Submitted:
12 December 2025
Posted:
15 December 2025
You are already at the latest version
Abstract
Drought stress is a primary constraint on global coffee production, threatening the industry's sustainability. This review provides a comprehensive synthesis of the determinants and physiological and agronomic impacts of drought stress on coffee plants, and critically evaluates current adaptation strategies. We delineate how drought severity is governed by the interplay of genetic factors (e.g., the greater susceptibility of Coffea arabica compared to C. canephora), soil properties, and local climatic conditions. The primary physiological disruptions triggered by drought—beginning with stomatal closure and oxidative stress, and culminating in severely reduced photosynthesis—compromise plant vigor and resource allocation. Consequently, these physiological insufficiencies manifest as significant agronomic losses, including stunted growth, yield reduction, bean quality deterioration, and heightened susceptibility to pests and diseases. Compounding these direct impacts, this work synthesizes climate modeling evidence indicating that prolonged drought risk may ultimately lead to a significant geographic shift in viable coffee production areas. By identifying the major determinants of drought stress—such as genetic variety, soil properties, and local climate patterns—this review establishes a foundation for targeted solutions. In response to these multifaceted challenges, we systematically analyze a suite of agronomic interventions—from drought-tolerant cultivars and irrigation to agroforestry and soil management—outlining their efficacy in building resilience. The review concludes by identifying critical knowledge gaps, establishing that future research must prioritize the development of integrated, scalable adaptation frameworks and advance breeding programs for drought tolerance, thereby enhancing climate-resilient coffee production.
Keywords:
coffea arabica
; drought resistant cultivar
; drought severity
; drought mitigation
; genetic factor
; irrigation
1. Introduction
Coffee is one of the most significant agricultural commodities in the world. It is the second-most globally traded commodity after petroleum [1] and a significant contributor for the socio-economic growth of many developing countries in the tropics. The coffee industry has a vast and a complex network, encompassing a wide range of activities from cultivation to consumption, generating a huge amount of revenues through the value chain and impacting the livelihoods of millions, worldwide [2]. For instance, coffee production and the entire coffee sector in 2022-2023 cropping year generated about 11.6 and 173.0 billion US dollar gross revenue, respectively [3]. Moreover, around 25 million farming families around the world, majority of them are smallholder farmers, produce coffee and deeply depend on this crop for their livelihoods [4,5]. In addition to this, coffee is deeply ingrained in cultural and social fabrics for the peoples of some countries like Ethiopia which is recognized as the birthplace of Arabica coffee. Coffee in Ethiopia is a symbol of hospitality and tradition of the nations and a backbone of the national economy, contributing to around 30% of the country’s foreign exchange earnings [6] and providing job opportunity and livelihood for about 25% of the country’s population [7].
Despite its global economic and social importance, coffee production is highly vulnerable to the impacts of climate change. Among these, drought stress represents a particularly critical threat, given the crop’s heavy reliance on predictable rainfall patterns. Drought, defined as a period of insufficient precipitation leading to soil moisture depletion that cannot meet plant water requirements [8], directly compromises coffee cultivation. In recent years, drought has becoming more frequent and severe, caused substantial declines in coffee yield as coffee production relies heavily on rain-fed agriculture [9]. The yield of Arabica coffee is projected to decline in many coffee-growing regions due to climate variability, particularly to extreme temperatures and water deficit [10,11] and a severe drought during the 2015–2016 cropping year in Brazil caused the yield reductions of 50–80% in un-irrigated Arabica coffee [12]. Particularly, water deficit during the key developmental stages of coffee, such as flowering and berry formation, leads to reduced productivity and bean quality [10]. Drought stress, however, affects coffee at multiple scales from gene expression and leaf gas exchange to whole-plant growth, yield and bean quality [13,14,15]. It affects growth and dry matter production by reducing relative water content, leaf water potential, turgor pressure, and stomatal aperture, and thereby reducing photosynthetic rate, and cell elongation and expansion [16,17,18]. Stomatal closure, which reduces the availability of CO2 in leaves and inhibits carbon fixation, exposes chloroplasts to excessive excitation energy, which in turn could increase the generation of reactive oxygen species (ROS) that can induce oxidative stress, leading to cellular damage [19,20]. Drought stress also negatively affects dry matter partitioning and temporal biomass distribution [21,22]. Reports indicate that changes in precipitation patterns will decrease yield, reduce quality, increase pest and disease pressure, and decrease coffee-suitable land by 2050 [13]. Yet, these drought effects vary with the genetics of coffee plants (e.g., species and cultivar), growing environment (e.g., soil and climate), land physiognomy (e.g., elevation, slope, aspect and shape), and farm management [23,24].
Over the past three decades, a considerable number of studies have been conducted to understand the morphological, physiological, and biochemical responses of coffee to drought stress [25,26,27,28,29,30,31] and identify the agronomic practices that mitigate the effects of drought on coffee [4,30,32]. Despite these efforts, significant challenges remain to exist and one of the key limitations is the slow pace of translating the research findings into practical solutions that can be widely adopted by farmers. The complex interactions between genetic, environmental, and management factors in determining the drought resistance in coffee hampers the progress of addressing the drought problem effectively [33]. Furthermore, while previous reviews have addressed plant physiology or specific adaptation techniques in isolation, a comprehensive and integrated analysis linking the determinants of drought stress to its physiological manifestations, agronomic consequences, and the practical scalability of adaptation strategies is lacking This situation underscores a critical need for a more integrated understanding of drought stress impacts and their interacting variables, which is essential for developing robust adaptive strategies to ensure the sustainability of coffee production in a changing climate. This review therefore seeks to fill this gap by providing a holistic synthesis that connects the root causes of drought stress to tangible on-the-ground solutions, critically integrating the latest evidence to inform future research and action. Hence, this review aims: 1) to synthesize the documented impacts of drought on coffee physiology and agronomic performance; 2) to identify the key factors that determine the severity of drought impacts on coffee plants; 3) to evaluate potential adaptation strategies to mitigate the effects of drought stress; and 4) to pinpoint critical knowledge gaps to guide and prioritize future research directions. To achieve these objectives, this review systematically addresses three critical areas. First, it analyzes the principal physiological and agronomic impacts of drought stress on coffee plants. Second, it examines the key genetic, environmental, and management factors that determine the severity of these impacts. Finally, it synthesizes and evaluates the spectrum of adaptation strategies available to mitigate the effects of drought on coffee production. Through this structured analysis, the review aims to consolidate existing knowledge and identify pivotal research gaps to inform future work on building resilient coffee systems.
2. Effects of Drought Stress on Coffee
The effects of drought stress on coffee plants are wide-ranging. It disrupts many cellular and whole-plant functions, which impact plant growth, yield and quality of coffee by disturbing the assimilate distribution balance between roots and shoots, reducing production of new leaves and stem elongation, increasing leaf chlorosis, branch die-back, impacting flowering, and reducing fruiting nodes per tree, number of fruits per node, seed size, seed density and seed uniformity, all of which lead to a yield loss that ranges from 40-80% [34,35,36]. Coffee plants under drought stress exhibit reduced plant and leaf growth, premature leaf shedding, wilting, and impaired photosynthesis due to stomatal closure, which can drastically reduce the net carbon assimilation rate [10]. Furthermore, drought influences the incidences of coffee pests and diseases, such as coffee leaf rust, coffee thrips and coffee berry borer which could reduce coffee yield and quality and increase production costs [37]. As indicated in Figure 1, this affects the plant’s overall health and ability to produce fruits. Drought also causes a substantial economic loss for coffee farmers, which leads to vulnerability for food insecurity particularly for smallholders who greatly rely on coffee for their livelihoods [38].
2.1. Physiological and Growth Impacts of Drought. Stress
Drought stress initiates a cascade of physiological disruptions in coffee plants that ultimately manifest as reduced growth and productivity. This section delineates the primary physiological effects from the subsequent impacts on growth and biomass allocation.
2.1.1. Primary Physiological Disruptions
The initial plant response to water deficit is a reduction in relative water content, leading to decreased leaf water potential and turgor pressure. This hydraulic change triggers stomatal closure to minimize water loss, which simultaneously limits CO₂ availability in the mesophyll. The direct consequence is a severe reduction in photosynthetic rate; recent studies indicate drought can reduce photosynthetic efficiency by up to 50% [15,16,18,39]. The impairment of photosynthesis subsequently curtails the synthesis of essential sugars and proteins, forming the foundational disruption for all downstream effects [40].
Concurrently, the restriction of photosynthetic electron transport under drought leads to an increase in oxidative stress. Chloroplasts exposed to excess excitation energy generate reactive oxygen species (ROS), causing oxidative damage to lipids, proteins, and DNA [19,41,42]. This damage, exemplified by significant increases in lipid peroxidation and protein oxidation, impairs cellular membranes and enzyme functions, further compromising plant health and resilience [43].
2.1.2. Consequences on Plant Growth and Biomass Partitioning
The primary physiological disruptions directly translate into significant reductions in growth and biomass accumulation. The relative growth rate (RGR) declines, primarily due to a decrease in the net assimilation rate (NAR)—a physiological adjustment—rather than morphological changes like specific leaf area [44,45]. This results in stunted growth and reduced dry matter production, with growth rates potentially declining by 20-30% [39].
Furthermore, drought stress profoundly alters dry matter partitioning. To enhance survival, plants shifts the balance of assimilate distribution, prioritizing root development over shoot growth to improve water uptake [21,46,47]. This strategic reallocation of resources often results in a decrease in leaf area by up to 30% [17], which in turn creates a negative feedback loop by further limiting the plant’s photosynthetic capacity [48]. The process is exacerbated as drought impairs photosynthesis and decreases sucrose content in leaves, reducing the export rate of assimilates from sources to sinks [49,50]. Additionally, drought limits the sink organs’ ability to utilize incoming assimilates efficiently and disrupts phloem loading/unloading through the inhibition of enzymes like acid invertase [51].
In summary, drought stress triggers a well-defined sequence of events, beginning with stomatal closure and oxidative stress, which severely inhibits photosynthesis. These physiological insufficiencies subsequently manifest as reduced vegetative growth and a strategic, yet ultimately limiting, reallocation of biomass away from the shoots.
2.2. Effects of Drought on Yield and Quality
Drought stress during the critical developmental stages of coffee, such as plant growth, flowering and fruit development, profoundly impacts coffee bean yield and quality. Reduced water availability during the plant growth period can limit plant growth and during fruiting impacts bean development, leading to lower yield and quality of coffee. Researches have demonstrated that water deficits during flowering and berry development can reduce coffee yield by up to 50% and negatively affect size, weight, and chemical composition of coffee beans [35,36]. Bunn et al. [32] found that drought stress during the flowering stage can reduce coffee yield by up to 30%, with significant decreases in bean size and weight. Reduced water availability during this stage can lead to poor flowering and fruit setting, and lower bean quality, resulting in decreased economic returns for farmers [52]. It is important to note that the magnitude of these reductions is highly context-dependent, varying significantly with the coffee cultivar (e.g., the greater sensitivity of C. arabica vs. C. canephora), soil water-holding capacity, the duration and severity of the drought stress, and local microclimatic conditions [15,22]. Therefore, these figures should be interpreted as indicative of a potential range of impacts rather than as universal constants.
Coffee plants under drought stress shows uneven ripening and produces smaller beans of uneven size and low density. Uneven ripening leads to defects in bean texture and flavor and beans with varying size are bad for specialty coffee roasters who greatly need uniform beans to get a uniform roast and smaller beans are not usually acceptable at export markets. Drought stress also alters the bean composition, such as chlorogenic acids, sucrose, and caffeine, which determines the cup quality attributes of the beans [35,36,53]. As it impacts their profit and market competitiveness, this decline in yield and quality is a critical concern for coffee producers.
2.3. Effects of Drought on Coffee Pests and Diseases
Drought stress not only directly stresses the plant but also indirectly reduces yields by exacerbating the incidence and infestation of some coffee pests and diseases, such as coffee leaf rust (Hemileia vastatrix), coffee berry borer (Hypothenemus hampei), and coffee thrips (Diarthrothrips coffeae). These coffee pests and diseases thrive under drought conditions, which can enhance their reproductive success and survival rates [54] and further reduce yield and quality of coffee [55]. Drought also influences the dynamics of plant-pathogen interactions, often leading to increased disease severity due to compromised plant immunity [56,57]. A study by Vázquez et al. [56] found that drought-stressed coffee plants have a 40% higher incidence of coffee leaf rust and a 30% increase in coffee berry borer infestation. Drought can also shift the distribution of pests, allowing them to invade new areas, particularly in regions where coffee is grown under suboptimal conditions [58,59]. In addition to the yield losses brought by direct impact of drought, the increased vulnerability of drought-stressed plants to these biotic stressors poses a significant challenge to coffee farmers, as it necessitates additional pest and disease management measures, which increases the costs of production.
2.4. Effects of Drought on Suitable Area for Coffee Cultivation
Several studies [9,32,59,60] predicted that suitable areas for coffee cultivation could significantly be decreased due to climate change. As temperature rises and precipitation pattern becomes more erratic, the land available for optimal coffee production is gradually shrinking. This contraction in suitable areas poses a threat to global coffee supply and necessitates the development of more resilient coffee varieties and cultivation practices [61]. Bunn et al. [32] predicted that by 2050, the suitable area for coffee cultivation in the common coffee-growing regions of the world could decrease by up to 50%. The common coffee-growing regions in countries like Brazil, Colombia, and Ethiopia are facing with increasing challenges due to drought and high temperature stresses. This leads to a shift in cultivation zones to higher altitudes and more temperate regions where the growing conditions are more favorable [32,61]. However, these new areas often come with their own set of challenges, including soil unsuitability and pests and diseases adapted to the highland or temperate areas [62]. To counteract the effects of drought and the associated shifts in coffee-growing regions, adaptation strategies, such as developing and using climate-resilient coffee varieties and climate smart cultivation practices, are essential.
3. Determinants of Drought Impacts on Coffee
The impacts of drought stress on coffee depends on multiple interacting factors that include the drought nature, plant genetics, soil and site characteristics, broader climatic context, farm management, and landscape elevation [23,24].
3.1. Duration, Severity, and Intensity of Drought
The negative impacts of drought are expected to become greater when the drought episodes associated with the present and ongoing climate changes are longer, and more frequent and intense [2,63]. The duration, severity, and intensity of drought, each play a critical role in determining the level of drought impacts on coffee plants. They determine the physiological, biochemical, and morphological responses of coffee, ultimately affecting the plant growth, yield, and quality (Table1).
The duration of drought refers to the length of time during which water availability is below the optimal levels for plant growth. Drought severity refers to the degree to which water is limited to plants and drought intensity refers to how quickly the water deficit develops and how extreme the conditions are. Prolonged drought can lead to extensive reductions in coffee yield and quality. For instance, extended water stress can cause severe leaf shedding to conserve water, significantly reducing the photosynthetic area [10]. Long-term drought also compromises the root function, reducing the plant’s ability to access available nutrients and water. Chronic water scarcity affects berry development, resulting in poor-quality beans with lower sugar content and impaired flavor [64]. Severe drought triggers oxidative stress, leading to cellular damage through the accumulation of ROS [65] and significantly reduces leaf water potential and photosynthetic activity, leading to leaf necrosis and plant wilting. It can also bring a reproductive failure by reducing fruit setting and causing a premature fruit drop [66]. A high-intensity drought causes a rapid reduction in soil water content, which significantly limits water availability to roots. This leads to stomatal closure, reduced carbon assimilation, and ultimately decreased plant growth [67]. The intense drought condition can also impair root hydraulic conductivity, limiting the plant’s ability to recover after rehydration, which exacerbates the damage and prolongs the recovery period. Understanding these drought situations is essential for developing effective mitigation strategies, which could be helpful for developing drought-tolerant varieties and management practices that ensure stable coffee production under the changing climatic conditions.

Table 1.
Determinants of Drought Impacts on Coffee.
| Determinant Factors | Outcomes of the Factors | References |
|---|---|---|
| Duration, Severity, Intensity | Longer, more severe or intense, lead to oxidative stress, reproductive failure, and lead to yield loss | [2,10,63,64,65,66,67] |
| Species and Cultivar | Tolerant varieties exhibit longer roots; close stomata and higher water use efficiency, osmolyte accumulation, and delayed wilting. | [15,23,28,29,30,31,68,69,70] |
| Temperature | Synergistically worsening drought impacts | [9,10,12,30,75,76] |
| Sunshine, Humidity & Wind | High solar radiation, low humidity, and strong winds increase atmospheric demand and evapotranspiration, rapidly depleting soil moisture. | [30,66,75,77,78,79,80] |
| Slope, Aspect, Curvature | Steep slopes and convex ridges increase runoff worsen drought impact; west-facing slopes are drier. Gentle, concave, and lower slopes retain more water better alleviate drought impacts. | [30,66,68,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102] |
| Soil Fertility | Degraded soils aggravate drought impacts as compared to fertile soil . | [104,105,106,107,108,109] |
| Farm Management | Practices like agroforestry, conservation, and precision soil and water management enhance drought resilience, while ignored fields are more prone to drought impacts. | [10,111,112,113,114,115] |
3.2. Genetic Factors
Differences in coffee genetics are vital for drought management and the diversity among coffee cultivars in morphological, anatomical, and physiological traits can confer resilience to drought stress [68]. Owing to its origin in warmer lowland areas of Africa and its typically deeper roots and vigorous growth, C. canephora (Robusta) is generally expedited to be more drought- and heat-tolerant than C. arabica. In comparative trials, Robusta clones maintained higher stomatal conductance and turgor at low water potentials than Arabica [23]. However, as Robusta often grows in hotter regions, it can suffer from extreme droughts combined with high heat stresses.
Several studies [15,28,29,30,31] also indicate notable varietal variations within coffee species in drought response. For example, some Arabica cultivars of Ethiopia maintained 2–3 times higher CO₂ assimilation and biomass accumulation under a prolonged drought period than the sensitive ones. The tolerant genotypes accumulated more osmolytes (glucose, maltose, and amino acids) to maintain cell turgor, a strategy not seen in sensitive varieties [15]. Similarly, Avila et al. [69] and Koutouleas et al. [70] reported elite Arabica hybrids and Robusta clones bred for drought resilience showed higher water use efficiency (WUE) and a delayed leaf wilting under the drought stress condition. Omics studies reveal that different coffee genotypes employ distinct molecular strategies, which explains their varying resilience levels. In Robusta coffee (e.g., CL153), shows a strong enrollment of ABA-signaling pathways, protein kinases, and phosphatases in response to drought stress [27]. A key feature is the marked up-regulation of aspartic protease genes, which are thought to be involved in protein recycling and stress signaling during severe water deficit [27,56]. In Arabica coffee, the superior resilience of the Icatu cultivar is linked to a more robust and coordinated activation of a protective network. This includes a greater abundance of transcripts and proteins related to heat shock proteins (HSPs), aquaporins, and antioxidant systems [63]. Icatu also shows a higher expression of genes involved in lipid metabolism and cuticle formation, such as lipid transfer proteins (LTPs), which help minimize water loss. Thus, cultivar choice strongly determines the impacts of drought responses on the coffee crop: farmers in dry regions often adopt locally adapted varieties (e.g., Sarchimor or Catimor hybrids) known for their drought stress tolerance. Breeding programs also now focus on water-stress traits of coffee, such as stomatal behavior, root depth, osmoprotectant metabolism and antioxidant capacity [15,23]. Such genotypic plasticity in response to drought stress involves inherent genetic traits that enhance the plant’s ability to cope with stress, regardless of environmental triggers.
3.3. Environmental Factors
A combination of low precipitation and high evapotranspiration (ET) causes a drought stress in plants [8,71]. Plants may experience transient drought stress during the noon hours of hot days even with adequate rainfall or irrigation [72] due to a higher rate of transpiration compared to that of plant absorption. A high evaporative demand, which is determined by low relative humidity and high irradiance, temperature, vapor pressure deficit (VPD) and wind, accelerates the drought stress development in plants [73]. An increase in soil temperature resulting from air temperature increase may be even stronger when it is accompanied by a drought-induced decline in soil water content [74]. These effects can be further aggravated by a progressive global warming.
3.3.1. Temperature
Temperature is a primary environmental factor that determines the growth and development of coffee plants. It influences the physiological processes of coffee that includes photosynthesis, respiration, and transpiration. Therefore, optimizing and minimizing the limitations of the thermal resource of the growing environment is required to meet the objectives of the coffee business, i.e., quantity and quality of crop production [75]. Optimal growth temperatures for Arabica coffee range from 18 °C to 22 °C. Deviations from this range, particularly in the form of elevated temperatures, can lead to increased transpiration rates, exacerbating water loss and contributing to drought stress [10]. Higher temperatures also accelerate soil moisture evaporation, leading to a quicker depletion of soil water reserves. This is particularly detrimental during the drought periods, as proper growth and development of coffee plants rely on adequate soil moisture [76]. The temperature increase combined with drought creates a synergistic stress particularly on Arabica coffee, leading to reduced plant growth, and bean yield and quality. The impact is especially severe in regions already vulnerable to climate variability, such as Harar in Ethiopia, where temperatures often exceed the optimal range, leading to accelerated water loss and reduced coffee yields [30]. The temperature-induced drought effects are particularly pronounced at lower altitudes, where temperature extremes are more common [9]. In general, temperature significantly influences the impact of drought stress on coffee through its direct effects on soil moisture dynamics, evapotranspiration, and plant physiology. Understanding these interactions is crucial for developing effective management strategies and breeding programs to enhance the resilience of coffee (particularly that of Arabica) in the face of climate change and increasing drought incidences. It is also important to know that the effects of temperature-induced drought stress on coffee can be mitigated by using heat-tolerant varieties and some management approaches that can buffer temperature extremes [12], e.g., agroforestry systems and mulching or cover crops.
3.3.2. Sunshine, Relative Humidity and Wind
Sunshine, relative humidity (RH), and wind are critical factors influencing plant growth, yield, and quality of coffee. While adequate sunlight is necessary for plant growth and development as it is vital for photosynthesis activity of plants, excessive sunlight, especially when combined with drought conditions can lead to increased water stress that negatively impacts plant growth, yield and quality of coffee. Coffee farms with a high sunshine exposure often face challenges related to excessive ET. This is particularly problematic during the dry season when the RH drops further increasing the crop’s demand for water and leading to pronounced drought stress [77]. Wind can also influence the microclimate around coffee plants. While moderate winds can aid in cooling and drying, excessive winds can increase the ET rate, which can in turn intensifies a drought condition, particularly in drought prone areas [78] and drought stress in coffee plants [66].
The interaction between sunshine and RH is also a significant determinant of drought stress in Arabica coffee [75]. Sunshine affects both photosynthesis and ET while RH influences the ET rate and water vapor in the atmosphere [79]. A low RH increases ET rate, enhancing the water needs of the crops and consequently, the risk of water stress conditions due to a lack of easily accessible water in the soil profile explored by roots. Therefore, a combination of a high sunshine and a low RH typically exacerbates drought stress by increasing water loss both from soils and plants [79,80]. The combination of an intense sunlight and a low humidity can also lead to leaf scorch and reduced photosynthetic efficiency, ultimately impacting coffee yield and quality [30]. This shows that understanding the balance between sunlight, atmospheric humidity, water availability, and coffee physiology is crucial for optimizing the cultivation practices, particularly in the face of climate variability and increasing drought occurrences [10]. It also essential to know the potential interaction effects between temperature, rainfall pattern, RH, soil moisture content, ET rate, and wind condition on severity of drought impacts on coffee production for developing and implementing effective management strategies that can enhance the resilience of coffee plants to drought stress. Implementing agricultural practices, such as agroforestry system, windbreak, and irrigation can help to mitigate the adverse effects of high solar radiation, strong winds, and drought on coffee plants.
3.4. Topographic and Edaphic Factors
The distribution and availability of soil moisture in coffee agro-ecosystems are not uniform but are critically shaped by the interplay of topographic relief and soil characteristics. These factors create a complex mosaic of micro-climates, which can either ameliorate or exacerbate drought stress, determining the success of coffee cultivation in hilly and mountainous regions.
3.4.1. Elevation
Elevation is a fundamental topographic factor that influences climate and soil property of the area. As elevation increases, temperature decreases which means the effect of drought stress on plants can decrease with elevation due to a reduction in ET rate with increasing elevation [33]. Higher elevations provide a cooler microclimate, allowing Arabica coffee plants to maintain a better water status during the periods of water deficit and a slower growth rate of berries, which can also enhance bean quality [10]. Furthermore, the highland areas provide a more favorable microclimate for Arabica coffee cultivation as compared to the lowland areas. This may imply that moving coffee cultivation from climatically unsuitable areas (lowlands) to more suitable areas (highlands) (if opportunities exist) and developing cultivars with broad adaptability to a range of elevations can be helpful for ensuring resilience to drought stress [81].
3.4.2. Slope, Aspect and Curvature of the Land
Topographic factors, such as slope, aspect (slope orientation) and land curvature are key determinants of this micro-climate, particularly in mountainous or hilly catchments [82,83,84,85,86,87,88,89,90]. These factors influence the distribution of sunlight, water runoff, and soil erosion in coffee-growing areas [91]. Although specific studies directly comparing the effects of different slopes and slope orientations (aspects) on drought stress in coffee cultivation appear to be lacking, some relevant indirect studies and reviews [91,92] provide broader insights into the impacts of slope and aspect on drought severity. The steepness of a slope is a primary determinant of water drainage dynamics. On steeper gradients, gravitational forces dominate, leading to increased surface runoff and reduced time for water infiltration. This results in lower soil moisture retention, directly intensifying drought stress for coffee plants [66]. In contrast, gentler slopes allow for greater water percolation, enhancing the soil’s moisture reservoir.
Aspect or the direction, the slope faces to sun affects the amount of sunlight and the heat load received [93]. While the traditional north- and south-facing aspect dichotomy is relevant in mid-latitudes, in equatorial regions, east- and west-facing slopes exhibit distinct micro-climatic regimes due to diurnal solar movement. East-facing slopes receive solar radiation during the cooler morning hours, which reduces evaporative loss while allowing effective warming, thus maintaining higher soil moisture and moderated temperature profiles. In contrast, west-facing slopes are exposed to more intense afternoon sun, leading to higher soil temperatures, greater evaporative demand, and drier conditions [92,94]. The west-facing slopes are favorably exposed for receiving more solar radiation, and the stored heat of soil warming facilitates a higher rate of soil moisture extraction by ET [88]. In general, an east-facing slope will experience a colder and wetter soil micro-climate, while a west-facing slope is commonly warmer and drier [86,87]. These variations are critical for crops like coffee, whose growth and productivity are highly sensitive to both soil moisture and temperature.
Slope position and curvature further control water redistribution. Lower slopes (toeslopes) accumulate moisture from lateral flow, while upper slopes are typically drier [90,95,96,97,98]. Concave slopes enhance water accumulation and infiltration, whereas convex slopes promote water shedding and rapid soil drying [88,98,99]. Consequently, depressions and valley bottoms have higher soil moisture, while ridges and hilltops experience more severe drought stress due to runoff and exposure [68]. This explains why in drought-prone areas of Ethiopia and Yemen, coffee is often cultivated in valley bottoms and terracing and contour planting are used to mitigate these effects on hillsides [30].
Overall, understanding the combined impact of these topographic features is essential for managing soil microclimate in coffee agro-ecosystems [89,94]. However, despite their recognized importance, specific studies directly linking slope, aspect, and curvature to drought stress effects in coffee are scarce [102]. Future research quantifying how these topographic factors modulate drought tolerance and productivity is needed.
3.4.3. Soil Type and Fertility Status
Different soil types exhibit varying capacities for moisture retention, which directly influence the impacts of drought stress on coffee. Sandy soils, characterized by their coarse texture, quickly drain water and may not retain adequate moisture during a drought period, leading to stress in coffee plants [103]. Conversely, clay soils have a higher moisture-holding capacity but can become compacted, potentially restricting root growth and water uptake from the deep soil layers [104]. Effective soil management can enhance moisture availability in these soils [105].
Soil fertility status, which is defined as the availability of essential nutrients, such as nitrogen, phosphorus, potassium, calcium, magnesium, sulfur and various micronutrients, also plays a critical role in enhancing the resilience of coffee plants to drought stress by promoting a robust root system that enables coffee plants to access the deeper moisture reserves [79,106]. In addition to improving root development, essential nutrients can also allow coffee plants to better cope with drought stress by improving physiological process and overall plant health. Furthermore, some nutrients, such as potassium, play a vital role in osmotic adjustment of plants that helps the plant to regulate stomatal opening and water loss [10]. Fertile soils rich in organic matter content could also enhance water-holding capacity of soils and physiological activity of plants, which is crucial for maintaining soil moisture and crop performance during the drought stress periods, respectively [107,108]. Researches indicate that coffee plants grown in nutrient-rich soils exhibit better physiological and growth performances (e.g., higher leaf area index and photosynthetic efficiency) under drought conditions [104,109]. Similarly, well-fertilized coffee plants tend to have a higher metabolic activity and protective compounds, which can improve their resistance to drought stress. However, drought conditions often reduce the availability of nutrients for plants, leading to nutrient deficiencies in plants that intensify drought stress [110]. Nutrient deficiencies can impair physiological processes, including photosynthesis and water use efficiency. For instance, nitrogen deficiency can lead to reduced chlorophyll content, negatively impacting the plant’s ability to perform photosynthesis effectively [10]. All in all, the results of several studies show that knowing the properties and fertilities of soils is essential to develop effective drought management methods tailored to specific soil types and fertility status.
However, one cannot understand a coffee plant’s water status by looking at just one of these factors. A coffee plant’s water status is an integrated outcome of the dynamic interplay between atmospheric demand, soil supply, and climatic context (Figure 2). Vapor Pressure Deficit (VPD) represents the atmospheric “pull” for water, forcing stomatal closure to conserve it at the cost of photosynthesis. Soil texture acts as the critical “buffer,” with sandy soils offering minimal plant-available water and clay-loam soils providing a vital reservoir [103]. Altitude functions as a key modulator, primarily by lowering temperature and thus VPD, creating a less stressful environment [33]. These factors are inseparable in their effect. A high VPD at low altitude can induce physiological stress even in moist soil, while the low VPD of high altitudes can partially compensate for poor soil water retention. The most severe stress and yield loss occur when high atmospheric demand coincides with low soil water supply—a “perfect storm” of high VPD and sandy soil. Consequently, effective adaptation strategies must be tailored to these interactions: in high-VPD, low-altitude regions, employing shade to reduce VPD and improving soil organic matter are critical, whereas at high altitudes, the priority shifts to maintaining soil health to leverage the naturally favorable climatic conditions.
3.5. Management Factors
Implementing sustainable management practices is crucial for enhancing resilience against drought stress, which threatens coffee production globally [111]. The resilience of coffee plants to drought stress can be significantly enhanced through effective management practices [112]. For instance, managing well the coffee farms by providing shade, improving soil fertility, and implementing soil and water conservation measures can enhance drought resilience as compared to unmanaged ones [113]. Employing specific nutrient management strategies, such as the use of biochar and growth-promoting bacteria, can enhance drought tolerance in coffee plants [114]. The choice of the optimal times for the execution of the different field activities (e.g., the times for soil cultivation, mulching, watering and weeding) is another crucial factor to limit the effects of drought stress. The adoption of effective management practices, including precision farming, can also mitigate the drought stress impacts on coffee. Such techniques address the micro-scale variability and optimize the resources, which enhance drought resilience and productivity of coffee under the water-limited conditions [10,115]. However, some of these management approaches can be costly for smallholder farmers, and calls for national and international supports.
4. Adaptation and Coping Measures to Drought Stress
Comprehensive adaptation and coping strategies are necessary to address the challenges of drought stress. Establishing robust breeding programs and developing drought-resistant coffee varieties, developing and applying innovative irrigation techniques, using soil and water conservation practices, using agroforestry systems, improving soil health through proper organic amendment and fertilization, implementing integrated pest and disease management practices, providing targeted training and extension services for farmers and institutional support and policy intervention can be essential components of a comprehensive approach to enhance drought resilience in coffee farming systems. Efforts towards solving the drought problem in plant production are primarily based on developing tolerant varieties and irrigation. On top of that, rational agricultural practices have been recognized worldwide as another basis for plant production planning in drought-prone regions [116,117]. For example, agroforestry systems reduces water loss and improves microclimates in the coffee agro-systems; organic amendments and soil conservation practices enhances soil health and water-holding capacity; and integrated pest and disease management strategies that account for the vulnerability of drought-stressed plants to pests and diseases improves plant health, all support the resilience of coffee plants to drought stress [118]. Institutional support and targeted training and extension services that can provide farmers with knowledge and tools they need to adapt to the changing climate also helps to combat the impacts of drought stress on coffee [119].
4.1. Breeding and Cultivar Selection
Breeding and drought-tolerant cultivar selection is a vital, long-term strategy for combating drought stress. Breeders have developed some drought-tolerant cultivars of Arabica and Robusta coffees. The research strongly indicates that different coffee cultivars possess distinct tolerances to environmental stresses [120]. The recent F1 Arabica hybrids (e.g., ‘Acaiá Cerrado’ derivatives) combine plant vigor with some drought tolerance and a Robusta clonal selection has produced strains like Conilon 120 with deeper roots and better WUE. Various Arabica coffee cultivars that exhibit superior drought resistance were also identified, e.g., Acauã [121], Siriema [45], Catuai SH3 [122], IPR 103, and SL28 [81]. The drought resistance of these cultivars is largely attributed to traits, such as root architecture [23,29,31], leaf morphology [28], stomatal behavior [123], and osmotic adjustment [124] (Table2). This varietal difference pinpoints the importance of drought resistance breeding programs of coffee for a sustainable coffee production under increasing the drought incidence conditions and in drought-prone coffee-growing regions of the world.
Moving beyond conventional methods, modern breeding is increasingly leveraging molecular tools to accelerate progress. Molecular breeding and marker-assisted selection (MAS) are now being employed to identify and select for key drought-resilience alleles more efficiently. For instance, quantitative trait loci (QTLs) and candidate genes associated with root depth (e.g., CaMYB20), water-use efficiency (e.g., CaERF1A), and osmotic adjustment (e.g., CaDHN1) have been identified, providing valuable targets for selection [125,126]. Furthermore, the emergence of genome editing, particularly CRISPR-Cas technology, holds transformative potential. While still in its early stages for perennial crops like coffee, proof-of-concept studies have successfully edited genes involved in caffeine biosynthesis and plant architecture. Targeted editing of genes regulating stomatal density (e.g., SDD1), abscisic acid (ABA) responsiveness (e.g., CaNCED3), and the accumulation of protective osmolytes like trehalose represents a promising frontier for the precise engineering of drought tolerance [127,128]. This varietal difference, now exploitable through both conventional and molecular means, pinpoints the importance of drought resistance breeding programs for a sustainable coffee production under increasing drought incidence in drought-prone coffee-growing regions of the world.
Table 2.
Drought-tolerant varieties of Arabica and Robusta coffee, along with their key traits.
| Variety/Clone (Species) | Key Drought-Resistance Traits | References |
|---|---|---|
| Apoatã IAC 2258 (C. canephora) | Deep & Prolific Root System (Primary trait) | [23,120] |
| Sarchimor (e.g., T5296, Costa Rica 95) (C. arabica) | Stomatal Regulation (Early closure) Osmotic Adjustment (Accumulation of solutes) Compact Structure (Lower leaf area) |
[12,129,135] |
| Icatu (C. arabica) | Leaf Traits (Thicker leaves, waxier cuticles) Robust Root System (From Robusta parentage) Control of oxidative stress |
[41,129,130] |
| Caturra | Stomatal Regulation (Early closure) High ware use efficiency |
[129,131,132] |
| Castillo / Centroamericano (C. arabica) | Osmotic Adjustment (Primary trait) Stomatal Regulation |
[131] |
| IAPAR 59 (C. arabica) | Vigorous Root System (From Robusta parentage) Leaf Shedding Mechanism (Reduces transpirational area) |
[68,133] |
| F1 Hybrids (e.g., Starmaya) (C. arabica) | Hydraulic Conductance (Efficient water transport - from heterosis) Robust Root System (From heterosis) |
[134,135] |
| F1 Hybrids (e.g., Mundo Maya) | Maintenance of photosynthesis Enhanced root density |
[134,135] |
4.2. Preconditioning
Preconditioning refers to agronomic practices designed to prepare the coffee plants for adverse environmental conditions, such as drought or high temperature. Preconditioning is a proactive approach for managing drought stress by enhancing the plant’s drought tolerance mechanisms. It represents a shift from reactive to preventive management, allowing plants to better cope with water deficits [136]. Under natural condition, repeated drought recovery cycles are far more common than a single (prolonged) drought event. By “training” the plants to endure water scarcity using a priori-exposure, it is possible to improve the physiological resistance of plants to drought stress. Plants submitted to multiple drought cycles are able to develop an acclimation which might help them to tolerate drought stress better than with plants suffering from drought stress for the first time [137,138,139]. This kind of differential acclimation to stress involves not only the expression of trainable genes related to drought tolerance [139], but also associates with an orchestrated reprogramming of metabolic and physiological processes. In this sense, plants that are previously exposed to a certain stress can develop a kind of acclimation, sometimes called stress memory, which would potentiate their defense responses in order to prime them to a subsequent exposure to the same stress [137,140,141].
Nevertheless if not managed properly, preconditioning can lead to reduced growth and yield, particularly if the stress is applied during the critical growth stages of the crop. Moreover, the effectiveness of preconditioning may vary with the time of the stress applied, severity of the stress applied, and the specific crop variety used. Therefore, integrating preconditioning in to the coffee farming systems could significantly reduce the impact of drought stress, although it requires careful monitoring and management to avoid its adverse effects on plant growth and yield. However, even if this strategy is being used in Robusta coffee [139] and in many other crops, responses of Arabica coffee to the consecutive cycles of drought stress and re-watering (recovery) treatments have scarcely been studied and it is difficult to extrapolate the information from Robusta and other crops for Arabica coffee. Therefore, it is necessary to investigate whether cyclic drought treatment could improve drought resistance in Arabica coffee or not.
4.3. Plant Nutrition
There are 17 elements which are currently considered essential for the plants to complete their life cycle [142]. These plant nutrients are not only required for better plant growth and development, but also for alleviating different abiotic stresses, such as drought stress. Evidences show that the mineral-nutrient status of plants plays a critical role in increasing plant resistance to environmental stresses [143] and inorganic fertilization mitigates the adverse effects of drought stress on crop growth and development [143,144,145]. This means that tailored adaptive strategies based on plant nutrition or soil fertility can increase the resilience capacity of coffee trees to drought stress.
Different essential mineral nutrients play various roles in alleviation of drought stress in plants. Nitrogen, for example, affects carbon partitioning and improves the accumulation of soluble sugars and starch, which in turn improves leaf growth [146,147]. Nitrogen fertilizer application, where light is not limiting, increases the plant’s antioxidative defense mechanisms [143] that results in reduced photooxidation of chloroplast pigments and leaf senescence. Nitrogen applied as fertilizers or in other forms is also closely related to the ability of plant roots to absorb water from the soil. As per Waraich et al. [142], improving water use efficiency is the possible mechanism of nitrogen to minimize the detrimental effects of drought stress on plants.
Turner, [148] pointed out that phosphorus (P) deficiency appears to be one of the earliest effects of mild to moderate drought stress in soil-grown plants and the application of P fertilizer considerably improves plant growth under drought conditions [149,150,151]. The positive effects of P on plant growth under drought have been attributed to an increase in stomatal conductance [152], photosynthetic rate [150], cell-membrane stability, and water relations [153]. Phosphorus also improves the root growth and maintains high leaf water potential. This improves the water and nutrient uptakes and the activity of nitrate reductase that in turn improves the assimilation of nitrate under drought condition [154]. Phosphorus also maintains cell turgidity by maintaining high leaf water potential which in turn increases the stomatal conductance and photosynthetic rate under drought condition.
Potassium (K) plays a key role for the survival of plants under environmental stress and plants suffering from environmental stress like drought have a large internal requirement for K [155]. Environmental stresses that enhance the requirement for K cause oxidative damage to cells by inducing ROS formation, especially during photosynthesis [156,157,158]. The reason for this appears to be related to the fact that K is required for the maintenance of photosynthetic CO2 fixation. For example, drought stress is associated with stomatal closure and thereby with decreased CO2 fixation. When plants are grown under a low K supply, drought-stress induced ROS production can be additionally enhanced, at least due to K-deficiency-induced disturbances in stomatal opening, water relation, and photosynthesis [143,159]. Decrease in photosynthesis caused by drought stress is particularly high in plants supplied with low K and minimal in plants supplied with sufficient K [160]. Moreover, alleviation of detrimental effects of drought stress, especially on photosynthesis, by sufficient K supply has also been shown in legume crops [161]. According to Waraich et al. [142], K minimizes the detrimental effect of drought on crop plants by improving water use efficiency of plants. K also improves the root growth that in turn improves water and nutrient uptakes and leaf water potential under drought condition [162]. Therefore, plant nutrition with K in drought condition increases the tolerance of plants to drought stress by utilizing the available soil moisture more efficiently than in K-deficient plants. However, as over-fertilization under drought stress condition can be counterproductive, a caution is needed when we use fertilization as means of drought stress management; for example, over-fertilization especially nitrogen can produce excess foliage that increases the water demand of plants. Thus, a balanced fertilization should be considered for drought management. However, evidence on the role of plant nutrition for drought stress alleviation in coffee plants is limited. This calls for studies on the drought alleviation roles of specific nutrients and fertilization.
4.4. Irrigation
Deficit irrigation is a water-saving strategy that involves applying less water than the crop’s full requirement during certain growth stages of the crop. This method can effectively reduce water use while maintaining acceptable yield levels [110]. By coinciding the irrigation time with critical growth periods of coffee, such as vegetative growth, flowering, and fruit setting, farmers can optimize water resources and improve coffee resilience to drought stress. In addition, accurate irrigation scheduling based on allowable soil moisture depletion is essential for maximizing water use efficiency in coffee cultivation. A study at Gera, southwest Ethiopia has shown that applying irrigation when the soil moisture reaches 40% depletion significantly enhances coffee yield compared to over-irrigation or under-irrigation practices [163]. This shows that implementing a precise irrigation schedule allows to use water resources and to reduce the impact of drought stress in a better way than other irrigation systems.
4.5. Soil and Water Conservation
Soil and water conservation practices are essential for preventing soil erosion, enhancing water infiltration, and maintaining soil fertility, particularly in hilly and drought-prone areas, e.g., some coffee growing-regions in Ethiopia, such as Harar. Techniques, such as contour plowing (planting), terracing, ridges and the construction of check dams are commonly used to reduce water runoff and conserve soil moisture, thereby improving the resilience of agricultural systems to drought. Terracing is widely practiced to slow down water runoff and increase water infiltration into the soil (Figure 3). In addition, check dams and contour plowing help to retain soil moisture and reduce the loss of the fertile topsoils during heavy rains, which is particularly important in maintaining soil health and productivity under drought condition [118]. These techniques are commonly used in some coffee growing countries like Yemen [164], mulching that involves covering of the soil surface with organic matter (i.e. pruned branches and leaves from coffee trees or other trees) or inorganic materials (i.e., plastic mulch) is another common practice applied to conserve soil moisture, suppress weeds, and enhance soil conditions [165]. Cover crops are also grown to protect the soils from erosion and drying and to improve soil properties and increase soil organic matter. In some Ethiopian coffee farms, mulching with coffee husks, leaves, and straw is practiced to maintain soil moisture and improve soil structure. The use of leguminous cover crops, such as Desmodium, Broad bean and Crotalaria spp., is also widespread, particularly in large coffee farms in the southern and southwestern regions of Ethiopia. These cover crops not only fix atmospheric nitrogen, thereby enriching the soil with nitrogen, but also help to retain soil moisture during the dry season [30]. However, there can be a competition between cover crops and coffee plants for water and nutrients. So, identification of cover crop species that are less competitive with coffee plants is needed to address these likelihood challenges related to cover crops. In anyway, integrating soil and soil moisture conservation practices including cover crops in to the coffee farming systems can maintain the productivity of coffee plants under water-limited conditions by enhancing the natural resource base. However, the successful implementation of these practices requires adequate technical support and community engagement [30].
4.6. Agroforestry and Shade Management
Coffee is normally an understory plant and deliberate shading can be an important drought mitigation strategy for coffee. Shade trees in a coffee agroforestry system modify the microclimate by reducing solar radiation, buffering the air temperature fluctuations (reducing temperature extremes), increasing humidity, and slow down the wind speed, all of which enhance soil moisture retention and lower the plant water demand and the severity of drought stress. In addition, shade trees improve soil properties through litter falls and root turnovers, which raise the organic matter content of soils and nutrient cycling, further helping soil moisture retention. A review by Koutouleas et al. [70] shows that shade trees produce “favorable microclimate modifications” by lowering the variability in air temperature and increasing relative humidity and wind protection. This review work also shows that agroforestry protected coffee plants from frost events in Vietnam. Similarly, shaded coffee farms in Ethiopia tend to have higher soil moisture levels and are less affected by drought compared to non-shaded farms [166].
Despite all these general thoughts and research findings on the importance of shade trees for drought stress alleviation, the existing empirical evidences are ambiguous and conflicting. There is a potential trade-off between the positive role of shade in alleviating the detrimental effects of drought stress by modifying the microclimate [167] and its negative effects through increasing the crop’s susceptibility to drought stress by reducing resource allocation to roots which in turn reduces root elongation for deep water absorption [46]. Besides, there could be a competition between shade trees and coffee plants for water and nutrients. It is hypothesized that drought may be more harmful to plants under low light or under shade because of the trade-off between the simultaneous requirements to allocate biomass in response to resource limitations both above and below the soil surface. Under water-deficit condition, there will also be an intense competition between shade trees and coffee plants for water and nutrients. The mixed results of the past studies could be due to the differences in shade tree type and shade level. A potential solution is, therefore, the use of a dynamic shade system, where the level of shade is adjusted based on seasonal and climatic conditions. Also, the choice of shade tree species, planting density, and pruning practices are all critical factors that need to be carefully managed to maximize the benefits of shade in alleviating the detrimental effects of drought stress on coffee while minimizing its potential drawbacks [168]. However, these contradictions call for studies that examine when, how and under which condition shade (agroforestry system) alleviates the impacts of drought stress on coffee.
4.7. Crop Management and Diversification
Improved crop management practices (e.g., regular pruning and effective disease and pest control) are equally important in sustaining coffee production under drought stress. Regular pruning of coffee plants helps maintain an optimal canopy size, reducing water demand and improving air circulation, which can lower the incidence of pests and diseases. Effective disease and pest management is also crucial, as drought-stressed plants are more susceptible to pest and disease attacks. Moreover, a diversified farm is inherently more resilient to environmental stress as engaging farmers in different farming activities enable them to respond more effectively to environmental challenges. Hence, crop diversification by integrating multiple crops with coffee could be a good economic strategy for farmers to reduce the economic risks associated with coffee farming under drought conditions.
4.8. Training and Extension Services
Giving targeted training and extension services to farmers are vital for disseminating knowledge and skills necessary for adapting to drought conditions. Many smallholder coffee farmers, particularly in the undeveloped countries, lack access to up-to-date information on climate-smart agricultural practices. This gap can be filled by implementing the well-programmed and participatory training and extension services on good agricultural practices that can alleviate the impact of drought stress on coffee, such as the use of drought-tolerant varieties and improved agronomic practices, e.g., pruning, shade, water and nutrient management, and soil and water conservation. These programs are most effective when they are participatory, allowing farmers to share their experiences and adapt the new knowledge and practices to their local contexts.
4.9. Policy Development and Institutional Support
Policy development and institutional support are crucial for creating an enabling environment for the adoption of adaptive strategies in coffee production. Governments play a pivotal role in shaping policies that support sustainable agricultural practices, including providing subsidies for drought-resistant varieties, financing soil and water conservation projects, and offering incentives for the adoption of climate-smart technologies. Policy frameworks that promote research and development in drought-resistant coffee varieties and innovative farming techniques can also drive progress.
In addition to national policies, international collaborations are essential in addressing the global challenges of coffee production posed by climate change. Initiatives like the Global Coffee Platform bring together stakeholders across the coffee value chain to develop and implement strategies that can enhance the resilience of coffee production to drought stress caused by climate change. Research and development are also critical in advancing our understanding of drought stress in coffee and in developing new technologies and practices that can mitigate its impact. Investments on research and development can lead to a breakthrough in biotechnology, water management, and climate modeling, which can provide farmers with tools they need to adapt to the changing climates [169]. While the climate-smart practices show a promise in enhancing coffee resilience to climate change, challenges remain in their widespread adoption, necessitating further research and supportive policies to ensure their effective implementation.
5. Conclusion
Drought poses a severe and growing threat to global coffee production, with far-reaching agronomic and economic consequences. This review paper highlights the impacts of drought stress on coffee physiology, growth, yield, and quality, which greatly vary with the genetics of coffee plants, and soil, climatic and farming contexts. It also underscores the potential of coffee breeding and drought-tolerant variety selection and agricultural practices (e.g., irrigation, soil and water conservation, shade, and balanced plant nutrition) and the importance of multidisciplinary, multi-sectorial and national and intentional efforts to sustain coffee production in drought condition. While this review has synthesized the current understanding of drought impacts and adaptations strategies in coffee, it has also revealed critical knowledge gaps in current research and proposes some research directions for future studies that are essential for developing effective drought management strategies for coffee production in a changing climate. Addressing these gaps requires future research efforts that employ integrative, systems-level approaches to overcome these constraints.
Current research has successfully identified key drought-tolerant coffee genotypes—such as the deep-rooted Robusta clone Apoatã IAC 2258, the osmotically adjusting Arabica varieties Sarchimor and Castillo, and the robust Icatu hybrids—each employing distinct physiological strategies from improved stomatal regulation and leaf traits to enhanced hydraulic conductance. However, this very diversity of mechanisms reveals critical research frontiers. Future efforts must pivot from singular trait identification to the integrated, field-based validation of trait combinations, defining the ideal “drought resilience ideotype” for specific growing environments. This requires leveraging high-throughput phenotyping and remote sensing to develop scalable drought indices, while genomics and predictive modeling must untangle complex Genotype-by-Environment (GxE) interactions. Furthermore, pressing questions remain regarding the synergistic role of shade trees and nutrient management in drought alleviation, as their interactions with coffee root and shoot physiology can be contradictory. Finally, unlocking the potential of drought “pre-conditioning” or hardening in C. arabica demands a deeper investigation into the underlying stress memory and epigenetic regulations that could prime plants for enhanced resilience.
This review concludes with several integrated recommendations to bolster the resilience of coffee production systems. First, it underscores the necessity for continuous breeding programs focused on developing cultivars that synergize drought tolerance with high yield and superior cup quality. Complementing these genetic efforts, the implementation of long-term agronomic field trials is critical to refine and integrate Good Agricultural Practices—including optimized irrigation, soil conservation, agroforestry, and nutrient management—into tailored packages for specific regional contexts. Furthermore, leveraging climate forecasts and early-warning systems can empower farmers to strategically time interventions such as irrigation and fertilization. The effective adoption of these strategies hinges on robust training and extension services to disseminate knowledge directly to growers. Finally, beyond on-farm measures, developing a coherent framework for policy interventions and institutional, sectoral, and governmental cooperation is essential to holistically address the multi-faceted challenge of drought resilience in coffee systems.
Author Contributions
Conceptualization, W.M and N.K.A; investigation, W.M and N.K.A writing—original draft, N.K.A.; writing—review and editing, W.M. and N.K.A.; visualization, W.M. and N.K.A.; supervision, W.M.Both authors have read and agreed to the published version of the manuscript.
Funding
Not applicable.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Davis, AP.; Gole, TW.; Baena, S.; Moat, J. The impact of climate change on indigenous Arabica coffee (Coffea arabica L.): predicting future trends and identifying priorities. PloS one 2012, 7(11), e47981. [Google Scholar] [CrossRef]
- DaMatta, FM.; Ronchi, CP.; Maestri, M.; Barros, RS. Coffee: Environment and Crop Physiology. In Ecophysiology of Tropical Tree Crops; DaMatta, F.M., Ed.; Nova Science Publishers: New York, NY, USA, 2010; pp. 181–216. [Google Scholar]
- ICO. World Coffee Production. 2023. Available online: www.ico.org (Accessed: 15 June 2024).
- DaMatta, FM.; Rahn, E; Läderach, P; Ghini, R; Ramalho, JC. Why could the coffee crop endure climate change and global warming to a greater extent than previously estimated? Climatic Change 2019, 152, 167–178. [Google Scholar] [CrossRef]
- Semedo, JN.; Rodrigues, AP.; Lidon, FC.; Pais, IP.; Marques, I.; Gouveia, D.; Armengaud, J.; Silva, MJ.; Martins, S.; Semedo, MC.; Dubberstein, D.; Partelli, FL.; Reboredo, FH.; Scotti-Campos, P.; Ribeiro-Barros, AI.; DaMatta, FM.; Ramalho, JC. Intrinsic non-stomatal resilience to drought of the photosynthetic apparatus in Coffea spp. is strengthened by elevated air [CO2]. Tree Physiology 2021, 41, 708–727. [Google Scholar] [CrossRef] [PubMed]
- ECTA, Annual Report 2023.
- Worku, M. Production, productivity, quality and chemical composition of Ethiopian coffee. Cogent Food and Agriculture 2023, 9, 2196868. [Google Scholar] [CrossRef]
- Larcher, W. Photosynthesis as a tool for indicating temperature stress events. In Ecophysiology of photosynthesis; Springer Berlin Heidelberg: Berlin, Heidelberg, 1995; pp. 261–277. [Google Scholar]
- Moat, J.; Williams, J.; Baena, S.; Wilkinson, T.; Demissew, S.; Challa, ZK.; Davis, AP. Resilience potential of the Ethiopian coffee sector under climate change. Nature Plants 2017, 3(4), 17081. [Google Scholar] [CrossRef] [PubMed]
- DaMatta, FM.; Ramalho, JDC. Impacts of drought and temperature stress on coffee physiology and production: a review. Brazilian Journal of Plant Physiology 2006, 18, 55–81. [Google Scholar] [CrossRef]
- Peruta, R.D.; Mereu, V.; Spano, D.; Marras, S.; Vezy, R.; Trabucco, A. Projecting trends of arabica coffee yield under climate change: A process-based modelling study at continental scale. Agricultural Systems 2025, 104353. [Google Scholar] [CrossRef]
- DaMatta, FM.; Avila, RT.; Cardoso, AA.; Martins, SC.; Ramalho, JC. Physiological and agronomic performance of the coffee crop in the context of climate change and global warming: A review. J. Agric Food Chem.;PubMed 2018, 66, 5264–5274. [Google Scholar] [CrossRef]
- Bilen, C.; El Chami, D.; Mereu, V.; Trabucco, A.; Marras, S.; Spano, D. A Systematic Review on Impacts of Climate Change on Coffee Agroecosystems. Plants 2023, 12, 102. [Google Scholar]
- Sánchez-Reinoso, AD.; Ávila-Pedraza, EÁ.; Lombardini, L.; Restrepo-Díaz, H. The application of coffee pulp biochar improves the physical, chemical, and biological characteristics of soil for coffee cultivation. Journal of Soil Science and Plant Nutrition 2023, 23(2), 2512–2524. [Google Scholar] [CrossRef]
- Chekol, H.; Warkineh, B.; Shimber, T.; Mierek-Adamska, A.; Dąbrowska, G.B.; Degu, A. Drought stress responses in Arabica coffee genotypes: Physiological and metabolic insights. Plants 2024, 13(6), 828. [Google Scholar] [CrossRef]
- Jaleel, CA.; Manivannan, P.; Wang, HZ. Water-deficit stress effects on plant growth, photosynthesis, and lipid peroxidation in coffee. Plant Physiology and Biochemistry 2009, 47(2), 127–133. [Google Scholar]
- Mingchi, Z.; Lin, H.; Zhao, F. The impact of drought on coffee growth and yield. Plant Growth Regulation 2010, 60(1), 27–35. [Google Scholar]
- Din, J.; Khan, SU.; Ali, I.; Gurmani, AR. Physiological and agronomic response of canola varieties to drought stress. The Journal of Animal and Plant Sciences 2011, 21(1), 78–82. [Google Scholar]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends in Plant Science 2002, 7(9), 405–410. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, M.E.A. ‘Photosynthesis and photoinhibition’. In Plant Responses to Air Pollution; 2008; pp. 129–152. [Google Scholar]
- Petropoulos, SA.; Sari, FM.; Papadopoulos, IA. Dry matter partitioning and stress adaptation in coffee plants. Journal of Agronomy and Crop Science 2008, 194(3), 189–197. [Google Scholar]
- Wu, QS.; Xia, RX.; Zou, YN. Improved soil structure and citrus growth after inoculation with three arbuscular mycorrhizal fungi under drought stress. European Journal of Soil Biology 2008, 44(1), 122–128. [Google Scholar] [CrossRef]
- Pinheiro, HA.; DaMatta, FM.; Chaves, ARM.; Loureiro, ME; Ducatti, C. Drought tolerance is associated with rooting depth and stomatal control of water use in clones of Coffea canephora. Annals of Botany 2005, 96, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Builes, VH.; Küsters, J.; Thiele, E.; Lopez-Ruiz, JC. Physiological and Agronomical Response of Coffee to Different Nitrogen Forms with and without Water Stress. Plants 2024, 13(10), 1387. [Google Scholar] [CrossRef]
- Venkataramanan, D; Ramaiah, PK. Osmotic adjustments under water stress in coffee. Proceeding of the12th Interna-tional Scientific Colloquium on coffee (ASIC) conferences, Montreux, Switzerland, 1987; pp. 493–499. [Google Scholar]
- Meinzer, FC; Saliendra, NZ; Crisosto, CH. Carbon iso-tope discrimination and gas exchange in Coffea arabica during adjustment to different soil moisture regimes. Australian J. Plant. Physiol. 1992, 19, 171–184. [Google Scholar] [CrossRef]
- DaMatta, F.M. Exploring drought tolerance in coffee: A physiological approach with some insights for plant breeding. Brazil Journal of Plant Physiology 2004, 16, 1–6. [Google Scholar] [CrossRef]
- Worku, M.; Astatkie, T. Dry matter partitioning and physiological responses of Coffea arabica varieties to soil moisture deficit stress at the seedling stage in Southwest Ethiopia. African Journal of Agricultural Research 2010a, 5(15), 2066–2072. [Google Scholar]
- Worku, M.; Astatkie, T. Growth responses of arabica coffee (Coffea arabica L.) varieties to soil moisture deficit at the seedling stage at Jimma, Southwest Ethiopia. Journal of Food, Agriculture and Environment 2010b, 8(1), 195–200. [Google Scholar]
- Tesfaye, M.; Ismail, S.; Asfaw, Z. Coffee production constraints and opportunities in Ethiopia. African Journal of Agricultural Research 2013, 8(50), 6674–6681. [Google Scholar]
- Aman, M.; Worku, M.; Shimbir, T.; Astatkie, T. Root traits and biomass production of drought-resistant and drought-sensitive Arabica coffee varieties growing under contrasting watering regimes. Agrosystems, Geosciences & Environment 2024, 7(2), p.e20488. no doi?? [Google Scholar]
- Bunn, C.; Läderach, P.; Ovalle Rivera, O.; Kirschke, D. A bitter cup: climate change profile of global production of Arabica and Robusta coffee. Climatic change 2015, 129(1), 89–101. [Google Scholar] [CrossRef]
- Jaramillo, J.; Muchugu, E.; Vega, FE.; Davis, A.; Borgemeister, C.; Chabi-Olaye, A. Some like it hot: the influence and implications of climate change on coffee berry borer (Hypothenemus hampei) and coffee production in East Africa. PLoS one 2013, 8(11), e88867. [Google Scholar] [CrossRef]
- Carr, MKV. The water relations and irrigation requirements of coffee. Experimental agriculture 2001, 37(1), 1–36. [Google Scholar] [CrossRef]
- Gay, C.; Estrada, F.; Conde, C.; Eakin, H.; Villers, L. Potential impacts of climate change on agriculture: A case of study of coffee production in Veracruz, Mexico. Climatic Change 2006, 79(3-4), 259–288. [Google Scholar] [CrossRef]
- Lin, CW. Effects of water stress on coffee bean quality. Journal of Coffee Science 2007, 9(1), 31–42. [Google Scholar]
- Damayanti, MH.; Rachman, MS. Drought Adaptation and Coping Strategies among Coffee Farmers in the Central Highlands of Vietnam. ResearchGate. 2019. Available online: https://www.researchgate.net/.
- Nuryanti, S.; Siregar, N.; Darusman, D.; Sudarsono, S. Impact of Climate Change on Coffee Quality and Yield. East African Scholars Journal of Agriculture and Veterinary Sciences 2021, 3(11), 1–8. Available online: https://www.easpublisher.com/get-articles/4388.
- Carmo-Silva, E.; Scales, JC.; Parker, P. The effects of drought on coffee plant physiology. Journal of Experimental Botany 2017, 68(16), 4623–4634. [Google Scholar]
- Flexas, J.; Bota, J.; Escalona, J. M. Effects of drought on photosynthesis in coffee plants. Journal of Experimental Botany 2006, 57(15), 3713–3722. [Google Scholar]
- Ramalho, JC.; Rodrigues, AP.; Lidon, FC.; Marques, LM.; Leitão, AE.; Fortunato, AS.; Pais, IP.; Silva, MJ.; Scotti-Campos, P.; Lopes, A.; Reboredo, FH. Stress cross-response of the antioxidative system promoted by superimposed drought and cold conditions in Coffea spp. PLoS one 2018, 13(6), e0198694. [Google Scholar] [CrossRef] [PubMed]
- Cruz de Carvalho, MH. Drought stress and reactive oxygen species: production, scavenging and signaling. Plant signaling & behavior 2008, 3(3), 156–165. [Google Scholar]
- Moller, IM.; Jensen, PE.; Hansson, A. Oxidative stress and antioxidants in plant cells. Annual Review of Plant Bi-ology 2007, 58, 161–185. [Google Scholar]
- Poorter, H.; Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review. Functional Plant Biology 2000, 27(12), 1191–1191. [Google Scholar]
- Dias, PC.; Araujo, WL.; Moraes, GABK.; Barros, RS.; DaMatta, FM. Morphological and Physiological Responses of Two Coffee Progenies to Soil Water Availability. J. Plant Physiol. 2007, 164, 1639–1647. [Google Scholar] [CrossRef]
- Canham, CD.; Berkowitz, AR.; Kelly, VR.; Lovett, GM.; Ollinger, SV.; Schnurr, J. Biomass allocation and multiple resource limitation in tree seedlings. Canadian Journal of Forest Research 1996, 26(9), 1521–1530. [Google Scholar] [CrossRef]
- Leport, L.; Turner, NC.; Davies, SL.; Siddique, KHM. Variation in pod production and abortion among chickpea cultivars under terminal drought. European Journal of Agronomy 2006, 24(3), 236–246. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, J.; Zhang, J. Effects of drought stress on root and shoot biomass in coffee plants. Plant Growth Regu-lation 2014, 73(2), 179–189. [Google Scholar]
- Komor, E. Source physiology and assimilate transport: the interaction of sucrose metabolism, starch storage and phloem export in source leaves and the effects on sugar status in phloem. Functional Plant Biology 2000, 27(6), 497–505. [Google Scholar] [CrossRef]
- Kim, SW.; Lee, SK.; Jeong, HJ.; An, G.; Jeon, JS.; Jung, KH. Crosstalk between diurnal rhythm and water stress reveals an altered primary carbon flux into soluble sugars in drought-treated rice leaves. Scientific Reports 2017, 7(1), 8214. [Google Scholar] [CrossRef]
- Zinselmeier, C.; Jeong, BR.; Boyer, JS. Starch and the control of kernel number in maize at low water potentials. Plant physiology 1999, 121(1), 25–36. [Google Scholar] [CrossRef]
- Tavares, JR.; Silva, CM.; de Almeida, RF. Climate change impacts on coffee yield and quality. Agricultural Meteorology 2018, 254, 60–72. [Google Scholar]
- Byrareddy, V.; Kouadio, L.; Mushtaq, S.; Kath, J.; Stone, R. Coping with drought: Lessons learned from Robusta coffee growers in Vietnam. Climate Services 2021, 22, 100229. [Google Scholar] [CrossRef]
- Liebig, TI. Abundance of pests and diseases in Arabica coffee production systems in Uganda-ecological mechanisms and spatial analysis in the face of climate change. Doctoral dissertation, 2017. [Google Scholar]
- Gadgil, M.; Rao, AN. Influence of drought on coffee pests and diseases. Tropical Agriculture 1998, 75(2), 81–90. [Google Scholar]
- Vázquez, S.; Gómez, M.; Fernández, A. Increased incidence of pests and diseases in drought-stressed coffee plants. Plant Pathology Journal 2018, 37(4), 213–224. [Google Scholar]
- Choudhary, A.; Senthil-Kumar, M. Drought: a context-dependent damper and aggravator of plant diseases. Plant, Cell & Environment 2024, 47(6), 2109–2126. [Google Scholar]
- Pham, Y.; Reardon-Smith, K.; Mushtaq, S.; Cockfield, G. The impact of climate change and variability on coffee production: a systematic review. Climatic Change 2019, 156, 609–630. [Google Scholar] [CrossRef]
- Magrach, A; Ghazoul, J. Climate and Pest-Driven Geographic Shifts in Global Coffee Production: Implications for Forest Cover, Biodiversity and Carbon Storage. PLoS One 2015, 15;10(7), e0133071. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ovalle-Rivera, O; Läderach, P; Bunn, C; Obersteiner, M; Schroth, G. Projected shifts in Coffea arabica suitability among major global producing regions due to climate change. PLoS One 2015, 14;10(4), e0124155. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Malhi, Y.; Roberts, J. T.; Rippel, C. The changing dynamics of coffee production in the context of climate change. Agricultural Systems 2017, 157, 141–150. [Google Scholar]
- Klaus, M.; Khoury, CK.; Andersson, M. Coffee cultivation shifts to higher altitudes in response to climate change. Agricultural Systems 2016, 144, 123–134. [Google Scholar]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Geneva, Switzerland, 2014. Available online: https://www.ipcc.ch/pdf/assessmentreport/ar5/syr/AR5_SYR_FINAL_SPM.pdf.
- Silva, PEM.; Cavatte, PC.; Morais, L.; Medina, EF.; DaMatta, FM. The functional divergence of biomass partitioning, carbon gain and water use in Coffea canephora in response to the water supply: Implications for breeding aimed at improving drought tolerance. Environ. Exp. Bot. 2013, 87, 49–57. [Google Scholar] [CrossRef]
- Chaves, SCL.; Soares, FF.; Rossi, TRA.; Cangussu, MCT.; Figueiredo, ACL.; Cruz, DN.; Cury, PR. Características do acesso e utilização de serviços odontológicos em municípios de médio porte. Ciência & Saúde Coletiva 2012, 17, 3115–3124. [Google Scholar]
- Martins, SC.; Galmes, J.; Cavatte, PC.; Pereira, LF.; Ventrella, MC.; DaMatta, FM. Understanding the low photo-synthetic rates of sun and shade coffee leaves: Bridging the gap on the relative roles of hydraulic, diffusive and biochemical constraints to photosynthesis. In PLoS ONE; PubMed, 2014; Volume 9. [Google Scholar]
- Rodrigues, W.P.; Martins, M.Q.; Fortunato, A.S.; Rodrigues, A.P.; Semedo, J.N.; Simões-Costa, M.C.; Pais, I.P.; Leitão, A.E.; Colwell, F.; Goulao, L.; Máguas, C. Long-term elevated air [CO2] strengthens photosynthetic functioning and mitigates the impact of supra-optimal temperatures in tropical Coffea arabica and C. canephora species. Global change biology 2016, 22(1), 415–431. [Google Scholar] [CrossRef]
- Silva, PCD.; Junior, WQR.; Ramos, MLG.; Rocha, OC.; Veiga, AD.; Silva, NH.; Brasileiro, LDO.; Santana, CC.; Soares, GF.; Malaquias, JV.; Vinson, CC. Physiological changes of Arabica coffee under different intensities and durations of water stress in the Brazilian Cerrado. Plants 2022, 11(17), 2198. [Google Scholar] [CrossRef] [PubMed]
- Avila, RT.; Cardoso, AA.; de Almeida, WL.; Costa, LC.; Machado, KL.; Barbosa, ML.; de Souza, RP.; Oliveira, LA.; Batista, DS.; Martins, SC.; Ramalho, JD. Coffee plants respond to drought and elevated [CO2] through changes in stomatal function, plant hydraulic conductance, and aquaporin expression. Environmental and Experimental Botany 2020, 177, 104148. [Google Scholar] [CrossRef]
- Koutouleas, A.; Sarzynski, T.; Bordeaux, M.; Bosselmann, AS.; Campa, C.; Etienne, H.; Turreira-García, N.; Rigal, C.; Vaast, P.; Ramalho, JC.; Marraccini, P. Shaded-coffee: A nature-based strategy for coffee production under climate change? A review. Frontiers in Sustainable Food Systems 2022, 6, 877476. [Google Scholar] [CrossRef]
- Salehi-Lisar, SY.; Bakhshayeshan-Agdam, H. Drought stress in plants: causes, consequences, and tolerance. In Drought stress tolerance in plants;physiology and biochemistry; Springer International Publishing: Cham, 2016; Vol 1, pp. 1–16. [Google Scholar]
- McKersie, BD.; Leshem, YY. Water and drought stress. In Stress and stress coping in cultivated plants; Kiuwer Academic Publishers: Dordrecht, Netherlands, 1994; pp. pp 148–180. [Google Scholar]
- López, J.; Way, D.A.; Sadok, W. Systemic effects of rising atmospheric vapor pressure deficit on plant physiology and productivity. Global Change Biology 2021, 27(9), 1704–1720. [Google Scholar] [CrossRef]
- Sekhon, HS.; Singh, G.; Sharma, P.; Bains, TS. Water Use Efficiency Under Stress Environments In: Climate Change and Management of Cool Season Grain Legume Crops; Yadav, SS., Mc Neil, DL., Redden, R., Patil, SA., Eds.; Springer Press: Dordrecht-Heidelberg-London-New York, 2010. [Google Scholar]
- Ferrante, A.; Mariani, L. Agronomic management for enhancing plant tolerance to abiotic stresses: High and low values of temperature, light intensity, and relative humidity. Horticulturae 2018, 4(3), 21. [Google Scholar] [CrossRef]
- Gustafson, A. Yale program on climate change communication. Journal of Applied Communication Research 2025, 53(1), 73–75. [Google Scholar] [CrossRef]
- Sarzynski, T.; Vaast, P.; Rigal, C.; Marraccini, P.; Delahaie, B.; Georget, F.; Nguyen, CTQ.; Nguyen, HP.; Nguyen, HTT.; Ngoc, QL.; Ngan, GK. Contrasted agronomical and physiological responses of five Coffea arabica genotypes under soil water deficit in field conditions. Frontiers in Plant Science 2024, 15, 1443900. [Google Scholar] [CrossRef] [PubMed]
- Camargo, MB.; Marcelo, BP. The impact of climatic variability in coffee crop. International Conference on Coffee Science, 22nd, Campinas, Brazil, 2009. [Google Scholar]
- Lince-Salazar, LA.; Sadeghian, KS. Soil taxonomy, consideration for the coffee regions of Colombia. Boletín Técnico Cenicafé (In Spanish). 2021, 45, 1–31. [Google Scholar] [CrossRef]
- Hao, Y.; Baik, J.; Fred, S.; Choi, M. Comparative analysis of two drought indices in the calculation of drought recovery time and implications on drought assessment: East Africa’s Lake Victoria Basin. Stochastic Environmental Research and Risk Assessment 2022, 36(7), 1943–1958. [Google Scholar] [CrossRef]
- Gichimu, B.M.; Cheserek, J.J. Drought and heat tolerance in coffee: a review. 2012. [Google Scholar]
- Mohanty, BP.; Klittich, WM.; Horton, R.; Vangenuchten, MT. Spatiotemporal variability of soil-temperature within land areas exposed to different tillage systems. Soil Sci. Soc. Am. J. 1995, 59(3), 752–759. [Google Scholar] [CrossRef]
- Famiglietti, JS.; Rudnicki, JW.; Rodell, M. Variability in surface moisture content along a hillslope transect: Rattlesnake Hill, Texas. J. Hydrol. 1998, 210(1-4), 259–281. [Google Scholar] [CrossRef]
- Qiu, Y.; Fu, BJ.; Wang, J.; Chen, LD. Soil moisture variation in relation to topography and land use in a hillslope catchment of the Loess Plateau. China. J. Hydrol. 2001, 240(3-4), 243–263. [Google Scholar] [CrossRef]
- Western, AW.; Zhou, SL.; Grayson, RB.; McMahon, TA.; Blöschl, G.; Wilson, DJ. Spatial correlation of soil moisture in small catchments and its relationship to dominant spatial hydrological processes. Journal of Hydrology 2004, 286(1-4), 113–134. [Google Scholar] [CrossRef]
- Burnett, BN.; Meyer, GA.; McFadden, LD. Aspect-related microclimatic influences on slope forms and processes, northeastern Arizona. J. Geophys. Res.-Earth 2008, 113(F3), 1–18. [Google Scholar] [CrossRef]
- Geroy, IJ.; et al. Aspect influences on soil water retention and storage. Hydrol. Process 2011, 25(25), 3836–3842. [Google Scholar] [CrossRef]
- Gutiérrez-Jurado, HA.; Vivoni, ER. Ecogeomorphic expressions of an aspect-controlled semiarid basin: I. Topo-graphic analyses with high-resolution data sets. Ecohydrology 2013, 6(1), 8–23. [Google Scholar] [CrossRef]
- Fan, B.; Tao, W.; Qin, G.; Hopkins, I.; et al. ‘Soil micro-climate variation in relation to slope aspect, position, and curvature in a forested catchment’. Agricultural and Forest Meteorology 2020, 290, 107999. [Google Scholar] [CrossRef]
- Araya, SN.; Strohmeier, S.; Klik, A.; Singh, RM. Advances in soil moisture retrieval from multispectral remote sensing using unoccupied aircraft systems and machine learning techniques. Hydrology and Earth System Sciences 2021, 25(3), 2739–2758. [Google Scholar] [CrossRef]
- Ndabasanze, P. Suitability analysis of coffee growing area in the Context of climate change in Nyaruguru District, Rwanda. Doctoral dissertation, University of Rwanda, 2018. [Google Scholar]
- Mallet, F.; Marc, V.; Douvinet, J.; Ruy, S. ‘Assessing soil water content variation in a small mountainous catchment over different time scales and land covers using geographical variables’. Journal of Hydrology 2020, 590, 125492. [Google Scholar] [CrossRef]
- Geiger, R.; Aron, RH.; Todhunter, P. The climate near the ground; Rowman & Littlefield, 2009. [Google Scholar]
- Ehrenbergerová, L.; Klimková, M.; Cano, Y.G.; Habrová, H.; Lvončík, S.; Volařík, D.; Khum, W.; Němec, P.; Kim, S.; Jelínek, P.; Maděra, P. ‘Does shade impact coffee yield, tree trunk, and soil moisture on Coffea canephora plantations in Mondulkiri, Cambodia?’. Sustainability 2021, 13(24), 13823. [Google Scholar] [CrossRef]
- Crave, A.; Gascuel-Odoux, C. The influence of topography on time and space distribution of soil surface water content. Hydrological processes 1997, 11(2), 203–210. [Google Scholar] [CrossRef]
- De Lannoy, GJM.; Verhoest, NEC.; Houser, PR.; Gish, TJ.; Van Meirvenne, M. Spatial and temporal characteristics of soil moisture in an intensively monitored agricultural field (OPE3). J. Hydrol. 2006, 331(3-4), 719–730. [Google Scholar] [CrossRef]
- Zhu, Q.; Nie, XF.; Zhou, XB.; Liao, KH.; Li, HP. Soil moisture response to rainfall at different topographic positions along a mixed land-use hillslope. Catena 2014, 119, 61–70. [Google Scholar] [CrossRef]
- Jiang, ZY.; Wang, XD.; Zhang, SY.; He, B.; Zhao, XL.; Kong, FL.; Feng, D.; Zeng, YC. ‘Response of soil water dynamics to rainfall on a collapsing gully slope: based on continuous multi-depth measurements’. Water 2020, 12(8), 2272. [Google Scholar] [CrossRef]
- Green, T.R.; Erskine, R.H. Measurement and inference of profile soil-water dynamics at different hillslope positions in a semiarid agricultural watershed. Water Resources Research 2011, 47(12). [Google Scholar] [CrossRef]
- Burt, TP.; Butcher, DP. Topographic controls of soil-moisture distributions. J. Soil Sci. 1985, 36(3), 469–486. [Google Scholar] [CrossRef]
- Moore, ID.; Burch, GJ.; Mackenzie, DH. Topographic effects on the distribution of surface soil-water and the location of ephemeral gullies. Trans. Am. Soc. Agric. Eng. 1988, 31(4), 1098–1107. [Google Scholar] [CrossRef]
- Castro-Tanzi, S.; Dietsch, T.; Urena, N.; Vindas, L.; Chandler, M. Analysis of management and site factors to improve the sustainability of smallholder coffee production in Tarrazú, Costa Rica. Agriculture, ecosystems & environment 2012, 155, 172–181. [Google Scholar]
- Kobusinge, J.; Twesigye, CK.; Kagezi, GH.; Ssembajwe, R.; Arinaitwe, G. Soil Moisture Content Suitability for Coffee Growing under Climate Change Scenarios in Uganda. PhD Thesis, 2024. [Google Scholar]
- Merga, D.; Beksisa, L. Mechanisms of drought tolerance in coffee (Coffea Arabica L.): implication for genetic im-provement program. American Journal of BioScience 2023, 11(3), 63–70. [Google Scholar] [CrossRef]
- Silva, BM.; de Oliveira, GC.; Serafim, ME.; Carducci, CE.; da Silva, ÉA.; Barbosa, SM.; de Melo, LBB.; dos Santos, WJR.; Reis, THP.; de Oliveira, CHC.; Guimarães, PTG. Soil management and water-use efficiency in Brazilian coffee crops. In Coffee-Production and Research; IntechOpen, 2019. [Google Scholar]
- Chemura, A. The growth response of coffee (Coffea arabica L) plants to organic manure, inorganic fertilizers and integrated soil fertility management under different irrigation water supply levels. International Journal of Recycling of Organic Waste in Agriculture 2014, 3(2), 59. [Google Scholar] [CrossRef]
- Boudjabi, S.; Ababsa, N.; Chenchouni, H. Enhancing soil resilience and crop physiology with biochar application for mitigating drought stress in durum wheat (Triticum durum). Heliyon 2023, 9(12), e22909. [Google Scholar] [CrossRef] [PubMed]
- Amir, R.; Jahan, S.; Zafar, M.; Naeem, M.B.; Usman, S.; Shah, A.A.; Nazim, M.; Alsahli, A.A. Organic amendments optimize Triticum aestivum var Zincol-16 performance under water stress through enhancement in physiobiochemical traits, secondary metabolites and yield. Scientific Reports 2025, 15, 17995. [Google Scholar] [CrossRef]
- DaMatta, FM.; Chaves, ARM.; Pinheiro, HA.; Ducatti, C.; Loureiro, ME. Drought tolerance of two field-grown clones of Coffea canephora. Plant Science 2003, 164, 111–117. [Google Scholar] [CrossRef]
- Asefa, A. Impact of Water Stress and Irrigation Scheduling on Arabica coffee at Jimma, Southwest Ethiopia: A Review. Int.J.Curr.Res.Aca.Rev 2023, 11(08), 20–24. [Google Scholar] [CrossRef]
- Bracken, P.; Burgess, PJ.; Girkin, NT. Opportunities for enhancing the climate resilience of coffee production through improved crop, soil and water management. Agroecology and Sustainable Food Systems 2023, 47(8), 1125–1157. [Google Scholar] [CrossRef]
- Rockström, J.; Barron, J. A reply to Lankford and Agol (2024). Irrigation is more than irrigating: agricultural green water interventions contribute to blue water depletion and the global water crisis. Water International 2025, 50(1), 11–21. [Google Scholar] [CrossRef]
- Mariani, L.; Ferrante, A. Agronomic management for enhancing plant tolerance to abiotic stresses—drought, salinity, hypoxia, and lodging. Horticulturae 2017, 3(4), 52. [Google Scholar] [CrossRef]
- Reyes-Herrera, DF.; Sánchez-Reinoso, AD.; Lombardini, L.; Restrepo-Díaz, H. Physiological responses of coffee (Coffea arabica L.) plants to biochar application under water deficit conditions. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 2023, 51(3), 12873–12873. [Google Scholar] [CrossRef]
- Sott, M.K.; Furstenau, L.B.; Kipper, L.M.; Giraldo, F.D.; Lopez-Robles, J.R.; Cobo, M.J.; Zahid, A.; Abbasi, Q.H.; Imran, M.A. Precision techniques and agriculture 4.0 technologies to promote sustainability in the coffee sector: state of the art, challenges and future trends. Ieee Access 2020, 8, 149854–149867. [Google Scholar] [CrossRef]
- Miletić, RPR.; Dodig, D.; Milutinović, S.; Mihajlović, I.; Nikodijević, SM. Strategies for solving the problem of drought in Eastern crop.icidonline.org/40doc.pdf. 2010.
- Faraz, M.; Mereu, V.; Spano, D.; Trabucco, A.; Marras, S.; El Chami, D. A systematic review of analytical and mod-elling tools to assess climate change impacts and adaptation on coffee agrosystems. Sustainability 2023, 15(19), 14582. [Google Scholar] [CrossRef]
- Vignoli, A.; Ferreira, SG.; Machado, SR. Strategies for managing coffee under drought conditions. Agricultural Water Management 2020, 239, 106248. [Google Scholar]
- Sisay, BT. Coffee production and climate change in Ethiopia. In Sustainable Agriculture Reviews; Climate Impact on Agriculture, 2018; Volume 33, pp. 99–113. [Google Scholar]
- Ramalho, JC; Marques, I; Pais, IP; Armengaud, J; Gouveia, D; Rodrigues, AP; Dubberstein, D; Leitão, AE; Rakočević, M; Scot-ti-Campos, P; Martins, S; Semedo, MC; Partelli, FL; Lidon, FC; DaMatta, FM; Ribeiro-Barros, AI. Stress resilience in Coffea arabica and Coffea canephora under harsh drought and/or heat conditions: selected genes, proteins, and lipid integrated responses. Front. Plant Sci. 2025, 16, 1623156. [Google Scholar] [CrossRef]
- De Souza, BP.; Martinez, HEP.; de Carvalho, FP.; Loureiro, ME.; Sturião, WP. Gas Exchanges and Chlorophyll Fluorescence of Young Coffee Plants Submitted to Water and Nitrogen Stresses. J. Plant Nutr. 2020, 43, 2455–2465. [Google Scholar] [CrossRef]
- Fazuoli, LC.; Braghini, MT.; Silvarolla, MB.; Gonçalves, W.; Mistro, JC.; Gallo, PB.; Guerreiro Filho, O. IAC. Catuaí SH3—A Dwarf Arabica Coffee Cultivar with Leaf Rust Resistance and Drought Tolerance. Crop Breed. Appl. Biotechnol. 2019, 19, 356–359. [Google Scholar] [CrossRef]
- Chaves, M.M.; Pereira, J.S.; Maroco, J. Understanding plant responses to drought from genes to the whole plant. Functional Plant Biology 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Tounekti, T.; Mahdhi, M.; Al-Turki, TA.; Khemira, H. Water relations and photo-protection mechanisms during drought stress in four coffee (Coffea Arabica L.) cultivars from southwestern Saudi Arabia. South African journal of botany 2018, 117, 17–25. [Google Scholar] [CrossRef]
- Zaidan, I.R.; da Silva Ferreira, M.F.; do Couto, D.P.; Santos, J.G.; Silva, M.A.; Canal, G.B.; de Oliveira Bernardes, C.; Azevedo, C.F.; Ferreira, A. Genome-wide association analysis of traits related to development, abiotic and biotic stress re-sistance in Coffea canephora. Scientia Horticulturae 2025, 341, 114004. [Google Scholar] [CrossRef]
- Santos, A.B.; Mazzafera, P. Dehydrins are highly expressed in water-stressed plants of two coffee species. Tropical plant biology 2012, 5(3), 218–232. [Google Scholar] [CrossRef]
- Prado, G.S.; Rocha, D.C.; Santos, L.N.D.; Contiliani, D.F.; Nobile, P.M.; Martinati-Schenk, J.C.; Padilha, L.; Maluf, M.P.; Lubini, G.; Pereira, T.C.; Monteiro-Vitorello, C.B. CRISPR technology towards genome editing of the perennial and semi-perennial crops citrus, coffee and sugarcane. Frontiers in Plant Science 2024, 14, 1331258. [Google Scholar] [CrossRef]
- Valencia-Lozano, E.L.I.A.N.A.; Barraza-Celis, A.A.R.O.N.; Ibarra-Rendon, J.O.R.G.E.; Cabrera-Ponce, J.L. Tran-scriptome Analysis of Coffee Coffea arabica L. Drought Tolerant Somatic Embryogenic Line, Mediated by Antisense Trehalase. 2024. [Google Scholar]
- Tezara, W.; Loyaga, D.W.; Reynel Chila, V.H.; Herrera, A. Photosynthetic Limitations and Growth Traits of Four Arabica Coffee (Coffea arabica L.) Genotypes under Water Deficit. Agronomy 2024, 14(8), 1713. [Google Scholar] [CrossRef]
- Marques, I.; Gouveia, D.; Gaillard, J.C.; Martins, S.; Semedo, M.C.; Lidon, F.C.; DaMatta, F.M.; Ribeiro-Barros, A.I.; Armengaud, J.; Ramalho, J.C. Next-generation proteomics reveals a greater antioxidative response to drought in Coffea arabica than in Coffea canephora. Agronomy 2022, 12(1), 148. [Google Scholar] [CrossRef]
- Hernández Cortés, S. A Coffee Hope Tale: Coffea arabica Castillo and Caturra varieties survive in a drier world. 2024. [Google Scholar]
- Molina, D.; Rivera, R.M. Identifying Coffea genotypes tolerant to water deficit. 2022. [Google Scholar] [CrossRef]
- Carvalho, F.G.; Sera, G.H.; Andreazi, E.; Sera, T.; Fonseca, I.D.B.; Carducci, F.C.; Shigueoka, L.H.; Holderbaum, M.M.; Costa, K.C. Drought tolerance in seedlings of coffee genotypes carrying genes of different species. 2017. [Google Scholar]
- Turreira-García, N. Farmers’ perceptions and adoption of Coffea arabica F1 hybrids in Central America. World De-velopment Sustainability 2022, 1, 100007. [Google Scholar] [CrossRef]
- Sarzynski, T.; Vaast, P.; Rigal, C.; Marraccini, P.; Delahaie, B.; Georget, F.; Nguyen, C.T.Q.; Nguyen, H.P.; Nguyen, H.T.T.; Ngoc, Q.L.; Ngan, G.K. Contrasted agronomical and physiological responses of five Coffea arabica genotypes under soil water deficit in field conditions. Frontiers in Plant Science 2024, 15, 1443900. [Google Scholar] [CrossRef] [PubMed]
- Sseremba, G.; Tongoona, P.B.; Musoli, P.; Eleblu, J.S.Y.; Melomey, L.D.; Bitalo, D.N.; Atwijukire, E.; Mulindwa, J.; Aryatwijuka, N.; Muhumuza, E.; Kobusinge, J. Viability of deficit irrigation pre-exposure in adapting robusta coffee to drought stress. Agronomy 2023, 13(3), 674. [Google Scholar] [CrossRef]
- Bruce, TJ.; Matthes, MC.; Napier, JA.; Pickett, JA. Stressful “memories” of plants: evidence and possible mechanisms. Plant science 2007, 173(6), 603–608. [Google Scholar] [CrossRef]
- Hu, X.; Wu, L.; Zhao, F.; Zhang, D.; Li, N.; Zhu, G.; Li, C.; Wang, W. Phosphoproteomic analysis of the response of maize leaves to drought, heat and their combination stress. Frontiers in plant science 2015, 6, 298. [Google Scholar] [CrossRef]
- Menezes-Silva, PE.; Sanglard, LM.; Ávila, RT.; Morais, LE.; Martins, SC.; Nobres, P.; Patreze, CM.; Ferreira, MA.; Araújo, WL.; Fernie, AR.; DaMatta, FM. Photosynthetic and metabolic acclimation to repeated drought events play key roles in drought tolerance in coffee. Journal of experimental botany 2017, 68(15), 4309–4322. [Google Scholar] [CrossRef]
- Iwasaki, M.; Paszkowski, J. Epigenetic memory in plants. The EMBO journal 2014, 33(18), 1987–1998. [Google Scholar] [CrossRef]
- Fleta-Soriano, E.; Munné-Bosch, S. Stress memory and the inevitable effects of drought: a physiological perspective. Frontiers in Plant Science 2016, 7, 143. [Google Scholar] [CrossRef]
- Waraich, EA.; Ahmad, R.; Ashraf, MY. Role of mineral nutrition in alleviation of drought stress in plants. Aust J Crop Sci 2011, 5(6), 764–777. [Google Scholar]
- Marschner, H. Mineral Nutrition of Higher Plants; Academic Press: San Diego, 1995. [Google Scholar]
- Payne, WA.; Hossner, LR.; Onken, AB.; Wedt, CW. Nitrogen and phosphorous uptake in pearl millet and its relation to nutrient and transpiration efficiency. Agron J 1995, 87, 425–431. [Google Scholar] [CrossRef]
- Raun, WR.; Johnson, GV. Improving nitrogen use efficiency for cereal production. Agron J 1999, 91, 357–363. [Google Scholar] [CrossRef]
- Rufty, TW.; Huber, SC.; Volk, RJ. Alterations in leaf carbohydrate metabolism in response to nitrate stress. Plant Physiol 1988, 88, 725–730. [Google Scholar] [CrossRef]
- Borgo, L.; Rabêlo, F.H.S.; Marchiori, P.E.R.; Guilherme, L.R.G.; Guerra-Guimarães, L.; Resende, M.L.V.D. Impact of drought, heat, excess light, and salinity on coffee production: strategies for mitigating stress through plant breeding and nutrition. Agriculture 2024, 15(1), 9. [Google Scholar] [CrossRef]
- Turner, LB. Changes in the phosphorus content of Capsicum annuum leaves during water-stress. J Plant Physiol 1985, 121, 429–439. [Google Scholar] [CrossRef]
- Studer, C. Interactive effects of N-P-K-nutrition and water stress on the development of young maize plants. Ph.D. Thesis, ETHZ, Zurich, Switzerland, 1993. [Google Scholar]
- Ackerson, RC. Osmoregulation in cotton in response to water-stress. Effects of phosphorus fertility. Plant Physiol 1985, 77, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Garg, BK.; Burman, U.; Kathju, S. The influence of phosphorus nutrition on the physiological response of mothbean genotypes to drought. J Plant Nutr Soil Sci 2004, 167, 503–508. [Google Scholar] [CrossRef]
- Bruck, H.; Payne, WA.; Sattelmacher, B. Effects of phosphorus and water supply on yield, transpirational water-use efficiency, and carbon isotope discrimination of pearl millet. Crop Sci 2000, 40, 120–125. [Google Scholar] [CrossRef]
- Sawwan, J.; Shibli, RA.; Swaidat, I.; Tahat, M. Phosphorus regulates osmotic potential and growth of African violet under in vitro-induced water deficit. J Plant Nutr 2000, 23, 759 771. [Google Scholar] [CrossRef]
- Pilbeam, DJ.; Cakmak, I.; Marschner, H.; Kikby, EA. Effect of withdrawal of phosphorous on nitrate assimilation and PEP carboxylase activity in tomato. Plant Soil 1993, 154, 111–117. [Google Scholar] [CrossRef]
- Cakmak, I.; Engels, C. Role of mineral nutrients in photosynthesis and yield formation, in Rengel, Zn Mineral Nutrition of Crops: Mechanisms and Implications; The Haworth Press: New York, USA, 1999; pp. 141–168. [Google Scholar]
- Bowler, C.; Van-Montagu, M.; Inze, D. Superoxide dismutase and stress tolerance. Ann. Rev. Plant Physiol. Plant Mol Biol 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Elstner, EF.; Osswald, W. Mechanism of oxygen activation during plant stress. Proc. Royal Soc. Edinburgh, Section-B 1994, 102, 131–154. [Google Scholar] [CrossRef]
- Foyer, CH.; Lelandais, M.; Kunert, KJ.; Gadgil. Photooxidative stress in plants. Physiol Plant 1994, 92, 696–717. [Google Scholar] [CrossRef]
- Mengel, K.; Kirkby, EA.; Kosegarten, H.; Appel, T. The soil as a plant nutrient medium. In Principles of plant nutrition; Springer Netherlands: Dordrecht, 2001; pp. 15–110. [Google Scholar]
- Sen Gupta, A.; Berkowitz, GA.; Pier, PA. Maintenance of photosynthesis at low leaf water potential in wheat. Plant Physiol 1989, 89, 1358–1365. [Google Scholar] [CrossRef]
- Sangakkara, UR.; Frehner, M.; Nosberger, J. Effect of soil moisture and potassium fertilizer on shoot water potential, photosynthesis and partitioning of carbon in mungbean and cowpea. J Agron Crop Sci 2000, 185, 201–207. [Google Scholar] [CrossRef]
- Sustr, M.; Soukup, A.; Tylova, E. Potassium in root growth and development. Plants 2019, 8, 435. [Google Scholar] [CrossRef]
- Tilahun, E.; Tadesse, M.; Asefa, A.; Admasu, R.; Shimbir, T. Determination of optimal irrigation scheduling for coffee (Coffee arabica L.) at Gera, South West of Ethiopia. International Journal of Research Studies in Agricultural Sciences 2021, 7(1), 37–42. [Google Scholar] [CrossRef]
- Wintgens, J.N. Coffee: growing, processing, sustainable production. A guidebook for growers, processors, traders, and researchers; WILEY-VCH Verlag GmbH & Co. KGaA, 2004. [Google Scholar]
- Silva, M.O.; Arruda, V.R.S.D.; Barbosa, F.R.S.; Firmino, M.W.M.; Pedrosa, A.W.; Cunha, F.F.D. Water Management of Arabica Coffee Seedlings Cultivated with a Hydroretentive Polymer. Agronomy 2025, 15(1), 218. [Google Scholar] [CrossRef]
- Jemal, A.; Haile, M.; Tesfaye, M. Coffee-based agroforestry systems for enhancing ecosystem services and resilience to climate change in Ethiopia. Sustainability 2019, 11(3), 683. [Google Scholar]
- Holmgren, M. Combined effects of shade and drought on tulip poplar seedlings: trade-off in tolerance or facilitation? Oikos 2000, 90(1), 67–78. [Google Scholar] [CrossRef]
- Vaast, P.; Somarriba, E. Trade-offs between crop intensification and ecosystem services: The role of agroforestry in coffee agroecosystems. Agricultural Systems 2014, 134, 64–73. [Google Scholar]
- da Silva, P.C.; Ribeiro Junior, W.Q.; Ramos, M.L.G.; Lopes, M.F.; Santana, C.C.; Casari, R.A.D.C.N.; Brasileiro, L.D.O.; Veiga, A.D.; Rocha, O.C.; Malaquias, J.V.; Souza, N.O.S. Multispectral Images for Drought Stress Evaluation of Arabica Coffee Genotypes Under Different Irrigation Regimes. Sensors 2024, 24(22), p7271. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Morphological symptoms of drought stress in coffee plants (Coffea arabica) observed in the Mana District, southwest Ethiopia, during the 2023 dry season. The image includes a scale bar. Visible effects are labeled as follows: (A) Branch Die-Back, and (B) leaf abscission (loss), which are characteristic responses to prolonged water deficit.
Figure 1.
Morphological symptoms of drought stress in coffee plants (Coffea arabica) observed in the Mana District, southwest Ethiopia, during the 2023 dry season. The image includes a scale bar. Visible effects are labeled as follows: (A) Branch Die-Back, and (B) leaf abscission (loss), which are characteristic responses to prolonged water deficit.
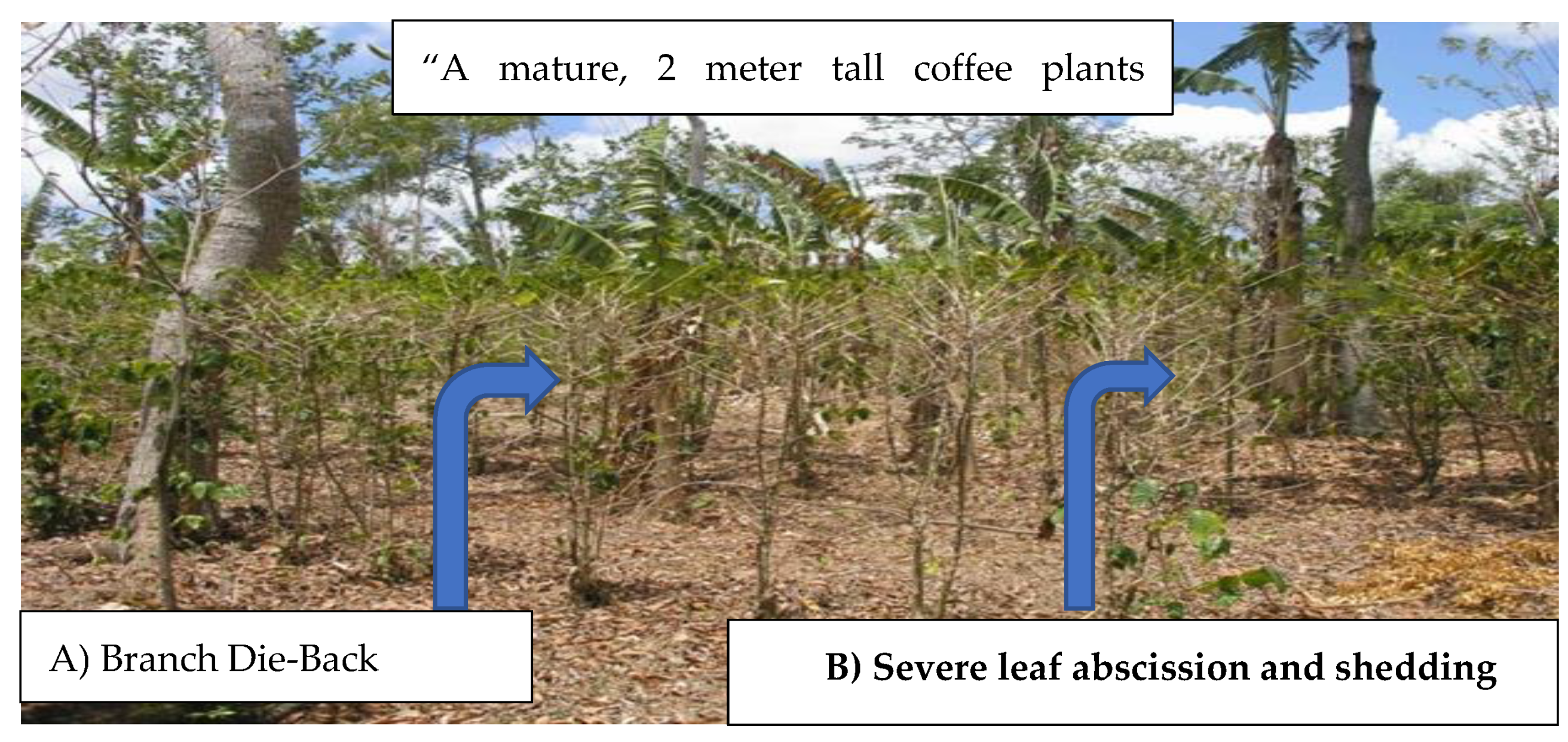
FigureF2.
A Systemic Framework of How VPD, Soil Texture, and Altitude Jointly Govern Coffee Plant Water Status and Physiological Outcomes.
FigureF2.
A Systemic Framework of How VPD, Soil Texture, and Altitude Jointly Govern Coffee Plant Water Status and Physiological Outcomes.
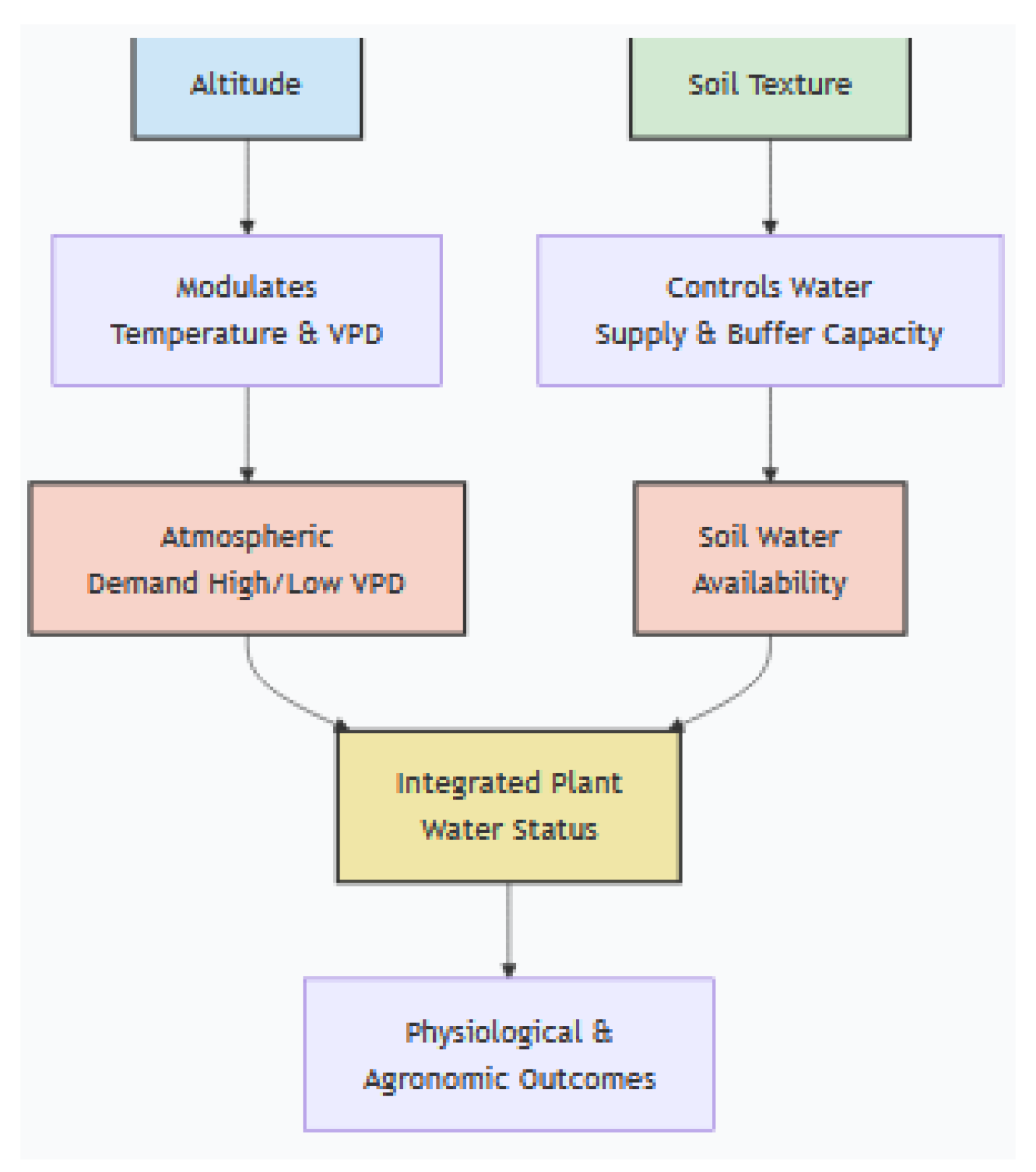
Figure 3.
Soil and water conservation practices in Yemen [158].
Figure 3.
Soil and water conservation practices in Yemen [158].

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



