Submitted:
19 November 2025
Posted:
20 November 2025
You are already at the latest version
Abstract
The circadian rhythm regulates the activity of the ants according to environmental conditions. We investigated the interplay among circadian rhythm, nest homeostasis, and worker morphology in Azteca chartifex spiriti, a Neotropical arboreal species that builds large polydomous nests suspended in trees. In ten colonies, we measured internal moisture and temperature gradients in the main nest, which houses most individuals, including the reproductive female, immatures, and numerous workers. In six colonies, we assessed the polymorphism of foraging workers over a 24-h cycle in relation to external temperature variation. The results show integrated thermoregulatory mechanisms that combine passive strategies, derived from nest architecture and moisture gradients from the suspension base to the lower extremity, with active strategies linked to foraging patterns and worker polymorphism. Internal temperature remained stable despite external fluctuations, and the humidity gradient accounted for both internal population distribution and heat absorption or release. Worker size displayed a bimodal distribution during the day that shifted to a unimodal pattern at night, indicating behavioral adjustments to thermal and operational demands. These findings demonstrate that the interaction between physical structure and worker behavior maintains colony homeostasis and enhances the ability of A. chartifex spiriti to persist in environments with fluctuating climatic conditions.
Keywords:
homeostasis
; foraging
; moisture
; nest architecture
; polymorphism
1. Introduction
Ants represent one of the most abundant and ecologically significant groups of terrestrial invertebrates, exhibiting wide geographic distribution and occurrence across diverse environments, which reflects their high taxonomic and functional diversity [1,2]. Their remarkable behavioral and ecological diversity includes complex collective behaviors that ensure colony homeostasis, a key factor for survival under varying environmental conditions. Among these behaviors, thermoregulation stands out as a fundamental mechanism that integrates nest architecture and social behavior to maintain stable microclimatic conditions within colonies [3,4,5]. Various species display distinct thermoregulatory strategies, including Solenopsis invicta [6], Acromyrmex heyeri [7], and Eciton burchellii [8], as well as other hypogeic ants [5,9,10,11]. In addition to temperature, humidity gradients and behavioral plasticity also shape nest architecture and worker spatial distribution, reflecting the integration of abiotic and behavioral factors in optimizing the colony microenvironment [5].
The foraging behavior of ants, regulated by diverse communication modes among colony members, is a key component of colony thermoregulation. Social signals and individual information determine when and where workers search for resources, which helps to reduce exposure to thermally adverse conditions [12]. During this activity, ants adjust their behavior to minimize exposure to thermally stressful environments, highlighting temperature as a major factor modulating foraging activity [13,14,15,16]. Thermal fluctuations influence both physiological functions and behavioral patterns, affecting individuals and the colony as a whole [17].
The reproductive cycle and the development of immature stages strongly depend on the thermal homeostasis within the nest, prompting the colony to adopt various strategies to maintain this homeostasis [18,19,20,21]. Adaptive responses such as thermal plasticity [22,23,24,25] and temporal niche partitioning [25,26,27] exemplify this process. These strategies allow colonies to adjust their foraging patterns [28,29,30] to the optimal thermal conditions, whether in terms of time periods or temperature ranges, for their activities [31,32,33].
Moreover, mature colonies exhibit morphological and ontogenetic variation among workers, resulting in different responses to environmental temperature conditions [6,34,35]. Such variation promotes the specialization of subgroups in specific tasks, including foraging, brood care, and the maintenance of the internal microclimate of the nest [17,36,37,38].
In addition to structural and behavioral adaptations, the daily rhythm of light and dark drives ants to adjust their activities in response to changes in the external environment [39,40,41]. These light–dark cycles modulate not only external behaviors like foraging [39], but also have a direct influence on the internal dynamics of the colony [42].
The response of biological rhythms to nycthemeral cycles can vary substantially among species that nest in contrasting substrates, such as the soil and the canopy. Microclimatic conditions and resource availability in these distinct environments affect the intensity and periodicity of their behaviors, which reflect specific adaptations to the thermal and ecological characteristics of each habitat [43,44,45,46].
Microclimatic studies conducted in forest canopies indicate that solar radiation, exposure, and the structural complexity of the arboreal layer generate spatial and temporal thermal and moisture variations, which influence the foraging activity windows of worker ants and nest placement due to the risk of overheating the resident population [45,47,48]. Despite the recognized ecological importance of ants, the specific influence of abiotic factors on the activity and survival of arboreal species remains less studied in comparison to ground-dwelling species, limiting predictions about their behavioral responses to the projected impacts of climate change [45,49].
Arboreal colonies may be particularly vulnerable to extreme thermal events and shifts in temperature regimes [50], as physiological and behavioral limits constrain foraging activity and the development of the immature stages within nests. Therefore, there is an urgent need for studies integrating nest architecture, microclimatology, and the behavioral ecology of thermal tolerance to guide conservation and management strategies for ants [51]. Different genera of arboreal carton-nesting species, such as Camponotus, Crematogaster, Oecophylla, Polyrhachis, and Azteca, face specific thermal challenges associated with canopy life. The structure of the supporting tree, the architectural complexity, as well as the materials used in nest construction play fundamental roles in microclimatic regulation within colonies, yet these roles remain insufficiently explored [47,48,52,53]. These considerations highlight the urgent need for in-depth studies on the thermal ecology and adaptive strategies of these species, particularly to better inform conservation and management practices under rapidly changing climatic conditions.
Our study focuses on Azteca chartifex spiriti Forel, 1912, a species widely distributed in the tropical rainforest environments in Brazil. Like other species of the genus, it exhibits a polydomous nesting system in which a single colony occupies multiple nests on one or more neighboring trees [54]. These colonies consist of a main nest surrounded by satellite and subordinate nests, as well as small structures called “shelters” that house associated hemipterans [55,56,57,58].
A. chartifex spiriti dominates the tree canopy where it nests, contributing to the mosaic of dominant arboreal ant species in the cocoa-producing region of Bahia, Brazil [59]. Colonies can contain thousands of highly aggressive workers which play an crucial ecological role in shaping the composition of canopy arthropod communities [59,60,61]. Evidence indicates that this species has strong potential as a biological control agent due to three key traits: (i) its presence is linked to reduced populations of harmful insects in cocoa plantations [55,62]; (ii) its colonies demonstrate spatial and temporal stability [63]; and (iii) they exhibit a high capacity for territorial expansion [59,62,63].
Due to these characteristics, A. chartifex spiriti emerges as a promising model for studies on integrated pest management and the conservation of ecosystem services provided by cocoa agroforestry systems. Understanding the thermal ecology of Azteca chartifex spiriti is essential, as this species faces specific thermoregulatory challenges that are inherent to its arboreal habitat and to the constant exposure of its nests to fluctuating wind, rainfall, and solar radiation. The forest canopy represents a highly dynamic thermal environment, subject to diurnal and nocturnal temperature variations, with amplitude that depend on seasonal conditions.
The ability of A. chartifex spiriti to regulate the internal temperature of its nests and optimize thermoregulatory mechanisms is critical for the species’ ecological success and the proper functioning of its colonies. Moreover, a detailed analysis of the species’ thermal ecology could provide valuable insights that are applicable to other arboreal ants, helping to elucidate how they cope with thermal fluctuations and how these factors may affect them in the context of climate change.
Using A. chartifex spiriti as a biological model organism, this study aimed to: (i) examine how environmental thermal fluctuations influence nest ecology; (ii) identify the thermal control mechanisms employed by colonies to maintain internal thermal homeostasis; (iii) assess the role of worker polymorphism in colony thermoregulation; and (iv) analyze how the physical and structural properties of nests contribute to the thermal regulation of colonies.
2. Materials and Methods
2.1. Study Area
The study was conducted in the municipality of Ilhéus, Bahia, Brazil (Figure 1). Located within the Atlantic Forest biome, the region exhibits a diverse vegetation structure, including “restinga” formations, mangrove forests, cacao plantations, wetlands, and floodplains [64]. The climate is humid tropical, with mean annual temperatures ranging from 20°C to 25°C and precipitation exceeding 1,200 mm.
Colonies of Azteca chartifex spiriti were collected in agroforestry cacao systems (for definitions of the different types of cacao cultivation in the region, see [65]: (i) on the campus of the Universidade Estadual de Santa Cruz (UESC) (14°47’44”S, 39°10’06”W), characterized by a “cabruca” system, where cacao trees are cultivated under the shade of the remnant Atlantic Forest canopy; and (ii) in the experimental areas of the Comissão Executiva do Plano da Lavoura Cacaueira (CEPLAC) (14°46’58”S, 39°13’16”W), representing a clear-cut cultivation system lacking arboreal cover.
2.2. Experimental Design
To investigate the interrelationship among the nycthemeral rhythm, nest homeostasis, and worker morphology within the polydomous nesting system of Azteca chartifex spiriti, the study was structured into two complementary experimental phases. The first focused on the internal humidity gradient of the nests, while the second evaluated how external temperature fluctuations influence nest temperature and worker polymorphism under various activity conditions.
2.2.1. Phase I: Internal Nest Humidity Patterns
Ten polydomous colonies were selected, each with a main nest that exceeds 60 cm in height. Only the principal nest from each colony was used in this phase. In each nest, four vertical layers were defined, each 15 cm thick, establishing a vertical sampling gradient (from bottom to top) across four levels: from level 1, at the lower extremity, to level 4, at the base where the nest is attached to the supporting branch (Figure 2A).
From each level, samples of the nest’s construction material were collected, fragmented, and stored in glass beakers of 100 mL. Samples were first weighed fresh using an analytical balance (APX200/Denver) and subsequently oven-dried (Tecnal TE- 393/2) at 45°C for 72 hours before they were reweighed. The moisture percentage for each level was calculated based on the difference between wet and dry weights.
2.2.2. Phase II: Thermal Ecology and Worker Morphometry
Six polydomous colonies were selected, each with a main nest taller than 60 cm in height. Two digital temperature sensors (Exbom FEPRO-MUT60OS, accuracy ± 1°C) were installed in each main nest to simultaneously monitor internal temperature: one at the basal attachment point and another at the lower extremity (Figure 2B). After a 40-minute acclimation period to allow for sensor calibration and worker adjustment, temperature monitoring was conducted continuously for 24 hours, from 15:00 on one day to 15:00 the following day.
Internal temperatures (from both the base and lower extremity) and external ambient temperatures were recorded every two hours, resulting in a total of 13 readings per colony across the six monitored colonies. Sampling times were categorized as morning (05:00–11:00), afternoon (13:00–17:00), and night (19:00–03:00 the following morning).
Simultaneously with thermal monitoring, five foraging workers were randomly collected from the trunk of the supporting tree every two hours. Workers were preserved and measured under a trinocular stereomicroscope (LEICA DMC2900) equipped with Leica Application Suite (LAS) software. The following morphometric traits (in millimeters) were analyzed: i) head width, which serves as a proxy for body size and strength; ii) Weber’s length, which is the largest rigid body measurement, which serves as an indicator of body size and metabolic traits [66]; and iii) femur length of one hind leg, which serves as a proxy for foraging speed and thermoregulatory strategy [67]. In total, 390 workers were used in this part of the experiment: six colonies × 13 samples of five workers collected every two hours.
2.3. Statistical Analysis
Moisture distribution in the main nest was assessed using the Kruskal-Wallis test. Sampling levels were defined as the predictor variable, and moisture content as the response variable. Pairwise comparisons were conducted using Dunn's test (1964). Differences in the internal temperature of the main nest were evaluated using Student's t-test for independent samples, comparing: i) the temperature at the supporting base versus the lower end of the structure; and ii) the internal nest temperature versus the external ambient temperature. Diurnal variation in temperature was investigated using analysis of variance (ANOVA).
Worker polymorphism across different activities and colonies was examined by correlating morphological parameters (head width, femur length and Weber’s length) using Pearson rank correlation coefficient. The relationship between foraging activity and worker polymorphism was tested with the Kruskal-Wallis test, followed by multiple comparisons using Dunn’s test (1964). In this analysis, the morphological parameters were treated as predictor variables and the periods of the day as the response variable. All statistical analyses and figures were produced with R v. 4.5.0 (R Core Team, 2025).
3. Results
3.1. Phase I: Internal Nest Humidity Patterns
The studied colonies exhibited a consistent pattern in moisture distribution: the lower end of the main nest showed consistently higher moisture content than the upper portion of the nest that attaches to the supporting branch. A significant difference in moisture content was found among sampling levels (Kruskal–Wallis: H = 17.61; df = 3; p < 0.001). Pairwise comparisons using Dunn’s test indicated that level 1 differed significantly from level 3 (Z = 3.90; p < 0.001) and level 4 (Z = 3.28; p < 0.05). Level 2 did not show significant differences when compared to the other levels (Figure 3).
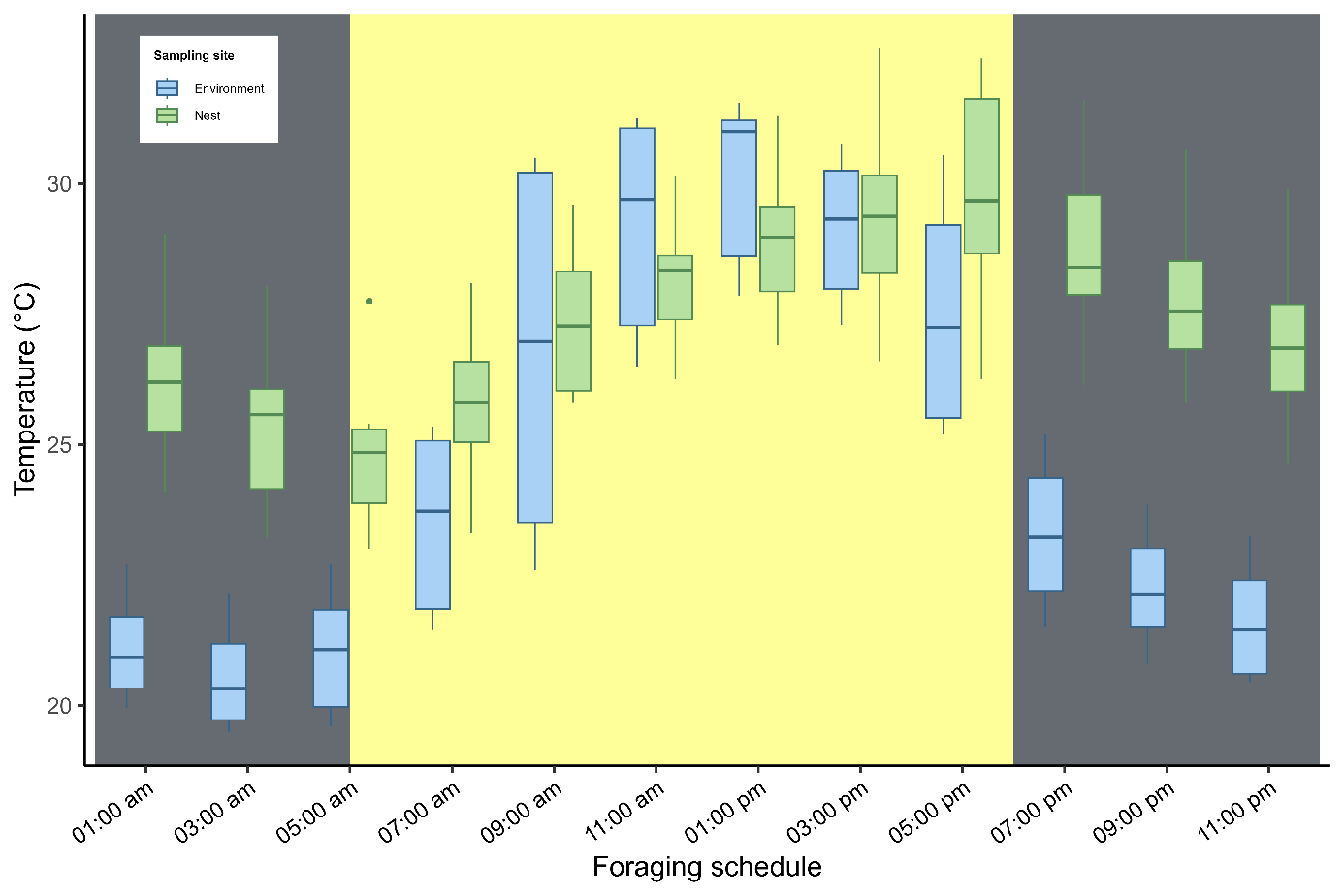
Internal nest temperature showed homogeneous variation between the supporting base (mean = 27.8 ± 2,41 °C) and the lower end (mean = 27.3 ± 2,40°C), with no significant difference between these points (t = −1.48; df = 154; p = 0.14; Figure 4A). In contrast, a significant difference was found between internal nest temperature and external ambient temperature (t = −5.06; df = 123; p < 0.001; Figure 4B). Significant thermal variation was also observed across different periods of the day (ANOVA: F2,153 = 51.06; R² = 0.40; p < 0.001; Figure 4C). Internal nest temperature tended to increase in parallel with rising external ambient temperature (linear model: F11,144 = 83.6; R² = 0.46; p < 0.001; Figure 5).
3.2. Phase II: Thermal Ecology and Worker Morphometry
In the preliminary study of worker polymorphism, strong correlations were observed between Weber’s length and hind femur length (r=0.86, p<0.001) (Figure 6A), and between Weber’s length and head width (r = 0.92, p < 0.001) (Figure 6B). In addition, head width was strongly correlated with hind femur length (r = 0.89, p < 0.001) (Figure 6C).
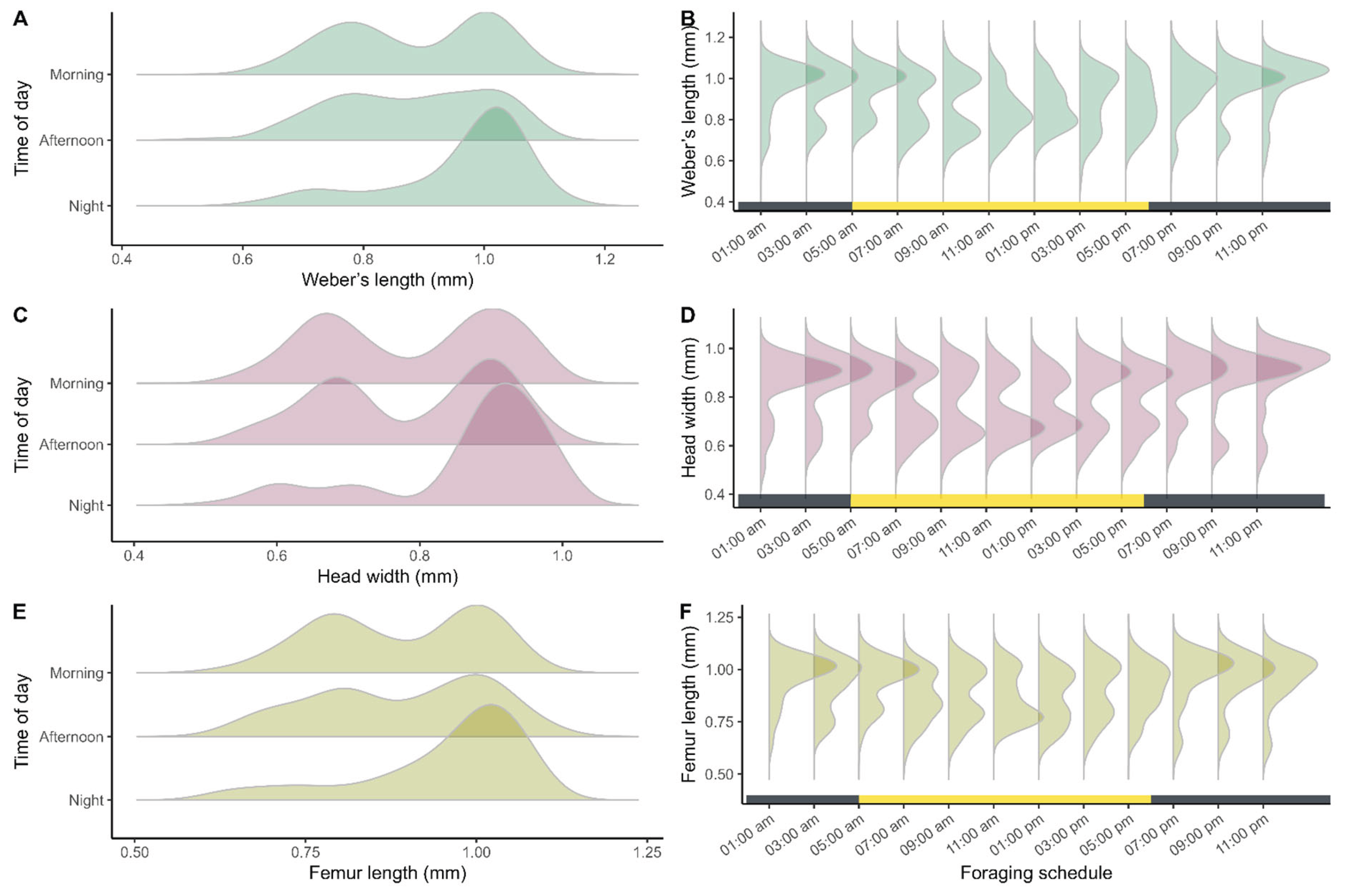
Significant differences in worker size were found among different periods of the day for all three traits analyzed: Weber’s length (Kruskal–Wallis: H = 36.6; df = 2; p < 0.001; Figure 7A), head width (H = 41.84; df = 2; p < 0.001; Figure 7C), and hind femur length (H = 27.6; df = 2; p < 0.001; Figure 7E). Multiple comparisons (Table S2, Supplementary Material) indicate that workers active during the night differ significantly from those active during other periods for all variables.
Based on the observed morphometric patterns, two distinct foraging patterns across the daily cycle were identified. During the night period, between 07:00 PM and 03:00 AM, foraging activity followed a unimodal pattern characterized by a predominance of larger workers (Figure 6B, D, F). During the daytime period, between 05:00 AM and 05:00 PM, foraging exhibited a bimodal structure that remained consistent throughout the day, indicating a greater diversity in the sizes of workers involved in these activities (Figure 6B, D, F).
4. Discussion
4.1. Phase I: Internal Nest Humidity Patterns
Our results indicate that temperature is evenly distributed within the main nest, with no significant differences between the basal attachment and the lower extremity, indicating a thermally stable microclimate. However, the moisture content was significantly higher in the lower portion of the nest. This variation is clearly influenced by nest morphology: while the basal attachment is broader and firmly anchored to the tree trunk, the lower extremity is narrower and elongated, creating a gradient that promotes the drainage of water collected by the structure (from dew, ambient humidity, or rainfall). Moreover, being positioned beneath a wide supporting branch, this portion remains partially shielded from direct solar radiation and wind exposure. Additionally, the network of interconnected tunnels and internal chambers that are linked by multiple openings, establishes a natural ventilation system that facilitates the inflow of fresh air and the expulsion of air saturated by moisture. This architectural configuration contributes to maintaining a stable internal microclimate suitable for colony homeostasis, as also observed in other social insects whose nest design plays a crucial role in thermal regulation and gas exchange [7,68,69]. These findings reinforce the idea that nest architecture, whether subterranean or arboreal, constitutes a fundamental component of the colony’s ability to buffer environmental fluctuations and maintain physiological homeostasis [68].
The insulating properties of the nest-building material represent another crucial factor for maintaining internal microclimate stability. Many Azteca species construct carton nests using a combination of sediments and cellulose fibers cemented by proteinaceous secretions from the workers’ maxillary glands [70]. These structural components are also found in the nests of other arboreal ants, such as Oecophylla, Camponotus and Polyrhachis [47,71,72], as well as in arboreal termites [73] and social wasp colonies [74]. Cellulose fibers provide mechanical strength to the nest, whereas the proteinaceous compounds secreted by workers form a hydrophobic barrier that reduces permeability and shields the nest’s interior from excessive moisture. These insulating properties are essential for maintaining the microclimatic stability of the colonies, ensuring favorable conditions for survival, brood development, and overall colony functioning [75].
The diurnal variation in temperature shows that during the afternoon period, the internal temperature of the nests tends to rise with increases in ambient temperature. Passive strategies may explain the colony’s ability to maintain a stable microclimate even during periods of the higher heat. In addition to nest architecture, the thermal diffusivity of the building material, combined with the humidity gradient, likely functions as an important thermal moderator, preventing colony overheating.
Similarly to the findings of Klingner [76] in the paper wasp nests, fluctuations in internal humidity may create pressure gradients between the intranidal and external environments, which are dissipated through the nest envelope and contribute to thermal stabilization. Another relevant mechanism is evaporation, an endothermic process that aids in temperature reduction. In A. chartifex spiriti nests, a comparable process may occur, where external air cools upon contact with the internal humidity of the nest, establishing a convective flow that helps maintain a cooler microclimate during temperature peaks.
In his thermodynamic analysis of Formica polyctena nests, Frouz [77] emphasized the central role of humidity in thermal regulation, noting contrasting effects between dry and moist nests. Additionally, Frouz [77] and Kadochová [4] demonstrated that nest humidity can influence microbial activity, increasing internal temperature or alter the insulating properties of the material, resulting in greater heat dissipation. Therefore, humidity not only modulates temperature but also determines the overall thermal balance within the nest, depending on both internal and environmental conditions.
4.2. Phase II: Thermal Ecology and Worker Morphometry
Despite the temperature reduction observed during the morning and nighttime periods, both inside and outside the nests, the intranidal temperature remained consistently higher than that of the external environment. Considering foraging patterns, worker polymorphism, and thermal fluctuations, it can be inferred that metabolic heat is the primary mechanism responsible for maintaining the temperature of the nest during periods of low external temperature. This heat, generated by the metabolic and behavioral activities of the organisms within the nest, acts as a key thermal modulator, ensuring the stability of the internal climate [8,78,79].
Studies conducted by Frouz [77] on Formica polyctena nests and by Korb [80] on Macrotermes bellicosus (Macrotermitinae) demonstrated the relevance of associated microorganisms in nest thermoregulation. In this context, A. chartifex spiriti stands out among arboreal ants because, in addition to hosting a highly diverse associated fauna [81,82,83], its colonies are exceptionally large and populous, containing tens of thousands of workers [56,58,84]. This high population density, combined with the continuous activity of workers, contributes to the generation and retention of metabolic heat, ensuring thermal homeostasis even under adverse environmental conditions.
The worker caste polymorphism in A. chartifex spiriti, previously documented by Wheeler [84] and Longino [85], may serve as an active thermoregulatory mechanism for the colony. Miranda [58] provided a detailed characterization of the spatial organization and population structure of this species, demonstrating that the polymorphic worker caste is functionally specialized within the nest. Larger workers perform defensive and maintenance activities, while smaller workers attend to the brood and maintain internal conditions. This segregation of tasks suggests that worker polymorphism is not only a behavioral adaptation linked to territorial dominance and resource defense, but may also indirectly contribute to microclimatic regulation. In the present study, polymorphic patterns were analyzed to support the hypothesis that metabolic heat and the spatial division of labor jointly mediate nest thermal stability during the nighttime period. Two plausible scenarios may explain how the shift in worker distribution patterns (from bimodal during the day to unimodal at night) coincides with temperature variations inside the nest: (i) the significant return of minor workers at night may lead to an increase in internal temperature, as residual heat from solar energy absorbed by their bodies contributes to nest warming [4,79]; and (ii) the higher activity of major workers during nighttime may raise nest temperature through the release of metabolic heat. These individuals, being more tolerant of thermal stress resulting from lower ambient temperatures, play a key role in maintaining elevated internal temperatures.
This phenomenon aligns with previous studies on polymorphic ant species [86,87,88,89], which highlights the contribution of worker polymorphism to thermal regulation and energy efficiency within colonies. Consequently, A. chartifex spiriti appears to dynamically adjust its activity and foraging patterns in response to thermal conditions, optimizing resource collection while minimizing the mortality risks associated with environmental stress [88].
Thermoregulation, both active and passive, depends on the coordinated activities of individual workers to maintain the thermal homeostasis of the colony [3]. Our results indicate that A. chartifex spiriti employs a combination of complementary thermoregulatory mechanisms. Passively, the architecture of the nest, along with internal humidity gradients, heat dissipation capacity, and evaporative processes, plays a key role in buffering thermal fluctuations, particularly during periods of elevated ambient temperature.
Actively, worker behavior, expressed through alternations in foraging patterns along the nycthemeral cycle, contributes to internal heat input when external temperatures decrease. Thus, this study not only advances our understanding of thermal adaptations in arboreal ant species but also highlights the importance of the interplay between behavioral and architectural factors in the microclimatic regulation of nests. This integration represents a critical component of colony survival and ecological success under thermally challenging conditions. The findings presented here provide a robust foundation for future research on the ecological mechanisms that sustain biodiversity and functional stability in arboreal nesting systems.
5. Conclusions
This study deepens our understanding of thermoregulatory mechanisms of A. chartifex spiriti, highlighting the complementarity between nest architecture and worker behavior in maintaining the colony’s thermal homeostasis. We demonstrated that this species employs a combination of passive and active strategies to regulate internal nest temperature, ensuring the stability necessary for survival in tropical environments.
Further studies may explore how different arboreal ant species adjust their thermoregulatory mechanisms in response to more extreme environmental fluctuations, including: (i) prolonged droughts and heat waves; and (ii) seasonal changes and variations in rainfall regimes. Another promising direction involves examining the relationship between nest architectural variation and nesting strata to gain a better understanding of how these interactions contribute to colony resilience across ecosystems.
In summary, our findings emphasize the importance of investigating biodiversity and ecological processes in arboreal ant nesting systems, providing a strong foundation for future research aimed at the conservation and sustainable management of these complex ecological structures.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, J.T.S., J.H.C.D. and C.S.F.M.; formal analysis, J.T.S. and E.B.A.K.; investigation J.T.S., J.L.S. and L.S.B.; writing – original draft preparation, J.T.S. and C.S.F.M.; writing – review and editing, J.T.S., E.B.A.K., J.H.C.D. and C.S.F.M. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by Coordination for the Improvement of Higher Education Personnel (CAPES, Brazil) project 88887.486528/2020-00 to Josieia Teixeira dos Santos; National Council for Scientific and Technological Development (CNPQ, Brazil) project 306885/2023-9 to Jacques Hubert Charles Delabie; Santa Cruz State University (UESC, Bahia, Brazil) project 073.110116.2025.0020483-37 to Cléa dos Santos Ferreira Mariano.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
We thank the staffs of the Myrmecology Laboratory of CEPLAC and Social Arthropod Laboratory (LABAS) of UESC for assistance in field works.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hölldobler, B.; Wilson, E.O. The Ants. Cambridge, Mass. Harvard Univ. Press 1990, xii +-732.
- Passera, L.; Aron, S. Les Fourmis: Comportement, Organisation Sociale et Évolution; 2005; ISBN 066097021X.
- Jones, J.C.; Oldroyd, B.P. Nest thermoregulation in social insects. In Advances in Insect Physiology; 2006; Vol. 33, pp. 153–191.
- Kadochová, Š.; Frouz, J. Thermoregulation strategies in ants in comparison to other social insects, with a focus on red wood ants (Formica rufa group). F1000Research 2014, 2, 280. [Google Scholar] [CrossRef]
- Sankovitz, M.; Purcell, J. Ant nest architecture is shaped by local adaptation and plastic response to temperature. Sci. Rep. 2021, 11, 23053. [Google Scholar] [CrossRef]
- Penick, C.A.; Tschinkel, W.R. Thermoregulatory brood transport in the fire ant, Solenopsis invicta. Insectes Soc. 2008, 55, 176–182. [Google Scholar] [CrossRef]
- Bollazzi, M.; Roces, F. The thermoregulatory function of thatched nests in the South American grass-cutting ant, Acromyrmex heyeri. J. Insect Sci. 2010, 10, 1–17. [Google Scholar] [CrossRef]
- Franks, N.R. Thermoregulation in army ant bivouacs. Physiol. Entomol. 1989, 14, 397–404. [Google Scholar] [CrossRef]
- Lapointe, S.L.; Serrano, M.S.; Jones, P.G. Microgeographic and vertical distribution of Acromyrmex landolti (Hymenoptera: Formicidae) nests in a Neotropical savanna. Environ. Entomol. 1998, 27, 636–641. [Google Scholar] [CrossRef]
- Bollazzi, M.; Kronenbitter, J.; Roces, F. Soil temperature, digging behaviour, and the adaptive value of nest depth in South American species of Acromyrmex leaf-cutting ants. Oecologia 2008, 158, 165–175. [Google Scholar] [CrossRef] [PubMed]
- García Ibarra, F.; Jouquet, P.; Bottinelli, N.; Bultelle, A.; Monnin, T. Experimental evidence that increased surface temperature affects bioturbation by ants. J. Anim. Ecol. 2024, 93, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Kolay, S.; Boulay, R.; D’Ettorre, P. Regulation of ant foraging: A review of the role of information use and personality. Front. Psychol. 2020, 11, 734. [Google Scholar] [CrossRef]
- Cerdá, X. Behavioural and physiological traits to thermal stress tolerance in two Spanish desert ants. Etologia 2001, 9, 15–27. [Google Scholar]
- Jayatilaka, P.; Narendra, A.; Reid, S.F.; Cooper, P.; Zeil, J. Different effects of temperature on foraging activity schedules in sympatric Myrmecia ants. J. Exp. Biol. 2011, 214, 2730–2738. [Google Scholar] [CrossRef] [PubMed]
- Bujan, J.; Yanoviak, S.P. Behavioral response to heat stress of twig-nesting canopy ants. Oecologia 2022, 198, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Roeder, D. V.; Paraskevopoulos, A.W.; Roeder, K.A. Thermal tolerance regulates foraging behaviour of ants. Ecol. Entomol. 2022, 47, 331–338. [Google Scholar] [CrossRef]
- Villalta, I.; Oms, C.S.; Angulo, E.; Molinas-González, C.R.; Devers, S.; Cerdá, X.; Boulay, R. Does social thermal regulation constrain individual thermal tolerance in an ant species? J. Anim. Ecol. 2020, 89, 2063–2076. [Google Scholar] [CrossRef]
- Porter, S.D. Impact of temperature on colony growth and developmental rates of the ant, Solenopsis invicta. J. Insect Physiol. 1988, 34. [Google Scholar] [CrossRef]
- Kipyatkov, V.E.; Lopatina, E.B.; Imamgaliev, A.A.; Shirokova, L.A. Effect of temperature on rearing of the first brood by the founder females of the ant Lasius niger (Hymenoptera, Formicidae) : Latitude-dependent variability of the response norm. J. Evol. Biochem. Physiol. 2004, 40, 165–175. [Google Scholar] [CrossRef]
- Weidenmüller, A.; Mayr, C.; Kleineidam, C.J.; Roces, F. Preimaginal and adult experience modulates the thermal response behavior of ants. Curr. Biol. 2009, 19, 1897–1902. [Google Scholar] [CrossRef]
- Penick, C.A.; Diamond, S.E.; Sanders, N.J.; Dunn, R.R. Beyond thermal limits: comprehensive metrics of performance identify key axes of thermal adaptation in ants. Funct. Ecol. 2017, 31, 1091–1100. [Google Scholar] [CrossRef]
- Andrew, N.R.; Hart, R.A.; Jung, M.P.; Hemmings, Z.; Terblanche, J.S. Can temperate insects take the heat? A case study of the physiological and behavioural responses in a common ant, Iridomyrmex purpureus (Formicidae), with potential climate change. J. Insect Physiol. 2013, 59. [Google Scholar] [CrossRef]
- Kaspari, M.; Clay, N.A.; Lucas, J.; Yanoviak, S.P.; Kay, A. Thermal adaptation generates a diversity of thermal limits in a rainforest ant community. Glob. Chang. Biol. 2015, 21, 1092–1102. [Google Scholar] [CrossRef]
- Bujan, J.; Roeder, K.A.; Yanoviak, S.P.; Kaspari, M. Seasonal plasticity of thermal tolerance in ants. Ecology 2020, 101, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Roeder, K.A.; Bujan, J.; de Beurs, K.M.; Weiser, M.D.; Kaspari, M. Thermal traits predict the winners and losers under climate change: An example from North American ant communities. Ecosphere 2021, 12, e03645. [Google Scholar] [CrossRef]
- Tavares, A. a; Bispo, P.C.; Zanzini, A.C. Efeito do turno de coleta sobre comunidades de formigas epigéicas (Hymenoptera: Formicidae) em áreas de Eucalyptus cloeziana e de cerrado. Neotrop. Entomol. 2008, 37, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Robledo, C.; Chuquillanqui, H.; Kuprewicz, E.K.; Escobrar-Sarria, F. Lower thermal tolerance in nocturnal than in diurnal ants: A Challenge for nocturnal ectotherms facing global warming. Ecol. Entomol. 2018, 43, 162–167. [Google Scholar] [CrossRef]
- Cerdá, X.; Retana, J.; Cros, S. Critical thermal limits in mediterranean ant species: Trade-off between mortality risk and foraging performance. Funct. Ecol. 1998, 12, 45–55. [Google Scholar] [CrossRef]
- Lessard, J.-P.; Dunn, R.R.; Sanders, N.J. Temperature-mediated coexistence in temperate forest ant communities. Insectes Soc. 2009, 56, 149–156. [Google Scholar] [CrossRef]
- Stuble, K.L.; Rodriguez-Cabal, M.A.; McCormick, G.L.; Juric, I.; Dunn, R.R.; Sanders, N.J. Tradeoffs, competition, and coexistence in eastern deciduous forest ant communities. Oecologia 2013, 171, 981–992. [Google Scholar] [CrossRef]
- Ruano, F.; Tinaut, A.; Soler, J.J. High surface temperatures select for individual foraging in ants. Behav. Ecol. 2000, 11, 396–404. [Google Scholar] [CrossRef]
- van Oudenhove, L.; Billoir, E.; Boulay, R.; Bernstein, C.; Cerdá, X. Temperature limits trail following behaviour through pheromone decay in ants. Naturwissenschaften 2011, 98, 1009–1017. [Google Scholar] [CrossRef]
- Nobua-Behrmann, B.E.; Lopez De Casenave, J.; Milesi, F.A.; Farji-Brener, A. Coexisting in harsh environments: temperature-based foraging patterns of two desert leafcutter ants (Hymenoptera: Formicidae: Attini). Myrmecological News 2017, 25, 41–49. [Google Scholar]
- Banschbach, V.S.; Levit, N.; Herbers, J.M. Nest temperatures and thermal preferences of a forest ant species: is seasonal polydomy a thermoregulatory mechanism ? Insectes Soc. 1997, 44, 109–122. [Google Scholar] [CrossRef]
- Wills, B.D.; Powell, S.; Rivera, M.D.; Suarez, A. V. Correlates and consequences of worker polymorphism in ants. Annu. Rev. Entomol. 2018, 63, 575–598. [Google Scholar] [CrossRef]
- Busher, C.E.; Calabi, P.; Traniello, J.F.A. Polymorphism and division of labor in the Neotropical ant Camponotus sericeiventris Guerin (Hymenoptera: Formicidae). Ann. Entomol. Soc. Am. 1985, 78, 221–228. [Google Scholar] [CrossRef]
- Crozier, R.H.; Newey, P.S.; Schlüns, E.A.; Robson, S.K.A. A masterpiece of evolution Oecophylla weaver ants (Hymenoptera: Formicidae). Myrmecological News 2009, 13, 57–71. [Google Scholar]
- Kamhi, J.F.; Nunn, K.; Robson, S.K.A.; Traniello, J.F.A. Polymorphism and division of labour in a socially complex ant: neuromodulation of aggression in the australian weaver ant, Oecophylla smaragdina. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150704. [Google Scholar] [CrossRef] [PubMed]
- Narendra, A.; Reid, S.F.; Raderschall, C.A. Navigational efficiency of nocturnal Myrmecia ants suffers at low light levels. PLoS One 2013, 8, e58801. [Google Scholar] [CrossRef] [PubMed]
- Kay, J.; Menegazzi, P.; Mildner, S.; Roces, F.; Helfrich-Förster, C. The circadian clock of the ant Camponotus floridanus is localized in dorsal and lateral neurons of the brain. J. Biol. Rhythms 2018, 33, 255–271. [Google Scholar] [CrossRef]
- Libbrecht, R.; Nadrau, D.; Foitzik, S. A role of histone acetylation in the regulation of circadian rhythm in ants. iScience 2020, 23, 100846. [Google Scholar] [CrossRef]
- Sharma, V.; Lone, S.; Goel, A.; Chandrashekaran, M.K. Circadian consequences of social organization in the ant species Camponotus compressus. Naturwissenschaften 2004, 91, 386–390. [Google Scholar] [CrossRef]
- Kaspari, M. Body Size and microclimate use in Neotropical granivorous ants. Oecologia 1993, 96, 500–507. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Morimoto, Y.; Widodo, E.S.; Mohamed, M.; Fellowes, J.R. Vertical habitat use and foraging activities of arboreal and ground ants (Hymenoptera: Formicidae) in a Bornean tropical rainforest. Sociobiology 2010, 56, 435–448. [Google Scholar]
- Stark, A.Y.; Adams, B.J.; Fredley, J.L.; Yanoviak, S.P. Out on a limb: Thermal microenvironments in the tropical forest canopy and their relevance to ants. J. Therm. Biol. 2017, 69, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Leahy, L.; Scheffers, B.R.; Williams, S.E.; Andersen, A.N. Arboreality drives heat tolerance while elevation drives cold tolerance in tropical rainforest ants. Ecology 2022, 103, e03549. [Google Scholar] [CrossRef] [PubMed]
- Devarajan, K. The antsy social network: Determinants of nest structure and arrangement in Asian weaver ants. PLoS One 2016, 11, e0156681. [Google Scholar] [CrossRef]
- Langshiang, E.S.; Hajong, S.R. Determination of structural features of the nest material of Crematogaster rogenhoferi (Mayr, 1879) (Hymenoptera: Myrmicinae). J. Entomol. Zool. Stud. 2018, 6, 1626–1631. [Google Scholar]
- Spicer, M.E.; Stark, A.Y.; Adams, B.J.; Kneale, R.; Kaspari, M.; Yanoviak, S.P. Thermal constraints on foraging of tropical canopy ants. Oecologia 2017, 183, 1007–1017. [Google Scholar] [CrossRef]
- Stuble, K.L.; Pelini, S.L.; Diamond, S.E.; Fowler, D.A.; Dunn, R.R.; Sanders, N.J. Foraging by forest ants under experimental climatic warming: A test at two sites. Ecol. Evol. 2013, 3, 482–491. [Google Scholar] [CrossRef]
- Mascarenhas, H.P. Utilidade das formiguinhas de S. Mateus. Rev. Agr. Imp. Inst. Flum. Agric. 1883, 14. [Google Scholar]
- Fowler, H.G.; Medeiros, M.A.; Delabie, J.H.C. Carton Nest allometry ans spatial patterning of the arboreal ant Azteca chartifex spiriti ( Hymenoptera, Formicidae). Rev. Bras. Entomol. 1996, 40, 337–339. [Google Scholar]
- Kulkarni, K.G.; Borkar, V.D. Architecture of arboreal nests of Crematogaster ants. In: Sediment Organism Interactions. A Multifaceted Ichnology; SEPM Society for Sedimentary Geology, 2007; Vol. 88, pp. 373–377.
- Jiménez-Soto, E.; Cruz-Rodríguez, J.A.; Vandermeer, J.; Perfecto, I. Hypothenemus hampei (Coleoptera: Curculionidae) and its interactions with Azteca instabilis and Pheidole synanthropica (Hymenoptera: Formicidae) in a shade coffee agroecosystem. Environ. Entomol. 2013, 42, 915–924. [Google Scholar] [CrossRef]
- Delabie, J.C. The ant problems of cocoa farms in Brazil. In: Applied Myrmecology: A World Perspective. Vander Meer, R.K., Jaffé, K., Cedeño, A., Eds., Boulder, Colorado, USA, 1990, 555-569.
- Delabie, J.H.C.; Benton, F.P.; Medeiros, M.A. La polydomie chez les Formicidae arboricoles dans les cacaoyères du Brésil: Optimisation de l’occupation de l’espace ou strategie défensive? Actes Coll. Insectes Soc. 1991, 7, 173–178. [Google Scholar]
- Delabie, J.H.C. Trophobiosis between Formicidae and Hemiptera (Sternorrhyncha and Auchenorrhyncha): An overview. Neotrop. Entomol. 2001, 30, 501–516. [Google Scholar] [CrossRef]
- Miranda, V.L.; Koch, E.; Delabie, J.H.C.; Bomfim, L.; Padre, J.; Mariano, C. Nest spatial structure and population organization in the Neotropical ant Azteca chartifex spiriti Forel, 1912 (Hymenoptera: Formicidae: Dolichoderinae). Ann. Soc.Entomol. Fr. 2021, 57, 499–508. [Google Scholar] [CrossRef]
- Majer, J.D.; Delabie, J.H.C.; Smith, M.R.B. Arboreal ant community patterns in Brazilian cocoa farms. Biotropica 1994, 26, 73. [Google Scholar] [CrossRef]
- Soares, G.R.; Anjos, D. V.; da Costa, F.V.; Lourenço, G.M.; Campos, R.I.; Ribeiro, S.P. Positive effects of ants on host trees are critical in years of low reproduction and not influenced by liana presence. Basic Appl. Ecol. 2022, 63, 93–103. [Google Scholar] [CrossRef]
- Soares, G.R.; Lourenço, G.M.; Costa, F. V.; Lopes, I.; Felisberto, B.H.; Pinto, V.D.; Campos, R.I.; Ribeiro, S.P. Territory and trophic cascading effects of the ant Azteca chartifex (Hymenoptera: Formicidae) in a tropical canopy. Myrmecological News 2022, 32, 103–113. [Google Scholar] [CrossRef]
- Delabie, J.H.C.; Mariano, C.S.F. Papel das formigas (Insecta: Hymenoptera : Formicidae) no controle biológico das pragas do cacaueiro na Bahia: síntese e limitações. In Proceedings of the International Cocoa Research Conference; Kota Kinabalu, Sabah, Malaysia, 2000; pp. 725–731.
- Medeiros, M.A. De; Fowler, H.G.; Delabie, J.H.C. O mosaico de formigas (Hymenoptera: Formicidae) em cacauais do Sul da Bahia. Cientifica 1995, 23, 291–300. [Google Scholar]
- Gouvea, J.B.S.; Silva, L.A.M.; Hori, M. 1. Fitogeografia. 2. Recursos Florestais. 3. Principais Vegetais Úteis. Comissão Executiva do Plano da Lavoura Cacaueira. Ilhéus. Ceplac/IICA. Diagnóstico Sócio-Econômico da Região Cacaueira; Ilheus, 1976; Vol. 7, 246 pp.
- Cassano, C.R.; Schroth, G.; Faria, D.; Delabie, J.H.C.; Bede, L. Landscape and farm scale management to enhance biodiversity conservation in the cocoa producing region of Southern Bahia, Brazil. Biodivers. Conserv. 2009, 18, 577–603. [Google Scholar] [CrossRef]
- Bishop, T.R.; Robertson, M.P.; van Rensburg, B.J.; Parr, C.L. Contrasting species and functional beta diversity in montane ant assemblages. J. Biogeogr. 2015, 42, 1776–1786. [Google Scholar] [CrossRef]
- Kaspari, M.; Weiser, M.D. The size–grain hypothesis and interspecific scaling in ants. Funct. Ecol. 1999, 13, 530–538. [Google Scholar] [CrossRef]
- Wakeham-Dawson, A. Ant architecture: The wonder, beauty and science of underground nests. Entomol. Mon. Mag. 2021, 157. [Google Scholar] [CrossRef]
- Römer, D.; Bollazzi, M.; Roces, F. Carbon dioxide sensing in an obligate insect-fungus symbiosis: CO2 preferences of leaf-cutting ants to rear their mutualistic fungus. PLoS One 2017, 12, e0174597. [Google Scholar] [CrossRef]
- Longino, J.T. Ants provide substrate for epiphytes. Selbyana 1986, 9, 100–103. [Google Scholar]
- Santos, J.C.; Del-Claro, K. Ecology and behaviour of the weaver ant Camponotus (Myrmobrachys) senex. J. Nat. Hist. 2009, 43, 1423–1435. [Google Scholar] [CrossRef]
- Tranter, C.; Hughes, W.O.H. A preliminary study of nest structure and composition of the weaver ant Polyrhachis (Cyrtomyrma) delecta (Hymenoptera: Formicidae). J. Nat. Hist. 2016, 50, 1197–1207. [Google Scholar] [CrossRef]
- Martius, C. Nest architecture of Nasutitermes termites in a white water floodplain forest in Central Amazonia, and a field key to species (Isoptera, Termitidae). Andrias 2001, 15, 163–171. [Google Scholar]
- Henrique-Simões, M.; Cuozzo, M.D.; Frieiro-Costa, F.A. Social wasps of Unilavras/Boqueirão Biological Reserve, Ingaí, state of Minas Gerais, Brazil. Check List 2011, 7, 656. [Google Scholar] [CrossRef]
- Schmolz, E.; Brüders, N.; Daum, R.; Lamprecht, I. Thermoanalytical investigations on paper covers of social wasps. Thermochim. Acta 2000, 361, 121–129. [Google Scholar] [CrossRef]
- Klingner, R.; Richter, K.; Schmolz, E.; Keller, B. The role of moisture in the nest thermoregulation of social wasps. Naturwissenschaften 2005, 92, 427–430. [Google Scholar] [CrossRef]
- Frouz, J. The effect of nest moisture on daily temperature regime in the nests of Formica polyctena wood ants. Insectes Soc. 2000, 47, 229–235. [Google Scholar] [CrossRef]
- Coenen-Stass, D.; Schaarschmidt, B.; Lamprecht, I. Temperature distribution and calorimetric determination of heat production in the nest of the wood ant, Formica polyctena (Hymenoptera, Formicidae). Ecology 1980, 61, 238–244. [Google Scholar] [CrossRef]
- Rosengren, R.; Fortelius, W.; Lindstrom, K.; Luther, A. Phenology and causation of nest heating and thermoregulation in red wood ants of the Formica rufa group studied in coniferous forest habitats in Southern Finland. Ann. Zool. Fennici 1987, 24, 147–155. [Google Scholar]
- Korb, J.; Linsenmair, K.E. Thermoregulation of termite mounds: what role does ambient temperature and metabolism of the colony play? Insectes Soc. 2000, 47, 357–363. [Google Scholar] [CrossRef]
- Pérez-Lachaud, G.; Lachaud, J.-P. Arboreal ant colonies as ‘hot-points’ of cryptic diversity for myrmecophiles: the weaver ant Camponotus sp. aff. textor and its interaction network with its associates. PLoS One 2014, 9, e100155. [Google Scholar] [CrossRef]
- Pérez-Lachaud, G.; Jahyny, B.J.B.; Ståhls, G.; Rotheray, G.; Delabie, J.H.C.; Lachaud, J.-P. Rediscovery and reclassification of the dipteran taxon Nothomicrodon Wheeler, an exclusive endoparasitoid of gyne ant larvae. Sci. Rep. 2017, 7, 45530. [Google Scholar] [CrossRef] [PubMed]
- Carvalho-Filho, F.S.; Barbosa, R.R.; Soares, M.M.M. Brakemyia, a new neotropical jackal fly genus of Milichiidae (Insecta: Diptera) associated with carton ant nest. Zool. Stud. 2023, 62, 1–11. [Google Scholar] [CrossRef]
- Wheeler, E. Polymorphism and division of labor in Azteca chartifex laticeps (Hymenoptera : Formicidae). J. Kansas Entomol. Soc. 1986, 59, 542–548. [Google Scholar]
- Longino, J.T. A taxonomic review of the genus Azteca (Hymenoptera: Formicidae) in Costa Rica and a global revision of the Aurita group. Zootaxa 2007, 1491, 1–63. [Google Scholar] [CrossRef]
- Clémencet, J.; Cournault, L.; Odent, A.; Doums, C. Worker thermal tolerance in the thermophilic ant Cataglyphis cursor (Hymenoptera, Formicidae). Insectes Soc. 2010, 57, 11–15. [Google Scholar] [CrossRef]
- O’Donnell, S.; Bulova, S.; Caponera, V.; Oxman, K.; Giladi, I. Species differ in worker body size effects on critical thermal limits in seed-harvesting desert ants (Messor ebeninus and M. arenarius). Insectes Soc. 2020, 67, 473–479. [Google Scholar] [CrossRef]
- Arnan, X.; Lázaro-González, A.; Beltran, N.; Rodrigo, A.; Pol, R. Thermal physiology, foraging pattern, and worker body size interact to influence coexistence in sympatric polymorphic harvester ants (Messor spp.). Behav. Ecol. Sociobiol. 2022, 76, 80. [Google Scholar] [CrossRef]
- Perez, R.; Benbachir, M.; Decroo, C.; Mascolo, C.; Wattiez, R.; Aron, S. Cataglyphis desert ants use distinct behavioral and physiological adaptations to cope with extreme thermal conditions. J. Therm. Biol. 2023, 111, 103397. [Google Scholar] [CrossRef]
Figure 1.
Map of South America highlighting the city of Ilhéus, located on the southern coast of the state of Bahia, Brazil. The map was created using public geospatial data to represent the study area where fauna associated with ant nest were sampled and analyzed.
Figure 1.
Map of South America highlighting the city of Ilhéus, located on the southern coast of the state of Bahia, Brazil. The map was created using public geospatial data to represent the study area where fauna associated with ant nest were sampled and analyzed.
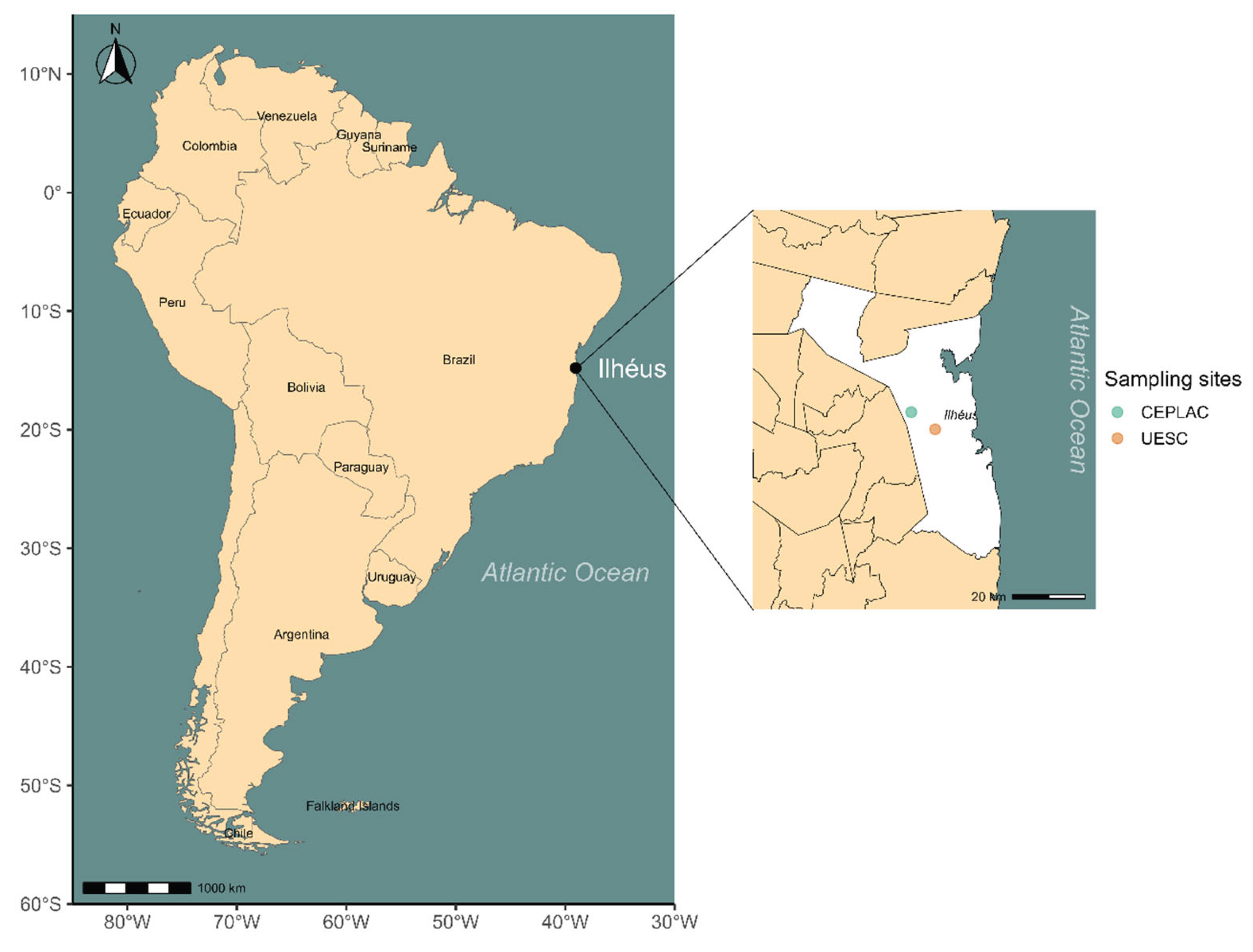
Figure 2.
Examples of Azteca chartifex spiriti nests located on the campus of the State University of Santa Cruz (UESC). (A) Vertical division of nests into sampling levels; white rectangles indicate the points at which nest material was collected at each level. (B) Installation of thermal sensors (TS) within the nests; white arrows indicate the positions of the two sensors (TS01 and TS02) installed on the nest structure.
Figure 2.
Examples of Azteca chartifex spiriti nests located on the campus of the State University of Santa Cruz (UESC). (A) Vertical division of nests into sampling levels; white rectangles indicate the points at which nest material was collected at each level. (B) Installation of thermal sensors (TS) within the nests; white arrows indicate the positions of the two sensors (TS01 and TS02) installed on the nest structure.

Figure 3.
Boxplots showing variation in moisture content across the four sampling levels in nests of Azteca chartifex spiriti. Boxes represent the median and interquartile range; whiskers indicate the range of the data while excluding outliers, and individual points denote discrepant observations.
Figure 3.
Boxplots showing variation in moisture content across the four sampling levels in nests of Azteca chartifex spiriti. Boxes represent the median and interquartile range; whiskers indicate the range of the data while excluding outliers, and individual points denote discrepant observations.
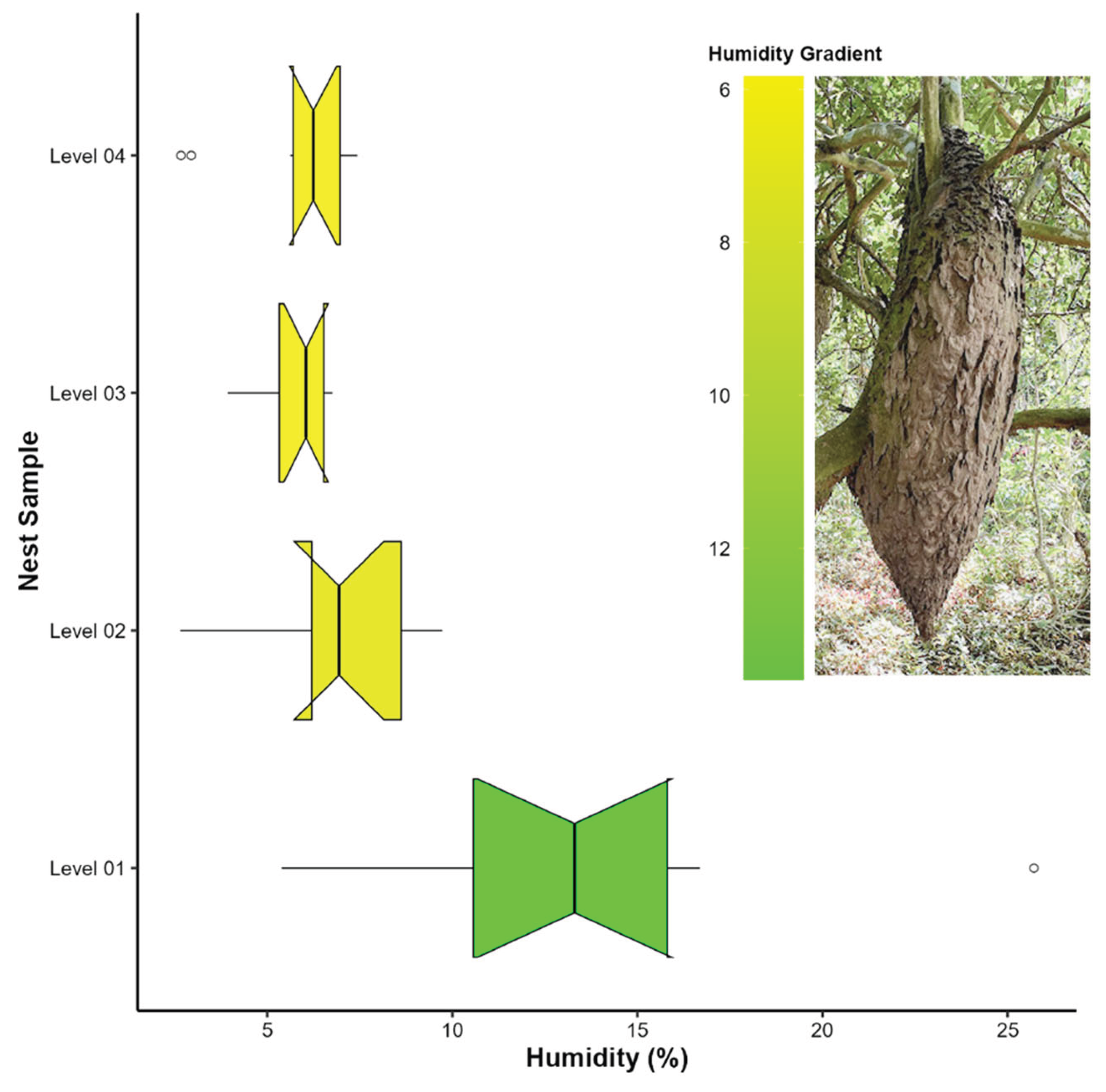
Figure 4.
Comparison of mean temperatures: (A) between the supporting base and the lower end of the nest; (B) between internal nest temperature and external ambient temperature; (C) among different periods of the day. Bars indicate the means and the corresponding error bars. Statistical tests: Student’s t-test for (A) and (B), and ANOVA for (C).
Figure 4.
Comparison of mean temperatures: (A) between the supporting base and the lower end of the nest; (B) between internal nest temperature and external ambient temperature; (C) among different periods of the day. Bars indicate the means and the corresponding error bars. Statistical tests: Student’s t-test for (A) and (B), and ANOVA for (C).
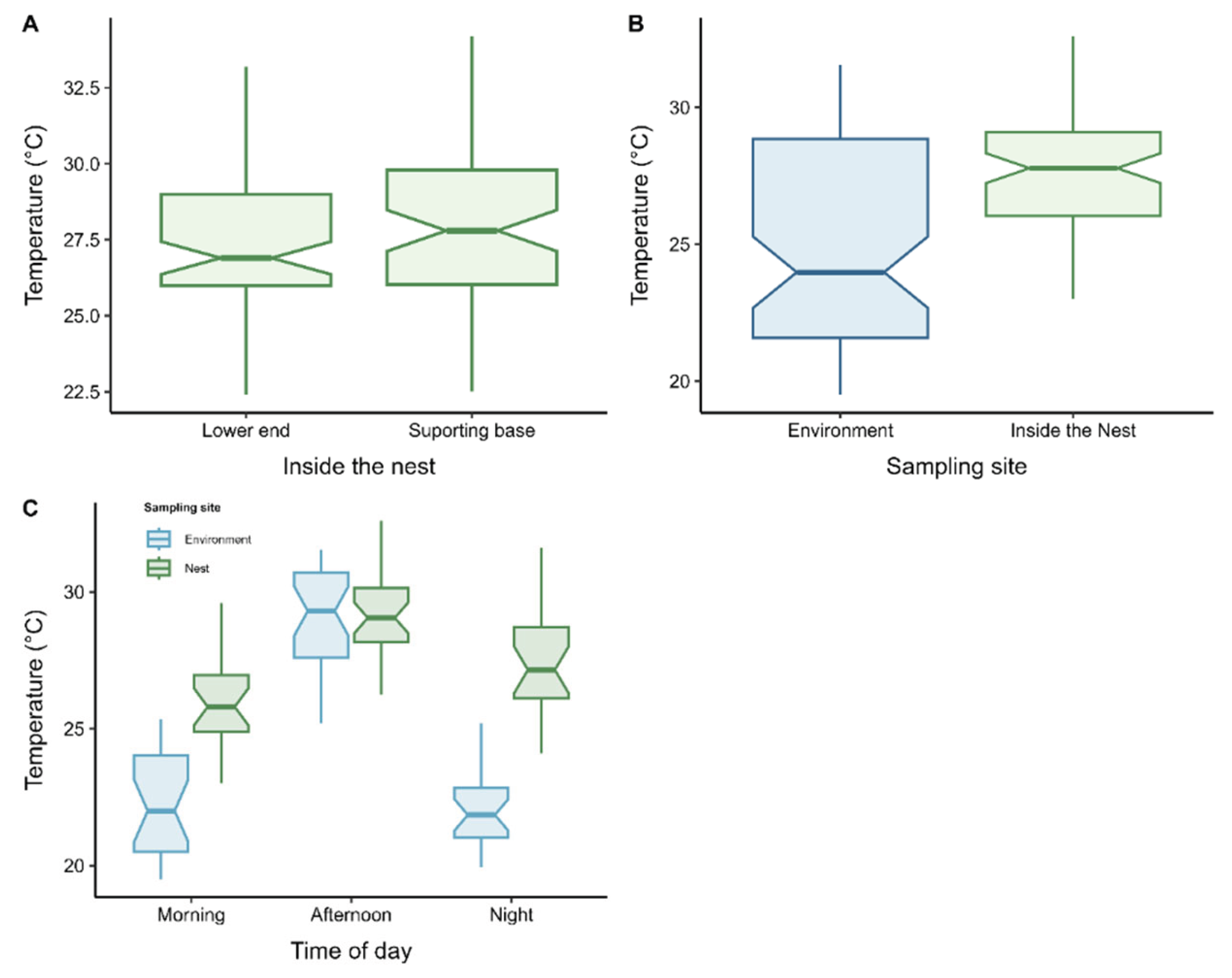
Figure 5.
Diurnal variation in temperature: comparison between the internal nest temperature and external ambient temperature. Vertical bars indicate the boundaries between the daytime period (shown in yellow) and the nighttime period (shown in gray). Twilight periods are quite short (approximately 20 minutes) in the study area.
Figure 5.
Diurnal variation in temperature: comparison between the internal nest temperature and external ambient temperature. Vertical bars indicate the boundaries between the daytime period (shown in yellow) and the nighttime period (shown in gray). Twilight periods are quite short (approximately 20 minutes) in the study area.
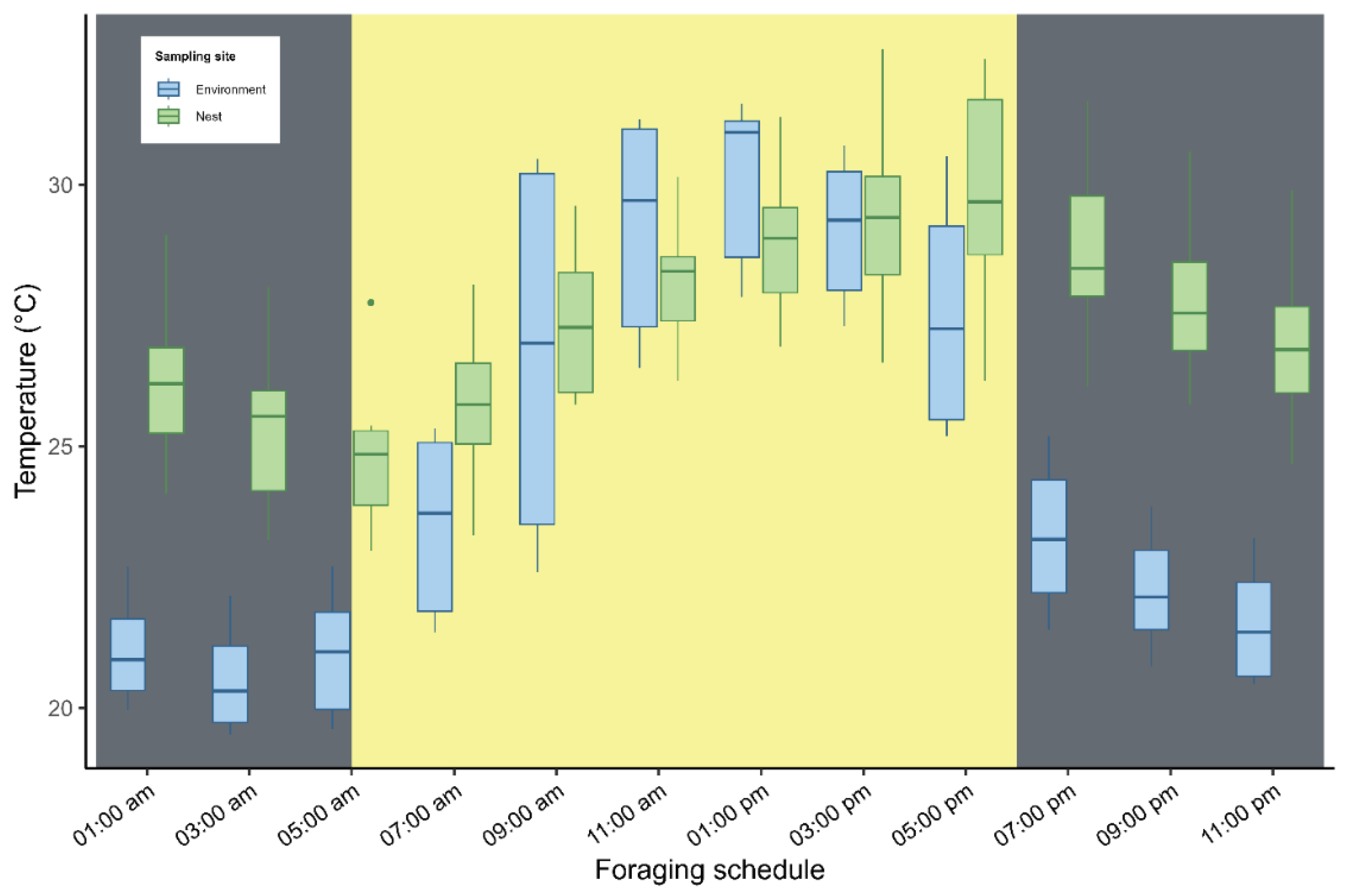
Figure 6.
Correlations among morphological parameters of Azteca chartifex spiriti workers: (A) Weber’s length vs. femur length; (B) Weber’s length vs. head width; (C) head width vs. femur length. Points represent individuals, and colors characterize the colonies analyzed (C01–C06).
Figure 6.
Correlations among morphological parameters of Azteca chartifex spiriti workers: (A) Weber’s length vs. femur length; (B) Weber’s length vs. head width; (C) head width vs. femur length. Points represent individuals, and colors characterize the colonies analyzed (C01–C06).
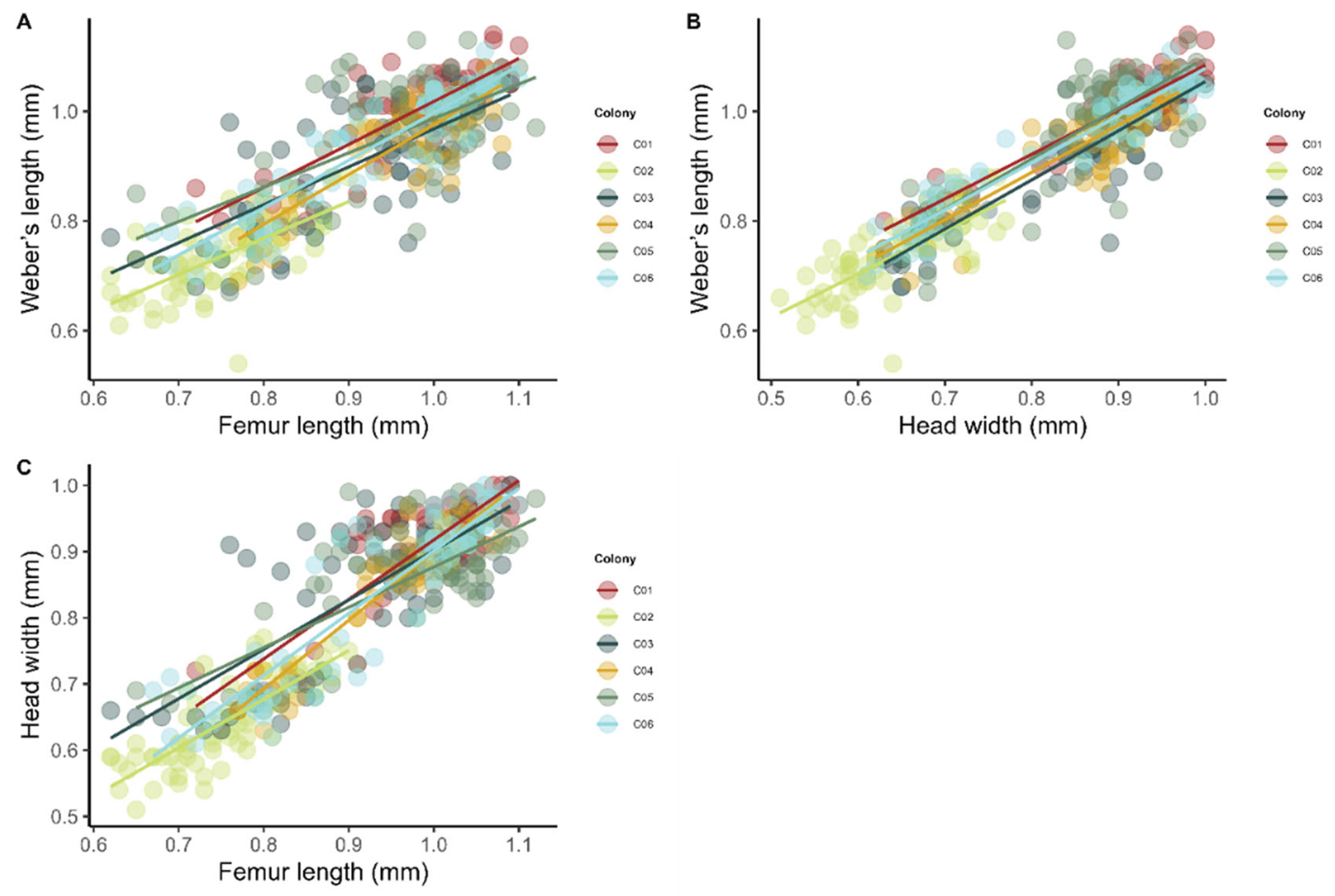
Figure 7.
Distribution of variation in morphological trait size of Azteca chartifex spiriti workers during foraging periods and across the daily cycle. (A-B) Weber’s length; (C-D) head width; (E-F) hind femur length. Each point represents an individual sampled in different activity periods; the lower horizontal bar indicates the alternation between the daytime period (yellow) and the nighttime period (gray).
Figure 7.
Distribution of variation in morphological trait size of Azteca chartifex spiriti workers during foraging periods and across the daily cycle. (A-B) Weber’s length; (C-D) head width; (E-F) hind femur length. Each point represents an individual sampled in different activity periods; the lower horizontal bar indicates the alternation between the daytime period (yellow) and the nighttime period (gray).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



