Submitted:
20 November 2025
Posted:
24 November 2025
You are already at the latest version
Abstract
Lactic acid accumulates in the tumour microenvironment (TME) at concentrations reaching up to 40 mM. Initially, lactic acid was regarded solely as a metabolic byproduct of aerobic glycolysis, a phenomenon commonly referred to as the Warburg effect and observed in the majority of tumours. Recent evidence, however, has demonstrated that lactic acid is not merely a waste product; rather, it plays a pivotal role in tumour biology. Elevated lactic acid levels in the TME have been shown to suppress antitumour immune responses, facilitate both metastasis and cellular senescence, and modulate gene expression through novel epigenetic mechanisms such as histone lactylation. This review aims to summarize current knowledge on the multifaceted impact of elevated lactic acid in the TME on tumour progression and biology.
Keywords:
cancer
; tumour microenvironment
; lactic acid
; lactate
; Warburg effect
; immune function
1. Introduction
Lactic acid (2-hydroxypropanoic acid) derives its common name from the Latin word lac, meaning milk. It was first identified in sour milk by Karl Wilhelm Scheele in 1780. Later, Louis Pasteur demonstrated that lactic acid results from the bacterial fermentation of sugars, while Justus von Liebig confirmed its consistent presence in the muscular tissue of deceased organisms [1].
In healthy individuals, lactic acid is found in the blood at concentrations of approximately 1–2 mM. Given its dissociation constant (pKa) of 3.86, it exists predominantly in its anionic form, lactate, at physiological pH. Unless otherwise stated, throughout this review, the terms lactic acid and lactate will be used interchangeably. Most human cells possess the enzymatic machinery to synthesize and efflux lactate into the extracellular milieu space a process that accounts for the baseline plasma concentration. Production can increase under specific conditions, such as limited oxygen availability during muscle exercise [2] or in rapidly proliferating tissues, including those during foetal development [3]. The excess, extracellularly secreted lactate is subsequently processed via hepatic gluconeogenesis in a pathway termed the Cori cycle [4]. Lactate is now recognized as a major circulating carbohydrate fuel and, together with pyruvate, serves as a redox buffer, helping regulate the NADH/NAD ratio [5].
Tumour cells exhibit a distinct metabolic signature [6,7], most notably the conversion of glucose to lactate even under aerobic conditions with functional mitochondria—a phenomenon known as the Warburg effect [8]. This metabolic shift can lead to lactic acid levels of up to 40 mM in the tumour microenvironment (TME) [9], accompanied by decreased pH (acidification). Elevated lactate concentrations in the tumour microenvironment correlate with increased metastatic potential, whereas tumours with lower lactate levels are associated with improved patient survival [9,10]. While common in many cancers, the Warburg effect is not universal. Documented examples include glioblastoma [11], pancreatic cancer [12], breast cancer [13], and cervical cancer [14] among others.
The TME is a complex, heterogeneous entity comprising immune cells, stromal cells, vasculature, and the extracellular matrix [15]. The TME actively contributes to the evasion of anti-tumour immunity, thereby diminishing the efficacy of immunotherapeutic strategies [16], and can alter the tumour’s responsiveness to chemotherapy and radiotherapy [17]. Within the TME, lactic acid plays multiple roles: it is utilized as an energy substrate, operates as a signalling molecule, and holds clinical significance. This review aims to explore the wide-ranging impacts of lactic acid on tumour biology.
2. The Origin of Lactic Acid in the Tumour Microenvironment
Metabolic dysregulation is a hallmark of cancer [6,7]. The most prevalent manifestation of this dysregulation is the uncoupling of glycolysis from mitochondrial oxidative phosphorylation (OXPHOS), a phenomenon known as the Warburg effect [18].
Upon cellular uptake, glucose is metabolized through glycolysis to pyruvate, producing two molecules of adenosine triphosphate (ATP) and two molecules of reducing equivalent NADH. While ATP provides energy for cellular processes, the intracellular accumulation of NADH can inhibit the continuous flux of glycolysis. To sustain glycolytic activity, NADH must be rapidly oxidized back to NAD+. This is achieved by the reduction of pyruvate to lactate, a reaction predominantly catalysed by the enzyme lactate dehydrogenase (LDH), thereby regenerating NAD+.
Empirical evidence, such as the observed correlation between lactate dehydrogenase A (LDHA) expression and elevated lactic acid concentration [19], supports the central role of this enzyme in TME lactate accumulation. It is important to note that LDH exists as two primary isoforms: LDHA, which favours the reduction of pyruvate to lactate, and LDHB, which primarily catalyses the reverse reaction, the oxidation of lactate back to pyruvate [20]. This bi-directional capacity enables lactate to be re-assimilated and utilized within cellular metabolic pathways.
Following its formation, lactate must be exported in order for the cell to sustain glycolytic flux. This is achieved by monocarboxylate transporters (MCTs), a family of proton-linked transporters that mediate lactate and pyruvate exchange [21]. Among them, MCT4 serves as the principal lactate exporter, capable of functioning against steep extracellular lactate gradients [22], leading to extracellular concentrations reaching up to 40 mM [23]. MCT1, by contrast, predominantly facilitates lactate uptake into cells [21]. However, both transporters can mediate bidirectional flux, as effective inhibition of lactate export requires simultaneous blockade of MCT1 and MCT4 [24]. Although the metabolic basis of the Warburg effect involves multiple glycolytic enzymes [8], the net outcome is the accumulation of lactic acid and subsequent acidification of the tumour microenvironment.
Tumour cells represent the primary source of lactic acid within the tumour microenvironment, but stromal components also contribute. For instance, macrophages undergo a metabolic shift from a resting to a differentiated state that includes the robust production and secretion of lactate [25]. Similarly, cancer-associated fibroblasts (CAFs) are known to release lactate into the extracellular space [26], further enriching the lactate pool that characterizes the tumour milieu.
3. Acidification of the Extracellular Media
Lactate is exported from cells via monocarboxylate transporters (MCTs) in symport with protons [27]. Consequently, the augmented release of lactate into the extracellular space results in a concomitant acidification of the extracellular medium [28] with in vivo measurements revealing extracellular pH values close to 6, as detected by hyperpolarized 13C MR imaging [29]; even at this low pH the predominant form of lactic acid is in the anionic form lactate. This acidic microenvironment contributes to multiple cancer-promoting processes, including epithelial-to-mesenchymal transition, extracellular matrix degradation and remodelling, altered vascularization, and enhanced immune evasion [30]. Cancer cells adapt to such conditions through metabolic reprogramming mediated by PPARα signalling, which facilitates metastasis [31].
Acidic pH can act as a signalling cue, activating various acid-sensing ion channels and downstream pathways. These include increased reactive oxygen species (ROS) production and the activation of RhoA, NFAT, PI3K, and AKT signalling cascades [32]. In addition, TME acidosis can trigger specific oncogenic signalling such as NF-κB activation, promoting cell invasion [33], and can drive immune escape through pathways involving SAT1 activation and interferon-γ–mediated upregulation of PD-L1 expression [34].
High lactate concentrations and low extracellular pH play key roles in shaping interactions between tumour and stromal or immune cells within the tumour microenvironment [35]. Chronic acidosis creates a hostile niche for immune effectors [36,37]. Notably, Colegio and colleagues [38] demonstrated that lactic acid—but not lactate at physiological pH—induces a pro-tumoral macrophage phenotype, while acidity promotes tumour progression by reprogramming macrophages in prostate cancer [39]. Acidic pH also significantly attenuates the cytotoxic functions of T cells [40,41]. Moreover, recent findings reveal that pH fluctuations can drive a reversible transition of fibroblasts into cancer-associated fibroblasts, further supporting tumour progression [42].
4. Lactic Acid as an Energy Source: The Reverse Warburg Effect
Lactate is a high-energy metabolite capable of yielding up to 32 molecules of ATP through mitochondrial oxidative phosphorylation (OXPHOS) when fully oxidized to CO₂. Lactate generated by one tissue or cell type can be used by another for energy production, a process known as the lactate shuttle. The most well-known example is the Cori cycle, first described by Carl and Gerty Cori. This cycle describes lactate transfer from muscle to liver, where lactate is converted to glucose that can be returned to muscle to sustain activity when energy demand exceeds oxygen supply [4]. Although the Cori cycle has a net energy cost of –2 ATP molecules, it enables prolonged muscle function under hypoxic conditions. Lactate shuttling also occurs between different cell populations within a tissue; for example, astrocytes produce lactate that is released extracellularly and subsequently metabolized by mitochondria in neurons [43].
Lactate shuttles have also been observed within the tumour microenvironment, where this phenomenon is frequently referred to as the reverse Warburg effect. A tumour can be viewed as an evolving metabolic ecosystem in which cancer cells adopt strategies to optimize the use of available resources [44]. One form of metabolic symbiosis involves tumour cells in oxygenated regions metabolizing lactate secreted by hypoxic tumour cells that rely on glycolysis and cannot perform mitochondrial OXPHOS [45]. In breast cancer, tumour cells preferentially use lactate in oxygen-rich areas [46], further supporting the concept of intra-tumour lactate shuttling based on variable oxygen availability. Evidence from head and neck cancers shows concurrent expression of MCT1 and MCT4 [47] further supporting the idea of metabolic symbiosis between lactate-producing and lactate-consuming tumour cell populations.
The use of lactate as a fuel source appears to be widespread among cancers. Studies have shown that non-small-cell lung cancers (NSCLCs) are capable of metabolizing plasma lactate by incorporating it into the Krebs cycle (TCA cycle), a process that is dependent on the expression of MCT1 [48]. KRAS-mutant lung adenocarcinomas display greater glycolytic dependence than KRAS wild-type tumours, with LDHB expression correlating with poor survival [49]. LDHB, which catalyses the conversion of lactate to pyruvate [20,50,51], is considered a key mediator of the reverse Warburg effect, suggesting that lactate utilization confers a metabolic advantage to tumours.
Metabolic symbiosis also occurs between cancer cells and stromal components. Cancer-associated fibroblasts (CAFs) often exhibit elevated glycolysis and increased lactate export into the microenvironment [52]. Co-culture experiments with MCF7 breast cancer cells and fibroblasts have shown reciprocal expression changes—MCT4 upregulation in CAFs and MCT1 upregulation in MCF7 cells—providing evidence of a stromal-epithelial lactate shuttle [53]. Similar patterns have been observed in human breast cancer samples, with CAFs from surgical resections showing higher MCT4 expression and lactate secretion compared to normal fibroblasts [54]. In prostate cancer, CAF-derived lactate is metabolized via the Krebs cycle to meet the high energetic demands of tumour cells [55]. Direct contact between cancer cells and CAFs can reprogram CAFs toward a Warburg-like glycolytic phenotype, while cancer cells switch to aerobic metabolism with increased lactate uptake through MCT1 [26]. In hepatocellular carcinoma, CAFs act as metabolic hubs, enhancing glycolysis and secreting lactic acid to fuel tumour growth [56].
5. Lactate as a Signalling Molecule
Within the tumour microenvironment, lactate functions not only as a metabolic by-product but also as a potent signalling molecule. It can interact with membrane-bound receptors, most notably the G protein–coupled receptor GPR81 (Figure 1), encoded by the HCAR1 gene [57,58]. GPR81 expression correlates positively with tumour growth and metastasis, and genetic deletion of GPR81 markedly suppresses tumour progression [59]. Activation of the GPR81 signalling axis induces a metabolic shift in cancer cells by enhancing glycolysis, creating an apparent autocrine feed-forward loop [60,61,62]. Notably, GPR81 expression is associated with the expression of primary lactate transporters MCT1 and MCT4 [63], supporting the concept of lactate-driven autocrine regulation. However, MCT4 has also been shown to activate signalling cascades independent of ambient lactate [64]. Lactate can promote metastasis in normoxic colorectal cancer stem cells through PGC-1α–mediated signalling [65] indicating that lactate signalling is not confined to hypoxic tumour regions.
GPR81 contributes to the creation of an immunosuppressive microenvironment. It facilitates the recruitment of immunosuppressive polymorphonuclear myeloid-derived suppressor cells by upregulating CCL2 and CCL7 [66]. In antigen-presenting cells within the TME, GPR81-mediated signalling supports breast cancer growth via paracrine mechanisms [67,68]. Additionally, in gastric cancer, it promotes regulatory T cell (Treg) migration through CX3CL1 secretion [69]. Furthermore, GPR81 has been postulated as a key mediator in the induction of cancer-associated cachexia [70].
Beyond receptor-mediated signalling, lactic acid can regulate gene expression through lactylation (Figure 1) —a post-translational modification in which lactate-derived groups are added to lysine residues on histones [71]. This mechanism has since been found to extend to non-histone proteins [72]. For instance, the protein High Mobility Group Box-1 (HMGB1) was observed to undergo lactylation in macrophages during septic conditions [73]. The extent of histone lactylation is directly proportional to intracellular lactate production, such as that driven by the Warburg effect, and has been shown to induce M2-like gene expression in macrophages [71]. Furthermore, TME derived lactic acid has been demonstrated to drive macrophage polarization via histone lactylation [74,75].
Lactate within the TME can also induce lactylation in tumour cells; for example, CAF-derived lactate drives the lactylation of histone H3 at lysine 18 (H3K18) in gastric cancer, promoting immune evasion [76]. In acute lymphoblastic leukemia, sphingomyelin-induced lactic acid production led to caspase-3 lactylation, which subsequently inhibited apoptosis [77]. Additionally, lactylation can facilitate the progression of colorectal cancer by establishing a positive feedback loop that further enhances lactate production [78].
6. Effects of Lactic Acid on the Immune System
The tumour microenvironment promotes immune evasion through multiple mechanisms [79], including the secretion of immunosuppressive cytokines such as TGF-β, [80] and LIF [81]. The behaviour of immune cells within the TME is shaped by various local factors, among which lactic acid plays a critical role [82]. Lactate, one of the most abundant metabolites in tumours, functions as both a metabolic substrate and a signalling molecule, acting through several receptors [57] to influence immune responses across diverse conditions [83]. Metabolic reprogramming of immune cells drives immunosuppressive phenotypes within both innate [84] and adaptive immune populations [85], impacting macrophages, T cells, myeloid-derived suppressor cells (MDSCs), and other stromal or immune-associated cells.
6.1. Macrophages
Macrophages are integral drivers of cancer pathophysiology, influencing tumour initiation, progression, and metastasis [86]. Their phenotypes and functions are tightly regulated by metabolic cues within the TME [87,88]. Lactic acid promotes the polarization of tumour-associated macrophages (TAMs) toward an immunosuppressive (M2-like) phenotype, enhancing tumour growth and survival [19,38]. Importantly, it is exogenous lactate imported via MCT1 that drives this polarization process [74]. This shift in macrophage phenotype is accompanied by a reciprocal metabolic reprogramming within the macrophages themselves [87]. Studies utilizing microfluidic systems demonstrated that lactate induces M2-like polarization in macrophages more rapidly than larger, slower diffusion, microenvironmental proteins [89].
Lactate-induced macrophage polarization has been reported across multiple cancer types. In head and neck cancer, lactate-dependent M2 polarization correlates with tumour aggressiveness [19]; in breast cancer, lactic acid induces M2 polarization through ERK/STAT3 pathway activation [90]; and in esophageal cancer, lactate enhances tumour growth via AKT/ERK signalling [91]. The lactate receptor GPR132 contributes to macrophage-mediated immune regulation within the TME, particularly in lung adenocarcinoma [92,93]. Moreover, lactate metabolism, through its conversion to pyruvate and subsequent mitochondrial oxidation—a process resembling the reverse Warburg effect—enhances histone acetylation and promotes the pro-tumoral macrophage phenotype [75]. Furthermore, in oral squamous cell carcinoma, tumour cell-derived lactic acid induces macrophage synthesis of glycoprotein non-metastatic protein B (GPNMB), which facilitates tumour cell migration and invasion [94]. Macrophages, in turn, interact with other immune cells, contributing to the suppression of T-cell recruitment [95].
6.2. T Cells
Effector T cells depend on glycolytic metabolism for their functions. Consequently, they are detrimentally affected in the TME due to the synergistic effects of low glucose availability and high lactate concentrations, which severely compromises their anti-tumour cytotoxic activity. Lactate accumulation impairs T-cell cytotoxicity and fosters the differentiation of regulatory T cells (Tregs) into pro-tumoral phenotypes. Inhibition of MCT1 disrupts lactate uptake and reduces the Treg-mediated suppression of antitumour responses [96]. In NSCLC, an LTB⁺LDHA⁺CD8⁺ T-cell subset exhibits enhanced glycolysis and lactate production, promoting tumour cell migration while impairing CD8⁺ cytotoxicity [97]. Acidic pH suppresses CD8⁺ T-cell activity via p38/JNK pathway inhibition and reduces interferon-γ production (INFγ) [98]. Furthermore, lactate stimulates PD-1 expression in Tregs [99] and activates TGF-β signalling, reinforcing their immunosuppressive profile [100].
6.3. Cancer-Associated Fibroblasts (CAFs)
The reciprocal contact between CAFs and neoplastic cells, as demonstrated in prostate cancer, induces a mutual metabolic rewiring: stromal fibroblasts increase GLUT1 expression, lactate production, and export via MCT4, adopting a glycolytic phenotype. Conversely, cancer cells shift toward an aerobic OXPHOS metabolism, exhibiting decreased GLUT1 expression and increased lactate uptake via MCT1 [26]. This process involves activation of the SIRT1/PGC-1α axis, enhancing mitochondrial respiration in tumour cells [101]. Beyond metabolic effects, lactate secreted by CAFs exerts direct immunosuppressive functions in pancreatic cancer TMEs [102].
6.4. Other Stromal and Immune Cells
The immunosuppressive composition of the TME also affects additional cell types. Lactic acid prevents monocyte differentiation into mature dendritic cells, promoting a tolerogenic phenotype and the secretion of anti-inflammatory cytokines [104]. Natural killer (NK) cell cytotoxicity is impaired under high lactate conditions through inhibition of the mTOR pathway [105]. Moreover, lactate can modulate NK cell activity via the proton-sensing receptor GPR132 [106].
7. Lactic Acid and Senescence
Cellular senescence is a stress-induced state characterized by permanent proliferative arrest and heterogeneous phenotypes that may exert both antitumour and tumour-promoting effects [107]. While senescence can suppress tumour growth by halting the proliferation of malignant cells, senescent cells often display enhanced resistance to apoptosis [108].
Metabolically, senescent cells retain mitochondrial activity but exhibit an increased glycolytic rate, resulting in elevated lactate production, which can contribute to tumour progression [109]. Lactate can modulate the senescence phenotype in lung cancer cell lines via Snail signalling [110], with a portion of this lactate originating from cancer-associated fibroblasts (CAFs), thereby supporting tumour growth and metastasis [111]. Conversely, lactate may also enable certain hepatocellular carcinoma cells to bypass or resist senescence, further promoting malignancy [112].
8. Effects Beyond the Microenvironment: Lactic Acid and Metastasis
The influence of elevated lactate concentrations within the tumour microenvironment extends beyond the primary site, contributing to metastatic dissemination [113]. Metastasis accounts for over 90% of cancer-related mortality and thus represents a major therapeutic challenge [114]. For metastasis to occur, cancer cells must undergo profound metabolic adaptations, with both lactate and pyruvate playing key roles in enhancing migratory and invasive capabilities [35].
In pancreatic cancer, increased lactate production—quantified by extracellular acidification rate (ECAR)—correlates positively with metastatic potential [115]. Conversely, highly metastatic melanomas display enhanced lactate uptake [116]. These findings, though apparently paradoxical, reflect the dual function of lactate in tumour biology. In breast cancer, activation of LDHA induces H3K18 lactylation and upregulates its own expression in a positive feedback loop, leading to increased lactate production, migration, and invasion [117]. In parallel, MCT1-dependent lactate transport and the reverse Warburg effect contribute to metastasis through lactylation of Rab7a [118].
As stated above, lactate also modulates the behaviour of stromal and immune components within the TME, thereby facilitating metastasis. It regulates T regulatory cell activity [100] and, in NSCLC patients, it has been shown that elevated LDHA expression is linked to increased lactate formation, CD8⁺ T-cell immune evasion, and enhanced metastatic progression [97]. Tumour-associated macrophages (TAMs) similarly drive tumour growth and metastatic spread [119,120]. Lactate secreted by cancer cells activates M2-like macrophages via GPR132 signalling, promoting migration and invasion through paracrine mechanisms [93]. Additionally, lactate induces PD-L1 expression in macrophages through activation of the NF-κB pathway, fostering formation of pre-metastatic niches [121].
Beyond immune modulation, lactate influences the extracellular matrix architecture critical for invasion. In prostate cancer, lactic acid stimulates collagen I production, enhancing cellular invasiveness [122]. Similarly, intracellular conversion of lactate to pyruvate promotes collagen deposition, which sustains cancer stem cell populations essential for metastatic outgrowth [123].
9. Clinical Implications
As reviewed above, lactate exerts multiple deleterious effects on tumour biology by promoting invasiveness, metastasis, and immune evasion. Consequently, therapeutic strategies aimed at limiting lactate accumulation within the tumour microenvironment have emerged as promising anticancer approaches [124].
One potential target is lactate dehydrogenase (LDH) [125], the key enzyme responsible for lactate synthesis. Inhibition of LDH has been shown to impede tumour progression [126], suggesting that interfering with lactate production may suppress tumour growth. However, subsequent studies proposed that the therapeutic benefit of LDH inhibition arises partly from increased glucose availability in the TME, which indirectly enhances T-cell function [127] or modulates immune activity through IL-21 signalling [128]. Although these studies noted reductions in intratumoral lactate, the specific contribution of lactate depletion to these effects has yet to be conclusively determined.
An alternative strategy focuses on disrupting lactate transport. Pharmacological inhibition of lactate export through monocarboxylate transporters (MCTs) has shown preclinical promise. The selective MCT1 inhibitor AZD3965 has advanced to phase I clinical trials, where it demonstrated good tolerability at doses sufficient to block lactate transport [129,130]. Further studies are needed to refine biomarkers for patient selection. Additionally, VB124—a recently developed MCT4-specific inhibitor—has shown efficacy in animal models [131,132]. Beyond monotherapy studies, preclinical data are the basis for trials assessing the combination of MCT inhibitors with other anticancer drugs such as immunotherapy agents.
Given that lactic acid accumulation is a common feature across diverse tumour types, its potential as a diagnostic and prognostic biomarker has been the focus of growing interest. Using 13C-hyperpolarization magnetic resonance spectroscopy, changes in TME lactate production can be detected within 24 hours after drug administration, often preceding conventional radiological indicators of therapeutic response [133].
Communication between the TME and the peripheral circulation may also allow tumour-derived metabolites to serve as minimally invasive biomarkers [134]. Although systemic lactate levels are tightly regulated [62], elevated plasma lactate could reflect tumour metabolic activity. Metabolomic profiling has revealed higher serum lactate levels in patients with non–small-cell lung cancer compared to healthy controls, with concentrations increasing alongside disease severity [135]. Retrospective analyses further support lactate’s prognostic value: in a study of 85 patients with stage IV NSCLC or small-cell lung cancer, plasma lactate correlated with metastatic burden even after adjusting for confounders such as intubation and ICU admission [136]. Elevated plasma lactate was also associated with reduced overall survival in lung cancer [136]. Similar findings have been reported in ovarian cancer [137], lymphoma [138], prostate cancer [109], and colorectal cancer, where lactate levels were significantly higher in metastatic versus non-metastatic patients [139]. In the case of breast cancer, lactate levels measured in whole tumour freshly excised samples correlated with tumour grade and Nottingham Prognostic Index but not with proliferation rate as measured by Ki67 expression [140]. On the other hand, a high correlation between Ki67 and lactate levels was found in gliomas [141]. Interestingly, lactate concentrations decline in multiple myeloma patients responding to therapy [142], suggesting that serum lactate could serve as a dynamic marker for treatment efficacy.
10. Concluding Remarks
Once regarded merely as a metabolic waste product, lactate is now recognized as a central player in cancer biology. Tumour cells can exploit lactate as an efficient energy source via mitochondrial oxidative phosphorylation (OXPHOS) and can induce stromal cells to enhance its production, fostering a metabolically supportive microenvironment. Beyond its metabolic role, lactic acid functions as a signalling molecule—acting through receptors such as GPR81 or via direct protein lactylation—to drive immunosuppression, promote metastasis, and modulate diverse aspects of tumour progression.
Therapeutic strategies aimed at reducing lactate abundance or blocking its downstream effects in the TME are gaining traction. Approaches include inhibiting lactate production, disrupting its transport, or targeting lactate-mediated signalling pathways. In parallel, lactate is being investigated as a prognostic biomarker and a potential indicator of treatment response, offering both mechanistic insights and clinical utility in oncology.
Author Contributions
Writing, reviewing and editing C.C. and I.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| TME | Tumour microenvironment |
| LDH | Lactate dehydrogenase |
| MCT | Monocarboxylate transporter |
| OXPHOS | Oxidative phosphorylation |
| CAF | Cancer associated fibroblast |
| TAM | Tumour associated macrophage |
| LIF | Leukemia inhibitory factor |
| NSCLC | Non-small cell lung cancer |
| ECAR | Extracellular acidification rate |
References
- Kompanje, E.J.O.; Jansen, T.C.; Van Der Hoven, B.; Bakker, J. The First Demonstration of Lactic Acid in Human Blood in Shock by Johann Joseph Scherer (1814-1869) in January 1843. Intensive Care Med 2007, 33, 1967–1971. [CrossRef]
- Katz, J.; Tayek, J.A. Gluconeogenesis and the Cori Cycle in 12-, 20-, and 40-h-Fasted Humans. Am. J. Physiol (Endocrinol Metab) 1998, 275, E537–E542.
- Perez-Ramirez, C.A.; Nakano, H.; Law, R.C.; Matulionis, N.; Thompson, J.; Pfeiffer, A.; Park, J.O.; Nakano, A.; Christofk, H.R. Atlas of Fetal Metabolism during Mid-to-Late Gestation and Diabetic Pregnancy. Cell 2024, 187, 204-215.e14. [CrossRef]
- Cori, C.F. Glycogen Bareakdown and Synthesis in Animal Tissues. Endocrinology 1940, 26, 285–296. [CrossRef]
- Rabinowitz, J.D.; Enerbäck, S. Lactate: The Ugly Duckling of Energy Metabolism. Nat Metab 2020, 2, 566–571. [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [CrossRef]
- Pavlova, N.N.; Thompson, C.B. The Emerging Hallmarks of Cancer Metabolism. Cell Metab 2016, 23, 27–47. [CrossRef]
- Barba, I.; Carrillo-Bosch, L.; Seoane, J. Targeting the Warburg Effect in Cancer: Where Do We Stand? Int J Mol Sci 2024, 25, 3142. [CrossRef]
- Walenta, S.; Wetterling, M.; Lehrke, M.; Schwickert, G.; Sundfor, K.; Rofstad, E.K.; Mueller-Klieser, W. High Lactate Levels Predict Likelihood of Metastases, Tumor Recurrence, and Restricted Patient Survival in Human Cervical Cancers. Cancer Res 2000, 60, 916–921.
- Walenta, S.; Mueller-Klieser, W.F. Lactate: Mirror and Motor of Tumor Malignancy. Semin Radiat Oncol 2004, 14, 267–274. [CrossRef]
- Autry, A.W.; Vaziri, S.; LaFontaine, M.; Gordon, J.W.; Chen, H.Y.; Kim, Y.; Villanueva-Meyer, J.E.; Molinaro, A.; Clarke, J.L.; Oberheim Bush, N.A.; et al. Multi-Parametric Hyperpolarized 13C/1H Imaging Reveals Warburg-Related Metabolic Dysfunction and Associated Regional Heterogeneity in High-Grade Human Gliomas. Neuroimage Clin 2023, 39, 103501. [CrossRef]
- Li, Q.; Zhang, D.; Sui, X.; Song, T.; Hu, L.; Xu, X.; Wang, X.; Wang, F. The Warburg Effect Drives Cachectic States in Patients with Pancreatobiliary Adenocarcinoma. FASEB Journal 2023, 37, e23144. [CrossRef]
- Woitek, R.; Brindle, K.M. Hyperpolarized Carbon-13 MRI in Breast Cancer. Diagnostics 2023, 13, 2311. [CrossRef]
- Guo, Y.; Meng, X.; Ma, J.; Zheng, Y.; Wang, Q.; Wang, Y.; Shang, H. Human Papillomavirus 16 E6 Contributes HIF-1α Induced Warburg Effect by Attenuating the VHL-HIF-1α Interaction. Int J Mol Sci 2014, 15, 7974–7986. [CrossRef]
- Anderson, N.M.; Simon, M.C. The Tumor Microenvironment. Current Biology 2020, 30, R921–R925. [CrossRef]
- Sadeghi Rad, H.; Monkman, J.; Warkiani, M.E.; Ladwa, R.; O’Byrne, K.; Rezaei, N.; Kulasinghe, A. Understanding the Tumor Microenvironment for Effective Immunotherapy. Med Res Rev 2021, 41, 1474–1498. [CrossRef]
- Liu, C.; Jin, Y.; Fan, Z. The Mechanism of Warburg Effect-Induced Chemoresistance in Cancer. Front Oncol 2021, 11, 698023. [CrossRef]
- Vaupel, P.; Multhoff, G. Revisiting the Warburg Effect: Historical Dogma versus Current Understanding. Journal of Physiology 2021, 599, 1745–1757. [CrossRef]
- Ohashi, T.; Aoki, M.; Tomita, H.; Akazawa, T.; Sato, K.; Kuze, B.; Mizuta, K.; Hara, A.; Nagaoka, H.; Inoue, N.; et al. M2-like Macrophage Polarization in High Lactic Acid-Producing Head and Neck Cancer. Cancer Sci 2017, 108, 1128–1134. [CrossRef]
- Urbańska, K.; Orzechowski, A. Unappreciated Role of LDHA and LDHB to Control Apoptosis and Autophagy in Tumor Cells. Int J Mol Sci 2019, 20, 2085. [CrossRef]
- Payen, V.L.; Mina, E.; Van Hée, V.F.; Porporato, P.E.; Sonveaux, P. Monocarboxylate Transporters in Cancer. Mol Metab 2020, 33, 48–66. [CrossRef]
- Contreras-Baeza, Y.; Sandoval, P.Y.; Alarcón, R.; Galaz, A.; Cortés-Molina, F.; Alegriá, K.; Baeza-Lehnert, F.; Arce-Molina, R.; Guequén, A.; Flores, C.A.; et al. Monocarboxylate Transporter 4 (MCT4) Is a High Affinity Transporter Capable of Exporting Lactate in High-Lactate Microenvironments. Journal of Biological Chemistry 2019, 294, 20135–20147. [CrossRef]
- Brizel, D.M.; Schroeder, T.; Scher, R.L.; Walenta, S.; Clough, R.W.; Dewhirst, M.W.; Mueller-Klieser, W. Elevated Tumor Lactate Concentrations Predict for an Increased Risk of Metastases in Head-and-Neck Cancer. Int. J. Radiation Oncology Biol. Phys. 2001, 51, 349–353.
- Benjamin, D.; Robay, D.; Hindupur, S.K.; Pohlmann, J.; Colombi, M.; El-Shemerly, M.Y.; Maira, S.M.; Moroni, C.; Lane, H.A.; Hall, M.N. Dual Inhibition of the Lactate Transporters MCT1 and MCT4 Is Synthetic Lethal with Metformin Due to NAD+ Depletion in Cancer Cells. Cell Rep 2018, 25, 3047-3058.e4. [CrossRef]
- Fuchs, A.L.; Schiller, S.M.; Keegan, W.J.; Ammons, M.C.B.; Eilers, B.; Tripet, B.; Copié, V. Quantitative 1H NMR Metabolomics Reveal Distinct Metabolic Adaptations in Human Macrophages Following Differential Activation. Metabolites 2019, 9. [CrossRef]
- Fiaschi, T.; Marini, A.; Giannoni, E.; Taddei, M.L.; Gandellini, P.; De Donatis, A.; Lanciotti, M.; Serni, S.; Cirri, P.; Chiarugi, P. Reciprocal Metabolic Reprogramming through Lactate Shuttle Coordinately Influences Tumor-Stroma Interplay. Cancer Res 2012, 79, 5130–5140. [CrossRef]
- Halestrap, A.P.; Wilson, M.C. The Monocarboxylate Transporter Family-Role and Regulation. IUBMB Life 2012, 64, 109–119. [CrossRef]
- Griffiths, J.R. Are Cancer Cells Acidic? Br. J. Cancer 1991, 64, 425–427.
- Gallagher, F.A.; Kettunen, M.I.; Day, S.E.; Hu, D.E.; Ardenkjær-Larsen, J.H.; In ’T Zandt, R.; Jensen, P.R.; Karlsson, M.; Golman, K.; Lerche, M.H.; et al. Magnetic Resonance Imaging of PH in Vivo Using Hyperpolarized 13C-Labelled Bicarbonate. Nature 2008, 453, 940–943. [CrossRef]
- Boedtkjer, E.; Pedersen, S.F. The Acidic Tumor Microenvironment as a Driver of Cancer. Annu Rev Physiol 2020, 82, 103–126. [CrossRef]
- Rolver, M.G.; Holland, L.K.K.; Ponniah, M.; Prasad, N.S.; Yao, J.; Schnipper, J.; Kramer, S.; Elingaard-Larsen, L.; Pedraz-Cuesta, E.; Liu, B.; et al. Chronic Acidosis Rewires Cancer Cell Metabolism through PPARα Signaling. Int J Cancer 2023, 152, 1668–1684. [CrossRef]
- Gründer, S.; Vanek, J.; Pissas, K.P. Acid-Sensing Ion Channels and Downstream Signalling in Cancer Cells: Is There a Mechanistic Link? Pflugers Arch 2024, 476, 659–672.
- Gupta, S.C.; Singh, R.; Pochampally, R.; Watabe, K.; Mo, Y.Y. Acidosis Promotes Invasiveness of Breast Cancer Cells through ROS-AKT-NF-ΚB Pathway. Oncotarget 2014, 5, 12070–12082. [CrossRef]
- Knopf, P.; Stowbur, D.; Hoffmann, S.H.L.; Hermann, N.; Maurer, A.; Bucher, V.; Poxleitner, M.; Tako, B.; Sonanini, D.; Krishnamachary, B.; et al. Acidosis-Mediated Increase in IFN-γ-Induced PD-L1 Expression on Cancer Cells as an Immune Escape Mechanism in Solid Tumors. Mol Cancer 2023, 22, 207. [CrossRef]
- Bergers, G.; Fendt, M. The Metabolism of Cancer Cells during Metastasis. Nat Rev Cancer 2021, 3, 162–180. [CrossRef]
- Hosonuma, M.; Yoshimura, K. Association between PH Regulation of the Tumor Microenvironment and Immunological State. Front Oncol 2023, 13, 1175563. [CrossRef]
- Zheng, Y.; Xu, R.; Chen, X.; Lu, Y.; Zheng, J.; Lin, Y.; Lin, P.; Zhao, X.; Cui, L. Metabolic Gatekeepers: Harnessing Tumor-Derived Metabolites to Optimize T Cell-Based Immunotherapy Efficacy in the Tumor Microenvironment. Cell Death Dis 2024, 15, 775. [CrossRef]
- Colegio, O.R.; Chu, N.-Q.; Szabo, A.L.; Chu, T.; Rhebergen, A.M.; Jairam, V.; Cyrus, N.; Brokowski, C.E.; Eisenbarth, S.C.; Phillips, G.M.; et al. Functional Polarization of Tumour-Associated Macrophages by Tumour-Derived Lactic Acid. Nature 2014, 513, 559–563. [CrossRef]
- El-Kenawi, A.; Gatenbee, C.; Robertson-Tessi, M.; Bravo, R.; Dhillon, J.; Balagurunathan, Y.; Berglund, A.; Visvakarma, N.; Ibrahim-Hashim, A.; Choi, J.; et al. Acidity Promotes Tumour Progression by Altering Macrophage Phenotype in Prostate Cancer. Br J Cancer 2019, 121, 556–566. [CrossRef]
- Fischer, K.; Hoffmann, P.; Voelkl, S.; Meidenbauer, N.; Ammer, J.; Edinger, M.; Gottfried, E.; Schwarz, S.; Rothe, G.; Hoves, S.; et al. Inhibitory Effect of Tumor Cell-Derived Lactic Acid on Human T Cells. Blood 2007, 109, 3812–3819. [CrossRef]
- Hirschhaeuser, F.; Sattler, U.G.A.; Mueller-Klieser, W. Lactate: A Metabolic Key Player in Cancer. Cancer Res 2011, 71, 6921–6925.
- Li, Y.; He, C.; Shen, A.N.; Wang, Y.; Xu, Z.P.; Zhang, L.; Wang, R. PH of Microenvironment Directly Modulates the Phenotype and Function of Cancer-Associated Fibroblasts. ACS Omega 2025, 10, 3937–3943. [CrossRef]
- Kim, Y.; Dube, S.E.; Park, C.B. Brain Energy Homeostasis: The Evolution of the Astrocyte-Neuron Lactate Shuttle Hypothesis. Korean Journal of Physiology and Pharmacology 2025, 29, 1–8.
- Nakajima, E.C.; Van Houten, B. Metabolic Symbiosis in Cancer: Refocusing the Warburg Lens. Mol Carcinog 2013, 52, 329–337. [CrossRef]
- Jayathilake, P.G.; Victori, P.; Pavillet, C.E.; Lee, C.H.; Voukantsis, D.; Miar, A.; Arora, A.; Harris, A.L.; Morten, K.J.; Buffa, F.M. Metabolic Symbiosis between Oxygenated and Hypoxic Tumour Cells: An Agent-Based Modelling Study. PLoS Comput Biol 2024, 20, e1011944. [CrossRef]
- Kennedy, K.M.; Scarbrough, P.M.; Ribeiro, A.; Richardson, R.; Yuan, H.; Sonveaux, P.; Landon, C.D.; Chi, J.T.; Pizzo, S.; Schroeder, T.; et al. Catabolism of Exogenous Lactate Reveals It as a Legitimate Metabolic Substrate in Breast Cancer. PLoS One 2013, 8, e75154. [CrossRef]
- Curry, J.M.; Tuluc, M.; Whitaker-Menezes, D.; Ames, J.A.; Anantharaman, A.; Butera, A.; Leiby, B.; Cognetti, D.M.; Sotgia, F.; Lisanti, M.P.; et al. Cancer Metabolism, Stemness and Tumor Recurrence: MCT1 and MCT4 Are Functional Biomarkers of Metabolic Symbiosis in Head and Neck Cancer. Cell Cycle 2013, 12, 1371–1384. [CrossRef]
- Faubert, B.; Li, K.Y.; Cai, L.; Hensley, C.T.; Kim, J.; Zacharias, L.G.; Yang, C.; Do, Q.N.; Doucette, S.; Burguete, D.; et al. Lactate Metabolism in Human Lung Tumors. Cell 2017, 171, 358-371.e9. [CrossRef]
- McCleland, M.L.; Adler, A.S.; Deming, L.; Cosino, E.; Lee, L.; Blackwood, E.M.; Solon, M.; Tao, J.; Li, L.; Shames, D.; et al. Lactate Dehydrogenase B Is Required for the Growth of KRAS-Dependent Lung Adenocarcinomas. Clinical Cancer Research 2013, 19, 773–784. [CrossRef]
- Ždralević, M.; Brand, A.; Ianni, L. Di; Dettmer, K.; Reinders, J.; Singer, K.; Peter, K.; Schnell, A.; Bruss, C.; Decking, S.M.; et al. Double Genetic Disruption of Lactate Dehydrogenases A and B Is Required to Ablate the “Warburg Effect” Restricting Tumor Growth to Oxidative Metabolism. Journal of Biological Chemistry 2018, 293, 15947–15961. [CrossRef]
- Doherty, J.R.; Cleveland, J.L. Targeting Lactate Metabolism for Cancer Therapeutics. Journal of Clinical Investigation 2013, 123, 3685–3692.
- Pavlides, S.; Whitaker-Menezes, D.; Castello-Cros, R.; Flomenberg, N.; Witkiewicz, A.K.; Frank, P.G.; Casimiro, M.C.; Wang, C.; Fortina, P.; Addya, S.; et al. The Reverse Warburg Effect: Aerobic Glycolysis in Cancer Associated Fibroblasts and the Tumor Stroma. Cell Cycle 2009, 8, 3984–4001. [CrossRef]
- Whitaker-Menezes, D.; Martinez-Outschoorn, U.E.; Lin, Z.; Ertel, A.; Flomenberg, N.; Witkiewicz, A.K.; Birbe, R.C.; Howell, A.; Pavlides, S.; Gandara, R.; et al. Evidence for a Stromal-Epithelial “Lactate Shuttle” in Human Tumors: MCT4 Is a Marker of Oxidative Stress in Cancer-Associated Fibroblasts. Cell Cycle 2011, 10, 1772–1783. [CrossRef]
- Affinito, A.; Quintavalle, C.; Chianese, R.V.; Roscigno, G.; Fiore, D.; D’Argenio, V.; Thomas, G.; Savarese, A.; Ingenito, F.; Cocca, L.; et al. MCT4-Driven CAF-Mediated Metabolic Reprogramming in Breast Cancer Microenvironment Is a Vulnerability Targetable by MiR-425-5p. Cell Death Discov 2024, 10, 140. [CrossRef]
- Pértega-Gomes, N.; Vizcaíno, J.R.; Attig, J.; Jurmeister, S.; Lopes, C.; Baltazar, F. A Lactate Shuttle System between Tumour and Stromal Cells Is Associated with Poor Prognosis in Prostate Cancer. BMC Cancer 2014, 14, 352. [CrossRef]
- Chen, P.; Geng, H.; Ma, B.; Zhang, Y.; Zhu, Z.; Li, M.; Chen, S.; Wang, X.; Sun, C. Integrating Spatial Omics and Single-Cell Mass Spectrometry Imaging Reveals Tumor–Host Metabolic Interplay in Hepatocellular Carcinoma. Proceedings of the National Academy of Sciences 2025, 122, e2505789122. [CrossRef]
- Certo, M.; Llibre, A.; Lee, W.; Mauro, C. Understanding Lactate Sensing and Signalling. Trends in Endocrinology and Metabolism 2022, 33, 722–735. [CrossRef]
- Mohammad Nezhady, M.A.; Modaresinejad, M.; Zia, A.; Chemtob, S. Versatile Lactate Signaling via HCAR1: A Multifaceted GPCR Involved in Many Biological Processes. Am J Physiol Cell Physiol 2023, 325, C1502–C1515. [CrossRef]
- Roland, C.L.; Arumugam, T.; Deng, D.; Liu, S.H.; Philip, B.; Gomez, S.; Burns, W.R.; Ramachandran, V.; Wang, H.; Cruz-Monserrate, Z.; et al. Cell Surface Lactate Receptor GPR81 Is Crucial for Cancer Cell Survival. Cancer Res 2014, 74, 5301–5310. [CrossRef]
- Jin, L.; Guo, Y.; Chen, J.; Wen, Z.; Jiang, Y.; Qian, J. Lactate Receptor HCAR1 Regulates Cell Growth, Metastasis and Maintenance of Cancerspecific Energy Metabolism in Breast Cancer Cells. Mol Med Rep 2022, 26, 268. [CrossRef]
- Ishihara, S.; Hata, K.; Hirose, K.; Okui, T.; Toyosawa, S.; Uzawa, N.; Nishimura, R.; Yoneda, T. The Lactate Sensor GPR81 Regulates Glycolysis and Tumor Growth of Breast Cancer. Sci Rep 2022, 12, 6261. [CrossRef]
- Lee, W.D.; Weilandt, D.R.; Liang, L.; MacArthur, M.R.; Jaiswal, N.; Ong, O.; Mann, C.G.; Chu, Q.; Hunter, C.J.; Ryseck, R.P.; et al. Lactate Homeostasis Is Maintained through Regulation of Glycolysis and Lipolysis. Cell Metab 2025, 37, 758–771. [CrossRef]
- Longhitano, L.; Vicario, N.; Tibullo, D.; Giallongo, C.; Broggi, G.; Caltabiano, R.; Barbagallo, G.M.V.; Altieri, R.; Baghini, M.; Di Rosa, M.; et al. Lactate Induces the Expressions of MCT1 and HCAR1 to Promote Tumor Growth and Progression in Glioblastoma. Front Oncol 2022, 12, 871798. [CrossRef]
- Payen, V.L.; Hsu, M.Y.; Rädecke, K.S.; Wyart, E.; Vazeille, T.; Bouzin, C.; Porporato, P.E.; Sonveaux, P. Monocarboxylate Transporter MCT1 Promotes Tumor Metastasis Independently of Its Activity as a Lactate Transporter. Cancer Res 2017, 77, 5591–5601. [CrossRef]
- Liu, S.; Zhao, H.; Hu, Y.; Yan, C.; Mi, Y.; Li, X.; Tao, D.; Qin, J. Lactate Promotes Metastasis of Normoxic Colorectal Cancer Stem Cells through PGC-1α-Mediated Oxidative Phosphorylation. Cell Death Dis 2022, 13, 651. [CrossRef]
- He, J.; Chai, X.; Zhang, Q.; Wang, Y.; Wang, Y.; Yang, X.; Wu, J.; Feng, B.; Sun, J.; Rui, W.; et al. The Lactate Receptor HCAR1 Drives the Recruitment of Immunosuppressive PMN-MDSCs in Colorectal Cancer. Nat Immunol 2025, 26, 391–403. [CrossRef]
- Brown, T.P.; Bhattacharjee, P.; Ramachandran, S.; Sivaprakasam, S.; Ristic, B.; Sikder, M.O.F.; Ganapathy, V. The Lactate Receptor GPR81 Promotes Breast Cancer Growth via a Paracrine Mechanism Involving Antigen-Presenting Cells in the Tumor Microenvironment. Oncogene 2020, 39, 3292–3304. [CrossRef]
- Yang, K.; Xu, J.; Fan, M.; Tu, F.; Wang, X.; Ha, T.; Williams, D.L.; Li, C. Lactate Suppresses Macrophage Pro-Inflammatory Response to LPS Stimulation by Inhibition of YAP and NF-ΚB Activation via GPR81-Mediated Signaling. Front Immunol 2020, 11. [CrossRef]
- Su, J.; Mao, X.; Wang, L.; Chen, Z.; Wang, W.; Zhao, C.; Li, G.; Guo, W.; Hu, Y. Lactate/GPR81 Recruits Regulatory T Cells by Modulating CX3CL1 to Promote Immune Resistance in a Highly Glycolytic Gastric Cancer. Oncoimmunology 2024, 13, 2320951. [CrossRef]
- Liu, X.; Li, S.; Cui, Q.; Guo, B.; Ding, W.; Liu, J.; Quan, L.; Li, X.; Xie, P.; Jin, L.; et al. Activation of GPR81 by Lactate Drives Tumour-Induced Cachexia. Nat Metab 2024, 6, 708–723. [CrossRef]
- Zhang, D.; Tang, Z.; Huang, H.; Zhou, G.; Cui, C.; Weng, Y.; Liu, W.; Kim, S.; Lee, S.; Perez-Neut, M.; et al. Metabolic Regulation of Gene Expression by Histone Lactylation. Nature 2019, 574, 575–580. [CrossRef]
- Shi, P.; Ma, Y.; Zhang, S. Non-Histone Lactylation: Unveiling Its Functional Significance. Front Cell Dev Biol 2025, 13, 1535611. [CrossRef]
- Yang, K.; Fan, M.; Wang, X.; Xu, J.; Wang, Y.; Tu, F.; Gill, P.S.; Ha, T.; Liu, L.; Williams, D.L.; et al. Lactate Promotes Macrophage HMGB1 Lactylation, Acetylation, and Exosomal Release in Polymicrobial Sepsis. Cell Death Differ 2022, 29, 133–146. [CrossRef]
- Fang, X.; Zhao, P.; Gao, S.; Liu, D.; Zhang, S.; Shan, M.; Wang, Y.; Herrmann, J.; Li, Q.; Wang, F. Lactate Induces Tumor-Associated Macrophage Polarization Independent of Mitochondrial Pyruvate Carrier-Mediated Metabolism. Int J Biol Macromol 2023, 237, 123810. [CrossRef]
- Noe, J.T.; Rendon, B.E.; Geller, A.E.; Conroy, L.R.; Morrissey, S.M.; Young, L.E.A.; Bruntz, R.C.; Kim, E.J.; Wise-Mitchell, A.; Barbosa De Souza Rizzo, M.; et al. Lactate Supports a Metabolic-Epigenetic Link in Macrophage Polarization. Sci. Adv 2021, 7, 8602.
- Zhou, S.; Xiao, L.; Hu, L.; Zuo, F.; Wang, Y.; Fei, B.; Dai, J.; Zhou, X. CAFs Promote Immune Evasion in Gastric Cancer through Histone Lactylation-Mediated Suppression of NCAPG Ubiquitination. J Transl Med 2025, 23, 989. [CrossRef]
- Lin, Z.; Long, F.; Liu, J.; Kang, R.; Klionsky, D.J.; Kroemer, G.; Tang, D.; Yang, M. Metabolic Reprogramming Promotes Apoptosis Resistance in Acute Lymphoblastic Leukemia through CASP3 Lactylation. Mol Cancer 2025, 24, 204. [CrossRef]
- Chen, B.; Deng, Y.; Hong, Y.; Fan, L.; Zhai, X.; Hu, H.; Yin, S.; Chen, Q.; Xie, X.; Ren, X.; et al. Metabolic Recoding of NSUN2-Mediated M5C Modification Promotes the Progression of Colorectal Cancer via the NSUN2/YBX1/M5C-ENO1 Positive Feedback Loop. Advanced Science 2024, 11, 2309840. [CrossRef]
- Tufail, M.; Jiang, C.H.; Li, N. Immune Evasion in Cancer: Mechanisms and Cutting-Edge Therapeutic Approaches. Signal Transduct Target Ther 2025, 10, 227. [CrossRef]
- Batlle, E.; Massagué, J. Transforming Growth Factor-β Signaling in Immunity and Cancer. Immunity 2019, 50, 924–940.
- Pascual-García, M.; Bonfill-Teixidor, E.; Planas-Rigol, E.; Rubio-Perez, C.; Iurlaro, R.; Arias, A.; Cuartas, I.; Sala-Hojman, A.; Escudero, L.; Martínez-Ricarte, F.; et al. LIF Regulates CXCL9 in Tumor-Associated Macrophages and Prevents CD8+ T Cell Tumor-Infiltration Impairing Anti-PD1 Therapy. Nat Commun 2019, 10, 2416. [CrossRef]
- Wegiel, B.; Vuerich, M.; Daneshmandi, S.; Seth, P. Metabolic Switch in the Tumor Microenvironment Determines Immune Responses to Anti-Cancer Therapy. Front Oncol 2018, 8, 1–9. [CrossRef]
- Caslin, H.L.; Abebayehu, D.; Pinette, J.A.; Ryan, J.J. Lactate Is a Metabolic Mediator That Shapes Immune Cell Fate and Function. Front Physiol 2021, 12, 688458. [CrossRef]
- Sangsuwan, R.; Thuamsang, B.; Pacifici, N.; Allen, R.; Han, H.; Miakicheva, S.; Lewis, J.S. Lactate Exposure Promotes Immunosuppressive Phenotypes in Innate Immune Cells. Cell Mol Bioeng 2020, 13, 541–557. [CrossRef]
- Morrot, A.; da Fonseca, L.M.; Salustiano, E.J.; Gentile, L.B.; Conde, L.; Filardy, A.A.; Franklim, T.N.; da Costa, K.M.; Freire-de-Lima, C.G.; Freire-de-Lima, L. Metabolic Symbiosis and Immunomodulation: How Tumor Cell-Derived Lactate May Disturb Innate and Adaptive Immune Responses. Front Oncol 2018, 8, 1–10. [CrossRef]
- Jin, R.; Neufeld, L.; McGaha, T.L. Linking Macrophage Metabolism to Function in the Tumor Microenvironment. Nat Cancer 2025, 6, 239–252. [CrossRef]
- Zhang, Q.; Wang, J.; Yadav, D.K.; Bai, X.; Liang, T. Glucose Metabolism: The Metabolic Signature of Tumor Associated Macrophage. Front Immunol 2021, 12, 702580. [CrossRef]
- Jin, X.; Zhang, N.; Yan, T.; Wei, J.; Hao, L.; Sun, C.; Zhao, H.; Jiang, S. Lactate-Mediated Metabolic Reprogramming of Tumor-Associated Macrophages: Implications for Tumor Progression and Therapeutic Potential. Front Immunol 2025, 16, 1573039. [CrossRef]
- Zhao, Y.; Wang, D.; Xu, T.; Liu, P.; Cao, Y.; Wang, Y.; Yang, X.; Xu, X.; Wang, X.; Niu, H. Bladder Cancer Cells Re-Educate TAMs through Lactate Shuttling in the Microfluidic Cancer Microenvironment. Oncotarget 2015, 6, 39196–39210.
- Mu, X.; Shi, W.; Xu, Y.; Xu, C.; Zhao, T.; Geng, B.; Yang, J.; Pan, J.; Hu, S.; Zhang, C.; et al. Tumor-Derived Lactate Induces M2 Macrophage Polarization via the Activation of the ERK/STAT3 Signaling Pathway in Breast Cancer. Cell Cycle 2018, 17, 428–438. [CrossRef]
- Zhang, C.; Cheng, W.; Yang, T.; Fang, H.; Zhang, R. Lactate Secreted by Esophageal Cancer Cells Induces M2 Macrophage Polarization via the AKT/ERK Pathway. Thorac Cancer 2023, 14, 2139–2148. [CrossRef]
- Chen, X.; Zhang, Z.; Wang, K. Lactate Released by Lung Adenocarcinoma (LUAD) Cells Promotes M2 Macrophage Polarization via the GPR132/CAMP/PKA Pathway. Genes Genomics 2025, 47, 521–531. [CrossRef]
- Chen, P.; Zuo, H.; Xiong, H.; Kolar, M.J.; Chu, Q.; Saghatelian, A.; Siegwart, D.J.; Wan, Y. Gpr132 Sensing of Lactate Mediates Tumor-Macrophage Interplay to Promote Breast Cancer Metastasis. Proc Natl Acad Sci U S A 2017, 114, 580–585. [CrossRef]
- Lin, Y.; Qi, Y.; Jiang, M.; Huang, W.; Li, B. Lactic Acid-Induced M2-like Macrophages Facilitate Tumor Cell Migration and Invasion via the GPNMB/CD44 Axis in Oral Squamous Cell Carcinoma. Int Immunopharmacol 2023, 124, 110972. [CrossRef]
- DeNardo, D.G.; Ruffell, B. Macrophages as Regulators of Tumour Immunity and Immunotherapy. Nat Rev Immunol 2019, 19, 369–382. [CrossRef]
- Watson, M.L.J.; Vignali, P.D.A.; Mullett, S.J.; Overacre-Delgoffe, A.E.; Peralta, R.M.; Grebinoski, S.; Menk, A. V.; Rittenhouse, N.L.; DePeaux, K.; Whetstone, R.D.; et al. Metabolic Support of Tumour-Infiltrating Regulatory T Cells by Lactic Acid. Nature 2021, 591, 645–651. [CrossRef]
- Wen, H.; Zhang, P.; Zhao, J.; Liu, Y.; Wan, L.; Li, H.; Yi, J.; Li, X. Metabolic Alterations Driven by LDHA in CD8 + T Cells Promote Immune Evasion and Therapy Resistance in NSCLC. Sci Rep 2025, 15, 24440. [CrossRef]
- Xia, H.; Wang, W.; Crespo, J.; Kryczek, I.; Li, W.; Wei, S.; Bian, Z.; Maj, T.; He, M.; Liu, R.J.; et al. Suppression of FIP200 and Autophagy by Tumor-Derived Lactate Promotes Naïve T Cell Apoptosis and Affects Tumor Immunity. Sci Immunol 2017, 17, 248–256. [CrossRef]
- Kumagai, S.; Koyama, S.; Itahashi, K.; Tanegashima, T.; Lin, Y. tzu; Togashi, Y.; Kamada, T.; Irie, T.; Okumura, G.; Kono, H.; et al. Lactic Acid Promotes PD-1 Expression in Regulatory T Cells in Highly Glycolytic Tumor Microenvironments. Cancer Cell 2022, 40, 201-218.e9. [CrossRef]
- Gu, J.; Zhou, J.; Chen, Q.; Xu, X.; Gao, J.; Li, X.; Shao, Q.; Zhou, B.; Zhou, H.; Wei, S.; et al. Tumor Metabolite Lactate Promotes Tumorigenesis by Modulating MOESIN Lactylation and Enhancing TGF-β Signaling in Regulatory T Cells. Cell Rep 2022, 39, 110986. [CrossRef]
- Ippolito, L.; Morandi, A.; Taddei, M.L.; Parri, M.; Comito, G.; Iscaro, A.; Raspollini, M.R.; Magherini, F.; Rapizzi, E.; Masquelier, J.; et al. Cancer-Associated Fibroblasts Promote Prostate Cancer Malignancy via Metabolic Rewiring and Mitochondrial Transfer. Oncogene 2019, 38, 5339–5355. [CrossRef]
- Kitamura, F.; Semba, T.; Yasuda-Yoshihara, N.; Yamada, K.; Nishimura, A.; Yamasaki, J.; Nagano, O.; Yasuda, T.; Yonemura, A.; Tong, Y.; et al. Cancer-Associated Fibroblasts Reuse Cancer-Derived Lactate to Maintain a Fibrotic and Immunosuppressive Microenvironment in Pancreatic Cancer. JCI Insight 2023, 8, e163022. [CrossRef]
- Gu, X.; Zhu, Y.; Su, J.; Wang, S.; Su, X.; Ding, X.; Jiang, L.; Fei, X.; Zhang, W. Lactate-Induced Activation of Tumor-Associated Fibroblasts and IL-8-Mediated Macrophage Recruitment Promote Lung Cancer Progression. Redox Biol 2024, 74, 103209. [CrossRef]
- Nasi, A.; Fekete, T.; Krishnamurthy, A.; Snowden, S.; Rajnavölgyi, E.; Catrina, A.I.; Wheelock, C.E.; Vivar, N.; Rethi, B. Dendritic Cell Reprogramming by Endogenously Produced Lactic Acid. The Journal of Immunology 2013, 191, 3090–3099. [CrossRef]
- Langin, D. Adipose Tissue Lipolysis Revisited (Again!): Lactate Involvement in Insulin Antilipolytic Action. Cell Metab 2010, 11, 242–243. [CrossRef]
- Hui, X.; Xue, M.; Ren, Y.; Chen, Y.; Chen, X.; Asad Farooq, M.; Ji, Y.; Zhan, W.; Huang, Y.; Du, B.; et al. GPR132 Regulates the Function of NK Cells through the Gαs/CSK/ZAP70/NF-ΚB Signaling Pathway as a Potential Immune Checkpoint. Sci. Adv 2025, 11, 9395.
- Schmitt, C.A.; Wang, B.; Demaria, M. Senescence and Cancer — Role and Therapeutic Opportunities. Nat Rev Clin Oncol 2022, 19, 619–636. [CrossRef]
- Colucci, M.; Sarill, M.; Maddalena, M.; Valdata, A.; Troiani, M.; Massarotti, M.; Bolis, M.; Bressan, S.; Kohl, A.; Robesti, D.; et al. Senescence in Cancer. Cancer Cell 2025, 43, 1204–1226. [CrossRef]
- Dou, X.; Fu, Q.; Long, Q.; Liu, S.; Zou, Y.; Fu, D.; Xu, Q.; Jiang, Z.; Ren, X.; Zhang, G.; et al. PDK4-Dependent Hypercatabolism and Lactate Production of Senescent Cells Promotes Cancer Malignancy. Nat Metab 2023, 5, 1887–1910. [CrossRef]
- Li, X.; Zhang, Z.; Zhang, Y.; Cao, Y.; Wei, H.; Wu, Z. Upregulation of Lactate-Inducible Snail Protein Suppresses Oncogene-Mediated Senescence through P16 INK4a Inactivation. Journal of Experimental and Clinical Cancer Research 2018, 37, 39. [CrossRef]
- Capparelli, C.; Guido, C.; Whitaker-Menezes, D.; Bonuccelli, G.; Balliet, R.; Pestell, T.G.; Goldberg, A.F.; Pestell, R.G.; Howell, A.; Sneddon, S.; et al. Autophagy and Senescence in Cancer-Associated Fibroblasts Metabolically Supports Tumor Growth and Metastasis, via Glycolysis and Ketone Production. Cell Cycle 2012, 11, 2285–2302. [CrossRef]
- Li, L.; Dong, J.; Xu, C.; Wang, S. Lactate Drives Senescence-Resistant Lineages in Hepatocellular Carcinoma via Histone H2B Lactylation of NDRG1. Cancer Lett 2025, 616. [CrossRef]
- Xing, W.; Li, X.; Zhou, Y.; Li, M.; Zhu, M. Lactate Metabolic Pathway Regulates Tumor Cell Metastasis and Its Use as a New Therapeutic Target. Explor Med 2023, 4, 541–549. [CrossRef]
- Ganesh, K.; Massagué, J. Targeting Metastatic Cancer. Nat Med 2021, 27, 34–44. [CrossRef]
- Wu, C.; Zheng, C.; Chen, S.; He, Z.; Hua, H.; Sun, C.; Yu, C. FOXQ1 Promotes Pancreatic Cancer Cell Proliferation, Tumor Stemness, Invasion and Metastasis through Regulation of LDHA-Mediated Aerobic Glycolysis. Cell Death Dis 2023, 14, 699. [CrossRef]
- Tasdogan, A.; Faubert, B.; Ramesh, V.; Ubellacker, J.M.; Shen, B.; Solmonson, A.; Murphy, M.M.; Gu, Z.; Gu, W.; Martin, M.; et al. Metabolic Heterogeneity Confers Differences in Melanoma Metastatic Potential. Nature 2020, 577, 115–120. [CrossRef]
- Hou, X.; Ouyang, J.; Tang, L.; Wu, P.; Deng, X.; Yan, Q.; Shi, L.; Fan, S.; Fan, C.; Guo, C.; et al. KCNK1 Promotes Proliferation and Metastasis of Breast Cancer Cells by Activating Lactate Dehydrogenase A (LDHA) and up-Regulating H3K18 Lactylation. PLoS Biol 2024, 22, e3002666. [CrossRef]
- Jiang, C.; He, X.; Chen, X.; Huang, J.; Liu, Y.; Zhang, J.; Chen, H.; Sui, X.; Lv, X.; Zhao, X.; et al. Lactate Accumulation Drives Hepatocellular Carcinoma Metastasis through Facilitating Tumor-Derived Exosome Biogenesis by Rab7A Lactylation. Cancer Lett 2025, 627, 217636. [CrossRef]
- Mantovani, A.; Marchesi, F.; Di Mitri, D.; Garlanda, C. Macrophage Diversity in Cancer Dissemination and Metastasis. Cell Mol Immunol 2024, 21, 1201–1214. [CrossRef]
- Lin, Y.; Xu, J.; Lan, H. Tumor-Associated Macrophages in Tumor Metastasis: Biological Roles and Clinical Therapeutic Applications. J Hematol Oncol 2019, 12, 76. [CrossRef]
- Morrissey, S.M.; Zhang, F.; Ding, C.; Montoya-Durango, D.E.; Hu, X.; Yang, C.; Wang, Z.; Yuan, F.; Fox, M.; Zhang, H. ge; et al. Tumor-Derived Exosomes Drive Immunosuppressive Macrophages in a Pre-Metastatic Niche through Glycolytic Dominant Metabolic Reprogramming. Cell Metab 2021, 33, 2040–2058. [CrossRef]
- Ippolito, L.; Duatti, A.; Iozzo, M.; Comito, G.; Pardella, E.; Lorito, N.; Bacci, M.; Pranzini, E.; Santi, A.; Sandrini, G.; et al. Lactate Supports Cell-Autonomous ECM Production to Sustain Metastatic Behavior in Prostate Cancer. EMBO Rep 2024, 25, 3506–3531. [CrossRef]
- Sun, Y.; Chen, Y.; Zhao, H.; Wang, J.; Liu, Y.; Bai, J.; Hu, C.; Shang, Z. Lactate-Driven Type I Collagen Deposition Facilitates Cancer Stem Cell-like Phenotype of Head and Neck Squamous Cell Carcinoma. iScience 2024, 27, 109340. [CrossRef]
- Wang, Z.; Gu, Z.; Mo, W.; Zhang, H. Lactate Metabolic Checkpoint in Immuno-Oncology: Mechanisms and Therapeutic Implications. Cancer Lett 2025, 633, 218038. [CrossRef]
- Wang, M.; Zhou, Q.; Cao, T.; Li, F.; Li, X.; Zhang, M.; Zhou, Y. Lactate Dehydrogenase A: A Potential New Target for Tumor Drug Resistance Intervention. J Transl Med 2025, 23, 713. [CrossRef]
- Le, A.; Cooper, C.R.; Gouw, A.M.; Dinavahi, R.; Maitra, A.; Deck, L.M.; Royer, R.E.; Vander Jagt, D.L.; Semenza, G.L.; Dang, C. V. Inhibition of Lactate Dehydrogenase A Induces Oxidative Stress and Inhibits Tumor Progression. Proc Natl Acad Sci U S A 2010, 107, 2037–2042. [CrossRef]
- Verma, S.; Budhu, S.; Serganova, I.; Dong, L.; Mangarin, L.M.; Khan, J.F.; Bah, M.A.; Assouvie, A.; Marouf, Y.; Schulze, I.; et al. Pharmacologic LDH Inhibition Redirects Intratumoral Glucose Uptake and Improves Antitumor Immunity in Solid Tumor Models. Journal of Clinical Investigation 2024, 134, e177606. [CrossRef]
- Hermans, D.; Gautam, S.; García-Cañaveras, J.C.; Gromer, D.; Mitra, S.; Spolski, R.; Li, P.; Christensen, S.; Nguyen, R.; Lin, J.-X.; et al. Lactate Dehydrogenase Inhibition Synergizes with IL-21 to Promote CD8+ T Cell Stemness and Antitumor Immunity. Proc Nat Acad Sci 2020, 117, 6047–6055. [CrossRef]
- Halford, S.; Veal, G.J.; Wedge, S.R.; Payne, G.S.; Bacon, C.M.; Sloan, P.; Dragoni, I.; Heinzmann, K.; Potter, S.; Salisbury, B.M. A Phase I Dose-Escalation Study of AZD3965, an Oral Monocarboxylate Transporter 1 Inhibitor, in Patients with Advanced Cancer. Clincial Cancer Research 2023, 29, 1429–1439. [CrossRef]
- Halford, S.E.R.; Walter, H.; McKay, P.; Townsend, W.; Linton, K.; Heinzmann, K.; Dragoni, I.; Brotherton, L.; Veal, G.; Siskos, A.; et al. Phase I Expansion Study of the First-in-Class Monocarboxylate Transporter 1 (MCT1) Inhibitor AZD3965 in Patients with Diffuse Large B-Cell Lymphoma (DLBCL) and Burkitt Lymphoma (BL). Journal of Clinical Oncology 2021, 39, 3115–3115. [CrossRef]
- Bonglack, E.N.; Messinger, J.E.; Cable, J.M.; Chng, J.; Mark Parnell, K.; Reinoso-Vizcaíno, N.M.; Barry, A.P.; Russell, V.S.; Dave, S.S.; Christofk, H.R.; et al. Monocarboxylate Transporter Antagonism Reveals Metabolic Vulnerabilities of Viral-Driven Lymphomas. Proc Natl Acad Sci U S A 2021, 118, 18–32. [CrossRef]
- Fang, Y.; Liu, W.; Tang, Z.; Ji, X.; Zhou, Y.; Song, S.; Tian, M.; Tao, C.; Huang, R.; Zhu, G.; et al. Monocarboxylate Transporter 4 Inhibition Potentiates Hepatocellular Carcinoma Immunotherapy through Enhancing T Cell Infiltration and Immune Attack. Hepatology 2023, 77, 109–123. [CrossRef]
- Day, S.E.; Kettunen, M.I.; Gallagher, F.A.; Hu, D.E.; Lerche, M.; Wolber, J.; Golman, K.; Ardenkjær-Larsen, J.H.; Brindle, K.M. Detecting Tumor Response to Treatment Using Hyperpolarized 13C Magnetic Resonance Imaging and Spectroscopy. Nat Med 2007, 13, 1382–1387. [CrossRef]
- Cao, Y.; Chang, T.; Schischlik, F.; Wang, K.; Sinha, S.; Hannenhalli, S.; Jiang, P.; Ruppin, E. Inferring Characteristics of the Tumor Immune Microenvironment of Patients with HNSCC from Single-Cell Transcriptomics of Peripheral Blood. Cancer Research Communications 2024, 4, 2335–2348. [CrossRef]
- Puchades-Carrasco, L.; Jantus-Lewintre, E.; Pérez-Rambla, C.; García-García, F.; Lucas, R.; Calabuig, S.; Blasco, A.; Dopazo, J.; Camps, C.; Pineda-Lucena, A. Serum Metabolomic Profiling Facilitates the Non-Invasive Identification of Metabolic Biomarkers Associated with the Onset and Progression of Non-Small Cell Lung Cancer. Oncotarget 2016, 7, 12904–12916.
- Vlachostergios, P.J.; Oikonomou, K.G.; Gibilaro, E.; Apergis, G. Elevated Lactic Acid Is a Negative Prognostic Factor in Metastatic Lung Cancer. Cancer Biomarkers 2015, 15, 725–734. [CrossRef]
- Kerslake, R.; Panfilov, S.; Mustafa, N.; Hall, M.; Kyrou, I.; Randeva, H.S.; Karteris, E.; Godfrey, R. Elevated Circulating Lactate Levels and Widespread Expression of Its Cognate Receptor, Hydroxycarboxylic Acid Receptor 1 (HCAR1), in Ovarian Cancer. J Clin Med 2023, 12, 217. [CrossRef]
- Atamna, B.; Rozental, A.; Haj Yahia, M.; Itchaki, G.; Gurion, R.; Yeshurun, M.; Raanani, P.; Wolach, O. Tumor-Associated Lactic Acidosis and Early Death in Patients With Lymphoma. Cancer Med 2025, 14, e70824. [CrossRef]
- Wei, Y.; Xu, H.; Dai, J.; Peng, J.; Wang, W.; Xia, L.; Zhou, F. Prognostic Significance of Serum Lactic Acid, Lactate Dehydrogenase, and Albumin Levels in Patients with Metastatic Colorectal Cancer. Biomed Res Int 2018, 2018, 1804086. [CrossRef]
- Cheung, S.M.; Husain, E.; Masannat, Y.; Miller, I.D.; Wahle, K.; Heys, S.D.; He, J. Lactate Concentration in Breast Cancer Using Advanced Magnetic Resonance Spectroscopy. Br J Cancer 2020, 123, 261–267. [CrossRef]
- Maldonado, F.; Fábregas, N.; Aldecoa, I.; González, J.; García-Orellana, M.; Belda, I.; Hurtado, P.; Gracia, I.; de Riva, N.; Tercero, J.; et al. Association between Preoperative Serum Lactate Concentrate with Tumor Cell Proliferative Index in Primary Brain Tumor. J Neurosurg Sci 2022, 66, 91–95. [CrossRef]
- Puchades-Carrasco, L.; Lecumberri, R.; Martínez-Ĺopez, J.; Lahuerta, J.J.; Mateos, M.V.; Prośper, F.; San-Miguel, J.F.; Pineda-Lucena, A. Multiple Myeloma Patients Have a Specific Serum Metabolomic Profile That Changes after Achieving Complete Remission. Clinical Cancer Research 2013, 19, 4770–4779. [CrossRef]
Figure 1.
Metabolism of lactic acid in the TME ( ). Glucose enters the cell via GLUT transporters where it is metabolized to pyruvate and lactate that is exported form the cell through MCT4 transporters together with protons. This leads to the accumulation of lactate and acidification of the extracellular medium. Stroma cells can also contribute to TME lactate accumulation. Lactic acid can also enter the cell through MCT1 transporter and can be incorporated into tumour cell OXPHOS metabolism, a process known as reverse Warburg effect. Lactic acid can induce signalling (
). Glucose enters the cell via GLUT transporters where it is metabolized to pyruvate and lactate that is exported form the cell through MCT4 transporters together with protons. This leads to the accumulation of lactate and acidification of the extracellular medium. Stroma cells can also contribute to TME lactate accumulation. Lactic acid can also enter the cell through MCT1 transporter and can be incorporated into tumour cell OXPHOS metabolism, a process known as reverse Warburg effect. Lactic acid can induce signalling ( ) mainly through lactate receptor GPR81 but also trough histone and other protein lactylation and MCT4 as a receptor. Lactic acid induced signalling pathways can affect tumoral and stromal cells where they tend to induce a pro-tumoral phenotype in immune cells.
) mainly through lactate receptor GPR81 but also trough histone and other protein lactylation and MCT4 as a receptor. Lactic acid induced signalling pathways can affect tumoral and stromal cells where they tend to induce a pro-tumoral phenotype in immune cells.
). Glucose enters the cell via GLUT transporters where it is metabolized to pyruvate and lactate that is exported form the cell through MCT4 transporters together with protons. This leads to the accumulation of lactate and acidification of the extracellular medium. Stroma cells can also contribute to TME lactate accumulation. Lactic acid can also enter the cell through MCT1 transporter and can be incorporated into tumour cell OXPHOS metabolism, a process known as reverse Warburg effect. Lactic acid can induce signalling () mainly through lactate receptor GPR81 but also trough histone and other protein lactylation and MCT4 as a receptor. Lactic acid induced signalling pathways can affect tumoral and stromal cells where they tend to induce a pro-tumoral phenotype in immune cells.
Figure 1.
Metabolism of lactic acid in the TME (). Glucose enters the cell via GLUT transporters where it is metabolized to pyruvate and lactate that is exported form the cell through MCT4 transporters together with protons. This leads to the accumulation of lactate and acidification of the extracellular medium. Stroma cells can also contribute to TME lactate accumulation. Lactic acid can also enter the cell through MCT1 transporter and can be incorporated into tumour cell OXPHOS metabolism, a process known as reverse Warburg effect. Lactic acid can induce signalling () mainly through lactate receptor GPR81 but also trough histone and other protein lactylation and MCT4 as a receptor. Lactic acid induced signalling pathways can affect tumoral and stromal cells where they tend to induce a pro-tumoral phenotype in immune cells.
). Glucose enters the cell via GLUT transporters where it is metabolized to pyruvate and lactate that is exported form the cell through MCT4 transporters together with protons. This leads to the accumulation of lactate and acidification of the extracellular medium. Stroma cells can also contribute to TME lactate accumulation. Lactic acid can also enter the cell through MCT1 transporter and can be incorporated into tumour cell OXPHOS metabolism, a process known as reverse Warburg effect. Lactic acid can induce signalling () mainly through lactate receptor GPR81 but also trough histone and other protein lactylation and MCT4 as a receptor. Lactic acid induced signalling pathways can affect tumoral and stromal cells where they tend to induce a pro-tumoral phenotype in immune cells.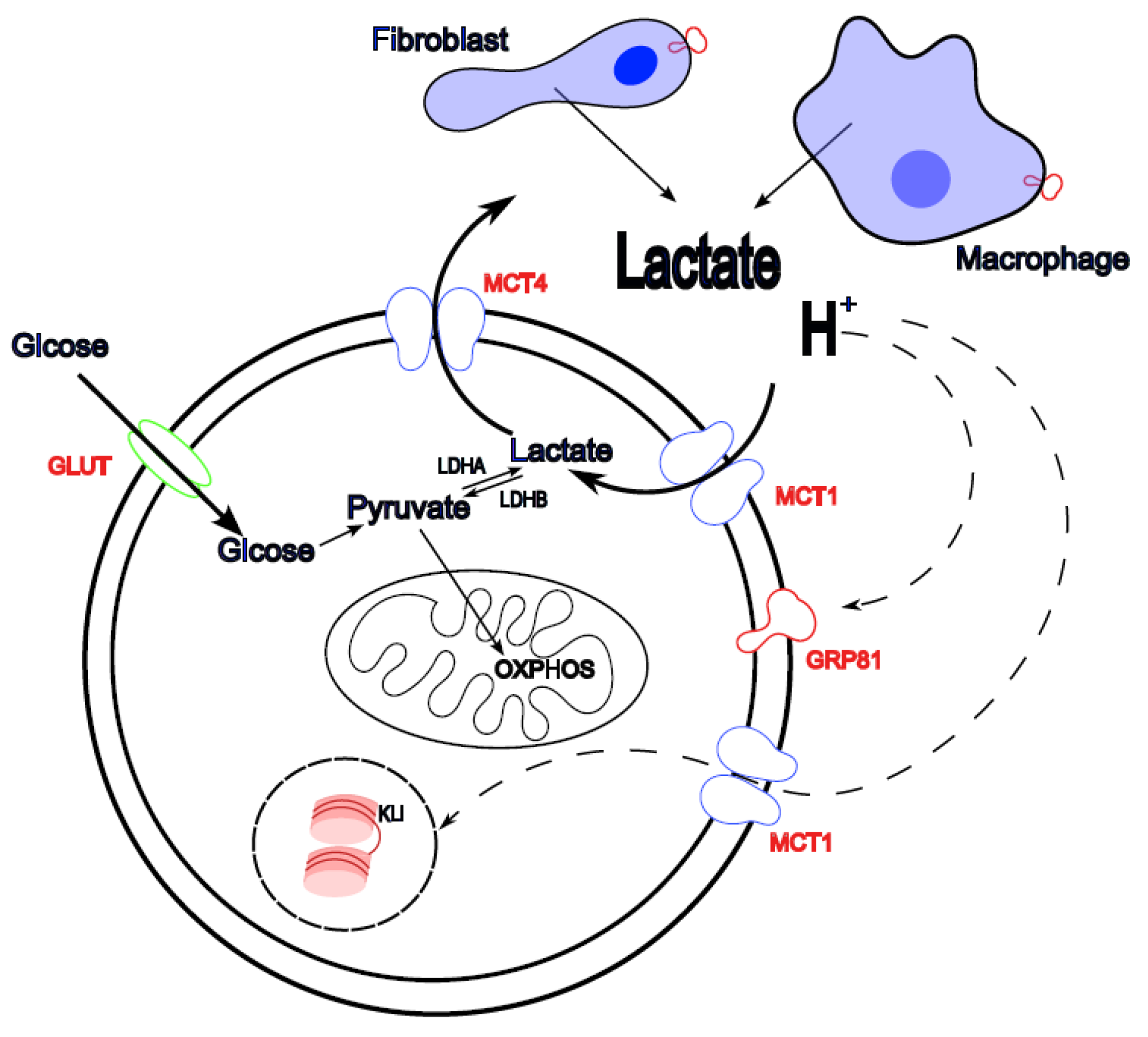
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



