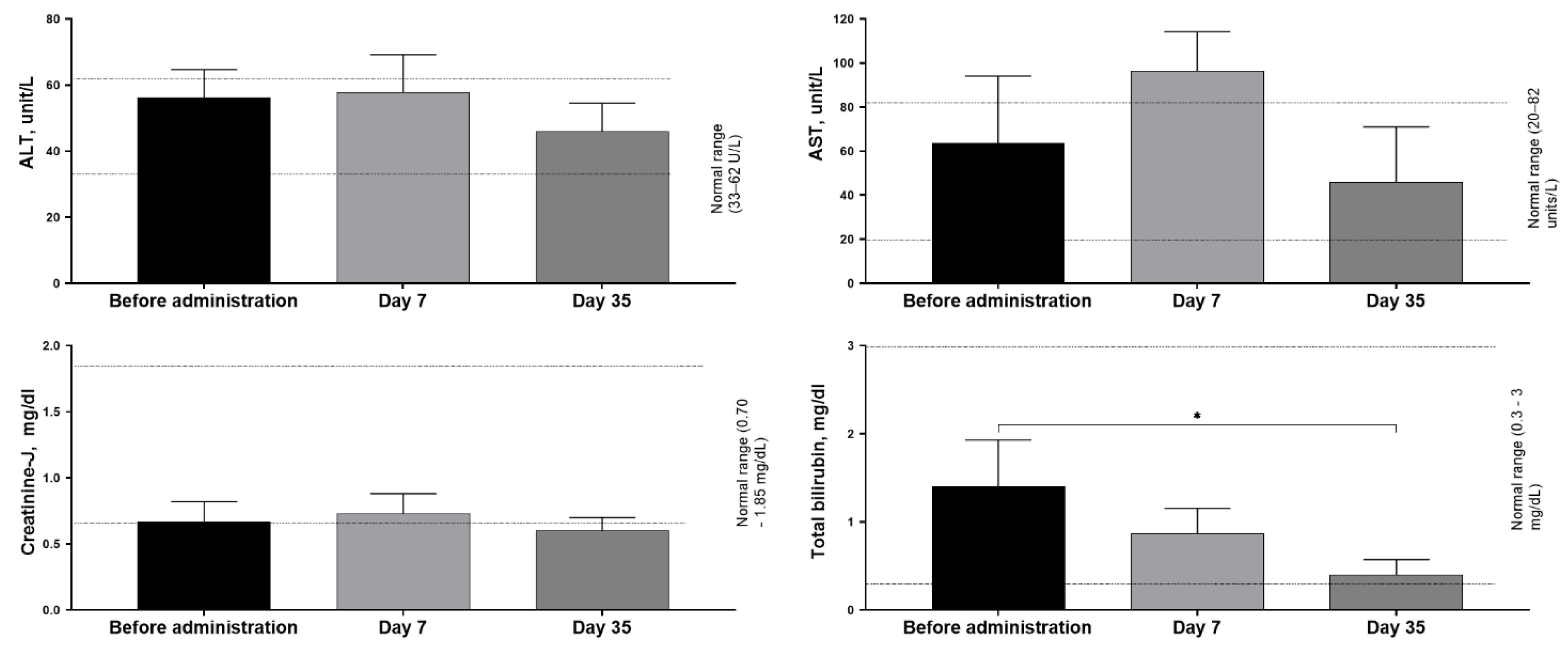
Figure 4.
Analysis of the levels of inflammatory cytokines and chemokines in the blood serum of pigs after intravenous administration of MSC+AAV9-ARSA.
Figure 4.
Analysis of the levels of inflammatory cytokines and chemokines in the blood serum of pigs after intravenous administration of MSC+AAV9-ARSA.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



