Submitted:
03 December 2025
Posted:
05 December 2025
You are already at the latest version
Abstract
The production of pear (Pyrus communis L.) nurseries is essential to providing high-quality planting material for the establishment of a successful orchard. Thus, encouraging early vegetative growth and seedling vigor during the nursery period requires optimal fertilization. Under temperate continental circumstances in northwest Romania, this study assessed the impact of various NPK fertilizer rates on the shoot fresh weight of pear nursery trees. The study was carried out in 2025 using a factorial design with two Romanian cultivars (‘Napoca’ and ‘Monica’) and four fertilization treatments (N0P0K₀, N8P8K8, N16P16K16, and N₂₄P₂₄K₂₄), set up in a randomized block system with five replications. At progressively higher rates of 50, 100, and 150 kg ha⁻¹, a totally water-soluble 16–16–16 fertilizer was applied. At the conclusion of the growing season, the fresh weight of the shoots was measured. The accumulation of shoot biomass was significantly and gradually impacted by fertilization. The fresh weight of the shoots rose by 29%, 45%, and 59% as compared to the unfertilized control (0.42 kg tree⁻¹) under the treatments of N8P8K8, N₁₆P16K16, and N₂₄P₂₄K₂₄. There were no discernible cultivar-dependent variations at any fertilization level, and both cultivars showed almost equal growth responses. These findings show how strongly the growth of pear nursery shoots depends on the availability of NPK and offer helpful advice for maximizing fertilization techniques to enhance the quality of planting materials.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Site description
2.2. Plant material
2.3. Experimental design
2.4. Growth measurements
2.5. Statistical analysis
3. Results
3.1. Effect of fertilization on shoot fresh weight
3.2. Interaction between fertilization rate and cultivar
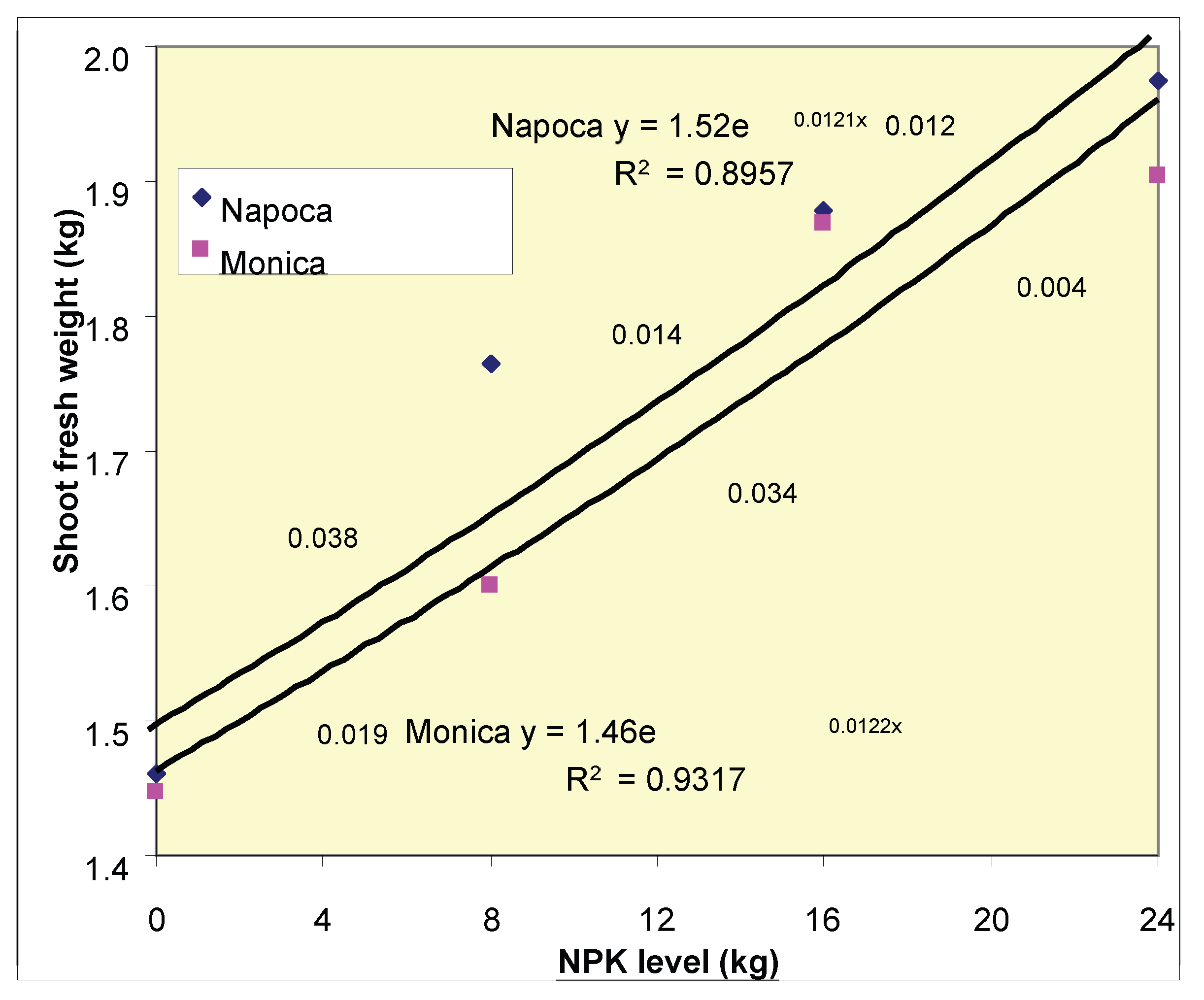
3.3. Relative increase in shoot fresh weight and cultivar differences
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, Y.; Zhang, X.; Li, H.; Chen, Z. Effects of nursery management practices on early growth and transplant survival of pear trees. Sci. Hortic. 2022, 293, 110651.
- Li, Y.; Wang, J.; Huang, Q.; Zhang, L. Nitrogen effects on shoot growth and photosynthesis of pear seedlings. J. Plant Nutr. 2017, 40, 2345–2356.
- Clark, J.R.; Smith, M.W.; Taylor, G. Nitrogen nutrition and growth of woody nursery plants. HortScience 2015, 50, 1442–1448.
- Razaq, M.; Zhang, P.; Shen, H.; Salahuddin. Influence of nitrogen and phosphorus on the growth and root morphology of plants. J. Plant Nutr. 2017, 40, 1514–1524.
- Brunetto, G.; Melo, G.W.; Toselli, M.; Quartieri, M.; Tagliavini, M. The role of nitrogen, phosphorus, and potassium fertilization on fruit tree nutrition. Rev. Bras. Frutic. 2015, 37, 1089–1104.
- Liao, X.; Zhao, W.; Liu, Y.; Zhang, C. Nitrogen availability regulates root architecture and shoot biomass in nursery woody perennials. Plant Soil 2025, 497, 215–229.
- Lindström, A.; Rune, G.; Larsson, S. Shoot and root biomass allocation in young forest plants as affected by fertilization. New For. 2005, 29, 87–98.
- Close, D.C.; Davidson, N.J.; Beadle, C.L. Physiological responses of nursery seedlings to nutrient supply. Tree Physiol. 2005, 25, 479–489.
- Haase, D.L. Understanding forest seedling quality: Measurements and interpretation. Tree Planters’ Notes 2008, 52, 24–30.
- Grossnickle, S.C. Why seedlings survive: Influence of plant attributes. New For. 2012, 43, 711–738.
- Grossnickle, S.C.; MacDonald, J.E. Seedling quality: History, application, and plant attributes. Forests 2018, 9, 283.
- Soti, P.G.; Jayachandran, K.; Koptur, S.; Volin, J.C. Effect of nutrient fertilization on growth of container-grown woody plants. HortTechnology 2014, 24, 583–591.
- Close, D.C.; Wilson, S.J. Provenance, and fertilizer effects on growth of nursery seedlings. Aust. For. 2002, 65, 242–248.
- Dong, Q.; Yang, Y.; Yu, K.; Feng, H. Biomass accumulation responses of fruit nursery plants to nutrient supply. Agronomy 2020, 10, 1241.
- Chen, S.; Zhang, X.; Zhao, Y.; Ma, Y. Excess nitrogen fertilization reduces stress tolerance and increases nutrient leaching in orchard systems. Environ. Pollut. 2018, 243, 563–572.
- Trubat, R.; Cortina, J.; Vilagrosa, A. Low nutrient availability delays nursery stock development and field establishment. Plant Soil 2010, 327, 29–42.
- Melo, G.W.; Brunetto, G.; Toselli, M.; Quartieri, M. Phosphorus and potassium fertilization in pear orchards. Acta Hortic. 2015, 1099, 273–280.
- Bordeanu, N.; Butcaru, A.; Sestras, R. Influence of organic and mineral fertilizers on pear tree growth. Fruits 2015, 70, 211–218.
- Ilić, R.; Aćimović, S.; Radičević, S. Fertilization effects on shoot growth and leaf nutrient status of pear. Hort. Sci. 2022, 49, 117–125.
- Liang, G.; He, P.; Jin, J.; Zhou, W. Nutrient surpluses and fertilization inefficiency in commercial pear orchards of China. Nutrients 2022, 14, 2465.
- Haifa Group. Pear Fertilization Guide; Haifa Chemicals: Haifa, Israel, 2020.
- ICL Growing Solutions. Pear Nutrition and Fertilizer Programmes; ICL: Tel-Aviv, Israel, 2021.
- Gill, P.P.S.; Verma, M.L.; Josan, J.S. Integrated nutrient management in pear orchards. Indian J. Hortic. 2017, 74, 258–264.
- Wu, L.; Lu, J.; Wang, J. Effects of balanced fertilization on vegetative growth and yield of pear. Sci. Hortic. 2019, 246, 112–118.
- Jiang, S.; Zhang, L.; Li, J. Long-term fertilization effects on pear orchard soil fertility and productivity. Soil Tillage Res. 2020, 198, 104542.
- Park, B.B.; Yanai, R.D.; Sahm, J.M. Nursery fertilization and post-transplant performance of deciduous trees. For. Ecol. Manag. 2015, 335, 150–158.
- Wang, Z.; Li, J.; Zhang, X. Fertilization effects on nutrient use efficiency of nursery-grown woody plants. J. Environ. Hortic. 2015, 33, 78–85.
- Rotowa, K.A.; Mensah, S.; Adu-Bredu, S. Nursery fertilization determines field performance of woody perennials. Agronomy 2025, 15, 312.
- Tagliavini, M.; Quartieri, M.; Millard, P. Remobilized nitrogen, and root uptake of nitrate for spring leaf growth, flowers and developing fruit in pear (Pyrus communis L.). Plant Soil 2000, 221, 281–290.
- Neilsen, D.; Neilsen, G.H.; Herbert, L.C.; Hogue, E.J. Response of apple to fertigation of N and K under conditions susceptible to nitrate leaching. Soil Sci. Soc. Am. J. 2004, 68, 1612–1620.
- Nava, G.; Dechen, A.R.; Nachtigall, G.R. Nitrogen and potassium fertilization affect apple fruit quality in southern Brazil. Sci. Hortic. 2008, 117, 344–349.
- Bauer, G.; Schulz, H.; Wieneke, J. Growth and nutrient uptake of nursery plants in response to different nitrogen fertilization regimes. J. Plant Nutr. 2004, 27, 985–999.
- Uscola, M.; Villar-Salvador, P.; Gross, P.; Maillard, P. Fast growth involves high dependence on stored resources in seedlings of Mediterranean woody species. New Phytol. 2015, 205, 169–180.
- Oliet, J.A.; Planelles, R.; Artero, F.; Jacobs, D.F. Nursery fertilization, outplanting stress, and field performance of Mediterranean oak seedlings. For. Ecol. Manag. 2009, 258, 2432–2440.
- Tsakaldimi, M.; Tsitsoni, T.; Ganatsas, P.; Zagas, T. Nursery stock quality and field performance of Quercus pubescens seedlings produced under different fertilization regimes. Ann. For. Sci. 2013, 70, 389–398.
- Lamhamedi, M.S.; Labbé, L.; Margolis, H.A.; Stowe, D.C.; Blais, L. Effects of nitrogen fertilization on growth and physiology of container-grown white spruce seedlings. New For. 2012, 43, 493–509.
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012.
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Functions of macronutrients. In Marschners Mineral Nutrition of Higher Plants; Marschner, P., Ed.; Academic Press: London, UK, 2012; pp. 135–189.
- Zörb, C.; Senbayram, M.; Peiter, E. Potassium in agriculture—Status and perspectives. J. Plant Physiol. 2014, 171, 656–669.
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2014, 202, 30–46.
- Atkinson, C.J.; Else, M.A.; Taylor, L.; Dover, C.J. Root and shoot growth of apple trees in response to different irrigation and fertilization regimes. Tree Physiol. 2010, 30, 123–131.
- Oliet, J.A.; Jacobs, D.F. Restoring forests: Advances in techniques and theory. New For. 2012, 43, 535–541.
- Landis, T.D.; Dumroese, R.K.; Haase, D.L. The Container Tree Nursery Manual, Volume 7: Seedling Processing, Storage, and Outplanting; USDA Forest Service: Washington, DC, USA, 2010.
- Tramontini, S.; Vitali, M.; Celi, L.; Certini, G. Fertilization effects on growth, nutrient uptake and nitrogen use efficiency in woody nursery seedlings. J. Plant Nutr. Soil Sci. 2013, 176, 267–276.
- Fernández-Escobar, R.; Benlloch, M.; Barranco, D.; Dueñas, A.; Gutiérrez-Gañán, J.A. Response of olive trees to nitrogen fertilization under drip irrigation. Plant Soil 2014, 381, 253–264.
- Jacobs, D.F.; Rose, R.; Haase, D.L.; Morgan, P.D. Influence of nursery fertilization and irrigation on early field performance of conifer seedlings. For. Sci. 2009, 55, 94–107.
- Villar-Salvador, P.; Puértolas, J.; Peñuelas, J.L.; Planelles, R. Effect of nursery fertilization on drought resistance and field performance of Mediterranean woody species. New For. 2012, 43, 721–737.
- Diacono, M.; Montemurro, F. Long-term effects of organic amendments on soil fertility: A review. Agron. Sustain. Dev. 2010, 30, 401–422.
- López-Bellido, L.; Muñoz-Romero, V.; López-Bellido, R.J. Energy use efficiency and sustainability of agricultural production systems. Agron. Sustain. Dev. 2014, 34, 955–963.

| Fertilization rate | Mean shoot fresh weight (kg tree⁻¹) | Relative value (%) | Significance |
| N₀P₀K₀ | 0.42 ± 0.02d | 100 | d |
| N₈P₈K₈ | 0.54 ± 0.03c | 129 | c |
| N₁₆P₁₆K₁₆ | 0.61 ± 0.03b | 145 | b |
| N₂₄P₂₄K₂₄ | 0.66 ± 0.03a | 159 | a |
| Fertilization rate (NPK) | ‘Napoca’ (kg tree⁻¹) |
Tukey letters |
CV (%) | ‘Monica’ (kg tree⁻¹) |
Tukey letters |
CV (%) |
| N₀P₀K₀ | 0.42 ± 0.03 | x | 14.3 | 0.42 ± 0.02 | x | 14.1 |
| N₈P₈K₈ | 0.53 ± 0.03 | y | 12.7 | 0.55 ± 0.03 | y | 13.2 |
| N₁₆P₁₆K₁₆ | 0.61 ± 0.03 | z | 11.2 | 0.60 ± 0.03 | z | 11.4 |
| N₂₄P₂₄K₂₄ | 0.66 ± 0.03 | z | 11.3 | 0.65 ± 0.03 | z | 11.7 |
| Fertilization rate (NPK) | Mean shoot fresh weight (kg tree⁻¹) | Increase vs. control (%) | ‘Napoca’ (kg tree⁻¹) | ‘Monica’ (kg tree⁻¹) |
| N₀P₀K₀ | 0.42 | - | 0.42 | 0.43 |
| N₈P₈K₈ | 0.54 | + 29 | 0.54 | 0.55 |
| N₁₆P₁₆K₁₆ | 0.61 | + 45 | 0.61 | 0.60 |
| N₂₄P₂₄K₂₄ | 0.66 | + 59 | 0.66 | 0.65 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



