
Submitted:
20 November 2025
Posted:
24 November 2025
You are already at the latest version
Abstract
This paper posits that the most significant long-term existential risk to human civilization is a chronic, systemic decay driven by the psychological and demographic consequences of a biologically capped lifespan. The entrenched expectation of mortality before 120 years fosters "temporal myopia," which cultivates cultural short-termism, consumerist nihilism, and demographic apathy. A critical aspect of this risk is the observed strong negative correlation between high cognitive ability and reproductive rates, leading to a systematic, dysgenic drain on humanity's problem-solving capacity. To counter this, we propose a novel biomedical paradigm: a strategy of continuous personal rejuvenation based on in-vitro gametogenesis (IVG) to generate autologous gametes, followed by auto-fertilization to create a new embryonic lineage. This protocol enables a comprehensive reset of cellular age, including the critical de novo formation of young centrioles, addressing the Centriolar Theory of Organismal Aging. The resulting young, perfectly matched adult stem cells are proposed for periodic autologous transplantation to maintain the body's regenerative potential indefinitely. We argue that this intervention transcends its medical purpose, acting as a necessary civilizational safeguard to prevent a slow-motion intellectual and demographic collapse.
Keywords:
biogerontology
; existential risk
; in-vitro gametogenesis
; centriolar theory of aging
; cognitive decline
; demographic transition
; rejuvenation biotechnology
; stem cell therapy
; temporal myopia
1. Introduction
For decades, the discourse on existential risks has been dominated by acute threats like nuclear annihilation and runaway climate change (Bostrom, 2014). These dangers overshadow a more insidious crisis: the civilizational consequence of a biologically capped human lifespan. The tacit acceptance of a maximum life expectancy, with a theoretical ceiling near 120 years (Dong et al., 2016), is not merely a medical statistic; it is a psychological and sociological paradigm that actively undermines humanity's long-term resilience. This paradigm, which we term "Temporal Myopia," fosters a culture of short-termism and demographic apathy, creating a feedback loop that systematically erodes our collective cognitive capital (Sandeep & Gopesh, 2017).
The specific danger is profound: the ingrained expectancy of mortality catalyzes cultural nihilism, drives demographic decline, and precipitates the systematic disappearance of the most intellectually capable individuals from the gene pool. Empirical evidence consistently demonstrates a significant negative correlation between cognitive ability and reproductive rates in developed societies (Beauchamp, 2016; Kanazawa, 2014). This creates a recursive, long-term civilizational risk far greater than any single technological threat, as it directly attacks the engine of human ingenuity (Lynn & Harvey, 2008).
The prevailing biomedical approach to aging, which focuses on combating individual age-related diseases, is akin to plugging holes in a sinking ship without addressing the corrosive seawater that is biological aging itself (López-Otín et al., 2013). Therefore, a paradigm shift is required. We must reconceptualize aging as a malleable biological process (de Magalhães, 2014). This paper proposes that the most effective countermeasure is a strategic intervention aimed at the root cause. We outline a detailed biotechnological strategy for continuous rejuvenation, a process designed to maintain an individual in a state of youth and health indefinitely, based on a novel application of in-vitro gametogenesis (IVG).
2. The Central Problem: Civilizational Nihilism from a Capped Lifespan
The tacit acceptance of a biological ceiling of ~120 years is an active agent shaping human psychology, culture, and demography (Dong et al., 2016). This "temporal myopia" manifests as a triad of pathologies:
2.1. The Psychology of Temporal Myopia and Short-Termism
The human brain is influenced by heuristics that favor immediate rewards, a phenomenon known as temporal discounting (Green & Myerson, 2004). When the "future" is constrained, this bias becomes a cultural paradigm. Neuroeconomics shows that finite time perception influences decision-making circuits, promoting choices with immediate payoffs (Peters & Büchel, 2011). This explains the dominance of short-term planning cycles in economics and politics (Sandeep & Gopesh, 2017; Jacobs, 2016). Terror Management Theory further illustrates how awareness of death can devalue the far future, creating a "legacy gap" (Pyszczynski et al., 2015; Wade-Benzoni, 2002).
2.2. Consumerist Nihilism and the Erosion of Meaning
A short-term perspective in a secular world leads to consumerist nihilism. If life is finite, the accumulation of experiences and possessions becomes a primary source of purpose (Kasser & Sheldon, 2000). Materialistic value orientation is linked to lower well-being and higher ecological footprints (Brown & Kasser, 2005), creating a feedback loop that depletes resources (Diener et al., 2006). This mindset is anathema to the long-term projects civilization requires (Bostrom, 2014).
2.3. Demographic Apathy and the Intellectual Drain
The most concrete consequence is the global shift towards sub-replacement fertility rates (Vollset et al., 2020). The decision to reproduce is an act of faith in the future; temporal myopia shifts this cost-benefit analysis against childbearing (Balbo et al., 2013). Crucially, this trend is qualitative: the opportunity costs of childbearing rise disproportionately for the most intellectually engaged individuals (Beauchamp, 2016; Kanazawa, 2014), creating a recursive, dysgenic loop (Lynn & Harvey, 2008). Underlying genetic factors may accelerate this decline (Barban et al., 2016).

Table 1.
The Triad of Civilizational Pathologies from a Capped Lifespan.
| Pathology | Manifestation | Key References |
|---|---|---|
| Temporal Myopia & Short-Termism | Quarterly profits, short political cycles, devaluation of long-term infrastructure and research. | Sandeep & Gopesh, 2017; Jacobs, 2016; Peters & Büchel, 2011 |
| Consumerist Nihilism | Materialism, hedonic treadmill, erosion of intergenerational meaning and projects. | Kasser & Sheldon, 2000; Brown & Kasser, 2005; Diener et al., 2006 |
| Demographic Apathy & Intellectual Drain | Sub-replacement fertility, negative correlation between cognitive ability and reproduction. | Vollset et al., 2020; Beauchamp, 2016; Kanazawa, 2014; Lynn & Harvey, 2008 |
3. The Mechanism of Intellectual Decline: Dysgenic Reproduction Patterns
The impact of temporal myopia is profoundly selective, creating a critical feedback loop that directly targets humanity's cognitive engine.
3.1. Empirical Evidence for the Negative Correlation
The inverse relationship between intelligence and fertility is well-replicated. Swedish data shows a negative association between IQ and probability of having children (Lynn & Van Court, 2004). UK data confirms higher childhood cognitive ability predicts lower fertility in women (Bajekal, 2005). Educational attainment shows a stark negative relationship with fertility (Skirbekk, 2008; Kravdal & Rindfuss, 2008), and individuals in cognitively demanding professions exhibit lower fertility rates (Hose et al., 2020).
3.2. The Causal Nexus
The mechanisms are multifactorial:
- Opportunity Cost: The high cost of pausing a demanding career for child-rearing (Hose et al., 2020; Balgopal, 2016).
- Existential Risk Awareness: Ethical concerns about overpopulation and environmental crises (Kellstedt et al., 2008).
- Delayed Gratification & Hyper-Agency: The trait of long-term planning paradoxically leads to delaying reproduction beyond the biological window (Shamosh & Gray, 2008; Balbo et al., 2013).
- Genetic Confound: Polygenic scores for educational attainment are negatively correlated with fertility, suggesting selective pressure against cognitive traits (Beauchamp, 2016; Conley, 2016).
3.3. The Long-Term Recursive Drain
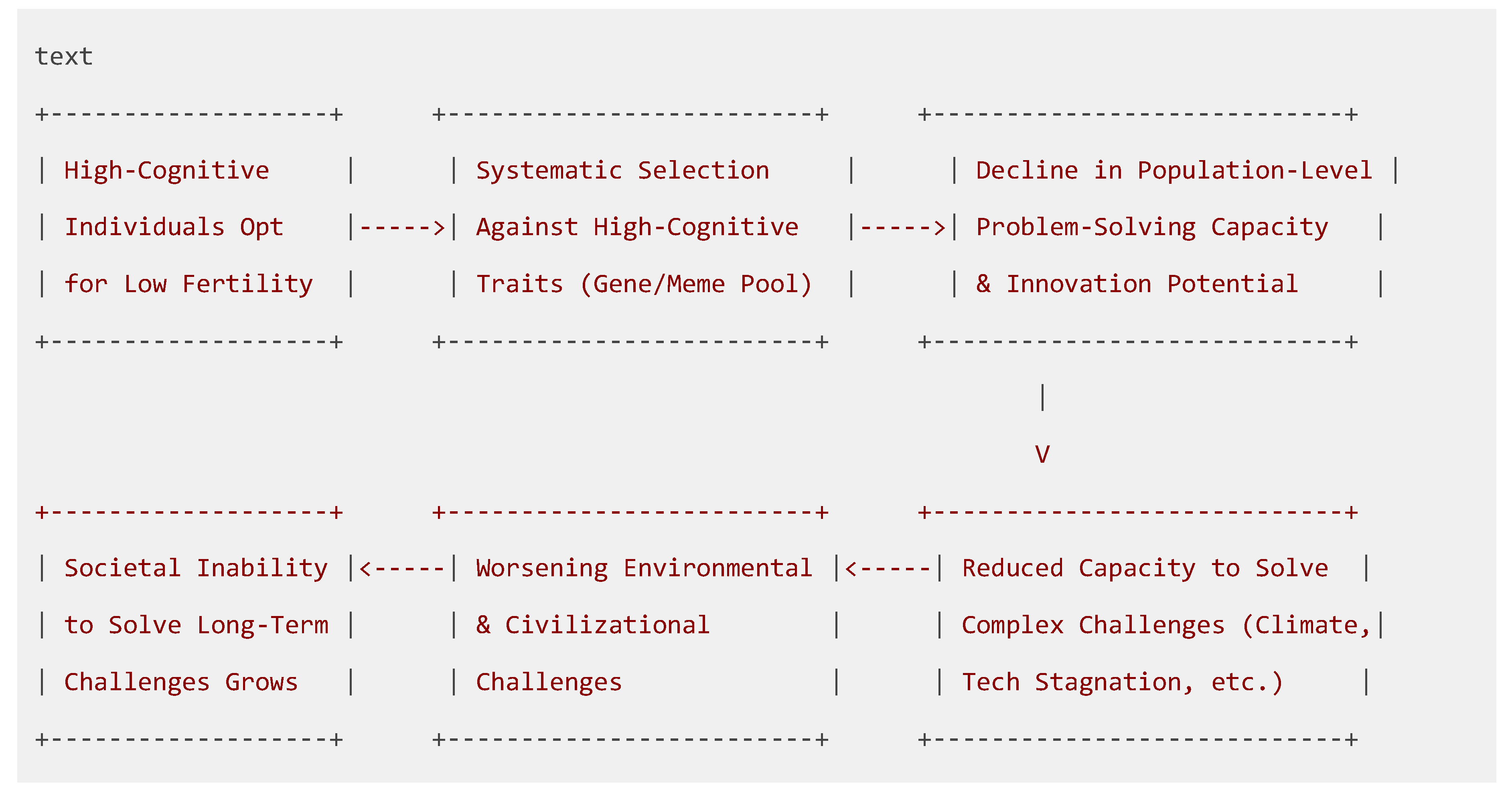
This is not a one-time loss but a recursive, civilizational-level decay. The effect compounds over time, creating a dangerous feedback loop where declining problem-solving capacity makes society less equipped to address the challenges that contributed to the dysgenic pressure (Lynn & Harvey, 2008; Woodley of Menie et al., 2017).
Figure 1.
The Dysgenic Feedback Loop of Intellectual Decline. (This figure illustrates the recursive cycle described in Section 3.3).
Figure 1.
The Dysgenic Feedback Loop of Intellectual Decline. (This figure illustrates the recursive cycle described in Section 3.3).
4. The Biotechnological Intervention: A Strategy of Continuous Rejuvenation
To counter this civilizational trajectory, we propose a paradigm shift from treating diseases to targeting aging itself. The strategy is to periodically replenish the body's aged stem cell populations with new, young, autologous stem cells.
4.1. The Scientific Premise: Stem Cell Exhaustion
The body's aging is driven largely by the functional decline of endogenous stem cell populations, which accumulate damage and enter senescence (Goodell & Rando, 2015; López-Otín et al., 2013).
4.2. The Protocol: In-Vitro Gametogenesis (IVG) and Auto-Fertilization
The key innovation is using IVG to generate autologous gametes (oocytes and sperm) from a patient's somatic cells (Hayashi et al., 2012; Hikabe et al., 2016; Zhou et al., 2016). These gametes are then used in auto-fertilization to create a new embryonic lineage.
- Advantage over SCNT (Cloning): Somatic Cell Nuclear Transfer (Wilmut et al., 1997) fails to reset the aged centriole from the donor somatic cell, carrying forward age-related defects (Simerly et al., 2003). Our IVG-based protocol recapitulates natural gametogenesis, where parental centrioles are eliminated, allowing for de novo formation of young centrioles upon fertilization (Szollosi et al., 1972; Fishman et al., 2017).
4.3. The Centriolar Theory of Organismal Aging and its Reversal
The Centriolar Theory of Organismal Aging posits that progressive centriolar dysfunction acts as a fundamental timer of aging (Tkemaladze, 2023). Aged centrioles lead to:
- Mitotic Errors: Chromosome mis-segregation and aneuploidy (Gönczy, 2015).
- Ciliopathy and Signaling Defects: Disruption of crucial signaling pathways (Anvarian et al., 2019).
- Stem Cell Exhaustion: Disruption of asymmetric cell division, depleting regenerative pools (Yamashita et al., 2010; Liang et al., 2020).
By generating new, pristine centrioles de novo, we directly target this proposed root cause.
Figure 2.
Comparative Schematic of SCNT vs. IVG-based Rejuvenation Protocol. (This figure illustrates the key technical difference and advantage of the proposed method).
Figure 2.
Comparative Schematic of SCNT vs. IVG-based Rejuvenation Protocol. (This figure illustrates the key technical difference and advantage of the proposed method).

4.4. The Rejuvenation Protocol: A Periodic Treatment
The therapeutic regimen would be periodic. The derived young stem cells (e.g., Hematopoietic Stem Cells (Ditadi et al., 2015) or Mesenchymal Stem Cells (Ullah et al., 2015)) are transplanted back into the patient. These cells, with renewed division potential and young centrioles, would engraft and outcompete resident aged stem cells, leading to systemic rejuvenation of blood, immune system, and tissues (Goodell & Rando, 2015).
Table 2.
Comparison of Rejuvenation Strategies.
| Feature | Current Approach (Treating Diseases) | Harvesting Own Adult Stem Cells | Proposed IVG-based Strategy |
| Target | Symptoms (e.g., cancer, dementia) | N/A (propagates aged state) | Root cause (cellular aging) |
| Epigenetic Age | No change | Aged | Reset to embryonic state |
| Centriolar Age | No change | Aged | Reset to young state (de novo) |
| Long-term Efficacy | Low (whack-a-mole) | None | High (periodic systemic reset) |
| Civilizational Impact | None | None | High (safeguard against intellectual decline) |
5. The Societal Benefit: Civilizational Safeguard
This intervention transcends individual medicine, offering a direct antidote to the systemic risks outlined.
5.1. Halting the Intellectual Drain and Accumulating Wisdom
By extending the healthy, productive lifespans of the most capable individuals, we arrest the recursive drain on cognitive capital. A scientist or philosopher with a 300-year lifespan could achieve unimaginable mastery (Maher & Baum, 2013), extending the period of peak cognitive output (Hartshorne & Germine, 2015). It also allows for the deep accumulation of wisdom, a trait correlated with age but cut short by mortality (Jeste et al., 2010), improving societal decision-making.
5.2. Transforming Demographics and Alleviating the Burden of Aging
Rejuvenation biotechnology fundamentally reshapes problematic demographics. By compressing morbidity, the concept of "retirement age" becomes obsolete (Farrelly, 2020). Individuals remain productive for centuries, transforming the dependency ratio, alleviating economic burdens, and reducing healthcare costs associated with aging (Goldman et al., 2013; Weber, 2020).
5.3. Enhancing Cultural and Scientific Continuity
A long-lived society possesses a stronger cultural and scientific memory. Lessons from past crises are retained in living memory, and research programs can be pursued with a consistency that outstrips a single lifetime (Maher & Baum, 2013; de Grey & Rae, 2007). The preservation of "tacit knowledge" accelerates progress (Weber, 2020).
5.4. Ethical and Equitable Implementation as a Prerequisite
The societal benefits are contingent upon equitable distribution. If restricted to a wealthy elite, it could create a biological caste system (Partridge et al., 2011). Therefore, development must be paired with policy frameworks ensuring broad availability as a universal human right (Farrelly, 2020). Public concerns about overpopulation, boredom, and equity must be addressed proactively (Partridge et al., 2009).
6. Discussion and Conclusion
6.1. Synthesis
Our thesis connects a microscopic biological mechanism (centriolar reset) to a macroscopic civilizational outcome (preservation of intellectual capital). The proposed IVG-based protocol offers a pathway to comprehensive cellular rejuvenation, directly addressing the Centriolar Theory of Aging (Tkemaladze, 2023). By decoupling healthspan from chronological age, we initiate a positive feedback loop: retained intellectual capital accelerates scientific progress, which further refines rejuvenation therapies.
6.2. Addressing Limitations
The vision faces significant challenges:
- Scientific Hurdles: Perfecting human IVG, ensuring genomic stability, and safe differentiation of stem cells are formidable tasks (Zhou et al., 2016; Ditadi et al., 2015).
- The Equity Problem: This is the most significant ethical challenge, requiring proactive policy to avoid a dystopian outcome (Partridge et al., 2011; Farrelly, 2020).
- Societal Adaptation: Society would need to rethink concepts of career, relationships, and meaning (Weber, 2020), though the goal is healthspan, not prolonged decrepitude.
- Overpopulation: This Malthusian objection ignores demographic trends; the societies adopting this will have low birth rates, and solutions lie in the innovation a long-lived society enables (Vollset et al., 2020).
6.3. Conclusion
The choice is clear: accept a slow, certain intellectual and demographic decline, or invest in a future where biological aging is a treatable condition. The pursuit of rejuvenation biotechnology is not a niche medical quest but a strategic, necessary, and urgent investment in the very foundations of a flourishing long-term civilization. It is the most profound safeguard we can devise against the quiet, creeping threat of our own biological limitations.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Anvarian, Z., Mykytyn, K., Mukhopadhyay, S., Pedersen, L. B., & Christensen, S. T. (2019). Cellular signalling by primary cilia in development, organ function and disease. Nature Reviews Nephrology, 15(4), 199–219. [CrossRef]
- Bajekal, M. (2005). Healthy life expectancy by area deprivation: magnitude and trends in England, 1994-1999. Health Statistics Quarterly, 25, 18–27. https://pubmed.ncbi.nlm.nih.gov/15804167/.
- Balbo, N., Billari, F. C., & Mills, M. (2013). Fertility in advanced societies: A review of research. European Journal of Population, 29(1), 1–38. [CrossRef]
- Balgopal, P. R. (2016). The impact of female education on fertility: A natural experiment from Nigeria. World Development, 87, 152–169. [CrossRef]
- Barban, N., Jansen, R., de Vlaming, R., Vaez, A., Mandemakers, J. J., Tropf, F. C., ... & Mills, M. C. (2016). Genome-wide analysis identifies 12 loci influencing human reproductive behavior. Nature Genetics, 48(12), 1462–1472. [CrossRef]
- Beauchamp, J. P. (2016). Genetic evidence for natural selection in humans in the contemporary United States. Proceedings of the National Academy of Sciences, 113(28), 7774–7779. [CrossRef]
- Bostrom, N. (2014). Superintelligence: Paths, dangers, strategies. Oxford University Press.
- Brown, K. W., & Kasser, T. (2005). Are psychological and ecological well-being compatible? The role of values, mindfulness, and lifestyle. Social Indicators Research, 74(2), 349–368. [CrossRef]
- Chatterji, S., Byles, J., Cutler, D., Seeman, T., & Verdes, E. (2015). Health, functioning, and disability in older adults—present status and future implications. The Lancet, 385(9967), 563–575. [CrossRef]
- Conley, D. (2016). Socio-genomic research using genome-wide molecular data. Annual Review of Sociology, 42, 275–299. [CrossRef]
- de Grey, A., & Rae, M. (2007). Ending Aging: The Rejuvenation Breakthroughs That Could Reverse Human Aging in Our Lifetime. St. Martin's Press.
- de Magalhães, J. P. (2014). The scientific quest for lasting youth: prospects for curing aging. Rejuvenation Research, 17(5), 458–467. [CrossRef]
- Diener, E., Lucas, R. E., & Scollon, C. N. (2006). Beyond the hedonic treadmill: Revising the adaptation theory of well-being. American Psychologist, 61(4), 305–314. [CrossRef]
- Ditadi, A., Sturgeon, C. M., & Keller, G. (2015). A view of human haematopoietic development from the Petri dish. Nature Reviews Molecular Cell Biology, 16(1), 56–67. [CrossRef]
- Dong, X., Milholland, B., & Vijg, J. (2016). Evidence for a limit to human lifespan. Nature, 538(7624), 257–259. [CrossRef]
- Farrelly, C. (2020). The case for optimism about human longevity. The Journals of Gerontology: Series A, 75(1), 35–38. [CrossRef]
- Fishman, E. L., Jo, K., Nguyen, Q. P. H., Kong, D., Royfman, R., Cekic, A. R., ... & Avidor-Reiss, T. (2017). A novel atypical sperm centriole is functional during human fertilization. Nature Communications, 8(1), 2210. [CrossRef]
- Goldman, D. P., Cutler, D., Rowe, J. W., Michaud, P. C., Sullivan, J., Peneva, D., & Olshansky, S. J. (2013). Substantial health and economic returns from delayed aging may warrant a new focus for medical research. Health Affairs, 32(10), 1698–1705. [CrossRef]
- Gönczy, P. (2015). Centrosomes and cancer: revisiting a long-standing relationship. Nature Reviews Cancer, 15(11), 639–652. [CrossRef]
- Goodell, M. A., & Rando, T. A. (2015). Stem cells and healthy aging. Science, 350(6265), 1199–1204. [CrossRef]
- Green, L., & Myerson, J. (2004). A discounting framework for choice with delayed and probabilistic rewards. Psychological Bulletin, 130(5), 769–792. [CrossRef]
- Hartshorne, J. K., & Germine, L. T. (2015). When does cognitive functioning peak? The asynchronous rise and fall of different cognitive abilities across the life span. Psychological Science, 26(4), 433–443. [CrossRef]
- Hayashi, K., Ogushi, S., Kurimoto, K., Shimamoto, S., Ohta, H., & Saitou, M. (2012). Offspring from oocytes derived from in vitro primordial germ cell-like cells in mice. Science, 338(6109), 971–975. [CrossRef]
- Hikabe, O., Hamazaki, N., Nagamatsu, G., Obata, Y., Hirao, Y., Hamada, N., ... & Saitou, M. (2016). Reconstitution in vitro of the entire cycle of the mouse female germ line. Nature, 539(7628), 299–303. [CrossRef]
- Hose, A., Küspert, S., & Schneider, M. V. (2020). The academic career landscape: A systematic review of the literature on academic work and life. Studies in Higher Education, 45(12), 2465–2482. [CrossRef]
- Jacobs, A. M. (2016). Policy making for the long term in advanced democracies. Annual Review of Political Science, 19, 433–454. [CrossRef]
- Jeste, D. V., Ardelt, M., Blazer, D., Kraemer, H. C., Vaillant, G., & Meeks, T. W. (2010). Expert consensus on characteristics of wisdom: a Delphi method study. The Gerontologist, 50(5), 668–680. [CrossRef]
- Kanazawa, S. (2014). Intelligence and childlessness. Social Science Research, 48, 157–170. [CrossRef]
- Kasser, T., & Sheldon, K. M. (2000). Of wealth and death: Materialism, mortality salience, and consumption behavior. Psychological Science, 11(4), 348–351. [CrossRef]
- Kellstedt, P. M., Zahran, S., & Vedlitz, A. (2008). Personal efficacy, the information environment, and attitudes toward global warming and climate change in the United States. Risk Analysis, 28(1), 113–126. [CrossRef]
- Kravdal, Ø., & Rindfuss, R. R. (2008). Changing relationships between education and fertility: A study of women and men born 1940 to 1964. American Sociological Review, 73(5), 854–873. [CrossRef]
- Liang, Y., Yang, N., Pan, G., Jin, L., & Liu, F. (2020). Centrosome dysfunction in hematopoietic stem cells and its clinical relevance. Stem Cell Reviews and Reports, 16(4), 637–647. [CrossRef]
- López-Otín, C., Blasco, M. A., Partridge, L., Serrano, M., & Kroemer, G. (2013). The hallmarks of aging. Cell, 153(6), 1194–1217. [CrossRef]
- Lynn, R., & Harvey, J. (2008). The decline of the world's IQ. Intelligence, 36(2), 112–120. [CrossRef]
- Lynn, R., & Van Court, M. (2004). New evidence of dysgenic fertility for intelligence in the United States. Intelligence, 32(2), 193–201. [CrossRef]
- Maher, T. M., & Baum, S. D. (2013). Adaptation to and recovery from global catastrophe. Sustainability, 5(4), 1461–1479. [CrossRef]
- Partridge, B., Lucke, J., & Hall, W. (2011). If we could live forever, should we? And who gets to decide? The American Journal of Bioethics, 11(9), 57–59.
- Partridge, B., Lucke, J., Bartlett, H., & Hall, W. (2009). Ethical, social, and personal implications of extended human lifespan identified by members of the public. Rejuvenation Research, 12(5), 351–357. [CrossRef]
- Peters, J., & Büchel, C. (2011). The neural mechanisms of inter-temporal decision-making: understanding variability. Trends in Cognitive Sciences, 15(5), 227–239. [CrossRef]
- Pyszczynski, T., Solomon, S., & Greenberg, J. (2015). Thirty years of terror management theory: From genesis to revelation. In Advances in Experimental Social Psychology (Vol. 52, pp. 1-70). Academic Press. [CrossRef]
- Sandeep, S., & Gopesh, A. (2017). The influence of temporal myopia on decision making. Journal of Neuroscience, Psychology, and Economics, 10(2-3), 89–104. [CrossRef]
- Shamosh, N. A., & Gray, J. R. (2008). Delay discounting and intelligence: A meta-analysis. Intelligence, 36(4), 289–305. [CrossRef]
- Simerly, C., Dominko, T., Navara, C., Payne, C., Capuano, S., Gosman, G., ... & Schatten, G. (2003). Molecular correlates of primate nuclear transfer failures. Science, 300(5617), 297. [CrossRef]
- Skirbekk, V. (2008). Fertility trends by social status. Demographic Research, 18, 145–180. [CrossRef]
- Szollosi, D., Calarco, P., & Donahue, R. P. (1972). Absence of centrioles in the first and second meiotic spindles of mouse oocytes. Journal of Cell Science, 11(2), 521–541. [CrossRef]
- Tkemaladze, J. (2023). Reduction, proliferation, and differentiation defects of stem cells over time: a consequence of selective accumulation of old centrioles in the stem cells?. Molecular Biology Reports, 50(3), 2751–2761. [CrossRef]
- Ullah, I., Subbarao, R. B., & Rho, G. J. (2015). Human mesenchymal stem cells - current trends and future prospective. Bioscience Reports, 35(2), e00191. [CrossRef]
- Vollset, S. E., Goren, E., Yuan, C. W., Cao, J., Smith, A. E., Hsiao, T., ... & Murray, C. J. L. (2020). Fertility, mortality, migration, and population scenarios for 195 countries and territories from 2017 to 2100: a forecasting analysis for the Global Burden of Disease Study. The Lancet, 396(10258), 1285-1306. [CrossRef]
- Wade-Benzoni, K. A. (2002). A golden rule over time: Reciprocity in intergenerational allocation decisions. Academy of Management Journal, 45(5), 1011–1028. [CrossRef]
- Weber, A. M. (2020). The social implications of longevity. Nature Aging, 1(1), 8–9. [CrossRef]
- Wilmut, I., Schnieke, A. E., McWhir, J., Kind, A. J., & Campbell, K. H. (1997). Viable offspring derived from fetal and adult mammalian cells. Nature, 385(6619), 810–813. [CrossRef]
- Woodley of Menie, M. A., Figueredo, A. J., Sarraf, M. A., Hertler, S., & Fernandes, H. B. (2017). The rhythm of the west: A biohistory of the modern era, AD 1600 to the present. Journal of Social, Political, and Economic Studies, 32(4), 1–40.
- Yamashita, Y. M., Yuan, H., Cheng, J., & Hunt, A. J. (2010). Polarity in stem cell division: asymmetric stem cell division in development, cancer and aging. Development, 137(1), 19–32.
- Zhou, Q., Wang, M., Yuan, Y., Wang, X., Fu, R., Wan, H., ... & Xie, W. (2016). Complete meiosis from embryonic stem cell-derived germ cells in vitro. Cell Stem Cell, 18(3), 330–340. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



