Submitted:
03 December 2025
Posted:
04 December 2025
You are already at the latest version
Abstract
The intestinal barrier is a complex and dynamic structure that plays a critical role in maintaining gut health. Though, pathological conditions such as, dysbiosis or inflammation that could damage the intestinal barrier to varying degrees, aggravating the primary disease. Recently, fermented plant-origin foods and probiotics were used as an alternative for health remedy. For this reason, in this current study, we made the inquiries about the effect of Tunisian zgougou juice obtained upon both spontaneous (J) and driven fermentation (J(A1+A2)) with autochthonous probiotic bacteria Lactiplantibacillus plantarum (LP) species A1 and A2, the whole bacteria (A1) and (A2) and their CFS. The cell integrity and permeability was assessed based on the TEER measurement on monolayer cell culture of Caco-2 cell, then the gene expression levels of the tight junction proteins (TJP) and the gene expression level of the inflammatory cytokine TNFα were evaluated using the RT-PCR. Our experimental findings exhibited that the driven fermented had a potential protective effects on intestinal barrier function in vitro shown by the great increase the electrical barrier detection (TEER) between Caco2 cell reaching ca. 690 to 963.33 Ω/cm2, although the regulatory mechanism has not been reported yet. Otherwise LP (A1 and A2) and their CFS showed moderate results. Furthermore, J(A1+A2) promoted the mRNA relative gene expression of the different tested tight TJPs and moderated the intestinal inflammation by influencing the TNFα expression. This study revealed that probiotics and especially probiotic-based plant-origin fermented functional foods (Tunisian zgougou juice) could improve and reinforce the intestinal barrier which provide new ideas for treating intestinal injury-related diseases, and have to be a revival in worldwide popularity, mostly as a consequence of claims made about their health benefits. So that, to completely reveal the health advantages of these probiotic LAB strains for humans and this new food matrix, additional in vivo and clinical investigations are necessary, as highlighted by the limitations of in vitro models.
Keywords:
zgougou
; fermentation
; L. plantarum group
; intestinal barrier integrity
; TNFα
; TEER
; tight junction proteins
1. Introduction
Aleppo pine (Pinus halepensis Mill) seeds represent the main ingredient for a Tunisian fermented food known as ‘Assidet zgougou’. Its putative health benefits are anecdotally known (Botella et al., 2011; Schiller, 2014; Kadri et al., 2015; Zaroual et al., 2019; Ait Atmane et al., 2021). It is postulated previously that these health benefits could be linked to its chemical composition and its richness on phenolic compounds that confer the analgesic, anti-inflammatory and antioxidant capacities (Minervini et al., 2020; Ait Atmane et al., 2024). Furthermore, zgougou nuts or juice obtained upon both driven and spontaneous fermentation are rich in bioactive molecules (polyunsaturated fatty acids, linolenic acid (Omega3), vitamins (B1, E…), iron, phosphorus…) also rich in nutrients and energy. In addition, zgougou is fermented with lactic acid bacteria with in vitro antimicrobial and antioxidant activities, which can positively influence the gut microbiota composition and function, leading to improved gut barrier function (Tukan et al., 2013; Missaoui et al., 2019; Minervini et al., 2020).
Intestinal epithelial barrier, having a pivotal role in maintaining the human body in good health, results from complex interactions that involve biochemical, immunological, microbial, and structural components (Han et al., 2023; Mosiej et al., 2025). Besides the mucous layer and microbial community, Tight Junction Proteins (TJP) of intestinal epithelium are of almost importance for its “gate and fence” function. In fact, the intercellular junction known as the tight junction or zona occludens are found at the tight junctions of endothelial and epithelial cells. They are essential for controlling cell permeability of ions, water, and solutes move through the paracellular route, coordinating the distribution of membrane proteins and lipids, and affecting the polarity, proliferation, and differentiation of cells in order to create selectively permeable cellular barriers that divide the body’s apical (luminal) and basolateral (abluminal) sides. This allows the tight junction to control the transport processes and preserve homeostasis (Balda & Matter, 2000; Suzuki, 2020; Paradis et al., 2021; Zhang et al., 2022). Occludin (OCLN), the first identified TJP, provides structural integrity to the tight junctions existing between the Intestinal Epithelial Cells (IECs) (Zhang et al., 2022). When over-synthesized, it limits the cytokine-induced increase of gut permeability (Weber et al., 2016; Wang, et al., 2021). Claudin-2 (CLDN2) is another TJP associated with improved permeability of intestinal epithelium (Colegio et al., 2003; Campbell et al., 2017; Zhang et al., 2022). Tumour necrosis factor-α (TNFα) is an inflammatory cytokine that regulates integrity of tight junctions through internalization of OCLN, which elevates gut permeability. Lipocalin-2 (LCN2) is a further protein component of the intestinal epithelial barrier, with antibacterial activity, which seems to be induced by inflammatory stimuli (Saha et al., 2017; Sahu et al., 2023).
Oxidative stress and several pathological conditions may damage intestinal barrier integrity, thus facilitating the transit of pathogenic microorganisms and their toxic metabolites from gut lumen to underlying layers (e.g., lamina propria, endowed with immune function) (Yuki et al., 2007; Ohland & Macnaughton, 2010; Heli et al., 2012; Zhang et al., 2022; Bae et al., 2022; Mosiej et al., 2025). Recently, some probiotic bacteria showed to benefit host health through restoration of the intestinal barrier function (Mosiej et al., 2025). For instance, probiotic strains of lactobacilli and Bifidobacterium infantis could restore the intestinal barrier by increasing the transcription level of genes involved in the tight junctions (Ohland & Macnaughton, 2010; Sharma et al., 2010; Ashida et al., 2011; La Fata et al., 2018; Wierzbicka et al., 2021; Wei et al., 2022; Zhang et al., 2022; Oudat, & Okour, 2025). Restoration of barrier function could also be obtained through administration of cell-free supernatant (CFS) from probiotic microorganisms, thus overlooking the “postbiotic” frontier (Nouri et al., 2016; Tiptiri-Kourpeti et al., 2016; Wierzbicka et al., 2021; Zhang et al., 2022; Dasriya et al., 2024; Mosiej et al., 2025). Furthermore, probiotics may display their beneficial activity also when administered as food ingredient (Nithya et al., 2023). However, in the latter case it is hard to define the borders between the food matrix and the probiotics, in terms of their contribution to the observed beneficial effect.
This study aims to understand the in vitro effect of two selected lactobacilli with probiotic traits on intestinal permeability and level of expression of OCLN, CLDN2, and LCN2. The lactobacilli were tested in different preparations: (i) cell biomass from single liquid culture; (ii) CFS from single liquid culture; (iii) zgougou obtained through the combination of lactobacilli as starters. In addition, spontaneously fermented zgougou was also subjected to the same assays, using human Caco-2 cells monolayer, as in vitro model for intestinal epithelium.
2. Material & Methods
2.1. Preparation of Epithelial Cell Monolayers (Cell Culture)
Caco-2 (Caucasian colon adenoarcinoma) cells, purchased from ATCC (ATCC, Rockville, USA), were seeded in the upper part of Corning® Transwell® 12 well plates with 0.4 μm pore size (Corning®, New York, USA) in Dulbecco Modified Eagles Medium (DMEM, Thermo Fisher Scientific, Massachusetts, USA) supplemented with L-Glutamine, 10% heat-inactivated Fetal Bovine Serum (Thermo Fisher Scientific, Massachusetts, USA), 1% of 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES, Sigma-Aldrich, St. Louis, USA) (1 M), 1% of sodium puryvate solution (100 mM) (Sigma-Aldrich, St. Louis, USA ), 1% of streptomycin solution (10 mg/mL) and penicillin solution (Thermo Fisher Scientific, Massachusetts, USA) (10000 U/mL). Cells were incubated at 37 °C in a 5% CO2 atmosphere for six days, feeding them, on alternate days, with fresh DMEM. Full differentiation of monolayer was confirmed through measurement of transepithelial electrical resistance (TEER), using the Millicell ERS Volt- Ohm-meter (Millipore, Eschborn, Germany) and visual control of cell layer integrity using a microscope, following the steps elsewhere reported (Huang, 2009; Talà et al., 2022).
2.2. Preparation of Bacterial Cultures and Zgougou Juices
Lactiplantibacillus paraplantarum A1 and Lactiplantibacillus plantarum A2, previously isolated from traditional zgougou (Missaoui et al., 2019) and belonging to the Culture Collection of the Department of Soil, Plant, and Food Sciences, University of Bari ALDO MORO, Italy and the Departement of Bio-resources: Integrative Biology & Valorization, Higher Institute of Biotechnology of Monastir, University of Monastir, Tunisia, were cultured in de Man Rogosa and Sharpe (MRS) broth (Oxoid, Basingstoke, Hamsphire, UK) at 37 °C for 24 h, under anaerobic conditions. The culture broth was then centrifuged (10,000×g, 20 min, 4° C) and CFS was separated from cell biomass. Zgougou juice was obtained from a mixture of Aleppo pine seeds and water according to a ratio (2:3; w:v), in a sampling container we puted 20 g of Aleppo pine’s seeds ground by coffee mill, and then 30 mL of tap water were added and mixed together, inoculated with L. paraplantarum A1 and L. plantarum A2 (J A1+A2) (both at cell density in the order of 8 log CFU/g) and fermented at 25 °C for 24 h, as previously detailed (Minervini et al., 2020). Additionally, juice from spontaneously fermented zgougou (J) was obtained after fermentation at 25 °C for 24 h, without addition of lactobacilli.
2.3. Exposure of Caco-2 Cell Line to Bacterial Biomass, CFS, and Zgougou Juices
Caco-2 cells became confluent after about 6 days, when confluence have been reached, cells were stimulated individually for 24 hours with: different samples probiotic bacterial strains A1, A2, CFS extracted from either strain, (J), and (J (A1+A2)) into the wells of the tissue culture plates. The blank was empty without cells. Untreated cells were used as negative controls in all experiments (vehicle). As a positive control 1µg/mL of Escherchia coli lipopolysaccharide (LPS) stimulation was used (Sigma-Aldrich, St. Louis, USA). Plates were then incubated at 37 °C in 10% CO2 and 90% air. Confluence was controlled by TEER measurement using a Millicell ERS Ohmmeter, (Millipore, Eschborn, Germany) and visual control of cell monolayer integrity under the microscope following the steps reported previously by (Talà et al., 2022). The integrity of the monolayer was assessed at various times ; after 10 ‒ 30 minutes and after 1‒ 2‒6 and 24 hours. Measurements were expressed in standard units (Ωcm2) after subtracting mean values for resistance obtained from cell-free inserts.
2.4. Gene Level Expression Related to Intestinal Barrier Integrity
The level of expression of genes related to intestinal barrier integrity was analysed through real-time PCR. Total RNA was extracted from cells exposed to the aforementioned treatments for 24 h using the Invitrogen™ TRIzol™ Reagent (Thermo Fisher Scientific, Massachusetts, USA), following the manufacturer’s instructions. Total RNA (500 ng) was reverse-transcribed to cDNA using iScript cDNA Synthesis kit (Biorad, California, USA) with random primers, as described elsewhere (Parlesak et al., 2004; Talà et al., 2022). The following TaqManTM (Thermo Fisher Scientific, Massachusetts, USA) gene expression assays were used: Hs00174128_m1for the TNF gene, coding for tumour necrosis factor; Hs00252666_s1 for the CLDN2 gene, coding for claudin 2; Hs05465837_g1 for the OCLN gene, coding for occludin; Hs01008571_m1 for the LCN2 gene, coding for lipocalin 2; and Hs02758991_g1 for the GAPDH gene, coding for glyceraldehyde-3-phosphate dehydrogenase. The latter was used as the housekeeping gene. PCR was performed on a CFX96 System (Biorad, California, USA), using the mixture reaction and conditions reported by Talà et al. (2022). The level of expression of the target genes was calculated using the ΔΔCt method, with respect to the expression level of the GAPDH gene.
2.5. Quantification of TNFα
The cell culture medium was recovered from the wells hosting the cells exposed to the different treatments and centrifuged for ten minutes. Supernatants were subjected to quantification of TNFα using the ELISA kit (RD Systems, Minneapolis, MN, USA) with 96-well plates, as described by Bermudez-Brito et al., 2015.
2.6. Statistical Analysis
Statistical analysis was performed using the Graphpad Prism 9.3.1 (471). All data were expressed as means± S.D. Statistical significance were assessed with two-tailed Student’s t-test and 2-way Anova test (*P<0.05; **P<0.01; ***P<0.001). Results were considered statistically significant at P<0.05. The experiments were conducted in triplicate except for ELISA where analyses were performed in duplicate.
3. Results & Discussions
3.1. Impact of Probiotics and Fermentation on Gut Integrity
In order to maintain a good intestinal barrier, human being must have a balanced diet high in fiber, exercise frequently, drink enough water, and control their stress levels. Rather, probiotics—particularly those belonging to the Lactoplantibacillus spp.— as well prebiotics, postbiotics, and fermented foods containing probiotic bacterial starter cultures, can also improve gut health by fostering a healthy microbiota (Dasriya et al., 2024; Mosiej et al., 2025). Thus, when the intestinal barrier is functioning effectively, nutrients and water can be absorbed selectively, while pathogens, toxins, and undigested food particles can’t enter. However, several factors, including chronic diseases, poor diet, stress, and infections, can weaken the barrier and result in “leaky gut”. Otherwise, persistent inflammation occurs when the immune system fails to regulate or control the immune response due to excessive stimulation by pro-inflammatory substances, impairing its ability to return to baseline (peripheral immunological tolerance) (Mosiej et al., 2025). Additionally, persistently high tissue or serum TNFα concentrations may be the primary inflammatory phenomenon with tissue consequences. This condition is linked to increased intestinal permeability and has been linked to various health issues, including autoimmune diseases, allergies, and inflammatory bowel diseases (Bamola et al., 2022). For this reason, human Caco-2 cells are a perfect monolayer system to explore this phenomenon in vitro. To the best of our knowledge, this is the first study that analysed the epithelial intestine cell integrity and to trigger the anti-inflammatory effect of two lactobacilli isolated from zgougou, their cell free supernatants (Missaoui et al., 2019), and the two samples of zgougou; spontaneous fermented Zgougou juice (J), and fermented Zgougou juice inoculated with probiotic LAB (J (A1+A2)) (Minervini et al., 2020). Those samples were tested in vitro for their adhesive capacities to Caco-2 monolayer cells, grown on trans-well membranes, at varying degrees and time of treatment. Their effects were evaluated by trans-epithelial electrical resistance (TEER) measurement and compared to the positive control LPS from Escherichia coli, focusing on and their anti-inflammatory activity and modulation of the tight junction protein gene expression.
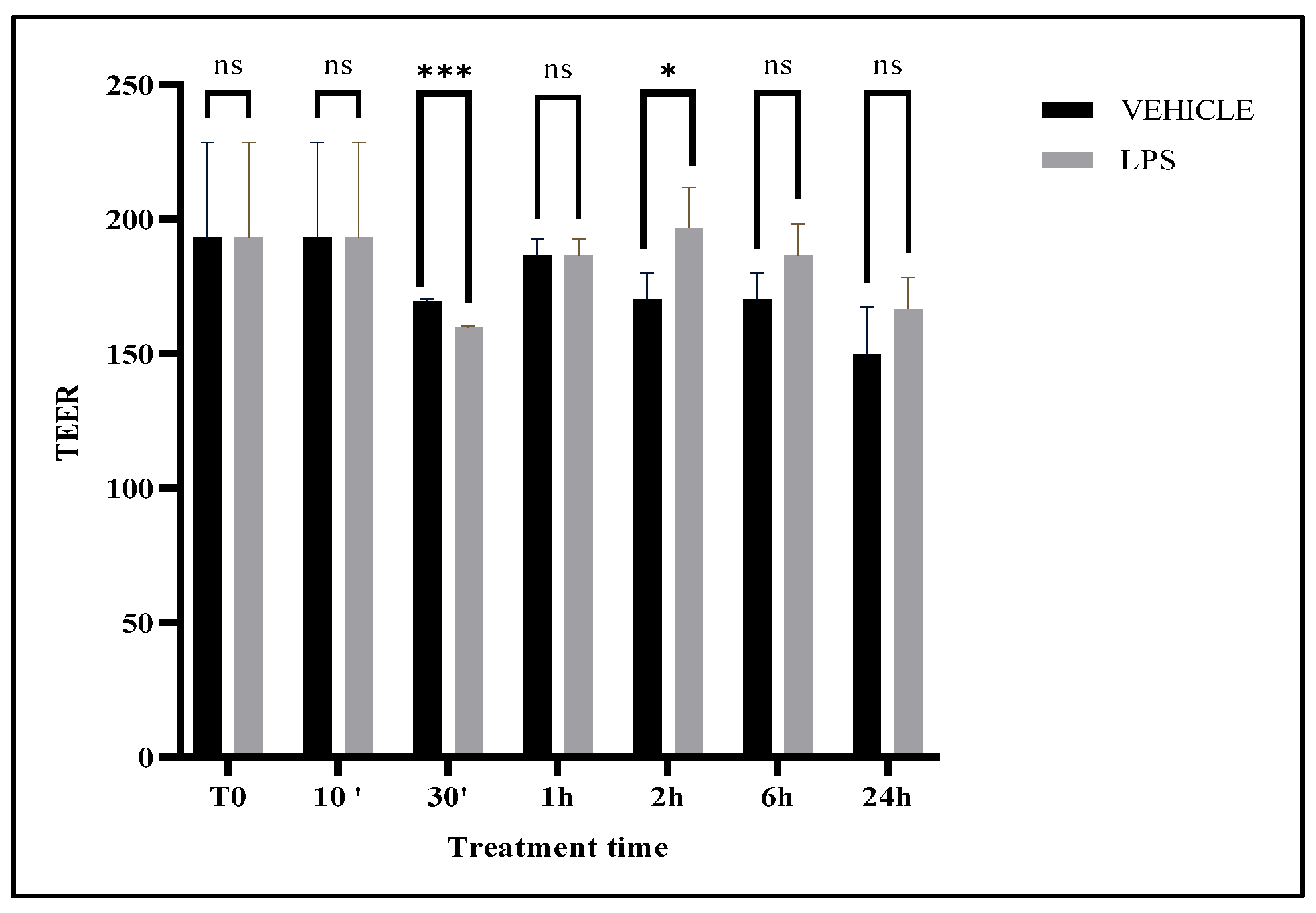
The outcomes of this study underscored that LPS (1 μg/ml) treatment caused a time-dependent decrease in TEER values, with the resistance dropping significantly over several hours. Indeed, after 30 min and 2 h of exposure to LPS the TEER value of Caco-2 monolayer was significantly (P <0.05) lower (ca. 160 Ω/cm2) than the untreated cells; however, no significant differences were found when TEER was measured at the subsequent times (1, 6, 24 h) (Figure 1). Thus, a lower TEER value directly correlates with increased permeability of the cell monolayer, indicating disruption of the intestinal barrier that onset of a “leaky”. This damage to the tight junctions can trigger the inflammatory pathways, leading to the release of pro-inflammatory cytokines, as previously reported (Tunisi et al., 2019).
Figure 1.
Average trans-epithelial electrical resistance (TEER; Ω/cm2) measurements of Caco-2 monolayer after 10’, 30’, 1h, 2h, 6h and 24h after treatment with LPS (positive control).
Figure 1.
Average trans-epithelial electrical resistance (TEER; Ω/cm2) measurements of Caco-2 monolayer after 10’, 30’, 1h, 2h, 6h and 24h after treatment with LPS (positive control).
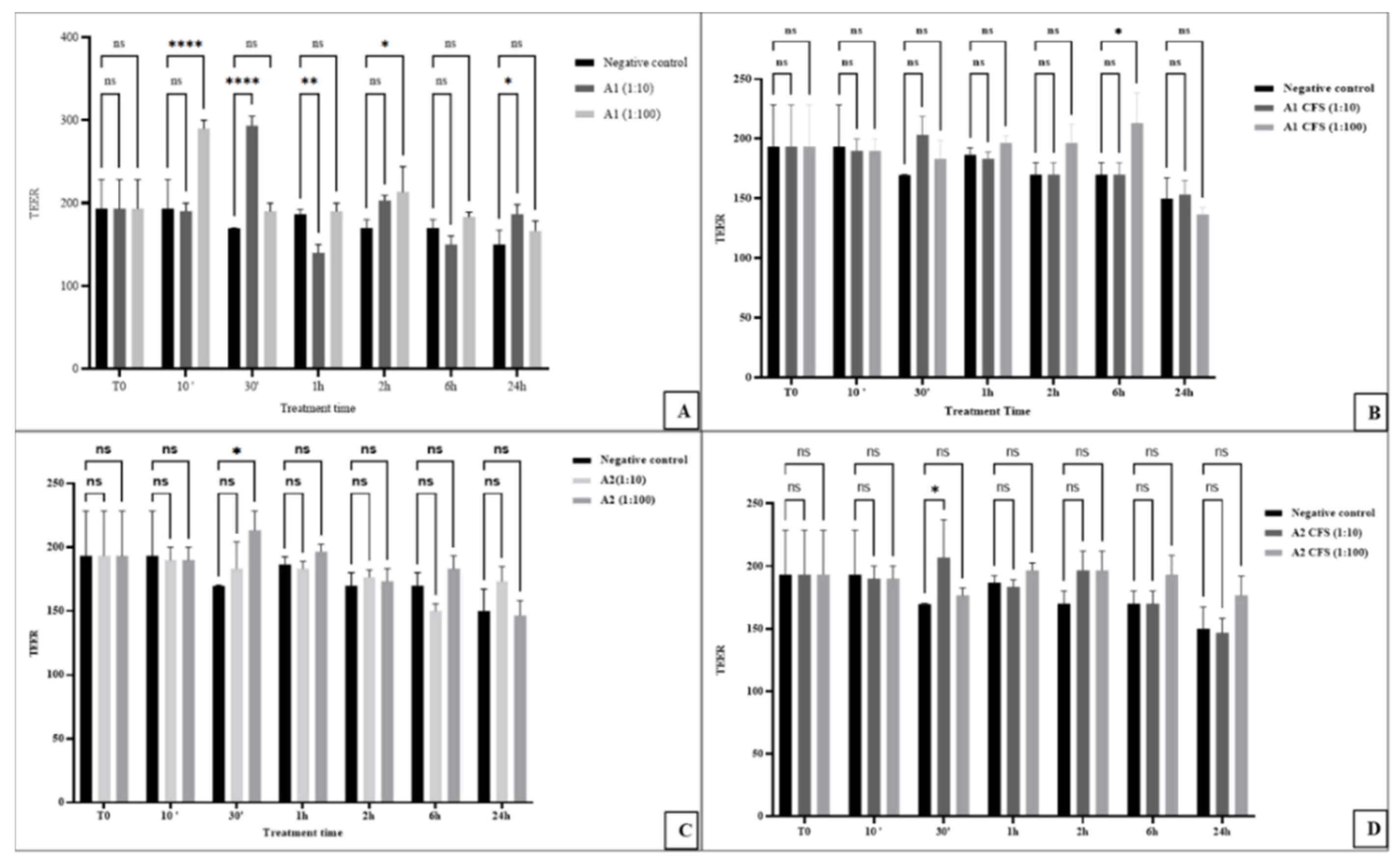
Otherwise, Caco-2 monolayer exposed to L. paraplantarum A1 biomass diluted 1:10 showed increased (P < 0.05) TEER values after 10 min and 2 h; afterwards no significant (P > 0.05) differences were found (Figure 2A). When the monolayer had been exposed to the biomass of the same LAB diluted 1:100, increased (P < 0.05) TEER values were found after 30 min, and 24 h. However, a decrease (P < 0.05) of TEER was strikingly found after 1 h (Figure 2A). When exposed to the CFS of L. paraplantarum A1, significant (P < 0.05) higher values of TEER of Caco-2 monolayer were found after 30 min (CFS of L. paraplantarum A1 diluted 1:10) and 6 h (diluted 1:100) than the untreated cells (Figure 2B). However, treating of Caco-2 monolayer by the biomass of L. plantarum A2 (at both dilutions) (1:100) resulted in higher (P < 0.05) TEER (183 – 213 Ω/cm2) only at 30 min, compared to the negative control (Figure 2C). At 30 min and 2 h, TEER of the monolayer treated with the diluted (1:10) CFS of L. plantarum A2 was significantly higher than the untreated cells (Figure 2D). Therefore, the treatment of Caco-2 cell line by probiotic LAB reported that the two tested lactobacilli could moderate and influence the cell-cell interaction and cell barrier integrity based on the TEER measurement, but in dose-strain-time-dependent manner. So, L. plantarum broth’s treated Caco-2 monolayers had significantly boosted the TEER level with both dilution (1:10, v/v) and (1:100, v/v) to approximately 294 Ω/cm2 and 290 Ω/cm2 at 30’ and 10’ respectively, however L. paraplantarum had moderate fundings of the TEER value after 30’ of incubation reaching approximately 213 Ω/cm2 and 183 Ω/cm2 for both dilution (1:100) and (1:10) respectively. Those results are consistent with previous studies that showed a noticeable increase in TEER values with L. plantarum species, which is not unexpected as different strains of L. plantarum may influence the cell membrane integrity in vitro in different ways. Indeed, some strains may not affect TEER values, while others can cause an increase, as reported in previous studies carried out before. Consequently, outcomes are typically evaluated through dose-time-effect safety experiments in Caco-2 cells (Klingberg et al., 2005; Anderson et al., 2010; Tunisi et al., 2019; Dasriya et al., 2024; Mosiej et al., 2025). Besides, when the CFS of the both lactobacilli were tested, the TEER value had significantly (P < 0.05) and gradually increased over time of co-culture, then the measurement decreased cautiously alike strain-dose-time dependent. These results accentuate the importance of bioactive molecules present in the CFS of lactobacilli and they put in value their crucial role in cell-cell interaction and the modulation of the cell integrity compared to untreated cells. Our outcomes were in concordance with other scientific reports (Dasriya et al., 2024; Mosiej et al., 2025).
Figure 2.
Average trans-epithelial electrical resistance (TEER; Ω/cm2) measurements of Caco-2 monolayer after 10’, 30’, 1h, 2h, 6h and 24h after treatment with L. paraplantarum cells A1 (1 :10) and (1 :100) (A); with A1 CFS (1 :10) and (1 :100) (B); L. plantarum cells A2 (1 :10) and (1 :100) (C) and with A2 CFS (1 :10) and (1 :100) (D).
Figure 2.
Average trans-epithelial electrical resistance (TEER; Ω/cm2) measurements of Caco-2 monolayer after 10’, 30’, 1h, 2h, 6h and 24h after treatment with L. paraplantarum cells A1 (1 :10) and (1 :100) (A); with A1 CFS (1 :10) and (1 :100) (B); L. plantarum cells A2 (1 :10) and (1 :100) (C) and with A2 CFS (1 :10) and (1 :100) (D).
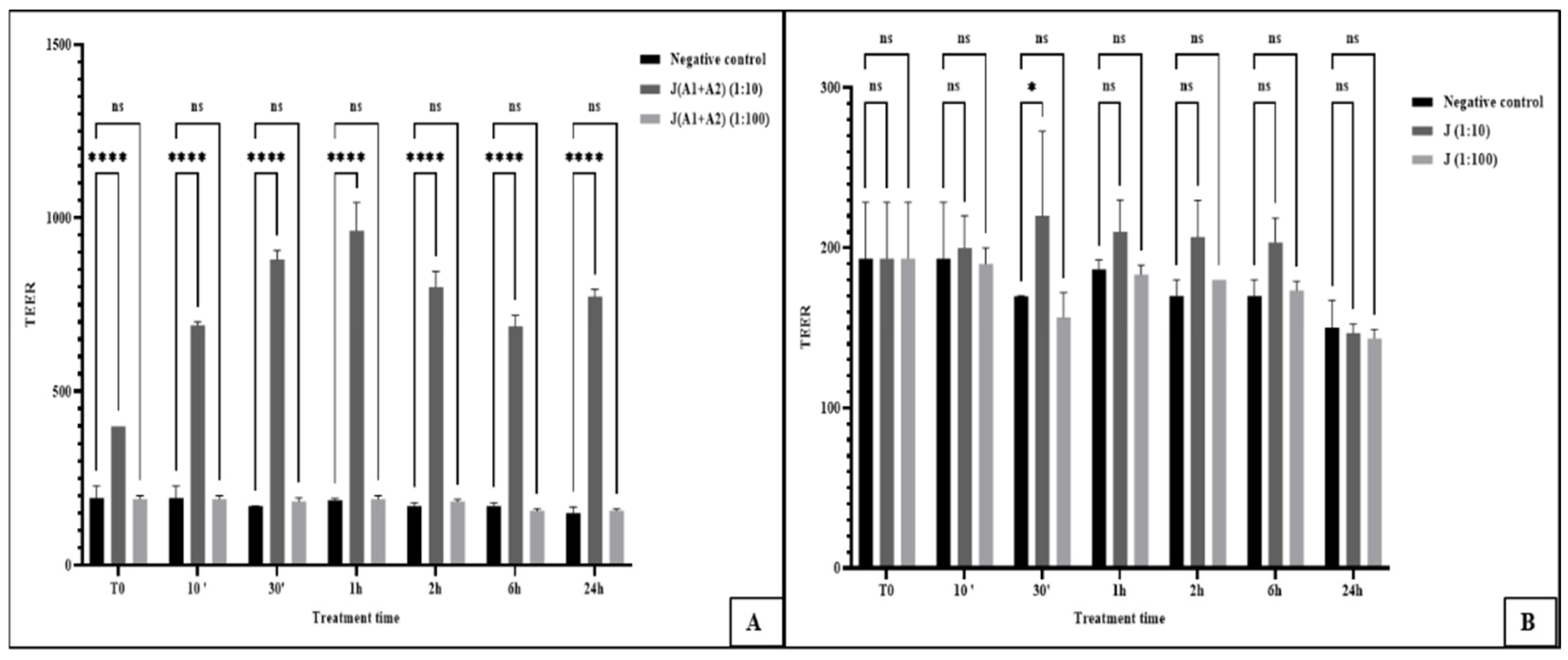
When the Caco-2 monolayer was exposed to the zgougou juice fermented by L. paraplantarum A1 and L. plantarum A2 (J A1+A2) and diluted 1:10, we found at all time-points higher (P < 0.05) values of TEER than the negative control. No significant (P > 0.05) differences were observed when cells were treated with J A1+A2 diluted 1:100 (Figure 3A). Treatment of Caco-2 monolayer with spontaneously fermented zgougou juice (J) resulted in higher (P < 0.05) TEER than the negative control, just at 30 min when cells were treated with J (1:10), (Figure 3B), however the treatment with J (1:100) doesn’t have any effect on TEER.
Figure 3.
Average trans-epithelial electrical resistance (TEER; Ω/cm2) measurements of Caco-2 monolayer after 10’, 30’, 1h, 2h, 6h and 24h after treatment with fermented zgougou juice driven with LAB starter culture J (A1+A2) at (1 :10) and (1 :100) (A) and zgougou juice spontaneously fermented J at (1 :10) and (1 :100) (B).
Figure 3.
Average trans-epithelial electrical resistance (TEER; Ω/cm2) measurements of Caco-2 monolayer after 10’, 30’, 1h, 2h, 6h and 24h after treatment with fermented zgougou juice driven with LAB starter culture J (A1+A2) at (1 :10) and (1 :100) (A) and zgougou juice spontaneously fermented J at (1 :10) and (1 :100) (B).
From the membrane integrity study, we concluded that the presence of fermented zgougou isolates did not affect the permeability of the intestinal lining. On the other hand, the non-dairy food isolate could strengthen intestinal barrier function, as demonstrated by an increase in TEER values over the incubation period. However, the intestinal epithelial system is a home of immune cells which function in close relation with the epithelial cells to maintain the homeostasis of the intestine. In fact, the intestinal epithelial cells are in permanent contact with the luminal contents, including the microbiota and they have a crucial role in the maintenance of barrier integrity, which enables the permeability of essential ions, nutrients, and water but circumscribes the entry of bacterial toxins and pathogens. The cellular integrity is maintained by the epithelial junction adhesion complex, the mucous layer, and released products by both host cells and bacteria (Yuki et al., 2007; Ohland & Macnaughton, 2010; Heli et al., 2012, Han et al., 2023). Our findings are in concordance with previous reports about plant-based fermented alternatives which are rich in microbes and their metabolites, in addition to their phytochemicals, so these metabolites can modulate the permeability of the intestinal barrier (Scott et al., 2020; Han et al., 2023). In the current study, zgougou could strengthen the intestinal barrier function, as demonstrated by a very great increase in TEER levels over all the incubation period. Different studies confirmed and exhibited that plants-based fermented foods such as kefir, dahi, and the korean kimchi, faba bean-using different probiotic LAB strains-are found to be advantageous against many diseases such as cancers, irritable bowel syndromes, anorexia nervosa, cardiovascular diseases, and diabetes (Chopra & Mathur, 2013; S¸engün & Güney, 2021; Nithya et al., 2023; Patel et al., 2023). Likewise, L. acidophilus C11 and L. plantarum C06, isolated from pickled cabbage, showed good in vitro performance in terms of duodenal cell adhesion and gut biotic stress tolerance (Wang et al., 2010, Aslam et al., 2020). It has also been shown that anchoring of microorganisms onto the vegetable surface and present in plant-origin ferment food (e.g., kimchi, sauerkraut, natto, tempeh, fermented soy, pickles, kombucha...) yield to cell integrity and stability during their passage through the gastrointestinal tract, until their subsequent release in the colon (Lavermicocca, 2006, Han et al., 2023).
3.2. Differential Modulation of Tight Junction Genes (CLDN2 and OCLN)
Traditional fermented beverages and products, predominantly dairy-based, may act as efficient vehicles for probiotics, ensuring their successful delivery to the host and contributing to the associated health benefits. But in last decades, an increase in demand for plant-based products, novel fermented drinks have been developed from plant-based alternatives such as soymilk kefir and fermented soymilk (Ashaolu, 2020; Tiss et al., 2020; Ilango & Antony, 2021). Similarly, the intensive use of Aleppo pine (Pinus halepensis Miller) seeds, named “zgougou”, particularly in Tunisia and generally in other Arabic countries, is not just linked to food. Zgougou is also highly valuated for its therapeutic potential, owing to its richness in bioactive compounds such as essential fatty acids, flavonoids, and phenolic compounds, which can function as antioxidant agents or supplementary additives in food fermentation and cosmetic formulations. The potential health benefits of these compounds, such as their anti-inflammatory properties, have been investigated. Nevertheless, the phenolic compounds found in Aleppo pine seeds - including catechins, flavonoids, and phenolic acids – exhibit strong antioxidant potential (Nasri et al., 2004; Cheikh-Rouhou et al., 2006; Missaoui et al., 2019; Aslam et al., 2020; Minervini et al., 2020; Han et al., 2023) and may contribute to the attenuation of inflammation through free radical scavenging and the inhibition of inflammatory pathways. Indeed, flavonoids and fatty acids - particularly linoleic acid and α-linolenic acid - have been shown to possess anti-inflammatory properties by inhibiting inflammatory enzymes such as cyclooxygenase (COX) and lipoxygenase (LOX), as well as by modulating the production of pro-inflammatory mediators and cytokines. Consequently, these compounds may help prevent or treat a variety of human and animal diseases and pathological conditions (Missaoui et al., 2019).
Many transmembrane proteins, including the occlusive protein (OCLN), synaptic connexin (CLDN), and junction adhesion molecules (JAMS), (Colegio et al., 2003; Campbell et al., 2017), are included in membrane proteins. Tight Junctions (TJs) between IECs have the ability to transport substances selectively, keep hazardous compounds and pathogenic bacteria out of the intestinal lumen, and preserve the normal function of the intestinal barrier. TJ-related complex proteins are membrane and intracellular proteins. (Suzuki, 2020; Paradis et al., 2021). However, probiotics influence the function of the intestinal barrier via controlling the expression of genes and proteins in IECs that are involved in TJs signaling. Overall, it seems that Lactobacillus plantarum affects tight junction proteins in the gastrointestinal tract, including OCLN, CLDN2, and LCN2, which helps to alter immune responses and preserve the integrity of the intestinal barrier. However, the precise processes and outcomes can differ based on the host, the gut microbiota, and the strain of L. plantarum. To completely understand L. plantarum’s effects on tight junction proteins and their consequences for gastrointestinal health and illness, more research is required (Ohland and Macnaughton, 2010; Sharma et al., 2010; Ashida et al., 2011; La Fata et al., 2018; Wierzbicka et al., 2021; Wei et al., 2022; Zhang et al., 2022; Oudat & Okour., 2025; Mosiej et al., 2025). Indeed, a healthy intestinal barrier prevents the translocation of harmful substances from the gut lumen into the bloodstream, thereby reducing systemic inflammation.
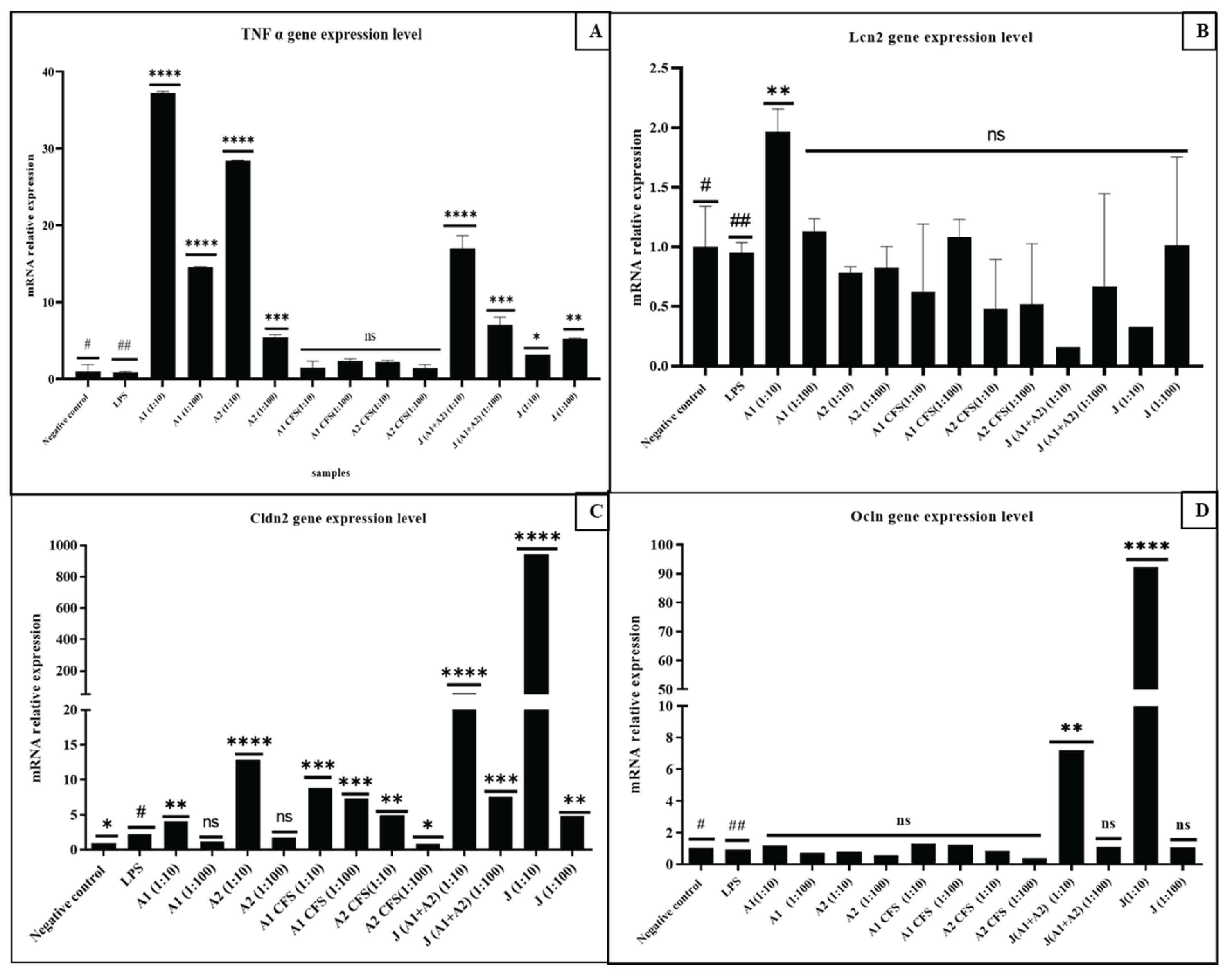
In the current research, Caco-2 monolayer exposed to L. paraplantarum A1 biomass, at both dilutions, showed an increased (P < 0.05) level of expression of TNF (Figure 4A) and LCN2 (Figure 4B) genes. In addition, the 1:10 diluted CFS of this strain increased (P < 0.05) the transcription level of CLDN2 gene of the monolayer (Figure 4D). When cells were treated with L. plantarum A2 biomass (diluted 1:10), they showed an increased (P < 0.05) expression of CLDN2 gene. Exposure of Caco-2 monolayer to J A1+A2 (diluted 1:10) favoured (P < 0.05) the transcription of TNF, OCLN, and CLDN2 genes (Figure 4A,C,D). The expression of the latter two genes was strongly increased (P < 0.05) in the monolayer co-incubated with the 1:10 diluted zgougou (J).
Our outcomes highlighted also that the used fermented plant-based matrix was the fermented zgougou in two types spontaneously and with starter culture LAB-based. Inspite, both fermented matrices diluted at (1:10) critically enhanced and increase the level of CLDN-2 mRNA relative gene expression which reach ~100pg/ml and ~1000pg/ml respectively (Figure 4C. Otherwise those two food matrices had the ability to significantly upregulate the mRNA relative gene expression of the OCLN to reach 8pg/ml and 100pg/ml when J(A1+A2) diluted (1:10) and J diluted (1:10) were tested respectively (Figure 4D). Those findings confirmed the TEER measurement and cell integrity results showed below which highlighted the capacity of both zgougou juices. This great activity of both samples J (A1+A2) at (1:10) and J (1:10) of fermented zgougou might be due to the presence of different bioactive compounds and probiotic strains which could alter the pH value of the environment, reduce the availability of oxygen, and produce a competitive environment in the gut against the growth of pathogens. It also alters the host’s immunological response thus strengthening the immune system and changing the composition of the microbiota of the host and their metabolic activity and reinforce the cell-cell integrity via the enhancement of TJPs via different signalling pathways.
In this present study, the outcomes showed that the tested strains have a moderate activity in the claudin-2 (CLDN-2) and occludin (OCLN) mRNA relative gene expression, and the response was strain-depending effects and these results are contradictory with previous reports that demonstrated that, L. plantarum MB452 upregulated the gene and protein expression of ZO1, ZO2, occludin, and cingulin and reinforced the expression of TJP-degrading genes (such as itchy E3 ubiquitin protein ligase and snail family transcriptional regressor 1), as an additional mechanism to enhance TJs integrity which improves intestinal barrier function (Anderson et al., 2010). Additionally, Karczewski et al., (2010) speculated that the injection of L. plantarum WCFS1 strain into the duodenum of healthy subjects enhanced the expression of occluding- and ZO1-coding genes, and that the TLR2 pathway activation affected the expression and distribution of TJ proteins. Another study revealed, in an in vitro model, that L. plantarum could regulate protein levels and distribution such as claudin1, occludin, JAM1, and ZO1, which allow to protect Caco-2 cells from entero-invasive E. coli and improve the intestinal barrier function (Qin et al., 2009).
Figure 4.
Differential mRNA levels of pro-inflammatory cytokine and tight junctions’ marker genes in Caco2 cells incubated with different samples as determined by RT-qPCR after 24 h of treatment. A. Normalized mRNA levels of pro-inflammatory cytokine TNFα seeded on multiwell plates. B Normalized mRNA levels of LCN2 marker genes in Caco-2 cells, C. Normalized mRNA levels of Cldn 2 marker genes in Caco-2 cells, D. Normalized mRNA levels of Ocln marker genes in Caco-2 cells which were seeded, respectively, on the top and the bottom of the Transwell system, after 24 h of incubation. #: negative control; ##: positive control; ns: no significance reported.
Figure 4.
Differential mRNA levels of pro-inflammatory cytokine and tight junctions’ marker genes in Caco2 cells incubated with different samples as determined by RT-qPCR after 24 h of treatment. A. Normalized mRNA levels of pro-inflammatory cytokine TNFα seeded on multiwell plates. B Normalized mRNA levels of LCN2 marker genes in Caco-2 cells, C. Normalized mRNA levels of Cldn 2 marker genes in Caco-2 cells, D. Normalized mRNA levels of Ocln marker genes in Caco-2 cells which were seeded, respectively, on the top and the bottom of the Transwell system, after 24 h of incubation. #: negative control; ##: positive control; ns: no significance reported.
3.3. Modulation of TNFalpha Gene Expression in Caco-2 Monolayers
TNFα is a cytokine with pro-inflammatory properties but its role in the immune response is complex, and in some cases, it may be involved in promoting immune responses against pathogens, while in other cases, it may contribute to inflammatory conditions. Although, the gut microbiome, which includes LAB, plays a crucial role in regulating immune responses, and the bacterial strains can influence the expression of various immune-related genes, including those involved in inflammation and immune cell development. Despite, different LAB strains can have varying effects on TNF-α production, highlighting the need for careful selection and characterization of probiotic strains, for this reason the ability of LAB to modulate TNF-α expression is strain-dependent. Depending on the context, LAB-induced TNF-α production could contribute to either pro-inflammatory or anti-inflammatory effects. In the present investigation, A1 (1:10), A2 (1:10) significantly increased the mRNA expression level of the pro-inflammatory cytokine TNFα. In addition, A1 (1:100) and J (A1+A2) (1:10) also enhanced TNFα gene expression (P<0.05) (Figure 4A). Although some treatments (exposure to lactobacilli biomass or to zgougou fermented with those lactobacilli) indicated a higher expression of TNF gene, TNFα was ever below the detection limit, as assessed through the ELISA assay (data not shown). The current findings showed that the tested LAB strains belonging to L. plantarum and L. paraplantarum groups could enhance the relative gene expression of TNF-α and also the gene expression level of the antimicrobial protein Lipocalin-2 (LCN-2) which is involved in host defence and inflammation regulation. This highlighted and confirmed the previous studies which reported that the inhibition or induction of TNF-α expression by probiotics might provide either immune-suppressant or immune-stimulating effects, and this is strain and context dependent. Probiotics that inhibit or induce TNF-α expression may have immune-stimulating or immune-suppressant effects, respectively. LAB modulates TNF-α signaling by regulating important pathways such as MAPK and NF-κB (Saha et al., 2017; Bae et al., 2022; Sahu et al., 2023; Kobayashi et al., 2023).
Recent studies reported that supernatants and cell extracts from L. plantarum and L. reuteri, when tested in vitro on human epithelium model (monolayers of caco-2 cells), can induce the translocation of the scaffold proteins to the tight junction region, enhance the toll like receptor (TLR2) signaling, and reduce the capacity for cell invasion and progression (Karczewski et al., 2010). Indeed, the colonization of Lactobacilli in the site of the primary tumor may be beneficial for prevention of metastases (Yang et al., 2014). In addition, probiotic lactobacilli (L. casei, L. rhamnosus, L. acidophilus, L. crispatus…) and their CFS could regulate the MMP levels in cancer cells, inhibit the cell invasion in vitro (colon cancer, HeLa cells, HT-29 cells) which speculate the anti-metastatic power of the bacterial bioactive substances (Escamilla et al., 2012; Nouri et al., 2016). L. acidophilus might be effective in metastasis suppression through their lipoteichoic acid (LTA) ability to up-regulate of the protein expression of ICAM5, RUNX3, TIMP2 and RASSF1A in human colon carcinoma cell line HT-29 (Lightfoot et al., 2013; Guo et al., 2018). Furthermore, another study, in vitro, showed that L. casei prompted the apoptotic cell death through up-regulation of TNFα and TRAIL ligand in both murine (CT26) and human (HT-29) cell lines model (Tiptiri-Kourpeti et al., 2016; Kobayashi et al., 2025).
Moreover, research has demonstrated the health benefits of fermented cabbage products like fermented soy, kimchi and sauerkraut, kombucha. Those fermented foods are well known for their anti-inflammatory, antioxidant, and cancer-preventive qualities due to their richness on phenolic and bioactive compounds (Di Cagno et al., 2016). Inspite, fermented soy products (miso, natto, and tempeh) exhibited, in vitro, a protective TJ proteins against oxidative stress and endotoxin challenge (Kadar et al., 2020). Similarly, kimchi and sauerkraut, wich contain LAB, were found to increase ZO-1 expression in colitis models (Lee et al., 2017), while kombucha polyphenols were reported to maintain gut permeability under oxidative stress conditions (Kim et al., 2023). Another study showed that fermented kiwi fruit exerts a protective effect on intestinal permeability, which is associated with reduced mitochondrial stress and the preservation of mitochondria morphology and respiratory function (Ma et al. 2023). In addition, fermentation of kiwi fruit using L. plantarum strain improved the LPS-induced leaky gut through the preservation of the intestinal tight junction structure and increase the expression of associated proteins (Bae et al., 2022; Ma et al., 2023; Haque et al., 2024).
Otherwise, the present study was contradictory to another previous report demonstrated L. plantarum inhibited TNF-α-induced decrease in TEER at 24 h and 36 h after treatment (P < 0.05), so that, the epithelial barrier function of TNF-α-stimulated Caco-2 cells was preserved by soy fermented by L. plantarum (Lin et al., 2016).
Otherwise, microbial metabolites produced in fermented food for dairy products also have immunomodulatory abilities. For example, oleamide, a microbial metabolite produced by Penicillium candidum, is present in products such as camembert cheese. Oleamide has been shown to suppress TNF α release from microglia by acting as an agonist for the P2Y and cannabinoid receptors (Kita et al., 2019). In addition, in the setting of inflammatory bowel disease (IBD), the effects of fermented food products on intestinal integrity have been the subject of numerous recent investigations. When fed fermented milk supplemented with Bacillus subtilis, mice with IBD demonstrated improved intestinal shape and tight junction protein levels compared to the disease control group (Zhang et al., 2021). Furthermore, in vitro and in a mouse model of IBD treated with fermented barley and soybean, this restoration of intestinal tight junction proteins has also been noted (Woo et al., 2016).
By this study we investigate the importance of the fermented zgougou (Aleppo pine seeds), even fermented spontaneously or with starter culture containing endogenous probiotic LAB strains, in the gut prevention and preservation from different problems and inflammatory diseases as an alternative remedy.
4. Conclusions and Perspectives
Fermented plant-origin foods are becoming wide interest worldwide as healthful alternatives because of their richness in phytochemicals and bioactive substances, and particularly their richness on probiotic microorganisms (LAB, yeasts …) and their by-products which highlight their potential health benefits and their superior biological activities. Reportedly, with an emphasis on their potential as probiotics this present innovative investigation explores the strain-specific modulatory characteristics of live Lactiplantibacillus plantarum strains A1 and A2 isolated for the first time from the Tunisian traditional fermented foods called “zgougou” and especially the main food matrix that may confer a health benefits. Thus, it is so important to consider in this present study that the interactions between the different components of the tested samples could enhance the intestinal barrier integrity and improve the cell-cell interactions based on the TEER measurement analysis. Indeed, fermentation by starter culture may increase the positive effects of Tunisian zgougou juice obtained upon both spontaneous (J) and driven fermentation (J(A1+A2)) with autochthonous probiotic bacteria Lactiplantibacillus plantarum (LP) species A1 and A2 due to an increase in bioactive compounds that highlight its importance in the reinforcement of the tight junction proteins (OCLN and CLDN-2), its modulatory effect on the antimicrobial protein (LCN-2) via the regulation of their mRNA relative gene expression. Furthermore, the results of this investigation are quite likely and consistent with previous studies on probiotics. Tunisian Zgougou juice (J (A1+A2)) and (J) has the qualities of a symbiotic food (containing both probiotics and prebiotics/beneficial metabolites) and can function as a powerful anti-inflammatory agent by influencing the TNF-α gene expression in intestinal cells, especially when fermented with specific LAB strains. This research could be an initiative a for screening process to identify new candidate probiotic bacteria and to investigate new fermented plant-origin food matrix that was traditionally and anecdotally evident and useful in Tunisia and some Arabic countries due to its potential health benefits. However, it’s so important to note that while Aleppo pine nuts (zgougou) showed promise for their anti-inflammatory effects, further research, including in vivo experiments and clinical trials, is needed to fully understand their mechanisms and potential therapeutic applications. As with any natural remedy, it’s advisable to consult with a healthcare professional before incorporating Aleppo pine seeds or any new supplement into your diet or treatment regimen, especially if you have existing health conditions or are taking medications.
Author Contributions
Conceptualization, M.D.A., and J.M; Formal analysis, L.A.; Investigation, J.M., M.L.; Writing-original draft, J.M., F.M.; Writing-review & editing, F.M., M.D.A.; Supervision, F.M & L.A.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Caco-2 cell | Caucasian Colon Adenoarcinoma Cells |
| CFS | Cell-Free Supernatant |
| CLDN2 | Claudin-2 |
| HT-29 | Human Colon Carcinoma Cell Line |
| IBD | Inflammatory Bowel Disease |
| IECs | Intestinal Epithelial Cells |
| JAMS | Junction Adhesion Molecules |
| J | Zgougou Juice Obtained Upon Both Spontaneous |
| J(A1+A2) | Zgougou Juice Driven Fermentation With Autochthonous Probiotic Bacteria |
| LAB | Lactic Acid Bacteria |
| LCN2 | Lipocalin-2 |
| LP | LactiplantibacillusPlantarum |
| LPS | Escherchia Coli Lipo Poly Saccharide |
| LTA | Lipoteichoic Acid |
| OCLN | Occludin |
| RT-PCR | Real Time- Polymorphism Chain Reaction |
| TEER | Trans Epithelial Electrical Resistance |
| TJs | Tight Junctions |
| TJP | Tight Junction Proteins |
| TLR2 | Toll Like Receptor |
| TNFα | Tumour Necrosis Factor Alpha |
References
- Ait Atmane, S.; Aksoylu Özbek, Z.; Günç Ergönül, P.; Khettal, B. Valorization of Pinus halepensis Mill. seed oil: Physicochemical characteristics, bioactive compounds, and antioxidant activity as affected by location and extraction method. J. Food Process. Preserv. 2021, 45, e15548. [Google Scholar] [CrossRef]
- Ait Atmane, S.; Batır, M. B.; Özbek, Z. A.; et al. Cold pressed Pinus halepensis Mill. seed oil for potential health applications: Analgesic, anti-inflammatory effects, and assessment of inflammatory mediators by RT-qPCR in skin wound healing. J. Ethnopharmacol. 2024, 319, 117157. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Cookson, A.L.; McNabb, W.C.; et al. Lactobacillus plantarum MB452 enhances the function of the intestinal barrier by increasing the expression levels of genes involved in tight junction formation. BMC Microbiol. 2010, 10, 316. [Google Scholar] [CrossRef]
- Ashaolu, T. J. Immune boosting functional foods and their mechanisms: A critical evaluation of probiotics and prebiotics. Biomed. Pharmacother. 2020, 130, 110625. [Google Scholar] [CrossRef]
- Ashida, N.; Yanagihara, S.; Shinoda, T.; Yamamoto, N. Characterization of adhesive molecule with affinity to Caco-2 cells in Lactobacillus acidophilus by proteome analysis. J. Biosci. Bioeng. 2011, 112, 333–337. [Google Scholar] [CrossRef]
- Aslam, H.; Green, J.; Jacka, F. N.; et al. Fermented foods, the gut and mental health: a mechanistic overview with implications for depression and anxiety. Nutr. Neurosci. 2020, 23, 659–671. [Google Scholar] [CrossRef]
- Bae, W.Y.; Jung, W.H.; Shin, S.L.; et al. Investigation of Immunostimulatory Effects of Heat-Treated Lacti-plantibacillus plantarum LM1004 and Its Underlying Molecular Mechanism. Food Sci. Anim. Resour. 2022, 2, 42–1031. [Google Scholar]
- Balda, M. S.; Matter, K. Transmembrane proteins of tight junctions. Semin. Cell Dev. Biol. 2000, 11, 281–289. [Google Scholar] [CrossRef]
- Bamola, V.D.; Dubey, D.; Samanta, P.; et al. Role of a probiotic strain in the modulation of gut microbiota and cytokines in inflammatory bowel disease. Anaerobe 2022, 78, 102652. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Muñoz-Quezada, S.; Gómez-Llorente, C.; et al. Lactobacillus paracasei CNCM I-4034 and its culture supernatant modulate Salmonella-induced inflammation in a novel transwell co-culture of human intestinal-like dendritic and Caco-2 cells. BMC Microbiol. 2015, 15, 79. [Google Scholar] [CrossRef]
- Botella, L.; Diez, J. J. Phylogenic diversity of fungal endophytes in Spanish stands of Pinus halepensis. Fungal Divers. 2011, 47, 9–18. [Google Scholar] [CrossRef]
- Campbell, H. K.; Maiers, J. L.; DeMali, K. A. Interplay between tight junctions & adherens junctions. Exp. Cell Res. 2017, 358, 39–44. [Google Scholar] [CrossRef]
- Cheikh-Rouhou, S.; Hentati, B.; Besbes, S.; et al. Chemical composition and lipid fraction characteristics of Aleppo pine (Pinus halepensis Mill.) seeds cultivated in Tunisia. Food Sci. Technol. Int. 2006, 12, 407–415. [Google Scholar] [CrossRef]
- Chopra, R.; Mathur, S. Probiotics in dentistry: A boon or sham. Dent. Res. J. 2013, 10, 302. [Google Scholar]
- Colegio, O. R.; Van Itallie, C.; Rahner, C.; Anderson, J. M. Claudin extracellular domains determine paracellular charge selectivity and resistance but not tight junction fibril architecture. Am. J. Physiol. Cell Physiol. 2003, 284, C1346–C1354. [Google Scholar] [CrossRef] [PubMed]
- Dasriya, V. L.; Samtiya, M.; Ranveer, S.; et al. Modulation of gut--microbiota through probiotics and dietary interventions to improve host health. J. Sci. Food Agric. 2024, 104, 6359–6375. [Google Scholar] [CrossRef]
- Di Cagno, R.; Filannino, P.; Gobbetti, M. Fermented foods: fermented vegetables and other products. In Encyclopedia of Food and Health; Elsevier Inc.: 2015; pp. 668–674.
- Escamilla, J.; Lane, M. A.; Maitin, V. Cell-free supernatants from probiotic Lactobacillus casei and Lactobacillus rhamnosus GG decrease colon cancer cell invasion in vitro. Nutr. Cancer 2012, 64, 871–878. [Google Scholar] [CrossRef]
- Guo, S.; Li, L.; Xu, B.; et al. A simple and novel fecal biomarker for colorectal cancer: ratio of Fusobacterium nucleatum to probiotics populations, based on their antagonistic effect. Clin. Chem. 2018, 64, 1327–1337. [Google Scholar] [CrossRef]
- Han, H.; You, Y.; Cha, S.; et al. Multi-Species Probiotic Strain Mixture Enhances Intestinal Barrier Function by Regulating Inflammation and Tight Junctions in Lipopolysaccharides Stimulated Caco-2 Cells. Microorganisms 2023, 11, 656. [Google Scholar] [CrossRef]
- Haque, M.; Kaminsky, L.; Abdulqadir, R.; et al. Lactobacillus acidophilus inhibits the TNF-α-induced increase in intestinal epithelial tight junction permeability via a TLR-2 and PI3K-dependent inhibition of NF-κB activation. Front. Immunol. 2024, 15, 1348010. [Google Scholar] [CrossRef]
- Huang, F.C. Upregulation of Salmonella-Induced IL-6 Production in Caco-2 Cells by PJ-34, PARP-1 Inhibitor: Involvement of PI3K, p38MAPK, ERK, JNK, and NF-κB. Mediators Inflamm. 2009, 103890. [Google Scholar] [CrossRef]
- Ilango, S.; Antony, U. Probiotic microorganisms from non-dairy traditional fermented foods. Trends Food Sci. Technol. 2021, 118, 617–638. [Google Scholar] [CrossRef]
- Kadar, A.D.; Astawan, M.; Putri, S.P.; Fukusaki, E.J.M. Metabolomics-Based Study of the Effect of Raw Materials to the End Product of Tempe—An Indonesian Fermented Soybean. Metabolites 2020, 10, 367. [Google Scholar] [CrossRef]
- Kadri, N.; Khettal, B.; Aid, Y.; Kherfellah, S.; Sobhi, W.; Barragan-Montero, V. Some physicochemical characteristics of pinus (Pinus halepensis Mill., Pinus pinea L., Pinus pinaster and Pinus canariensis) seeds from North Algeria, their lipid profiles and volatile contents. Food Chem. 2015, 188, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, J.; Troost, F. J.; Konings, I.; et al. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef]
- Kim, H.; Hur, S.; Lim, J.; et al. Enhancement of the phenolic compounds and antioxidant activities of Kombucha prepared using specific bacterial and yeast. Food Biosci. 2023, 56, 103431. [Google Scholar] [CrossRef]
- Kita, M.; Ano, Y.; Inoue, A.; Aoki, J. Identification of P2Y receptors involved in oleamide-suppressing inflammatory responses in murine microglia and human dendritic cells. Sci. Rep. 2019, 9, 3135. [Google Scholar] [CrossRef]
- Klingberg, T. D.; Pedersen, M. H.; Cencic, A.; Budde, B. B. Application of measurements of transepithelial electrical resistance of intestinal epithelial cell monolayers to evaluate probiotic activity. Appl. Environ. Microbiol. 2005, 71, 7528–7530. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Imai, Y.; Shimizu, Y.; et al. Lactobacillus delbrueckii subsp. bulgaricus 2038 and Streptococcus thermophilus 1131 ameliorate barrier dysfunction in human induced pluripotent stem cell-derived crypt-villus structural small intestine. Front. Immunol. 2025, 16, 1585007. [Google Scholar] [CrossRef]
- Kobayashi, K.; Mochizuki, J.; Yamazaki, F.; Sashihara, T. Yogurt starter strains ameliorate intestinal barrier dysfunction via activating AMPK in Caco-2 cells. Tissue Barriers 2023, 12. [Google Scholar] [CrossRef]
- La Fata, G.; Weber, P.; Mohajeri, M. H. Probiotics and the gut immune system: indirect regulation. Probiotics Antimicrob. Proteins 2018, 10, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Lavermicocca, P. Highlights on new food research. Dig. Liver Dis. 2006, 38, S295–S299. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, D. S.; Cho, J.; et al. Perilla frutescens Britton: A comprehensive study on flavor/taste and chemical properties during the roasting process. Molecules 2019, 24, 1374. [Google Scholar] [CrossRef]
- Lee, S.Y.; Sekhon, S.S.; Kim, H.C.; et al. Anti-inflammatory effect of lactic acid bacteria isolated from kimchi on acid-induced acute colitis in model mice. Toxicol. Environ. Health Sc. 2017, 9, 279–283. [Google Scholar] [CrossRef]
- Lightfoot, Y. L.; Yang, T.; Sahay, B.; Mohamadzadeh, M. Targeting aberrant colon cancer-specific DNA methylation with lipoteichoic acid-deficient Lactobacillus acidophilus. Gut Microbes 2013, 4, 84–88. [Google Scholar] [CrossRef]
- Lin, Q.; Mathieu, O.; Tompkins, T. A.; et al. Modulation of the TNF$\alpha$-induced gene expression profile of intestinal epithelial cells by soy fermented with lactic acid bacteria. J. Funct. Foods 2016, 23, 400–411. [Google Scholar] [CrossRef]
- Ma, Y.; Chen, X.; Xu, R.; et al. Lactiplantibacillus plantarum fermentation enhanced the protective effect of kiwifruit on intestinal injury in rats: Based on mitochondrial morphology and function. Food Chem. X 2023, 20, 101025. [Google Scholar] [CrossRef]
- Minervini, F.; Missaoui, J.; Celano, G.; et al. Use of autochthonous lactobacilli to increase the safety of zgougou. Microorganisms 2020, 8, 29. [Google Scholar] [CrossRef]
- Missaoui, J.; Saidane, D.; Mzoughi, R.; Minervini, F. Fermented Seeds (“Zgougou”) From Aleppo Pine as Novel Source of Potentially Probiotic Lactic Acid Bacteria. Microorganisms 2019, 7, 709. [Google Scholar] [CrossRef]
- Mosiej, W.; Dorota, Z.; Marcin, K.; et al. The Impact of food-derived lactic acid bacteria (LAB) Supplementation on Gut Microbiota Modulation in Elderly Individuals With Cognitive Impairment: Insights from the SHIME® Model. Int. J. Psychophysiol. 2025, 213, 113147. [Google Scholar] [CrossRef]
- Nasri, N.; Triki, S. Analyse des lipides des graines de pins de Tunisie (Pinus halepensis Mill. et Pinus pinea L.). Riv. Ital. Sostanze Grasse 2004, 81, 244–247. [Google Scholar]
- Nithya, A.; Misra, S.; Panigrahi, C.; et al. Probiotic potential of fermented foods and their role in non-communicable diseases management: An understanding through recent clinical evidences. Food Chem. Adv. 2023, 3, 100381. [Google Scholar] [CrossRef]
- Nouri, Z.; Karami, F.; Neyazi, N.; et al. Dual anti-metastatic and anti-proliferative activity assessment of two probiotics on HeLa and HT-29 cell lines. Cell J. (Yakhteh) 2016, 18, 127. [Google Scholar]
- Ohland, C.L.; Mac Naughton, W.K. Probiotic bacteria and intestinal epithelial barrier function. Am. J. Physiol. Ganstrointest. Liver Physiol. 2010, 298, G807–G819. [Google Scholar] [CrossRef]
- Oladejo, B.O.; Akingbade, G.T.; Adeboboye, C.F.; Adeoye, S.B. Lactobacillus species modulates TNF-α and Nrf2 expression in carrageenan-induced rat paw oedema. The Microbe 2024, 2, 100050. [Google Scholar] [CrossRef]
- Oudat, Q.; Okour, A. The Role of Probiotics in Modulating Gut Microbiota and Metabolic Health for Weight Management: A Mini Review. Acta Microbiol. Hell. 2025, 70, 5. [Google Scholar] [CrossRef]
- Paradis, T.; Bègue, H.; Basmaciyan, L.; Dalle, F.; Bon, F. Tight junctions as a key for pathogens invasion in intestinal epithelial cells. Int. J. Mol. Sci. 2021, 22, 2506. [Google Scholar] [CrossRef]
- Parlesak, A.; Haller, D.; Brinz, S.; et al. Modulation of cytokine release by differentiated CACO-2 cells in a compartmentalized coculture model with mononuclear leucocytes and nonpathogenic bacteria. Scand. J. Immunol. 2004, 60, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.; Butani, K.; Kumar, A.; et al. Effects of fermented food consumption on non-communicable diseases. Foods 2023, 12, 687. [Google Scholar] [CrossRef]
- Qin, H.; Zhang, Z.; Hang, X.; Jiang, Y. L. L. plantarum prevents enteroinvasive Escherichia coli-induced tight junction proteins changes in intestinal epithelial cells. BMC Microbiol. 2009, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Saha, P.; Chassaing, B.; Yeoh, B.S.; et al. Ectopic expression of innate immune protein, lipocalin-2, in Lactococcus lactis protects against gut and environmental stressors. Inflamm. Bowel Dis. 2017, 23, 1120–1132. [Google Scholar] [CrossRef] [PubMed]
- Sahu, B.; Johnson, L.M.; Sohrabi, M.; et al. Effects of Probiotics on Colitis-Induced Exacerbation of Alzheimer’s Disease in AppNL-G-F Mice. 2023, 24, 11551.
- Sharma, R.; Young, C.; Neu, J. Molecular modulation of intestinal epithelial barrier: contribution of microbiota. BioMed Res. Int. 2010, 2010, 305879. [Google Scholar] [CrossRef]
- Şengün, İ. Y.; Güney, D. Probiotic potential of fermented foods and their effects on immune system. Turk. J. Agric. Food Sci. Technol. 2021, 9, 1744–1750. [Google Scholar] [CrossRef]
- Schiller, G. Therapeutic Use of Aleppo Pine (Pinus halepensis Mill.). In Medicinal and Aromatic Plants of the Middle-East; Yaniv, Z., Dudai, N., Eds.; Springer Netherlands: Dordrecht, 2014; pp. 215–224. [Google Scholar]
- Suzuki, T. Regulation of the intestinal barrier by nutrients: The role of tight junctions. Anim. Sci. J. 2020, 91, e13357. [Google Scholar] [CrossRef]
- Talà, A.; Guerra, F.; Resta, S. C.; et al. Phenotyping of fecal microbiota of Winnie, a rodent model of spontaneous chronic colitis, reveals specific metabolic, genotoxic, and pro-inflammatory properties. Inflammation 2022, 45, 2477–2497. [Google Scholar] [CrossRef]
- Tiptiri-Kourpeti, A.; Spyridopoulou, K.; Santarmaki, V.; et al. Lactobacillus casei exerts anti-proliferative effects accompanied by apoptotic cell death and up-regulation of TRAIL in colon carcinoma cells. PloS One 2016, 11, e0147960. [Google Scholar] [CrossRef]
- Tiss, M.; Souiy, Z.; Ben Abdeljelil, N.; et al. Fermented soy milk prepared using kefir grains prevents and ameliorates obesity, type 2 diabetes, hyperlipidemia and Liver-Kidney toxicities in HFFD-rats. J. Funct. Foods 2020, 67, 103869. [Google Scholar] [CrossRef]
- Tukan, S. K.; Al-Ismail, K.; Ajo, R. Y.; Al-Dabbas, M. M. Seeds and seed oil compositions of Aleppo pine (Pinus halepensis Mill.) grown in Jordan. Riv. Ital. Delle Sostanze Grasse 2013, 90, 87–93. [Google Scholar]
- Tunisi, L.; Forte, N.; Fernández-Rilo, A. C.; et al. Orexin-A prevents lipopolysaccharide-induced neuroinflammation at the level of the intestinal barrier. Front. Endocrinol. 2019, 10, 219. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Lin, P.R.; Ng, C.C.; Shyu, Y.T. Probiotic properties of Lactobacillus strains isolated from the feces of breast-fed infants and Taiwanese pickled cabbage. Anaerobe 2010, 16, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.-K.; Wu, Y.-Y.; Yang, Y.-F.; et al. The effect of probiotics on serum levels of cytokine and endotoxin in peritoneal dialysis patients: a randomised, double blind, placebo-controlled trial. Benef. Microbes 2015, 6, 423–430. [Google Scholar] [CrossRef]
- Wang, I.K.; Yen, T.H.; Hsieh, P.S.; et al. Effect of a probiotic combination in an experimental mouse model and clinical patients with chronic kidney disease: a pilot study. Front. Nutr. 2021, 8, 661794. [Google Scholar] [CrossRef] [PubMed]
- Weber, K.T.; Alipui, D.O.; Sison, C.P.; et al. Serum levels of the proinflammatory cytokine interleukin-6 vary based on diagnoses in individuals with lumbar intervertebral disc diseases. Arthritis Res. Ther. 2016, 18, 3. [Google Scholar] [CrossRef]
- Wei, C.X.; Wu, J.H.; Huang, Y.H.; et al. Lactobacillus plantarum improves LPS-induced Caco2 cell line intestinal barrier damage via cyclic AMP-PKA signaling. PLoS One 2022, 17, e0267831. [Google Scholar] [CrossRef] [PubMed]
- Wierzbicka, A.; Ma’nkowska Wierzbicka, D.; Mardas, M.; Stelmach Mardas, M. Role of Probiotics in Modulating Human Gut Microbiota Populations and Activities in Patients with Colorectal Cancer—A Systematic Review of Clinical Trials. Nutrients 2021, 13, 1160. [Google Scholar] [CrossRef] [PubMed]
- Woo, J. K.; Choi, S.; Kang, J. H.; et al. Fermented barley and soybean (BS) mixture enhances intestinal barrier function in dextran sulfate sodium (DSS)-induced colitis mouse model. BMC Complement. Altern. Med. 2016, 16, 498. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, X.; Le, M.H.; et al. Reutericyclin producing Lactobacillus reuteri modulates development of fecal microbiota in weanling pigs. Front. Microbiol. 2015, 6, 762. [Google Scholar] [CrossRef]
- Yuki, T.; Haratake, A.; Koishikawa, H.; et al. Tight junction proteins in keratinocytes: localization and contribution to barrier function. Exp. Dermatol. 2007, 16, 324–330. [Google Scholar] [CrossRef]
- Zaroual, H.; Boughattas, F.; Karoui, R. Traditional foods in Maghreb: Production and research progress. In Traditional Foods: History, Preparation, Processing and Safety; 2019; pp. 51–113.
- Zhang, X. F.; Guan, X. X.; Tang, Y. J.; et al. Clinical effects and gut microbiota changes of using probiotics, prebiotics or synbiotics in inflammatory bowel disease: a systematic review and meta-analysis. Eur. J. Nutr. 2021, 60, 2855–2875. [Google Scholar] [CrossRef]
- Zhang, X.; Li, Y.; Zhang, C.; et al. Postbiotics derived from Lactobacillus plantarum 1.0386 ameliorate lipopolysaccharide-induced tight junction injury via MicroRNA-200c-3p mediated activation of the MLCK-MLC pathway in Caco-2 cells. Food Funct. 2022, 13, 11008–11020. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



