Submitted:
02 December 2025
Posted:
02 December 2025
You are already at the latest version
Abstract
The Chongming Dongtan wetland, a representative coastal wetland in East Asia, is subject to a significant ecological threat from the invasive species Spartina alterniflora. The mixed ecotone formed between this invasive species and the native Phragmites australis serves as a highly sensitive and critical indicator of alterations in wetland ecosystem structure and function. Using spring and autumn Sentinel-2 imagery from 2016 to 2023, this study developed a method that integrates a three-dimensional feature space with multi-threshold Otsu segmentation to accurately extract the mixed S. alterniflora–P. australis ecotone. The spatiotemporal dynamics of the mixed ecotone were analyzed at multiple temporal scales using a centroid migration model and the Seasonal Area Ratio (SAR) index. The results suggest that: (1) Near-infrared reflectance and NDVI were identified as the optimal spectral indices for spring and autumn, respectively, which led to a classification achieving an overall accuracy of 87.3±1.4% and a Kappa coefficient of 0.84±0.02. Notably, the mixed ecotone was mapped with producer’s and user’s accuracies of 85.2% and 83.6%. (2) The vegetation followed a distinct land-to-sea ecological sequence of “pure P. australis–mixed ecotone–pure S. alterniflora”, predominantly distributed as an east–west trending belt. This pattern was fragmented by tidal creeks and micro-topography in the northwest, contrasting with geometrically regular linear anomalies in the central area, indicative of human engineering. (3) The ecotone saw continuous seaward expansion throughout the 2016–2023 period. Spring exhibited a consistent annual area growth of 13.93% and a stable seaward centroid migration, whereas autumn exhibited significant intra-annual fluctuations in both area and centroid due to extreme climate events. (4) The SAR index uncovered a fundamental transition in the seasonal competition pattern in 2017, initiating a seven-year spring-dominant phase after a single year of autumn dominance. This spring-dominated era exhibited a distinctive sawtooth fluctuation pattern, indicative of competitive dynamics arising from the phenological advancement of P. australis combined with the niche penetration of S. alterniflora. This study elucidates the multi-scale competition and succession mechanisms between S. alterniflora and P. australis, thus providing a scientific underpinning for effective invasive species control and ecological restoration in coastal wetlands.
Keywords:
1. Introduction
2. Study Area, Data Sources, and Methodology
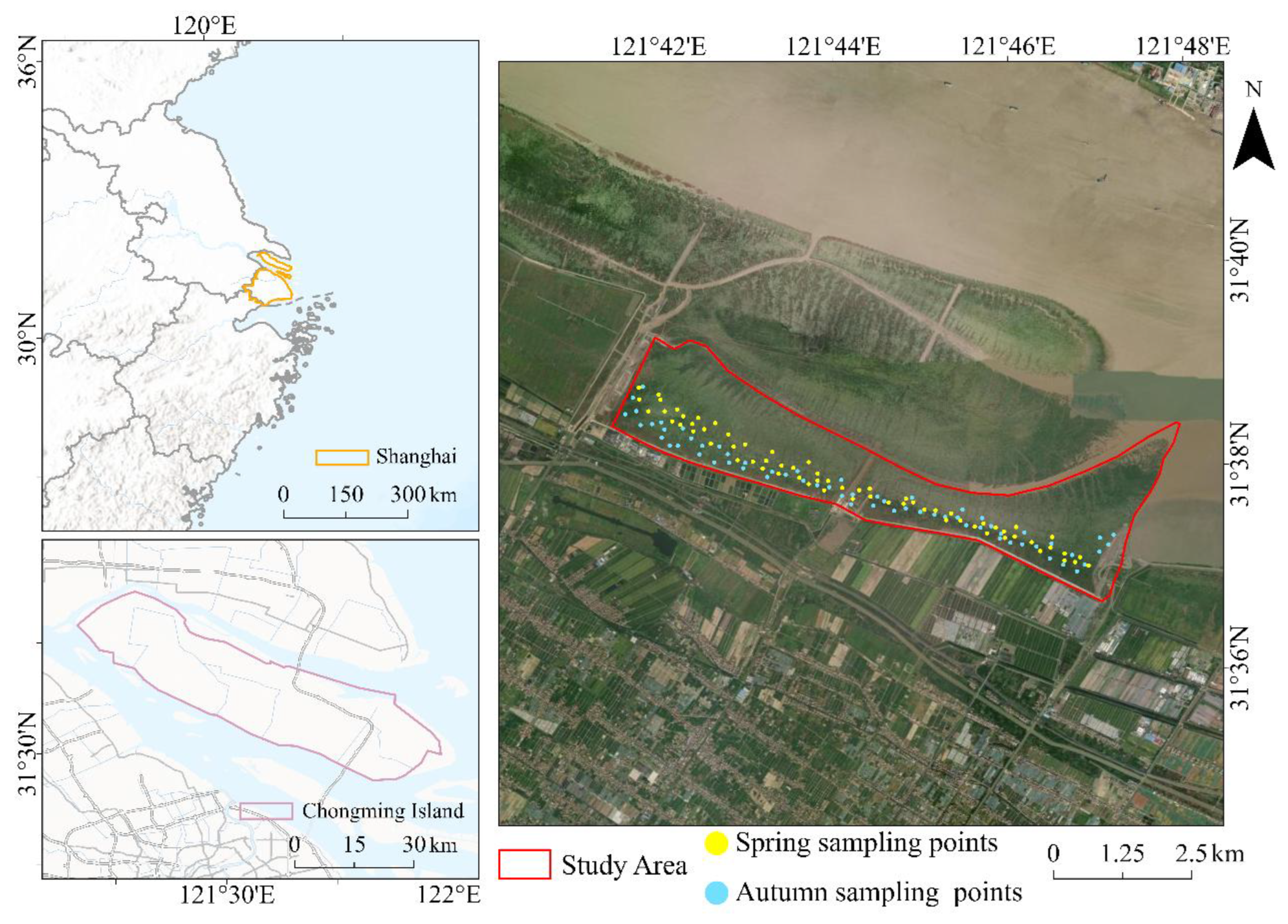
2.1. Study Area
2.2. Data Source and Preprocessing
2.3. Methodology
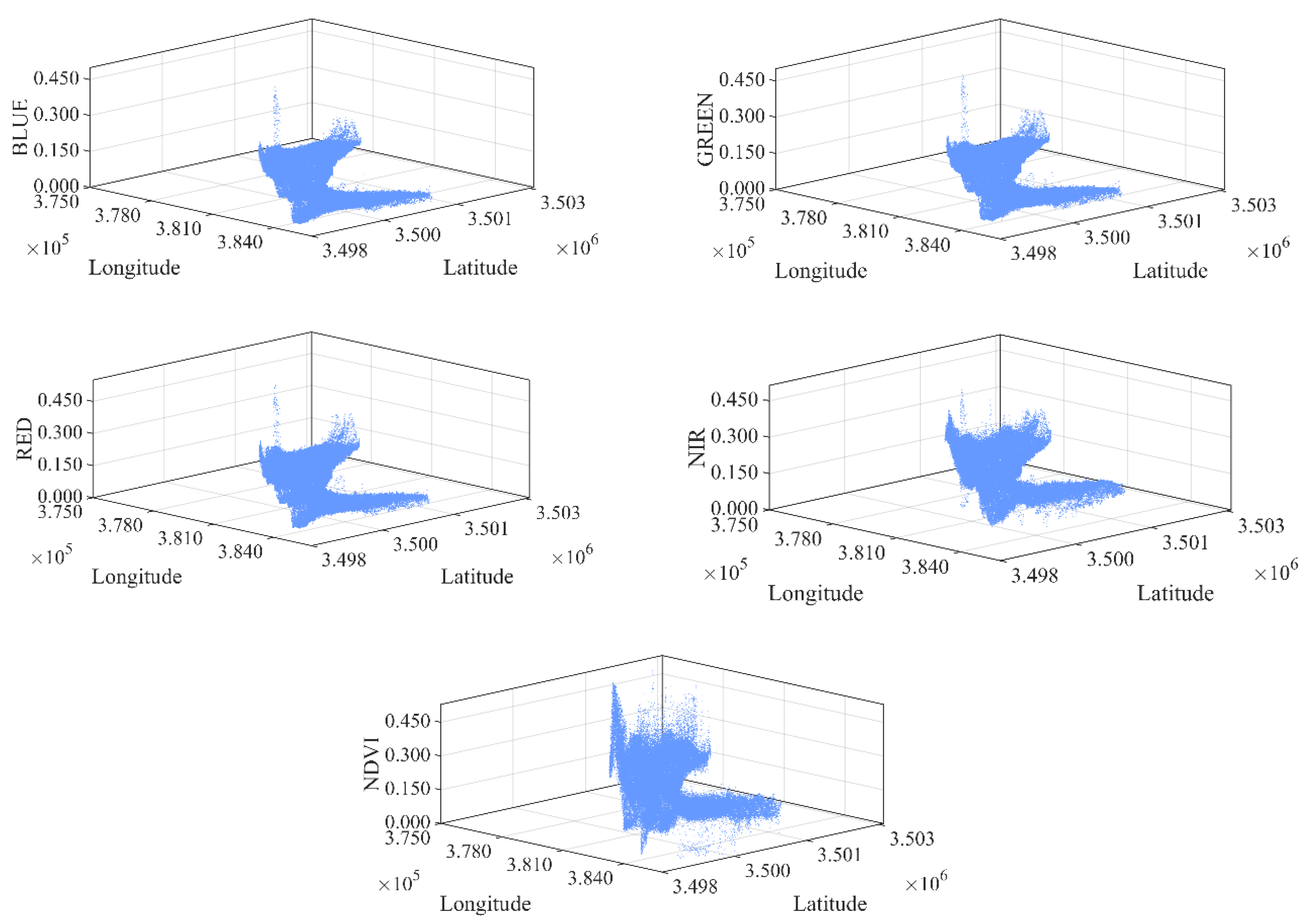
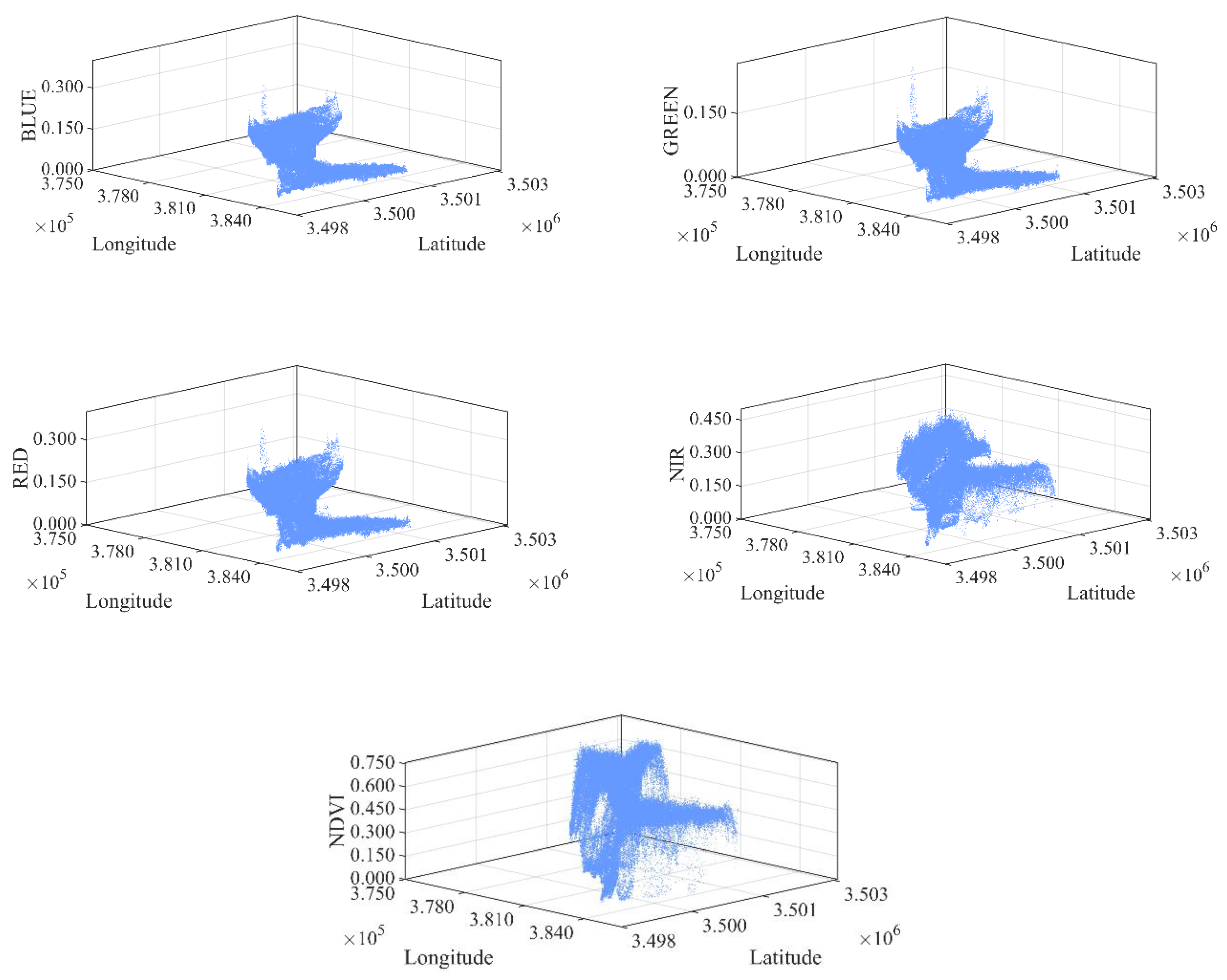
2.3.1. Delineation of the Mixed Ecotone
- Spectral Feature Extraction and 3D Feature Space Construction.
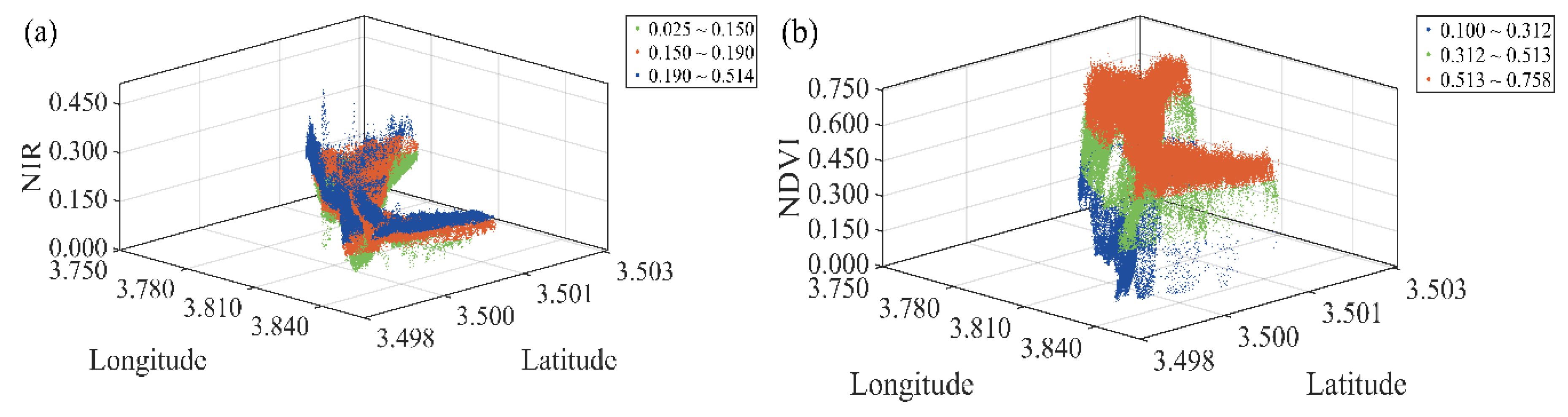
- Optimal Spectral Index Selection and Multi-threshold Otsu Segmentation.
- Time-Series Classification and Accuracy Validation.
2.3.2. Spatiotemporal Dynamics Monitoring of the Mixed Ecotone
- Centroid Migration Model.
- Seasonal Area Ratio (SAR) Index.
3. Results and Analysis
3.1. Selection of Optimal Spectral Indices
3.2. Accuracy Validation of Classification Results
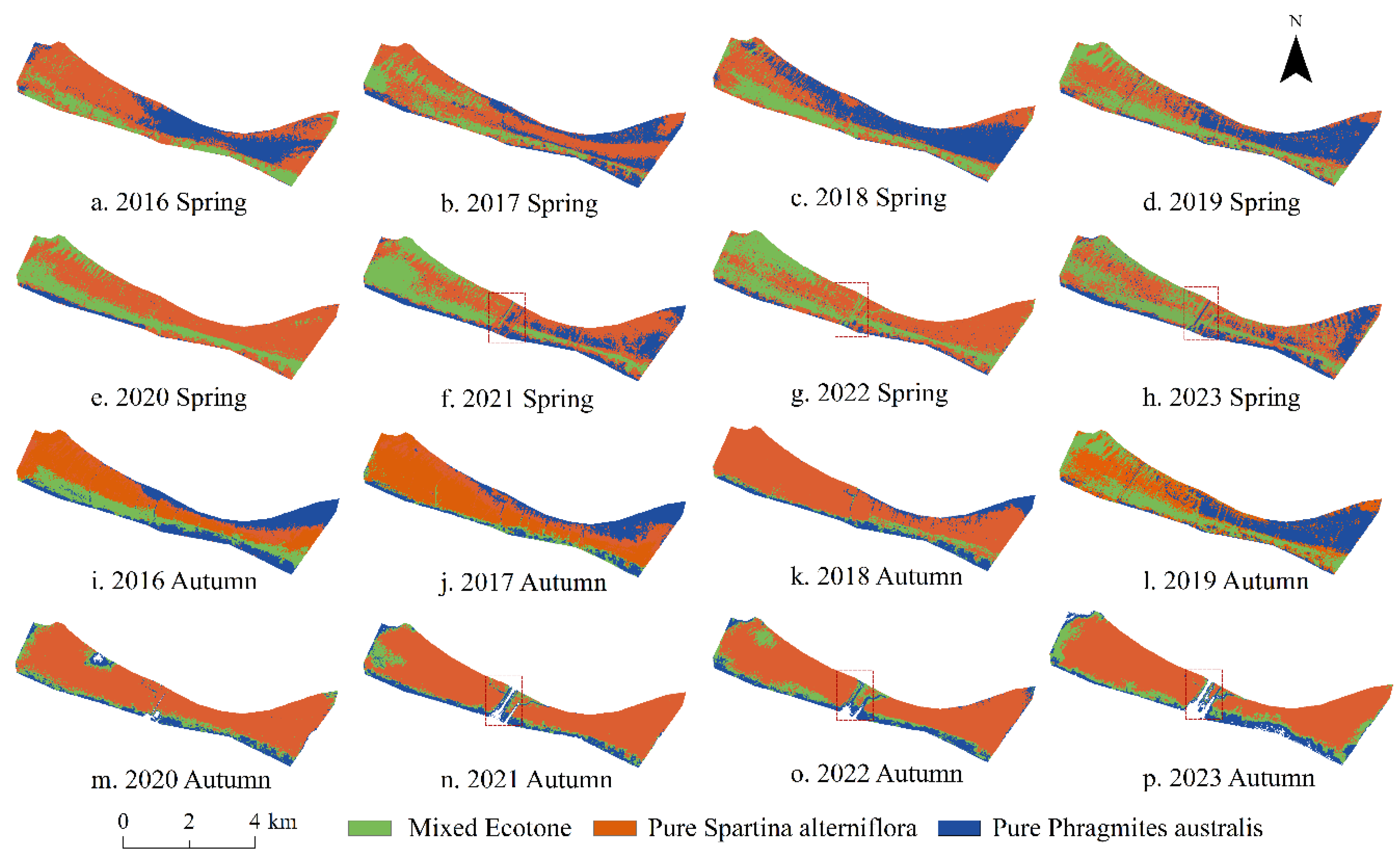
3.3. Spatial Distribution Pattern of the Mixed Ecotone
3.4. Spatiotemporal Dynamics of the Mixed Ecotone
3.4.1. Inter-Annual Variations (2016–2023)
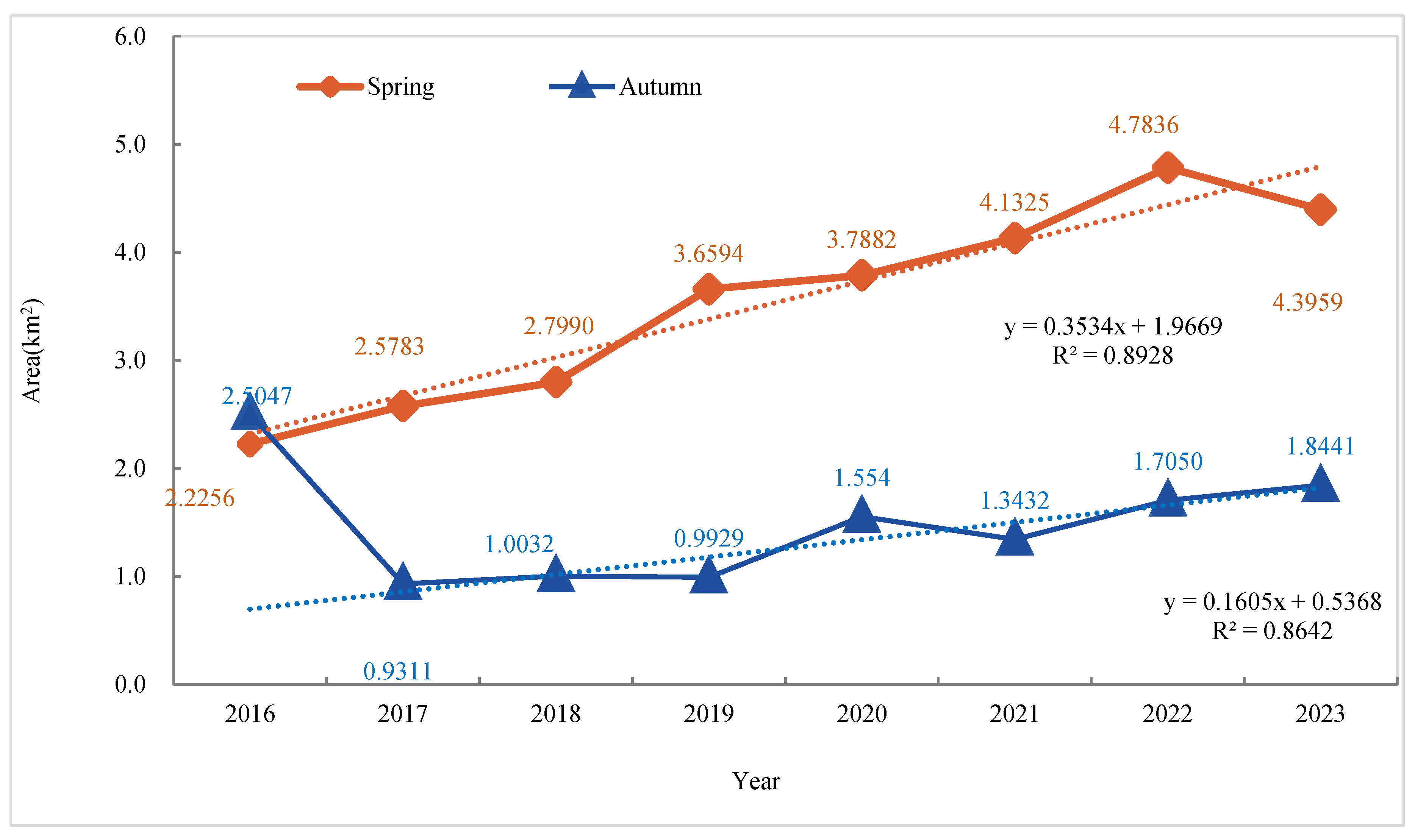
- Analysis of Area Change Trends.
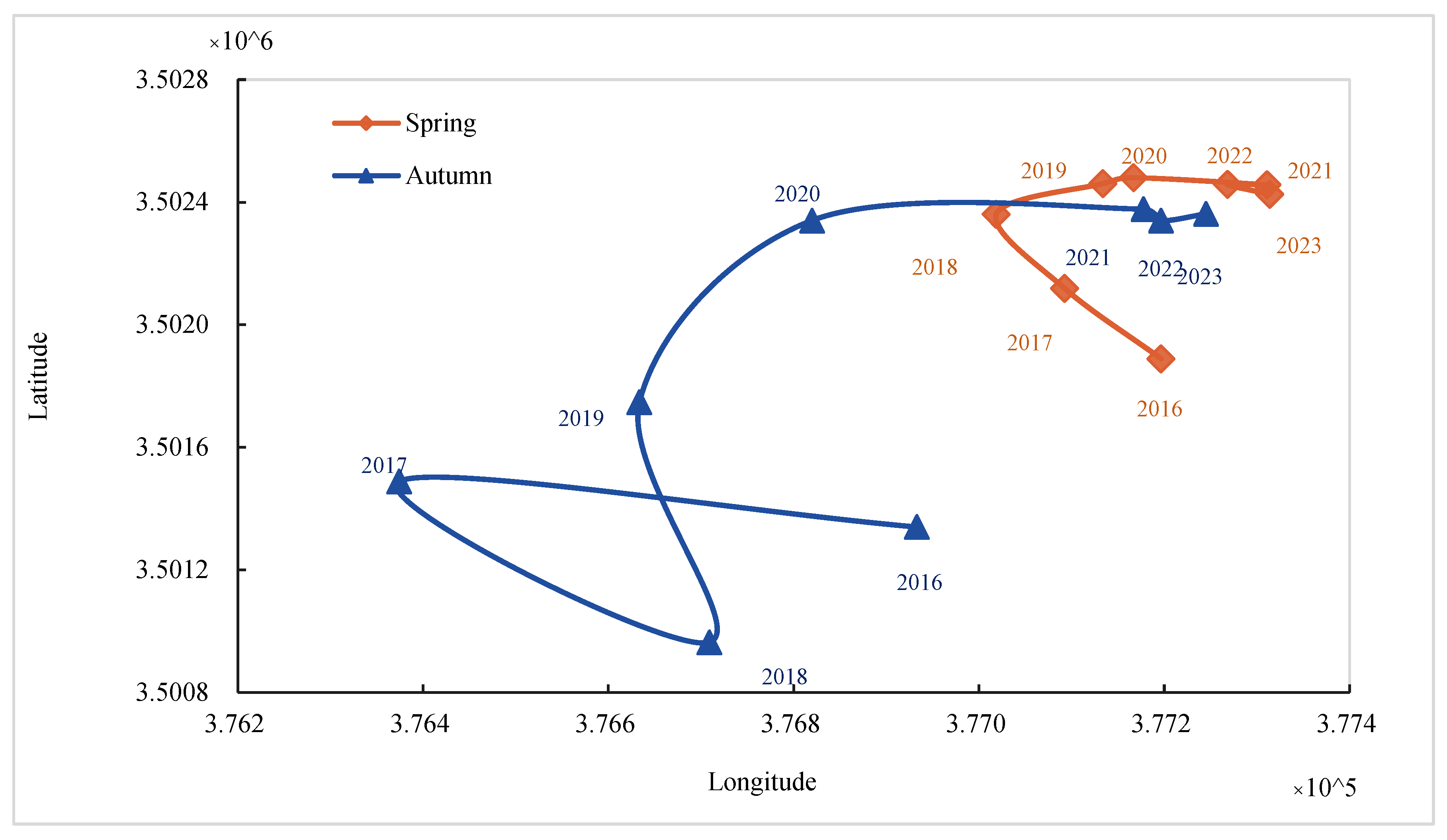
- Centroid Migration Trajectory and Spatial Expansion Pattern.
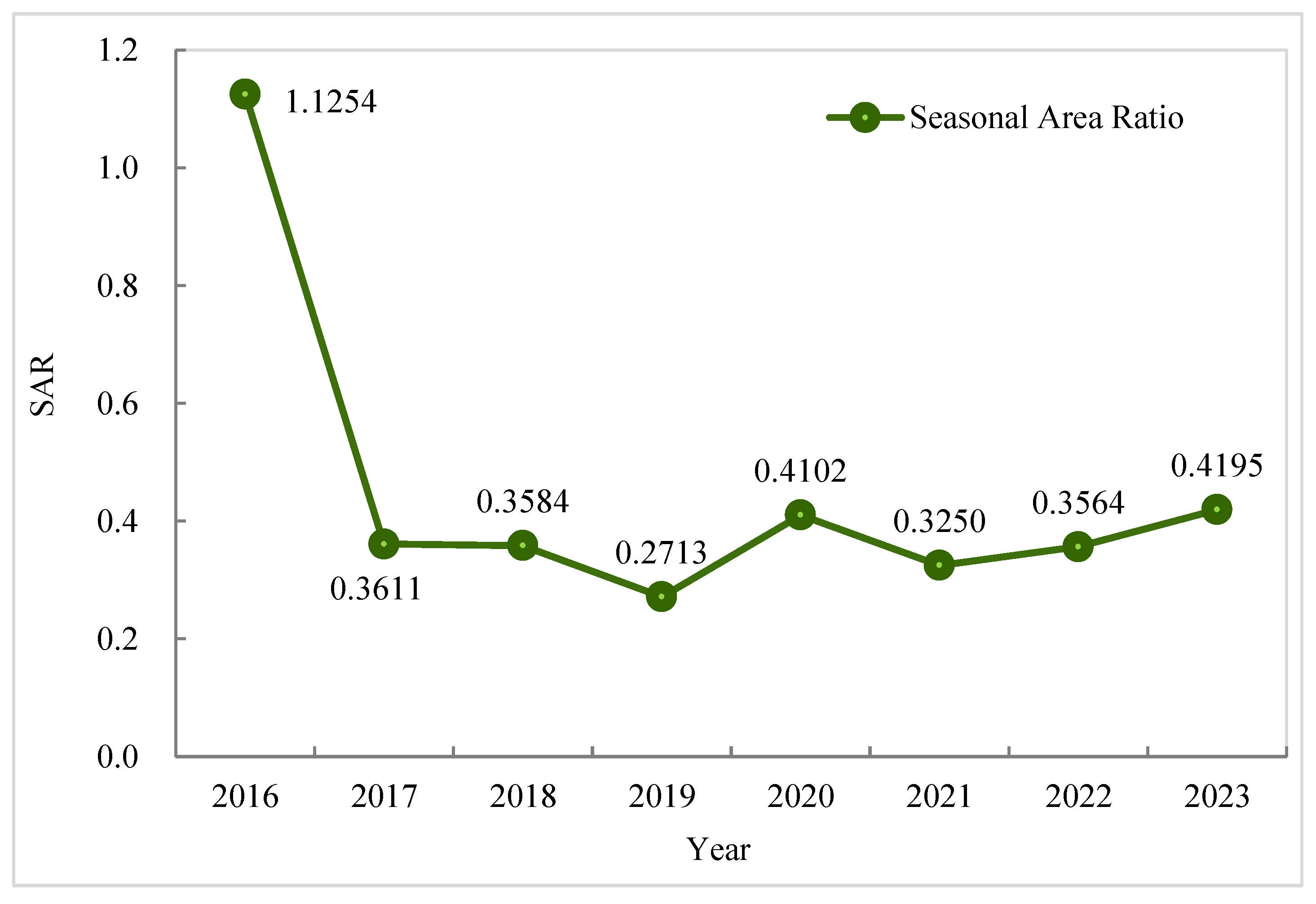
3.4.2. Intra-Annual Dynamics (Seasonal Variations)
4. Discussion
4.1. Methodological Innovations and Comparative Analysis
4.2. Ecological Implications and Mechanistic Insights
4.3. Management Implications and Recommendations
- Develop differentiated seasonal management strategies based on phenological asynchrony. This involves implementing precise control measures during S. alterniflora’s physiologically vulnerable periods (e.g., green-up and flowering/fruiting stages) [55], and conducting ecological restoration projects during the spring dominance phase of P. australis. Implementing these strategies significantly offers a synergistic benefit, enhancing management efficacy with minimal ecological disruption.
- Adopt spatially-differentiated management strategies. This requires accounting for micro-topographic and salinity gradients [56] to implement tailored strategies for the patchy northwestern ecotone, whereas the central region—affected by human activities—must undertake ecological engineering assessments and adaptive restoration planning to mitigate counter infrastructure impacts on vegetation patterns.
- Establish a dynamic monitoring and early-warning system. By integrating multi-source remote sensing data with ground observation networks [57], develop a model that captures spatial, phenological, and competitive dynamics to track the expansion of S. alterniflora and issue early risk alerts, thereby providing essential decision-support for coastal wetland conservation.
4.4. Management Implications and Recommendations
- Enhancement of multi-temporal validation and in-situ monitoring through integrated long-term plots and UAV-based hyperspectral observations for improved classification reliability and ecological process analysis.
- Employ spatially explicit models to decipher the interactive effects of anthropogenic and natural drivers to quantify the impact of human disturbances—particularly engineering infrastructure—on ecotone dynamics.
5. Conclusion
- A seasonally adaptive spectral index framework, incorporating a three-dimensional feature space, was developed to extract the mixed ecotone. Identification of the optimal spectral features—near-infrared reflectance (spring) and NDVI (autumn)—followed by adoption of the multi-threshold Otsu algorithm, enabled high-precision vegetation community classification. The validation results yielded high accuracy, with an overall accuracy of 87.3±1.4% and a Kappa coefficient of 0.84±0.02. The mixed ecotone was accurately delineated, achieving producer’s and user’s accuracies of 85.2% and 83.6%, respectively, which greatly enhances the identification capability and temporal stability for narrow transition zones.
- This study documented a distinct land-to-sea ecological sequence—pure P. australis–mixed ecotone–pure S. alterniflora—in the Chongming Dongtan wetland, with the vegetation arranged in an overall east–west belt. The northwestern sector exhibited a patchy distribution influenced by tidal creeks and micro-topography, related to salinity gradients and hydrological differentiation induced by elevation heterogeneity. Meanwhile, regular linear anomalies identified in the central area were attributable to engineering infrastructure, underscoring the spatial heterogeneity shaped by coupled natural-anthropogenic drivers.
- The mixed ecotone showed consistent seaward expansion from 2016 to 2023. During spring, it exhibited an average annual growth rate of 13.93%, with the centroid migrating steadily seaward at an azimuth of 112°. The migration rate showed a notable deceleration in later years, interspersed with periodic reversals, suggesting control by hydrological resistance or interspecific competition. Influenced by extreme climate events, the ecotone area exhibited marked fluctuations in autumn—most notably a 62.83% decline from 2016 to 2017. Paralleling this instability, the centroid migration path was similarly complex, unfolding in a three-phase sequence of “retreat–leap–expansion.” The spatiotemporal dynamics were shaped by the long-term drivers of sediment deposition and climate warming, with short-term modifiers including typhoons, salt stress, and hydrological disturbances.
- Analysis of the SAR index revealed a tipping point in 2017, when the seasonal competition pattern of the ecotone transitioned from a single year of autumn dominance to a subsequent seven-year phase of spring dominance. During the spring-dominated phase, the SAR values demonstrated a characteristic sawtooth-like fluctuation (0.27–0.42) with cyclical low-medium-high shifts, thereby indicating a dynamic equilibrium wherein P. australis establishes dominance through phenological advancement, whereas S. alterniflora ensures competitive resilience via niche penetration. The SAR index serves as a powerful tool for capturing spatiotemporal competition dynamics and deciphering the mechanisms driving wetland vegetation succession.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SAR | Seasonal Area Ratio |
| JM | Jeffries-Matusita |
| 3D | Three-Dimensional |
| S. alterniflora | Spartina alterniflora |
| P. australis | Phragmites australis |
| B. mariqueter | Bolboschoenoplectus mariqueter |
| NIR | Near-Infrared Reflectance |
| NDVI | Normalized Difference Vegetation Index |
| GIS | Geographic Information System |
| NLSD | National Land Survey Data |
| CLCD | China Land Cover Dataset |
| GF-1 | Gaofen-1satellite |
| UAV | Unmanned Aerial Vehicle |
| MSI | Multispectral Instrument |
| GPS | Global Positioning System |
| RTK | Real-Time Kinematic |
| UN | United Nations |
| SDG | Sustainable Development Goal |
References
- Zhu, W.Q.; Ren, G.B.; Wang, J.P.; Wang, J.B.; Hu, Y.B; Lin, Z.Y.; Li, W.; Zhao, Y.J.; Li, S.B.; Wang, N. Monitoring the Invasive Plant Spartina alterniflora in Jiangsu Coastal Wetland Using MRCNN and Long-Time Series Landsat Data. Remote Sens. 2022, 14, 2630. [Google Scholar] [CrossRef]
- Zhang, G.L.; Bai, J.H.; Tebbe, C.C.; Huang, L.B.; Jia, J.; Wang, W.; Wang, X.; Yu, L.; Zhao, Q.Q. Spartina alterniflora invasions reduce soil fungal diversity and simplify co-occurrence networks in a salt marsh ecosystem. Sci. Total Environ. 2021, 758, 143667. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Huang, Y.Y.; Du, X.L.; Li, Y.M.; Tian, J.T.; Chen, Q.; Huang, Y.H.; Lv, W.W.; Yang, Y.; Liu, Z.Q.; Zhao, Y.L. Assessment of macrobenthic community function and ecological quality after reclamation in the Changjiang (Yangtze) River Estuary wetland. Acta Oceanol. Sin. 2022, 41, 96–107. [Google Scholar] [CrossRef]
- Syed, S.; Xu, M.; Wang, Z.F.; Yu, C.F.; Lian, B. Invasive Spartina alterniflora in controlled cultivation: Environmental implications of converging future technologies. Ecol. Indic. 2021, 130, 108027. [Google Scholar] [CrossRef]
- Chen, W.J.; Shi, C. Fine-scale mapping of Spartina alterniflora-invaded mangrove forests with multi-temporal WorldView-Sentinel-2 data fusion. Remote Sens. Environ. 2023, 295, 113690. [Google Scholar] [CrossRef]
- Chang, D.; Wang, Z.Y.; Ning, X.G.; Li, Z.J.; Zhang, L.; Liu, X.T. Vegetation changes in Yellow River Delta wetlands from 2018 to 2020 using PIE-Engine and short time series Sentinel-2 images. Front. Mar. Sci. 2022, 9, 977050. [Google Scholar] [CrossRef]
- Döweler, F.; Fransson, J.E.S.; Bader, M.K.F. Linking High-Resolution UAV-Based Remote Sensing Data to Long-Term Vegetation Sampling—A Novel Workflow to Study Slow Ecotone Dynamics. Remote Sens. 2024, 16, 840. [Google Scholar] [CrossRef]
- Zhou, H.; Tang, M.D.; Huang, J.; Mei, X.X.; Zhao, H.J. Driving mechanisms and multi-scenario simulation of land use change based on National Land Survey Data: a case in Jianghan Plain, China. Front. Environ. Sci. 2024, 12, 1422987. [Google Scholar] [CrossRef]
- Shi, Z.Y.; Pu, W.P.; Dong, J.H. Spatiotemporal Changes of Land Use and Their Impacts on Ecosystem Service Value inthe Agro-pastoral Ecotone of Northern China. Environ. Sci. 2025, 46, 2373–2384. [Google Scholar]
- Yao, H.Y.; Liu, P.D.; Shi, R.H.; Zhang, C. Extracting the transitional zone of Spartina alterniflora and Phragmites australis in the wetland using high-resolution remotely sensed images. J. Geo-inf. Sci. 2017, 19, 1375–1381. [Google Scholar]
- Dong, D.; Huang, H.M.; Gao, Q.; Zhang, S.P.; Li, K.; Wei, Z.; Sun, Y.C. Combing satellite and UAV remote sensing to monitor a mangrove-saltmarsh ecotone: a case study from Dandou Sea in Guangxi. J. Appl. Oceanogr. 2025, 44, 167–176. [Google Scholar]
- Li, C.W.; Fang, S.B.; Geng, X.L.; Yuan, Y.; Zheng, X.W.; Zhang, D.; Li, R.X.; Sun, W.; Wang, X.R. Coastal ecosystem service in response to past and future land use and land cover change dynamics in the Yangtze river estuary. J. Clean. Prod. 2023, 385, 135601. [Google Scholar] [CrossRef]
- Mohammed Rabiul Alam, S.; Shawkat Hossain, M. Using a water index approach to mapping periodically inundated saltmarsh land-cover vegetation and eco-zonation using multi-temporal Landsat 8 imagery. J. Coast. Conserv. 2024, 28, 19. [Google Scholar] [CrossRef]
- Wittstruck, L.; Jarmer, T.; Waske, B. Multi-Stage Feature Fusion of Multispectral and SAR Satellite Images for Seasonal Crop-Type Mapping at Regional Scale Using an Adapted 3D U-Net Model. Remote Sens. 2024, 16, 3115. [Google Scholar] [CrossRef]
- Zhou, N.; Zhou, M.T.; Sui, H.G. DepthCD: Depth prompting in 2D remote sensing imagery change detection. ISPRS J. Photogramm. Remote Sens. 2025, 227, 145–169. [Google Scholar] [CrossRef]
- Liu, P.D.; Shi, R.H.; Meng, F.; Liu, J.T.; Yao, G.B.; Fu, P.J. Combining Multi-Indices by Neural Network Model for Estimating Canopy Chlorophyll Content: a Case Study of Interspecies Competition between Spartina alterniflora and Phragmites australis. Pol. J. Environ. Stud. 2022, 31, 199–217. [Google Scholar] [CrossRef]
- Najjar Khodabakhsh, Z.; Moradi, H.; Rahimzadeh-Bajgiran, P.; Pourmanafi, S.; Ahmadi, M. A 30-year phenological study of mangrove forests at the species level as a function of climatic drivers using multispectral remote sensing satellites. Ecol. Indic. 2025, 178, 114038. [Google Scholar] [CrossRef]
- Klehr, D.; Stoffels, J.; Hill, A.; Pham, V.D.; van der Linden, S.; Frantz, D. Mapping tree species fractions in temperate mixed forests using Sentinel-2 time series and synthetically mixed training data. Remote Sens. Environ. 2025, 323, 114740. [Google Scholar] [CrossRef]
- Lomeo, D.; Simis, S.G.H.; Selmes, N.; Jungblut, A.D.; Tebbs, E.J. Colour-informed ecoregion analysis highlights a satellite capability gap for spatially and temporally consistent freshwater cyanobacteria monitoring. ISPRS J. Photogramm. Remote Sens. 2025, 228, 323–339. [Google Scholar] [CrossRef]
- Kumbier, K.; Hughes, M.G.; Rogers, K.; Woodroffe, C.D. Inundation characteristics of mangrove and saltmarsh in micro-tidal estuaries. Estuar. Coast. Shelf Sci. 2021, 261, 107553. [Google Scholar] [CrossRef]
- Deijns, A.A.J.; Michéa, D.; Déprez, A.; Malet, J.P.; Kervyn, F.; Thiery, W.; Dewitte, O. A semi-supervised multi-temporal landslide and flash flood event detection methodology for unexplored regions using massive satellite image time series. ISPRS J. Photogramm. Remote Sens. 2024, 215, 400–418. [Google Scholar] [CrossRef]
- Lin, Y.X.; Hu, H.W.; Yang, P.; Ye, G.P. Spartina alterniflora invasion has a greater impact than non-native species, Phragmites australis and Kandelia obovata, on the bacterial community assemblages in an estuarine wetland. Sci. Total Environ. 2022, 822, 153517. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.T.; Yang, Z.Q.; Chen, C.P.; Tian, B. Tracking the environmental impacts of ecological engineering on coastal wetlands with numerical modeling and remote sensing. J. Environ. Manage. 2022, 302, 113957. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Hu, Z.; Grandjean, T.J.; Bing Wang, Z.; van Zelst, V.T.M.; Qi, L.; Xu, T.P.; Young Seo, J.; Bouma, T.J. Dynamics and drivers of tidal flat morphology in China. Nat. Commun. 2025, 16, 2153. [Google Scholar] [CrossRef]
- Cao, Y.; Dong, B.; Xu, H.F.; Xu, Z.L.; Wei, Z.Z.; Lu, Z.P.; Liu, X. Landscape ecological risk assessment of chongming dongtan wetland in shanghai from 1990 to 2020. Environ. Res. Commun. 2023, 5, 105016. [Google Scholar] [CrossRef]
- Chen, X.H.; Yang, Q.Q.; Chen, Z.Z.; Lei, J.R.; Wu, T.T.; Li, Y.L.; Pan, X.Y. The interaction between temperature and rainfall determines the probability of tropical forest fire occurrence in Hainan Island. Front. Vet. Sci. 2025, 8, 1495699. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, B. Field practice of Scirpus mariqueter restoration in the bird habitats of Chongming Dongtan Wetland, China. Chin. J. Appl. Ecol. 2023, 34, 2663–2671. [Google Scholar]
- Suwanprasit, C. ; Shahnawaz. Mapping burned areas in Thailand using Sentinel-2 imagery and OBIA techniques. Sci. Rep. 2024, 14, 9609. [Google Scholar]
- Zhang, J.Y.; Ning, Y.L.; Li, J.X.; Shi, Z.L.; Zhang, Q.Z.; Li, L.Q.; Kang, B.Y.; Du, Z.B.; Luo, J.C.; He, M.X.; Li, H.Y. Invasion stage and competition intensity co-drive reproductive strategies of native and invasive saltmarsh plants: Evidence from field data. Sci. Total Environ. 2024, 954, 176383. [Google Scholar] [CrossRef]
- Sun, C.; Li, J.L.; Liu, Y.X.; Liu, Y.C.; Liu, R.Q. Plant species classification in salt marshes using phenological parameters derived from Sentinel-2 pixel-differential time-series. Remote Sens. Environ. 2021, 256, 112320. [Google Scholar] [CrossRef]
- Zhao, Q.; Qu, Y.H. The Retrieval of Ground NDVI (Normalized Difference Vegetation Index) Data Consistent with Remote-Sensing Observations. Remote Sens. 2024, 16, 1212. [Google Scholar] [CrossRef]
- Zhang, Q.J.; Guo, Z.H.; Liu, L.; Mei, J.C.; Wang, L. Lithological classification using SDGSAT-1 TIS data and three-dimensional spectral feature space model. Int. J. Digit. Earth 2025, 18, 2467983. [Google Scholar] [CrossRef]
- Li, J.; Wu, Z.C.; Hu, Z.W.; Jian, C.L.; Luo, S.J.; Mou, L.C.; Zhu, X.X.; Molinier, M. A Lightweight Deep Learning-Based Cloud Detection Method for Sentinel-2A Imagery Fusing Multiscale Spectral and Spatial Features. IEEE Trans. Geosci. Remote Sens. 2021, 60, 1–19. [Google Scholar] [CrossRef]
- Turker, M.; Rahimzadeganasl, A. Agricultural Field Detection from Satellite Imagery Using the Combined Otsu’s Thresholding Algorithm and Marker-Controlled Watershed-Based Transform. J. Indian Soc. Remote Sens. 2021, 49, 1035–1050. [Google Scholar] [CrossRef]
- Kahaer, Y.; Shi, Q.D.; Shi, H.B.; Peng, L.; Abudureyimu, A.; Wan, Y.B.; Li, H.; Zhang, W.Q.; Yang, N.J. What Is the Effect of Quantitative Inversion of Photosynthetic Pigment Content in Populus euphratica Oliv. Individual Tree Canopy Based on Multispectral UAV Images? Forests 2022, 13, 542. [Google Scholar]
- Yang, X.Z.; Hong, L. A New Classification Rule-Set for Mapping Surface Water in Complex Topographical Regions Using Sentinel-2 Imagery. Water 2024, 16, 943. [Google Scholar] [CrossRef]
- Zhao, J.Q.; Wang, Z.X.; Zhang, Q.J.; Niu, Y.F.; Lu, Z.; Zhao, Z. A novel feature selection criterion for wetland mapping using GF-3 and Sentinel-2 Data. Ecol. Indic. 2025, 171, 113146. [Google Scholar] [CrossRef]
- Zhang, X.M.; Lin, X.F.; Fu, D.J.; Wang, Y.; Sun, S.B.; Wang, F.; Wang, C.P.; Xiao, Z.Y.; Shi, Y.Q. Comparison of the Applicability of J-M Distance Feature Selection Methods for Coastal Wetland Classification. Water 2023, 15, 2212. [Google Scholar] [CrossRef]
- Seo, D.M.; Woo, H.J.; Hong, W.H.; Seo, H.; Na, W.J. Optimization of Number of GCPs and Placement Strategy for UAV-Based Orthophoto Production. Appl. Sci. 2024, 14, 3163. [Google Scholar] [CrossRef]
- Xu, Q.; Jin, M.T.; Guo, P. A High-Precision Crop Classification Method Based on Time-Series UAV Images. Agriculture 2023, 13, 97. [Google Scholar] [CrossRef]
- Motiee, H.; Ahrari, S.; Motiee, S.; McBean, E. Assessment of Climate Change with Remote Sensing Data on Snow and Ice Cover in the Rocky Mountains Glaciers. J. Environ. Inf. 2025, 46, 544. [Google Scholar] [CrossRef]
- Hei Wong, C.; Li, Z.Y.; Hung Lam Yim, S.; Chang, T.Y.; Ling Man, C.; Ho Cheng, C.; Chi-Yui Kwok, T.; Fai Ho, K. The comparison between multiple linear regression and random forest model in predicting environmental noise and its frequency components level in Hong Kong using a land-use regression approach. Environ Res. 2025, 286, 122919. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.M.; Huang, Q.; Huang, S.Z.; Leng, G.Y.; Bai, Q.J.; Liang, H.; Wang, L.; Zhao, J.; Fang, W. Spatial-temporal dynamics of agricultural drought in the Loess Plateau under a changing environment: Characteristics and potential influencing factors. Agric. Water Manag. 2021, 244, 106540. [Google Scholar] [CrossRef]
- Qiu, T.Z.; Cong, N.; Rong, L.; Qi, G.; Du, L.; Ren, P.; Xiao, J.T. Spatiotemporal dynamics and driving mechanisms of alpine peatland wetlands in the eastern Qinghai–Tibet Plateau based on a Vision Transformer model. Ecol. Indic. 2025, 178, 114136. [Google Scholar] [CrossRef]
- Yan, S.Y.; Yang, G.H.; Li, Q.Y.; Zhang, B.; Wang, Y.; Zhang, Y.; Wang, C.H. Distance–Intensity Image Strategy for Pulsed LiDAR Based on the Double-Scale Intensity-Weighted Centroid Algorithm. Remote Sens. 2021, 13, 432. [Google Scholar] [CrossRef]
- Haight, J.D.; Hall, S.J.; Lewis, J.S. Landscape modification and species traits shape seasonal wildlife community dynamics within an arid metropolitan region. Landsc. Urban Plan. 2025, 259, 105346. [Google Scholar] [CrossRef]
- Wang, J.B.; Lin, Z.Y.; Ma, Y.Q.; Ren, G.B.; Xu, Z.J.; Song, X.K.; Ma, Y.; Wang, A.D.; Zhao, Y.J. Distribution and invasion of Spartina alterniflora within the Jiaozhou Bay monitored by remote sensing image. Acta Oceanol. Sin. 2022, 41, 31–40. [Google Scholar] [CrossRef]
- Yu, H.L.; Zhao, X.Y.; Huang, W.D.; Zhan, J.; He, Y.Z. Drought Stress Influences the Growth and Physiological Characteristics of Solanum rostratum Dunal Seedlings From Different Geographical Populations in China. Front. Plant Sci. 2021, 12, 733268. [Google Scholar] [CrossRef]
- Yin, S.; Liang, G.P.; Wang, C.K.; Zhou, Z.H. Asynchronous seasonal patterns of soil microorganisms and plants across biomes: A global synthesis. Soil Biol. Biochem. 2022, 175, 108859. [Google Scholar] [CrossRef]
- Lisner, A.; Segrestin, J.; Konečná, M.; Blažek, P.; Janíková, E.; Applová, M.; Švancárová, T.; Lepš, J. Why are plant communities stable? Disentangling the role of dominance, asynchrony and averaging effect following realistic species loss scenario. J. Ecol. 2024, 112, 1832–1841. [Google Scholar]
- Feng, J.; Grandjean, T.J.; Wu, X.R.; van de Koppel, J.; van der Wal, D. Long-term phenological shifts in coastal saltmarsh vegetation reveal complex responses to climate change. Ecol. Indic. 2025, 179, 114219. [Google Scholar] [CrossRef]
- Ai, J.Q.; Chen, L.J.; He, H.Q.; Han, X.X. Revealing the long-term impacts of plant invasion and reclamation on native saltmarsh vegetation in the Yangtze River estuary using multi-source time series remote sensing data. Ecol. Eng. 2024, 208, 107362. [Google Scholar] [CrossRef]
- Bobrowski, M.; Bechtel, B.; Böhner, J.; Oldeland, J.; Weidinger, J.; Schickhoff, U. Application of Thermal and Phenological Land Surface Parameters for Improving Ecological Niche Models of Betula utilis in the Himalayan Region. Remote Sens. 2018, 10, 814. [Google Scholar] [CrossRef]
- Wei, F.; Han, M.; Han, G.X.; Wang, M.; Tian, L.X.; Zhu, J.Q.; Kong, X.L. Reclamation-oriented spatiotemporal evolution of coastal wetland along Bohai Rim, China. Acta Oceanol. Sin. 2022, 41, 192–204. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, Q.; Zhang, H.; Zheng, Y.L.; Yuan, L.; Zhong, Q.C. Climate warming increases the invasiveness of the exotic Spartina alterniflora in a coastal salt marsh: Implications for invasion management. J. Environ. Manage. 2025, 380, 124765. [Google Scholar] [CrossRef]
- Wang, D.W.; Bai, J.H.; Wang, W.; Ma, X.; Guan, Y.N.; Gu, C.H.; Zhang, S.Y.; Lu, F. Micro-Topography Manipulations Facilitate Suaeda Salsa Marsh Restoration along the Lateral Gradient of a Tidal Creek. Wetlands 2020, 40, 1657–1666. [Google Scholar] [CrossRef]
- Ma, X.S.; Wang, Z.L.; Liu, Y. Ecological risk assessment of a coastal area using multi-source remote sensing images and in-situ sample data. Ecol. Indic. 2024, 158, 111470. [Google Scholar] [CrossRef]
- Han, Y.L.; Dong, Y.Y.; Zhu, Y.N. , Huang, W. J. Remote Sensing Inversion of Vegetation Parameters With IPROSAIL-Net. IEEE Trans. Geosci. Remote Sens. 2023, 62, 1–16. [Google Scholar]

| Year | Spring Image Date | Cloud Cover | Autumn Image Date | Cloud Cover |
|---|---|---|---|---|
| 2016 | 20160325 | 7.8% | 20161207 | 0.2% |
| 2017 | 20170429 | 6.9% | 20171115 | 3.1% |
| 2018 | 20180401 | 7.3% | 20181212 | 6.0% |
| 2019 | 20190406 | 0.6% | 20191115 | 0.0% |
| 2020 | 20200413 | 4.9% | 20201129 | 1.4% |
| 2021 | 20210418 | 4.6% | 20211114 | 0.0% |
| 2022 | 20220420 | 2.8% | 20221116 | 8.4% |
| 2023 | 20230428 | 6.5% | 20231124 | 8.7% |
| Season | Spectral Index | Threshold T1 | Threshold T2 | JM distance | ||
|---|---|---|---|---|---|---|
| P. australis–ecotone | P. australis–ecotone | P. australis–ecotone | ||||
| Spring | ρBLUE | 0.040600 | 0.094005 | 1.50 | 1.50 | 1.50 |
| ρGREEN | 0.066416 | 0.123075 | 1.60 | 1.60 | 1.60 | |
| ρRED | 0.075696 | 0.129511 | 1.70 | 1.70 | 1.70 | |
| ρNIR | 0.149945 | 0.190183 | 1.99 | 1.99 | 1.99 | |
| NDVI | 0.174555 | 0.282613 | 1.86 | 1.86 | 1.86 | |
| Autumn | ρBLUE | 0.066783 | 0.104831 | 1.40 | 1.40 | 1.40 |
| ρGREEN | 0.043947 | 0.074789 | 1.50 | 1.50 | 1.50 | |
| ρRED | 0.072376 | 0.119112 | 1.60 | 1.60 | 1.60 | |
| ρNIR | 0.181992 | 0.258286 | 1.80 | 1.80 | 1.80 | |
| NDVI | 0.311753 | 0.513176 | 1.93 | 1.93 | 1.93 | |
| Year | Season | Overall Accuracy (%) |
Kappa Coefficient (%) |
Mixed ecotone | |
|---|---|---|---|---|---|
| Producer’s accuracy (%) | User’s accuracy (%) | ||||
| 2016 | Spring | 85.5 | 0.82 | 83.8 | 82.0 |
| Autumn | 86.0 | 0.83 | 84.5 | 82.8 | |
| 2017 | Spring | 86.8 | 0.83 | 85.0 | 83.5 |
| Autumn | 85.2 | 0.81 | 82.5 | 80.9 | |
| 2018 | Spring | 87.2 | 0.84 | 85.5 | 84.0 |
| Autumn | 87.8 | 0.85 | 86.0 | 84.5 | |
| 2019 | Spring | 88.0 | 0.85 | 86.3 | 85.0 |
| Autumn | 87.0 | 0.84 | 85.0 | 83.2 | |
| 2020 | Spring | 88.5 | 0.86 | 87.0 | 85.8 |
| Autumn | 87.3 | 0.84 | 85.2 | 83.5 | |
| 2021 | Spring | 88.9 | 0.86 | 87.5 | 86.2 |
| Autumn | 88.2 | 0.85 | 86.3 | 84.8 | |
| 2022 | Spring | 89.2 | 0.87 | 88.0 | 86.5 |
| Autumn | 88.5 | 0.86 | 86.8 | 85.0 | |
| 2023 | Spring | 89.5 | 0.86 | 86.2 | 85.0 |
| Autumn | 88.7 | 0.87 | 85.8 | 84.5 | |
| Average ± StdDev (%) | 87.3±1.4 | 0.84±0.02 | 85.2±1.5 | 83.6±1.7 | |
| Year | Spring Area | Annual Spring Change Rate | Autumn Area | Annual Autumn Change Rate |
|---|---|---|---|---|
| 2016 | 2.2256 | — | 2.5047 | — |
| 2017 | 2.5783 | 15.85 | 0.9311 | -62.83 |
| 2018 | 2.7990 | 8.56 | 1.0032 | 7.74 |
| 2019 | 3.6594 | 30.74 | 0.9929 | -1.03 |
| 2020 | 3.7882 | 3.52 | 1.5540 | 56.51 |
| 2021 | 4.1325 | 9.09 | 1.3432 | -13.56 |
| 2022 | 4.7836 | 15.76 | 1.7050 | 26.94 |
| 2023 | 4.3959 | -8.10 | 1.8441 | 8.16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



