Submitted:
02 December 2025
Posted:
03 December 2025
You are already at the latest version
Abstract
The kelp genus Eisenia Areschoug is represented by two species in the Southeast Pacific: Eisenia cokeri, distributed in Peru and Chile, and E. gracilis, endemic to Peru. However, the taxonomic distinction between these two species has been questioned. Additionally, it is uncertain whether E. cokeri is conspecific with the Northeast Pacific species E. arborea. To address these issues, we examine, for the first time, the morphology and the molecular phylogenetics of E. cokeri and E. gracilis across their geographic range. Sampling for morphological and molecular analyses was conducted at subtidal zones in five locations in Peru. Furthermore, a locality in Chile was sampled for molecular studies, where E. cokeri was documented. Peruvian Eisenia species exhibited notable differences in morphological characters, including size, holdfast diameter, stipe modifications (e.g., longitudinal division and pseudostipe formation), frond division, frond margins, and roughness, as evidenced by differences in thallus morphometric variables. Phylogenetic analyses using mitochondrial and chloroplast markers revealed three distinct genetic lineages and two endemic species: E. cokeri and E. gracilis from Peru, and a new Eisenia sp. from Chile. Moreover, we confirmed the distinctiveness of E. cokeri from E. arborea and provided updated information on the distribution of E. cokeri and E. gracilis in the Southeast Pacific. These findings are crucial for the management and conservation of these endemic, foundational species.
Keywords:
endemic
; genetic delimitation
; genetic diversity
; morphology
; taxonomy
1. Introduction
Kelps are structurally complex brown seaweeds, mainly of the order Laminariales [1], widely distributed across more than 25% of coastal areas worldwide [1,2]. These algae can cover extensive benthic substrates and form highly productive and dynamic ecosystems called kelp beds or kelp forests that physically and biologically modify their surrounding environments [3,4]. Indeed, kelps are foundation species that facilitate the occurrence of a great variety of life forms, including mammals, fish, and invertebrates [6,7], and harbor many economically valuable species [5,6]. Despite the fact that these ecosystems provide a multitude of goods and services to society [7], valued at billions of dollars each year [10], basic aspects such as their taxonomy and phylogeny require greater attention.
Advances in molecular systematics have redefined the phylogenetic framework of the Laminariales [4], prompting significant revisions to their taxonomic circumscription. One major change was the reassignment of the genus Eisenia to the family Lessoniaceae, alongside Ecklonia, Eckloniopsis, Egregia, and Lessonia [4,8]. This assemblage is distinguished by the presence of sporophylls arising from the blades, a trait considered phylogenetically informative [9]. Multiple phylogenetic studies have further highlighted the close affinity between Eisenia and Ecklonia [4,9,10,11], two genera that exhibit striking morphological similarities, differing mainly in the mode of meristem division at the distal end of the stipe [12]. Based on mitochondrial and chloroplast markers, Rothman et al. proposed that Eisenia arborea Areschoug—the type species of the genus—should be regarded as conspecific with Ecklonia, raising the possibility that other Eisenia species might also require transfer [12]. However, subsequent multilocus analyses by Kawai et al. resolved Ecklonia and Eisenia as distinct, well-supported clades, thereby reaffirming Eisenia as a valid genus and retaining E. arborea as its generitype [13].
Despite significant progress in Eisenia phylogenetics, the taxonomic status of multiple species remains in need of formal revision. Previous molecular phylogenetic studies have primarily targeted conspicuous Northern Hemisphere representatives of the genus, including E. arborea and E. bicyclis [4,9,10,11,12], as well as E. nipponica [13]. In contrast, Southeast Pacific endemics such as E. desmarestioides, E. galapagensis, E. cokeri, and E. gracilis have remained unexamined at the molecular level since their original taxonomic descriptions.
Currently, Eisenia comprises seven species that are geographically restricted to the warm−temperate waters of the Pacific Ocean: Eisenia arborea, Eisenia bicyclis (Kjellman) Setchell, Eisenia desmarestioides Setchell & N. L. Gardner, Eisenia galapagensis W. R. Taylor, Eisenia nipponica H. Kawai, S. Akita, K. Hashimoto & T. Hanyuda, Eisenia cokeri M. Howe, and Eisenia gracilis E. Y. Dawson, Acleto & Foldvik [13,14,15,16,17,18]. Additionally, Buglass et al. reported a potential new species of Eisenia at depths exceeding 40 m near the Galapagos Islands, potentially increasing the number of recognized species within this genus to eight [19].
Subcanopy kelp Eisenia is found along the Peruvian coast, where it contributes to structuring subtidal beds and supports a diverse benthic community (see Carbajal et al. (2021) and Uribe et al. (2024) for further details) [6,20]. Eisenia cokeri has been recorded in several localities in Peru, between Máncora (5°S) and Paracas (15°S) [14,16,21], inhabiting hard substrates at depths of 0 and 12 m [6,22]. Additionally, it has been found in Chile, in the Desventuradas Islands and San Felix and San Ambrosio Islands (26°S) [15,17] from the intertidal zone to depths of up to 34 m [17]. In contrast, E. gracilis has been recorded in a single locality in Peru, Bahía San Juan (15° S), where it was dredged from sandy and shelly bottoms at 27 and 36 meters during the Allan Hancock Expedition in 1938 [16]. Based on the predictions of the potential occurrence of kelps in the deep waters of the eastern Pacific [23], E. gracilis may also be present in other locations along the central and southern Peruvian coast. However, this hypothesis has not yet been explored. Thus, Eisenia from Peru includes a low latitude shallow-water species (E. cokeri) and a mid-latitude deep species, potentially mesophotic (>30 m) (E. gracilis), influenced by the oceanographic conditions of the Humboldt Current Ecosystem. This system is characterized by permanent coastal upwelling that brings cold, low-oxygen, and nutrient-rich water to the surface, generating high coastal productivity [24,25]. Furthermore, the interannual phenomenon El Niño Southern Oscillation (ENSO) modifies these climatic conditions by reducing or canceling upwelling, and consequently increasing coastal temperatures [24,25].
Despite morphological differences exhibited by E. cokeri and E. gracilis [16], some doubts have emerged regarding the taxonomic identity of these species and other Eisenia species. In the past, the species status of E. cokeri and E. gracilis was questioned, with the suggestion that both could be conspecific [16,26]. Additionally, Howe highlighted the morphological resemblance of E. cokeri to E. arborea from North America [14] and suggested the conspecificity of these two taxa [16,17]. Also, the morphological differences between E. cokeri from Peru and those from the Desventuradas Islands [27], together with the large geographical distance separating these populations, raise doubts about the taxonomic unity of E. cokeri.
In this study, we conducted integrated molecular phylogenetic and morphological analyses of Eisenia species from the Peruvian coast to test the taxonomic distinctiveness of E. cokeri and E. gracilis. Furthermore, using multilocus molecular data, we evaluated the genetic cohesion of E. cokeri populations from Peru and Chile and assessed their divergence from the southern sea palm kelp, E. arborea.
2. Materials and Methods
2.1. Sampling Sites and Collection
Sampling was conducted between March and July 2018 across five localities spanning ~1,600 km of the Peruvian coastline. The sites were selected based on historical records [16,21] of E. cokeri and E. gracilis. This was complemented with one additional locality where the occurrence of E. gracilis was previously reported (Figure 1).
For E. cokeri, samples were collected from depths of 1 to 9 m at Lobos de Tierra Island, Casma, and Paracas, which together encompass 88% of the species geographic distribution in Peru (Figure 2A). For E. gracilis, we collected samples from Marcona, at a 22 m depth, and from Ilo (Figure 3A), at depths ranging from 10 to 15 m (Figure 1, Table 1).
In each locality, we randomly collected plants and thallus fragments via semiautonomous diving. A minimum of 30 adult Eisenia sporophytes were sampled for morphometric measurements, and 30 epibiont-free fronds, from plants separated by at least 1 meter, were obtained and immediately preserved in silica gel for phylogenetic analysis. Also, a total of 14 thallus samples of E. cokeri were collected from the San Ambrosio Islands, Chile (hereafter referred to as Eisenia from the Desventuradas Islands or Chilean Eisenia) for molecular analysis.
Additionally, qualitative observations of the Eisenia habitat (bathymetric range and bottom type) were made. The depth range was estimated from perpendicular dives from the intertidal zone to the depth at which the Eisenia record was null for at least three contiguous meters, or up to a maximum depth of 30 m, due to diving safety restrictions. Sampling at the Desventuradas Islands was not possible due to this site’s inaccessibility.
Sporophyte habit descriptions were carried out using fresh and preserved specimens (i.e., herbarium exsiccate) deposited in the Instituto del Mar del Perú Herbarium, collected between 2008 and 2019 across Peruvian coastal localities. Morphological features were complemented by field observations and photographs of E. cokeri from Lobos de Tierra Island (6°S) and Rancherío (14°S), between 2018 and 2019. Additionally, unpublished data from maximum holdfast diameter, stipe length, total length, and total fresh weight of E. cokeri from Casma (2009 and 2015) and total length of E. gracilis from Marcona (2015) were included (Supplementary Table S1).
We compared our information with the original descriptions of E. cokeri [14] and E. gracilis [16]. Due to logistical constraints associated with the remoteness of the Desventuradas Islands, no morphological observations of Eisenia could be made; therefore, descriptions were based on previous studies.
We randomly collected a minimum of 30 adult Eisenia sporophytes per locality to measure in situ the following morphometric variables: maximum holdfast diameter (MHD), holdfast height (HH), stipe length (SL), stipe diameter (SDI), and total length (TL). In addition, biomass was quantified as the total fresh weight (TFW). Descriptive statistics (mean, standard deviation, minimum, and maximum) were computed for each species. Differences in morphometric variables and biomass between E. cokeri and E. gracilis were evaluated using the pairwise Mann–Whitney U test. This non-parametric test was used due to non-normal and heteroscedastic distributions of most variables, as evidenced by the Shapiro–Wilk and Levene tests, respectively. The threshold for statistical significance was set at a p-value of 0.05. To graphically explore morphometric differences between the two Eisenia species, Principal Component Analysis (PCA) was applied after square root transformation and data normalization. A multivariate non-parametric permutational ANOVA (PERMANOVA) [31] was performed to assess significant differences in morphometric variables between the two kelps. For this purpose, similarity matrices based on Euclidean distances of the untransformed data were used, with 9999 random permutations generated using the unrestricted permutation method applied to the raw data. Statistical analyses were performed using R statistical software (version 4.4.1, Core Team 2024) and the PRIMER v7 program [28].
2.2. Phylogenetic Analyses
DNA extraction was performed from the pulverized algae using a modified protocol of the Thermo Scientific GeneJET Genomic DNA Purification Kit (Thermo Fisher Scientific). Five genetic markers from two cellular compartments were evaluated: (i) three mitochondrial markers: atp8-trnS or ATP8S (intergenic spacer between the atp8 and trnS genes), trnW-trnI or WI (intergenic spacer between the trnW and trnI genes), and COI (cytochrome oxidase C subunit I gene), and (ii) two chloroplast markers: rbcL (RuBisCo or ribulose-1,5-bisphosphate carboxylase/oxygenase large subunit) and the rbcL spacer or RS (spacer between the rbcL and rbcS genes) (Supplementary Table S2). For the ATP8S and WI markers, we used the primers described by Engel et al. [33] and the PCR mixture and program used by Voisin et al. [29]. For COI, we used the GazF1 and GazR2 primers designed by Lane et al. [35], and the PCR mixture and program described by Macaya & Zuccarello [30]. For rbcL, we used the KL2 and KL8 primers [4] and the PCR recipe and program described by Rothman et al. [12]. The RS1 and RS2 primers [10] were used for amplification of RS (Supplementary Table S2). PCR products were verified by 1% agarose gel electrophoresis, purified, and sequenced using an ABI 3730xl automatic DNA sequencer from Macrogen Inc. (Korea). For each marker, the obtained sequences were edited using Chromas v.2.5.1 [31] and multiple sequence alignments were performed with the CLUSTALW function of BioEdit 7.2.5 [32].
For each marker, the sequence alignment was performed using Eisenia, Ecklonia, and Lessonia sequences available in GenBank. To infer phylogenetic relationships between Eisenia and Ecklonia species, the trees were rooted with Laminariales species (Supplementary Table S3). The construction of the trees was based on the options and parameters described by Tellier et al. [33]. The JModelTest2 program [34] was employed to determine the optimal model of sequence evolution for each marker and the consensus (all markers concatenated), using the Akaike Information Criterion (AIC). The GTR+I+G model was identified as the best fit for both markers. Phylogenetic relationships were inferred using Maximum Likelihood (ML) in the IQ-TREE program [35] with 1,000 bootstrap replications, and Bayesian Inference (BI) implemented in MrBayes 3.2.1 [36] with 10 million generations. Trees were sampled every 1,000 generations during the run, and the first 30% were discarded as burn-in. The resulting phylogenetic trees were visualized and edited using Figtree v1.4.3 (http://tree.bio.ed.ac.uk/software/figtree/). Sequences of the Laminariales order were used for the outgroup (Macrocystis pyrifera) selection, as performed by Rothman et al. [12] and Kawai et al. [13].
For each marker, molecular diversity indices were estimated at the species and locality level: the number of haplotypes, private haplotypes, and polymorphic sites, the haplotype diversity (H), and the nucleotide diversity (π) in the program DnaSP 5.10.01 [37]. We calculated differences between groups as mean pairwise p-distances for intraspecific and interspecific comparisons using MEGA 7.0.14, excluding positions with indel polymorphisms. Haplotype networks were constructed for each marker using the Median Joining algorithm of NETWORK v.5.0 [38]. The reticulations and loops were solved according to Crandall & Templeton [39].
2.3. DNA-Based Species Delimitation Method with GMYC
Prior to delimitation analyses, sequences were collapsed into unique haplotypes. For the GMYC analysis, an ultrametric tree was constructed in BEAST v.1.8 [40] for each marker (Supplementary Table S3) using the appropriate model selected by JModelTest [40] and assuming an uncorrelated lognormal relaxed molecular clock under the constant-size coalescent model. Fifty million generations were implanted for independent runs, sampling every 1000 trees. Runs were inspected for convergence using Tracer v.1.5, and trees were summarized from the MCMC analyses after discarding the first 25% of trees generated. GMYC analyses were run using a single threshold following Fujisawa & Barraclough [41] using the SPLITS package in R [42].
3. Results
3.1. Morphological Observations
3.1.1. Eisenia cokeri
Additional specimens examined: Lambayeque, Lobos de Tierra Island, subtidal, coll. A. Indacochea, IMARPE 05-000263, 06° 24’ 25.8'' S, 80° 51′ 50.5″ W, 08.x.2008. Ancash, Huarmey, Islote Patillos, Culebras, subtidal, coll. A. Gamarra, IMARPE 05-000343, 09° 53′ 44.0″ S, 78° 14′ 14.2″ W, 03. x.2015. Ica, Paracas, Rancherío, drift, coll. J. Zavala, IMARPE 05-000279, 16.vii.2008.
Description: Sporophyte thalli up to 1.8 m high and up to 1.2 kg in weight. Holdfast of densely intertwined haptera (Figure 2A−B, D), with a diameter of up to 34 cm and a height of 16 cm. The sporophyte exhibited a solid stipe, measuring 1.6 cm in diameter and reaching lengths of up to 50 cm, terete at the base and transitioning to a flattened and compressed form in the upper portion (Figure 2C). Juvenile fronds are lanceolate–oblong, initially smooth and then longitudinally roughened (rugose) (Figure 2E), reaching widths of up to 11 cm and exhibiting falcate lanceolate lateral bladelets with whole margins (Figure 2F−H). The main blade, thickened and elongated, included lanceolate, entire-edged bladelets that transformed into secondary blades, which reduced in size toward the apex (Figure 2I−J). The main blade began to erode at the apex while its base thickened to form entire lanceolate blades (Figure 2K−L), with serrated margins and spines of different sizes (Figure 2M). The mature main blade splits into two false stipes or compressed, flattened branches, which twist, culminating in a broad, compressed base that supports several simple, long blades reaching up to 1.6 m in length, which are occasionally divided. Mature blades are thick serrated, and exhibit longitudinal rugosity (Figure 2N). Sporangia are clustered in sori that form confluent, subcontinuous bands.
Remarks: Previous measurements of E. cokeri conducted in 2009 and 2015 in Casma attained maximum values exceeding those reported here for total thallus height (2.7 m, measured specimens = 103), total fresh weight (2.5 kg, measured specimens = 37), maximum holdfast diameter (28 cm, measured specimens = 51), and stipe length (92 cm, n=16).
Habitat:Eisenia cokeri beds (Figure 2A) were observed in Peru in the lower intertidal zone to depths of approximately 10 m, in locations protected from wave exposure (Lobos de Tierra Island and Paracas) as well as in places with moderate level of wave exposure (Casma). Sporophytes were mainly observed on platforms and irregular rocky blocks, pebbles, and small rocks, surrounded by coarse sand or sandy-shingle bottoms. In all assessed locations, E. cokeri formed monospecific kelp beds.
3.1.2. Eisenia gracilis
Additional specimens examined: Ica, Carrizales, subtidal, coll. A. Gamarra, IMARPE 05-001683, 15° 26′ 15.7″ S, 75° 04’ 46.0″ W, 29.xi.2015. Ica, San Juan de Marcona, subtidal, coll. J. Zavala, IMARPE 05-000279, 16.vii.2008.
Description: Sporophytes reach a height of 1.1 m and a weight of 0.09 kg. Holdfasts exhibiting sparsely branched haptera and rhizomatous development from which new stipes (stolon-like) emerged (Figure 3A−C). The stipe is subcylindrical at the base, measuring up to 0.8 cm in diameter, then becomes flattened, attaining a length of up to 52 cm. Juvenile fronds are initially lanceolate and smooth (Figure 3D), thereafter developing marginal bladelets that are spread apart, some falcate, placed alternately, with smooth margins (Figure 3E−F). Some mature individuals have primary blades and secondary blades with strongly serrated margins (Figure 3G). Most individuals exhibit a compressed primary blade base, generating 3−6 additional linear, lanceolate, serrate blades of similar width (Figure 3H). Sporangia aggregated in sori, forming confluent, subcontinuous bands (Figure 3I). Fronds featuring thick, serrated mature blades concentrated at the distal region of the thallus, exhibiting an elongated stipe reaching up to 52 cm (Figure 3J).
Remarks: Measurements of E. gracilis obtained in Marcona in 2011 and 2015 exhibited greater total thallus height (1.5 m, measured specimens = 38) and total fresh weight (0.5 kg, measured specimens = 21) than those recorded in this study.
Habitat: Eisenia gracilis (Figure 3A) was located at depths ranging from 10 to 30 m (Marcona: 20−30 m, Ilo: 10−15 m), in regions with moderate to high wave exposure, on both rocky surfaces and biogenic substrates. In Marcona, this species was observed to grow on mytilid Aulacomya ater communities, while in Ilo, it was found on small, scattered blocks, pebbles, and rocks located between sandy shell substrates and boulders. At both sites, E. gracilis was observed producing monospecific or mixed aggregations, cohabiting with Lessonia trabeculata. In the latter case, E. gracilis grew on the flanks of the rocky blocks already colonized by L. trabeculata (Ilo). The discovery of E. gracilis in Ilo extends the known distribution area of this species.
3.1.2. Eisenia from Desventuradas Islands
Remarks: Thalli attained a maximum length of 1 m [17], characterized by a holdfast including intertwined haptera and a robust, compressed, and flattened stipe. The blades were oblong at the base and linear–lanceolate, measuring 1.3 to 2.1 cm wide and 20 to 30 cm long. All blades showed pronounced longitudinal grooves and submarginal toothed margins [15,27]. The blades of Eisenia from Desventuradas exhibit an intermediate morphology between the Peruvian E. cokeri and E. gracilis, characterized by longitudinal grooves and a narrow linear–lanceolate shape, respectively [27].
3.2. Morphometric Analysis
Descriptive statistics of the morphometric variables measured in situ in 101 specimens of E. cokeri and in 67 of E. gracilis are presented in Table 2. Morphometric descriptors, including maximum holdfast diameter, holdfast height, stipe diameter, total length, and total fresh weight, were significantly higher in E. cokeri than in E. gracilis (Mann–Whitney, p<0.05; see Supplementary Table S4). Four variables (maximum holdfast diameter, holdfast height, stipe diameter, and total fresh weight) of E. cokeri exceeded twice the metrics of E. gracilis, yet the most notable difference among both species was found in their biomass (Table 2). The morphological differentiation between the two Eisenia species was further evidenced by the PCA plot (Figure 4), which showed that each species formed a well-defined group. The first and second principal components adequately captured the variation in thallus morphology, explaining 86% (PC1: 69%, PC2: 17%) of the variance across all observations. E. gracilis plants were associated with lower values of morphometric variables and also showed less morphological variation, forming a more cohesive cluster of points compared to the more dispersed observations of E. cokeri (Figure 4). The morphological distinctiveness of both Eisenia was significantly supported by PERMANOVA analysis (Pseudo-F = 169.04, P < 0.001).
3.3. Delimitation of Species by GMYC Analyses
GMYC analyses (Table 3) across the five genetic markers (ATP8S, WI, COI, RS, and rbcL) consistently supported the presence of three Eisenia species in the Southeastern Pacific (SEP), including the Peruvian coast. Although the number of putative GMYC clusters varied among markers—ranging from 9 to 14 for COI, RS, and rbcL, and 10 for ATP8S—this variation mainly reflects intraspecific structure rather than additional species. WI showed no significant support for the GMYC model, indicating lower resolution for lineage discrimination. Overall, despite differences in clustering depth, all markers converge on the same biological conclusion: three well-delimited Eisenia species occur in Peru and the broader SEP region.
3.4. Phylogenetic Analysis
A total of 277 sequences were generated (Table 4), comprising five molecular markers. Sequences for ATP8S and WI were trimmed to 162 and 174 bp, respectively. For COI, the full alignment was 602 bp, although some sequences were shorter and trimmed to 564 bp. Similarly, RS alignments were 669 bp in length, while the reduced dataset was 427 bp; rbcL alignments were 932 bp, with a reduced dataset of 522 bp.
The concatenated molecular dataset yielded a well-supported phylogeny under both Bayesian inference (BI) and maximum likelihood (ML) (Figure 5). The genera Ecklonia and Eisenia were recovered as strongly supported clades; within Eisenia, E. arborea was clearly segregated from the remaining taxa. The Peruvian and Chilean Eisenia exhibited low overall genetic diversity. Across all markers, the number of haplotypes ranged from three to five, with haplotype diversity (H) values spanning 0.629–0.732 and nucleotide diversity (π) ranging from 0.003 to 0.026 (Table 5). No haplotypes were shared among E. cokeri and E. gracilis from Peru, nor with the Eisenia population from the Desventuradas Islands. In contrast, E. gracilis exhibited no haplotype variation across the two sampled localities, regardless of the marker analyzed. A similar pattern was observed in E. cokeri from Peru, with a single haplotype across three localities, except for the WI marker, where all individuals from the northernmost population at Lobos de Tierra Island possessed a private haplotype distinct from that observed in the other sites. Finally, Eisenia from the Desventuradas Islands harbored two haplotypes for WI and ATP8S, but only a single haplotype across the remaining markers.
Inferences regarding the phylogenetic relationships among E. cokeri, E. gracilis, and Eisenia from the Desventuradas Islands are constrained by the overall low genetic diversity. In the concatenated phylogeny (Figure 5), haplotypes of E. gracilis and the Desventuradas Eisenia formed a closer association than those of E. cokeri. However, analyses based on individual markers yielded incongruent topologies, complicating the interpretation of interspecific relationships (Figure S1–S5).
The reconstructed haplotype networks (Figure 6) revealed no haplotype sharing among E. cokeri, E. gracilis, and Eisenia from the Desventuradas Islands, supporting their genetic distinctiveness. Nevertheless, the inferred evolutionary affinities varied among markers. In the rbcL network, Desventuradas haplotypes were positioned equidistantly between E. cokeri and E. gracilis. In contrast, they clustered more closely with E. gracilis in the ATP8S, COI, and WI networks, whereas in the RS network, they were more closely associated with E. cokeri.
The interspecific divergences among our sequences of E. cokeri, E. gracilis, and Eisenia from the Desventuradas Islands consistently exceeded the minimum genetic distance threshold established for Laminariales (Concatenated > 0.0024; ATP8S > 0.0030; COI > 0.0036; WI > 0.0154; RS > 0.0035; rbcL > 0.0025). The highest levels of divergence were observed between E. cokeri and E. gracilis (Concatenated = 0.0260; ATP8S = 0.1148; COI = 0.0336; WI = 0.0697; RS = 0.0157; rbcL = 0.0012) (Table S5). In contrast, both species exhibited lower pairwise distances than Eisenia from the Desventuradas Islands.
4. Discussion
The integration of morphological and molecular approaches to address, for the first time, the taxonomic uncertainties of Eisenia species in the southeastern Pacific allowed us to confirm the distinction between two Eisenia species from Peru, E. cokeri and E. gracilis, described by Howe [14] and Dawson et al. [16], respectively. Furthermore, our molecular findings resolved the ambiguity between E. cokeri and the Northeast Pacific species E. arborea and suggest that Eisenia from the Desventuradas Islands, Chile, represents a new species. This study establishes three distinct endemic species in the region: two in Peruvian mainland waters and one in oceanic Chilean islands. These results lay the groundwork for the management and conservation of these foundational species.
4.1. Morphological Phylogenetic Position of Eisenia within Laminariales
Previous molecular studies consistently reported a close phylogenetic relationship between the laminarian genera Eisenia and Ecklonia [4,9,10,11] (Saunders & Druehl 1993, Yoon & Boo 1999; Yoon et al. 2001, Lane et al. 2006), to the point of suggesting that both might constitute a single genus. This hypothesis was reinforced by the limited morphological differentiation between them, with the main diagnostic trait being the pattern of meristem division at the distal end of the stipe [12]. More recently, Kawai et al. [13] rejected the proposed synonymy and reinstated Eisenia as a genus distinct from Ecklonia, although they did not include representatives from the southeastern Pacific.
Our concatenated phylogeny resolves two strongly supported clades corresponding to Ecklonia and Eisenia, confirming the placement of southeastern Pacific taxa within Eisenia and aligning with previous molecular studies (e.g., Kawai et al. [13], Buglass et al. [19]). Nevertheless, further sequencing efforts, especially phylogenomic datasets and the inclusion of additional Pacific species such as E. galapagensis, E. desmarestioides, and Eisenia sp., would refine our understanding of intergeneric boundaries and clarify species-level relationships within Eisenia.
4.2. Resolving the E. cokeri–E. arborea Ambiguity
Our results also clarify the long-standing taxonomic uncertainty between E. cokeri from Peru and E. arborea from North America. Historically, their high morphological similarity led several authors to question whether they represented distinct species. Howe [14], in the original description of E. cokeri, and Dawson et al. [16] both suggested a possible conspecificity, noting that the only diagnostic difference was the presence of muciferous canals in E. arborea. Later, specimens from the Desventuradas Islands (Chile), initially identified as E. cokeri, were also hypothesized to correspond to E. arborea [17]. Our phylogenetic analyses clearly separate E. arborea from both E. cokeri (Peru) and the Desventuradas lineage, providing the first molecular evidence to reject their synonymy and confirming that they represent independent evolutionary entities.
4.3. How Many Species Are in the Southeast Pacific? Evidence from Molecular Delimitation
Eisenia gracilis and E. cokeri were traditionally differentiated solely through morphology [16,17]. However, delimiting species based on genetic data can be challenging, particularly in recently diverged lineages or in cases involving incomplete lineage sorting, hybridization, or introgression [44]. In such situations, congruence among multiple lines of evidence—monophyly in phylogenetic reconstructions, species delimitation models (e.g., GMYC), and multilocus datasets—is generally required to support species boundaries [45,46]. Despite its usefulness, GMYC may be sensitive to incomplete sampling, variation in effective population size, or the presence of rare taxa [47].
In our analysis, three independent genetic entities (E. cokeri, E. gracilis, and Eisenia from the Desventuradas Islands) were consistently recovered across molecular delimitation methods, with two of them (E. cokeri and E. gracilis) also showing distinct morphological traits. Although the genetic divergence between E. cokeri and E. gracilis is relatively low for COI (0.36%) and rbcL (0.73%), these values still exceed the intraspecific divergence observed within each species and fall within thresholds reported for species-level differentiation in other kelps. For comparison, COI divergence between Eisenia species ranges from 2.37 to 3.12% [48], and rbcL divergence between Ecklonia radiata and Ecklonia maxima ranges from 3 to 7% [15].
The resolving power of molecular markers varies among studies and low divergence may obscure species boundaries. Rothman et al. found that E. bicyclis and E. arborea did not form monophyletic clades when rbcL or ITS was used, likely because these markers are not highly informative phylogenetically [12]. Conversely, more informative loci (e.g., atp8–trnS and WI) resolve Eisenia relationships more clearly in concatenated datasets. Likewise, Kawai et al. demonstrated that single-marker analyses could not distinguish E. nipponica, whereas multilocus datasets strongly supported its species-level status [13]. Therefore, additional markers with higher polymorphisms, such as SNPs or microsatellites, would further increase resolution and robustness in delimiting Eisenia species [49,50,51]. Across phylogenetic trees, haplotype networks, and pairwise genetic distances, E. cokeri, E. gracilis, and Eisenia from Desventuradas consistently formed separate and well-supported evolutionary lineages, with no haplotype sharing across markers. Pairwise divergences exceeded the minimum threshold for interspecific differentiation in Laminariales, and lineage-specific mutations (e.g., indels) indicate restricted or absent gene flow, particularly in the Desventuradas lineage. Although this lineage clearly represents an independent evolutionary unit, confirming its species status will require additional data—ideally, genome-wide SNPs—to test this delimitation under an integrative framework.
The Desventuradas Islands are located ~1300 km from the southernmost Peruvian population and are separated from continental Chile by an oceanographic barrier driven by contrasting temperature and salinity gradients [52]. Their biota are known for high endemism and Indo-Pacific affinities [43,53]. Taken together, the molecular, morphological, and biogeographical evidence indicates the presence of a previously unrecognized species of Eisenia endemic to the Desventuradas Islands and supports restricting E. cokeri as an endemic species of Peruvian coastal waters.
4.4. Morphological and Morphometric Distinctiveness
The morphological characterization of E. cokeri and E. gracilis presented here refines and expands upon the original taxonomic descriptions [14,16] by providing quantitative measurements of external structures, including total thallus length, holdfast dimensions, and blade size. Adult sporophytes of the two species exhibit marked differences in overall morphology (Figure 2 and Figure 3). E. cokeri develops a robust thallus with a well-formed holdfast, stipe, and blades, whereas E. gracilis forms smaller sporophytes characterized by a rhizomatous holdfast, cord-like stipe, and narrow, single-lamina blades. Additional diagnostic traits—such as stipe differentiation (longitudinal divisions, pseudostipe formation), degree of frond branching, margin serration, and surface roughness—further differentiate the two species.
Although seaweed systematics increasingly relies on molecular phylogenetics, the quantitative assessment of morphological traits remains a critical component for supporting phylogenetic hypotheses and enabling reliable field identification [54,55]. Our morphometric comparison strongly supports the distinction between the two Peruvian species: E. cokeri is significantly larger and exhibits a more complex architecture, while E. gracilis is consistently smaller, slender, and lightweight. The PCA revealed distinct morphometric clusters for each species and highlighted greater intraspecific variation in E. cokeri. Individuals from Lobos de Tierra Island were generally smaller—with shorter thalli, shorter stipes, and smaller conical holdfasts—than those from Casma and Paracas, yet still exhibited higher biomass than the Paracas plants. Conversely, individuals from Casma displayed the largest measurements across all traits. This morphological variability within E. cokeri is consistent with its occurrence in shallow, highly dynamic environments, where temperature fluctuations, wave exposure, and substrate instability can promote phenotypic plasticity. In contrast, the more uniform morphology of E. gracilis likely reflects the comparatively stable conditions of deeper subtidal habitats.
4.5. Spatial and Ecological Segregation
Our sampling of E. cokeri covered nearly its entire known latitudinal range (6°–26° S), including northern and central Peru (Lobos de Tierra, Casma, Paracas) and the Desventuradas Islands (Chile). Its distribution along the Peruvian coast appears stable since its original description [14], although it is no longer present in historical locations such as Callao and Pucusana (12° S). For E. gracilis, we confirm its presence in Marcona—the type locality and the only site previously reported [16]—and extend its distribution southward to Ilo (17° S). Together, these results reveal a disjunct latitudinal pattern: E. cokeri predominates in northern and central Peru (up to ~14° S), whereas E. gracilis inhabits central and southern Peru, with its northern limit around 15° S.
Bathymetric distribution further distinguishes the species. Eisenia cokeri occurs in the lower intertidal and shallow subtidal (0–10 m), while E. gracilis occupies deeper subtidal habitats (10–30 m). Eisenia from the Desventuradas Islands forms beds from 0 to 34 m [17]. These depth patterns align with the ecological breadth reported for the genus, which includes species restricted to shallow waters (<12 m) and others extending to mesophotic depths (up to 60 m). Depth partitioning may reflect the interplay of light availability and interspecific competition [56,57], with deeper habitats (>10 m) likely reducing substrate competition with kelps such as Macrocystis pyrifera and Lessonia trabeculata. Given the scarcity of depth records for E. gracilis, its occurrence below 30 m suggests the potential presence of deep-water kelp forests, a habitat type proposed for this region by Graham et al. [23].
Eisenia from the Desventuradas Islands, previously misidentified as E. cokeri, shows consistent morphological differences—including holdfast shape and a wider, compressed stipe—and narrower, lanceolate blades [27]. Its occurrence in highly exposed, high-energy environments suggests that stipe thickening may be an adaptation to hydrodynamic stress, as reported for other kelps.
5. Conclusion
This study resolves long-standing taxonomic uncertainties within Eisenia in the southeastern Pacific. By integrating multilocus phylogenetic analyses, molecular species delimitation, detailed morphology, and ecological (spatial and depth) segregation, we demonstrate that E. cokeri and E. gracilis represent two distinct species in Peruvian coastal waters. At the same time, the lineage from the Desventuradas Islands in Chile constitutes a third, previously unrecognized evolutionary unit. Phylogenetic analyses separate this oceanic lineage and E. cokeri from the North American species E. arborea, rejecting historical assumptions of conspecificity. Morphological and morphometric analyses corroborate these findings by revealing clear diagnostic differences between species, and the contrasting depth and latitudinal distributions reflect ecological partitioning across environments, from shallow, hydrodynamically variable habitats (E. cokeri) to deeper mesophotic zones (E. gracilis), with the Desventuradas lineage adapted to exposed oceanic conditions.
Together, genetic, morphological, and ecological evidence converge to consistently support the existence of three independent species of Eisenia in the southeastern Pacific—two endemic to Peru and one endemic to the Desventuradas Islands—highlighting previously unrecognized regional biodiversity and providing a foundation for future conservation efforts.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Phylogenetic tree constructed from RS marker. Support values were calculated using ML (top) and BI (bottom) methods. Displayed are only those values exceeding the thresholds of 70 for ML and 0.80 BI, and the asterisk (*) denotes values falling below these specified minimum thresholds; Figure S2: Phylogenetic tree constructed from WI marker. Support values were calculated using ML (bootstrap values on the left) and BI (posterior probabilities on the right) methods. Displayed are only those values exceeding the thresholds of 70 for ML and 0.80 BI and the asterisk (*) denotes values falling below these specified minimum threshold; Figure S3: Phylogenetic tree constructed from COI marker. Support values were calculated using ML (bootstrap values on the left) and BI (posterior probabilities on the right) methods. Displayed are only those values exceeding the thresholds of 70% for ML and 0.80 PP and the asterisk (*) denotes values falling below these specified minimum thresholds; Figure S4: Phylogenetic tree constructed from rbcL marker. Support values were calculated using ML (bootstrap values on the left) and BI (posterior probabilities on the right) methods. Displayed are only those values exceeding the thresholds of 70% for ML and 0.80 PP for BI; Figure S5: Phylogenetic tree constructed from ATP8S marker. Support values were calculated using ML (bootstrap values on the left) and BI (posterior probabilities on the right) methods. Displayed are only those values exceeding the thresholds of 70 for ML and 0.80 BI and the asterisk (*) denotes values falling below these specified minimum thresholds; Table: S1. Morphometric data for Eisenia cokeri and Eisenia gracilis obtained from previous research conducted in 2009 and 2015 (P. Carbajal, unpublished data); Table S2: Molecular markers, primers, and PCR protocols used for PCR reactions and DNA sequencing. The genomic compartment of origin, the abbreviations used in this study, and the full name of each marker are provided, along with the name, sequence, and bibliographic reference of each partition. In addition, references to the protocols specifying reagent concentrations and PCR cycling conditions are included; Table S3. Specimens of GenBank included in the molecular analysis with their accession numbers. ‘*’ indicates the sequences that were used for the concatenated tree, ‘-’ indicates sequence not available in GenBank. Each species of Ecklonia and Eisenia was used for GMYC analyses; Table S4. Mann–Whitney tests of morphometric variables of Eisenia cokeri and E. gracilis measured in five locations along the Peruvian coast; Table S5. Genetic distances for species of Eisenia, Ecklonia, and the Macrocystis pyrifera. The average number of nucleotide base differences per site between species sequences is shown. ‘n/c’ means not calculated.
Author Contributions
Conceptualization and resources, P.C., N.A. and F.T; methodology, investigation, and writing—original draft preparation, P.C., N.A., D.M.C and F.T.; formal analysis, P.C., D.M.C and F.T.; writing—review and editing, P.C., N.A., D.M.C and F.T. All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank the Convenio de Subvención N° 129-2015-FONDECYT, CONCYTEC-Peru for logistical support in sample collection on Lobos de Tierra Island, and for providing specimens used in this study. We are very grateful to Pablo Manríquez Ángulo, Felipe Paredes Vergara, Aldo Racabarren López and Lucas Mekis for the sample collection in the Desventuradas Islands, and to Iván A. Hinojosa for his support. We thank Sara Clemente for her generous assistance with sampling and taking morphometric data in the field. The valuable comments from three anonymous referee and the editor were very helpful in improving the original manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Krumhansl, K.A.; Okamoto, D.K.; Rassweiler, A.; Novak, M.; Bolton, J.J.; Cavanaugh, K.C.; Connell, S.D.; Johnson, C.R.; Konar, B.; Ling, S.D.; et al. Global Patterns of Kelp Forest Change over the Past Half-Century. Proceedings of the National Academy of Sciences 2016, 113, 13785–13790. [Google Scholar] [CrossRef]
- Wernberg, T.; Krumhansl, K.; Filbee-Dexter, K.; Pedersen, M.F. Chapter 3 - Status and Trends for the World’s Kelp Forests. In World Seas: an Environmental Evaluation (Second Edition); Sheppard, C., Ed.; Academic Press, 2019; pp. 57–78. ISBN 978-0-12-805052-1. [Google Scholar]
- Dayton, P.K. The Structure and Regulation of Some South American Kelp Communities. Ecological Monographs 1985, 55, 447–468. [Google Scholar] [CrossRef]
- Lane, C.E.; Mayes, C.; Druehl, L.D.; Saunders, G.W. A Multi-Gene Molecular Investigation of the Kelp (Laminariales, Phaeophyceae) Supports Substantial Taxonomic Re-Organization. Journal of Phycology 2006, 42, 493–512. [Google Scholar] [CrossRef]
- Almanza, V.; Buschmann, A.H. The Ecological Importance of Macrocystis Pyrifera (Phaeophyta) Forests towards a Sustainable Management and Exploitation of Chilean Coastal Benthic Co-Management Areas. International Journal of Environment and Sustainable Development 2013, 12, 341–360. [Google Scholar] [CrossRef]
- Carbajal, P.; Gamarra Salazar, A.; Moore, P.J.; Pérez-Matus, A. Different Kelp Species Support Unique Macroinvertebrate Assemblages, Suggesting the Potential Community-Wide Impacts of Kelp Harvesting along the Humboldt Current System. Aquatic Conservation: Marine and Freshwater Ecosystems 2022, 32, 14–27. [Google Scholar] [CrossRef]
- Steneck, R.S.; Graham, M.H.; Bourque, B.J.; Corbett, D.; Erlandson, J.M.; Estes, J.A.; Tegner, M.J. Kelp Forest Ecosystems: Biodiversity, Stability, Resilience and Future. Environmental Conservation 2002, 29, 436–459. [Google Scholar] [CrossRef]
- Bringloe, T.T.; Starko, S.; Wade, R.M.; Vieira, C.; Kawai, H.; De Clerck, O.; Cock, J.M.; Coelho, S.M.; Destombe, C.; Valero, M.; et al. Phylogeny and Evolution of the Brown Algae. Critical Reviews in Plant Sciences 2020, 39, 281–321. [Google Scholar] [CrossRef]
- Saunders, G.W.; Druehl, L.D. Revision of the Kelp Family Alariaceae and the Taxonomic Affinities of Lessoniopsis Reinke (Laminariales, Phaeophyta). Hydrobiologia 1993, 260, 689–697. [Google Scholar] [CrossRef]
- Yoon, H.S.; Boo, S.M. Phylogeny of Alariaceae (Phaeophyta) with Special Reference to Undaria Based on Sequences of the RuBisCo Spacer Region. In Proceedings of the Sixteenth International Seaweed Symposium; Kain, J.M., Brown, M.T., Lahaye, M., Eds.; Springer Netherlands: Dordrecht, 1999; pp. 47–55. [Google Scholar]
- Yoon, H.S.; Lee, J.Y.; Boo, S.M.; Bhattacharya, D. Phylogeny of Alariaceae, Laminariaceae, and Lessoniaceae (Phaeophyceae) Based on Plastid-Encoded RuBisCo Spacer and Nuclear-Encoded ITS Sequence Comparisons. Molecular Phylogenetics and Evolution 2001, 21, 231–243. [Google Scholar] [CrossRef]
- Rothman, M.D.; Mattio, L.; Wernberg, T.; Anderson, R.J.; Uwai, S.; Mohring, M.B.; Bolton, J.J. A Molecular Investigation of the Genus Ecklonia (Phaeophyceae, Laminariales) with Special Focus on the Southern Hemisphere. J Phycol 2015, 51, 236–246. [Google Scholar] [CrossRef]
- Kawai, H.; Akita, S.; Hashimoto, K.; Hanyuda, T. A Multigene Molecular Phylogeny of Eisenia Reveals Evidence for a New Species, Eisenia Nipponica (Laminariales), from Japan. European Journal of Phycology 2020, 55, 234–241. [Google Scholar] [CrossRef]
- Howe, M.A. The Marine Algae of Peru. Memoirs of the Torrey Botanical Club 1914, 15, 1–185. [Google Scholar]
- Etcheverry, D.H. Algas Marinas de Las Islas Oceánicas Chilenas. Revista de Biología Marina. Revista de Biología Marina 1960, 10, 83–132. [Google Scholar]
- Dawson, Y.E.; Acleto, C.; Foldvik, N. The Seaweeds of Peru; Beihefte zur Nova Hedwigia, 1964; Vol. 13, ISBN 978-3-7682-5413-7. [Google Scholar]
- Silva, P.C.; Chacana, M.E. Marine Algae from Islas San Félix y San Ambrosio (Chilean Oceanic Islands). Cryptogamie, Algologie 2005, 26, 103–118. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: https://www.algaebase.org/ (accessed on 16 November 2025).
- Buglass, S.; Kawai, H.; Hanyuda, T.; Harvey, E.; Donner, S.; De la Rosa, J.; Keith, I.; Bermúdez, J.R.; Altamirano, M. Novel Mesophotic Kelp Forests in the Galápagos Archipelago. Mar Biol 2022, 169, 156. [Google Scholar] [CrossRef]
- Uribe, R.A.; Smale, D.A.; Morales, R.; Aleman, S.; Atoche-Suclupe, D.; Burrows, M.T.; Earp, H.S.; Hinostroza, J.D.; King, N.G.; Perea, A.; et al. Spatiotemporal Variability in the Structure and Diversity of Understory Faunal Assemblages Associated with the Kelp Eisenia Cokeri (Laminariales) in Peru. Mar Biol 2024, 171, 62. [Google Scholar] [CrossRef]
- Acleto, C. Algas marinas del Perú de importancia económica. Serie de Divulgación, 1st ed.; Museo de Historia Natural - UNMSM: Lima, Peru, 1986. [Google Scholar]
- Atoche-Suclupe, D.; Alemán, S.; Perea, Á.; Uribe, R. Variabilidad espacio temporal de la estructura poblacional, morfología y morfometría de Eisenia cokeri M.A. Howe, 1914 (Phaeophycea: Laminariales) en el nor-centro de Perú. Inf Inst Mar Perú 2021, 48, 414–429. [Google Scholar]
- Graham, M.H.; Kinlan, B.P.; Druehl, L.D.; Garske, L.E.; Banks, S. Deep-Water Kelp Refugia as Potential Hotspots of Tropical Marine Diversity and Productivity. Proceedings of the National Academy of Sciences 2007, 104, 16576–16580. [Google Scholar] [CrossRef]
- Zuta, S.; Guillén, O. Oceanografía de las aguas costeras del Perú. Boletin Instituto del Mar del Perú 1970, 2, 157–324. [Google Scholar]
- Tarazona, J.; Arntz, W. The Peruvian Coastal Upwelling System. In Coastal Marine Ecosystems of Latin America; Seeliger, U., Kjerfve, B., Eds.; Springer: Berlin, Heidelberg, 2001; pp. 229–244. ISBN 978-3-662-04482-7. [Google Scholar]
- Hollenberg, G.J. Culture Studies of Marine Algae. I. Eisenia Arborea. American Journal of Botany 1939, 26, 34. [Google Scholar] [CrossRef]
- Meneses, I.; Hoffmann, A.J. Contribution to the Marine Algal Flora of San Felix Island, Desventuradas Archipelago, Chile; 1994. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. Getting Started with PRIMER V7; PRIMER-E: Plymouth Marine Laboratory: Plymouth, United Kingdom, 2015; p. 20. [Google Scholar]
- Voisin, M.; Engel, C.R.; Viard, F. Differential Shuffling of Native Genetic Diversity across Introduced Regions in a Brown Alga: Aquaculture vs. Maritime Traffic Effects. Proceedings of the National Academy of Sciences 2005, 102, 5432–5437. [Google Scholar] [CrossRef]
- Macaya, E.C.; Zuccarello, G.C. Genetic Structure of the Giant Kelp Macrocystis Pyrifera along the Southeastern Pacific. Marine Ecology Progress Series 2010, 420, 103–112. [Google Scholar] [CrossRef]
- McCarthy, C. Chromas; Version 1.41; School of Biomolecular and Biomedical Science; Griffith University: Brisbane, Queensland, 1997. [Google Scholar]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic acids symposium series 1999, 41, 95–98. [Google Scholar]
- Tellier, F.; Meynard, A.P.; Correa, J.A.; Faugeron, S.; Valero, M. Phylogeographic Analyses of the 30°S South-East Pacific Biogeographic Transition Zone Establish the Occurrence of a Sharp Genetic Discontinuity in the Kelp Lessonia Nigrescens: Vicariance or Parapatry? Molecular Phylogenetics and Evolution 2009, 53, 679–693. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More Models, New Heuristics and Parallel Computing. Nat Methods 2012, 9, 772–772. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Molecular Biology and Evolution 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A Software for Comprehensive Analysis of DNA Polymorphism Data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-Joining Networks for Inferring Intraspecific Phylogenies. Mol Biol Evol 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Crandall, K.A.; Templeton, A.R. Empirical Tests of Some Predictions from Coalescent Theory with Applications to Intraspecific Phylogeny Reconstruction. Genetics 1993, 134, 959–969. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian Phylogenetics with BEAUti and the BEAST 1.7. Mol Biol Evol 2012, 29, 1969–1973. [Google Scholar] [CrossRef]
- Fujisawa, T.; Barraclough, T.G. Delimiting Species Using Single-Locus Data and the Generalized Mixed Yule Coalescent Approach: A Revised Method and Evaluation on Simulated Data Sets. Syst Biol 2013, 62, 707–724. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing 2025.
- Friedlander, A.M.; Ballesteros, E.; Caselle, J.E.; Gaymer, C.F.; Palma, A.T.; Petit, I.; Varas, E.; Muñoz Wilson, A.; Sala, E. Marine Biodiversity in Juan Fernández and Desventuradas Islands, Chile: Global Endemism Hotspots. PLoS ONE 2016, 11, e0145059. [Google Scholar] [CrossRef] [PubMed]
- Mols-Mortensen, A.; Neefus, C.D.; Nielsen, R.; Gunnarsson, K.; Egilsdóttir, S.; Pedersen, P.Mø.; Brodie, J. New Insights into the Biodiversity and Generic Relationships of Foliose Bangiales (Rhodophyta) in Iceland and the Faroe Islands. European Journal of Phycology 2012, 47, 146–159. [Google Scholar] [CrossRef]
- Carstens, B.C.; Pelletier, T.A.; Reid, N.M.; Satler, J.D. How to Fail at Species Delimitation. Molecular Ecology 2013, 22, 4369–4383. [Google Scholar] [CrossRef]
- Modica, M.V.; Puillandre, N.; Castelin, M.; Zhang, Y.; Holford, M. A Good Compromise: Rapid and Robust Species Proxies for Inventorying Biodiversity Hotspots Using the Terebridae (Gastropoda: Conoidea). PLOS ONE 2014, 9, e102160. [Google Scholar] [CrossRef]
- Fujisawa, T.; Barraclough, T.G. Delimiting Species Using Single-Locus Data and the Generalized Mixed Yule Coalescent Approach: A Revised Method and Evaluation on Simulated Data Sets. Syst Biol 2013, 62, 707–724. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, H.; Yamaji, F.; Ohsawa, T.A. Genetic Structure and Dispersal Patterns in Limnoria Nagatai (Limnoriidae, Isopoda) Dwelling in Non-Buoyant Kelps, Eisenia Bicyclis and E. Arborea, in Japan. PLOS ONE 2018, 13, e0198451. [Google Scholar] [CrossRef] [PubMed]
- Provan, J.; Glendinning, K.; Kelly, R.; Maggs, C.A. Levels and Patterns of Population Genetic Diversity in the Red Seaweed Chondrus Crispus (Florideophyceae): A Direct Comparison of Single Nucleotide Polymorphisms and Microsatellites. Biol J Linn Soc 2013, 108, 251–262. [Google Scholar] [CrossRef]
- Camus, C.; Faugeron, S.; Buschmann, A.H. Assessment of Genetic and Phenotypic Diversity of the Giant Kelp, Macrocystis Pyrifera, to Support Breeding Programs. Algal Research 2018, 30, 101–112. [Google Scholar] [CrossRef]
- Evankow, A.; Christie, H.; Hancke, K.; Brysting, A.K.; Junge, C.; Fredriksen, S.; Thaulow, J. Genetic Heterogeneity of Two Bioeconomically Important Kelp Species along the Norwegian Coast. Conserv Genet 2019, 20, 615–628. [Google Scholar] [CrossRef]
- Pequeño, G.; Lamilla, J. The Littoral Fish Assemblage of the Desventuradas Islands (Chile) Has Zoogeographical Affinities with the Western Pacific. Global Ecology and Biogeography 2000, 9, 431–437. [Google Scholar] [CrossRef]
- Retamal, M.A.; Moyano, H.I. Zoogeography of Chilean Marine and Freshwater Decapod Crustaceans. Latin American Journal of Aquatic Research 2010, 38, 302–328. [Google Scholar] [CrossRef]
- Verbruggen, H.; De Clerck, O.; Cocquyt, E.; Kooistra, W.H.C.F.; Coppejans, E. Morphometric Taxonomy of Siphonous Green Algae: A Methodological Study Within the Genus Halimeda (Bryopsidales). Journal of Phycology 2005, 41, 126–139. [Google Scholar] [CrossRef]
- Zanolla, M.; Carmona, R.; De la Rosa, J.; Salvador, N.; Sherwood, A.R.; Andreakis, N.; Altamirano, M. Morphological Differentiation of Cryptic Lineages within the Invasive Genus Asparagopsis (Bonnemaisoniales, Rhodophyta). Phycologia 2014, 53, 233–242. [Google Scholar] [CrossRef]
- Smith, K.E.; Moore, P.J.; King, N.G.; Smale, D.A. Examining the Influence of Regional-Scale Variability in Temperature and Light Availability on the Depth Distribution of Subtidal Kelp Forests. Limnology and Oceanography 2022, 67, 314–328. [Google Scholar] [CrossRef]
- Witman, J.D. Subtidal Coexistence: Storms, Grazing, Mutualism, and the Zonation of Kelps and Mussels. Ecological Monographs 1987, 57, 167–187. [Google Scholar] [CrossRef]
Figure 1.
Map of sampling localities for Eisenia cokeri (circles) and Eisenia gracilis (stars) in Peru, and Eisenia specimens in Chile (square).
Figure 1.
Map of sampling localities for Eisenia cokeri (circles) and Eisenia gracilis (stars) in Peru, and Eisenia specimens in Chile (square).
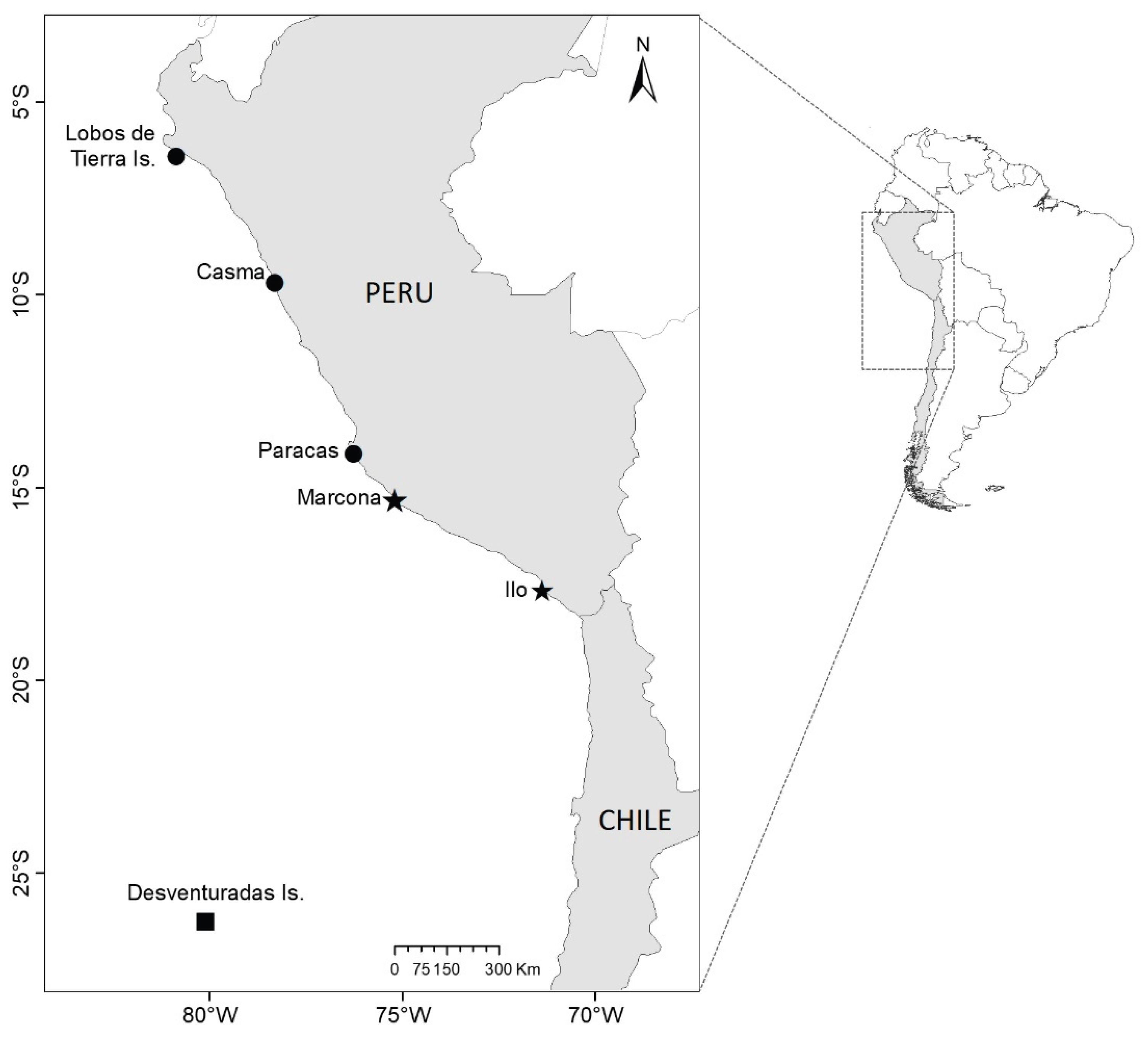
Figure 2.
Habitat and morphology of Eisenia cokeri sporophytes from Peru. (A) Subtidal bed, Lobos de Tierra Island. (B) Holdfast attached to a rocky substrate, Lobos de Tierra Island. (C) Small adult specimen at a rocky intertidal shelf, showing a holdfast and a short stipe, Casma. (D) Detail of a solid stipe (arrow) and branched holdfast, Casma. (E,F) Entire juvenile fronds with scarce roughness, Casma. (G) and (H) Rough juvenile fronds bearing marginal pinnae of different sizes and smooth margins, Casma. (I,J) Juvenile specimens showing main and lanceolate secondary blades, Paracas and Huarmey (IMARPE 05-000343). (K,L) Juvenile fronds with a divided primary blade, a thickened base and serrated margins, Paracas. (M) Detail of a sporophyll, with serrated edges and sorus, Casma. (N) Specimen detached from subtidal populations, with a long stipe and blades grouped in two distal lateral branches, Rancherío. Scale bar = 5 cm. Photographs from P. Carbajal & N. Arakaki.
Figure 2.
Habitat and morphology of Eisenia cokeri sporophytes from Peru. (A) Subtidal bed, Lobos de Tierra Island. (B) Holdfast attached to a rocky substrate, Lobos de Tierra Island. (C) Small adult specimen at a rocky intertidal shelf, showing a holdfast and a short stipe, Casma. (D) Detail of a solid stipe (arrow) and branched holdfast, Casma. (E,F) Entire juvenile fronds with scarce roughness, Casma. (G) and (H) Rough juvenile fronds bearing marginal pinnae of different sizes and smooth margins, Casma. (I,J) Juvenile specimens showing main and lanceolate secondary blades, Paracas and Huarmey (IMARPE 05-000343). (K,L) Juvenile fronds with a divided primary blade, a thickened base and serrated margins, Paracas. (M) Detail of a sporophyll, with serrated edges and sorus, Casma. (N) Specimen detached from subtidal populations, with a long stipe and blades grouped in two distal lateral branches, Rancherío. Scale bar = 5 cm. Photographs from P. Carbajal & N. Arakaki.

Figure 3.
Habitat and morphology of Eisenia gracilis sporophytes from Peru. (A) Subtidal bed, Ilo. (B) Holdfast attached to a rocky substrate, Ilo. (C) Detail of a branched holdfast with new stipes at the ends, Ilo. (D) Entire juvenile frond, Marcona. (E) Juvenile frond with short, spaced marginal pinnae, Marcona. (F) Juvenile specimen with alternately arranged and smooth-edged secondary blades, Marcona. (G) Adult uncommon specimen with short stipe, primary blade and serrated-edge secondary blades, Ilo. (H) Juvenile specimen with a divided primary blade, thickened base and long secondary blades, Marcona. (I) Blade detail with serrated edges with spaced teeth (right), and sorus (left), Ilo. (J) Adult specimen appearing fork-like, showing a long stipe, divided primary blade, and long lateral fronds, Ilo. Photographs from P. Carbajal & N. Arakaki.
Figure 3.
Habitat and morphology of Eisenia gracilis sporophytes from Peru. (A) Subtidal bed, Ilo. (B) Holdfast attached to a rocky substrate, Ilo. (C) Detail of a branched holdfast with new stipes at the ends, Ilo. (D) Entire juvenile frond, Marcona. (E) Juvenile frond with short, spaced marginal pinnae, Marcona. (F) Juvenile specimen with alternately arranged and smooth-edged secondary blades, Marcona. (G) Adult uncommon specimen with short stipe, primary blade and serrated-edge secondary blades, Ilo. (H) Juvenile specimen with a divided primary blade, thickened base and long secondary blades, Marcona. (I) Blade detail with serrated edges with spaced teeth (right), and sorus (left), Ilo. (J) Adult specimen appearing fork-like, showing a long stipe, divided primary blade, and long lateral fronds, Ilo. Photographs from P. Carbajal & N. Arakaki.

Figure 4.
Principal Component Analysis of morphological variables and biomass of Eisenia cokeri (green) and Eisenia gracilis (blue). MHD: maximum holdfast diameter; HH: holdfast height; SL: stipe length; SDI: stipe diameter; TL: total length; TFW: total fresh weight.
Figure 4.
Principal Component Analysis of morphological variables and biomass of Eisenia cokeri (green) and Eisenia gracilis (blue). MHD: maximum holdfast diameter; HH: holdfast height; SL: stipe length; SDI: stipe diameter; TL: total length; TFW: total fresh weight.
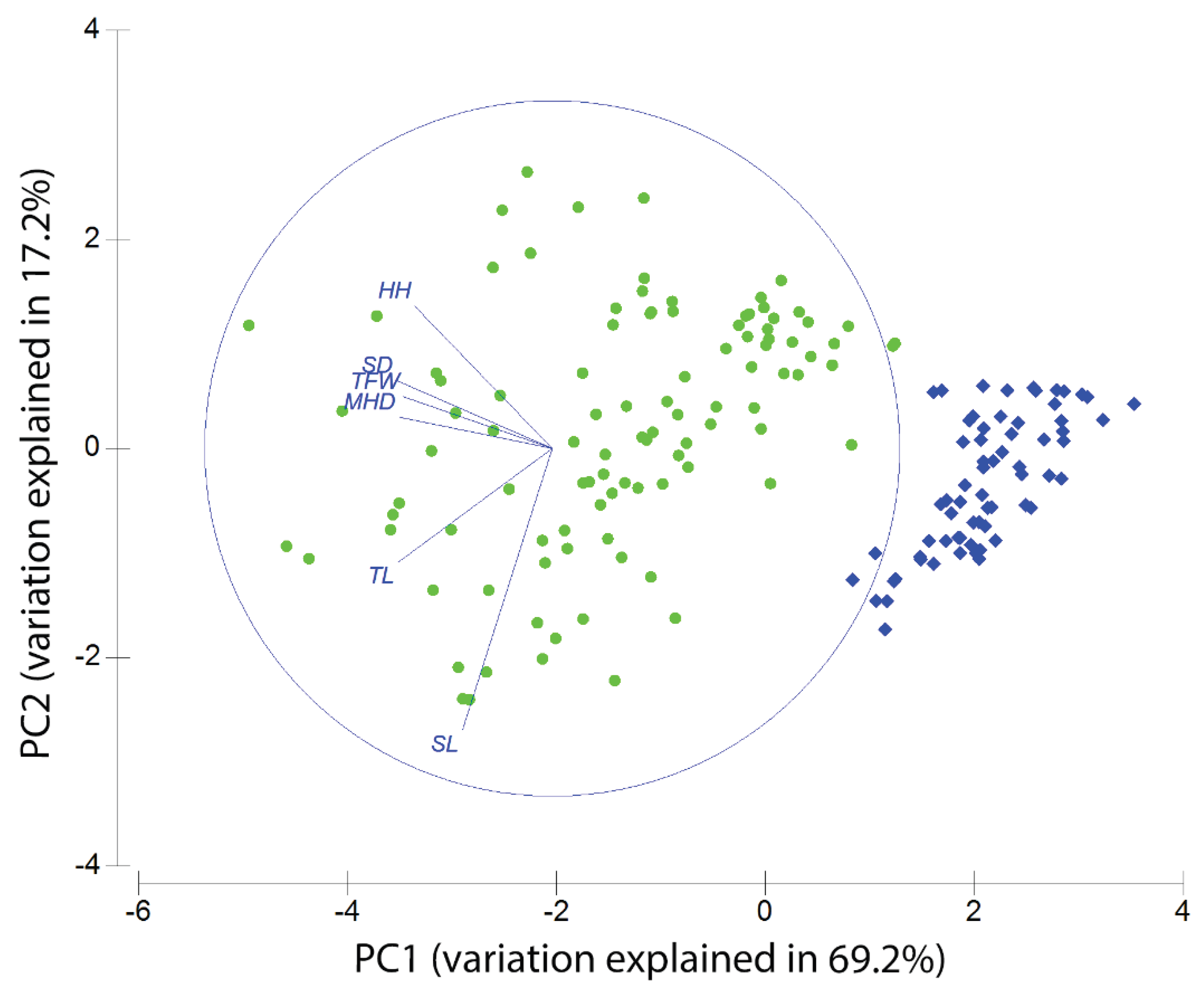
Figure 5.
Phylogenetic tree of Eisenia DNA sequences constructed from concatenated markers. Support values were calculated using ML (bootstrap values on the left) and BI (posterior probabilities on the right) methods. Displayed are only those values exceeding the thresholds of 70% for ML and 0.80 PP.
Figure 5.
Phylogenetic tree of Eisenia DNA sequences constructed from concatenated markers. Support values were calculated using ML (bootstrap values on the left) and BI (posterior probabilities on the right) methods. Displayed are only those values exceeding the thresholds of 70% for ML and 0.80 PP.
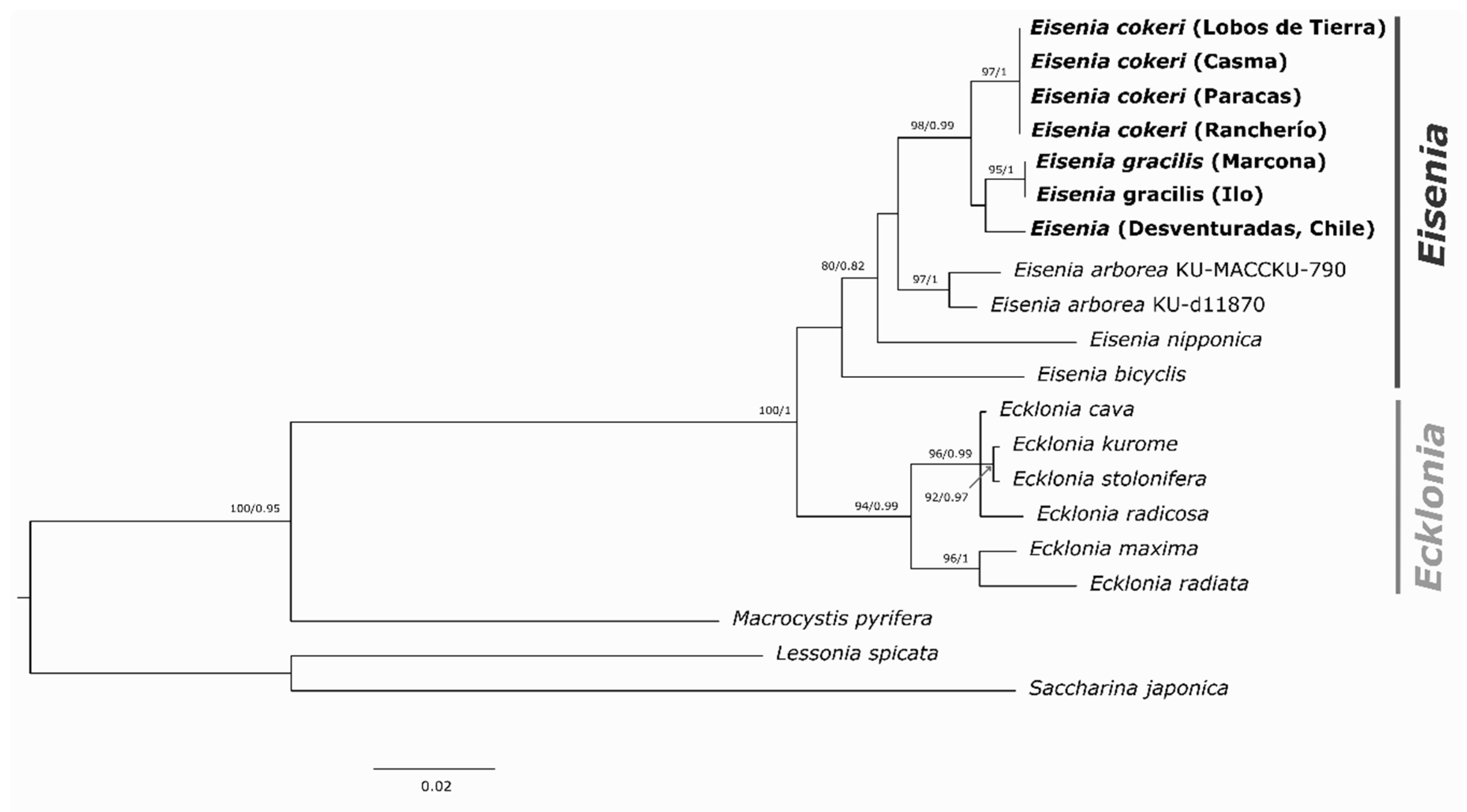
Figure 6.
Haplotype networks for Eisenia based on DNA sequences obtained in this study for the markers ATP8S, COI, WI, rbcL, and RS. Circle size is proportional to the number of individuals per haplotype, and pie-chart colors indicate species assignment (red: E. cokeri [Peru]; blue: E. gracilis [Peru]; gray: Eisenia [Desventuradas Islands]). Line connecting haplotypes represent mutational pathways: circles along the lines denote single nucleotide substitutions, perpendicular bars indicate indels, and boxed bars correspond to blocks of 10 tandem indels. Unlabeled yellow nodes represent inferred intermediate haplotypes not detected in the sampled populations.
Figure 6.
Haplotype networks for Eisenia based on DNA sequences obtained in this study for the markers ATP8S, COI, WI, rbcL, and RS. Circle size is proportional to the number of individuals per haplotype, and pie-chart colors indicate species assignment (red: E. cokeri [Peru]; blue: E. gracilis [Peru]; gray: Eisenia [Desventuradas Islands]). Line connecting haplotypes represent mutational pathways: circles along the lines denote single nucleotide substitutions, perpendicular bars indicate indels, and boxed bars correspond to blocks of 10 tandem indels. Unlabeled yellow nodes represent inferred intermediate haplotypes not detected in the sampled populations.
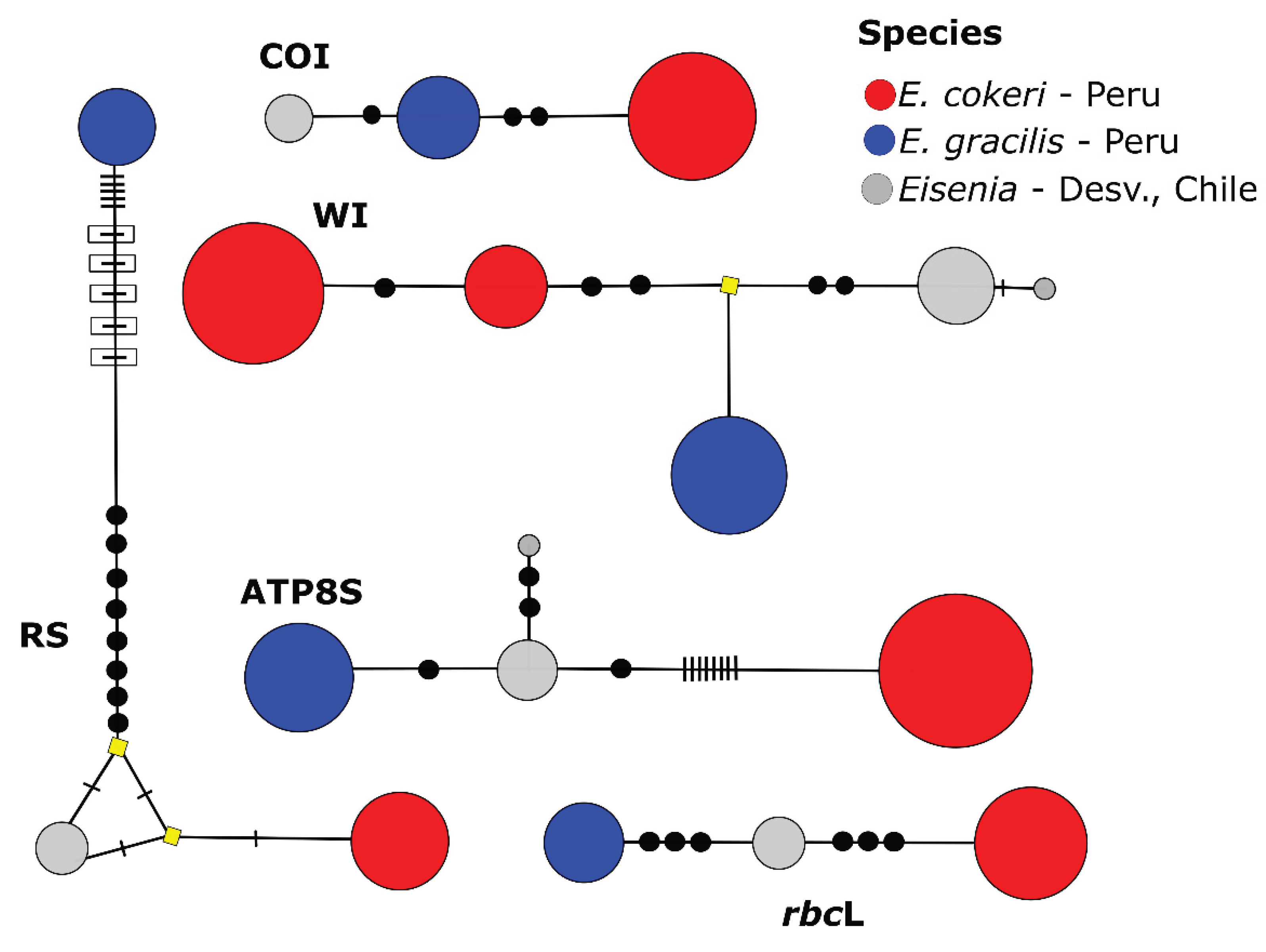

Table 1.
Collection information and GenBank accession numbers for the specimens of Eisenia from Peru and Desventuradas Islands, Chile, included in the molecular analysis. Eisenia from Desventuradas Islands is considered as a species not a priori determined.
Table 1.
Collection information and GenBank accession numbers for the specimens of Eisenia from Peru and Desventuradas Islands, Chile, included in the molecular analysis. Eisenia from Desventuradas Islands is considered as a species not a priori determined.
| Species (Area) |
Locality | Accession Number | ||||
|---|---|---|---|---|---|---|
| ATP8S | COI | WI | RS | rbcL | ||
| E. cokeri (Peru) | Lobos de Tierra Island, Lambayeque, coll. P. Carbajal & Iván Gómez, 06°25'30.4" S, 80°51'58.9" W |
OR361924 | OR361839 | OR362016 | OR362119 | OR361884 |
| OR361925 | OR361840 | OR362017 | OR362120 | OR361885 | ||
| OR361926 | OR361841 | OR362018 | OR362121 | OR361886 | ||
| OR361927 | OR361842 | OR362019 | OR362122 | OR361887 | ||
| OR361928 | OR361843 | OR362020 | OR362123 | OR361888 | ||
| OR361929 | OR361844 | OR362021 | OR362124 | OR361889 | ||
| OR361930 | OR362022 | |||||
| OR361931 | OR362023 | |||||
| OR361932 | OR362024 | |||||
| OR361933 | OR362025 | |||||
| OR361934 | OR362026 | |||||
| OR361935 | OR362027 | |||||
| OR361936 | OR362028 | |||||
| OR361937 | OR362029 | |||||
| OR361938 | OR362030 | |||||
| E. cokeri (Peru) | Casma, Ancash, coll. D. Hinostroza, 09°42'22.8" S, 78°17'49.8" W |
OR361939 | OR361845 | OR362031 | OR362125 | OR361890 |
| OR361940 | OR361846 | OR362032 | OR362126 | OR361891 | ||
| OR361941 | OR361847 | OR362033 | OR362127 | OR361892 | ||
| OR361942 | OR361848 | OR362034 | OR362128 | OR361893 | ||
| OR361943 | OR361849 | OR362035 | OR362129 | OR361894 | ||
| OR361944 | OR362036 | |||||
| OR361945 | OR362037 | |||||
| OR361946 | OR362038 | |||||
| OR361947 | OR362039 | |||||
| OR361948 | OR362040 | |||||
| OR361949 | OR362041 | |||||
| OR361950 | OR362042 | |||||
| OR361951 | OR362043 | |||||
| OR361952 | OR362044 | |||||
| OR361953 | ||||||
| E. cokeri (Peru) | El Ancla, Paracas, Ica, coll. P. Carbajal, 14°09'10.5" S, 76°14'52.9" W |
OR361954 | OR361850 | OR362045 | OR362130 | OR361895 |
| OR361955 | OR361851 | OR362046 | OR362131 | OR361896 | ||
| OR361956 | OR361852 | OR362047 | OR362132 | OR361897 | ||
| OR361957 | OR361853 | OR362048 | OR362133 | OR361898 | ||
| OR361958 | OR361854 | OR362049 | OR362134 | OR361899 | ||
| OR361959 | OR362050 | |||||
| OR361960 | OR362051 | |||||
| OR361961 | OR362052 | |||||
| OR361962 | OR362053 | |||||
| OR361963 | OR362054 | |||||
| OR361964 | OR362055 | |||||
| OR361965 | OR362056 | |||||
| OR361966 | OR362057 | |||||
| OR361967 | OR362058 | |||||
| OR361968 | OR362059 | |||||
| E. cokeri (Peru) | Rancherío, Laguna Grande, Ica, coll. P. Carbajal, 14°15'10.5" S, 76°25'52.9" W |
OR361969 | OR361855 | OR362060 | OR362135 | OR361900 |
| OR361970 | OR361856 | OR362061 | OR362136 | OR361901 | ||
| OR361971 | OR361857 | OR362062 | OR362137 | OR361902 | ||
| OR361972 | OR361858 | OR362063 | OR362138 | OR361903 | ||
| OR361973 | OR361859 | OR362064 | OR362139 | OR361904 | ||
| OR361974 | OR361860 | OR362065 | ||||
| OR361975 | OR361861 | OR362066 | ||||
| OR361862 | OR362067 | |||||
| OR361863 | OR362068 | |||||
| OR361864 | OR362069 | |||||
| OR362070 | ||||||
| OR362071 | ||||||
| OR362072 | ||||||
| OR362073 | ||||||
| OR362074 | ||||||
| Eisenia (Desventuradas Is.) | San Ambrosio Island, Desventuradas Island, coll. P. Manríquez Ángulo, 26°20'12.8" S, 79°53'31.4" W |
OR361976 | OR361865 | OR362075 | OR362140 | OR361905 |
| OR361977 | OR361866 | OR362076 | OR362141 | OR361906 | ||
| OR361978 | OR361867 | OR362077 | OR362142 | OR361907 | ||
| OR361979 | OR361868 | OR362078 | OR362143 | OR361908 | ||
| OR361980 | OR361869 | OR362079 | OR362144 | OR361909 | ||
| OR361981 | OR361870 | OR362080 | OR362145 | OR361910 | ||
| OR361982 | OR362081 | |||||
| OR361983 | OR362082 | |||||
| OR361984 | OR362083 | |||||
| OR361985 | OR362084 | |||||
| OR361986 | OR362085 | |||||
| OR361987 | OR362086 | |||||
| OR361988 | OR362087 | |||||
| OR361989 | OR362088 | |||||
| E. gracilis (Peru) | Tres Hermanas, Marcona, Ica, coll. P. Carbajal, 15°26'29.1" S, 75°04'32.5" W |
OR361990 | OR361871 | OR362089 | OR362146 | OR361911 |
| OR361991 | OR361872 | OR362090 | OR362147 | OR361912 | ||
| OR361992 | OR361873 | OR362091 | OR362148 | OR361913 | ||
| OR361993 | OR361874 | OR362092 | OR362149 | OR361914 | ||
| OR361994 | OR361875 | OR362093 | OR362150 | OR361915 | ||
| OR361995 | OR361876 | OR362094 | OR362151 | OR361916 | ||
| OR361996 | OR361877 | OR362095 | OR362152 | OR361917 | ||
| OR361997 | OR361878 | OR362096 | OR362153 | OR361918 | ||
| OR361998 | OR362097 | |||||
| OR361999 | OR362098 | |||||
| OR362000 | OR362099 | |||||
| OR362001 | OR362100 | |||||
| OR362101 | ||||||
| OR362102 | ||||||
| OR362103 | ||||||
| E. gracilis (Peru) | Leonas, Ilo, Moquegua, coll. P. Carbajal, 17°40'39.3" S, 71°22'15.9" W |
OR362002 | OR361879 | OR362104 | OR362154 | OR361919 |
| OR362003 | OR361880 | OR362105 | OR362155 | OR361920 | ||
| OR362004 | OR361881 | OR362106 | OR362156 | OR361921 | ||
| OR362005 | OR361882 | OR362107 | OR362157 | OR361922 | ||
| OR362006 | OR361883 | OR362108 | OR362158 | OR361923 | ||
| OR362007 | OR362109 | |||||
| OR362008 | OR362110 | |||||
| OR362009 | OR362111 | |||||
| OR362010 | OR362112 | |||||
| OR362011 | OR362113 | |||||
| OR362012 | OR362114 | |||||
| OR362013 | OR362115 | |||||
| OR362014 | OR362116 | |||||
| OR362015 | OR362117 | |||||
| OR362118 | ||||||
Table 2.
Morphometric variables and biomass of Eisenia cokeri and Eisenia gracilis collected in Peru during 2018 and 2019.
Table 2.
Morphometric variables and biomass of Eisenia cokeri and Eisenia gracilis collected in Peru during 2018 and 2019.
| Variable | E. cokeri | E. gracilis | |
|---|---|---|---|
| Maximum holdfast diameter (cm) |
Mean | 18,8 | 7,1 |
| DE | 4,0 | 2,8 | |
| Min | 11,0 | 2,0 | |
| Max | 33,5 | 13,0 | |
| Holdfast height (cm) | Mean | 7,4 | 2,7 |
| DE | 2,7 | 1,0 | |
| Min | 3,0 | 1,0 | |
| Max | 16,0 | 6,0 | |
| Stipe length (cm) | Mean | 32,5 | 24,8 |
| DE | 20,9 | 13,2 | |
| Min | 4,4 | 6,0 | |
| Max | 84,5 | 52,0 | |
| Stipe diameter (cm) | Mean | 1,4 | 0,5 |
| DE | 0,3 | 0,1 | |
| Min | 0,9 | 0,3 | |
| Max | 2,3 | 0,8 | |
| Total length (cm) | Mean | 132,4 | 70,4 |
| DE | 41,6 | 21,9 | |
| Min | 1,3 | 24,0 | |
| Max | 220,0 | 114,0 | |
| Total fresh weight (g) | Mean | 815 | 25 |
| DE | 536 | 16 | |
| Min | 80 | 5 | |
| Max | 2780 | 90 |
Table 3.
Species delimited using different gene regions implemented in GMYC using a single threshold.
Table 3.
Species delimited using different gene regions implemented in GMYC using a single threshold.
| Laminariales | |||||
|---|---|---|---|---|---|
| ATP8S | WI | COI | RS | rbcL | |
| Likelihood of null model | 538 | 815 | 569 | 1651 | 667 |
| Maximum likelihood of GMYC model | 543 | 815 | 597 | 1678 | 668 |
| Likelihood ratio | 11** | 0.14 n.s | 6*** | 574*** | 15*** |
| Number of ML clusters (95% CI) | 10 (8–12) | 13 (6–21) | 9 (8–10) | 12 (11–13) | 14 (12-16) |
| Number of Eisenia species in SEP | 3 | 3 | 3 | 3 | 3 |
Table 4.
Number of DNA sequences generated for each genomic marker and locality. Eisenia from Desventuradas Islands, Chile, is considered as a species not a priori determined.
Table 4.
Number of DNA sequences generated for each genomic marker and locality. Eisenia from Desventuradas Islands, Chile, is considered as a species not a priori determined.
| Species/Area | Locality | Genomic compartment | |||||
|---|---|---|---|---|---|---|---|
| Mitochondrial | Chloroplast | ||||||
| ATP8S | COI | WI | RS | rbcL | |||
| E. cokeri (Peru) | Lobos de Tierra I. | 15 | 6 | 15 | 6 | 6 | |
| Casma | 15 | 5 | 14 | 5 | 5 | ||
| Paracas | 15 | 5 | 15 | 5 | 5 | ||
| Rancherío | 7 | 10 | 15 | 5 | 5 | ||
| Eisenia (Desventuradas Is.) | Desventuradas Is. | 14 | 6 | 14 | 6 | 6 | |
| E. gracilis (Peru) | Marcona | 12 | 8 | 15 | 8 | 8 | |
| Ilo | 14 | 5 | 15 | 5 | 5 | ||
| Total number of specimens | 85 | 34 | 88 | 35 | 35 | ||
Table 5.
Genetic diversity of Eisenia at the species or species group level (Eisenia from Desventuradas Islands, Chile, is considered as a species not a priori determined), for ATP8S, COI, WI, RS and rbcL markers. Complete alignments are considered.
Table 5.
Genetic diversity of Eisenia at the species or species group level (Eisenia from Desventuradas Islands, Chile, is considered as a species not a priori determined), for ATP8S, COI, WI, RS and rbcL markers. Complete alignments are considered.
| Marker | ATP8S | COI | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Group |
E. cokeri (Peru) |
E. gracilis (Peru) |
E. cokeri + E. gracilis (Peru) |
Eisenia (Desv.) |
Total Eisenia |
E. cokeri (Peru) |
E. gracilis (Peru) |
E. cokeri + E. gracilis (Peru) |
Eisenia (Desv.) |
Total Eisenia |
|
| bp | 162 | 162 | 162 | 162 | 162 | 602 | 602 | 602 | 602 | 602 | |
| N | 45 | 26 | 71 | 14 | 85 | 16 | 13 | 29 | 6 | 34 | |
| Nhap | 1 | 1 | 2 | 2 | 4 | 1 | 1 | 2 | 1 | 3 | |
| Nhpriv | 1 | 1 | 2 | 2 | 4 | 1 | 1 | 2 | 1 | 3 | |
| S | 0 | 0 | 9 | 2 | 11 | 0 | 0 | 2 | 0 | 4 | |
| Ssubst | 0 | 0 | 2 | 2 | 4 | 0 | 0 | 2 | 0 | 4 | |
| Sindels | 0 | 0 | 7 | 0 | 7 | 0 | 0 | 0 | 0 | 0 | |
| Nindels | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | |
| H | 0 | 0 | 0.471 | 0.143 | 0.610 | 0 | 0 | 0.512 | 0 | 0.629 | |
| H (SD) | 0 | 0 | 0.032 | 0.119 | 0.033 | 0 | 0 | 0.031 | 0 | 0.042 | |
| π | 0 | 0 | 0.006 | 0.176 | 0.006 | 0 | 0 | 0.002 | 0 | 0.003 | |
| π (SD) | 0 | 0 | 4E-04 | 0.001 | 4E-04 | 0 | 0 | 1E-04 | 0 | 3E-04 | |
| Π | 0 | 0 | 0.942 | 0.286 | 0.981 | 0 | 0 | 1.025 | 0 | 1.544 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



