
Submitted:
24 November 2025
Posted:
25 November 2025
You are already at the latest version
Abstract
Curcumin, a natural polyphenol derived from Curcuma longa, is widely recognized for its therapeutic properties. However, its clinical utility is limited because of poor solubility, rapid degradation and hence low bioavailability. To overcome these issues, nanoformulation approaches, especially PEGylated liposomes, have been explored as advanced delivery systems. PEGylation, which involves attaching polyethylene glycol (PEG) to the liposomal surface, enhances circulation time by creating a steric shield that reduces protein interactions and clearance by the mononuclear phagocyte system (MPS). However, PEG can alter lipid membrane properties, which may in turn affect curcumin’s solubility and distribution within the liposomal bilayer, ultimately reducing its loading efficiency. To ensure that PEG-modified liposomes can be effectively loaded with curcumin, we investigated curcumin–membrane interactions in saturated (DMPC) and unsaturated (POPC) liposomes, both in the presence and absence of PEG. Based on dissociation constants (Kd) obtained from fluorescence spectroscopy measurements, we found that PEGylated DMPC liposomes exhibit the strongest binding affinity for curcumin. Fluorescence quenching experiments showed that curcumin adopts a transbilayer orientation in all membranes examined. Curcumin’s location within PEGylated and non-PEGylated liposomal membranes was further confirmed by examining its effects on membrane properties, including fluidity, polarity, and oxygen transport. These effects were investigated using electron paramagnetic resonance (EPR) spectroscopy with spin labels. The results indicate that PEG does not impose major changes on membrane properties. Curcumin, however, was found to reinforce the liposomal membranes, increase their polarity, and reduce oxygen availability. Overall, the findings suggest that liposomes, particularly those composed of PEGylated DMPC, are effective vehicles for curcumin delivery.
Keywords:
membrane
; liposome
; curcumin
; PEG
; electron paramagnetic resonance (EPR)
; spin labels
; fluorescence
1. Introduction
Curcumin, a polyphenolic compound derived from Curcuma longa, has garnered widespread interest for its diverse pharmacological effects, notably its anti-inflammatory, antioxidant, and anticancer potential [1]. Nonetheless, its therapeutic use is hindered by poor aqueous solubility, low chemical stability, and limited bioavailability [2,3]. To address these limitations, several nano formulation approaches have been investigated, with liposomes standing out as one of the most promising drug delivery systems [4].
Liposomes are nanoscale vesicles composed of phospholipids, known for their biocompatibility, biodegradability, and low toxicity [5]. They are extensively explored as drug delivery systems and have led to several clinical applications, particularly in cancer chemotherapy and the treatment of serious fungal infections [6]. However, conventional liposomes are rapidly cleared from circulation through uptake by cells of the mononuclear phagocyte system (MPS), primarily in the liver and spleen. This rapid MPS uptake greatly limits their distribution to other tissues and may even exert toxic effects on MPS organs [7]. Therefore, to prolong liposome circulation time in the bloodstream, various modifications have been explored. One of the most effective strategies is PEGylation, in which polyethylene glycol (PEG) chains are covalently attached to phosphatidylethanolamine (PE) lipids incorporated into the liposomal membrane [8]. Another critical challenge in liposome technology is achieving high drug loading efficiency. The extent to which liposomes encapsulate therapeutic compounds is largely governed by the biophysical and structural properties of their lipid bilayers, including fluidity, micropolarity, and internal free volume. These membrane characteristics can be modified through the incorporation of specific lipid additives [9]. In this regard, PEGylated lipids have been shown to influence bilayer organization, reduce interfacial hydration, and enhance overall vesicle stability [10,11,12]. Also, curcumin’s effects on various membrane properties were investigated in liposomes [13]. In our previous study [3], we examined in detail the effects of curcumin on membrane properties, such as fluidity, polarity, and phase transition. The main aim of this research was to determine curcumin’s solubility and orientation within the liposomal membrane. The results indicated a transbilayer orientation perpendicular to the membrane surface, with two curcumin molecules likely forming dimers through hydrogen bonding within the bilayer. These findings were compared with the orientation and effects of carotenoids on membranes and discussed in terms of possible antioxidant properties. Interestingly, curcumin (10 mol %) exerted a rigidifying effect on 1,2-dimyristoyl-sn-glycero-3-phosphocholine (DMPC) and 1,2-distearoyl-sn-glycero-3-phosphocholine (DSPC) membranes, while strongly increasing polarity both in the polar headgroup region and at the membrane center. The first effect was like that observed for carotenoids, but the latter was not, raising questions about curcumin’s protective role against lipid peroxidation.
Knowing that curcumin is located within the lipid membrane, in the present work we focus on curcumin-containing liposomes as potential carriers for curcumin delivery in vivo. Encapsulation of curcumin within liposomes has been shown to significantly improve its aqueous solubility and chemical stability, while prolonging systemic circulation and enhancing therapeutic efficacy. Additionally, liposomal delivery protects curcumin from premature degradation, promotes accumulation at target sites, and enables controlled release, making it a highly effective strategy to overcome its pharmacokinetic limitations [14]. Considering that PEGylation of liposomes extends their circulation time and reduces immune clearance and may also alter lipid membrane properties, we chose to focus on curcumin-containing PEGylated liposomes. This study aims to understand how PEGylation influences the biophysical characteristics of liposomal membranes and, consequently, how these changes affect their interaction with curcumin and its loading efficiency. To achieve this, we employed two complementary approaches. First, leveraging the intrinsic fluorescence of curcumin (emission at 460–550 nm depending on the local environment) [15], we applied fluorescence spectroscopy to directly estimate the binding affinity of curcumin to different liposomes (saturated DMPC vs. unsaturated 1-palmitoyloyl-2-oleoyl-sn-glycero-3-phosphocholine POPC, PEGylated vs. non-PEGylated). Second, we investigated the location of curcumin within the bilayer indirectly by analyzing its fluorescence quenching in the presence of paramagnetic agents positioned at defined depths along lipid acyl chains. Finally, using electron paramagnetic resonance (EPR) spectroscopy and lipid spin labels, we evaluated the impact of curcumin on membrane fluidity, polarity, and oxygen transport in PEGylated and non-PEGylated DMPC and POPC liposomes. By integrating these approaches, our work aims to elucidate the mechanistic impact of PEGylation on drug–liposome interactions and contribute to the rational design of more effective liposomal carriers for poorly soluble drugs such as curcumin.
2. Materials and Methods
2.1. Liposome Preparation
DMPC, POPC, 14:0-PEG2000-PE, 18:0-PEG2000-PE, curcumin (1,7-bis(4-hydroxy-3-methoxyphenyl)1,6-heptadiene-3,5-dione) and spin labels: 5- and 16-doxyl stearic acid (5- and 16-SASL) and 1-palmitoyl-2-stearoyl-(n-doxyl)-sn-glycero-3-phosphocholine n-PC, where n = 5 or 16) and TEMPO-PC (1,2-dipalmitoyl-sn-glycero-3-phospho(tempo)choline) were purchased from Merck, Poland. Multilamellar liposomes for EPR measurements were prepared of DMPC or POPC. If required, curcumin was added in the final concentration of 10 mol %, and PEGylated PE in the final concentration of 7 mol%. To match the lengths of host lipids with PEGylated ones, 14:0-PEG2000-PE was added to DMPC, and 18:0-PEG2000-PE to POPC liposomes. The liposomes were prepared by the thin film hydration method [16]. Briefly, chloroform solutions of lipids (containing 2.5 μmol of total lipid), PEGylated PE and/or curcumin (if applicable), and spin labels (1 mol%) were mixed, chloroform was evaporated with a stream of nitrogen, and the lipid film on the bottom of the test tube was thoroughly dried under reduced pressure (about 0.1 mm Hg) for 12 h. A phosphate buffered saline pH 7.4 (usually 0.5 mL) was added to the dried film at the temperature well above the lipid phase transition temperature (TM) and vortexed vigorously. Then, the multilamellar liposome suspension underwent five freeze–thaw cycles, after which it was centrifuged at 14,000 × g for 15 min at 4 °C, and the resulting pellet was used for EPR measurements. For fluorescence measurements, unilamellar liposomes were used. They were prepared by extrusion of multilamellar liposome suspensions (consisting of DMPC or POPC and, if applicable, adequate PEGylated PE) through the polycarbonate filters with the pore size of 100 nm using the mini extruder (Avanti Polar Lipids, USA). For unilamellar liposomes, curcumin was subsequently added from a concentrated ethanol solution.
2.2. Fluorescence Measurements
2.2.1. Curcumin-Liposomes Dissociation Constant
Dissociation constant values Kd was calculated based on titration curves obtained by adding concentrated ethanol solution of curcumin to unilamellar liposome suspensions of increasing lipid concentration (in the range of 0.0019 to 2 mM). The final concentration of curcumin was 2 mM. Fluorescence intensity of curcumin was measured using a spectrofluorimeter (PerkinElmer LS 55 Luminescence Spectrometer) with a quartz cuvette of 1 mL volume. The excitation wavelength was 425 nm, and the maximum emission was observed at 525 nm. The data from three independent experimental series were globally fitted to a single-site binding model using the DynaFit program (version 4.09.019; BioKin, Watertown, MA, USA) [17]. The global analysis was performed separately for POPC and DMPC liposomes, each under two conditions: with and without PEG.
2.2.2. Curcumin’s Fluorescence Quenching by n-SASL
Unilamellar liposomes of POPC, POPC-PEG, DMPC and DMPC-PEG containing 15 mol% of 5- or 16-SASL were prepared, and curcumin was then added from a concentrated ethanol solution to achieve a final concentration of 2 μM. The samples were then allowed to equilibrate on a shaker (PRO-BLOT Rocker, UJKR, model UJ60102) for 0.5 and 1 hour. Liposomes without spin labels served as a control. Curcumin fluorescence intensity was measured as described above - using a PerkinElmer LS 55 Luminescence Spectrometer with a quartz cuvette of 1 mL volume. The excitation wavelength was 425 nm, and the maximum emission was observed at 525 nm.
2.3. EPR Measurements
2.3.1. Continuous Wave (CW)
CW EPR measurements were performed using a Bruker EMX spectrometer equipped with a temperature control unit (EMX ER 4141 VT). Suspensions of multilamellar liposomes containing 1 mol% spin label were placed in gas-permeable TPX capillaries (i.d. 0.7 mm) and positioned inside the EPR dewar insert within the resonant cavity of the spectrometer. Samples were thoroughly deoxygenated with nitrogen gas for approximately 10 minutes, which was also used to maintain temperature control. For measurements of lipid order and correlation times, the EPR spectra were recorded at 20, 25 and at 37 °C for POPC, and at 25 and 37 °C for DMPC. For polarity measurements (maximum splitting 2Az), samples were frozen to 120 K (−153 °C) using liquid nitrogen vapor [3].
2.3.2. Saturation Recovery (SR)
SR EPR measurements were performed on a home-made pulse spectrometer [18] operating at X-band and equipped with a 1-mm loop gap resonator [19]. Liposome samples were prepared and placed in TPX capillaries within the resonant cavity of the spectrometer, following the same procedure as for CW EPR measurements. For measurements of the oxygen transport parameter (OTP), the oxygen concentration in the sample was controlled by equilibration with the same gas used for temperature regulation, either air or nitrogen [20]. The spin-lattice relaxation times (T1) of the spin labels were determined by analyzing the saturation recovery signal of the central EPR line. SR curves were digitized with 2048 points at various rates depending on gas atmosphere and relaxation rate. The number of accumulations varied from 10,000 to 50,000 to obtain a satisfactory signal-to-noise ratio. Acquired SR curves were then fitted by single exponential functions using the Eleana computer program (https://sourceforge.net/projects/eleana/).
3. Results
3.1. Effect of PEGylation on Curcumin Binding to DMPC and POPC Liposomes
Figure 1 presents titration curves obtained for POPC and POPC-PEG (Figure 1a), and DMPC and DMPC-PEG (Figure 1b) liposomes. Curcumin fluorescence intensity was measured for increasing concentrations of lipids. The data were globally fitted to a single-site binding model using the DynaFit program and dissociation constants (Kd) for curcumin with various liposomes were calculated (Table 1).
Kd constants of curcumin are comparable for POPC and DMPC membranes, but for both types of liposomes, PEGylation clearly increases the binding abilities of curcumin to liposomes. PEG seems to increase curcumin-lipid interaction more in DMPC membranes.
3.2. Quenching of Curcumin Fluorescence by Lipid Spin Labels
To assess the location of curcumin within the liposomal membranes, its fluorescence intensity was measured in the presence of 5- and 16-SASL and compared with a control sample without the quenchers. In 5-SASL the paramagnetic nitroxide moiety responsible for fluorescence quenching is in the region close to the membrane surface whereas the free radical moiety of 16-SASL is embedded deep within the membrane center [21,22].
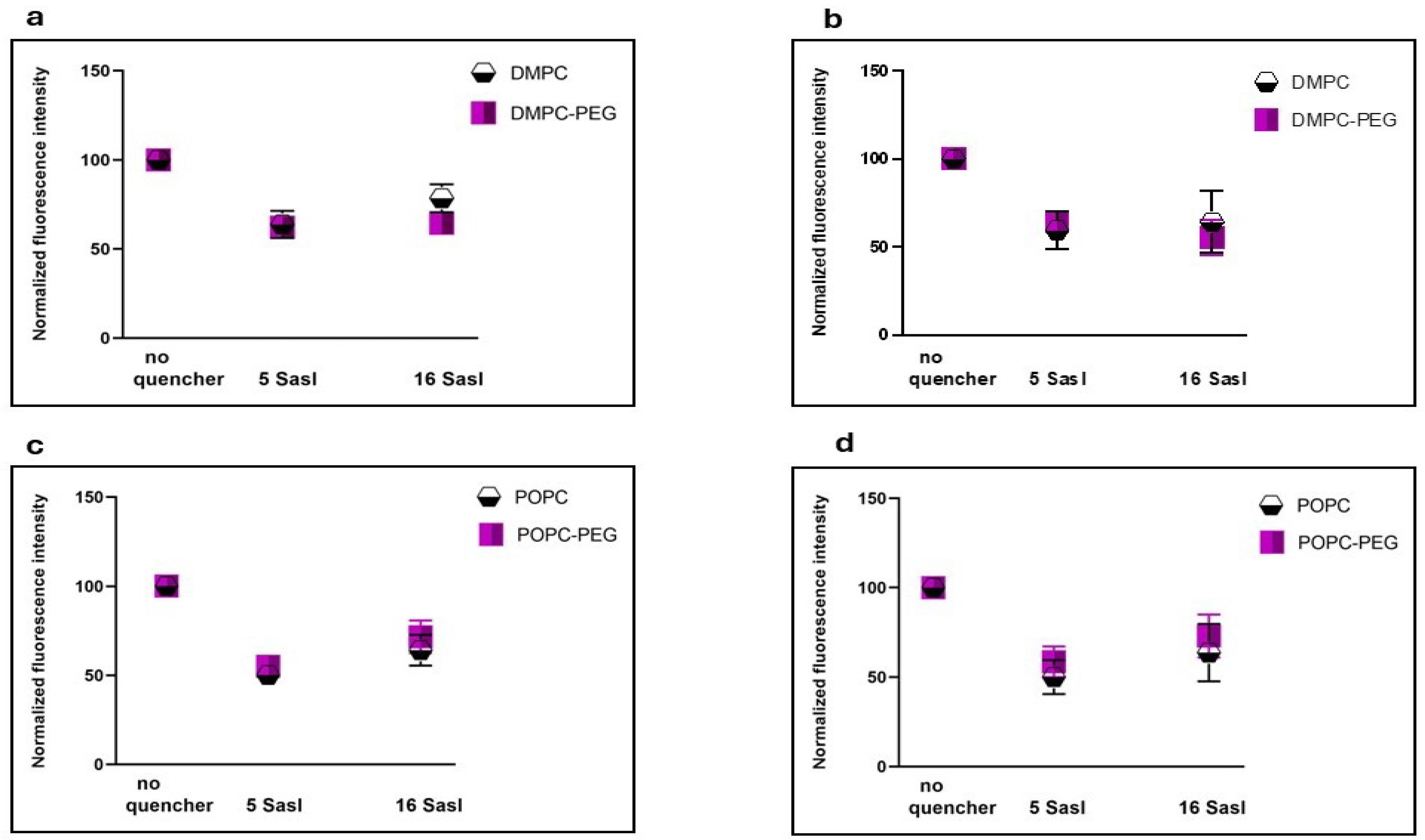
Figure 2.
Quenching effect of lipid spin labels (SASL) having quenching moieties attached at two different positions (5 and 16) on the fluorescence of curcumin incubated with DMPC, DMPC-PEG (a,b), POPC, POPC-PEG (c,d) liposomes for 0,5 h (left panel) and 1 h (right panel).
Figure 2.
Quenching effect of lipid spin labels (SASL) having quenching moieties attached at two different positions (5 and 16) on the fluorescence of curcumin incubated with DMPC, DMPC-PEG (a,b), POPC, POPC-PEG (c,d) liposomes for 0,5 h (left panel) and 1 h (right panel).
Kd constants of curcumin are comparable for POPC and DMPC membranes, but for both types of liposomes, PEGylation clearly increases the binding abilities of curcumin to liposomes. PEG seems to increase curcumin-lipid interaction more in DMPC membranes.
3.3. Effect of Curcumin on Lipid Mobility in PEGylated and Non-PEGylated Liposomes
3.3.1. Polar Headgroup Region
To investigate the membrane properties in the polar headgroup region, the T-PC spin label was used. The parameters obtained from the EPR spectra of T-PC at 25 °C and 37 °C: peak-to-peak width of the low-field line (dh+) and the ratio of the height of central and high-field peaks (h0/h−) give information about the headgroup motional freedom [3]. The increase in motional freedom leads to a decrease in these parameters, as evidenced by the comparison of values at 25°C and 37°C (Figure 3). In DMPC membranes, both spectral parameters have smaller values in the presence of curcumin, especially at 25°C, indicating an increase in lipid headgroups' mobility, and the difference in smaller in the PEGylated membranes. In POPC membranes, both PEGylated and non-PEGylated, the effect is negligible. The results suggest that in the presence of curcumin the polar headgroups of DMPC molecules are more separated from each other and have more motional freedom than in pure DMPC membrane, while PEG reduces this effect.
3.3.2. Alkyl Chain Region
Membrane fluidity may be described in terms of static or dynamic parameters. The static one, order parameter S, reflects the segmental order parameter of the hydrocarbon chain segment to which the nitroxide fragment is attached [23]. It was calculated based on spectral parameters of 5- and 16-PC spin labels according to Marsh [24]. As shown in Figure 4, curcumin increases the order of acyl chains in both positions within the DMPC membrane (Figure 4A). Interestingly, the effect of PEG can also be observed, but only close to the membrane headgroup region (5-PC). In this position, the ordering effect of curcumin is more pronounced in PEGylated DMPC membranes, particularly at 25 °C, whereas the effect of PEG alone is negligible. In POPC membranes, the curcumin ordering effect is observed only close to the membrane headgroup region (5-PC) and there is no difference between PEGylated and non-PEGylated membranes (Figure 4B).
The relatively fast motion of the 16-PC spin label produces more isotropic spectra, enabling the calculation of dynamic parameters, specifically the rotational correlation times τ₂B and τ₂C, using the formulas provided by Berliner [25]. As shown in Figure 5, curcumin clearly increases the rotational correlation times of 16-PC in DMPC and POPC membranes, both PEGylated and non-PEGylated. Additionally, membranes containing curcumin show a greater difference between τ₂B and τ₂C, indicating that in the membrane center, the motion of lipid acyl chains is not only slower but also more anisotropic compared to membranes without curcumin. At lower temperatures (25°C in DMPC and 20°C in POPC) and in DMPC membranes, the effect is more pronounced. PEG alone exerts a slight but noticeable ordering effect in the membrane center and further enhances the ordering effect of curcumin.
3.4. Effect of Curcumin on Polarity Profiles of PEGylated and Non-PEGylated Liposomes
2Az parameter (z-component of the hyperfine interaction tensor) obtained from the EPR spectra of spin labels in frozen suspensions of liposomes is sensitive to the local polarity[26]. Higher values of this parameter indicate higher polarity. Based on this dependance, polarity profiles across lipid bilayer membranes can be obtained [3,16,26]. Figure 6 depicts polarity profiles obtained for DMPC (a), DMPC-PEG (b), POPC (c) and POPC-PEG (d) in the presence and absence of 10 mol% curcumin. Curcumin exerts a strong effect on all investigated membranes. In DMPC, curcumin significantly increases water penetration to the polar headgroup region, as monitored by T-PC, and, to a lesser extent, to the alkyl chain region (positions 5th and 16th ) (Figure 6a). PEG alone does not alter the polarity profile of DMPC significantly, although a slight decrease in polarity can be observed in the center of the PEGylated membrane (Figure 6b). In PEGylated DMPC membranes, curcumin increases polarity more than in non-PEGylated membranes, but only in the membrane center, where the difference in 2Az values is 0.62 G compared to 0.39 G in the absence of PEG. However, in PEGylated DMPC membranes, curcumin does not induce an increase in polarity near the 5th carbon. In POPC membrane, the effect of curcumin is also significant, but different than in DMPC (Figure 6 c,d). First, the curcumin-induced increase in polarity within the headgroup region is weaker in both PEGylated and non-PEGylated POPC membranes compared to DMPC. Second, a strong increase in polarity is observed in the membrane center (Figure 6c). This effect is especially noticeable because the POPC membrane is inherently more hydrophobic at the center than DMPC. PEG alone has no significant effect on the polarity of the headgroup region, but, unlike in DMPC, it markedly reduces the hydrophobicity barrier in the center of POPC membranes. In this region of the membrane, PEG also diminishes the effect of curcumin. In POPC membranes, the difference in 2Az values with and without curcumin is approximately 2 G, whereas in PEGylated POPC it is only 0.3 G.
3.5. Effect of Curcumin on Oxygen Transport Parameter of PEGylated and Non-PEGylated Liposomes
Saturation recovery (SR) EPR technique allows to obtain spin-lattice (T1) relaxation times of paramagnetic probes, such as nitroxide spin labels [27]. T1 of spin labels is significantly shorter in the presence of oxygen due to strong Heisenberg exchange between spin label and oxygen. Therefore, the method is used to measure the oxygen diffusion-concentration product, Do[O2], within the membrane, which is based on the bimolecular collision rate of spin label with oxygen [28]. The effect of oxygen on the spin-lattice relaxation of spin labels is generally much greater than the motional effects. Thus, oxygen transport parameter (OTP), W(x), was introduced as a convenient quantitative measure of the rate of the collision between the spin probe and molecular oxygen by Kusumi et al. [29] as:
where the T1s are the spin-lattice relaxation times of the nitroxides in samples equilibrated with atmospheric air and nitrogen, respectively. The collision rate is also proportional to the local oxygen concentration, C(x), and the local oxygen diffusion coefficient, D(x) at a ‘‘depth’’ x in the membrane that is in equilibrium with atmospheric air.
W(x) = T1-1 (air, x) – T1-1 (N2, x),
Figure 7 shows the profiles of OTP obtained for DMPC (a), DMPC-PEG (b), POPC (c) and POPC-PEG (d) in the presence and absence of 10 mol% curcumin. Membranes lacking curcumin exhibit distinct changes in the oxygen transport parameter (OTP) along the lipid alkyl chains. All investigated membranes, both PEGylated and non-PEGylated, exhibit significant oxygen permeability in their central regions, whereas oxygen penetration is reduced near the polar headgroup region. The presence of curcumin significantly alters these profiles, leading to a pronounced decrease in the oxygen transport parameter at the membrane center. The effect is stronger in DMPC. In the polar headgroup region, where OTP is lower already in pure membranes, the curcumin presence only slightly reduces its values. Again, the effect is stronger in DMPC. PEG does not affect oxygen penetration into the membranes; OPT profiles look very similar in PEGylated and non-PEGylated DMPC and POPC membranes.
4. Discussion
Among different pharmaceutical nanocarriers, PEGylated liposomes are quite commonly used. Incorporating a polymer to form a corona around liposomes significantly prolongs their circulation time in the bloodstream by reducing their clearance by the mononuclear phagocyte system (MPS) [12]. However, the exact mechanism of this prevention is not completely understood. Moreover, growing evidence indicates that the polymeric corona is not entirely inert, and PEGylated nanoparticles can engage with the biological environment through Van der Waals forces, hydrophobic interactions, and hydrogen-bond formation [30]. PEG presence may also impact liposome interactions with the cell membrane after delivery and influence their subsequent biological fate [31]. For example, the fusion of liposomes with cell membranes may be influenced by PEG [32]. Studies on PEG effects on liposome membranes may help to understand more about these interactions. In this case, liposomes can be studied not only as nanocarriers but also can serve as models of cell membranes. Another feature of PEG is its ability to enhance the partitioning of hydrophobic compounds into lipid membranes. The presence of a PEG corona around liposomes has been shown to increase the binding constant (Kb) of these compounds to a lipid bilayer [10]. Therefore, PEGylation may increase the loading efficiency of liposomes for hydrophobic molecules.
In the present work, we focused on curcumin-loaded liposomes and examined the potential effects of PEG on curcumin–liposome interactions and loading efficiency.
Fluorescence Experiments
First, taking advantage of curcumin’s intrinsic fluorescence, we examined its interaction with both PEGylated and non-PEGylated DMPC and POPC liposomes. The calculated dissociation constant (Kd) values were markedly lower for PEGylated liposomes, particularly for PEGylated DMPC, compared with their non-PEGylated counterparts. The Kd values decreased from 263 μM for DMPC to 88 μM for PEGylated DMPC and from 270 μM for POPC to 156 μM for PEGylated POPC. This indicates that PEG enhances curcumin binding, and potentially its partitioning, into lipid membranes, with the length of the PEG lipid anchor also appearing to play a role. A similar effect of increased partitioning in PEGylated membranes was observed previously for hydrophobic porphyrin p-THPP [10,12]. On the other hand, we have shown previously that curcumin adopts a transbilayer orientation within the lipid bilayer and affects membrane properties in a concentration-dependent manner [3]. Therefore, we expected that PEG-induced increase of curcumin partitioning to lipid bilayer will enhance the effect of curcumin on bilayer properties. To explore this further, we conducted a series of experiments using fluorescence quenching and EPR spectroscopy to investigate curcumin’s location within PEGylated and non-PEGylated membranes, as well as its effects on lipid order, local polarity, and oxygen transport in these membranes.
The vertical location of curcumin in lipid membranes was examined using nitroxide-labeled molecules (5-SASL and 16-SASL), which are known as efficient quenchers of fluorescence [12,22,33]. In PEGylated DMPC, after 30 minutes of incubation with curcumin, the fluorescence intensity was reduced to a similar extent by both 5- and 16-SASL (62% and 64%, respectively) compared with liposomes without quenchers. In contrast, for non-PEGylated DMPC, 16-SASL was less effective than 5-SASL, reducing the signal to 78% versus 63% for 5-SASL. This indicates that curcumin penetrates both types of DMPC membranes and is oriented perpendicular to the bilayer surface; however, PEG enhances curcumin penetration toward the membrane center. In POPC membranes, both PEGylated and non-PEGylated, quenching was significant at both positions, but slightly more pronounced near the 5th carbon than at the membrane center (fluorescence reduced to 55% vs. 71% in PEGylated POPC, and 46% vs. 58% in non-PEGylated). This indicates that, also in unsaturated membranes, curcumin adopts a transbilayer orientation, although it predominantly resides near the membrane surface. Interestingly, POPC curcumin’s partition was not increased by PEG. The enhanced curcumin penetration into DMPC membranes in the presence of PEG is consistent with the calculated Kd values, which indicate that PEGylated DMPC binds curcumin to the greatest extent. Generally, fluorescence experiments, including curcumin binding and quenching, proved that curcumin not only binds to liposome membranes, but also penetrates and fully spans the lipid bilayer, although accumulates preferably in the region close to the membrane surface. In DMPC, however, PEGylation induces more uniform distribution of curcumin within the membrane.
EPR Experiments
To study the effects of curcumin on PEGylated and non-PEGylated liposome membranes EPR spectroscopy with lipid spin labels was employed. The conventional, continuous wave (CW) EPR technique was used to investigate the mobility of polar headgroups, order parameter of lipid alkyl chains, rotational correlation times and polarity, whereas the pulse EPR technique, saturation recovery (SR) was employed to acquire oxygen transport parameter.
The polar headgroup region of DMPC and POPC membranes was investigated using T-PC spin label. In DMPC, the changes in the spectral parameters of T-PC, especially the linewidth of the low-field line (dh+), indicate that there is a slight increase in lipid headgroups mobility in the presence of curcumin, which is similar in both PEGylated and non-PEGylated membranes. In POPC the effects are negligible. The increase of lipid headgroups mobility is most probably caused by their separation by terminal rings of curcumin molecules containing polar hydroxyl and methoxyl groups [3]. Similar effects were observed for other membrane modifiers containing ring structures, such as cholesterol or xanthophylls [34,35,36]. Also, these effects were more pronounced in saturated membranes than in unsaturated ones, which may explain the minimal impact observed in POPC. The thickness of the membrane compared with the length of the curcumin molecule may also matter. In a thicker membrane, such as POPC, curcumin may sink deeper into the lipid part, which results in its smaller effect on polar headgroups. Similarly, unlike its effect on the motional freedom of DMPC polar headgroups, curcumin was previously found to have no impact on the thicker distearoylphosphatidylcholine (DSPC) membrane [3].
The alkyl chain order and lipid mobility in DMPC and POPC membranes were investigated using 5- and 16-PC spin labels which probe regions near the membrane surface and at the membrane center, respectively. The values of order parameter S significantly differ between the 5th and 16th positions in all investigated membranes, which confirms the existence of a fluidity gradient across the lipid bilayer [3,23,34,37]. As already observed previously [3] at 25°C curcumin increases the values of S parameter in both regions of DMPC membrane (from 0.63 to 0.66 at the 5th position, and from 0.13 to 0.18 at the 16th position). Also, at 37°C the effect is observed, although the difference is smaller. The ordering effect of curcumin in DMPC seems to be slightly enhanced by PEG, particularly at 25°C, whereas the effect of PEG alone is negligible. In POPC, curcumin’s ordering effect is observed only at the 5th position, and PEG does not further enhance this effect. This suggests that curcumin partitions to the DMPC membrane more efficiently when PEG is present, which is consistent with fluorescence data. A similar conclusion can be drawn from the analysis of the 16-PC rotational correlation time data. Both, τ2B and τ2C values are higher in the presence of curcumin in both DMPC and POPC membranes; however, the effects are stronger in DMPC. Additionally, in the membranes containing curcumin there is a bigger difference between τ2B and τ2C, which indicates that the motion of lipid alkyl chains is more anisotropic in the presence of curcumin than in the membranes without it. The ordering effect of curcumin, as reflected in correlation times, has already been demonstrated in DMPC [3]. Here, we confirmed this effect and demonstrated it also in unsaturated membranes (POPC) as well as in PEGylated membranes. At lower temperatures (25°C in DMPC and 20°C in POPC), the effect of curcumin on correlation times is more pronounced than at 37°C. As previously shown, the impact of various compounds on lipid motion decreases with increasing temperature [3,23,34,37]. PEG alone exerts a slight but noticeable ordering effect in the membrane center, making it difficult to determine whether it further enhances curcumin’s effect. Nonetheless, the effects remain stronger in DMPC than in POPC membranes, consistent with the observations above. Regarding the effect of PEG alone, correlation times were the only parameters in our study that revealed a direct impact of PEG on lipid mobility. Lemaalem et al investigated how PEGylation affects the mechanical properties of the lipid bilayer. Their MD simulations showed that PEGylated membranes possess increased bending rigidity and reduced lateral diffusion and explained that the mobility of lipids in PEGylated membranes was reduced due to PEG-induced crowding and increased interfacial viscosity [38]. Giakoumatos et al. studied lipid of colloid supported lipid bilayers ordering via differential scanning calorimetry and fluidity using fluorescence recovery after photobleaching (FRAP) technique and found that up to 5 mol% of PEGylated lipids could be incorporated into studied membranes without any pronounced effects. However, the fluorescence recovery of the more fluid DOPC membrane was markedly decelerated upon incorporating 10 mol% of PEGylated lipids, whilst insertion of the anchoring lipids (DOPE and DSPE without PEG2000) had no detectable impact. Therefore, they concluded that the quantity of incorporated PEG stabilizers, rather than the chemical nature of the lipid anchor, should be carefully optimized to ensure adequate colloidal stability without compromising membrane dynamics [39]. However, the results on flat bilayers immobilized on silica support cannot be fully compared to liposomes in suspension.
It is well established that compounds that alter membrane fluidity can also influence its polarity. In our previous work we studied the effect of curcumin on polarity of DMPC and DSPC membranes, and we showed that curcumin significantly changes the polarity profiles across these membranes, making them more water permeable [3]. Considering PEG’s hydrophilic nature, in the present work we investigated how PEG affects the polarity profiles of saturated (DMPC) and unsaturated (POPC) membranes, and how it modulates the curcumin-induced increase in polarity. We used 2Az as a measure of local membrane polarity, which is directly obtained from the EPR spectra of spin labels in frozen liposome suspensions [26]. As expected, curcumin increases polarity within both membranes, however in DMPC the effect is more pronounced in the polar headgroup region, whereas in POPC, in the membrane center. Interestingly, PEG alters curcumin’s effect on DMPC polarity – near the 5th position, curcumin’s impact is reduced compared to its effect in the absence of PEG. In the membrane center PEG alone affects local polarity, making the membrane less polar (2Az value in PEGylated DMPC is 67.5 G compared to 68.25 G in non-PEGylated one). Similar effect of PEGylated lipids on membrane polarity was shown by Aloia and Bartucci in dipalmitoylphosphatidylcholine (DPPC) membranes [40]. They reported sigmoidal water penetration and polarity profiles in sterically stabilized liposomes (SSL) formed with submicellar amounts of PEG₋2000–DPPE incorporated into DPPC, and suggested that, compared to DPPC bilayers, SSL exhibit increased hydrophobicity at both the polar/apolar interface and the chain termini [40]. In POPC, however, we observed the opposite effect of PEG on membrane polarity: PEG clearly reduces the hydrophobicity barrier and increases polarity throughout the membrane. At both positions, 5th and 16th, the 2Az values are higher in PEGylated POPC than in non-PEGylated one (69.31 G vs 68.9 G for 5-PC and 68.35 G vs 67.5 G for 16-PC, respectively). The effect on curcumin on polarity profile in POPC is significant in the membrane center. The pronounced effects of curcumin and PEG can be attributed to the higher hydrophobicity of POPC, as an unsaturated membrane, compared to DMPC [26]. Also, the polarity profiles in our study differ between DMPC and POPC, with POPC exhibiting a higher hydrophobicity barrier (2Az values are lower at all positions across the POPC membrane compared to the DMPC membrane). As a result, curcumin may have a more pronounced effect on the 2Az parameter, making the difference more noticeable. Also, because PEG clearly increases the polarity across the membrane, curcumin’s effect is less noticeable in the presence of PEG. Apart from its modest impact on correlation times, polarity seems to be another membrane feature changed by PEG alone. However, because the effect varies with the host lipid, drawing a definitive conclusion is difficult. In the polar headgroup region the effect of curcumin on water accessibility is very strong in DMPC, which proves that curcumin separates effectively the headgroups from each other. This is also consistent with the curcumin’s effect on T-PC parameters measured at room temperature, which was more pronounced in DMPC. Considering the structure of the curcumin molecule, the polarity profiles support its transbilayer orientation in all the membranes studied. Assuming this orientation, the effect of increased water penetration is observed in the places where the terminal rings containing polar hydroxyl and methoxyl groups of curcumin are located. A comparison of the curcumin’s effects on the polarity profiles of DMPC and POPC membranes allows us to conclude that the membrane thickness and unsaturation does not affect curcumin’s orientation within the membrane. Notably, the pronounced effect of curcumin on polarity at the center of the POPC membrane suggests that hydrogen bonding between two curcumin molecules spanning opposite leaflets is strong enough to stabilize the dimer, even when the membrane thickness and curcumin dimer length do not perfectly match. This is consistent with our previous work comparing the polarity profiles of DMPC and DSPC [3].
The final membrane characteristic obtained from EPR measurements was the oxygen transport parameter (OTP). Previous studies have shown that the oxygen collision rate – a product of the local concentration and diffusion coefficient of molecular oxygen within the membrane – is a highly sensitive measure of the presence and dynamics of free volume, which can be extremely small, just enough to accommodate a single oxygen molecule within the membrane [41]. The small size and appropriate level of hydrophobicity of molecular oxygen allows it to enter the small vacant pockets that are transiently formed in the lipid bilayer membrane [20]. Therefore, collision rates between molecular oxygen and nitroxide spin labels at specific locations in the membrane are sensitive to the dynamics of gauche-trans isomerization of lipid hydrocarbon chains and to the structural nonconformability of neighboring lipids [41]. Our data show that curcumin significantly reduces the oxygen transport parameter at all locations in all membranes studied, with the effect being more pronounced in DMPC. This indicates that curcumin reduces the dynamics of gauche-trans isomerization of lipid hydrocarbon chains which is consistent with the data on order parameter and correlation times. Interestingly, curcumin also reduces OTP in the polar headgroup region, even though our other data indicate increased mobility and greater headgroup separation in this region. A similar effect has been observed for cholesterol, which markedly reduces OTP in the polar headgroup region of lipid bilayers [41] while simultaneously increasing water penetration into this region [26]. It was found that incorporation of cholesterol into lipid bilayers greatly reduced the oxygen collision rate at the place where cholesterol’s rigid steroid ring structure was located. Assuming a transbilayer location of curcumin, as suggested by Duda et al. [3] and confirmed in this work through quenching experiments and polarity profiles, it is likely that curcumin’s rigid rings positioned in the headgroup region as well as at the membrane center create a rigidity barrier that limits the penetration of small hydrophobic molecules such as oxygen. The effect of curcumin on OTP in POPC membranes is smaller, consistent with the order parameter and correlation times data, which indicates a relatively negligible impact of curcumin in these membranes. PEG alone has no significant effect on OTP, nor does it noticeably alter curcumin’s impact in either membrane.
It is worth comparing the polarity and OTP profiles, as curcumin clearly alters both – but in opposite directions. While curcumin increases water penetration into the membrane, it simultaneously decreases oxygen penetration. Thus, curcumin appears to reduce the hydrophobicity barrier for polar molecules while increasing the rigidity barrier for apolar molecules. Both effects are more pronounced in a saturated DMPC membrane. PEG alone influences polarity profiles, as discussed above, but has no effect on oxygen penetration. This suggests that OTP reflects the rigidity barrier, which is not affected by PEG, as further supported by the absence of PEG’s effect on the order parameter.
The rigidity barrier formed by curcumin, which also reduces oxygen diffusion within the membrane, may be important for liposome stability. Membranes containing curcumin are not only ordered but also have lower oxygen content, limiting the potential for reactive oxygen species formation and thereby helping to prevent lipid peroxidation.
5. Conclusions
Curcumin effectively interacts with liposome membranes, both saturated (DMPC) and unsaturated (POPC). PEG enhances the curcumin–lipid interaction, particularly in DMPC membranes, although it does not significantly alter the overall biophysical properties of the membrane. Curcumin penetrates deeply into the membrane, adopting a perpendicular orientation relative to the membrane surface, and in DMPC – especially when PEGylated—it reaches greater depths. The presence of curcumin increases lipid order and substantially enhances water penetration across the membrane, while simultaneously reducing oxygen diffusion at all membrane locations. Among the four types of liposomes investigated, PEGylated DMPC appears to be the most suitable for curcumin delivery.
Author Contributions
Conceptualization, A.W-B. and N.F.; methodology, A.W-B., A.G. and N.F.; software, A.W-B., A.G. and N.F.; validation, A.W-B., A.G. and N.F.; formal analysis, A.W-B.; A.G.; investigation, A.W-B., and N.F.; resources, A.W-B., and N.F; data curation, A.W-B., and N.F,; writing—original draft preparation, A.W-B., and N.F.; writing—review and editing, A.W-B., and N.F.; visualization, A.W-B.; supervision, A.W-B; project administration, A.W-B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.:
Data Availability Statement
Data will be available from the corresponding author on request or publicly at: https://ruj.uj.edu.pl/home (Jagiellonian University Repository).
Acknowledgments
The authors gratefully acknowledge the technical support of Jerzy Kozioł, in connection with the home-built EPR pulse spectrometer.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wang, G.; Zhou, X.; Pang, X.; Ma, K.; Li, L.; Song, Y.; Hou, D.; Wang, X. Pharmacological Effects, Molecular Mechanisms and Strategies to Improve Bioavailability of Curcumin in the Treatment of Neurodegenerative Diseases. Front Pharmacol 2025, 16. [Google Scholar] [CrossRef]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of Curcumin: Problems and Promises. Mol Pharm 2007, 4, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Duda, M.; Cygan, K.; Wisniewska-Becker, A. Effects of Curcumin on Lipid Membranes: An EPR Spin-Label Study. Cell Biochem Biophys 2020, 78, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Hu, S.; Sun, M.; Shi, J.; Zhang, H.; Yu, H.; Yang, Z. Recent Advances and Clinical Translation of Liposomal Delivery Systems in Cancer Therapy. European Journal of Pharmaceutical Sciences 2024, 193. [Google Scholar] [CrossRef] [PubMed]
- Pisani, S.; Di Martino, D.; Cerri, S.; Genta, I.; Dorati, R.; Bertino, G.; Benazzo, M.; Conti, B. Investigation and Comparison of Active and Passive Encapsulation Methods for Loading Proteins into Liposomes. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Gerasimov, O.V.; Boomer, J.A.; Qualls, M.M.; Thompson, D.H. Cytosolic Drug Delivery Using PH- and Light-Sensitive Liposomes. Adv Drug Deliv Rev 1999, 38, 317–338. [Google Scholar] [CrossRef]
- Allen, T.M.; Cullis, P.R. Liposomal Drug Delivery Systems: From Concept to Clinical Applications. Adv Drug Deliv Rev 2013, 65, 36–48. [Google Scholar] [CrossRef]
- Mohamed, M.; Alaaeldin, E.; Hussein, A.; Sarhan, H.A. Liposomes and PEGylated Liposomes as Drug Delivery Systems, 2020.
- Liu, P.; Chen, G.; Zhang, J. A Review of Liposomes as a Drug Delivery System: Current Status of Approved Products, Regulatory Environments, and Future Perspectives. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Kepczyński, M.; Nawalany, K.; Kumorek, M.; Kobierska, A.; Jachimska, B.; Nowakowska, M. Which Physical and Structural Factors of Liposome Carriers Control Their Drug-Loading Efficiency? Chem Phys Lipids 2008, 155, 7–15. [Google Scholar] [CrossRef]
- Nakamura, K.; Yamashita, K.; Itoh, Y.; Yoshino, K.; Nozawa, S.; Kasukawa, H. Comparative Studies of Polyethylene Glycol-Modified Liposomes Prepared Using Different PEG-Modification Methods. Biochim Biophys Acta Biomembr 2012, 1818, 2801–2807. [Google Scholar] [CrossRef]
- Dzieciuch, M.; Rissanen, S.; Szydłowska, N.; Bunker, A.; Kumorek, M.; Jamróz, D.; Vattulainen, I.; Nowakowska, M.; Róg, T.; Kepczynski, M. Pegylated Liposomes as Carriers of Hydrophobic Porphyrins. Journal of Physical Chemistry B 2015, 119, 6646–6657. [Google Scholar] [CrossRef] [PubMed]
- Gudyka, J.; Ceja-Vega, J.; Ivanchenko, K.; Morocho, Z.; Panella, M.; Gamez Hernandez, A.; Clarke, C.; Perez, E.; Silverberg, S.; Lee, S. Concentration-Dependent Effects of Curcumin on Membrane Permeability and Structure. ACS Pharmacol Transl Sci 2024, 7, 1546–1556. [Google Scholar] [CrossRef] [PubMed]
- Kokkinis, S.; Paudel, K.R.; De Rubis, G.; Yeung, S.; Singh, M.; Singh, S.K.; Gupta, G.; Panth, N.; Oliver, B.; Dua, K. Liposomal Encapsulated Curcumin Attenuates Lung Cancer Proliferation, Migration, and Induces Apoptosis. Heliyon 2024, 10. [Google Scholar] [CrossRef] [PubMed]
- Gangemi, C.M.A.; Mirabile, S.; Monforte, M.; Barattucci, A.; Bonaccorsi, P.M. Bioimaging and Sensing Properties of Curcumin and Derivatives. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef]
- Wisniewska, A.; Subczynski, W.K. Effects of Polar Carotenoids on the Shape of the Hydrophobic Barrier of Phospholipid Bilayers; 1998; Vol. 1368.
- Kuzmič, P. Program DYNAFIT for the Analysis of Enzyme Kinetic Data: Application to HIV Proteinase. Anal Biochem 1996, 237, 260–273. [Google Scholar] [CrossRef]
- Ilnicki, J., O.T., K.J., G.W., G.R.J., & F.W. SATURATION RECOVERY EPR SPECTROMETER. Molecular physics reports 1994.
- Froncisz W and James S Hyde The Loop-Gap Resonator: A New Microwave Lumped Circuit ESR Sample Structure. Journal of Magnetic Resonance 1982, 47, 515–521.
- Subczynski, W.K.; Wisniewska, A.; Hyde, J.S.; Kusumi, A. Three-Dimensional Dynamic Structure of the Liquid-Ordered Domain in Lipid Membranes as Examined by Pulse-EPR Oxygen Probing. Biophys J 2007, 92, 1573–1584. [Google Scholar] [CrossRef]
- Wisniewska, A.; Widomska, J.; Subczynski, W.K. Carotenoid-Membrane Interactions in Liposomes: Effect of Dipolar, Mo-Nopolar, and Nonpolar Carotenoids *; 2006. Interactions in Liposomes.
- Kepczynski, M.; Kumorek, M.; Stepniewski, M.; Rog, T.; Kozik, B.; Jamroz, D.; Bednar, J.; Nowakowska, M. Behavior of 2,6-Bis(Decyloxy)Naphthalene inside Lipid Bilayer. J Phys Chem B 2010, 114, 15483–15494. [Google Scholar] [CrossRef]
- Stepien, P.; Polit, A.; Wisniewska-Becker, A. Comparative EPR Studies on Lipid Bilayer Properties in Nanodiscs and Liposomes. Biochim Biophys Acta Biomembr 2015, 1848, 60–66. [Google Scholar] [CrossRef]
- Marsh, D. Electron Spin Resonance: Spin Labels. 1981. [Google Scholar]
- Berliner, L.J. Spin Labeling in Enzymology: Spin-Labeled Enzymes and Proteins. Methods Enzymol 1978, 49, 418–480. [Google Scholar]
- Subczynski, W.K., W.A., Y.J.J., H.J.S., & K.A. Hydrophobic Barriers of Lipid Bilayer Membranes Formed by Reduction of Water Penetration by Alkyl Chain Unsaturation and Cholesterol?; 1994; Vol. 33.
- Yin, J.J.; Feix, J.B.; Hyde, J.S. Mapping of Collision Frequencies for Stearic Acid Spin Labels by Saturation-Recovery Electron Paramagnetic Resonance. Biophys J 1990, 58, 713–720. [Google Scholar] [CrossRef]
- Subczynski, W.K.; Hyde, J.S. The Diffusion-Concentration Product of Oxygen in Lipid Bilayers Using the Spin-Label Method. Biochimica et Biophysica Acta (BBA) - Biomembranes 1981, 643, 283–291. [Google Scholar] [CrossRef]
- Kusumi, A.; Subczynski, W.K.; Hydet, J.S. Oxygen Transport Parameter in Membranes as Deduced by Saturation Recovery Measurements of Spin-Lattice Relaxation Times of Spin Labels (Pulse ESR/Diffusion and Concentration of Oxygen/Phase Transitions); 1982; Vol. 79.
- Moghimi, S.M.; Szebeni, J. Stealth Liposomes and Long Circulating Nanoparticles: Critical Issues in Pharmacokinetics, Opsonization and Protein-Binding Properties. Prog Lipid Res 2003, 42, 463–478. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Ye, H.; Kröger, M.; Li, Y. Aggregation of Polyethylene Glycol Polymers Suppresses Receptor-Mediated Endocytosis of PEGylated Liposomes. Nanoscale 2018, 10, 4545–4560. [Google Scholar] [CrossRef] [PubMed]
- Tomatsu, I.; Marsden, H.R.; Rabe, M.; Versluis, F.; Zheng, T.; Zope, H.; Kros, A. Influence of Pegylation on Peptide-Mediated Liposome Fusion. J Mater Chem 2011, 21, 18927–18933. [Google Scholar] [CrossRef]
- Merkle, H.; Subczynski, W.K.; Kusumi, A. Dynamic Fluorescence Quenching Studies on Lipid Mobilities in Phosphatidylcholine-Cholesterol Membranes. Biochimica et Biophysica Acta (BBA) - Biomembranes 1987, 897, 238–248. [Google Scholar] [CrossRef]
- Mainali, L.; Hyde, J.S.; Subczynski, W.K. Using Spin-Label W-Band EPR to Study Membrane Fluidity Profiles in Samples of Small Volume. Journal of Magnetic Resonance 2013, 226, 35–44. [Google Scholar] [CrossRef]
- Subczynski, W.K.; Markowska, E.; Gruszecki, W.I.; Sielewiesiuk, J. Effects of Polar Carotenoids on Dimyristoylphosphatidylcholine Membranes: A Spin-Label Study. Biochimica et Biophysica Acta (BBA) - Biomembranes 1992, 1105, 97–102. [Google Scholar] [CrossRef]
- Subczynski, W.K.; Hyde, J.S.; Kusumi, A. Oxygen Permeability of Phosphatidylcholine-Cholesterol Membranes (Pulse ESR/Permeability Coefficient/Oxygen Transport); 1989; Vol. 86.
- Subczynski, W.K.; Markowska, E.; Gruszecki, W.I.; Sielewiesiuk, J. Effects of Polar Carotenoids on Dimyristoylphosphatidylcholine Membranes: A Spin-Label Study. Biochimica et Biophysica Acta (BBA) - Biomembranes 1992, 1105, 97–108. [Google Scholar] [CrossRef]
- Lemaalem, M.; Hadrioui, N.; Derouiche, A.; Ridouane, H. Structure and Dynamics of Liposomes Designed for Drug Delivery: Coarse-Grained Molecular Dynamics Simulations to Reveal the Role of Lipopolymer Incorporation. RSC Adv 2020, 10, 3745–3755. [Google Scholar] [CrossRef]
- Giakoumatos, E.C.; Gascoigne, L.; Gumí-Audenis, B.; García, Á.G.; Tuinier, R.; Voets, I.K. Impact of Poly(Ethylene Glycol) Functionalized Lipids on Ordering and Fluidity of Colloid Supported Lipid Bilayers. Soft Matter 2022, 18, 7569–7578. [Google Scholar] [CrossRef]
- Aloi, E.; Bartucci, R. Cryogenically Frozen PEGylated Liposomes and Micelles: Water Penetration and Polarity Profiles. Biophys Chem 2020, 266. [Google Scholar] [CrossRef]
- Subczynski Witold, J.S.H. and A.K. Effect of Alkyl Chain Unsaturation and Cholesterol Intercalation on Oxygen Transport in Membranes: A Pulse ESR Spin Labeling Study?; 1991; Vol. 30.
Figure 1.
Fluorescence measurements of curcumin binding to liposomes composed of: (a) POPC (black circles) or POPC-PEG (red circles) and (b) DMPC (black circles) or DMPC-PEG (red circles). The phosphatidylcholine concentration is shown on the x-axis, and the fraction of bound curcumin on the y-axis. The solid lines represent the best fits obtained using the DynaFit software.
Figure 1.
Fluorescence measurements of curcumin binding to liposomes composed of: (a) POPC (black circles) or POPC-PEG (red circles) and (b) DMPC (black circles) or DMPC-PEG (red circles). The phosphatidylcholine concentration is shown on the x-axis, and the fraction of bound curcumin on the y-axis. The solid lines represent the best fits obtained using the DynaFit software.
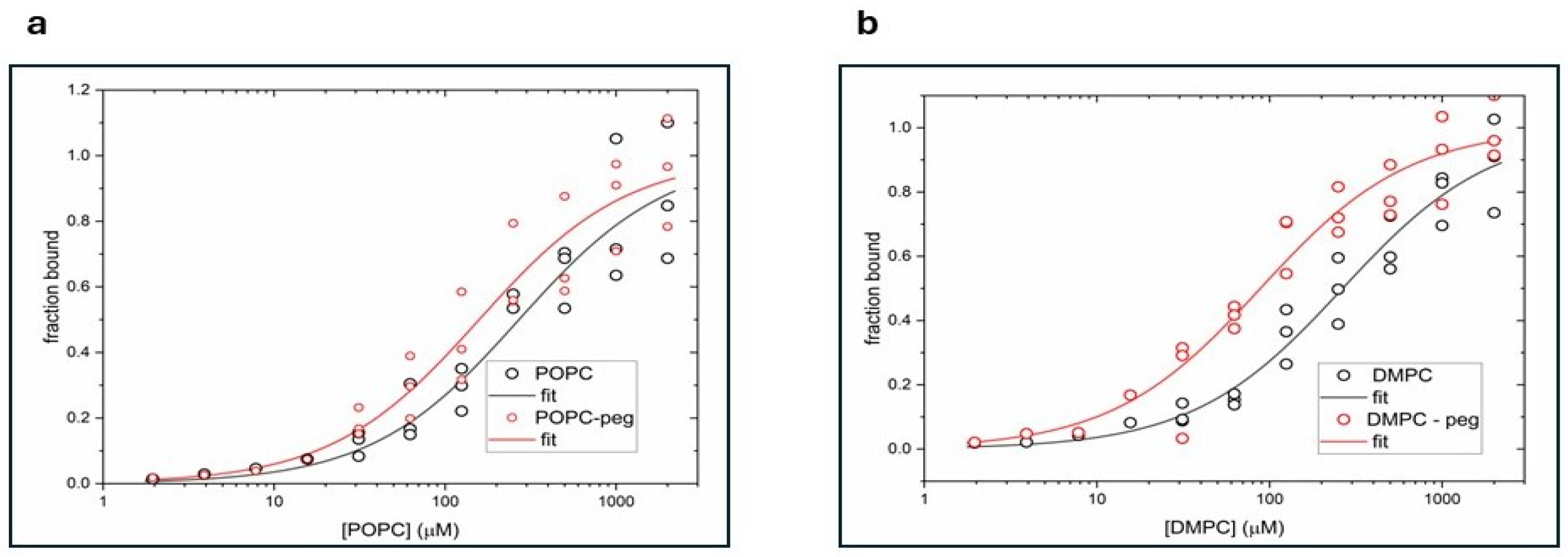
Figure 3.
EPR spectral parameters of T-PC spin probe measured for DMPC, DMPC-PEG, POPC and POPC-PEG membranes in the presence and absence of 10 mol% curcumin at 25°C and 37 °C. h0/h− is a ratio of the height of central and high-field peaks and dh+ is a peak-to-peak width of the low-field line.
Figure 3.
EPR spectral parameters of T-PC spin probe measured for DMPC, DMPC-PEG, POPC and POPC-PEG membranes in the presence and absence of 10 mol% curcumin at 25°C and 37 °C. h0/h− is a ratio of the height of central and high-field peaks and dh+ is a peak-to-peak width of the low-field line.
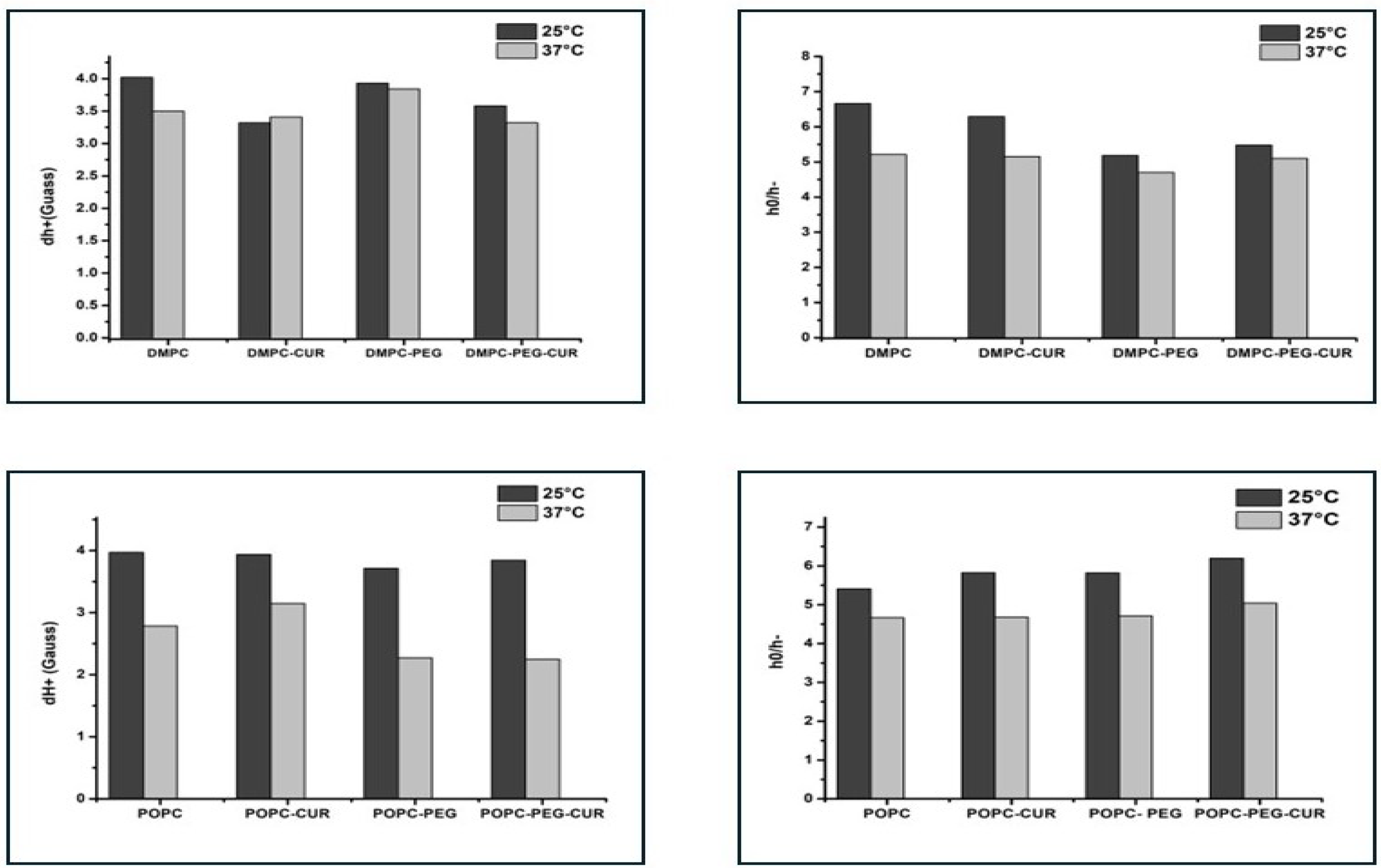
Figure 4.
Order parameter S of 5- and 16-PC in DMPC (A) and POPC (B) membranes, at 25 and 37°C, in the presence and absence of 10 mol% curcumin and/or 7 mol% PEG.
Figure 4.
Order parameter S of 5- and 16-PC in DMPC (A) and POPC (B) membranes, at 25 and 37°C, in the presence and absence of 10 mol% curcumin and/or 7 mol% PEG.
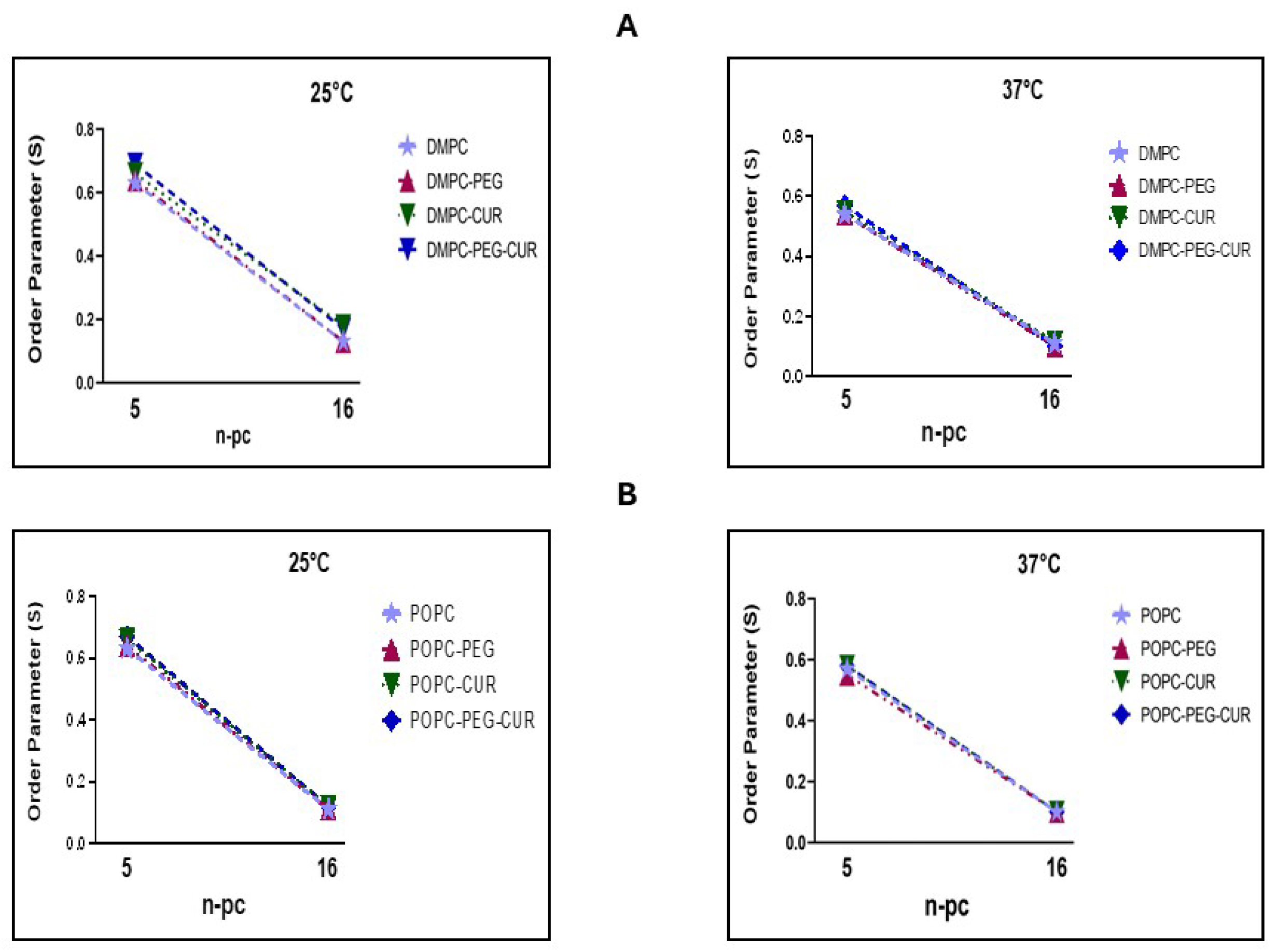
Figure 5.
Rotational correlation times τ2B and τ2C of 16-PC in (a): DMPC (at 25 and 37 °C) and (b): POPC (at 20, 25 and 37°C) membranes in the presence and absence of 10 mol% curcumin and/or 7 mol% PEG.
Figure 5.
Rotational correlation times τ2B and τ2C of 16-PC in (a): DMPC (at 25 and 37 °C) and (b): POPC (at 20, 25 and 37°C) membranes in the presence and absence of 10 mol% curcumin and/or 7 mol% PEG.
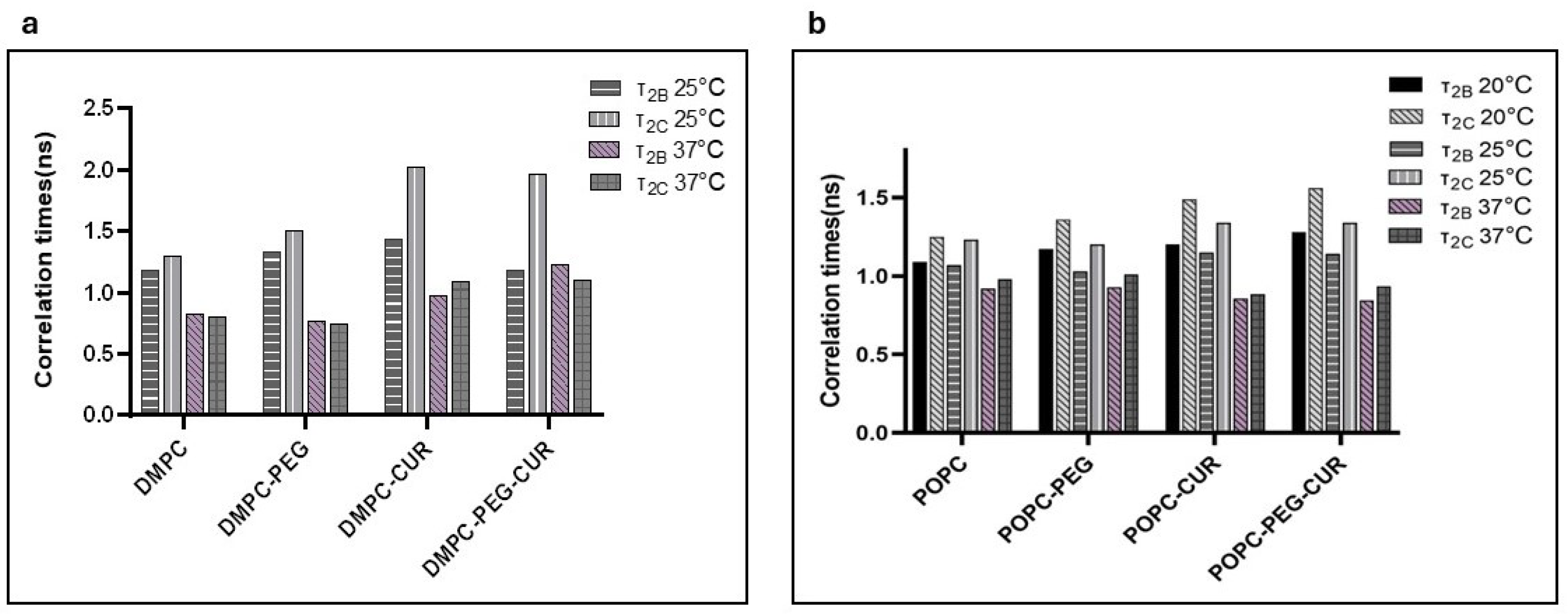
Figure 6.
Polarity profiles across DMPC (a), DMPC-PEG (b), POPC (c) and POPC-PEG (d) membranes in the presence and absence of 10 mol% curcumin. Upward changes of 2Az indicate a decrease in polarity. Approximate locations of the nitroxide moieties of spin labels are indicated by arrows.
Figure 6.
Polarity profiles across DMPC (a), DMPC-PEG (b), POPC (c) and POPC-PEG (d) membranes in the presence and absence of 10 mol% curcumin. Upward changes of 2Az indicate a decrease in polarity. Approximate locations of the nitroxide moieties of spin labels are indicated by arrows.
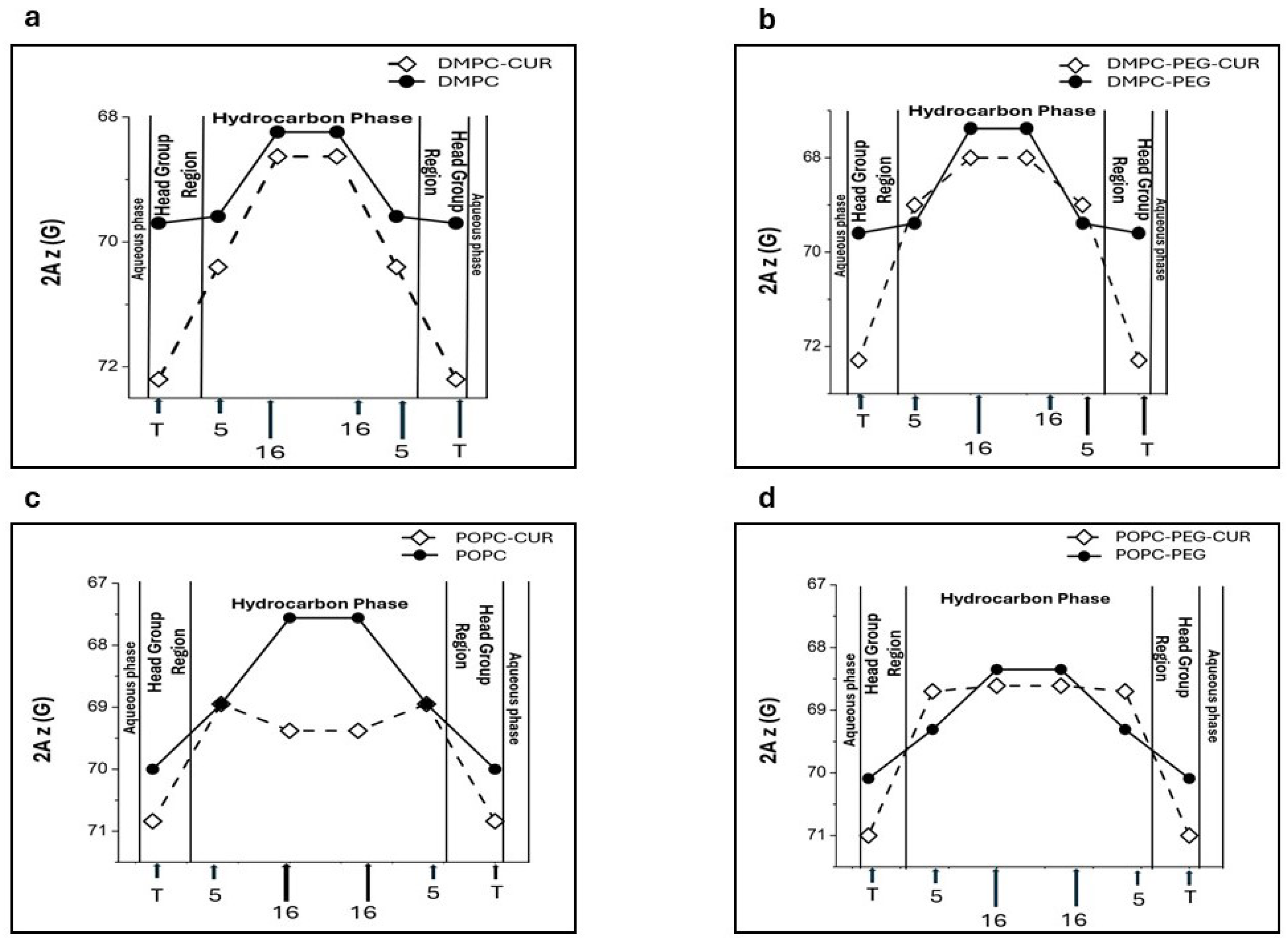
Figure 7.
Profiles of oxygen transport parameters across DMPC (a), DMPC-PEG (b), POPC (c), and POPC-PEG (d).
Figure 7.
Profiles of oxygen transport parameters across DMPC (a), DMPC-PEG (b), POPC (c), and POPC-PEG (d).
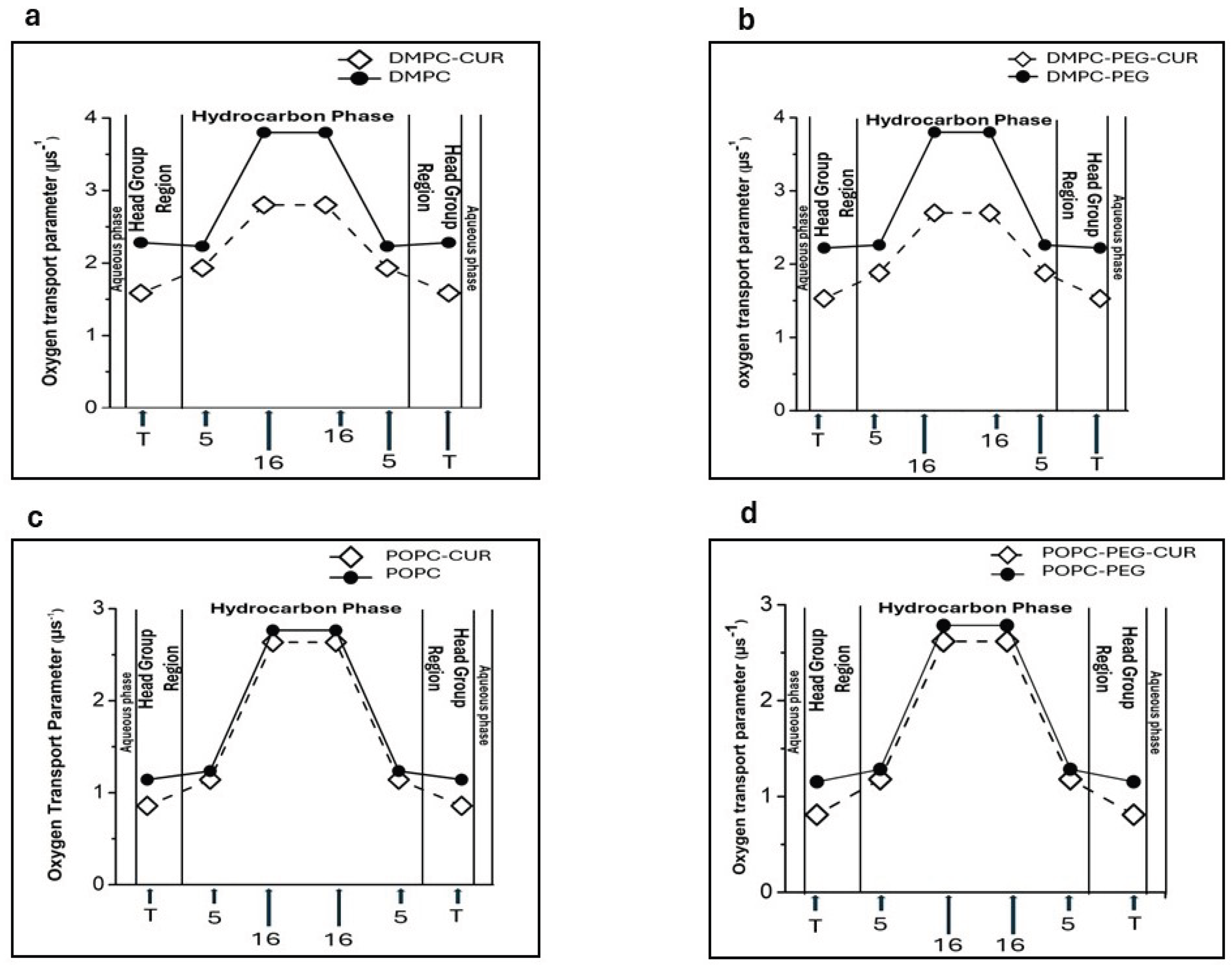

Table 1.
Binding constants obtained from the global fitting of curcumin interaction with POPC, POPC-PEG, DMPC, and DMPC-PEG liposomes.
Table 1.
Binding constants obtained from the global fitting of curcumin interaction with POPC, POPC-PEG, DMPC, and DMPC-PEG liposomes.
| Samples | Kd* (μM) |
|---|---|
| POPC | 270 ± 89 |
| POPC-peg | 156 ± 46 |
| DMPC | 263 ± 58 |
| DMPC-peg | 88 ± 21 |
1 Values represent the best-fit parameters ± standard errors (SE) obtained from global fitting using the DynaFit program.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



