Submitted:
10 December 2025
Posted:
11 December 2025
You are already at the latest version
Abstract
Prosopis juliflora is a highly invasive tree species in semi-arid and arid regions in eastern Africa. Its ability to displace herbaceous and woody species has been attributed to allelopathic effects, but this has rarely been tested in competition experiments on natural soil and experimentally binding potentially allelopathic substances. We tested the effect of soil collected underneath and outside of P. juliflora canopies, or treated with P.julilfora leaf litter, on the survival, growth and competitive ability of three resident tree species in the presence and absence of activated carbon. Survival and growth of tree seedlings was reduced on, compared to seedlings growing on soil collected outside P. juliflora canopies. When activated carbon was added to the soils, seedling performance significantly increased and did not differ anymore from that on soil collected outside P. juliflora canopies. Competition significantly reduced seedling height irrespective of the type of competitor (P. juliflora or resident tree species). There was no significant interaction between soil type and competition, suggesting that the effect of competition was independent of soil type. The results suggest that P. juliflora releases allelochemicals into the soil which have allelopathic effects on resident tree species and that at least part of these allelochemicals originate from leaf material.
Keywords:
allelopathic effects
; activated carbon
; resident trees
; Prosopis juliflora
; survival
; growth
1. Introduction
According to the “novel weapons hypothesis,” some invasive non-native species possess novel biochemical weapons that function as powerful allelopathic agents exerting a direct inhibitory effect on the germination or growth of native plants, or they mediate plant-soil microbial interactions [1,2,3,4]. The inhibitory effect of invasive plants on the growth and development of other plant species may be mediated by organic compounds that are released into the environment as exudates, leachates, or volatiles. Allelochemicals are important mediator in plant-plant interaction if they accumulate in the soil, creating persisting effects on the establishment and growth of interspecific and conspecific plants [5].
The invasive non-native tree Prosopis juliflora (Sw.) DC. (Fabaceae) is considered to be one of the worst invasive non-native species worldwide [6]. It reduces biodiversity and forage for livestock, consumes a lot of water, and increases densities of disease-transmitting insects (Muller et al. 2017; Schaffner et al. 2025). Prosopis juliflora has been found to increase mortality of native trees, and this effect has been attributed to competition for limited resources such as water, or to allelopathy [7,8]. Aqueous extracts from different parts of P. juliflora, including the leaves, bark, and the roots, exhibit phytotoxic effects on various plant species, including economically significant crops such as alfalfa, sesame and sorghum [9]; Ratnayake, 2021). [11]t al. (2021) compared the inhibitory effects of leaf, bark, and root aqueous extracts on the germination of native grass species and found that leaf extracts showed the highest effect. The leaf leachates of P. juliflora contain high concentrations of phenolic compounds and L-tryptophan, both of which may contribute to the plant’s allelopathic properties (Endris, 2008). Large and dense stands of P. juliflora negatively influence growth of plant species richness compared to outside P. juliflora canopy (Mehar [13,14].
However, most of the studies on allelopathy of P. juliflora published so far were conducted under artificial conditions, e.g., using petri dishes (e.g., [11,13,15,16]), artificial potting media or soil of unspecified origin [11,16,17,18]. Experiments conducted under artificial conditions may be confounded by experimental artifacts of the methodology and thus make it difficult to draw conclusions on the role of allelopathy in natural ecosystems. Also, to our knowledge, methodological approaches to differentiate allelopathy from competition for limiting resources, such as water [19], have not been used in experiments assessing allelopathic effects of P. juliflora. This differentiation is particularly important because the effects of allelopathy can be difficult to separate from other more pronounced impacts of competition for resources [20,21].
Activated carbon (AC) has been widely used to differentiate between allelopathy and other processes that can influence plant–plant interactions due to its ability to absorb allelochemicals, thereby neutralizing their potential inhibitory effects on plant growth [22,23,24,25]. Adding AC to soils can, however, also affect nutrient availability and plant growth directly (Lau et al. 2008). Because other techniques to study allelopathy also have their limitations, ref. [26] suggested that using a set of techniques (e.g., AC, watering with plant extracts, root chamber experiments) provides a robust approach to study allelopathy.
In this study, we assessed the role of allelopathy in the interaction between P. juliflora and common tree species in the invaded range in eastern Africa by assessing the impact of soils from underneath and outside the Prosopis canopy and of soil mixed with leaf litter on the survival and growth of seedlings in the presence and absence of AC. We hypothesized that a) in the absence of AC, soil collected underneath P. juliflora cover (UPC) reduces the survival, growth and competitive ability of seedlings from resident tree species, b) adding leaf litter to soil collected outside the Prosopis canopy (OPCLL) also reduces survival, growth and competitive ability of resident tree seedlings, c) the effects of UPC and OPCLL disappear when AC is added to the soil. In our experiments we used three tree species (Vachellia tortilis (Forssk.) Galasso & Banfi, Balanites aegyptica (L.) Delile, and Ziziphus mauritania Lam.) which are characteristic for habitats invaded by P. juliflora in eastern Africa. Both V. tortilis and B. aegyptica are native to eastern Africa while Z. mauritania is native to Asia but widely naturalized in eastern Africa.
2. Material and Methods
2.1. Plant and Soil Samples
The seeds of the three resident tree species (V. tortilis, B. aegyptica and Z. mauritania) and P. juliflora were collected from locally available trees one month before the experiment. The seeds of V. tortilis and B. aegyptica were soaked in warm water for 24 h before sowing to improve germination rates. Immediately after the treatment, the seeds of the two species and of Z. mauritania were sown in seed beds prepared at the KEFRI tree nursery. Four days after sowing the seeds of the three resident species, seeds of P. juliflora were also sown in the same nursery. This procedure made it possible to obtain seedlings of all the test species at the same time. Seeds were watered twice every day, in the morning and in the evening. Any weeds which sprouted in the potting tubes containing seedlings of the test species were removed continuously.
Potting media used for raising the seedlings were sourced from KEFRI Marigat Centre in Baringo County, Kenya, either underneath P. juliflora canopy (UPC) or outside P. juliflora canopy (OPC). A total of 140 soil cores (20 cm in depth, 5 cm in diameter) were collected underneath and outside the canopy of P. juliflora, representing 70 soil cores for each soil type. The soil from underneath P. juliflora canopy was sampled from more than 40 P. juliflora trees of different sizes. All the soil samples were collected a day before setting up the experiment.
Fully developed leaves of P. juliflora of 3-10 cm length were collected a fortnight before the experiment was set up. The leaves were cleaned using fresh water, sun-dried, crushed and blended to form fine powder [27]. The powdered leaf samples were stored in a darkness at room temperature. On the day the experiment was set up, half of the soil collected outside P. juliflora canopy was mixed with the powdered P. juliflora leaf litter at a ratio of 60 g leaf litter/ kg soil [28].
2.2. Experimental Design
The study encompassed three soil treatments: soil from underneath the Prosopis canopy (UPC), soil from outside the Prosopis canopy (OPC), and soil from outside the Prosopis canopy mixed with Prosopis leaf litter (OPCLL). We added AC bought from Ecospot Chemicals Africa Ltd-Jacobi Aquasorb to half of the pots from each soil type at a concentration of 20 mL/L substrate [29].
Seedlings of V. tortilis, B. aegyptica, Z. mauritania and P. juliflora were transplanted at the 2-leaf stage (approximately 4-10 cm tall) into pots filled with the six soil type-AC combinations. Sets of 18 pots were set up containing one seedling of three resident species (V. tortilis, B. aegyptica, Z. mauritania), resulting in six soil type-AC combinations with a seedling of one of the three resident species. In a first set, the resident seedlings were grown alone. In a second set, a second resident seedling was transplanted alongside the first resident seedling; the second seedling was never a conspecific of the first resident seedling. In a third set, a seedling of P. juliflora was transplanted alongside the resident seedling. Lastly, one P. juliflora seedling was transplanted alone in each of the six soil type-AC combinations. This resulted in 60 different soil type-AC-tree species combination. Each soil type-AC-tree species combination was replicated 4 times, resulting in a total of 240 pots.
All pots were transferred to an empty garden bed in the nursery at the KEFRI Centre in Marigat, Kenya. The pots were arranged in a randomized block design, with each block comprising one replicate of each of the soil type-AC-tree species combinations. On the day the potting tubes were transferred to the garden bed, height was measured for all seedlings. The newly transplanted seedlings were shaded for a period of two weeks to protect against shock and enable them to adjust to the new conditions. The pots were evenly watered using a fine nozzle watering can. The watering was done twice a day, early in the morning and late in the afternoon. After three months, all seedlings were harvested and their survival and height recorded.
2.3. Data Analysis
The statistical software R (version 4.4.2; R Core Team, 2024) with the interface R Studio (version 2024.9.0.375; Posit team, 2024) was used. Two samples were excluded, because of extreme outliers in height baseline that could not be explained. A Rosner’s Test from the package ‘EnvStats’ (version 3.0.0; Millard SP, 2013) was used to confirm that they were extreme outliers. Additionally, one sample was excluded, as it stood out as an influential data point in the model on height after three months in the QQ-plot, the comparison of residuals and fitted values and in Cook’s distance. The dataset used for the analysis of the binary variable ‘survival after three months’ included 213 samples. A logistic model was fitted including potting media (UPC, OPC, OPCLL), AC (yes, no), competition (none, resident, Prosopis), the three two-way interactions of the aforementioned variables, height baseline, and resident species as fixed factors.
For the analysis of height after three months, all the samples that were still alive at this point were considered (N = 129). A linear regression model was fitted including the same fixed factors as for the model on survival. Heteroscedasticity was detected using the Breusch-Pragan-Test from the package ‘lmtest’ (version 0.9.40; [30], thus heteroscedasticity consistent standard errors were calculated, using HC3 as suggested by [31]. A Wilcoxon rank sum test was conducted to assess the direct effect of AC on seedling of resident tree species after three months in the absence of competition.
3. Results
3.1. Effect of Soil Origin and Activated Carbon on Seedling Survival
The survival of resident seedlings differed depending on the soil types and the presence and absence of AC. In the absence of AC, survival of resident seedlings was 75% in OPC soil but only 40% in IPC and 12% in OPCLL soils (Figure 1). When AC was added, survival of resident seedlings was 70-75% in UPC and OPC soils and somewhat lower in OPCLL soils. In particular, adding AC to soil collected outside P. juliflora canopy mixed with leaf litter significantly increased the survival of resident seedlings (Table S1). The figure below shows the effect of soil origin and activated carbon on seedlings survival (Figure 1).
3.2. Effect of Soil Type and AC on Resident Seedlings Height
The height of resident seedlings was significantly influenced by the soil type and by the presence/absence of AC. In the absence of AC, resident seedling height was highest in OPC soils, followed by UPC and OPCLL soil types. With addition of AC, the increase in seedling height was significantly larger in OPCLL and UPC soils, relative to OPC soil. The figure below indicates the effect of soil type and activated carbon on resident seedlings height (Figure 2).
3.3. Effect of Soil Origin and Competition on Seedling Height
Competition significantly reduced resident seedling height irrespective of the type of competitor (P. juliflora or resident seedlings) (Table S2). Additionally, there is no significant interaction between the soil type and competition, suggesting that the effect of resident or P.julifora competitors was independent of soil type (Table S2). Overall, the lowest seedling height was found in OPCLL soil, followed by UPC and OPC soils, both in the presence and absence of competitor species. Below is the figure that visualizes the effects of soil origin and competition on resident seedlings height (Figure 3).
3.4. Effect of Competition and Activated Carbon (AC) on Seedling Height
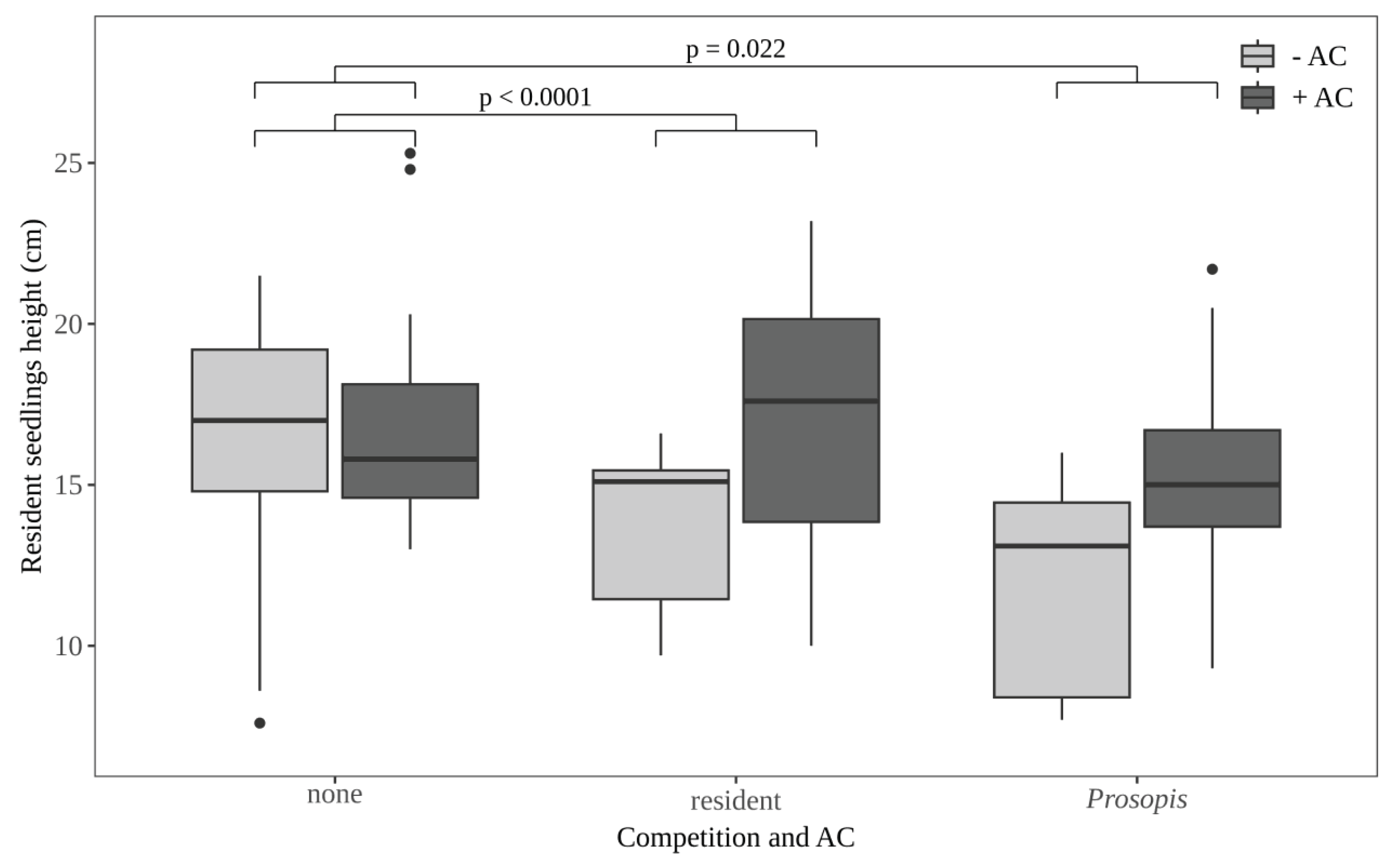
The effect of AC differs between different competition treatments. While there was no direct effect of AC on seedling growth in the absence of competition (Wilcoxon rank sum test: P-value= 0.7526), addition of AC increased seedling growth in the presence of both resident and P. juliflora seedlings (Table S2). The effects of competition of resident seedlings and activated carbon on resident seedlings height is indicated in Figure below (Figure 4).
4. Discussion
Our results provide evidence that soil collected from underneath the P. juliflora canopy and leaf litter added to soil incur allelopathic effects on seedlings of resident tree species by negatively affecting their survival and growth. Seedlings grown on UPC and OPCLL soils suffered increased mortality, relative to OPC soil. Similarly, growth of the resident seedlings was suppressed in UPC and OPCLL soils. When AC was added to UPC and OPCLL soils, the survival and growth of the resident seedlings increased significantly. This suggests that P. juliflora releases allelochemicals into the soil which have allelopathic effects on resident trees and that at least part of these allelochemicals originate from leaf material.
4.1. The Effect of Soil Origin
We found that seedling survival and growth of resident t differed between soil collected from underneath P. juliflora canopy and soil collected outside P. juliflora canopy. These results are consistent with a study on the impact of the invasive tree Acacia pennatula (Benth) on the performance of co-occurring tree species which revealed suppressed growth of tree seedlings underneath the A. pennatula canopy [32]. Similarly, a study on allelopathy of invasive tamarisk species (Tamarix ramosissima Ledeb., T. chinensis Lour.) revealed negative effects of tamarisks on co-occurring plants due to allelopathic effects mediated by the soil in which tamarisks grow [33]. The low survival and stunted growth of the resident tree species grown in UPC soils may be explained by the release of toxic plant metabolites that are directly inhibitory to growth [34]. Alternatively, this effect can also stem from the accumulation of soil microorganisms which render the living conditions unfavorable for seedlings of con- or heterospecific species. For example, Packer & Clay [35] found that sterilization of soil collected underneath trees improved seedling survival relative to unsterilized soil, whereas sterilization of distant soil did not affect survival; inoculation of healthy seedlings with isolates obtained from dying seedlings reduced seedling growth by 65%.
4.2. Leaf Litter as a Source of Allelopathic Substances
Various parts of invasive alien plants, including leaves, roots and bark, have been assessed as potential sources of allelopathic substances [11,36]. Overall, leaf litter and root exudates appear to be the main sources of allelochemicals, while bark contributes relatively little to the allelopathic effects of invasive alien plant species [34,37]. A previous study [38] had indicated that leaves of invasive species are the main source of phytotoxic compounds due to fast decomposition, which enhances the release of allelochemicals. This notion is in line with our findings that adding P. juliflora leaf litter to soil caused a strong allelopathic effect on seedlings of resident tree species. Similarly, ref. [39] found that leaf litter of invasive knotweeds elicited more inhibitory effects on seed germination of native plant species than soil collected underneath the invader’s canopy [39].
The more pronounced inhibitory effects of soil mixed with leaf litter compared to soil collected underneath P. juliflora canopy may be due to a greater concentration of active compounds released from decomposing leaf material, or the chemical composition differs between leaves and roots [40,41]. Alternatively, exudates of P. juliflora may degrade relatively rapidly in the soil, thus leaving only a short-term legacy effect in the absence of continuous influx from plant material [42].
In studies assessing the allelopathic effects of leaves of invasive alien plant species, either leaf litter aqueous extracts or leaf litter fragments were mixed with soil. A study by Huang [40] compared the two treatments when testing the allelopathic effects of Cinnamomum septentrionale Hand. Mazz leaves on tree saplings. They found that both treatments negatively influenced the growth, chlorophyll synthesis and photosynthesis of Eucalyptus grandis Hill ex Maid saplings, but that leaf litter aqueous extracts had a slightly stronger effect than leaf litter fragments.
4.3. Assessing Allelopathy of P. juliflora Using Multiple Techniques Under Semi-Natural Conditions
We combined three different techniques to assess allelopathic effects of P. julilfora, i.e., use of soil with a P. juliflora history, the use of AC and competitive interactions of resident plants with P. juliflora. This was done deliberately to reduce the risk of methodological artifacts in experimental designs as suggested in previous allelopathy studies [26]. Applying a set of techniques and comparing the results provides compelling evidence for the role of allelopathy in species interactions. In particular, we found a significant increase in height of resident species grown in competition with seedlings of another resident tree species or with P. juliflora when AC was added to UPC and OPCLL soils.
The significant increase in height development of resident seedlings in competition with P. juliflora in the presence of AC can be attributed to the ability of AC to bind allelopathic substances exuded by P. juliflora [43,44]. Similarly, the fact that height of resident seedlings grown in competition with a second resident seedlings also increased when AC was mixed with soil may be explained by allelopathic properties of at least some of the test plant species. In particular, Vachellia tortilis might have influenced result of the experiment as aqueous extracts of different plant parts exhibit allelopathic effects on seedlings of crop plants [45].
In summary, our study provides evidence for allelopathic effects of P. julilfora on resident tree species and thereby corroborates results from other studies in which the role of allelopathy in plant invasions was assessed by combining multiple experimental techniques [29,46]. The sensitivity of resident plant species to allelochemicals exuded by invasive alien plants can vary, which complicates the interpretation of results when using a single test species [47]. Therefore, further studies are needed to better understand the allelopathic potential of P. juliflora against a wider range of plant species. Also, identification of the allelopathic substances exuded by P. juliflora and monitoring their concentrations in the soil over time will help to manage the allelopathic interactions between P. juliflora and resident plant species in restoration projects [48,49].
5. Conclusion
The results suggests that leaf litter and soil from underneath Prosopis juliflora canopy inhibited survival and height development of resident tree seedlings. Therefore, the findings suggests that soil collected from underneath Prosopis juliflora canopy and the leaf litter of the species possess allelopathic properties.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Figures and Tables, The regression analysis results are provided in supplementary figures and tables section. This included the results of the logistic regression on seedling survival and height.
Acknowledgments
Special thanks to Kenya Forestry Research Institute (KEFRI), Baringo Sub-Centre for supported for technical and financial support during data collection. We are sincerely grateful to John Amiani and Reuben Lekurle for their assistance in recording and measuring the study parameters.
ACRONYMN and Abbreviation
UPC- underneath Prosopis juliflora canopy
OPC- outside Prosopis juliflora canopy
OPCLL- outside Prosopis juliflora canopy mixed with leaf litter
IAS - invasive alien species
References
- R. M. Callaway and W. M. Ridenour, “Novel weapons: Invasive success and the evolution of increased competitive ability,” Front Ecol Environ, vol. 2, no. 8, pp. 436–443, 2004. [CrossRef]
- T. Qu, X. Du, Y. Peng, W. Guo, C. Zhao, and G. Losapio, “Invasive species allelopathy decreases plant growth and soil microbial activity,” PLoS One, vol. 16, no. 2 February, pp. 1–12, 2021. [CrossRef]
- S. W. Choudhari, T. Chopde, V. P. Mane, and V. B. Shambharkar, “Allelopathic effects of Acacia nilotica (L.) Leaf leachate with emphasis on Trigonella foenum graceum L. (fenugreek),” vol. 8, no. 1, pp. 500–506, 2019.
- L. Yuan, J. M. Li, F. H. Yu, A. M. O. Oduor, and M. van Kleunen, “Allelopathic and competitive interactions between native and alien plants,” Biol Invasions, vol. 23, no. 10, pp. 3077–3090, 2021. [CrossRef]
- Y. Xu, X. Chen, L. Ding, and C. H. Kong, “Allelopathy and Allelochemicals in Grasslands and Forests,” Mar. 01, 2023, MDPI. [CrossRef]
- R. M. U. K. Gunarathne and G. A. D. Perera, “Does the invasion of Prosopis juliflora cause the die-back of the native Manilkara hexandra in seasonally dry tropical forests of Sri Lanka?,” Trop Ecol, vol. 57, no. 3, pp. 475–488, 2016.
- S. Getachew, S. Demissew, and T. Woldemariam, “Allelopathic effects of the invasive Prosopis juliflora (Sw.) DC. on selected native plant species in mIddle Awash, Southern Afar Rift of Ethiopia,” Management of Biological Invasions, vol. 3, no. 2, pp. 105–114, 2012. [CrossRef]
- J. Mungoche, O. V. Wasonga, D. Ikiror, H. Akala, C. Gachuiri, and G. Gitau, “Prosopis juliflora (sw.) DC in the drylands: A review of invasion, impacts and management in Eastern Africa,” 2025, Taylor and Francis Ltd. [CrossRef]
- A. Noha, L. Nigatu, and R. Manikandan, “Allelopathy effect of prosopis juliflora on selected grass species (Cenchrus ciliaris, paspalidium desertorum and lintonia nutans),” Journal of Phytology, vol. 13, pp. 75–78, 2021. [CrossRef]
- R. M. C. S. Ratnayake, “Allelopathic effects of Prosopis juliflora (Sw.) Dc. on selected crops and native plants,” no. November, 2021.
- A. Noha, L. Nigatu, and R. Manikandan, “Allelopathy effect of prosopis juliflora on selected grass species (Cenchrus ciliaris, paspalidium desertorum and lintonia nutans),” Journal of Phytology, vol. 13, pp. 75–78, 2021. [CrossRef]
- S. Endris, “Gene Conserve - Articles - Articles - - Articles - - Articles - Gene Conserve - Articles - Articles - Volume 7 - Issue 30 - October / December, 2008.,” Production, vol. 7, no. 27, pp. 2008–2010, 2008. [CrossRef]
- Z. H. Mehari, “The invasion of Prosopis juliflora and Afar pastoral livelihoods in the Middle Awash area of Ethiopia,” Ecol Process, vol. 4, no. 1, pp. 1–9, 2015. [CrossRef]
- A. El-Keblawy and A. Al-Rawai, “Impacts of the invasive exotic Prosopis juliflora (Sw.) D.C. on the native flora and soils of the UAE,” Plant Ecol, vol. 190, no. 1, pp. 23–35, 2007. [CrossRef]
- G. Asrat and A. Seid, “Allelopathic effect of meskit (Prosopis juliflora (Sw.) DC) aqueous extracts on tropical crops tested under laboratory conditions,” Momona Ethiopian Journal of Science, vol. 9, no. 1, p. 32, 2017. [CrossRef]
- S. Bibi, A. Bibi, M. A. Al-Ghouti, and M. H. Abu-Dieyeh, “Allelopathic Effects of the Invasive Prosopis juliflora (Sw.) DC. on Native Plants: Perspectives toward Agrosystems,” Agronomy, vol. 13, no. 2, 2023. [CrossRef]
- R. M. U. K. Gunarathne and G. A. D. Perera, “Does the invasion of Prosopis juliflora cause the die-back of the native Manilkara hexandra in seasonally dry tropical forests of Sri Lanka?,” Trop Ecol, vol. 57, no. 3, pp. 475–488, 2016.
- D. Biológica et al., “TESIS DOCTORAL CONGENERIC APPROACH FOR UNDERSTANDING INVASIVE ABILITY OF THE EXOTIC PROSOPIS JULIFLORA UNDER THE HYPER-ARID DESERT OF THE UNITED ARAB EMIRATES (UAE) Programa de Doctorado,” 2022. [Online]. Available: https://orcid.org/0000-0001-9740-894X.
- H. Shiferaw, T. Alamirew, S. Dzikiti, W. Bewket, G. Zeleke, and U. Schaffner, “Water use of Prosopis juliflora and its impacts on catchment water budget and rural livelihoods in Afar Region, Ethiopia,” Sci Rep, vol. 11, no. 1, pp. 1–14, 2021. [CrossRef]
- Z. Zhang, Y. Liu, L. Yuan, E. Weber, and M. van Kleunen, “Effect of allelopathy on plant performance: a meta-analysis,” Ecol Lett, vol. 24, no. 2, pp. 348–362, 2021. [CrossRef]
- Q. Wang, M. Kong, J. Wang, B. Gao, and X. Ping, “The Specific Impacts of Allelopathy and Resource Competition from Artemisia frigida on the Growth of Three Plant Species in Northern China,” Plants, vol. 13, no. 23, 2024. [CrossRef]
- D. Cipollini and K. Cipollini, “A review of garlic mustard (Alliaria petiolata, Brassicaceae) as an allelopathic plant,” Journal of the Torrey Botanical Society, vol. 143, no. 4, pp. 339–348, 2016. [CrossRef]
- M. Parepa and O. Bossdorf, “Testing for allelopathy in invasive plants: it all depends on the substrate!,” Biol Invasions, vol. 18, no. 10, pp. 2975–2982, 2016. [CrossRef]
- M. Parepa and O. Bossdorf, “Testing for allelopathy in invasive plants: it all depends on the substrate!,” Biol Invasions, vol. 18, no. 10, pp. 2975–2982, 2016. [CrossRef]
- Z. Zhang, Y. Liu, L. Yuan, E. Weber, and M. van Kleunen, “Effect of allelopathy on plant performance: a meta-analysis,” Ecol Lett, vol. 24, no. 2, pp. 348–362, 2021. [CrossRef] [PubMed]
- J. A. Lau et al., “Inference of allelopathy is complicated by effects of activated carbon on plant growth,” pp. 412–423, 2003.
- G. Asrat and A. Seid, “Allelopathic effect of meskit (Prosopis juliflora (Sw.) DC) aqueous extracts on tropical crops tested under laboratory conditions,” Momona Ethiopian Journal of Science, vol. 9, no. 1, p. 32, 2017. [CrossRef]
- S. Getachew, S. Demissew, and T. Woldemariam, “Allelopathic effects of the invasive Prosopis juliflora (Sw.) DC. on selected native plant species in mIddle Awash, Southern Afar Rift of Ethiopia,” Management of Biological Invasions, vol. 3, no. 2, pp. 105–114, 2012. [CrossRef]
- C. Murrell, E. Gerber, C. Krebs, M. Parepa, U. Schaffner, and O. Bossdorf, “Nvasive knotweed affects native plants through allelopathy 1,” vol. 98, no. 1, pp. 38–43, 2011. [CrossRef]
- A. Pechci’nskapechci’nska, “The Robustness of Heteroscedasticity Tests,” 2023.
- A. F. Hayes and L. I. Cai, “Using heteroskedasticity-consistent standard error estimators in OLS regression: An introduction and software implementation,” 2007.
- G. Peguero, O. R. Lanuza, R. Savé, and J. M. Espelta, “Allelopathic potential of the neotropical dry-forest tree Acacia pennatula Benth.: Inhibition of seedling establishment exceeds facilitation under tree canopies,” Plant Ecol, vol. 213, no. 12, pp. 1945–1953, 2012. [CrossRef]
- P. Lesica and T. H. Deluca, “Is tamarisk allelopathic?,” 2004.
- M. Dorning and D. Cipollini, “Leaf and root extracts of the invasive shrub, Lonicera maackii, inhibit seed germination of three herbs with no autotoxic effects,” Plant Ecol, vol. 184, no. 2, pp. 287–296, Jun. 2006. [CrossRef]
- “packer2000”.
- H. Kato-Noguchi, “Allelopathy and Allelochemicals of Imperata cylindrica as an Invasive Plant Species,” Oct. 01, 2022, MDPI. [CrossRef]
- Z. Gul, A. Akbar, and S. K. Leghari, “Elucidating Therapeutic and Biological Potential of Berberis baluchistanica Ahrendt Bark, Leaf, and Root Extracts,” Front Microbiol, vol. 13, no. March, pp. 1–14, 2022. [CrossRef]
- D. Zhang et al., “The Dynamics of Allelochemicals and Phytotoxicity in Eisenia fetida during the Decomposition of Eucalyptus grandis Litter,” Plants, vol. 13, no. 17, Sep. 2024. [CrossRef]
- M. Parepa, U. Schaffner, and O. Bossdorf, “Sources and modes of action of invasive knotweed allelopathy: the effects of leaf litter and trained soil on the germination and growth of native plants,” NeoBiota, vol. 13, pp. 15–30, 2012. [CrossRef]
- W. Huang, G. V. P. Reddy, P. Shi, J. Huang, H. Hu, and T. Hu, “Allelopathic effects of Cinnamomum septentrionale leaf litter on Eucalyptus grandis saplings,” Glob Ecol Conserv, vol. 21, p. e00872, 2020. [CrossRef]
- M. Parepa and O. Bossdorf, “Testing for allelopathy in invasive plants: it all depends on the substrate!,” Biol Invasions, vol. 18, no. 10, pp. 2975–2982, 2016. [CrossRef]
- D. Cipollini and K. Cipollini, “A review of garlic mustard (Alliaria petiolata, Brassicaceae) as an allelopathic plant,” Journal of the Torrey Botanical Society, vol. 143, no. 4, pp. 339–348, 2016. [CrossRef]
- A. I. Arroyo et al., “Evidence for chemical interference effect of an allelopathic plant on neighboring plant species: A field study,” PLoS One, vol. 13, no. 2, 2018. [CrossRef]
- S. Shannon-Firestone and J. Firestone, “Allelopathic potential of invasive species is determined by plant and soil community context,” Plant Ecol, vol. 216, no. 3, pp. 491–502, 2015. [CrossRef]
- Z. Noumi and M. Chaieb, “Allelopathic effects of Acacia tortilis (Forssk.) hayne subsp. Raddiana (Savi) brenan in North Africa,” Pak J Bot, vol. 43, no. 6, pp. 2801–2805, 2011.
- R. Kaur et al., “Community Impacts of Prosopis juliflora Invasion: Biogeographic and Congeneric Comparisons,” PLoS One, vol. 7, no. 9, 2012. [CrossRef]
- C. Wang et al., “Effects of autotoxicity and allelopathy on seed germination and seedling growth in Medicago truncatula,” Front Plant Sci, vol. 13, no. July, pp. 1–11, 2022. [CrossRef]
- C. Murrell, E. Gerber, C. Krebs, M. Parepa, U. Schaffner, and O. Bossdorf, “Nvasive knotweed affects native plants through allelopathy 1,” vol. 98, no. 1, pp. 38–43, 2011. [CrossRef]
- S. Shannon-Firestone and J. Firestone, “Allelopathic potential of invasive species is determined by plant and soil community context,” Plant Ecol, vol. 216, no. 3, pp. 491–502, 2015. [CrossRef]
Figure 1.
The interacting effect of soil type and activated carbon (AC) on survival of seedlings of resident tree species. OPC = soil from outside P. juliflora canopy; UPC = soil from underneath P. juliflora canopy; OPCLL = soil from outside P. juliflora canopy mixed with P. juliflora leaf litter.
Figure 1.
The interacting effect of soil type and activated carbon (AC) on survival of seedlings of resident tree species. OPC = soil from outside P. juliflora canopy; UPC = soil from underneath P. juliflora canopy; OPCLL = soil from outside P. juliflora canopy mixed with P. juliflora leaf litter.
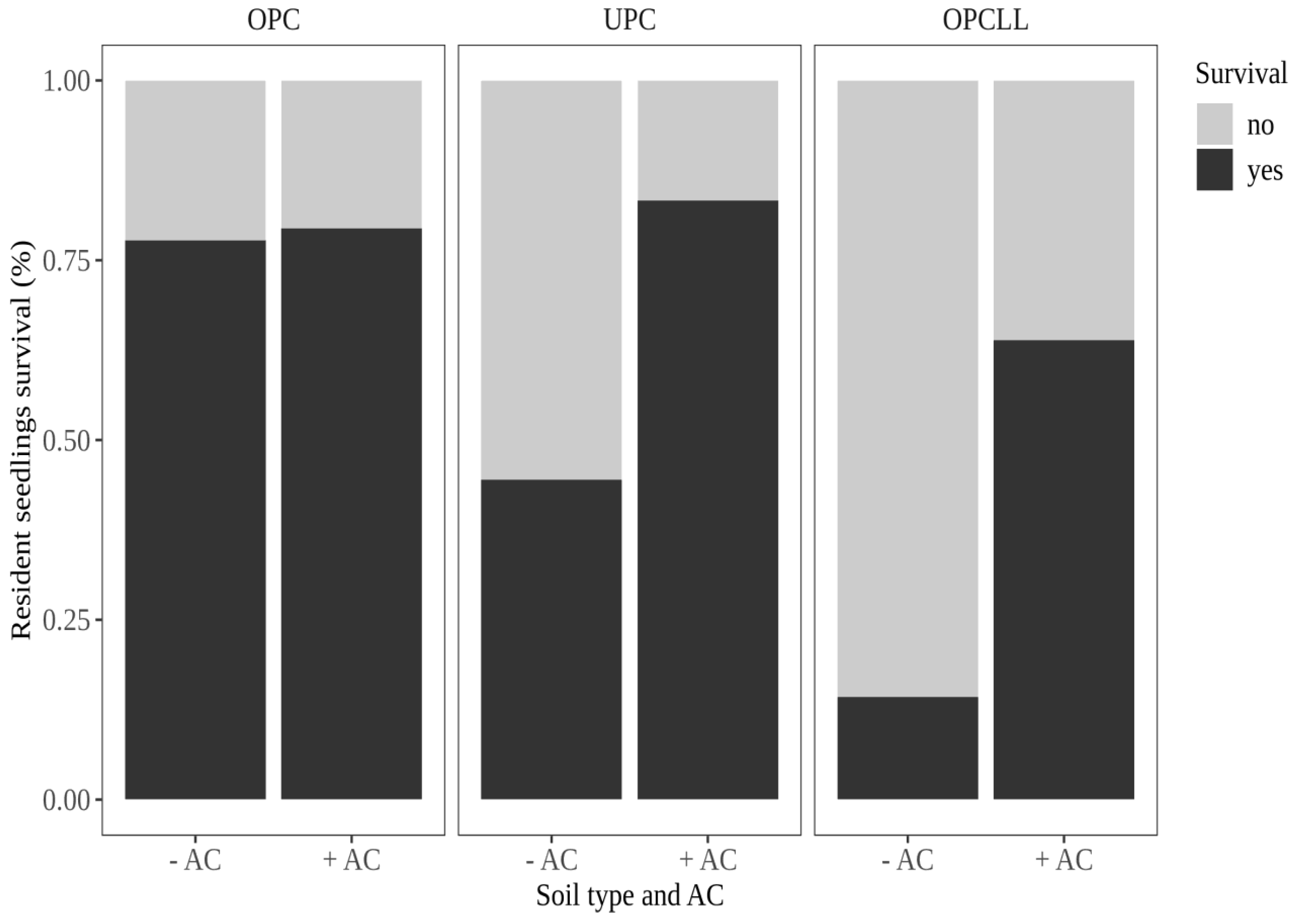
Figure 2.
Effect of soil type and activated carbon (AC) on seedling height of resident tree species. OPC = soil from outside P. juliflora canopy; UPC = soil from underneath P. juliflora canopy; OPCLL = soil from outside P. juliflora canopy mixed with P. juliflora leaf litter.
Figure 2.
Effect of soil type and activated carbon (AC) on seedling height of resident tree species. OPC = soil from outside P. juliflora canopy; UPC = soil from underneath P. juliflora canopy; OPCLL = soil from outside P. juliflora canopy mixed with P. juliflora leaf litter.
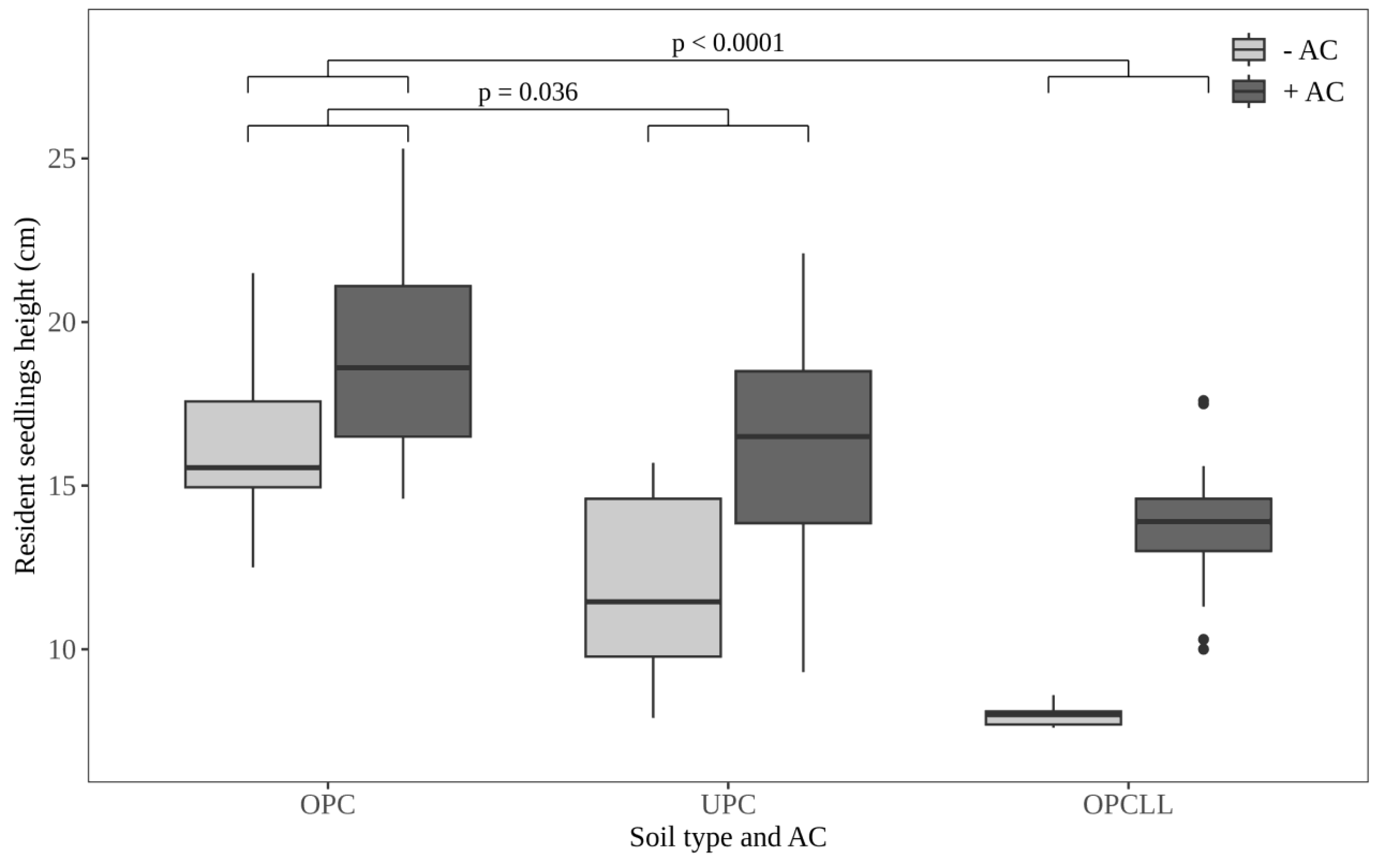
Figure 3.
Effect of soil type and competition on seedling height of resident tree species. OPC = soil from outside P. juliflora canopy; UPC = soil from underneath P. juliflora canopy; OPCLL = soil from outside P. juliflora canopy mixed with P. juliflora leaf litter.
Figure 3.
Effect of soil type and competition on seedling height of resident tree species. OPC = soil from outside P. juliflora canopy; UPC = soil from underneath P. juliflora canopy; OPCLL = soil from outside P. juliflora canopy mixed with P. juliflora leaf litter.
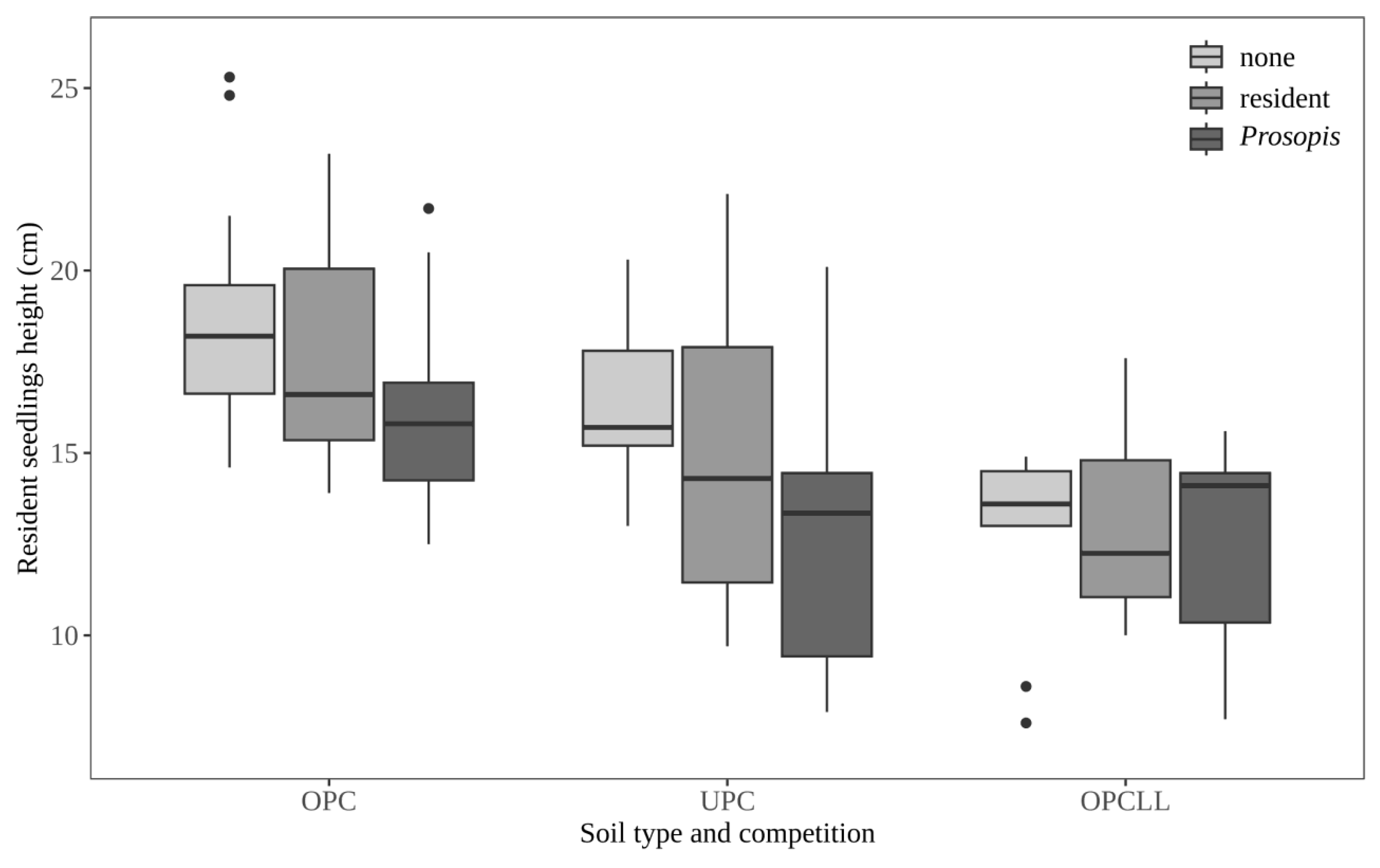
Figure 4.
The interacting effect of competition and activated carbon (AC) on seedling height of resident tree species. None = resident seedlings were grown without competitor; resident = seedlings of resident tree species were grown in competition with a seedling of a different resident tree species; Prosopis = seedlings of resident tree species were grown in competition with a seedling of P. juliflora.
Figure 4.
The interacting effect of competition and activated carbon (AC) on seedling height of resident tree species. None = resident seedlings were grown without competitor; resident = seedlings of resident tree species were grown in competition with a seedling of a different resident tree species; Prosopis = seedlings of resident tree species were grown in competition with a seedling of P. juliflora.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



