
Submitted:
29 December 2025
Posted:
30 December 2025
You are already at the latest version
Abstract
In 2015 a polymorphic variant of Bactericidal/permeability-increasing protein (BPI) Fold containing Family B Member 4 (BPIFB4) was genetically associated with longevity. Following on from this intriguing observation, a literature has developed that suggests that this poorly characterised secreted protein, plays a pleiotropic role in maintaining human health and extending lifespan. In this article we briefly review what is known about BPIFB4 and discuss how its sites of expression may impact on these proposed functions.
Keywords:
BPIFB4
; longevity
; health lifespan
; gene expression
; protein localisation
; olfactory epithelium
; nasal
Perspectives
Genetics play a clear role in determination of lifespan and understanding the genetic components of healthy longevity will be important in improving quality of life and combating chronic diseases.
Variations in BPIFB4, a gene initially identified in the nasal cavity, have been associated with longevity and a number of studies have implicated the secreted protein with functions that may contribute to healthy longevity.
We review the distribution of mammalian BPIFB4 and propose that its restricted distribution may not allow this protein to perform the pleiotropic roles that have recently been ascribed to it. Instead we believe that the true function of BPIFB4 will reside in the nasal cavity and remains unresolved.
Introduction
BPIFB4 is a member of the BPI/LBP/PLUNC family of secreted proteins [1,2]. Members of the family contain either one, (BPIFAn), or two (BPIFBn) structural domains that were first identified in BPI and Lipopolysaccharide binding protein (LBP), key host defence lipid transfer proteins. Similar domains are also found in cholesteryl ester transfer protein (CETP) and phospholipid transfer protein (PLTP). These domains form a twisted, anti-parallel beta-sheet barrel that has the ability to bind lipids. Subsequently the BPI fold containing proteins were assigned to the larger Tubular Lipid Binding Protein (TULIP) family [3]. Although the functions of BPI, LBP, CETP and PLTP are well recognised, the function of members of the BPIF branch of the protein family remains unclear, despite the original suggestion that they function in host defence [1].
The BPIF protein family has been described in the past three decades, with members of the family predominantly being found in the respiratory, nasopharyngeal, and oral epitheliums [4]. BPIF genes are rapidly evolving and are highly divergent between species. Human BPIF genes are located on a gene locus on chromosome 20q11, and comprise eight functional genes along with four pseudogenes (Figure 1A). The mouse locus on chromosome 2 contains thirteen functional members and a single pseudogene. All mammals have single loci containing differing numbers of genes with the greatest diversity being seen in the BPIFA subfamily [5].
BPIFs generally show low protein homology between both orthologs and paralogs, with for example, human and mouse BPIFB1 having around 50% identity [6]. However, BPIFs maintain a conserved intron/exon structure and similar protein length. BPIFAs generally have 8-9 exons and encode proteins between 220-270 amino acids whereas BPIFBs generally have 15-16 exons and encode proteins between 450-500 amino acids in length. The shorter BPIF proteins are homologous to the N-terminal BPI (BPI1) domain, whilst the BPIFB proteins are homologous to both the N- and C-terminal BPI (BPI2) domains [6]. It has been suggested that a loss of the C-terminal domain of a BPIFB gene gave rise to the BPIFA gene branch early in mammalian evolution [6]. The “complete” BPIF family (composed of BPIFA and BPIFB genes) is found in placental mammals, whereas some BPIFB genes (including BPIFB4) are found in birds and amphibians [7,8].
Background on BPIFB4
BPIFB4 (as Ry2g5) was initially identified by subtractive hybridization of genes highly-expressed in rat olfactory mucosa and in situ hybridization showed that the gene was localised to the Bowman’s glands [9]. It was later shown that BPIFB4 was expressed in human nasal mucosa [1], and murine olfactory epithelium (OE) [10]. Consistent with this localised expression, BPIFB4 has been identified in olfactory cleft mucus [11]. BPIFB4 exhibits some significant differences in both sequence conservation and genomic organisation when compared to other BPIFB genes. Principally, the gene includes two exons towards the 5’ end that are significantly different to those of other family members. One is much shorter than that seen in the other BPIF genes and there is also the inclusion of large exon (508bp in the human gene) that encodes a peptide sequence that is unique to BPIFB4 (Figure 1B), resulting in the production of a protein that is 614 amino acids long. This sequence is inserted into the BPI1 domain and is predicted to form unstructured loops that are outside of the well-defined BPIF structural fold [6] (Figure 1C). The functional significance of this difference remains unclear. The other striking difference between BPIFB4 and other BPIFB proteins is the very high level of sequence similarity between orthologs. For example, the pairwise identity between human and mouse proteins is 85% with the highest level of similarity seen in the BPI1 domain. These two observations identify BPIFB4 as being somewhat different to other BPIF family members.
A Role for BPIFB4 in Longevity and Human Health?
BPIFB4 was essentially unstudied until a paper was published in 2015 suggesting that the protein may function to regulate longevity [12]. This suggestion arose following the validation of a common non-synonymous polymorphic variant in BPIFB4 (rs20700325, V268I), identified in a genome wide association study in long lived individuals (LLIs) in Italy, as being associated with longevity in two independent cohorts from Germany and the USA [12]. This SNP, located the N-terminal BPI1 domain was associated with three additional common non-synonymous variants (rs2889732, T320N; rs11699009, F527L and rs11696307, T533I) (Figure 1B) and generates a haplotype that the authors called a longevity associated variant (LAV). These SNPs have not been identified in other cohort studies,[13]. In the original paper the authors provide some biochemical evidence from in vivo and in vitro studies, for the action of the LAV in modulating endothelial function and angiogenesis [12]. The same group reported that serum BPIFB4 levels were elevated in LLIs and could be used to stratify their health status [14]. Since that time a literature has developed that appears to suggest that BPIFB4 plays a pleiotropic role in regulating human cardiovascular health, frailty and longevity (reviewed by Montella et al 2021 [15]). Some of this has focused on therapeutic effects of LAV-BPIFB4, with the hypothesis that gene therapy of the LAV could promote longevity. For example, overexpression of LAV was shown to prevent progression of frailty in old mice [16], prevents type 2 diabetes complications in mice [17], and protects against cardiac ischaemia in mice [18]. Endogenous BPIFB4 staining has also been shown to be reduced in tissue from ischemic heart failure compared to non-diseased tissue [19]. In addition, it has also been shown to be protective in a mouse model of Huntington’s disease [20]. Studies have also linked BPIFB4 with Chronic Obstructive Pulmonary Disease (COPD) [21,22] and nonsteroidal anti-inflammatory drug (NSAID)-induced enteropathy [23]. High circulating levels of BPIFB4 in LLIs may be associated with an anti-inflammatory myeloid cell profile [24]. The majority of this data has been published by a related research group
Although this and other data is intriguing perhaps it is important to revisit what is known about the localisation of BPIFB4 to see if there is a rational basis for these suggested roles in the maintenance of wellbeing.
Simply put, does localisation of BPIFB4 support a potential role in longevity and lifespan?
Expression and Localisation of BPIFB4
As outlined above Bpifb4, was identified as being differentially expressed in rat OE [9] and was subsequently shown to be expressed in the human and mouse nasal mucosa [1,10]. The protein was also identified in human nasal cleft mucus by proteomics [11] but has not been localised in these regions at the protein level. Unbiased transcriptional analysis of human OE showed that BPIFB4, as well as BPIFB3 and BPIFB6 were amongst the most tissue enriched and highly expressed genes when compared against 17 other tissues, including blood and heart [25]. A second transcriptional study showed that BPIFB4 was the second most highly expressed transcript in human OE and very highly expressed in the OE of multiple additional species, including macaques, dogs, rats and mice [26]. Although this data confirms that BPIFB4 is highly expressed in the nose, the olfactory regions are not well studied compared with most other tissues and so direct comparisons of expression levels between tissues cannot easily be made. For example, the Human Protein Atlas (HPA) [27] contains no olfactory tissues. The same is true for single cell RNA sequencing (scRNAseq) data in the Chan Zuckerberg CELLxGENE data repository [28] which contains no specific data from the human OE. However, these databases do enable assessment of BPIFB4 expression in a very wide range of other tissues.
So Where Is BPIFB4 Expressed in This Data?
In bulk RNAseq data in the HPA, BPIFB4 is shown to be expressed at low levels in pituitary (8.2 normalised transcripts per million (nTPM)) and <1 nTPM in parathyroid, choroid plexus, salivary gland, testes, heart muscle, and adipose tissue. scRNAseq data in the HPA show low expression of BPIFB4 in oocytes (3.7 nTPM) and respiratory epithelial cells (<2.6 nTPM) but three distinct sets in the HPA show no expression of BPIFB4 in immune cells.
scRNAseq and single nuclei (sn)RNAseq offers unparalleled power to detect gene expression in individual cells and have transformed our understanding to tissue and cell-type specific gene expression. CELLxGENE contains expression data from over 90 million single cells from over 60 tissues and represents the largest repository of scRNAseq data [28]. If BPIFB4 is abundantly expressed in tissue/cell types represented in the data sets then this should be clear to see.
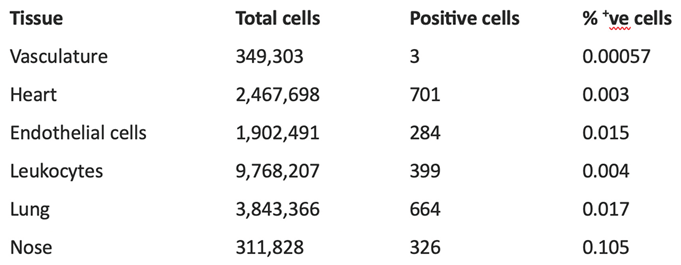
In CELLxGENE, there is very little evidence of BPIFB4 being expressed above background levels. In cells where BPIFB4 has been suggested by the recent literature to be functional/expressed, less than 0.02% of cells express the gene (Table 1). In the vasculature BPIFB4 is expressed in 3 of 349,303 cells, in endothelial cells 284 of >1.9 million cells are positive and in heart tissue, which has been suggested to contain abundant levels of BPIFB4 [17,19], 701 of >2.4 million cells express the gene. Less than 0.004% of total leukocytes are positive and even when these are broken down into distinct cell lineages no specific leukocyte cell population is shown to express the gene. In nose tissue (which may contain some OE) 0.1% of total cells are positive. When numbers of cells expressing a gene are so low in this type of data, it is difficult to be confident that the gene is truly expressed. It remains a formal possibility that BPIFB4 expression is both context and disease responsive and conditions that induce expression of the gene are not captured in these databases. However, the gene has not been noted to be differentially expressed in experimental studies, for example it was not found to be differentially expressed in scRNAseq data from individuals with heart failure [29].
Although CELLxGENE contains no OE tissues, a number of scRNAseq data sets have been generated from nasal and olfactory tissue that show BPIFB4 expression. In data from cells isolated from the human olfactory cleft, BPIFB4 is enriched in Bowman’s gland cells [30]. Bpifb4 is also highly enriched in Bowman’s gland ductular cells from the mouse OE [31]. More recently snRNAseq data from porcine OE also identified BPIFB4 in Bowman’s gland cells [32]. This data is all consistent with the first paper describing localisation of rat Bpifb4 [9].
At the protein level, the HPA does not display any localisation data for BPIFB4 and there are no robustly validated commercial antibodies for the protein. BPIFB4 contains a signal peptide (Figure 1B) and is predicted to be secreted. Does the protein, at approximately 65kDa enter the circulation? Although BPIFB4 has been reported in serum, with levels determined by ELISA [14], it is only found in one of 158 data sets in the human plasma peptide atlas [33]. It is also not present in heart, blood vessel or blood cell data sets in the Peptide Atlas [34], which shows only 20 of >800 studies contain BPIFB4 (Figure 2). Proteomics studies have shown however, that BPIFB4 is present in human nasal tissue [35,36], nasal cleft mucus [11,37], and nasal lavage [38]. It has also been identified in nasal secretions in mice [39]. So, available proteomic data also supports the view that BPIFB4 is predominantly found in the nasal/olfactory regions.
Closing Thoughts
Although some population studies have demonstrated that there is a correlation between LAV-BPIFB4 and longevity and there is an emerging literature suggesting that BPIFB4 plays a role in maintaining health, it remains unclear if the protein is expressed in the appropriate places to function in this manner. Objective analysis of existing data supports the contention that BPIFB4 is highly expressed in the nasal and OE and is not highly expressed in the vasculature, the heart and in leucocytes. Even though absence of evidence is not evidence of absence, it seems unlikely that BPIFB4 is highly expressed or localised in these tissues and therefore it calls into question some of the proposed functional roles of BPIFB4. We believe that the true function of BPIFB4 remains to be identified but will likely be associated with the OE. This will require further study.
Author Contributions
Conceptulization: CDB. Supervision: CDB. Investigation: LM, OB. Formal analysis: CDB. Validation: DB. Writing - original draft: LM, OB. Writing - review and editing: CDB
Funding
This research received no external funding. LM and OB are current MBChB students at the University of Sheffield School of Medicine and Population Health.
Data Availability Statement
Peptide Atlas: https://peptideatlas.org/. The Human Protein Atlas: https://www.proteinatlas.org/. CellXGene, Gene Expression application: https://cellxgene.cziscience.com/gene-expression
Acknowledgements
The authors thank Professors Sarah Teichmann and Kory Lavine for discussion of heart scRNAseq data, and Professor Bradly Goldstein for discussion of BPIF genes in cells of the human OE. They also wish to thank Dr Joris Deelen for his thoughts about the role of BPIFB4 variants in longevity.
Conflict of Interest
The authors report no conflicts of interest
References
- Bingle, CD; Craven, CJ. PLUNC: a novel family of candidate host defence proteins expressed in the upper airways and nasopharynx. Hum Mol Genet. 2002, 11, 937–43. [Google Scholar] [CrossRef]
- Bingle, CD; Seal, RL; Craven, CJ. Systematic nomenclature for the PLUNC/PSP/BSP30/SMGB proteins as a subfamily of the BPI fold-containing superfamily. Biochem Soc Trans. 2011, 39, 977–83. [Google Scholar] [CrossRef]
- Alva, V; Lupas, AN. The TULIP superfamily of eukaryotic lipid-binding proteins as a mediator of lipid sensing and transport. Biochim Biophys Acta 2016, 1861, 913–923. [Google Scholar] [CrossRef]
- Bingle, L; Bingle, CD. Distribution of human PLUNC/BPI fold-containing (BPIF) proteins. Biochem Soc Trans. 2011, 39, 1023–7. [Google Scholar] [CrossRef] [PubMed]
- Bingle, CD; Bingle, L; Craven, CJ. Distant cousins: genomic and sequence diversity within the BPI fold-containing (BPIF)/PLUNC protein family. Biochem Soc Trans. 2011, 39, 961–5. [Google Scholar] [CrossRef]
- Bingle, CD; LeClair, EE; Havard, S; Bingle, L; Gillingham, P; Craven, CJ. Phylogenetic and evolutionary analysis of the PLUNC gene family. Protein Sci. 2004, 13, 422–30. [Google Scholar] [CrossRef]
- Chiang, SC; Veldhuizen, EJ; Barnes, FA; Craven, CJ; Haagsman, HP; Bingle, CD. Identification and characterisation of the BPI/LBP/PLUNC-like gene repertoire in chickens reveals the absence of a LBP gene. Dev Comp Immunol. 2011, 35, 285–95. [Google Scholar] [CrossRef]
- Le Roy, N; Stapane, L; Gautron, J; Hincke, MT. Evolution of the Avian Eggshell Biomineralization Protein Toolkit - New Insights From Multi-Omics. Front Genet. 2021, 12, 672433. [Google Scholar] [CrossRef] [PubMed]
- Dear, TN; Boehm, T; Keverne, EB; Rabbitts, TH. Novel genes for potential ligand-binding proteins in subregions of the olfactory mucosa. EMBO J. 1991, 10, 2813–9. [Google Scholar] [CrossRef]
- Andrault, JB; Gaillard, I; Giorgi, D; Rouquier, S. Expansion of the BPI family by duplication on human chromosome 20: characterization of the RY gene cluster in 20q11.21 encoding olfactory transporters/antimicrobial-like peptides. Genomics 2003, 82, 172–84. [Google Scholar] [CrossRef] [PubMed]
- Débat, H; Eloit, C; Blon, F; Sarazin, B; Henry, C; Huet, JC; et al. Identification of human olfactory cleft mucus proteins using proteomic analysis. J Proteome Res. 2007, 6, 1985–96. [Google Scholar] [CrossRef]
- Villa, F; Carrizzo, A; Spinelli, CC; Ferrario, A; Malovini, A; Maciąg, A; Damato, A; et al. Genetic Analysis Reveals a Longevity-Associated Protein Modulating Endothelial Function and Angiogenesis. Circ Res. 2015, 117, 333–45. [Google Scholar] [CrossRef]
- Smulders, L; Deelen, J. Genetics of human longevity: From variants to genes to pathways. J Intern Med. 2024, 295, 416–435. [Google Scholar] [CrossRef] [PubMed]
- Villa, F; Malovini, A; Carrizzo, A; Spinelli, CC; Ferrario, A; Maciąg, A; et al. Serum BPIFB4 levels classify health status in long-living individuals. Immun Ageing 2015, 12, 27. [Google Scholar] [CrossRef] [PubMed]
- Montella, F; Lopardo, V; Cattaneo, M; Carrizzo, A; Vecchione, C; Ciaglia, E; et al. The Role of BPIFB4 in Immune System and Cardiovascular Disease: The Lesson from Centenarians. Transl Med UniSa 2021, 24, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Malavolta, M; Dato, S; Villa, F; Rango, F; Iannone, F; Ferrario, A; et al. LAV-BPIFB4 associates with reduced frailty in humans and its transfer prevents frailty progression in old mice. Aging (Albany NY) Erratum in: Aging (Albany NY). 2019, 11, 9220. doi: 10.18632/aging.102398. 2019, 11, 6555–6568. [Google Scholar] [CrossRef]
- Dang, Z; Avolio, E; Thomas, AC; Faulkner, A; Beltrami, AP; Cervellin, C; et al. Transfer of a human gene variant associated with exceptional longevity improves cardiac function in obese type 2 diabetic mice through induction of the SDF-1/CXCR4 signalling pathway. Eur J Heart Fail. 2020, 22, 1568–1581. [Google Scholar] [CrossRef]
- Cattaneo, M; Aleksova, A; Malovini, A; Avolio, E; Thomas, A; Alvino, VV; et al. BPIFB4 and its longevity-associated haplotype protect from cardiac ischemia in humans and mice. Cell Death Dis. 2023, 14, 523. [Google Scholar] [CrossRef]
- Cattaneo, M; Beltrami, AP; Thomas, AC; Spinetti, G; Alvino, VV; Avolio, E; et al. The longevity-associated BPIFB4 gene supports cardiac function and vascularization in ageing cardiomyopathy. Cardiovasc Res. 2023, 119, 1583–1595. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, M; Maciag, A; Milella, MS; Ciaglia, E; Bruno, A; Puca, AA. Longevity-Associated Variant of BPIFB4 Confers Neuroprotection in the STHdh Cell Model of Huntington Disease. Int J Mol Sci. 2022, 23, 15313. [Google Scholar] [CrossRef]
- Xue, L; Xu, J; Gong, H; Abudureheman, Z; ALimu, A; Chen, Y; et al. Overexpression of BPIFB4 Alleviates COPD Inflammatory Damage by Inhibiting M1 Macrophage Activation via the PI3K/AKT Pathway. Lung 2025, 203, 69. [Google Scholar] [CrossRef]
- Xu, J; Tao, L; Shi, Y; Gong, H; Abudureheman, Z; Zheng, A; et al. Sci Rep. 2025, 15, 14515. [CrossRef]
- Fujimori, S; Fukunaga, K; Takahashi, A; Mushiroda, T; Kubo, M; Hanada, R; et al. Bactericidal/Permeability-Increasing Fold-Containing Family B Member 4 May Be Associated with NSAID-Induced Enteropathy. Dig Dis Sci. 2019, 64, 401–408. [Google Scholar] [CrossRef]
- Ciaglia, E; Montella, F; Lopardo, V; Scala, P; Ferrario, A; Cattaneo, M; et al. Circulating BPIFB4 Levels Associate With and Influence the Abundance of Reparative Monocytes and Macrophages in Long Living Individuals. Front Immunol. 2020, 11, 1034. [Google Scholar] [CrossRef] [PubMed]
- Olender, T; Keydar, I; Pinto, JM; Tatarskyy, P; Alkelai, A; Chien, MS; Fishilevich, S; Restrepo, D; Matsunami, H; Gilad, Y; Lancet, D. The human olfactory transcriptome. BMC Genomics 2016, 17, 619. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Saraiva, LR; Riveros-McKay, F; Mezzavilla, M; Abou-Moussa, EH; Arayata, CJ; Makhlouf, M; et al. A transcriptomic atlas of mammalian olfactory mucosae reveals an evolutionary influence on food odor detection in humans. Sci Adv. 2019, 5, eaax0396. [Google Scholar] [CrossRef]
- Uhlén, M; Fagerberg, L; Hallström, BM; Lindskog, C; Oksvold, P; Mardinoglu, A. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Abdulla, S; Aevermann, B; Assis, P; Badajoz, S; Bell, SM; et al.; CZI Cell Science Program CZ CELLxGENE Discover: a single-cell data platform for scalable exploration, analysis and modeling of aggregated data. Nucleic Acids Res. 2025, 53, D886–D900. [Google Scholar] [CrossRef]
- Koenig, AL; Shchukina, I; Amrute, J; Andhey, PS; Zaitsev, K; Lai, L; et al. Single-cell transcriptomics reveals cell-type-specific diversification in human heart failure. Nat Cardiovasc Res. 2022, 1, 263–280. [Google Scholar] [CrossRef]
- Durante, MA; Kurtenbach, S; Sargi, ZB; Harbour, JW; Choi, R; Kurtenbach, S; et al. Single-cell analysis of olfactory neurogenesis and differentiation in adult humans. Nat Neurosci. 2020, 23, 323–326. [Google Scholar] [CrossRef] [PubMed]
- Ualiyeva, S; Lemire, E; Wong, C; Perniss, A; Boyd, AA; Avilés, EC; et al. A nasal cell atlas reveals heterogeneity of tuft cells and their role in directing olfactory stem cell proliferation. Sci Immunol. 2024, 9, eabq4341. [Google Scholar] [CrossRef]
- Caballero, I; Mbouamboua, Y; Weise, S; López-Gálvez, R; Couralet, M; Fleurot, I; et al. Cystic fibrosis alters the structure of the olfactory epithelium and the expression of olfactory receptors affecting odor perception. Sci Adv. 2025, 11, eads1568. [Google Scholar] [CrossRef]
- Pernemalm, M; Palaniappan, KK; Letunica, N; Vandenbrouck, Y; Brun, V; Tao, SC; et al. Advances and Utility of the Human Plasma Proteome. J Proteome Res. 2021, 20, 5241–5263. [Google Scholar] [CrossRef]
- Omenn, GS; Lane, L; Overall, CM; Pineau, C; Packer, NH; Cristea, IM; et al. The 2022 Report on the Human Proteome from the HUPO Human Proteome Project. J Proteome Res. 2023, 22, 1024–1042. [Google Scholar] [CrossRef]
- Ndika, J; Airaksinen, L; Suojalehto, H; Karisola, P; Fyhrquist, N; Puustinen, A; et al. Epithelial proteome profiling suggests the essential role of interferon-inducible proteins in patients with allergic rhinitis. J Allergy Clin Immunol. 2017, 140, 1288–1298. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H; Jeong, JE; Lee, HK; Yun, KN; An, HJ; Lee, B; et al. Identification of Missing Proteins in Human Olfactory Epithelial Tissue by Liquid Chromatography-Tandem Mass Spectrometry. J Proteome Res. 2018, 17, 4320–4324. [Google Scholar] [CrossRef]
- Yoshikawa, K; Wang, H; Jaen, C; Haneoka, M; Saito, N; Nakamura, J; et al. The human olfactory cleft mucus proteome and its age-related changes. Sci Rep. 2018, 8, 17170. [Google Scholar] [CrossRef] [PubMed]
- Kim, YS; Han, D; Kim, J; Kim, DW; Kim, YM; Mo, JH. In-Depth, Proteomic Analysis of Nasal Secretions from Patients With Chronic Rhinosinusitis and Nasal Polyps doi: 10.4168/aair.2019.11.5.691. Allergy Asthma Immunol Res. Erratum in: Allergy Asthma Immunol Res. 2020, 12, 744. doi: 10.4168/aair.2020.12.4.744. 2019, 11, 691–708. [Google Scholar] [CrossRef] [PubMed]
- Kuntová, B; Stopková, R; Stopka, P. Transcriptomic and Proteomic Profiling Revealed High Proportions of Odorant Binding and Antimicrobial Defense Proteins in Olfactory Tissues of the House Mouse. Front Genet. 2018, 9, 26. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Genomic context, domain organisation and structure of BPIFB4. Organisation of the human BPIF gene locus showing the position of BPIFB4. BPIFB genes are in blue (pseudogenes are in turquoise) and BPIFA genes are in red (pseudogenes are pink). A. Schematic representation of human BPIFB4 protein. The position of the BPI1 (green) and BPI2 (red) domains are shown along with the position of the polymorphic variants seen in LAV-BPIFB4. The insertion of the unstructured region is shown in the BPI1 domain. B. The structure of human BPIFB4 from alphafold (https://alphafold.ebi.ac.uk/entry/AF-P59827-F1) is presented above that of human BPI (https://alphafold.ebi.ac.uk/entry/AF-P17213-F1). The unstructured region is predicted with very low confidence. The two domains of BPI are shown in different colours. C.
Figure 1.
Genomic context, domain organisation and structure of BPIFB4. Organisation of the human BPIF gene locus showing the position of BPIFB4. BPIFB genes are in blue (pseudogenes are in turquoise) and BPIFA genes are in red (pseudogenes are pink). A. Schematic representation of human BPIFB4 protein. The position of the BPI1 (green) and BPI2 (red) domains are shown along with the position of the polymorphic variants seen in LAV-BPIFB4. The insertion of the unstructured region is shown in the BPI1 domain. B. The structure of human BPIFB4 from alphafold (https://alphafold.ebi.ac.uk/entry/AF-P59827-F1) is presented above that of human BPI (https://alphafold.ebi.ac.uk/entry/AF-P17213-F1). The unstructured region is predicted with very low confidence. The two domains of BPI are shown in different colours. C.
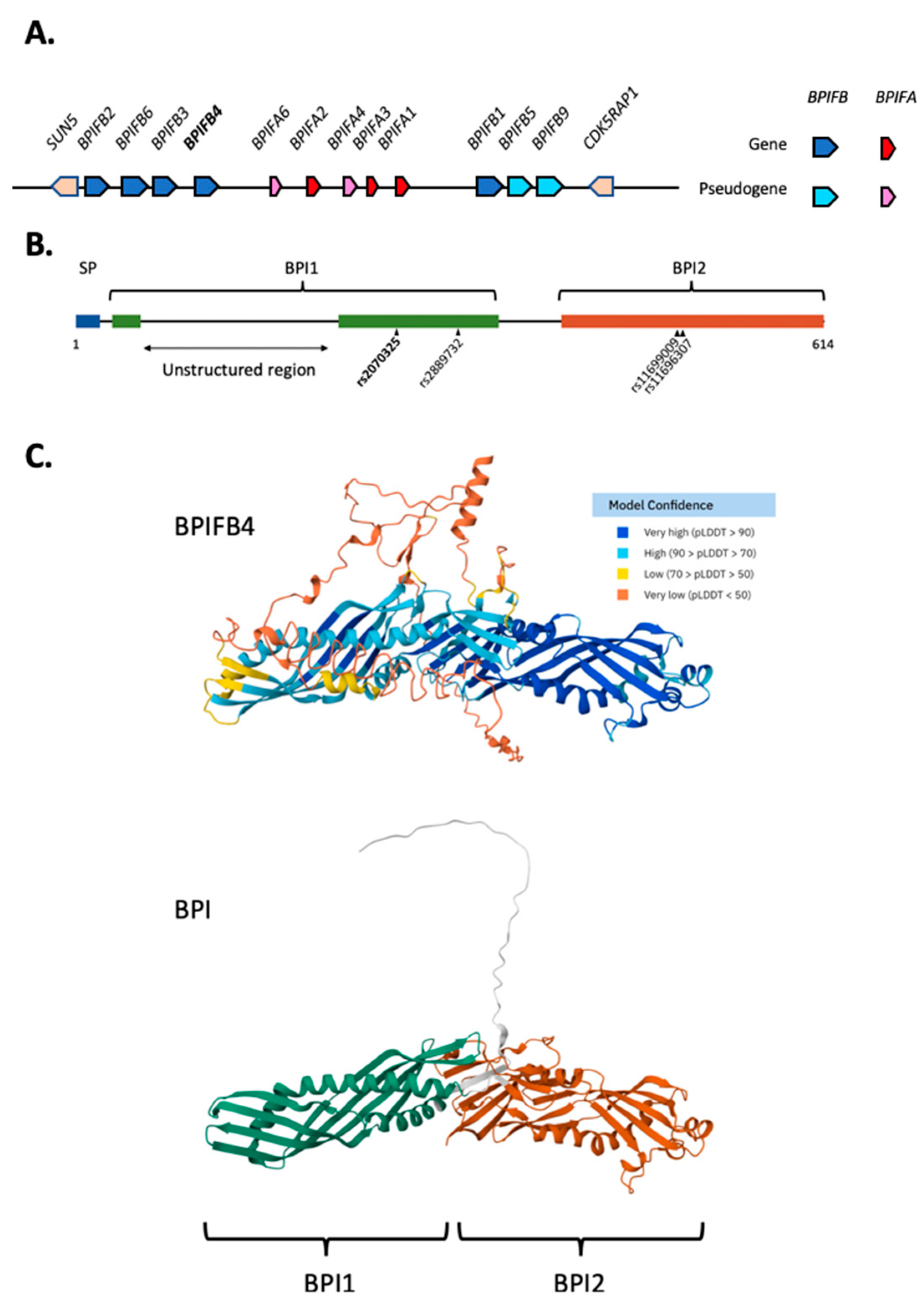
Figure 2.
Distribution of BPIFB4 in data sets represented in the Human Peptide Atlas. BPIFB4 (P59827) is present in 20 of 837 data sets in the 2025-1 build of the Human Peptide Atlas. https://peptideatlas.org/builds/human/.
Figure 2.
Distribution of BPIFB4 in data sets represented in the Human Peptide Atlas. BPIFB4 (P59827) is present in 20 of 837 data sets in the 2025-1 build of the Human Peptide Atlas. https://peptideatlas.org/builds/human/.
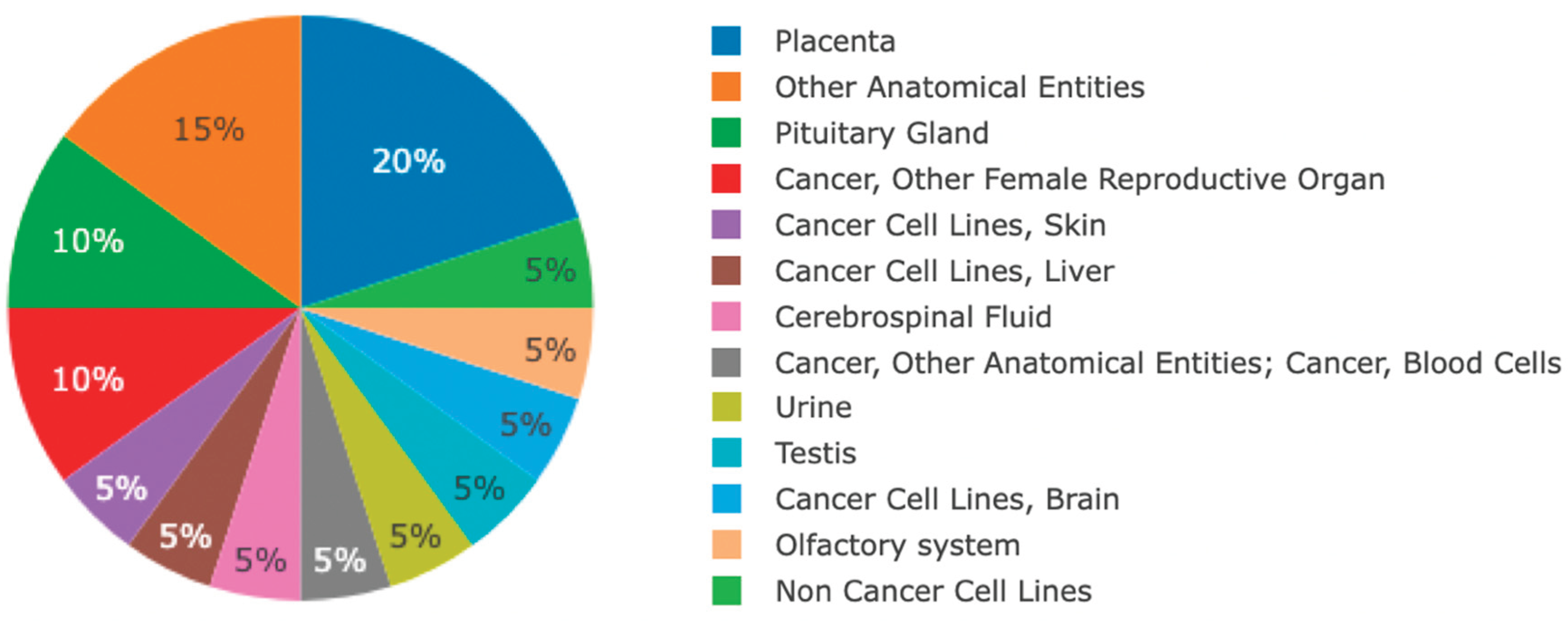

Table 1.
BPIFB4 positive cell numbers in selected single cell populations. BPIFB4 positivity data was extracted from selected cells/tissues in the Gene Expression application of CellXGene. https://cellxgene.cziscience.com/gene-expression.
Table 1.
BPIFB4 positive cell numbers in selected single cell populations. BPIFB4 positivity data was extracted from selected cells/tissues in the Gene Expression application of CellXGene. https://cellxgene.cziscience.com/gene-expression.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



