
Submitted:
08 December 2025
Posted:
10 December 2025
You are already at the latest version
Abstract
Multiple myeloma is a malignant tumor disease that affects plasma cells and is one of the most common tumors of lymphoid origin. In the process of oncogenesis, that is, the formation of a tumor, there is a significant change in the immune balance in the body, which leads to the suppression of the immune response to the tumor. This suppression is one of the reasons why the tumor can progress and cause serious health problems for the patient. One of the key factors that affect immune balance is innate lymphoid cells (ILCs). ILCs play an important role in regulating the immune response and can both promote and hinder the development of tumor processes, depending on their functional state and interaction with other cells of the immune system. It has been shown that the number of immature CD5+ILC2s cells in the peripheral blood of patients with multiple myeloma is comparable to that of healthy individuals. However, autologous hematopoietic stem cell transplantation in MM patients leads to an increase in the relative number of CD5+ILC2s.
Keywords:
innate lymphoid cells
; immune balance
; innate immunity
; oncology
; multiple myeloma
1. Introduction
The introduction lymphoid cells of innate immunity (ILCs), capable of synthesizing cytokines, were discovered relatively recently in comparison with other immunocompetent cells - at the end of the noughties of the twenty-first century in several laboratories independently of each other [1,2,3,4]. Due to the fragmentation of studies and the relatively short period of time that has passed since the discovery, in addition to the currently generally recognized nomenclature for groups of cytokine-producing ILCs (ILC1s, ILC2s, ILC3s), other names for these cells are currently used – nuocytes (for ILC2), natural helpers [5,6,7].
Helper ILCs produce a similar cytokine profile to T-helper cells, as T-helper cells are divided into the main subsets of Th1, Th2, and Th17, with ILC1s, ILC2s, and ILC3s considered their analogues, respectively [8,9]. Unlike T-helper cells, ILCs lack a T-cell receptor and are unable to recognize antigens. Furthermore, helper ILCs are predominantly found in tissues, while their numbers in peripheral blood are extremely low. Due to their tissue location, helper ILCs quickly respond to changes in the tissue microenvironment and are capable of modulating the immune response.
ILC1s are known to produce interferon gamma (IFNγ), which exhibits pronounced inhibiting activity on myeloma cell proliferation [10]. In a study by Kellermeier et al., neutralization of IFNγ significantly increased myeloma development and progression in C57Bl/6 mice, demonstrating the importance of this pathway in early disease control [11].In turn, interleukin-4 (IL-4), one of the main cytokines for ILC2s, was also able to suppress the growth of malignant plasma cells in vitro via suppression of IL-6 expression [12]. However, another study demonstrated opposite results: IL-4 not only failed to suppress tumor plasma cell proliferation but also caused a slight increase in the number of cells in S-phase compared to baseline and increased the number of chromosomal abnormalities in plasma cells from patients with MM, which may serve as a basis for the development of malignancy [13]. Furthermore, IL-4 has been shown to promote the differentiation of myeloma cell precursors in multiple myeloma in vitro [14].
Like different types of T helper cells, different subsets of ILCs are capable of both promoting tumor growth and exerting an antitumor effect [15]. In the case of hematologic malignancies, ILCs, on the one hand, enhance the immunosuppressive effect of the tumor microenvironment, and on the other hand, promote tissue homeostasis and reparation, which is especially important in the case of antitumor therapy [16]. The antitumor therapy, be it chemotherapy or radiotherapy, as well as ablation of hematopoietic stem cells before transplantation, leads to a decrease in helper ILCs. The restoration of helper ILC numbers, unlike NK cells, is slow and comparable to T cell reconstitution [16,17,18].
In a mouse model of multiple myeloma, tumor growth was associated with phenotypic and functional changes in bone marrow ILC2s, characterized by increased expression of mature ILC2 markers GATA3, CD117, CD25, and KLRG1, and decreased levels of Sca1 and CD127, as well as reduced cytokine production upon cell stimulation with IL-2/IL-33 [19]. These phenotypic changes might indicate a progressive maturation of ILC2s in the presence of myeloma. Administration of IL-33, one of the cytokines capable of activating ILC2s, induces apoptosis of circulating KLRG1hi ILC2s in this model, which inhibits protective type 1 immune responses against MM.
In patients with plasma cell dyscrasias, a decrease in the number of ILC2s in the bone marrow was noted with a simultaneous increase in the circulating subset [20,21]. It is known that in monoclonal gammopathy of unknown genesis (MGUS), a condition preceding multiple myeloma (MM), an increase in the proportion of ILC2s in the peripheral blood was observed, while in the bone marrow, on the contrary, the proportion of ILC1s increased, but the functional activity of ILC1s and ILC2s, determined by the intracellular content of cytokines, was reduced [21]. In patients with MGUS, ILC2s demonstrated the ability to secrete IL13, which was not observed in patients with asymptomatic MM. In patients with a recently established diagnosis of MM, a decrease in the proportion of ILC2s among Lin–CD127+ cells in the peripheral blood was observed before therapy [22]. In another study, in a group of patients with MM who achieved remission after therapy, the proportion of ILC2s among Lin–CD127+ in the blood was comparable to the values in healthy volunteers [23], while ILC1s and ILC3s were reduced. Presumably, the differences may be associated with the therapy.
Despite significant advances in modern therapies for MM, autologous hematopoietic stem cell transplantation (auto-HSCT) with hi-dose chemiotherapy remains one of the standard methods [24,25]. Helper ILCs restoration after HSCT can occur in various ways [16]. Circulating ILCs are probably derived from common lymphoid progenitor cells in the bone marrow and possibly from mature ILCs co-transfused with the graft. ILCs are considered tissue resident, depending on local self-renewal. However, given the plasticity of ILCs and their ability to migrate, the peripheral blood after HSCT may contain both immature ILCs derived from the hematopoietic stem cell of the graft and mature ILCs from the graft or from various non-lymphoid tissues.
CD5 is a cell surface protein expressed on T lymphocytes that functions as an inhibitor of antigen receptor signaling [26,27,28]. CD5 was initially considered a T lymphocyte marker and was even used in some studies to exclude T cells in ILC analysis [26,29,30,31]. However, ILC1s were also found to express T lymphocyte-associated molecules, including CD5 [26,32,33]. Furthermore, CD5+ ILCs were found to be present in human thymus and cord blood [33]. Moreover, CD5+ ILCs were functionally immature but could further differentiate into mature CD5- cytokine-secreting ILCs [34]. Recently, there has been evidence that CD5+ ILC2s can also be formed in the thymus [35]. The work of Alisjahbana at al found that CD5+ ILCs had a distinct ontogeny compared to conventional CD5- ILCs because they first appeared in the thymus, spleen and liver rather than in the bone marrow after transplantation of MISTRG mice with human CD34+ hematopoietic stem and progenitor cells [36].
It has been shown that ILC2 types expressing CD5 on their surface are mainly formed in different ways, while the role of these cells and possible participation in pathological processes are not yet fully understood [35]. Here, we evaluate the hypothesis that auto-HSCT performed in MM leads to changes in subpopular composition and CD5 expression on peripheral blood ILC.
2. Materials and Methods
2.1. Sample Collection
Blood samples from conditionally healthy volunteers (n=9) and patients with multiple myeloma (MM) undergoing treatment at the Clinic of Immunopathology of the Research Institute of Fundamental and Clinical Immunogy (n=19) were used as study material.
Study inclusion criteria: patients with MM aged 18–65 years; clinical stages II and III of the disease according to the Durie–Salmon classification; patients in remission at the time of inclusion; written informed consent. Control group inclusion criteria: healthy donors aged 18–65 years; absence of autoimmune, oncological, and chronic recurrent viral infections. Blood samples were collected from apparently healthy individuals once.Auto-HSCT was performed using peripheral blood stem cells obtained by apheresis. Cyclophosphamide (2–4 g/m2) was used for mobilization, followed by granulocyte colony-stimulating factor (5 μg/kg/day) until a concentration of 104 CD34+CD45+ hematopoietic cells/mL was achieved in the blood of patients with MM. Apheresis was performed in 1-2 procedures on ASTEC 204 (Fresenius, Germany) and Spectra LRS 07 (COBE BCT, Lakewood, Colorado, United States ) blood cell separators to obtain 2.0×106 CD34+CD45+ cells/kg. The resulting cells were frozen and stored until use. High-dose chemotherapy with melphalan followed by HSCT was performed 2-6 months after separation. Blood samples were obtained twise: first in 1 day before high-dose chemotherapy and second after autoHSCT after recovery from leukopenia (day +12–+16, leukocytes >1×109L).
2.2. Cell Isolation
Peripheral blood mononuclear cells (PBMCs) were isolated from heparinized venous blood of healthy donors or patients using the ficoll-urographin density gradient centrifugation method (1.077 g/cm3). Briefly, 5 ml of fresh heparinized venous blood from healthy donors was layered in a test tube of 3 ml of ficoll-urographin (1.077 g/cm3) (Biolot, Russia). The tubes were centrifuged at 3,000 rpm for 25 minutes. After centrifugation, mononuclear rings were selected in separate tubes followed by double washing in 10 mL of PBS + Na2 EDTA (0,02% Na2EDTA in PBS) and centrifugating at 1,200 rpm for 10 minutes. Next, cells were stained with monoclonal antibodies for 24 hours and the data were analyzed using flow cytometry.
2.3. Cell Staining and Flow Cytometry
To assess ILCs, isolated PBMCs were stained with monoclonal antibodies conjugated with fluorochromes: anti-Lineage (CD2/3/14/16/19/20/56/235a) (eBioscience, San Diego, CA), antiCD11c (BioLegend, San Diego, CA, USA), anti-FceR1 alpha-FITC (eBioscience, San Diego, CA), and anti-CD294-PE, anti-CD127-PerCP/Cy5.5, anti-CD117-APC and anti-CD5-BV421 (all from BioLegend, San Diego, CA, USA). The total number of ILC cells was defined as Lin-CD127+, since these cells do not carry linear markers, but have an alpha chain of the IL-7 receptor on their surface. ILC2 was determined by the presence of CD294 (CRTH2) on the surface of the cell (Figure 1). ILC1 were defined as CD294-CD117-, and ILC3 as CD294-CD117+.To the labelled tubes, antibody cocktail was added to the sample and incubated in the dark for 15 min. The sample was then directly centrifuged at 1,200 rpm for 5 min. After decanting the supernatant, 1 ml of staining buffer (0,5% fetal calf serum and 0,02% Na2EDTA in PBS) was added for washing. The sample was centrifuged at 1,200 rpm for 5 min. Again after decanting the supernatant, the final suspension in 0.2 ml of staining buffer was ready for acquisition. The phenotype of the cells was analyzed using the LongCyte flow cytometer (Challenbio, China). At least 100,000 events were acquired and analyzed per tube.
2.4. Statistical Analysis
Statistical analysis of the results was performed using GraphPadPrizm 9.0.0 (GraphPad Software, Inc., USA). The nonparametric Kruskal-Wallis test with subsequent post-hoc analysis was used to assess differences between groups. The Mann-Whitney test was used to compare parameters between the control group and the patient groups before and after HSCT. The Wilcoxon test was used to compare patient parameters before and after therapy. Results are presented as median with interquartile range. Results were considered statistically significant at p < 0.05.
3. Results
3.1. Subpopulation Composition of ILC Before and After Auto-HSCT.
In patients with MM, the proportion of ILCs among PBMCs was reduced before auto-HSCT compared to healthy volunteers (Figure 2). However, in patients with MM after auto-HSCT, no significant differences were observed compared to healthy controls. This change is not due to an increase in the absolute number of ILCs, but rather occurs due to a change in proportions against the background of lymphopenia and a more pronounced decrease in the number of T and B lymphocytes after auto-HSCT.
According to the literature, different studies report different numbers of ILC2 subsets in patients with MM and MGUS, which may depend on the stage of the disease and the therapy [21,22,23]. In this study, patients before HSCT are in complete or very good partial remission after therapy, and the number of ILC2 in patients is increased compared to healthy controls (Figure 3).
Moreover, in patients with MM, the relative number of ILC1 before HSCT decreases, while ILC3 does not change compared to healthy individuals. We have previously shown that the proportion of ILC2 increases in patients with MM in complete or partial remission [21]. AutoHSCT leads to an increase in the level of ILC1, a decrease in the proportion of ILC2, and does not affect the number of ILC3. The relative number of different ILC subpopulations in patients after HSCT does not differ significantly from that in healthy controls. Therefore, in patients with MM, there are differences in the subpopulation composition before auto-HSCT with high-dose chemotherapy, but none after high-dose chemotherapy and auto-HSCT.
3.2. Relative Number of CD5+ILC2 Before and After Auto-HSCT.
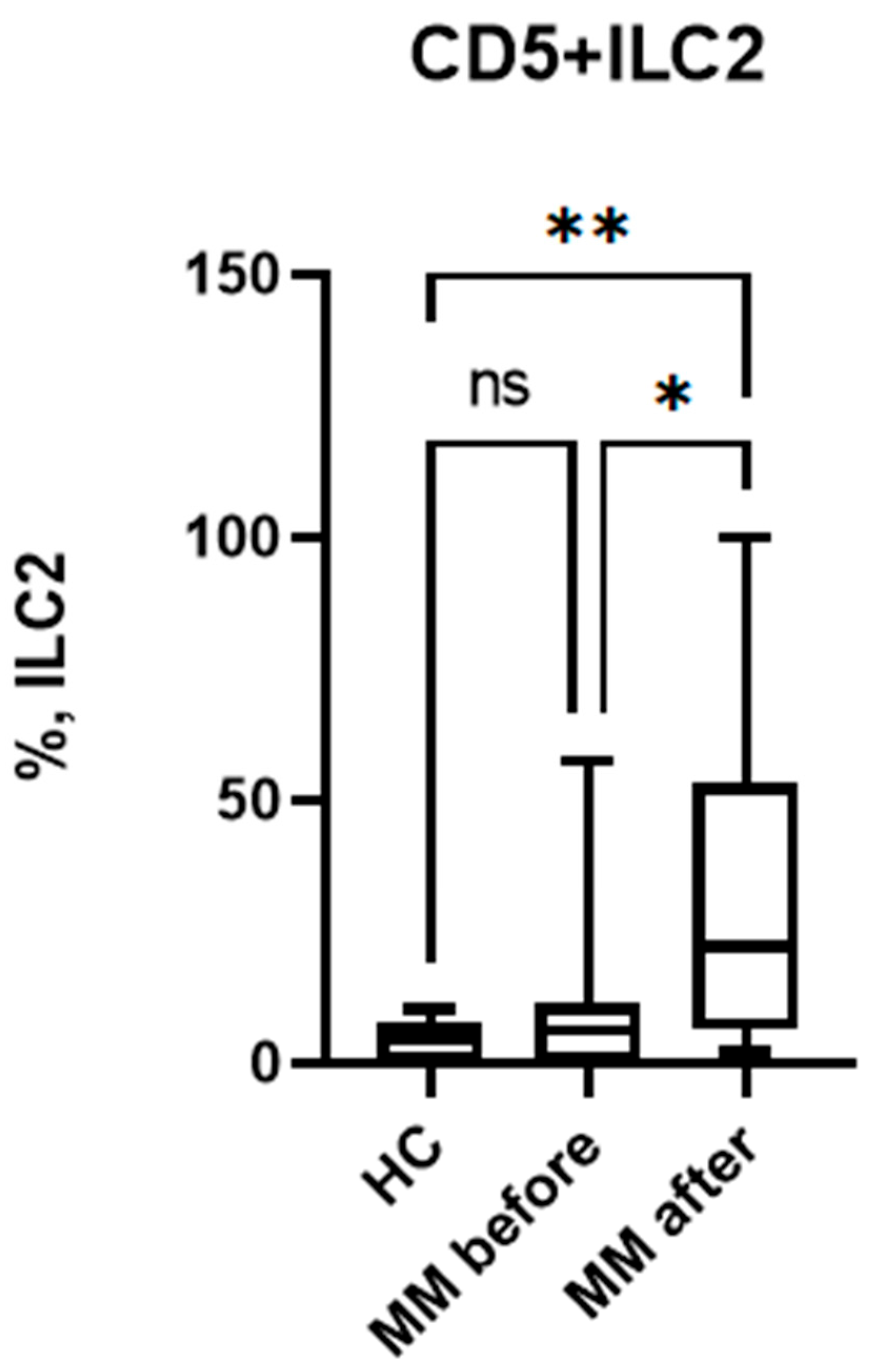
In the course of the study, we determined the content of immature CD5+ILC2 in the peripheral blood of MM patients. As a result, it was found that the proportion of these cells among ILC2 is approximately 5% and is almost identical in patients with MM and conditionally healthy individuals (Figure 4).
The next step was a comparative analysis of CD5+ ILC levels before and after auto-HSCT. It was shown that the proportion of immature CD5+ ILC2 increases sharply after auto-HSCT (Figure 4) both compared with healthy controls and with the values in patients before auto-HSCT. The increase in immature ILCs may be aimed at replenishing the ILC pool during immune reconstitution after recovery from leukopenia.
4. Discussion
Currently, it is known that there are 3 main types of cell-mediated effector immunity, namely type 1, type 2 and type 3 [37]. Type 1 immunity consists of T-bet+ IFN-γ–producing group 1 ILCs (ILC1 and natural killer cells), CD8+ cytotoxic T cells (TC1), and CD4+ Th1 cells, which protect against intracellular microbes through activation of mononuclear phagocytes. Type 2 immunity consists of GATA-3+ ILC2s, TC2 cells, and Th2 cells producing IL-4, IL-5, and IL-13, which induce mast cell, basophil, and eosinophil activation, as well as IgE antibody production, thus protecting against helminthes and venoms. Type 3 immunity is mediated by retinoic acid–related orphan receptor γt+ ILC3s, TC17 cells, and Th17 cells producing IL-17, IL-22, or both, which activate mononuclear phagocytes but also recruit neutrophils and induce epithelial antimicrobial responses, thus protecting against extracellular bacteria and fungi.
Traditionally, type 1 immunity is considered to be antitumor, suppressing tumor growth and killing tumor cells by activating NK and T cells [38]. Type 2 immunity is key in tissue regeneration processes and has a pro-tumor effect. However, it has been established that classical type 2 immune cytokines—IL-4, IL-5, and IL-13—may play conflicting roles in cancer development [38,39]. Moreover, type 1 cytokines, IFNγ and TNF, have also been found to have pro-tumor effects under certain circumstances, further complicating the overall picture. Thus, T1 and T2 cytokines can either stimulate or suppress tumor development depending on the context.
In our study, we identified elevated levels of IL-4-producing ILC2s in patients in remission before auto-HSCT. According to published data, serum IL-4 levels were low at diagnosis in 75% of patients with MM (median 4 pg/ml) and then increased during remission (median 25 pg/ml). IL-4 levels remained stable throughout the disease in chemotherapy-resistant patients [40]. MM patients with IL-4 levels above 3.382 pg/ml also had higher overall survival [41], but IL-4 levels had no prognostic significance.After recovery from leukopenia, the ratio of ILC subsets changed: the proportion of ILC2s decreased, and the number of ILC1s increased. No significant differences were observed between the ILC ratios in patients with MM after HSCT and in healthy individuals. Therefore, HSCT may normalize the ratio between different ILC subsets.
Previous studies have shown that Th2 cell cytokines like IL-4 decreased, while Th1 cell cytokines like IFN-γ increased after auto-HSCT [42,43,44]. Therefore, the change in the T1/T2 balance after auto-HSCT may be associated with a change in the ILC1/ILC2 balance. Since IFNγ can inhibit the growth and proliferation of myeloma cells, an increase in the proportion of its producers, ILC1, can enhance the antitumor response after auto-HSCT.
However, there is evidence of possible changes in the functional activity of ILC2s in patients with MM. In patients with MGUS and MM, ILC2s in the peripheral blood acquired cytotoxic activity towards tumor cells, which may be associated with the differentiation of ILC2s into ILC1-like cells, the antitumor activity of which may increase due to the polarization of the immune response towards T1 and the acquisition of cytotoxic activity.
The mechanisms underlying changes in ILC balance during auto-HSCT are currently unknown and require further research. It is possible that the rate of subpopulation regeneration varies, partly due to replenishment from different sources and the different subpopulation composition of ILCs in these organs and tissues. Alternatively, polarization toward T1 occurs deliberately due to changes in internal factors. In this case, enhanced transdifferentiation of ILC2s into ILC1-like cells is possible, similar in mechanism to that observed in the mouse model of allo-HSCT with ILC2 co-transplantation [45].
We then determined the content of immature CD5+ ILC2 in the peripheral blood of patients with MM. We found that the proportion of these cells among ILC2 cells was nearly identical in both patients with MM and healthy controls. Therefore, neither the disease itself nor previous therapy to achieve remission affected the relative number of CD5+ ILC2 cells in the peripheral blood of patients with MM.
One possible source of CD5+ILC2 is the thymus [34,35]. It is known that in MM, thymic atrophy can develop, which, in turn, leads to a decrease in the formation of a number of cells in the body, primarily various subpopulations of T-lymphocytes [46]. Such disorders entail a change in the ratio of effector T lymphocytes and T-regulatory cells, and, as a result, a violation of the immune balance. CD5+ILC2 does not change, which may indicate that in MM only the formation of T-lymphocytes is impaired, while the differentiation of ILC2 is maintained at the initial level corresponding to that in healthy individuals. Alternatively, these cells in MM are replenished from a source other than the thymus.
The HSCT graft consists of hematopoietic stem cells and other different cells, include ILC. Circulating in blood ILC originate from common lymphoid progenitor (CLP) cells located in the bone marrow, as well as from mature ILC that are co-transferred with the graft during transplantation [16]. In the case of thymus-derived ILC, they will most likely be formed from lymphoid precursors differentiated from the graft stem cell. Also, ILCs are considered tissue-resident cells that rely on local self-renewal within tissues. However, since donor-derived ILC have been observed in non-lymphoid tissues after allog- HSCT, it suggests that circulating ILC, may also migrate to tissues. Conversely, tissue-resident ILCs can enter the bloodstream. Therefore, further studies are needed to determine the source of the increase in CD5+ILC after auto-HSCT.
It is known that CD5+ILC cells are found intravascularly in the organ after transplantation, while CD5-ILC are located outside the vasculature within the organ itself [36]. Moreover, CD5+ILC vascular CD5+CD7+ ILC may perform functions similar to blood monocytes that patrol blood vessels and gain access to tissue niches during altered organ homeostasis. This subpopulation may play an important role in organ and tissue regeneration after high-dose chemotherapy administered before auto-HSCT.
The increase in the proportion of immature CD5+ ILC2 after HSCT may be associated, on the one hand, with a decrease in the functional activity of the cells and, consequently, a decrease in the intensity of the T2 immune response. On the other hand, immature ILC2 may exhibit greater plasticity and more effectively transdifferentiate into ILC1-like cells, which exhibit cytotoxic activity and are capable of effectively participating in the antitumor immune response.
5. Conclusions
Thus, before HSCT, patients with MM exhibit a change in the ILC population composition, with an increased proportion of ILC2s and a decrease in ILC1s. However, no significant differences in the relative number of immature CD5+ ILC2s cells were found in patients with multiple myeloma and conditionally healthy donors. Autologous HSCT, in turn, results in an ILC ratio similar to that of the control group, namely, an increased relative number of ILC1s and a decrease in ILC2s.Also, autologous HSCT increases the proportion of immature CD5+ ILC2s in patients with MM. Therefore, autologous HSCT in MM may, on the one hand, weaken the function of helper ILCs, leading to a reduction in T1/T2/T17 immune responses and impaired tissue reparative processes. On the other hand, an increased proportion of immature ILC2s may enhance ILC2 plasticity and transdifferentiation, ncluding into cytotoxic ILC1-like cells, which may contribute to a stronger antitumor response. Thus, the impact of autologous HSCT on the ratio of different ILC populations and the antitumor response requires further study.
Author Contributions
Conceptualization, V.A.K.. and E.A.P.; writing—original draft preparation, E.A.P.; investigation – E.A.P., O.S.B., I.P.S. and V.V.D.; writing—review and editing, E.A.P and V.A.K; funding acquisition, E.A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Russian Science Foundation according to research project No.25-15-00558.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Institutional Local Ethical Committee of Research Institute of Fundamental and Clinical Immunology (protocol No. 145, 04 April 2024).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Satoh-TakayamaN, VosshenrichCA, Lesjean-Pottier S, Sawa S, Lochner M, Rattis F. et al. Microbial flora drives interleukin 22 production in intestinal NKp46 +cells that provide innate mucosal immune defense. Immunity 2008; 29:958–70 . [CrossRef]
- Luci C, Reynders A, Ivanov II, Cognet C, Chiche L, Chasson L. et al. Influence of the transcription factor RORgammat on the development of NKp46(+) cell populations in gut and skin. Nat Immunol 2009; 10:75–82. [CrossRef]
- Moro K, Yamada T, Tanabe M, Takeuchi T, Ikawa T, Kawamoto H, et al. Innate production of T(H)2 cytokines by adipose tissue-associated c-Kit(+)Sca-1(+) lymphoid cells. Nature 2010; 463:540–4. [CrossRef]
- Neill DR, Wong SH, Bellosi A, Flynn RJ, Daly M, Langford TK, et al. Nuocytes represent a new innate effector leukocyte that mediates type-2 immunity. Nature 2010; 464:1367–70. [CrossRef]
- Ji X., Bie Q., Liu Y., Chen J., Su Z., Wu Y., Ying X., Yang H., Wang S., Xu H. Increased frequencies of nuocytes in peripheral blood from patients with Graves' hyperthyroidism. Int J Clin Exp Pathol. 2014 Oct 15; 7(11):7554-62.
- Liu J, Wu J, Qi F, Zeng S, Xu L, Hu H, Wang D, Liu B. Natural helper cells contribute to pulmonary eosinophilia by producing IL-13 via IL-33/ST2 pathway in a murine model of respiratory syncytial virus infection. Int Immunopharmacol. 2015 Sep; 28(1):337-43. [CrossRef]
- Spits H, Artis D, Colonna M, Diefenbach A, Di Santo JP, Eberl G. et al. Innate lymphoid cells--a proposal for uniform nomenclature. Nat Rev Immunol. 2013 Feb; 13(2):145-9. [CrossRef]
- Mehrani Y, Morovati S, Keivan F, Tajik T, Forouzanpour D, Shojaei S, Bridle BW, Karimi K. Dendritic Cells and Their Crucial Role in Modulating Innate Lymphoid Cells for Treating and Preventing Infectious Diseases. Pathogens. 2025 Aug 8;14(8):794. [CrossRef]
- Jan-Abu SC, Kabil A, McNagny KM. Parallel origins and functions of T cells and ILCs. Clin Exp Immunol. 2023 Jul 5;213(1):76-86. [CrossRef]
- Palumbo A, Bruno B, Boccadoro M, Pileri A. Interferon-gamma in multiple myeloma. Leuk Lymphoma. 1995 Jul;18(3-4):215-9. PMID: 8535185. [CrossRef]
- Kellermayer Z, Tahri S, de Jong MME, Papazian N, Fokkema C, Stoetman ECG, Hoogenboezem R, van Beek G, Sanders MA, Boon L, Den Hollander C, Broijl A, Sonneveld P, Cupedo T. Interferon gamma-mediated prevention of tumor progression in a mouse model of multiple myeloma. Hemasphere. 2024 Dec 2;8(12):e70047. [CrossRef]
- Herrmann F, Andreeff M, Gruss HJ, Brach MA, Lübbert M, Mertelsmann R. Interleukin-4 inhibits growth of multiple myelomas by suppressing interleukin-6 expression. Blood. 1991 Oct 15;78(8):2070-4.
- Hernández JM, Gutiérrez NC, Almeida J, García JL, Sánchez MA, Mateo G, Ríos A, San Miguel JF. IL-4 improves the detection of cytogenetic abnormalities in multiple myeloma and increases the proportion of clonally abnormal metaphases. Br J Haematol. 1998 Oct;103(1):163-7. [CrossRef]
- Sawamura M, Murakami H, Tamura J, Matsushima T, Sato S, Naruse T, Tsuchiya J. Tumour necrosis factor-alpha and interleukin 4 promote the differentiation of myeloma cell precursors in multiple myeloma. Br J Haematol. 1994 Sep;88(1):17-23. [CrossRef]
- Sugimura R, Wang CY. The Role of Innate Lymphoid Cells in Cancer Development and Immunotherapy. Front Cell Dev Biol. 2022 Apr 26;10:803563. [CrossRef]
- Blom B, van Hoeven V, Hazenberg MD. ILCs in hematologic malignancies: Tumor cell killers and tissue healers. Semin Immunol. 2019 Feb;41:101279. [CrossRef]
- Vély F, Barlogis V, Vallentin B, Neven B, Piperoglou C, Ebbo M, Perchet T, Petit M, Yessaad N, Touzot F, Bruneau J, Mahlaoui N, Zucchini N, Farnarier C, Michel G, Moshous D, Blanche S, Dujardin A, Spits H, Distler JH, Ramming A, Picard C, Golub R, Fischer A, Vivier E. Evidence of innate lymphoid cell redundancy in humans. Nat Immunol. 2016 Nov;17(11):1291-1299. [CrossRef]
- Laurie, S.J., Foster, J.P., Bruce, D.W. et al. Type II innate lymphoid cell plasticity contributes to impaired reconstitution after allogeneic hematopoietic stem cell transplantation. Nat Commun 15, 6000 (2024). [CrossRef]
- Guillerey C, Stannard K, Chen J, Krumeich S, Miles K, Nakamura K, Smith J, Yu Y, Ng S, Harjunpää H, Teng MW, Engwerda C, Belz GT, Smyth MJ. Systemic administration of IL-33 induces a population of circulating KLRG1hi type 2 innate lymphoid cells and inhibits type 1 innate immunity against multiple myeloma. Immunol Cell Biol. 2021 Jan;99(1):65-83. [CrossRef]
- Szudy-Szczyrek A, Ahern S, Kozioł M, Majowicz D, Szczyrek M, Krawczyk J, Hus M. Therapeutic Potential of Innate Lymphoid Cells for Multiple Myeloma Therapy. Cancers (Basel). 2021 Sep 26;13(19):4806. [CrossRef]
- Kini Bailur J, Mehta S, Zhang L, Neparidze N, Parker T, Bar N, et al. Changes in bone marrow innate lymphoid cell subsets in monoclonal gammopathy: target for IMiD therapy. Blood Adv. 2017; 1 (25): 2343–7. [CrossRef]
- Drommi F, Calabr– A, Pezzino G, Vento G, Freni J, Costa G, et al. Multiple Myeloma Cells Shift the Fate of Cytolytic ILC2s Towards TIGIT-Mediated Cell Death. Cancers. 2025; 17 (2): 263. Available from: . [CrossRef]
- Pashkina EA, Boeva OS, Borisevich VI, Abbasova VS, Skachkov IP, Lazarev YA, et al. Assessment of the features of innate lymphoid cells in patients with multiple myeloma. Bulletin of RSMU. 2025; (1): 42–6. [CrossRef]
- Hughes CFM, Shah GL, Paul BA. Autologous hematopoietic stem cell transplantation for multiple myeloma in the age of CAR T cell therapy. Front Oncol. 2024 Mar 27;14:1373548. [CrossRef]
- Rocchi S, Zannetti BA, Marconi G, Lanza F. Multiple Myeloma: The Role of Autologous Stem Cell Transplantation in the Era of Immunotherapy. Cells. 2024 May 16;13(10):853. [CrossRef]
- Alisjahbana A, Gao Y, Sleiers N, Evren E, Brownlie D, von Kries A, Jorns C, Marquardt N, Michaëlsson J, Willinger T. CD5 Surface Expression Marks Intravascular Human Innate Lymphoid Cells That Have a Distinct Ontogeny and Migrate to the Lung. Front Immunol. 2021 Nov 18;12:752104. [CrossRef]
- Voisinne G, Gonzalez de Peredo A, Roncagalli R. CD5, an Undercover Regulator of TCR Signaling. Front Immunol (2018) 9:2900. doi: 10.3389/fimmu.2018.02900.
- Burgueno-Bucio E, Mier-Aguilar CA, Soldevila G. The Multiple Faces of CD5. J Leukoc Biol (2019) 105(5):891–904. [CrossRef]
- Simoni Y, Fehlings M, Kløverpris HN, McGovern N, Koo SL, Loh CY, Lim S, Kurioka A, Fergusson JR, Tang CL, Kam MH, Dennis K, Lim TKH, Fui ACY, Hoong CW, Chan JKY, Curotto de Lafaille M, Narayanan S, Baig S, Shabeer M, Toh SES, Tan HKK, Anicete R, Tan EH, Takano A, Klenerman P, Leslie A, Tan DSW, Tan IB, Ginhoux F, Newell EW. Human Innate Lymphoid Cell Subsets Possess Tissue-Type Based Heterogeneity in Phenotype and Frequency. Immunity. 2017 Jan 17;46(1):148-161. Epub 2016 Dec 13. Erratum in: Immunity. 2018 May 15;48(5):1060. doi: 10.1016/j.immuni.2018.04.028. [CrossRef]
- Yudanin NA, Schmitz F, Flamar AL, Thome JJC, Tait Wojno E, Moeller JB, Schirmer M, Latorre IJ, Xavier RJ, Farber DL, Monticelli LA, Artis D. Spatial and Temporal Mapping of Human Innate Lymphoid Cells Reveals Elements of Tissue Specificity. Immunity. 2019 Feb 19;50(2):505-519.e4. [CrossRef]
- hen L, Youssef Y, Robinson C, Ernst GF, Carson MY, Young KA, Scoville SD, Zhang X, Harris R, Sekhri P, Mansour AG, Chan WK, Nalin AP, Mao HC, Hughes T, Mace EM, Pan Y, Rustagi N, Chatterjee SS, Gunaratne PH, Behbehani GK, Mundy-Bosse BL, Caligiuri MA, Freud AG. CD56 Expression Marks Human Group 2 Innate Lymphoid Cell Divergence from a Shared NK Cell and Group 3 Innate Lymphoid Cell Developmental Pathway. Immunity. 2018 Sep 18;49(3):464-476.e4. [CrossRef]
- Björklund ÅK, Forkel M, Picelli S, Konya V, Theorell J, Friberg D, Sandberg R, Mjösberg J. The heterogeneity of human CD127(+) innate lymphoid cells revealed by single-cell RNA sequencing. Nat Immunol. 2016 Apr;17(4):451-60. [CrossRef]
- Nagasawa M, Germar K, Blom B, Spits H. Human CD5+ Innate Lymphoid Cells Are Functionally Immature and Their Development from CD34+ Progenitor Cells Is Regulated by Id2. Front Immunol. 2017 Aug 31;8:1047. [CrossRef]
- Shin SB, McNagny KM. ILC-You in the Thymus: A Fresh Look at Innate Lymphoid Cell Development. Front Immunol. 2021 May 6;12:681110. [CrossRef]
- Cupedo T. ILC2: at home in the thymus. Eur J Immunol. 2018 Sep; 48(9):1441-1444. [CrossRef]
- Alisjahbana A, Gao Y, Sleiers N, Evren E, Brownlie D, von Kries A, Jorns C, Marquardt N, Michaëlsson J, Willinger T. CD5 Surface Expression Marks Intravascular Human Innate Lymphoid Cells That Have a Distinct Ontogeny and Migrate to the Lung. Front Immunol. 2021 Nov 18;12:752104. [CrossRef]
- Annunziato F, Romagnani C, Romagnani S. The 3 major types of innate and adaptive cell-mediated effector immunity. J Allergy Clin Immunol. 2015 Mar;135(3):626-35. [CrossRef]
- Feng B, Tang L. A new paradigm for cancer immunotherapy: Orchestrating type 1 and type 2 immunity for curative response. Clin Transl Med. 2025 Jan;15(1):e70154. [CrossRef]
- Wagner M, Nishikawa H, Koyasu S. Reinventing type 2 immunity in cancer. Nature. 2025 Jan;637(8045):296-303. Epub 2025 Jan 8. PMID: 39780006. [CrossRef]
- Kyrstsonis M.-C., Dedou S.G., Bax E.C., Stamate Lou M., Maniatis A. Serum Interleukin-6 (IL-6) and Interleukin-4 (IL-4) in Patients with Multiple Myeloma (MM) Br. J. Haematol. 1996;92:420–422. [CrossRef]
- Mikulski D, Robak P, Perdas E, Węgłowska E, Łosiewicz A, Dróżdż I, Jarych D, Misiewicz M, Szemraj J, Fendler W, Robak T. Pretreatment Serum Levels of IL-1 Receptor Antagonist and IL-4 Are Predictors of Overall Survival in Multiple Myeloma Patients Treated with Bortezomib. J Clin Med. 2021 Dec 26;11(1):112. [CrossRef]
- Traynor AE, Schroeder J, Rosa RM, Cheng D, Stefka J, Mujais S, et al. Treatment of severe systemic lupus erythematosus with high-dose chemotherapy and haemopoietic stem-cell transplantation: a phase I study. Lancet. 2000;356:701–707. [CrossRef]
- Leung PSC, Shuai Z, Liu B, Shu SA, Sun L. Stem Cell Therapy in the Treatment of Rheumatic Diseases and Application in the Treatment of Systemic Lupus Erythematosus. Next-Generation Therapies and Technologies for Immune-Mediated Inflammatory Diseases. 2016 Sep 19:167–98. [CrossRef]
- Fagoaga, Omar & Nehlsen-Cannarella, Sandra & Ratanatharatorn, Voravit & Martinelli, Christine & Lum, Lawrence & Uberti, Joseph. (2007). Sequential Serum Cytokine Levels after Autologous, Related and Unrelated Allogeneic Stem Cell Transplantation: Reflection of Immune Reactivity and Reconstitution.. Blood. 110. 3885-3885. 10.1182/blood.V110.11.3885.3885.
- Laurie SJ, Foster JP 2nd, Bruce DW, Bommiasamy H, Kolupaev OV, Yazdimamaghani M, Pattenden SG, Chao NJ, Sarantopoulos S, Parker JS, Davis IJ, Serody JS. Type II innate lymphoid cell plasticity contributes to impaired reconstitution after allogeneic hematopoietic stem cell transplantation. Nat Commun. 2024 Jul 17;15(1):6000. [CrossRef]
- Chen H, Wang X, Wang Y, Chang X. What happens to regulatory T cells in multiple myeloma. Cell Death Discov. 2023 Dec 21; 9(1):468. [CrossRef]
Figure 1.
Flow cytometry gating strategy used for ILC2.
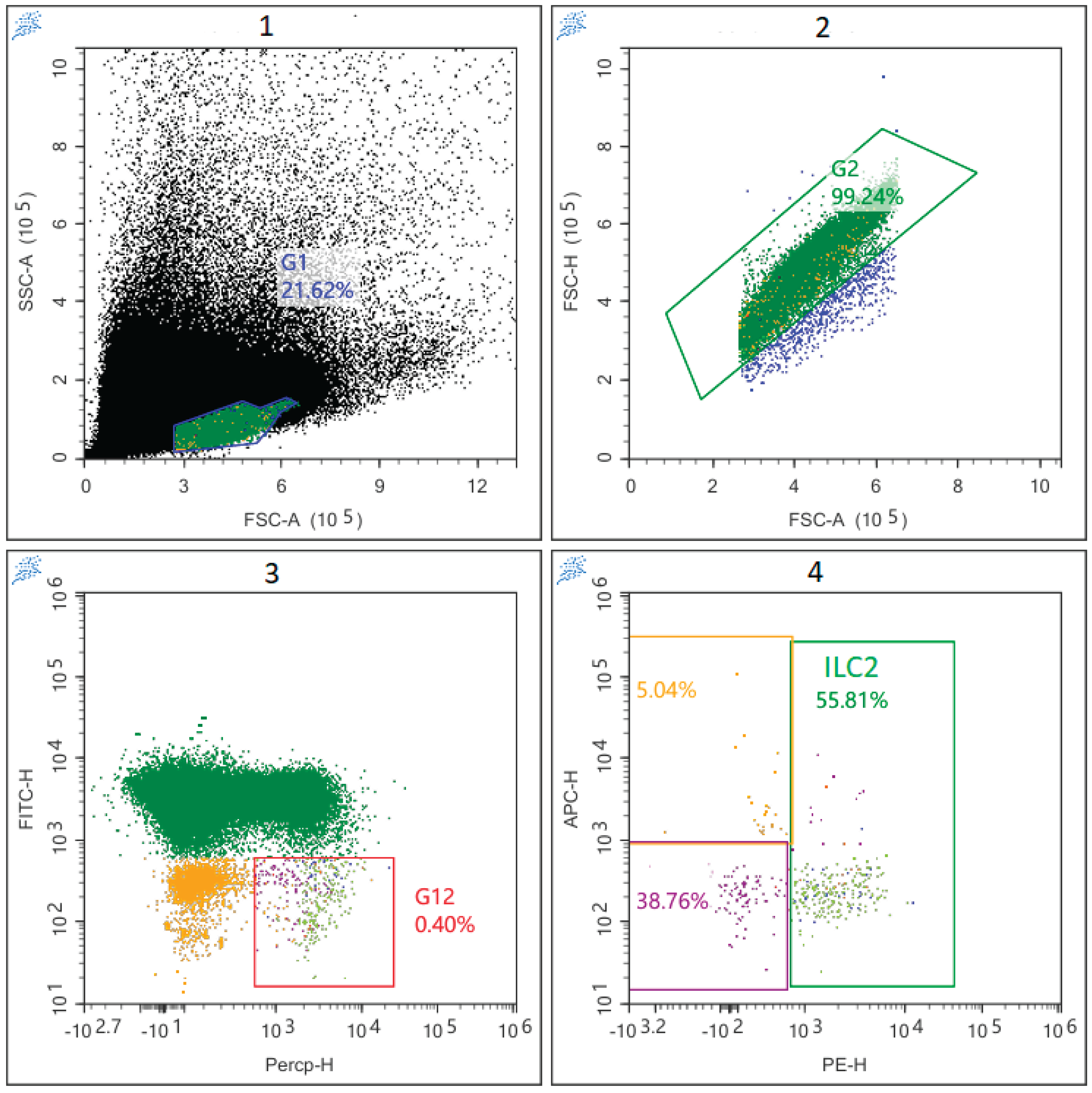
Figure 2.
Relative number of ILC among PBMC in patients with multiple myeloma and healthy volunteers; HC – healthy control group, MM before – MM patients before auto-HSCT, MM after - MM patients after auto-HSCT. Data are presented as box-and-whisker plots, with boxes extending from the 25th to the 75th percentile, with a horizontal line at the median, while the whiskers extend to the lowest and highest data points. * Indicates a significant difference (p < 0.05). ** Indicates a significant difference (p < 0.01) . ns Indicates no significant differences. .
Figure 2.
Relative number of ILC among PBMC in patients with multiple myeloma and healthy volunteers; HC – healthy control group, MM before – MM patients before auto-HSCT, MM after - MM patients after auto-HSCT. Data are presented as box-and-whisker plots, with boxes extending from the 25th to the 75th percentile, with a horizontal line at the median, while the whiskers extend to the lowest and highest data points. * Indicates a significant difference (p < 0.05). ** Indicates a significant difference (p < 0.01) . ns Indicates no significant differences. .
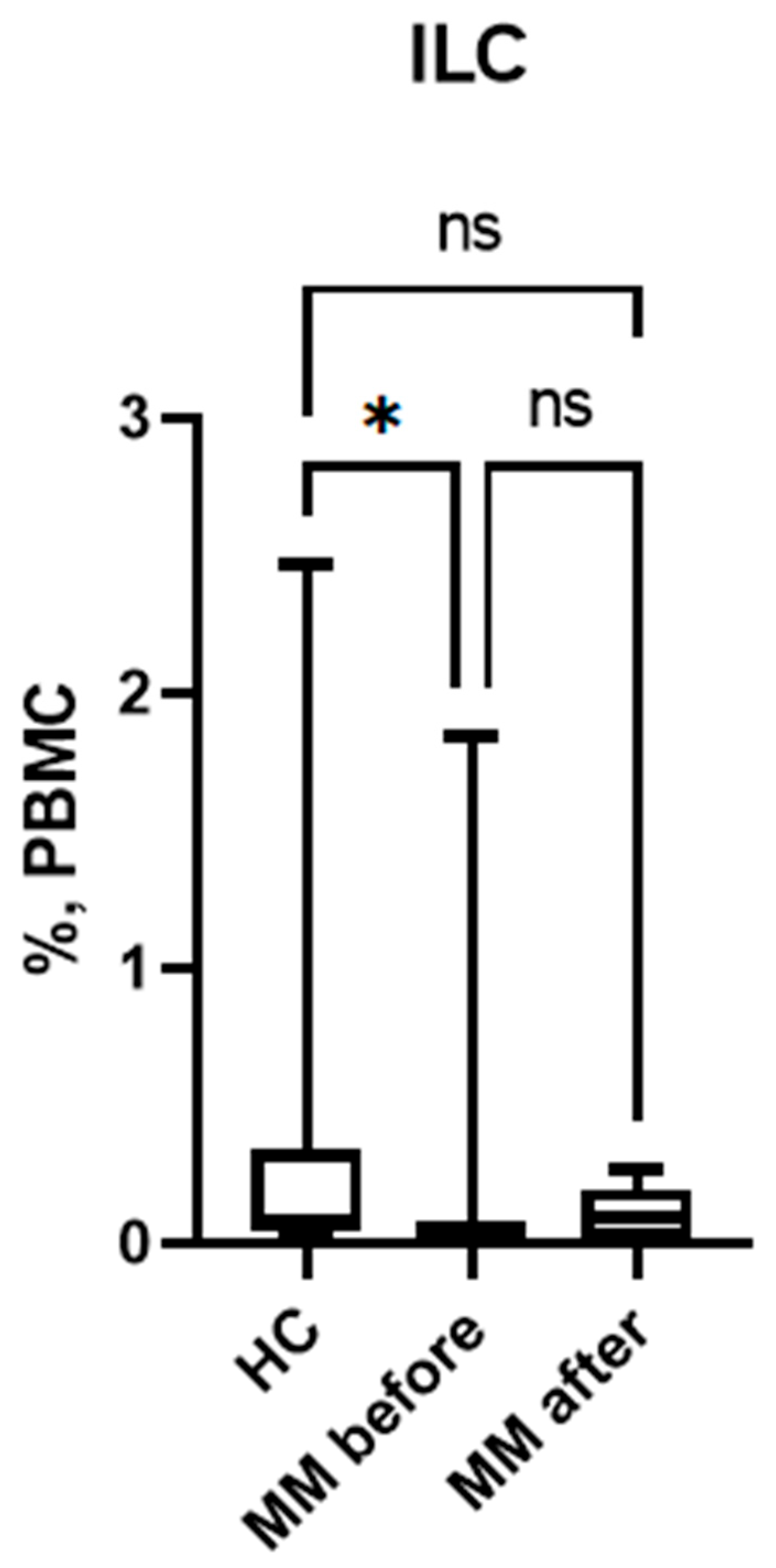
Figure 3.
Relative number of different subpopulations of ILC in patients with multiple myeloma and healthy volunteers; HC – healthy control group, MM before – MM patients before auto-HSCT, MM after - MM patients after auto-HSCT; (a) relative number of ILC1, (b) relative number of ILC2, (c) relative number of ILC3. Data are presented as box-and-whisker plots, with boxes extending from the 25th to the 75th percentile, with a horizontal line at the median, while the whiskers extend to the lowest and highest data points. * Indicates a significant difference (p < 0.05). ** Indicates a significant difference (p < 0.01) . *** Indicates a significant difference (p < 0.001). ns Indicates no significant differences.
Figure 3.
Relative number of different subpopulations of ILC in patients with multiple myeloma and healthy volunteers; HC – healthy control group, MM before – MM patients before auto-HSCT, MM after - MM patients after auto-HSCT; (a) relative number of ILC1, (b) relative number of ILC2, (c) relative number of ILC3. Data are presented as box-and-whisker plots, with boxes extending from the 25th to the 75th percentile, with a horizontal line at the median, while the whiskers extend to the lowest and highest data points. * Indicates a significant difference (p < 0.05). ** Indicates a significant difference (p < 0.01) . *** Indicates a significant difference (p < 0.001). ns Indicates no significant differences.
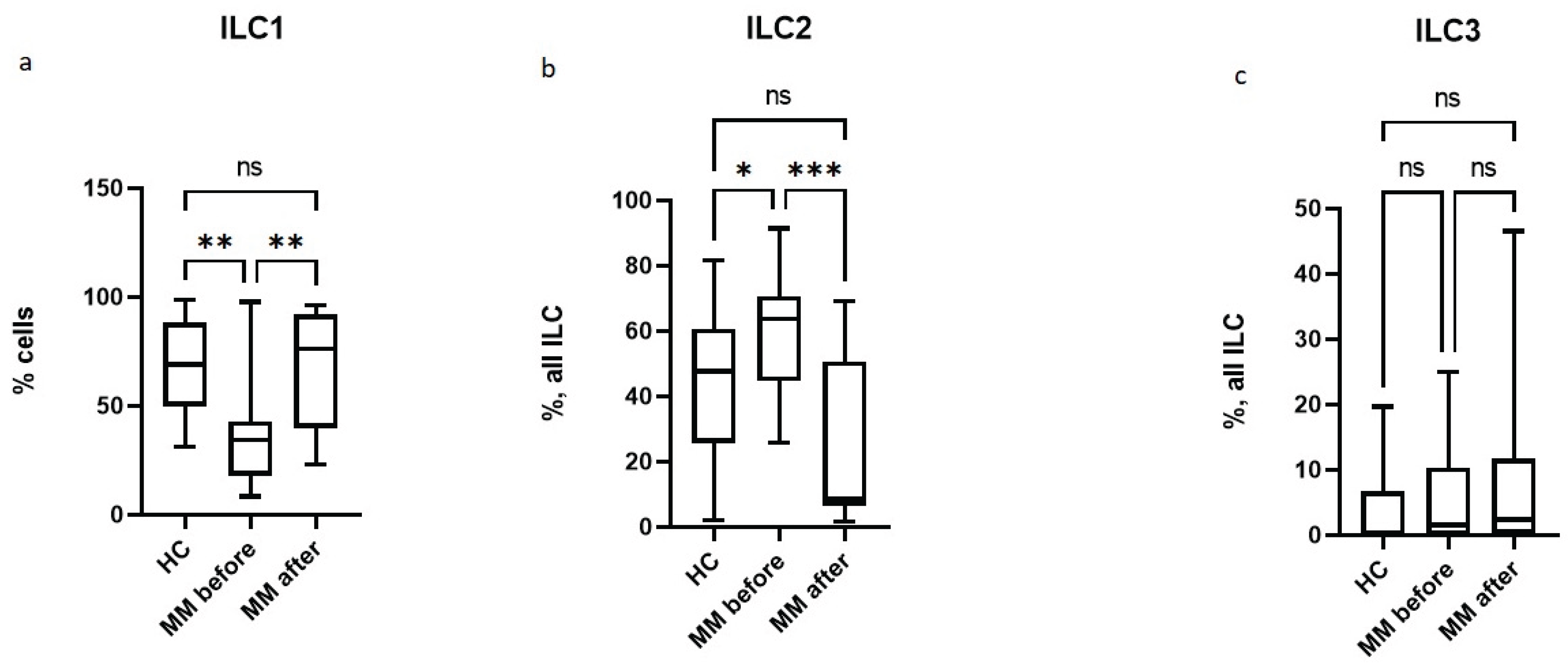
Figure 4.
Relative number of CD5-expressing ILC2 in patients with multiple myeloma and healthy volunteers; HC – healthy control group, MM before – MM patients before auto-HSCT, MM after - MM patients after auto-HSCT. Data are presented as box-and-whisker plots, with boxes extending from the 25th to the 75th percentile, with a horizontal line at the median, while the whiskers extend to the lowest and highest data points. * Indicates a significant difference (p < 0.05). ** Indicates a significant difference (p < 0.01) . ns Indicates no significant differences. .
Figure 4.
Relative number of CD5-expressing ILC2 in patients with multiple myeloma and healthy volunteers; HC – healthy control group, MM before – MM patients before auto-HSCT, MM after - MM patients after auto-HSCT. Data are presented as box-and-whisker plots, with boxes extending from the 25th to the 75th percentile, with a horizontal line at the median, while the whiskers extend to the lowest and highest data points. * Indicates a significant difference (p < 0.05). ** Indicates a significant difference (p < 0.01) . ns Indicates no significant differences. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



