Submitted:
18 November 2025
Posted:
19 November 2025
You are already at the latest version
Abstract
Low temperature is one of the major environmental challenges for most crops, especially from tropical and semitropical origin. The present work aimed to study low temperatures tolerance in tomato plants when previously these were inoculated with Trichoderma asperellum. Here, it was demonstrated that tomato plants inoculated with fungal isolate exhibited an alleviation of the injuries caused by low temperatures. The increasement of chilling tolerance was accompanied by a strong reduction in oxidative stress, but also by an enhancement of proline and soluble sugar accumulation. Additionally, leaf stomata features were also measured, and found that both bioinoculant and low temperatures increased leaf stomatal densities indexes but decreased the stomatal area, suggesting leaf traits may also contribute to alleviating the damage caused by the low temperatures in tomato plants. The results of the present study demonstrate that T. asperellum provokes physiological and biochemical changes in tomato plants that together enhance tolerance to low temperature, proposing the use of T. asperellum as an agroecological strategy confronting crop damage to low temperatures.
Keywords:
bioinoculants
; chilling
; cold
; proline
; soluble sugars
; reactive oxygen species
; stomatal density
; stomatal index
; stomata size
; tolerance
1. Introduction
Currently, global warming has intensified the frequency and severity exerted by both biotic and abiotic stressors on crops, posing a significant threat to agricultural productivity and food security [1]. Among these, cold stress stands out as one of the primary abiotic factors limiting crop growth worldwide, negatively impacting yield and quality [2]. Currently, it is estimated that extreme temperatures together with other abiotic stresses reduce average crop yields by more than 50% and remain to be a risk to agricultural and forest production [3].
Geographic location and climate significantly impact plant cold tolerance. For instance, native plants from warmer climates tend to be more susceptible to low temperatures (LT) than those from colder regions. Plants can be classified in three groups depending on their sensitivity to subzero and chilling (0–10 °C) temperatures: freezing-tolerant, chilling-tolerant and chilling-sensitive [4]. In this context, tomato (Solanum lycopersicum)—a semi-tropical crop—is classified as chilling-sensitive [5], whereas most commercially cultivated tomato cultivars are sensitive to temperatures below 15 °C, and a drop below 12 °C typically hampers its growth [6,7]. Depending on the intensity and duration of LT, it can negatively affect tomato plants by reducing growth traits, biochemical parameters, fruit yield, and the nutritional values of the fruits, including lycopene and β-carotene [8]. Among tomato agronomic traits affected by LT are found the relative growth rate, leaf area, number of new leaves, stem diameter, and the fresh:dry weight [9,10]. In LT-sensitive tomato plants are also observed various physiological and biochemical changes, including a decrease in chlorophyll content, low levels in photosynthetic parameters, an increase of electrolyte leakage provoked by a loss in the membrane stability, and a strong oxidative burst caused by Reactive Oxygen Species (ROS) production. Depending on ROS concentration, they may have antagonist roles in the cells. While low concentrations of them are related to signaling pathways associated with plant physiological processes and resistance to stresses, high concentrations result toxic and lethal for the cells [11].
Given that tomatoes are among the most widely cultivated horticultural crops, with a production exceeding 192 million tonnes [12], geneticists have actively worked to develop tomato varieties with enhanced tolerance to cold conditions to meet the growing demand for this important crop. Nevertheless, recent research has been developed to explore other alternatives to mitigate the injuries caused by LT in tomato plants. These includes the foliar application of phenolic compounds [13], selenium and brassinosteroids [14]. However, the use of microorganisms of soil also has been proposed as a promising alternative to plants facing extreme temperatures, representing a valuable tool for sustainable agriculture in the context of global change [6,15].
Trichoderma spp. is considered plant growth promoting fungi (PGPF) due to its rhizosphere residence and beneficial interactions with host plants [16]. Several Trichoderma species are well known to supress pathogenic fungi through nutrient competition, mycoparasitism, and antibiosis [17]. Trichoderma spp. also induces plant defense through local and systemic responses in the host plants, enhancing tolerance to both biotic and abiotic stresses [18].
It is known that Trichoderma species have the capability to produce small amounts of phytohormones, including indol-3-acetic acid (IAA), salicylic acid (SA), cytokinin (CTK), gibberellin (GA), jasmonic acid (JA), ethylene (ET) and abscisic acid (ABA), which can activate signaling events in plants during biotic and abiotic stresses [19,20]. Moreover, some Trichoderma species also secrete elicitor molecules that initiate defense signalling cascades in the plant inducing systemic resistance to biotic and abiotic stresses [21,22,23].
Synthesis of phytohormones such as SA and JA is induced during Trichoderma interactions with the host plants whereas have been related to the systemic immunity activation [24,25,26,27,28]. The interconnections between signaling pathways activated by molecules from plants and Trichoderma origin share a common approach to fine-tunes ROS levels by different mechanisms including enzymatic or nonenzymatic systems and modulation of phytohormones levels [29,30,31] in order to prevent overaccumulation of ROS to harmful levels.. Low accumulation of ROS is crucial to alleviate the injuries caused by stressful factors.
The present study was aimed at studying the effect of T. asperellum on ROS production, accumulation of the osmoprotectant proline and soluble sugars, and morphological stomatal traits (stomatal density and index, and stoma size) in tomato plants stressed with LT.
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
Commercial tomato (Solanum lycopersicum) cultivar Vita seeds were used for the study. The tomato seeds were germinated in peat moss:vermiculite (7:3) mixture under 24–30 °C/18 °C day/night cycle with natural light, and a relative humidity of 60%.
The seedlings and plants were irrigated with tap water or nutritive solution on alternate days until stress treatment application.
2.2. Fungal Growth and Inoculation of Tomato Plants
T. asperellum was grown as previously described [32]. For the T. asperellum treatment of tomato plants, T. asperellum was cultivated in a sterile solid media (oat 9%, kaolin 22%, vermiculite 43%, water 26%) and incubated at 25 °C for 5 days and shaken daily. Three- week-old tomato plants were treated with T. asperellum grown in the solid media at a final concentration of 1 × 107 Colony Forming Units (CFU)/g soil by mixing it with peat moss and vermiculite (7:3 v/v) [32].
2.3. Low Temperature Treatment
Six-week-old tomato plants were transferred to another growth chamber to apply LT at 4 °C. The control plants were kept in a growth chamber at room temperatures (RT). The collection of plant material was carried out at 0, 3-, 6-, 9- and 12-days post-treatment.
2.4. Visual Damage
Visual differences of tomato plants exposed by 12 uninterrupted days to RT or LT were analyzed. For damage severity, five levels of damage were established based on the loss of leaf turgor: level 0 (healthy), level 1 (minor loss of turgor), level 2 (medium loss of turgor), level 3 (severe loss of turgor) and level 4 (total loss of leaves turgor or dead). The phenotype was determined by calculating the percentage of damaged leaflets in relation to the total number of leaflets of each plant. The survival rate was calculated by dividing the survived plants by the total number of plants that were originally tested.
2.5. Histochemical Detection of Hydrogen Peroxide
Hydrogen peroxide detection was performed using 3,3′-diaminobenzidine (DAB) staining, as previously described [33], with minor modifications. Six-week-old tomato plants, as non-inoculated as inoculated with T. asperellum, were subjected to treatments at LT or RT for 3, 6, 9 and 12 days. The third leaf from the base was collected and immersed in 5 ml of DAB buffer solution (1 mg mL-1) for 2 hours. After staining, the DAB solution was replaced with 95% ethanol, and the samples were subsequently heated in boiling water for 10–15 minutes. The samples were then bleached in 95% ethanol three times until all chlorophyll was completely removed. Finally, the tissues were fixed in a solution of ethanol:glycerol:acetic acid at a ratio of 3:1:1 (v:v:v) and photographed. H2O2 is visualized as a reddish-brown coloration.
2.6. Catalase Activity
Six-week-old tomato leaves (0.5 g), either inoculated or non-inoculated with T. asperellum, and from both stressed and non-stressed plants, were homogenized in an ice-cold 100 mM sodium phosphate buffer (pH 7.6) supplemented with 1.0 mM EDTA and 2% (w/v) polyvinylpyrrolidone. The homogenate was then centrifuged at 4 °C and 12,000 rpm for 20 min, and the supernatant was collected for enzymatic assays.
CAT activity was measured using the method described by Aebi et al. [34]. The reaction mixture for CAT activity consisted of 2 mL of 50 mM phosphate-buffered saline (PBS, pH 7.0), 1 mL of 30 mM H2O2, and 50 μg of enzyme extract. The change in absorbance of the reaction mixture at 240 nm was recorded every 1–3 min and expressed as μmol H2O2 min-1 mg protein-1.
2.7. Soluble Sugar Content
All the collected leaf samples from the treatment and control were dried and weighed. The dried sample (0.1 g) was placed in glass vials which contained 5 ml of 80% (v/v) ethanol, and then placed in a water bath heated at 30 °C for 30 min. The supernatant was recuperated after centrifugation at 4,500 rpm and the pellet was again in contact with 2.5 mL 80% ethanol in a water bath for 30 min at 30 °C. After centrifugation the supernatant was mixed with the other supernatant. Therefore, 0.2% anthrone solution was added to each sample, and the mixture was heated at 100 °C for 10 min, followed by cooling at room temperature. The absorbance of the reaction solution was measured at 620 nm. Soluble sugar concentration in the extract was determined by comparison with a standard curve using the criterion of glucose. Results were expressed as mg soluble sugar/g DW [35].
2.8. Proline Content
The content of free proline was determined using the modified Bates et al. method [36]. A leaf sample (0.1 g) was homogenized in 2 mL of 3% sulfosalicylic acid using a mortar and pestle. The homogenate was centrifuged at maximum speed in a benchtop centrifuge for 5 min. Aliquots of 0.1 mL from the supernatant were mixed with 0.1 mL of 3% sulfosalicylic acid, 0.1 mL of acid ninhydrin solution (25 mg mL-1) and 0.2 mL of acetic acid, and the mixture was placed on a shaker for 20 minutes. The samples were heated at 90 °C for 60 min and then cooled in an ice water bath. After cooling, the mixture was thoroughly mixed with 1 mL of toluene and allow to stand at RT for 5 min to facilitate phase separation. The upper layer of the resulting mixture was used to measure absorbance at 520 nm using a UV–VIS spectrophotometer (T60 UV/VIS, PG instruments). Six plants were used as independent samples for each treatment. Proline concentration was determined from a calibration curve and expressed as µg proline g−1 fresh weight (FW).
2.9. Stomatal Traits
For the analysis of stomatal traits, we selected the third leaf from the base. Imprints were obtained using the nail polish method (NP method) [37], which involves applying nail polish to the leaf surface to create an imprint. After allowing the polish to dry, we carefully peeled it off using adhesive cellophane tape and then examined the leaf imprint under a light microscope.
We obtained one imprint from the abaxial leaf surface of each of the three plants per treatment. For every imprint, we took ten photographs at different locations on the slide. In each image, the number of stomata was recorded, excluding those that were cut off by the edges of the frame. Stomatal density was defined as the number of stomata per unit leaf area, expressed as stomata per mm2 (pores mm−2). The stomatal index was calculated by the formula described by Poole and Kürschner (1999) [38]
For each photograph, we selected five stomata for manual measurement of its length and width using the image processing software ImageJ with the Rectangle selection tool. Stomatal size was defined as the area of an ellipse, calculated from the lengths of the major and minor axes, including the size of the pore within.
2.10. Colony Forming Units (CFU) Counting
To quantify T. asperellum population in the growing media, one gram of a soil mixture containing the solid substrate inoculated with T. asperellum was collected from pots with plants maintained at cold temperatures. These samples were then diluted in an isotonic solution in a serial way, and aliquots were plated on potato dextrose agar (PDA) supplemented with 50 mg L−1 of rose Bengal and 10% streptomycin sulfate. The medium was adjusted to a pH of 4.9 using lactic acid. The plates were incubated at 28 °C for 5 days, after which colony-forming units (CFUs) were counted. Dilutions were selected to ensure that the number of colonies on the agar plates fell within the optimal range of 30 to 300 colonies per plate. The data were expressed as Log CFU per gram of dry substrate [39].
2.11. Evaluation of the Effect of Temperature on the Mycelial Growth Rate of Trichoderma asperellum
Mycelial plugs (6 mm in diameter) obtained from the actively growing margins of 7-day-old colonies were transferred to the center of Petri dishes containing PDA (one plug per plate). The plates were incubated in the dark at 4 and 28 °C in a controlled-environment growth chamber. After 7 days of incubation, the extension of the colony’s diameter was measured. The plates incubated to LT were placed at 28 °C for 3 days to measure again the colony diameter [40].
2.12. Statistical Analysis
The data were expressed as mean ± standard deviation (SD) for three-six replicates in each group The significance of differences among the treatments was evaluated by using the Tukey HSD test (p< 0.05). The analyses were performed by using Minitab®14 statistical software for windows.
3. Results
3.1. Trichoderma asperellum Enhances Low Temperature Tolerance in Tomato
To investigate the phenotypical changes of tomatoes during LT stress tolerance in combination with T. asperellum treatment, three-week-old tomato plants were inoculated and three weeks later the plants were exposed to LT conditions. After 12 days of LT stress, both T. asperellum-inoculated and -uninoculated plants exhibited symptoms of LT stress, including chlorosis and wilting. However, the T. asperellum-inoculated plants showed fewer symptoms to those of the non-inoculated plants (Figure 1a). To determine how LT stress affected the tomato plants—with or without T. asperellum inoculation—leaf damage severity was evaluated 12 days post-treatment (Figure 1b). Results were expressed as the percentage of damaged leaflets per plant. Leaf damage was quantified using a five-level severity scale that classified damage based on the loss of leaflets turgor. As shown in Figure 1b, inoculation with T. asperellum mitigated leaf damage severity provoked by LT compared to non-inoculated plants. The predominant damage level in inoculated plants was Level 1, observed in 61% of the cases, whereas non-inoculated plants exhibited higher damage levels, primarily in Levels 3 and 4, with incidences of 28% and 32%, respectively. When these plants were returned to optimal growth conditions for 14 days, tomato plants inoculated with T. asperellum exhibited an approximately 2.2-fold increase in survival rate compared to that of non-inoculated plants (Figure 1c).
3.2. Trichoderma asperellum Decreases LT-Induced ROS Accumulation
Since T. asperellum improves LT stress tolerance in tomato, it was expected that the bioinoculant could be affecting ROS accumulation in tomato plants under LT stress conditions. To investigate this, it was visualized ROS accumulation using DAB staining. As shown in Figure 2a, DAB staining showed that H2O2 accumulation kept at low levels under normal conditions of growth and no differences could be observed between non- and inoculated plants. However, after exposure to LT for 3, 6, 9 and 12 days, the H2O2 accumulation increased to higher levels in non-inoculated plants, but it remained at lower levels in inoculated plants. These results indicated that T. asperellum decreased LT-induced ROS accumulation.
To understand the mechanism by which T. asperellum regulates ROS production, the role of the antioxidant activity of the catalase (CAT) following exposure to LT was analyzed. Figure 2b shows that lowest activity of CAT was observed in the plants grown at RT with or without T. asperellum, while the highest CAT activity was observed in non-inoculated plants exposed to LT, followed by the inoculated plants under LT stress.
3.3. Trichoderma asperellum Promotes Overaccumulation of Proline and Total Soluble Sugars Content in Tomato Plants Under Low Temperature
Proline is an amino acid that is positively correlated with plant tolerance in response to various stress conditions [41]. The measurement of proline content of tomato plants revealed a significant increase in plants exposed to LT stress, which was further enhanced by inoculation with T. asperellum (Figure 3a). The highest increase of the proline content was observed after 12 days of LT treatment, where plants pretreated with T. asperellum showed a 2.1-fold increase in proline content in stressed leaves, compared to stressed but non-inoculated leaves.
On the other hand, the content of total soluble sugars, which is considered as osmoprotectant, increased by 41% when the plants were treated with T. asperellum alone and increase by 50% when treated in combination with LT for 12 days, in comparison with their respective control plants, indicating T. asperellum induces the production of osmolytes to promote homeostasis maintenance during LT stress.
3.4. Analysis of Stomatal Traits in Tomato Inoculated with Trichoderma asperellum and Treated with Low Temperatures
As previously we reported that LT dramatically diminished leaf area and succulence of non-inoculated tomato plants, but in a minor level in inoculated plants [9], so this study investigated whether stoma cells could be contributing to enhance LT-tolerance induced by T. asperellum. For that, it was analysed stomatal traits, including stomatal density, stomatal index and stomatal size, of tomato plants treated with or without T. asperellum and under LT stress conditions.
As shown the Figure 4, T. asperellum, in the absence of abiotic stress, increased the stomata number and decreased the stomata size, compared with uninoculated plants, indicating that T. asperellum by itself is modifying the leaf morphological traits of tomato plants. Regarding application of stress for 9 days, the stomata density significantly increased in both uninoculated and inoculated plants by 44.2% and 29.3%, compared with their respective controls (Figure 4a), while the increase of stomata index was lower (Figure 4b). Notably, LT stress significantly reduced stomatal size after 9 days of treatment in uninoculated plants while stomatal area was slightly affected by LT stress in inoculated plants (Figure 4c).
3.5. Trichoderma asperellum Survives in Low Temperatures Conditions
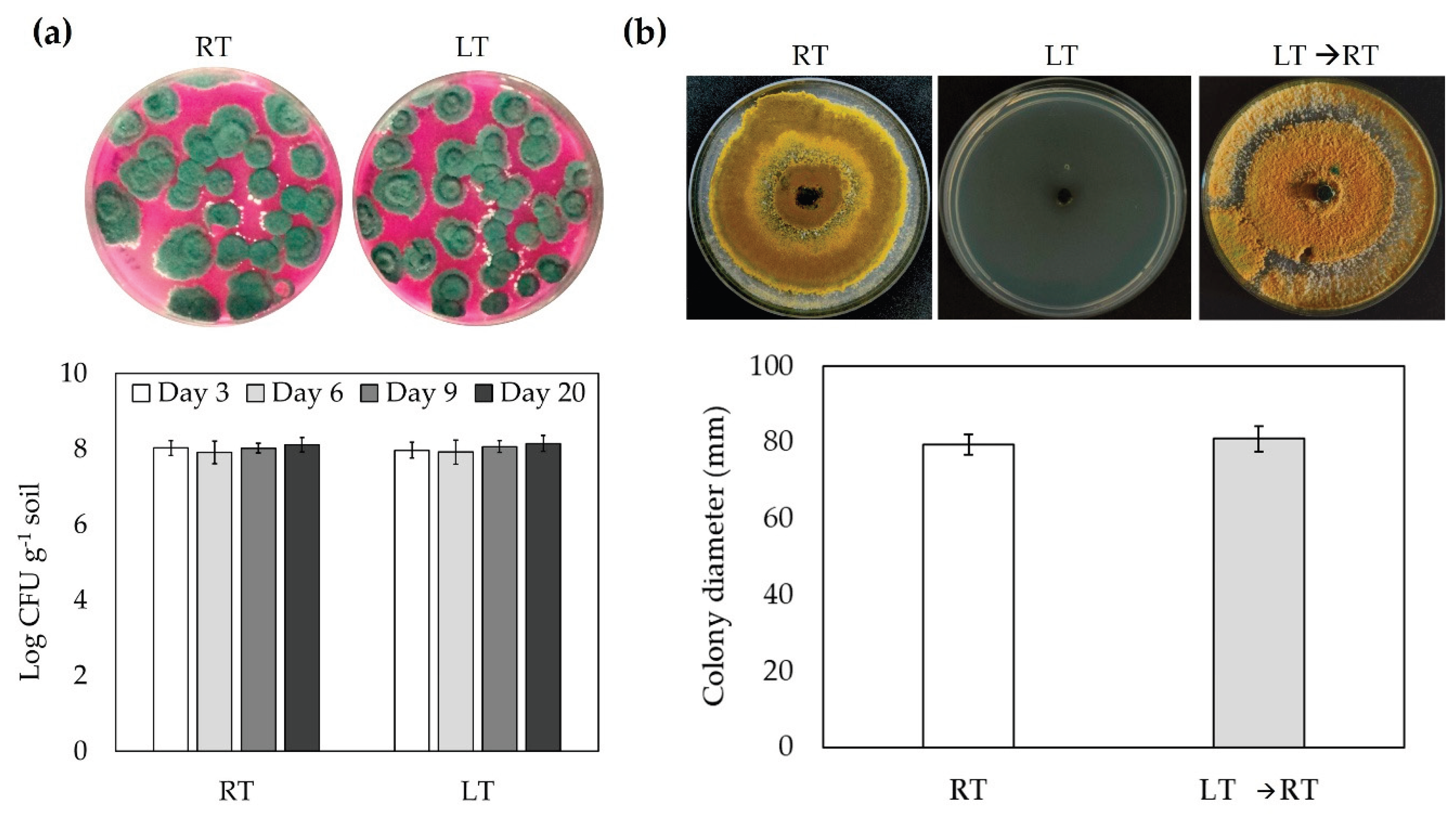
The CFU counting assay is a widely used technique for estimating the number of viable fungal cells in any sample. As was appreciated in Figure 5a, the CFU of T. asperellum in the soil was conserved at LT in the studied times. The growth of the fungus in PDA revealed a strong growth inhibition at 4 °C; however, when the plates were returned to 28 °C no differences in fungal growth was observed in reference to control conditions (28 °C), indicating that T. asperellum did not have lethal effects at LT.
4. Discussion
Trichoderma spp. is an important fungal genus that could improve tolerance against LT stress in plants by inducing systemic resistance and stimulating plant growth.
Low temperatures are known to elevate the production of ROS, particularly in species with lower antioxidant capacities. The present study reported that the non-inoculated tomato plants experienced high levels of oxidative stress during LT stress, contrary to plants inoculated with T. asperellum, suggesting that tolerance of tomato plants to LT stress is closely related to the accumulation of ROS.
Plants use antioxidant machinery to counter hazardous ROS levels and improve tolerance to adverse environments. The ROS can be scavenged by low molecular weight antioxidative metabolites e.g., glutathione, ascorbic acid, α-tocopherol or by antioxidative enzymes e.g., CAT, ascorbate peroxidase (APX) and superoxide dismutase (SOD). In this study it was found the CAT activity was activated in tomato plants under LT stress. The fact that uninoculated plants under LT stress showed a strong ROS accumulation even when the CAT activity was also increased, suggests that oxidative burst may have exceeded the potential of antioxidant capacity. In contrast, the CAT activity was moderate in inoculated plants under LT. This result may be explained by 1) involvement of other cellular sources of detoxify ROS (such the production of proline as we described below), 2) because of a lesser production of ROS, 3) production of SA by the fungus and/or by the plant, which has the ability to inhibiting CAT activity [42,43].
Proline is an amino acid that plays beneficial roles in stressed plants since it acts as an osmolyte, a metal chelator, and an antioxidative and signaling molecule [41]. Non-toxic accumulation of proline leads to the improvement in the osmotic adjustment and the integrity of membranes and proteins. Additionally, proline accumulation prevents electrolyte leakage and the oxidative burst by increasing the antioxidant enzymatic activities such as CAT, SOD, APX, and peroxidase [41,44].
Proline action on modulation of ROS levels remains under study, however it has been documented that proline can directly interact with ROS (mainly free radicals) [45,46] and that during proline biosynthesis is induced the production of antioxidant molecules through the ascorbate/glutathione (AsA-GSH) pathway that reduce ROS like H2O2 [47].
Under adverse conditions, the plants rapidly lead to an increase in the transcriptional regulation of Δ1-pyrroline-5-carboxylate synthetase (P5CS) and Δ1-pyrroline-5-carboxylate reductase (P5CR), which encode the two enzymes involved in proline biosynthesis. Furthermore, the prevalence of high levels of proline also is explained by the downregulation in expression of genes whose products are enzymes that act on proline catabolism [48,49]. In tomato plants under chilling stress, inoculation with Trichoderma harzianum strain AK20G led to increased P5CS expression levels over time, which correlated with enhanced cold tolerance and proline accumulation.
In the present study, the plants inoculated with T. asperellum exhibited a marked increase by 50% in leaf proline content under low-temperature stress (Figure 3). This result is in accordance with the effect of T. harzianum on proline accumulation and improvement salt and LT tolerance in cucumber and tomato plants, respectively [50,51]. Interestingly, T. harzianum increased the expression of P5CS in tomato plants after cold treatment [51]. Furthermore, the observed results are consistent with the increase in proline content and activation of the antioxidant machinery in plants inoculated with psychrotolerant bacterial strains Pseudomonas vancouverensis OB155-gfp, P. frederiksbergensis OS261-gfp, P. frederiksbergensis OS211 and Flavobacterium glaciei OB146 to mitigate the damages provoked by LT in tomato plants [6,15].
The increase in proline accumulation could positively correlate to the low levels of ROS accumulation observed in tomato inoculated plants, since it has been postulated that proline can function in the ROS detoxifying process, as it has been reported in response to metal treatment [52]. Also, this data reinforces our findings that it seems CAT activity is not the main source of detoxification during T. asperellum treatment.
On the other hand, soluble sugars (sucrose glucose, fructose and oligosaccharides) are another kind of metabolites that are related to LT tolerance. Soluble sugars are well documented molecules as osmolytes; however, these can also protect cell membranes from dehydration [53], and act as second messenger in signal transduction pathways to regulate de expression for genes related to hormone biosynthesis (such as ABA, ET and GA) and cold stress tolerance, like COR (COLD REGULATED) and RD (RESPONSIVE TO DEHYDRATION) [54,55]. The mechanism by which soluble sugars rise could be caused by the activation of invertase enzymes and sucrose synthases as was shown wheat seedlings during LT treatment [56].
The tomato plants treated with T. asperellum alone, increased soluble sugar content in tomato plants, like the observed for proline, suggesting that Trichoderma could be inducing the gene expression or activity of plant enzymes related to the biosynthesis of both osmolytes, which would be greater in plants under adverse environmental conditions to improve tolerance.
Phenotypic plasticity is one of the main characteristics of a plant that is used to acclimatize to different growth conditions [57]. A reduction in leaf area and plant growth allows plants to reduce their transpiration, thus increasing water use efficiencies (WUE) [58,59]. At the same time, leaf expansion and transpiration reduction is directly related to stomatal density and stomatal size [60], which in turn affects ecosystem function [61].
To optimize carbon fixation per unit water loss, plants can adjust stomatal pore aperture in the short term and modify stomatal traits such as stomatal density and stomatal size [62], where these last two have a negative relationship in many species [60,63,64]. Every plant can rapidly regulate the stomata aperture within minutes, but changing density takes days or weeks [57].
There are several studies revealing that drought stress leads to an increase in stomatal density and a decrease in stomatal size to minimize water loss while maintaining CO2 assimilation [58,65]; however, little is known concerning this relationship regarding LT and to bioinoculants.
Like drought stress, stomata density increased in tomato leaves under LT stress, suggesting that an increase in stomatal density might be associated with the decrease in leaf size previously observed in tomato plants under LT stress [9] to optimize carbon fixation per unit water loss. By increasing stomatal density, the plant may be able to increase stomatal conductance and maximize CO2 uptake, which could be beneficial for plant nutrient uptake and photosynthesis [57].
Previous studies have proposed that small stomata can adjust stomatal pore area and regulate stomatal conductance faster and, thus, respond rapidly to environmental change improving long-term water use efficiency and diminishing risk of disruption to the leaf hydraulic system [66,67,68]. In leaves with many small pores, there are better gas conductance and a greater potential for photosynthesis than for leaves with fewer large stomata with the same pore area per unit leaf area [64]. In this study, LT stress led to a reduction of stomata size at 9 days, suggesting tomato plant under stress is modifying the phenotype of guard cell size as an adaptation way to protect themselves from damage caused by environmental variations for restricting transpiration, increasing WUE and improving carbon gain for photosynthesis.
Inoculation of tomato plants with T. asperellum also led to the negative correlation between stomatal density and stomatal size in optimal growth conditions and it was further under stress conditions. These results suggest that T. asperellum can alter the anatomy and patterning of stomata. Recently, it was found that beneficial microorganisms, particularly endophytes, alter stomatal traits. For example, Salicacea inoculated Populus trichocarpa had more compact stomata and an increase in stomatal density prior and after water-deficit [69]. Additionally, it has been shown that inoculation with Burkholderia sp LD-11 or with Xerophyte-Derived Synthetic Bacterial Communities in maize improved WUE and increased the sensitivity of stomatal gas conductance, and decreased transpiration rate promoting tolerance to water deficits [70,71].
The involved mechanism in stomatal density and stomata size provoked by T. asperellum inoculation is unknown, but it could hypothesize that the bioinoculant may induce expression or activation of transcription factors that regulate the cellular divisions and cell fate transitions necessary for stomatal development through regulation of epidermal patterning factor (EPF) [57], or by STOMAGEN action, a positive regulator of stomatal development [72], through hormonal signaling pathways to control the stomatal density. In this sense is already known that exogenous ABA induce the production of smaller stomata and higher stomatal density [73], and also, is already known, that some Trichoderma species synthesizing ABA [20] so in the future will be necessary to analyze the phytohormone profile of the strain used in this study.
Soil microbial communities either respond immediately to changes in the environment or can adapt to prolonged periods of stress through physiological adjustments [74]. Recently, it has been demonstrated that cooling soils decreased microbial processes such as respiration and growth but had little to no effect on the uptake and respiration rate of amended glucose. This result is of great relevance because sugars are precursors of osmolytes and act also as osmolytes themselves. Additionally, the rise of unsaturated phospholipid fatty acids was also observed, which can be interpreted as a specific microbial community adaptation to cool temperatures by enhancing the flexibility of cell membranes [75].
In this study, the prevalence of T. asperellum in soils from plants exposed to LT was evaluated. The results did not show any significant changes in the fungal densities during abiotic stress compared to optimal conditions, suggesting that viability of the fungus is maintained in cold soils for long periods (20 days); however, it remains unclear whether it can colonize the plants enough in cold conditions.
The temperature has a significant influence on the in vitro growth of T. asperellum, which has an optimal mycelial growth of 30 °C [40]. The growth of fungi in PDA at 4 °C was totally inhibited after 7 days, but when the fungi-containing plates were returned to optimal temperatures, the fungal growth was restored with the same rate that those plates grown at 28 °C (as was previously observed [40]), indicating LT did not have lethal effects on the fungus as was previously observed [40].
5. Conclusions
Tomato plants inoculated with T. asperellum exhibited an improvement of LT tolerance. Increased ROS scavenging, maintaining the osmotic balance and changes in stomatal traits may be relying in the enhancement LT tolerance. Our results revealed that T. asperellum could be used to develop sustainable strategies to induce LT tolerance by maintaining food security in response to climate change.
Author Contributions
Conceptualization, M.S.-G.; results and method development: M.d.P.O.-S.; statistical analysis, M.I.R.-S.; writing—original draft, M.S.-G.; writing—review and editing, D.S.-R. and A.M.-N.; supervision, E.A.-T., I.H.-S., M.I.R.-S., D.S.-R., and A.M.-N..; funding acquisition, M.S.-G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding. M.d.P.O.-S. received CONACyT (822722) fellowships.
Acknowledgments
The authors offer special thanks to Pip Eitel (Peace Corps) for editing work and to the four anonymous reviewers for their comments and critiques.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ABA | Abscisic acid |
| APX | Ascorbate peroxidase |
| CAT | Catalase |
| CFU | Colony Forming Units |
| ET | Ethylene |
| GA | Gibberellin |
| JA | Jasmonic acid |
| LT | Low Temperatures |
| ROS | Reactive Oxygen Species |
| RT | Room Temperature |
| SA | Salicylic acid |
| SOD | Superoxide Dismutase |
References
- Tanaka, K.; Mudgil, Y.; Tunc-Ozdemir, M. Editorial: Abiotic stress and plant immunity – a challenge in climate change. Front Plant Sci 2023, 14, 1197435. [Google Scholar] [CrossRef]
- Guan, Y.; Hwarari, D.; Korboe, H.M.; Ahmad, B.; Cao, Y.; Movahedi, A.; et al. Low temperature stress-induced perception and molecular signaling pathways in plants. Environ Exp Bot 2023, 207, 105190. [Google Scholar] [CrossRef]
- Devireddy, A.R.; Tschaplinski, T.J.; Tuskan, G.A.; Muchero, W.; Chen, J.-G. Role of reactive oxygen species and hormones in plant responses to temperature changes. Int J Mol Sci 2021, 22, 8843. [Google Scholar] [CrossRef] [PubMed]
- Guy, C.L. Cold acclimation and freezing stress tolerance: role of protein metabolism. Annu Rev Plant Biol 1990, 41, 187–223. [Google Scholar] [CrossRef]
- Lyons, J.M. Chilling injury in plants. Annu Rev Plant Biol 1973, 24, 445–466. [Google Scholar] [CrossRef]
- Subramanian, P.; Kim, K.; Krishnamoorthy, R.; Mageswari, A.; Selvakumar, G.; Sa, T. Cold stress tolerance in psychrotolerant soil bacteria and their conferred chilling resistance in tomato (Solanum lycopersicum Mill.) under low temperatures. PLOS ONE 2016, 11, e0161592. [Google Scholar] [CrossRef]
- Van Ploeg, D.; Heuvelink, E. Influence of sub-optimal temperature on tomato growth and yield: a review. J Hortic Sci Biotechnol 2005, 80, 652–659. [Google Scholar] [CrossRef]
- Khan, T.A.; Fariduddin, Q.; Yusuf, M. Lycopersicon esculentum under low temperature stress: an approach toward enhanced antioxidants and yield. Environ Sci Pollut Res 2015, 22, 14178–14188. [Google Scholar] [CrossRef]
- Cornejo-Ríos, K.; Osorno-Suárez, M. delP.; Hernández-León, S.; Reyes-Santamaría, M.I.; Juárez-Díaz, J.A.; Pérez-España, V.H.; et al. Impact of Trichoderma asperellum on chilling and drought stress in tomato (Solanum lycopersicum). Horticulturae 2021, 7, 385. [Google Scholar] [CrossRef]
- Sherzod, R.; Yang, E.Y.; Cho, M.C.; Chae, S.Y.; Kim, J.H.; Nam, C.W.; et al. Traits affecting low temperature tolerance in tomato and its application to breeding orogram. Plant Breed Biotechnol 2019, 7, 350–359. [Google Scholar] [CrossRef]
- Quan, L,-J.; Zhang, B.; Shi, W.-W.; Li, H.-Y. Hydrogen peroxide in plants: a versatile molecule of the reactive oxygen species network. J. Integr. Plant Biol. 2008, 50, 2–18. [CrossRef] [PubMed]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 11 June 2025).
- Meena, Y.K.; Khurana, D.S.; Kaur, N.; Singh, K. Phenolic compounds enhanced low temperature stress tolerance in tomato (Solanum lycopersicum L.). Curr J Appl Sci Technol 2017, 20, 1–9. [Google Scholar] [CrossRef]
- Saeed, T.; Khan, T.A.; Ahmad, A.; Yusuf, M.; Kappachery, S.; Fariduddin, Q.; et al. Exploring the effects of selenium and brassinosteroids on photosynthesis and protein expression patterns in tomato plants under low temperatures. Plants 2023, 12, 3351. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, P.; Mageswari, A.; Kim, K.; Lee, Y.; Sa, T. Psychrotolerant endophytic Pseudomonas sp. strains OB155 and OS261 induced chilling resistance in tomato plants (Solanum lycopersicum Mill.) by activation of their antioxidant capacity. Mol Plant Microbe Interact 2015, 28, 1073–1081. [Google Scholar] [CrossRef]
- Hossain, M.d.M.; Sultana, F.; Islam, S. Plant Growth-Promoting Fungi (PGPF): phytostimulation and induced systemic resistance. In Plant-Microbe Interact Agro-Ecol Perspect.; Singh, D.P., Singh, H.B., Prabha, R., Eds.; Interact. Agro-Ecol. Impacts; Springer: Singapore, 2017; Vol. 2, pp. 135–191. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species - opportunistic, avirulent plant symbionts. Nat Rev Microbiol 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Zhang, S.; Gan, Y.; Xu, B. Application of plant-growth-promoting fungi Trichoderma longibrachiatum T6 enhances tolerance of wheat to salt stress through improvement of antioxidative defense system and gene expression. Front Plant Sci 2016, 7, 1405. [Google Scholar] [CrossRef]
- Chanclud, E.; Morel, J.-B. Plant hormones: a fungal point of view. Mol Plant Pathol 2016, 17, 1289–1297. [Google Scholar] [CrossRef]
- Illescas, M.; Pedrero-Méndez, A.; Pitorini-Bovolini, M.; Hermosa, R.; Monte, E. Phytohormone production profiles in Trichoderma species and their relationship to wheat plant responses to water stress. Pathogens 2021, 10, 991. [Google Scholar] [CrossRef]
- Lamdan, N.-L.; Shalaby, S.; Ziv, T.; Kenerley, C.M.; Horwitz, B.A. Secretome of Trichoderma interacting with maize roots: role in induced systemic resistance. Mol Cell Proteomics 2015, 14, 1054–1063. [Google Scholar] [CrossRef]
- Zaid, R.; Koren, R.; Kligun, E.; Gupta, R.; Leibman-Markus, M.; Mukherjee, P.K.; et al. Gliotoxin, an immunosuppressive fungal metabolite, primes plant immunity: evidence from Trichoderma virens-tomato interaction. mBio 2022, 13, e00389-22. [Google Scholar] [CrossRef]
- Yao, X.; Guo, H.; Zhang, K.; Zhao, M.; Ruan, J.; Chen, J. Trichoderma and its role in biological control of plant fungal and nematode disease. Front Microbiol 2023, 14, 1160551. [Google Scholar] [CrossRef]
- Jogaiah, S.; Abdelrahman, M.; Tran, L.-S.P.; Ito, S.-I. Different mechanisms of Trichoderma virens-mediated resistance in tomato against Fusarium wilt involve the jasmonic and salicylic acid pathways. Mol Plant Pathol 2018, 19, 870–882. [Google Scholar] [CrossRef]
- Yuan, M.; Huang, Y.; Ge, W.; Jia, Z.; Song, S.; Zhang, L.; et al. Involvement of jasmonic acid, ethylene and salicylic acid signaling pathways behind the systemic resistance induced by Trichoderma longibrachiatum H9 in cucumber. BMC Genomics 2019, 20, 144. [Google Scholar] [CrossRef]
- Agostini, R.B.; Postigo, A.; Rius, S.P.; Rech, G.E.; Campos-Bermudez, V.A.; Vargas, W.A. Long-lasting primed state in maize plants: salicylic acid and steroid signaling pathways as key players in the early activation of immune responses in silks. Mol Plant-Microbe Interactions 2019, 32, 95–106. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Beltrán-Peña, E.; Herrera-Estrella, A.; López-Bucio, J. Trichoderma-induced plant immunity likely involves both hormonal- and camalexin-dependent mechanisms in Arabidopsis thaliana and confers resistance against necrotrophic fungi Botrytis cinerea. Plant Signal Behav 2011, 6, 1554–1563. [Google Scholar] [CrossRef]
- Kottb, M.; Gigolashvili, T.; Großkinsky, D.K.; Piechulla, B. Trichoderma volatiles effecting Arabidopsis: from inhibition to protection against phytopathogenic fungi. Front Microbiol 2015, 6, 995. [Google Scholar] [CrossRef]
- Chen, S.-C.; Ren, J.-J.; Zhao, H.-J.; Wang, X.-L.; Wang, T.-H.; Jin, S.-D.; et al. Trichoderma harzianum improves defense against Fusarium oxysporum by regulating ROS and RNS metabolism, redox balance, and energy flow in cucumber roots. Phytopathology 2019, 109, 972–982. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Liu, Z.; Li, Z.; Wang, Y.; Yang, K. Alleviation of the effects of saline-alkaline stress on maize seedlings by regulation of active oxygen metabolism by Trichoderma asperellum. PloS One 2017, 12, e0179617. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Trejo, J.; Aquino-Torres, E.; Reyes-Santamaría, M.I.; Islas-Pelcastre, M.; Pérez-Ríos, S.R.; Madariaga-Navarrete, A.; et al. Plant defensive responses triggered by Trichoderma spp. as tools to face stressful conditions. Horticulturae 2022, 8, 1181. [Google Scholar] [CrossRef]
- Herrera-Téllez, V.I.; Cruz-Olmedo, A.K.; Plasencia, J.; Gavilanes-Ruíz, M.; Arce-Cervantes, O.; Hernández-León, S.; et al. The protective effect of Trichoderma asperellum on tomato plants against Fusarium oxysporum and Botrytis cinerea diseases involves inhibition of reactive oxygen species production. Int J Mol Sci 2019, 20, 2007. [Google Scholar] [CrossRef]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley—powdery mildew interaction. Plant J 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol 1984, 121–126. [Google Scholar] [CrossRef]
- Khan, A.A.; McNeilly, T.; Collins, J.C. Accumulation of amino acids, proline, and carbohydrates in response to aluminum and manganese stress in maize. J Plant Nutr 2000, 23, 1303–1314. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ceulemans, R.; Van Praet, L.; Jiang, X.N. Effects of CO2 enrichment, leaf position and clone on stomatal index and epidermal cell density in poplar (Populus). New Phytol 1995, 131, 99–107. [Google Scholar] [CrossRef]
- Poole, I.; Kürschner, W. Stomatal density and index: The practice. In Foss. Plants Spores Mod. Tech.; 1999; pp. 257–260. [Google Scholar]
- Martínez-Medina, A.; Roldán, A.; Pascual, J.A. Interaction between arbuscular mycorrhizal fungi and Trichoderma harzianum under conventional and low input fertilization field condition in melon crops: growth response and Fusarium wilt biocontrol. Appl Soil Ecol 2011, 47, 98–105. [Google Scholar] [CrossRef]
- Cavalcante, A.L.A.; Negreiros, A.M.P.; Melo, N.J. de A.; Santos, F.J.Q.; Soares Silva, C.S.A.; Pinto, P.S.L.; et al. Adaptability and sensitivity of Trichoderma spp. isolates to environmental factors and fungicides. Microorganisms 2025, 13, 1689. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments. Plant Signal Behav 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Klessig, D.F.; Tian, M.; Choi, H.W. Multiple targets of salicylic acid and its derivatives in plants and animals. Front Immunol 2016, 7, 206. [Google Scholar] [CrossRef]
- Durner, J.; Klessig, D.F. Salicylic acid is a modulator of tobacco and mammalian catalases. J Biol Chem 1996, 271, 28492–28501. [Google Scholar] [CrossRef]
- Ritonga, F.N.; Chen, S. Physiological and molecular mechanisms involved in cold stress tolerance in plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef]
- Smirnoff, N.; Cumbes, Q.J. Hydroxyl radical scavenging activity of compatible solutes. Phytochemistry 1989, 28, 1057–1060. [Google Scholar] [CrossRef]
- Signorelli, S.; Coitiño, E.L.; Borsani, O.; Monza, J. Molecular mechanisms for the reaction between •OH radicals and proline: insights on the role as reactive oxygen species scavenger in plant stress. J Phys Chem B 2014, 118, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Hare, P.D.; Cress, W.A. Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul 1997, 21, 79–102. [Google Scholar] [CrossRef]
- Forlani, G.; Trovato, M.; Funck, D.; Signorelli, S. Regulation of proline accumulation and its molecular and physiological functions in stress defence. In Osmoprotectant-Mediat. Abiotic Stress Toler. Plants Recent Adv. Future Perspect; Hossain, M.A., Kumar, V., Burritt, D.J., Fujita, M., Mäkelä, P.S.A., Eds.; Springer International Publishing: Amsterdam, The Netherlands, 2019; pp. 73–97. [Google Scholar] [CrossRef]
- Renzetti, M.; Funck, D.; Trovato, M. Proline and ROS: a unified mechanism in plant development and stress response? Plants 2025, 14, 2. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, Y.; Liu, C.; Chen, F.; Ge, H.; Tian, F.; et al. Trichoderma harzianum mitigates salt stress in cucumber via multiple responses. Ecotoxicol Environ Saf 2019, 170, 436–445. [Google Scholar] [CrossRef]
- Ghorbanpour, A.; Salimi, A.; Ghanbary, M.A.T.; Pirdashti, H.; Dehestani, A. The effect of Trichoderma harzianum in mitigating low temperature stress in tomato (Solanum lycopersicum L.) plants. Sci. Hortic. 2018, 230, 134–141. [Google Scholar] [CrossRef]
- Wang, Y.; Tan, P.; Chang, L.; Yue, Z.; Zeng, C.; Lo, M.; Liu, Z.; Dong, X.; Yan, M. Exogenous proline mitigates toxic effects of cadmium via the decrease of cadmium accumulation and reestablishment of redox homeostasis in Brassica juncea. BMC Plant Biology 2022, 22, 182. [Google Scholar] [CrossRef]
- Anchordoguy, T.J.; Rudolph, A.S.; Carpenter, J.F.; Crowe, J.H. Modes of interaction of cryoprotectants with membrane phospholipids during freezing. Cryobiology 1987, 24, 324–331. [Google Scholar] [CrossRef]
- Sakr, S.; Wang, M.; Dédaldéchamp, F.; Perez-Garcia, M.-D.; Ogé, L.; Hamama, L.; et al. The sugar-signaling Hub: overview of regulators and interaction with the hormonal and metabolic network. Int J Mol Sci 2018, 19, 2506. [Google Scholar] [CrossRef]
- Yuanyuan, M.; Yali, Z.; Jiang, L.; Hongbo, S. Roles of plant soluble sugars and their responses to plant cold stress. Afr J Biotechnol 2009, 8, 2004–2010. [Google Scholar]
- Abdel-Latif, A. Activity of sucrose synthase and acid invertase in wheat seedlings during a cold-shock using micro plate reader assays. Aust. J. Basic Appl. Sci. 2008, 2, 53–56. [Google Scholar]
- Hasanuzzaman, M.; Zhou, M.; Shabala, S. How does stomatal density and residual transpiration contribute to osmotic stress tolerance? Plants 2023, 12, 494. [Google Scholar] [CrossRef]
- Xu, Z.; Zhou, G. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass. J Exp Bot 2008, 59, 3317–3325. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.Y.; Han, J.M.; Yi, X.P.; Zhang, W.F.; Zhang, Y.L. Coordinated variation between veins and stomata in cotton and its relationship with water-use efficiency under drought stress. Photosynthetica 2018, 56, 1326–1335. [Google Scholar] [CrossRef]
- Franks, P.J.; Beerling, D.J. Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time. Proc Natl Acad Sci U S A 2009, 106, 10343–10347. [Google Scholar] [CrossRef]
- Liu, C.; Li, Y.; Zhang, J.; Baird, A.S.; He, N. Optimal community assembly related to leaf economic- hydraulic-anatomical traits. Front Plant Sci 2020, 11, 341. [Google Scholar] [CrossRef]
- Liu, C.; Sack, L.; Li, Y.; Zhang, J.; Yu, K.; Zhang, Q.; et al. Relationships of stomatal morphology to the environment across plant communities. Nat Commun 2023, 14, 6629. [Google Scholar] [CrossRef] [PubMed]
- Hetherington, A.M.; Woodward, F.I. The role of stomata in sensing and driving environmental change. Nature 2003, 424, 901–908. [Google Scholar] [CrossRef]
- Franks, P.J.; Drake, P.L.; Beerling, D.J. Plasticity in maximum stomatal conductance constrained by negative correlation between stomatal size and density: an analysis using Eucalyptus globulus. Plant Cell Environ 2009, 32, 1737–1748. [Google Scholar] [CrossRef]
- Quarrie, S.A.; Jones, H.G. Effects of abscisic acid and water stress on development and morphology of wheat. J Exp Bot 1977, 28, 192–203. [Google Scholar] [CrossRef]
- Drake, P.L.; Froend, R.H.; Franks, P.J. Smaller, faster stomata: scaling of stomatal size, rate of response, and stomatal conductance. J Exp Bot 2013, 64, 495–505. [Google Scholar] [CrossRef]
- Raven, J.A. Speedy small stomata? J Exp Bot 2014, 65, 1415–1424. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-B.; Guan, Z.-J.; Sun, M.; Zhang, J.-J.; Cao, K.-F.; Hu, H. Evolutionary association of stomatal traits with leaf vein density in Paphiopedilum, Orchidaceae. PLoS ONE 2012, 7, e40080. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, M.; Banan, D.; Tournay, R.; Bakker, J.D.; Doty, S.L.; Kim, S.-H. Salicaceae endophyte inoculation alters stomatal patterning and improves the intrinsic water-use efficiency of Populus trichocarpa after a water deficit. J Exp Bot 2025, 76, 3499–3514. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Hu, H.; Huang, G.; Huang, F.; Li, Y.; Palta, J. Soil inoculation with Burkholderia sp. LD-11 has positive effect on water-use efficiency in inbred lines of maize. Plant Soil 2015, 390, 337–349. [Google Scholar] [CrossRef]
- Chukwudi, U.P.; Babalola, O.O.; Glick, B.R.; Santoyo, G.; Rigobelo, E.C. Field application of beneficial microbes to ameliorate drought stress in maize. Plant Soil 2025. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sugano, S.S.; Shimada, T.; Hara-Nishimura, I. Enhancement of leaf photosynthetic capacity through increased stomatal density in Arabidopsis. New Phytol 2013, 198, 757–764. [Google Scholar] [CrossRef]
- Franks, P.J.; Farquhar, G.D. Effect of exogenous abscisic acid on stomatal development, stomatal mechanics, and leaf gas exchange in Tradescantia virginiana. Plant Physiol 2001, 125, 935–942. [Google Scholar] [CrossRef]
- Jansson, J.K.; Hofmockel, K.S. Soil microbiomes and climate change. Nat Rev Microbiol 2020, 18, 35–46. [Google Scholar] [CrossRef]
- Schnecker, J.; Spiegel, F.; Li, Y.; Richter, A.; Sandén, T.; Spiegel, H.; et al. Microbial responses to soil cooling might explain increases in microbial biomass in winter. Biogeochemistry 2023, 164, 521–535. [Google Scholar] [CrossRef]
Figure 1.
Low temperature tolerance is induced by T. asperellum in tomato plants. (a) Representative plants grown under room temperature (RT) or exposed to 4 °C (LT) for 12 days. The inoculation of tomato plants with T. asperellum was performed 3 weeks prior to LT treatment. (b) The severity of leaf damage was assessed using a five-level scale based on the loss of leaf turgor: Level 0 (healthy), Level 1 (minor loss of turgor), Level 2 (medium loss of turgor), Level 3 (severe loss of turgor) and Level 4 (total loss of turgor). The accompanying photographs illustrate a representative example of each damage level. LT-induced leaf damage was quantified as the percentage of the leaflets exhibiting symptoms relative to the total number of leaves in each plant 12 days after treatment. The data presented are the average of five independent experiments with each consisting of 3 plants. (c) Percent survival rates of tomato plants exposed 12 days at 4 °C and then returned to RT for 14 days for their recuperation. Mean and Standard Deviation (SD) from three independent experiments are presented. The asterisk indicates statistically significant differences, as determined by a Student’s t-test (* p < 0.01).
Figure 1.
Low temperature tolerance is induced by T. asperellum in tomato plants. (a) Representative plants grown under room temperature (RT) or exposed to 4 °C (LT) for 12 days. The inoculation of tomato plants with T. asperellum was performed 3 weeks prior to LT treatment. (b) The severity of leaf damage was assessed using a five-level scale based on the loss of leaf turgor: Level 0 (healthy), Level 1 (minor loss of turgor), Level 2 (medium loss of turgor), Level 3 (severe loss of turgor) and Level 4 (total loss of turgor). The accompanying photographs illustrate a representative example of each damage level. LT-induced leaf damage was quantified as the percentage of the leaflets exhibiting symptoms relative to the total number of leaves in each plant 12 days after treatment. The data presented are the average of five independent experiments with each consisting of 3 plants. (c) Percent survival rates of tomato plants exposed 12 days at 4 °C and then returned to RT for 14 days for their recuperation. Mean and Standard Deviation (SD) from three independent experiments are presented. The asterisk indicates statistically significant differences, as determined by a Student’s t-test (* p < 0.01).
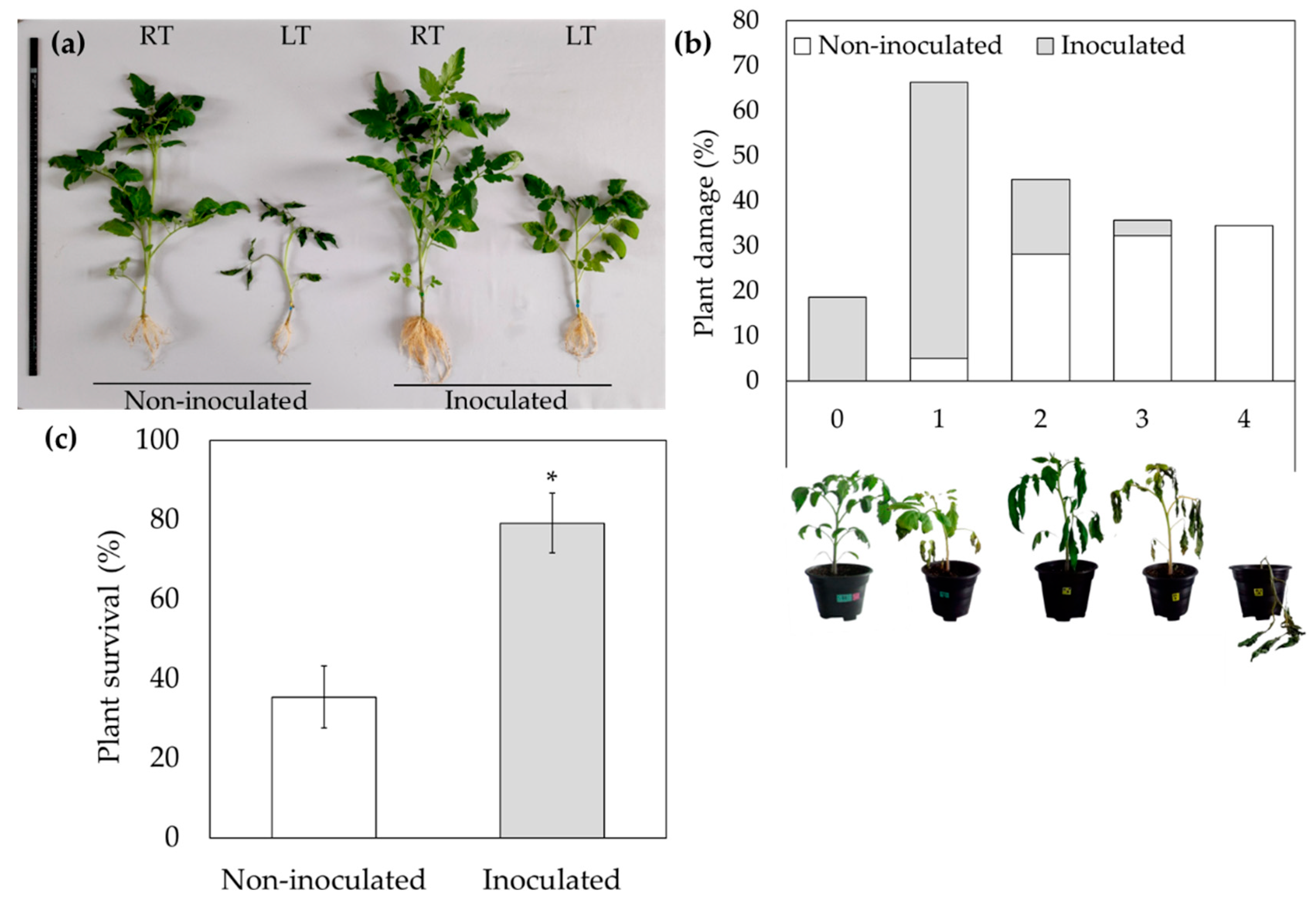
Figure 2.
Trichoderma asperellum reduces LT-induced ROS accumulation in tomato plants. (a) Analyses of DAB staining in six-week-old tomato leaves treated under RT or LT conditions. (b) Catalase activity in tomato plants under LT stress. The values are the means of three independent experiments with three replicates. Error bars indicate SD. The statistical significance (p < 0.05) was calculated with a Tukey test. The columns with different letters indicate significant difference.
Figure 2.
Trichoderma asperellum reduces LT-induced ROS accumulation in tomato plants. (a) Analyses of DAB staining in six-week-old tomato leaves treated under RT or LT conditions. (b) Catalase activity in tomato plants under LT stress. The values are the means of three independent experiments with three replicates. Error bars indicate SD. The statistical significance (p < 0.05) was calculated with a Tukey test. The columns with different letters indicate significant difference.
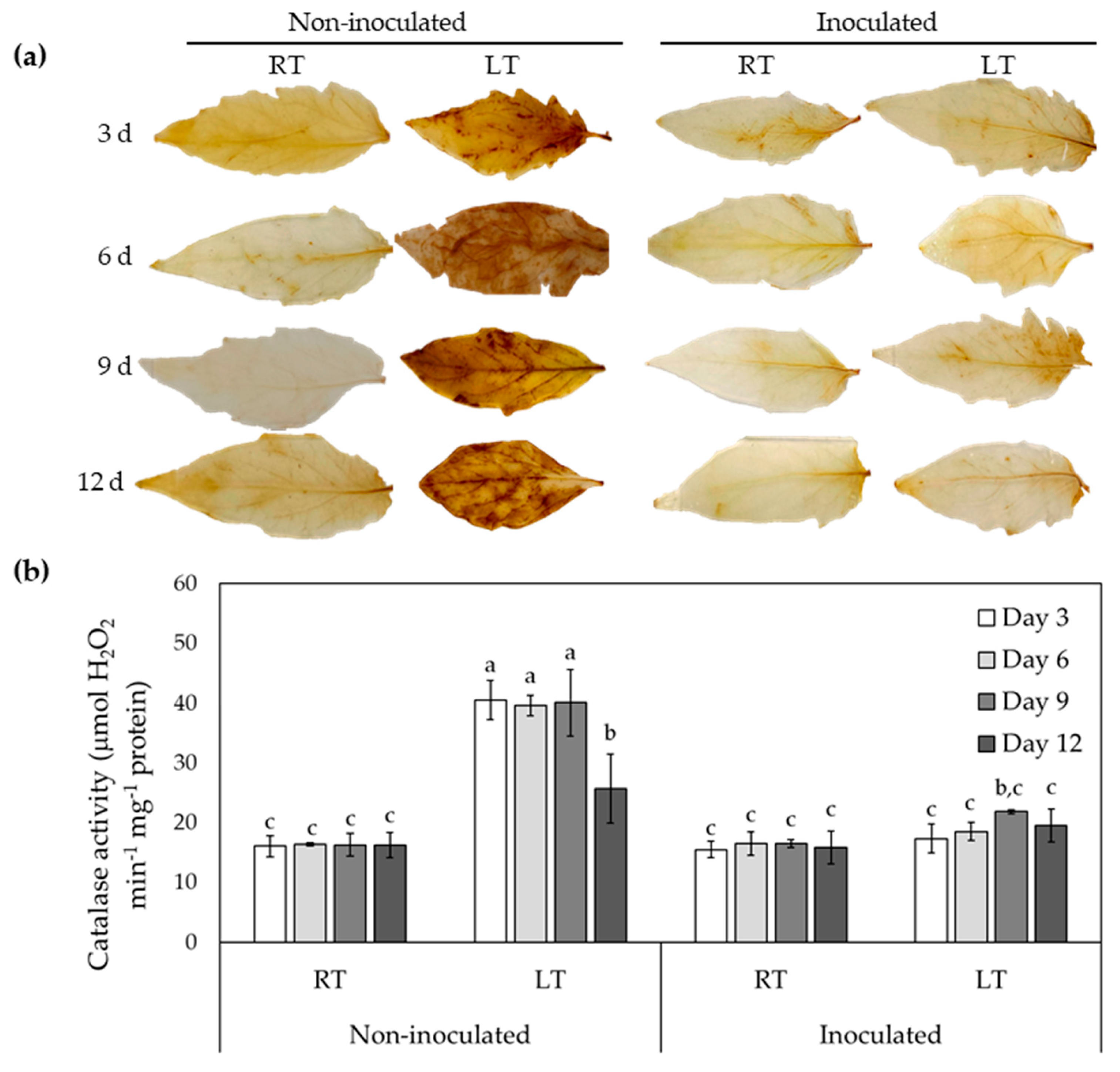
Figure 3.
Trichoderma asperellum enhances the accumulation of osmolytes during LT stress. Content of (a) Proline and (b) total soluble sugar in tomato leaves under low temperatures. All experiments were performed in triplicate for three independent experiments. Values are presented as means ± SD. Different letters denote significant differences between means according to Tukey HSD post hoc test p ≤ 0.05.
Figure 3.
Trichoderma asperellum enhances the accumulation of osmolytes during LT stress. Content of (a) Proline and (b) total soluble sugar in tomato leaves under low temperatures. All experiments were performed in triplicate for three independent experiments. Values are presented as means ± SD. Different letters denote significant differences between means according to Tukey HSD post hoc test p ≤ 0.05.
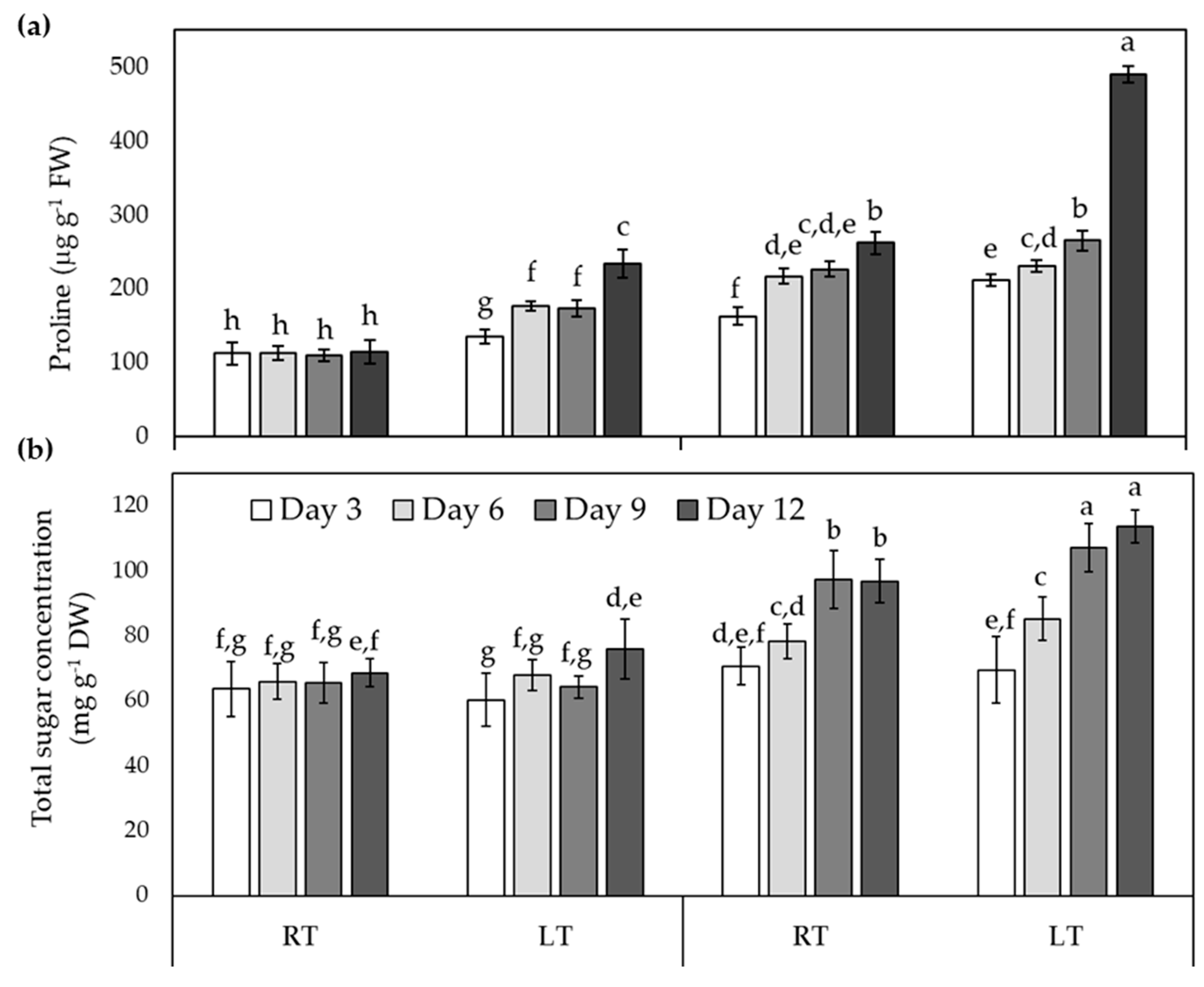
Figure 4.
Trichoderma asperellum modifies stomatal traits related to the photosynthetic optimization in tomato plants under LT. Responses of (a) Stomatal density, (b) Stomatal index and (c) Stomatal area on abaxial leaf surface from tomato plant under room- (RT) or low-temperatures (LT). Values are presented as means ± SD. Different letters above bars indicate significant differences (n=30).
Figure 4.
Trichoderma asperellum modifies stomatal traits related to the photosynthetic optimization in tomato plants under LT. Responses of (a) Stomatal density, (b) Stomatal index and (c) Stomatal area on abaxial leaf surface from tomato plant under room- (RT) or low-temperatures (LT). Values are presented as means ± SD. Different letters above bars indicate significant differences (n=30).
Figure 5.
Trichoderma asperellum remains viable under LT exposure. Fungal viability in the growing media of tomato plants (a) or semi-synthetic growth medium (b). (a) CFU per gram was quantified in soil samples collected from tomato plants under LT treatment. The population was expressed as Log CFU/gram of soil. (b) Mycelial growth of T. asperellum at 28 °C (5 days), 4 °C (5 days) and 4 °C (5 days) + 28 °C (5 days). Values are presented as means ± SD from three independent experiments.
Figure 5.
Trichoderma asperellum remains viable under LT exposure. Fungal viability in the growing media of tomato plants (a) or semi-synthetic growth medium (b). (a) CFU per gram was quantified in soil samples collected from tomato plants under LT treatment. The population was expressed as Log CFU/gram of soil. (b) Mycelial growth of T. asperellum at 28 °C (5 days), 4 °C (5 days) and 4 °C (5 days) + 28 °C (5 days). Values are presented as means ± SD from three independent experiments.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



