Submitted:
17 December 2025
Posted:
18 December 2025
You are already at the latest version
Abstract
Systems favoring cross-pollination such as male sterility and female flowering type are of great importance in the development of new hybrid cultivars and their seed production. The advantages of male sterility are expressed in production of cheaper and competitive seeds. The presence of this characteristic in watermelon is not common, and in some cases, it is accompanied by negative manifestations. A collection of 150 watermelon genotypes was tested at the Maritsa Vegetable Crops Research Institute, Bulgaria, over the past nine years to search for a genetic source of male sterility. The results revealed that two mutations were found. The first mutation was in a plant of the Asar variety, which formed completely degenerated structures in the place of male and female flowers that were completely sterile. The other mutation affected male flowers, female flowers and leaf shape. Male flowers produced a small amount of pollen. Female flowers were formed but they were sterile and aborted at an early stage. The genotype can be propagated by pollination of the normal plants, which in the next generation segregate into mutant - 25%, and normal - 75%. The gene source is phenotyped according to the main characteristics of the fruits and the vegetation period. The mutation found cannot be directly used in a breeding program, but it is of interest for studying this important trait. The success of detecting flowers that are sterile depends on the number of watermelon plants, which for the conditions of the experiment amounted to a minimum of 4492 plants at a probability level of P3 – 0.95.
Keywords:
watermelon
; mutant
; male
; sterility
; probability
; segregation
1. Introduction
Watermelon (Citrullus lanatus (Thunb.) Matsum. & Nakai) is one of the important crops grown in Bulgaria and worldwide. Its importance is determined by the taste and dietary qualities of the fruits. Similar to other members of the Cucurbitaceae family, watermelon is characterized by broad polymorphism in several plant, flower, and fruit characteristics [1]. A characteristic feature of cucurbits is the formation of three types of flowers - male, female, and hermaphroditic. The presence of different flower types on one plant allows for the formation of seven sexual types withing the family. The great variety of sexual types is a prerequisite for easy cross-pollination and also for using the heterosis method of breeding. The genetic systems that favor cross-pollination in watermelons are monoecious and andromonoecious flowering types.
The heterosis method is well known and widely applied to other cucurbit crops such as cucumbers (Cucumis sativus L.), melons (C. melo L.) and zucchini (Cucurbita pepo var gyromontia), where its positive effect is expressed in early maturity, productivity and plant growth [2,3,4]. In addition to the pronounced heterotic effect, the facilitation of seed production and the reduction of seed cost are of great importance for commercial hybrid development. Seed production in these crops is based on the use of different flowering types, with female flowering type in cucumber, male sterility in melon and monoecious flowering type in zucchini being used for the maternal components of F1 hybrids. Pollination between the two parental components and the maintenance of the parental lines are typically carried out by bees, which ensures efficient and economically feasible hybrid seed production. Several reports have described the occurrence of male-sterile forms in watermelon. The first report of a male sterile form was in 1962, obtained by gamma radiation treatment of the cultivar ‘Sugar Baby’ and it was characterized as glabrous male sterile (gms) due to the associated absence of hairs on the leaves of the plant [5]. Along with male sterility, female fertility is reduced, limiting the use of this genetic source. In 1983 in China, another male sterility mutant (ms-1) which resulted from a spontaneous plant mutation in variety Nongmei 100 was found [6]. A third male sterile (ms-2) mutant was identified in Russia from a natural mutation of the variety Kamyzyatsky in 1984. The ms-2 mutant produces only female flowers, while male flowers remain underdeveloped and fail to open; however, female fertility is not affected, making it a suitable donor for breeding purposes [7]. A fourth male sterility mutant was found in 1998 in Korea in the progeny of a line originating from a cross between the commercial hybrid ‘Fiesta’ (Syngenta) and a high-quality breeding line, HL229 (Hungnong Seed Co.). This mutant was named male sterile dwarf (ms-dw) and differs from the sterility mentioned above [8]. All known male sterility mutants in watermelon are monogenic recessive, but their practical use in hybrid seed production is remains limited [9,10].
Pollen fertility and germination are critical factors for successful fertilization and fruit and seed formation in watermelon. Studies on pollen viability, germination rates, and pollen tube growth provide valuable information about the reproductive system and are essential for evaluating the performance of male sterile and fertile lines. [11,12]. In most male-sterile mutants, pollen grains are deformed, low viability, or fail to germinate on the stigma surface, leading to reduced fertilization success. Understanding the physiological and genetic factors affecting pollen development and germination can therefore facilitate the identification of stable male-sterile genotypes and aid in optimizing hybrid seed production systems [13,14].
Male sterility has also been identified in melon, where it is successfully utilized for the production of hybrid seeds. In Bulgaria, this practice has been applied for more than 45 years, when a natural mutant (ms-4) was discovered in the local variety Vidinski koravaci [15]. In melon, four more types of male sterility (ms-1, ms-2, ms-3 and ms-5) have been established, which differ in their manifestation and whose genes are not allelic [16]. All five male-sterile mutants originated from natural mutations and do not affect female fertility.
Male sterility has also been reported in cucumbers, but the female type of flowering is most successfully used. The expression of the female flowering type is controlled by the action and interaction of three genes, but can also be modified by the use of growth regulators [17]. Often, two female lines are used in seed production of some cultivars, which are maintained and hybridized by treatment with gibberellin acid (GA-3) or silver nitrate to produce male flowers. A female type of flowering, which is controlled by two genes, has also been identified in melon, but its use in the development of new F1 hybrids and in seed production is so far insignificant [18]. The apetalous gynoecious mutant (gy) was found in the watermelon variety ‘Mi Guo’ in China in 2006 [19]. Inheritance of this trait is monogenic recessive and female flowers have normal fertility, making it suitable for efficient seed production [20]. Maintenance of the mutant is achieved by crossing with the wild-type monoecious or with an andromonoecious pollinator, which necessitates the discovery of an approach to self-pollinate the female line, a notable disadvantage.
In considering the issues raised, it is important to note that an efficient, simple and cost-effective system for the seed production of hybrid watermelon varieties has not yet been developed. Systems promising cross-pollination, such as male sterility and female type flowering, have been successfully applied in other cucurbit crops; however, in watermelon, male sterile genes often reduce female fertility. Therefore, identifying new genetic sources that confer male sterility without reducing female flower fertility is a crucial prerequisite for initiating new breeding programs. Plant genetic collections represent a valuable resource for identifying novel donors of male sterility, which is the primary objective of the present research.
2. Materials and Methods
2.1. Plant Material and Growing Conditions
The experiments were carried out at the Maritsa VCRI, Plovdiv, Bulgaria, during 2015-2024. A collection of 150 watermelon genotypes (PI’s, cultivars, landraces, breeding lines, and introduced materials) was studied in order to find out mutations affected male flowers or male sterility.
Plants were grown in greenhouse conditions. Seeds were sown a in perlite substrate on 18-20 March; plantlets were pricked in 0.5 L pots on 27-28 March; plants were transplanted on 20-25 April; a double-row system was used; the scheme of transplanting was 240 cm between the centers of each pair of rows, 80 cm between the two rows within a pair, 50 cm between plants in the rows. Plant density was 0.8 plants/m2; fruits were harvested until 2-4 August.
2.2. Phenotype Description
Phenotyping was performed on progeny of the mutant variety Concurrent.
The following characters were measured: days to 50% flowering, days to ripening (from pollination to mature fruit), vegetation period (from emergence of plants to harvest fruits), fruit length (cm), fruit diameter (cm), rind thickness (cm), fruit weight (kg), and total soluble solids TSS (oBx) (measured by using a digital refractometer KERN ORA 32 BA/BB).
Characterization according to UPOV [21]
Fruit: shape in longitudinal section (1 Flattened, 2 Round, 3 Broad elliptical, 4 Elliptical, 5 Pyriform, 6 Oblong);
Fruit: depression at base (1 absent or very shallow, 2 shallow, 3 medium, 4 deep, 5 very deep);
Fruit: depression at apex (1 absent or very shallow, 2 shallow, 3 medium, 4 deep, 5 very deep);
Fruit: ground color of skin (1 light green, 2 medium green, 3 dark green, 4 white, 5 yellow, 6 brown);
Fruit: margin of stripes (1 diffuse, 2 medium, 3 contrasting);
Fruit: width of stripes (1 very narrow, 2 narrow, 3 medium, 4 wide, 5 very wide); Fruit: main color of stripes (1 yellow, 2 very light green, 3 light green, 4 medium
green, 5 dark green, 6 very dark green);
Fruit: main color of flesh (1 white, 2 yellow, 3 orange, 4 pink, 5 pink red, 6 red, 7 dark red);
Fruit: waxy layer (1 absent or very weak, 2 medium, 3 very strong).
The organoleptic characteristics were determined by tasting the fruits by expert assessors.
Taste (1 foul, 2 unremarkable pleasant taste, 3 pleasant taste, 4 superb taste);
Sweetness (1 absent, 2 slightly expressed, 3 sweet, 4 very sweet);
Juiciness (1 weak, 2 medium, 3 strong);
Texture (1 fibrous (stringy), 2 soft, 3 grainy melting, 4 fine grain crunchy).
Pollen Fertility and Germination Rate
Pollen fertility and germination were assessed using standard cytological and in vitro techniques. Temporary pollen preparations were stained with a 1:1 mixture of 4% acetocarmine and glycerine, and at least 100 pollen grains per sample were examined across multiple microscopic fields to determine the proportion of fertile (stained) versus sterile (unstained) pollen. For germination assays, fresh pollen collected one day prior to anthesis was incubated on a medium containing 250 ppm boric acid, 1% agar, and 10% sucrose [12] in darkness at 25 °C for 2 days. A total of 25 flowers were used, with five Petri dishes prepared per accession. The pollen fertility and germination were observed under a light microscope.
2.3. Probability of Occurrence of Mutations in a Collection of Watermelons and Determination of the Minimum Number of Plants to Obtain at Least One Mutant Plant
Probability of mutations occurring:
p = m/n;
p – determining the probability of mutation
m – number of mutated plants
n – total number of plants
Determination of the minimum number of plants to obtain at least one mutant plant [22]
nmin = log (1 – P3) / log (1 - p)
P3 – 0.95 – probability
P3 – 0.99 – probability
P3 = 0.999 – probability
2.4. Experimental Design and Statistical Analysis
Plots were arranged in a randomized complete block design, with two replications, ten plants per plot, and a total of twenty plants per genotype. The trait expression was estimated using all plants per genotype. For analyzing the data, Descriptive statistics and Chi-square test were used. Results were processed by statistical program Genes [23].
3. Results
3.1. Screening a Watermelon Collection for Natural Mutants
The results of the last nine years show that it is possible to obtain natural mutants by screening a collection of watermelons. The frequency of mutations obtained is not high, two genotypes show mutational variability out of a total of 150 genotypes examined. The resulting mutations affect morphological changes in flower buds and changes in leaf shape. The first mutant was registered in Asar variety originating from Kazakhstan in 2015. Of the 20 plants tested, one was distinguished by a mutation that was expressed in a change in the structure of flower buds (Figure 1a). A granular structure surrounded by numerous sepals formed on the flower stem (Figure 1b). Pollination of the mutant flowers was not possible and the grain structure died after seven days (Figure 1c). Throughout the growing season, the plant continued to grow and form mutant flowers, and at the end of the growing season, it failed to form normal flowers and reproduce (Figure 1d). Normal watermelon flowers are characterized by the formation of five sepals, five petals, five stamens in male flowers, pistil in female flowers or stamens and pistil in hermaphrodite flowers (Figure 1e,f). The Asar variety is characterized by an andromonoecious type of flowering, which allowed plants to self-pollinate, but no mutation was found in subsequent generations.
A second mutation was found in the Concurrent variety originating from Russia in 2019. Several plants were found to have changes in flower and leaf characteristics (Figure 1g). Most plants of the cultivar form normal male and female flowers (Figure 1h). Mutant plants can be distinguished in the seedling stage by the formation of strongly dissected leaf lobes, being densely spaced and generally having a smaller habit than normal plants. Later in the growing season, modified male flowers appear that form stamens. The stamens are initially tightly enveloped by the sepals, but they open as the pollen matures (Figure 1i). The amount of pollen produced is scarce and attempts to pollinate other flowers are not successful. Mutant plants produce female flowers, but they are not fertile. The pistil of the carpel is not developed and cannot be pollinated, whereupon it turns yellow and dies (Figure 1j). No progeny was obtained from the mutant plants. Self-pollination of normally developed plants allowed subsequent offspring to be produced.
3.2. Inheritance of Mutation
The resulting progeny of the self-pollinated normal plants of the Concurrent variety showed that they segregated into fertile and sterile, and the sterile plants failed to reproduce. Self-pollination of the next generation of fertile plants again resulted in the emergence of fertile and sterile plants. The analysis of the self-pollinated generations showed segregation in the ratio of 3:1 fertile to sterile plants (Table 1). Fertile plants do not differ phenotypically from the original, and sterile plants also do not differ from the previous generation. Chi-square shows a good match of the experimentally obtained with the expected results, which is 3:1 fertile to sterile plants. It is obvious from the obtained data that the causative mutation is inherited monogenically recessively. Interestingly, this segregation results from self-pollination of fertile plants. These plants are assumed to be in the heterozygous state to cause such segregation. Established segregation was seen in several offspring in 2022 and 2023, with the results having a high degree of probability P (%).
Figure 1.
Mutant plants obtained: (a) cv. Asar mutated flower; (b) Structural change of the flower; (c) Dead flower; (d) The plant with dead flowers; (e) Fertile male flower; (f) Fertile female flower; (g) cv. Concurrent mutant plants; (h) cv. Concurrent fertile plant with normal leaves; (i) Mutated male flowers; (j) Mutated female flower.
Figure 1.
Mutant plants obtained: (a) cv. Asar mutated flower; (b) Structural change of the flower; (c) Dead flower; (d) The plant with dead flowers; (e) Fertile male flower; (f) Fertile female flower; (g) cv. Concurrent mutant plants; (h) cv. Concurrent fertile plant with normal leaves; (i) Mutated male flowers; (j) Mutated female flower.

3.3. Phenotyping of Some Important Traits of cv. Concurrent
The reported metric indicators of the individual offspring show that the Concurrent variety belongs to the group of medium-ripe varieties with a vegetation period of 90-110 days (Table 2). The fruits of fertile plants are of medium size (16.9 cm by 16.7 cm) and weight (2.7 kg). The rind thickness has average parameters (0.8 cm). The Total soluble solids content (TSS) is within good fruit quality, averaging 8.4%. The results of the two years on the reported characteristics differ. In 2022 the indicators days to mass flowering, days to fruit ripening and vegetation period were shorter. Fruit sizes were larger but dry matter content was lower compared to 2023.
In general, the fruits are of good quality, characterized by a round shape, red flesh color, pleasant taste, great sweetness, extensive juiciness, the texture is grainy melting (Figure 2). These qualities of the variety make it particularly suitable for breeding, which facilitates the development of a hybrid based on male sterility.
Pollen Fertility and Germination Rate
Pollen fertility in all evaluated watermelon genotypes was high, varying within a narrow range from 82.3% to 88.5% (Figure 3). This indicates a generally stable reproductive capacity among the tested genotypes. In contrast, mutant line 4-1-3 produced a lower number of pollen grains compared to the other genotypes (Figure 3a).
Pollen germination rates varied considerably among the studied genotypes. The control cultivar ‘Bojura’ showed the highest germination rate (73.3%), followed by the mutants 4-6-6 and 4-6-8 with values of 55.4% and 54.0%, respectively. In the mutant line 4-1-3, pollen germination was significantly reduced, reaching only 10.4% (Figure 3b). The pronounced decrease in germination in this mutant suggests impaired pollen viability and potential disorders in pollen tube development.
3.4. Probability of Mutation
Although mutations rarely occur, with what probability and with what number of examined plants they are found is a question that contributes to clarifying the breeding strategy in general. In the present experiment, 150 progenies of different watermelon genotypes with a total number of about 3000 plants were tested. In the last nine years we have been able to discover two mutations, of which the second mutational form can be reproduced by pollination of fertile plants. The breeding program was aimed at detecting mutations affecting male sterility and the screening was directed specifically at such plants. Calculations made to determine the probability of mutation occurring amounted to 0.067% (Table 3). The probability is not high, but it is still enough to select those with the desired changes by careful examination of the plants.
The data obtained allow for the determination of the minimum number of plants to select at least one mutant plant. According to the results, one mutant plant can be obtained by examining a minimum of 4492 plants at a probability of P3 – 0.95; over 6905 plants at P3 – 0.99 and over 10358 plants at P3 = 0.999.
4. Discussion
4.1. Screening a Watermelon Collection for Natural Mutants
Screening of watermelon collections is essential for selecting forms of interest and represents the first and most critical step toward developing effective breeding programs. Identification of useful traits can be achieved either by screening of existing gemplasm collections or through mutational breeding. Male sterility is one of the key characteristics important for obtaining new F1 watermelon varieties, as it facilitates controlled pollination and hybrid seed production. In the present study, two spontaneous mutations were identified in the available collection, originating from two different varieties. The first mutation found in the variety Asar was characterized by dramatic changes to parts of the flower bud, and it was not possible to obtain seed progeny with which to continue the breeding of hybrid varieties based on male sterility (Figure 1a–d). The second mutation identified in the Concurrent variety, also showed significant changes in both male and female flowers (Figure 1g–j). However, the strong modifications in female flowers, combined with low germination rates did not allow offspring to be reproduced.
Previous studies have reported male sterility in watermelon, often accompanied by reduced female flower fertility. In some cases, sterility was induced artificially (e.g., mutagenesis), while in others it was discovered by screening collections. In both cases, reduced fertility or low seed yield has been reported, making it difficult to use these gene sources in breeding programs [24]. In rare instances, male sterility is not accompanied by negative side effects [7]. However, there is no data on the introduction of male sterility in watermelon in hybrid breeding schemes.
Despite female sterility, the resulting new mutant form can be maintained by self-pollination of fertile plants. Generally, sterile plants in the recessive state are reproduced by hybridization with fertile plants in the heterozygous state, where half of the plants in the next generation are sterile. Hybridization between sterile and fertile plants is typically facilitated by insect pollinators, usually bees, enabling efficient and cost-effective seed production.
In contrast, five types of male sterility are known in melon, which do not affect female fertility [25]. Moreover, for the conditions of Bulgaria, the use of ms-4 is important for breeding and has been used intensively since its discovery more than 45 years ago [26]. The five male sterilities in melon are used in seed production because they differ in their expression and are not allelic [27].
4.2. Inheritance of Mutation
Inheritance of male sterility has been shown to be monogenic recessive in almost all studies [28]. In some cases, it is accompanied by the appearance of another mutant trait such as glabrous foliage [29], juvenile albino [30], dwarfism [31], which is used as a genetic marker. The presence of a distinct morphological marker by which male sterile plants can be identified at an early stage of their development helps in efficient seed production. However, all male-sterile genes are often associated with reduced female fertility and low seed yield.
In the present study, the inheritance pattern observed also corresponds to a monogenic recessive trait, accompanied by changes in leaf morphology and female fertility. It is possible to observe a pleiotropic effect affecting several traits such as leaves and flowers. Furthermore, the new mutant form was maintained by self-pollination of the fertile plants in a ratio of 3:1. This indicates that the fertile plants were heterozygous (Msms). Although the results indicate a monogenic recessive trait, the current data are not conclusive and require further investigation. Future studies should evaluate the response of progeny from crosses with genotypes producing normal male and female flowers, to enable a comprehensive hybridological analysis. It is also important to establish the degree of sterility of the mutated male flowers, as well as other accompanying traits. The research has more fundamental significance for the study of sterility and, to a lesser extent, practical significance. Their use can be aimed at obtaining hybrids with a small number of seeds or without seeds by limited pollination with sterile or not fully fertile pollen. Similar approaches have been reported in diploid watermelon, where seedless fruits were obtained by pollination with non-viable pollen treated with soft γ-radiation [32].
4.3. Phenotyping of Some Important Traits of cv. Concurrent
The quality of the cultivars is very important in determining the parental components for hybridization. The mutational breeding also selects the best quality and most promising genotypes with which the experiment will be carried out. Within our collection, we identified a variety exhibiting relatively good characteristics, except for male sterility. The Concurrent variety belongs to the group of mid-early watermelons, forms medium-sized fruits with good organoleptic qualities, red flesh and monoecious flowering type. The combination of male sterility with good qualities of watermelon fruits greatly facilitates the selection process. In some studies, poor fruit quality has been reported in association with male sterility [30]. Considering the polygenic nature of productivity-related traits, early ripening and fruit quality it is often challenging to combine early ripening, high fruit quality, and male sterility within a single genotype.
4.4. Probability of Mutation
The occurrence of a mutation, and especially one with a positive effect, is extremely important in plant improvement. Mutations are occurred rarely in nature, yet they offer considerable potential for qualitative progress. Spontaneous mutations are often observed when growing a significantly large number of plants. Induced mutations, through physical or chemical mutagens, can significantly increase the frequency of occurrence, but in a number of cases they also cause other unwanted effects. Importantly, there is no guarantee that a desired trait or quality will arise, and the effectiveness of positive mutations is unclear, as they remain more of a random variable and success is not guaranteed. In this study, an attempt was made to define parameters for estimating the probability of mutation. The probability of a mutation affecting floral traits in the tested watermelon collection was calculated as 0.067%. Based on this probability and a confidence level of P₃ = 0.95, the minimum number of plants required for screening was estimated at 4,492. To date, no studies have been conducted that use a probabilistic method to detect random events in watermelons or other cucurbit crops. In previous experiments with the Bojura watermelon variety, using gamma radiation and EMS treatments, mutational changes were observed in plants and seeds; however, the male-sterility trait of interest did not occur. In the individual treatments, the efficiency coefficient varied from 0 to 1.2%, and the number of M2 plants studied was 1395 [33]. In the context of the obtained results of the present study, it is evident that it was necessary to examine a significantly larger volume of plant material – at least 4,492 plants.
5. Conclusions
The study of a large watermelon collection allowed the detection of a spontaneously occurring mutation affecting floral traits. The new mutation is characterized by female sterility, male flowers produce a small amount of pollen, changes in leaf lobes and a less plant growth than normal. Inheritance of male-sterility is monogenic recessive, and the maintenance of the Concurrent variety is carried out by self-pollination of normal plants. Importantly, fruits from fertile plants retain good organoleptic qualities, highlighting the potential value of this mutation in breeding programs. The successful identification of such a mutation strongly depends on both the size of the plant population and the genotypes included in the study, which for the conditions of the experiment amounts to a minimum of 4492 plants at a probability of P3 – 0.95.
Author Contributions
Conceptualization N.V. and S.G.; methodology, N.V. and S.G.; investigation, N.V. and S.G.; resources, N.V.; data curation, N.V.; writing—original draft preparation, N.V.; writing—review and editing, N.V. and S.G.; visualization, N.V. and S.G.; supervision, S.G.; project administration, N.V.; funding acquisition, N.V. and S.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Science Fund, Bulgaria and the Financial Support Competition for Bilateral Projects 2024 – Bulgaria – China by the project KP-06-K/5.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of the data; in the writing of the manuscript; or in the decision to publish the results.
References
- Whitaker, T.; Davis, G. Cucurbits; Interscience Publishers, Inc.: New York, NY, USA, 1962. [Google Scholar]
- Ene, C.O.; Ogbonna, P.E.; Agbo, C.U.; Chukwudi, U.P. Heterosis and combining ability in cucumber (Cucumis sativus L.). Inform. Process. Agric. 2019, 6, 150–157. [Google Scholar] [CrossRef]
- Napolitano, M.; Terzaroli, N.; Kashyap, S.; Russi, L.; Jones-Evans, E.; Albertini, E. Exploring Heterosis in Melon (Cucumis melo L.). Plants 2020, 9, 282. [Google Scholar] [CrossRef] [PubMed]
- Darrudi, R.; Nazeri, V.; Soltani, F.; Shokrpour, M.; Ercolano, M.R. Evaluation of combining ability in Cucurbita pepo L. and Cucurbita moschata Duchesne accessions for fruit and seed quantitative traits. J. Appl. Res. Med. Aromat. Plants 2018, 9, 70–77. [Google Scholar] [CrossRef]
- Watts, V.M. A marked male-sterile mutant in watermelon. Proc. Amer. Soc. Hort. Sci. 1962, 81, 498–505. [Google Scholar]
- Zhang, X.P.; Wang, M. A genetic male-sterile (ms) watermelon from China. Cucurbit Genetics Coop. Rpt 1990, 13, 45–46. [Google Scholar]
- Dyutin, K.E.; Sokolov, S.D. Spontaneous mutant of watermelon with male sterility. Cytology and Genetics 1990, 24, 56–57. [Google Scholar]
- Yang, D.; Paek, K.; Hwang, J.; Park, H. Characterization of a New Male Sterile Mutant in Watermelon. Cucurbit Genetics Cooperative Report 2001, 24, 52–58. [Google Scholar]
- Dong, W.; Wu, D.; Yan, C.; Wu, D. Mapping and analysis of a novel genic male sterility gene in watermelon (Citrullus lanatus). Front. Plant Sci. 2021, 12, 639431. [Google Scholar] [CrossRef]
- Zhang, G.; Ding, Q.; Wei, B.; Yang, H. Transcriptome analysis of sterile and fertile floral buds from recessive genetic male sterility lines in watermelon (Citrullus lanatus L.). J. Plant Growth Regul. 2023, 42, 2800–2812. [Google Scholar] [CrossRef]
- Adıgüzel, P.; Solmaz, İ.; Karabıyık, Ş.; Sarı, N. Comparison on flower, fruit and seed characteristics of tetraploid and diploid watermelons (Citrullus lanatus Thunb. Matsum. & Nakai). Int. J. Agric. Environ. Food Sci. 2022, 6, 704–710. [Google Scholar]
- Kara, E.; Ta¸skın, H.; Karabıyık, ¸S.; Solmaz, ˙I.; Sarı, N.; Karaköy, T.; Baktemur, G. Are Cytological and Morphological Analyses Sufficient in Ploidy Determination of Watermelon Haploid Plants? Horticulturae 2024, 10, 818. [Google Scholar] [CrossRef]
- Wijesinghea, S.A.E.C.; Evans, L.J.; Kirkland, L.; Rader, R. A global review of watermelon pollination biology and ecology: The increasing importance of seedless cultivars. Sci. Hortic. 2020, 271, 109493. [Google Scholar] [CrossRef]
- Zhang, R.; Chang, J.; Li, J.; Lan, G.; Xuan, C.; Li, H.; Ma, J.; Zhang, Y.; Yang, J.; Tian, S.; Yuan, L.; Zhang, X. Disruption of the bHLH transcription factor Abnormal Tapetum 1 causes male sterility in watermelon. Hortic. Res. 2021, 8, 258. [Google Scholar] [CrossRef]
- Lozanov, P. Breeding of male-sterile parental components for facilitating the production of hybrid seeds from melon. In Proceedings of the First Scientific Conference on Genetics and Breeding, Razgrad, Bulgaria, 1983. [Google Scholar]
- Lecouviour, M.; Pitrat, M.; Risser, G. A Fifth Gene for Male Sterility in Cucumis melo. Cucurbit Genetics Cooperative Report 1990, 13, 34–35. [Google Scholar]
- Weng, Y.; Wehner, T.C. Cucumber Gene Catalog 2017. Cucurbit Genet. Coop. Rep. 2017, 39–40. [Google Scholar]
- Pitrat, M. Melon. In: Handbook of plant breeding. Vol. 1, Vegetables I: Asteraceae, Brassicaceae, Chenopodicaceae, and Cucurbitaceae. Ed. by Prohens J and Nuez F. Springer, 2008; p. 283-316.
- Li, Y.; Yang, H.; Yang, J.; Yang, D. A Apetalous Gynoecious Mutant in Watermelon. Cucurbit Genet. Coop. Rep. 2009, 31–32, 15–16. [Google Scholar]
- Xiantao, J.; Depei, L. Discovery of Watermelon Gynoecious Gene gy. Acta Hortic. Sin. 2007, 34, 141–142. [Google Scholar]
- UPOV Code: CTRLS_LAN, (Citrullus lanatus (Thunb.) Matsum. et Nakai), International union for the protection of new varieties of plants, Geneva, TG/142/5 Rev. Watermelon, 2013-03-20 + 2019-10-29.
- Lidanski, T. Statistical methods in biology and agriculture; Zemizdat: Sofia, Bulgaria, 1988; p. 375. [Google Scholar]
- Cruz, C.D. GENES - a software package for analysis in experimental statistics and quantitative genetics. Acta Sci Agron 2013, 35, 271–276. [Google Scholar] [CrossRef]
- Tonfack, L.B.; Foamouhoue, E.N.; Tchoutang, D.N.; Youmbi, E. Application of pesticide combinations on watermelon affects pollen viability, germination, and storage. J. App. Biol. Biotech. 2019, 7, 35–39. [Google Scholar]
- Dogimont, C. Gene List for Melon. Cucurbit Genet. Coop. Rep. 2011, 33-34, 104–133. [Google Scholar]
- Lozanov, P. Heterosis in watermelon, melon and squashes. In The Heterosis and Their Use in Vegetable Production; Publisher house Hristo G. Danov: Plovdiv, Bulgaria, 1974; pp. 254–322. [Google Scholar]
- Lecouviour, M.; Pitrat, M.; Risser, G. A fifth gene for male sterility in Cucumis melo. Cucurbit Genet. Coop. Rep. 1990, 13, 34–35. [Google Scholar]
- Wehner, T.C. Overview of the genes of watermelon, Cucurbitaceae 2008, Proceedings of the IXth EUCARPIA meeting on genetics and breeding of Cucurbitaceae, (Pitrat M, ed), INRA, Avignon (France), May 21-24th, 2008.
- Ray, D.T.; Sherman, J.D. Desynaptic chromosome behavior of the gms mutant in watermelon. J. Hered 1988, 79, 397–399. [Google Scholar] [CrossRef]
- Zhang, X.P.; Rhodes, B.B.; Baird, W.V.; Skorupska, H.T.; Bridges, W.C. Phenotype, Inheritance, and Regulation of Expression of a New Virescent Mutant in Watermelon: Juvenile Albino. J. Amer. Soc. Hort. Sci. 1996, 121, 609–615. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, X.; Wei, Z.; Li, Q.; Li, X. Inheritance of male-sterility and dwarfism in watermelon [Citrullus lanatus (Thunb.) Matsum. and Nakai]. Sci. Hortic. 1998, 74, 175–181. [Google Scholar] [CrossRef]
- Sugyama, K.; Morishita, M. Production of seedless watermelon using soft-X-irradiated pollen. Sci. Hortic. 2000, 84, 255–264. [Google Scholar] [CrossRef]
- Velkov, N.; Tomlekova, N.; Sarsu, F. Sensitivity of watermelon variety Bojura to mutant agents 60Co and EMS. J. BioSci. Biotech. 2016, 5, 105–110. [Google Scholar]
Figure 2.
Morphological and organoleptic characteristics of cv. Concurrent fruits according to UPOV descriptor. Legend: Fruit: shape in longitudinal section (1 Flattened, 2 Round, 3 Broad elliptical, 4 Elliptical, 5 Pyriform, 6 Oblong); Fruit: depression at base (1 absent or very shallow, 2 shallow, 3 medium, 4 deep, 5 very deep); Fruit: depression at apex (1 absent or very shallow, 2 shallow, 3 medium, 4 deep, 5 very deep); Fruit: ground color of skin (1 light green, 2 medium green, 3 dark green, 4 white, 5 yellow, 6 brown); Fruit: margin of stripes (1 diffuse, 2 medium, 3 contrasting); Fruit: width of stripes (1 very narrow, 2 narrow, 3 medium, 4 wide, 5 very wide); Fruit: main color of stripes (1 yellow, 2 very light green, 3 light green, 4 medium green, 5 dark green, 6 very dark green); Fruit: main color of flesh (1 white, 2 yellow, 3 orange, 4 pink, 5 pink red, 6 red, 7 dark red); Fruit: waxy layer (1 absent or very weak, 2 medium, 3 very strong); Taste (1 foul, 2 unremarkable pleasant taste, 3 pleasant taste, 4 superb taste); Sweetness (1 absent, 2 slightly expressed, 3 sweet, 4 very sweet); Juiciness (1 weak, 2 medium, 3 strong); Texture (1 fibrous (stringy), 2 soft, 3 grainy melting, 4 fine grain crunchy).
Figure 2.
Morphological and organoleptic characteristics of cv. Concurrent fruits according to UPOV descriptor. Legend: Fruit: shape in longitudinal section (1 Flattened, 2 Round, 3 Broad elliptical, 4 Elliptical, 5 Pyriform, 6 Oblong); Fruit: depression at base (1 absent or very shallow, 2 shallow, 3 medium, 4 deep, 5 very deep); Fruit: depression at apex (1 absent or very shallow, 2 shallow, 3 medium, 4 deep, 5 very deep); Fruit: ground color of skin (1 light green, 2 medium green, 3 dark green, 4 white, 5 yellow, 6 brown); Fruit: margin of stripes (1 diffuse, 2 medium, 3 contrasting); Fruit: width of stripes (1 very narrow, 2 narrow, 3 medium, 4 wide, 5 very wide); Fruit: main color of stripes (1 yellow, 2 very light green, 3 light green, 4 medium green, 5 dark green, 6 very dark green); Fruit: main color of flesh (1 white, 2 yellow, 3 orange, 4 pink, 5 pink red, 6 red, 7 dark red); Fruit: waxy layer (1 absent or very weak, 2 medium, 3 very strong); Taste (1 foul, 2 unremarkable pleasant taste, 3 pleasant taste, 4 superb taste); Sweetness (1 absent, 2 slightly expressed, 3 sweet, 4 very sweet); Juiciness (1 weak, 2 medium, 3 strong); Texture (1 fibrous (stringy), 2 soft, 3 grainy melting, 4 fine grain crunchy).
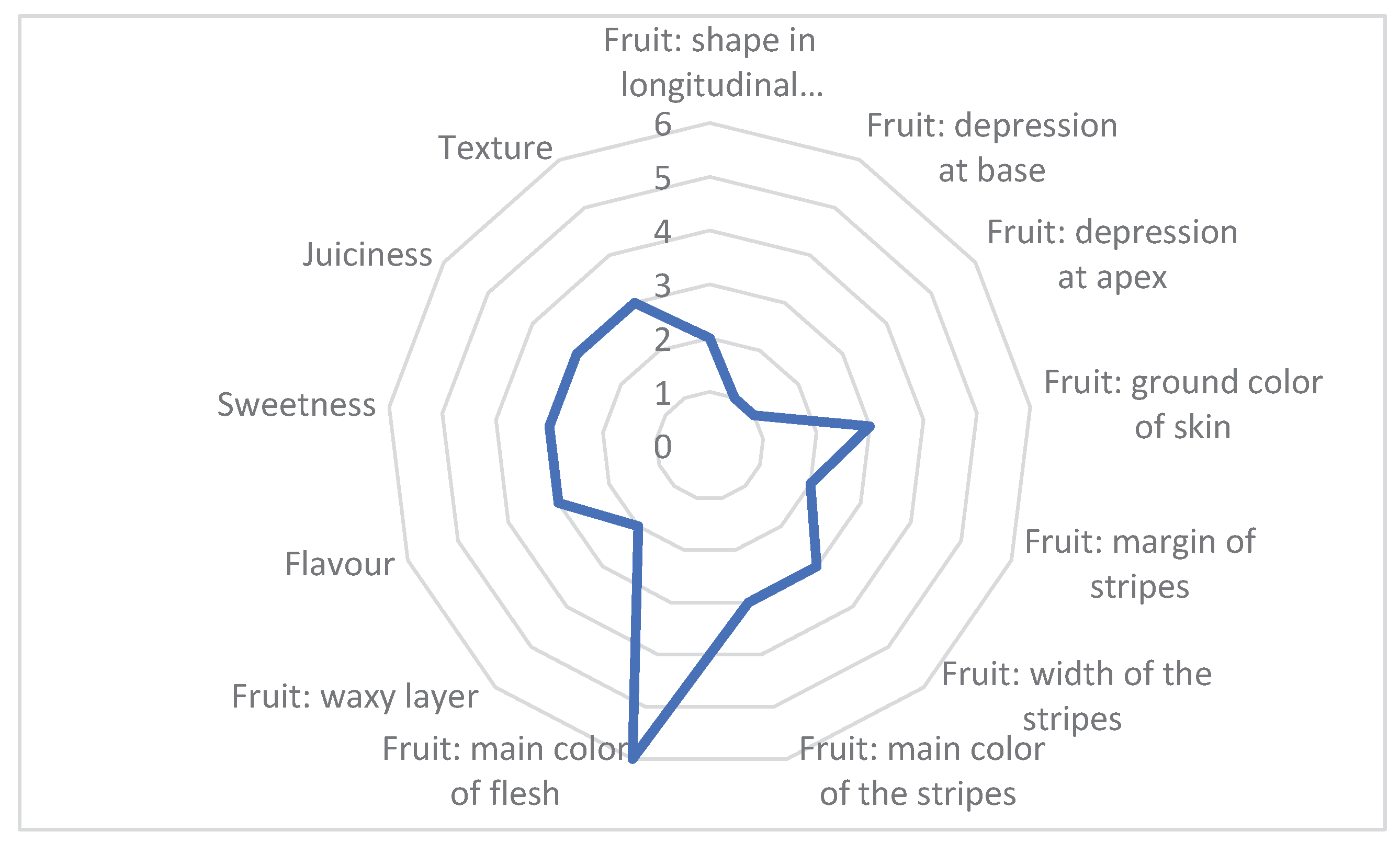
Figure 3.
Pollen fertility (%) and germination rates (%) in three watermelon mutants and variety ‘Bozhura’; a. fertile pollen and b. germination rete in mutant 4-1-3.
Figure 3.
Pollen fertility (%) and germination rates (%) in three watermelon mutants and variety ‘Bozhura’; a. fertile pollen and b. germination rete in mutant 4-1-3.
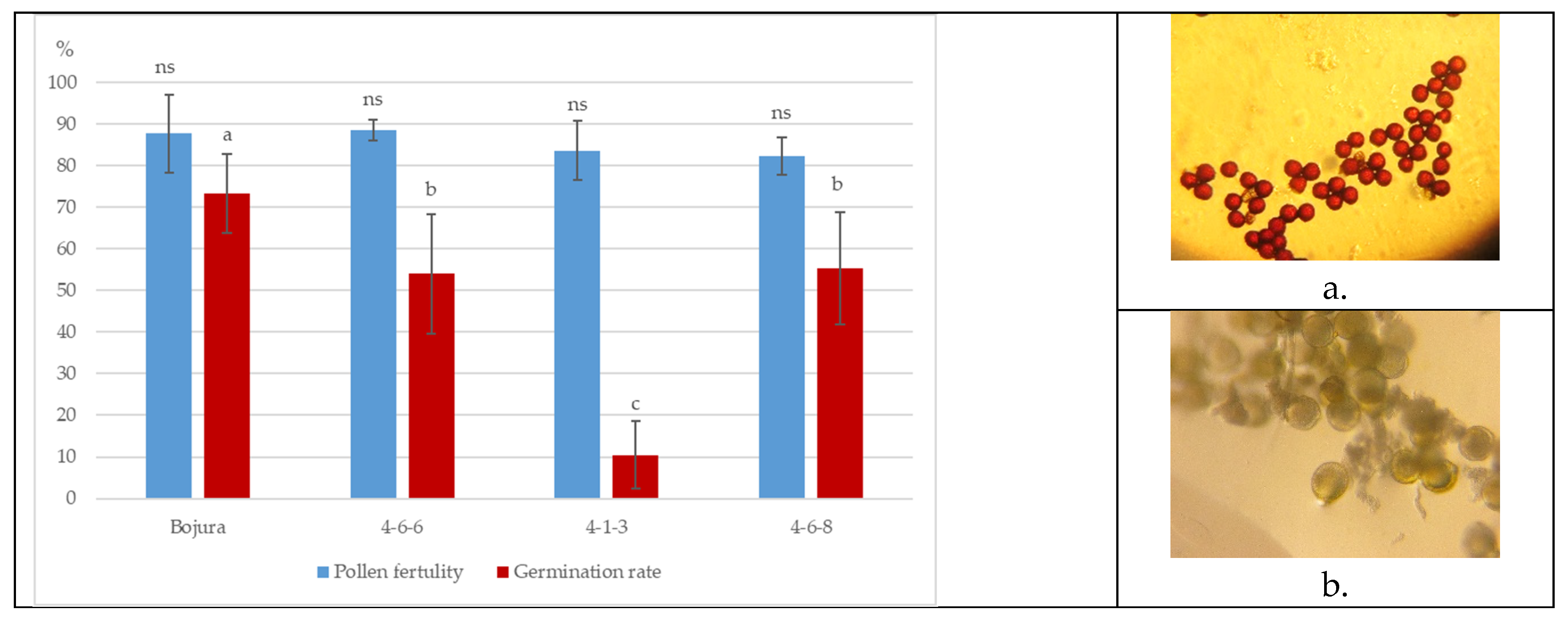

Table 1.
Segregation of self-mated progeny of cv. Concurrent from fertile plants.
| Self-pollinated progeny | Total | Male fertile |
Male sterile |
Ratio Obs:Exp | Chi square | Probability P(%) |
|---|---|---|---|---|---|---|
| 2022 | ||||||
| Concurrent 4-1 | 18 | 14 | 4 | 3:1 | 0.0741 | 78.55 |
| Concurrent 4-4 | 18 | 12 | 6 | 3:1 | 0.6667 | 41.42 |
| Concurrent 4-6 | 17 | 13 | 4 | 3:1 | 0.0196 | 88.86 |
| 2023 | ||||||
| Concurrent 4-1-3 | 10 | 7 | 3 | 3:1 | 0.1333 | 71.50 |
| Concurrent 4-4-6 | 12 | 8 | 4 | 3:1 | 0.4444 | 50.50 |
| Concurrent 4-6-8 | 14 | 10 | 4 | 3:1 | 0.0952 | 75.76 |
| Total | ||||||
| Concurrent | 89 | 64 | 25 | 3:1 | 0.4532 | 50.08 |
Table 2.
Important fruit characteristics of cv. Concurrent.
| Genotype | Days to mass flowering | Days to ripening | Vegetation period (days) | Fruit weight (kg) | Fruit length (cm); | Fruit diameter (cm); | Rind thickness (cm) | TSS (%) |
|---|---|---|---|---|---|---|---|---|
| 2022 | ||||||||
| Concurrent 4-1 | 56 | 35 | 91 | 3.414 | 18 | 19 | 0.9 | 8.2 |
| Concurrent 4-4 | 55 | 42 | 98 | 4.316 | 22 | 20 | 0.6 | 7.0 |
| Concurrent 4-6 | 56 | 33 | 89 | 4.156 | 19.5 | 20 | 0.9 | 8.2 |
| 2023 | ||||||||
| Concurrent 4-1-3 | 61 | 50 | 111 | 1.754 | 14 | 15 | 1 | 9.0 |
| Concurrent 4-4-6 | 59 | 44 | 103 | 1.140 | 14 | 12 | 1 | 9.8 |
| Concurrent 4-6-8 | 60 | 55 | 115 | 1.446 | 14 | 14 | 0.6 | 8.0 |
| Mean | 57.8 | 43.2 | 101.2 | 2.7 | 16.9 | 16.7 | 0.8 | 8.4 |
| Standard Error± | 1.0 | 3.5 | 4.3 | 0.6 | 1.4 | 1.4 | 0.1 | 0.4 |
| Confidence Level (95.0%) | 2.6 | 8.9 | 11.0 | 1.5 | 3.6 | 3.6 | 0.2 | 1.0 |
Table 3.
Probability of occurrence of a mutation in a collection of watermelons and determination of the minimum number of plants (nmin) to obtain at least one mutant plant.
Table 3.
Probability of occurrence of a mutation in a collection of watermelons and determination of the minimum number of plants (nmin) to obtain at least one mutant plant.
| Probability of mutation occurring | Confidence probability | nmin |
|---|---|---|
| 0.00067 | P3 – 0.95 | 4492 |
| 0.067% | P3 – 0.99 | 6905 |
| P3 = 0.999 | 10358 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



