
Submitted:
18 November 2025
Posted:
19 November 2025
You are already at the latest version
Abstract
Drought is a major threat to global food security, with climate change enhancing its severity. Living organisms employ various adaptation strategies against loss of water, with desiccation tolerance representing the most extreme ones. The present study provides an analysis of desiccation response mechanisms in Pisum sativum L., focusing on the early seedling development. The experiments were carried out on 3-day-old seedlings with radicle up to 20 mm and similar seedlings with cotyledons removed, each subjected to the 'desiccation and rehydration' treatment. We assessed stress responses using growth and electrolyte leakage tests, and by measuring ascorbate content and lipid peroxidation products. The results demonstrated that while all desiccated seedlings experienced oxidative stress, seedlings with cotyledons exhibited superior recovery capacity, e.g. maintaining membrane integrity, ascorbate status and the ability to form the adventitious roots. We also monitored the ABA-dependent stress response by registering expression of several genes associated with "response to water deprivation" (ABI3, ABI4, ABI5, HVA22, PER1, LEA14, RD22-1/2/3, LTI65, and LTP4). Intrudingly, 'desiccation and rehydration' treatment resulted in the 40-fold up-regulation of ABI5 in hypocotyls of seedlings both with removed and intact cotyledons and the 108-fold up-regulation of this gene only in roots of seedlings with intact cotyledons. We propose that the post-germination stage represents a ‘resilience window’ where embryonic protective mechanisms remain partially active while the plant prepares for autotrophic growth. Herewith, the role of cotyledons extends beyond their classical role as nutrient reserves by coordinating both developmental and stress responses under conditions of severe water stress.
Keywords:
desiccation tolerance
; post-germination
; seedling development
; cotyledons
; ABA signaling
Abstract
Drought is a major threat to global food security, with climate change enhancing its severity. Living organisms employ various adaptation strategies against loss of water, with desiccation tolerance representing the most extreme ones. The present study provides an analysis of desiccation response mechanisms in Pisum sativum L., focusing on the early seedling development. The experiments were carried out on 3-day-old seedlings with radicle up to 20 mm and similar seedlings with cotyledons removed, each subjected to the 'desiccation and rehydration' treatment. We assessed stress responses using growth and electrolyte leakage tests, and by measuring ascorbate content and lipid peroxidation products. The results demonstrated that while all desiccated seedlings experienced oxidative stress, seedlings with cotyledons exhibited superior recovery capacity, e.g. maintaining membrane integrity, ascorbate status and the ability to form the adventitious roots. We also monitored the ABA-dependent stress response by registering expression of several genes associated with "response to water deprivation" (ABI3, ABI4, ABI5, HVA22, PER1, LEA14, RD22-1/2/3, LTI65, and LTP4). Intrudingly, 'desiccation and rehydration' treatment resulted in the 40-fold up-regulation of ABI5 in hypocotyls of seedlings both with removed and intact cotyledons and the 108-fold up-regulation of this gene only in roots of seedlings with intact cotyledons. We propose that the post-germination stage represents a ‘resilience window’ where embryonic protective mechanisms remain partially active while the plant prepares for autotrophic growth. Herewith, the role of cotyledons extends beyond their classical role as nutrient reserves by coordinating both developmental and stress responses under conditions of severe water stress.
1. Introduction
Drought represents one of the most significant threats to global food security, with drought stress becoming increasingly severe due to climate change [1]. Even moderate dehydration significantly reduces plant productivity, forcing organisms to employ diverse adaptive survival mechanisms [2,3,4]. Among these adaptations, desiccation tolerance defined as the ability to survive drying below −100 MPa and resume metabolic activity upon rehydration [5], stands out as one of the most extreme responses to water limitation [6,7].
This remarkable trait likely originated in early land plants as an adaptation to periodic drying, playing a pivotal role in life's transition from aquatic to terrestrial environments [8,9]. In modern plants, desiccation tolerance persists in two main forms including resurrection plants inhabiting arid regions and orthodox seeds [6,10,11,12,13]
The maturation of orthodox seeds is accompanied by water loss of up to 5–10% w/w, enabling them to withstand extreme environmental conditions during the period of dormancy [12,14,15]. The master regulators of seed maturation include the LAFL network, which includes the transcription factors LEAFY COTYLEDON1 (LEC1), ABSCISIC ACID INSENSITIVE3 (ABI3), FUSCA3 (FUS3), and LEC2, along with the primary dormancy regulator DELAY OF GERMINATION1 (DOG1) and ABSCISIC ACID INSENSITIVE5 (ABI5) [12,16,17,18]. This specific group of proteins plays a pivotal role in establishing and maintaining embryonic cell identity through precise regulation of genes controlling embryogenesis, germination repression, reserve substance accumulation and stress resistance including tolerance to desiccation.
Mechanisms of desiccation tolerance are activated during the late maturation of orthodox seeds and involve the accumulation of late embryogenesis abundant (LEA) proteins, small heat shock proteins (sHSPs), lipid droplets, non-reducing oligosaccharides, and low-molecular-weight antioxidants [7,14,19,20,21,22,23,24,25]. These reactions allow seeds to survive during dry storage without losing their viability [12]. The key mechanism in response to desiccation, as well as part of the adaptive strategy, is the synthesis of reactive oxygen species (ROS) [9,26]. ROS can damage plants during osmotic stress, but they are also capable of switching genetic programs [27].
Notably, orthodox seeds maintain desiccation tolerance not only during dormancy but also throughout germination, persisting until the transition to the post-germination stage [28]. This tolerance loss is widely attributed to the repression of LAFL network genes [16,29,30]. However, our experiments with pea seeds revealed that only PsABI3 exhibited detectable expression in embryonic axes prior to radicle protrusion, while LEC1, FUS3, and LEC2 were suppressed early, likely at the late maturation [31].
The transition from germination to post-germination involves a comprehensive reprogramming of stress resistance response, including downregulation of ABA signaling, dehydration and cold adaptation genes, reduced synthesis of LEA proteins, small heat shock proteins (sHSPs), and raffinose-family oligosaccharides with up-regulation of genes associated with resistance to pathogens, oxidative stress, and secondary metabolism [31,32]. Thus, the transition from seeds to seedlings signifies a critical shift in stress adaptation. Cells lose the ability to survive water potentials below −5 MPa and develop a dehydration sensitivity that is suited for vegetative growth.
Seeds germinating in the soil bank frequently experience multiple imbibition-drying cycles due to alternating rain and drought events. Water in the topsoil evaporates rapidly, creating diurnal moisture fluctuations [33]. To adapt, plants have evolved unique strategies to resume not only germination but also seedling growth after imbibition-drying cycles, though this capacity is restricted to the post-germination stage following radicle emergence. Bruggink and van der Toorn (1995) were the first to report on the re-induction of desiccation tolerance in seedlings by priming germinating seeds with polyethylene glycol (PEG) [34]. Subsequent studies showed that the ability to regain desiccation tolerance is limited to radicles lengths: 1–3 mm in Medicago sativa [35], 2 mm in Pisum sativum [36], 4 mm in Solanum lycopersicum [37] and Vigna unguiculata [38].
The present study aimed to decipher the desiccation response in seedlings of Pisum sativum L. at the later stage of development, with embryonic roots up to 20 mm-long. We dried intact seedlings and seedlings with removed cotyledons, thereby revealing the role of cotyledons in the drought tolerance. To understand the role of ABA in the response to seedling desiccation, changes in the expression of abscisic acid (ABA)-dependent genes associated with "response to water deprivation" were studied in hypocotyls and roots after drying followed by rehydration. Thus, we examined the possibility of activating ABA-dependent desiccation tolerance genes that are normally suppressed during the transition from seed to seedling stage at post-germination.
2. Results
2.1. Effects of ‘Drying and Rehydration’ Treatment on Growth of Pisum sativum L. Seedlings
This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
At the first stage, we examined the response of 3-day-old pea seedlings to desiccation. Seeds were germinated between layers of moist filter paper for 72 h, after which the seedlings were subjected to drying in a desiccator over silica gel for 24 h – either with or without cotyledons (Figure S1). Following desiccation, the seedlings were rehydrated between moist filter paper for another 24 h. Figure 1 demonstrates the appearance of the 3-day-old pea seedlings in control (Figure 1a), after drying and rehydration without cotyledons (Figure 1b) and after drying and rehydration with cotyledons (Figure 1c).
The viability of embryonic axes isolated from seedlings was assessed using a tetrazolium-topographic test (Figure 2). In control seedlings, embryonic axes displayed uniform staining in hypocotyls and roots, with more intense staining at the root tips, indicating higher dehydrogenase activity (Figure 2a). In embryonic axes dried and rehydrated without cotyledons, staining was weaker, suggesting reduced metabolic activity (Figure 2b). These seedlings remained viable for only 3 days after post-rehydration, likely due to nutrient deprivation. In embryonic axes dried and rehydrated with cotyledons, the primary roots showed damage, but adventitious roots formed (Figure 1c, 2c). By 3rd day after post-rehydration, the primary roots died, but at the same time new adventitious roots have grown (Figure 1d).
2.2. Physiological and Biochemical Responses of Pisum sativum L. Seedlings to ‘Drying and Rehydration’ Treatment
Effect of ‘drying and rehydration’ treatment on the water potential of plant tissues was studied with using a Dewpoint PotentiaMeter WP4-C (Decagon Devices, USA). Water potential of hypocotyls and roots isolated from control seedlings was -3.3 ± 0.3 MPa and -2.8 ± 0.1 MPa, correspondingly (Figure 3a). Drying and rehydration of seedlings with cotyledons resulted in a 1.4-fold increase of water potential in hypocotyls with no changes in roots. Drying and rehydration of seedlings without cotyledons resulted in a 2-fold increase of water potential in hypocotyls and a 2.5-fold increase of water potential in roots.
The integrity of cell membranes was assessed by measurement of total amount of electrolytes released from embryonic axes into the incubation solution [39]. This method serves as a sensitive indicator of membrane damage under various stresses including drought, salinity, pathogen attack, and extreme temperatures [40]. Our experiments revealed that drying and subsequent rehydration, whether with or without cotyledons, consistently increased electrolyte leakage by 1.7-fold in hypocotyls and 1.5–1.6-fold in roots, in comparison to control (Figure 3b).
The levels of MDA and other TBA-reactive lipid peroxidation products in hypocotyls and roots of pea seedlings are presented in Figure 4a. Drying followed by rehydration of 3-day-old seedlings with intact cotyledons did not alter lipid peroxidation levels in either hypocotyls or roots. However, drying of seedlings with removed cotyledons led to a 1.7-fold increase in lipid peroxidation products in the roots.
Ascorbate was present in both hypocotyls and roots of 3-day-old pea seedlings in its reduced form (Figure 4b). The ascorbate content was 22 ± 0.4 μmol/g DW in hypocotyls and 24 ± 0.8 μmol/g DW in roots of control seedlings. Drying and subsequent rehydration of seedlings, either with and without cotyledons, resulted to decreased ascorbate levels in both hypocotyls and roots.
2.3. Changes in Expression of ABA-Depended Genes
To reveal the effect of the ‘drying and rehydration’ treatment on ABA signaling, we analyzed the expression of genes associated with "response to water deprivation" and "response to abscisic acid (ABA)," as classified in the Gene Ontology (GO) database [41]. From the "response to water deprivation" group, we selected the following genes, identified in our previous work for pea embryonic axes [31]: ABA INSENSITIVE 3/4/5 (ABI3, ABI4, ABI5), HORDEUM VULGARE ABUNDANT 22 (HVA22), LATE EMBRYOGENESIS ABUNDANT 14 (LEA14), 1-CYSTEINE PEROXIREDOXIN (PER1), RESPONSIVE TO DEHYDRATION 22 (RD22-1/2/3), LOW-TEMPERATURE INDUCED 65 (LTI65) and LIPID TRANSFER PROTEIN 4 (LTP4). Previously, we demonstrated that the expression of these genes decreases in pea embryonic axes during the transition from germination stage to the post-germination stage [31,32]. Here, we analyzed the role of cotyledons in the relative expression of these genes in hypocotyls and roots from 3-day-old seedlings after their ‘drying and rehydration’. Figure 5 shows the behavior of the ABI3, ABI4, and ABI5 genes, which encode transcription factors significant for seed maturation, longevity, and germination [42,43,44]. The constitutive expression of these genes was very low both in hypocotyls and roots of control seedlings. The ‘drying and rehydration’ treatment induced an increase in level of expression, indicating gene reactivation under water loss. The ABI3 and ABI4 were up-regulated in the hypocotyls of treated seedlings, both with cotyledons (4-fold and 3-fold, respectively) and without cotyledons (5-fold and 7-fold, respectively), with no significant changes observed in the roots (Figure 5). The most significant changes were observed for ABI5 gene, the expression of which increased 40-fold in hypocotyls and 108-fold in the roots of seedlings after ‘drying and rehydration’ with cotyledons.
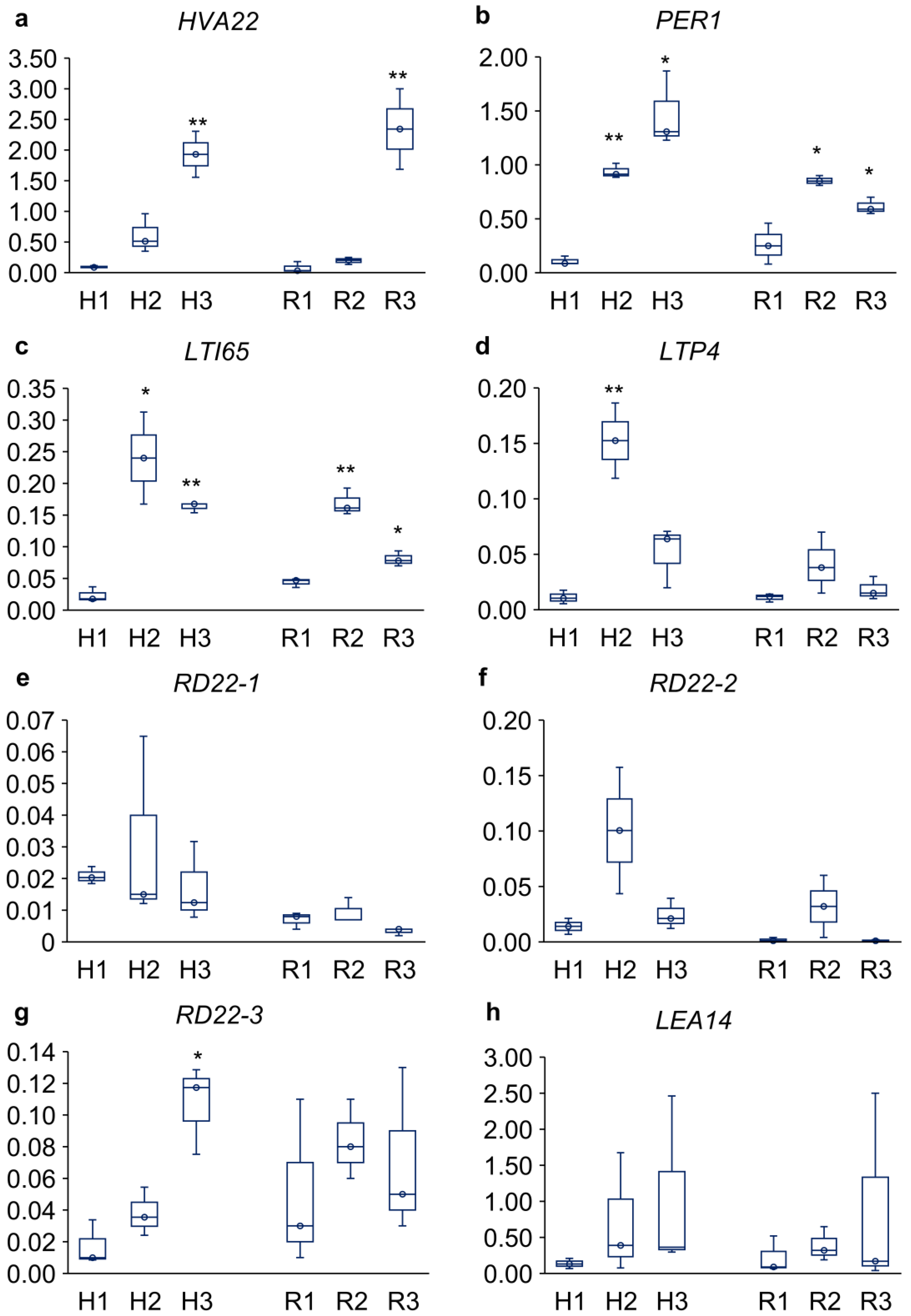
The changes in expression levels of HVA22E, PER1, LTI65, LTP4, three RD22 copies, and LEA14 are presented in Figure 6. The expression level of HVA22 increased in both the hypocotyls and roots of seedlings dried and rehydrated without cotyledons (21-fold and 30-fold, respectively; Figure 6a). Up-regulation of PER1 and LTI65 was detected in both hypocotyls and roots after drying and rehydration, regardless of cotyledon presence. In hypocotyls, PER1 was up-regulated 9-fold in seedlings treated with cotyledons and 14-fold in those without cotyledons (Figure 6b). Similarly, in hypocotyls, expression of LTI65 was increased tenfold in seedlings treated with cotyledons and 7-fold in those without cotyledons (Figure 6c). In the roots, this effect was weaker. LTP4 expression was altered only in the hypocotyls of seedlings with intact cotyledons, showing a 14-fold increase (Figure 6d). Among the three RD22 copies, only RD22-3 was up-regulated in the hypocotyls of seedlings without cotyledons (6-fold; Figure 6g). In all tests, LEA14 showed no significant response to the ‘drying and rehydration’ treatment (Figure 6h).
3. Discussion
The ability to withstand drying represents one of biology's most astonishing survival strategies. While vascular plants typically require substantial water to maintain structure and function, their seeds (orthodox type) exhibit the extraordinary ability to withstand near-total water loss, not only during dormancy but also during germination [5,13]. This "window of desiccation tolerance" closes as plants transit from the seed to seedling stage of ontogenesis [45]. However, the molecular mechanisms governing this transition remain poorly understood.
In our model system, we investigated the desiccation response in 3-day-old pea seedlings. Seedlings with emerged roots up to 20 mm-long were subjected to drying followed by rehydration, either with or without cotyledons. Previous studies have demonstrated the ability to re-establish desiccation tolerance in young seedlings of some species, including Cucumis sativus, Impatiens walleriana, Solanum lycopersicum, Medicago sativa, Arabidopsis thaliana and Pisum sativum [34,35,36,37,46] These studies also reported that the phenotypic ‘window’ for supporting the dehydration tolerance is typically limited to 2–4 mm of radicle elongation. Intriguingly, we observed that P. sativum seedlings with radicles up to 20 mm in length survived desiccation, maintaining cell viability (Figure 2) and resuming growth after rehydration (Figure 1). However, further seedling development depended critically on cotyledon presence. Without cotyledons, embryonic axes remained viable for about three days after post-rehydration and then died (Figure 1b). But with cotyledons seedlings dropped their primary roots, developed adventitious roots (Figure 1c), and continued growth via the newly developed root system (Figure 1d). In dicotyledon plants, such as peas, cotyledons serve as storage organs for reserve nutrients that support germination and initial growth until the true leaves form [47]. Stored macromolecules are hydrolyzed into soluble sugars, amino acids, and fatty acids, which provide energy and necessary components for the development of the embryonic axis [48,49,50]. Thus, the survival of dried and rehydrated embryonic axes primarily depends on the metabolic resources derived from cotyledons.
To assess the physiological response of embryonic axes to drying, we measured water potential, cell membrane integrity, TBA-reactive lipid peroxidation product levels, and ascorbate content in hypocotyls and roots taken from 3-day-old pea seedlings after their drying and rehydration (Figure 3).
In plant biology, water potential serves as a key indicator of water's capacity to participate in metabolic processes [1]. Leaves typically exhibit water potentials ranging from -1.5 to -0.7 MPa, while dry seeds can show values from -50 to -350 MPa [47]. During germination, as seed moisture content increases, water potential becomes less negative [51]. In our study, hypocotyls and roots isolated from control seedlings showed water potential around -3 MPa, approaching values typical of adult plants (Figure 3a). However, drying without cotyledons followed by rehydration caused a 2–2.5-fold increase in water potential. This was accompanied by approximately 1.5-fold increases in both electrolyte leakage and lipid peroxidation products, indicating some damage of cell membranes (Figure 3b and 4a). Lipids represent very important targets for some ROS [52], and, if oxidized, serve as crucial markers of oxidative stress in plants [53].
L-ascorbic acid is a major antioxidant in higher plants, which plays crucial roles in protection against ROS, redox metabolism and signaling both in photosynthesizing tissues and roots [54,55,56]. In our experiments, the ascorbate level in hypocotyl and root cells of control seedlings was similar (22–24 μmol/g DW) (Figure 4b). Following desiccation and rehydration of embryonic axes, this value decreased which can be considered an indicator of ROS generation and oxidative stress.
It is well-known fact that seed germination is characterized by a shift in phytohormone balance, primarily between abscisic acid (ABA) and gibberellins (GA) [47]. While ABA maintains seed dormancy and suppresses germination, GA breaks dormancy and activates the germination program [28,57,58,59]. ABA also controls plant stress responses, with particularly well-characterized roles in drought adaptation [60,61].
Our previous studies provided insight into the involvement of ABA in the transition of Pisum sativum from the germination to post-germination stages [31,32]. A decrease in the ABA level with an accumulation of its catabolites (phaseic acid, dihydrophaseic acid, and neo-phaseic acid) was observed [32]. The changes in the level of ABA metabolites were accompanied with the downregulation of genes controlling “water deprivation” [32]. Additionally, the expression of ABI3, ABI4, and ABI5, which encode crucial transcription factors during late maturation [42,43,44], was 20-fold down-regulated with the high level of their methylation [31,32]. To elucidate the molecular mechanisms underlying desiccation response in embryonic axes under early pea seedling development, we analyzed the expression of ABA-associated genes linked to both "response to water deprivation" and "response to abscisic acid" (ABI3, ABI4, ABI5, HVA22, LEA14, PER1, RD22-1, RD-22-2, RD22-3, LTI65 and LTP4) (Figure 5 and 6). Our study revealed that desiccation significantly altered expression patterns of the target genes in both hypocotyls and roots of 3-day-old pea seedlings.
The bZIP transcription factor ABI5 plays a key role in ABA signaling during seed development by integrating hormonal crosstalk between ABA and other phytohormones [46,62,63,64]. ABI5 transcripts and proteins accumulate during seed maturation but decline sharply during seed germination [59,65,66]. Lopes-Molina et al. [67] has established that ABI5 expression defines a developmental checkpoint post-germination, enabling environmental osmotic status monitoring before vegetative growth commences. Recent evidence suggested that ABI5 can function as a terminal inhibitor of multiple signals regulating seed germination, including not only ABA and GA but also other hormones and light [59,65]. In our study, we observed a striking 108-fold up-regulation of ABI5 in roots of seedlings subjected to drying-rehydration cycles with cotyledons (Figure 5). Notably, these seedlings subsequently dropped their primary roots and developed a new adventitious root system.
Among the studied genes, HVA22 exhibited the highest constitutive expression (Figure 6a). The HVA22 gene was originally discovered in barley aleurone layer cells [68] and initially showed limited homology with ABA-responsive genes such as LEA and RAB (Responsive to ABA) [69]. Induced by ABA and various stressors, HVA22 gene family encodes stress-responsive proteins with a conserved TB2/DP1/HVA22 domain that plays crucial roles in adaptation [70,71,72,73]. HVA22 family proteins, characterized by conserved TB2/DP1/HVA22 domains, are ubiquitous in eukaryotes [70,74]. Sequence analysis revealed high similarity between plant HVA22s and human TB2/DP1 proteins [70,74]. Expression of HVA22 : green fluorescent protein fusion proteins in barley aleurone layer cells revealed that HVA22 is an endoplasmic reticulum (ER)-Golgi-localized protein that negatively regulates gibberellic acid (GA)-mediated vacuolization and programmed cell death [75]. Authors proposed that accumulation of HVA22 proteins may inhibit the vesicular trafficking involved in nutrient mobilization in germinated seed, thereby delaying the coalescence of protein storage vacuoles [75]. In Arabidopsis thaliana, HVA22 gene family exhibit responsiveness to multiple abiotic stresses, including low-temperature exposure, salt stress, ABA treatment and dehydration [76]. Our experiments revealed HVA22 induction in cotyledon-excised pea seedlings following ‘desiccation and rehydration’ treatment, with 21-fold up-regulation in hypocotyls and 30-fold up-regulation in roots (Figure 6a). These findings highlight the crucial role of HVA22 in ABA-mediated signaling pathways potentially enhancing drought tolerance in juvenile seedlings of P. sativum.
Plant peroxiredoxins (Prxs) are a family of thiol-dependent peroxidases essential for antioxidant defense and ROS signaling [77]. Localized in cellular compartments, including chloroplasts and mitochondria, they scavenge ROS, shaping redox signal. Among these, a seed-specific 1-cysteine peroxiredoxin (PER1) serves as a key antioxidant protein responsible for ROS scavenging in seeds [78]. PER1 gene is predominantly expressed during seed developmental stages associated with ROS production, particularly during late maturation and desiccation [78]. While PER1 expression is typically suppressed during germination, evidence indicates its reactivation under abiotic stress conditions [79,80]. Adverse environmental conditions may stabilize 1-Cys Prx by inhibiting its degradation, leading to reduced ROS accumulation, attenuated ABA catabolism and consequently, delayed germination [80]. PER1 expression showed significant up-regulation in response to ‘desiccation and rehydration’ treatment. Embryonic axes of seedlings with intact cotyledons and seedlings with removed cotyledons demonstrated 8.6-fold and 13.5-fold increase after ‘drying and rehydration’ treatment, respectively (Figure 6b). Roots also showed moderate PER1 up-regulation (~3-fold increase) both in seedlings with cotyledons and seedlings without them.
Altogether, our study demonstrated that Pisum sativum L. seedlings maintain a remarkable capacity to withstand desiccation during early post-germination stages, depending on the presence of intact cotyledons and involving ROS accumulation and ABA-responsive pathways.
4. Materials and Methods
4.1. Plant Material and Growth Conditions
Pea (Pisum sativum L., cv. Prima) seedlings were used in the study. Seeds were germinated between layers of moist filter paper at 22°C for 3 days until radicle emergence. Seedlings with emerged roots up to 20 mm long were then divided into two groups including one with intact cotyledons and another with excised cotyledons (cut by a razor blade). Both intact and cotyledon-removed seedlings were dried in a desiccator over blue silica gel (Central Drug House LTD, India) at 22°C for 24 h. After desiccation, the seedlings were rehydrated between layers of moist filter paper for 24 h at 22°C. For experiments embryonic axes from three sets of 3-day-old seedlings were isolated including untreated seedlings (control), seedlings dried and rehydrated with cotyledons, and seedlings dried and rehydrated without cotyledons.
4.2. Tissue Viability
Embryonic axes isolated from 3-day-old pea seedlings were incubated in a 1% (w/v) aqueous solution of tetrazolium chloride (pH 7.0) for 1 h at room temperature, followed by visual evaluation of staining intensity. The method relies on the reduction of colorless tetrazolium chloride to red formazan by dehydrogenases in metabolically active cells [81]. Viable tissues stain red due to formazan accumulation, whereas nonviable tissues remain colorless.
4.3. Water Potential and Cell Membrane Integrity
The water potential of plant tissues was determined using a Dewpoint PotentiaMeter WP4-C (Decagon Devices, USA). The device determines water potential by measuring the relative humidity of air within a sealed chamber above the sample.
Cell membrane integrity was evaluated by measuring the total amount of electrolytes released from plant tissue into the incubation solution [39]. Embryonic axes were incubated in distilled water for 30 minutes, and electrical conductivity was subsequently measured using the HI8733 conductivity meter (HANNA, Germany).
4.4. Biochemical Stress Markers
Stress markers evaluation included measurements of the level of lipid peroxidation and concentration of L-ascorbic acid in embryonic axes of pea seedlings. The embryonic axes were frozen in liquid nitrogen and homogenized with in a Mixer Mill MM 400 ball mill (Retsch, Haan, Germany) with two stainless steel balls (ø 15 mm) at a vibration frequency of 30 Hz for 2 × 30 s. Approximately 30 and 55 mg of frozen grinded plant material were used for the lipid peroxidation and ascorbate analyses, respectively. Lipid peroxidation was measured using the thiobarbituric acid (TBA) test, which quantified malondialdehyde (MDA), a product of lipid peroxidation, as described elsewhere [82]. Ascorbic acid was quantified using the method described by Huang and co-workers [83] with modifications [84].
4.5. RNA Isolation and Real-Time PCR Analysis
The embryonic axes were frozen in liquid nitrogen and homogenized with a pestle in a mortar. Total RNA was isolated using the RNeasy Plant Mini Kit (QIAGEN) according to the manufacturer's protocol, followed by DNase treatment with the RNase-free DNase set (QIAGEN) (www.qiagen.com). Electrophoretic analysis was performed in a 1% agarose gel prepared in TAE buffer with the addition of ethidium bromide as an intercalating dye. The DNA molecular weight marker "Sky-High 250 bp - 10 kb" (BioLabMix, Novosibirsk) was used as a size standard. The isolation products were visualized using the BioRad ChemiDoc MP gel documentation system (Bio-Rad Laboratories, Moscow). The concentration of isolated RNA was measured using a NanoDrop™ 2000/2000c spectrophotometer (Thermo Fisher Scientific Inc.). Single-stranded cDNA was synthesized from the RNA template using the OT M-MuLV-RH kit (Biolabmix, Novosibirsk). Quantitative PCR was performed using the SYNTOL SYBR Green I+ROX kit (Syntol) on a CFX-96 instrument (BioRad). Primer design was performed using IDT PrimerQuest software (http://eu.idtdna.com/PrimerQuest/Home) (Table S1). The gene encoding phosphoprotein phosphatase 2A was selected as the reference gene [85].
4.6. Statistical Analysis
The statistical analysis was conducted utilizing an MS Excel add-in. The significance of differences between control and treatments was estimated using Students' two-tailed t-test (α = 0.05, p ≤ 0.05). The calculation of biochemical parameters was conducted using five biological replicates. The gene expression levels in the qPCR analysis were derived from the three biological and three technical replicates.
5. Conclusions
The presented work primarily declares the importance of intact cotyledons for young pea seedlings to survive during desiccation. Cotyledons removal severely compromises viability of seedlings at the post-germination stage. Embryonic axes with cotyledons exhibit a remarkable capacity to survive desiccation even after radicle elongation up to 20 mm in length with formation of adventitious roots instead of the damaged primary root. Essential to this adaptation is apparently the activation of ABA-mediated signal pathways. The massive 40-fold up-regulation of ABI5 in hypocotyls and 108-fold up-regulation in roots of seedlings, which were dried and rehydrated with intact cotyledons, points to the pivotal role of this transcription factor in the desiccation response. Increase of expression of HVA22 (21-30-fold) and PER1 (8-14-fold) genes indicates the importance of vesicular transport, programmed cell death and ROS detoxication in the tolerance to desiccation. These findings redefine our understanding of drought stress tolerance during plant growth, positioning post-germination stage as a unique niche for deciphering the resilience mechanisms activated by severe water loss.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/doi/s1, Figure S1: Appearance of embryonic axes of 3-day-old pea seedlings after drying with and without cotyledons; Table S1: List of primers used in this work.
Author Contributions
Conceptualization, G.S.; formal analysis, A.L. and P.V.; investigation, T.B. and E.K.; data curation, A.L., P.V. and E.K.; writing—original draft preparation, G.S., V.D. and S.M.; writing—review and editing, G.S., V.D. and S.M.; supervision, S.M.; project administration, G.S.; funding acquisition, S.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation, grant no. 25-16-00215.
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Juenger, T.E.; Verslues, P.E. Time for a drought experiment: Do you know your plants’ water status? Plant Cell 2023, 35, 10–23. [Google Scholar] [CrossRef]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants: implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Farrant, J.M.; Hilhorst, H. Crops for dry environments. Curr. Opin. Biotechnol. 2022, 74, 84–91. [Google Scholar] [CrossRef]
- Cooper, M.; Messina, C.D. Breeding crops for drought-affected environments and improved climate resilience. Plant Cell 2023, 35, 162–186. [Google Scholar] [CrossRef]
- Marks, R.A.; Ekwealor, J.T.B.; Artur, M.A.S.; Bondi, L.; Boothby, T.C.; Carmo, O.M.S.; Centeno, D.C.; Coe, K.K.; Dace, H.J.W.; Field, S.; et al. Life on the dry side: a roadmap to understanding desiccation tolerance and accelerating translational applications. Nat. Commun. 2025, 16, 3284. [Google Scholar] [CrossRef]
- Farrant, J.M.; Moore, J.P. Programming desiccation-tolerance: from plants to seeds to resurrection plants. Curr. Opin. Plant Biol. 2011, 14, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Rosete-Enríquez, M.; Juárez-González, V.R.; Escobar-Muciño, E.; Muñoz-Rojas, J.; Quintero-Hernández, V. Surviving desiccation: key factors underlying tolerance in prokaryotes and eukaryotes. Protoplasma 2025. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, A.L.; Castelle, C.J.; Banfield, J.F. Habitat transition in the evolution of bacteria and archaea. Annu. Rev. Microbiol. 2023, 77, 193–212. [Google Scholar] [CrossRef] [PubMed]
- Oliver, M.J.; Farrant, J.M.; Hilhorst, H.W.M.; Mundree, S.; Williams, B.; Bewley, J.D. Desiccation tolerance: Avoiding cellular damage during drying and rehydration. Annu. Rev. Plant Biol. 2020, 71, 435–460. [Google Scholar] [CrossRef]
- Costa, M.-C.D.; Cooper, K.; Hilhorst, H.W.M.; Farrant, J.M. Orthodox seeds and resurrection plants: two of a kind? Plant Physiol. 2017, 175, 589–599. [Google Scholar] [CrossRef]
- Gao, B.; Li, X.; Liang, Y.; Chen, M.; Liu, H.; Liu, Y.; Wang, J.; Zhang, J.; Zhang, Y.; Oliver, M.J.; et al. Drying without dying: A genome database for desiccation-tolerant plants and evolution of desiccation tolerance. Plant Physiol. 2023, kiad672. [Google Scholar] [CrossRef]
- Smolikova, G.; Leonova, T.; Vashurina, N.; Frolov, A.; Medvedev, S. Desiccation tolerance as the basis of long-term seed viability. Int. J. Mol. Sci. 2020, 22, 101. [Google Scholar] [CrossRef]
- Marques, A.; Buijs, G.; Ligterink, W.; Hilhorst, H. Evolutionary ecophysiology of seed desiccation sensitivity. Funct. Plant Biol. 2018, 45, 1083. [Google Scholar] [CrossRef] [PubMed]
- Leprince, O.; Pellizzaro, A.; Berriri, S.; Buitink, J. Late seed maturation: drying without dying. J. Exp. Bot. 2016, 68, erw363. [Google Scholar] [CrossRef] [PubMed]
- Matilla, A.J. The Orthodox dry seeds are alive: A clear example of desiccation tolerance. Plants 2021, 11, 20. [Google Scholar] [CrossRef]
- Lepiniec, L.; Devic, M.; Roscoe, T.J.; Bouyer, D.; Zhou, D.-X.; Boulard, C.; Baud, S.; Dubreucq, B. Molecular and epigenetic regulations and functions of the LAFL transcriptional regulators that control seed development. Plant Reprod. 2018, 31, 291–307. [Google Scholar] [CrossRef]
- Alizadeh, M.; Hoy, R.; Lu, B.; Song, L. Team effort: Combinatorial control of seed maturation by transcription factors. Curr. Opin. Plant Biol. 2021, 63, 102091. [Google Scholar] [CrossRef]
- Li, Q.; Chen, X.; Zhang, S.; Shan, S.; Xiang, Y. DELAY OF GERMINATION 1, the master regulator of seed dormancy, integrates the regulatory network of phytohormones at the transcriptional level to control seed dormancy. Curr. Issues Mol. Biol. 2022, 44, 6205–6217. [Google Scholar] [CrossRef]
- Artur, M.A.S.; Zhao, T.; Ligterink, W.; Schranz, E.; Hilhorst, H.W.M. Dissecting the genomic diversification of late embryogenesis abundant (LEA) protein gene families in plants. Genome Biol. Evol. 2019, 11, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Li, Y.; Fang, T.; Shi, X.; Chen, X. Specific roles of tocopherols and tocotrienols in seed longevity and germination tolerance to abiotic stress in transgenic rice. Plant Sci. 2016, 244, 31–39. [Google Scholar] [CrossRef]
- Nagaraju, M.; Kumar, S.A.; Reddy, P.S.; Kumar, A.; Rao, D.M.; Kavi Kishor, P.B. Genome-scale identification, classification, and tissue specific expression analysis of late embryogenesis abundant (LEA) genes under abiotic stress conditions in Sorghum bicolor L. PLoS One 2019, 14, e0209980. [CrossRef]
- Roach, T.; Nagel, M.; Börner, A.; Eberle, C.; Kranner, I. Changes in tocochromanols and glutathione reveal differences in the mechanisms of seed ageing under seedbank conditions and controlled deterioration in barley. Environ. Exp. Bot. 2018, 156, 8–15. [Google Scholar] [CrossRef]
- Sano, N.; Rajjou, L.; North, H.M.; Debeaujon, I.; Marion-Poll, A.; Seo, M. Staying alive: molecular aspects of seed longevity. Plant Cell Physiol. 2016, 57, 660–674. [Google Scholar] [CrossRef]
- de Vries, J.; Ischebeck, T. Ties between stress and lipid droplets pre-date seeds. Trends Plant Sci. 2020, 25, 1203–1214. [Google Scholar] [CrossRef]
- Bouchnak, I.; Coulon, D.; Salis, V.; D’Andréa, S.; Bréhélin, C. Lipid droplets are versatile organelles involved in plant development and plant response to environmental changes. Front. Plant Sci. 2023, 14, 1–17. [Google Scholar] [CrossRef]
- Pagano, A.; Folini, G.; Pagano, P.; Sincinelli, F.; Rossetto, A.; Macovei, A.; Balestrazzi, A. ROS accumulation as a hallmark of dehydration stress in primed and overprimed Medicago truncatula seeds. Agronomy 2022, 12, 268. [Google Scholar] [CrossRef]
- Demidchik, V. Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Smolikova, G.; Strygina, K.; Krylova, E.; Leonova, T.; Frolov, A.; Khlestkina, E.; Medvedev, S. Transition from seeds to seedlings: hormonal and epigenetic aspects. Plants 2021, 10, 1884. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Attuluri, V.P.S.; Robert, H.S. Transcriptional control of Arabidopsis seed development. Planta 2022, 255, 1–19. [Google Scholar] [CrossRef]
- Han, J.-D.; Li, X.; Jiang, C.-K.; Wong, G.K.-S.; Rothfels, C.J.; Rao, G.-Y. Evolutionary analysis of the LAFL genes involved in the land plant seed maturation program. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Smolikova, G.; Strygina, K.; Krylova, E.; Vikhorev, A.; Bilova, T.; Frolov, A.; Khlestkina, E.; Medvedev, S. Seed-to-seedling transition in Pisum sativum L.: a transcriptomic approach. Plants 2022, 11, 1686. [Google Scholar] [CrossRef] [PubMed]
- Smolikova, G.; Krylova, E.; Petřík, I.; Vilis, P.; Vikhorev, A.; Strygina, K.; Strnad, M.; Frolov, A.; Khlestkina, E.; Medvedev, S. Involvement of abscisic acid in transition of pea (Pisum sativum L.) seeds from germination to post-germination stages. Plants 2024, 13, 206. [Google Scholar] [CrossRef]
- Agam, N.; Berliner, P.R. Dew formation and water vapor adsorption in semi-arid environments—A review. J. Arid Environ. 2006, 65, 572–590. [Google Scholar] [CrossRef]
- Bruggink, T.; van der Toorn, P. Induction of desiccation tolerance in germinated seeds. Seed Sci. Res. 1995, 5, 1–4. [Google Scholar] [CrossRef]
- Buitink, J.; Ly Vu, B.; Satour, P.; Leprince, O. The re-establishment of desiccation tolerance in germinated radicles of Medicago truncatula Gaertn. seeds. Seed Sci. Res. 2003, 13, 273–286. [Google Scholar] [CrossRef]
- Sahu, B.; Naithani, S.C. Role of reactive oxygen species and antioxidative enzymes in the loss and re-establishment of desiccation tolerance in germinated pea seeds. South African J. Bot. 2023, 163, 75–86. [Google Scholar] [CrossRef]
- Sano, N.; Verdier, J. The re-establishment of desiccation tolerance in germinated tomato (Solanum lycopersicum) seeds. Seed Sci. Res. 2024, 1–9. [Google Scholar] [CrossRef]
- da Silva, A.C.; Davide, L.C.; Braz, G.T.; Maia, J.; de Castro, E.M.; da Silva, E.A.A. Re-induction of desiccation tolerance in germinated cowpea seeds. South African J. Bot. 2017, 113, 34–39. [Google Scholar] [CrossRef]
- Matthews, S.; Powell, A. Electrical conductivity vigour test: Physiological basis and use. Seed Test. Int. 2006, 32–35. [Google Scholar]
- Demidchik, V.; Straltsova, D.; Medvedev, S.S.; Pozhvanov, G. a; Sokolik, A.; Yurin, V. Stress-induced electrolyte leakage: the role of K+-permeable channels and involvement in programmed cell death and metabolic adjustment. J. Exp. Bot. 2014, 65, 1259–1270. [Google Scholar] [CrossRef]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. agriGO v2.0: a GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef]
- Zinsmeister, J.; Lalanne, D.; Terrasson, E.; Chatelain, E.; Vandecasteele, C.; Vu, B.L.; Dubois-Laurent, C.; Geoffriau, E.; Signor, C. Le; Dalmais, M.; et al. ABI5 is a regulator of seed maturation and longevity in legumes. Plant Cell 2016, 28, 2735–2754. [Google Scholar] [CrossRef]
- Zinsmeister, J.; Lalanne, D.; Ly Vu, B.; Schoefs, B.; Marchand, J.; Dang, T.T.; Buitink, J.; Leprince, O. ABSCISIC ACID INSENSITIVE 4 coordinates eoplast formation to ensure acquisition of seed longevity during maturation in Medicago truncatula. Plant J. 2023, 113, 934–953. [Google Scholar] [CrossRef]
- Gazzarrini, S.; Song, L. LAFL factors in seed development and phase transitions. Annu. Rev. Plant Biol. 2024, 75, 459–488. [Google Scholar] [CrossRef]
- Smolikova, G.; Medvedev, S. Seed-to-seedling transition : Novel aspects. Plants 2022, 11, 1988. [Google Scholar] [CrossRef]
- Maia, J.; Dekkers, B.J.W.; Provart, N.J.; Ligterink, W.; Hilhorst, H.W.M. The re-establishment of desiccation tolerance in germinated Arabidopsis thaliana seeds and its associated transcriptome. PLoS One 2011, 6, e29123. [Google Scholar] [CrossRef] [PubMed]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: physiology of development, germination and dormancy; 3rd Ed.; Springer New York: New York, NY, 2013; ISBN 978-1-4614-4692-7. [Google Scholar]
- Gonçalves, J. de P.; Gasparini, K.; Picoli, E.A. de T.; Costa, M.D.-B.L.; Araujo, W.L.; Zsögön, A.; Ribeiro, D.M. Metabolic control of seed germination in legumes. J. Plant Physiol. 2024, 295, 154206. [Google Scholar] [CrossRef]
- Diaz-Mendoza, M.; Diaz, I.; Martinez, M. Insights on the proteases involved in barley and wheat grain germination. Int. J. Mol. Sci. 2019, 20, 2087. [Google Scholar] [CrossRef] [PubMed]
- Graham, I.A. Seed storage oil mobilization. Annu. Rev. Plant Biol. 2008, 59, 115–142. [Google Scholar] [CrossRef] [PubMed]
- Bradford, K.J. A water relations analysis of seed germination rates. Plant Physiol. 1990, 94, 840–849. [Google Scholar] [CrossRef]
- Valgimigli, L. Lipid peroxidation and antioxidant protection. Biomolecules 2023, 13, 1291. [Google Scholar] [CrossRef] [PubMed]
- Jové, M.; Mota-Martorell, N.; Pradas, I.; Martín-Gari, M.; Ayala, V.; Pamplona, R. The Advanced lipoxidation end-product malondialdehyde-lysine in aging and longevity. Antioxidants 2020, 9, 1132. [Google Scholar] [CrossRef] [PubMed]
- Sharova, E.; Romanova, A. Ascorbate in the apoplast of elongating plant cells. Biol. Commun. 2018, 63, 77–86. [Google Scholar] [CrossRef]
- Smirnoff, N.; Wheeler, G.L. The ascorbate biosynthesis pathway in plants is known, but there is a way to go with understanding control and functions. J. Exp. Bot. 2024, 75, 2604–2630. [Google Scholar] [CrossRef]
- Makavitskaya, M.; Svistunenko, D.; Navaselsky, I.; Hryvusevich, P.; Mackievic, V.; Rabadanova, C.; Tyutereva, E.; Samokhina, V.; Straltsova, D.; Sokolik, A.; et al. Novel roles of ascorbate in plants: induction of cytosolic Ca2+ signals and efflux from cells via anion channels. J. Exp. Bot. 2018, 69, 3477–3489. [Google Scholar] [CrossRef]
- Tuan, P.A.; Kumar, R.; Rehal, P.K.; Toora, P.K.; Ayele, B.T. Molecular mechanisms underlying abscisic acid/gibberellin balance in the control of seed dormancy and germination in cereals. Front. Plant Sci. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Shu, K.; Zhou, W.; Chen, F.; Luo, X.; Yang, W. Abscisic acid and gibberellins antagonistically mediate plant development and abiotic stress responses. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhang, Y.; Zheng, Y. Integration of ABA, GA, and light signaling in seed germination through the regulation of ABI5. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Zheng, X.; Mo, W.; Zuo, Z.; Shi, Q.; Chen, X.; Zhao, X.; Han, J. From regulation to application: the role of abscisic acid in seed and fruit development and agronomic production strategies. Int. J. Mol. Sci. 2024, 25. [Google Scholar] [CrossRef]
- Mo, W.; Zheng, X.; Shi, Q.; Zhao, X.; Chen, X.; Yang, Z.; Zuo, Z. Unveiling the crucial roles of abscisic acid in plant physiology: implications for enhancing stress tolerance and productivity. Front. Plant Sci. 2024, 15, 1–23. [Google Scholar] [CrossRef]
- Zhao, H.; Nie, K.; Zhou, H.; Yan, X.; Zhan, Q.; Zheng, Y.; Song, C. ABI5 modulates seed germination via feedback regulation of the expression of the PYR/PYL/RCAR ABA receptor genes. New Phytol. 2020, 228, 596–608. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Han, X.; Yang, M.; Zhang, M.; Pan, J.; Yu, D. The transcription factor INDUCER OF CBF EXPRESSION1 interacts with ABSCISIC ACID INSENSITIVE5 and DELLA proteins to fine-tune abscisic acid signaling during seed germination in Arabidopsis. Plant Cell 2019, 31, 1520–1538. [Google Scholar] [CrossRef]
- Wei, J.; Li, X.; Song, P.; Wang, Y.; Ma, J. Studies on the interactions of AFPs and bZIP transcription factor ABI5. Biochem. Biophys. Res. Commun. 2022, 590, 75–81. [Google Scholar] [CrossRef]
- Skubacz, A.; Daszkowska-Golec, A.; Szarejko, I. The role and regulation of ABI5 (ABA-insensitive 5) in plant development, abiotic stress responses and phytohormone crosstalk. Front. Plant Sci. 2016, 7, 1–17. [Google Scholar] [CrossRef]
- Collin, A.; Daszkowska-Golec, A.; Szarejko, I. Updates on the role of ABSCISIC ACID INSENSITIVE 5 (ABI5) and ABSCISIC ACID-RESPONSIVE ELEMENT BINDING FACTORs (ABFs) in ABA signaling in different developmental stages in plants. Cells 2021, 10, 1996. [Google Scholar] [CrossRef]
- Lopez-Molina, L.; Mongrand, S.; Chua, N.-H. A postgermination developmental arrest checkpoint is mediated by abscisic acid and requires the ABI5 transcription factor in Arabidopsis. Proc. Natl. Acad. Sci. 2001, 98, 4782–4787. [Google Scholar] [CrossRef]
- Shen, Q.; Uknes, S.J.; Ho, T.H. Hormone response complex in a novel abscisic acid and cycloheximide-inducible barley gene. J. Biol. Chem. 1993, 268, 23652–23660. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Chen, C.N.; Brands, A.; Pan, S.M.; David Ho, T.H. The stress- and abscisic acid-induced barley gene HVA22: Developmental regulation and homologues in diverse organisms. Plant Mol. Biol. 2001, 45, 327–340. [Google Scholar] [CrossRef]
- Zhang, H.; Yuan, Y.; Xing, H.; Xin, M.; Saeed, M.; Wu, Q.; Wu, J.; Zhuang, T.; Zhang, X.; Mao, L.; et al. Genome-wide identification and expression analysis of the HVA22 gene family in cotton and functional analysis of GhHVA22E1D in drought and salt tolerance. Front. Plant Sci. 2023, 14. [Google Scholar] [CrossRef]
- Brands, A.; Ho, T.D. Function of a plant stress-induced gene, HVA22. Synthetic enhancement screen with its yeast homolog reveals its role in vesicular traffic. Plant Physiol. 2002, 130, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, L. Genome-wide identification, evolution, and expression analysis of HVA22 gene family in Brassica napus L. Genet. Resour. Crop Evol. 2025, 72, 7495–7515. [Google Scholar] [CrossRef]
- Chen, Q.; Huang, L.; Li, X.; Ma, Y.; Wang, Z.; Zhang, C.; Lin, F.; Liu, C. Comprehensive genome-wide analysis and expression profiling of the HVA22 gene family unveils their potential roles in soybean responses to abiotic stresses. J. Plant Growth Regul. 2025, 44, 2122–2138. [Google Scholar] [CrossRef]
- Sharon, K. Cloning of HVA22 homolog from Aloe vera and preliminary study of transgenic plant development. Int. J. Pure Appl. Biosci. 2017, 5, 1113–1121. [Google Scholar] [CrossRef]
- Guo, W.-J.J.; David Ho, T.-H.; Ho, T.H.D. An abscisic acid-induced protein, HVA22, inhibits gibberellin-mediated programmed cell death in cereal aleurone cells. Plant Physiol. 2008, 147, 1710–1722. [Google Scholar] [CrossRef]
- Chen, C.N.; Chu, C.C.; Zentella, R.; Pan, S.M.; Ho, T.H.D. AtHVA22 gene family in Arabidopsis: Phylogenetic relationship, ABA and stress regulation, and tissue-specific expression. Plant Mol. Biol. 2002, 49, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Dietz, K.-J. Plant Peroxiredoxins. Annu. Rev. Plant Biol. 2003, 54, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chu, P.; Zhou, Y.; Ding, Y.; Li, Y.; Liu, J.; Jiang, L.; Huang, S. Ectopic expression of NnPER1, a Nelumbo nucifera 1-cysteine peroxiredoxin antioxidant, enhances seed longevity and stress tolerance in Arabidopsis. Plant J. 2016, 88, 608–619. [Google Scholar] [CrossRef]
- Haslekås, C.; Viken, M.K.; Grini, P.E.; Nygaard, V.; Nordgard, S.H.; Meza, T.J.; Aalen, R.B. Seed 1-cysteine peroxiredoxin antioxidants are not involved in dormancy, but contribute to inhibition of germination during stress. Plant Physiol. 2003, 133, 1148–1157. [Google Scholar] [CrossRef]
- Chen, H.; Ruan, J.; Chu, P.; Fu, W.; Liang, Z.; Li, Y.; Tong, J.; Xiao, L.; Liu, J.; Li, C.; et al. AtPER1 enhances primary seed dormancy and reduces seed germination by suppressing the ABA catabolism and GA biosynthesis in Arabidopsis seeds. Plant J. 2020, 101, 310–323. [Google Scholar] [CrossRef]
- Moore, R.P. Handbook on tetrazolium testing; 2nd ed.; International Seed Testing Association: Zurich, Switzerland, 1993; ISBN 3906549259.
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Huang, C.; He, W.; Guo, J.; Chang, X.; Su, P.; Zhang, L. Increased sensitivity to salt stress in an ascorbate-deficient Arabidopsis mutant. J. Exp. Bot. 2005, 56, 3041–3049. [Google Scholar] [CrossRef] [PubMed]
- Frolov, A.; Bilova, T.; Paudel, G.; Berger, R.; Balcke, G.U.G.U.; Birkemeyer, C.; Wessjohann, L.A.L.A. Early responses of mature Arabidopsis thaliana plants to reduced water potential in the agar-based polyethylene glycol infusion drought model. J. Plant Physiol. 2017, 208, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Knopkiewicz, M.; Wojtaszek, P. Validation of reference genes for gene expression analysis using quantitative polymerase chain reaction in pea lines (Pisum sativum) with different lodging susceptibility. Ann. Appl. Biol. 2019, 174. [Google Scholar] [CrossRef]
Figure 1.
Typical effects of ‘drying and rehydration’ treatment on the P. sativum seedlings. (a) 3-day-old intact seedling. (b) 3-day-old seedling with excised cotyledons. (c) 3-day-old intact seedling after ‘drying and rehydration’ treatment. (d) 6-day-old intact seedling after ‘drying and rehydration’ treatment. H – hypocotyl. R – primary root. Black arrows show the formation of adventitious roots.
Figure 1.
Typical effects of ‘drying and rehydration’ treatment on the P. sativum seedlings. (a) 3-day-old intact seedling. (b) 3-day-old seedling with excised cotyledons. (c) 3-day-old intact seedling after ‘drying and rehydration’ treatment. (d) 6-day-old intact seedling after ‘drying and rehydration’ treatment. H – hypocotyl. R – primary root. Black arrows show the formation of adventitious roots.
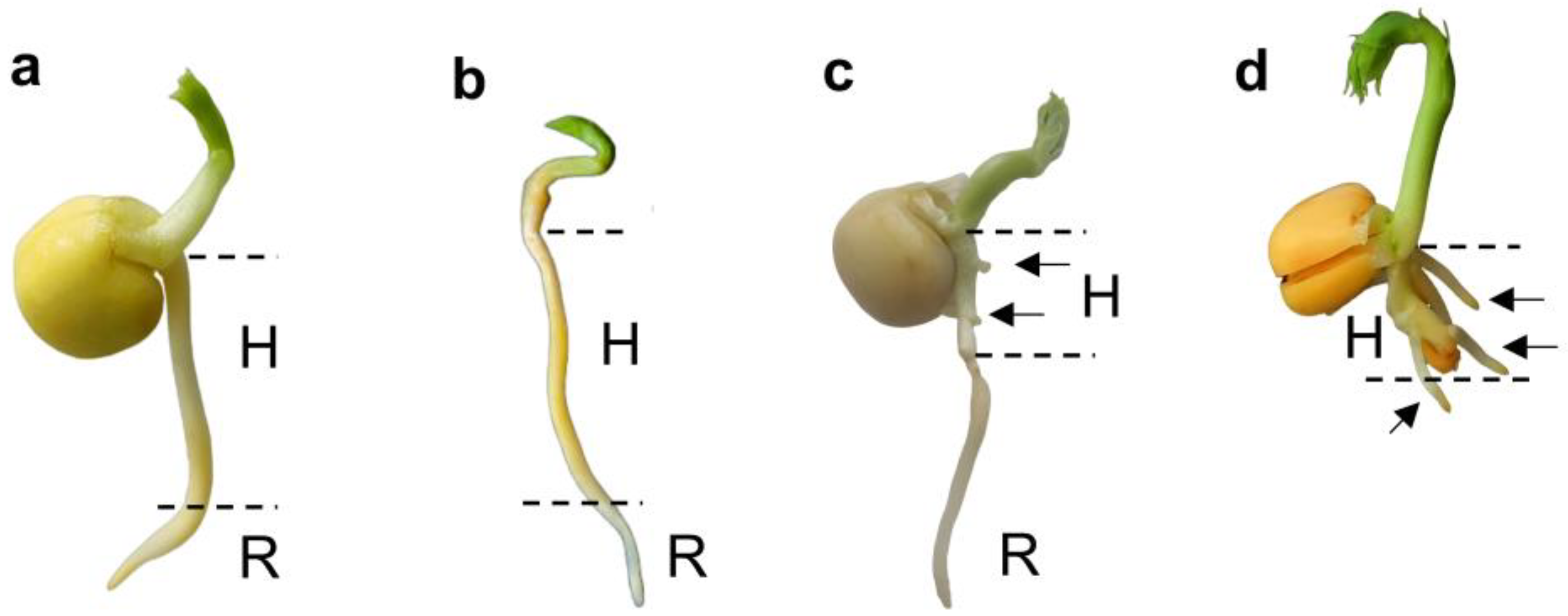
Figure 2.
Viability test with tetrazolium chloride (1%) of P. sativum embryonic axes isolated from 3-day-old intact seedling (a), 3-day-old seedlings with excised cotyledons (b), and 3-day-old intact seedlings after ‘drying and rehydration’ treatment (c). H – hypocotyl. R – primary root. Black arrows show the formation of adventitious roots. The red coloration (reduction of colorless tetrazolium to formazan) indicates dehydrogenase activity, demonstrating active metabolic reactions and cell viability.
Figure 2.
Viability test with tetrazolium chloride (1%) of P. sativum embryonic axes isolated from 3-day-old intact seedling (a), 3-day-old seedlings with excised cotyledons (b), and 3-day-old intact seedlings after ‘drying and rehydration’ treatment (c). H – hypocotyl. R – primary root. Black arrows show the formation of adventitious roots. The red coloration (reduction of colorless tetrazolium to formazan) indicates dehydrogenase activity, demonstrating active metabolic reactions and cell viability.

Figure 3.
Changes in water potential (a) and electrolyte leakage (b) of hypocotyls (H) and roots (R) from 3-day-old P. sativum seedlings in control (1), ‘drying and rehydration’ with cotyledons (2), ‘drying and rehydration’ without cotyledons (3). Box plots show high, low, and median values (n = 5). Outliers are represented as white dots. Asterisks over the box plots denote significant differences between treatments and control (one-way ANOVA test, p ≤ 0.05 [*], p ≤ 0.01 [**] or p ≤ 0.001 [***]).
Figure 3.
Changes in water potential (a) and electrolyte leakage (b) of hypocotyls (H) and roots (R) from 3-day-old P. sativum seedlings in control (1), ‘drying and rehydration’ with cotyledons (2), ‘drying and rehydration’ without cotyledons (3). Box plots show high, low, and median values (n = 5). Outliers are represented as white dots. Asterisks over the box plots denote significant differences between treatments and control (one-way ANOVA test, p ≤ 0.05 [*], p ≤ 0.01 [**] or p ≤ 0.001 [***]).
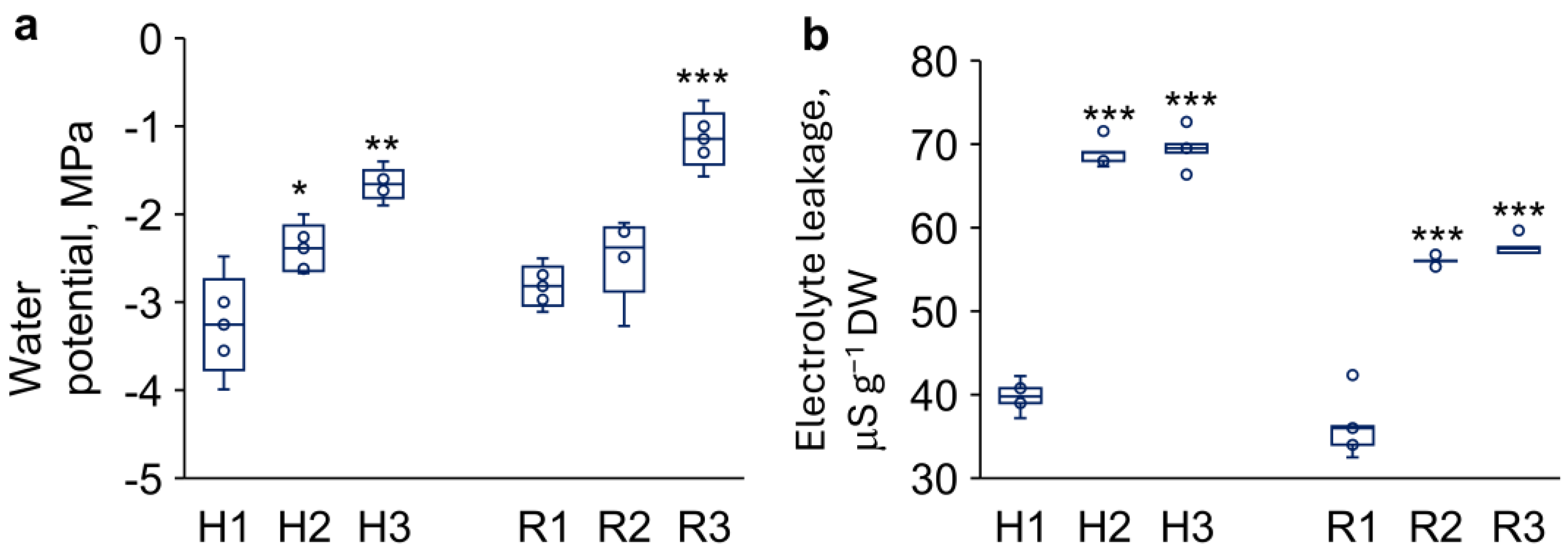
Figure 4.
Changes in content of malondialdehyde (MDA) and other thiobarbituric acid-reactive lipid peroxidation products (a) and L-ascorbic acid (b) in hypocotyls (H) and roots (R) from 3-day-old P. sativum seedlings in response to ‘drying and rehydration’ treatment. H1–3 and R1–3 denote experimental variants as follows: H1 and R1 – control, H2 and R2 – ‘drying and rehydration’ treatment with cotyledons, H3 and R3 – ‘drying and rehydration’ treatment without cotyledons. Box plots show high, low, and median values (n = 5). Outliers are represented as dots beyond the bounds of box plot whiskers. Asterisks over the box plots denote significant differences between treatments and control (one-way ANOVA test, p ≤ 0.05 [*], p ≤ 0.01 [**], and p ≤ 0.001 [***]).
Figure 4.
Changes in content of malondialdehyde (MDA) and other thiobarbituric acid-reactive lipid peroxidation products (a) and L-ascorbic acid (b) in hypocotyls (H) and roots (R) from 3-day-old P. sativum seedlings in response to ‘drying and rehydration’ treatment. H1–3 and R1–3 denote experimental variants as follows: H1 and R1 – control, H2 and R2 – ‘drying and rehydration’ treatment with cotyledons, H3 and R3 – ‘drying and rehydration’ treatment without cotyledons. Box plots show high, low, and median values (n = 5). Outliers are represented as dots beyond the bounds of box plot whiskers. Asterisks over the box plots denote significant differences between treatments and control (one-way ANOVA test, p ≤ 0.05 [*], p ≤ 0.01 [**], and p ≤ 0.001 [***]).
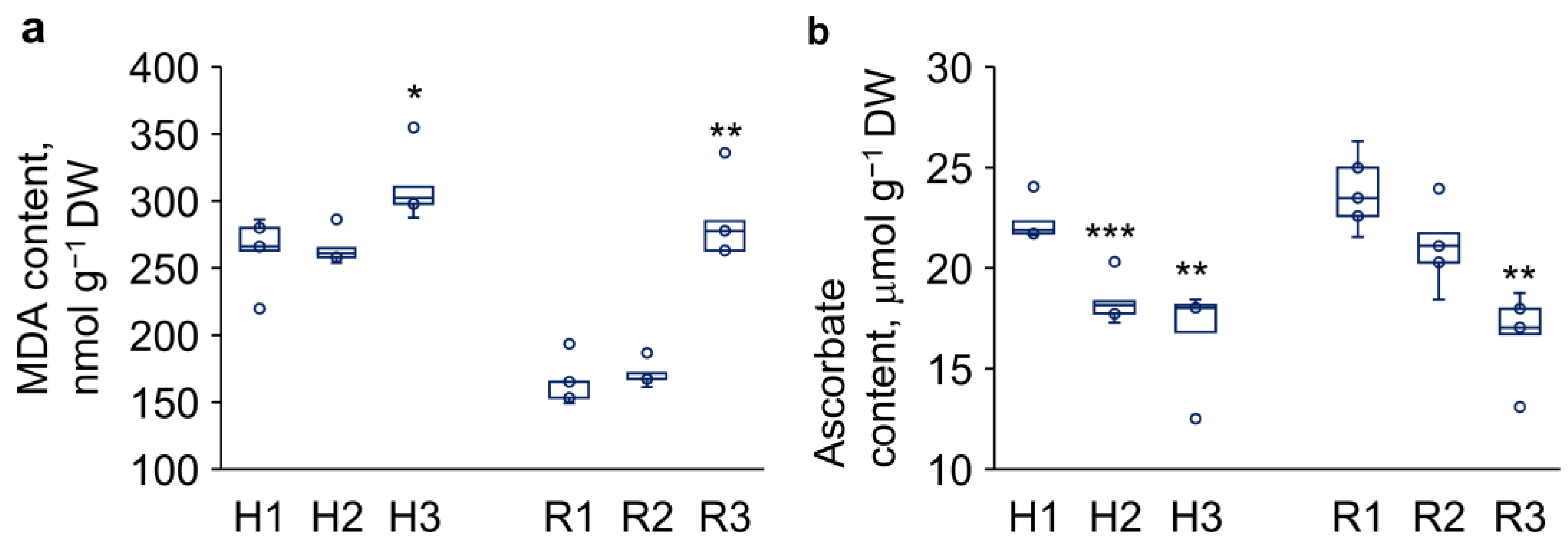
Figure 5.
Relative expression levels of the ABA-dependent genes encoded the transcription factors ABSCISIC ACID INSENSITIVE3 (ABI3), ABSCISIC ACID INSENSITIVE4 (ABI4), and ABSCISIC ACID INSENSITIVE5 (ABI5) in hypocotyls (H) and roots (R) from 3-day-old P. sativum seedlings. H1–3 and R1–3 denote experimental variants as follows: H1 and R1 – control, H2 and R2 – ‘drying and rehydration’ treatment with cotyledons, H3 and R3 – ‘drying and rehydration’ treatment without cotyledons. Box plots show high, low, and median values (n = 3). Asterisks over the box plots denote significant differences between treatments and control (one-way ANOVA test, p ≤ 0.05 [*], p ≤ 0.01 [**]).
Figure 5.
Relative expression levels of the ABA-dependent genes encoded the transcription factors ABSCISIC ACID INSENSITIVE3 (ABI3), ABSCISIC ACID INSENSITIVE4 (ABI4), and ABSCISIC ACID INSENSITIVE5 (ABI5) in hypocotyls (H) and roots (R) from 3-day-old P. sativum seedlings. H1–3 and R1–3 denote experimental variants as follows: H1 and R1 – control, H2 and R2 – ‘drying and rehydration’ treatment with cotyledons, H3 and R3 – ‘drying and rehydration’ treatment without cotyledons. Box plots show high, low, and median values (n = 3). Asterisks over the box plots denote significant differences between treatments and control (one-way ANOVA test, p ≤ 0.05 [*], p ≤ 0.01 [**]).
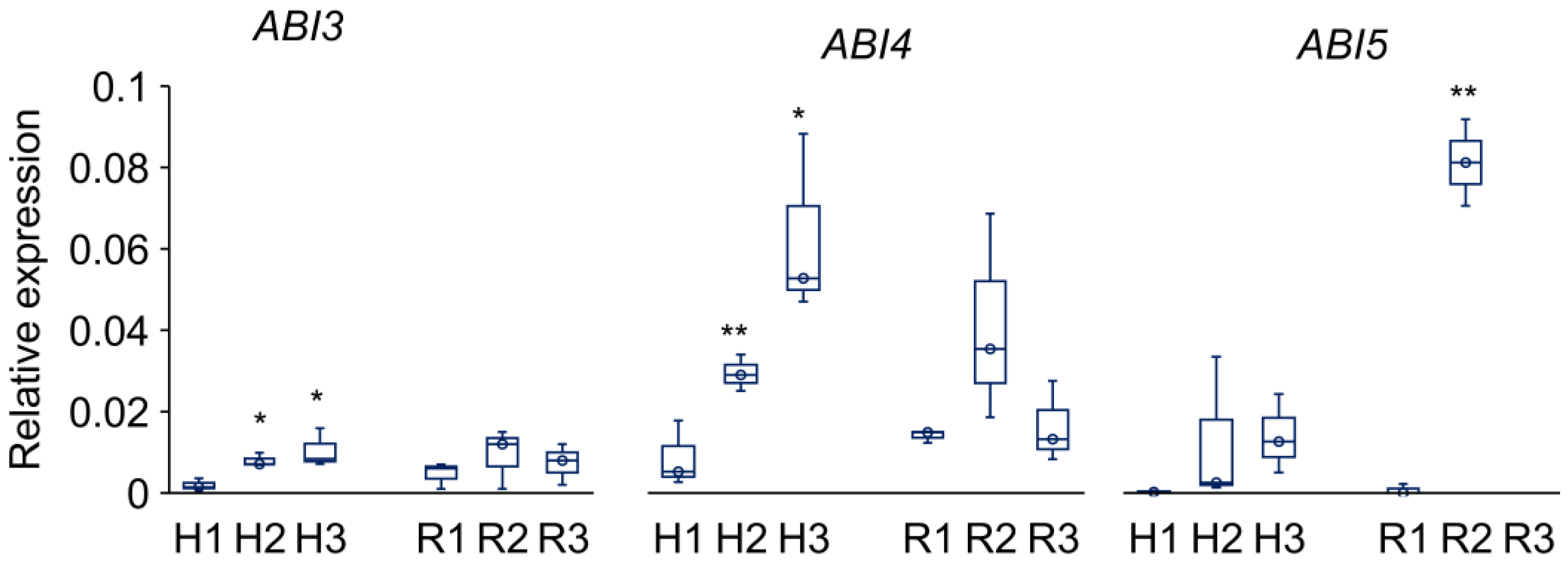
Figure 6.
Relative expression levels (Y-axes) of the ABA-dependent genes (selected from the ‘response to water deprivation’ group in the Gene Ontology database) in hypocotyls (H) and roots (R) from 3-day-old P. sativum seedlings. (a) HORDEUM VULGARE ABUNDANT 22E (HVA22), (b) 1-CYSTEINE PEROXIREDOXIN (PER1), (c) LOW-TEMPERATURE INDUCED 65 (LTI65), (d) LIPID TRANSFER PROTEIN 4 (LTP4), (e-g) RESPONSIVE TO DEHYDRATION22-1/2/3 (RD22-1/2/3), (h) LATE EMBRYOGENESIS ABUNDANT 14 (LEA14). H1–3 and R1–3 denote experimental variants as follows: H1 and R1 – control, H2 and R2 – ‘drying and rehydration’ treatment with cotyledons, H3 and R3 – ‘drying and rehydration’ treatment without cotyledons. Box plots show high, low, and median values (n = 3). Asterisks over the box plots denote significant differences between treatments and control (one-way ANOVA test, p ≤ 0.05 [*], p ≤ 0.01 [**]).
Figure 6.
Relative expression levels (Y-axes) of the ABA-dependent genes (selected from the ‘response to water deprivation’ group in the Gene Ontology database) in hypocotyls (H) and roots (R) from 3-day-old P. sativum seedlings. (a) HORDEUM VULGARE ABUNDANT 22E (HVA22), (b) 1-CYSTEINE PEROXIREDOXIN (PER1), (c) LOW-TEMPERATURE INDUCED 65 (LTI65), (d) LIPID TRANSFER PROTEIN 4 (LTP4), (e-g) RESPONSIVE TO DEHYDRATION22-1/2/3 (RD22-1/2/3), (h) LATE EMBRYOGENESIS ABUNDANT 14 (LEA14). H1–3 and R1–3 denote experimental variants as follows: H1 and R1 – control, H2 and R2 – ‘drying and rehydration’ treatment with cotyledons, H3 and R3 – ‘drying and rehydration’ treatment without cotyledons. Box plots show high, low, and median values (n = 3). Asterisks over the box plots denote significant differences between treatments and control (one-way ANOVA test, p ≤ 0.05 [*], p ≤ 0.01 [**]).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



