
Submitted:
13 December 2025
Posted:
15 December 2025
You are already at the latest version
Abstract
Research on gut-microbiota (GM) changes in infants and children with intestinal surgical conditions, and how these changes might leverage to improve outcomes, is progressing rapidly. Nonetheless, there is a lack of information regarding the role of GM following surgery for the most prevalent intestinal pediatric disorders that necessitate surgical intervention, despite it receiving considerable interest. Pre-surgical dysbiosis and post-surgical GM assessment of these disorders are still poorly understood. This detailed review has gathered insights into the current understanding of unique alterations in GM colonization following surgery for significant childhood conditions, such as necrotizing enterocolitis (NEC), Hirschsprung’s disease (HD), inflammatory bowel disease (IBD), and short bowel syndrome (SBS).In particular, surgery for NEC may result in a reduction in GM diversity. Surgical procedures for HD can alter the GM, potentially causing shifts in GM compositions and increasing the likelihood of complications such as Hirschsprung’s associated enterocolitis (HAEC). For children diagnosed with IBD, surgical interventions can bring about changes in the diversity and structure of GM, which may lead to disease recurrence or affect the success of treatment. Children with SBS, following extensive bowel resections, display abnormal GM profiles when compared to healthy children. These GM patterns may include diminished. GM diversity, an increase in inflammation-related bacteria, and a decrease in beneficial bacteria. Moreover, the most common complications that occur after surgical procedures for the above-mentioned intestinal disorders are greatly affected by the GM. Current research provides an initial understanding of the possible post-surgical implications for outcomes of these intestinal disorders. Future studies could clarify GM alterations associated with various intestinal paediatric surgical procedures and their complications, which may influence the evaluation of GM-targeted treatments.
Keywords:
gut microbiota
; children
; intestinal disorders
; dysbiosis
; surgery
; post-surgical alterations
; post-surgical complications
1. Introduction
The human gut microbiota (GM), a complex ecosystem of trillions of microorganisms, is now recognizedas a critical determinant of human health, playing a critical role in digestion, immune modulation, metabolism, and barrier function [1]. This delicate community is particularly vulnerable in early life, and its establishment and stability are crucial for normal growth and development in paediatric patients [2].
Surgical procedures are often a life-saving necessity for a spectrum of pediatric intestinal disorders, such as necrotizing enterocolitis (NEC), Hirschsprung's disease (HD), intestinal atresia, anorectal malformations, and inflammatory bowel disease (IBD)[3,4,5,6,7]. While these procedures aim to restore anatomical function and resolve the primary pathology, they represent a major physiological insult. The process of surgery itself in combination with factors like antibiotic administration, bowel preparation, alterations in diet, and post-operative fasting, can cause profound and potentially long-lasting disruptions to the GM. This post-surgical dysbiosis, characterized by a loss of beneficial commensals and an expansion of pathobionts, is increasingly implicated in a range of post-surgical complications, including infections, anastomotic leaks, and prolonged recovery [8,9].
Despite its clinical significance, the unique nature of post-surgical GM alterations in the paediatric intestinal patient population remains poorly characterized [10,11,12]. A comprehensive understanding of these shifts of microorganisms is essential to move beyond simple observational studies and towards developing targeted interventions,such as using antibiotics, probiotics, prebiotics, or tailored nutritional strategies, aimed at restoring a healthy microbiota and improving clinical outcomes [13,14,15,16,17,18].
This narrative review aims to synthesize current evidence on the impact of intestinal surgery on the GM of paediatric patients. We will explore common patterns of dysbiosis across different disorders and procedures, discuss the potential clinical implications of these alterations, and highlight avenues for future therapeutic GM modulation.
2. Methodology
2.1. Search Strategy
An extensive literature search was conducted across electronic databases, including Pubmed/Medline, Scopus and Google Scholar, to find pertinent studies published from January 1, 2010 to August 31, 2025. The focus was on articles that studied alterations of the GM following surgery for pediatric intestinal disorders, namely NEC, HD, IBD, and short bowel syndrome (SBS). The search employed Boolean operators (AND, OR) to combine keywords including “gut microbiota”, “paediatric surgery”, “intestinal disorders”, “dysbiosis”, “children”, “infants”, “neonates”, necrotizing enterocolitis”, Hirschsprung's disease”, “Hirschsprung -associated enterocolitis”, “inflammatory bowel disease”, “small bowel syndrome”, “post-surgical infection”, “post-surgical intestinal obstruction”, “anastomotic leak”.
2.2. Inclusion Criteria
Studies were chosen based on established inclusion and exclusion criteria. Randomized controlled trials, meta-analyses, prospective and retrospective observational studies, systematic reviews and review articles were incorporated in the analysis. In addition, this narrative review included studies performed in animal subjects. Clinical guidelines and expert recommendations were also taken into account. Only articles published in English language were assessed. The exclusion criteria included articles published in languages other than English, case reports, editorials and conference abstracts lacking sufficient data.
2.3. Process of Selecting Studies
Two reviewers (N.V. and N.Z.) conducted the data search and analysis. The selection of studies was driven by their relevance to the review’s objectives rather than strictly by the systematic inclusion criteria. Titles and abstracts were first screened, followed by a thorough examination of full texts. Moreover, the reference lists of the selective articles were further screened for additional relevant studies.
2.4. Synthesis of Data
Studies were synthesized narratively. An overview of GM alterations following surgery for paediatric intestinal disorders, namely NEC, HD, IBD, and SBS, was included to clarify and underscore the significance of the study’s aim. Key arguments from the studies (e.g. GM-related post-surgical alterations and complications of intestinal surgery in pediatric patients) were extracted to create several thematic sections. This approach aided in the synthesis of heterogeneous evidence.
3. Physiology and Function of Gut Microbiota
The GM refers to the diverse ecosystem of microorganisms, including bacteria, fungi, viruses, and protozoa, that reside in the human intestinal tract [19]. Based on culture-independent molecular methods, the genetic material (the microbiome) contains around 3.3 million genes which is about 100 times more than the human genome. Of the identified genes, 99% were of bacterial origin. The entire cohort harbored a collective pool of 1000 to 1150 prevalent bacterial species, with each individual contributing at least 160 such species. The substantial sharing of these species highlights the vast functional potential of the GM to influence human health [20]. Key factors influencing the composition and function of the GM encompass host genetics, dietary patterns, age, lifestyle, geographic location, and medication use, most notably antibiotics [21].
The advent of novel culture-independed techniques, most notably high throughput sequencies of bacteria 16S ribosomal RNA (rRNA) gene, has fundamentally transformed our comprehension of the GM [22]. While the 16S rRNA gene is a powerful taxonomic tool due to its universal conservation, shotgun metagenomics overcomes its limitations by providing robust, amplification-free estimates of entire communities of microorganisms [23]. Initiatives like Metagenomics of the Human Intestinal Tractand the Human Microbiome Project have leveraged this method to comprehensively map the human GM [24].
Based on new molecular techniques and methodologies, our understanding of the gut microbiota's taxonomic landscape has been refined. It is now established that the adult gastrointestinal tract is dominated by a few key bacterial phyla. Among these, Firmicutes (primarily Gram-positive clostridia) and Bacteroidetes (primarily Gram-negative bacteria) are the most prevalent, collectively accounting for the 90% of species [25,26]. Together with the less abundant but critically important phyla Actinobacteria and Proteobacteria, Fusobacteria and Verrumicrobia, they constitute not only the most numerous but also the most functionally diverse microorganisms in the gut ecosystem [27]. Furthermore, the Firmicutes/Bacteroidetes ratio is dynamic across the human lifespan, reported at approximately 0.4 in infants, 0.9 in adults, and 0.6 in the elderly, with no significant difference observed between infant and elderly ratios [28].
A healthy and diverse GM is critical for numerous essential functions, such as intestinal homeostasis, host immunity and systemic health, extending far beyond a purely structural role [29]. For instance, key species of microorganisms, mainly from the phyla Bacteroidetes and Firmicutes, perform this role by fermenting indigestible fibers into various products. These products include, short-chain fatty acids (SCFAs), hydrogen, vitamin K, carbon dioxide, ammonia, amines, and phenolic compounds [30].SCFAs, primarily acetate, propionate, and butyrate, at a molar ratio 60:20:20 respectively, account for approximately 95% of the total SCFAs and are the GM’s main metabolites, produced in the colon by the bacterial fermentation of dietary fibers and other non-digestible carbohydrates [31,32,33]. Main functions include providing cellular energy, exerting anti-inflammatory and anti-tumor effects, regulating appetite, and fortifying the intestinal barrier through the upregulation of tight junction proteins [34,35,36].In addition, SCFAs through sophisticated, bidirectional communication networks, known as gut-organ axes,maintain the systemic physiological balance [37]. In the gut-brain axis, SCFAs can stimulate the synthesis of serotonin from enterochromaffin cells, a process that influences cognitive functions such as learning and memory [38]. In the gut-lung axis, SCFAs modulates respiratory health by mitigating pulmonary inflammation and protecting against infections like pneumonia through systemic immunomodulatory effects [39]. In the gut-liver axis, butyrate ameliorates metabolic diseases like insulin resistance and obesity and directly protects against hepatic ailments, including non-alcoholic liver disease, non-alcoholic steatohepatitis, inflammation, cancer, and liver injury [40]. In the gut-kidney axis, SCFAs play a protective role in kidney health. In the chronic kidney disease (CKD), however, a disruption of GM decreases the population of SCFA-producing bacteria such as Bifidobacterium and Lactobacillus, undermining this protective effect [41]. Lower SCFA concentrations compromise the gut barrier, leading to increase permeability and presence of endotoxins in the blood stream, which subsequently triggers inflammation and oxidative stress in the kidney [42]. Importantly, a recent systematic review confirmed a distinct gut dysbiosis in CKD patients, characterized by a loss of beneficial bacteria and an expansion of pathogens [43]. In the gut-heart axis, SCFAs have become key-mediators for comprehending the complex interplay between gut health and cardiovascular function, There is strong evidence that the cardiovascular health may begin in the gut, as early as during infancy. Any imbalance during this period may increase the cardiovascular dysfunction later in adult life [44]. For example, elevated levels of the phylum Pseudomonadota observed in preterm infants are also found in some adults with atherosclerotic cardiovascular disease [45].
4. Unique Characteristics of the Pediatric Gut Microbiota
The GM of an infant differs from that of an adult in terms of both its composition and its function, characterized by greater instability and heterogeneity [46,47]. Actually, the GM development in early infancy is influenced by three factors, that contribute during the prenatal period, the delivery, and the postnatal period. During these phases, the GM development is shaped by a multitude of factors, including maternal transmission during birth, delivery mode, maternal health, antibiotic exposure, and diet [48].
The long-held notion of a sterile fetal environment is now contested [49]. Evidence of microorganisms in meconium, the placenta, and amniotic fluid supports that infant gut colonization begins in utero [50,51,52], potentially via translocation of maternal gut microorganisms through the bloodstream. The clinical significance of this process is underscored by its association with pregnancy outcomes like preterm birth and low birth weight full-term infants [53]. The most dramatic shift of microorganisms, however, occurs at birth, establishing a foundation that is shaped by a convergence of early-life factors. The mode of delivery is a primary determinant: vaginally delivered infants acquire a microbiota resembling maternal vaginal flora including Lactobacilli, Prevotella, and group B Streptococci, while those born via Caesarian section are initially colonized by maternal skin-associated bacteria (e.g., Staphylococcus, Corynebacterium) [54,55,56,57]. This initial inoculum is then profoundly modified by postnatal exposures, most notably feeding type. Breastfed infants typically exhibit a GM dominated by beneficial Bifidobacterium species, whereas formula-fed infants is associated with a higher prevalence of taxa like Escherichia coli, Clostridium difficile, Bacteroides, and lactobacilli [58,59]. However, for preterm and very low birth weight infants, these natural processes are often disrupted by the clinical necessities of NICU hospitalization. Practices such as antibiotic use, respiratory support, and altered feeding protocols can lead to a dysbiotic state characterized by reduced Bifidobacteria and elevated levels of opportunistic pathogens like Enterococcus and Staphylococcus [60,61]. Contrary to this common clinical expectation, Li et al. [62], in a prospective cohort study,analyzing the GM of 46 term and 23 preterm infants over the first three months of life, revealed a more complex and ultimately positive trajectory. The authors confirmed that gestational age was the primary factor shaping the initial meconium microbiota. However, instead of a persistently dysbiotic state, the preterm infant GM demonstrated a notable drive towards normalization from one month of postnatal age. Indeed, homogeneous and increasing abundance of the beneficial bacteria Bifidobacterium and Lactobacillus, alongside a gradual decrease in potentially pathogenic genera was noted. This suggests a resilient succession of microorganisms in preterm infants that can, under certain conditions, overcome early clinical insults.
Following its initial establishment at birth, the GM embarks on a predictable successional journey through early childhood. Stewart et al. [63] provided the foundational roadmap for this process, defining three sequential phases: a developmental phase (3–14 months) of high volatility, a transitional phase (15–30 months) of increasing stability, and a final stable phase (31–46 months). Their multinational multi-center study identified that progression along this timeline is primarily driven by dietary changes, specifically the cessation of breastfeeding, which acts as a switch to reduce Bifidobacterium and promote a Firmicute-rich, more mature community. However, this core developmental program is then modulated by other powerful factors. For instance, birth mode sets the initial trajectory, with vaginal delivery enriching for Bacteroides and accelerating early maturation. Moreover, Yatsunenko et al. [64] showed that, while the functional progression is universal, the specific species of microorganisms that constitute the stable adult-like state differ dramatically between industrialized and non-industrialized populations. Finally, the functional programming of this assembly process is age-specific: unlike adults, whose GM was oriented towards inflammatory functions, the paediatric GM is primed to support ongoing growth and development. [65]
5. Disruption of the Gut Microbiota in Intestinal Paediatric Surgical Diseases
The physiological trauma of surgery introduces a multitude of disruptive factors across the perioperative timeline, including the underlying disease itself [66], the composition of GM [67], the production of key metabolites from species of microorganisms [68,69], and iatrogenic elements such as mechanical bowel preparation [70], antibiotics [71], anesthetic regimens [72], postoperative pain [73], and anastomotic techniques [74]. These collective insults profoundly disrupt the symbiotic milieu, leading to a persistent dysbiosis that can be described as a long-term "scarring" community of microorganisms, which hinders the restoration of its pre-surgical composition [75].
In paediatric patients, the disruption of the GM (dysbiosis) is a cornerstone of intestinal pathology. This is well-established in a spectrum of functional disorders including celiac disease, IBD, and irritable bowel syndrome, where specific dysbiotic signatures actively contribute to disease [76,77]. This inherent vulnerability is profoundly amplified in the context of surgical disease [78]. A striking example is found in neonates with congenital gastrointestinal surgical conditions. Research demonstrates that these infants undergo a severe and rapid ecological collapse of the GM within the first two weeks of life, a period that encompasses both their underlying pathology and initial surgical intervention [79]. While their GM is initially comparable to healthy infants, it fails to develop normally. By the second week, a dramatic dysbiosis emerges, characterized by a significant decrease of beneficial genera like Bifidobacterium and Bacteroides and a bloom of pathobionts such as Escherichia,Shigella and Pseudomonas. This failure of microorganisms is coupled with a profound and persistent deficiency in beneficial SCFAs, which stagnate instead of increasing the microorganisms, as they do in healthy infants. This combination, a pathobiont-dominated community of microorganisms and a critical metabolic deficit, presents a fundamental failure in the establishment of a healthy gut ecosystem, highlighting the extreme fragility of the GM in the paediatric surgical patient [79].
The above-mentioned pattern of harmful bacteria is more evident in surgical disorders such as NEC, HD, Hirschsprung’s-associated enterocolitis (HAEC), IBD and SBS. In NEC, dysbiosis is characterized by a bloom of bacteria such as GammaProteobacteria, Klebsiella, Actinobacteria, Clostridia, Enterococcus and Staphylococcus and a decrease of Firmicutes [80]. This leads to a damaging metabolic profile, including an overproduction of butyric acid that induces mucosal damage. In this “butyrate paradox” the butyric acid becomes pathogenic in the immature gut of infants [69,81]. This is coupled with a deficiency in secondary bile acids, which disrupts the Toll-like Receptor 4 pathway, exacerbating inflammation and disease progression [82]. In HD, the risk of HAEC is defined by a distinct shift of microorganisms and characterized by a dominance of Enterobacteriaceae, followed by Enterococcus and Acinetobacter [83]. In an iatrogenic rectosigmoid hypoganglionosis rat model, Budianto et al. [84] showed that neuronal disruption triggers a progressive alteration of the GM, characterized by an increasing abundance of Proteobacteria and a decrease of Firmicutes over 12 weeks. This escalating dysbiosis was accompanied by a worsening of enterocolitis, indicating a time-dependent relationship between the shift of microorganisms and disease severity. Furthermore, Arnaud et al. [85] in a neonatal porcine model of inatrogenicaganglionosis, evaluated the functional consequences of aganglionosis on epithelial barrier function and GM compared to healthy control. The results showed decreased tight junction protein and greater levels of the pro-inflammatory bacteria Fusobacterium (Fusobacteria), and Biophilia (Proteobacteria), and Mogibacterium (Firmicutes) and Biophilia (Proterobacteria) while the control group characterized by greater levels of Firmicutes and bacteria of Lachnospiracae family. In contrast, HD patients without enterocolitis maintain a more balanced GM where the Bacteroides genus is most prevalent. This indicates that a shift away from Bacteroides and toward an Enterobacteriaceae-driven community is a key risk factor for inflammatory complications. This specific dysbiotic profile now finds support from genomic evidence. Indeed, recent Mendelian randomization studies [86,87] have demonstrated that this microbial imbalance is not merely associative but causal, identifying taxa like the protective Peptococcus and Ruminococcusand the risk-associated Eggerthella as direct genetic contributors to HD susceptibility. Furthermore, research has uncovered that the depletion of beneficial bacteria like Peptococcusleads to a critical deficiency in protective metabolites, such as stearoyl sphingomyelin and lysine, which in turn facilitate disease development [87]. Furthermore, in IBD, a characteristic shift of microorganisms is observed. For instance, pediatric Crohn's disease (CD) is marked by an increased abundance of Proteobacteria (specifically Enterobacteriaceae) alongside decreased levels of Actinobacteria and Bacteroidetes [88]. A similar pattern in ulcerative colitis (UC) features a rise in Proteobacteria and a concurrent decline in Firmicutes. Conversely, paediatric patients with SBS may show alterations in GM for factors including delayed enteral diet, surgical resection of intestine, intestinal dysmotility, bacterial translocation, and bacteremia [89,90]. The exact mechanism of GM alterations has not fully understood, but factors such as mucosal inflammation and bacterial by-products, has been shown to implicate [91,92,93,94,95], Indeed, several articles have documented this pattern of GM diversity in paediatric patients with SBS [91,92,93,94,95]. Lilja et al. [91], compared neonates diagnosed with SBS with healthy siblings, by using 16S rRNA gene sequencing in fecal samples. They reported an overall decreased bacteria diversity and an abundance of Enterobacteriacaein neonates with SBS. In another study, it was found an increase ofEnterobacteriacae (p=0.038) in children experiencing poor growth compared with children with SBS and adequate growth [92]. Of interest is also a study that reported alterations in the GM composition in children with SBS who receive prophylactic antibiotics for a long period [95].
6. Impact of Surgical Stress on Gut Microbiota in Paediatric Patients
Surgical stress induces a physiological stress response that directly disrupts the GM, with significant implications for recovery [66]. This is demonstrated by a consistent pattern of dysbiosis across various procedures. For example, in rat models, surgery triggers a localized bloom of pathogens like Enterococcus, Escherichia and Shigella at the anastomotic site [96]. Moreover, surgery-induced gut instability extends beyond abdominal procedures, as demonstrated in infants with congenital heart disease undergoing cardiopulmonary bypass. Infants with a spectrum of congenital heart diseases exhibit a predictable pre-surgical dysbiosis, characterized by a significantly greater abundance of Proteobacteria and Actinobacteria and a depletion of Bacteroides compared to controls. This process is then exacerbated by the surgical stress of cardiopulmonary bypass [97]. The disruption of microorganisms was directly linked to clinical risk, as these patients also developed significant post-surgical intestinal barrier dysfunction, measured by elevated serum markers [98,99,100]. Since this barrier damage occurred specifically after cardiopulmonary bypass and not in control infants, it emphasizes that the procedure itself rather than the cardiac defect alone is mainly associated with the gut injury. This paradigm of early surgical stress setting the stage for long-term sequelae is further evidenced by studies showing that children with histories of major surgery harbor altered GM years later, which are linked to persistent pathologies ranging from chronic diarrhea to anxiety [101,102]. The above-describedevidence establishes that the initial physiological insult of surgery can trigger a cascade of dysfunction of microorganisms, critically influencing both short-term recovery and long-term health.
7. Alterations of Gut Microbiota After Intestinal Surgery in Paediatric Diseases
7.1. Necrotizing Enterocolitis
NEC remains a devastating disease in newborns. For infants who require surgery, mortality rates persist at a tragically high 20-30%, despite advancements in neonatal care [103]. The pathogenesis of NEC is influenced by many factors such as prematurity, enteral feeding, and dysbiosis, the latest seeming to play a critical role [104]. While the role of the GM in the initial onset of NEC has been extensively investigated [105,106,107,108], the landscape of microorganisms after surgical intervention remains almost entirely unexplored. Within this research gap, Lin et al. [10] prospectively investigated the dynamic evolution of the GM following surgical intervention for NEC. Their study enrolled a cohort of preterm infants diagnosed with NEC who required surgical management, alongside a matched control group of preterm infants. Fecal specimens were collected at different clinical periods: at the time of NEC diagnosis, upon initiation of refeeding, and after the establishment of full enteral nutrition. Parallel samples were obtained from control infants at corresponding periods. Analysis of the GM revealed significantly lower alpha-diversity, as measured by the Shann on and Simpson indices, in the NEC patients after achieving full enteral nutrition compared to matched controls at the same nutritional period (p <.05).Taxonomic analysis identified a higher relative abundance of Methylobacterium, Clostridium butyricum, and Acidobacteria in the NEC group at diagnosis. Methylobacterium and Acidobacteria remained significantly more abundant in the NEC group. Furthermore, the abundance of these bacterial taxa demonstrated a significant positive correlation with serum CRP levels and a significant negative correlation with platelet count. At the 12-month corrected age follow-up, the rate of delayed growth was higher in the NEC group (25%) than in the control group (7.1%). However, this difference was not statistically significant. Furthermore, functional analysis revealed a significantly heightened activity in ketone body metabolism among infants with NEC, offering valuable insights into a potential metabolic link to the disease. Ketone bodies, particularly β-hydroxybutyrate, are not merely energy substrates but also key signaling molecules known to influence intestinal stem cell fate and modulate the GM by selectively inhibiting beneficial bacteria like Bifidobacterium, which can suppress protective Th17 immune responses [109]. This suggests that the observed metabolic shift is not just a consequence but may actively contribute to NEC pathogenesis by disrupting epithelial regeneration and immune homeostasis [110]. Key findings include a long-term reduction in diversity of microorganisms, enrichment of resilient taxa like Methylobacterium, and a distinct metabolic profile marked by elevated ketone body metabolism and diminished sphingolipid activity.
7.2. Hirschsprung’s Disease and Hirschsprung’s-associated Enterocolitis
HD is a congenital disorder causing functional intestinal obstruction from aganglionosis [111]. While surgical resection of the aganglionic segment is the definitive treatment, approximately one-third of patients suffer postoperative complications. The most severe of these complications is HAEC, a life-threatening inflammatory condition with a mortality rate of approximately 5% [112].
Even in the absence of HAEC, patients with HD exhibit a persistent gut dysbiosis. This is supported by a study, that identified a persistent state of subclinical gut inflammation in HD patients, even though no variation in overall diversity of microorganisms was observed. In this study, significantly higher fecal calprotectin levels that correlated with an abundance of Fusobacteria was found [113]. Neuvonen et al. [11] further documented an enrichment of Escherichia coli and Pseudomonas and a depletion of beneficial Bacteroidales and Ruminococcaceaecompared to healthy controls. In contrast, children with a history of HAEC demonstrate a more severe dysbiosis, characterized by a pronounced expansion of Escherichia and Lactococcus, suggesting an increased risk for inflammatory complications [11]. Furthermore, the extent of aganglionosis directly shapes the gut ecosystem. Patients with total colonic aganglionosis exhibit a marked decrease in alpha diversity and significantly altered beta diversity compared to those with rectosigmoid involvement [114].
Emerging research reframes HAEC from a mere surgical complication to a state of distinct ecological failure of GM. Experimentally, Ward et al. [115] found that colorectal aganglionosis in mice causes early and persistent disruption of microorganisms, characterized by increased Bacteroidetes, decreased Firmicutes, and altered metabolite profiles. Clinically, Yan et al. [83] showed that the HAEC gut is defined by a Proteobacteria-dominated landscape, while Frykman et al. [116] revealed a concurrent disruption of the fungal community, marked by a bloom of Candida. Crucially, these shifts of microorganisms have predictive power; Tang et al. [117] identified a specific microbiota signature at the time of surgery that could predict postoperative HAEC with approximately 85% accuracy. They also identified exclusive breast feeding as a protective factor, potentially via microbiota modulation that reduces pro-inflammatory Gram-negative bacteria, particularly Enterobacteriaceae. More recently, metabolomics was used to identify distinct metabolic profiles in HD, revealing impaired pathways like tyrosine metabolism in HAEC that suggest a breakdown of the intestinal mucosal barrier. This collective evidence firmly establishes HAEC as a complex state of gut ecological failure [118].
Importantly, the consequences of this dysbiotic foundation may extend beyond classic HAEC. Emerging clinical evidence describes a rare but severe late-onset complication: a secondary, Crohn's-like disease of the residual intestine years after HD surgery [119]. This suggests that the persistent, surgically-altered dysbiotic state can predispose patients to chronic inflammation that manifests as a different clinical syndrome later in life. Unfortunately, the authors did not investigate the GM species in these cases, leaving a critical area for upcoming research. Similar refractory, IBD-like inflammation has been observed in paediatric patients with a history of other major intestinal procedures, including surgery for NEC and intestinal atresia.
7.3. Inflammatory Bowel Disease
Research on post-surgical alterations of GM, following procedures for IBD has almost exclusively focused on adults. Indeed, surgical intervention is ultimately required in a significant proportion of IBD cases, with the cumulative risk for surgery estimated at 24% for CD patients and 9% for UC patients [120,121]. While surgery is often necessary to manage complications or treat refractory disease, it induces significant and lasting alterations to the gut ecosystem. In adult population, several studies have demonstrated that procedures such as ileocolonic resection or colectomy lead to persistent decreases or alterations in the diversity of both the GM and the metabolome . For example, Fang et al. [122], compared GM and metabolome alterations after different types of surgery for IBD patients with those who did not undergo surgery. The authors noticed a dysbiotic state characterized by a marked decline in beneficial, anti-inflammatory bacteria, such as Faecalibacteriumprausnitzii, alongside an expansion of potential pathobionts like Escherichia coli,Klebsiella pneumoniae, Enterococcus faecium and Veillonellaatypicawhen compared to control cohort. Metabolically, a key characteristic of the post-surgical condition is a marked increase in primary bile acids, which are pro-inflammatory and create a microenvironment that may contribute to disease recurrence. The specific alterations reported by the authors are supported by several independent studies, reinforcing that such disruptions of microorganisms and metabolomes are a consistent consequence of intestinal surgery [123,124,125,126]. For instance, Mondot et al. [126] examined the GM following ileocolonic resection in patients with CD. The authors documented a shift in the composition of microorgasnisms, specifically a decline in Lachnospiraceae pre-surgically to a post-surgical dominance of Enterococcus durans in half of the patients who experienced endoscopic recurrence. This crucial finding enhances the potential of targeting specific pathobionts to guide future therapeutic strategies.It is critical to note, however, that the vast majority of this evidence comes from adult populations. A significant gap exists in the paediatric literature, with a lack of studies specifically investigating how the developing GM of a child responds to and is reshaped by surgical intervention for IBD.
7.4. Short Bowel Syndrome
SBS is defined as intestinal failure following massive bowel resection, most commonly of the small intestine [127]. Clinical manifestations range from mild malabsorption to complete intestinal failure, depending entirely on the length, portion, and function of the residual intestine, as well as various superimposed factors such as total parenteral nutrition, antibiotic exposure, and variable enteral intake [128,129]. A critical, unresolved question is the directionality of this relationship: do the characteristic shifts of microorganisms in SBS occur as a maladaptive consequence of the altered anatomy, or does a pre-existing dysbiotic GM increase susceptibility to the initial conditions, such as NEC or volvulus, that necessitate the massive resection leading to SBS?
Experimental evidence from a controlled mouse study provides crucial insight [130]. Investigating the specific effects of intestinal resection independent of other diseases, they found that a 50% small bowel resection induced long-term, site-specific alterations in the ileum community of microorganisms, but not in the cecum or stool. The primary shift was an increase in Lactobacillus and a decrease in Enterobacteriaceae in the resected ileum, suggesting that the altered gut anatomy creates a new microenvironment that favors beneficial, fermenting bacteria, potentially as an adaptive mechanism to maximize nutrient absorption. These findings strongly support the first pathway, that the anatomical change itself is a primary driver of restructuring of microorganisms.However, the lack of significant diversity alterations in this healthy mouse model leaves open the question of how a preexisting dysbiosis might interact with surgical stress, highlighting the need for studies on vulnerable hosts. Furtermore, another study demonstrated that children with SBS exhibited a higher abundance of the phylum of Proteobacteria and the class Gammaproteobacteria, alongside an increasedabudance of the genera Escjerichia coli/Shigella and Streptococcus from the class of Gammaproteobacteria and Bacilli respectively, when compared to healthy controls. It is important to mention therelatve abundance of Lactobacillus in this study among children with SBS experiencing diarrhea. [102]. These findings are consistent with other other research, indicating a condition linked to the generation of high ammounts of D-lactose, resulting in notable metabolic acidosis [141].
A recent systematic review by Cleminson et al. [132] further delineated the SBS dysbiosis, revealing a depletion of beneficial SCFA-producing genera from the Lachnospiraceae family such as Dorea, Ruminococcus, Blautia in children dependent on total parenteral nutrition when compared to control subjects. This depletion, likely driven by antibiotics and overgrowth of microorganisms, creates a deficit of protective metabolites, thereby exacerbating mucosal inflammation [132]. This evidence strongly positions anatomical loss as the primary driver of this dysbiosis. However, a more compelling, cyclical pathogenesis is likely: a pre-existing dysbiotic GM may increase susceptibility to initial intestinal insults, and the subsequent massive resection then acts as a powerful amplifier, locking the gut into a self-perpetuating state of pathological dysbiosis, chronic inflammation, and failed adaptation.
In summary, surgery in NEC may result is a decrease in GM diversity. Surgical intervention for HD can impact the GM, potentially leading to alterations in its compositions and icreasing the risk of complications, such HAEC. In children with IBD, surgical procedures can result in alterations to the diversity and structure of GM, which may contribute to disease recurrence or affect the effectiveness of treatment. Children with SBS, after extensive bowel resections, exhibit abnormal GM profiles in comparison to health peers. These GM characteristics may feature reduce diversity, an increase in bacteria linked to inflammation, and a decrease in beneficial bacteria. Examples of various post-surgical GM alterations and outcomes in intestinal paediatric diseases, are illustrated in Table 1.
8. GM-Related Post-surgical Complications of Intestinal Surgery in Paediatric Patients
Post-surgical complications represent a significant challenge in adult visceral surgery and exhibit a similar pattern in paediatric cases, with risk fundamentally linked to the complexity of the procedures involved [133,134,135,136]. The GM is increasingly recognized as a crucial factor in this risk, especially through its influence on inflammation and tissue healing [137]. An individual's vulnerability is not solely influenced by the operating conditions but is instead pre-determined by their underlying GM status, which is shaped by various elements such as diet, physical activity, and existing health conditions [138]. This environment of microorganisms before surgery undergoes drastic alterations throughout the pre-surgical period. Elements such as pre-surgical preparations, surgical injury, and physiological changes work together to disrupt an already fragile ecosystem, leading to a dominance of pathobionts within the community. As a result, this chain of events exacerbates exiting dysbiosis, favoring the survival of resilient pathogens and weaking the host’s immune defense. This relationship is essential, as the most prevalent complications following surgery, such as infections, intestinal obstruction (IO), and anastomotic leak (AL) [139,140], are significantly influenced by the GM across all processes (Figure 1). The underlying mechanisms are still only partially understood and the specific degree of alterations remains largely unknown.
8.1. Infection
Despite advances in surgical techniques, pre-surgical skin antisepsis, and intraoperative antimicrobial prophylaxis, surgical infections, remain a major cause of morbidity and mortality in the post-surgical period [140]. Experimental and adult studies have shown that the GM is implicated as a primary reservoir for causative pathogens through several key-proposed mechanisms. For example, Kreazalek et al. [141] proposed the "Trojan Horse" hypothesis, that refers to how immune cells or other carriers can move bacteria like methicillin resistant Staphylococcus aureus (MRSA) from the gut to a surgical site, leading to infection. Based on this assumption, the authors showed that MRSA labeled with Green Fluorescent Protein originated from the gut of a mouse model, were taken up by neutrophils andsubsequently colonized the surgical wound causing clinical infection by the MRSA. On the other hand, Donaldson et al. [142] revealed that commensal bacteria such as Bacteroides fragilis and Bifidobacterium release metabolites that provide essential tonic stimulation to the immune system through epithelial receptors. Importantly, this immunomodulation does not require bacterial translocation; it is efficiently mediated by the sampling processes of dendritic cells. This mechanism clarifies the significant influence of the length of surgery. A brief surgical trauma allows for swift manifestation of microorganisms, helping to uphold immune defenses. However, an extended operation can lead to drastic reduction in communities of microorganisms within hours, resulting in the overgrowth of harmful microorganisms such as Enterococcus faecalis, Pseudomonas aeruginosa and Clostridium difficile which adapt to survive in challenging environmentalconditions [9,143], thereby continuing to support immune defense. This directly links shorter, meticulous surgeries with excellent outcomes, and longer, complex procedures with increased antibiotic needs and extended hospitalizations. This scenario is supported by a growing body of literature linking the re-establishment of a healthy GM to improved recovery in critically ill patients [144,145]. Recently, the concept of enhanced recovery after surgery is becoming widely used, as enhances the early mobilization, enteral nutrition, and discontinuation of opioid analgesia [9]. The translation of these compelling mechanisms into evidence-based paediatric practice is hindered by a pronounced paucity of literature. However, studies have shown the beneficial results of minimally invasive surgery in paediatric population. For example, in a meta-analysis, comparing outcomes between open and laparoscopic appendectomy in children with complicated acute appendicitis the results demonstrated a clear advantage for the laparoscopic approach, which was associated with a lower overall complication rate, a reduced incidence of infections, and a shorter hospital length of stay [146]. In another meta-analysis that compared the open approach versus laparoscopy in neonates with duodenal atresia, statistically shorter hospital stay, faster enteral feeding and less infection were reported (p=0.004, p<0.001, p=0.02 in the cohort of laparoscopic surgery [147]
8.2. Intestinal Obstruction
IO is a major problem for surgeons because it results in increased hospital stay and morbidity, and readmissions in the hospital [148,149,150]. In the adult literature, the reported incidence varies from 10% to 30%, depending on the patient population and surgical technique [148]. Recently,a large paediatric multicenter study reported a 7% incidence of post-surgical IO following various intestinal reconstructions performed through open and laparoscopic approaches. Notably, the authors found that patients who developed IO had received significantly higher volumes and higher rates of intraoperative fluid administration compared to those who did not (p=0.02 and p=0.03, respectively) [150].
The clinical presentation of IO is influenced by different factors that include GM, metabolites, inhibitory neural reflexes and neurohumoral peptides. In a study of colorectal cancer patients, among causative pathogens that implicated in the pathogenesis of IO reported a decrease of Fermicutes, Bacteroidetes and Fusobacteria and an increase of Proteobacteria and Actinobacteria, when compared with the control group [151]. Furthermore, another study demonstrated a decrease of important commensal bacteria such as Faecalibacteriumprausnitzii , which could serve as a predictor of post-surgical IO [152] Indeed, Faecalibacteriumprausntzii is one of the most abundant butyrate-producing bacterial species that participate in host epithelial homeostasis by modulating the intestinal mucus barrier after modifying goblet cells and glycosylation [153,154]. Furthermore, studies involving animal models have shown that a neuronal inhibition of motility may be activated by monocyte-derived macrophages that produce various cytokines and chemokines such as TNFα, IL6, CCL2. These substances hinder the motility of smooth gut cells, positioning macrophages as central players in the sequence of cellular and molecular processes that may result in IO [155]. Currently, these findings have not been applicable to the treatment of IO in human population.
8.3. Anastomotic Leak
Despite surgical advances in techniques and the introduction of innovative technology, anastomotic leak (AL) is a serious complication that can occur after a gastrointestinal anastomosis,a surgical procedure to restore continuity of the intestinal tract, typically healing within 7-10 days [156,157]. Risk factors for AL differ between adult and paediatric populations. In adults, particularly those with colorectal cancer, major risk factors include male gender, obesity, malnutrition, chronic renal disease, smoking, a history of abdominal surgery, radiation, and neoadjuvant chemotherapy [158]. For paediatric patients, risk factors vary by age and condition. Non-invasive ventilation may be a potential risk factor for neonates undergoing surgery for intestinal disorders [159]. For children undergoing ileal-pouch anal anastomosis for UC, risk factors include corticosteroid use, poor nutritional status, a prior surgical approach, and treatment with infliximab [160].
The etiology of AL is multifactorial, with factors identified in both the pre-surgical and intra-surgical stages. Pre-surgical susceptibilities arise from interventions such as multimodal bowel preparation (antibiotics and mechanical cleansing), neoadjuvant radiotherapy, and the use of non-steroidal anti-inflammatory drugs. Subsequently, intra-surgical challenges- including impaired tissue perfusion, hypoxic conditions, and mechanical injury, directly disrupt the wound healing process at the anastomotic site [161]. Across these diverse conditions, a common pathway leading to AL often involves local GM dysbiosis. In a rodent study, Sjogam et al. [96] found that although stool microbiota remained unchanged after colon surgery, the community of microorganisms adherent to the anastomotic tissue was significantly altered. By post-surgical day 6, they observed a marked increase in opportunistic bacteria and a concurrent decrease in beneficial taxa, indicating an injury-specific dysbiotic response. Furthermore, Okada et al. [162] investigated the impact of GM on the healing of intestinal anastomoses. Using germ-free, conventional, monocontaminated with Lactobacillus acidophilus or Escherichia coli, and ex-germ-free rat models, ileal and colonic anastomoses were created and healing was assessed after 7 days by measuring bursting pressure. Results demonstrated that the presence of a conventional GM significantly enhanced anastomotic strength compared to germ-free or monocontaminated conditions. In addition, Christley et al. [163] examined a rat anastomotic leak to analyze the genetic variations between pre-surgical and post-surgical isolates of Enterococcus faecaliswithin the intestinal wall. They reported increased collagenolytic activity of Enterococcus feacalis at anastomotic sites, potentially influenced by epigenetic factors affecting collagenase production of Enterococcus faecalis. Zamorano et al. [164] in a rat model of AL, discovered that oxygen exposure to intestinal mucus resulting in significantly reduced α-diversity, and a relative abundance of facultative anaerobes, species that better characterize the AL phenotype. Finally, Boatman et al. [165] reported that changing mice from a Western diet to standard diet resulted in a lower rate of AL. These results reinforce the important function of the GM in facilitating the healing of anastomoses, an influence that relies not only on the existence of specific species of microorganisms but is also shaped by a pro-inflammatory environment that they help to establish.
Based on foundational evidence from animal models, recent clinical studies, mainly derived from patients with colorectal cancer, have solidified the role of specific GM in anastomotic leakage (AL) in humans [166,167,168,169,170]. A stable risk profile has emerged, characterized by reduced diversity of microorganisms and genus-specific alterations. For example, examination of human anastomotic tissue has directly linked AL to an increase in the Lachnospiraceae and Bacteroidaceae families [166]. This genus-specificity is further supported by a recent systematic review, that identified bacteria such as Lachnospiraceae and Faecalibacterium as risk factors, while others like Prevotella and Ruminococcus were found to be protective [167]. In addition to genus-level associations, the temporal patterns of particular pathobionts are also highly indicative. In a study of 243 patients with both benign and malignant diseases, the concentration of Escherichia coli and Enterococcus faecalis in the early posto-surgical days was a significant indicator of symptomatic colorectal AL, with Enterococcus faecalis exhibiting a particularly high diagnostic odds ratio of 31.6 on day 3 [168]. In another study, Palmisano et al. [169] compared the GM between healthy individuals and colorectal cancer patients, examining those with and without AL, both before and after neoadjuvant treatment. They discovered that all patients who had surgery showed a moderate increase in Bacteroidetes and Proteobacteria, coupled with a decrease in the beneficial Faecalibacteriumprausnitzii. However, patients who developed AL were distincly characterized by an aboundance of specific inflammatory pathogens, including Acinetobacter lwoffii, Acinetobacter jhonsonii, and Hafnia alvei. All of them are part of the phylunProteobacteria, a microorganism that is associated with hospital-aquired infections. Moreover Mima et al. [170] proposed a possible mechanism, stating that the presence of Bifidobacterium in resected surgical specimens may hinder anastomotic healing, resulting in inadequate vascularization and tissue hypoxia incolorectal tissues. Interestingly, Hajjar et al. [171] found that fecal microbiota transplantation from patients who subsequently experienced AL notably impaired anastomotic healing in mice. This influence was linked to pre-surgical dysbiosis in patients prone to AL, which was characterized by an increase in specific bacteria such as Alistipesonderdonkii and Parabacteroides goldsteini, leading to a pre-existing inflammatory condition and a weakened immune response. Importantly, administering Parabacteroides goldsteini orally was later shown to improve healing, indicating a complex interaction at the species level and highlighting the potential for targeted therapeutics of microorganisms.
Besides these clinical observations, the proposed mechanisms, especially the disruption of the extracellular matrix, which includes collagen, fibronectin, laminin [172,173,174], along to a significant decrease in beneficial metabolites of microorganisms, like butyrate, are thought to form a common pathway that leads to compromised healing in anastomoses across the gastrointestinal tract [175]. This growing body of convincingly evidence shows that GM dysbiosis, through the dual pathways of collagenolytic activity and a lack of protective metabolites such as butyrate, is a contributor to AL. Although this paradigm is increasingly accepted in adult colorectal surgery, its validation and application in the unique context of the developing gut in paediatric patients represent a critical and necessary area for research.
9. Future Research
The study of post-surgical GM alterations in children with intestinal disorders is a rapidly evolving field. Although current research has effectively described these alterations of microorganisms and their correlations with clinical outcomes, future investigations must move beyond past correlations to establish causation and creat effective interventions. Key promising directions for future research consist of:
9.1. Elucidating Causal Mechanisms Through Germ-free Models
A primary limitation of observational studies is the inability to prove causation. Future research should employ germ-free organisms [176]colonized with a completely defined microbiome, to establish direct cause-and-effect relationships. For instance, an investigation was conducted to study if ileocecal resection promotes post-surgical inflammation using interleukin-10 null and wild-type mice [177]. The authors found that germ-free IL-10 knockout mice undergoing ileocecal resection developed only a low incidence of significantly milder fibrosis. This provides direct in vivo evidence that bacteria or their products are essential for producing post-surgical fibrosis.
9.2. The Pre-surgical Microbiota Profile: An Indicator for Surgical Readiness
Future research might focus on the development of a pre-surgical readiness evaluation, integrating a patient's pre-surgical microbiota profile with key clinical metrics to predict complication risk [178]. This could be accomplished by analyzing communities of microorganisms from the gut, oral cavity, and skin, allowing the early identification at risk patients. This enables a shift from reactive treatment to preventive pre-surgical care, using targeted approaches such as particular antibiotics or probiotics to optimize a patient's condition before surgery.
9.2.1. Dietary Prehabilitation and Stools Biomarkers
Pre-surgical nutrition significantly influences surgical outcomes through its impact on the GM. Studies have shown that the consumption of highly processed, low-fiber, and high-fat diets induces dysbiosis, reducing the population of microorganisms responsible for the production of crucial immunomodulatory metabolites, like butyrate [179]. Experimental evidence from Keskey et al. [180] demonstrates this connection: mice on a low-fiber, high-fat diet prior to surgery had a significantly low survival rate (29%), coupled with a dysbiotic GM and reduced butyrate. Importantly, a short 7-day pre-surgical dietary intervention was sufficient to improve survival to 79% and restore butyrate levels, confirming the plasticity of GM. Consequently, future research should focus on applying these findings in clinical practice.
9.2.2. Precision Antimicrobial Therapy
This strategy utilizes narrow-spectrum antibiotics that selectively target known pathogenic bacteria while minimizing collateral damage to the beneficial GM [181]. A recent study demonstrated that lolamicin, selectively targets gram-negative bacteria and spares the GM in mice, thus preventing secondary Clostridioides difficileinfection [182]. Although direct evidence in paediatric surgical groups remains limited, future clinical trials should investigate whether combining rapid pathogen diagnostics with targeted narrow-spectrum therapies can enhance outcomes in surgical paediatric patients by preserving their developing gut ecosystems.
9.2.3. Probiotics, Prebiotics, Synbiotics
A literature search has shown the changing the GM in targeted ways perioperatively through the administration of probiotics and synbiotics before and after surgery, can lead to better surgical results [183,184]. For instance, Huang et al [185] showed that probiotics and synbiotics reduce post-surgical complications such as post-surgical IO or diarrhea.
Kanamori et al. [186] and Uchida et al. [187] demonstrated that the long-term use of probiotics andsynbiotics effectively inhibited pathogenic GM while promoting physical growth in pediatric patients with SBS and various congenital anomalies. More recently, Rao et al. [188] conducted a randomized-controlled trial and found that after 2 weeks of supplementation with a three-strains Bifidobacterium, neonates with various congenital surgical diseases such as HD, SBS, exomphalos, gastroschisis, and congenital diaphragmatic hernia, exhibited a decreased relative abundance of potentially pathogenic bacterial families, an increased abundance of Bifidobacterium, and elevated SCFA levels in their stools. However, these clinical studies are still imitated, and the majority concentrates on preventive strategies against NEC [189]. Therefore, extensive clinical studies are necessary to confirm the safety and efficacy of probiotics in post-surgical complications, such as infection, IO, and AL.
9.2.4. Immunonutrition
Immunonutrition has been shown to be safe and effective in perioperative use as it reduces post-surgical complications and length of hospital stay [190]. Moreover, a recent study highlighted the critical role of breast milk and certain dietary amino acids in managing paediatric intestinal failure and SBS mainly by enhancing intestinal adaptation to reduce dependence on parenteral nutrition [191]. Future studies are required to determine if these targeted nutritional strategies can also help to reduce particular post-surgical complications.
9.3. Precision Microbiota Engineering
Precision microbiota engineering represents a developing therapeutic strategy that utilizes targeted approaches, including engineered probiotics, selective antimicrobials, and phage therapy, to achieve specific therapeutic outcomes [192]. For instance, in the context of IBD, this strategy enables the development of probiotics that can precisely adjust immune responses, correct dysbiosis, enhance intestinal barrier integrity and SCFAs production and reduce the symptoms of IBD [193]. Furthermore, exogenous bacteriophage administration serves as a complementary strategy to selectively target and inhibit pathogenic bacteria while preserving commensal microbiota. [194]. Ultimately, future studies must now focus on translating these sophisticated tools into validated clinical protocols that can predict and proactively address post-surgical complications in at risk surgical patients.
10. Conclusions
The GM plays a crucial role in post-surgical outcomes, and its disturbance triggers harmful consequences that can result in serious complications like surgical infections, IO, and ALs. This occurs because of the failure of protective functions and growth of microorganisms that hinder the healing process. Although the pathophysiological role of the GM in surgical outcomes is increasingly defined in adults, dedicated studies in children are exceptionally rare. In paediatric surgery, this point of view emphasizes the need for a prompt research schedule to identify age-specific dysbiosis, create diagnostics based on the GM, and develop therapies targeting the GM, with the ultimate goal to improve surgical outcomes for children with intestinal diseases.
Author Contributions
Conceptualization, N.V.,N.Z and G.C.V.; methodology, N.V.; N.Z.and G.C.V.; formal analysis, N.V., N.Z. andC.S.; investigation, N.V., N.Z.; resources, N.V.,N.Z and G.C.V.; data curation, N.V., N.Z .andC.S; writing—original draft preparation, N.V., and N.Z.,writing—review and editing, C.V., G.V., A.T. and G.C.V..; supervision, G.C.V.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| GM | Gut microbiota |
| NEC | Necrotizing enterocolitis |
| HD | Hirschsprung’s disease |
| IBD | Inflammatory bowel disease |
| SBS | Short bowel syndrome |
| HAEC | Hirschsprung’s-associated-enterocolitis |
| 16S rRNA | 16S ribosomal RNA |
| SCFAs | Short-chains fatty acids |
| CKD | Chronic kidney disease |
| CD | Chron’s disease |
| UC | Ulcerative colitis |
| IO | Intestinal obstruction |
| AL | Anastomotic leak |
| MRSA | Methicillin-resistant Staphylococcus aureous |
References
- Takisha, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef]
- Bankole, T.; Li, Y. The early-life gut microbiome in common pediatric diseases: roles and therapeutic implications. Front Nutr. 2025, 12, 1597206. [Google Scholar] [CrossRef] [PubMed]
- Kamanga, N.H.B.; Thomas, R.; Thandrayen, K.; Velaphi, S.C. Risk factors and outcomes of infants with necrotizing enterocolitis: a case-control study. Front Pediatr. 2025, 13, 1611111. [Google Scholar] [CrossRef]
- Pini Prato, A.; Rossi, V.; Avanzini, S.; Mattioli, G.; Disma, N.; Jasonni, V. Hirschsprung's disease: what about mortality? Pediatr Surg Int. 2011, 27, 473–478. [Google Scholar] [CrossRef]
- Kolho, KL.; Nikkonen, A.; Merras-Salmio, L.; Molander, P. The need for surgery in pediatric patients with inflammatory bowel disease treated with biologicals. Int J Colorectal Dis. 2024, 39, 58. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Z.; O'Daniel, E.L. Updates in intestinal failure management. J Clin Med. 2025, 14, 3031. [Google Scholar] [CrossRef]
- Decembrino, N.; Scuderi, M.G.; Betta, P.M.; Leonardi, R.; Bartolone, A.; Marsiglia, R.; Marangelo, C.; Pane, S.; De Rose, D.U.; Salvatori, G.; Grosso, G.; Di Domenico, F.M.; Dotta, A.; Putignani, L.; Capolupo, I.; Di Benedetto, V. Microbiota-Modulating Strategies in Neonates Undergoing Surgery for Congenital Gastrointestinal Conditions: A Narrative Review. Nutrients 2025, 17, 2234. [Google Scholar] [CrossRef] [PubMed]
- Gibiino, G.; Binda, C.; Cristofaro, L.; Sbrancia, M.; Coluccio, C.; Petraroli, C.; Jung, C.F.M.; Cucchetti, A.; Cavaliere, D.; Ercolani, G.; Sambri, V.; Fabbri, C. Dysbiosis and Gastrointestinal Surgery: Current Insights and Future Research. Biomedicines 2022, 10, 2532. [Google Scholar] [CrossRef]
- Shi, Y.; Cui, H.; Wang, F.; Zhang, Y.; Xu, Q.; Liu, D.; Wang, K.; Hou, S. Role of gut microbiota in postoperative complications and prognosis of gastrointestinal surgery: A narrative review. Medicine (Baltimore) 2022, 101, e29826. [Google Scholar] [CrossRef]
- Lin, H.; Xu, C.; Chen, J.; Ma, X.; Shi, L.; Shi, W.; Du, L.; Ni, Y. Alteration of the gut microbiota after surgery in preterm infants with necrotizing enterocolitis. Front Pediatr. 2023, 11, 993759. [Google Scholar] [CrossRef]
- Neuvonen, MI.; Korpela, K.; Kyrklund, K.; Salonen, A.; de Vos, W.; Rintala, R.J.; Pakarinen, M.P. Intestinal Microbiota in Hirschsprung Disease. J Pediatr Gastroenterol Nutr. 2018, 67, 594–600. [Google Scholar] [CrossRef]
- Thänert, R.; Thänert, A.; Ou, J.; Bajinting, A.; Burnham, C.D.; Engelstad, H.J.; Tecos, M.E.; Ndao, I.M.; Hall-Moore, C.; Rouggly-Nickless, C.; Carl, M.A.; Rubin, D.C.; Davidson, N.O.; Tarr, P.I.; Warner, B.B.; Dantas, G.; Warner, B.W. Antibiotic-driven intestinal dysbiosis in pediatric short bowel syndrome is associated with persistently altered microbiome functions and gut-derived bloodstream infections. Gut Microbes 2021, 13, 1940792. [Google Scholar] [CrossRef]
- Cleminson, J.S.; Thomas, J.; Stewart, C.J.; Campbell, D.; Gennery, A.; Embleton, N.D.; Köglmeier, J.; Wong, T.; Spruce, M.; Berrington, J.E. Gut microbiota and intestinal rehabilitation: a prospective childhood cohort longitudinal study of short bowel syndrome (the MIRACLS study): study protocol. BMJ Open Gastroenterol. 2024, 11, e001450. [Google Scholar] [CrossRef]
- Packey, C.D.; Sartor, R.B. Commensal bacteria, traditional and opportunistic pathogens, dysbiosis and bacterial killing in inflammatory bowel diseases. CurrOpin Infect Dis. 2009, 22, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Mimura, T.; Rizzello, F.; Helwig, U.; Poggioli, G.; Schreiber, S.; Talbot, I.C.; Nicholls, R.J.; Gionchetti, P.; Campieri, M.; Kamm, M.A. Once daily high dose probiotic therapy (VSL#3) for maintaining remission in recurrent or refractory pouchitis. Gut. 2004, 53, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Z.; Xu, Z.; Wang, Z.; Feng, J. Probiotics prevent Hirschsprung's disease-associated enterocolitis: a prospective multicenter randomized controlled trial. Int J Colorectal Dis. 2015, 30, 105–110. [Google Scholar] [CrossRef]
- D'Haens, G.R.; Vermeire, S.; Van Assche, G.; Noman, M.; Aerden, I.; Van Olmen, G.; Van Olmen, G.; Rutgeerts, P. Therapy of metronidazole with azathioprine to prevent postoperative recurrence of Crohn's disease: a controlled randomized trial. Gastroenterology 2008, 135, 1123–1129. [Google Scholar] [CrossRef]
- Premkumar, M.H.; Soraisham, A.; Bagga, N.; Massieu, L.A.; Maheshwari, A. Nutritional Management of Short Bowel Syndrome. Clin Perinatol. 2022, 49, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Nishida, A.; Inoue, R.; Inatomi, O; Bamba, S.; Naito, Y.; Andoh, A. Gut microbiota in the pathogenesis of inflammatory bowel disease. Clin J Gastroenterol. 2018, 11, 1–10. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464(7285), 59–65. [Google Scholar] [CrossRef]
- Ma, Z.F.; Lee, Y.Y. The Role of the Gut Microbiota in Health, Diet, and Disease with a Focus on Obesity. Foods 2025, 14, 492. [Google Scholar] [CrossRef] [PubMed]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- Bazinet, AL; Cummings, MP. A comparative evaluation of sequence classification programs. BMC Bioinformatics 2012, 13, 92. [Google Scholar] [CrossRef]
- Hugon, P; Dufour, JC; Colson, P; Fournier, PE; Sallah, K; Raoult, D. A comprehensive repertoire of prokaryotic species identified in human beings. Lancet Infect Dis. 2015, 15, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef]
- Arumugam, M; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473(7346), 174–180. [Google Scholar] [CrossRef]
- Laterza, L.; Rizzatti, G.; Gaetani, E.; Chiusolo, P.; Gasbarrini, A. The Gut Microbiota and Immune System Relationship in Human Graft-versus-Host Disease. Mediterr J Hematol Infect Dis. 2016, 8, e2016025. [Google Scholar] [CrossRef]
- Mariat, D; Firmesse, O; Levenez, F; Guimarăes, V; Sokol, H; Doré, J; Corthier, G; Furet, JP. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009, 9, 123. [Google Scholar] [CrossRef]
- Hooper, L.V.; Macpherson, A.J. Immune adaptations that maintain homeostasis with the intestinal microbiota. Nat Rev Immunol. 2010, 10, 59–169. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J; Thiele, I; Tuohy, K. Gut microbiota functions: metabolism of nutrients and other food components. Eur J Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Krautkramer, K.A.; Fan, J.; Bäckhed, F. Gut microbial metabolites as multi-kingdom intermediates. Nat Rev Microbiol. 2021, 19, 77–94. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut. 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
- Pascale, A; Marchesi, N; Marelli, C; Coppola, A; Luzi, L; Govoni, S; Giustina, A; Gazzaruso, C. Microbiota and metabolic diseases. Endocrine 2018, 61, 357–371. [Google Scholar] [CrossRef]
- Wang, L.Y.; He, L.H.; Xu, L.J.; Li, S.B. Short-chain fatty acids: bridges between diet, gut microbiota, and health. J Gastroenterol Hepatol. 2024, 39, 1728–1736. [Google Scholar] [CrossRef]
- Mansuy-Aubert, V.; Ravussin, Y. Short chain fatty acids: the messengers from down below. Front Neurosci. 2023, 17, 1197759. [Google Scholar] [CrossRef]
- Liu, P.; Wang, Y.; Yang, G.; Zhang, Q.; Meng, L.; Xin, Y.; Jiang, X. The role of short-chain fatty acids in intestinal barrier function, inflammation, oxidative stress, and colonic carcinogenesis. Pharmacol Res. 2021, 165, 105420. [Google Scholar] [CrossRef]
- MogoşGFR; ManciuleaProfir, M; Enache, RM; Pavelescu, LA; Popescu Roşu, OA; Cretoiu, SM; Marinescu, I. Intestinal Microbiota in Early Life: Latest Findings Regarding the Role of Probiotics as a Treatment Approach for Dysbiosis. Nutrients 2025, 17, 2071. [Google Scholar] [CrossRef]
- De Palma, G.; Collins, S.M.; Bercik, P. The microbiota-gut-brain axis in functional gastrointestinal disorders. Gut Microbes 2014, 5, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Enaud, R.; Prevel, R.; Ciarlo, E.; Beaufils, F.; Wieërs, G.; Guery, B.; Delhaes, L. The Gut-Lung Axis in Health and Respiratory Diseases: A Place for Inter-Organ and Inter-Kingdom Crosstalks. Front Cell Infect Microbiol. 2020, 10, 9. [Google Scholar] [CrossRef]
- Paul, J.K.; Azmal, M.; Haque, A.S.N.B.; Meem, M.; Talukder, O.F.; Ghosh, A. Unlocking the secrets of the human gut microbiota: Comprehensive review on its role in different diseases. World J Gastroenterol. 2025, 31, 99913. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.W.; Chen, M.J. Exploring the Preventive and Therapeutic Mechanisms of Probiotics in Chronic Kidney Disease through the Gut-Kidney Axis. J Agric Food Chem. 2024, 72, 8347–8364. [Google Scholar] [CrossRef] [PubMed]
- Magliocca, G.; Mone, P.; Di Iorio, B.R.; Heidland, A.; Marzocco, S. Short-Chain Fatty Acids in Chronic Kidney Disease: Focus on Inflammation and Oxidative Stress Regulation. Int J Mol Sci. 2022, 23, 5354. [Google Scholar] [CrossRef]
- Zhao, J.; Ning, X.; Liu, B.; Dong, R.; Bai, M.; Sun, S. Specific alterations in gut microbiota in patients with chronic kidney disease: an updated systematic review. Ren Fail. 2021, 43, 102–112. [Google Scholar] [CrossRef]
- Indrio, F; Martini, S; Francavilla, R; Corvaglia, L; Cristofori, F; Mastrolia, SA; Neu, J; Rautava, S; Russo Spena, G; Raimondi, F; Loverro, G. Epigenetic Matters: The Link between Early Nutrition, Microbiome, and Long-term Health Development. Front Pediatr. 2017, 5, 178. [Google Scholar] [CrossRef]
- Bavineni, M.; Wassenaar, T.M.; Agnihotri, K.; Ussery, D.W.; Lüscher, T.F.; Mehta, JL. Mechanisms linking preterm birth to onset of cardiovascular disease later in adulthood. Eur Heart J. 2019, 40, 1107–1112. [Google Scholar] [CrossRef]
- Barker-Tejeda, T.C.; Zubeldia-Varela, E.; Macías-Camero, A.; Alonso, L.; Martín-Antoniano, I.A.; Rey-Stolle, M.F.; Mera-Berriatua, L.; Bazire, R.; Cabrera-Freitag, P; Shanmuganathan, M.; Britz-McKibbin, P.; et al. Comparative characterization of the infant gut microbiome and their maternal lineage by a multi-omics approach. Nat Commun. 2024, 15, 3004. [Google Scholar] [CrossRef]
- Yang, R.; Shi, Z.; Li, Y.; Huang, X.; Li, Y.; Li, X.; Chen, Q.; Hu, Y.; Li, X. Research focus and emerging trends of the gut microbiome and infant: a bibliometric analysis from 2004 to 2024. Front Microbiol. 2024, 15, 1459867. [Google Scholar] [CrossRef]
- Yang, R; Wang, Y; Ying, Z; Shi, Z; Song, Y; Yan, J; Hou, S; Zhao, Z; Hu, Y; Chen, Q; Peng, W; Li, X. Inspecting mother-to-infant microbiota transmission: disturbance of strain inheritance by cesarian section. FrontMicrobiol 2024, 15, 1292377. [Google Scholar] [CrossRef] [PubMed]
- Schoenmakers, S.; Steegers-Theunissen, R.; Faas, M. The matter of the reproductive microbiome. Obstet Med. 2019, 12, 107–115. [Google Scholar] [CrossRef]
- Agaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The placenta harbors a unique microbiome. Sci Transl Med. 2014, 6(237), 237ra65. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, E.; Fernández, L.; Marín, M.L.; Martín, R.; Odriozola, J.M.; Nueno-Palop, C.; Narbad, A.; Olivares, M.; Xaus, J.; Rodríguez, J.M. Isolation of commensal bacteria from umbilical cord blood of healthy neonates born by cesarean section. CurrMicrobiol 2005, 51, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Stinson, L.F.; Payne, M.S.; Keelan, J.A. Planting the seed: Origins, composition, and postnatal health significance of the fetal gastrointestinal microbiota. Crit Rev Microbiol. 2017, 43, 352–369. [Google Scholar] [CrossRef]
- Zheng, J.; Xiao, X.; Zhang, Q.; Mao, L.; Yu, M.; Xu, J. The Placental Microbiome Varies in Association with Low Birth Weight in Full-Term Neonates. Nutrients 2015, 7, 6924–6937. [Google Scholar] [CrossRef]
- Derrien, M.; Alvarez, A.S.; de Vos, W.M. The Gut Microbiota in the First Decade of Life. Trends Microbiol. 2019, 27, 997–1010. [Google Scholar] [CrossRef]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc Natl Acad Sci U S A 2010, 107, 11971–11975. [Google Scholar] [CrossRef] [PubMed]
- Carlisle, E.M.; Morowitz, M.J. Pediatric surgery and the human microbiome. J Pediatr Surg. 2011, 46, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Konya, T.; Maughan, H.; Guttman, D.S.; Field, C.J.; Chari, R.S.; Sears, M.R.; Becker, A.B.; Scott, J.A.; Kozyrskyj, A.L. CHILD Study Investigators. Gut microbiota of healthy Canadian infants: profiles by mode of delivery and infant diet at 4 months. CMAJ 2013, 185, 385–394. [Google Scholar] [CrossRef]
- Penders, J.; Thijs, C.; Vink, C.; Stelma, F.F.; Snijders, B.; Kummeling, I.; van den Brandt, P.A.; Stobberingh, EE. Factors influencing the composition of the intestinal microbiota in early infancy. Pediatrics 2006, 118, 511–521. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe. 2015, 17, 690–703, Erratum in: Cell Host Microbe. 2015,17, 852. Jun, Wang [corrected to Wang, Jun]. Erratum in: Cell Host Microbe. 2015,17,852. [Google Scholar] [CrossRef]
- Rozé, J.C.; Ancel, P.Y.; Marchand-Martin, L.; Rousseau, C.; Montassier, E.; Monot, C.; Le Roux, K.; Butin, M.; Resche-Rigon, M.; Aires, J.; et al. Assessment of Neonatal Intensive Care Unit Practices and Preterm Newborn Gut Microbiota and 2-Year Neurodevelopmental Outcomes. JAMA Netw Open. 2020, 3, e2018119. [Google Scholar] [CrossRef]
- Jia, J.; Xun, P.; Wang, X.; He, K.; Tang, Q.; Zhang, T.; Wang, Y.; Tang, W.; Lu, L.; Yan, W.; Wang, W.; Hu, T; Cai, W. Impact of Postnatal Antibiotics and Parenteral Nutrition on the Gut Microbiota in Preterm Infants During Early Life. JPEN J Parenter Enteral Nutr. 2020, 44, 639–654. [Google Scholar] [CrossRef]
- Li, F.; Hooi, S.L.; Choo, Y.M.; The, C.S.J.; Toh, K.Y.; Lim, L.W.Z.; Lee, Y.Q.; Chong, C.W.; Ahmad Kamar, A. Progression of gut microbiome in preterm infants during the first three months. Sci Rep. 2025, 15, 12104. [Google Scholar] [CrossRef]
- Stewart, C.J.; Ajami, N.J.; O'Brien, J.L.; Hutchinson, D.S.; Smith, D.P.; Wong, M.C.; Ross, M.C.; Lloyd, R.; Doddapaneni, H.; Metcalf, G.A.; et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature 2018, 562(7728), 583–588. [Google Scholar] [CrossRef]
- Yatsunenko, T; Rey, FE; Manary, MJ; Trehan, I; Dominguez-Bello, MG; Contreras, M; Magris, M; Hidalgo, G; Baldassano, RN; Anokhin, AP; Heath, AC; Warner, B; Reeder, J; Kuczynski, J; Caporaso, JG; Lozupone, CA; Lauber, C; Clemente, JC; Knights, D; Knight, R; Gordon, JI. Human gut microbiome viewed across age and geography. Nature. 2012, 486(7402), 222–227. [Google Scholar] [CrossRef]
- Hollister, E.B.; Riehle, K.; Luna, R.A.; Weidler, E.M.; Rubio-Gonzales, M.; Mistretta, T.A.; Raza, S.; Doddapaneni, H.V.; Metcalf, G.A.; Muzny, D.M.; et al. Structure and function of the healthy pre-adolescent pediatric gut microbiome. Microbiome 2015, 3, 36. [Google Scholar] [CrossRef]
- Guyton, K; Alverdy, J.C. The gut microbiota and gastrointestinal surgery. Nat Rev Gastroenterol Hepatol. 2017, 14, 43–54. [Google Scholar] [CrossRef]
- Morowitz, MJ; Babrowski, T; Carlisle, EM; Olivas, A; Romanowski, KS; Seal, JB; Liu, DC; Alverdy, JC. The human microbiome and surgical disease. Ann Surg. 2011, 253, 1094–1101. [Google Scholar] [CrossRef]
- Liu, L.; Fu, Q.; Li, T.; Shao, K.; Zhu, X.; Cong, Y.; Zhao, X. Gut microbiota and butyrate contribute to nonalcoholic fatty liver disease in premenopause due to estrogen deficiency. PLoS One 2022, 17, e0262855. [Google Scholar] [CrossRef] [PubMed]
- Alsharairi, N.A. Therapeutic Potential of Gut Microbiota and Its Metabolite Short-Chain Fatty Acids in Neonatal Necrotizing Enterocolitis. Life (Basel) 2023, 13, 561. [Google Scholar] [CrossRef] [PubMed]
- Weaver, L.; Troester, A.; Jahansouz, C. The Impact of Surgical Bowel Preparation on the Microbiome in Colon and Rectal Surgery. Antibiotics (Basel) 2024, 23(13), 580. [Google Scholar] [CrossRef] [PubMed]
- Nalluri, H; Kizy, S; Ewing, K; Luthra, G; Leslie, DB; Bernlohr, DA; Sadowsky, MJ; Ikramuddin, S; Khoruts, A; Staley, C; Jahansouz, C. Peri-operative antibiotics acutely and significantly impact intestinal microbiota following bariatric surgery. Sci Rep. 2020, 10, 20340. [Google Scholar] [CrossRef]
- Zhang, R.; Li, L.; Yu, G.; Li, Y.; Wei, K.; Lin, L.; Ye, Y. Interaction between gut microbiota and anesthesia: mechanism exploration and translation challenges focusing on the gut-brain-liver axis. Front Cell Infect Microbiol. 2025, 15, 1626585. [Google Scholar] [CrossRef] [PubMed]
- Brenner, D.; Shorten, G.D.; O'Mahony, S.M. Postoperative pain and the gut microbiome. Neurobiol Pain 2021, 10, 100070. [Google Scholar] [CrossRef]
- Ilhan, Z.E.; DiBaise, J.; Dautel, S.E.; Isern, N.G.; Kim, Y.M.; Hoyt, D.W.; Schepmoes, A.A.; Brewer, H.M.; Weitz, K.K.; et al. Temporospatial shifts in the human gut microbiome and metabolome after gastric bypass surgery. NPJ Biofilms Microbiomes 2020, 6, 12. [Google Scholar] [CrossRef] [PubMed]
- Paine, H.; Jones; Kinross, J. Preparing the Bowel (Microbiome) for Surgery: Surgical Bioresilience. Clin Colon Rectal Surg. 2023, 36, 138–145. [Google Scholar] [CrossRef]
- Saeed, NK.; Al-Beltagi, M.; Bediwy, A.S.; El-Sawaf, Y.; Toema, O. Gut microbiota in various childhood disorders: Implication and indications. World J Gastroenterol. 2022, 28, 1875–1901. [Google Scholar] [CrossRef]
- Ihekweazu, F.D.; Versalovic, J. Development of the Pediatric Gut Microbiome: Impact on Health and Disease. Am J Med Sci. 2018, 356, 413–423. [Google Scholar] [CrossRef]
- Wu, Y.; LI, S.; Liu, L.; Zhang, X.; Lv, J.; Li, Q.; L, Y. Intestinal microecology in pediatric surgery-related diseases: Current insights and future perspectives. J Pediatr Surg Open. 2024, 6, 100134. [Google Scholar] [CrossRef]
- Rao, S.C.; Esvaran, M.; Patole, S.K.; Simmer, K.N.; Gollow, I.; Keil, A.; Wemheuer, B.; Chen, L.; Conway, P.L. Gut microbiota in neonates with congenital gastrointestinal surgical conditions: a prospective study. Pediatr Res. 2020, 88, 878–886. [Google Scholar] [CrossRef]
- Zhu, L.; Chen, F.; Gao, T.; Liu, T.; Zhu, H.; Sheng, Q.; Liu, J.; Lu, L.; Lv, Z. Bacteroidetes/Firmicutes ratio as an early biomarker for NEC development in preterm infants. J PediatrSurg 2025, 60, 162606. [Google Scholar] [CrossRef]
- Salvi, P.S.; Cowles, R.A. Butyrate and the Intestinal Epithelium: Modulation of Proliferation and Inflammation in Homeostasis and Disease. Cells 2021, 10, 1775. [Google Scholar] [CrossRef]
- Huang, K.; Mukherjee, S.; DesMarais, V.; Albanese, J.M.; Rafti, E.; Draghi, Ii. A.; Maher, L.A.; Khanna, K.M.; Mani, S.; Matson, A.P. Targeting the PXR-TLR4 signaling pathway to reduce intestinal inflammation in an experimental model of necrotizing enterocolitis. Pediatr Res. 2018, 83, 1031–1040. [Google Scholar] [CrossRef]
- Yan, Z.; Poroyko, V.; Gu, S.; Zhang, Z.; Pan, L.; Wang, J.; Bao, N.; Hong, L. Characterization of the intestinal microbiome of Hirschsprung's disease with and without enterocolitis. BiochemBiophys Res Commun. 2014, 445, 269–274. [Google Scholar] [CrossRef]
- Budianto, I.R.; Kusmardi, K.; Maulana, A.M.; Arumugam, S.; Afrin, R.; Soetikno, V. Paneth-like cells disruption and intestinal dysbiosis in the development of enterocolitis in an iatrogenic rectosigmoid hypoganglionosis rat model. Front Surg. 2024, 11, 1407948. [Google Scholar] [CrossRef] [PubMed]
- Arnaud, A.P.; Hascoet, J.; Berneau, P.; LeGouevec, F.; Georges, J.; Randuineau, G.; Formal, M.; Henno, S.; Boudry, G. A piglet model of iatrogenic rectosigmoid hypoganglionosis reveals the impact of the enteric nervous system on gut barrier function and microbiota postnatal development. J Pediatr Surg. 2021, 56, 337–345. [Google Scholar] [CrossRef]
- Liu, W.; Yan, H.; Jia, W.; Huang, J.; Fu, Z.; Xu, W.; Yu, H.; Yang, W.; Pan, W.; Zheng, B.; et al. Association between gut microbiota and Hirschsprung disease: a bidirectional two-sample Mendelian randomization study. Front Microbiol. 2024, 15, 1366181. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gao, B.; Liu, X.; Li, A. The mediating role of metabolites between gut microbiome and Hirschsprung disease: a bidirectional two-step Mendelian randomization study. Front Pediatr. 2024, 12, 1371933. [Google Scholar] [CrossRef]
- Zhuang, X; Liu, C; Zhan, S; Tian, Z; Li, N; Mao, R; Zeng, Z; Chen, M. Gut Microbiota Profile in Pediatric Patients with Inflammatory Bowel Disease: A Systematic Review. Front Pediatr. 2021, 9, 626232. [Google Scholar] [CrossRef]
- Sondheimer, JM; Asturias, E; Cadnapaphornchai, M. Infection and cholestasis in neonates with intestinal resection and long-term parenteral nutrition. J Pediatr Gastroenterol Nutr. 1998, 27, 131–197. [Google Scholar] [PubMed]
- Meehan, JJ; GeorgesonKE. Prevention of liver failure in parenteral nutrition-dependent children with short bowel syndrome. J Pediatr Surg. 1997, 32, 473–475. [Google Scholar] [CrossRef]
- Engstrand, Lilja. H; Wefer, H.; Nyström, N.; Finkel, Y.; Engstrand, L. Intestinal dysbiosis in children with short bowel syndrome is associated with impaired outcome. Microbiome 2015, 3, 18. [Google Scholar] [CrossRef]
- Piper, H.G.; Fan, D.; Coughlin, L.A.; Ho, E.X.; McDaniel, M.M.; Channabasappa, N.; Kim, J.; Kim, M.; Zhan, X.; Xie, Y.; Koh, A.Y. Severe Gut Microbiota Dysbiosis Is Associated With Poor Growth in Patients With Short Bowel Syndrome. JPEN J Parenter Enteral Nutr. 2017, 41, 1202–1212. [Google Scholar] [CrossRef]
- Davidovics, Z.H.; Carter, B.A.; Luna, R.A.; Hollister, E.B.; Shulman, R.J.; Versalovic, J. The Fecal Microbiome in Pediatric Patients With Short Bowel Syndrome. JPEN J Parenter EnteralNutr 2016, 40, 1106–1113. [Google Scholar] [CrossRef]
- Zeichner, S.; Mongodin, E.F.; Hittle, L.; Huang, S.H.; Torres, C. The bacterial communities of the small intestine and stool in children with short bowel syndrome. PLoS One. 2019, 14, e0215351. [Google Scholar] [CrossRef] [PubMed]
- Phyo, L.Y.; Singkhamanan, K; Laochareonsuk, W.; Surachat, K.; Phutong, N.; Boonsanit, K.; Chiengkriwate, P.; Sangkhathat, S. Fecal microbiome alterations in pediatric patients with short bowel syndrome receiving a rotating cycle of gastrointestinal prophylactic antibiotics. Pediatr Surg Int. 2021, 37, 1371–1381. [Google Scholar] [CrossRef] [PubMed]
- Shogan, BD; Smith, DP; Christley, S; Gilbert, JA; Zaborina, O; Alverdy, JC. Intestinal anastomotic injury alters spatially defined microbiome composition and function. Microbiome 2014, 2, 35. [Google Scholar] [CrossRef]
- Salomon, J; Ericsson, A; Price, A; Manithody, C; Murry, DJ; Chhonker, YS; Buchanan, P; Lindsey, ML; Singh, AB; Jain, AK. Dysbiosis and Intestinal Barrier Dysfunction in Pediatric Congenital Heart Disease Is Exacerbated Following Cardiopulmonary Bypass. JACC Basic Transl Sci. 2021, 6, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Derikx, JP; Luyer, MD; Heineman, E; Buurman, WA. Non-invasive markers of gut wall integrity in health and disease. World J Gastroenterol. 2010, 16, 5272–5279. [Google Scholar] [CrossRef]
- Lu, Z; Ding, L; Lu, Q; Chen, YH. Claudins in intestines: Distribution and functional significance in health and diseases. Tissue Barriers 2013, 1, e24978. [Google Scholar] [CrossRef]
- Rhoads, JM; Plunkett, E; Galanko, J; Lichtman, S; Taylor, L; Maynor, A; Weiner, T; Freeman, K; GuariscoJL; Wu, GY. Serum citrulline levels correlate with enteral tolerance and bowel length in infants with short bowel syndrome. J Pediatr. 2005, 146, 542–547. [Google Scholar] [CrossRef]
- Yang, X.; Wu, Y.; Xu, X.; Gao; Xie, J.; Li, Z.; Zhou, X.; Feng, X. Impact of Repeated Infantile Exposure to Surgery and Anesthesia on Gut Microbiota and Anxiety Behaviors at Age 6-9. J Pers Med. 2023, 13, 823. [Google Scholar] [CrossRef] [PubMed]
- Traini, I.; Chan, S.Y.; Menzies, J.; Hughes, J.; Coffey, M.J.; McKay, I.R.; Ooi, C.Y.; Leach, S.T.; Krishnan, U. Intestinal dysbiosis and inflammation in children with repaired esophageal atresia. J Pediatr Gastroenterol Nutr. 2023, 78, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Doğan, G.; İpek, H. The Development of Necrotizing Enterocolitis Publications: A Holistic Evolution of Global Literature with Bibliometric Analysis. Eur J Pediatr Surg. 2020, 30, 293–303. [Google Scholar] [CrossRef]
- Claud, E.C.; Walker, W.A. Hypothesis: inappropriate colonization of the premature intestine can cause neonatal necrotizing enterocolitis. FASEB J. 2001, 15, 1398–1403. [Google Scholar] [CrossRef] [PubMed]
- Coggins, S.A.; Wynn, J.L.; Weitkamp, J.H. Infectious causes of necrotizing enterocolitis. Clin Perinatol. 2015, 42, 133–154. [Google Scholar] [CrossRef]
- Boccia, D.; Stolfi, I.; Lana, S.; Moro, M.L. Nosocomial necrotizing enterocolitis outbreaks: epidemiology and control measures. Eur J Pediatr. 2001, 160, 385–391. [Google Scholar] [CrossRef]
- Morrow, A.L.; Lagomarcino, A.J.; Schibler, K.R.; Taft, D.H.; Yu, Z.; Wang, B.; Altaye, M.; Wagner, M.; Gevers, D.; Ward, D.V.; et al. Early microbial and metabolomic signatures predict later onset of necrotizing enterocolitis in preterm infants. Microbiome 2013, 1, 13. [Google Scholar] [CrossRef]
- Wang, Y.; Hoenig, J.D.; Malin, K.J.; Qamar, S.; Petrof, E.O.; Sun, J.; Antonopoulos, D.A.; Chang, E.B.; Claud, E.C. 16S rRNA gene-based analysis of fecal microbiota from preterm infants with and without necrotizing enterocolitis. ISME J. 2009, 3, 944–954. [Google Scholar] [CrossRef]
- Cheng, C.W.; Biton, M.; Haber, A.L.; Gunduz, N.; Eng, G.; Gaynor, L.T.; Tripathi, S.; Calibasi-Kocal, G.; Rickelt, S.; Butty, V.L.; et al. Ketone Body Signaling Mediates Intestinal Stem Cell Homeostasis and Adaptation to Diet. Cell. 2019, 178, 1115–1131. [Google Scholar] [CrossRef]
- Carr, T.M.; Wheaton, J.D.; Houtz, G.M.; Ciofani, M. JunB promotes Th17 cell identity and restrains alternative CD4+ T-cell programs during inflammation. Nat Commun. 2017, 8, 301. [Google Scholar] [CrossRef]
- Obermayr, F.; Hotta, R.; Enomoto, H.; Young, H.M. Development and developmental disorders of the enteric nervous system. Nat Rev Gastroenterol Hepatol. 2013, 10, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Gosain, A. Established and emerging concepts in Hirschsprung's-associated enterocolitis. Pediatr Surg Int. 2016, 32, 313–320. [Google Scholar] [CrossRef]
- Murphy, C.E.; Coffey, M.J.; Chen, Q.; Adams, S.; Ooi, C.Y.; van Dorst, J. Altered Development of Gut Microbiota and Gastrointestinal Inflammation in Children with Post-Operative Hirschsprung's Disease. Int J Mol Sci. 2025, 26, 10570. [Google Scholar] [CrossRef]
- Pini Prato, A.; Bartow-McKenney, C.; Hudspeth, K.; Mosconi, M.; Rossi, V.; Avanzini, S.; et al. A Metagenomics Study on Hirschsprung's Disease Associated Enterocolitis: Biodiversity and Gut Microbial Homeostasis Depend on Resection Length and Patient's Clinical History. Front Pediatr. 2019, 7, 326. [Google Scholar] [CrossRef]
- Ward, N.L.; Pieretti, A.; Dowd, S.E.; Cox, S.B.; Goldstein, A.M. Intestinal aganglionosis is associated with early and sustained disruption of the colonic microbiome. NeurogastroenterolMotil 2012, 24, 874–e400. [Google Scholar] [CrossRef]
- Frykman, P.K.; Nordenskjöld, A.; Kawaguchi, A.; Hui, T.T.; Granström, A.L.; Cheng, Z.; Tang, J.; Underhill, D.M.; Iliev, I.; Funari, V.A.; et al. HAEC Collaborative Research Group (HCRG). Characterization of Bacterial and Fungal Microbiome in Children with Hirschsprung Disease with and without a History of Enterocolitis: A Multicenter Study. PLoS One 2015, 10, e0124172. [Google Scholar] [CrossRef]
- Tang, W; Su, Y; Yuan, C; Zhang, Y; Zhou, L; Peng, L; Wang, P; Chen, G; Li, Y; Li, H; Zhi, Z; Chang, H; Hang, B; Mao, JH; Snijders, AM; Xia, Y. Prospective study reveals a microbiome signature that predicts the occurrence of post-operative enterocolitis in Hirschsprung disease (HSCR) patients. Gut Microbes 2020, 11, 842–854. [Google Scholar] [CrossRef]
- Plekhova, V.; De Paepe, E.; Van Renterghem, K.; Van Winckel, M.; Hemeryck, L.Y.; Vanhaecke, L. Disparities in the gut metabolome of post-operative Hirschsprung's disease patients. Sci Rep. 2021, 11, 16167. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J.J.; Rabah, R.; Hirschl, R.B.; Maspons, A.; Meier, D.; Teitelbaum, D.H. Anti-TNF-α treatment for post-anastomotic ulcers and inflammatory bowel disease with Crohn's-like pathologic changes following intestinal surgery in pediatric patients. Pediatr Surg Int. 2015, 31, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, R.S.; Sanderson, I.R.; Claesson, M.J. Paediatric Inflammatory Bowel Disease and its Relationship with the Microbiome. Microb Ecol. 2021, 82, 833–844. [Google Scholar] [CrossRef]
- Kelay, A.; Tullie, L.; Stanton, M. Surgery and paediatric inflammatory bowel disease. TranslPediatr 2019, 8, 436–448. [Google Scholar] [CrossRef]
- Fang, X.; Vázquez-Baeza, Y.; Elijah, E.; Vargas, F.; Ackermann, G.; Humphrey, G.; Lau, R.; Weldon, K.C.; Sanders, J.G.; Panitchpakdi, M; et al. Gastrointestinal Surgery for Inflammatory Bowel Disease Persistently Lowers Microbiome and Metabolome Diversity. Inflamm Bowel Dis. 2021, 27, 603–616. [Google Scholar] [CrossRef]
- Neut, C.; Bulois, P.; Desreumaux, P.; Membré, J.M.; Lederman, E.; Gambiez, L.; Cortot, A.; Quandalle, P.; van Kruiningen, H.; Colombel, J.F. Changes in the bacterial flora of the neoterminal ileum after ileocolonic resection for Crohn's disease. Am J Gastroenterol. 2002, 97, 939–946. [Google Scholar] [CrossRef]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacteriumprausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc Natl Acad Sci U S A 2008, 105, 16731–16736. [Google Scholar] [CrossRef]
- Marteau, P.; Lémann, M.; Seksik, P.; Laharie, D.; Colombel, J.F.; Bouhnik, Y.; Cadiot, G.; Soulé, J.C.; Bourreille, A.; Metman, E.; et al. Ineffectiveness of Lactobacillus johnsonii LA1 for prophylaxis of postoperative recurrence in Crohn's disease: a randomised, double blind, placebo controlled GETAID trial. Gut. 2006, 55, 842–847. [Google Scholar] [CrossRef]
- Mondot, S.; Lepage, P.; Seksik, P.; Allez, M.; Tréton, X.; Bouhnik, Y.; Colombel, J.F.; Leclerc, M.; Pochart, P.; Doré, J.; et al. GETAID. Structural robustness of the gut mucosal microbiota is associated with Crohn's disease remission after surgery. Gut 2016, 65, 954–62. [Google Scholar] [CrossRef] [PubMed]
- Goulet, O.; Abi Nader, E.; Pigneur, B.; Lambe, C. Short Bowel Syndrome as the Leading Cause of Intestinal Failure in Early Life: Some Insights into the Management. Pediatr Gastroenterol Hepatol Nutr. 2019, 22, 303–329. [Google Scholar] [CrossRef] [PubMed]
- Massironi, S.; Cavalcoli, F.; Rausa, E.; Invernizzi, P.; Braga, M.; Vecchi, M. Understanding short bowel syndrome: Current status and future perspectives. Dig Liver Dis. 2020, 52, 253–261. [Google Scholar] [CrossRef]
- Kastl, A.J, Jr.; Terry, N.A.; Wu, G.D.; Albenberg, L.G. The Structure and Function of the Human Small Intestinal Microbiota: Current Understanding and Future Directions. Cell Mol Gastroenterol Hepatol. 2020, 9, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Sommovilla, J.; Zhou, Y.; Sun, R.C.; Choi, P.M.; Diaz-Miron, J.; Shaikh, N.; Sodergren, E.; Warner, B.B.; Weinstock, G.M.; Tarr, P.I.; et al. Small bowel resection induces long-term changes in the enteric microbiota of mice. J Gastrointest Surg. 2015, 19, 56–64, discussion 64. [Google Scholar] [CrossRef]
- Kaneko, T.; Bando, Y.; Kurihara, H.; Satomi, K.; Nonoyama, K.; Matsuura, N. Fecal microflora in a patient with short-bowel syndrome and identification of dominant lactobacilli. J Clin Microbiol. 1997, 35, 3181–3185. [Google Scholar] [CrossRef]
- Cleminson, J.S.; Young, G.R.; Campbell, D.I.; Campbell, F.; Gennery, A.R.; Berrington, J.E.; Stewart, C.J. Gut microbiome in paediatric short bowel syndrome: a systematic review and sequencing re-analysis. Pediatr Res 2025. [Google Scholar] [CrossRef]
- van der Steeg, H.J.J.; Hageman, I.C.; Morandi, A.; Amerstorfer, E.E.; Sloots, C.E.J.; Jenetzky, E.; Stenström, P.; Samuk, I.; Fanjul, M.; Midrio, P.; et al. ARM-Net Consortium. Complications After Surgery for Anorectal Malformations: An ARM-Net consortium Registry Study. J Pediatr Surg. 2025, 60, 162403. [Google Scholar] [CrossRef] [PubMed]
- Wilms, M.; Mãrzheuser, S.; Jenetzky, E.; Busse, R.; Nimptsch, U. Treatment of Hirschsprung's Disease in Germany: Analysis of National Hospital Discharge Data From 2016 to 2022. J Pediatr Surg. 2024, 59, 161574. [Google Scholar] [CrossRef] [PubMed]
- Bickler, S.W.; Rode, H. Surgical services for children in developing countries. Bull World Health Organ. 2002, 80, 829–835. [Google Scholar]
- Rajabaleyan, P.; Kaalby, L.; Deding, U.; Al-Najami, I.; Ellebæk, M.B. Mortality in patients with secondary peritonitis treated by primary closure or vacuum-assisted closure: nationwide register-based cohort study. BJS Open. 2025, 9, zraf118. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, Z.; Wu, B.X. Promoting wound recovery through stable intestinal flora: Reducing post-operative complications in colorectal cancer surgery patients. Int Wound J. 2024 Mar;21(3):e14501. Epub 2023 Dec 4. Retraction in: Int Wound J. 2025, 22, e70472. [CrossRef]
- Zheng, Z.; Hu, Y.; Tang, J.; Xu, W.; Zhu, W.; Zhang, W. The implication of gut microbiota in recovery from gastrointestinal surgery. Front Cell Infect Microbiol. 2023, 13, 1110787. [Google Scholar] [CrossRef]
- Agnes, A.; Puccioni, C.; D'Ugo, D.; Gasbarrini, A.; Biondi, A.; Persiani, R. The gut microbiota and colorectal surgery outcomes: facts or hype? A narrative review. BMC Surg. 2021, 21, 83. [Google Scholar] [CrossRef]
- Tsigalou, C.; Paraschaki, A.; Bragazzi, N.L.; Aftzoglou, K.; Bezirtzoglou, E.; Tsakris, Z.; Vradelis, S.; Stavropoulou, E. Alterations of gut microbiome following gastrointestinal surgical procedures and their potential complications. Front Cell Infect Microbiol 2023, 13, 1191126, Erratum in: Front Cell Infect Microbiol. 2023, 13,1281527. [Google Scholar] [CrossRef]
- Krezalek, M.A.; Hyoju, S.; Zaborin, A.; Okafor, E.; Chandrasekar, L.; Bindokas, V.; Guyton, K.; Montgomery, C.P.; Daum, R.S.; Zaborina, O.; et al. Can Methicillin-resistant Staphylococcus aureus Silently Travel From the Gut to the Wound and Cause Postoperative Infection? Modeling the "Trojan Horse Hypothesis. Ann Surg. 2018, 267, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat Rev Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef]
- Zaborin, A.; Smith, D.; Garfield, K.; Quensen, J.; Shakhsheer, B.; Kade, M.; Tirrell, M.; Tiedje, J.; Gilbert, J.A.; Zaborina, O.; et al. Membership and behavior of ultra-low-diversity pathogen communities present in the gut of humans during prolonged critical illness. mBio 2014, 5, e01361-14. [Google Scholar] [CrossRef]
- Krezalek, M.A.; DeFazio, J.; Zaborina, O.; Zaborin, A.; Alverdy, J.C. The Shift of an Intestinal "Microbiome" to a "Pathobiome" Governs the Course and Outcome of Sepsis Following Surgical Injury. Shock. 2016, 45, 475–82. [Google Scholar] [CrossRef]
- Dickson, R.P. The microbiome and critical illness. Lancet Respir Med. 2016, 4, 59–72. [Google Scholar] [CrossRef]
- Neogi, S.; Banerjee, A.; Panda, S.S.; Ratan, S.K.; Narang, R. Laparoscopic versus open appendicectomy for complicated appendicitis in children: A systematic review and meta-analysis. J Pediatr Surg. 2022, 57, 394–405. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, X.; Wang, X.; Zhao, L.; Lv, Y.; Chen, K. Laparoscopic versus open repair of congenital duodenal obstruction: a systematic review and meta-analysis. Pediatr Surg Int. 2022, 38, 1507–1515. [Google Scholar] [CrossRef]
- Venara, A.; Neunlist, M.; Slim, K.; Barbieux, J.; Colas, P.A.; Hamy, A.; Meurette, G. Postoperative ileus: Pathophysiology, incidence, and prevention. J Visc Surg. 2016, 153, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Alverdy, J.C.; Hyoju, S.K.; Weigerinck, M.; Gilbert, J.A. The gut microbiome and the mechanism of surgical infection. Br J Surg. 2017, 104, e14–e23. [Google Scholar] [CrossRef] [PubMed]
- Lehane, A.; Perez, M.; Smith, C.; Tian, Y.; Holl, J.L.; Raval, M.V. Slow Down Fluids to Speed Up the Intestine - Exploring Postoperative Ileus in Pediatric Gastrointestinal Surgery. J Pediatr Surg. 2025, 60, 162153. [Google Scholar] [CrossRef]
- Jin, Y.; Geng, R.; Liu, Y.; Liu, L.; Jin, X.; Zhao, F.; Feng, J.; Wei, Y. Prediction of Postoperative Ileus in Patients With Colorectal Cancer by Preoperative Gut Microbiota. Front Oncol. 2020, 10, 526009. [Google Scholar] [CrossRef]
- Yeh, A.; Rogers, M.B.; Firek, B.; Neal, M.D.; Zuckerbraun, B.S.; Morowitz, M.J. Dysbiosis Across Multiple Body Sites in Critically Ill Adult Surgical Patients. Shock. 2016, 46, 649–654. [Google Scholar] [CrossRef]
- Miquel, S.; Martín, R.; Bridonneau, C.; Robert, V.; Sokol, H.; Bermúdez-Humarán, L.G.; Thomas, M.; Langella, P. Ecology and metabolism of the beneficial intestinal commensal bacterium Faecalibacteriumprausnitzii. Gut Microbes 2014, 5, 146–51. [Google Scholar] [CrossRef]
- Wrzosek, L.; Miquel, S.; Noordine, M.L.; Bouet, S.; Joncquel Chevalier-Curt, M.; Robert, V.; Philippe, C.; Bridonneau, C.; Cherbuy, C.; Robbe-Masselot, C.; et al. Bacteroides thetaiotaomicron and Faecalibacteriumprausnitzii influence the production of mucus glycans and the development of goblet cells in the colonic epithelium of a gnotobiotic model rodent. BMC Biol. 2013, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Türler, A.; Schwarz, N.T.; Türler, E.; Kalff, J.C.; Bauer, A.J. MCP-1 causes leukocyte recruitment and subsequently endotoxemic ileus in rat. Am J PhysiolGastrointest Liver Physiol. 2002, 282, G145–55. [Google Scholar] [CrossRef]
- Jørgensen, A.B.; Jonsson, I.; Friis-Hansen, L.; Brandstrup, B. Collagenase-producing bacteria are common in anastomotic leakage after colorectal surgery: a systematic review. Int J Colorectal Dis. 2023, 38, 275. [Google Scholar] [CrossRef]
- Gershuni, V.M.; Friedman, E.S. The Microbiome-Host Interaction as a Potential Driver of Anastomotic Leak. Curr Gastroenterol Rep. 2019, 21, 4. [Google Scholar] [CrossRef] [PubMed]
- Dias, V.E.; Castro, P.A.S.V.; Padilha, H.T.; Pillar, L.V.; Godinho, L.B.R.; Tinoco, A.C.A.; Amil, R.D.C.; Soares, A.N.; Cruz, G.M.G.D.; BezerraJMT; et al. Preoperative risk factors associated with anastomotic leakage after colectomy for colorectal cancer: a systematic review and meta-analysis. Rev Col Bras Cir. 2022, 49, e20223363. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Gauda, E.B.; Chiu, P.P.L.; Moore, A.M. Risk factors for prolonged mechanical ventilation in neonates following gastrointestinal surgery. TranslPediatr 2022, 11, 617–624. [Google Scholar] [CrossRef]
- Gonzalez, D.O.; Nwomeh, B.C. Complications in children with ulcerative colitis undergoing ileal pouch-anal anastomosis. Semin Pediatr Surg. 2017, 26, 384–390. [Google Scholar] [CrossRef]
- Liu, Y.; Li, B.; Wei, Y. New understanding of gut microbiota and colorectal anastomosis leak: A collaborative review of the current concepts. Front Cell Infect Microbiol 2022, 12, 1022603. [Google Scholar] [CrossRef]
- Okada, M.; Bothin, C.; Kanazawa, K.; Midtvedt, T. Experimental study of the influence of intestinal flora on the healing of intestinal anastomoses. Br J Surg. 1999, 86, 961–665. [Google Scholar] [CrossRef]
- Christley, S.; Shogan, B.; Levine, Z.; Koo, H.; Guyton, K.; Owens, S.; Gilbert, J.; Zaborina, O.; Alverdy, J.C. Comparative genetics of Enterococcus faecalis intestinal tissue isolates before and after surgery in a rat model of colon anastomosis. PLoS One 2020, 15, e0232165. [Google Scholar] [CrossRef]
- Zamorano, D.; Ivulic, D.; Viver, T.; Morales, F.; López-Kostner, F.; Vidal, R.M. Microbiota Phenotype Promotes Anastomotic Leakage in a Model of Rats with Ischemic Colon Resection. Microorganisms 2023, 11, 680. [Google Scholar] [CrossRef] [PubMed]
- Boatman, S.; Kaiser, T.; Nalluri-Butz, H.; Khan, M.H.; Dietz, M.; Kohn, J.; Johnson, A.J.; Gaertner, W.B.; Staley, C.; Jahansouz, C. Diet-induced shifts in the gut microbiota influence anastomotic healing in a murine model of colonic surgery. Gut Microbes. 2023, 15, 2283147. [Google Scholar] [CrossRef]
- van Praagh, J.B.; de Goffau, M.C.; Bakker, I.S.; van Goor, H.; Harmsen, H.J.M.; Olinga, P.; Havenga, K. Mucus Microbiome of Anastomotic Tissue During Surgery Has Predictive Value for Colorectal Anastomotic Leakage. Ann Surg. 2019, 269, 911–916. [Google Scholar] [CrossRef]
- Lianos, G.D.; Frountzas, M.; Kyrochristou, I.D.; Sakarellos, P.; Tatsis, V.; Kyrochristou, G.D.; Bali, C.D.; Gazouli, M.; Mitsis, M.; Schizas, D. What Is the Role of the Gut Microbiota in Anastomotic Leakage After Colorectal Resection? A Scoping Review of Clinical and Experimental Studies. J Clin Med. 2024, 13, 6634. [Google Scholar] [CrossRef] [PubMed]
- Komen, N.; Slieker, J.; Willemsen, P.; Mannaerts, G.; Pattyn, P.; Karsten, T.; de Wilt, H.; van der Harst, E.; van Leeuwen, W.; Decaestecker, C.; et al. Polymerase chain reaction for Enterococcus faecalis in drain fluid: the first screening test for symptomatic colorectal anastomotic leakage. The Appeal-study: analysis of parameters predictive for evident anastomotic leakage. Int J Colorectal Dis. 2014, 29. [Google Scholar] [CrossRef] [PubMed]
- Palmisano, S.; Campisciano, G.; Iacuzzo, C.; Bonadio, L.; Zucca, A.; Cosola, D.; Comar, M.; de Manzini, N. Role of preoperative gut microbiota on colorectal anastomotic leakage: preliminary results. Updates Surg. 2020, 72, 1013–1022. [Google Scholar] [CrossRef]
- Mima, K.; Sakamoto, Y.; Kosumi, K.; Ogata, Y.; Miyake, K.; Hiyoshi, Y.; Ishimoto, T.; Iwatsuki, M.; Baba, Y.; Iwagami, S.; et al. Mucosal cancer-associated microbes and anastomotic leakage after resection of colorectal carcinoma. Surg Oncol. 2020, 32, 63–68. [Google Scholar] [CrossRef]
- Hajjar, R.; Gonzalez, E.; Fragoso, G.; Oliero, M.; Alaoui, A.A.; Calvé, A.; VenninRendos, H.; Djediai, S.; Cuisiniere, T.; Laplante, P.; et al. Gut microbiota influence anastomotic healing in colorectal cancer surgery through modulation of mucosal proinflammatory cytokines. Gut. 2023, 72, 1143–1154. [Google Scholar] [CrossRef]
- Wiegerinck, M.; Hyoju, S.K.; Mao, J.; Zaborin, A.; Adriaansens, C.; Salzman, E.; Hyman, N.H.; Zaborina, O.; van Goor, H.; Alverdy, J.C. Novel de novo synthesized phosphate carrier compound ABA-PEG20k-Pi20 suppresses collagenase production in Enterococcus faecalis and prevents colonic anastomotic leak in an experimental model. Br J Surg. 2018, 105, 1368–1376. [Google Scholar] [CrossRef]
- Kuusela, P. Fibronectin binds to Staphylococcus aureus. Nature 1978, 276(5689), 718–720. [Google Scholar] [CrossRef]
- Singh, B.; Fleury, C.; Jalalvand, F.; Riesbeck, K. Human pathogens utilize host extracellular matrix proteins laminin and collagen for adhesion and invasion of the host. FEMSMicrobiol Rev. 2012, 36, 1122–1180. [Google Scholar] [CrossRef] [PubMed]
- Wildeboer, A.C.L.; van Helsdingen, C.P.M.; Gallé, C.G.; de Vries, R.B.M.; Derikx, J.P.M.; Bouvy, N.D. Enhancing intestinal anastomotic healing using butyrate: Systematic review and meta-analysis of experimental animal studies. PLoS One. 2023, 18, e0286716. [Google Scholar] [CrossRef] [PubMed]
- Aghighi, F.; Salami, M. What we need to know about the germ-free animal models. AIMS Microbiol. 2024, 10, 107–147. [Google Scholar] [CrossRef] [PubMed]
- Rigby, R.J.; Hunt, M.R.; Scull, B.P.; Simmons, J.G.; Speck, K.E.; Helmrath, M.A.; Lund, P.K. A new animal model of postsurgical bowel inflammation and fibrosis: the effect of commensal microflora. Gut 2009, 58, 1104–1112. [Google Scholar] [CrossRef]
- Hyoju, S.; Machutta, K.; Krezalek, M.A.; Alverdy, J.C. What Is the Role of the Gut in Wound Infections? Adv Surg. 2023, 57, 31–46. [Google Scholar] [CrossRef]
- Włodarczyk, J.; Dziki, Ł.; Harmon, J.; Fichna, J. The role of short-chain fatty acids in the prehabilitation before colorectal surgery. CurrProbl Surg. 2025, 69, 101810. [Google Scholar] [CrossRef]
- Keskey, R.; Papazian, E.; Lam, A.; Toni, T.; Hyoju, S.; Thewissen, R.; Zaborin, A.; Zaborina, O.; Alverdy, JC. Defining Microbiome Readiness for Surgery: Dietary Prehabilitation and Stool Biomarkers as Predictive Tools to Improve Outcome. Ann Surg. 2022, 276, e361–e369. [Google Scholar] [CrossRef]
- Avis, T.; Wilson, F.X.; Khan, N.; Mason, C.S.; Powell, D.J. Targeted microbiome-sparing antibiotics. Drug Discov Today 2021, 26, 2198–2203. [Google Scholar] [CrossRef]
- Muñoz, K.A.; Ulrich, R.J.; Vasan, A.K.; Sinclair, M.; Wen, P.C.; Holmes, J.R.; Lee, H.Y.; Hung, C.C.; Fields, C.J.; Tajkhorshid, E.; et al. A Gram-negative-selective antibiotic that spares the gut microbiome. Nature. 2024, 630, 429–436. [Google Scholar] [CrossRef]
- Yang, Y.; Xia, Y.; Chen, H.; Hong, L.; Feng, J.; Yang, J.; Yang, Z.; Shi, C.; Wu, W.; Gao, R.; et al. The effect of perioperative probiotics treatment for colorectal cancer: short-term outcomes of a randomized controlled trial. Oncotarget. 2016, 7, 8432–8440. [Google Scholar] [CrossRef]
- Ruiz, N.I.; Herrera Giron, C.G.; Arragan Lezama, C.A.; Frias Redroban, S.J.; Ventura Herrera, M.O.; SanicCoj, G.A. Timing and Protocols for Microbiome Intervention in Surgical Patients: A Literature Review of Current Evidence. Cureus 2025, 17, e86104. [Google Scholar] [CrossRef]
- Huang, F.; Li, S.; Chen, W.; Han, Y.; Yao, Y.; Yang, L. Postoperative Probiotics Administration Attenuates Gastrointestinal Complications and Gut Microbiota Dysbiosis Caused by Chemotherapy in Colorectal Cancer Patients. Nutrients 2023, 15, 356. [Google Scholar] [CrossRef] [PubMed]
- Kanamori, Y; Sugiyama, M; Hashizume, K; Yuki, N; Morotomi, M; Tanaka, R. Experience of long-term synbiotic therapy in seven short bowel patients with refractory enterocolitis. J Pediatr Surg. 2004, 39, 1686–92. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Takahashi, T.; Inoue, M.; Morotomi, M.; Otake, K.; Nakazawa, M.; Tsukamoto, Y.; Miki, C.; Kusunoki, M. Immunonutritional effects during synbiotics therapy in pediatric patients with short bowel syndrome. Pediatr Surg Int. 2007, 23, 243–248. [Google Scholar] [CrossRef]
- Rao, S.; Esvaran, M.; Chen, L.; Keil, A.D.; Gollow, I.; Simmer, K.; Wemheuer, B.; Conway, P.; Patole, S. Probiotic supplementation in neonates with congenital gastrointestinal surgical conditions: a pilot randomised controlled trial. Pediatr Res. 2022, 92, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.L.; Sun, Y.; Dong, X.; Ren, Z.; Li, B.; Yang, P.; et al. Infant feces-derived Lactobacillus gasseriFWJL-4 mitigates experimental necrotizing enterocolitis via acetate production. Gut Microbes 2024, 16, 2430541. [Google Scholar] [CrossRef]
- Matsui, R.; Sagawa, M.; Sano, A.; Sakai, M.; Hiraoka, S.I.; Tabei, I.; et al. Impact of Perioperative Immunonutrition on Postoperative Outcomes for Patients Undergoing Head and Neck or Gastrointestinal Cancer Surgeries: A Systematic Review and Meta-analysis of Randomized Controlled Trials. Ann Surg. 2024, 279, 419–428. [Google Scholar] [CrossRef]
- Sukhotnik, I.; Levi, R.; Moran-Lev, H. Impact of Dietary Protein on the Management of Pediatric Short Bowel Syndrome. Nutrients 2023, 15, 2826. [Google Scholar] [CrossRef]
- Mousa, W.K.; Al Ali, A. The Gut Microbiome Advances Precision Medicine and Diagnostics for Inflammatory Bowel Diseases. Int J Mol Sci. 2024, 25, 11259. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Gan, G.; Cheng, J.; Shen, Z.; Zhang, G.; Liu, S.; Hu, J. Engineered Probiotics Enable Targeted Gut Delivery of Dual Gasotransmitters for Inflammatory Bowel Disease Therapy. Angew Chem Int Ed Engl. 2025, 64, e202502588. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, Y. Phage therapy as a revitalized weapon for treating clinical diseases. Microbiome Res Rep. 2025, 4, 35. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The relation between surgery and GM-related complications.
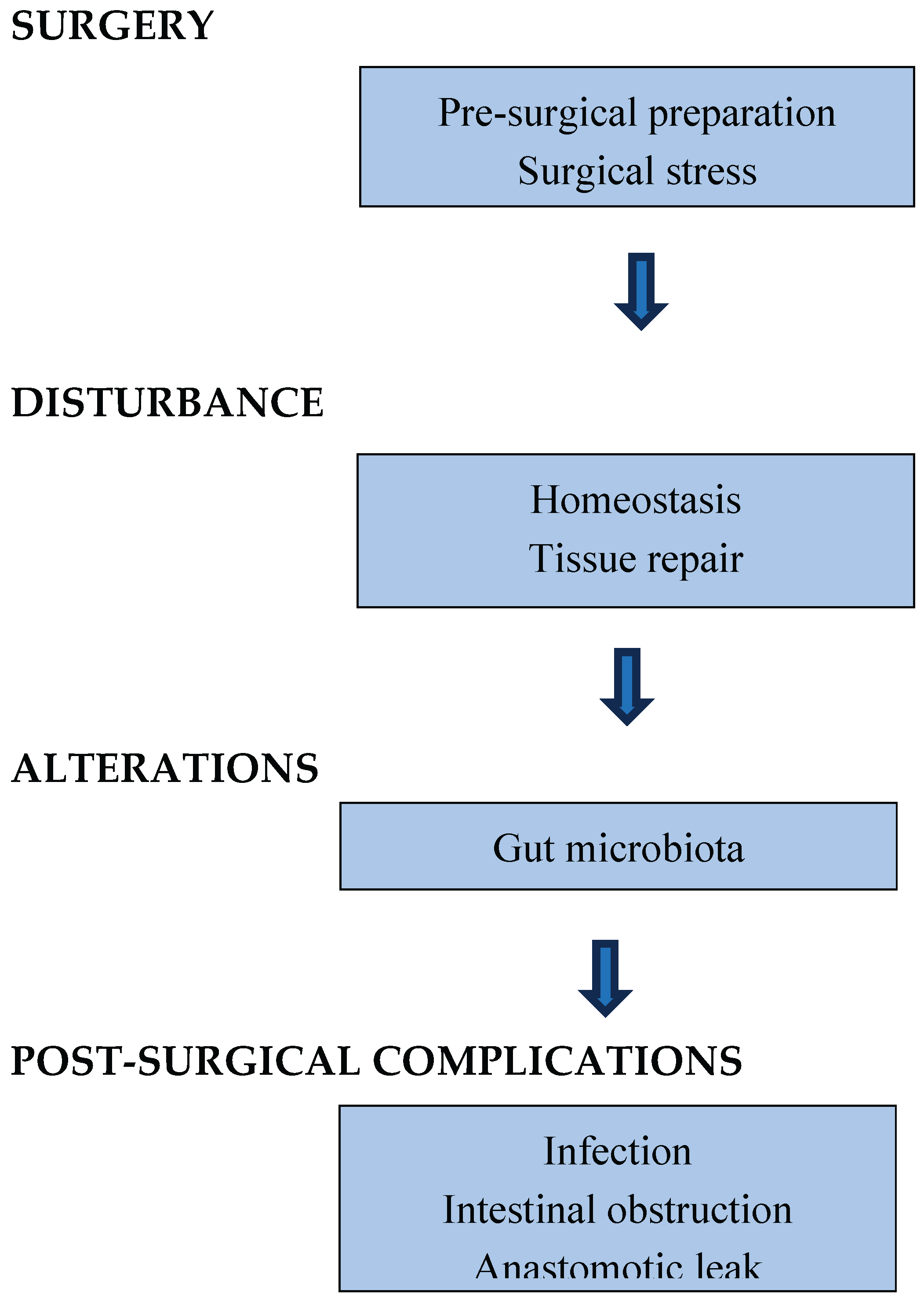

Table 1.
Post-surgical GM alterations and outcomes in paediatric patients with intestinal diseases.
| Authors (year) |
Type of study | Disease | Gut alterations | Outcomes | Reference |
|---|---|---|---|---|---|
|
Lin et al. (2023) |
Prospective case control |
NEC |
↓ Alpha-diversity after full enteral nutrition. ↑ Methylobacterium, Clostridiumbutyricum, Acidobacteria. |
Enriched Escherichia coli, Pseudomonas; depleted Bacteroidales, Ruminococcaceae |
[10] |
|
Neuvonen et al. (2018) |
Prospective observational |
HD |
Enriched Escherichia coli, Pseudomonas; depleted Bacteroidales, Ruminococcaceae |
Baseline dysbiosis persists post-surgery, defining a state of ecological failure. |
[11] |
|
Pini Prato et al. (2019) |
Retrospective |
HD |
↓ Alpha diversity and altered beta diversity in total colonic aganglionosisvs rectosigmoid. |
The extent of aganglionosis directly shapes the GM ecosystem. |
[124] |
| Yan et al. (2014) | Cross -sectional | HAEC | Proteobacteria-dominated GM | Bacteroides-rich community in stable patients. | [91] |
|
Frykman et al. (2015) |
Cross-sectional |
HAEC |
Separate fungal dysbiosis: loss of diversity, bloom of Candida |
HAEC involves a disruption of both bacterial and fungal communities |
[126] |
|
Murphy et al. (2025) |
Prospective observational |
HD |
Absence of healthy developmental trajectory: No increase in alpha diversity with age vs healthy controls ↑Fusobacteria linkedtoinflammation. |
Disrupted GM maturation is linked to persistent GI inflammation and symptoms post-pull-through |
[123] |
|
Davidocs et al. (2016) |
Prospective observational |
SΒS |
↑ Escherichia coli/Shigella ↑ Streptococcus Relative abundance of Lactobacillus noted in patients with diarrhea. |
Associated with D-lactic acidosis due to high D-lactate production, leading to metabolic acidosis. |
[102] |
|
Cleminson et al. (2025) |
Systematic review | SBS |
Depletion of beneficial SCFA-producers: ↓ Dorea, ↓ Ruminococcus, ↓ Blautia (genera from Lachnospiraceae family) in children on TPN. |
Deficit of protective microbial metabolites, exacerbating mucosal inflammation. Likely driven by antibiotics and bacterial overgrowth. |
[141] |
NEC: necrotizing enterocolitis, HD: Hirschsprung’s disease, HAEC: Hirschsprung’s -associated enterocolitis, SBS: short bowel syndrome.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



