
Submitted:
28 November 2025
Posted:
03 December 2025
You are already at the latest version
Abstract
This review examines convergent neurobiological mechanisms linking stress and drugs that drives stress-induced drug-related behaviors. It first outlines main theoretical frameworks explaining substance use disorders (SUDs), emphasizing vulnerability factors—particularly stressful life events—that increase addiction risk. The analysis integrates preclinical evidence demonstrating that chronic stress facilitates cross-sensitization to psychostimulants and accelerates drug self-administration, underscoring how stress and drugs converge on glutamatergic and dopaminergic transmission within the Nucleus Accumbens (NAc). Special attention is given to the glial cells, particularly microglia and astrocytes, in mediating stress-induced neuroimmune activation and glutamate dysregulation in the NAc. Three major themes related to microglia–astrocyte crosstalk are addressed: (i) the contribution of these glial cells to neuroimmune and glutamatergic alterations induced by stress; (ii) their role in synaptic and structural plasticity changes within the NAc; and (iii) the mechanisms by which stress and drug exposure reshape glial–neuronal communication, driving the comorbidity between stress and SUDs. A dedicated section focuses on key neuroimmune signaling pathways—particularly the TNF-α/NF-κB axis—and their involvement in stress-induced vulnerability to cocaine addiction. Finally, the review discusses preclinical evidence supporting the therapeutic potential of repurposed glutamate-modulating agents, as promising pharmacological candidates for treating comorbid stress and cocaine use disorder.
Keywords:
microglia
; astrocytes
; glutamate
; nucleus accumbens
; neuroimmune signaling
; cocaine
; substance use disorders
; stress
1. Introduction: A Neurobiological Framework for the Comorbidity of Stress and Substance Use Disorders
Stress is a physiological and psychological state that arises when environmental demands are perceived to exceed an individual’s capacity to cope. While the stress response is inherently adaptive, enabling the organism to manage acute challenges effectively, its chronic or unpredictable activation imposes significant bioenergetics and neurobiological costs. Prolonged engagement of the stress response can shift the brain from a state of adaptive regulation to a maladaptive and pathophysiological condition, thereby increasing vulnerability to psychiatric disorders, including substance use disorders (SUDs) [1,2].
SUDs are severe neuropathological conditions representing the culmination of a series of neurobehavioral events that define the addictive process. This trajectory involves a progression from voluntary, controlled drug use to a gradual loss of control, culminating in compulsive drug-seeking behavior. A defining feature of addiction is the persistent vulnerability to relapse, even after extended periods of abstinence [3,4]. This enduring, relapsing nature reflects long-lasting neurobiological alterations within the reward circuit, often accompanied by cognitive impairments, motivational disturbances, and physiological adaptations that drive continued drug use despite adverse consequences [5]. From a diagnostic perspective, the term addiction is now encompassed within the broader construct of SUDs. In 2013, the DSM-5 unified the previously distinct diagnoses of substance abuse and substance dependence into a single continuum, defining SUDs as ranging from mild to severe based on the number of diagnostic criteria met.
Clinical evidence indicates that stressful life events are critical vulnerability factors for the development of SUDs, with a high degree of comorbidity observed between drug use and stressful or traumatic experiences [6]. Between 50% and 65% of individuals with post-traumatic stress disorders (PTSD) also suffer from SUDs, prevalence three to five times higher than in the general population [7,8]. Traumatic experiences are not only major risk factors for initiating SUDs but also potent triggers for relapse [9]. Preclinical studies using drug self-administration models have demonstrated that stress exposure facilitates drug-taking behavior [10,11]. Moreover, reinstatement models have demonstrated that stress, drug exposure, and conditioned cues can each independently trigger the reinstatement of heroin- or cocaine-seeking behavior in animals previously trained and subjected to extinction procedures [12,13]. These findings suggest that stress-, drug-, and cue-related stimuli converge on shared neural circuits that drive drug-seeking behavior.
An expanding body of clinical and experimental evidence further demonstrates that neuroadaptations triggered by chronic, uncontrollable stress, especially traumatic stress capable of inducing PTSD, and repeated drug exposure converge within the mesocorticolimbic system, a core component of the addiction circuitry. These shared pathological mechanisms reflect the profound impact of both stress and drugs on neural systems governing reward, emotional regulation, and executive control [14,15,16]. A hallmark of these alterations is the disruption of top-down control exerted by the prefrontal cortex over limbic and striatal regions. Dysfunction within this circuitry contributes to maladaptive behavioral phenotypes, including heightened impulsivity, emotional dysregulation, and compulsive drug-seeking, core features of both stress-related disorders and addiction [17,18].
Importantly, chronic stress is known to induce profound structural and functional changes in astrocytes and microglia [19]. While traditionally viewed as support cells, it is now evident that astrocytes and microglia actively regulate neuronal function by modulating glutamatergic neurotransmission, synaptic plasticity, and neuroimmune signaling [20]. Both chronic stress and exposure to addictive drugs produce overlapping alterations in glial activity, indicating that glial dysregulation may represent a central mechanism underlying the dual pathology of stress-related disorders and SUDs [21]. Of particular interest is the emerging evidence of astrocyte–microglia cross-talk mediated by cytokines such as interleukin 1-beta (IL-1β), Tumor necrosis factor-alpha (TNF-α), and complement protein C1q, which can reshape neuronal circuits under both physiological and pathological conditions, including those induced by chronic stress and/or drug exposure [22,23,24].
Historically, addiction research has focused predominantly on neuronal adaptations [25,26,27], often neglecting the critical role of glial cells. However, it is increasingly recognized that glial-mediated dysregulation of glutamate homeostasis, neuroinflammation, and maladaptive synaptic remodeling within the mesocorticolimbic system plays a central role in the molecular and cellular processes underlying stress-induced drug-seeking, drug-taking, and relapse behaviors [21]. Integrating glial mechanisms into the neurobiological framework of stress-SUDs comorbidity is thus essential for advancing our understanding of these overlapping psychiatric and addictive conditions and for developing novel therapeutic strategies that specifically target glial dysfunction.
This review focuses primarily on psychostimulants, particularly cocaine, as they represent the most extensively studied class of drugs in preclinical models addressing stress-induced glutamatergic and neuroimmune alterations. Central to this discussion is the Nucleus Accumbens (NAc), a key hub within the motivation circuitry, where converging neurobiological and immune-related adaptations emerge in response to both stress and drug exposure. Throughout the review, we adopt a mechanistic perspective to examine stress–drug comorbidity, with particular emphasis on the proactive effects of stress in promoting drug-related behaviors. Specifically, we highlight its role in facilitating the acquisition of drug self-administration and in enhancing sensitivity to the psychostimulant effects of cocaine through cross-sensitization. This focus is guided by the depth and consistency of available preclinical evidence in these areas. Accordingly, although models investigating stress-induced reinstatement of drug-seeking behavior are not explored in detail, they are referenced when relevant findings contribute to the broader conceptual framework.
2. Stress as Vulnerability Factor in Substance Use Disorders: Theoretical Approaches and Interacting Risk Factors
2.1. Vulnerability Factors
The development of SUDs results from a complex interaction between multiple vulnerability factors that predispose individuals to compulsive drug use. Evidence from preclinical, clinical, and epidemiological studies has identified a range of risk domains, including genetic predisposition, neurobiological traits, psychological characteristics, comorbid psychiatric disorders, and environmental, developmental, and social factors that shape an individual’s susceptibility to addiction [15,28,29,30,31]. Among these, environmental stressors, particularly exposure to stressful life events, have been consistently identified as major contributors to increased risk of drug use and the development of SUDs in humans [9,32,33,34,35,36,37,38]. Stress not only increases the probability of initial drug use but also exacerbates the transition to compulsive use and heightens the risk of relapse. Moreover, the persistent vulnerability to relapse even after extended periods of abstinence suggests that addiction is maintained not only by neuroadaptations resulting from repeated drug exposure but also by enduring interactions between genetic factors, stress responsiveness, and environmental drug-associated cues [3,39]. These findings underscore the importance of considering stress not simply as a trigger but as a core component in the development and maintenance of SUDs.
2.2. Theoretical Approaches
A central issue in addiction neuroscience is understanding why, despite the widespread use of psychoactive substances, only 10-15% of individuals develop a compulsive, addiction-like pattern of drug use [40]. This observation has prompted the development of two major theoretical perspectives: the drug-centered and individual-centered models of addiction. The drug-centered perspective posits that addiction arises primarily as a consequence of the drug’s neurobiological impact. According to this view, repeated drug exposure induces long-lasting alterations in brain function, such as tolerance, dependence, behavioral sensitization, conditioned reinforcement, and withdrawal, which collectively shift the individual toward compulsive drug-seeking and drug-taking behavior [25,41,42]. These neuroadaptations are proposed to underlie not only the initiation and escalation of drug use, but also the persistence of addiction and the high probability of relapse after abstinence [26,43,44].
In contrast, the individual-centered perspective emphasizes the role of biological predispositions—shaped by genetic, developmental, and environmental factors, such as stress exposure—that interact to determine an individual’s susceptibility to addiction [45,46]. This model is supported by extensive preclinical data showing individual differences in vulnerability to psychostimulant self-administration, including strain-specific differences in drug-seeking behavior. These behavioral differences have been correlated with drug intake during the acquisition phase, suggesting a trait-like predisposition to compulsive use.
2.3. Emergent Integrative Perspective of Substance Use Disorders
Beyond the direct reinforcing properties of drugs, it is well established that chronic drug use leads to long-lasting dysregulation of the brain’s reward system, resulting in the emergence of an anti-reward state [47]. This state is characterized by heightened stress responsiveness and a diminished capacity to experience reward, which together promote negative emotional states that drive continued drug use and relapse. In this framework, stress plays a central role at all stages of the addiction cycle, from initiation to maintenance and relapse. However, despite its importance, the molecular and neurophysiological mechanisms that mediate the interaction between stress and addiction, particularly in the context of comorbid conditions such as PTSD and SUDs, are not yet fully understood. A growing body of research is now dedicated to clarifying how stress-related molecular pathways contribute to the maladaptive reorganization of reward circuits, and how genetic and epigenetic factors modulate this vulnerability [48,49].
Emerging evidence indicates that life experiences, including exposure to chronic stress, can produce long-lasting changes in the brain’s epigenetic architecture, thereby altering gene expression patterns that influence vulnerability to addiction. These epigenetic modifications may sensitize neural circuits to drug exposure, facilitating the transition from voluntary use to compulsive drug-seeking behavior [50]. Recent transcriptomic studies have begun to identify the molecular underpinnings of this process. Browne et al. (2023) [51] mapped heroin-induced transcriptional changes across the brain’s reward circuitry, uncovering gene networks linked to drug intake, seeking, and relapse. Similarly, Mews et al. (2023)[48] characterized cocaine-related transcriptional alterations, revealing both overlapping and distinct molecular profiles compared to opioid use disorder. Importantly, the gene expression signatures identified in both studies show strong correspondence with human data, reinforcing the translational relevance of preclinical models and suggesting common molecular targets for future interventions.
Taken together, these findings support the view that SUDs are complex, multifactorial conditions that cannot be understood solely as consequences of chronic drug exposure. Instead, they emerge from a dynamic interplay between genetic predispositions, phenotypic traits, environmental stressors–particularly chronic or repeated stress experienced in daily life– and drug-induced neuroplasticity [3]. From this perspective, stress emerges as a pivotal factor that interacts with individual genetic background to shape early drug responses. These initial responses may trigger neuroadaptive processes that, when sustained by ongoing psychological and environmental pressures, significantly increase the risk of developing compulsive drug use. Although a detailed analysis of genetic and epigenetic contributions lies beyond the scope of this review, it is important to acknowledge that gene–environment interactions likely contribute to individual vulnerability and warrant further investigation. Here, our primary objective is to examine the enduring neuroadaptations through which stress modulates drug action within the mesolimbic reward circuitry, while situating genetic factors within a broader conceptual framework. This integrative perspective may help explain why only a subset of individuals transitions from drug use to addiction and underscores the need for personalized prevention and treatment strategies that consider both neurobiological and environmental determinants of addiction risk.
3. Motivational Neural Circuits Implicated in Stress and Substance Use Disorders Vulnerability
From a neurobiological perspective, mesencephalic dopaminergic neurons projecting from the Ventral tegmental area (VTA) to the NAc and various cortical regions constitute a key substrate modulated by stress-related hormones, such as corticotropin-releasing factor (CRF) and glucocorticoids. During stressful experiences, these hormones are released and potently enhance dopaminergic transmission [46,52]. Activation of this mesocorticolimbic pathway (Figure 1) mediates the reinforcing properties of drugs of abuse [53]. Consequently, stress-induced alterations in this system can facilitate the initiation and maintenance of drug self-administration [46,54,55]. At the molecular level, exposure to various stressors or repeated drug administration activates both glutamatergic and dopaminergic systems, triggering intracellular signaling cascades that alter gene expression within motivational circuits [56,57]. These transcriptional changes promote long-lasting neurochemical and structural adaptations underlying neural sensitization, a process that manifests behaviorally as psychomotor sensitization [58,59]. Notably, psychomotor sensitization is tightly linked to enhanced drug self-administration and relapse vulnerability [59,60].
As noted by Pierce and Kalivas (1997) [56], and by Scofield et al. (2016) [61] more recently, the NAc receives convergent glutamatergic inputs from cortical, allocortical, thalamic, midbrain, and brainstem regions, and projects to basal ganglia structures including the ventral pallidum and substantia nigra pars reticulata, as well as to mesencephalic, hypothalamic, and extended amygdala areas. Afferent projections often innervate both the core and shell of the NAc along defined topographic gradients (e.g., dorsoventral hippocampal projections terminate from lateral to medial regions of the NAc). Within this circuit, dopaminergic, glutamatergic, GABAergic, endocannabinoid, and opioid signaling systems interact in complex and dynamic ways. In particular, the mesocorticolimbic and nigrostriatal dopamine (DA) pathways underlie both voluntary motor function and motivated behaviors, and they are co-opted during drug exposure to drive compulsive drug seeking and consumption. Further, Nall et al., (2021) [62] has pointed out that distinct circuit motifs have been implicated in specific aspects of drug-seeking behavior, including: (i) an Action/Reward subcircuit comprising the NAc, ventral pallidum, and VTA; (ii) a Prefrontal subcircuit involving the prelimbic, infralimbic, and insular cortices; (iii) a stress subcircuit encompassing the central amygdala and bed nucleus of the stria terminalis; and (iv) a Diencephalic subcircuit involving the lateral hypothalamus. While the precise contributions of the NAc shell, insular cortex, and ventral pallidum remain partially unresolved, extensive evidence across species supports a critical role for these structures in mediating both drug- and natural-reward seeking.
The NAc itself—a key target of both stress and drugs of abuse—is subdivided into two functionally distinct compartments: the core and the shell [63,64], with differential anatomical afferents and efferent projections [65], which may account for different aspects of the drug rewarding process [66]. The shell plays a critical role in assigning salience to motivationally relevant stimuli and is particularly involved in the formation of stimulus-reward associations, especially during early phases of drug conditioning [67,68]. In contrast, the core is essential for sensorimotor integration [69] and supports the expression of learned behaviors elicited by cues predicting salient events [64,70]. It is also a central node for the long-term expression of drug- and stress-induced sensitization and for cue-triggered drug-seeking behavior [66,71,72]. In this context, glutamatergic projections from the prelimbic prefrontal cortex to the NAc core are critical for the enduring control of cocaine seeking and reinstatement [73].
Physiologically, the NAc integrates pharmacological and environmental stimuli to generate adaptive or maladaptive behavioral responses [56,74,75]. Natural rewards such as food, sex, and social interaction increase DA levels in the NAc, eliciting hedonic states that reinforce survival-related behaviors. Drugs of abuse, however, usurp this system, inducing supraphysiological DA release that is both more intense and more sustained than that produced by natural reinforcers, but without conferring adaptive value [76,77,78]. Despite differing pharmacological targets, chronic exposure to addictive substances—including alcohol, nicotine, opioids, cannabinoids, and psychostimulants—converges on the dysregulation of the mesocorticolimbic system, resulting in persistently elevated extracellular DA levels in the NAc [79,80]. This dopaminergic hyperactivity contributes to both the heightened reinforcing properties of drugs and the exaggerated locomotor responses characteristic of psychostimulant exposure [81].
These interconnected systems form the neurobiological foundation of addiction. Their modulation by stress underscores the critical importance of understanding how environmental factors interact with neural circuits to promote vulnerability to SUDs.
4. Neurobiological Mechanisms Linking Stress and Psychostimulants: Dopaminergic and Glutamatergic Interactions in the Nucleus Accumbens
Although the precise mechanisms by which stress promotes SUDs remain unclear [14,82], convergent preclinical evidence demonstrates that chronic stress reliably increases vulnerability to psychostimulant self-administration in rodents (see Box 1) [83]. Specifically, exposure to stressors such as intermittent social defeat or restraint (acute or chronic) leads to long-lasting neuroadaptations—including locomotor and dopaminergic cross-sensitization—which result in escalated cocaine intake during extended-access sessions [84,85,86,87]. These findings are consistent with both clinical and preclinical observations indicating that certain forms of stress can promote the initiation, escalation, and/or relapse of drug use [30,88,89,90].
CRF and glucocorticoids (corticosterone in rodents) orchestrate many of the hormonal interactions between stress and drugs of abuse. Acting through CRFR1 and CRFR2, CRF is a major driver of stress-evoked drug-seeking and relapse [11,13,88,91,92,93,94]. Although CRF’s canonical role is to activate the hypothalamic-pituitary-adrenal axis [95], it also modulates extrahypothalamic nuclei—including the amygdala, bed nucleus of the stria terminalis and VTA—that govern motivation and reward [96,97]. The VTA serves as a major site of convergence where CRF and psychostimulants synergistically enhance DA release within mesolimbic and cortical circuits [98,99]. Dopaminergic neurons in the VTA, projecting to the NAc, prefrontal cortex, and amygdala, are pivotal for mediating both the acute reinforcing effects of drugs of abuse and the long-term neuroadaptations underlying addiction [100,101]. Within this circuitry, CRF signaling in the VTA promotes neuroadaptive alterations in dopaminergic activity through CRFR1 activation [102,103]. However, studies directly assessing the role of VTA CRF in regulating mesocortical DA output have yielded mixed results [104]. Strengthening this causal link, Refojo et al. (2011) [105] demonstrated that selective deletion of CRFR1 from VTA DA neurons markedly attenuated stress-induced elevations in prefrontal DA, underscoring the contribution of CRF-dependent mechanisms to stress-driven dopaminergic adaptations that enhance addiction vulnerability.
In parallel, foundational rodent studies demonstrated behavioral cross-sensitization between stress and amphetamine exposure [106]. Subsequent research further revealed that individual differences in stress reactivity—such as novelty-induced locomotion—and prior drug experience synergistically increase vulnerability to the reinforcing effects of psychostimulants, including amphetamines and cocaine [107,108]. Importantly, corticosterone—the primary glucocorticoid in rodents—also emerges as a key hormonal mediator of this interaction. While sustained corticosterone signaling following stress enhances psychostimulant intake, adrenalectomy or physiological corticosterone replacement significantly attenuates the stress-induced potentiation of drug use [109,110,111]. Furthermore, a variety of stressors—including electric foot shock, restraint, social defeat, and food deprivation—promote cocaine self-administration, often through elevations in corticosterone [112,113].
The rewarding properties of these drugs, assessed via conditioned place preference [114,115,116], and their reinforcing efficacy in self-administration paradigms [46,117], are likewise amplified in stress–drug cross-sensitization paradigms. Social defeat stress similarly sensitizes animals to cocaine and amphetamine, promoting binge-like intake patterns [118,119], while environmental factors such as social housing conditions modulate the magnitude of stress-evoked drug seeking in mice [120]. Notably, when stress is experienced within the drug-taking context—for example, during intra-session electric footshock—animals show an escalation of cocaine intake that mirrors the excessive consumption observed in extended-access models of addiction [121,122].
Likewise to observed following stress, repeated, non-contingent administration of cocaine, amphetamine, nicotine, ethanol, morphine, or Δ⁹-tetrahydrocannabinol (THC) reliably induces locomotor sensitization in rodents [123,124,125,126,127,128]. Although a single drug injection can elicit a transient increase in locomotor activity [129,130], stable, long-term sensitization generally requires repeated exposure. Likewise, acute or chronic stress exposure can cross-sensitize organisms to the psychomotor-activating and reinforcing effects of drugs of abuse. Social defeat, food restriction, foot-shock, prenatal stress, and restraint stress each enhance locomotor cross-sensitization to cocaine [11,13,86,131,132] and to amphetamine [133,134,135]. Crucially, repeated exposure to these stressors does not diminish but rather consolidates the sensitized response, underscoring the pathological significance of chronic, everyday stress. These findings point to overlapping neural substrates engaged by both stress and psychostimulant exposure. Consistent with this view, various stressors—foot-shock [136], restraint [52,133], and prenatal stress [137]—activate mesolimbic DA transmission and heighten locomotor responses to cocaine and amphetamine. Nevertheless, although stress and psychostimulants produce parallel changes in behavior and dopaminergic signaling, a direct causal link between these two domains has yet to be fully established [56,128,138,139].
Glutamatergic afferents tightly control VTA DA neurons and mediate both rapid and long-term adaptations [140]. In cocaine-experienced animals, CRF magnifies excitatory transmission onto these neurons [141]. In vitro, CRFR2 activation strengthens NMDA currents, underscoring glutamate as a common substrate for stress and psychostimulant action [142]. CRF-dependent amplification of AMPA drive further escalates mesocorticolimbic DA output [143]. Consistent with this, acute stress and diverse addictive substances—including cocaine, amphetamine, morphine, ethanol, and nicotine—increase the AMPA/NMDA ratio in VTA DA neurons [102]. Interestingly, direct CRF infusion into the VTA produces enduring synaptic plasticity via CRFR1, potentiating NMDA-receptor currents, up-regulating AMPA-receptor function, and elevating the AMPA/NMDA ratio after stress or drug exposure [102,103,142,143]. At the same time, CRF diminishes GABA-B and D2 receptor signaling, further boosting DA-neuron excitability [144].
Interestingly, stress not only increases extracellular glutamate concentrations in the VTA by enhancing synaptic glutamate release, but also exerts similar effects in other brain regions, including the medial prefrontal cortex, striatum, and NAc [145]. In parallel, both acute and chronic administrations of psychostimulants, such as cocaine and amphetamine, have been shown to elevate glutamate release in the VTA and NAc [146,147,148]. These effects are largely mediated by glutamatergic projections from the prefrontal cortex and amygdala to mesolimbic structures [149,150,151]. One of the key mechanistic implications of this corticolimbic glutamatergic activation is its ability to modulate the mesolimbic DA system, a process fundamental to the neurobiology of addiction [145,152]. Although the NAc is often treated as a single entity, accumulating evidence highlights functional divergence between its core and shell subregions in shaping behavioral and neurochemical responses to stress and drugs [66,71,72]. Pacchioni et al. (2007) [134] showed that a single restraint session potentiates amphetamine-evoked DA release in both compartments at 24 h, yet this effect endures for at least eight days only in the core, waning over time in the shell. Extending this dissociation, García-Keller et al. (2013) [132] found that acute restraint stress produces cross-sensitization to cocaine exclusively within the NAc core, where elevated dopaminergic responses persist up to 21 days post-stress. Chronic restraint stress produces a similar pattern: Avalos et al. (2022) [153] reported a sustained, stress-induced enhancement of cocaine-evoked DA release that is restricted to the NAc core, confirming a persistent core–shell asymmetry. This asymmetry is also reflected in structural remodeling, with an increase in mushroom spine density observed in the NAc core but not in the shell, following chronic restraint stress.
Glutamatergic mechanisms are central to this behavioral phenomenon of cross-sensitization between stress and psychostimulants (cocaine and amphetamine). For instance, pharmacological blockade of NMDA or AMPA receptors reverses stress-induced behavioral sensitization to cocaine and amphetamine, respectively [132,134]. Acute restraint elevates AMPA-receptor surface expression in the NAc core, heightening sensitivity to intra-NAc AMPA injection and to cocaine-induced locomotion—effects abolished by NMDA or AMPA receptor antagonists [132]. Likewise, chronic stress induced cross sensitization to cocaine psychostimulant effect and escalating of drug-self administration along 10 days. Further, it correlates with increased AMPA-receptor expression and structural plasticity—namely, greater mushroom-type spine density—within the core [86,131,153,154]. Together, these findings underscore a persistent, core-specific glutamatergic remodeling that underlies stress-induced vulnerability to psychostimulant effects (see details of synaptic plasticity in section 8).
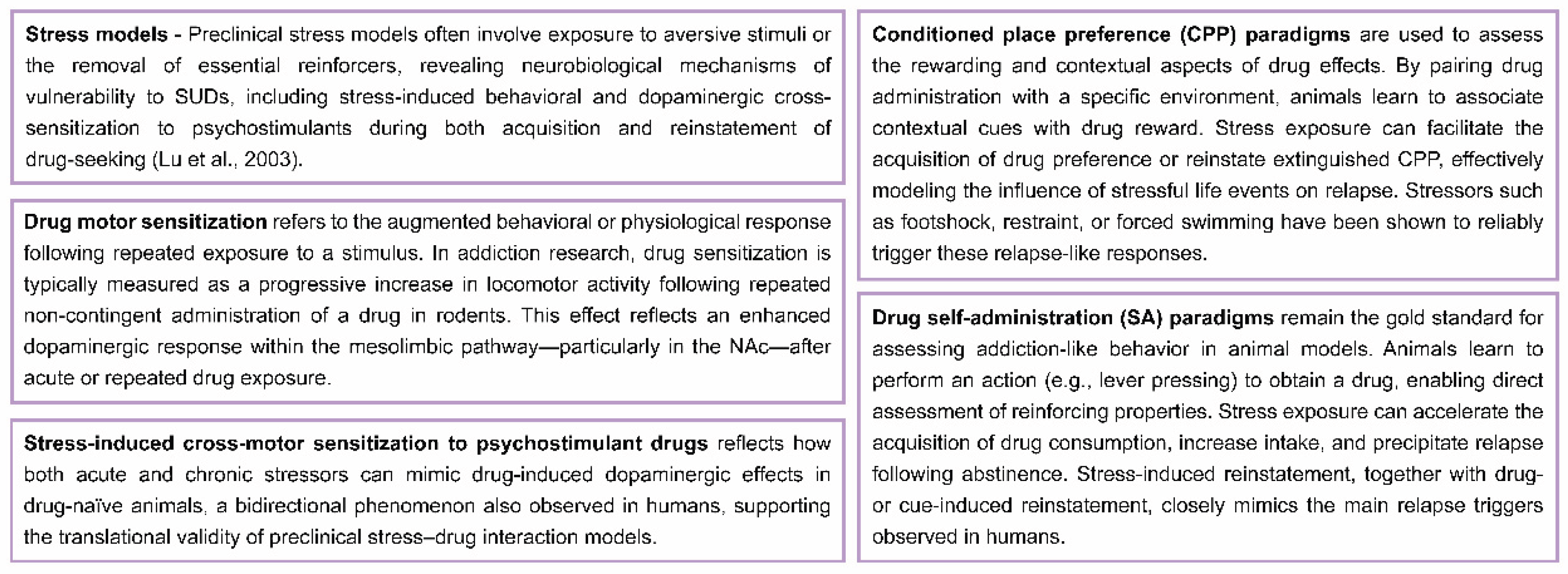
Box 1. Preclinical paradigms of stress-induced vulnerability to SUDs.
5. Dysregulation of Glutamate Homeostasis in the Nucleus Accumbens Core as a Key Mechanism of Stress-Induced Cocaine Vulnerability
Repeated stress exposure is thought to produce a persistent hyperglutamatergic state within key nodes of the reward circuit, such as the NAc, thereby facilitating the development of behavioral sensitization and drug-seeking behavior [86,94,131,132,153,155,156]. A central mechanism underlying these adaptations involves the downregulation of the glutamate transporter 1 (GLT-1) in the NAc core, a pathogenic hallmark proposed to mediate the heightened vulnerability to addictive behaviors induced by drugs, and by acute or chronic restraint stress exposure [17,86,132].
It is well established that cocaine-evoked increases in glutamate release within the NAc core occur exclusively in sensitized animals, but not in non-sensitized controls or saline-treated subjects after prolonged withdrawal [147,157]. Similar results have been reported in the VTA, where Kalivas and Duffy (1998) [158] showed that a cocaine challenge selectively elevated glutamate release in previously sensitized animals after 21 days of withdrawal. Basal extrasynaptic glutamate concentrations in the NAc core critically regulates synaptic glutamatergic activity through presynaptic metabotropic glutamate receptors (mGluR2/3), which exert inhibitory feedback on glutamate release [159,160]. Notably, following chronic cocaine exposure, withdrawal reduces basal extracellular glutamate levels in the NAc core [161], while cocaine re-exposure elicits a marked glutamate surge [147,157]. These adaptations have been linked to impaired cystine–glutamate exchange via system Xc⁻ and decreased GLT-1-mediated uptake [161,162,163]. Consistently with this, current evidence indicates that most extracellular glutamate detected by microdialysis originates from astrocytic mechanisms rather than vesicular neuronal release [160,164,165,166]. Compelling evidence links both stress and psychostimulant exposure to persistent impairments in glutamate clearance, particularly via downregulation of astrocytic GLT-1 across motivation-related brain regions. For example, GLT-1 decreases have been observed in the hippocampus and cortex [167] and in corticolimbic slices following inescapable footshock [168], as well as in the prefrontal cortex and striatum after chronic social defeat stress [169]. Consistent findings have emerged from addiction models, in which prolonged self-administration and withdrawal from cocaine, amphetamine, heroin, nicotine, and ethanol result in robust GLT-1 reductions, particularly in the NAc core [162,163,170,171,172,173,174,175,176]. Fischer-Smith et al. (2012) [177] further demonstrated that this downregulation persists throughout cocaine abstinence and is most pronounced in the NAc core, regardless of prior drug intake levels.
Consistently, both acute and chronic restraint stress lead to long-lasting disruptions in glutamate homeostasis within the NAc core. Using no-net-flux microdialysis method, elevated basal glutamate concentration were detected in the NAc core—but not the shell—up to 21 days after the first restraint session (acute restraint stress: García-Keller et al., 2013 [132]; chronic restraint stress: Ávalos et al., 2022 [153]). Importantly, this imbalance contributed to long-term postsynaptic neuroadaptations within the NAc core and to associated behavioral consequences. Specifically, chronic restraint stress promoted cross-sensitization to cocaine and facilitated the acquisition of cocaine self-administration when assessed 21 days after the initial restraint stress exposure (2 h daily for seven consecutive days) [87,131,154,178]. Remarkably, even a single 2-hour restraint session was sufficient to induce behavioral sensitization and enhance cocaine self-administration [86]. Critically, treatment with the β-lactam antibiotic ceftriaxone, which upregulate GLT-1 expression, reversed both the stress-induced behavioral alterations and the associated GLT-1 downregulation. It should be noted that acute 2-hour restraint stress did not affect Na+-independent 3H-glutamate uptake, which estimates cystine-glutamate exchanger activity, but significantly reduced Na+-dependent uptake, which reflects GLT-1 function. This reduction in glutamate uptake was further supported by whole-cell patch-clamp recordings of NAc core medium spiny neurons (MSNs), demonstrating greater synaptic glutamate spillover in stress compared with control animals [86]. Together, these findings support the notion that stress-induced dysregulation of glutamate homeostasis via GLT-1 in the NAc core is a central mechanism driving increased sensitivity to the psychomotor and reinforcing effects of cocaine. In line with this, Guzmán et al. (2020) [116] showed increased NAc core glutamate levels associated with a cocaine-paired context during restraint stress-induced reinstatement of extinguished cocaine-conditioned place preference in rats.
GLT-1 is strategically localized on astrocytic processes adjacent to the synaptic cleft, enabling rapid and efficient clearance of synaptically released glutamate to maintain extracellular homeostasis [179,180]. Disruption of GLT-1-mediated uptake following restraint stress results in glutamate spillover into the extrasynaptic space, leading to two key pathophysiological consequences: i) activation of mGluR2/3 and loss of GLT-1’s neuroprotective constraint against excessive postsynaptic glutamate stimulation, ii) a prolonged decay time of NR2B-containing NMDA receptor-mediated currents in the NAc core, detectable up to 21 days after stress, which strengthens and prolongs glutamatergic transmission in this nucleus [86]. Given the predominantly perisynaptic localization of NR2B subunits [175], these findings provide strong support for the glutamate spillover hypothesis. Notably, ceftriaxone treatment this stress-induced increases in NMDA decay time within the NAc core. This electrophysiological evidence derives from a collaborative study between Peter Kalivas’s laboratory and our own [86]. Consistent with earlier reports [162,163,181], more recent evidence demonstrate that ceftriaxone also normalizes basal extracellular glutamate levels in the NAc core [86,87,153], thereby reestablishing mGluR2/3-mediated inhibitory feedback and preventing the stress-induced behavioral consequences.
Moreover, the persistent elevation in basal extracellular glutamate resulting from reduced GLT-1 function contributes to maladaptive postsynaptic changes, including structural plasticity in MSNs [86,87,131,153] (see section 8 for further details). Together these findings highlight astrocytic GLT-1 dysregulation as a unifying mechanism underlying stress and drugs-induced glutamatergic maladaptations within the reward circuit, ultimately predisposing individuals to SUDs.
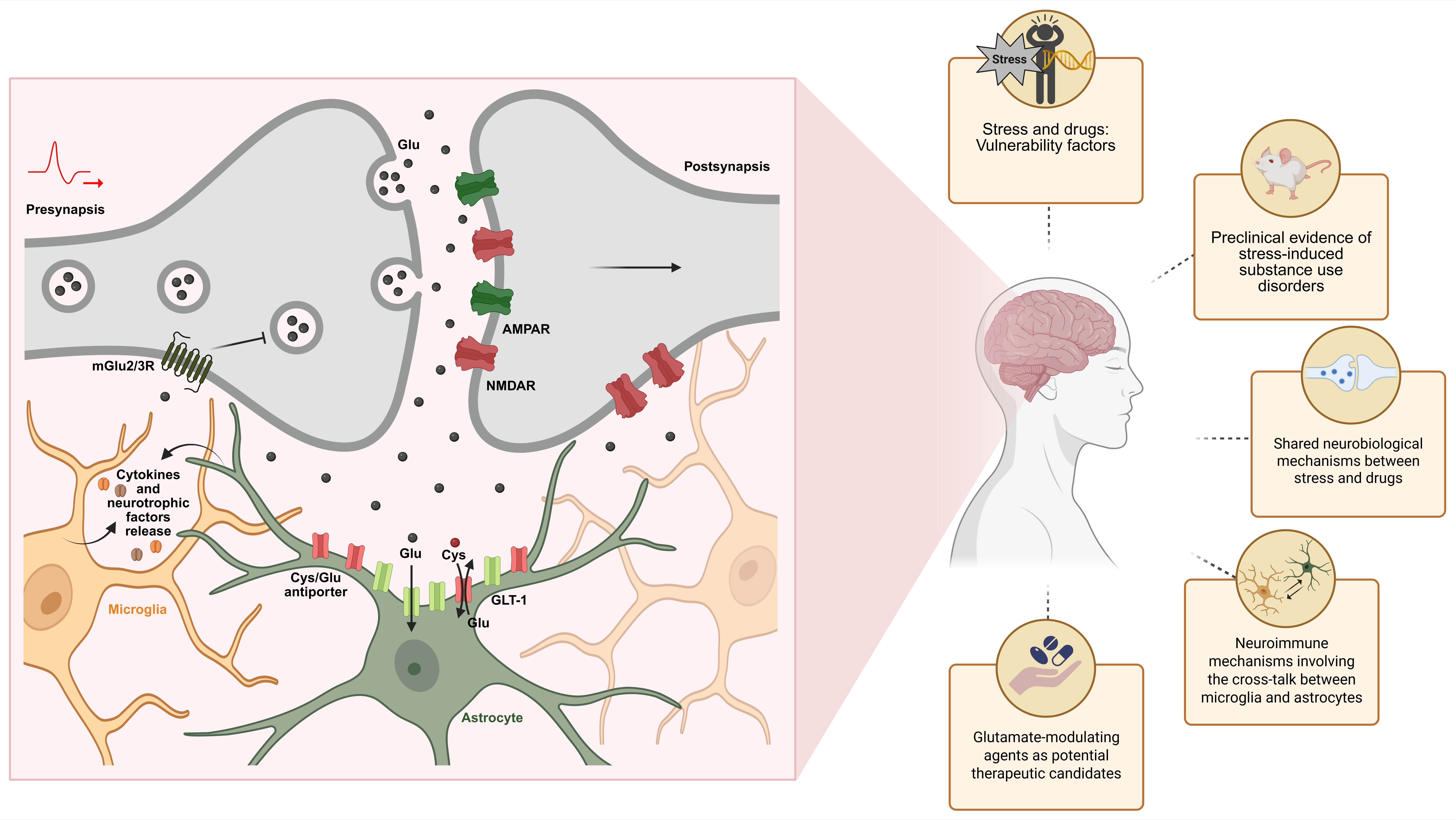
6. Glial Cells: Microglia and Astrocytes in Glutamate and Neuroimmune Regulation
Glial cells, once considered merely immune sentinels or passive structural elements, are now recognized as active regulators of neuronal function and plasticity. Among them, astrocytes have been extensively studied for their essential roles in synaptic transmission and in maintaining extracellular glutamate homeostasis. More recently, microglia have been identified as dynamic modulators of neuronal circuits and synaptic plasticity [182]. Although oligodendrocytes also participate in neuronal regulation, their functions are beyond the scope of this review.
Both microglia and astrocytes are central players in neuroimmune signaling and the structural remodeling of synaptic connections. This section provides an overview of their defining characteristics and physiological functions, with particular emphasis on their interactive roles in glutamatergic regulation, neuroimmune communication, and the reorganization of neuronal circuits in response to stress and drugs of abuse. This framework supports the primary aim of this review: to examine the neurobiological mechanisms underlying stress-induced vulnerability to SUDs. By focusing on glial cell contributions, we highlight how microglia and astrocytes not only monitor the neural microenvironment but also actively shape synaptic connectivity, regulate glutamatergic transmission, and orchestrate immune responses—core processes involved in both stress- and drug-induced neuroadaptations.
6.1. Microglia: Characteristics, Role in Motivation Circuits, Synaptic Plasticity, Phenotypic Markers, and Activation States
6.1.1. Characteristics and Neurophysiology
Microglia account for approximately 10–20% of the total brain cell population, with a higher density in gray matter than in white matter [183]. These immunocompetent cells serve as the brain’s resident macrophages, constituting the innate immune defense of the central nervous system (CNS) [184]. Sharing phenotypic markers with peripheral monocytes and macrophages [185], microglia express a wide array of receptors, including those for neurotransmitters (glutamate, GABA, acetylcholine, DA, norepinephrine), neuromodulators (cannabinoids, opioids), pattern recognition molecules (e.g., TLR2, TLR4), purinergic receptors, glucocorticoid and mineralocorticoid receptors, and receptors for neurotrophins, cytokines, chemokines, complement proteins, and CSF-1 [186]. This diverse receptor repertoire enables microglia to respond to an extensive range of environmental signals.
Even in their highly ramified “resting” state, microglia exhibit continuous motility, extending and retracting processes to monitor the local environment without moving their soma or disrupting neuronal circuitry [187,188,189]. This constant surveillance reflects a vigilant immunological state. In response to injury or developmental cues, microglia can transition into an amoeboid, migratory phenotype [190].
6.1.2. Microglia in Motivation Circuits
Microglia have a physiological role within motivational circuits, particularly in key nuclei such as the NAc, where they participate in synaptogenesis and activity-dependent synaptic pruning under normal conditions. They eliminate synapses during development [191,192] and modulate cognitive functions such as learning and memory [193,194]. Notably, synaptic pruning in the NAc during adolescence exhibits sex-specific patterns, targeting distinct synaptic inputs in males and females [195]. Microglial elimination of DA receptors during adolescence influences sex-specific development of NAc circuitry and social behavior. Supporting this, Csf1rΔFIRE/ΔFIRE mice lacking microglia show reduced excitatory synapse formation, altered presynaptic release, and changes in postsynaptic kinetics, underscoring their essential role in sculpting neural networks underlying adult behavior [196].
6.1.3. Synaptic Activity and Plasticity
Microglial processes form direct physical contacts with synapses [197,198,199,200], facilitating bidirectional communication with neurons and maintaining synaptic homeostasis [201,202]. They influence synaptic plasticity through the release of a variety of signaling molecules, including ATP [203], neurotransmitters [204,205,206,207], extracellular matrix components [208,209] and cytokines [210]. Among these, TNF-α is particularly significant, modulating synaptic strength [211], activity-dependent plasticity [212,213], and astrocytic function [214,215,216].
Microglia also participate in glutamate signaling. Under specific conditions, they express the system Xc⁻ [217,218,219] and, although capable of expressing glutamate transporters like GLT-1, they account for only ~10% of glutamate uptake under physiological conditions [186]. Their transporter expression increases in pathological states, suggesting a compensatory role during glutamatergic dysfunction, whereas astrocytes remain the primary regulators of extracellular glutamate clearance [220,221,222]. Thus, microglia and astrocytes work in tandem to preserve excitatory balance, and dysfunction in either cell type can profoundly impact circuit integrity.
6.1.4. Phenotypic Markers
Microglia are cells characterized by specific markers, including CSF-1R (CD115), CX3CR1, CD68, F4/80, CD11b, and Iba-1. The expression of CD11b and Iba-1 is often upregulated upon activation [223,224,225], making them useful tools for assessing microglial states in physiological and pathological conditions.
6.1.5. Activation, Priming and Morphological Changes
Once seen merely as sensors of injury [184], microglia are now known to actively initiate and propagate brain pathologies. Their activation can be triggered by diverse stimuli, including ATP [189], cannabinoids [226](Walter et al., 2003), opioids [227], proinflammatory cytokines, glutamate [20], cocaine [228], and stress-related signals [229,230,231]. Microglial activation is a graded and context-dependent process, not a binary switch, and varies across brain regions according to microenvironmental cues [232,233]. Microglial morphological changes associated with activation include process retraction, soma hypertrophy, and transformation into an amoeboid shape. Intermediate forms, like hyper-ramified microglia, also occur [188,189,234]. Depending on their functional state, microglia release a variety of bioactive molecules, including pro- and anti-inflammatory cytokines, chemokines, prostaglandins, reactive oxygen/nitrogen species, and neurotrophic factors, all of which can influence neuronal survival and synaptic remodeling [235].
Importantly, activated microglia can return to a resting-like morphology while retaining epigenetic changes that leave them in a “primed” state [229,230]. These “experienced” microglia exhibit exaggerated responses to subsequent stimuli and may contribute to the etiology of neuropsychiatric conditions involving glial dysfunction. Chronic stress, in particular, primes microglia toward a proinflammatory phenotype, increasing their sensitivity to future insults, including drug exposure [230].
As mentioned, microglial functions are closely integrated with astrocytes. The following section therefore focuses on astrocytes, detailing their neurophysiology and roles in synaptic regulation within stress- and drug-sensitive brain circuits.
6.2. Astrocytes: Characteristics, Role in Motivation Circuits, Synaptic Plasticity, Neurotransmitter Regulation, and Immune Functions
6.2.1. Characteristics and Neurophysiology
Astrocytes are the predominant non-neuronal cells in the CNS, outnumbering neurons by approximately ten to one and accounting for more than 50% of all CNS cells [236]. In subcortical regions, they represent about 20% of the total cellular population [237,238,239]. Morphologically, astrocytes exhibit a star-like shape, with a central soma and numerous branching processes. Although they do not generate action potentials, they display changes in membrane potential and intracellular calcium (Ca²⁺) dynamics that are essential for modulating neuronal activity and maintaining homeostasis [240].
Far from being passive support cells, astrocytes participate in a broad spectrum of critical functions, including formation and regulation of the blood–brain barrier, protection against excitotoxicity, promotion of synaptic plasticity, coordination of neural network activity, metabolic support, and participation in bidirectional communication with neurons, other astrocytes, and microglia [241,242,243,244]. Each astrocyte occupies a non-overlapping spatial domain and extends fine processes that contact thousands of synapses and neuronal soma [245,246], thus playing a key role in sustaining synaptic function and metabolic coupling [247]. This intimate association forms the so-called “tripartite synapse,” which integrates astrocytes into the functional structure of synaptic transmission [240,248]. Astrocytes are also strategically positioned at the interface between neurons and the vasculature. Their perivascular end feet envelop blood vessels, enabling glucose uptake and the delivery of metabolic substrates to neurons [247,249].
6.2.2. Astrocytic Markers
Glial fibrillary acidic protein (GFAP) is the most widely used marker for identifying astrocytes and plays diverse roles in cell migration, proliferation, synaptic remodeling, and blood–brain barrier maintenance [243]. GFAP has also been implicated in regulating the expression and function of glutamate transporters. In GFAP-deficient mice, impaired glutamate uptake and downregulation of both astrocytic (GLAST/EAAT1) and neuronal (EAAC1/EAAT3) transporters have been observed in cortical and hippocampal regions [243].
6.2.3. Astrocytes in Motivation Circuits
Within the NAc, astrocytes are essential regulators of synaptic transmission and neural excitability. They influence motivational and reward-related behavior by releasing gliotransmitters and neuromodulators [250]. They maintain neurotransmitter balance through Ca²⁺-dependent uptake and release of glutamate and GABA [251,252], ensuring proper synaptic termination and recycling. In addition to glutamate clearance, astrocytes can also release small quantities of glutamate, coordinating neuronal firing and modulating excitatory/inhibitory balance via mechanisms resembling synaptic exocytosis.
Recent evidence highlights astrocyte–DA interactions in the NAc. For instance, astrocytes respond to synaptically released DA with increases in intracellular Ca²⁺, leading to ATP/adenosine release and subsequent suppression of excitatory transmission via adenosine A1 receptor activation [253]. Similarly, DA D2 receptor signaling modulates Ca²⁺ activity in midbrain astrocytes, indicating regional heterogeneity in astrocyte–DA interactions [254]. These findings underscore the importance of astrocytes in tuning dopaminergic and glutamatergic signaling within reward circuits.
Astrocytes also participate in neuroimmune regulation. They express receptors for cytokines and other immune signals, allowing them to sense and modulate inflammation [255,256]. Upon injury or stress, they undergo reactive gliosis—a hypertrophic response associated with the release of pro- and anti- inflammatory mediators, recruitment of microglia, and regulation of immune cell infiltration [257,258,259].
6.2.4. Astrocytes and Synaptic Activity
Astrocytes contribute to extracellular homeostasis by regulating potassium and glutamate concentrations [247]. They detect synaptic activity through glutamate receptor activation [20,260], triggering Ca²⁺ transients that induce gliotransmitter release [261,262]. Gliotransmitters such as glutamate, ATP, D-serine, cytokines, and neuropeptides [211,262,263,264] influence neuronal excitability and network synchronization. Astrocytes also generate Ca²⁺ waves across glial networks, coordinating activity within and across brain regions [265] and potentially activating microglia as early signals of glutamate dysregulation [187,266,267].
Glutamatergic gliotransmission from astrocytes typically targets extrasynaptic NMDA receptors containing NR2B subunits, affecting plasticity and excitability [263]. In addition, astrocytes supply neurons with glutamine for glutamate synthesis [268], reinforcing their role in maintaining synaptic balance. Astrocytic regulation of extracellular glutamate is particularly relevant in excitotoxicity, epilepsy, neurodegeneration [221,269,270,271], and addiction. Within the cortico-accumbal circuit, astrocytes modulate glutamatergic plasticity by controlling glutamate tone: system Xc⁻ exchanges intracellular glutamate for extracellular cystine in a 1:1 ratio, accounting for over 50% of extracellular glutamate in the NAc core [272,273]. In parallel, astrocytic glutamate uptake via GLT-1 is crucial for synaptic fidelity and neuroprotection, as GLT-1 mediates over 90% of total brain glutamate clearance [274,275].
7. Glial Contributions to Stress-Induced Glutamate and Neuroimmune Dysregulation in the Nucleus Accumbens Core: Microglia and Astrocyte Cross-Talk
Increasingly, microglia—the resident innate immune cells of the CNS—are recognized as pivotal regulators of neurobiological adaptations to both stress [188,189] and drug exposure [21], as well as their interaction [276]. This convergence may represent a critical mechanism driving heightened vulnerability to addiction. While microglial activation has been investigated across multiple mesocorticolimbic regions, studies specifically addressing their role within the NAc—a central hub of the brain’s reward circuitry—remain relatively scarce.
In parallel, a growing body of evidence—outlined in the preceding sections—highlights the active involvement of both, astrocytes and microglia, as well as their dynamic interactions, in regulating neuronal function following stress. A major advance has been the discovery of bidirectional astrocyte–microglia communication mediated by cytokines such as IL-1α, IL-1β, and TNF-α, which collectively shape neuronal circuitry under both physiological and pathological conditions [20].
Emerging evidence further supports a close link between maladaptive glutamatergic plasticity and neuroimmune processes in drug-experienced animals [277,278,279]. In this context, stress and glucocorticoids have been shown to sensitize neuroinflammatory responses to subsequent drug exposure, indicating that immune mechanisms may play a crucial role in mediating stress-induced vulnerability to addiction [229,280,281]. Moreover, several studies have documented a robust association between neuroinflammatory states and altered glutamate homeostasis in specific brain regions, including the NAc [20,231,282], thereby reinforcing the hypothesis that neuroimmune alterations may serve as upstream modulators of glutamatergic dysregulation in addiction.
7.1. Microglial Activation by Stress and Drugs: Neuroimmune Modulation in Mesolimbic Circuits and Peripheral Crosstalk
Since stress is a significant risk factor in the development of SUDs, understanding the role of microglia in the biological mechanisms that mediate the interaction between stress and drugs of abuse is a crucial step toward developing novel therapeutics for dual pathology, such as comorbid stress-related disorders (e.g., PTSD) and addiction. Notably, microglial cells rapidly adopt reactive phenotypes in response to various stimuli. In this activated state, they release pro-inflammatory cytokines, contribute to disrupt glutamate homeostasis, and participate in the remodeling dendritic spine architecture [21,193,283,284,285].
Their dynamic ability to shift activation states (see section 6.1) has attracted increasing attention, as stress exposure can lead to both immediate and long-lasting neuroimmune and structural alterations in stress-sensitive brain regions [21,24,188,189,286]. In this context, it becomes important to explore how exposure to stress leads to microglial activation and the subsequent release of pro-inflammatory mediators within stress-sensitive brain regions. For instance, stress robustly activates microglia and elevates pro-inflammatory cytokines, including TNF-α and IL-1β, in the prefrontal cortex and hippocampus. These neuroimmune changes persist well beyond the cessation of stress. Specifically, a six-day chronic electric shock stress protocol—previously shown to induce behavioral alterations reminiscent of PTSD—elicited these molecular and cellular changes, which remained detectable nine days after the final stress session. Notably, a six-day regimen of intragastric minocycline –an inhibitor of microglial activation-, administered 30 minutes prior to each stress exposure, effectively prevented both the behavioral and molecular changes. This protective effect has been attributed to the anti-inflammatory properties of minocycline [287]. Similarly, inescapable shock stress upregulates IL-1β and IL-6 mRNA in the hypothalamus and hippocampus, effects also prevented by minocycline [288,289,290]. Human studies similarly link psychological stress to immune dysfunction and increased prevalence of mood disorders [291,292,293,294,295].
In addition to these findings, stress appears to prime microglial inflammatory responses. Hyper-ramification is often considered a primed intermediate state between resting and hypertrophic microglial activation [197,234]. Consistent with enhanced neuroinflammatory signaling, chronic stress induces profound morphological changes in microglia. Several paradigms—including restraint stress, repeated social defeat, and footshock—promote hyper-ramification, characterized by increased secondary branching without alterations in primary processes [296], along with elevated Iba-1 expression in the prefrontal cortex, amygdala, and dentate gyrus of the hippocampus [287,297,298,299,300,301,302,303].
Although the functional implications of this hyper-ramified morphology are not fully understood, it is generally considered an intermediate state between resting and fully activated microglia, often interpreted as an early stage of hypertrophy. This phenotype may reflect subtle homeostatic shifts [234] or responses to non-pathological, experience-dependent stimuli [197,296,298,304]. Supporting this concept, glucocorticoids released in response to inescapable stress can prime microglia to exhibit exaggerated inflammatory responses to subsequent challenges [230]. Notably, these studies focused on hippocampal microglia, raising the question of whether similar glucocorticoid-driven priming occurs in other stress-sensitive regions such as the NAc. Overall, these findings highlight the critical role of glucocorticoid regulation in maintaining brain homeostasis and demonstrate how stress-induced neuroendocrine dysregulation can lead to region-specific microglial activation and neuroinflammation [305,306,307].
Exposure to addictive drugs can also profoundly affect microglial function and the release of inflammatory mediators, although the nature of these responses varies depending on the drug type, dose, and brain region [308]. In vitro studies have further demonstrated that cocaine exposure enhances the transcription of TNF-α and IL-6 in microglial cultures [309]. In vivo, ethanol has been shown to sensitize microglia, inducing a hyper-ramified activation state characterized by modest morphological changes and increased release of pro-inflammatory cytokines [277]. In cocaine models, microglia are also activated through Toll-like receptor 4 (TLR4), receptor for advanced glycation end-products (RAGE), and high-mobility group box 1 (HMGB1) signaling, as well as through NLRP3 inflammasome pathways, thereby linking psychostimulant action to neuroimmune cascades [21,228,310,311]. Consistent with these findings, psychostimulant administration increases Iba-1 expression across multiple brain regions, including the striatum [309]. Additionally, a recent study reported a long-lasting elevation of TNF-α in the NAc core following nicotine self-administration and extinction [279]. Collectively, these findings indicate that microglial morphological and functional alterations are common neuroimmune responses triggered by both stress and exposure to drugs of abuse.
Interestingly, compelling evidence for microglia’s central role in mediating the effects of chronic stress on cocaine vulnerability comes from Ávalos et al. (2022) [87]. Male rats were exposed to chronic restraint stress or sham treatment and then underwent a cocaine self-administration protocol. Chronic stress led to a progressive escalation of cocaine intake over ten days, an effect that was completely prevented by daily administration of minocycline during the self-administration phase, maintaining intake at levels comparable to non-stressed controls. Notably, neither stress nor minocycline altered sucrose consumption in a separate cohort, indicating that the stress-induced enhancement was specific to cocaine-seeking behavior. At the cellular level, chronic restraint stress induced hyper-ramified microglial morphology in the NAc core of vehicle-treated animals, consistent with a primed, pro-inflammatory state. Minocycline effectively blocked this morphological activation. Molecular analyses revealed that stressed animals exhibited elevated TNF-α mRNA and protein, reduced GLT-1 expression, and showed signs of astrocyte hypoactivity, reflected by decreased GFAP immunoreactivity in the NAc core. Remarkably, all these molecular alterations were reversed by minocycline, highlighting the tight link between microglial activation, astrocytic dysfunction, and stress-induced vulnerability to cocaine. Furthermore, these findings strongly support the notion that microglia are key mediators of the behavioral and neurobiological consequences of combined stress and cocaine exposure. Taken together, this body of evidence indicates that microglia play a pivotal role in the stress-induced escalation of cocaine intake, as well as in the associated cellular and molecular neuroadaptations—underscoring their critical involvement in the shared pathophysiology of stress-related disorders and SUDs.
Stress-Induced Recruitment of Peripheral Monocytes to the Brain: Role of Corticosterone, NMDA Receptors, and IL-6 Signaling
Complementing these findings, studies using restraint and social defeat stress models have demonstrated neuroimmune alterations, including microglial proliferation within the brain parenchyma and the recruitment of peripheral monocytes to regions such as the prefrontal cortex and hippocampus [301]. Notably, this redistribution and increased trafficking of peripheral monocytes have been associated with neuroimmune disturbances triggered by repeated stress exposure, events that have direct implications for synaptic plasticity and may promote the emergence of stress-related behavioral alterations [312]. In mice subjected to restraint stress, flow cytometry analyses based on CD11b/CD45 markers revealed microglial proliferation, attributed to corticosterone-induced activation of NMDA receptors. Blocking corticosterone synthesis, glucocorticoid receptors, or NMDA receptors mitigated this stress-induced microglial expansion, and administration of the NMDA receptor antagonist MK-801 prevented proliferation even following exogenous corticosterone treatment in non-stressed animals [231]. These results align with earlier work from our laboratory, which demonstrated that amphetamine-induced immune sensitization—paralleling phenomena in the limbic system—was blocked by systemic MK-801 administration, suggesting that both processes are mediated by glutamate-dependent mechanisms [313]. While NMDA receptor blockade effectively prevents microglial proliferation induced by stress or psychostimulants, the effectiveness of other anti-inflammatory strategies remains to be fully established; thus far, additional studies are needed to determine whether minocycline or related compounds can effectively reverse microglial proliferation under stress and drug exposure. However, due to the non-specific actions of minocycline and the complexity of distinguishing central from peripheral immune responses definitive conclusions regarding peripheral contributions under stress conditions are difficult to draw by using this drug [186,314].
Collectively, these observations suggest that chronic stress engages both central and peripheral immune mechanisms. Cytokines such as IL-6 play a central role in this immune-to-brain communication, acting as mediators between peripheral immune cells and the CNS. Repeated social defeat stress robustly increases plasma IL-6 levels in mice, promoting recruitment of proinflammatory monocytes to the brain, which adopt a primed profile and facilitate IL-1-mediated inflammatory responses [315]. Translationally, elevated IL-6 levels have been observed in cocaine users [316]. Stress-induced IL-6 can also cross the brain barrier following loss of tight junction proteins such as claudin-5, thereby modulating brain parenchymal inflammation and behavior [317]. Importantly, although IL-6 contributes to peripheral-to-central immune signaling, microglial morphological activation in regions such as the NAc can occur through IL-6–independent mechanisms, consistent with findings from our laboratory showing that chronic stress-driven increases in cocaine intake do not alter IL-6 expression in the NAc, despite marked microglial activation [87,178]. Additional studies corroborate this region- and paradigm-specific pattern, reporting both unaltered and increased IL-6 levels in different brain regions and stress models [318,319,320,321].
7.2. Astrocyte Reactivity Under Stress and Drug Exposure: Glutamate and Immune Adaptations
Astrocytes respond to both acute and chronic injuries in the CNS [322,323]. In acute pathological conditions involving cell death—such as ischemia, oxidative stress, or trauma—astrocytes can become pro-inflammatory and adopt phagocytic roles. This activation involves increased expression of inflammatory markers and the release of inflammatory mediators, including IL-1β, TNF, IL-6, and various chemokines. This pro-inflammatory state impairs glutamate clearance and promotes oxidative stress, contributing to excitotoxic neuronal damage. In contrast, during chronic CNS injury or disease, astrocytes undergo reactive astrogliosis, characterized by hypertrophy, proliferation, and migration, with elevated expression of markers such as GFAP, commonly observed in many brain disorders [324].
As discussed in this review, a key hallmark of cocaine-induced astrocytic alterations is the downregulation of the GLT-1 in the NAc. Typically, reduced GLT-1 levels are linked to reactive astrogliosis in response to brain injury, ischemia, or neurodegeneration, suggesting that astrocytes might adopt a reactive phenotype following drug exposure. In support of this notion, increased astrocyte reactivity has been reported after non-contingent exposure to psychostimulants such as cocaine [325]. This reactive state is often marked by elevated expression of GFAP. In vitro studies demonstrated that methamphetamine induces robust activation of purified cortical astrocytes via a protein kinase C (PKC)-dependent mechanism. Furthermore, sustained astrocytic activation was observed in cortical neuron–glia cocultures following methamphetamine exposure [326,327]. These in vitro findings are consistent with in vivo evidence showing that repeated administration of methamphetamine leads to behavioral sensitization, which is accompanied by PKC-dependent astrocytic activation in the cingulate cortex and NAc. Similarly, morphine also induced astrocytic activation through PKC signaling in cortical neuron–glia cocultures, although purified astrocytes did not respond directly to morphine [328].
Despite evidence of astrocyte activation during experimenter-administered drug exposure, cocaine self-administration followed by extinction training leads to astrocytes in the NAc exhibiting reduced GFAP expression, decreased surface area and volume, and diminished colocalization with the presynaptic marker synapsin I [61], while no such changes occur in the prelimbic cortex or basolateral amygdala [329]. Importantly, astrocyte morphology and synaptic proximity remain unchanged after self-administration alone, with alterations emerging only after extinction/abstinence, underscoring the region-specific and time-dependent nature of astrocyte adaptations to cocaine exposure. Similarly, following methamphetamine self-administration and extinction, astrocytes in the NAc core show reduced contact of their perisynaptic processes (PAPs) with synapses, indicative of astrocytic process retraction [330], although this morphological change does not impair GLT-1 expression or glutamate uptake capacity. Together, these findings emphasize the complexity of psychostimulant-induced neuroadaptations and underscore the contribution of astrocytic dysfunction to synaptic alterations that may underlie relapse vulnerability.
Consistent with this evidence, this population of glial cells does not exhibit a reactive phenotype under stress conditions, proving on the contrary a hypo-reactive state [87,331]. For example, early or juvenile stress has been shown to adversely affect astrocytic function, leading to diminished GFAP expression, less complex astrocyte morphology, and impaired uptake of key neurotransmitters such as glutamate and GABA. Additionally, disruptions in the glutamate–glutamine cycle have been reported under these conditions [332]. Experimental models of early life stress, such as repeated maternal separation or deprivation during lactation, consistently show marked reductions in GFAP-immunoreactive astrocytes across multiple brain regions, including the prefrontal cortex, hippocampus, striatum, and amygdala [318,333,334]. These astrocytic alterations induced by stress are frequently accompanied by decreased levels of other astrocytic markers, such as S100β, as well as structural changes including reduced dendritic spine density in the prefrontal cortex [335]. Although this consistency in GFAP reduction and astrocyte complexity across different brain regions following stress, recent evidence from chronic stress and systemic inflammation paradigms reveals a more complex picture. Using the unpredictable chronic mild stress model and LPS injections, astrocyte activation increased—evidenced by higher GFAP fluorescence, greater branch bifurcation, and enhanced arborization—in multiple brain regions, with females showing more pronounced effects in the hippocampus and amygdala than males [336]. These findings highlight the region- and sex-specific nature of astrocyte responses to diverse stress paradigms and inflammatory stimuli (i.e., LPS), potentially contributing to differential vulnerability to stress-related disorders. Consistent with this idea, our laboratory demonstrated that chronic restraint stress, which underlies stress-induced cocaine self-administration, is associated with reduced astrocyte reactivity in the NAc core evidenced by decreased GFAP immunoreactivity [87].
7.3. Microglia-Astrocytes Crosstalk: Proinflammatory Signaling, GLT-1 Downregulation and Stress-Induced Vulnerability to Cocaine Use Disorder
Microglia–astrocyte crosstalk plays a central role in the neuroadaptations induced by chronic stress that enhance vulnerability to cocaine addiction. Stress induces profound structural and functional changes in both astrocytes and microglia—two glial populations essential for glutamatergic regulation and synaptic homeostasis—particularly in the NAc core [86,87,132]. Although neurons and microglia can express GLT-1 in pathological conditions, their contribution under physiological states is minimal [186,337,338,339,340,341]. In this sense, astrocytes are the principal cells responsible for glutamate clearance, playing a predominant role in maintaining extracellular glutamate homeostasis [221,274,342].
Microglial cells are exquisitely sensitive sentinels of the parenchyma, capable of detecting subtle microenvironmental changes and mounting rapid responses to preserve homeostasis [184]. Their participation in glutamate regulation is supported by the presence of ionotropic and metabotropic glutamate receptors, enabling direct detection of extracellular glutamate fluctuations [343,344]. Under stress conditions, excessive extracellular glutamate not only disrupts synaptic signaling but also acts as a danger-associated signal that triggers microglial activation, including hyper-ramification, process extension, and proliferation via NMDA receptor activation—particularly following corticosterone release [20,87,132,145,231,345,346,347,348]. This extracellular glutamate accumulation may reflect both enhanced glutamatergic drive and reduced astrocytic reuptake [87,145,349].
Once activated, microglia—as the brain’s innate immune cells—release pro-inflammatory cytokines such as TNF-α that contribute to the disruption of glutamate homeostasis by modulating astrocytic function [350,351,352,353,354,355] and postsynaptic outcomes [24]. Although astrocytes can produce TNF-α at low levels, transcriptomic analyses consistently identify microglia as the primary CNS source [356,357,358,359,360,361]. Stress and drugs of abuse elevate TNF-α, which act at glial and postsynaptic levels to perpetuate glutamatergic dysregulation [20,24,279,362,363]. Microglia thus appear to occupy an upstream position in regulating glutamate uptake and excitatory transmission, while astrocytes can reciprocally influence microglial activation through the release of ATP, glutamate, and other modulatory signals, supporting bidirectional crosstalk [364,365]. Mechanistically, TNF-α downregulates astrocytic GLT-1 expression and impairs its functional activity through transcriptional repression [214,216,366,367,368,369,370], consistent with a glutamate-to-cytokine feed-forward loop, as restraint stress increases cortical TNF-α in a glutamate-dependent manner, an effect blunted by NMDA receptor antagonism [371]. IL-1β may likewise reduce GLT-1 through post-translational or GFAP-independent mechanisms [372], although in our studies chronic stress capable of impairing glutamate homeostasis and facilitating cocaine self-administration does not necessarily increase IL-1β expression in the NAc [87].
Chronic stress not only produces a sustained decrease in GLT-1, but also reduces astrocytic GFAP immunoreactivity in the NAc core [87,346]. GFAP scaffolds GLT-1: GFAP-knockout astrocytes show impaired GLT-1 trafficking to the membrane and reduced uptake capacity [373]. Neuron–astrocyte communication further governs GLT-1 transcription and membrane dynamics, shaping transporter clustering and synaptic regulation [374,375,376]. Mirroring stress effects, chronic exposure to cocaine or nicotine not only reduces GLT-1, but also GFAP in the NAc core [162,279,377]. These findings emphasize that stress- or drug-evoked alterations in glutamatergic signaling critically impair astrocytic function and extracellular glutamate control.
Importantly, microglial hyperactivation appears to precede and drive astrocytic dysfunction. Microglia become activated before astrocytes, releasing signals that tune astrocytic responses and positioning microglia upstream in the control of glutamate uptake and excitatory transmission [203,236]. In summary, chronic stress disrupts the finely tuned interactions among astrocytes, microglia, and neurons in the NAc core, resulting in impaired glutamate clearance and its extracellular accumulation. In this context, TNF-α signaling has emerged as a key factor, being implicated both in GLT-1 downregulation [214,215,216,368] and in the regulation of synaptic plasticity [212,378]. Despite significant advances, the specific microglial mechanisms governing glutamate homeostasis within the NAc core under conditions of stress–drug comorbidity remain to be fully elucidated.
TNF-α/NF-κB Pathway: A key Signaling Axis Driving Astrocyte–Microglia Crosstalk in Stress-Induced Cocaine Vulnerability
As discussed above, and consistent with the general alterations in cytokine levels induced by stress and drugs, elevated TNF-α levels have been reported in several rodent models of chronic stress and in individuals with mood disorders [379,380], highlighting its role as a key mediator in astrocyte–microglia communication. Importantly, the transcription factor NF-κB acts as a pivotal regulator of this inflammatory response by controlling the transcription of TNF-α. Through its activation, NF-κB mediates stress-induced synaptic plasticity changes driven by cytokine signaling, thus linking neuroinflammation to behavioral adaptations [381,382].
The signaling pathway involving TNF-α/NF-κB has been widely investigated in the brain under both normal and disease conditions [381,383]. In the CNS, TNF-α commonly initiates the canonical NF-κB pathway, which depends on the phosphorylation of IKKα/β subunits. This event promotes the degradation of IκBα, allowing NF-κB heterodimers to translocate into the nucleus [384]. Within the nucleus, NF-κB interacts with κB elements in the DNA to modulate the transcription of numerous genes, particularly those related to inflammatory processes and glutamate signaling [385,386]. Because of the influence of NF-κB-regulated genes on drug-seeking behavior, it has been identified as a crucial mediator of the neuroadaptations triggered by chronic drug exposure [387]. Chronic cocaine exposition increases both total protein levels of the p50/p65 NF-κB subunits and overall NF-κB activity in the NAc [388,389]. Reducing NF-κB activity in this region diminishes cocaine reward and blocks the increase in dendritic spine density typically observed after prolonged exposure [389]. Inhibition of NF-κB signaling in the NAc core also reduces cue-induced cocaine seeking in a sex-dependent manner [390]. Recent evidence demonstrates robust activation of the cortical microglial TLR4/NF-κB signaling pathway after methamphetamine self-administration. Inhibition of this pathway effectively attenuates neuroinflammation and reduces addiction-related behaviors [391]. Stress, on the other hand, is also a well-established activator of NF-κB. Persistent NF-κB activation has been reported in the hippocampus following predator scent stress, and in the prefrontal cortex after exposure to immobilization/acoustic stress [392,393]. Manipulation of NF-κB signaling in the ventral striatum via viral vectors leads to a reduction in behavioral sensitivity to chronic social defeat stress, underscoring the involvement of IKK in the neuroplastic changes driven by stress [394]. In line with these findings, recent data from our lab demonstrated a marked activation of NF-κB in the NAc core from chronic stressed rats. Consistently, viral NF-κB inhibition prevented stress-induced facilitation of cocaine self-administration and cross sensitization [178]. The impact of viral NF-κB inhibition on stress-induced enhancement of cocaine intake is, at least in part, mediated by the restoration of GLT-1 levels within the NAc, leading to the normalization of glutamate homeostasis. In this sense, these results notably support that NF-κB regulates the expression of astrocytic GLT-1 [369]. TNF-α is typically recognized as an activator of NF-κB-dependent gene expression; however, it can also promote NF-κB-mediated transcriptional repression, as observed by the TNF-α/NF-κB-induced downregulation of GLT-1—a process in which N-myc proto-oncogene protein (N-myc) is involved [368]. Specifically, in the context of motor neuron injury, microglial TNF-α has been shown to downregulate astrocytic GLT-1 through the NF-κB pathway, enhancing glutamate-mediated excitotoxicity and contributing to motor neuron death in amyotrophic lateral sclerosis [216]. Under conditions of stress and drug exposure, TNF-α/NF-κB signaling plays a critical role in chronic stress-induced GLT-1 downregulation within the NAc core and the increased escalation of drug self-administration of cocaine self-administration, with N-myc appearing to contribute to NF-κB-mediated transcriptional repression [178]. These findings are consistent with those reported by Namba et al., 2022 [390] following nicotine exposure. We further showed that stress-induced NF-κB signaling occurs specifically in astrocytes within the NAc core. Interestingly, the NF-κB transcriptomic profile did not correlate with GLT-1 expression changes in the prefrontal cortex following extinction training after cocaine self-administration [395], suggesting that NF-κB may exert region-specific regulatory control over GLT-1 expression under conditions of drug exposure. Notwithstanding, these results support a pivotal role for TNF-α/NF-κB signaling in mediating astrocyte–microglia crosstalk, potentially driving the neuroplastic changes associated with stress-related vulnerability to cocaine addiction.
8. Microglia-Astrocyte Cross-Talk in the Regulation of Structural Synaptic Plasticity in the Nucleus Accumbens Core and Its Role in Stress-Induced Cocaine Vulnerability
Stress-induced plasticity shifts are often maladaptive, contributing to craving, depressive-like behaviors, cross-sensitization to psychostimulants, and the escalation of drug self-administration [396,397,398,399]. As both stress and drugs of abuse engage overlapping mechanisms—characterized by glial disturbances and disrupted synaptic plasticity—and given the close interplay between glial function and postsynaptic plasticity, this section reviews evidence from animal models highlighting glial alterations. Particular emphasis is placed on immune-regulatory processes primarily mediated by microglia, including the release of proinflammatory cytokines such as TNF-α and IL-1β, which critically modulate synaptic strength and structural plasticity within reward-related circuits. These microglial-derived signals influence astrocytic glutamate transporters and receptor dynamics, thereby linking stress-induced neuroimmune activation to maladaptive synaptic remodeling and, ultimately, to heightened vulnerability to SUDs.
In addition to the well-established role of astrocytes in glutamatergic mechanisms of synaptic plasticity shared by stress and SUDs, microglia—beyond their classical immune surveillance function—have emerged as key regulators of neurobiological processes underlying addiction and stress-related behaviors [277,363,400,401,402]. Early developmental studies first demonstrated their essential role in sculpting synaptic plasticity [192]. Microglial processes are highly dynamic, continuously extending and retracting to contact synapses during brain development and experience-dependent remodeling; however, their precise role in modulating synaptic circuits in the adult brain remains incompletely understood [187,191,197,403].
Over the past decade, compelling evidence has shown that microglia undergo marked morphological remodeling and secrete diverse signaling molecules—including cytokines, prostaglandins, and growth factors—in response to stress, suggesting their active involvement in stress-induced neuronal dysfunction [193]. Chronic stress has been shown to alter both microglial morphology and functionality [297,300], and these microglial alterations often coincide with neuronal structural and functional changes that may underlie long-lasting emotional disturbances associated with mood disorders such as major depression [404]. Consistently, Gaspar et al. (2021) [405] reported that unpredictable chronic mild stress induces morphological modifications in microglia and neurons within the NAc, with distinct sex-dependent features. More recent studies indicate that microglia actively shape neuronal networks by directly interacting with neurons or by releasing bioactive molecules such as cytokines—a mechanism susceptible to disruption by both stress and addictive drugs [276,406]. In line with this, inhibition of stress-induced microglial remodeling prevents the associated increases in NAc spine density and excitability in cocaine-exposed mice [407]. Consistently, evidence from our laboratory [87,178] supports a link between stress-induced microglial activation and structural plasticity that ultimately modulates cocaine-induced behavioral responses.
Delving into the features of both stress- and drug-induced alterations in synaptic plasticity, it is important to emphasize that the glutamatergic postsynapse represents a critical convergence point for mechanisms underlying stress- and addiction-related adaptations in MSNs of the NAc. Dendritic spine remodeling—reflected in the balance between mature and immature spines—serves as a structural correlate of experience-dependent synaptic strength [17,408,409,410]. Drugs of abuse, such as cocaine, robustly increase dendritic spine density in NAc MSNs [411,412,413,414,415]. These structural changes are reflected in enhanced actin cytoskeleton remodeling, increased surface expression of AMPA (GluR1) receptors, and elevated AMPA/NMDA receptor ratios—synaptic adaptations that underlie potentiated excitatory transmission [17,171,416,417,418]. Following prolonged abstinence, re-exposure to psychostimulants or drug-associated cues induces a rapid and transient enlargement of dendritic spine heads in NAc MSNs, accompanied by dynamic changes in actin polymerization and AMPA receptor trafficking; these effects typically normalize within approximately two hours [419,420,421].
Similarly, stress induces convergent structural adaptations within the NAc core, including increased densities of total and mushroom-type spines, with specific patterns depending on the stress paradigm, MSNs subtype, and timing of assessment [86,394,422,423]. Larger “mushroom” spines form stronger excitatory synapses [424] and are associated with enhanced AMPA receptor insertion and remodeling of the postsynaptic density [425,426,427,428,429,430]. Chronic stress also elevates basal extracellular glutamate levels [145,345,346] and promotes cytoskeletal remodeling, possibly as a homeostatic compensatory adaptation [17,431]. Within this framework, stress may “prime” the NAc core by expanding the pool of structurally mature spines, thereby facilitating subsequent cocaine-induced synaptic potentiation. This primed state may enable cocaine exposure to transform latent structural potential into functional plasticity through AMPA receptor trafficking and insertion into the postsynaptic membrane [86,131,132,154,398].
Microglial sensing of glutamate can drive cytokine-mediated forms of synaptic plasticity. Microglia express functional glutamate receptors [343,344] and are capable of detecting even subtle fluctuations in extracellular glutamate as signals of alarm or danger, leading to hyper-ramification, process extension, and proliferation [20,231,347,348]. Glutamate-induced microglial activation promotes the release of several signaling molecules [350,351,353,354,432,433]. Among these, TNF-α and IL-1β exert modulatory effects on glutamate homeostasis both directly at the postsynaptic level and indirectly through glial-mediated mechanisms involving microglia–astrocyte crosstalk. Concerning their direct actions on postsynaptic function, these pro-inflammatory cytokines influence long-term potentiation (LTP) and synaptic scaling. TNF-α, for instance, enhances synaptic efficacy by increasing surface AMPA receptor expression, whereas blockade of TNF-α signaling reduces synaptic strength [434]. Moreover, TNF-α mediates homeostatic synaptic scaling, adjusting synaptic weights to stabilize network activity following prolonged activity blockade [212]. Nonetheless, TNF-α appears to be dispensable for acute forms of plasticity, as most studies report no significant effect on LTP after TNF-α manipulation [213,435].
Elevated IL-1β levels, in turn, affect both the induction and maintenance phases of LTP [436,437]. Importantly, IL-1β can disrupt neuronal function and synaptic plasticity by interfering with both LTP and long-term depression (LTD), two complementary mechanisms underlying learning and memory [438,439]. Mechanistically, IL-1β selectively regulates AMPA receptor phosphorylation and surface expression through extracellular calcium- and NMDA receptor-dependent signaling in hippocampal neurons [440]. Additionally, IL-1β induces the loss of PSD-95—a scaffold protein critical for synapse maturation and stability—in neuronal cultures [441]. Altogether, these findings underscore the central role of inflammatory cytokines in regulating neuronal plasticity, bridging adaptive homeostatic processes with maladaptive neuroinflammatory responses.
These inflammatory mediators also play pivotal roles in microglia–astrocyte crosstalk, which drives a complex dysregulation of glutamate homeostasis with enduring postsynaptic consequences that underlie stress- and drug-related behavioral adaptations. TNF-α has been implicated in the disruption of glutamate balance by (1) downregulating astrocytic GLT-1 via NF-κB activation and (2) promoting postsynaptic AMPA receptor insertion, thereby altering synaptic efficacy [212,214,216,366,367,368,369,370,378]. Similarly, IL-1β has been shown to contribute to glutamate imbalance and excitotoxicity [362,372].
Crucially, Avalos et al. (2022) [87] demonstrated that chronic restraint stress is associated with increased NAc core microglial hyper-ramification and elevated mushroom-type spine density, along with higher TNF-α (mRNA/protein) levels and decreased GLT-1 and GFAP expression—changes consistent with astrocytic hypoactivity. Daily minocycline administration prevented all these adaptations, indicating a microglia-dependent cascade linking stress to glutamatergic dysregulation and structural remodeling, ultimately facilitating cocaine self-administration.
Collectively, these findings support a mechanistic framework in which chronic stress engages microglia–astrocyte interactions to regulate and sense extracellular glutamate while promoting the release of proinflammatory cytokines—particularly TNF-α and IL-1β. Through these inflammatory and non-inflammatory pathways, microglia contribute to both homeostatic and maladaptive forms of plasticity that drive stress-induced vulnerability to drug addiction.
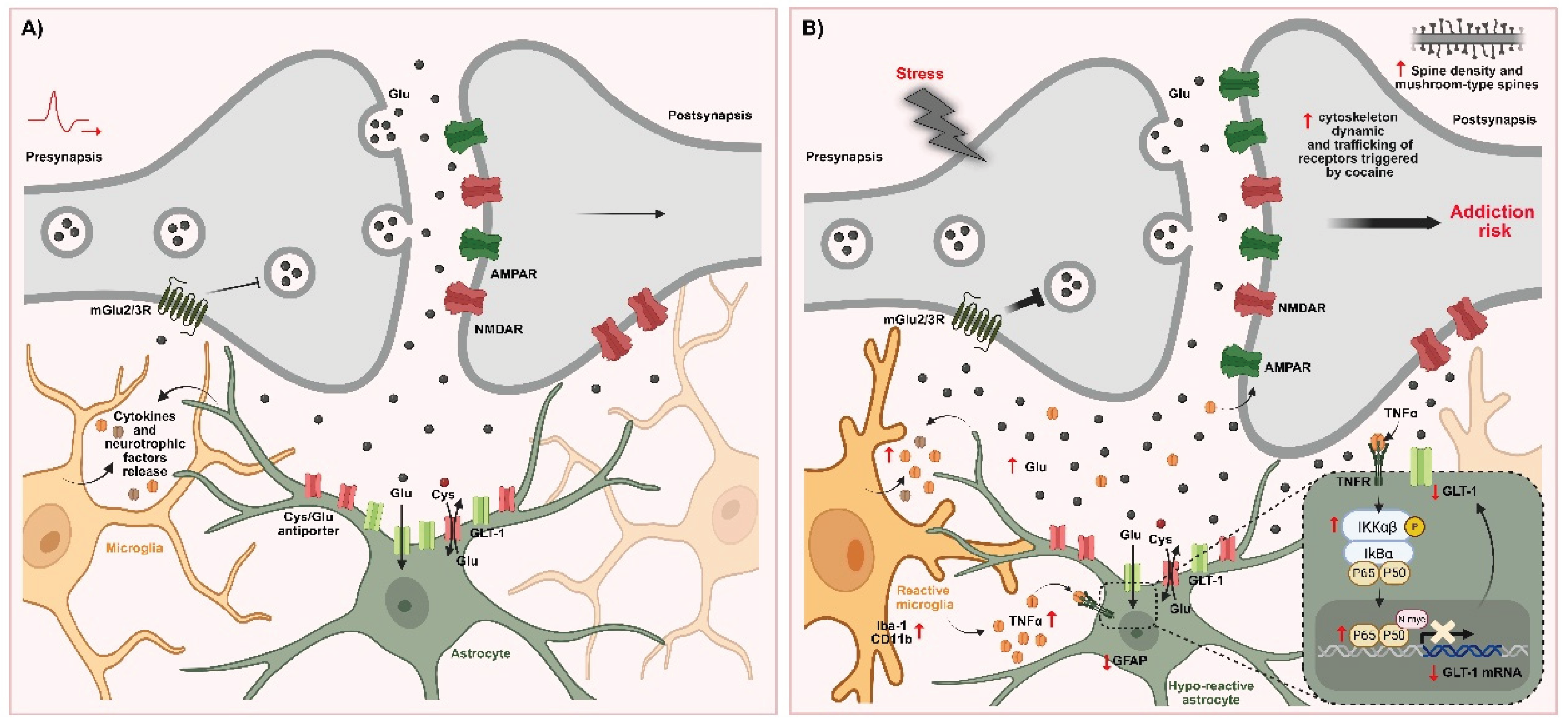
Figure 2.
Microglia–Astrocyte Crosstalk in Stress-Induced Vulnerability to Cocaine Use Disorder. Panel a) depicts a tripartite synapse within the NAc core under control conditions, illustrating the normal interactions among presynaptic terminals, postsynaptic medium spiny neurons, and perisynaptic astrocytic processes that collectively maintain the glutamate homeostasis in the region. Panel b) illustrates the cascade of events initiated by chronic stress, which elevates extracellular glutamate in the NAc core and activates microglia through ionotropic and metabotropic receptors. Activated microglia undergo hyper-ramification and release proinflammatory cytokines, primarily TNF-α. This cytokine promotes the nuclear recruitment of NF-κB, resulting in transcriptional repression of astrocytic GLT-1. TNF-α also diminishes GFAP-associated structural integrity, further compromising glutamate clearance. The resulting glutamate spillover drives extrasynaptic NMDA/mGluR activation, leading to postsynaptic remodeling of MSNs, characterized by increased mushroom-type dendritic spines and increased AMPA receptor trafficking. These adaptations heighten vulnerability to cocaine reinforcement and relapse-like behaviors, which can be prevented by pharmacological restoration of microglial homeostasis with minocycline. This figure was created by BioRender.com.
Figure 2.
Microglia–Astrocyte Crosstalk in Stress-Induced Vulnerability to Cocaine Use Disorder. Panel a) depicts a tripartite synapse within the NAc core under control conditions, illustrating the normal interactions among presynaptic terminals, postsynaptic medium spiny neurons, and perisynaptic astrocytic processes that collectively maintain the glutamate homeostasis in the region. Panel b) illustrates the cascade of events initiated by chronic stress, which elevates extracellular glutamate in the NAc core and activates microglia through ionotropic and metabotropic receptors. Activated microglia undergo hyper-ramification and release proinflammatory cytokines, primarily TNF-α. This cytokine promotes the nuclear recruitment of NF-κB, resulting in transcriptional repression of astrocytic GLT-1. TNF-α also diminishes GFAP-associated structural integrity, further compromising glutamate clearance. The resulting glutamate spillover drives extrasynaptic NMDA/mGluR activation, leading to postsynaptic remodeling of MSNs, characterized by increased mushroom-type dendritic spines and increased AMPA receptor trafficking. These adaptations heighten vulnerability to cocaine reinforcement and relapse-like behaviors, which can be prevented by pharmacological restoration of microglial homeostasis with minocycline. This figure was created by BioRender.com.
9. Repurposing Glutamatergic Therapies for the Treatment of Substance Use Disorders Comorbidity
In recent years, research on treatments for stress and SUDs has grown substantially. Current initiatives continue to explore and assess innovative therapeutic options, including both psychological interventions and pharmacological strategies. There is increasing interest in discovering pharmacological treatments that can reduce the symptoms associated with PTSD and SUDs. For alcohol use disorder, the U.S. Food and Drug Administration (FDA) have approved medications such as naltrexone, acamprosate, and disulfiram. In the case of opioid use disorder, approved options include buprenorphine, methadone, and extended-release naltrexone. Regarding PTSD, the FDA has authorized two selective serotonin reuptake inhibitors—paroxetine and sertraline— for clinical use. To date, no pharmacological treatment has been approved for cocaine addiction, or for the comorbid presentation of PTSD, despite the high clinical need. Efforts to develop effective pharmacotherapies for this comorbidity have focused on evaluating compounds that target shared neurobiological mechanisms underlying both disorders. Thus, in recent years, the repurposing of drugs that modulate glutamatergic transmission has emerged as a promising therapeutic avenue for treating the comorbidity between cocaine addiction and stress-related disorders. In the following section, we discuss how this approach has expanded to include pharmacotherapies aimed at modulating glial components as a potential strategy for addressing comorbid PTSD and SUDs, with a particular focus on cocaine.
9.1. N-Acetylcysteine (NAC)
N-acetylcysteine (chemical formula C₅H₉NO₃S) (NAC) is a cysteine prodrug. For more than 30 years, NAC has been used clinically as an antidote for acetaminophen poisoning [442]. More recently, its therapeutic use has expanded to include mucolytic therapy for a range of respiratory disorders.
NAC exerts its effects in the brain through various mechanisms. As an acetylated derivative of cysteine, it has good bioavailability and can cross the blood–brain barrier. Cystine acts as the substrate for the system Xc⁻, which facilitates the import of cystine into the cell in exchange for glutamate. Through this exchange, the transporter plays a key role in regulating extracellular glutamate levels. Once internalized, cystine is reduced to cysteine, the rate-limiting precursor for the synthesis of glutathione (GSH), a crucial endogenous antioxidant [443]. Several preclinical studies have demonstrated that oral administration of NAC increases glutathione levels in the brain [444,445]. Numerous preclinical and clinical studies report elevated oxidative stress following exposure to various drugs of abuse, including psychostimulants [189,446,447,448]. Accordingly, strategies aimed at enhancing glutathione synthesis, such as NAC, have been investigated as potential therapeutic agents, given their ability to restore redox balance in the brain [444,445].
Due to its action on system Xc⁻ capable of influencing extracellular glutamate levels, another proposed mechanism involves the modulation of neurotransmission, which is commonly altered across various psychiatric pathologies. NAC has shown potential in regulating not only glutamatergic system but also dopaminergic transmission. Since system Xc⁻ and glutamate transporter work in coordinated manner, deficits in system Xc⁻ or GLT-1 expression have been associated with excessive synaptic glutamate levels, which in turn can lead to reduced mGlu2/3 receptor signaling as occurs after exposure to stress and addictive drugs as described in earlier sections. Thus, NAC treatment has been consistently investigated in preclinical studies as a potential therapy for cocaine dependence, and its evaluation has also extended to promising clinical trials.
In preclinical models, pretreatment with NAC (60 mg/kg, i.p.) -administered after each self-administration session or prior to each injection during a 7-day non-contingent cocaine regimen- effectively prevented cocaine-induced escalation of drug intake, behavioral sensitization, and drug-primed reinstatement [449]. Importantly, NAC (30-90 mg/kg) reduce cocaine seeking in a dose-dependent manner during both early and late addiction stages, without affecting cocaine reinforcement, supporting its potential for relapse prevention [450]. However, when administered after the establishment of escalated cocaine intake, chronic NAC treatment (60mg/kg) had no measurable effect on further escalation or on motivation for cocaine under a progressive ratio schedule. Instead, NAC promoted abstinence by increasing sensitivity to punishment, suggesting a selective role in restoring behavioral control in the face of adverse consequences [451]. Consistent with these behavioral findings, NAC restores glutamate homeostasis by enhancing the activity of the cystine–glutamate exchanger, thereby normalizing extracellular glutamate levels in animals with a history of drug exposure [161]. Moreover, administration of NAC (100 mg/kg; i.p.) for five consecutive days prior to reinstatement testing attenuated both stress-enhanced alcohol and cocaine consumption, as well as the reinstatement of alcohol- and cocaine-seeking behaviors triggered by conditioned stress cues in rats [452].
Mechanistically, stimulation of mGluR2/3 receptors by non-synaptic glutamate derived from the cystine-glutamate exchanger inhibits synaptic glutamate release and, consequently, excitatory synaptic activity, an effect believed to underlie the ability of NAC to suppress cocaine-associated behaviors [161]. Accordingly, NAC treatment prevented the persistent reduction in system Xc⁻ activity, preserving cystine transport, basal extracellular glutamate levels, and cocaine-evoked glutamate release in the NAc [453]. NAC (33 mg/kg, i.p., administered 20 min prior to a cocaine priming injection) also restored synaptic plasticity in the NAc of cocaine-exposed rats by stimulating mGluR2/3 and mGluR5, reversing cocaine-induced metaplasticity and supporting its anti-relapse effects [454]. Consistently, administration of the mGluR2/3 antagonist LY341495 blocked the inhibitory effect of NAC on reinstatement in animals trained to self-administer cocaine [455]. In addition to its action on the cystine-glutamate exchange, evidence suggests that the restoration of GLT-1 may also constitute a key mechanism by which repeated NAC treatment reduces cue-induced cocaine reinstatement [173].
Several clinical studies have explored the effects of NAC on cocaine use, with mixed results. Early short-term, placebo-controlled trials in hospitalized, non-treatment-seeking cocaine-dependent individuals found that NAC (1200 mg/day) reduced cue-reactivity and showed trends toward decreased craving and withdrawal symptoms, although some effects were not statistically significant, likely due to small sample sizes [456,457]. Open-label studies further demonstrated that NAC was well tolerated at doses up to 3600 mg/day and associated with reduced self-reported cocaine use and craving [458,459]. Neuroimaging studies indicated that a single dose of NAC normalized elevated glutamate levels in the dorsal anterior cingulate cortex of cocaine users, suggesting that NAC may restore glutamate homeostasis linked to impulsivity [460]. However, a larger double-blind placebo-controlled trial in 111 treatment-seeking adults did not find significant effects of NAC on abstinence, although subgroup analysis revealed that NAC increased time to relapse and reduced craving in already abstinent individuals [461], supporting the idea that NAC may be more effective as a relapse prevention strategy rather than a treatment for ongoing use. Recently, a randomized, placebo-controlled crossover trial demonstrated that short-term NAC administration (2400 mg/day for 2 days) significantly reduced cocaine cue-related reactivity in the superior prefrontal cortex of individuals with cocaine use disorder. Notably, this effect occurred without changes in subjective craving [462].
In line with findings from preclinical research, a double-blind, 8-week clinical trial was conducted in veterans (n = 35) diagnosed with PTSD and SUDs, who received either NAC (2.4 g/day) or placebo in combination with cognitive-behavioral therapy for SUDs. NAC treatment led to reductions in PTSD symptoms, as reported by both the participants and clinicians, as well as in drug craving. Notably, these improvements persisted for at least one month after discontinuation of the pharmacological treatment [463].
As another relevant mechanism, NAC may play a role in modulating immune responses. NAC modulates the expression and secretion of inflammatory cytokines under various inflammatory conditions both in vitro and in vivo. As a precursor of GSH and direct antioxidant, NAC inhibits NF-κB activation and reduces the expression of pro-inflammatory genes. It significantly suppresses IL-1β release induced by lipopolysaccharides. Additionally, NAC reverses cytokine alterations triggered by social isolation, including elevated IL-6 and IL-4 and reduced TNF-α and IFN-γ levels [464,465,466]. As extensively discussed in this review, increased levels of inflammatory mediators—such as TNF-α/ NF-κB signaling—have been reported in individuals with comorbid stress and SUDs. Considering that both neuroinflammation and stress-induced adaptations at glutamatergic synapses play a key role in the pathophysiology of PTSD and SUDs [467], NAC may hold therapeutic potential for treating this comorbid condition, potentially also by modulating its neuroinflammatory component.
9.2. Ceftriaxone
Ceftriaxone is a third-generation cephalosporin antibiotic known not only for its antimicrobial properties but also for its ability to modulate the glutamatergic system within the CNS. Cephalosporins, derived from the fungus Acremonium, belong to the β-lactam class of antibiotics, characterized by a β-lactam ring, and are widely used to treat infections caused by both Gram-positive and Gram-negative bacteria [468]. Ceftriaxone is capable of crossing the blood–brain barrier, making it suitable for the treatment of CNS infections, such as meningitis [469].
Rothstein et al., 2005 [470] was the first to identify β-lactam antibiotics as potent inducers of GLT-1 expression, primarily by promoting its gene transcription. In this study, ceftriaxone-induced upregulation of astrocyte GLT-1 expression and functional activity conferred neuroprotective effects in both in vitro and in vivo models of ischemia and amyotrophic lateral sclerosis (ALS), two conditions characterized by glutamate-induced toxicity.
Given the capacity of ceftriaxone to restore GLT-1 levels in the NAc, resulted in a reduction in drug-seeking behavior in animals with a history of cocaine intake [162]. Specifically, ceftriaxone treatment (200 mg/kg/day for 7 consecutive days), administered following each extinction session, attenuates the reinstatement of cocaine-seeking behavior induced by either cocaine-associated cues or the drug. Importantly, this effect appears to be specific to cocaine-exposed animals, as ceftriaxone does not enhance GLT-1 expression in the NAc core of drug-naïve rats. The ability of ceftriaxone to modulate reinstatement behavior and GLT-1 expression after cocaine exposure is dose-dependent. Doses of 100 and 200 mg/kg effectively elevate GLT-1 levels and reduce cue-induced reinstatement, whereas a lower dose of 50 mg/kg given over 5 days fails to produce these effects [471]. Supporting this, glutamate uptake studies have shown that administering 200 mg/kg of ceftriaxone for 5 days restores both GLT-1 and also system Xc⁻ activity in the NAc core following cocaine self-administration [163]. This restoration leads to elevated basal extracellular glutamate and a reduction in glutamate release during cocaine-primed reinstatement. The enhancement of glutamate clearance prevents extracellular glutamate overflow, thereby attenuating drug-induced neuroplasticity and behavioral sensitization [17].
Interestingly, although ceftriaxone significantly reduced reinstatement even weeks after treatment had ended its administration for 5 days prior to the initiation of cocaine self-administration did not affect the acquisition of drug-taking behavior. Moreover, ceftriaxone attenuated the locomotor response to the first cocaine injection and prevented the development of cocaine sensitization [472]. These findings extend to behavioral studies, in which chronic ceftriaxone administration has been shown to attenuate psychomotor sensitization induced by amphetamine [473]. Ceftriaxone reduces cocaine-seeking in rats after 21 to 45 days of abstinence without extinction, increasing GLT-1 expression in both the NAc core and shell [474,475]. However, only GLT-1 in the core is necessary to attenuate cocaine relapse [475]. At doses of 100 and 200 mg/kg, ceftriaxone decreases context-induced relapse, but only 200 mg/kg elevates GLT-1 in the core [474]. Thus, GLT-1 regulation in the core is critical for its protective effects in the absence of extinction. In line with this preclinical data, findings from our lab demonstrated that ceftriaxone restores NAc core GLT-1 expression and function and reduces stress stress-induced locomotion in rats exposed to acute and chronic immobilization stress, although it does not reverse the stress induced increases in AMPA)/NMDA ratio [86,153]. Importantly, the restoration of glutamate transport in the NAc core by ceftriaxone prevented the capacity of acute stress to augment the acquisition of cocaine self-administration [86]. Mechanistically, the upregulation of GLT-1 expression by ceftriaxone is mediated through the canonical NF-κB transcriptional pathway [476]. Consistent with this mechanism, data from our laboratory indicate that genetic inhibition of NF-κB activity in the NAc core is sufficient to restore GLT-1 levels reduced by chronic stress exposure and to prevent the stress-induced facilitation of cocaine self-administration [178]. Despite its preclinical efficacy, ceftriaxone has not yet been clinically evaluated for the treatment of stress and comorbid substance use disorder in a consistent way.
9.3. Minocycline
Minocycline is a second-generation tetracycline antibiotic and has been explored as a potential treatment for a wide range of CNS disorders, including schizophrenia, depression, stroke, Parkinson’s disease, and multiple sclerosis [477,478,479,480,481,482,483,484,485]. Minocycline confers neuroprotection primarily through the inhibition of microglial activation through multiple mechanisms [486], including the suppression of the pro-inflammatory NF-κB intracellular signaling pathway [487]. It has been shown, for instance, to markedly reduce LPS-induced microglial activation—as indicated by Iba1 expression—and to decrease the release of pro-inflammatory cytokines such as IL-6 [488,489], TNF-α [490], and chemokines like CCL2 [491]. Moreover, minocycline engages additional molecular targets, further contributing to its broad therapeutic potential. For example, minocycline inhibits pro-apoptotic enzymes such as caspase-1 and caspase-3, thereby preventing neuronal cell death, as shown in a transgenic mouse model of Huntington’s disease where minocycline delayed disease progression by suppressing caspase activation [492]. Additionally, minocycline inhibits matrix metalloproteinases (MMPs), particularly MMP-12, which are involved in blood-brain barrier breakdown and neuroinflammation [493].
Based on its pharmacological profile, several studies have evaluated its therapeutic potential in the context of psychostimulant use disorders. Thus, minocycline (40 mg/kg) reduced hyperlocomotion in mice following a single methamphetamine administration (3 mg/kg) and attenuated the development of behavioral sensitization induced by repeated exposure to the drug (3 mg/kg/day for 5 days) [494]. This effect appears to be dose-dependent, as lower doses (10 and 20 mg/kg) were ineffective. Importantly, the effective 40 mg/kg dose did not produce locomotor effects on its own, indicating that the attenuation was not due to general sedation or motor impairment. Moreover, minocycline may attenuate methamphetamine-induced neurotoxicity, as evidenced by its ability to reduce the depletion of striatal DA and its major metabolite, 3,4-dihydroxyphenylacetic acid (DOPAC), following repeated methamphetamine administration, as well as by preventing the loss of DA transporter (DAT) immunoreactivity in the striatum [494]. In line with this report, a previous study showed that a high dose of minocycline (100 mg/kg, s.c.) reduced the locomotor-stimulating effects of a moderate dose of amphetamine (0.5 mg/kg, i.p.) in rats [495]. Similarly, minocycline has been shown to suppress methamphetamine-induced glial activation and reduce intravenous methamphetamine self-administration in Long-Evans rats. When administered intraperitoneally once daily at doses of 10, 30, or 60 mg/kg for three consecutive days during self-administration sessions, minocycline significantly decreased responding maintained by the 0.03 mg/kg/infusion methamphetamine dose under a fixed ratio 1 (FR1) schedule [401].
Minocycline interferes with the development of cocaine-induced locomotor sensitization but not with its expression in male C57BL/6J mice. A high dose (80 mg/kg) reduced locomotor activity both when given alone and when administered 30 minutes before cocaine. Conversely, a lower dose (40 mg/kg), administered 3 hours prior to cocaine, prevented the progressive increase in locomotor activity typically observed after repeated cocaine exposure, without affecting the acute locomotor response [496]. The ability of minocycline to inhibit cocaine-induced locomotor sensitization appears to depend on the 5-lipoxygenase (5-LOX) pathway, as minocycline failed to prevent sensitization or alter GluR1 phosphorylation in the frontal cortex of 5-LOX-deficient mice [497].
The inhibition of microglia reactivity by minocycline treatment (40 mg/kg/day) administered three days prior stress exposure, significantly alleviates PTSD-related anxiety and contextual fear in mice [498]. Similarly, minocycline (35mg/kg) attenuated anxious-like behaviors in stress-exposed rats and stress-induced elevations of pro-inflammatory cytokines (IL-1, IL-6, TNF-α) in the hippocampus, frontal cortex, and hypothalamus [499]. Consistent with these findings, our laboratory demonstrated that minocycline (30 mg/kg, i.p., every 12 hours for 5 days), when administered 16 days after the initial stress episode, prevents chronic restraint stress-induced increase in vulnerability to cocaine self-administration, along with the associated glutamatergic and immune alterations [87].
Clinical research on minocycline for PTSD and anxiety related-disorders, with or without comorbid depression, is still limited but promising. In a small open-label pilot study, minocycline (100 – 200 mg/day) reduced PTSD symptoms and lowered CRP levels, although IL-6 and TNF-α responses were variable, suggesting complex inflammatory kinetics [500]. In patients with comorbid anxiety and depression, minocycline showed a trend toward reducing anxiety symptoms without significantly affecting depression scores in individuals primarily diagnosed with Major Depressive Disorder, with effects persisting post-treatment [501]. Another randomized controlled trial supported anxiolytic and antidepressant benefits in generalized anxiety disorder [502], though other trials in treatment-resistant depression showed no overall effect unless stratified by inflammation levels, which revealed improved antidepressant but not anxiolytic responses [503].
While these preclinical and clinical findings support the therapeutic promise of minocycline in modulating neuroinflammation and glutamatergic imbalance associated with stress-related SUDs, its clinical efficacy in addressing addiction and stress comorbidity remains to be determined.
9.4. Ampicillin/Sulbactam (AMP/SUL)
Sulbactam is a β-lactamase inhibitor that lacks intrinsic antibacterial activity when used alone. Recent studies have demonstrated that sulbactam can upregulate GLT-1 expression and enhance glutamate uptake, thereby exerting neuroprotective effects in models of cerebral ischemia [504,505]). Moreover, sulbactam protects neurons from the combined neurotoxicity of amyloid beta and glutamate by upregulating GLT1 expression, highlighting its potential as a therapeutic agent for Alzheimer’s disease by ameliorating glutamate overload [506].
On the other hand, ampicillin, a β-lactam antibiotic previously identified as a GLT-1 upregulator [470], has also been evaluated in preclinical studies for its therapeutic potential in various SUDs, particularly alcohol consumption [507,508]. Specifically, ampicillin (100 mg/kg, i.p.), administered for 5 consecutive days to rats with high ethanol intake, effectively reduced alcohol consumption and restored GLT-1 levels in the NAc and PFC. This effect was mediated through AKT phosphorylation, a signaling mechanism previously implicated in the upregulation of GLT-1 [507]. Notably, ampicillin also increased the expression of xCT and both GLT-1 isoforms (GLT-1a and GLT-1b) in the NAc, further supporting its potential to modulate glutamate homeostasis and attenuate ethanol consumption [508]. The combination of ampicillin and sulbactam (AMP/SUL) exerts synergistic effects by enhancing glutamate transporter regulation and reducing neuroinflammation [509]. Thus, AMP/SUL (200 mg/kg i.p.) reduced cocaine-induced reinstatement [510]. These effects were linked to restoration of GLT-1 and xCT in the NAc core and shell, together with mGluR1 upregulation in the NAc. Similar results were observed in a model examining the facilitatory effect of cocaine on ethanol relapse-like behavior in male alcohol-preferring rats [511], further highlighting the effect of AMP/SUL on glutamatergic signaling during cocaine and ethanol co-exposure [511].
To date, no clinical trials in humans have evaluated the use of AMP/SUL for the treatment of comorbidity between stress and SUDs, or for any specific SUDs. The evidence available regarding AMP/SUL in this context is limited to preclinical research. Moreover, its effects have not been assessed in preclinical models of comorbidity, nor in models of stress or post-traumatic stress. Nevertheless, its potential to modulate glutamatergic signaling in brain reward areas makes it a promising target for further investigation as a therapeutic approach for these disorders.
10. Conclusions
Microglia-astrocyte interaction emerges as a pivotal cellular substrate linking stress and addiction through both inflammatory and non-inflammatory mechanisms. Stressors and drugs of abuse consistently induce microglial activation, triggering pro-inflammatory cytokine release, glutamate dysregulation, and dendritic spine remodeling. These effects converge in the NAc core, where microglial sensitivity to extracellular glutamate appears particularly important for amplifying stress-induced neuroadaptations that favor drug seeking. Importantly, chronic stress primes microglia into a hyper-ramified state, sensitizing them to subsequent cocaine exposure and promoting exaggerated inflammatory and synaptic responses—a mechanism that may underlie heightened addiction vulnerability following prior stress.
In parallel, addictive substances directly engage microglial-astrocyte communication. Cocaine, ethanol, nicotine, and psychostimulants induce morphological and inflammatory changes across reward-related brain regions, highlighting microglia as common effectors of drug-induced neuroadaptations. Notably, studies combining chronic stress with cocaine self-administration demonstrate that microglia mediate the escalation of intake and associated molecular and structural plasticity. Pharmacological inhibition with minocycline reverses these alterations, reinforcing the translational potential of targeting microglial function.
Beyond inflammatory effects, microglia also regulates neuronal plasticity through non-inflammatory mechanisms such as synaptic pruning, gliotransmission, and trophic support. These functions are particularly relevant as maladaptive synaptic plasticity represents a shared substrate for both stress and SUDs. Taken together, microglia occupy a central position at the intersection of stress and addiction neurobiology, and future studies should clarify the balance between their inflammatory and non-inflammatory roles, regional specificity, and the contribution of peripheral immune cells.
10.1. Mechanistic Synthesis
- -
- Stress-related vulnerability to cocaine use disorders arises from the interplay of HPA axis activation, neuroimmune signaling, and glial dysfunction.
- -
- Microglial TNF-α release and subsequent downregulation of astrocytic GLT-1 emerge as central pathways linking stress to glutamatergic dysregulation in the NAc core.
- -
- Glial cross-talk between microglia and astrocytes critically shapes synaptic plasticity, structural remodeling, and ultimately behavioral vulnerability to cocaine.
10.2. Weaknesses and Knowledge Gaps
- -
- Most preclinical evidence derives from male rodents; studies in females are limited despite evidence that gonadal hormones (e.g., estrogens) modulate neurobiological processes underlying stress and drug responses.
- -
- Clinical validation of glutamate-modulating drugs (minocycline, ceftriaxone, NAC, AMP/SUL) remains scarce, leaving a translational gap between promising preclinical findings and patient applications.
- -
- Integration of mechanistic rodent data with human neuroimaging and biomarker studies remains insufficient.
10.3. Future Directions
- -
- Mechanistic studies dissecting inflammatory versus non-inflammatory microglial functions, as well as peripheral–central immune crosstalk, in the comorbidity between stress and SUDs.
- -
- Systematic inclusion of females in preclinical models to capture sex-specific neuroimmune and glutamatergic adaptations.
- -
- Electrophysiological studies should determine whether the microglial and structural alterations induced by chronic stress and cocaine are accompanied by changes in neuronal excitability and synaptic transmission within the NAc, and define the temporal window and mechanisms through which microglial modulation can restore glutamatergic balance and circuit function.
- -
- Clinical trials assessing glial-targeting interventions, stratified by stress history and sex hormone status.
- -
- Early intervention strategies for stress-exposed populations aimed at modulating neuroimmune and glutamatergic mechanisms before maladaptive plasticity becomes established.
Author Contributions
Liliana Marina Cancela: Conceptualization; Formal Analysis; Investigation; Methodology; Resources; Data Curation; Funding Acquisition; Supervision; Visualization; Writing-Original Draft; Writing-Review & Editing. Bethania Mongi-Bragato: Conceptualization; Methodology; Investigation; Resources; Funding Acquisition; Writing-Draft of Section 7.2, 7.3.1, and 9; Writing-Review & Editing. María Paula Ávalos: Conceptualization; Methodology; Investigation; Resources; Visualization (Graphical Abstract, Figure 1 and Figure 2); Writing-Draft of Section 6.1, 6.2, and 7.1; Writing-Review & Editing. Flavia Andrea Bollati: Methodology; Investigation; Resources; Funding Acquisition; Bibliography Management (verification and integration of all references into the journal template); Review & Editing. All authors have read and agreed to the published version of the manuscript.
Funding
The authors acknowledge research funding from FONCyT BID PICT 2019–3560 (to LMC), PUE 22920180100029CO (to LMC), SECyT Res., 411/18 (to LMC), PRIMAR 2019 Res. 34,020,190,100,036 CB (to LMC), PIP Grant CONICET Res., 2021–1639 (to LMC), SECyT 337 202,301 00073CB, (to BMB), SECyT 33620230100084CB (to FAB), FONCyT BID PICT 2020–3875 (to FAB). Apart from providing financial support, all these institutions had no further impact in this study and played no role in manuscript preparation or submission.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
During the preparation of this study, the authors used ChatGPT (OpenAI, GPT-5.1) exclusively to assist with English grammar supervision. The authors have critically reviewed, revised and incorporated all the AI-assisted suggestions, and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AMP/SULB | Ampicillin/Sulbactam |
| GFAP | Glial fibrillary acidic protein |
| GLT-1 | Glutamate transporter-1 |
| IL-1β | Interleukin 1 beta |
| LTP | long-term potentiation |
| LTD | long term depression |
| mGluR2/3 | presynaptic metabotropic glutamate receptors 2/3 |
| MSNs | Medium Spiny Neurons |
| NAC | N-Acetylcysteine |
| NAc | Nucleus Accumbens |
| N-myc | N-myc proto-oncogene protein |
| PTSD | Post-Traumatic Stress Disorder |
| SUDs | Substance Use Disorders |
| TNF-α | Tumor Necrosis Factor-alpha |
References
- McEwen, B.S. Protective and Damaging Effects of the Mediators of Stress and Adaptation: Allostasis and Allostatic Load. In Allostasis, Homeostasis, and the Costs of Physiological Adaptation; Schulkin, J., Ed.; Cambridge University Press, 2004; pp. 65–98. ISBN 978-0-521-81141-5. [Google Scholar]
- McEwen, B.S. Physiology and Neurobiology of Stress and Adaptation: Central Role of the Brain. Physiological Reviews 2007, 87, 873–904. [Google Scholar] [CrossRef]
- Koob, G.F.; Volkow, N.D. Neurobiology of Addiction: A Neurocircuitry Analysis. The Lancet Psychiatry 2016, 3, 760–773. [Google Scholar] [CrossRef]
- Belin, D.; Belin-Rauscent, A.; Murray, J.E.; Everitt, B.J. Addiction: Failure of Control over Maladaptive Incentive Habits. Current Opinion in Neurobiology 2013, 23, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Camí, J.; Farré, M. Drug Addiction. N Engl J Med 2003, 349, 975–986. [Google Scholar] [CrossRef] [PubMed]
- Brady, K.T.; Sinha, R. Co-Occurring Mental and Substance Use Disorders: The Neurobiological Effects of Chronic Stress. AJP 2005, 162, 1483–1493. [Google Scholar] [CrossRef]
- Breslau, N.; Davis, G.C.; Schultz, L.R. Posttraumatic Stress Disorder and the Incidence of Nicotine, Alcohol, and Other Drug Disorders in Persons Who Have Experienced Trauma. Arch Gen Psychiatry 2003, 60, 289. [Google Scholar] [CrossRef]
- Mills, K.L.; Teesson, M.; Ross, J.; Peters, L. Trauma, PTSD, and Substance Use Disorders: Findings From the Australian National Survey of Mental Health and Well-Being. AJP 2006, 163, 652–658. [Google Scholar] [CrossRef]
- Ouimette, P.; Coolhart, D.; Funderburk, J.S.; Wade, M.; Brown, P.J. Precipitants of First Substance Use in Recently Abstinent Substance Use Disorder Patients with PTSD. Addictive Behaviors 2007, 32, 1719–1727. [Google Scholar] [CrossRef]
- Piazza, P.V.; Deminiere, J.M.; Le Moal, M.; Simon, H. Stress- and Pharmacologically-Induced Behavioral Sensitization Increases Vulnerability to Acquisition of Amphetamine Self-Administration. Brain Research 1990, 514, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Boyson, C.O.; Miguel, T.T.; Quadros, I.M.; DeBold, J.F.; Miczek, K.A. Prevention of Social Stress-Escalated Cocaine Self-Administration by CRF-R1 Antagonist in the Rat VTA. Psychopharmacology 2011, 218, 257–269. [Google Scholar] [CrossRef]
- Mueller, D.; Stewart, J. Cocaine-Induced Conditioned Place Preference: Reinstatement by Priming Injections of Cocaine after Extinction. Behavioural Brain Research 2000, 115, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Boyson, C.O.; Holly, E.N.; Shimamoto, A.; Albrechet-Souza, L.; Weiner, L.A.; DeBold, J.F.; Miczek, K.A. Social Stress and CRF–Dopamine Interactions in the VTA: Role in Long-Term Escalation of Cocaine Self-Administration. J. Neurosci. 2014, 34, 6659–6667. [Google Scholar] [CrossRef]
- Sinha, R. How Does Stress Increase Risk of Drug Abuse and Relapse? Psychopharmacology 2001, 158, 343–359. [Google Scholar] [CrossRef]
- Sinha, R.; Garcia, M.; Paliwal, P.; Kreek, M.J.; Rounsaville, B.J. Stress-Induced Cocaine Craving and Hypothalamic-Pituitary-Adrenal Responses Are Predictive of Cocaine Relapse Outcomes. Arch Gen Psychiatry 2006, 63, 324. [Google Scholar] [CrossRef]
- Koob, G.F.; Schulkin, J. Addiction and Stress: An Allostatic View. Neuroscience & Biobehavioral Reviews 2019, 106, 245–262. [Google Scholar] [CrossRef]
- Kalivas, P.W. The Glutamate Homeostasis Hypothesis of Addiction. Nat Rev Neurosci 2009, 10, 561–572. [Google Scholar] [CrossRef]
- Myers-Schulz, B.; Koenigs, M. Functional Anatomy of Ventromedial Prefrontal Cortex: Implications for Mood and Anxiety Disorders. Mol Psychiatry 2012, 17, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Woodburn, S.C.; Bollinger, J.L.; Wohleb, E.S. The Semantics of Microglia Activation: Neuroinflammation, Homeostasis, and Stress. J Neuroinflammation 2021, 18, 258. [Google Scholar] [CrossRef] [PubMed]
- Mayhew, J.; Beart, P.M.; Walker, F.R. Astrocyte and Microglial Control of Glutamatergic Signalling: A Primer on Understanding the Disruptive Role of Chronic Stress. J Neuroendocrinology 2015, 27, 498–506. [Google Scholar] [CrossRef]
- Lacagnina, M.J.; Rivera, P.D.; Bilbo, S.D. Glial and Neuroimmune Mechanisms as Critical Modulators of Drug Use and Abuse. Neuropsychopharmacol 2017, 42, 156–177. [Google Scholar] [CrossRef]
- Wang, J.; Chen, H.-S.; Li, H.-H.; Wang, H.-J.; Zou, R.-S.; Lu, X.-J.; Wang, J.; Nie, B.-B.; Wu, J.-F.; Li, S.; et al. Microglia-Dependent Excessive Synaptic Pruning Leads to Cortical Underconnectivity and Behavioral Abnormality Following Chronic Social Defeat Stress in Mice. Brain, Behavior, and Immunity 2023, 109, 23–36. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.-S.; Peterson, T.C.; et al. Neurotoxic Reactive Astrocytes Are Induced by Activated Microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef]
- Lewitus, G.M.; Konefal, S.C.; Greenhalgh, A.D.; Pribiag, H.; Augereau, K.; Stellwagen, D. Microglial TNF-α Suppresses Cocaine-Induced Plasticity and Behavioral Sensitization. Neuron 2016, 90, 483–491. [Google Scholar] [CrossRef]
- Robinson, T. The Neural Basis of Drug Craving: An Incentive-Sensitization Theory of Addiction. Brain Research Reviews 1993, 18, 247–291. [Google Scholar] [CrossRef]
- White, F.J.; Kalivas, P.W. Neuroadaptations Involved in Amphetamine and Cocaine Addiction. Drug and Alcohol Dependence 1998, 51, 141–153. [Google Scholar] [CrossRef]
- Nestler, E.J.; Aghajanian, G.K. Molecular and Cellular Basis of Addiction. Science 1997, 278, 58–63. [Google Scholar] [CrossRef]
- Kreek, M.J.; Nielsen, D.A.; Butelman, E.R.; LaForge, K.S. Genetic Influences on Impulsivity, Risk Taking, Stress Responsivity and Vulnerability to Drug Abuse and Addiction. Nat Neurosci 2005, 8, 1450–1457. [Google Scholar] [CrossRef] [PubMed]
- Müller, D.J.; Likhodi, O.; Heinz, A. Neural Markers of Genetic Vulnerability to Drug Addiction. In Behavioral Neuroscience of Drug Addiction;Current Topics in Behavioral Neurosciences; Self, D.W., Staley Gottschalk, J.K., Eds.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2010; Vol. 3, pp. 277–299. ISBN 978-3-642-03000-0. [Google Scholar]
- Sinha, R. Stress and Addiction: A Dynamic Interplay of Genes, Environment, and Drug Intake. Biological Psychiatry 2009, 66, 100–101. [Google Scholar] [CrossRef] [PubMed]
- Swendsen, J.; Conway, K.P.; Degenhardt, L.; Glantz, M.; Jin, R.; Merikangas, K.R.; Sampson, N.; Kessler, R.C. Mental Disorders as Risk Factors for Substance Use, Abuse and Dependence: Results from the 10-year Follow-up of the National Comorbidity Survey. Addiction 2010, 105, 1117–1128. [Google Scholar] [CrossRef] [PubMed]
- Boden, J.M.; Fergusson, D.M.; Horwood, L.J. Alcohol Misuse and Violent Behavior: Findings from a 30-Year Longitudinal Study. Drug and Alcohol Dependence 2012, 122, 135–141. [Google Scholar] [CrossRef]
- Clark, H.W.; Masson, C.L.; Delucchi, K.L.; Hall, S.M.; Sees, K.L. Violent Traumatic Events and Drug Abuse Severity. Journal of Substance Abuse Treatment 2001, 20, 121–127. [Google Scholar] [CrossRef]
- Montoya, I.D.; Covarrubias, L.D.; Patek, J.A.; Graves, J.A. Posttraumatic Stress Disorder Among Hispanic and African-American Drug Users. The American Journal of Drug and Alcohol Abuse 2003, 29, 729–741. [Google Scholar] [CrossRef]
- Seal, K.H. Bringing the War Back Home: Mental Health Disorders Among 103 788 US Veterans Returning From Iraq and Afghanistan Seen at Department of Veterans Affairs Facilities. Arch Intern Med 2007, 167, 476. [Google Scholar] [CrossRef] [PubMed]
- Seal, K.H.; Metzler, T.J.; Gima, K.S.; Bertenthal, D.; Maguen, S.; Marmar, C.R. Trends and Risk Factors for Mental Health Diagnoses Among Iraq and Afghanistan Veterans Using Department of Veterans Affairs Health Care, 2002–2008. Am J Public Health 2009, 99, 1651–1658. [Google Scholar] [CrossRef]
- Tuerk, P.W.; Grubaugh, A.L.; Hamner, M.B.; Foa, E.B. Diagnosis and Treatment of PTSD-Related Compulsive Checking Behaviors in Veterans of the Iraq War: The Influence of Military Context on the Expression of PTSD Symptoms. AJP 2009, 166, 762–767. [Google Scholar] [CrossRef] [PubMed]
- Ullman, S.E.; Relyea, M.; Peter-Hagene, L.; Vasquez, A.L. Trauma Histories, Substance Use Coping, PTSD, and Problem Substance Use among Sexual Assault Victims. Addictive Behaviors 2013, 38, 2219–2223. [Google Scholar] [CrossRef]
- Kalivas, P.W.; Volkow, N.D. The Neural Basis of Addiction: A Pathology of Motivation and Choice. AJP 2005, 162, 1403–1413. [Google Scholar] [CrossRef]
- O’Brien, C.P.; Childress, A.R.; Ehrman, R.; Robbins, S.J. Conditioning Factors in Drug Abuse: Can They Explain Compulsion? J Psychopharmacol 1998, 12, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E. Molecular Mechanisms of Drug Addiction [Published Erratum Appears in J Neurosci 1992 Aug;12(8):Following Table of Contents]. J. Neurosci. 1992, 12, 2439–2450. [Google Scholar] [CrossRef]
- White, N.M. Addictive Drugs as Reinforcers: Multiple Partial Actions on Memory Systems. Addiction 1996, 91, 921–949; discussion 951-965. [Google Scholar] [CrossRef]
- Nestler, E.J.; Aghajanian, G.K. Molecular and Cellular Basis of Addiction. Science 1997, 278, 58–63. [Google Scholar] [CrossRef]
- Nestler, E.J. Molecular Neurobiology of Addiction. American J Addict 2001, 10, 201–217. [Google Scholar] [CrossRef]
- Deminiere, J.M.; Piazza, P.V.; Le Moal, M.; Simon, H. Experimental Approach to Individual Vulnerability to Psychostimulant Addiction. Neuroscience & Biobehavioral Reviews 1989, 13, 141–147. [Google Scholar] [CrossRef]
- Piazza, P.V.; Le Moal, M. The Role of Stress in Drug Self-Administration. Trends in Pharmacological Sciences 1998, 19, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Le Moal, M. Plasticity of Reward Neurocircuitry and the “dark Side” of Drug Addiction. Nat Neurosci 2005, 8, 1442–1444. [Google Scholar] [CrossRef] [PubMed]
- Mews, P.; Cunningham, A.M.; Scarpa, J.; Ramakrishnan, A.; Hicks, E.M.; Bolnick, S.; Garamszegi, S.; Shen, L.; Mash, D.C.; Nestler, E.J. Convergent Abnormalities in Striatal Gene Networks in Human Cocaine Use Disorder and Mouse Cocaine Administration Models. Sci. Adv. 2023, 9, eadd8946. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Buck, C.L.; Cohen, A.; Edwards, S.; Park, P.E.; Schlosburg, J.E.; Schmeichel, B.; Vendruscolo, L.F.; Wade, C.L.; Whitfield, T.W.; et al. Addiction as a Stress Surfeit Disorder. Neuropharmacology 2014, 76, 370–382. [Google Scholar] [CrossRef]
- Nestler, E.J.; Lüscher, C. The Molecular Basis of Drug Addiction: Linking Epigenetic to Synaptic and Circuit Mechanisms. Neuron 2019, 102, 48–59. [Google Scholar] [CrossRef]
- Browne, C.J.; Futamura, R.; Minier-Toribio, A.; Hicks, E.M.; Ramakrishnan, A.; Martínez-Rivera, F.J.; Estill, M.; Godino, A.; Parise, E.M.; Torres-Berrío, A.; et al. Transcriptional Signatures of Heroin Intake and Relapse throughout the Brain Reward Circuitry in Male Mice. Sci. Adv. 2023, 9, eadg8558. [Google Scholar] [CrossRef]
- Kalivas, P.W.; Stewart, J. Dopamine Transmission in the Initiation and Expression of Drug- and Stress-Induced Sensitization of Motor Activity. Brain Research Reviews 1991, 16, 223–244. [Google Scholar] [CrossRef]
- Berridge, K.C.; Robinson, T.E. What Is the Role of Dopamine in Reward: Hedonic Impact, Reward Learning, or Incentive Salience? Brain Research Reviews 1998, 28, 309–369. [Google Scholar] [CrossRef]
- Koob, G.F.; Bloom, F.E. Cellular and Molecular Mechanisms of Drug Dependence. Science 1988, 242, 715–723. [Google Scholar] [CrossRef]
- Wise, R.A. Addictive Drugs and Brain Stimulation Reward. Annu. Rev. Neurosci. 1996, 19, 319–340. [Google Scholar] [CrossRef] [PubMed]
- Pierce, R.C.; Kalivas, P.W. A Circuitry Model of the Expression of Behavioral Sensitization to Amphetamine-like Psychostimulants. Brain Research Reviews 1997, 25, 192–216. [Google Scholar] [CrossRef]
- Pierce, R.C.; Kalivas, P.W. Repeated Cocaine Modifies the Mechanism by Which Amphetamine Releases Dopamine. J. Neurosci. 1997, 17, 3254–3261. [Google Scholar] [CrossRef]
- Robinson, T. The Neural Basis of Drug Craving: An Incentive-Sensitization Theory of Addiction. Brain Research Reviews 1993, 18, 247–291. [Google Scholar] [CrossRef]
- Robinson, T.E.; Berridge, K.C. Incentive-sensitization and Addiction. Addiction 2001, 96, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, M.; Le Moal, M.; Piazza, P.V. Sensitization to the Motor Effects of Contingent Infusions of Heroin but Not of κ Agonist RU 51599. Psychopharmacology 1998, 139, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Scofield, M.D.; Heinsbroek, J.A.; Gipson, C.D.; Kupchik, Y.M.; Spencer, S.; Smith, A.C.W.; Roberts-Wolfe, D.; Kalivas, P.W. The Nucleus Accumbens: Mechanisms of Addiction across Drug Classes Reflect the Importance of Glutamate Homeostasis. Pharmacological Reviews 2016, 68, 816–871. [Google Scholar] [CrossRef]
- Nall, R.W.; Heinsbroek, J.A.; Nentwig, T.B.; Kalivas, P.W.; Bobadilla, A. Circuit Selectivity in Drug versus Natural Reward Seeking Behaviors. Journal of Neurochemistry 2021, 157, 1450–1472. [Google Scholar] [CrossRef]
- Heimer, L.; Zahm, D.S.; Churchill, L.; Kalivas, P.W.; Wohltmann, C. Specificity in the Projection Patterns of Accumbal Core and Shell in the Rat. Neuroscience 1991, 41, 89–125. [Google Scholar] [CrossRef]
- Kelley, A.E. Ventral Striatal Control of Appetitive Motivation: Role in Ingestive Behavior and Reward-Related Learning. Neuroscience & Biobehavioral Reviews 2004, 27, 765–776. [Google Scholar] [CrossRef]
- Heimer, L.; Alheid, G.F.; de Olmos, J.S.; Groenewegen, H.J.; Haber, S.N.; Harlan, R.E.; Zahm, D.S. The Accumbens: Beyond the Core-Shell Dichotomy. J Neuropsychiatry Clin Neurosci 1997, 9, 354–381. [Google Scholar] [CrossRef]
- Ito, R.; Robbins, T.W.; Everitt, B.J. Differential Control over Cocaine-Seeking Behavior by Nucleus Accumbens Core and Shell. Nat Neurosci 2004, 7, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Bassareo, V.; Di Chiara, G. Differential Responsiveness of Dopamine Transmission to Food-Stimuli in Nucleus Accumbens Shell/Core Compartments. Neuroscience 1999, 89, 637–641. [Google Scholar] [CrossRef] [PubMed]
- Sellings, L.H.L.; Clarke, P.B.S. Segregation of Amphetamine Reward and Locomotor Stimulation between Nucleus Accumbens Medial Shell and Core. J. Neurosci. 2003, 23, 6295–6303. [Google Scholar] [CrossRef]
- Zahm, D.S.; Brog, J.S. On the Significance of Subterritories in the “Accumbens” Part of the Rat Ventral Striatum. Neuroscience 1992, 50, 751–767. [Google Scholar] [CrossRef] [PubMed]
- Di Ciano, P.; Cardinal, R.N.; Cowell, R.A.; Little, S.J.; Everitt, B.J. Differential Involvement of NMDA, AMPA/Kainate, and Dopamine Receptors in the Nucleus Accumbens Core in the Acquisition and Performance of Pavlovian Approach Behavior. J. Neurosci. 2001, 21, 9471–9477. [Google Scholar] [CrossRef]
- Di Chiara, G. Nucleus Accumbens Shell and Core Dopamine: Differential Role in Behavior and Addiction. Behavioural Brain Research 2002, 137, 75–114. [Google Scholar] [CrossRef]
- Meredith, G.E.; Baldo, B.A.; Andrezjewski, M.E.; Kelley, A.E. The Structural Basis for Mapping Behavior onto the Ventral Striatum and Its Subdivisions. Brain Struct Funct 2008, 213, 17–27. [Google Scholar] [CrossRef]
- McFarland, K.; Lapish, C.C.; Kalivas, P.W. Prefrontal Glutamate Release into the Core of the Nucleus Accumbens Mediates Cocaine-Induced Reinstatement of Drug-Seeking Behavior. J. Neurosci. 2003, 23, 3531–3537. [Google Scholar] [CrossRef]
- Kalivas, P.W.; Churchill, L.; Klitenick, M.A. The Circuitry Mediating the Translation of Motivational Stimuli Into Adaptive Motor Responses. In Limbic Motor Circuits and Neuropsychiatry; Kalivas, P.W., Barnes, C.D., Eds.; CRC Press, 2019; pp. 237–288. ISBN 978-0-429-27441-1. [Google Scholar]
- Kalivas, P.W. Neurotransmitter Regulation of Dopamine Neurons in the Ventral Tegmental Area. Brain Research Reviews 1993, 18, 75–113. [Google Scholar] [CrossRef]
- Koob, G.F.; Nestler, E.J. The Neurobiology of Drug Addiction. J Neuropsychiatry Clin Neurosci 1997, 9, 482–497. [Google Scholar] [CrossRef]
- Koob, G.F.; Moal, M.L. Drug Abuse: Hedonic Homeostatic Dysregulation. Science 1997, 278, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J. Is There a Common Molecular Pathway for Addiction? Nat Neurosci 2005, 8, 1445–1449. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wang, G.-J.; Fowler, J.S.; Tomasi, D.; Telang, F. Addiction: Beyond Dopamine Reward Circuitry. Proc. Natl. Acad. Sci. U.S.A. 2011, 108, 15037–15042. [Google Scholar] [CrossRef]
- Di Chiara, G.; Acquas, E.; Tanda, G.; Cadoni, C. Drugs of Abuse: Biochemical Surrogates of Specific Aspects of Natural Reward? Biochem Soc Symp 1993, 59, 65–81. [Google Scholar]
- Kalivas, P.W.; Duffy, P. Effect of Acute and Daily Cocaine Treatment on Extracellular Dopamine in the Nucleus Accumbens. Synapse 1990, 5, 48–58. [Google Scholar] [CrossRef]
- Sinha, R. Chronic Stress, Drug Use, and Vulnerability to Addiction. Annals of the New York Academy of Sciences 2008, 1141, 105–130. [Google Scholar] [CrossRef]
- Lu, L.; Shepard, J.D.; Scott Hall, F.; Shaham, Y. Effect of Environmental Stressors on Opiate and Psychostimulant Reinforcement, Reinstatement and Discrimination in Rats: A Review. Neuroscience & Biobehavioral Reviews 2003, 27, 457–491. [Google Scholar] [CrossRef] [PubMed]
- Nikulina, E.M.; Covington, H.E.; Ganschow, L.; Hammer, R.P.; Miczek, K.A. Long-Term Behavioral and Neuronal Cross-Sensitization to Amphetamine Induced by Repeated Brief Social Defeat Stress: Fos in the Ventral Tegmental Area and Amygdala. Neuroscience 2004, 123, 857–865. [Google Scholar] [CrossRef]
- Miczek, K.A.; Nikulina, E.M.; Shimamoto, A.; Covington, H.E. Escalated or Suppressed Cocaine Reward, Tegmental BDNF, and Accumbal Dopamine Caused by Episodic versus Continuous Social Stress in Rats. Journal of Neuroscience 2011, 31, 9848–9857. [Google Scholar] [CrossRef]
- Garcia-Keller, C.; Kupchik, Y.M.; Gipson, C.D.; Brown, R.M.; Spencer, S.; Bollati, F.; Esparza, M.A.; Roberts-Wolfe, D.J.; Heinsbroek, J.A.; Bobadilla, A.C.; et al. Glutamatergic Mechanisms of Comorbidity between Acute Stress and Cocaine Self-Administration. Molecular Psychiatry 2016, 21, 1063–1069. [Google Scholar] [CrossRef]
- Avalos, M.P.; Guzman, A.S.; Rigoni, D.; Gorostiza, E.A.; Sanchez, M.A.; Mongi-Bragato, B.; Garcia-Keller, C.; Perassi, E.M.; Virgolini, M.B.; Peralta Ramos, J.M.; et al. Minocycline Prevents Chronic Restraint Stress-Induced Vulnerability to Developing Cocaine Self-Administration and Associated Glutamatergic Mechanisms: A Potential Role of Microglia. Brain, Behavior, and Immunity 2022, 101, 359–376. [Google Scholar] [CrossRef]
- Shaham, Y.; Erb, S.; Stewart, J. Stress-Induced Relapse to Heroin and Cocaine Seeking in Rats: A Review. Brain Research Reviews 2000, 33, 13–33. [Google Scholar] [CrossRef]
- Goeders, N.E. Stress and Cocaine Addiction. Journal of Pharmacology and Experimental Therapeutics 2002, 301, 785–789. [Google Scholar] [CrossRef]
- Mantsch, J.R.; Baker, D.A.; Funk, D.; Lê, A.D.; Shaham, Y. Stress-Induced Reinstatement of Drug Seeking: 20 Years of Progress. Neuropsychopharmacol 2016, 41, 335–356. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F. Stress, Corticotropin-Releasing Factor, and Drug Addiction. Annals of the New York Academy of Sciences 1999, 897, 27–45. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Zorrilla, E.P. Neurobiological Mechanisms of Addiction: Focus on Corticotropin-Releasing Factor. Curr Opin Investig Drugs 2010, 11, 63–71. [Google Scholar] [PubMed]
- Blacktop, J.M.; Seubert, C.; Baker, D.A.; Ferda, N.; Lee, G.; Graf, E.N.; Mantsch, J.R. Augmented Cocaine Seeking in Response to Stress or CRF Delivered into the Ventral Tegmental Area Following Long-Access Self-Administration Is Mediated by CRF Receptor Type 1 But Not CRF Receptor Type 2. J. Neurosci. 2011, 31, 11396–11403. [Google Scholar] [CrossRef]
- Wang, B.; Shaham, Y.; Zitzman, D.; Azari, S.; Wise, R.A.; You, Z.-B. Cocaine Experience Establishes Control of Midbrain Glutamate and Dopamine by Corticotropin-Releasing Factor: A Role in Stress-Induced Relapse to Drug Seeking. J. Neurosci. 2005, 25, 5389–5396. [Google Scholar] [CrossRef]
- Bale, T.L.; Vale, W.W. CRF and CRF Receptors: Role in Stress Responsivity and Other Behaviors. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 525–557. [Google Scholar] [CrossRef]
- Swanson, L.W.; Sawchenko, P.E.; Rivier, J.; Vale, W.W. Organization of Ovine Corticotropin-Releasing Factor Immunoreactive Cells and Fibers in the Rat Brain: An Immunohistochemical Study. Neuroendocrinology 1983, 36, 165–186. [Google Scholar] [CrossRef] [PubMed]
- Sawchenko, P.E.; Imaki, T.; Potter, E.; Kovács, K.; Imaki, J.; Vale, W. The Functional Neuroanatomy of Corticotropin-Releasing Factor. Ciba Found Symp discussion 21-29. 1993, 172, 5–21. [Google Scholar] [CrossRef]
- Dunn, A.J.; Berridge, C.W.; Lai, Y.I.; Yachabach, T.L. CRF-Induced Excessive Grooming Behavior in Rats and Mice. Peptides 1987, 8, 841–844. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Heinrichs, S.C. A Role for Corticotropin Releasing Factor and Urocortin in Behavioral Responses to Stressors. Brain Research 1999, 848, 141–152. [Google Scholar] [CrossRef]
- Koob, G.F.; Weiss, F. Pharmacology of Drug Self-Administration. Alcohol 1990, 7, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Spanagel, R.; Weiss, F. The Dopamine Hypothesis of Reward: Past and Current Status. Trends in Neurosciences 1999, 22, 521–527. [Google Scholar] [CrossRef]
- Saal, D; Dong, Y; Bonci, A; Malenka, RC. Drugs of Abuse and Stress Trigger a Common Synaptic Adaptation in Dopamine Neurons. Neuron 2003, 37, 577–582. [Google Scholar] [CrossRef]
- Wanat, M.J.; Hopf, F.W.; Stuber, G.D.; Phillips, P.E.M.; Bonci, A. Corticotropin-releasing Factor Increases Mouse Ventral Tegmental Area Dopamine Neuron Firing through a Protein Kinase C-dependent Enhancement ofIh. The Journal of Physiology 2008, 586, 2157–2170. [Google Scholar] [CrossRef]
- Kalivas, P.W.; Duffy, P.; Latimer, L.G. Neurochemical and Behavioral Effects of Corticotropin-Releasing Factor in the Ventral Tegmental Area of the Rat. J Pharmacol Exp Ther 1987, 242, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Refojo, D.; Schweizer, M.; Kuehne, C.; Ehrenberg, S.; Thoeringer, C.; Vogl, A.M.; Dedic, N.; Schumacher, M.; Von Wolff, G.; Avrabos, C.; et al. Glutamatergic and Dopaminergic Neurons Mediate Anxiogenic and Anxiolytic Effects of CRHR1. Science 2011, 333, 1903–1907. [Google Scholar] [CrossRef]
- Antelman, S.M.; Eichler, A.J.; Black, C.A.; Kocan, D. Interchangeability of Stress and Amphetamine in Sensitization. Science 1980, 207, 329–331. [Google Scholar] [CrossRef] [PubMed]
- Piazza, P.Vincenzo.; Deminière, J.-Marie.; Le Moal, Michel.; Simon, Hervé. Factors That Predict Individual Vulnerability to Amphetamine Self-Administration. Science 1989, 245, 1511–1513. [Google Scholar] [CrossRef]
- Mantsch, J.; Ho, A.; Schlussman, S.; Kreek, M. Predictable Individual Differences in the Initiation of Cocaine Self-Administration by Rats under Extended-Access Conditions Are Dose-Dependent. Psychopharmacology 2001, 157, 31–39. [Google Scholar] [CrossRef]
- Piazza, P.V.; Maccari, S.; Deminière, J.M.; Le Moal, M.; Mormède, P.; Simon, H. Corticosterone Levels Determine Individual Vulnerability to Amphetamine Self-Administration. Proc. Natl. Acad. Sci. U.S.A. 1991, 88, 2088–2092. [Google Scholar] [CrossRef]
- Deroche, V.; Piazza, P.V.; Casolini, P.; Maccari, S.; Le Moal, M.; Simon, H. Stress-Induced Sensitization to Amphetamine and Morphine Psychomotor Effects Depend on Stress-Induced Corticosterone Secretion. Brain Research 1992, 598, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Deroche, V.; Piazza, P.V.; Maccari, S.; Le Moal, M.; Simon, H. Repeated Corticosterone Administration Sensitizes the Locomotor Response to Amphetamine. Brain Research 1992, 584, 309–313. [Google Scholar] [CrossRef]
- Mantsch, J.R.; Saphier, D.; Goeders, N.E. Corticosterone Facilitates the Acquisition of Cocaine Self-Administration in Rats: Opposite Effects of the Type II Glucocorticoid Receptor Agonist Dexamethasone. J Pharmacol Exp Ther 1998, 287, 72–80. [Google Scholar] [CrossRef]
- Goeders, N.E.; Guerin, G.F. Non-Contingent Electric Footshock Facilitates the Acquisition of Intravenous Cocaine Self-Administration in Rats. Psychopharmacology 1994, 114, 63–70. [Google Scholar] [CrossRef]
- Capriles, N.; Cancela, L.M. Effect of Acute and Chronic Stress Restraint on Amphetamine-Associated Place Preference: Involvement of Dopamine D1 and D2 Receptors. European Journal of Pharmacology 1999, 386, 127–134. [Google Scholar] [CrossRef]
- De Giovanni, L.N.; Guzman, A.S.; Virgolini, M.B.; Cancela, L.M. NMDA Antagonist MK 801 in Nucleus Accumbens Core but Not Shell Disrupts the Restraint Stress-Induced Reinstatement of Extinguished Cocaine-Conditioned Place Preference in Rats. Behavioural Brain Research 2016, 315, 150–159. [Google Scholar] [CrossRef]
- Guzman, A.S.; Avalos, M.P.; De Giovanni, L.N.; Euliarte, P.V.; Sanchez, M.A.; Mongi-Bragato, B.; Rigoni, D.; Bollati, F.A.; Virgolini, M.B.; Cancela, L.M. CB1R Activation in Nucleus Accumbens Core Promotes Stress-Induced Reinstatement of Cocaine Seeking by Elevating Extracellular Glutamate in a Drug-Paired Context. Sci Rep 2021, 11, 12964. [Google Scholar] [CrossRef]
- Shaham, Y.; Shalev, U.; Lu, L.; De Wit, H.; Stewart, J. The Reinstatement Model of Drug Relapse: History, Methodology and Major Findings. Psychopharmacology 2003, 168, 3–20. [Google Scholar] [CrossRef]
- Covington, H.; Miczek, K. Repeated Social-Defeat Stress, Cocaine or Morphine. Psychopharmacology 2001, 158, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Miczek, K.; Yap, J.; Covingtoniii, H. Social Stress, Therapeutics and Drug Abuse: Preclinical Models of Escalated and Depressed Intake. Pharmacology & Therapeutics 2008, 120, 102–128. [Google Scholar] [CrossRef] [PubMed]
- Engeln, M.; Fox, M.E.; Lobo, M.K. Housing Conditions during Self-Administration Determine Motivation for Cocaine in Mice Following Chronic Social Defeat Stress. Psychopharmacology 2021, 238, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Mantsch, J.R.; Katz, E.S. Elevation of Glucocorticoids Is Necessary but Not Sufficient for the Escalation of Cocaine Self-Administration by Chronic Electric Footshock Stress in Rats. Neuropsychopharmacol 2007, 32, 367–376. [Google Scholar] [CrossRef]
- Mantsch, J.R.; Yuferov, V.; Mathieu-Kia, A.-M.; Ho, A.; Kreek, M.J. Neuroendocrine Alterations in a High-Dose, Extended-Access Rat Self-Administration Model of Escalating Cocaine Use. Psychoneuroendocrinology 2003, 28, 836–862. [Google Scholar] [CrossRef]
- Vezina, P.; Giovino, A.A.; Wise, R.A.; Stewart, J. Environment-Specific Cross-Sensitization between the Locomotor Activating Effects of Morphine and Amphetamine. Pharmacology Biochemistry and Behavior 1989, 32, 581–584. [Google Scholar] [CrossRef]
- Borowsky, B.; Kuhn, C.M. Chronic Cocaine Administration Sensitizes Behavioral but Not Neuroendocrine Responses. Brain Research 1991, 543, 301–306. [Google Scholar] [CrossRef]
- Cadoni, C.; Di Chiara, G. Differential Changes in Accumbens Shell and Core Dopamine in Behavioral Sensitization to Nicotine. European Journal of Pharmacology 2000, 387, R23–R25. [Google Scholar] [CrossRef]
- Cadoni, C.; Pisanu, A.; Solinas, M.; Acquas, E.; Chiara, G. Behavioural Sensitization after Repeated Exposure to Δ 9 -Tetrahydrocannabinol and Cross-Sensitization with Morphine. Psychopharmacology 2001, 158, 259–266. [Google Scholar] [CrossRef]
- Quadros, P.S.; Pfau, J.L.; Goldstein, A.Y.N.; De Vries, G.J.; Wagner, C.K. Sex Differences in Progesterone Receptor Expression: A Potential Mechanism for Estradiol-Mediated Sexual Differentiation. Endocrinology 2002, 143, 3727–3739. [Google Scholar] [CrossRef]
- Paulson, P.E.; Robinson, T.E. Amphetamine-Induced Time-dependent Sensitization of Dopamine Neurotransmission in the Dorsal and Ventral Striatum: A Microdialysis Study in Behaving Rats. Synapse 1995, 19, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Post, R.M.; Weiss, S.R. Psychomotor Stimulant vs. Local Anesthetic Effects of Cocaine: Role of Behavioral Sensitization and Kindling. NIDA Res Monogr 1988, 88, 217–238. [Google Scholar]
- Valjent, E.; Bertran-Gonzalez, J.; Aubier, B.; Greengard, P.; Hervé, D.; Girault, J.-A. Mechanisms of Locomotor Sensitization to Drugs of Abuse in a Two-Injection Protocol. Neuropsychopharmacol 2010, 35, 401–415. [Google Scholar] [CrossRef]
- Esparza, M.A.; Bollati, F.; Garcia-Keller, C.; Virgolini, M.B.; Lopez, L.M.; Brusco, A.; Shen, H.-W.; Kalivas, P.W.; Cancela, L.M. Stress-Induced Sensitization to Cocaine: Actin Cytoskeleton Remodeling within Mesocorticolimbic Nuclei. European Journal of Neuroscience 2012, 36. [Google Scholar] [CrossRef]
- Garcia-Keller, C.; Martinez, S.A.A.; Esparza, M.A.A.; Bollati, F.; Kalivas, P.W.W.; Cancela, L.M.M. Cross-Sensitization between Cocaine and Acute Restraint Stress Is Associated with Sensitized Dopamine but Not Glutamate Release in the Nucleus Accumbens. European Journal of Neuroscience 2013, 37, 982–995. [Google Scholar] [CrossRef] [PubMed]
- Deroche, V.; Marinelli, M.; Maccari, S.; Le Moal, M.; Simon, H.; Piazza, P. Stress-Induced Sensitization and Glucocorticoids. I. Sensitization of Dopamine-Dependent Locomotor Effects of Amphetamine and Morphine Depends on Stress-Induced Corticosterone Secretion. J. Neurosci. 1995, 15, 7181–7188. [Google Scholar] [CrossRef] [PubMed]
- Pacchioni, A.M.; Cador, M.; Bregonzio, C.; Cancela, L.M. A Glutamate–Dopamine Interaction in the Persistent Enhanced Response to Amphetamine in Nucleus Accumbens Core but Not Shell Following a Single Restraint Stress. Neuropsychopharmacol 2007, 32, 682–692. [Google Scholar] [CrossRef]
- Kõiv, K.; Vares, M.; Kroon, C.; Metelitsa, M.; Tiitsaar, K.; Laugus, K.; Jaako, K.; Harro, J. Effect of Chronic Variable Stress on Sensitization to Amphetamine in High and Low Sucrose-Consuming Rats. J Psychopharmacol 2019, 33, 1512–1523. [Google Scholar] [CrossRef] [PubMed]
- Sorg, B.A.; Kalivas, P.W. Effects of Cocaine and Footshock Stress on Extracellular Dopamine Levels in the Ventral Striatum. Brain Research 1991, 559, 29–36. [Google Scholar] [CrossRef]
- Kippin, T.E.; Szumlinski, K.K.; Kapasova, Z.; Rezner, B.; See, R.E. Prenatal Stress Enhances Responsiveness to Cocaine. Neuropsychopharmacol 2008, 33, 769–782. [Google Scholar] [CrossRef]
- Di Chiara, G. The Role of Dopamine in Drug Abuse Viewed from the Perspective of Its Role in Motivation. Drug and Alcohol Dependence 1995, 38, 95–137. [Google Scholar] [CrossRef]
- Robinson, T.E.; Jurson, P.A.; Bennett, J.A.; Bentgen, K.M. Persistent Sensitization of Dopamine Neurotransmission in Ventral Striatum (Nucleus Accumbens) Produced by Prior Experience with (+)-Amphetamine: A Microdialysis Study in Freely Moving Rats. Brain Research 1988, 462, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Bonci, A.; Malenka, R.C. Properties and Plasticity of Excitatory Synapses on Dopaminergic and GABAergic Cells in the Ventral Tegmental Area. J. Neurosci. 1999, 19, 3723–3730. [Google Scholar] [CrossRef] [PubMed]
- Hahn, J.; Hopf, F.W.; Bonci, A. Chronic Cocaine Enhances Corticotropin-Releasing Factor-Dependent Potentiation of Excitatory Transmission in Ventral Tegmental Area Dopamine Neurons. Journal of Neuroscience 2009, 29, 6535–6544. [Google Scholar] [CrossRef]
- Ungless, M.A.; Singh, V.; Crowder, T.L.; Yaka, R.; Ron, D.; Bonci, A. Corticotropin-Releasing Factor Requires CRF Binding Protein to Potentiate NMDA Receptors via CRF Receptor 2 in Dopamine Neurons. Neuron 2003, 39, 401–407. [Google Scholar] [CrossRef]
- Ungless, M.A.; Argilli, E.; Bonci, A. Effects of Stress and Aversion on Dopamine Neurons: Implications for Addiction. Neuroscience and Biobehavioral Reviews 2010, 35, 151–156. [Google Scholar] [CrossRef]
- Beckstead, M.J.; Phillips, T.J. Mice Selectively Bred for High- or Low-Alcohol-Induced Locomotion Exhibit Differences in Dopamine Neuron Function. The Journal of Pharmacology and Experimental Therapeutics 2009, 329, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, B. Stress Preferentially Increases Extraneuronal Levels of Excitatory Amino Acids in the Prefrontal Cortex: Comparison to Hippocampus and Basal Ganglia. Journal of Neurochemistry 1993, 60, 1650–1657. [Google Scholar] [CrossRef]
- Kalivas, P.W.; Duffy, P. Selective Activation of Dopamine Transmission in the Shell of the Nucleus Accumbens by Stress. Brain Research 1995, 675, 325–328. [Google Scholar] [CrossRef]
- Reid, M.S.; Berger, S.P. Evidence for Sensitization of Cocaine-Induced Nucleus Accumbens Glutamate Release. NeuroReport 1996, 7, 1325–1329. [Google Scholar] [CrossRef]
- Reid, M.S.; Hsu, K.; Berger, S.P. Cocaine and Amphetamine Preferentially Stimulate Glutamate Release in the Limbic System: Studies on the Involvement of Dopamine. Synapse 1997, 27, 95–105. [Google Scholar] [CrossRef]
- Groenewegen, H.J.; Wright, C.I.; Beijer, A.V.J.; Voorn, P. Convergence and Segregation of Ventral Striatal Inputs and Outputs. Annals of the New York Academy of Sciences 1999, 877, 49–63. [Google Scholar] [CrossRef]
- Sesack, S.R.; Deutch, A.Y.; Roth, R.H.; Bunney, B.S. Topographical Organization of the Efferent Projections of the Medial Prefrontal Cortex in the Rat: An Anterograde Tract-tracing Study with Phaseolus Vulgaris Leucoagglutinin. J of Comparative Neurology 1989, 290, 213–242. [Google Scholar] [CrossRef]
- Wright, C.I.; Groenewegen, H.J. Patterns of Overlap and Segregation between Insular Cortical, Intermediodorsal Thalamic and Basal Amygdaloid Afferents in the Nucleus Accumbens of the Rat. Neuroscience 1996, 73, 359–373. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, B. Stress Activation of Glutamate Neurotransmission in the Prefrontal Cortex: Implications for Dopamine-Associated Psychiatric Disorders. Biological Psychiatry 2002, 51, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Avalos, M.P.; Guzman, A.S.; Garcia-Keller, C.; Mongi-Bragato, B.; Esparza, M.A.; Rigoni, D.; Sanchez, M.A.; Calfa, G.D.; Bollati, F.A.; Cancela, L.M. Impairment of Glutamate Homeostasis in the Nucleus Accumbens Core Underpins Cross-Sensitization to Cocaine Following Chronic Restraint Stress. Front. Physiol. 2022, 13, 896268. [Google Scholar] [CrossRef]
- Rigoni, D.; Avalos, M.P.; Boezio, M.J.; Guzmán, A.S.; Calfa, G.D.; Perassi, E.M.; Pierotti, S.M.; Bisbal, M.; Garcia-Keller, C.; Cancela, L.M.; et al. Stress-Induced Vulnerability to Develop Cocaine Addiction Depends on Cofilin Modulation. Neurobiology of Stress 2021, 15, 100349. [Google Scholar] [CrossRef]
- McFarland, K.; Davidge, S.B.; Lapish, C.C.; Kalivas, P.W. Limbic and Motor Circuitry Underlying Footshock-Induced Reinstatement of Cocaine-Seeking Behavior. J. Neurosci. 2004, 24, 1551–1560. [Google Scholar] [CrossRef]
- Campioni, M.R.; Xu, M.; McGehee, D.S. Stress-Induced Changes in Nucleus Accumbens Glutamate Synaptic Plasticity. Journal of Neurophysiology 2009, 101, 3192–3198. [Google Scholar] [CrossRef]
- Pierce, R.; Bell, K.; Duffy, P.; Kalivas, P. Repeated Cocaine Augments Excitatory Amino Acid Transmission in the Nucleus Accumbens Only in Rats Having Developed Behavioral Sensitization. J. Neurosci. 1996, 16, 1550–1560. [Google Scholar] [CrossRef]
- Kalivas, P.W.; Duffy, P. Repeated Cocaine Administration Alters Extracellular Glutamate in the Ventral Tegmental Area. Journal of Neurochemistry 1998, 70, 1497–1502. [Google Scholar] [CrossRef]
- Pendyam, S.; Mohan, A.; Kalivas, P.W.; Nair, S.S. Computational Model of Extracellular Glutamate in the Nucleus Accumbens Incorporates Neuroadaptations by Chronic Cocaine. Neuroscience 2009, 158, 1266–1276. [Google Scholar] [CrossRef]
- Baker, D.A.; Xi, Z.-X.; Shen, H.; Swanson, C.J.; Kalivas, P.W. The Origin and Neuronal Function of In Vivo Nonsynaptic Glutamate. J. Neurosci. 2002, 22, 9134–9141. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.A.; McFarland, K.; Lake, R.W.; Shen, H.; Tang, X.-C.; Toda, S.; Kalivas, P.W. Neuroadaptations in Cystine-Glutamate Exchange Underlie Cocaine Relapse. Nat Neurosci 2003, 6, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Knackstedt, L.A.; Moussawi, K.; Lalumiere, R.; Schwendt, M.; Klugmann, M.; Kalivas, P.W. Extinction Training after Cocaine Self-Administration Induces Glutamatergic Plasticity to Inhibit Cocaine Seeking. Journal of Neuroscience 2010, 30, 7984–7992. [Google Scholar] [CrossRef]
- Trantham-Davidson, H.; LaLumiere, R.T.; Reissner, K.J.; Kalivas, P.W.; Knackstedt, L.A. Ceftriaxone Normalizes Nucleus Accumbens Synaptic Transmission, Glutamate Transport, and Export Following Cocaine Self-Administration and Extinction Training. Journal of Neuroscience 2012, 32, 12406–12410. [Google Scholar] [CrossRef] [PubMed]
- Moussawi, K.; Zhou, W.; Shen, H.; Reichel, C.M.; See, R.E.; Carr, D.B.; Kalivas, P.W. Reversing Cocaine-Induced Synaptic Potentiation Provides Enduring Protection from Relapse. Proceedings of the National Academy of Sciences 2011, 108, 385–390. [Google Scholar] [CrossRef]
- Pow, D.V. Visualising the Activity of the Cystine-glutamate Antiporter in Glial Cells Using Antibodies to Aminoadipic Acid, a Selectively Transported Substrate. Glia 2001, 34, 27–38. [Google Scholar] [CrossRef]
- Sato, H.; Tamba, M.; Okuno, S.; Sato, K.; Keino-Masu, K.; Masu, M.; Bannai, S. Distribution of Cystine/Glutamate Exchange Transporter, System x(c)-, in the Mouse Brain. J Neurosci 2002, 22, 8028–8033. [Google Scholar] [CrossRef] [PubMed]
- Zink, M.; Vollmayr, B.; Gebicke-Haerter, P.J.; Henn, F.A. Reduced Expression of Glutamate Transporters vGluT1, EAAT2 and EAAT4 in Learned Helpless Rats, an Animal Model of Depression. Neuropharmacology 2010, 58, 465–473. [Google Scholar] [CrossRef]
- Almeida, R.F.; Thomazi, A.P.; Godinho, G.F.; Saute, J.A.M.; Wofchuk, S.T.; Souza, D.O.; Ganzella, M. Effects of Depressive-Like Behavior of Rats on Brain Glutamate Uptake. Neurochem Res 2010, 35, 1164–1171. [Google Scholar] [CrossRef]
- Rappeneau, V.; Blaker, A.; Petro, J.R.; Yamamoto, B.K.; Shimamoto, A. Disruption of the Glutamate–Glutamine Cycle Involving Astrocytes in an Animal Model of Depression for Males and Females. Front. Behav. Neurosci. 2016, 10. [Google Scholar] [CrossRef]
- Das, S.C.; Yamamoto, B.K.; Hristov, A.M.; Sari, Y. Ceftriaxone Attenuates Ethanol Drinking and Restores Extracellular Glutamate Concentration through Normalization of GLT-1 in Nucleus Accumbens of Male Alcohol-Preferring Rats. Neuropharmacology 2015, 97, 67–74. [Google Scholar] [CrossRef]
- Gipson, C.D.; Reissner, K.J.; Kupchik, Y.M.; Smith, A.C.W.; Stankeviciute, N.; Hensley-Simon, M.E.; Kalivas, P.W. Reinstatement of Nicotine Seeking Is Mediated by Glutamatergic Plasticity. Proc Natl Acad Sci U S A 2013, 110, 9124–9129. [Google Scholar] [CrossRef] [PubMed]
- Melendez, R.I.; Hicks, M.P.; Cagle, S.S.; Kalivas, P.W. Ethanol Exposure Decreases Glutamate Uptake in the Nucleus Accumbens. Alcoholism Clin & Exp Res 2005, 29, 326–333. [Google Scholar] [CrossRef]
- Reissner, K.J.; Gipson, C.D.; Tran, P.K.; Knackstedt, L.A.; Scofield, M.D.; Kalivas, P.W. Glutamate Transporter GLT-1 Mediates N-acetylcysteine Inhibition of Cocaine Reinstatement. Addiction Biology 2015, 20, 316–323. [Google Scholar] [CrossRef]
- Sari, Y.; Sreemantula, S.N. Neuroimmunophilin GPI-1046 Reduces Ethanol Consumption in Part through Activation of GLT1 in Alcohol-Preferring Rats. Neuroscience 2012, 227, 327–335. [Google Scholar] [CrossRef]
- Shen, H.; Scofield, M.D.; Boger, H.; Hensley, M.; Kalivas, P.W. Synaptic Glutamate Spillover Due to Impaired Glutamate Uptake Mediates Heroin Relapse. J. Neurosci. 2014, 34, 5649–5657. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Ng, J.P.; Li, Y.; Wolf, M.E. Acute and Repeated Systemic Amphetamine Administration: Effects on Extracellular Glutamate, Aspartate, and Serine Levels in Rat Ventral Tegmental Area and Nucleus Accumbens. Journal of Neurochemistry 1996, 67, 352–363. [Google Scholar] [CrossRef] [PubMed]
- Fischer-Smith, K.D.; Houston, A.C.W.; Rebec, G.V. Differential Effects of Cocaine Access and Withdrawal on Glutamate Type 1 Transporter Expression in Rat Nucleus Accumbens Core and Shell. Neuroscience 2012, 210, 333–339. [Google Scholar] [CrossRef]
- Mongi-Bragato, B.; Sánchez, M.A.; Avalos, M.P.; Boezio, M.J.; Guzman, A.S.; Rigoni, D.; Perassi, E.M.; Mas, C.R.; Bisbal, M.; Bollati, F.A.; et al. Activation of Nuclear Factor-Kappa B in the Nucleus Accumbens Core Is Necessary for Chronic Stress-Induced Glutamate and Neuro-Immune Alterations That Facilitate Cocaine Self-Administration. Brain, Behavior, and Immunity 2025, 128, 1–15. [Google Scholar] [CrossRef]
- Minelli, A.; Barbaresi, P.; Reimer, R.J.; Edwards, R.H.; Conti, F. The Glial Glutamate Transporter GLT-1 Is Localized Both in the Vicinity of and at Distance from Axon Terminals in the Rat Cerebral Cortex. Neuroscience 2001, 108, 51–59. [Google Scholar] [CrossRef]
- Cholet, N. Similar Perisynaptic Glial Localization for the Na+,K+-ATPase Alpha2 Subunit and the Glutamate Transporters GLAST and GLT-1 in the Rat Somatosensory Cortex. Cerebral Cortex 2002, 12, 515–525. [Google Scholar] [CrossRef]
- Moran, M.M.; McFarland, K.; Melendez, R.I.; Kalivas, P.W.; Seamans, J.K. Cystine/Glutamate Exchange Regulates Metabotropic Glutamate Receptor Presynaptic Inhibition of Excitatory Transmission and Vulnerability to Cocaine Seeking. J. Neurosci. 2005, 25, 6389–6393. [Google Scholar] [CrossRef]
- Béchade, C.; Cantaut-Belarif, Y.; Bessis, A. Microglial Control of Neuronal Activity. Front. Cell. Neurosci. 2013, 7. [Google Scholar] [CrossRef]
- Lynch, M.A. The Multifaceted Profile of Activated Microglia. Mol Neurobiol 2009, 40, 139–156. [Google Scholar] [CrossRef] [PubMed]
- Kreutzberg, G.W. Microglia: A Sensor for Pathological Events in the CNS. Trends in Neurosciences 1996, 19, 312–318. [Google Scholar] [CrossRef]
- Mcgeer, P.; Mcgeer, E. The Inflammatory Response System of Brain: Implications for Therapy of Alzheimer and Other Neurodegenerative Diseases. Brain Research Reviews 1995, 21, 195–218. [Google Scholar] [CrossRef]
- Kettenmann, H.; Hanisch, U.-K.; Noda, M.; Verkhratsky, A. Physiology of Microglia. Physiological Reviews 2011, 91, 461–553. [Google Scholar] [CrossRef] [PubMed]
- Davalos, D.; Grutzendler, J.; Yang, G.; Kim, J.V.; Zuo, Y.; Jung, S.; Littman, D.R.; Dustin, M.L.; Gan, W.-B. ATP Mediates Rapid Microglial Response to Local Brain Injury in Vivo. Nat Neurosci 2005, 8, 752–758. [Google Scholar] [CrossRef]
- Walker, F.; Nilsson, M.; Jones, K. Acute and Chronic Stress-Induced Disturbances of Microglial Plasticity, Phenotype and Function. CDT 2013, 14, 1262–1276. [Google Scholar] [CrossRef]
- Walker, F.R.; Beynon, S.B.; Jones, K.A.; Zhao, Z.; Kongsui, R.; Cairns, M.; Nilsson, M. Dynamic Structural Remodelling of Microglia in Health and Disease: A Review of the Models, the Signals and the Mechanisms. Brain, Behavior, and Immunity 2014, 37, 1–14. [Google Scholar] [CrossRef]
- Jinno, S.; Fleischer, F.; Eckel, S.; Schmidt, V.; Kosaka, T. Spatial Arrangement of Microglia in the Mouse Hippocampus: A Stereological Study in Comparison with Astrocytes. Glia 2007, 55, 1334–1347. [Google Scholar] [CrossRef]
- Stevens, B.; Allen, N.J.; Vazquez, L.E.; Howell, G.R.; Christopherson, K.S.; Nouri, N.; Micheva, K.D.; Mehalow, A.K.; Huberman, A.D.; Stafford, B.; et al. The Classical Complement Cascade Mediates CNS Synapse Elimination. Cell 2007, 131, 1164–1178. [Google Scholar] [CrossRef] [PubMed]
- Paolicelli, R.C.; Bolasco, G.; Pagani, F.; Maggi, L.; Scianni, M.; Panzanelli, P.; Giustetto, M.; Ferreira, T.A.; Guiducci, E.; Dumas, L.; et al. Synaptic Pruning by Microglia Is Necessary for Normal Brain Development. Science 2011, 333, 1456–1458. [Google Scholar] [CrossRef] [PubMed]
- Delpech, J.-C.; Madore, C.; Nadjar, A.; Joffre, C.; Wohleb, E.S.; Layé, S. Microglia in Neuronal Plasticity: Influence of Stress. Neuropharmacology 2015, 96, 19–28. [Google Scholar] [CrossRef]
- Blank, T.; Prinz, M. Microglia as Modulators of Cognition and Neuropsychiatric Disorders. Glia 2013, 61, 62–70. [Google Scholar] [CrossRef]
- Kopec, A.M.; Smith, C.J.; Ayre, N.R.; Sweat, S.C.; Bilbo, S.D. Microglial Dopamine Receptor Elimination Defines Sex-Specific Nucleus Accumbens Development and Social Behavior in Adolescent Rats. Nat Commun 2018, 9, 3769. [Google Scholar] [CrossRef] [PubMed]
- Gongwer, M.W.; Etienne, F.; Moca, E.N.; Chappell, M.S.; Blagburn-Blanco, S.V.; Riley, J.P.; Enos, A.S.; Haratian, M.; Qi, A.; Rojo, R.; et al. Microglia Regulate Nucleus Accumbens Synaptic Development and Circuit Function Underlying Threat Avoidance Behaviors 2025.
- Tremblay, M.-È.; Lowery, R.L.; Majewska, A.K. Microglial Interactions with Synapses Are Modulated by Visual Experience. PLoS Biol 2010, 8, e1000527. [Google Scholar] [CrossRef]
- Tremblay, M.-È.; Stevens, B.; Sierra, A.; Wake, H.; Bessis, A.; Nimmerjahn, A. The Role of Microglia in the Healthy Brain: Figure 1. J. Neurosci. 2011, 31, 16064–16069. [Google Scholar] [CrossRef] [PubMed]
- Schafer, D.P.; Lehrman, E.K.; Kautzman, A.G.; Koyama, R.; Mardinly, A.R.; Yamasaki, R.; Ransohoff, R.M.; Greenberg, M.E.; Barres, B.A.; Stevens, B. Microglia Sculpt Postnatal Neural Circuits in an Activity and Complement-Dependent Manner. Neuron 2012, 74, 691–705. [Google Scholar] [CrossRef]
- Wake, H.; Moorhouse, A.J.; Jinno, S.; Kohsaka, S.; Nabekura, J. Resting Microglia Directly Monitor the Functional State of Synapses In Vivo and Determine the Fate of Ischemic Terminals. J. Neurosci. 2009, 29, 3974–3980. [Google Scholar] [CrossRef]
- Costello, D.A.; Lyons, A.; Denieffe, S.; Browne, T.C.; Cox, F.F.; Lynch, M.A. Long Term Potentiation Is Impaired in Membrane Glycoprotein CD200-Deficient Mice. Journal of Biological Chemistry 2011, 286, 34722–34732. [Google Scholar] [CrossRef]
- Kierdorf, K.; Prinz, M. Factors Regulating Microglia Activation. Front. Cell. Neurosci. 2013, 7. [Google Scholar] [CrossRef]
- Pascual, O.; Ben Achour, S.; Rostaing, P.; Triller, A.; Bessis, A. Microglia Activation Triggers Astrocyte-Mediated Modulation of Excitatory Neurotransmission. Proc. Natl. Acad. Sci. U.S.A. 2012, 109. [Google Scholar] [CrossRef]
- Piani, D.; Fontana, A. Involvement of the Cystine Transport System Xc- in the Macrophage-Induced Glutamate-Dependent Cytotoxicity to Neurons. J Immunol 1994, 152, 3578–3585. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.K.; Ames, H.M.; Hayashi, Y. Tetrameric Hub Structure of Postsynaptic Scaffolding Protein Homer. J. Neurosci. 2006, 26, 8492–8501. [Google Scholar] [CrossRef]
- Flierl, M.A.; Rittirsch, D.; Nadeau, B.A.; Chen, A.J.; Sarma, J.V.; Zetoune, F.S.; McGuire, S.R.; List, R.P.; Day, D.E.; Hoesel, L.M.; et al. Phagocyte-Derived Catecholamines Enhance Acute Inflammatory Injury. Nature 2007, 449, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Elkabes, S.; DiCicco-Bloom, E.; Black, I. Brain Microglia/Macrophages Express Neurotrophins That Selectively Regulate Microglial Proliferation and Function. J. Neurosci. 1996, 16, 2508–2521. [Google Scholar] [CrossRef]
- Chamak, B.; Morandi, V.; Mallat, M. Brain Macrophages Stimulate Neurite Growth and Regeneration by Secreting Thrombospondin. J of Neuroscience Research 1994, 38, 221–233. [Google Scholar] [CrossRef]
- Dityatev, A.; Rusakov, D.A. Molecular Signals of Plasticity at the Tetrapartite Synapse. Current Opinion in Neurobiology 2011, 21, 353–359. [Google Scholar] [CrossRef]
- Hanisch, U. Microglia as a Source and Target of Cytokines. Glia 2002, 40, 140–155. [Google Scholar] [CrossRef]
- Santello, M.; Bezzi, P.; Volterra, A. TNFα Controls Glutamatergic Gliotransmission in the Hippocampal Dentate Gyrus. Neuron 2011, 69, 988–1001. [Google Scholar] [CrossRef]
- Stellwagen, D.; Malenka, R.C. Synaptic Scaling Mediated by Glial TNF-α. Nature 2006, 440, 1054–1059. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.; Stellwagen, D.; Malenka, R.C.; Stryker, M.P. Tumor Necrosis Factor-α Mediates One Component of Competitive, Experience-Dependent Plasticity in Developing Visual Cortex. Neuron 2008, 58, 673–680. [Google Scholar] [CrossRef]
- Carmen, J.; Rothstein, J.D.; Kerr, D.A. Tumor Necrosis Factor-α Modulates Glutamate Transport in the CNS and Is a Critical Determinant of Outcome from Viral Encephalomyelitis. Brain Research 2009, 1263, 143–154. [Google Scholar] [CrossRef]
- Tilleux, S.; Hermans, E. Neuroinflammation and Regulation of Glial Glutamate Uptake in Neurological Disorders. J of Neuroscience Research 2007, 85, 2059–2070. [Google Scholar] [CrossRef] [PubMed]
- Tolosa, L.; Caraballo-Miralles, V.; Olmos, G.; Lladó, J. TNF-α Potentiates Glutamate-Induced Spinal Cord Motoneuron Death via NF-κB. Molecular and Cellular Neuroscience 2011, 46, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Barger, S.W.; Goodwin, M.E.; Porter, M.M.; Beggs, M.L. Glutamate Release from Activated Microglia Requires the Oxidative Burst and Lipid Peroxidation. Journal of Neurochemistry 2007, 101, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Piani, D.; Spranger, M.; Frei, K.; Schaffner, A.; Fontana, A. Macrophage-induced Cytotoxicity of N-methyl-D-aspartate Receptor Positive Neurons Involves Excitatory Amino Acids Rather than Reactive Oxygen Intermediates and Cytokines. Eur J Immunol 1992, 22, 2429–2436. [Google Scholar] [CrossRef]
- Qin, S.; Colin, C.; Hinners, I.; Gervais, A.; Cheret, C.; Mallat, M. System Xc− and Apolipoprotein E Expressed by Microglia Have Opposite Effects on the Neurotoxicity of Amyloid-β Peptide 1–40. J. Neurosci. 2006, 26, 3345–3356. [Google Scholar] [CrossRef]
- Liang, J.; Takeuchi, H.; Doi, Y.; Kawanokuchi, J.; Sonobe, Y.; Jin, S.; Yawata, I.; Li, H.; Yasuoka, S.; Mizuno, T.; et al. Excitatory Amino Acid Transporter Expression by Astrocytes Is Neuroprotective against Microglial Excitotoxicity. Brain Research 2008, 1210, 11–19. [Google Scholar] [CrossRef]
- Danbolt, N.C.; Furness, D.N.; Zhou, Y. Neuronal vs Glial Glutamate Uptake: Resolving the Conundrum. Neurochemistry International 2016, 98, 29–45. [Google Scholar] [CrossRef]
- Zhang, H. Microglia-Friend or Foe. Front Biosci 2011, S3, 869. [Google Scholar] [CrossRef]
- Ito, D.; Imai, Y.; Ohsawa, K.; Nakajima, K.; Fukuuchi, Y.; Kohsaka, S. Microglia-Specific Localisation of a Novel Calcium Binding Protein, Iba1. Molecular Brain Research 1998, 57, 1–9. [Google Scholar] [CrossRef]
- Morrison, H.W.; Filosa, J.A. A Quantitative Spatiotemporal Analysis of Microglia Morphology during Ischemic Stroke and Reperfusion. J Neuroinflammation 2013, 10, 782. [Google Scholar] [CrossRef] [PubMed]
- Ito, D.; Tanaka, K.; Suzuki, S.; Dembo, T.; Fukuuchi, Y. Enhanced Expression of Iba1, Ionized Calcium-Binding Adapter Molecule 1, After Transient Focal Cerebral Ischemia In Rat Brain. Stroke 2001, 32, 1208–1215. [Google Scholar] [CrossRef]
- Walter, L.; Franklin, A.; Witting, A.; Wade, C.; Xie, Y.; Kunos, G.; Mackie, K.; Stella, N. Nonpsychotropic Cannabinoid Receptors Regulate Microglial Cell Migration. J. Neurosci. 2003, 23, 1398–1405. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, H.; Li, Y.; Sun, X.; Zhu, M.; Hanley, G.; Lesage, G.; Yin, D. Essential Role of Toll-like Receptor 2 in Morphine-Induced Microglia Activation in Mice. Neurosci Lett 2011, 489, 43–47. [Google Scholar] [CrossRef]
- Northcutt, A.L.; Hutchinson, M.R.; Wang, X.; Baratta, M.V.; Hiranita, T.; Cochran, T.A.; Pomrenze, M.B.; Galer, E.L.; Kopajtic, T.A.; Li, C.M.; et al. DAT Isn’t All That: Cocaine Reward and Reinforcement Require Toll-like Receptor 4 Signaling. Mol Psychiatry 2015, 20, 1525–1537. [Google Scholar] [CrossRef]
- Frank, M.G.; Watkins, L.R.; Maier, S.F. Stress- and Glucocorticoid-Induced Priming of Neuroinflammatory Responses: Potential Mechanisms of Stress-Induced Vulnerability to Drugs of Abuse. Brain, Behavior, and Immunity 2011, 25, S21–S28. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.G.; Thompson, B.M.; Watkins, L.R.; Maier, S.F. Glucocorticoids Mediate Stress-Induced Priming of Microglial pro-Inflammatory Responses. Brain, Behavior, and Immunity 2012, 26, 337–345. [Google Scholar] [CrossRef]
- Nair, A.; Bonneau, R.H. Stress-Induced Elevation of Glucocorticoids Increases Microglia Proliferation through NMDA Receptor Activation. Journal of Neuroimmunology 2006, 171, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, U.-K.; Kettenmann, H. Microglia: Active Sensor and Versatile Effector Cells in the Normal and Pathologic Brain. Nat Neurosci 2007, 10, 1387–1394. [Google Scholar] [CrossRef]
- Schwartz, M.; Butovsky, O.; Brück, W.; Hanisch, U.-K. Microglial Phenotype: Is the Commitment Reversible? Trends in Neurosciences 2006, 29, 68–74. [Google Scholar] [CrossRef]
- Streit, W.J.; Walter, S.A.; Pennell, N.A. Reactive Microgliosis. Progress in Neurobiology 1999, 57, 563–581. [Google Scholar] [CrossRef]
- Smith, J.A.; Das, A.; Ray, S.K.; Banik, N.L. Role of Pro-Inflammatory Cytokines Released from Microglia in Neurodegenerative Diseases. Brain Research Bulletin 2012, 87, 10–20. [Google Scholar] [CrossRef]
- Liu, W.; Tang, Y.; Feng, J. Cross Talk between Activation of Microglia and Astrocytes in Pathological Conditions in the Central Nervous System. Life Sciences 2011, 89, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Herculano-Houzel, S. The Glia/Neuron Ratio: How It Varies Uniformly across Brain Structures and Species and What That Means for Brain Physiology and Evolution. Glia 2014, 62, 1377–1391. [Google Scholar] [CrossRef]
- Kardos, J.; Dobolyi, Á.; Szabó, Z.; Simon, Á.; Lourmet, G.; Palkovits, M.; Héja, L. Molecular Plasticity of the Nucleus Accumbens Revisited—Astrocytic Waves Shall Rise. Mol Neurobiol 2019, 56, 7950–7965. [Google Scholar] [CrossRef]
- Keller, D.; Erö, C.; Markram, H. Cell Densities in the Mouse Brain: A Systematic Review. Front. Neuroanat. 2018, 12, 83. [Google Scholar] [CrossRef] [PubMed]
- Perea, G.; Navarrete, M.; Araque, A. Tripartite Synapses: Astrocytes Process and Control Synaptic Information. Trends in Neurosciences 2009, 32, 421–431. [Google Scholar] [CrossRef]
- Bezzi, P.; Volterra, A. A Neuron–Glia Signalling Network in the Active Brain. Current Opinion in Neurobiology 2001, 11, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Magistretti, P.J. Neuron–Glia Metabolic Coupling and Plasticity. Journal of Experimental Biology 2006, 209, 2304–2311. [Google Scholar] [CrossRef]
- Middeldorp, J.; Hol, E.M. GFAP in Health and Disease. Progress in Neurobiology 2011, 93, 421–443. [Google Scholar] [CrossRef]
- Perea, G.; Araque, A. Astrocytes Potentiate Transmitter Release at Single Hippocampal Synapses. Science 2007, 317, 1083–1086. [Google Scholar] [CrossRef]
- Bushong, E.A.; Martone, M.E.; Jones, Y.Z.; Ellisman, M.H. Protoplasmic Astrocytes in CA1 Stratum Radiatum Occupy Separate Anatomical Domains. J. Neurosci. 2002, 22, 183–192. [Google Scholar] [CrossRef]
- Halassa, M.M.; Fellin, T.; Takano, H.; Dong, J.-H.; Haydon, P.G. Synaptic Islands Defined by the Territory of a Single Astrocyte. Journal of Neuroscience 2007, 27, 6473–6477. [Google Scholar] [CrossRef]
- Parpura, V.; Verkhratsky, A. Astroglial Amino Acid-Based Transmitter Receptors. Amino Acids 2013, 44, 1151–1158. [Google Scholar] [CrossRef]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Tripartite Synapses: Glia, the Unacknowledged Partner. Trends in Neurosciences 1999, 22, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Simard, M.; Arcuino, G.; Takano, T.; Liu, Q.S.; Nedergaard, M. Signaling at the Gliovascular Interface. J. Neurosci. 2003, 23, 9254–9262. [Google Scholar] [CrossRef]
- D’Ascenzo, M.; Fellin, T.; Terunuma, M.; Revilla-Sanchez, R.; Meaney, D.F.; Auberson, Y.P.; Moss, S.J.; Haydon, P.G. mGluR5 Stimulates Gliotransmission in the Nucleus Accumbens. Proc. Natl. Acad. Sci. U.S.A. 2007, 104, 1995–2000. [Google Scholar] [CrossRef]
- Anderson, C.M.; Swanson, R.A. Astrocyte Glutamate Transport: Review of Properties, Regulation, and Physiological Functions. Glia 2000, 32, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Schousboe, A. Pharmacological and Functional Characterization of Astrocytic GABA Transport: A Short Review. Neurochem Res 2000, 25, 1241–1244. [Google Scholar] [CrossRef] [PubMed]
- Corkrum, M.; Covelo, A.; Lines, J.; Bellocchio, L.; Pisansky, M.; Loke, K.; Quintana, R.; Rothwell, P.E.; Lujan, R.; Marsicano, G.; et al. Dopamine-Evoked Synaptic Regulation in the Nucleus Accumbens Requires Astrocyte Activity. Neuron 2020, 105, 1036–1047.e5. [Google Scholar] [CrossRef] [PubMed]
- Xin, W.; Schuebel, K.E.; Jair, K.; Cimbro, R.; De Biase, L.M.; Goldman, D.; Bonci, A. Ventral Midbrain Astrocytes Display Unique Physiological Features and Sensitivity to Dopamine D2 Receptor Signaling. Neuropsychopharmacol 2019, 44, 344–355. [Google Scholar] [CrossRef]
- Farina, C.; Aloisi, F.; Meinl, E. Astrocytes Are Active Players in Cerebral Innate Immunity. Trends in Immunology 2007, 28, 138–145. [Google Scholar] [CrossRef]
- Jensen, C.J.; Massie, A.; De Keyser, J. Immune Players in the CNS: The Astrocyte. J Neuroimmune Pharmacol 2013, 8, 824–839. [Google Scholar] [CrossRef]
- Lau, L.T.; Yu, A.C.-H. Astrocytes Produce and Release Interleukin-1, Interleukin-6, Tumor Necrosis Factor Alpha and Interferon-Gamma Following Traumatic and Metabolic Injury. Journal of Neurotrauma 2001, 18, 351–359. [Google Scholar] [CrossRef]
- Pekny, M.; Pekna, M. Astrocyte Reactivity and Reactive Astrogliosis: Costs and Benefits. Physiological Reviews 2014, 94, 1077–1098. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Ma, L.; Kaarela, T.; Li, Z. Neuroimmune Crosstalk in the Central Nervous System and Its Significance for Neurological Diseases. J Neuroinflammation 2012, 9. [Google Scholar] [CrossRef] [PubMed]
- Spampinato, S.F.; Copani, A.; Nicoletti, F.; Sortino, M.A.; Caraci, F. Metabotropic Glutamate Receptors in Glial Cells: A New Potential Target for Neuroprotection? Front. Mol. Neurosci. 2018, 11. [Google Scholar] [CrossRef]
- Haydon, P.G. Glia: Listening and Talking to the Synapse. Nat Rev Neurosci 2001, 2, 185–193. [Google Scholar] [CrossRef]
- Volterra, A.; Meldolesi, J. Astrocytes, from Brain Glue to Communication Elements: The Revolution Continues. Nat Rev Neurosci 2005, 6, 626–640. [Google Scholar] [CrossRef] [PubMed]
- Halassa, M.M.; Haydon, P.G. Integrated Brain Circuits: Astrocytic Networks Modulate Neuronal Activity and Behavior. Annu. Rev. Physiol. 2010, 72, 335–355. [Google Scholar] [CrossRef]
- Volterra, A.; Magistretti, P. The Tripartite Synapse: Glia in Synaptic Transmission; Oxford University Press: Oxford, 2002; ISBN 978-0-19-850854-0. [Google Scholar]
- Poskanzer, K.E.; Yuste, R. Astrocytic Regulation of Cortical UP States. Proc. Natl. Acad. Sci. U.S.A. 2011, 108, 18453–18458. [Google Scholar] [CrossRef] [PubMed]
- Schipke, C.G.; Boucsein, C.; Ohlemeyer, C.; Kirchhoff, F.; Kettenmann, H. Astrocyte Ca2+Waves Trigger Responses in Microglial Cells in Brain Slices. FASEB j. 2002, 16, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sieger, D.; Moritz, C.; Ziegenhals, T.; Prykhozhij, S.; Peri, F. Long-Range Ca2+ Waves Transmit Brain-Damage Signals to Microglia. Developmental Cell 2012, 22, 1138–1148. [Google Scholar] [CrossRef]
- Albrecht, J.; Sidoryk-Węgrzynowicz, M.; Zielińska, M.; Aschner, M. Roles of Glutamine in Neurotransmission. Neuron Glia Biol. 2010, 6, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, S.; Gharagozloo, M.; Simard, C.; Gris, D. Astrocytes Maintain Glutamate Homeostasis in the CNS by Controlling the Balance between Glutamate Uptake and Release. Cells 2019, 8, 184. [Google Scholar] [CrossRef]
- Petr, G.T.; Sun, Y.; Frederick, N.M.; Zhou, Y.; Dhamne, S.C.; Hameed, M.Q.; Miranda, C.; Bedoya, E.A.; Fischer, K.D.; Armsen, W.; et al. Conditional Deletion of the Glutamate Transporter GLT-1 Reveals That Astrocytic GLT-1 Protects against Fatal Epilepsy While Neuronal GLT-1 Contributes Significantly to Glutamate Uptake into Synaptosomes. J. Neurosci. 2015, 35, 5187–5201. [Google Scholar] [CrossRef]
- Rothstein, J.D.; Dykes-Hoberg, M.; Pardo, C.A.; Bristol, L.A.; Jin, L.; Kuncl, R.W.; Kanai, Y.; Hediger, M.A.; Wang, Y.; Schielke, J.P.; et al. Knockout of Glutamate Transporters Reveals a Major Role for Astroglial Transport in Excitotoxicity and Clearance of Glutamate. Neuron 1996, 16, 675–686. [Google Scholar] [CrossRef]
- Hascup, K.N.; Hascup, E.R.; Pomerleau, F.; Huettl, P.; Gerhardt, G.A. Second-by-Second Measures of L-Glutamate in the Prefrontal Cortex and Striatum of Freely Moving Mice. J Pharmacol Exp Ther 2008, 324, 725–731. [Google Scholar] [CrossRef]
- Van Der Zeyden, M.; Oldenziel, W.H.; Rea, K.; Cremers, T.I.; Westerink, B.H. Microdialysis of GABA and Glutamate: Analysis, Interpretation and Comparison with Microsensors. Pharmacology Biochemistry and Behavior 2008, 90, 135–147. [Google Scholar] [CrossRef]
- Danbolt, N.C. Glutamate Uptake. Progress in Neurobiology 2001, 65, 1–105. [Google Scholar] [CrossRef]
- Williams, S.M.; Sullivan, R.K.P.; Scott, H.L.; Finkelstein, D.I.; Colditz, P.B.; Lingwood, B.E.; Dodd, P.R.; Pow, D.V. Glial Glutamate Transporter Expression Patterns in Brains from Multiple Mammalian Species. Glia 2005, 49, 520–541. [Google Scholar] [CrossRef]
- McGrath, A.G.; Briand, L.A. A Potential Role for Microglia in Stress- and Drug-Induced Plasticity in the Nucleus Accumbens: A Mechanism for Stress-Induced Vulnerability to Substance Use Disorder. Neuroscience & Biobehavioral Reviews 2019, 107, 360–369. [Google Scholar] [CrossRef]
- Crews, F.T.; Lawrimore, C.J.; Walter, T.J.; Coleman, L.G. The Role of Neuroimmune Signaling in Alcoholism. Neuropharmacology 2017, 122, 56–73. [Google Scholar] [CrossRef] [PubMed]
- Gipson, C.D.; Rawls, S.; Scofield, M.D.; Siemsen, B.M.; Bondy, E.O.; Maher, E.E. Interactions of Neuroimmune Signaling and Glutamate Plasticity in Addiction. J Neuroinflammation 2021, 18. [Google Scholar] [CrossRef] [PubMed]
- Namba, M.D.; Kupchik, Y.M.; Spencer, S.M.; Garcia-Keller, C.; Goenaga, J.G.; Powell, G.L.; Vicino, I.A.; Hogue, I.B.; Gipson, C.D. Accumbens Neuroimmune Signaling and Dysregulation of Astrocytic Glutamate Transport Underlie Conditioned Nicotine-seeking Behavior. Addiction Biology 2020, 25, e12797. [Google Scholar] [CrossRef]
- Frank, M.G.; Weber, M.D.; Watkins, L.R.; Maier, S.F. Stress-Induced Neuroinflammatory Priming: A Liability Factor in the Etiology of Psychiatric Disorders. Neurobiology of Stress 2016, 4, 62–70. [Google Scholar] [CrossRef]
- Lo Iacono, L.; Catale, C.; Martini, A.; Valzania, A.; Viscomi, M.T.; Chiurchiù, V.; Guatteo, E.; Bussone, S.; Perrone, F.; Di Sabato, P.; et al. From Traumatic Childhood to Cocaine Abuse: The Critical Function of the Immune System. Biological Psychiatry 2018, 84, 905–916. [Google Scholar] [CrossRef]
- Boldyrev, A.A.; Carpenter, D.O.; Johnson, P. Emerging Evidence for a Similar Role of Glutamate Receptors in the Nervous and Immune Systems. Journal of Neurochemistry 2005, 95, 913–918. [Google Scholar] [CrossRef]
- Salter, M.W.; Beggs, S. Sublime Microglia: Expanding Roles for the Guardians of the CNS. Cell 2014, 158, 15–24. [Google Scholar] [CrossRef]
- Tay, T.L.; Savage, J.C.; Hui, C.W.; Bisht, K.; Tremblay, M. Microglia across the Lifespan: From Origin to Function in Brain Development, Plasticity and Cognition. The Journal of Physiology 2017, 595, 1929–1945. [Google Scholar] [CrossRef]
- Weinhard, L.; Di Bartolomei, G.; Bolasco, G.; Machado, P.; Schieber, N.L.; Neniskyte, U.; Exiga, M.; Vadisiute, A.; Raggioli, A.; Schertel, A.; et al. Microglia Remodel Synapses by Presynaptic Trogocytosis and Spine Head Filopodia Induction. Nat Commun 2018, 9, 1228. [Google Scholar] [CrossRef]
- Kreisel, T.; Frank, M.G.; Licht, T.; Reshef, R.; Ben-Menachem-Zidon, O.; Baratta, M.V.; Maier, S.F.; Yirmiya, R. Dynamic Microglial Alterations Underlie Stress-Induced Depressive-like Behavior and Suppressed Neurogenesis. Mol Psychiatry 2014, 19, 699–709. [Google Scholar] [CrossRef]
- Wang, W.; Wang, R.; Xu, J.; Qin, X.; Jiang, H.; Khalid, A.; Liu, D.; Pan, F.; Ho, C.S.H.; Ho, R.C.M. Minocycline Attenuates Stress-Induced Behavioral Changes via Its Anti-Inflammatory Effects in an Animal Model of Post-Traumatic Stress Disorder. Front. Psychiatry 2018, 9, 558. [Google Scholar] [CrossRef]
- Johnson, J.D.; O’Connor, K.A.; Watkins, L.R.; Maier, S.F. The Role of IL-1β in Stress-Induced Sensitization of Proinflammatory Cytokine and Corticosterone Responses. Neuroscience 2004, 127, 569–577. [Google Scholar] [CrossRef]
- Blandino, P.; Barnum, C.J.; Deak, T. The Involvement of Norepinephrine and Microglia in Hypothalamic and Splenic IL-1β Responses to Stress. Journal of Neuroimmunology 2006, 173, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Blandino, P.; Barnum, C.J.; Solomon, L.G.; Larish, Y.; Lankow, B.S.; Deak, T. Gene Expression Changes in the Hypothalamus Provide Evidence for Regionally-Selective Changes in IL-1 and Microglial Markers after Acute Stress. Brain, Behavior, and Immunity 2009, 23, 958–968. [Google Scholar] [CrossRef] [PubMed]
- Dowlati, Y.; Herrmann, N.; Swardfager, W.; Liu, H.; Sham, L.; Reim, E.K.; Lanctôt, K.L. A Meta-Analysis of Cytokines in Major Depression. Biological Psychiatry 2010, 67, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Gilman, S.E.; Trinh, N.-H.; Smoller, J.W.; Fava, M.; Murphy, J.M.; Breslau, J. Psychosocial Stressors and the Prognosis of Major Depression: A Test of Axis IV. Psychol. Med. 2013, 43, 303–316. [Google Scholar] [CrossRef]
- Kendler, K.S.; Karkowski, L.M.; Prescott, C.A. Causal Relationship Between Stressful Life Events and the Onset of Major Depression. AJP 1999, 156, 837–841. [Google Scholar] [CrossRef]
- McLaughlin, K.A.; Kubzansky, L.D.; Dunn, E.C.; Waldinger, R.; Vaillant, G.; Koenen, K.C. Childhood Social Environment, Emotional Reactivity to Stress, and Mood and Anxiety Disorders across the Life Course. Depress. Anxiety 2010, 27, 1087–1094. [Google Scholar] [CrossRef]
- Miller, G.E.; Chen, E.; Sze, J.; Marin, T.; Arevalo, J.M.G.; Doll, R.; Ma, R.; Cole, S.W. A Functional Genomic Fingerprint of Chronic Stress in Humans: Blunted Glucocorticoid and Increased NF-κB Signaling. Biological Psychiatry 2008, 64, 266–272. [Google Scholar] [CrossRef]
- Hinwood, M.; Tynan, R.J.; Charnley, J.L.; Beynon, S.B.; Day, T.A.; Walker, F.R. Chronic Stress Induced Remodeling of the Prefrontal Cortex: Structural Re-Organization of Microglia and the Inhibitory Effect of Minocycline. Cerebral Cortex 2013, 23, 1784–1797. [Google Scholar] [CrossRef]
- Tynan, R.J.; Naicker, S.; Hinwood, M.; Nalivaiko, E.; Buller, K.M.; Pow, D.V.; Day, T.A.; Walker, F.R. Chronic Stress Alters the Density and Morphology of Microglia in a Subset of Stress-Responsive Brain Regions. Brain Behav Immun 2010, 24, 1058–1068. [Google Scholar] [CrossRef]
- Hinwood, M.; Morandini, J.; Day, T.A.; Walker, F.R. Evidence That Microglia Mediate the Neurobiological Effects of Chronic Psychological Stress on the Medial Prefrontal Cortex. Cerebral Cortex 2012, 22, 1442–1454. [Google Scholar] [CrossRef]
- Gerecke, K.M.; Kolobova, A.; Allen, S.; Fawer, J.L. Exercise Protects against Chronic Restraint Stress-Induced Oxidative Stress in the Cortex and Hippocampus. Brain Research 2013, 1509, 66–78. [Google Scholar] [CrossRef]
- Wohleb, E.S.; Hanke, M.L.; Corona, A.W.; Powell, N.D.; Stiner, L.M.; Bailey, M.T.; Nelson, R.J.; Godbout, J.P.; Sheridan, J.F. β-Adrenergic Receptor Antagonism Prevents Anxiety-Like Behavior and Microglial Reactivity Induced by Repeated Social Defeat. J. Neurosci. 2011, 31, 6277–6288. [Google Scholar] [CrossRef]
- Wohleb, E.S.; Powell, N.D.; Godbout, J.P.; Sheridan, J.F. Stress-Induced Recruitment of Bone Marrow-Derived Monocytes to the Brain Promotes Anxiety-Like Behavior. J. Neurosci. 2013, 33, 13820–13833. [Google Scholar] [CrossRef] [PubMed]
- Wohleb, E.S.; Patterson, J.M.; Sharma, V.; Quan, N.; Godbout, J.P.; Sheridan, J.F. Knockdown of Interleukin-1 Receptor Type-1 on Endothelial Cells Attenuated Stress-Induced Neuroinflammation and Prevented Anxiety-Like Behavior. J. Neurosci. 2014, 34, 2583–2591. [Google Scholar] [CrossRef] [PubMed]
- Wohleb, E.S.; McKim, D.B.; Shea, D.T.; Powell, N.D.; Tarr, A.J.; Sheridan, J.F.; Godbout, J.P. Re-Establishment of Anxiety in Stress-Sensitized Mice Is Caused by Monocyte Trafficking from the Spleen to the Brain. Biological Psychiatry 2014, 75, 970–981. [Google Scholar] [CrossRef] [PubMed]
- Fontainhas, A.M.; Wang, M.; Liang, K.J.; Chen, S.; Mettu, P.; Damani, M.; Fariss, R.N.; Li, W.; Wong, W.T. Microglial Morphology and Dynamic Behavior Is Regulated by Ionotropic Glutamatergic and GABAergic Neurotransmission. PLoS ONE 2011, 6, e15973. [Google Scholar] [CrossRef]
- Silverman, M.N.; Sternberg, E.M. Glucocorticoid Regulation of Inflammation and Its Functional Correlates: From HPA Axis to Glucocorticoid Receptor Dysfunction. Annals of the New York Academy of Sciences 2012, 1261, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Sorrells, S.F.; Caso, J.R.; Munhoz, C.D.; Sapolsky, R.M. The Stressed CNS: When Glucocorticoids Aggravate Inflammation. Neuron 2009, 64, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Sorrells, S.F.; Sapolsky, R.M. An Inflammatory Review of Glucocorticoid Actions in the CNS. Brain, Behavior, and Immunity 2007, 21, 259–272. [Google Scholar] [CrossRef]
- Coller, J.K.; Hutchinson, M.R. Implications of Central Immune Signaling Caused by Drugs of Abuse: Mechanisms, Mediators and New Therapeutic Approaches for Prediction and Treatment of Drug Dependence. Pharmacology and Therapeutics 2012, 134, 219–245. [Google Scholar] [CrossRef]
- Liao, K.; Guo, M.; Niu, F.; Yang, L.; Callen, S.E.; Buch, S. Cocaine-Mediated Induction of Microglial Activation Involves the ER Stress-TLR2 Axis. J Neuroinflammation 2016, 13, 33. [Google Scholar] [CrossRef]
- Guo, M.-L.; Roodsari, S.K.; Cheng, Y.; Dempsey, R.E.; Hu, W. Microglia NLRP3 Inflammasome and Neuroimmune Signaling in Substance Use Disorders. Biomolecules 2023, 13, 922. [Google Scholar] [CrossRef]
- Ye, J.; Gao, S.; Liu, Z.; Chen, X.; He, J.; Hu, Z. The HMGB1–RAGE Axis in Nucleus Accumbens Facilitates Cocaine-induced Conditioned Place Preference via Modulating Microglial Activation. Brain and Behavior 2024, 14, e3457. [Google Scholar] [CrossRef]
- Wohleb, E.S.; Delpech, J.-C. Dynamic Cross-Talk between Microglia and Peripheral Monocytes Underlies Stress-Induced Neuroinflammation and Behavioral Consequences. Progress in Neuro-Psychopharmacology and Biological Psychiatry 2017, 79, 40–48. [Google Scholar] [CrossRef]
- Assis, M.A.; Hansen, C.; Lux-Lantos, V.; Cancela, L.M. Sensitization to Amphetamine Occurs Simultaneously at Immune Level and in Met-Enkephalin of the Nucleus Accumbens and Spleen: An Involved NMDA Glutamatergic Mechanism. Brain, Behavior, and Immunity 2009, 23, 464–473. [Google Scholar] [CrossRef]
- Jurga, A.M.; Paleczna, M.; Kuter, K.Z. Overview of General and Discriminating Markers of Differential Microglia Phenotypes. Front. Cell. Neurosci. 2020, 14, 198. [Google Scholar] [CrossRef] [PubMed]
- Niraula, A.; Wang, Y.; Godbout, J.P.; Sheridan, J.F. Corticosterone Production during Repeated Social Defeat Causes Monocyte Mobilization from the Bone Marrow, Glucocorticoid Resistance, and Neurovascular Adhesion Molecule Expression. J. Neurosci. 2018, 38, 2328–2340. [Google Scholar] [CrossRef]
- Moreira, F.P.; Medeiros, J.R.C.; Lhullier, A.C.; Souza, L.D.D.M.; Jansen, K.; Portela, L.V.; Lara, D.R.; Silva, R.A.D.; Wiener, C.D.; Oses, J.P. Cocaine Abuse and Effects in the Serum Levels of Cytokines IL-6 and IL-10. Drug and Alcohol Dependence 2016, 158, 181–185. [Google Scholar] [CrossRef]
- Menard, C.; Pfau, M.L.; Hodes, G.E.; Kana, V.; Wang, V.X.; Bouchard, S.; Takahashi, A.; Flanigan, M.E.; Aleyasin, H.; LeClair, K.B.; et al. Social Stress Induces Neurovascular Pathology Promoting Depression. Nat Neurosci 2017, 20, 1752–1760. [Google Scholar] [CrossRef] [PubMed]
- Banqueri, M.; Méndez, M.; Gómez-Lázaro, E.; Arias, J.L. Early Life Stress by Repeated Maternal Separation Induces Long-Term Neuroinflammatory Response in Glial Cells of Male Rats. Stress 2019, 22, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Aniszewska, A.; Chłodzińska, N.; Bartkowska, K.; Winnicka, M.M.; Turlejski, K.; Djavadian, R.L. The Expression of Interleukin-6 and Its Receptor in Various Brain Regions and Their Roles in Exploratory Behavior and Stress Responses. Journal of Neuroimmunology 2015, 284, 1–9. [Google Scholar] [CrossRef]
- Shirokova, O.M.; Kuzmina, D.M.; Zaborskaya, O.G.; Shchelchkova, N.A.; Kozliaeva, E.V.; Korotchenko, S.A.; Pershin, V.I.; Vasilchikov, P.I.; Mukhina, I.V. The Long-Term Effects of Chronic Unpredictable Mild Stress Experienced During Adolescence Could Vary Depending on Biological Sex. IJMS 2025, 26, 1251. [Google Scholar] [CrossRef]
- Bodemeier Loayza Careaga, M.; Wu, T.J. Chronically Stressed Male and Female Mice Show a Similar Peripheral and Central Pro-Inflammatory Profile after an Immune Challenge. PLoS ONE 2024, 19, e0297776. [Google Scholar] [CrossRef]
- Khakh, B.S.; Sofroniew, M.V. Diversity of Astrocyte Functions and Phenotypes in Neural Circuits. Nat Neurosci 2015, 18, 942–952. [Google Scholar] [CrossRef]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and Pathology. Acta Neuropathol 2010, 119, 7–35. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Butt, A.; Li, B.; Illes, P.; Zorec, R.; Semyanov, A.; Tang, Y.; Sofroniew, M.V. Astrocytes in Human Central Nervous System Diseases: A Frontier for New Therapies. Sig Transduct Target Ther 2023, 8, 396. [Google Scholar] [CrossRef]
- Bowers, M.S.; Kalivas, P.W. Forebrain Astroglial Plasticity Is Induced Following Withdrawal from Repeated Cocaine Administration. Eur J of Neuroscience 2003, 17, 1273–1278. [Google Scholar] [CrossRef] [PubMed]
- Narita, M.; Miyatake, M.; Shibasaki, M.; Tsuda, M.; Koizumi, S.; Narita, M.; Yajima, Y.; Inoue, K.; Suzuki, T. Long-lasting Change in Brain Dynamics Induced by Methamphetamine: Enhancement of Protein Kinase C-dependent Astrocytic Response and Behavioral Sensitization. Journal of Neurochemistry 2005, 93, 1383–1392. [Google Scholar] [CrossRef] [PubMed]
- Narita, M.; Suzuki, M.; Kuzumaki, N.; Miyatake, M.; Suzuki, T. Implication of Activated Astrocytes in the Development of Drug Dependence: Differences between Methamphetamine and Morphine. Annals of the New York Academy of Sciences 2008, 1141, 96–104. [Google Scholar] [CrossRef]
- Narita, M.; Miyatake, M.; Narita, M.; Shibasaki, M.; Shindo, K.; Nakamura, A.; Kuzumaki, N.; Nagumo, Y.; Suzuki, T. Direct Evidence of Astrocytic Modulation in the Development of Rewarding Effects Induced by Drugs of Abuse. Neuropsychopharmacol 2006, 31, 2476–2488. [Google Scholar] [CrossRef]
- Testen, A.; Sepulveda-Orengo, M.T.; Gaines, C.H.; Reissner, K.J. Region-Specific Reductions in Morphometric Properties and Synaptic Colocalization of Astrocytes Following Cocaine Self-Administration and Extinction. Front. Cell. Neurosci. 2018, 12, 246. [Google Scholar] [CrossRef] [PubMed]
- Siemsen, B.M.; Reichel, C.M.; Leong, K.C.; Garcia-Keller, C.; Gipson, C.D.; Spencer, S.; McFaddin, J.A.; Hooker, K.N.; Kalivas, P.W.; Scofield, M.D. Effects of Methamphetamine Self-Administration and Extinction on Astrocyte Structure and Function in the Nucleus Accumbens Core. Neuroscience 2019, 406, 528–541. [Google Scholar] [CrossRef]
- Tynan, R.J.; Beynon, S.B.; Hinwood, M.; Johnson, S.J.; Nilsson, M.; Woods, J.J.; Walker, F.R. Chronic Stress-Induced Disruption of the Astrocyte Network Is Driven by Structural Atrophy and Not Loss of Astrocytes. Acta Neuropathol 2013, 126, 75–91. [Google Scholar] [CrossRef]
- Çalışkan, G.; Müller, A.; Albrecht, A. Long-Term Impact of Early-Life Stress on Hippocampal Plasticity: Spotlight on Astrocytes. IJMS 2020, 21, 4999. [Google Scholar] [CrossRef]
- Leventopoulos, M.; Rüedi-Bettschen, D.; Knuesel, I.; Feldon, J.; Pryce, C.R.; Opacka-Juffry, J. Long-Term Effects of Early Life Deprivation on Brain Glia in Fischer Rats. Brain Research 2007, 1142, 119–126. [Google Scholar] [CrossRef]
- Musholt, K.; Cirillo, G.; Cavaliere, C.; Rosaria Bianco, M.; Bock, J.; Helmeke, C.; Braun, K.; Papa, M. Neonatal Separation Stress Reduces Glial Fibrillary Acidic Protein- and S100β-immunoreactive Astrocytes in the Rat Medial Precentral Cortex. Developmental Neurobiology 2009, 69, 203–211. [Google Scholar] [CrossRef]
- Woodburn, S.C.; Bollinger, J.L.; Wohleb, E.S. Synaptic and Behavioral Effects of Chronic Stress Are Linked to Dynamic and Sex-Specific Changes in Microglia Function and Astrocyte Dystrophy. Neurobiology of Stress 2021, 14, 100312. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.Y.; Elias, E.; Manners, M.T. Sex-Dependent Astrocyte Reactivity: Unveiling Chronic Stress-Induced Morphological Changes across Multiple Brain Regions. Neurobiology of Disease 2024, 200, 106610. [Google Scholar] [CrossRef]
- Berger, U.V.; Hediger, M.A. Comparative Analysis of Glutamate Transporter Expression in Rat Brain Using Differential Double in Situ Hybridization. Anatomy and Embryology 1998, 198, 13–30. [Google Scholar] [CrossRef]
- Berger, U.V.; Hediger, M.A. Distribution of the Glutamate Transporters GLAST and GLT-1 in Rat Circumventricular Organs, Meninges, and Dorsal Root Ganglia. J Comp Neurol 2000, 421, 385–399. [Google Scholar] [CrossRef]
- Berger, U.V.; Hediger, M.A. Differential Distribution of the Glutamate Transporters GLT-1 and GLAST in Tanycytes of the Third Ventricle. J of Comparative Neurology 2001, 433, 101–114. [Google Scholar] [CrossRef]
- Persson, M.; Brantefjord, M.; Hansson, E.; Rönnbäck, L. Lipopolysaccharide Increases Microglial GLT-1 Expression and Glutamate Uptake Capacity in Vitro by a Mechanism Dependent on TNF-α. Glia 2005, 51, 111–120. [Google Scholar] [CrossRef]
- Shaked, I.; Tchoresh, D.; Gersner, R.; Meiri, G.; Mordechai, S.; Xiao, X.; Hart, R.P.; Schwartz, M. Protective Autoimmunity: Interferon-γ Enables Microglia to Remove Glutamate without Evoking Inflammatory Mediators. Journal of Neurochemistry 2005, 92, 997–1009. [Google Scholar] [CrossRef]
- Lehre, K.; Levy, L.; Ottersen, O.; Storm-Mathisen, J.; Danbolt, N. Differential Expression of Two Glial Glutamate Transporters in the Rat Brain: Quantitative and Immunocytochemical Observations. J. Neurosci. 1995, 15, 1835–1853. [Google Scholar] [CrossRef] [PubMed]
- Murugan, M.; Ling, E.-A.; Kaur, C. Glutamate Receptors in Microglia. CNSNDDT 2013, 12, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Leak, R.K.; Hu, X. Neurotransmitter Receptors on Microglia. Stroke Vasc Neurol 2016, 1, 52–58. [Google Scholar] [CrossRef]
- Moghaddam, B.; Bolinao, M.L.; Stein-Behrens, B.; Sapolsky, R. Glucocortcoids Mediate the Stress-Induced Extracellular Accumulation of Glutamate. Brain Research 1994, 655, 251–254. [Google Scholar] [CrossRef]
- Popoli, M.; Yan, Z.; McEwen, B.S.; Sanacora, G. The Stressed Synapse: The Impact of Stress and Glucocorticoids on Glutamate Transmission. Nat Rev Neurosci 2012, 13, 22–37. [Google Scholar] [CrossRef]
- Fontainhas, A.M.; Wang, M.; Liang, K.J.; Chen, S.; Mettu, P.; Damani, M.; Fariss, R.N.; Li, W.; Wong, W.T. Microglial Morphology and Dynamic Behavior Is Regulated by Ionotropic Glutamatergic and GABAergic Neurotransmission. PLoS ONE 2011, 6, e15973. [Google Scholar] [CrossRef] [PubMed]
- Eyo, U.B.; Peng, J.; Swiatkowski, P.; Mukherjee, A.; Bispo, A.; Wu, L.-J. Neuronal Hyperactivity Recruits Microglial Processes via Neuronal NMDA Receptors and Microglial P2Y12 Receptors after Status Epilepticus. Journal of Neuroscience 2014, 34, 10528–10540. [Google Scholar] [CrossRef]
- Virgin, C.E.; Ha, T.P. -T.; Packan, D.R.; Tombaugh, G.C.; Yang, S.H.; Homer, H.C.; Sapolsky, R.M. Glucocorticoids Inhibit Glucose Transport and Glutamate Uptake in Hippocampal Astrocytes: Implications for Glucocorticoid Neurotoxicity. Journal of Neurochemistry 1991, 57, 1422–1428. [Google Scholar] [CrossRef]
- Chaparro-Huerta, V.; Rivera-Cervantes, M.C.; Torres-Mendoza, B.M.; Beas-Zárate, C. Neuronal Death and Tumor Necrosis Factor-α Response to Glutamate-Induced Excitotoxicity in the Cerebral Cortex of Neonatal Rats. Neuroscience Letters 2002, 333, 95–98. [Google Scholar] [CrossRef]
- Noda, M.; Nakanishi, H.; Nabekura, J.; Akaike, N. AMPA–Kainate Subtypes of Glutamate Receptor in Rat Cerebral Microglia. J. Neurosci. 2000, 20, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.L.; Jones, F.; Kubota, E.S.F.C.S.; Pocock, J.M. Stimulation of Microglial Metabotropic Glutamate Receptor mGlu2 Triggers Tumor Necrosis Factor α-Induced Neurotoxicity in Concert with Microglial-Derived Fas Ligand. J. Neurosci. 2005, 25, 2952–2964. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Xie, W.; Le, W.; Beers, D.R.; He, Y.; Henkel, J.S.; Simpson, E.P.; Yen, A.A.; Xiao, Q.; Appel, S.H. Activated Microglia Initiate Motor Neuron Injury by a Nitric Oxide and Glutamate-Mediated Mechanism. J Neuropathol Exp Neurol 2004, 63, 964–977. [Google Scholar] [CrossRef]
- Zhu, W.; Zheng, H.; Shao, X.; Wang, W.; Yao, Q.; Li, Z. Excitotoxicity of TNFα Derived from KA Activated Microglia on Hippocampal Neurons in Vitro and in Vivo. Journal of Neurochemistry 2010, 114, 386–396. [Google Scholar] [CrossRef]
- De, A.; Krueger, J.M.; Simasko, S.M. Glutamate Induces the Expression and Release of Tumor Necrosis Factor-α in Cultured Hypothalamic Cells. Brain Research 2005, 1053, 54–61. [Google Scholar] [CrossRef]
- Cahoy, J.D.; Emery, B.; Kaushal, A.; Foo, L.C.; Zamanian, J.L.; Christopherson, K.S.; Xing, Y.; Lubischer, J.L.; Krieg, P.A.; Krupenko, S.A.; et al. A Transcriptome Database for Astrocytes, Neurons, and Oligodendrocytes: A New Resource for Understanding Brain Development and Function. J Neurosci 2008, 28, 264–278. [Google Scholar] [CrossRef]
- Doyle, J.P.; Dougherty, J.D.; Heiman, M.; Schmidt, E.F.; Stevens, T.R.; Ma, G.; Bupp, S.; Shrestha, P.; Shah, R.D.; Doughty, M.L.; et al. Application of a Translational Profiling Approach for the Comparative Analysis of CNS Cell Types. Cell 2008, 135, 749–762. [Google Scholar] [CrossRef]
- Foo, L.C.; Allen, N.J.; Bushong, E.A.; Ventura, P.B.; Chung, W.-S.; Zhou, L.; Cahoy, J.D.; Daneman, R.; Zong, H.; Ellisman, M.H.; et al. Development of a Method for the Purification and Culture of Rodent Astrocytes. Neuron 2011, 71, 799–811. [Google Scholar] [CrossRef] [PubMed]
- Meissner, A.; Mikkelsen, T.S.; Gu, H.; Wernig, M.; Hanna, J.; Sivachenko, A.; Zhang, X.; Bernstein, B.E.; Nusbaum, C.; Jaffe, D.B.; et al. Genome-Scale DNA Methylation Maps of Pluripotent and Differentiated Cells. Nature 2008, 454, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.K.; Mansur, D.B.; Reifenberger, G.; Perry, A.; Leonard, J.R.; Aldape, K.D.; Albin, M.G.; Emnett, R.J.; Loeser, S.; Watson, M.A.; et al. Distinct Genetic Signatures among Pilocytic Astrocytomas Relate to Their Brain Region Origin. Cancer Research 2007, 67, 890–900. [Google Scholar] [CrossRef] [PubMed]
- Zamanian, J.L.; Xu, L.; Foo, L.C.; Nouri, N.; Zhou, L.; Giffard, R.G.; Barres, B.A. Genomic Analysis of Reactive Astrogliosis. J. Neurosci. 2012, 32, 6391–6410. [Google Scholar] [CrossRef]
- Clark, A.K.; Gruber-Schoffnegger, D.; Drdla-Schutting, R.; Gerhold, K.J.; Malcangio, M.; Sandkühler, J. Selective Activation of Microglia Facilitates Synaptic Strength. J. Neurosci. 2015, 35, 4552–4570. [Google Scholar] [CrossRef]
- Wohleb, E.S.; Terwilliger, R.; Duman, C.H.; Duman, R.S. Stress-Induced Neuronal Colony Stimulating Factor 1 Provokes Microglia-Mediated Neuronal Remodeling and Depressive-like Behavior. Biological Psychiatry 2018, 83, 38–49. [Google Scholar] [CrossRef]
- Bianco, F.; Pravettoni, E.; Colombo, A.; Schenk, U.; Möller, T.; Matteoli, M.; Verderio, C. Astrocyte-Derived ATP Induces Vesicle Shedding and IL-1β Release from Microglia. The Journal of Immunology 2005, 174, 7268–7277. [Google Scholar] [CrossRef]
- Jha, M.K.; Jo, M.; Kim, J.-H.; Suk, K. Microglia-Astrocyte Crosstalk: An Intimate Molecular Conversation. Neuroscientist 2019, 25, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Fine, S.M.; Angel, R.A.; Perry, S.W.; Epstein, L.G.; Rothstein, J.D.; Dewhurst, S.; Gelbard, H.A. Tumor Necrosis Factor α Inhibits Glutamate Uptake by Primary Human Astrocytes. Journal of Biological Chemistry 1996, 271, 15303–15306. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Pekarskaya, O.; Bencheikh, M.; Chao, W.; Gelbard, H.A.; Ghorpade, A.; Rothstein, J.D.; Volsky, D.J. Reduced Expression of Glutamate Transporter EAAT2 and Impaired Glutamate Transport in Human Primary Astrocytes Exposed to HIV-1 or Gp120. Virology 2003, 312, 60–73. [Google Scholar] [CrossRef]
- Sitcheran, R.; Gupta, P.; Fisher, P.B.; Baldwin, A.S. Positive and Negative Regulation of EAAT2 by NF-κB: A Role for N-Myc in TNFα-Controlled Repression. EMBO J 2005, 24, 510–520. [Google Scholar] [CrossRef]
- Zou, J.Y.; Crews, F.T. TNFα Potentiates Glutamate Neurotoxicity by Inhibiting Glutamate Uptake in Organotypic Brain Slice Cultures: Neuroprotection by NFκB Inhibition. Brain Research 2005, 1034, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Leszczyniecka, M.; Kang, D.; Sarkar, D.; Chao, W.; Volsky, D.J.; Fisher, P.B. Insights into Glutamate Transport Regulation in Human Astrocytes: Cloning of the Promoter for Excitatory Amino Acid Transporter 2 (EAAT2). Proc. Natl. Acad. Sci. U.S.A. 2003, 100, 1955–1960. [Google Scholar] [CrossRef]
- Madrigal, J. The Increase in TNF-α Levels Is Implicated in NF-κB Activation and Inducible Nitric Oxide Synthase Expression in Brain Cortex after Immobilization Stress. Neuropsychopharmacology 2002, 26, 155–163. [Google Scholar] [CrossRef]
- Prow, N.A.; Irani, D.N. The Inflammatory Cytokine, Interleukin-1 Beta, Mediates Loss of Astroglial Glutamate Transport and Drives Excitotoxic Motor Neuron Injury in the Spinal Cord during Acute Viral Encephalomyelitis. Journal of Neurochemistry 2008, 105, 1276–1286. [Google Scholar] [CrossRef]
- Hughes, E.G.; Maguire, J.L.; McMinn, M.T.; Scholz, R.E.; Sutherland, M.L. Loss of Glial Fibrillary Acidic Protein Results in Decreased Glutamate Transport and Inhibition of PKA-Induced EAAT2 Cell Surface Trafficking. Molecular Brain Research 2004, 124, 114–123. [Google Scholar] [CrossRef]
- Yang, Y.; Gozen, O.; Watkins, A.; Lorenzini, I.; Lepore, A.; Gao, Y.; Vidensky, S.; Brennan, J.; Poulsen, D.; Won Park, J.; et al. Presynaptic Regulation of Astroglial Excitatory Neurotransmitter Transporter GLT1. Neuron 2009, 61, 880–894. [Google Scholar] [CrossRef]
- Nakagawa, T.; Otsubo, Y.; Yatani, Y.; Shirakawa, H.; Kaneko, S. Mechanisms of Substrate Transport-induced Clustering of a Glial Glutamate Transporter GLT-1 in Astroglial–Neuronal Cultures. Eur J of Neuroscience 2008, 28, 1719–1730. [Google Scholar] [CrossRef]
- Zhou, J.; Sutherland, M.L. Glutamate Transporter Cluster Formation in Astrocytic Processes Regulates Glutamate Uptake Activity. J. Neurosci. 2004, 24, 6301–6306. [Google Scholar] [CrossRef]
- Scofield, M.D.; Li, H.; Siemsen, B.M.; Healey, K.L.; Tran, P.K.; Woronoff, N.; Boger, H.A.; Kalivas, P.W.; Reissner, K.J. Cocaine Self-Administration and Extinction Leads to Reduced Glial Fibrillary Acidic Protein Expression and Morphometric Features of Astrocytes in the Nucleus Accumbens Core. Biological Psychiatry 2016, 80, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Stellwagen, D.; Beattie, E.C.; Seo, J.Y.; Malenka, R.C. Differential Regulation of AMPA Receptor and GABA Receptor Trafficking by Tumor Necrosis Factor-α. J. Neurosci. 2005, 25, 3219–3228. [Google Scholar] [CrossRef]
- Pandey, G.N.; Rizavi, H.S.; Zhang, H.; Ren, X. Abnormal Gene and Protein Expression of Inflammatory Cytokines in the Postmortem Brain of Schizophrenia Patients. Schizophrenia Research 2018, 192, 247–254. [Google Scholar] [CrossRef]
- Uzzan, S.; Azab, A.N. Anti-TNF-α Compounds as a Treatment for Depression. Molecules 2021, 26, 2368. [Google Scholar] [CrossRef]
- Gupta, S.; Guleria, R.S. Involvement of Nuclear Factor-κB in Inflammation and Neuronal Plasticity Associated with Post-Traumatic Stress Disorder. Cells 2022, 11, 2034. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Meffert, M.K. Roles for NF-κB in Nerve Cell Survival, Plasticity, and Disease. Cell Death Differ 2006, 13, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, T.G. Role of Nuclear Factor Kappa B (NF-κB) Signalling in Neurodegenerative Diseases: An Mechanistic Approach. CN 2020, 18, 918–935. [Google Scholar] [CrossRef]
- Kaltschmidt, B.; Widera, D.; Kaltschmidt, C. Signaling via NF-κB in the Nervous System. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research 2005, 1745, 287–299. [Google Scholar] [CrossRef]
- Dresselhaus, E.C.; Meffert, M.K. Cellular Specificity of NF-κB Function in the Nervous System. Front. Immunol. 2019, 10, 1043. [Google Scholar] [CrossRef]
- Oeckinghaus, A.; Ghosh, S. The NF- B Family of Transcription Factors and Its Regulation. Cold Spring Harbor Perspectives in Biology 2009, 1, a000034–a000034. [Google Scholar] [CrossRef]
- Nennig, S.E.; Schank, J.R. The Role of NFkB in Drug Addiction: Beyond Inflammation. Alcohol and Alcoholism 2017, 52, 172–179. [Google Scholar] [CrossRef]
- Ang, E.; Chen, J.; Zagouras, P.; Magna, H.; Holland, J.; Schaeffer, E.; Nestler, E.J. Induction of Nuclear factor-κB in Nucleus Accumbens by Chronic Cocaine Administration. Journal of Neurochemistry 2001, 79, 221–224. [Google Scholar] [CrossRef]
- Russo, S.J.; Wilkinson, M.B.; Mazei-Robison, M.S.; Dietz, D.M.; Maze, I.; Krishnan, V.; Renthal, W.; Graham, A.; Birnbaum, S.G.; Green, T.A.; et al. Nuclear Factor κB Signaling Regulates Neuronal Morphology and Cocaine Reward. J. Neurosci. 2009, 29, 3529–3537. [Google Scholar] [CrossRef]
- Namba, M.D.; Phillips, M.N.; Neisewander, J.L.; Olive, M.F. Nuclear Factor Kappa B Signaling within the Rat Nucleus Accumbens Core Sex-Dependently Regulates Cue-Induced Cocaine Seeking and Matrix Metalloproteinase-9 Expression. Brain, Behavior, and Immunity 2022, 102, 252–265. [Google Scholar] [CrossRef]
- Wang, F.; Liu, H.; Ke, Y.; Huang, X.; Chen, S.; Zhuang, D.; Zhou, Y.; Wu, M.; Wang, Y.; Lai, M.; et al. Ibudilast-Mediated Suppression of Neuronal TLR4 in the Prefrontal Cortex Mitigates Methamphetamine-Induced Neuroinflammation and Addictive Behaviours. Addiction Biology 2025, 30, e70033. [Google Scholar] [CrossRef] [PubMed]
- Cohen, H.; Kozlovsky, N.; Matar, M.A.; Zohar, J.; Kaplan, Z. The Characteristic Long-Term Upregulation of Hippocampal NF-κB Complex in PTSD-Like Behavioral Stress Response Is Normalized by High-Dose Corticosterone and Pyrrolidine Dithiocarbamate Administered Immediately after Exposure. Neuropsychopharmacol 2011, 36, 2286–2302. [Google Scholar] [CrossRef]
- Zoppi, S.; Pérez Nievas, B.G.; Madrigal, J.L.M.; Manzanares, J.; Leza, J.C.; García-Bueno, B. Regulatory Role of Cannabinoid Receptor 1 in Stress-Induced Excitotoxicity and Neuroinflammation. Neuropsychopharmacol 2011, 36, 805–818. [Google Scholar] [CrossRef] [PubMed]
- Christoffel, D.J.; Golden, S.A.; Dumitriu, D.; Robison, A.J.; Janssen, W.G.; Ahn, H.F.; Krishnan, V.; Reyes, C.M.; Han, M.-H.; Ables, J.L.; et al. IκB Kinase Regulates Social Defeat Stress-Induced Synaptic and Behavioral Plasticity. J. Neurosci. 2011, 31, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Smaga, I.; Fierro, D.; Mesa, J.; Filip, M.; Knackstedt, L.A. Molecular Changes Evoked by the Beta-Lactam Antibiotic Ceftriaxone across Rodent Models of Substance Use Disorder and Neurological Disease. Neuroscience & Biobehavioral Reviews 2020, 115, 116–130. [Google Scholar] [CrossRef] [PubMed]
- Bessa, J.M.; Morais, M.; Marques, F.; Pinto, L.; Palha, J.A.; Almeida, O.F.X.; Sousa, N. Stress-Induced Anhedonia Is Associated with Hypertrophy of Medium Spiny Neurons of the Nucleus Accumbens. Transl Psychiatry 2013, 3, e266–e266. [Google Scholar] [CrossRef]
- Loweth, J.A.; Tseng, K.Y.; Wolf, M.E. Adaptations in AMPA Receptor Transmission in the Nucleus Accumbens Contributing to Incubation of Cocaine Craving. Neuropharmacology 2014, 76, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Nestler, E.J. The Neural Rejuvenation Hypothesis of Cocaine Addiction. Trends in Pharmacological Sciences 2014, 35, 374–383. [Google Scholar] [CrossRef]
- Taylor, S.B.; Anglin, J.M.; Paode, P.R.; Riggert, A.G.; Olive, M.F.; Conrad, C.D. Chronic Stress May Facilitate the Recruitment of Habit- and Addiction-Related Neurocircuitries through Neuronal Restructuring of the Striatum. Neuroscience 2014, 280, 231–242. [Google Scholar] [CrossRef]
- Brown, K.T.; Bachtell, R.K. Activation of the Immune System During a Developmental Window May Provide a Link Between Early Life Stress and Future Susceptibility to Cocaine Abuse. Biological Psychiatry 2018, 84, 865–866. [Google Scholar] [CrossRef]
- Snider, S.E.; Hendrick, E.S.; Beardsley, P.M. Glial Cell Modulators Attenuate Methamphetamine Self-Administration in Therat. European Journal of Pharmacology 2013, 701, 124–130. [Google Scholar] [CrossRef]
- Weber, M.D.; Frank, M.G.; Tracey, K.J.; Watkins, L.R.; Maier, S.F. Stress Induces the Danger-Associated Molecular Pattern HMGB-1 in the Hippocampus of Male Sprague Dawley Rats: A Priming Stimulus of Microglia and the NLRP3 Inflammasome. J. Neurosci. 2015, 35, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, A.; Kirchhoff, F.; Helmchen, F. Resting Microglial Cells Are Highly Dynamic Surveillants of Brain Parenchyma in Vivo. Science 2005, 308, 1314–1318. [Google Scholar] [CrossRef]
- Kessler, R.C. THE EFFECTS OF STRESSFUL LIFE EVENTS ON DEPRESSION. Annu. Rev. Psychol. 1997, 48, 191–214. [Google Scholar] [CrossRef]
- Gaspar, R.; Soares-Cunha, C.; Domingues, A.V.; Coimbra, B.; Baptista, F.I.; Pinto, L.; Ambrósio, A.F.; Rodrigues, A.J.; Gomes, C.A. Resilience to Stress and Sex-Specific Remodeling of Microglia and Neuronal Morphology in a Rat Model of Anxiety and Anhedonia. Neurobiology of Stress 2021, 14, 100302. [Google Scholar] [CrossRef]
- Nowak, D.B.; Taborda-Bejarano, J.P.; Chaure, F.J.; Mantsch, J.R.; Garcia-Keller, C. Understanding Microglia in Mesocorticolimbic Circuits: Implications for the Study of Chronic Stress and Substance Use Disorders. Cells 2025, 14, 1014. [Google Scholar] [CrossRef]
- Reverte, I.; Marchetti, C.; Pezza, S.; Zenoni, S.F.; Scaringi, G.; Ferrucci, L.; D’Ottavio, G.; Pignataro, A.; Andolina, D.; Raspa, M.; et al. Microglia-Mediated Calcium-Permeable AMPAR Accumulation in the Nucleus Accumbens Drives Hyperlocomotion during Cocaine Withdrawal. Brain, Behavior, and Immunity 2024, 115, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Spiga, S.; Mulas, G.; Piras, F.; Diana, M. The €œaddicted†Spine. Front. Neuroanat. 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Keck, T.; Mrsic-Flogel, T.D.; Vaz Afonso, M.; Eysel, U.T.; Bonhoeffer, T.; Hübener, M. Massive Restructuring of Neuronal Circuits during Functional Reorganization of Adult Visual Cortex. Nat Neurosci 2008, 11, 1162–1167. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Pan, F.; Gan, W.-B. Stably Maintained Dendritic Spines Are Associated with Lifelong Memories. Nature 2009, 462, 920–924. [Google Scholar] [CrossRef]
- Cahill, M.E.; Walker, D.M.; Gancarz, A.M.; Wang, Z.J.; Lardner, C.K.; Bagot, R.C.; Neve, R.L.; Dietz, D.M.; Nestler, E.J. The Dendritic Spine Morphogenic Effects of Repeated Cocaine Use Occur through the Regulation of Serum Response Factor Signaling. Mol Psychiatry 2018, 23, 1474–1486. [Google Scholar] [CrossRef]
- Kim, J.; Park, B.-H.; Lee, J.H.; Park, S.K.; Kim, J.-H. Cell Type-Specific Alterations in the Nucleus Accumbens by Repeated Exposures to Cocaine. Biological Psychiatry 2011, 69, 1026–1034. [Google Scholar] [CrossRef]
- Lee, K.-W.; Kim, Y.; Kim, A.M.; Helmin, K.; Nairn, A.C.; Greengard, P. Cocaine-Induced Dendritic Spine Formation in D1 and D2 Dopamine Receptor-Containing Medium Spiny Neurons in Nucleus Accumbens. Proc. Natl. Acad. Sci. U.S.A. 2006, 103, 3399–3404. [Google Scholar] [CrossRef]
- Robinson, T.E.; Kolb, B. Alterations in the Morphology of Dendrites and Dendritic Spines in the Nucleus Accumbens and Prefrontal Cortex Following Repeated Treatment with Amphetamine or Cocaine. Eur J of Neuroscience 1999, 11, 1598–1604. [Google Scholar] [CrossRef]
- Toda, S.; Shen, H.; Kalivas, P.W. Inhibition of Actin Polymerization Prevents Cocaine-Induced Changes in Spine Morphology in the Nucleus Accumbens. Neurotox Res 2010, 18, 410–415. [Google Scholar] [CrossRef]
- Kourrich, S.; Rothwell, P.E.; Klug, J.R.; Thomas, M.J. Cocaine Experience Controls Bidirectional Synaptic Plasticity in the Nucleus Accumbens. J. Neurosci. 2007, 27, 7921–7928. [Google Scholar] [CrossRef]
- Shen, H.; Moussawi, K.; Zhou, W.; Toda, S.; Kalivas, P.W. Heroin Relapse Requires Long-Term Potentiation-like Plasticity Mediated by NMDA2b-Containing Receptors. Proceedings of the National Academy of Sciences 2011, 108, 19407–19412. [Google Scholar] [CrossRef]
- Gipson, C.D.; Kupchik, Y.M.; Kalivas, P.W. Rapid, Transient Synaptic Plasticity in Addiction. Neuropharmacology 2014, 76, 276–286. [Google Scholar] [CrossRef]
- Anderson, S.M.; Famous, K.R.; Sadri-Vakili, G.; Kumaresan, V.; Schmidt, H.D.; Bass, C.E.; Terwilliger, E.F.; Cha, J.-H.J.; Pierce, R.C. CaMKII: A Biochemical Bridge Linking Accumbens Dopamine and Glutamate Systems in Cocaine Seeking. Nat Neurosci 2008, 11, 344–353. [Google Scholar] [CrossRef]
- Boudreau, A.C.; Reimers, J.M.; Milovanovic, M.; Wolf, M.E. Cell Surface AMPA Receptors in the Rat Nucleus Accumbens Increase during Cocaine Withdrawal But Internalize after Cocaine Challenge in Association with Altered Activation of Mitogen-Activated Protein Kinases. J. Neurosci. 2007, 27, 10621–10635. [Google Scholar] [CrossRef]
- Spencer, S.; Garcia-Keller, C.; Roberts-Wolfe, D.; Heinsbroek, J.A.; Mulvaney, M.; Sorrell, A.; Kalivas, P.W. Cocaine Use Reverses Striatal Plasticity Produced During Cocaine Seeking. Biological Psychiatry 2017, 81, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Kasai, H.; Fukuda, M.; Watanabe, S.; Hayashi-Takagi, A.; Noguchi, J. Structural Dynamics of Dendritic Spines in Memory and Cognition. Trends in Neurosciences 2010, 33, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.E.; Figueiredo, A.; Menken, M.S.; Lobo, M.K. Dendritic Spine Density Is Increased on Nucleus Accumbens D2 Neurons after Chronic Social Defeat. Sci Rep 2020, 10, 12393. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.M.; Kater, S.B. Dendritic Spines: Cellular Specializations Imparting Both Stability and Flexibility to Synaptic Function. Annu. Rev. Neurosci. 1994, 17, 341–371. [Google Scholar] [CrossRef]
- Golden, S.A.; Russo, S.J. Mechanisms of Psychostimulant-Induced Structural Plasticity. Cold Spring Harbor Perspectives in Medicine 2012, 2, a011957–a011957. [Google Scholar] [CrossRef] [PubMed]
- Kopec, C.D.; Li, B.; Wei, W.; Boehm, J.; Malinow, R. Glutamate Receptor Exocytosis and Spine Enlargement during Chemically Induced Long-Term Potentiation. J. Neurosci. 2006, 26, 2000–2009. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Penick, E.C.; Edwards, J.G.; Kauer, J.A.; Ehlers, M.D. Recycling Endosomes Supply AMPA Receptors for LTP. Science 2004, 305, 1972–1975. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Salgado, J.M.; Ostroff, L.; Helton, T.D.; Robinson, C.G.; Harris, K.M.; Ehlers, M.D. Plasticity-Induced Growth of Dendritic Spines by Exocytic Trafficking from Recycling Endosomes. Neuron 2006, 52, 817–830. [Google Scholar] [CrossRef]
- Kasai, H. Unraveling the Mysteries of Dendritic Spine Dynamics: Five Key Principles Shaping Memory and Cognition. Proc. Jpn. Acad., Ser. B 2023, 99, 254–305. [Google Scholar] [CrossRef]
- Matsuzaki, M.; Ellis-Davies, G.C.R.; Nemoto, T.; Miyashita, Y.; Iino, M.; Kasai, H. Dendritic Spine Geometry Is Critical for AMPA Receptor Expression in Hippocampal CA1 Pyramidal Neurons. Nat Neurosci 2001, 4, 1086–1092. [Google Scholar] [CrossRef]
- Russo, S.J.; Dietz, D.M.; Dumitriu, D.; Morrison, J.H.; Malenka, R.C.; Nestler, E.J. The Addicted Synapse: Mechanisms of Synaptic and Structural Plasticity in Nucleus Accumbens. Trends in Neurosciences 2010, 33, 267–276. [Google Scholar] [CrossRef]
- De, A.; Krueger, J.M.; Simasko, S.M. Glutamate Induces the Expression and Release of Tumor Necrosis Factor-α in Cultured Hypothalamic Cells. Brain Research 2005, 1053, 54–61. [Google Scholar] [CrossRef]
- Taylor, D.L.; Jones, F.; Kubota, E.S.F.C.S.; Pocock, J.M. Stimulation of Microglial Metabotropic Glutamate Receptor mGlu2 Triggers Tumor Necrosis Factor α-Induced Neurotoxicity in Concert with Microglial-Derived Fas Ligand. J. Neurosci. 2005, 25, 2952–2964. [Google Scholar] [CrossRef]
- Beattie, E.C.; Stellwagen, D.; Morishita, W.; Bresnahan, J.C.; Ha, B.K.; Von Zastrow, M.; Beattie, M.S.; Malenka, R.C. Control of Synaptic Strength by Glial TNFα. Science 2002, 295, 2282–2285. [Google Scholar] [CrossRef]
- Albensi, B.C.; Mattson, M.P. Evidence for the Involvement of TNF and NF-?B in Hippocampal Synaptic Plasticity. Synapse 2000, 35, 151–159. [Google Scholar] [CrossRef]
- Loscher, C.E.; Mills, K.H.G.; Lynch, M.A. Interleukin-1 Receptor Antagonist Exerts Agonist Activity in the Hippocampus Independent of the Interleukin-1 Type I Receptor. Journal of Neuroimmunology 2003, 137, 117–124. [Google Scholar] [CrossRef]
- Schmid, A.W.; Lynch, M.A.; Herron, C.E. The Effects of IL-1 Receptor Antagonist on Beta Amyloid Mediated Depression of LTP in the Rat CA1 in Vivo. Hippocampus 2009, 19, 670–676. [Google Scholar] [CrossRef]
- Vereker, E.; O’Donnell, E.; Lynch, M.A. The Inhibitory Effect of Interleukin-1β on Long-Term Potentiation Is Coupled with Increased Activity of Stress-Activated Protein Kinases. J. Neurosci. 2000, 20, 6811–6819. [Google Scholar] [CrossRef] [PubMed]
- Ikegaya, Y.; Delcroix, I.; Iwakura, Y.; Matsuki, N.; Nishiyama, N. Interleukin-1β Abrogates Long-term Depression of Hippocampal CA1 Synaptic Transmission. Synapse 2003, 47, 54–57. [Google Scholar] [CrossRef]
- Lai, A.Y.; Swayze, R.D.; El-Husseini, A.; Song, C. Interleukin-1 Beta Modulates AMPA Receptor Expression and Phosphorylation in Hippocampal Neurons. Journal of Neuroimmunology 2006, 175, 97–106. [Google Scholar] [CrossRef]
- Mir, S.; Sen, T.; Sen, N. Cytokine-Induced GAPDH Sulfhydration Affects PSD95 Degradation and Memory. Molecular Cell 2014, 56, 786–795. [Google Scholar] [CrossRef]
- Dean, O.; Giorlando, F.; Berk, M. N -Acetylcysteine in Psychiatry: Current Therapeutic Evidence and Potential Mechanisms of Action. J Psychiatry Neurosci 2011, 36, 78–86. [Google Scholar] [CrossRef]
- Viña, J.; Romero, F.J.; Saez, G.T.; Pallardó, F.V. Effects of Cysteine and N-Acetyl Cysteine on GSH Content of Brain of Adult Rats. Experientia 1983, 39, 164–165. [Google Scholar] [CrossRef]
- Yin, J.; Ren, W.; Yang, G.; Duan, J.; Huang, X.; Fang, R.; Li, C.; Li, T.; Yin, Y.; Hou, Y.; et al. l -Cysteine Metabolism and Its Nutritional Implications. Molecular Nutrition Food Res 2016, 60, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Pallanti, S.; Grassi, G. Pharmacologic Treatment of Obsessive-Compulsive Disorder Comorbidity. Expert Opinion on Pharmacotherapy 2014, 15, 2543–2552. [Google Scholar] [CrossRef]
- Ramkissoon, A.; Wells, P.G. Methamphetamine Oxidative Stress, Neurotoxicity, and Functional Deficits Are Modulated by Nuclear Factor-E2-Related Factor 2. Free Radical Biology and Medicine 2015, 89, 358–368. [Google Scholar] [CrossRef]
- Huang, Y.-N.; Yang, L.-Y.; Wang, J.-Y.; Lai, C.-C.; Chiu, C.-T.; Wang, J.-Y. L-Ascorbate Protects Against Methamphetamine-Induced Neurotoxicity of Cortical Cells via Inhibiting Oxidative Stress, Autophagy, and Apoptosis. Mol Neurobiol 2017, 54, 125–136. [Google Scholar] [CrossRef]
- Jang, E.Y.; Yang, C.H.; Hedges, D.M.; Kim, S.P.; Lee, J.Y.; Ekins, T.G.; Garcia, B.T.; Kim, H.Y.; Nelson, A.C.; Kim, N.J.; et al. The Role of Reactive Oxygen Species in Methamphetamine Self-Administration and Dopamine Release in the Nucleus Accumbens: ROS in METH-Taking Behavior. Addiction Biology 2017, 22, 1304–1315. [Google Scholar] [CrossRef] [PubMed]
- Madayag, A.; Lobner, D.; Kau, K.S.; Mantsch, J.R.; Abdulhameed, O.; Hearing, M.; Grier, M.D.; Baker, D.A. Repeated N -Acetylcysteine Administration Alters Plasticity-Dependent Effects of Cocaine. J. Neurosci. 2007, 27, 13968–13976. [Google Scholar] [CrossRef]
- Murray, J.E.; Lacoste, J.; Belin, D. N-Acetylcysteine as a Treatment for Addiction. In Addictions—from pathophysiology to treatment; InTech: Rijeka, Croatia, 2012; pp. 355–380. [Google Scholar]
- Ducret, E.; Puaud, M.; Lacoste, J.; Belin-Rauscent, A.; Fouyssac, M.; Dugast, E.; Murray, J.E.; Everitt, B.J.; Houeto, J.-L.; Belin, D. N-Acetylcysteine Facilitates Self-Imposed Abstinence After Escalation of Cocaine Intake. Biological Psychiatry 2016, 80, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Keller, C.; Smiley, C.; Monforton, C.; Melton, S.; Kalivas, P.W.; Gass, J. N -Acetylcysteine Treatment during Acute Stress Prevents Stress-induced Augmentation of Addictive Drug Use and Relapse. Addiction Biology 2020, 25, e12798. [Google Scholar] [CrossRef]
- Kau, K.S.; Madayag, A.; Mantsch, J.R.; Grier, M.D.; Abdulhameed, O.; Baker, D.A. Blunted Cystine–Glutamate Antiporter Function in the Nucleus Accumbens Promotes Cocaine-Induced Drug Seeking. Neuroscience 2008, 155, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Moussawi, K.; Pacchioni, A.; Moran, M.; Olive, M.F.; Gass, J.T.; Lavin, A.; Kalivas, P.W. N-Acetylcysteine Reverses Cocaine-Induced Metaplasticity. Nat Neurosci 2009, 12, 182–189. [Google Scholar] [CrossRef]
- Kupchik, Y.M.; Moussawi, K.; Tang, X.-C.; Wang, X.; Kalivas, B.C.; Kolokithas, R.; Ogburn, K.B.; Kalivas, P.W. The Effect of N-Acetylcysteine in the Nucleus Accumbens on Neurotransmission and Relapse to Cocaine. Biological Psychiatry 2012, 71, 978–986. [Google Scholar] [CrossRef]
- LaRowe, S.D.; Mardikian, P.; Malcolm, R.; Myrick, H.; Kalivas, P.; McFarland, K.; Saladin, M.; McRae, A.; Brady, K. Safety and Tolerability of N-Acetylcysteine in Cocaine-Dependent Individuals. American J Addict 2006, 15, 105–110. [Google Scholar] [CrossRef]
- LaRowe, S.D.; Myrick, H.; Hedden, S.; Mardikian, P.; Saladin, M.; McRae, A.; Brady, K.; Kalivas, P.W.; Malcolm, R. Is Cocaine Desire Reduced by N -Acetylcysteine? AJP 2007, 164, 1115–1117. [Google Scholar] [CrossRef] [PubMed]
- Mardikian, P.N.; LaRowe, S.D.; Hedden, S.; Kalivas, P.W.; Malcolm, R.J. An Open-Label Trial of N-Acetylcysteine for the Treatment of Cocaine Dependence: A Pilot Study. Progress in Neuro-Psychopharmacology and Biological Psychiatry 2007, 31, 389–394. [Google Scholar] [CrossRef]
- Amen, S.L.; Piacentine, L.B.; Ahmad, M.E.; Li, S.-J.; Mantsch, J.R.; Risinger, R.C.; Baker, D.A. Repeated N-Acetyl Cysteine Reduces Cocaine Seeking in Rodents and Craving in Cocaine-Dependent Humans. Neuropsychopharmacol 2011, 36, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Schmaal, L.; Veltman, D.J.; Nederveen, A.; Van Den Brink, W.; Goudriaan, A.E. N-Acetylcysteine Normalizes Glutamate Levels in Cocaine-Dependent Patients: A Randomized Crossover Magnetic Resonance Spectroscopy Study. Neuropsychopharmacol 2012, 37, 2143–2152. [Google Scholar] [CrossRef]
- LaRowe, S.D.; Kalivas, P.W.; Nicholas, J.S.; Randall, P.K.; Mardikian, P.N.; Malcolm, Robert.J. A Double-blind Placebo-controlled Trial of N-acetylcysteine in the Treatment of Cocaine Dependence. American J Addict 2013, 22, 443–452. [Google Scholar] [CrossRef]
- Engeli, E.J.E.; Preller, K.H.; Rieser, N.M.; Klar, J.; Staempfli, P.; Hulka, L.M.; Kirschner, M.; Seifritz, E.; Herdener, M. N-Acetylcysteine Reduces Prefrontal Reactivity to Cocaine Cues in Individuals with Cocaine Use Disorder. Front. Psychiatry 2025, 15, 1489194. [Google Scholar] [CrossRef] [PubMed]
- Back, S.E.; McCauley, J.L.; Korte, K.J.; Gros, D.F.; Leavitt, V.; Gray, K.M.; Hamner, M.B.; DeSantis, S.M.; Malcolm, R.; Brady, K.T.; et al. A Double-Blind, Randomized, Controlled Pilot Trial of N-Acetylcysteine in Veterans With Posttraumatic Stress Disorder and Substance Use Disorders. J. Clin. Psychiatry 2016, 77, e1439–e1446. [Google Scholar] [CrossRef]
- Palacio, J.R.; Markert, U.R.; Martínez, P. Anti-Inflammatory Properties of N-Acetylcysteine on Lipopolysaccharide-Activated Macrophages. Inflamm. Res. 2011, 60, 695–704. [Google Scholar] [CrossRef]
- Zheng, W.; Zhang, Q. -E.; Cai, D. -B.; Yang, X. -H.; Qiu, Y.; Ungvari, G.S.; Ng, C.H.; Berk, M.; Ning, Y. -P.; Xiang, Y. -T. N -acetylcysteine for Major Mental Disorders: A Systematic Review and Meta-analysis of Randomized Controlled Trials. Acta Psychiatr Scand 2018, 137, 391–400. [Google Scholar] [CrossRef]
- Möller, M.; Du Preez, J.L.; Viljoen, F.P.; Berk, M.; Harvey, B.H. N-Acetyl Cysteine Reverses Social Isolation Rearing Induced Changes in Cortico-Striatal Monoamines in Rats. Metab Brain Dis 2013, 28, 687–696. [Google Scholar] [CrossRef]
- Baker, D.G.; Nievergelt, C.M.; O’Connor, D.T. Biomarkers of PTSD: Neuropeptides and Immune Signaling. Neuropharmacology 2012, 62, 663–673. [Google Scholar] [CrossRef]
- Reiner, R.; Weiss, U.; Brombacher, U.; Lanz, P.; Montavon, M.; Furlenmeier, A.; Angehrn, P.; Probst, P.J. Ro 13-9904/001, a Novel Potent and Long-Acting Parenteral Cephalosporin. J. Antibiot. 1980, 33, 783–786. [Google Scholar] [CrossRef]
- Beskid, G.; Christenson, J.G.; Cleeland, R.; DeLorenzo, W.; Trown, P.W. In Vivo Activity of Ceftriaxone (Ro 13-9904), a New Broad-Spectrum Semisynthetic Cephalosporin. Antimicrob Agents Chemother 1981, 20, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Rothstein, J.D.; Patel, S.; Regan, M.R.; Haenggeli, C.; Huang, Y.H.; Bergles, D.E.; Jin, L.; Dykes Hoberg, M.; Vidensky, S.; Chung, D.S.; et al. β-Lactam Antibiotics Offer Neuroprotection by Increasing Glutamate Transporter Expression. Nature 2005, 433, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Sari, Y.; Smith, K.D.; Ali, P.K.; Rebec, G.V. Upregulation of Glt1 Attenuates Cue-Induced Reinstatement of Cocaine-Seeking Behavior in Rats. J. Neurosci. 2009, 29, 9239–9243. [Google Scholar] [CrossRef]
- Sondheimer, I.; Knackstedt, L.A. Ceftriaxone Prevents the Induction of Cocaine Sensitization and Produces Enduring Attenuation of Cue- and Cocaine-Primed Reinstatement of Cocaine-Seeking. Behavioural Brain Research 2011, 225, 252–258. [Google Scholar] [CrossRef]
- Rasmussen, B.; Unterwald, E.M.; Rawls, S.M. Glutamate Transporter Subtype 1 (GLT-1) Activator Ceftriaxone Attenuates Amphetamine-Induced Hyperactivity and Behavioral Sensitization in Rats. Drug and Alcohol Dependence 2011, 118, 484–488. [Google Scholar] [CrossRef]
- LaCrosse, A.L.; Hill, K.; Knackstedt, L.A. Ceftriaxone Attenuates Cocaine Relapse after Abstinence through Modulation of Nucleus Accumbens AMPA Subunit Expression. European Neuropsychopharmacology 2016, 26, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.D.; Houston, A.C.W.; Rebec, G.V. Role of the Major Glutamate Transporter GLT1 in Nucleus Accumbens Core Versus Shell in Cue-Induced Cocaine-Seeking Behavior. J. Neurosci. 2013, 33, 9319–9327. [Google Scholar] [CrossRef]
- Lee, S.-G.; Su, Z.-Z.; Emdad, L.; Gupta, P.; Sarkar, D.; Borjabad, A.; Volsky, D.J.; Fisher, P.B. Mechanism of Ceftriaxone Induction of Excitatory Amino Acid Transporter-2 Expression and Glutamate Uptake in Primary Human Astrocytes. Journal of Biological Chemistry 2008, 283, 13116–13123. [Google Scholar] [CrossRef]
- Abdel-Salam, O. Drugs Used to Treat Parkinsons Disease, Present Status and Future Directions. CNSNDDT 2008, 7, 321–342. [Google Scholar] [CrossRef]
- Blum, D.; Chtarto, A.; Tenenbaum, L.; Brotchi, J.; Levivier, M. Clinical Potential of Minocycline for Neurodegenerative Disorders. Neurobiology of Disease 2004, 17, 359–366. [Google Scholar] [CrossRef]
- Buller, K.M.; Carty, M.L.; Reinebrant, H.E.; Wixey, J.A. Minocycline: A Neuroprotective Agent for Hypoxic-ischemic Brain Injury in the Neonate? J of Neuroscience Research 2009, 87, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ma, X.; Jiang, Y.; Pi, R.; Liu, Y.; Ma, L. The Prospects of Minocycline in Multiple Sclerosis. Journal of Neuroimmunology 2011, 235, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-S.; Suh, Y.-H. Minocycline and Neurodegenerative Diseases. Behavioural Brain Research 2009, 196, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Miyaoka, T.; Yasukawa, R.; Yasuda, H.; Hayashida, M.; Inagaki, T.; Horiguchi, J. Possible Antipsychotic Effects of Minocycline in Patients with Schizophrenia. Progress in Neuro-Psychopharmacology and Biological Psychiatry 2007, 31, 304–307. [Google Scholar] [CrossRef]
- Soczynska, J.K.; Mansur, R.B.; Brietzke, E.; Swardfager, W.; Kennedy, S.H.; Woldeyohannes, H.O.; Powell, A.M.; Manierka, M.S.; McIntyre, R.S. Novel Therapeutic Targets in Depression: Minocycline as a Candidate Treatment. Behavioural Brain Research 2012, 235, 302–317. [Google Scholar] [CrossRef]
- Stirling, D.P.; Koochesfahani, K.M.; Steeves, J.D.; Tetzlaff, W. Minocycline as a Neuroprotective Agent. Neuroscientist 2005, 11, 308–322. [Google Scholar] [CrossRef]
- Yenari, M.A.; Kauppinen, T.M.; Swanson, R.A. Microglial Activation in Stroke: Therapeutic Targets. Neurotherapeutics 2010, 7, 378–391. [Google Scholar] [CrossRef]
- Möller, T.; Bard, F.; Bhattacharya, A.; Biber, K.; Campbell, B.; Dale, E.; Eder, C.; Gan, L.; Garden, G.A.; Hughes, Z.A.; et al. Critical Data-based Re-evaluation of Minocycline as a Putative Specific Microglia Inhibitor. Glia 2016, 64, 1788–1794. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Imagama, S.; Ohgomori, T.; Hirano, K.; Uchimura, K.; Sakamoto, K.; Hirakawa, A.; Takeuchi, H.; Suzumura, A.; Ishiguro, N.; et al. Minocycline Selectively Inhibits M1 Polarization of Microglia. Cell Death Dis 2013, 4, e525–e525. [Google Scholar] [CrossRef] [PubMed]
- Moini-Zanjani, T.; Ostad, S.-N.; Labibi, F.; Ameli, H.; Mosaffa, N.; Sabetkasaei, M. Minocycline Effects on IL-6 Concentration in Macrophage and Microglial Cells in a Rat Model of Neuropathic Pain. Iran Biomed J 2016, 20, 273–279. [Google Scholar] [CrossRef]
- Switzer, J.A.; Sikora, A.; Ergul, A.; Waller, J.L.; Hess, D.C.; Fagan, S.C. Minocycline Prevents IL-6 Increase after Acute Ischemic Stroke. Transl. Stroke Res. 2012, 3, 364–368. [Google Scholar] [CrossRef]
- Wang, H.-T.; Huang, F.-L.; Hu, Z.-L.; Zhang, W.-J.; Qiao, X.-Q.; Huang, Y.-Q.; Dai, R.-P.; Li, F.; Li, C.-Q. Early-Life Social Isolation-Induced Depressive-Like Behavior in Rats Results in Microglial Activation and Neuronal Histone Methylation That Are Mitigated by Minocycline. Neurotox Res 2017, 31, 505–520. [Google Scholar] [CrossRef]
- Piotrowska, A.; Kwiatkowski, K.; Rojewska, E.; Slusarczyk, J.; Makuch, W.; Basta-Kaim, A.; Przewlocka, B.; Mika, J. Direct and Indirect Pharmacological Modulation of CCL2/CCR2 Pathway Results in Attenuation of Neuropathic Pain — In Vivo and in Vitro Evidence. Journal of Neuroimmunology 2016, 297, 9–19. [Google Scholar] [CrossRef]
- Chen, M.; Ona, V.O.; Li, M.; Ferrante, R.J.; Fink, K.B.; Zhu, S.; Bian, J.; Guo, L.; Farrell, L.A.; Hersch, S.M.; et al. Minocycline Inhibits Caspase-1 and Caspase-3 Expression and Delays Mortality in a Transgenic Mouse Model of Huntington Disease. Nat Med 2000, 6, 797–801. [Google Scholar] [CrossRef]
- Wasserman, J.K.; Schlichter, L.C. Minocycline Protects the Blood–Brain Barrier and Reduces Edema Following Intracerebral Hemorrhage in the Rat. Experimental Neurology 2007, 207, 227–237. [Google Scholar] [CrossRef]
- Zhang, L.; Kitaichi, K.; Fujimoto, Y.; Nakayama, H.; Shimizu, E.; Iyo, M.; Hashimoto, K. Protective Effects of Minocycline on Behavioral Changes and Neurotoxicity in Mice after Administration of Methamphetamine. Progress in Neuro-Psychopharmacology and Biological Psychiatry 2006, 30, 1381–1393. [Google Scholar] [CrossRef]
- Kofman, O.; Klein, E.; Newman, M.; Hamburger, R.; Kimchi, O.; Nir, T.; Shimon, H.; Belmaker, R.H. Inhibition by Antibiotic Tetracyclines of Rat Cortical Noradrenergic Adenylate Cyclase and Amphetamine-Induced Hyperactivity. Pharmacology Biochemistry and Behavior 1990, 37, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Uz, T.; Manev, H. Minocycline Affects Cocaine Sensitization in Mice. Neuroscience Letters 2009, 452, 258–261. [Google Scholar] [CrossRef]
- Chen, H.; Manev, H. Effects of Minocycline on Cocaine Sensitization and Phosphorylation of GluR1 Receptors in 5-Lipoxygenase Deficient Mice. Neuropharmacology 2011, 60, 1058–1063. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liao, Y.; Dong, Y.; Li, X.; Li, J.; Cheng, Y.; Cheng, J.; Yuan, Z. Microglial Deletion and Inhibition Alleviate Behavior of Post-Traumatic Stress Disorder in Mice. J Neuroinflammation 2021, 18, 7. [Google Scholar] [CrossRef] [PubMed]
- Levkovitz, Y.; Fenchel, D.; Kaplan, Z.; Zohar, J.; Cohen, H. Early Post-Stressor Intervention with Minocycline, a Second-Generation Tetracycline, Attenuates Post-Traumatic Stress Response in an Animal Model of PTSD. European Neuropsychopharmacology 2015, 25, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Gerst, A.; Murthy, T.; Elson, A.; Driscoll, D.; Bittner, M.J.; Ramaswamy, S. Adjunctive Minocycline for Treatment of Posttraumatic Stress Disorder. WMJ 2021, 120, 321–324. [Google Scholar]
- Dean, O.M.; Kanchanatawan, B.; Ashton, M.; Mohebbi, M.; Ng, C.H.; Maes, M.; Berk, L.; Sughondhabirom, A.; Tangwongchai, S.; Singh, A.B.; et al. Adjunctive Minocycline Treatment for Major Depressive Disorder: A Proof of Concept Trial. Aust N Z J Psychiatry 2017, 51, 829–840. [Google Scholar] [CrossRef]
- Husain, M.I.; Chaudhry, I.B.; Husain, N.; Khoso, A.B.; Rahman, R.R.; Hamirani, M.M.; Hodsoll, J.; Qurashi, I.; Deakin, J.F.; Young, A.H. Minocycline as an Adjunct for Treatment-Resistant Depressive Symptoms: A Pilot Randomised Placebo-Controlled Trial. J Psychopharmacol 2017, 31, 1166–1175. [Google Scholar] [CrossRef]
- Nettis, M.A. Minocycline in Major Depressive Disorder: And Overview with Considerations on Treatment-Resistance and Comparisons with Other Psychiatric Disorders. Brain, Behavior, & Immunity - Health 2021, 17, 100335. [Google Scholar] [CrossRef]
- Cui, X.; Li, L.; Hu, Y.-Y.; Ren, S.; Zhang, M.; Li, W.-B. Sulbactam Plays Neuronal Protective Effect Against Brain Ischemia via Upregulating GLT1 in Rats. Mol Neurobiol 2015, 51, 1322–1333. [Google Scholar] [CrossRef]
- Qi, J.; Xian, X.-H.; Li, L.; Zhang, M.; Hu, Y.-Y.; Zhang, J.-G.; Li, W.-B. Sulbactam Protects Hippocampal Neurons Against Oxygen-Glucose Deprivation by Up-Regulating Astrocytic GLT-1 via P38 MAPK Signal Pathway. Front. Mol. Neurosci. 2018, 11, 281. [Google Scholar] [CrossRef]
- Li, L.; Li, W.; Jiang, W.; Xu, R. Sulbactam Protects Neurons against Double Neurotoxicity of Amyloid Beta and Glutamate Load by Upregulating Glial Glutamate Transporter 1. Cell Death Discov. 2024, 10, 64. [Google Scholar] [CrossRef]
- Rao, P.S.S.; Goodwani, S.; Bell, R.L.; Wei, Y.; Boddu, S.H.S.; Sari, Y. Effects of Ampicillin, Cefazolin and Cefoperazone Treatments on GLT-1 Expressions in the Mesocorticolimbic System and Ethanol Intake in Alcohol-Preferring Rats. Neuroscience 2015, 295, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Alasmari, F.; Abuhamdah, S.; Sari, Y. Effects of Ampicillin on Cystine/Glutamate Antiporter and Glutamate Transporter 1 Isoforms as Well as Ethanol Drinking in Male P Rats. Neuroscience Letters 2015, 600, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Alasmari, F.; Alhaddad, H.; Wong, W.; Bell, R.L.; Sari, Y. Ampicillin/Sulbactam Treatment Modulates NMDA Receptor NR2B Subunit and Attenuates Neuroinflammation and Alcohol Intake in Male High Alcohol Drinking Rats. Biomolecules 2020, 10, 1030. [Google Scholar] [CrossRef] [PubMed]
- Hammad, A.M.; Alasmari, F.; Althobaiti, Y.S.; Sari, Y. Modulatory Effects of Ampicillin/Sulbactam on Glial Glutamate Transporters and Metabotropic Glutamate Receptor 1 as Well as Reinstatement to Cocaine-Seeking Behavior. Behavioural Brain Research 2017, 332, 288–298. [Google Scholar] [CrossRef]
- Hammad, A.M.; Alasmari, F.; Sari, Y. Effect of Modulation of the Astrocytic Glutamate Transporters’ Expression on Cocaine-Induced Reinstatement in Male P Rats Exposed to Ethanol. Alcohol and Alcoholism 2021, 56, 210–219. [Google Scholar] [CrossRef]
Figure 1.
The motivation circuit. This panel presents the organization of the motivation circuit, detailing the key nuclei and the directional projections that link limbic processing with pyramidal and extrapyramidal motor outputs. As illustrated, the NAc core functions as a central hub where reward- and aversion-related information converges. Motivational states are translated into action through the integration of inputs from regions such as the amygdala, hippocampus, and prefrontal cortex, with DA contributing to the assignment of motivational significance to goals. VP: ventral pallidum; PFC: prefrontal cortex; SN: substantia nigra; VTA: ventral tegmental area; Thal: thalamus. Modified from Pierce and Kalivas, 1997. This figure was created by BioRender.com.
Figure 1.
The motivation circuit. This panel presents the organization of the motivation circuit, detailing the key nuclei and the directional projections that link limbic processing with pyramidal and extrapyramidal motor outputs. As illustrated, the NAc core functions as a central hub where reward- and aversion-related information converges. Motivational states are translated into action through the integration of inputs from regions such as the amygdala, hippocampus, and prefrontal cortex, with DA contributing to the assignment of motivational significance to goals. VP: ventral pallidum; PFC: prefrontal cortex; SN: substantia nigra; VTA: ventral tegmental area; Thal: thalamus. Modified from Pierce and Kalivas, 1997. This figure was created by BioRender.com.
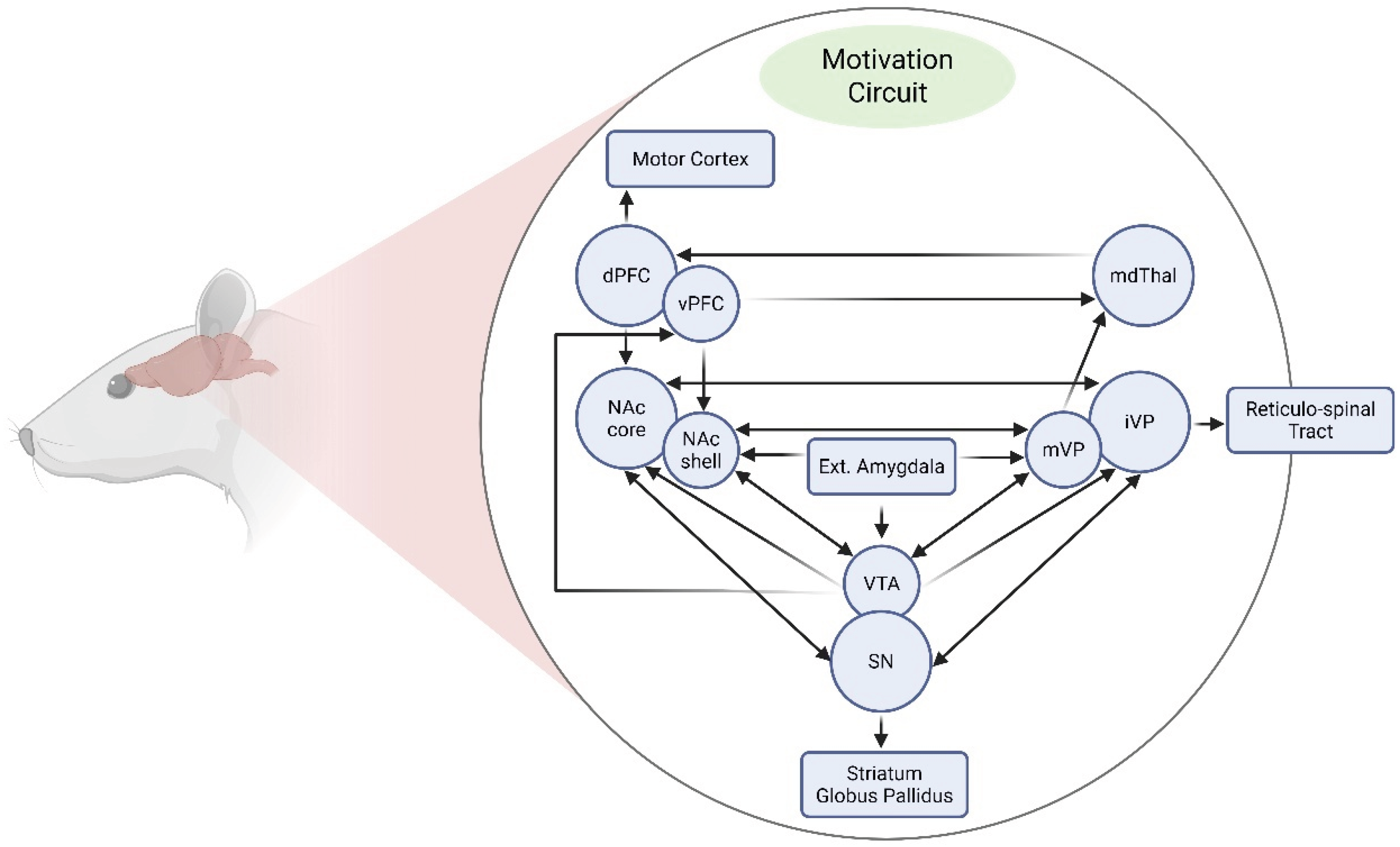
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



