Submitted:
11 December 2025
Posted:
12 December 2025
You are already at the latest version
Abstract
Multiple sclerosis (MS) pathogenesis involves not only immune-mediated myelin injury but also glial responses. We examined how three charge isomers of myelin basic protein (MBP)—native (C1), phosphorylated (C4), and citrullinated (C8)—modulate rat astrocytes. Cytokines were quantified and grouped (pro/anti-inflammatory, chemotactic, neurotrophic, angiogenic, tissue remodeling), and regulatory markers assessed. C1 strongly upregulated the lipid-sensing receptor LXR, and reduced global DNA methylation; C4 moderately enhanced LXR; C8 failed to activate LXR or alter methylation. Functionally, C1 attenuated IL-1β, IL-6 and GM-CSF while increasing IL-10 and certain chemokines. C4 elicited an intermediate pattern, inducing CX3CL1 (fractalkine), CCL20, VEGF-A and TIMP-1 with minor effects on classical cytokines. In contrast, C8 triggered a robust pro-inflammatory phenotype, increasing IL-1α/β, TNF-α and GM-CSF, with higher IL-10, fractalkine, CCL20, VEGF-A and TIMP-1. All isomers suppressed IFN-γ, IL-4 and CNTF. These data indicate that MBP post-translational modifications drive distinct astrocyte phenotypes through integrated cytokine, metabolic and epigenetic pathways: C1 favours immune regulation and repair, C4 blends inflammatory and reparative cues, and C8 amplifies neuroinflammation. Understanding how modified MBP shapes astrocyte behaviour provides mechanistic insight into lesion evolution in MS and suggests astrocyte-directed strategies to modulate neuroinflammation and promote remyelination.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Animal Sourcing and Euthanasia
2.2. Primary Astrocyte Cultures
2.3. MBP Isomers and Treatments:
2.4. Cytokine Profiling
2.5. DNA Extraction and PCR
2.6. Data Analysis
3. Results
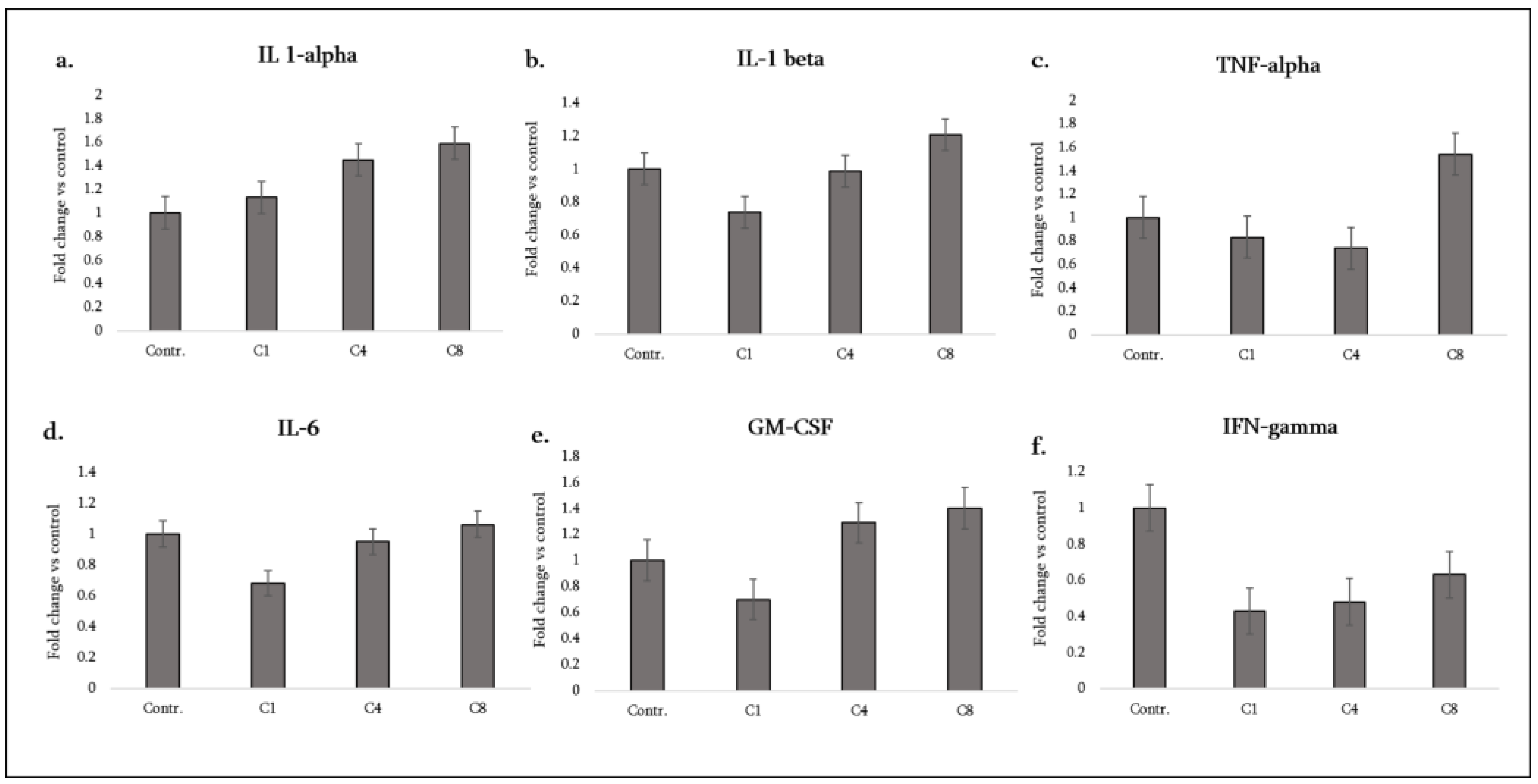
3.1. Pro-Inflammatory Cytokines:
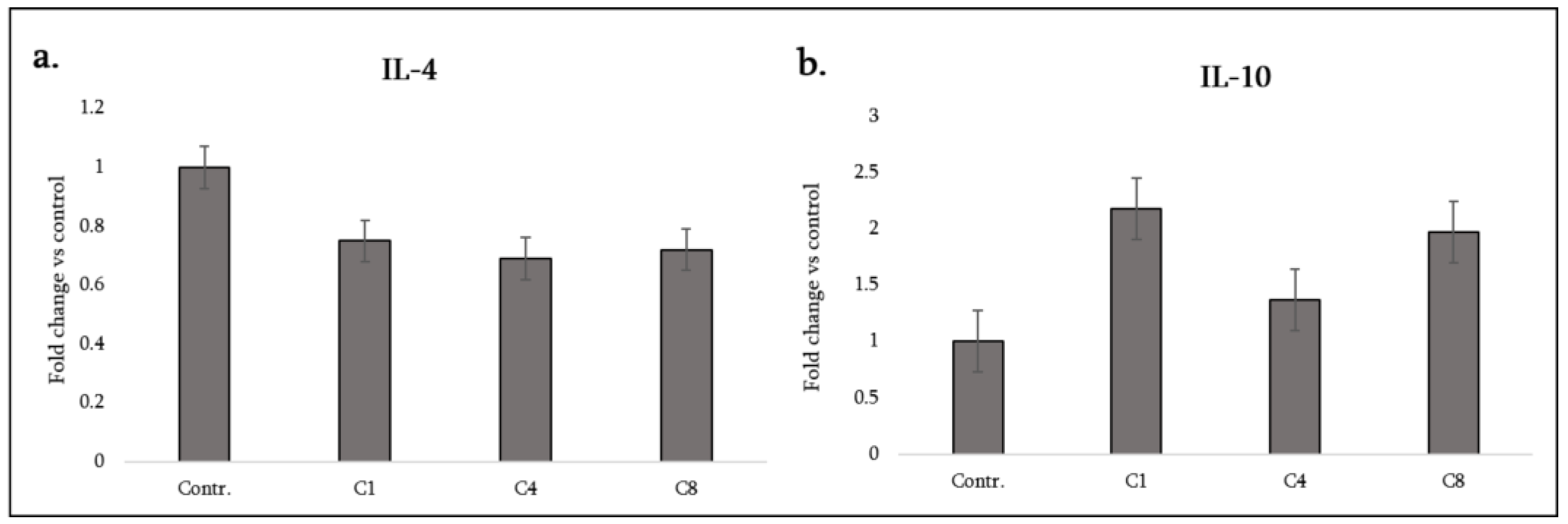
3.2. Anti-Inflammatory Cytokines
3.3. Neutrophil-Attracting CXC Chemokines
3.4. Monocyte/Macrophage- and Lymphocyte-Attracting Chemokines
3.5. Neurotrophic and Growth Factors
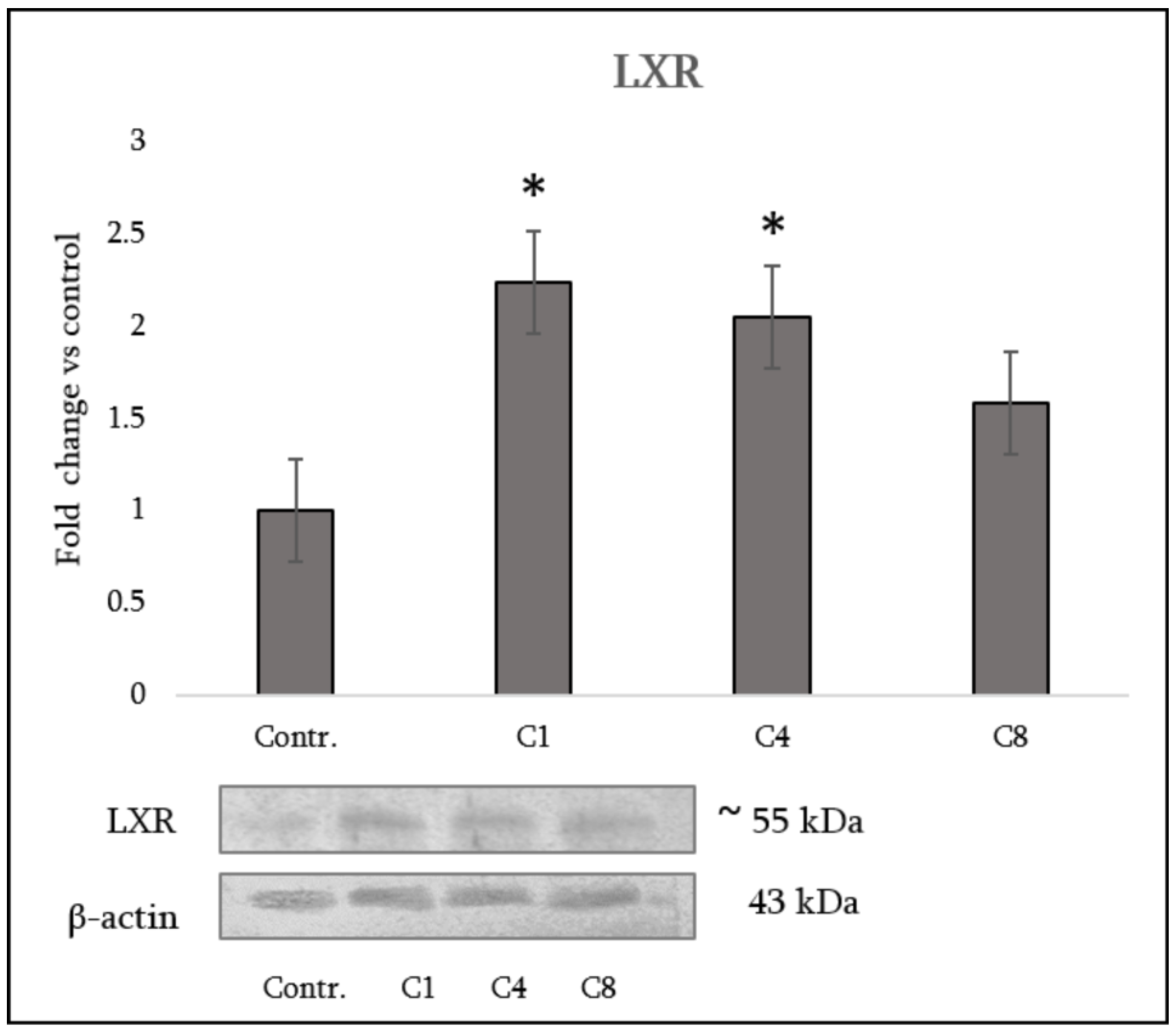
3.5. LXR Expression in Astrocytes
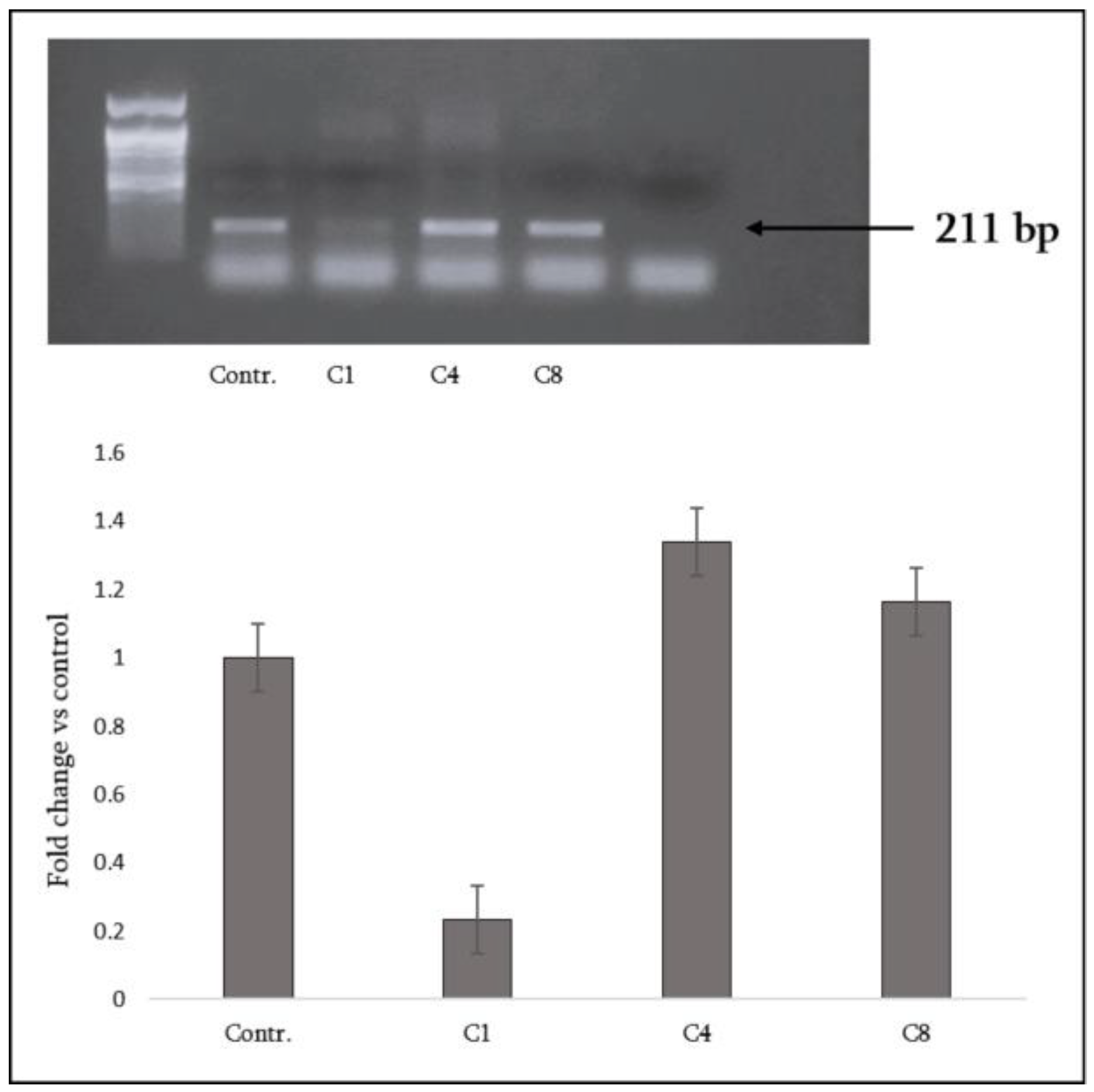
3.6. Global DNA Methylation in Astrocytes
4. Discussion
4.1. Astrocyte Activation by Modified MBP
4.2. Recruitment of Immune Cells
4.3. Relevance to Remyelination
5. Conclusions
- Native MBP (C1) promoted an anti-inflammatory and pro-remyelinating phenotype, with increased IL-10, strong LXR upregulation and markedly reduced DNA methylation;
- Phosphorylated MBP (C4) elicited a mixed response, combining moderate LXR with increased methylation and partial induction of repair-associated factors;
- Citrullinated MBP (C8) drove a pro-inflammatory, neurotoxic profile with impaired LXR activation, stable methylation, and loss of neurotrophic support.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CINC | Cytokine-induced neutrophil chemoattractants |
| CNS | Central nervous system |
| CNTF | Ciliary neurotrophic factor |
| CX3CR1 | Fractalkine receptor |
| EAE | Model experimental autoimmune encephalomyelitis |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| IFN-γ | Interferon-gamma |
| IL-1β | Interleukin-1β |
| LXRs | Liver X receptors |
| MBP | Myelin basic protein |
| MS | Multiple sclerosis |
| NF-κB | Nuclear factor-kappa B |
| OPCs | Oligodendrocyte precursor cells |
| PTMs | Post-translational modifications |
| TIMP-1 | Tissue Inhibitor of Metalloproteinases-1 |
| TNF-α | Tumour necrosis factor-α |
| VEGF-A | Vascular endothelial growth factor-A |
| β-NGF | beta nerve growth factor |
References
- Gaudet, A.D.; Fonken, L.K. Glial Cells Shape Pathology and Repair After Spinal Cord Injury. Neurotherapeutics 2018, 15, 554–577. [Google Scholar] [CrossRef]
- Kıray, H.; Lindsay, S.L.; Hosseinzadeh, S.; Barnett, S.C. The Multifaceted Role of Astrocytes in Regulating Myelination. Exp Neurol 2016, 283, 541–549. [Google Scholar] [CrossRef]
- Li, J.; Zhang, L.; Chu, Y.; Namaka, M.; Deng, B.; Kong, J.; Bi, X. Astrocytes in Oligodendrocyte Lineage Development and White Matter Pathology. Front Cell Neurosci 2016, 10, 119. [Google Scholar] [CrossRef]
- Shanshiashvili, L.; Mikeladze, D. Some aspects of myelin basic protein. J Biol Phys Chem 2003, 3, 96–99. [Google Scholar] [CrossRef]
- Boggs, J.M. Myelin Basic Protein: A Multifunctional Protein. Cell Mol Life Sci 2006, 63, 1945–1961. [Google Scholar] [CrossRef]
- Shanshiashvili, L.V.; Kalandadze, I.V.; Ramsden, J.J.; Mikeladze, D.G. Different Isoforms of Deiminated Myelin Basic Protein Have Different Adsorption Capacities to the Myelin Lipids. J Biol Phys Chem 2014, 14, 57–63. [Google Scholar] [CrossRef]
- Standiford, M.M.; Grund, E.M.; Howe, C.L. Citrullinated Myelin Induces Microglial TNFα and Inhibits Endogenous Repair in the Cuprizone Model of Demyelination. J Neuroinflammation 2021, 18, 305. [Google Scholar] [CrossRef]
- Shanshiashvili, L.V.; Suknidze, N.C.; Machaidze, G.G.; Mikeladze, D.G.; Ramsden, J.J. Adhesion and Clustering of Charge Isomers of Myelin Basic Protein at Model Myelin Membranes. Arch Biochem Biophys 2003, 419, 170–177. [Google Scholar] [CrossRef]
- Chikviladze, M.; Mamulashvili, N.; Sepashvili, M.; Narmania, N.; Ramsden, J.; Shanshiashvili, L.; Mikeladze, D. Citrullinated Isomer of Myelin Basic Protein Can Induce Inflammatory Responses in Astrocytes. IBRO Neurosci Rep 2024, 16, 127–134. [Google Scholar] [CrossRef]
- Moscarello, M.A.; Mastronardi, F.G.; Wood, D.D. The Role of Citrullinated Proteins Suggests a Novel Mechanism in the Pathogenesis of Multiple Sclerosis. Neurochem Res 2007, 32, 251–256. [Google Scholar] [CrossRef]
- Calderon, T.M.; Eugenin, E.A.; Lopez, L.; Kumar, S.S.; Hesselgesser, J.; Raine, C.S.; Berman, J.W. A Role for CXCL12 (SDF-1α) in the Pathogenesis of Multiple Sclerosis: Regulation of CXCL12 Expression in Astrocytes by Soluble Myelin Basic Protein. J Neuroimmunol 2006, 177, 27–39. [Google Scholar] [CrossRef]
- Chikviladze, M.; Mamulashvili, N.; Shanshiashvili, L.; Mikeladze, D. Influence of Myelin Basic Protein Charge Isomers on IL-33 and Galectin-3 Expression in astrocytes. J Biol Phys Chem 2022, 22, 59–60. [Google Scholar] [CrossRef]
- Aharoni, R.; Eilam, R.; Arnon, R. Astrocytes in Multiple Sclerosis—Essential Constituents with Diverse Multifaceted Functions. IJMS 2021, 22, 5904. [Google Scholar] [CrossRef]
- Lotfi, N.; Thome, R.; Rezaei, N.; Zhang, G.-X.; Rezaei, A.; Rostami, A.; Esmaeil, N. Roles of GM-CSF in the Pathogenesis of Autoimmune Diseases: An Update. Front Immunol 2019, 10, 1265. [Google Scholar] [CrossRef]
- Zhang-Gandhi, C.X.; Drew, P.D. Liver X Receptor and Retinoid X Receptor Agonists Inhibit Inflammatory Responses of Microglia and Astrocytes. J Neuroimmunol 2007, 183, 50–59. [Google Scholar] [CrossRef]
- Richartz, J.; Yam, S.C.; Zhan, N.; Schepers, M.; Tiane, A.; Mulder, M.T.; Wens, I.; Vanmierlo, T. Liver X Receptors: A Therapeutic Target in Demyelinating Disorders. Pharmacol Res 2025, 219, 107861. [Google Scholar] [CrossRef]
- Kremer, L.P.M.; Cerrizuela, S.; El-Sammak, H.; Al Shukairi, M.E.; Ellinger, T.; Straub, J.; Korkmaz, A.; Volk, K.; Brunken, J.; Kleber, S.; et al. DNA Methylation Controls Stemness of Astrocytes in Health and Ischaemia. Nature 2024, 634, 415–423. [Google Scholar] [CrossRef]
- Pamies, D.; Sartori, C.; Schvartz, D.; González-Ruiz, V.; Pellerin, L.; Nunes, C.; Tavel, D.; Maillard, V.; Boccard, J.; Rudaz, S.; et al. Neuroinflammatory Response to TNFα and IL1β Cytokines Is Accompanied by an Increase in Glycolysis in Human Astrocytes In Vitro. IJMS 2021, 22, 4065. [Google Scholar] [CrossRef]
- Joseph, S.B.; Castrillo, A.; Laffitte, B.A.; Mangelsdorf, D.J.; Tontonoz, P. Reciprocal Regulation of Inflammation and Lipid Metabolism by Liver X Receptors. Nature Med 2003, 9, 213–219. [Google Scholar] [CrossRef]
- Daskalaki, M.G.; Lapi, I.; Hurst, A.E.; Al-Qahtani, A.; Vergadi, E.; Tsatsanis, C. Epigenetic and Metabolic Regulation of Macrophage Responsiveness and Memory. J Immunol 2025, vkaf135. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.-S.; Peterson, T.C.; et al. Neurotoxic Reactive Astrocytes Are Induced by Activated Microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef]
- Musse, A.A.; Harauz, G. Molecular “Negativity” May Underlie Multiple Sclerosis: Role of the Myelin Basic Protein Family in the Pathogenesis of MS. Intl Rev Neurobiol 2007, 79, 149–172. [Google Scholar]
- Cua, D.J.; Hutchins, B.; LaFace, D.M.; Stohlman, S.A.; Coffman, R.L. Central Nervous System Expression of IL-10 Inhibits Autoimmune Encephalomyelitis. J Immunol 2001, 166, 602–608. [Google Scholar] [CrossRef]
- Song, Y.; Li, X.; Huang, D.; Song, H. Macrophages in Periapical Lesions: Potential Roles and Future Directions. Front Immunol 2022, 13, 949102. [Google Scholar] [CrossRef]
- Matejuk, A.; Matejuk, S.; Offner, H.; Vandenbark, A.A. Altered Lipid Metabolism in CNS Demyelination and Remyelination Are Key Elements Driving Progressive MS. IJMS 2025, 26, 8314. [Google Scholar] [CrossRef]
- Wu, C.-H.; Chen, C.-C.; Lai, C.-Y.; Hung, T.-H.; Lin, C.-C.; Chao, M.; Chen, S.-F. Treatment with TO901317, a Synthetic Liver X Receptor Agonist, Reduces Brain Damage and Attenuates Neuroinflammation in Experimental Intracerebral Hemorrhage. J Neuroinflammation 2016, 13, 62. [Google Scholar] [CrossRef]
- Berghoff, S.A.; Spieth, L.; Saher, G. Local Cholesterol Metabolism Orchestrates Remyelination. Trends Neurosci 2022, 45, 272–283. [Google Scholar] [CrossRef]
- Pineda-Torra, I.; Siddique, S.; Waddington, K.E.; Farrell, R.; Jury, E.C. Disrupted Lipid Metabolism in Multiple Sclerosis: A Role for Liver X Receptors? Front. Endocrinol. 2021, 12, 639757. [Google Scholar] [CrossRef]
- Bilotta, M.T.; Petillo, S.; Santoni, A.; Cippitelli, M. Liver X Receptors: Regulators of Cholesterol Metabolism, Inflammation, Autoimmunity, and Cancer. Front. Immunol. 2020, 11, 584303. [Google Scholar] [CrossRef]
- Babaalizadeh, B.; Kalaki-Jouybari, F.; Abarghooi-Kahaki, F.; Afkhami, H.; Razavi, Z.-S. Role of Nuclear Receptors on the Progression of Multiple Sclerosis: A Review. Cell Mol Neurobiol 2025, 45, 58. [Google Scholar] [CrossRef]
- Kaluski, S.; Portillo, M.; Besnard, A.; Stein, D.; Einav, M.; Zhong, L.; Ueberham, U.; Arendt, T.; Mostoslavsky, R.; Sahay, A.; et al. Neuroprotective Functions for the Histone Deacetylase SIRT6. Cell Rep 2017, 18, 3052–3062. [Google Scholar] [CrossRef]
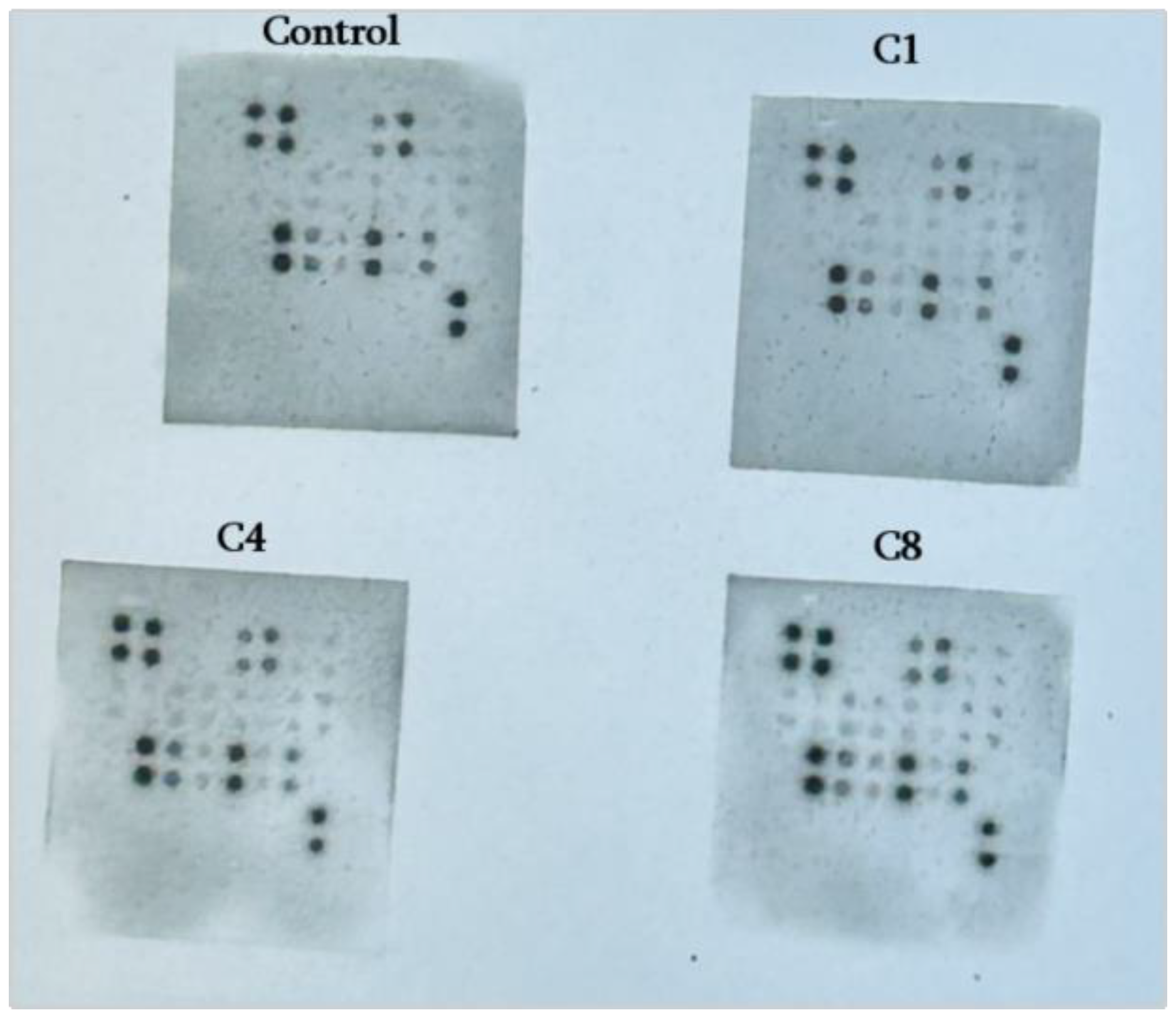
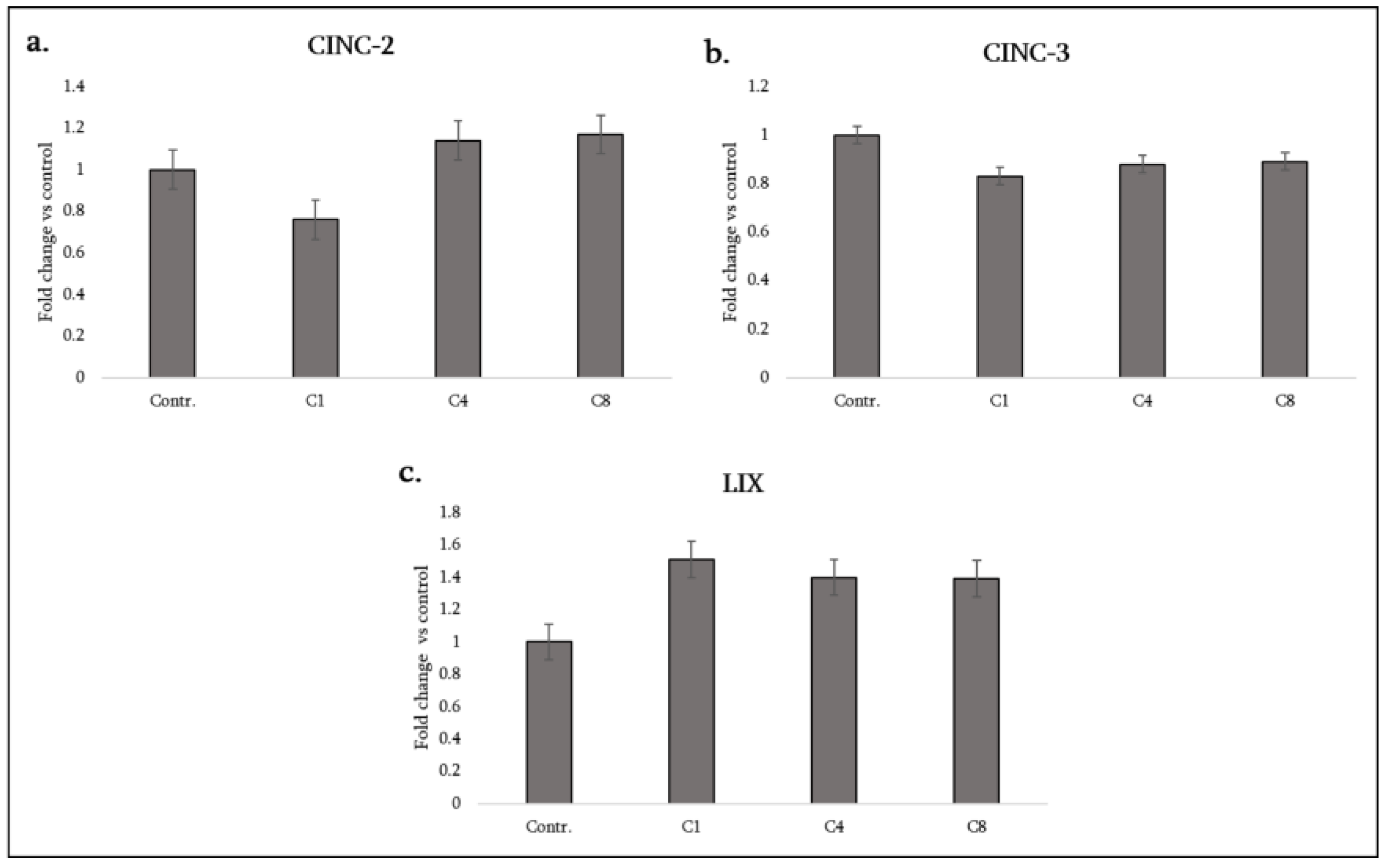
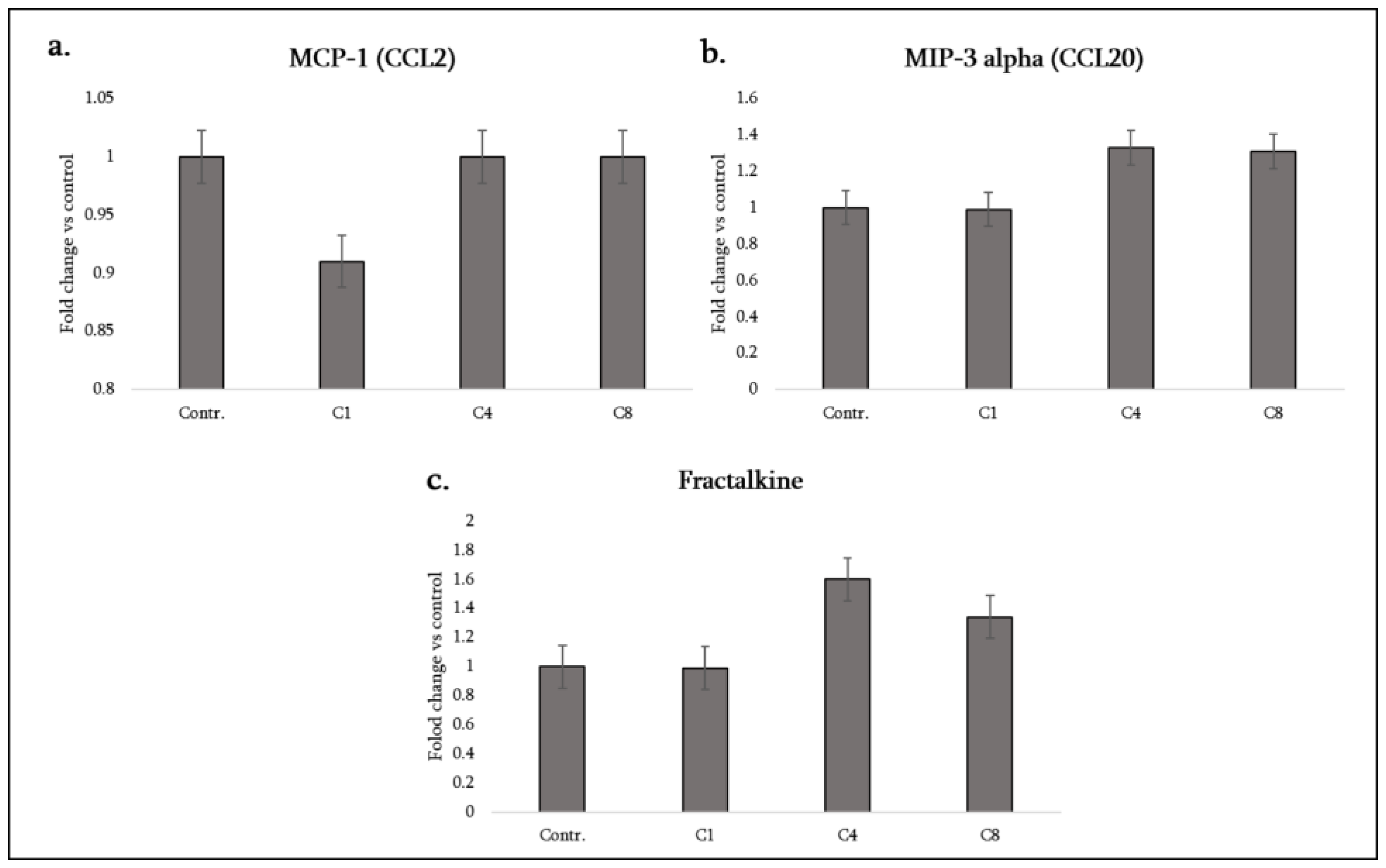
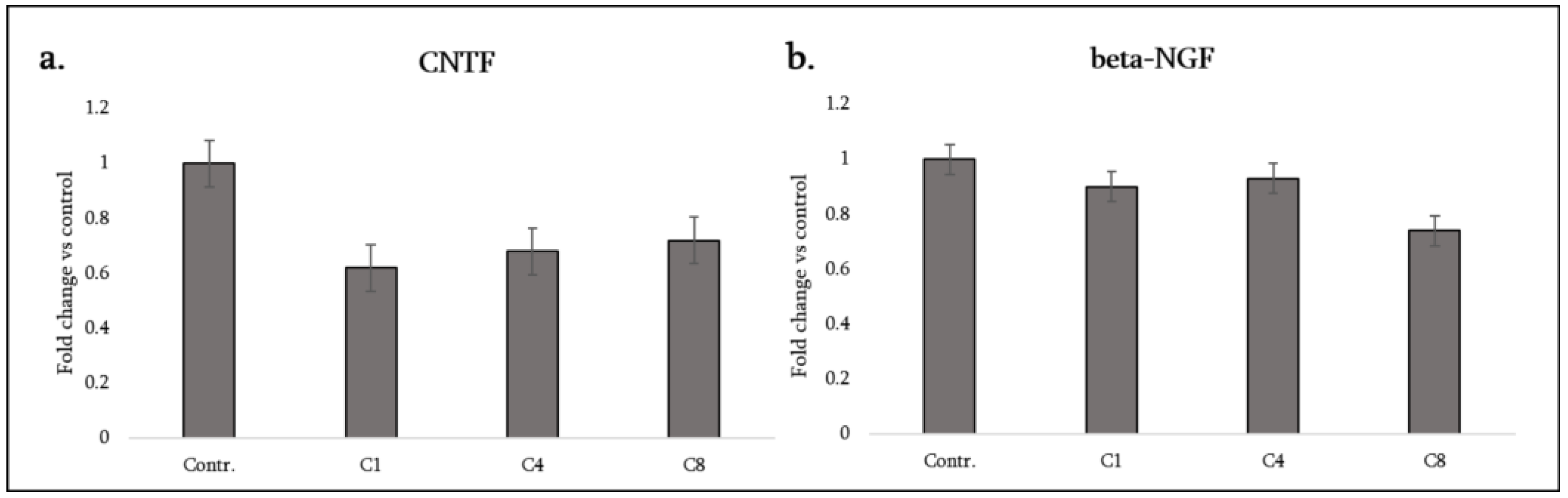
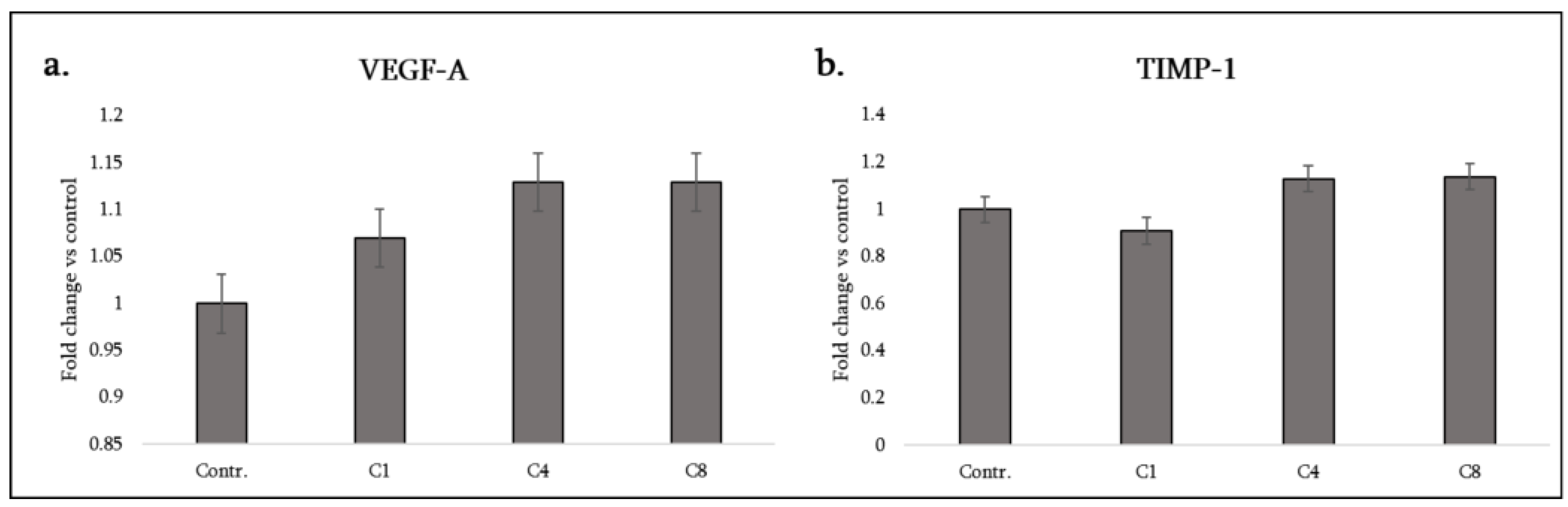

| Cytokine (Synonyms) | Functional Group | C1 | C4 | C8 |
|---|---|---|---|---|
| CINC-2 (CXCL2) | Chemokine (pro-inflammatory) | ↓ | – | – |
| CINC-3 (CXCL3/MIP-2) | Chemokine (pro-inflammatory) | ↓ | ↓ | ↓ |
| CNTF (Ciliary neurotrophic factor) | Neurotrophic cytokine | ↓ | ↓ | ↓ |
| Fractalkine (CX3CL1) | Chemotactic factor (atypical CX3C) | – | ↑ | ↑ |
| GM-CSF (CSF2) | Pro-inflammatory cytokine/myeloid activator | ↓ | – | ↑ |
| IFN-γ | Pro-inflammatory cytokine | ↓ | ↓ | ↓ |
| IL-1α | Pro-inflammatory cytokine (alarmin) | – | ↑ | ↑ |
| IL-1β | Pro-inflammatory cytokine | ↓ | – | ↑ |
| IL-4 | Anti-inflammatory (Th2 cytokine) | ↓ | ↓ | ↓ |
| IL-6 | Pro-inflammatory cytokine/Th17 | ↓ | – | – |
| IL-10 | Anti-inflammatory cytokine | ↑ | – | ↑ |
| LIX (CXCL5) | Chemokine (pro-inflammatory) | ↑ | ↑ | ↑ |
| MCP-1 (CCL2) | Chemotactic factor (pro-inflammatory) | ↓ | – | – |
| MIP-3α (CCL20) | Chemotactic factor (pro-inflammatory) | – | ↑ | ↑ |
| β-NGF | Neurotrophic factor | – | – | ↓ |
| TIMP-1 | Tissue remodeling factor | – | ↑ | ↑ |
| TNF-α | Pro-inflammatory cytokine/cytotoxic | – | – | ↑ |
| VEGF-A | Angiogenic factor | ↑ | ↑ | ↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



