
Submitted:
17 December 2025
Posted:
18 December 2025
You are already at the latest version
Abstract
Programmable organoids are emerging as a powerful new class of engineered developmental systems in which genetic circuits, epigenetic memory architectures, synthetic organizers, and closed-loop control frameworks converge to enable precise regulation of morphogenesis. Traditional organoids rely on spontaneous self-organization, but this intrinsic variability limits reproducibility, causal inference, and translational relevance. Recent advances in CRISPR-based transcriptional and epigenetic engineering, optogenetic and chemogenetic patterning technologies, reaction–diffusion design, and real-time biosensing now allow developmental trajectories to be scripted with increasing precision. This review synthesizes these developments into a unified framework spanning genetic circuit construction, epigenetic programming, synthetic morphogenesis, multi-scale sensing, adaptive regulation, and AI-guided design. Applications across human developmental biology, disease modeling, and regenerative medicine are highlighted, alongside the technical, biosafety, and ethical considerations associated with building increasingly autonomous, self-regulating developmental systems. Collectively, these advances establish programmable organoids as a foundation for developmental synthetic biology and outline a roadmap toward fully engineered human developmental architectures.
Keywords:
programmable organoids
; synthetic gene circuits
; epigenetic engineering
; morphogenesis
; synthetic organizers
; closed-loop developmental control
; cybergenetic systems
; developmental synthetic biology
1. Introduction
Organoids have transformed the study of human biology by providing self-organizing, three-dimensional systems that recapitulate key features of embryonic and tissue development. Built from pluripotent or adult stem cells, these structures recreate the intrinsic patterning logic of morphogenesis, including symmetry breaking, lineage diversification, and the emergence of spatially ordered architectures [1,2]. Their rise has made it possible to observe human developmental processes at a level of physiological relevance and ethical accessibility that traditional in vivo models cannot offer. Yet despite these advances, organoid development remains largely governed by spontaneous self-organization [3]. While powerful, this uncontrolled process often leads to heterogeneity, batch-to-batch variability, incomplete maturation, loss of axial fidelity, and limited reproducibility across laboratories [4]. These constraints restrict the ability to interrogate causality in developmental pathways and limit the translational potential of organoid platforms.
The next frontier in the field involves shifting from passive observation to active control of organoid development. Achieving this requires tools that can tune lineage decisions, sculpt spatial domains, regulate morphogen exposure, and shape emergent architectures with precision. The idea of programmable development captures this transition. Rather than allowing organoids to follow intrinsic yet unpredictable trajectories, researchers are beginning to embed instructions directly into their genetic and epigenetic regulatory layers. Through the integration of synthetic biology, CRISPR-based logic systems, epigenomic engineering, optogenetic control, and inducible signaling modules, organoids are evolving into platforms where developmental outcomes can be scripted, refined, and optimized [5,6].
Central to this transformation is the convergence of several previously distinct technological streams. Synthetic gene circuits provide the computational logic needed to control cell behavior, enabling Boolean decision-making, feedback regulation, and multicellular coordination. CRISPR interference and activation expand this logic space by allowing programmable modulation of specific transcriptional programs. Epigenetic engineering introduces a heritable layer of control, enabling circuits that record signals, stabilize fates, or impose synthetic competence windows. Optogenetic and chemogenetic tools enable spatiotemporal precision, turning organoids into systems where morphogenetic cues can be triggered at defined times and locations [5,7,8,9,10]. Together, these technologies form a growing toolkit for steering development in ways that were previously impossible.
Within this emerging landscape, the term programmable organoid refers to an organoid system in which developmental trajectories are shaped not only by intrinsic self-organization but also by engineered genetic and epigenetic control modules that impose defined rules, responses, or feedback. A programmable organoid can sense its internal state, react to environmental inputs, store information through epigenomic modifications, and execute controlled developmental sequences through synthetic circuitry [5,10,11,12]. This concept represents a profound shift from organoids as passive developmental models to organoids as dynamic, tunable systems with designed behaviors.
This review synthesizes and unifies these emerging directions, focusing on the integration of genetic circuits, epigenomic memory systems, and synthetic developmental control strategies to actively engineer morphogenesis. While prior reviews have examined genetic manipulation, organoid patterning, or synthetic biology in isolation, none have combined these fields into a coherent framework for programmable developmental engineering. By articulating the principles, tools, and future opportunities of this convergence, this work aims to outline a conceptual foundation for the next generation of organoid technology—an era where development becomes not only observable but designable.
2. Engineering the Genetic Landscape with Synthetic Circuits for Cell Fate Control
2.1. Synthetic Gene Circuits in Stem Cells and Organoids
Synthetic gene circuits provide a foundational framework for transforming pluripotent or multipotent stem cells into programmable developmental systems. These circuits operate by linking regulatory DNA elements such as promoters, enhancers, and untranslated regions with engineered transcriptional regulators to create predictable input–output relationships [13,14]. Similar to electronic components, circuit motifs such as logic gates, toggle switches, excitable loops, and feedback controllers allow cells to process, store, and respond to information in controlled ways (Figure 1).
In developmental contexts, logic gates integrate combinations of morphogens or intracellular signals to define lineage outcomes only when precise conditions are met. Toggle switches drive bistable transitions that stabilize cell fates against environmental fluctuations, while excitable or pulse-generating loops create transient activation patterns that reflect natural oscillatory behaviors such as segmentation rhythms [15]. Feedback controllers, both positive and negative, ensure stability, sharpen boundaries, and buffer stochastic noise, which is especially critical in three-dimensional organoid environments characterized by variable cell density and heterogeneous microenvironments [16,17].
CRISPR-based platforms have rapidly expanded the sophistication and modularity of synthetic circuits. CRISPR interference (CRISPRi) and CRISPR activation (CRISPRa) enable fine-grained transcriptional control of endogenous pathways, allowing developmental regulators to be modulated without exogenous overexpression [9,18,19]. Catalytically inactive Cas9 (dCas9) can be fused to activators, repressors, or chromatin modifiers and programmed with multiple guide RNAs, permitting construction of scalable logic circuits capable of implementing AND, NOR, NAND, and multi-input regulatory architectures [9,20,21]. In parallel, base editors introduce irreversible single-nucleotide changes to create genomic “recording” modules, whereas prime editors enable precise rewriting of regulatory sequences to reshape enhancer logic or correct developmental regulators with minimal genomic disruption [22,23,24]. These tools create a regulatory landscape in which entire gene networks can be rewired to control cell fate, responsiveness, and competence windows.
A major advantage of synthetic circuits is their ability to integrate diverse multi-modal inputs that are relevant to organoid development. Cells can be engineered to sense canonical morphogens such as WNT, BMP, SHH, and FGF, to interpret mechanical cues including stiffness, strain, or compression, and to respond to external stimuli such as light or small molecules [25,26,27]. This combinatorial flexibility allows circuits to reproduce the complex information-processing tasks carried out by early embryos, where spatial gradients, mechanical constraints, and temporal cues converge to define identity. In three-dimensional organoids, where morphogen diffusion, mechanical patterning, and stochastic interactions strongly influence developmental trajectories, multi-input circuits provide a way to impose reproducibility, introduce defined structure, and formalize control over emergent morphogenetic events [27,28].
2.2. Inducible and Spatiotemporal Control
Temporal and spatial precision is essential for programming morphogenesis, and inducible systems provide a powerful set of tools to impose such control in developing organoids. Optogenetic systems enable developmental pathways to be regulated with millisecond responsiveness and micron-level spatial resolution. By linking critical regulators of WNT, SHH, or BMP signaling to light-responsive proteins, investigators can generate synthetic morphogen gradients with tunable amplitude, geometry, and duration [29]. Spatially patterned illumination can guide symmetry breaking, define neural tube like dorsal and ventral domains, or induce localized axis formation [30,31]. Because optogenetic cues can be pulsed, stepped, or oscillated, these systems permit experimental reconstruction of dynamic developmental processes such as transient competence windows, morphogen decay profiles, or traveling waves of differentiation. In addition, combining optogenetics with live imaging enables real-time, closed-loop modulation of organoid behavior, which brings organoid engineering closer to a controllable and feedback-regulated developmental platform [29,30,32].
Chemogenetic and drug-inducible systems complement optical control by offering robust, scalable, and reversible mechanisms to modulate gene expression. Tet-On and Tet-Off systems allow transcriptional activation or repression through doxycycline, enabling researchers to titrate differentiation factor expression with fine temporal precision [33,34]. CreERT2 recombination provides tamoxifen-dependent genetic switching for lineage tracing, fate restriction, or the induction of irreversible developmental transitions [35]. Rapalog-inducible modules can assemble or disassemble signaling complexes, activating developmental pathways only in the presence of defined synthetic ligands [36,37]. Newer inducible platforms, including red light responsive regulators, UV sensitive repressors, and small molecule stabilized protein domains, further expand the range of regulatory mechanisms available for developmental programming [38,39]. These systems are especially powerful for probing processes that depend on precise timing, such as stage specific germ layer specification or tightly defined patterning intervals that are otherwise difficult to control in self-organizing tissues.
At a higher organizational level, inducible systems enable the creation of synthetic organizers, which are engineered analogs of the signaling centers that orchestrate embryonic patterning. Engineered cells implanted within organoids can be programmed to secrete morphogens such as WNT3A, SHH, BMP4, or FGF8 under defined inducible conditions. Alternatively, endogenous cells can be reprogrammed in situ using CRISPRa driven secretion circuits to convert them into temporally controlled signaling hubs [27,30,40,41]. These synthetic organizers recapitulate key developmental roles such as axial induction, competence specification, and boundary formation, while allowing their activity to be externally modulated or dynamically tuned. Incorporating feedback modules allows synthetic organizers to adjust their morphogen output based on real-time tissue states, mirroring the self-correcting properties of natural embryonic organizers. As a result, synthetic organizers provide a powerful means to overcome the variability inherent to spontaneous organoid development and to impose reproducible, geometrically defined patterning outcomes.
2.3. Synthetic Morphogenetic Programs
Synthetic morphogenetic programs represent the convergence of genetic circuit design, inducible systems, and developmental biology to actively sculpt organoid architecture. A foundational application is guided symmetry breaking, where synthetic circuits introduce controlled anisotropy into initially isotropic stem-cell aggregates. Inducible activation of WNT/β-catenin, NODAL, or LEFTY pathways can steer germ-layer formation toward precise spatial domains, while optogenetic gradients introduce directional cues that orient early axes [30,42,43]. These approaches allow researchers to systematically vary spatial and temporal parameters of symmetry breaking, revealing how early asymmetries propagate and shape downstream patterning. More advanced circuit designs incorporate symmetry sensors, enabling cells to detect uniformity in gene expression, tension, or morphogen levels and trigger asymmetry only when appropriate [27,42,44]. These programs provide a mechanistic handle on one of the most fundamental yet elusive events in embryogenesis.
Synthetic circuits also enable engineering of axis induction and spatial pattern formation, recreating key organizational principles of multicellular development. Engineered signaling centers that produce region-defining morphogens such as SHH for ventral identities, WNT for posteriorization, and FGF for axial elongation can establish reproducible anterior to posterior or dorsal to ventral axes within organoids [45,46]. Optogenetically sculpted gradients can create graded or sharply bordered domains, while chemogenetic control allows temporal gating of axis induction to mimic developmental progression [30,47]. Incorporating feedback circuits that reinforce or sharpen boundary formation ensures stability of synthetic axes even as organoids grow and undergo morphogenetic remodeling [48,49]. These systems provide a unique framework to dissect and reconstruct the logic of positional information and to explore in a systematic way how tissue level patterns emerge from molecular cues.
Finally, directed lineage specification uses synthetic circuits to guide cells through multi step differentiation cascades. Bistable SOX2 and OCT4 toggle switches can drive neural induction, while inducible expression of mesodermal regulators such as T, EOMES, or MIXL1 initiates orderly mesoderm formation [50,51]. CRISPRa based activation cascades guide sequential induction of endodermal programs without dependence on exogenous morphogens, improving reproducibility and reducing protocol variability [52,53]. When combined with spatial cues, such as patterned illumination or engineered organizers, these circuits enable the simultaneous formation of multiple germ layers or region specific subdomains within a single organoid. This capability transforms organoids into programmable developmental platforms where lineage interactions, patterning hierarchies, and morphogenetic rules can be reconstructed or redesigned with high precision.
3. Epigenetic Circuitry in Organoids for Building Heritable and Context-Specific Developmental Programs
Epigenetic circuitry adds a critical dimension to organoid engineering by enabling cells to store past information, stabilize fate choices, and participate in multicellular patterning through heritable chromatin states. Whereas genetic circuits operate through transcription factor logic and promoter architecture, epigenetic systems act through modifications that persist across cell divisions, integrate environmental cues over time, and define the competence of individual cells and tissues [54,55,56]. These properties make epigenetic engineering uniquely suited to developmental systems, where timing, history, and spatial context govern morphogenesis. Recent advances in dCas9-based epigenetic tools, enhancer engineering, and multicellular chromatin patterning now position organoids as programmable systems capable of executing complex, memory-guided developmental programs (Figure 2) [8,57].
3.1. Epigenome Engineering as a Programmable Memory Layer
A central strategy for engineering epigenetic control in organoids relies on dCas9 fused to “writer” or “eraser” enzymes that deposit or remove chromatin marks at precise loci. Targeted DNA methylation using DNMT3A or locus-specific demethylation via TET1 can durably shift the regulatory state of key developmental genes, while histone-directed fusions such as KRAB (H3K9me3 deposition), p300 (H3K27 acetylation), and LSD1 (removal of H3K4me1/2) provide highly tunable regulation of enhancers and promoters [8,9,58]. These tools allow precise reconfiguration of regulatory landscapes that dictate lineage competence, germ-layer commitment, and spatial identity within organoids.
An equally important design axis is the distinction between persistent and reversible epigenetic states. Transient chromatin opening can generate competence windows during which cells are sensitive to morphogen cues, recapitulating temporally constrained phenomena such as gastrulation, neural induction, or axial patterning [59,60]. In contrast, stable marks create epigenetic memory that endures through multiple divisions, converting past morphogen exposure or mechanical signals into long-term lineage commitments [54,55]. Balancing reversibility and persistence is essential for engineering orderly developmental sequences.
Epigenetic circuits can also operate as biological recorders, encoding the duration, intensity, or temporal ordering of morphogen signals. For example, “write-once” methylation marks can permanently record transient WNT or SHH signals, enabling reconstruction of developmental trajectories long after the original cues have dissipated [8,61]. Multistep dCas9-based systems can encode graded or cumulative exposure, offering a means to understand how early signaling history constrains later morphogenesis. These recorders are particularly valuable in organoids, where spatial heterogeneity makes it challenging to interpret developmental events from endpoint phenotypes alone [62,63].
Finally, epigenetic memory can be intentionally integrated into synthetic circuits as a design principle. Memory modules can serve as developmental checkpoints that restrict progression unless specific chromatin states are met; they can prevent premature lineage switching; and they can enforce sequential transitions by locking cells into intermediate states until circuit-defined conditions are satisfied [64,65]. In this way, epigenome engineering creates programmable, heritable rules that guide how organoids progress through complex developmental landscapes.
3.2. Chromatin State Control for Fate Stabilization
Precise chromatin organization is essential for establishing and maintaining stable lineage identities, and synthetic biology now provides tools for controlling chromatin architecture with high specificity. Synthetic nucleosome-positioning frameworks allow researchers to tune accessibility at promoters and enhancers, thereby regulating transcriptional responsiveness without altering the underlying DNA sequence. Targeted chromatin remodelers recruited through dCas9 or other DNA-binding platforms can shift nucleosome arrays to prime developmental genes, silence alternative fates, or modify transcription factor access during critical differentiation events [66,67].
ATAC-seq guided engineering expands this level of control by linking synthetic circuits to the endogenous chromatin landscape. Enhancers that become accessible during neural, mesodermal, or endodermal differentiation can be selectively activated using p300 based systems, while competing enhancer programs can be silenced using KRAB or LSD1. These approaches enable rational engineering of lineage specific enhancer hierarchies [58]. In organoids, where spatial domains often correspond to differential enhancer activation, chromatin guided engineering ensures that regional identities emerge consistently and remain resistant to disruption caused by local variations in microenvironmental signals [68,69].
A key application of chromatin editing is the generation of synthetic competence windows. Temporarily opening lineage-specific enhancers (e.g., SOX17 for endoderm, NEUROG2 for neural induction) before reclosing them at later stages allows organoids to recapitulate the narrow developmental intervals during which cells respond to specific cues [70,71]. This ensures that differentiation proceeds in a controlled, stage-aligned manner, improving reproducibility and reducing lineage ambiguity. Synthetic competence windows mirror natural mechanisms governing early embryonic transitions, making them powerful tools for engineering high-fidelity developmental programs.
Beyond competence, synthetic epigenetic circuits can lock in lineage trajectories, reinforcing chromatin states compatible with chosen fates while suppressing alternative programs. Stable deposition of activating marks at lineage-defining enhancers, coupled with inhibitory marks on competing networks, prevents lineage drift and ensures that regional identities persist as organoids expand, fold, and mature [57,72]. This stabilization is crucial for multi-domain organoids such as patterned brain or gut models where early specification must be maintained over extended periods of morphological remodeling.
3.3. Multicellular Epigenetic Patterning
Epigenetic programming does not operate only at the level of individual cells; it also organizes multicellular assemblies by creating spatially distinct chromatin states that encode positional identity. Region-specific chromatin landscapes, engineered using spatially restricted delivery of dCas9 modifiers through optogenetics, ligand gradients, or synthetic organizers, allow organoids to develop compartments that mirror embryonic domains long before transcriptional divergence is detectable [73,74]. These epigenetically encoded territories form a scaffold upon which tissue-level patterning unfolds, enabling early establishment of neural, mesodermal, or endodermal zones.
A critical feature of multicellular epigenetic structure is the formation of epigenetic boundaries, which are sharp transitions in chromatin state that prevent inappropriate mixing of developmental programs. Synthetic circuits can generate such boundaries by activating one chromatin state in a defined region, for example rostral HOX genes, while enforcing a mutually exclusive state in another region, for example caudal HOX repression through KRAB [75]. This engineered opposition stabilizes anterior to posterior or dorsal to ventral patterning and shields developing domains from the effects of morphogen diffusion or growth related distortion.
Epigenetic circuits also contribute to emergent pattern formation through feedback loops that propagate or restrict chromatin states across cell populations. Positive feedback can reinforce domain identity, allowing spatial domains to expand or consolidate, while negative feedback sharpens borders and prevents lineage intermixing [76]. These emergent dynamics resemble natural mechanisms such as lateral inhibition, mutual repression, and community effects, all of which generate robust developmental patterns in embryos. In organoids, such synthetic feedback loops allow pattern formation to proceed even in the absence of exogenous gradients, creating self-organizing structures guided by heritable epigenetic logic [77,78].
As spatially resolved epigenome editing becomes more precise, organoid systems are approaching the construction of multicellular epigenetic architectures, which function as blueprints that encode positional cues, lineage boundaries, and long term identity in chromatin rather than relying solely on signaling gradients. These architectures provide a strategy for engineering tissues with stable developmental trajectories, consistent compartmentalization, and predictable patterning outcomes. Integration of epigenetic, genetic, and signaling based networks is likely to produce a new generation of organoids in which developmental logic is encoded across multiple layers of regulation, enabling robust and highly controlled morphogenesis with reproducibility that approaches in vivo development.
3.4. Integrating Epigenetic Logic with Genetic and Morphogenetic Programs
Across organoid development, epigenetic circuitry interacts continuously with genetic circuits, signaling pathways, and mechanical forces. As synthetic organoid engineering advances, a central challenge is designing systems where these layers operate cooperatively rather than independently. Genetic circuits provide rapid, signal-responsive regulation; signaling pathways set spatial gradients; mechanical cues shape tissue geometry; and epigenetic programs provide stability, memory, and heritable identity [49,79]. A coherent synthetic framework integrates these modalities so that transient genetic inputs produce long-term chromatin commitments, morphogen gradients modify epigenetic signatures, and mechanical forces are translated into chromatin remodeling. Such multi-layered integration enables organoids to become not only reactive but predictive systems, where developmental outcomes follow programmable, heritable rules [80,81]. The emergence of this integrative logic marks a conceptual transition toward fully programmable developmental systems in which epigenetic architecture acts as a durable, context-aware foundation for engineered morphogenesis.
4. Programmable Morphogenesis Enabling Spatial and Temporal Architectural Control
Organoid morphogenesis arises from a dynamic interplay between intrinsic self-organizing principles and extrinsic molecular or physical cues. While this spontaneous behavior reveals fundamental aspects of human development, it also introduces variability, incomplete patterning, and divergence from canonical embryonic trajectories [69,82]. Programmable morphogenesis aims to replace this inherent unpredictability with engineered control systems that endow organoids with defined, reproducible developmental logic. These strategies rely on synthetic gene and epigenetic circuits that modulate responsiveness to environmental signals, optogenetic and chemogenetic systems that precisely regulate temporal and spatial patterning, and engineered signaling hubs that recapitulate organizer-like functions [73,77]. Collectively, these approaches shift organoids from passively self-organizing assemblies to actively designed developmental systems, enabling researchers to sculpt spatial and temporal architecture with high fidelity (Figure 3). Taken together, they coalesce into two overarching conceptual domains that frame the discussion that follows, encompassing the deliberate engineering of self-organizing behaviors and the systematic reconstruction of canonical human developmental events.
4.1. Engineering Self-Organization and Pattern Formation
Intrinsic self-organization in organoids reflects the natural capacity of stem cells to interpret local morphogen signals, mechanical forces, and cell to cell interactions to generate symmetry breaking, axial specification, and spatial patterning. However, these processes are often stochastic, sensitive to small fluctuations, and difficult to replicate across experiments [83,84]. Programmable morphogenesis introduces engineered regulatory systems that override or redirect these endogenous programs, enabling consistent and predictable developmental outcomes. Synthetic gene circuits can modulate a cell’s sensitivity to key morphogens such as WNT, BMP, SHH, and FGF, or impose synthetic thresholds that restrict developmental transitions to defined conditions. By dampening or amplifying endogenous feedback loops, these circuits reduce noise and stabilize desired outcomes [85,86]. This level of control allows researchers to guide organoid development toward defined trajectories rather than relying on the probabilistic unfolding of intrinsic signaling networks.
A major conceptual framework underlying pattern formation is rooted in reaction–diffusion mechanisms described by Turing, in which interacting activators and inhibitors generate spatial patterns such as stripes, spots, and periodic domains. Synthetic biology enables explicit construction of these systems by engineering cells to produce activators, inhibitors, or both in precisely controlled ratios. For instance, cells can be programmed to secrete WNT agonists in conjunction with DKK inhibitors, creating synthetic landscapes that generate reproducible periodicity or spatial differentiation within organoids [43,87]. Manipulating production rates, diffusion dynamics, and degradation parameters allows reconstruction of natural patterning events and exploration of theoretical ones that do not exist in vivo. These engineered reaction–diffusion networks serve not only as patterning tools but also as experimental platforms for testing mathematical models of morphogenesis in a biologically realistic context.
Beyond reaction–diffusion systems, programmable morphogenesis enables the construction of synthetic spatial gradients that define positional information with high precision. Optogenetic tools can generate sharply defined, customizable illumination patterns that activate or repress signaling pathways with spatial specificity. By adjusting wavelength, intensity, or exposure duration, synthetic SHH, BMP, or WNT gradients can be sculpted with nearly arbitrary geometry [30,88]. Chemogenetic systems complement this approach by producing morphogen gradients through controlled diffusion of ligands or small-molecule inducers [89]. These synthetic gradients provide a foundational scaffold for reproducible regionalization, enabling organoids to acquire stable polarity, directional identity, and multi-domain patterning without reliance on endogenous variability.
Finally, programmable morphogenesis incorporates engineered signaling hubs, also known as synthetic organizers, that replicate the role of embryonic organizers in orchestrating large scale tissue patterning [77,86,90]. These signaling centers can be built from engineered cells implanted into organoids or generated in situ through inducible expression systems. Synthetic organizers impose spatially restricted secretion of morphogens, directing symmetry breaking, axis initiation, and boundary formation. Their activity can be regulated by light, chemical inducers, or genetic circuits, allowing highly precise temporal control. Most importantly, synthetic organizers transform organoid patterning from a probabilistic process into a designed developmental sequence, enabling researchers to define the architecture of the developing tissue through strategic placement and timing of organizer activation.
4.2. Reconstructing Canonical Human Developmental Events
Programmable morphogenesis enables organoids to reenact developmental events that are otherwise inaccessible in human embryos, particularly early processes such as gastrulation, germ layer specification, and axial formation. Through engineered control of WNT, BMP, and NODAL pathways, researchers can induce spatially asymmetric signaling that results in organized mesendoderm formation, an essential hallmark of gastrulation like transitions. Unlike spontaneous organoid systems, which often display inconsistent or incomplete germ layer segregation, synthetically guided gastrulation produces predictable and anatomically interpretable structures [49,77,79]. This creates a powerful model for studying early human development and for investigating congenital abnormalities that originate during gastrulation but cannot be ethically examined in vivo.
The ability to induce and orient body axes with precision is a central goal of developmental engineering. Synthetic organizers, together with optogenetic and chemogenetic tools, allow reliable induction of anterior–posterior, dorsal–ventral, and medial–lateral axes within organoids. For example, localized activation of SHH pathways can generate stable ventral domains, while spatially confined WNT stimulation can posteriorize tissues or induce primitive streak–like structures [49,91,92]. Synthetic manipulation of activator–inhibitor dynamics further refines these axes by sharpening boundaries and maintaining stable domain identities. These engineered processes replicate the logic of human axis formation, providing an experimentally tractable system for testing hypotheses about symmetry breaking, positional information, and the integration of signaling cues across developing tissues.
Reconstructing neural tube patterning is another achievement made possible through programmable morphogenesis. The neural tube emerges from gradients of SHH, BMP, and WNT signaling, which establish dorsal–ventral neural progenitor domains. By engineering cells within neural organoids to respond to light-controlled SHH induction or CRISPRa-mediated BMP repression, researchers can create controlled dorsal or ventral regions that recapitulate key features of neural tube development [77,93]. Manipulating the steepness or duration of these gradients reveals how progenitor populations interpret positional cues and provides an experimental platform for understanding congenital disorders of neural tube closure or patterning.
At a higher organizational level, programmable morphogenesis enables precise regionalization of the forebrain, midbrain, and hindbrain within cerebral organoids. Controlled activation of FGF8, SHH, WNT, and retinoic acid signaling pathways allows researchers to impose distinct anterior–posterior identities [94,95,96]. Epigenetic boundary engineering can further stabilize the segregation of these regions, ensuring that forebrain, midbrain, and hindbrain compartments develop in proper proportion and orientation. These strategies permit the creation of patterned brain organoids with region-specific architectures, enabling the study of inter-regional communication, developmental timing, and mechanisms underlying region-specific neurodevelopmental disorders.
5. Integrating Multi-Scale Control Systems for Feedback, Sensing, and Closed-Loop Regulation
As organoid engineering advances toward increasingly programmable and predictable developmental systems, the field is beginning to integrate the concepts of biosensing, adaptive regulation, and closed-loop control into living tissues. Traditional organoid protocols rely on exogenous interventions delivered according to fixed schedules, with cells responding passively to environmental cues [84,97,98]. Yet organoid development itself is dynamic and nonlinear, shaped by stochastic molecular fluctuations, variable mechanical environments, and emergent multicellular interactions [83,99]. Achieving reliable and reproducible control of morphogenesis therefore requires a paradigm in which organoids not only receive developmental instructions but also monitor their own state, evaluate deviations from desired trajectories, and autonomously initiate corrective responses.
This emerging framework integrates several layers of control, including biosensors that monitor molecular, metabolic, or mechanical states, internal genetic circuits that interpret this information and implement regulatory actions, and external computational systems powered by machine learning that analyze complex developmental data and recommend optimal interventions. Together, these capabilities form the foundations of cybergenetic developmental regulation, a field that merges synthetic biology, developmental biomechanics, systems control, and artificial intelligence.
5.1. Biosensors and Reporting Circuits
Biosensors constitute the primary interface through which programmable organoids become aware of their internal physiological state. Classical reporter systems, including fluorescent, luminescent, and transcriptionally encoded indicators, provide real-time visibility into the activation of key signaling pathways such as WNT, BMP, SHH, Notch, Ras–ERK, or Hippo. Fluorescent protein based reporters reveal spatial domains of pathway activity, allowing visualization of symmetry breaking events or lineage specification boundaries [100,101]. Fluorescent timers further enrich these systems by encoding the temporal history of gene expression through predictable shifts in emission spectra, enabling reconstruction of lineage dynamics with temporal resolution.
Luminescent reporters extend these capabilities by offering long-term, low background measurements suitable for monitoring slow developmental processes or large three dimensional structures where phototoxicity may compromise viability [102,103]. When coupled to enhancer specific barcodes or immediate early regulatory elements, these systems allow researchers to map the dynamics of developmental enhancers and track the emergence of spatial heterogeneity across complex tissues.
Beyond protein based reporters, the field is now rapidly adopting probe free genetic sensors that operate entirely within endogenous molecular networks. RNA based regulatory circuits, including toehold switches, riboregulators, and synthetic RNA logic gates, can detect the presence of specific transcripts or small molecules and convert these signals into transcriptional or translational outputs. Aptamer based sensors recognize intracellular ligands such as ATP, NAD+, acetyl CoA, or cAMP, providing real-time readouts of metabolic states or signaling flux [104,105,106,107]. Light up RNA aptamers, such as Broccoli, Mango, and Pepper, enable fluorescent reporting without protein intermediates, reducing metabolic burden and allowing deep integration into endogenous RNA dynamics [108,109,110].
Together, these systems transform organoids into self-reporting developmental platforms that can reveal their internal states with spatial and temporal precision and lay the groundwork for more advanced and autonomous control systems (Table 1).
5.2. Adaptive and Closed-Loop Control
Once biosensors provide insight into organoid state, adaptive regulatory systems translate this information into dynamic modulation of developmental processes. Natural tissues maintain functional integrity through networks of regulatory feedback loops that stabilize signaling activity and coordinate growth. Synthetic biology now offers the ability to engineer analogous regulatory architectures within organoids, enabling them to self-correct during morphogenesis [27,111].
Adaptive circuits modulate gene expression or signaling output in response to fluctuations detected by biosensors. For example, engineered pathways can increase morphogen production in regions where signaling becomes insufficient, preventing collapse of axis formation, or reduce proliferative activity when tissue density surpasses a desirable threshold [10,112,113]. Other designs stabilize differentiation trajectories by buffering against stochastic pulses of transcription factor activity that would otherwise push cells into inappropriate lineages [87,114,115]. Such circuits allow organoids to maintain developmental trajectories despite perturbations, whether arising from intrinsic noise or external influences.
Recent advances have demonstrated the feasibility of constructing complex cybergenetic loops in mammalian cells, where molecular sensing is directly linked to signal processing and corrective actuation. These systems can preserve stable gene-expression states under sustained perturbations and maintain morphogen gradients with remarkable robustness [86,116]. When embedded into organoids, they enable more deterministic control over spatial patterning, prevent aberrant lineage diversification, and sustain desired tissue organization across time [117,118].
The integration of machine learning guided intervention further enhances these capabilities. High resolution imaging, multimodal reporters, and real-time transcriptomic or epigenomic profiling generate rich datasets that capture the evolving state of an organoid. Machine learning models, including predictive neural networks and reinforcement learning agents, can interpret these data streams to forecast developmental trajectories, detect early divergence from intended outcomes, and recommend targeted interventions [119,120,121,122]. These interventions may include optogenetic stimulation, timed induction of signaling pathways, or modulation of synthetic circuit activity.
The result is a hybrid closed-loop framework in which biological systems perform local, autonomous regulation while computational systems provide global oversight and predictive decision-making. Such integration paves the way for organoids that can maintain stable developmental programs even under complex dynamic conditions.
5.3. Merging AI with Programmable Organoids
Artificial intelligence plays a pivotal role in elevating programmable organoids from engineered tissues to predictive and designable developmental systems. AI-driven modeling platforms enable researchers to simulate how synthetic gene circuits, signaling networks, or mechanical perturbations influence morphogenesis across scales [123,124]. By incorporating single-cell transcriptomics, chromatin landscapes, proteomic profiles, and live-imaging data, these models achieve increasingly accurate representations of cellular decision-making and tissue morphodynamics.
Generative AI models further expand the design space by proposing novel circuit architectures or morphogenetic strategies that align with specified developmental outcomes. Deep generative networks, including diffusion models, graph neural networks, and transformer based sequence design algorithms, analyze high dimensional datasets from both natural embryos and synthetic organoid systems to infer robust and generalizable design rules [125,126,127]. These systems can recommend unexpected combinations of signaling cues, feedback loops, or circuit topologies that are capable of producing complex patterning behaviors that would otherwise be difficult to engineer manually.
AI also optimizes circuit performance through evolutionary search and reinforcement-learning frameworks [128,129,130]. These algorithms iteratively refine promoter strengths, enhancer configurations, transcription factor ratios, and optogenetic stimulation patterns, converging on parameter sets that produce stable and reproducible outcomes. When coupled with real-time biosensing, such systems can conduct closed-loop optimization directly on living organoids, adjusting circuit activity or environmental conditions as tissues develop.
Collectively, the integration of AI with programmable organoids establishes a new class of cybergenetic developmental systems that function as biologically embodied machines capable of interpreting sensory information, executing synthetic genetic logic, and evolving under computational guidance. This convergence of synthetic biology and artificial intelligence positions programmable organoids as powerful platforms for modeling human development, dissecting disease mechanisms, and designing entirely new morphogenetic processes with unprecedented precision.
6. Applications in Developmental Biology, Disease Modeling, and Regenerative Medicine
Programmable organoids are emerging not only as platforms for synthetic morphogenesis but as versatile systems for interrogating human biology with a level of precision previously unattainable. By integrating genetic circuits, epigenetic programming, synthetic organizers, and closed-loop regulatory architectures, organoids can reconstruct developmental processes, model disease mechanisms, and engineer therapeutic tissues in a manner that is both mechanistically rigorous and experimentally controllable, a direction supported by recent analyses emphasizing the expanding role of organoid systems in neurodevelopmental disease research [5,28,131,132]. These advances position programmable organoids as a cornerstone technology for developmental biology, mechanistic disease modeling, and regenerative medicine, with broad implications for understanding human-specific biology, dissecting complex disorders, and building next-generation therapeutic systems (Figure 4).
6.1. Deep Mechanistic Interrogation of Human Development
Programmable organoids create opportunities to explore aspects of human embryogenesis that cannot be ethically or technically accessed in vivo. Classical animal models diverge markedly from humans in critical developmental parameters, including enhancer logic, chromatin remodeling tempo, morphogen sensitivity thresholds, and neurodevelopmental patterning, which limits their ability to capture human specific regulatory grammars [133,134]. Organoids derived from human pluripotent stem cells retain this species restricted architecture, enabling the study of primate specific developmental programs, human germ layer patterning dynamics, and region specific enhancer promoter interactions [135,136].
Synthetic control systems allow causal replay of developmental hypotheses. Engineered organizers can initiate gastrulation-like events with precise spatial localization; optogenetic gradients can reproduce morphogenetic waves with tunable strength and duration; and synthetic epigenetic circuits can impose competence windows that mirror temporally restricted embryonic transitions [31,137]. These capabilities enable rigorous testing of long-standing questions regarding the sequence of axial specification, the necessity of symmetry breaking signals, or the thresholds that govern lineage bifurcation.
Programmable organoids further permit systematic perturbation across molecular, cellular, and tissue scales, revealing how gene regulatory networks, chromatin states, mechanical cues, and spatial signaling domains converge to construct human form. Emerging directions include incorporation of immune–developmental interfaces, such as fetal macrophage signaling or early thymic epithelial interactions, which provide insights into how immune elements participate in shaping organ architecture during early human life [138,139].
6.2. Disease Modeling with Programmable Perturbations
Programmable organoids provide a transformative framework for modeling congenital, genetic, and acquired diseases by enabling precise perturbation of developmental pathways in a controlled and human specific context. Traditional organoid models often rely on passive observation or isolated gene disruptions, whereas programmable systems allow explicit reconstruction of multi gene network dysfunctions, temporal mispatterning, and disrupted intercellular communication, which are features central to many developmental disorders [69,140].
Synthetic epigenetic editors enable faithful modeling of epigenetic disease states, such as imprinting disorders, enhanceropathies, and chromatin-boundary defects [72,141,142]. Locus-specific methylation, histone modification, or chromatin-loop engineering can recreate pathogenic epigenomic landscapes and allow longitudinal tracking of their consequences for lineage specification and morphogenesis.
Programmable organoids also permit circuit-guided reconstruction of maldevelopment, such as neural tube closure defects, cardiac outflow tract malformations, or regionalization errors in the developing forebrain [143,144]. By modulating organizer function, adjusting reaction–diffusion dynamics, or altering cell–cell feedback loops, researchers can simulate pathological contexts with unprecedented mechanistic precision [145,146].
A rapidly advancing frontier involves metabolic disease modeling, where engineered circuits control intracellular levels of acetyl-CoA, NAD+, or ROS to mimic metabolic congenital disorders. Similarly, synthetic immune–epithelial interfaces allow modeling of autoimmune and inflammatory developmental diseases by programming immune-like signals within organoids [147,148,149].
Together, these approaches enable programmable organoids to dissect disease mechanisms in a manner that unites molecular precision with tissue-level phenotypic relevance.
6.3. Regenerative Medicine and Cell Therapy Engineering
Programmable organoids offer powerful opportunities for regenerative medicine by enabling the construction of tissues that recapitulate developmental logic rather than relying solely on terminal differentiation cues. Regeneration often requires niche environments rich in spatially patterned signals, mechanical constraints, and lineage-supportive microstructures. Synthetic organizers and engineered gradients allow reconstruction of developmental niches in vitro, promoting accurate cell fate specification and functional maturation [42,150,151,152].
Programmable epigenetic and signaling circuits allow long-term stabilization of therapeutic cell fates, addressing a major challenge in cell therapy where transplanted cells frequently lose identity or respond aberrantly to inflammatory cues [10,72]. Lineage-locking circuits, enhancer reinforcement modules, and self-stabilizing feedback programs help maintain differentiation states and prevent lineage drift after transplantation [153,154].
Advances in programmable morphogenesis also enable the assembly of multi-tissue constructs incorporating vasculature, stromal components, and innervation. Synthetic pro-angiogenic circuits coordinate vascular network formation; axon-guiding signaling architectures facilitate organized neuronal wiring; and programmed stromal cells support matrix remodeling and tissue integration [155,156,157]. These developments move beyond isolated organoids toward integrated organ assemblies capable of supporting physiological function, modeling organ–organ interactions, or serving as pre-transplantation therapeutic units.
At the translational interface, layered safety circuits, including inducible kill switches, growth restriction modules, and immune evasive epigenetic programs, provide essential control for clinical applications. These synthetic safeguards help ensure compatibility, prevent uncontrolled proliferation, and enable external regulation of therapeutic constructs in vivo [158,159].
Collectively, programmable organoids establish a new paradigm for regenerative medicine: therapeutic tissues built not only from differentiated cell types but from engineered developmental programs that ensure structural integrity, functional stability, and long-term physiological performance.
7. Challenges and Ethical Considerations
As programmable organoids move toward increasingly sophisticated developmental capabilities, their scientific promise is accompanied by technical challenges, biosafety risks, and important ethical considerations. Engineering tissues that contain synthetic gene circuits, epigenetic programs, and self-organizing developmental logic requires careful evaluation of stability, predictability, and regulatory oversight [160,161,162]. This section outlines the major scientific and ethical questions that must be addressed to ensure responsible advancement of programmable organoid systems (Table 2).
7.1. Safety of Engineered Gene and Epigenetic Circuits
The introduction of synthetic regulatory modules into organoids brings inherent biosafety concerns, particularly regarding off-target effects, genetic stability, epigenetic integrity, and interactions between engineered components and surrounding cellular populations [163,164,165].
Off-target activity remains a central challenge, especially for CRISPR-based transcriptional regulators, base editors, and epigenetic modifiers. Even modest levels of unintended promoter activation, enhancer modification, or chromatin remodeling can alter lineage trajectories or generate aberrant cell states. As organoids grow in 3D environments, these off-target effects may propagate through cell–cell interactions, amplifying subtle errors into large-scale patterning defects [24,166,167].
Genetic instability represents an additional risk. Long-term culture, repeated circuit activation, and high transcriptional load may promote DNA damage, structural variants, or loss of synthetic constructs. Some complex circuits exert metabolic or proteostatic burden, selecting for escape mutants that disable circuit components or rewire regulatory modules [163,168]. Ensuring genetic robustness will require design of circuits that minimize fitness penalties and incorporate fail-safe mechanisms.
Epigenetic drift poses unique concerns in systems that rely on heritable chromatin states. Synthetic epigenetic programs, including methylation based memories, enhancer locking modules, and histone modification circuits, may lose fidelity over time as cells divide, differentiate, or experience mechanical stress [72,169]. Drift can destabilize lineage commitment, impair long term reproducibility, or generate mosaic epigenetic states within a tissue.
Horizontal transfer of circuits in co-culture environments or multi-lineage systems represents an emerging issue. Circuit-bearing DNA or RNA elements may theoretically spread between cells through extracellular vesicles, viral vectors, or fusion events, potentially altering intended compartmentalization [170,171]. As programmable organoids are increasingly used in human–animal chimeric models to study integration, vascularization, or in vivo maturation, preventing unintended circuit dissemination across species boundaries becomes particularly critical.
7.2. Predictability, Controllability, and Emergent Behavior
Despite careful engineering, organoids remain inherently complex adaptive systems with emergent properties that cannot always be fully predicted from the behavior of their individual components [133]. This poses biological and design challenges for programmable morphogenesis.
One major concern is the risk of unintended self-organization. Synthetic organizers, reaction–diffusion networks, or feedback-modulated gradients may generate patterns that deviate from intended designs, especially when interacting with endogenous signaling pathways [172]. Complex 3D geometries can amplify small stochastic differences in initial conditions, producing divergent morphogenetic outcomes across replicates [173].
Synthetic gene circuits embedded within organoids may evolve over time, especially during prolonged growth or differentiation. Mutations, epigenetic silencing, and circuit–host interactions can reshape circuit behavior, altering response thresholds, introducing instability, or reversing intended logic [174]. Evolutionary drift is particularly problematic in circuits governing growth, survival, or metabolic regulation, where selective pressures may favor aberrant states.
Moreover, engineered circuits may interact nonlinearly with mechanical forces, metabolic gradients, and spatial constraints, giving rise to emergent behaviors that are difficult to anticipate computationally. Morphogenetic feedback loops, once inserted into a 3D system, may spontaneously generate new spatial domains, unexpected pattern boundaries, or novel modes of symmetry breaking [49,175].
As closed-loop regulatory systems incorporate machine learning, predictive algorithms, and autonomous control architectures, additional considerations arise related to algorithmic transparency, data provenance, and the governance of AI-driven decision frameworks. Ensuring predictability in programmable organoids will therefore require advances in modeling, high-resolution monitoring, and circuit designs that prioritize robustness, modularity, and containment of emergent dynamics.
7.3. Ethical Dimensions
Increasing sophistication in programmable organoid systems raises important ethical questions, particularly as engineered developmental programs enable reconstructions of events previously restricted to intact human embryos [161,176]. These concerns span lineage identity, developmental potential, cross-species integration, and governance of synthetic biological systems.
Lineage rewiring in human tissues raises questions about the moral status of constructs engineered to acquire complex or unique developmental identities [161,177,178]. Programming neural, germline-like, or early embryonic structures requires clear ethical boundaries on what types of lineage transformations are permissible, especially when circuits enable higher-order patterning or quasi-embryonic features.
The generation of synthetic developmental events that approach embryo like organization, including gastrulation, axial formation, or early neural tube patterning, pushes existing regulatory frameworks to their limits. Although current organoids lack the full integrated architecture or totipotent potential of embryos, the convergence of multiple engineered features may require a reevaluation of conceptual and legal definitions, particularly regarding embryo models, integrated human models, and permissible limits on developmental fidelity [179,180].
The use of programmable organoids in chimeric transplantation studies, where human engineered tissues integrate into animal hosts [181,182], poses additional ethical challenges regarding species boundaries, functional integration, and potential neurodevelopmental implications.
Finally, the regulation of engineered organoid systems lags behind their technical capabilities. There is limited consensus on oversight for constructs containing synthetic gene circuits, epigenetic memory systems, or autonomous closed-loop regulatory modules [178,180,183]. As organoid systems move closer to therapeutic applications, new considerations arise regarding clinical-grade manufacturing, long-term patient safety, and the responsible deployment of tissues with engineered developmental logic.
Collectively, these issues emphasize the need for ethical foresight, transparent governance structures, and interdisciplinary dialogue to ensure that programmable organoids advance in a manner aligned with societal values and biomedical responsibility.
8. Future Directions and Conclusion Toward Fully Programmable Human Developmental Systems
The integration of synthetic biology, stem cell engineering, computational modeling, and organoid technology is steering the field toward an ambitious new horizon in which fully programmable human developmental systems become feasible. These systems will not simply emulate embryogenesis but will execute engineered developmental programs with precision, tunability, and robustness. Achieving this vision requires coordinated advances in genetic circuitry, epigenetic memory frameworks, spatial patterning technologies, autonomous regulatory systems, and artificial intelligence–guided developmental design. Together, these innovations will redefine the capabilities of organoid platforms, enabling the construction of biological systems that can be directed, shaped, and stabilized according to engineered developmental logic (Figure 5).
A central direction for future research involves multi-circuit developmental programming in which several synthetic regulatory networks operate in parallel to control lineage specification, morphogen signaling, mechanical feedback, metabolic states, and temporal progression. Current organoid systems largely depend on single-input perturbations, but next-generation architectures will employ hierarchical and layered control reminiscent of natural developmental hierarchies. These multi-circuit frameworks will enable sequential activation of developmental modules, context-dependent fate decisions guided by local microenvironments, division-of-labor regulatory designs distributed across specific lineages, and coordinated tissue-level governance across complex three-dimensional assemblies. Such integrated programming will allow organoids to progress through intricate developmental sequences with engineered reliability and reduced stochastic variability.
In parallel, substantial advances are anticipated in the development of synthetic organizers with higher-order spatial and temporal control. Early organizers functioned primarily as engineered morphogen sources, but future constructs will incorporate sensing, computation, and adaptive responses. These organizers may transition between anteriorizing and posteriorizing influences, impose orthogonal patterning axes within a single structure, reposition themselves according to dynamic tissue gradients, or function collectively as distributed networks that sculpt multi-domain architectures. Through these mechanisms, it will become possible to generate controlled axes, compartment boundaries, regionalized neural territories, and complex tissue interfaces that closely mirror early human organogenesis.
Equally transformative are emerging epigenetic memory systems designed to encode, stabilize, and propagate developmental instructions across extended temporal windows. Because long-term pattern fidelity depends on durable and heritable regulatory states, synthetic epigenetic architectures offer essential tools for controlling developmental history. Future designs will include multi-state chromatin switches capable of storing graded information, synthetic epigenetic timers that record the duration or order of signaling events, spatially encoded enhancer landscapes that define and maintain positional identity, and feedback mechanisms that correct drifted chromatin states. These systems will reinforce lineage trajectories, preserve spatial domains, and markedly improve reproducibility of long-duration morphogenesis.
As these technologies evolve, organoids will increasingly adopt features of autonomous and self-regulating developmental entities. Through integration of real-time biosensing with embedded computation and feedback-adjusted gene expression, future organoids will be capable of identifying deviations from intended developmental pathways and initiating corrective responses without external intervention. Embedded robustness modules will buffer against fluctuations in morphogen levels, metabolic stresses, or mechanical perturbations, and safety systems will prevent uncontrolled proliferation, aberrant differentiation, or pathological patterning. Over time, these capabilities will produce engineered tissues that maintain stable organization across diverse conditions, bringing organoid systems closer to controlled developmental machines.
Collectively, these advances point toward the emergence of developmental synthetic biology, an interdisciplinary field dedicated to engineering the principles that govern multicellular organization in human-specific contexts. This discipline will draw together synthetic biology, developmental and stem cell biology, systems engineering, control theory, machine learning, computational morphodynamics, and epigenetic and evolutionary science. Its overarching aim is to shift developmental biology from a descriptive discipline to a programmable engineering framework capable of designing new developmental trajectories, constructing synthetic body plans, and generating multicellular systems with emergent structural and functional complexity.
Programmable organoids therefore occupy a pivotal position at the intersection of developmental research, disease modeling, and regenerative medicine. Through the integration of synthetic circuits, engineered epigenetic memory, synthetic organizers, and multi-scale feedback systems, these platforms now provide causally interpretable and developmentally informed models that far exceed the capabilities of conventional organoids. This progress enables precise reconstruction of early human developmental pathways, mechanistic dissection of congenital and epigenetic disorders, and engineering of therapeutic tissues with stable identity and long-term function. However, the increasing sophistication of programmable organoids also introduces significant responsibilities, including the need for rigorous biosafety evaluation, management of emergent behaviors, careful consideration of ethical boundaries, and development of adaptive regulatory frameworks capable of evolving alongside technological capability.
Looking ahead, the convergence of synthetic biology, artificial intelligence, and programmable developmental systems suggests a future in which human development is not only observed but intentionally designed. The coming decade will determine how this unprecedented capability is used, regulated, and integrated into both scientific and clinical practice. Programmable organoids thus stand at the threshold of a fundamentally new era in which the processes that shape human form become programmable substrates for discovery, therapeutic innovation, and regenerative medicine.
Author Contributions
M.N. contributed to the conceptualization, design, and writing of the main manuscript.
Conflict of Interest
The authors declare no conflict of interest.
References
- Lancaster, MA; Knoblich, JA. Generation of cerebral organoids from human pluripotent stem cells. Nat Protoc 2014, 9, 2329–2340. [Google Scholar] [CrossRef] [PubMed]
- Sasai, Y. Next-generation regenerative medicine: organogenesis from stem cells in 3D culture. Cell Stem Cell 2013, 12, 520–530. [Google Scholar] [CrossRef]
- Brassard, JA; Lutolf, MP. Engineering Stem Cell Self-organization to Build Better Organoids. Cell Stem Cell 2019, 24, 860–876. [Google Scholar] [CrossRef]
- Zhao, Z; Chen, X; Dowbaj, AM; et al. Organoids. Nat Rev Methods Primers 2 2022. [Google Scholar] [CrossRef] [PubMed]
- Trentesaux, C; Yamada, T; Klein, OD; et al. Harnessing synthetic biology to engineer organoids and tissues. Cell Stem Cell 2023, 30, 10–19. [Google Scholar] [CrossRef] [PubMed]
- McNamara, HM; Ramm, B; Toettcher, JE. Synthetic developmental biology: New tools to deconstruct and rebuild developmental systems. Semin Cell Dev Biol 2023, 141, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Santorelli, M; Lam, C; Morsut, L. Synthetic development: building mammalian multicellular structures with artificial genetic programs. Curr Opin Biotechnol 2019, 59, 130–140. [Google Scholar] [CrossRef]
- Liu, XS; Wu, H; Ji, X; et al. Editing DNA Methylation in the Mammalian Genome. Cell 2016, 167, 233–247 e217. [Google Scholar] [CrossRef]
- Gilbert, LA; Larson, MH; Morsut, L; et al. CRISPR-mediated modular RNA-guided regulation of transcription in eukaryotes. Cell 2013, 154, 442–451. [Google Scholar] [CrossRef]
- Morsut, L; Roybal, KT; Xiong, X; et al. Engineering Customized Cell Sensing and Response Behaviors Using Synthetic Notch Receptors. Cell 2016, 164, 780–791. [Google Scholar] [CrossRef] [PubMed]
- Prochazka, L; Michaels, YS; Lau, C; et al. Synthetic gene circuits for cell state detection and protein tuning in human pluripotent stem cells. Mol Syst Biol 2022, 18, e10886. [Google Scholar] [CrossRef] [PubMed]
- Perkins, ML; Benzinger, D; Arcak, M; et al. Cell-in-the-loop pattern formation with optogenetically emulated cell-to-cell signaling. Nat Commun 2020, 11, 1355. [Google Scholar] [CrossRef]
- Nissim, L; Perli, SD; Fridkin, A; et al. Multiplexed and programmable regulation of gene networks with an integrated RNA and CRISPR/Cas toolkit in human cells. Mol Cell 2014, 54, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Xie, M; Fussenegger, M. Designing cell function: assembly of synthetic gene circuits for cell biology applications. Nat Rev Mol Cell Biol 2018, 19, 507–525. [Google Scholar] [CrossRef]
- Sonnen, KF; Lauschke, VM; Uraji, J; et al. Modulation of Phase Shift between Wnt and Notch Signaling Oscillations Controls Mesoderm Segmentation. Cell 2018, 172, 1079–1090 e1012. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y; Wang, L; Wang, B. Customizing cellular signal processing by synthetic multi-level regulatory circuits. Nat Commun 2023, 14, 8415. [Google Scholar] [CrossRef]
- De Carluccio, G; Fusco, V; di Bernardo, D. Engineering a synthetic gene circuit for high-performance inducible expression in mammalian systems. Nat Commun 2024, 15, 3311. [Google Scholar] [CrossRef] [PubMed]
- Chavez, A; Scheiman, J; Vora, S; et al. Highly efficient Cas9-mediated transcriptional programming. Nat Methods 2015, 12, 326–328. [Google Scholar] [CrossRef] [PubMed]
- Kampmann, M. CRISPRi and CRISPRa Screens in Mammalian Cells for Precision Biology and Medicine. ACS Chem Biol 2018, 13, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Zalatan, JG; Lee, ME; Almeida, R; et al. Engineering complex synthetic transcriptional programs with CRISPR RNA scaffolds. Cell 2015, 160, 339–350. [Google Scholar] [CrossRef]
- Gao, Y; Xiong, X; Wong, S; et al. Complex transcriptional modulation with orthogonal and inducible dCas9 regulators. Nat Methods 2016, 13, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Chen, PJ; Liu, DR. Prime editing for precise and highly versatile genome manipulation. Nat Rev Genet 2023, 24, 161–177. [Google Scholar] [CrossRef]
- Anzalone, AV; Randolph, PB; Davis, JR; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Komor, AC; Kim, YB; Packer, MS; et al. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Teng, F; Cui, T; Zhou, L; et al. Programmable synthetic receptors: the next-generation of cell and gene therapies. Signal Transduct Target Ther 2024, 9, 7. [Google Scholar] [CrossRef]
- Nims, RJ; Pferdehirt, L; Ho, NB; et al. A synthetic mechanogenetic gene circuit for autonomous drug delivery in engineered tissues. Sci Adv 2021, 7. [Google Scholar] [CrossRef]
- Toda, S; McKeithan, WL; Hakkinen, TJ; et al. Engineering synthetic morphogen systems that can program multicellular patterning. Science 2020, 370, 327–331. [Google Scholar] [CrossRef]
- Velazquez, JJ; Su, E; Cahan, P; et al. Programming Morphogenesis through Systems and Synthetic Biology. Trends Biotechnol 2018, 36, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K; Cui, B. Optogenetic control of intracellular signaling pathways. Trends Biotechnol 2015, 33, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Repina, NA; Johnson, HJ; Bao, X; et al. Optogenetic control of Wnt signaling models cell-intrinsic embryogenic patterning using 2D human pluripotent stem cell culture. Development 150 2023. [Google Scholar] [CrossRef]
- Beyer, HM; Kumar, S; Nieke, M; et al. Genetically-stable engineered optogenetic gene switches modulate spatial cell morphogenesis in two- and three-dimensional tissue cultures. Nat Commun 2024, 15, 10470. [Google Scholar] [CrossRef]
- Komatsu, N; Terai, K; Imanishi, A; et al. A platform of BRET-FRET hybrid biosensors for optogenetics, chemical screening, and in vivo imaging. Sci Rep 2018, 8, 8984. [Google Scholar] [CrossRef]
- Sim, X; Cardenas-Diaz, FL; French, DL; et al. A Doxycycline-Inducible System for Genetic Correction of iPSC Disease Models. Methods Mol Biol 2016, 1353, 13–23. [Google Scholar]
- Gossen, M; Bujard, H. Tight control of gene expression in mammalian cells by tetracycline-responsive promoters. Proc Natl Acad Sci U S A 1992, 89, 5547–5551. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, K; Watt, FM. Lineage tracing. Cell 2012, 148, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Kim, S; Park, J; Jeon, BW; et al. Chemical control of receptor kinase signaling by rapamycin-induced dimerization. Mol Plant 2021, 14, 1379–1390. [Google Scholar] [CrossRef] [PubMed]
- Liu, L; Chen, L; Chung, J; et al. Rapamycin inhibits F-actin reorganization and phosphorylation of focal adhesion proteins. Oncogene 2008, 27, 4998–5010. [Google Scholar] [CrossRef]
- Fan, LZ; Lin, MZ. Optical control of biological processes by light-switchable proteins. Wiley Interdiscip Rev Dev Biol 2015, 4, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Shamala, LF; Zhou, HC; Han, ZX; et al. UV-B Induces Distinct Transcriptional Re-programing in UVR8-Signal Transduction, Flavonoid, and Terpenoids Pathways in Camellia sinensis. Front Plant Sci 2020, 11, 234. [Google Scholar] [CrossRef] [PubMed]
- Li, L; Klim, JR; Derda, R; et al. Spatial control of cell fate using synthetic surfaces to potentiate TGF-beta signaling. Proc Natl Acad Sci U S A 2011, 108, 11745–11750. [Google Scholar] [CrossRef] [PubMed]
- Pandelakis, M; Delgado, E; Ebrahimkhani, MR. CRISPR-Based Synthetic Transcription Factors In Vivo: The Future of Therapeutic Cellular Programming. Cell Syst 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Cheng, D; Clark, CT; Smith, Q. Advances in engineered models of peri-gastrulation. iScience 2025, 28, 112659. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, S; Liu, L; Goh, R; et al. Dissecting the dynamics of signaling events in the BMP, WNT, and NODAL cascade during self-organized fate patterning in human gastruloids. PLoS Biol 2019, 17, e3000498. [Google Scholar] [CrossRef] [PubMed]
- Heisenberg, CP; Bellaiche, Y. Forces in tissue morphogenesis and patterning. Cell 2013, 153, 948–962. [Google Scholar] [CrossRef]
- Feuerstein, M; Chleilat, E; Khakipoor, S; et al. Expression patterns of key Sonic Hedgehog signaling pathway components in the developing and adult mouse midbrain and in the MN9D cell line. Cell Tissue Res 2017, 370, 211–225. [Google Scholar] [CrossRef]
- Scuderi, S; Khouri-Farah, N; Rauthan, R; et al. Engineering human neuronal diversity: Morphogens and stem cell technologies for neurodevelopmental biology. Stem Cell Reports 2025, 20, 102615. [Google Scholar] [CrossRef]
- Repina, NA; McClave, T; Johnson, HJ; et al. Engineered Illumination Devices for Optogenetic Control of Cellular Signaling Dynamics. Cell Rep 2020, 31, 107737. [Google Scholar] [CrossRef]
- Sagner, A; Briscoe, J. Morphogen interpretation: concentration, time, competence, and signaling dynamics. In Wiley Interdiscip Rev Dev Biol; 2017; Volume 6. [Google Scholar]
- Warmflash, A; Sorre, B; Etoc, F; et al. A method to recapitulate early embryonic spatial patterning in human embryonic stem cells. Nat Methods 2014, 11, 847–854. [Google Scholar] [CrossRef]
- Strebinger, D; Deluz, C; Friman, ET; et al. Endogenous fluctuations of OCT4 and SOX2 bias pluripotent cell fate decisions. Mol Syst Biol 2019, 15, e9002. [Google Scholar] [CrossRef] [PubMed]
- Schroder, CM; Zissel, L; Mersiowsky, SL; et al. EOMES establishes mesoderm and endoderm differentiation potential through SWI/SNF-mediated global enhancer remodeling. Dev Cell 2025, 60, 735–748 e735. [Google Scholar] [CrossRef] [PubMed]
- Weltner, J; Balboa, D; Katayama, S; et al. Human pluripotent reprogramming with CRISPR activators. Nat Commun 2018, 9, 2643. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y; Yu, C; Daley, TP; et al. CRISPR Activation Screens Systematically Identify Factors that Drive Neuronal Fate and Reprogramming. Cell Stem Cell 2018, 23, 758–771 e758. [Google Scholar] [CrossRef]
- Bragdon, MDJ; Patel, N; Chuang, J; et al. Cooperative assembly confers regulatory specificity and long-term genetic circuit stability. Cell 2023, 186, 3810–3825 e3818. [Google Scholar] [CrossRef]
- Schuettengruber, B; Bourbon, HM; Di Croce, L; et al. Genome Regulation by Polycomb and Trithorax: 70 Years and Counting. Cell 2017, 171, 34–57. [Google Scholar] [CrossRef] [PubMed]
- Nashun, B; Hill, PW; Hajkova, P. Reprogramming of cell fate: epigenetic memory and the erasure of memories past. EMBO J 2015, 34, 1296–1308. [Google Scholar] [CrossRef] [PubMed]
- Black, JB; Adler, AF; Wang, HG; et al. Targeted Epigenetic Remodeling of Endogenous Loci by CRISPR/Cas9-Based Transcriptional Activators Directly Converts Fibroblasts to Neuronal Cells. Cell Stem Cell 2016, 19, 406–414. [Google Scholar] [CrossRef]
- Hilton, IB; D’Ippolito, AM; Vockley, CM; et al. Epigenome editing by a CRISPR-Cas9-based acetyltransferase activates genes from promoters and enhancers. Nat Biotechnol 2015, 33, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Zaret, KS; Mango, SE. Pioneer transcription factors, chromatin dynamics, and cell fate control. Curr Opin Genet Dev 2016, 37, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Du, Z; Zhang, K; Xie, W. Epigenetic Reprogramming in Early Animal Development. Cold Spring Harb Perspect Biol 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Cambuli, F; Murray, A; Dean, W; et al. Epigenetic memory of the first cell fate decision prevents complete ES cell reprogramming into trophoblast. Nat Commun 2014, 5, 5538. [Google Scholar] [CrossRef]
- Shipman, SL; Nivala, J; Macklis, JD; et al. CRISPR-Cas encoding of a digital movie into the genomes of a population of living bacteria. Nature 2017, 547, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Chen, W; Choi, J. Molecular circuits for genomic recording of cellular events. Trends Genet 2025, 41, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Tomljanovic, M; Muflihah, CH; Rajkovski, D; et al. The epigenetic circle: feedback loops in the maintenance of cellular memory. Epigenetics Chromatin 2025, 18, 56. [Google Scholar] [CrossRef]
- Bell, CC; Faulkner, GJ; Gilan, O. Chromatin-based memory as a self-stabilizing influence on cell identity. Genome Biol 2024, 25, 320. [Google Scholar] [CrossRef] [PubMed]
- Wang, H; Xu, X; Nguyen, CM; et al. CRISPR-Mediated Programmable 3D Genome Positioning and Nuclear Organization. Cell 2018, 175, 1405–1417 e1414. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y; Sun, J; Zhu, R. CRISPR-epigenetic crosstalk: From bidirectional regulation to therapeutic potential. Comput Struct Biotechnol J 2025, 27, 4496–4504. [Google Scholar] [CrossRef]
- Pollen, AA; Bhaduri, A; Andrews, MG; et al. Establishing Cerebral Organoids as Models of Human-Specific Brain Evolution. Cell 2019, 176, 743–756 e717. [Google Scholar] [CrossRef]
- Velasco, S; Kedaigle, AJ; Simmons, SK; et al. Individual brain organoids reproducibly form cell diversity of the human cerebral cortex. Nature 2019, 570, 523–527. [Google Scholar] [CrossRef]
- Hawkins, RD; Hon, GC; Yang, C; et al. Dynamic chromatin states in human ES cells reveal potential regulatory sequences and genes involved in pluripotency. Cell Res 2011, 21, 1393–1409. [Google Scholar] [CrossRef]
- Tsankov, AM; Gu, H; Akopian, V; et al. Transcription factor binding dynamics during human ES cell differentiation. Nature 2015, 518, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Nunez, JK; Chen, J; Pommier, GC; et al. Genome-wide programmable transcriptional memory by CRISPR-based epigenome editing. Cell 2021, 184, 2503–2519 e2517. [Google Scholar] [CrossRef] [PubMed]
- Polstein, LR; Gersbach, CA. A light-inducible CRISPR-Cas9 system for control of endogenous gene activation. Nat Chem Biol 2015, 11, 198–200. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y; Bartosovic, M; Ma, S; et al. Spatial profiling of chromatin accessibility in mouse and human tissues. Nature 2022, 609, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Noordermeer, D; Leleu, M; Schorderet, P; et al. Temporal dynamics and developmental memory of 3D chromatin architecture at Hox gene loci. Elife 2014, 3, e02557. [Google Scholar] [CrossRef] [PubMed]
- Amberg, N; Laukoter, S; Hippenmeyer, S. Epigenetic cues modulating the generation of cell-type diversity in the cerebral cortex. J Neurochem 2019, 149, 12–26. [Google Scholar] [CrossRef]
- Manfrin, A; Tabata, Y; Paquet, ER; et al. Engineered signaling centers for the spatially controlled patterning of human pluripotent stem cells. Nat Methods 2019, 16, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Moris, N; Anlas, K; van den Brink, SC; et al. An in vitro model of early anteroposterior organization during human development. Nature 2020, 582, 410–415. [Google Scholar] [CrossRef]
- Etoc, F; Metzger, J; Ruzo, A; et al. A Balance between Secreted Inhibitors and Edge Sensing Controls Gastruloid Self-Organization. Dev Cell 2016, 39, 302–315. [Google Scholar] [CrossRef]
- Mammoto, T; Mammoto, A; Ingber, DE. Mechanobiology and developmental control. Annu Rev Cell Dev Biol 2013, 29, 27–61. [Google Scholar] [CrossRef] [PubMed]
- Treutlein, B; Brownfield, DG; Wu, AR; et al. Reconstructing lineage hierarchies of the distal lung epithelium using single-cell RNA-seq. Nature 2014, 509, 371–375. [Google Scholar] [CrossRef]
- Kadoshima, T; Sakaguchi, H; Nakano, T; et al. Self-organization of axial polarity, inside-out layer pattern, and species-specific progenitor dynamics in human ES cell-derived neocortex. Proc Natl Acad Sci U S A 2013, 110, 20284–20289. [Google Scholar] [CrossRef]
- Sasai, Y. Cytosystems dynamics in self-organization of tissue architecture. Nature 2013, 493, 318–326. [Google Scholar] [CrossRef]
- Lancaster, MA; Renner, M; Martin, CA; et al. Cerebral organoids model human brain development and microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef]
- Anand, AA; Khan, M; V, M; et al. The Molecular Basis of Wnt/beta-Catenin Signaling Pathways in Neurodegenerative Diseases. Int J Cell Biol 2023, 2023, 9296092. [Google Scholar] [CrossRef] [PubMed]
- Toda, S; Blauch, LR; Tang, SKY; et al. Programming self-organizing multicellular structures with synthetic cell-cell signaling. Science 2018, 361, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Sekine, R; Shibata, T; Ebisuya, M. Synthetic mammalian pattern formation driven by differential diffusivity of Nodal and Lefty. Nat Commun 2018, 9, 5456. [Google Scholar] [CrossRef] [PubMed]
- Bugaj, LJ; Choksi, AT; Mesuda, CK; et al. Optogenetic protein clustering and signaling activation in mammalian cells. Nat Methods 2013, 10, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Lende-Dorn, BA; Atkinson, JC; Bae, Y; et al. Chemogenetic tuning reveals optimal MAPK signaling for cell-fate programming. Cell Rep 2025, 44, 116226. [Google Scholar] [CrossRef]
- Liu, TL; Upadhyayula, S; Milkie, DE; et al. Observing the cell in its native state: Imaging subcellular dynamics in multicellular organisms. Science 360 2018. [Google Scholar] [CrossRef] [PubMed]
- Martyn, I; Brivanlou, AH; Siggia, ED. A wave of WNT signaling balanced by secreted inhibitors controls primitive streak formation in micropattern colonies of human embryonic stem cells. Development 146 2019. [Google Scholar] [CrossRef]
- Meinhardt, A; Eberle, D; Tazaki, A; et al. 3D reconstitution of the patterned neural tube from embryonic stem cells. Stem Cell Reports 2014, 3, 987–999. [Google Scholar] [CrossRef] [PubMed]
- Cao, J; Spielmann, M; Qiu, X; et al. The single-cell transcriptional landscape of mammalian organogenesis. Nature 2019, 566, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Muguruma, K; Nishiyama, A; Kawakami, H; et al. Self-organization of polarized cerebellar tissue in 3D culture of human pluripotent stem cells. Cell Rep 2015, 10, 537–550. [Google Scholar] [CrossRef]
- Cederquist, GY; Asciolla, JJ; Tchieu, J; et al. Specification of positional identity in forebrain organoids. Nat Biotechnol 2019, 37, 436–444. [Google Scholar] [CrossRef]
- Qian, X; Nguyen, HN; Song, MM; et al. Brain-Region-Specific Organoids Using Mini-bioreactors for Modeling ZIKV Exposure. Cell 2016, 165, 1238–1254. [Google Scholar] [CrossRef] [PubMed]
- Huch, M; Knoblich, JA; Lutolf, MP; et al. The hope and the hype of organoid research. Development 2017, 144, 938–941. [Google Scholar] [CrossRef]
- Rossi, G; Manfrin, A; Lutolf, MP. Progress and potential in organoid research. Nat Rev Genet 2018, 19, 671–687. [Google Scholar] [CrossRef]
- Gao, Q; Yang, Y; Yang, H; et al. Beyond biochemical patterning: How mechanical bistability governs robust organoid morphogenesis. Mechanobiol Med 2025, 3, 100134. [Google Scholar] [CrossRef]
- Depry, C; Mehta, S; Zhang, J. Multiplexed visualization of dynamic signaling networks using genetically encoded fluorescent protein-based biosensors. Pflugers Arch 2013, 465, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Zou, F; Bai, L. Using time-lapse fluorescence microscopy to study gene regulation. Methods 2019, 159-160, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Hall, MP; Unch, J; Binkowski, BF; et al. Engineered luciferase reporter from a deep sea shrimp utilizing a novel imidazopyrazinone substrate. ACS Chem Biol 2012, 7, 1848–1857. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A; Sharkey, J; Plagge, A; et al. Multicolour In Vivo Bioluminescence Imaging Using a NanoLuc-Based BRET Reporter in Combination with Firefly Luciferase. Contrast Media Mol Imaging 2018, 2018, 2514796. [Google Scholar] [CrossRef] [PubMed]
- Wang, T; Simmel, FC. Riboswitch-inspired toehold riboregulators for gene regulation in Escherichia coli. Nucleic Acids Res 2022, 50, 4784–4798. [Google Scholar] [CrossRef]
- Takahashi, K; Galloway, KE. RNA-based controllers for engineering gene and cell therapies. Curr Opin Biotechnol 2024, 85, 103026. [Google Scholar] [CrossRef] [PubMed]
- Paige, JS; Wu, KY; Jaffrey, SR. RNA mimics of green fluorescent protein. Science 2011, 333, 642–646. [Google Scholar] [CrossRef]
- Chappell, J; Takahashi, MK; Lucks, JB. Creating small transcription activating RNAs. Nat Chem Biol 2015, 11, 214–220. [Google Scholar] [CrossRef]
- Chen, Z; Chen, W; Reheman, Z; et al. Genetically encoded RNA-based sensors with Pepper fluorogenic aptamer. Nucleic Acids Res 2023, 51, 8322–8336. [Google Scholar] [CrossRef]
- Dolgosheina, EV; Jeng, SC; Panchapakesan, SS; et al. RNA mango aptamer-fluorophore: a bright, high-affinity complex for RNA labeling and tracking. ACS Chem Biol 2014, 9, 2412–2420. [Google Scholar] [CrossRef] [PubMed]
- Filonov, GS; Moon, JD; Svensen, N; et al. Broccoli: rapid selection of an RNA mimic of green fluorescent protein by fluorescence-based selection and directed evolution. J Am Chem Soc 2014, 136, 16299–16308. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A; Keshara, R; Kim, YH; et al. Self-organization of organoids from endoderm-derived cells. J Mol Med (Berl) 2021, 99, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Del Vecchio, D; Abdallah, H; Qian, Y; et al. A Blueprint for a Synthetic Genetic Feedback Controller to Reprogram Cell Fate. Cell Syst 2017, 4, 109–120 e111. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y; Budde, MW; Mayalu, MN; et al. Synthetic mammalian signaling circuits for robust cell population control. Cell 2022, 185, 967–979 e912. [Google Scholar] [CrossRef]
- Kotula, JW; Kerns, SJ; Shaket, LA; et al. Programmable bacteria detect and record an environmental signal in the mammalian gut. Proc Natl Acad Sci U S A 2014, 111, 4838–4843. [Google Scholar] [CrossRef]
- Holmes, WR; Reyes de Mochel, NS; Wang, Q; et al. Gene Expression Noise Enhances Robust Organization of the Early Mammalian Blastocyst. PLoS Comput Biol 2017, 13, e1005320. [Google Scholar] [CrossRef]
- Milias-Argeitis, A; Rullan, M; Aoki, SK; et al. Automated optogenetic feedback control for precise and robust regulation of gene expression and cell growth. Nat Commun 2016, 7, 12546. [Google Scholar] [CrossRef]
- Amin, ND; Kelley, KW; Kaganovsky, K; et al. Generating human neural diversity with a multiplexed morphogen screen in organoids. Cell Stem Cell 2024, 31, 1831–1846 e1839. [Google Scholar] [CrossRef]
- Johnson, MB; March, AR; Morsut, L. Engineering multicellular systems: using synthetic biology to control tissue self-organization. Curr Opin Biomed Eng 2017, 4, 163–173. [Google Scholar] [CrossRef]
- Monzel, AS; Hemmer, K; Kaoma, T; et al. Machine learning-assisted neurotoxicity prediction in human midbrain organoids. Parkinsonism Relat Disord 2020, 75, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Bai, L; Wu, Y; Li, G; et al. AI-enabled organoids: Construction, analysis, and application. Bioact Mater 2024, 31, 525–548. [Google Scholar] [CrossRef] [PubMed]
- Huang, K; Li, M; Li, Q; et al. Image-based profiling and deep learning reveal morphological heterogeneity of colorectal cancer organoids. Comput Biol Med 2024, 173, 108322. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z; Chen, C; Wang, S; et al. scRL: Utilizing Reinforcement Learning to Evaluate Fate Decisions in Single-Cell Data. In Biology (Basel); 2025; Volume 14. [Google Scholar]
- Etcheverry, M; Moulin-Frier, C; Oudeyer, PY; et al. AI-driven automated discovery tools reveal diverse behavioral competencies of biological networks. Elife 13 2025. [Google Scholar]
- Zhu, D; Jerby, L. Gradient-aware modeling advances AI-driven prediction of genetic perturbation effects. bioRxiv 2025. [Google Scholar]
- Ali, M; Richter, S; Erturk, A; et al. Graph neural networks learn emergent tissue properties from spatial molecular profiles. Nat Commun 2025, 16, 8419. [Google Scholar] [CrossRef] [PubMed]
- Chen, K; Qin, KR; Na, J; et al. Deep manifold learning reveals hidden developmental dynamics of a human embryo model. Sci Adv 2025, 11, eadr8901. [Google Scholar] [CrossRef] [PubMed]
- Stillman, NR; Mayor, R. Generative models of morphogenesis in developmental biology. Semin Cell Dev Biol 2023, 147, 83–90. [Google Scholar] [CrossRef]
- Li, Z; Zhang, Y; Peng, B; et al. A novel interpretable deep learning-based computational framework designed synthetic enhancers with broad cross-species activity. Nucleic Acids Res 2024, 52, 13447–13468. [Google Scholar] [CrossRef]
- Peleke, FF; Zumkeller, SM; Gultas, M; et al. Deep learning the cis-regulatory code for gene expression in selected model plants. Nat Commun 2024, 15, 3488. [Google Scholar] [CrossRef] [PubMed]
- Moeckel, C; Mouratidis, I; Chantzi, N; et al. Advances in computational and experimental approaches for deciphering transcriptional regulatory networks: Understanding the roles of cis-regulatory elements is essential, and recent research utilizing MPRAs, STARR-seq, CRISPR-Cas9, and machine learning has yielded valuable insights. Bioessays 2024, 46, e2300210. [Google Scholar]
- Ho, C; Morsut, L. Novel synthetic biology approaches for developmental systems. Stem Cell Reports 2021, 16, 1051–1064. [Google Scholar] [CrossRef]
- Naffaa, MM. Bridging molecular mechanisms and therapeutic innovations: The role of brain organoids in neurodevelopmental disorder research. 2025. [Google Scholar] [CrossRef]
- Organoid Research 1.
- Kanton, S; Boyle, MJ; He, Z; et al. Organoid single-cell genomic atlas uncovers human-specific features of brain development. Nature 2019, 574, 418–422. [Google Scholar] [CrossRef]
- Won, H; Huang, J; Opland, CK; et al. Human evolved regulatory elements modulate genes involved in cortical expansion and neurodevelopmental disease susceptibility. Nat Commun 2019, 10, 2396. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, AE; Santos, SD. The ever-growing world of gastruloids: autogenous models of mammalian embryogenesis. Curr Opin Genet Dev 2023, 82, 102102. [Google Scholar] [CrossRef] [PubMed]
- Klein, JC; Keith, A; Agarwal, V; et al. Functional characterization of enhancer evolution in the primate lineage. Genome Biol 2018, 19, 99. [Google Scholar] [CrossRef]
- De Santis, R; Rice, E; Croft, G; et al. The emergence of human gastrulation upon in vitro attachment. Stem Cell Reports 2024, 19, 41–53. [Google Scholar] [CrossRef]
- Mezu-Ndubuisi, OJ; Maheshwari, A. Role of macrophages in fetal development and perinatal disorders. Pediatr Res 2021, 90, 513–523. [Google Scholar] [CrossRef]
- Park, JE; Botting, RA; Dominguez Conde, C; et al. A cell atlas of human thymic development defines T cell repertoire formation. Science 2020, 367. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y; Pan, W. Brain organoids: a new paradigm for studying human neuropsychiatric disorders. Front Neurosci 2025, 19, 1699814. [Google Scholar] [CrossRef] [PubMed]
- Li, K; Liu, Y; Cao, H; et al. Interrogation of enhancer function by enhancer-targeting CRISPR epigenetic editing. Nat Commun 2020, 11, 485. [Google Scholar] [CrossRef]
- Wang, K; Escobar, M; Li, J; et al. Systematic comparison of CRISPR-based transcriptional activators uncovers gene-regulatory features of enhancer-promoter interactions. Nucleic Acids Res 2022, 50, 7842–7855. [Google Scholar] [CrossRef] [PubMed]
- Karzbrun, E; Khankhel, AH; Megale, HC; et al. Human neural tube morphogenesis in vitro by geometric constraints. Nature 2021, 599, 268–272. [Google Scholar] [CrossRef]
- Rao, KS; Kameswaran, V; Bruneau, BG. Modeling congenital heart disease: lessons from mice, hPSC-based models, and organoids. Genes Dev 2022, 36, 652–663. [Google Scholar] [CrossRef]
- Abdel Fattah, AR; Daza, B; Rustandi, G; et al. Actuation enhances patterning in human neural tube organoids. Nat Commun 2021, 12, 3192. [Google Scholar] [CrossRef]
- Lee, JH; Shin, H; Shaker, MR; et al. Production of human spinal-cord organoids recapitulating neural-tube morphogenesis. Nat Biomed Eng 2022, 6, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Suhito, IR; Sunil, C; Tay, A. Engineering human immune organoids for translational immunology. Bioact Mater 2025, 44, 164–183. [Google Scholar] [CrossRef] [PubMed]
- Harter, MF; Recaldin, T; Gjorevski, N. Organoids as models of immune-organ interaction. Cell Rep 2025, 44, 116214. [Google Scholar] [CrossRef] [PubMed]
- Gunther, C; Winner, B; Neurath, MF; et al. Organoids in gastrointestinal diseases: from experimental models to clinical translation. Gut 2022, 71, 1892–1908. [Google Scholar] [CrossRef] [PubMed]
- Nikolaev, M; Mitrofanova, O; Broguiere, N; et al. Homeostatic mini-intestines through scaffold-guided organoid morphogenesis. Nature 2020, 585, 574–578. [Google Scholar] [CrossRef]
- Lewis-Israeli, YR; Wasserman, AH; Gabalski, MA; et al. Self-assembling human heart organoids for the modeling of cardiac development and congenital heart disease. Nat Commun 2021, 12, 5142. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M. Signaling Boundary Conditions Drive Self-Organization of Human “Gastruloids”. Dev Cell 2016, 39, 279–280. [Google Scholar] [CrossRef] [PubMed]
- Del Vecchio, D. Epigenetic memory: The role of the crosstalk between histone modifications and DNA methylation. Comput Struct Biotechnol J 2025, 27, 4019–4025. [Google Scholar] [CrossRef] [PubMed]
- Maier, JAH; Mohrle, R; Jeltsch, A. Design of synthetic epigenetic circuits featuring memory effects and reversible switching based on DNA methylation. Nat Commun 2017, 8, 15336. [Google Scholar] [CrossRef]
- Cakir, B; Xiang, Y; Tanaka, Y; et al. Engineering of human brain organoids with a functional vascular-like system. Nat Methods 2019, 16, 1169–1175. [Google Scholar] [CrossRef]
- Andersen, J; Revah, O; Miura, Y; et al. Generation of Functional Human 3D Cortico-Motor Assembloids. Cell 2020, 183, 1913–1929 e1926. [Google Scholar] [CrossRef]
- Takebe, T; Sekine, K; Enomura, M; et al. Vascularized and functional human liver from an iPSC-derived organ bud transplant. Nature 2013, 499, 481–484. [Google Scholar] [CrossRef]
- Di Stasi, A; Tey, SK; Dotti, G; et al. Inducible apoptosis as a safety switch for adoptive cell therapy. N Engl J Med 2011, 365, 1673–1683. [Google Scholar] [CrossRef]
- Deuse, T; Hu, X; Gravina, A; et al. Hypoimmunogenic derivatives of induced pluripotent stem cells evade immune rejection in fully immunocompetent allogeneic recipients. Nat Biotechnol 2019, 37, 252–258. [Google Scholar] [CrossRef]
- Simsek, E; Yao, Y; Lee, D; et al. Toward predictive engineering of gene circuits. Trends Biotechnol 2023, 41, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Hyun, I; Scharf-Deering, JC; Lunshof, JE. Ethical issues related to brain organoid research. Brain Res 2020, 1732, 146653. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, SO; Cappuccio, G; Kruth, K; et al. Rigor and reproducibility in human brain organoid research: Where we are and where we need to go. Stem Cell Reports 2024, 19, 796–816. [Google Scholar] [CrossRef] [PubMed]
- Lamm, N; Ben-David, U; Golan-Lev, T; et al. Genomic Instability in Human Pluripotent Stem Cells Arises from Replicative Stress and Chromosome Condensation Defects. Cell Stem Cell 2016, 18, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Lackner, M; Helmbrecht, N; Paabo, S; et al. Detection of unintended on-target effects in CRISPR genome editing by DNA donors carrying diagnostic substitutions. Nucleic Acids Res 2023, 51, e26. [Google Scholar] [CrossRef]
- Su, Z; Dong, H; Fang, X; et al. Frontier progress and translational challenges of pluripotent differentiation of stem cells. Front Genet 2025, 16, 1583391. [Google Scholar] [CrossRef]
- Hanna, RE; Doench, JG. Design and analysis of CRISPR-Cas experiments. Nat Biotechnol 2020, 38, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Li, A; Mitsunobu, H; Yoshioka, S; et al. Cytosine base editing systems with minimized off-target effect and molecular size. Nat Commun 2022, 13, 4531. [Google Scholar] [CrossRef] [PubMed]
- Henry, MP; Hawkins, JR; Boyle, J; et al. The Genomic Health of Human Pluripotent Stem Cells: Genomic Instability and the Consequences on Nuclear Organization. Front Genet 2018, 9, 623. [Google Scholar] [CrossRef]
- Andrews, PW; Barbaric, I; Benvenisty, N; et al. The consequences of recurrent genetic and epigenetic variants in human pluripotent stem cells. Cell Stem Cell 2022, 29, 1624–1636. [Google Scholar] [CrossRef]
- Ghosh, S; Brown, AM; Jenkins, C; et al. Viral Vector Systems for Gene Therapy: A Comprehensive Literature Review of Progress and Biosafety Challenges. Appl Biosaf 2020, 25, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H; Ekstrom, K; Bossios, A; et al. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Arendt, D; Musser, JM; Baker, CVH; et al. The origin and evolution of cell types. Nat Rev Genet 2016, 17, 744–757. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, MA; Corsini, NS; Wolfinger, S; et al. Guided self-organization and cortical plate formation in human brain organoids. Nat Biotechnol 2017, 35, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Helenek, C; Krzyszton, R; Petreczky, J; et al. Synthetic gene circuit evolution: Insights and opportunities at the mid-scale. Cell Chem Biol 2024, 31, 1447–1459. [Google Scholar] [CrossRef]
- Gjorevski, N; Sachs, N; Manfrin, A; et al. Designer matrices for intestinal stem cell and organoid culture. Nature 2016, 539, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Lavazza, A; Massimini, M. Cerebral organoids: ethical issues and consciousness assessment. J Med Ethics 2018, 44, 606–610. [Google Scholar] [CrossRef]
- Nicolas, P; Etoc, F; Brivanlou, AH. The ethics of human-embryoids model: a call for consistency. J Mol Med (Berl) 2021, 99, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Rivron, NC; Martinez Arias, A; Pera, MF; et al. An ethical framework for human embryology with embryo models. Cell 2023, 186, 3548–3557. [Google Scholar] [CrossRef]
- De Miguel Beriain, I; Rueda, J; Villalba, A. Re-defining the human embryo: A legal perspective on the creation of embryos in research. EMBO Rep 2024, 25, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Pennings, G; Dondorp, W; et al.; Writing Group of the EEC Ethical considerations on the moral status of the embryo and embryo-like structuresdagger. Hum Reprod 2024, 39, 2387–2391. [Google Scholar] [PubMed]
- Koplin, JJ. Response to the ISSCR guidelines on human-animal chimera research. Bioethics 2023, 37, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Real, R; Peter, M; Trabalza, A; et al. In vivo modeling of human neuron dynamics and Down syndrome. Science 2018, 362. [Google Scholar] [CrossRef]
- Ou, Y; Guo, S. Safety risks and ethical governance of biomedical applications of synthetic biology. Front Bioeng Biotechnol 2023, 11, 1292029. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Synthetic Gene Circuit Architectures Underlying Programmable Developmental Control.

Figure 2.
Epigenetic Engineering Strategies for Stable Lineage and Pattern Control.
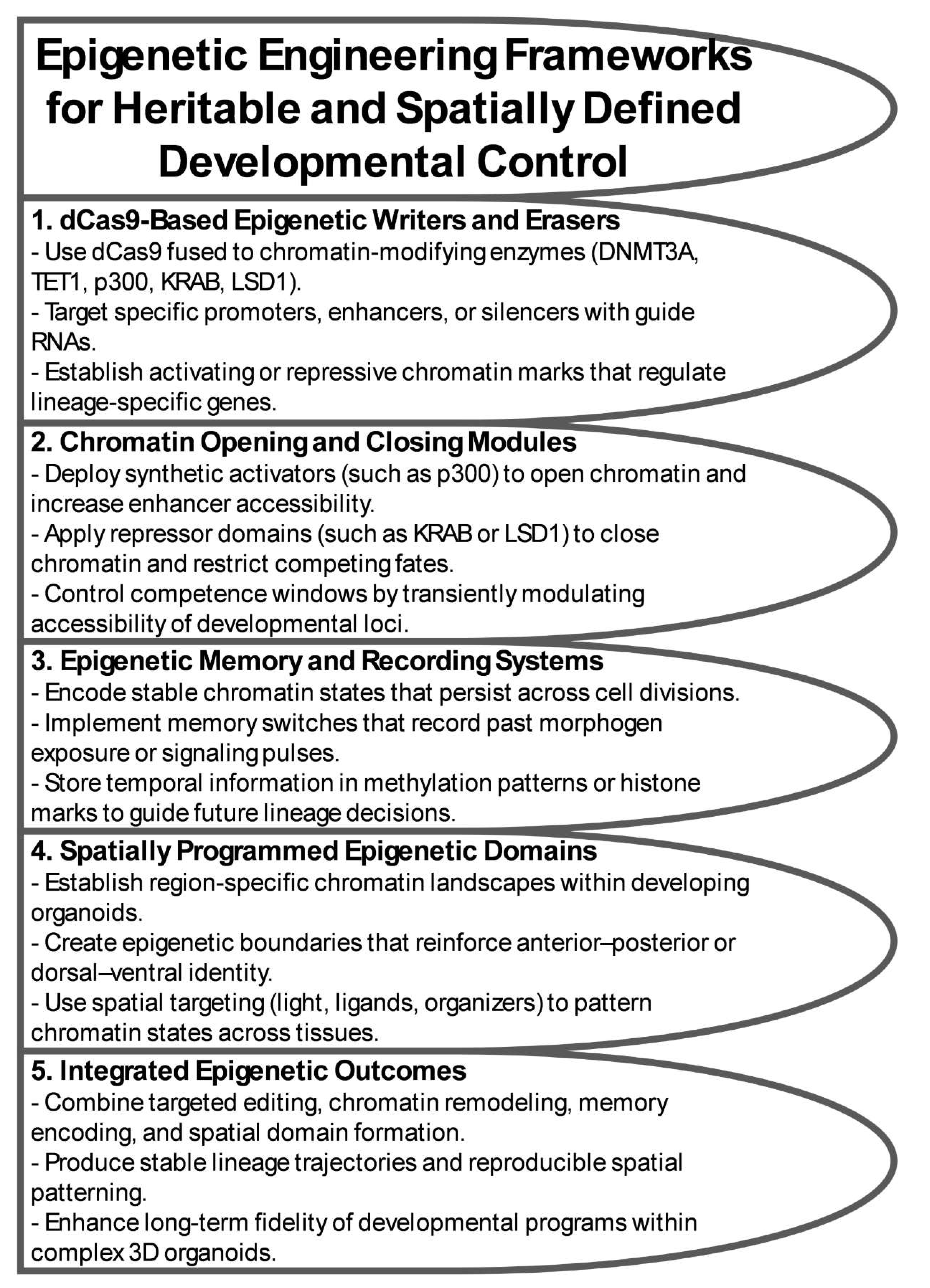
Figure 3.
Engineered Spatial and Temporal Patterning in Organoids.
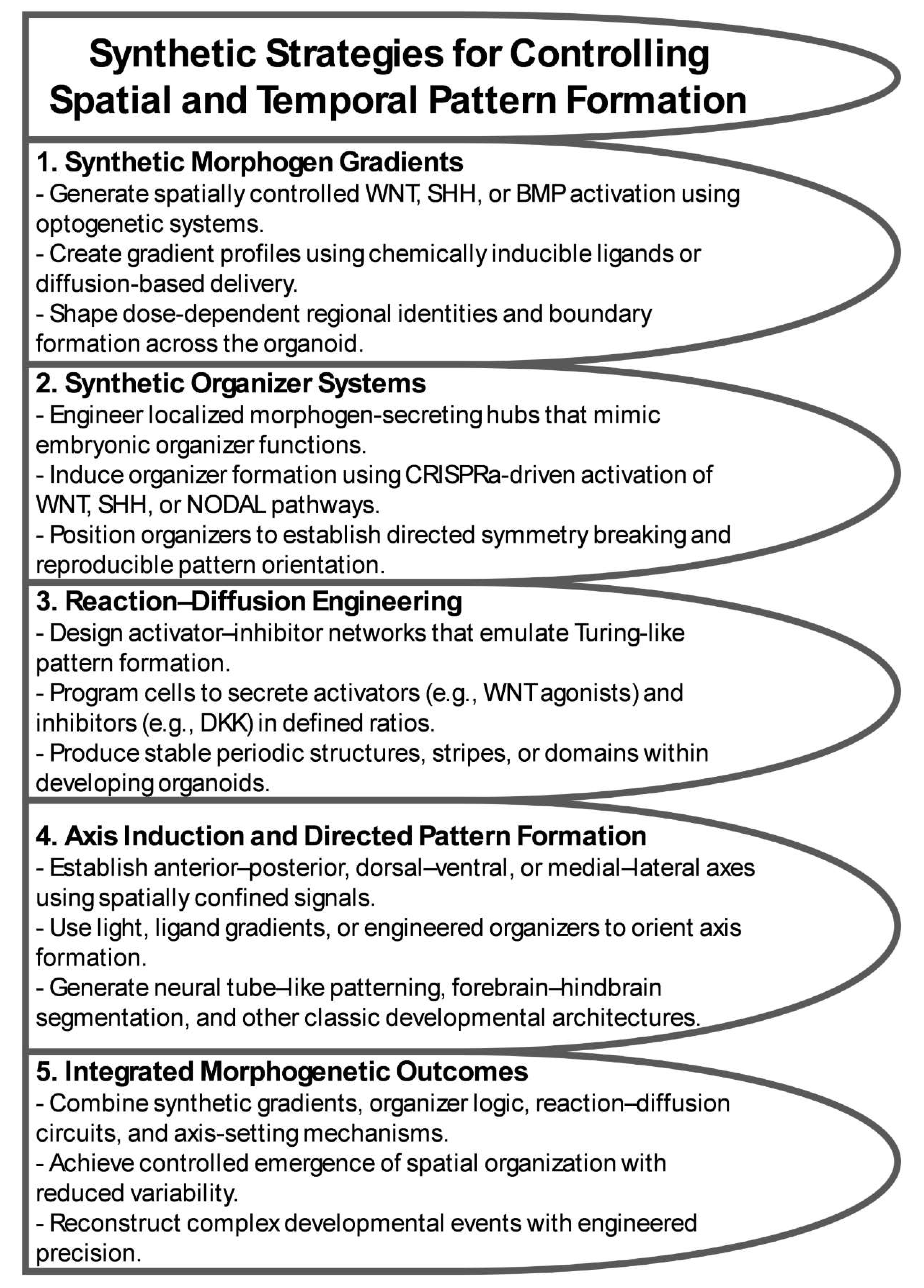
Figure 4.
Applications of Programmable Organoids in Human Development, Disease Modeling, and Regenerative Medicine.
Figure 4.
Applications of Programmable Organoids in Human Development, Disease Modeling, and Regenerative Medicine.

Figure 5.
Roadmap Toward Fully Programmable Human Developmental Systems.
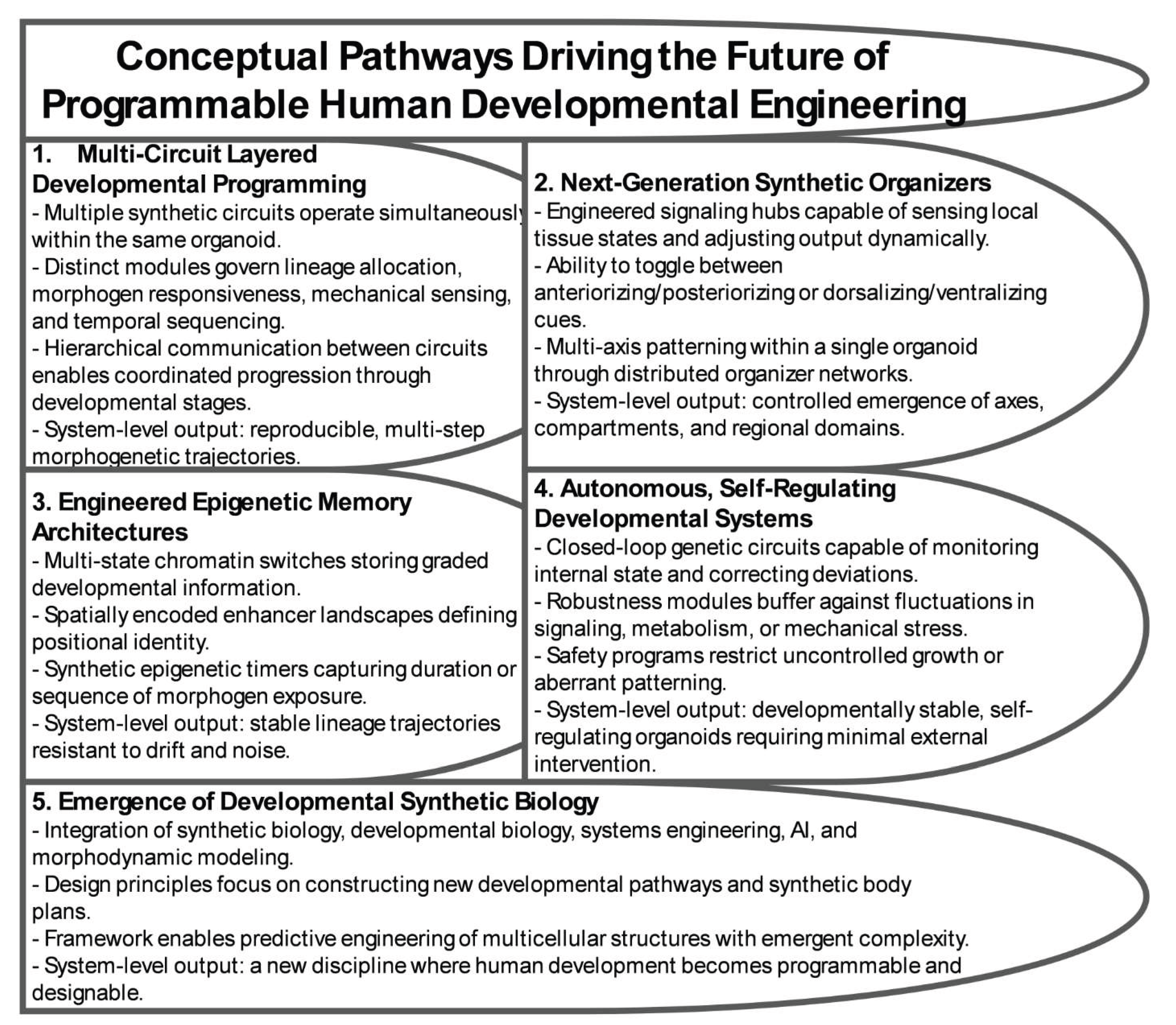

Table 1.
Biosensors and Feedback Control Modules for Programmable Development.
| Sensor / Control Module | Detection Modality | Response Speed | Integration with Synthetic Circuits | Developmental Role |
|---|---|---|---|---|
| Fluorescent Pathway Reporters (e.g., WNT, SHH, BMP, ERK reporters) | Fluorescent protein expression downstream of pathway-responsive elements | Minutes to hours | Provides real-time readout of morphogen activity; can feed signals into downstream logic gates | Tracks lineage decisions, monitors signaling gradients, validates circuit performance |
| Luminescent Reporters | Luciferase expression controlled by signaling-responsive promoters | Minutes to hours (low noise) | Compatible with long-term monitoring and closed-loop algorithms | Non-invasive tracking of slow developmental processes, stability assessments |
| RNA-Based Toehold Switches / Riboregulators | RNA hybridization, endogenous transcript detection | Seconds to minutes | Direct coupling to CRISPRa/i or translational control modules | Context-dependent activation of lineage programs, sensing endogenous developmental states |
| Metabolic Aptamer Sensors (e.g., ATP, SAM, NAD+, acetyl-CoA) | Metabolite binding to RNA aptamers | Seconds | Link metabolic state to gene circuit activation; modulate fate transitions | Synchronizes metabolic state with developmental progression, models metabolic disorders |
| Mechanosensitive Gene Reporters (e.g., YAP/TAZ activity sensors) | Nuclear localization or transcription driven by mechanical forces | Seconds to minutes | Feeds mechanical information into gene-expression circuits | Regulates morphogenesis, symmetry breaking, and niche sensing |
| Synthetic Feedback Controllers (negative or positive feedback) | Internal sensing of circuit output or key developmental genes | Fast to intermediate | Maintains gene expression within defined ranges; buffers noise | Stabilizes lineage identity, shapes gradients, enforces temporal order |
| Integral Feedback Controllers | Accumulates deviations over time; adjusts circuit output | Slow to intermediate | Ensures robust homeostasis even under perturbation | Maintains stable morphogen thresholds and timing of transitions |
| Optogenetic Feedback Loops | Light-controlled sensing and actuation | Milliseconds to seconds | Enables precision tuning of pathway activity using ML-guided stimulation | Fine control of spatial patterning, gradient sharpening, and axis formation |
| Chemogenetic Control Modules (e.g., Tet-On, CreERT2, rapalog switches) | Small-molecule activation | Minutes to hours | Couples external control inputs to developmental regulators | Inducible lineage switching, timed activation of transient competence windows |
| Hybrid Cybergenetic Systems (cell–computer feedback) | Live sensing + computational prediction | Variable; usually fast | Reinforcement learning or ML algorithms adjust circuit inputs | Predictive control of morphogenesis, pattern stabilization, correction of deviations |
Table 2.
Technical and Ethical Challenges in Programmable Organoid Systems.
| Challenge | Description | Risk | Mitigation Strategy |
|---|---|---|---|
| Off-target genetic or epigenetic activity | Unintended CRISPRa/i activation, base-editing changes, or ectopic chromatin modification | Aberrant lineage specification, mispatterning, unpredictable morphogenesis | High-fidelity Cas variants, genome-wide off-target profiling, orthogonal control systems |
| Genetic instability of synthetic circuits | Mutations, silencing, or loss of constructs during long-term culture or high transcriptional load | Circuit collapse, emergence of escape mutants, divergence across replicates | Safe-harbor integration, low-burden design, redundancy in key modules |
| Epigenetic drift and memory erosion | Loss or blurring of engineered chromatin states during division or stress | Loss of lineage fidelity, mosaic chromatin patterns, reduced reproducibility | Error-correcting epigenetic circuits, re-establishment modules, longitudinal chromatin profiling |
| Horizontal transfer of circuit components | DNA/RNA elements spreading via EVs, viral vectors, or cell fusion | Mislocalized circuit function, loss of spatial specificity, unintended lineage rewiring | Genetic insulation motifs, compartment-specific promoters, containment sequences |
| Unintended emergent self-organization | Nonlinear interactions between endogenous and synthetic signals produce unplanned patterning | Aberrant domains, spontaneous symmetry breaking, uncontrolled morphogenetic events | Real-time imaging, predictive modeling, negative-feedback designs |
| Evolution or drift of embedded circuits | Adaptive mutation or silencing that alters thresholds or logic | Loss of developmental control, variability between replicates | Evolutionarily stable design principles, periodic reseeding, multi-layer redundancy |
| Lineage rewiring in human-derived tissues | Engineering neural, germline-like, or embryo-like domains raises conceptual concerns | Ambiguity regarding biological status or moral boundaries | Defined lineage limits, ethical oversight, transparent reporting |
| Embryo-like developmental features | Synthetic organizers and patterned epigenetic systems approach early embryonic structure | Regulatory uncertainty, potential reclassification as embryo models | Adherence to ISSCR guidelines, developmental boundary definitions |
| Governance gaps for autonomous/AI-coupled systems | Lack of regulatory frameworks for self-regulated or ML-guided organoids | Biosafety concerns, ungoverned autonomy, reduced public trust | Algorithmic transparency, data-provenance safeguards, interdisciplinary governance |
| Clinical translation risks | Uncertain long-term stability of transplanted engineered tissues | Uncontrolled proliferation, mispatterning, immune activation | Safety switches, growth-restriction modules, staged preclinical testing |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



