
Submitted:
20 November 2025
Posted:
20 November 2025
You are already at the latest version
Abstract
Rosemary (Salvia rosmarinus) has been linked to improvements in psychological wellbeing through cholinergic mechanisms. However, this study investigated whether individual differences in eye blink rate (EBR) and blink variability (EBV), which are proxies of dopaminergic activity and attentional control, influence the cognitive and mood-enhancing properties of a rosemary-containing drink. Forty-eight healthy adults completed a 3-stimulus odd-ball cognitive task under rosemary or control conditions, while vertical electrooculograms were recorded. Event-related brain potentials (ERPs) were also measured using the P3a component at the Cz scalp electrode as an additional index of dopaminergic activity. Subjective mood and arousal (alert, contented, calm) were collected pre- and post-task using Bond–Lader visual analogue scales. Reaction times during the task were modelled with ex-Gaussian parameters (μ, σ, τ). Rosemary ingestion led to increased alertness and contentedness following the task. Cognitive effects were moderated by blink metrics, with significant interactions between rosemary and blink metrics for mean reaction time μ and response variability σ. Rosemary also increased P3a amplitudes, indicative of dopaminergic contribution. The effects of rosemary on cognition and mood were moderated by individual blink profiles, indicating that baseline neurocognitive state plays a role. Although cholinergic accounts are well established, this study highlights the use of proxies of dopamine to investigate broader neurotransmitter involvement in rosemary's enhancing properties.
Keywords:
rosemary
; eye blinks
; dopamine
; acetylcholine
; attention
; arousal
; mood
; wellbeing
; ex-Gaussian
; P3a ERP
; attentional control
; cognitive flexibility
1. Introduction
Rosemary is a herb native to the Mediterranean region and has long been associated with cognitive enhancement and psychological wellbeing. Historically, rosemary was used in Egyptian burial rituals and symbolised fidelity and remembrance in ancient Greek ceremonies, including weddings and funerals (see [1,2]). The association with memory and cognition is not purely symbolic. In Hamlet, Ophelia remarks, "There’s rosemary, that’s for remembrance” (4.5.175), reflecting early cultural intuitions of its mnemonic effects [3]. Its presence in such traditions, alongside evidence of physical and psychological benefits, has contributed to its reputation as a cognitive enhancer [4]. Carnosic and rosmarinic acid are two of the active ingredients of rosemary, which have known antioxidant and anti-inflammatory properties. These compounds support the immune system, offer relief from pain, and are linked to cardiovascular protection [5]. Rosemary's anxiolytic and stimulatory effects have also been linked to reduced stress, improved emotional regulation, and particularly relevant here, cognitive facilitation [6]. In the present study, we aim to identify the specific mood and attentional components that improve during cognitive task engagement and examine the moderating effects of dopamine linked blink metrics.
Consider first acetylcholine (ACh), a neuromodulator central to attention and memory [7]. ACh is typically released from the basal forebrain, notably the nucleus basalis of Meynert, with widespread cholinergic projections targeting the cortex [8,9]. These projections support both synaptic and volume transmission, allowing broad modulation of cortical activity thought to be involved in attentional control, including the regions of the frontoparietal and default mode networks [10]. The active compounds of rosemary, including 1,8-cineole and carnosic acid, have been shown to inhibit acetylcholinesterase (AChE) and may increase ACh availability [11]. This finding is consistent with the action of pharmaceutical cholinesterase inhibitors used in the treatment of Alzheimer's disease [12]. Animal models also suggest rosemary may have anxiolytic and antidepressant effects alongside AChE inhibition properties [12]. These findings suggest cholinergic involvement. However, given rosemary's influence on attentional and affective processes, examining dopaminergic pathways may offer a theoretically grounded and potentially worthwhile route to examine the enhancing properties of rosemary further.
In addition to involvement in emotional processing, dopamine plays a critical role in the regulation of attention and executive function, particularly in the filtering of distractions and the processing of novelty [13]. Disruptions in dopamine signalling in the prefrontal cortex and basal ganglia may impair a variety of aspects of attention functioning and behaviour (see [14] for a review). This neurochemical framework has informed a range of pharmacological interventions. For instance, psychostimulants such as methylphenidate and amphetamines, commonly used in the treatment of ADHD, enhance dopaminergic transmission and improve attentional ability [15]. Similarly, dopaminergic agents have been explored in cognitive enhancement studies targeting episodic memory and attention in clinical populations, including Parkinson's disease [16] and schizophrenia [17]. Nutritional interventions have also demonstrated modulatory effects on dopamine. Tyrosine, a dietary precursor to dopamine, has been shown to influence cognitive performance under stress and fatigue [18]. Therefore, it is hardly surprising that compounds found in a variety of herbs, such as rosemary, may exert dopaminergic effects possibly by interacting with brain receptors. Specifically, rosemary's antidepressant-like properties have been linked to activity at dopamine D1 and D2 receptors, as well as noradrenergic and serotonergic systems [19]. These interactions suggest a broader neuromodulatory profile that may contribute to its cognitive and mood-related effects. From a neuropsychological perspective, eye blink rate (EBR) has emerged as a non-invasive proxy for central dopamine activity and is employed in the present study.
While the relationship is unclear, EBR has been shown to vary across clinical populations in ways that may reflect dopaminergic status. For example, elevated blink rates are often reported in schizophrenia, while reduced rates are typically observed in Parkinson's disease [20], which is consistent with underlying dopamine dysregulation. Our previous work on the enhancing properties of music, mind wandering, and creativity (e.g., [21]), alongside studies elsewhere [22], has established the use of blink metrics as indicators of dopaminergic function, attentional control, and emotional processing. Importantly, blink rate is sensitive to cognitive and motivational demands, and may increase during internally focused states involved in self-generated thoughts [23]; [24]. Blink variability (EBV), though less commonly discussed, may reflect transient shifts in attentional engagement but is included due to the partial exploratory nature of the present work.
The Current Study
The present study aimed to investigate the cognitive and mood-enhancing effects of rosemary ingestion, with a focus on the moderating role of eye blink metrics. Eye blinks were used as putative non-invasive proxies of dopamine function and attentional control, allowing us to explore individual differences in responsiveness to rosemary. Building on previous work linking rosemary to cholinergic mechanisms, this study examined whether dopaminergic tone influences cognitive performance and subjective mood. Reaction time data were analysed using ex-Gaussian modelling (Mu, Sigma, Tau) to provide a comprehensive examination of attentional engagement during task and mirrors previous work on Stroop task performance [25]. In addition, the P3a ERP component was included as a secondary and confirmatory index of dopaminergic activity to aid in the interpretation of the blink-related findings. While P3a is often linked to dopaminergic activity, it may also reflect broader physiological aspects of dopamine function beyond baseline activity. By integrating behavioural and electrophysiological measures, the study aimed to clarify how rosemary interacts with baseline neurocognitive states to influence attention, arousal and mood.
2. Methods
2.1. Participants
Forty-eight adults participated in the study (32 women; Mean age = 30.7; Range 18-63; SD = 11.2). All participants were right-handed and reported no history of neurological disorders. Screening excluded individuals with a current diagnosis of depression or anxiety, substance use, migraines, or any of the following: anaemia, heart disorder, high blood pressure, respiratory disorder, diabetes, pregnancy, history of seizures, or current use of prescribed, illicit, or herbal medication. Participants with food allergies or sensitivities were also excluded. Allocation to treatment condition was randomised prior to testing. Ethical approval was granted by the Department of Psychology Ethics Board at Northumbria University (Reference-882), and all procedures were conducted in accordance with the Declaration of Helsinki. Written informed consent was obtained from all participants. Further details and a description of part one of this program of work can be found in [26].
2.2. Treatment
Participants consumed either 330ml of rosemary water or plain still water. The rosemary drink was supplied by No1 Botanicals and contained both a volatile oil extract and a hydrolat derived from fresh rosemary via steam distillation. The extract included terpenes such as 1,8-cineole (0.025 mg/ml) and rosmarinic acid (0.13 mg/ml), while the hydrolat contained lower levels of terpenes (1,8-cineole at 0.012 mg/ml) and no rosmarinic acid. Production and analysis were carried out by Blue Sky Botanics, Castle Farm, Upton Bishop, Ross-on-Wye, UK.
To maintain blinding, participants were informed that both drinks had been matched for taste. Those in the rosemary condition were told the placebo had been flavoured to match, while those in the control condition were told the rosemary extract had been taste-neutralised. The researcher was also blind to condition allocation.
3. Measures
3.1. The 3-Stimulus Odd-Ball Task
Participants completed a 3-stimulus visual odd-ball task designed to assess attentional control [27]. Stimuli were presented using E-Prime software on a 17.5-inch monitor. The task involved discriminating between three types of stimuli: a frequent standard stimulus (green square, area = 16 cm²; 74% of trials), a rare target stimulus (red circle, area = 12.6 cm²; 13% of trials), and a novel distractor stimulus (green square, area = 256 cm²; 13% of trials). Each stimulus was displayed for 100 ms, followed by a variable interstimulus interval between 830 and 930 ms. Participants responded to target stimuli using the spacebar on a PC keyboard. A short practice block preceded the main task to ensure participants understood the nature of the task.
3.2. Electrooculograms (Eye Blinks)
Vertical electrooculograms (VEOG) were recorded using electrodes placed above and below the eye to capture blink activity. Participants were asked to minimise excessive blinking and movement during the task. Researchers responsible for blink identification were trained in the characteristics of ocular EEG data, including blink duration, waveform shape and tempo. Eye blink rate (EBR) was calculated as the total number of blinks occurring during the 8-minute odd-ball task. Eye blink variability (EBV) was assessed by dividing the task into eight 60-second intervals and calculating the standard deviation of blink counts across these intervals.
3.3. Event-Related Potentials
Although ERP data were recorded during the task, only the P3a (measured at the Cz scalp electrode) was used as a secondary dopamine metric, as the current paper focuses on blink metrics, subjective arousal and cognition. A Biosemi electrode cap (biosemi.com) with 32 channels of EEG recordings were employed to measure ERP's. Signals were digitised at a rate of 2048Hz, with a recording epoch of 1200ms. Full details of the methodology can be found in [26], where the neuroimaging aspects of this program of work are described.
3.4. Mood and Arousal Measures
Subjective mood and arousal were assessed using the Bond–Lader visual analogue scales. This measure comprises a series of bipolar items presented as 100 mm horizontal lines anchored by opposing adjectives (e.g. "alert–drowsy", "calm–excited", "contented–discontented"). Participants were asked to mark a point along each line that best represented their current state. Ratings were collected immediately before and after completion of the odd-ball task. Scoring involved the grouping into three composite dimensions: alertness, calmness, and contentedness. These composites were derived by averaging relevant items within each dimension, as outlined in the original Bond–Lader methodology [28]. The resulting scores were used to examine changes in subjective state across the testing session and to evaluate potential treatment effects.
4. Results
4.1. Subjective Arousal and Mood Ratings
Participants completed ratings of alertness, calmness, and contentedness before treatment and after the task. Paired-samples t-tests (1-tailed) were conducted to assess pre–post changes. In the rosemary condition, there was a significant increase in alertness from pre- to post-task (t(22) = –2.41, p = .013). Participants also reported feeling significantly more content after the task compared to baseline (t(22) = –2.63, p = .008). There was no significant change in calmness. In the control condition, no significant changes were observed for alertness or calmness. However, there was a significant increase in contentedness, with scores rising from pre- to post-task (t(22) = –1.97, p = .031). Overall, these results indicate that rosemary significantly enhanced subjective alertness and contentedness, while the control condition only showed a modest improvement in contentedness, likely due to task completion effects. Treatment effects are summarised in Table 1.
4.2. Blink Metrics (Total Rate and Variability)
An ANCOVA was conducted to examine whether the effect of condition (rosemary vs control) on mean response time μ was moderated by individual differences in EBR. The overall model was significant, F(3, 38) = 3.08, p = .039, partial η² = .20. There was a trend-level main effect of condition, F(1, 38) = 3.52, p = .068, partial η² = .09. The interaction between condition and blink rate was significant, F(1, 38) = 8.08, p = .007, partial η² = .18, indicating that the relationship between blink rate and mu differed depending on condition. Specifically, higher blink rates were associated with faster responses in the control group, but slower responses in the rosemary group, reflecting a reversal in slope across conditions (see Figure 1).
An ANCOVA was conducted to examine the effect of aroma drink condition (control vs. rosemary) on response time variability (σ), with EBR entered as a continuous between-subjects factor. The interaction between condition and blink rate was significant, F(1, 38) = 4.69, p = .037, η² = .110, indicating that the relationship between blink rate and σ differed by condition (see Figure 2), with the association restricted to rosemary.
A further ANCOVA revealed that the main effects of condition and blink rate, as well as their interaction, were not significant, indicating no meaningful differences in τ (reflecting lapses of attention; [24]) across conditions or as a function of blink rate. Similarly, an ANCOVA examining the proportion of hits revealed no significant main effect of treatment, nor a significant interaction with blink rate.
To explore whether P3a amplitude (measured at Cz scalp electrode) was associated with blink rate, an ANCOVA was conducted. The main effect of condition approached significance, F(1,34) = 3.77, p=.061, η² =.10, with higher P3a amplitudes in the rosemary condition.
The next series of ANCOVAs considered EBV. When considering mu, there were no significant main effects or interaction effects observed for mean response time μ when including blink variability in the model. An ANCOVA revealed a significant main effect of condition on σ, F(1, 38) = 4.92, p = .033, η² = .115, with greater response variability in the rosemary condition than control. Importantly, a significant interaction between condition and blink variability emerged, F(1, 38) = 4.83, p = .013, η² = .203, indicating that the effect of the drink on σ depended on individual differences in blink variability (see Figure 3).
An ANCOVA examining τ revealed no significant main effect of condition, nor a significant interaction with blink variability. Similarly, the ANCOVA examining hit rates and the P3a amplitude both revealed no significant main effect of condition, nor a significant interaction with blink variability.
4.3. Bivariate Correlations
Discussion
The present study tested not only the domains enhanced by rosemary but also whether these benefits depend on individual differences in spontaneous blinking, a proxy for dopaminergic tone and attentional control. By combining ex-Gaussian modelling of response times, subjective mood ratings, and electrophysiological measures of attentional orienting (P3a), we aimed to clarify whether rosemary exerts broad effects or whether its influence is better understood as state-dependent. The central finding was that rosemary's effects are not uniform across individuals but depend on blink profiles, supporting a state-dependent model of cognitive enhancement.
The results revealed that rosemary ingestion significantly enhanced subjective alertness and contentedness, consistent with previous behavioural and neurophysiological investigations (e.g. [29]). These changes occurred from baseline to post-task, indicating that rosemary promoted mood facilitation during active cognitive engagement. The control drink also produced a modest increase in contentedness, likely due to task completion and satisfaction, but did not influence alertness, which underscores the specificity of rosemary's impact. Rosemary has traditionally been associated with cholinergic mechanisms, with active constituents such as 1,8-cineole and rosmarinic acid inhibiting acetylcholinesterase and increasing cholinergic signalling. These mechanisms are well established in supporting memory and attentional processes. However, the observed enhancement in subjective alertness and contentedness may reflect broader arousal mechanisms, including increased sympathetic nervous system activity (see [30]). Indeed, rosemary and other botanicals such as peppermint [31] and citrus oils [32] have been shown to increase arousal and alertness. Recent longitudinal findings concur and suggest that nightly exposure to rotating botanical aromas, including rosemary, modulates central arousal pathways, potentially via noradrenergic and limbic system engagement [33]. The current findings support this on a self-report level, and future work can confirm whether the subjective experience coincides with central arousal activation.
The main consideration in the present work was the moderating effect of blink dynamics. Our ANCOVAs revealed a significant rosemary × blink rate interaction for μ and σ, indicating that individual differences in blink rate moderate rosemary's effects on response speed and variability. Specifically, participants with higher blink rates showed slower and more variable responses under rosemary, possibly reflecting enhanced dopaminergic sensitivity. The reversal in blink–μ slope pattern may reflect two overlapping processes. First, individuals with lower blink rates, potentially indexing reduced dopaminergic tone, may have had greater scope for improvement, consistent with inverted-U models of neuromodulation and enhancement (e.g. glucose facilitation; [34]). While a higher blink rate is associated with greater cognitive flexibility [35], its relationship with performance follows an inverted U-shaped curve, where an extremely high blink rate is linked to distractibility, and an extremely low blink rate to perseveration [36]. Second, the crossover pattern does not reflect impaired performance, but rather a shift in cognitive strategy. As slower responses can accompany more diffuse or exploratory attentional states, rosemary may have modulated the balance between focused and flexible cognition, particularly in high-blink individuals. Riby and colleagues (e.g. [26]) argue that such profiles may reflect increased engagement with task-irrelevant as well as relevant information. Previous evidence supports the notion that blink rate reflects modulation between task-oriented and default-mode attention allocation, reflecting updating of information and transitions between attentional phases [37]. Rosemary may therefore modulate attentional control not by enhancing speed or consistency, but by altering the balance between focused and diffuse cognitive states. No significant interactions were observed for τ or hit rate, reinforcing the specificity of these blink condition effects.
While blink rate is the better-established marker of dopaminergic function, EBV is now recognised as a valuable measure in its own right. Considering that overall blink rate has complex temporal relationships with task phases [38], it is valuable to account for blink variability. In the current data, blink variability moderated the effect of rosemary on σ only. It demonstrated that individuals with greater EBV exhibited increased response variability in the rosemary condition, consistent with a shift toward more flexible attentional control. EBV reflects fluctuations in control and arousal, providing an index of attentional stability over time. A key demonstration comes from [39], who showed that resting EBV was positively correlated with IQ and that high-IQ participants displayed greater variability than those with lower scores. Our study builds on this by demonstrating that rosemary's effects are moderated by baseline EBV, while more broadly advancing the use of blink metrics in botanical and olfactory cognitive neuroscience. Considering rosemary’s involvement in attention regulation through acetylcholine availability, exploring EBV provides relevant insight into the complexity of attentional mechanisms.
Correlational analyses (Table 2 and Table 3) showed consistent links between blink measures, mood, and task performance across both treatment conditions. Though exploratory, these patterns complement the ANCOVA results and suggest that rosemary modulated how blink behaviour relates to attentional control.
The electrophysiological data offer further support for a state-dependent interpretation. Rosemary ingestion was associated with increased P3a amplitude at Cz, a component linked to involuntary attentional orienting and novelty detection. Polich's integrative model [27] attributes P3a to frontal–central engagement with salient stimuli, modulated by dopaminergic activity in prefrontal and cingulate regions. Its amplitude varies with neurochemical state, task relevance, and individual differences in control. Reduced P3a is observed in conditions marked by dopaminergic dysfunction (e.g., Parkinson's, schizophrenia), while elevated amplitudes have been linked to novelty seeking and intact executive function. In this light, rosemary's enhancement of P3a may reflect increased dopaminergic facilitation of attentional orienting, particularly in individuals with greater baseline flexibility as indexed by EBV. Notably, P3a amplitude did not correlate with either EBR or EBV (see Table 2 and Table 3), supporting the view that blink metrics and ERP components index distinct facets of dopaminergic function. Blinks likely reflect tonic attentional readiness over extended intervals, while P3a captures phasic activity. This is consistent with evidence that blink rates, unlike P3a, are not stimulus-dependent and reflect broader primary consciousness phenomena and attention mode modulation [37]. This dissociation underscores the value of convergent physiological markers in characterising individual responsiveness to botanical interventions.
The present findings underscore the importance of jointly considering behavioural performance and physiological responses when evaluating the cognitive effects of botanical interventions. As previously discussed, cognitive performance has a complex relationship with dopamine, and thus blink rate as an indicator of dopaminergic activity. The current literature also focuses extensively on overall blink rates, leaving room for better understanding of how blink rate variability fluctuates in response to phases of attention, stimulus encoding and exposure to interventions. Future research should also utilise working memory tasks to explore dopamine-modulated attention as a ‘gate-switching’ mechanism for memory encoding and consolidation. Eye blink rate has been linked to shifts between updating and maintenance mode during working memory performance [40]. In light of the current findings, rosemary may offer a low-cost, accessible tool for flexibly engaging and disengaging task-oriented attention. In realistic, ecologically valid settings such as the workplace, attention fluctuates in relation to task type, rote work and context [41]. As such, cognitive focus is vulnerable to habituation and fatigue, which is reflected in longer intervals between blinks [42]. Botanical interventions, such as rosemary, may be particularly useful for applied uses beyond the laboratory, where sustaining attention over prolonged periods would benefit from a flexible ability to shift between attentional states.
Lastly, the relationship between rosemary exposure and blink rates can inform a wellbeing-oriented approach to attention in applied practice. In a laboratory context, mind-wandering and deviations in attention away from task stimuli lower performance. Indeed, mind-wandering is linked to decoupling attention from perceptual information and engaging with internal, default-mode responses [43]. Task-unrelated thoughts are associated with higher eye blink rates, suggesting that increased dopaminergic activity corresponds to a diffuse centre of attention [24]. However, ecologically valid contexts that require sustained focus also inevitably involve dynamic changes to attention, including managing interruptions and own affective states alongside work-relevant tasks [44]. Thus, stable and highly concentrated attentional states may be difficult to sustain, while flexible and diffuse attention would permit shifting focus as needed, engaging and decoupling self-referential and task-oriented thought, thereby ensuring a smoother transition between states. This can also be related to the construct of mindfulness, a state of consciousness oriented toward the here and now. Mindfulness is characterised by breadth of attention, i.e. receptivity to a wide range of stimuli, which can be detrimental to performance during static tasks, but helpful during dynamic work responsibilities that require adapting to changes and improvisation [44]. In the context of mindfulness in the workplace, rosemary-modulated shifts in attention can provide a suitable balance of scope and focus, allowing for the desired flexibility and consequent improvements in wellbeing. This finding is consistent with the present study’s results, which indicate that participants in the rosemary condition feel more content and alert even after completing an attention-demanding task, and offers promising avenues for maintaining a balance between performance-related and wellbeing concerns in botanical intervention research.
5. Conclusions
This study demonstrates that rosemary’s cognitive and mood-enhancing effects are state-dependent, varying systematically with individual blink profiles that index dopaminergic tone and attentional control. Theoretically, this extends existing cholinergic accounts by introducing a dual-neuromodulator framework in which rosemary influences both acetylcholine- and dopamine-related mechanisms of attention and arousal. The integration of blink dynamics, ex-Gaussian modelling, and electrophysiological measures offers a novel and reproducible approach for examining how botanical interventions interact with baseline neurocognitive state in cognitive enhancement.
Author Contributions
Conceptualization, L.R., D.K., S.F., S.Fo., and M.M.; Methodology, L.R., D.K., and S.Fo.; Data curation, D.K. and S.Fo.; Formal analysis, L.R.; Writing-original draft preparation, L.R.; Writing- review and editing, all authors; Funding acquisition, L.R. and M.M.; Project administration, L.R.
Funding
This project was supported by a funding award to MM and LR from No 1 Botanicals, Bischheim House, 19-20 Berners Street, London, W1T 3NW. The funders did not play a role in the study design, data collection, analysis, decision to publish or the preparation of the manuscript.
Data Availability Statement
The data supporting the findings of this study will be made available via the Northumbria Figshare repository upon publication (DOI to be provided upon acceptance).
References
- Ribeiro-Santos, R.; Carvalho-Costa, D.; Cavaleiro, C.; Costa, H.S.; Albuquerque, T.G.; Castilho, M.C.; Ramos, F.; Melo, N.R.; Sanches-Silva, A. A novel insight on an ancient aromatic plant: The rosemary (Rosmarinus officinalis L.). Trends in Food Science & Technology 2015, 45, 355–368. [Google Scholar] [CrossRef]
- Hanson, J.R. Rosemary, the beneficial chemistry of a garden herb. Science progress 2016, 99, 83–91. [Google Scholar]
- Shakespeare, W. Hamlet. 1603.
- Moss, M.; Cook, J.; Wesnes, K.; Duckett, P. Aromas of rosemary and lavender essential oils differentially affect cognition and mood in healthy adults. International Journal of Neuroscience 2003, 113, 15–38. [Google Scholar] [CrossRef]
- Singletary, K. Rosemary: an overview of potential health benefits. Nutrition Today 2016, 51, 102–112. [Google Scholar]
- Hongratanaworakit, T. Simultaneous aromatherapy massage with rosemary oil on humans. Scientia Pharmaceutica 2009, 77, 375–388. [Google Scholar] [CrossRef]
- Hasselmo, M.E. The role of acetylcholine in learning and memory. Current opinion in neurobiology 2006, 16, 710–715. [Google Scholar] [PubMed]
- Mesulam, M.-M. The cholinergic innervation of the human cerebral cortex. Progress in brain research 2004, 145, 67–78. [Google Scholar] [PubMed]
- Ballinger, E.C.; Ananth, M.; Talmage, D.A.; Role, L.W. Basal forebrain cholinergic circuits and signaling in cognition and cognitive decline. Neuron 2016, 91, 1199–1218. [Google Scholar] [CrossRef] [PubMed]
- Sarter, M.; Gehring, W.J.; Kozak, R. More attention must be paid: the neurobiology of attentional effort. Brain research reviews 2006, 51, 145–160. [Google Scholar]
- Savelev, S.; Okello, E.; Perry, N.; Wilkins, R.; Perry, E. Synergistic and antagonistic interactions of anticholinesterase terpenoids in Salvia lavandulaefolia essential oil. Pharmacology Biochemistry and Behavior 2003, 75, 661–668. [Google Scholar] [CrossRef]
- Colovic, M.B.; Krstic, D.Z.; Lazarevic-Pasti, T.D.; Bondzic, A.M.; Vasic, V.M. Acetylcholinesterase inhibitors: pharmacology and toxicology. Current neuropharmacology 2013, 11, 315–335. [Google Scholar] [CrossRef]
- Duszkiewicz, A.J.; McNamara, C.G.; Takeuchi, T.; Genzel, L. Novelty and dopaminergic modulation of memory persistence: a tale of two systems. Trends in neurosciences 2019, 42, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Matzel, L.D.; Sauce, B. A multi-faceted role of dual-state dopamine signaling in working memory, attentional control, and intelligence. Frontiers in Behavioral Neuroscience 2023, 17, 1060786. [Google Scholar] [CrossRef]
- Pitzianti, M.B.; Spiridigliozzi, S.; Bartolucci, E.; Esposito, S.; Pasini, A. New insights on the effects of methylphenidate in attention deficit hyperactivity disorder. Frontiers in Psychiatry 2020, 11, 531092. [Google Scholar] [CrossRef] [PubMed]
- Stanzione, A.; Melchiori, F.M.; Costa, A.; Leonardi, C.; Scalici, F.; Caltagirone, C.; Carlesimo, G.A. Dopaminergic Treatment and Episodic Memory in Parkinson’s Disease: A Meta-analysis of the Literature. Neuropsychology Review 2024, 1–22. [Google Scholar]
- Howes, O.D.; Kapur, S. The dopamine hypothesis of schizophrenia: version III—the final common pathway. Schizophrenia bulletin 2009, 35, 549–562. [Google Scholar] [CrossRef]
- Jongkees, B.J.; Hommel, B.; Kühn, S.; Colzato, L.S. Effect of tyrosine supplementation on clinical and healthy populations under stress or cognitive demands—A review. Journal of psychiatric research 2015, 70, 50–57. [Google Scholar] [CrossRef]
- Rahbardar, M.G.; Hosseinzadeh, H. Therapeutic effects of rosemary (Rosmarinus officinalis L.) and its active constituents on nervous system disorders. Iranian journal of basic medical sciences 2020, 23, 1100. [Google Scholar]
- Reddy, V.C.; Patel, S.V.; Hodge, D.O.; Leavitt, J.A. Corneal sensitivity, blink rate, and corneal nerve density in progressive supranuclear palsy and Parkinson disease. Cornea 2013, 32, 631–635. [Google Scholar] [CrossRef]
- Riby, L.M.; Fenwick, S.K.; Kardzhieva, D.; Allan, B.; McGann, D. Unlocking the beat: Dopamine and eye blink response to classical music. NeuroSci 2023, 4, 152–163. [Google Scholar] [CrossRef]
- Müller, J.; Dreisbach, G.; Brocke, B.; Lesch, K.-P.; Strobel, A.; Goschke, T. Dopamine and cognitive control: The influence of spontaneous eyeblink rate, DRD4 exon III polymorphism and gender on flexibility in set-shifting. Brain research 2007, 1131, 155–162. [Google Scholar] [CrossRef]
- Hollander, J.; Huette, S. Extracting blinks from continuous eye-tracking data in a mind wandering paradigm. Consciousness and Cognition 2022, 100, 103303. [Google Scholar] [CrossRef]
- Riby, L.M.; Marr, L.; Barron-Millar, L.; Greer, J.; Hamilton, C.J.; McGann, D.; Smallwood, J. Elevated blink rates predict mind wandering: Dopaminergic insights into attention and task focus. Journal of Integrative Neuroscience 2025, 24, 26508. [Google Scholar] [CrossRef]
- Parris, B.A.; Dienes, Z.; Hodgson, T.L. Application of the ex-Gaussian function to the effect of the word blindness suggestion on Stroop task performance suggests no word blindness. Frontiers in psychology 2013, 4, 647. [Google Scholar] [CrossRef]
- Riby, L.M.; Edwards, S.; McDonald, H.; Moss, M. The impact of a rosemary containing drink on event-related potential neural markers of sustained attention. PLoS One 2023, 18, e0286113. [Google Scholar] [CrossRef]
- Polich, J. Updating P300: an integrative theory of P3a and P3b. Clinical neurophysiology 2007, 118, 2128–2148. [Google Scholar] [CrossRef] [PubMed]
- Bond, A.; Lader, M. The use of analogue scales in rating subjective feelings. British Journal of Medical Psychology 1974, 47, 211–218. [Google Scholar] [CrossRef]
- Sayorwan, W.; Ruangrungsi, N.; Piriyapunyporn, T.; Hongratanaworakit, T.; Kotchabhakdi, N.; Siripornpanich, V. Effects of inhaled rosemary oil on subjective feelings and activities of the nervous system. Scientia pharmaceutica 2012, 81, 531. [Google Scholar] [CrossRef] [PubMed]
- Sattayakhom, A.; Wichit, S.; Koomhin, P. The effects of essential oils on the nervous system: A scoping review. Molecules 2023, 28, 3771. [Google Scholar] [CrossRef] [PubMed]
- Moss, M.; Hewitt, S.; Moss, L.; Wesnes, K. Modulation of cognitive performance and mood by aromas of peppermint and ylang-ylang. International Journal of Neuroscience 2008, 118, 59–77. [Google Scholar] [CrossRef]
- Hawiset, T.; Sriraksa, N.; Somwang, P.; Inkaew, P. Effect of orange essential oil inhalation on mood and memory in female humans. Journal of Physiological and Biomedical Sciences 2016, 29, 5–11. [Google Scholar]
- Woo, C.C.; Miranda, B.; Sathishkumar, M.; Dehkordi-Vakil, F.; Yassa, M.A.; Leon, M. Overnight olfactory enrichment using an odorant diffuser improves memory and modifies the uncinate fasciculus in older adults. Frontiers in Neuroscience 2023, 17, 1200448. [Google Scholar] [CrossRef] [PubMed]
- Riby, L.M. The impact of age and task domain on cognitive performance: a meta-analytic review of the glucose facilitation effect. Brain Impairment 2004, 5, 145–165. [Google Scholar] [CrossRef]
- Jongkees, B.J.; Colzato, L.S. Spontaneous eye blink rate as predictor of dopamine-related cognitive function—A review. Neuroscience & Biobehavioral Reviews 2016, 71, 58–82. [Google Scholar]
- Dreisbach, G.; Müller, J.; Goschke, T.; Strobel, A.; Schulze, K.; Lesch, K.-P.; Brocke, B. Dopamine and cognitive control: the influence of spontaneous eyeblink rate and dopamine gene polymorphisms on perseveration and distractibility. Behavioral neuroscience 2005, 119, 483. [Google Scholar] [CrossRef]
- Callara, A.L.; Greco, A.; Scilingo, E.P.; Bonfiglio, L. Neuronal correlates of eyeblinks are an expression of primary consciousness phenomena. Scientific Reports 2023, 13, 12617. [Google Scholar] [CrossRef]
- Ortega, J.; Plaska, C.R.; Gomes, B.A.; Ellmore, T.M. Spontaneous eye blink rate during the working memory delay period predicts task accuracy. Frontiers in Psychology 2022, 13, 788231. [Google Scholar] [CrossRef] [PubMed]
- Paprocki, R.; Lenskiy, A. What does eye-blink rate variability dynamics tell us about cognitive performance? Frontiers in human neuroscience 2017, 11, 620. [Google Scholar] [CrossRef] [PubMed]
- Rac-Lubashevsky, R.; Slagter, H.A.; Kessler, Y. Tracking real-time changes in working memory updating and gating with the event-based eye-blink rate. Scientific reports 2017, 7, 2547. [Google Scholar] [CrossRef]
- Mark, G.; Iqbal, S.T.; Czerwinski, M.; Johns, P. Bored mondays and focused afternoons: the rhythm of attention and online activity in the workplace. . In Proceedings of the Proceedings of the SIGCHI Conference on Human Factors in Computing Systems, 2014; pp. 3025-3034.
- Veltman, J.; Gaillard, A. Physiological workload reactions to increasing levels of task difficulty. Ergonomics 1998, 41, 656–669. [Google Scholar] [CrossRef]
- Barron, E.; Riby, L.M.; Greer, J.; Smallwood, J. Absorbed in thought: the effect of mind wandering on the processing of relevant and irrelevant events. Psychol Sci 2011, 22, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Dane, E. Paying attention to mindfulness and its effects on task performance in the workplace. Journal of management 2011, 37, 997–1018. [Google Scholar] [CrossRef]
Figure 1.
Relationship between EBR and Mu (μ) for control and rosemary conditions.
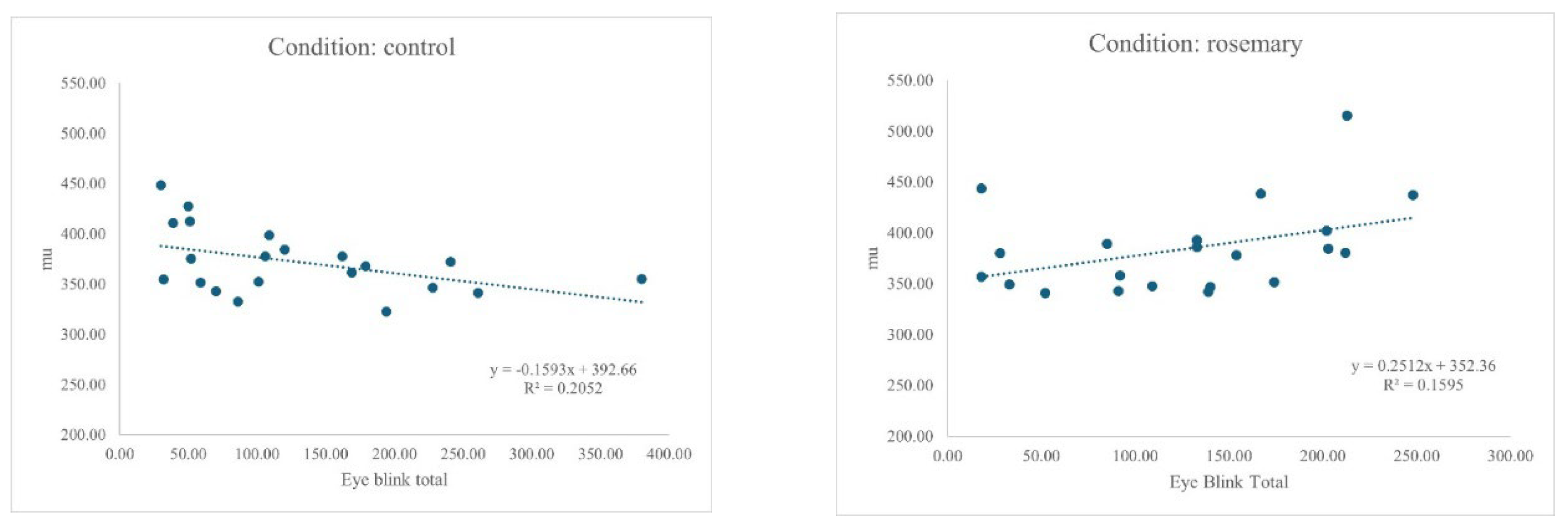
Figure 2.
Relationship between EBR and Sigma (σ) for control and rosemary conditions.
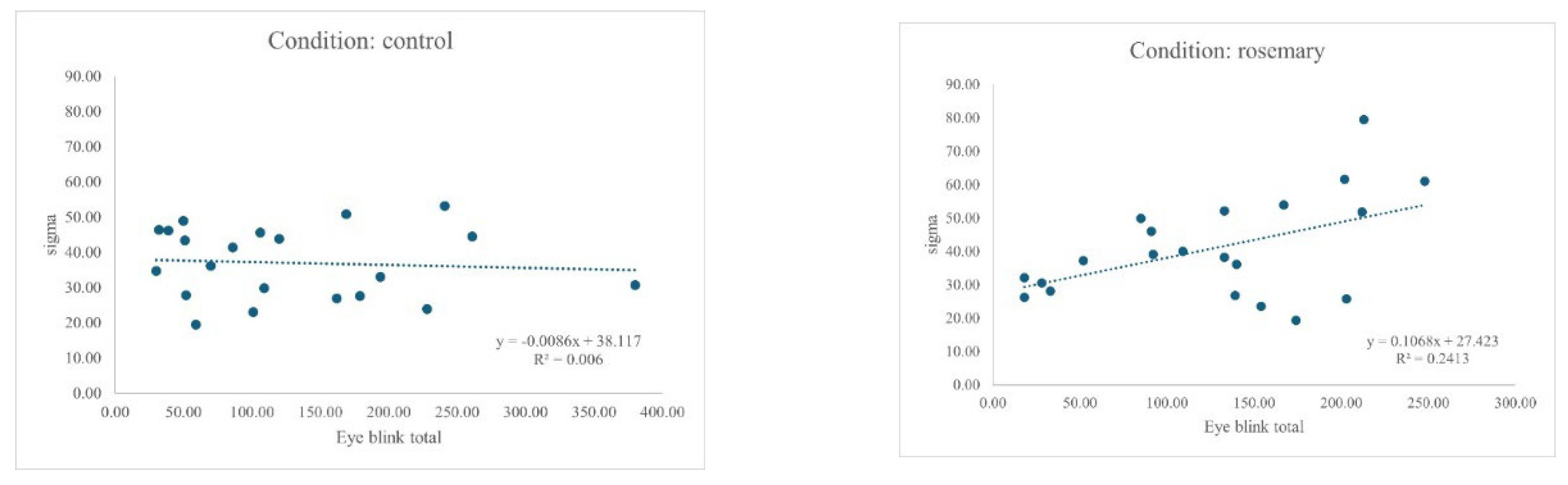
Figure 3.
Relationship between EBV and Sigma (σ) for control and rosemary condition.
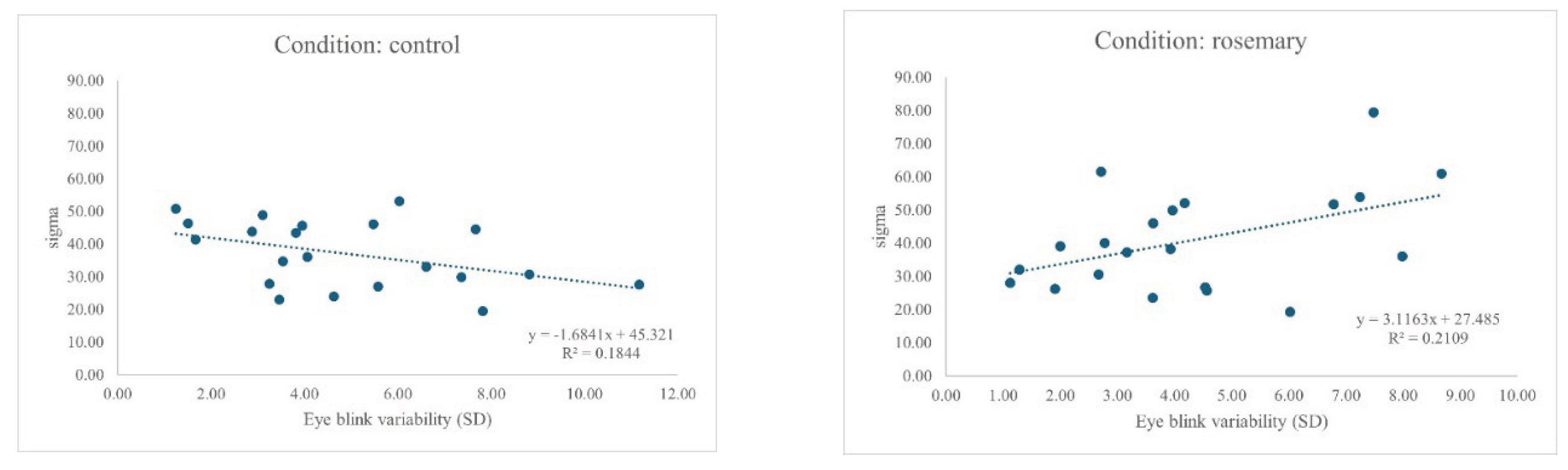

Table 1.
Means and Standard Deviations for Key Variables by Condition (Control vs. Rosemary), with Cohen's d Effect Sizes.
Table 1.
Means and Standard Deviations for Key Variables by Condition (Control vs. Rosemary), with Cohen's d Effect Sizes.
| Measure | Control M (SD) | Rosemary M (SD) | Cohen's d |
| Eye Blink Rate (EBR) | 120.24 ± 90.21 | 122.91 ± 67.77 | 0.03 |
| Blink Variability (EBV) | 4.69 ± 2.52 | 4.14 ± 2.21 | -0.23 |
| Alertness (Pre) | 35.11 ± 12.08 | 36.91 ± 11.56 | 0.15 |
| Calmness (Pre) | 30.91 ± 14.43 | 33.54 ± 17.75 | 0.17 |
| Contentedness (Pre) | 24.22 ± 9.48 | 26.67 ± 10.97 | 0.25 |
| Alertness (Post) | 38.32 ± 18.48 | 45.06 ± 18.01 | 0.37 |
| Calmness (Post) | 32.92 ± 13.99 | 32.87 ± 14.00 | -0.00 |
| Contentedness (Post) | 28.34 ± 10.94 | 30.97 ± 15.06 | 0.19 |
| Mu (μ) | 372.04 ± 32.40 | 383.98 ± 44.06 | 0.31 |
| Sigma (σ) | 37.01 ± 10.15 | 40.87 ± 15.24 | 0.30 |
| Tau (τ) | 33.86 ± 12.74 | 36.81 ± 10.61 | 0.25 |
| Hits | 75.75 ± 21.53 | 78.37 ± 14.67 | 0.13 |
| P3a Amplitude (Cz) | 4.79 ± 2.86 | 6.44 ± 2.89 | 0.63 |
Table 2.
Pearson's correlations (control condition) between blink proxies (total and variability), alertness (alert, calm and content), cognitive performance (μ, τ, σ and hits) and the P3a ERP (Cz scalp electrode).
Table 2.
Pearson's correlations (control condition) between blink proxies (total and variability), alertness (alert, calm and content), cognitive performance (μ, τ, σ and hits) and the P3a ERP (Cz scalp electrode).
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | |
| 1. Eye blink total | 1 | .588** | .555** | .232 | .316 | .309 | .051 | .063 | -.453* | -.078 | .042 | -.281 | .176 |
| 2. Eye blink variability | .588** | 1 | .462* | .164 | .302 | .416* | .174 | .243 | -.138 | -.429 | -.062 | .114 | -.319 |
| 3. Alertness (pre) | .555** | .462* | 1 | -.117 | .249 | .692** | -.132 | .266 | -.296 | -.428 | .396 | .857 | .498 |
| 4. Calmness (pre) | .232 | .164 | -.117 | 1 | .237 | .048 | .220 | -.049 | .084 | -.063 | -.342 | .162 | -.086 |
| 5. Contentedness (pre) | .316 | .302 | .249 | .237 | 1 | .496* | .265 | .267 | -.312 | -.480* | -.061 | -.113 | -.040 |
| 6. Alertness (post) | .309 | .416* | .692** | .048 | .496* | 1 | -.133 | .572** | -.146 | -.234 | -.330 | -.036 | .424 |
| 7. Calmness (post) | .051 | .174 | -.132 | .220 | .265 | -.133 | 1 | .511* | -.197 | -.197 | -.438 | .205 | .446 |
| 8. Contentedness (post) | .063 | .243 | .243 | .266 | .267 | .572** | .511* | 1 | -.269 | -.104 | -.400 | -.314 | .436 |
| 9. mu (μ) | -.453* | -.138 | -.296 | .084 | -.312 | -.146 | -.197 | -.269 | 1 | .242 | .136 | .448* | -.252 |
| 10. sigma (σ) | -.078 | -.429 | -.428 | -.063 | -.480* | -.234 | -.197 | -.104 | .242 | 1 | -.002 | -.250 | .262 |
| 11. tau (τ) | .042 | -.062 | .396 | -.342 | -.061 | -.330 | -.438 | -.400 | .136 | -.002 | 1 | .046 | -.163 |
| 12. hits | -.281 | .114 | .857 | .162 | -.113 | -.036 | .205 | -.314 | .448* | -.250 | .046 | 1 | -.559* |
| 13. P3a ERP | .176 | -.319 | .498 | -.086 | -.040 | .424 | .446 | .436 | -.252 | .262 | -.163 | -.559* | 1 |
Note: *p<.05, **p<.01.
Table 3.
Pearson's correlations (rosemary condition) between blink proxies (total and variability), alertness (alert, calm and content), cognitive performance (μ, τ, σ and hits) and the P3a ERP (Cz scalp electrode).
Table 3.
Pearson's correlations (rosemary condition) between blink proxies (total and variability), alertness (alert, calm and content), cognitive performance (μ, τ, σ and hits) and the P3a ERP (Cz scalp electrode).
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | |
| 1. Eye blink total | 1 | .776** | .228 | .201 | .158 | .056 | .092 | .077 | .399 | .491* | .118 | .001 | -.227 |
| 2. Eye blink variability | .776** | 1 | .215 | .262 | .426* | .221 | -.049 | .306 | .388 | .459* | .096 | .345 | -.310 |
| 3. Alertness (pre) | .228 | .215 | 1 | .337 | .467* | .465* | -.275 | .475* | .242 | .388 | .104 | -.091 | -.251 |
| 4. Calmness (pre) | .201 | .262 | .337 | 1 | .826** | .375 | .498* | .743** | .291 | .433 | .290 | .323 | .138 |
| 5. Contentedness (pre) | .158 | .426* | .467* | .826** | 1 | .469* | .267 | .865** | .302 | .371 | .188 | .388 | -.095 |
| 6. Alertness (post) | .056 | .221 | .465* | .375 | .469* | 1 | -.043 | .739** | -.038 | -.134 | -.117 | .195 | .015 |
| 7. Calmness (post) | .092 | -.049 | -.275 | .498* | .267 | -.043 | 1 | .241 | .041 | .155 | -.149 | -.221 | .275 |
| 8. Contentedness (post) | .077 | .306 | .475* | .743** | .865** | .739** | .241 | 1 | .023 | .182 | .182 | .318 | .019 |
| 9. mu (μ) | .399 | .388 | .242 | .291 | .302 | -.038 | .155 | .182 | 1 | .691** | -.004 | .181 | -.026 |
| 10. sigma (σ) | .491* | .459* | .388 | .433 | .371 | -.134 | .155 | .182 | .691** | 1 | .108 | .258 | -.032 |
| 11. tau (τ) | .118 | .096 | .104 | .290 | .188 | -.117 | -.149 | .182 | -.004 | .108 | 1 | .531* | -.247 |
| 12. hits | .001 | .345 | -.091 | .323 | .388 | .195 | -.221 | .318 | .181 | .258 | .531* | 1 | -.120 |
| 13. P3a ERP | -.227 | -.310 | -.251 | .138 | -.095 | .015 | .275 | .019 | -.026 | -.032 | -.247 | -.120 | 1 |
Note: *p<.05, **p<.01.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



