
Submitted:
17 November 2025
Posted:
18 November 2025
You are already at the latest version
Abstract
Climate changes caused by anthropogenic actions can directly and indirectly affect living beings, including parasites and their hosts. Several factors are involved in climatic changes. Some of them are water temperature, salinity, pH, distribution of nutrients in the aquatic environment and those can interfere with the fish community, whether in open water or in production systems. In this review, we will show how climate change factors can affect the hosts and parasites in the aquatic environment.
Keywords:
climate change
; host-parasite relationships
1. Introduction
The threats of climate change to human society and natural ecosystems were highlighted in the fourth Assessment Report of the Intergovernmental Panel on Climate Change (IPCC) in 2007. A scientific consensus has been reached, among the scientific community, that finally admit that human activity causes the accumulation of greenhouse gases in the atmosphere, leading to increasing temperatures and causing environmental changes[1,2].
The theme on Climate Change has recently been addressed more frequently, due to the direct and indirect environmental impacts that affect the planet and causes several ecological changes, which affect all life forms[3,4,5]. Among the actions that contribute to climatic changes we can mention: the increase in the use of fossil fuels, deforestation, industrial activity and biomass burning, which results in an increase in the concentration of CO2 and volatile organic compounds such as butane and propane among others and will destroy the ozone layer that protect the planet[5,6].Activities such as livestock, agriculture, hydroelectric construction and highways are directly related to the occurrence of climate change, due to their environmental impacts [3,6,7,8,9]. The main results of climatic change include temperature rise, and variations in other climate factors such as precipitation and humidity[6,10,11].
The effects mentioned above affect all groups of living beings, their ecological relationships with the environment and also the relationships between trophic levels, leading to biodiversity loss in many taxonomic groups[3]. The resulting environmental changes, caused by both natural phenomena and human intervention, have a marked influence on the emergence and proliferation of zoonotic parasitic diseases[6,12]. Many infections, vector organisms, non-human reservoir species, and rate of pathogen replication are sensitive to climatic conditions[13,14]. For example, the blue crab (Collinectes sapidus) and its parasite, the dinoflagellate Hematodinium perezi, the later has its optimal growth temperature between 23 and 31 ⁰C, while this temperature range causes a decline in the host's immune response. This effect is also known among the host Chionoecetes opilio which decreases its immune response around 4 ⁰C and the parasite Hematodinium sp. has the reproduction increased around of 3 ⁰C, being stable until 10 ⁰C[15] .
The relationship between climate change and the incidence/prevalence of infectious/parasitic diseases has been addressed by different authors, considering certain regions of the planet[7,8,16,17,18,19,20,21] or using more general approaches, without considering the spatial distribution or any specific geographic region[9,10,11,12,22].
Parasitic species are associated with their hosts, and these with their ecological niche, this ecological niche suffers from climate change, directly affecting the environmental behavior of host species, and consequently the adaptations of parasitic species associated with their hosts. It is well known that seasonal changes in the environment can cause modifications in the parasite-host relationship. The presence or abundance of parasites is directly influenced by both the environment within the host, their micro-environment and/or the condition of the ecosystem, their macro-environment[23].
The repercussions of climate change are not limited to temperature effects on hosts and their parasites, but also include: alterations in water levels and flow regimes, eutrophication, stratification, changes in acidification and salinity, reduced ice cover, changes in ocean currents, increased ultra-violet (UV) light penetration, runoff and weather extremes[24].
One of the factors pointed out as temperature, directly influenced by species distribution patterns, favors the growth of host rates reservoirs of parasites and vectors, associated with the prevalence of parasites[17] besides affecting the life cycles of parasites[11] and consequently, something that can be linked to the emergence or reemergence of some parasitic diseases[21].
The interaction between climate change and the occurrence of diseases raises questions about the difficulties involved especially those that, highlights the fact of the cause-effect relationship between climate change and the incidence/prevalence of diseases, caused by parasites, where biotic or abiotic factors can interfere in host/parasite interaction[25]. Host-parasite systems with community-level interactions make them complex systems; in this complexity, it is difficult to assume climatic factors as the only cause, without taking into account the other related factors [25]. It is important to contextualize the difficulties raised[25], in which climate change attributed to global warming, can interfere in the dynamics of disease transmission, including parasite and host populations[11,21,22]. The aim this revision was understand as climate changes can interfere with parasite-hosts interactions.
2. Materials and Methods
This article is a bibliographic review. A survey was made on the “Web of Science” database a platform available for researching scientific references on the proposed topic. A survey of articles using the key words: climate change, parasites and fishes. Accessed at day June 2025.
More than 629,403 papers with the main topic "Climate change" have been found on the search platform for scientific articles "Web of Science" for the most varied areas of Science (Figure 1).
However, only 3,064 articles were found relating climate change and parasites on the same platform. The first publication date back to the 1920s, with one article published in 1926, intituled “Effect of temperature and moisture on nematode root knot “, where the author, Godfrey, observes different levels of humidity and temperature and the development of the nematoda Heterodera radicicola plant parasite and the root development of plants. The author concludes that climatic variations are determinant for the development of the parasite, which in turn interferes with the root growth of plants[26]. Only, from 1999 onwards there was an annual continuity of publications involving climate change and parasites. In 2024, there were 275 articles published and 12,573 citations made.
Many articles have been published annually since the 2000s. However, most of the published articles were literature reviews and/or indirectly had climatic factors as a reference in the parasite-host interaction. And even less, 446 articles have related information between “climate change”, “parasite” and “fish” in freshwater or marine environment, the majority of the works were revisions on the subject proposed. The first paper was published in 2001, intitled “The status of the grey seal, Halicopterus grypus, in the northwest Atlantic”, that mention fishes in indirect way[27]. There were 45 articles published in 2024 and 2,086 citations (Figure 2).
In fact, only in 18 papers the authors are discussing the climate changes as the main causes of interference in the interaction fish/parasite relationship, in our survey (Table 1).
This table presents some studies that conclude that climatic factors directly interfered in the parasite-host relationship, with the factors being water temperature, air temperature, rainfall, salinity, dissolved oxygen, CO2 concentration. It was noted that in the central part of the European continent, the increase in river water temperature is related to the higher incidence of Proliferative Kidney Disease in salmonid species[29,30,31,32]. Showing that the high temperature in the waters of the rivers favors the proliferation of Myxozoa and increases the susceptibility of the host species to disease in lentic environment. The increase in temperature favored the proliferation of the myoliquefaction-causing Myxozoa in swordfish off the coast of Australia in a marine environment[33]. Another important factor is related to the biological cycle of parasites. Monoxene parasites tend to increase adaptability to climate change in the aquatic environment, reflecting in greater parasite intensity in their hosts[34,35]. For heteroxene parasites, they are subject to the number of intermediate hosts to close the biological cycle. Species that require more than one invertebrate intermediate host tend to decrease the population in the environment, caused by the dispersion, decrease or disappearance of the first or second intermediate invertebrate hosts[36]. The temperature of water, air and rainfall, when favorable to the first intermediate invertebrate hosts, favor the population of trematode parasites, reflecting in the greater intensity and prevalence in the environment. However, this condition is not a rule, there is also the decrease or disappearance of parasitic species, because of climate change, reflected in the increase in water temperature, air and rainfall, with the decrease or loss of the ecological niche associated with parasitic species.
3. Effect of Climate Change on Host Fish
In ectothermic animals, such as fish, the impact of climate change, is more severe, their body temperature and metabolism are closely regulated by their environmental water that make them especially vulnerable to temperature fluctuations inducing organic responses at all levels of their biology[46,47].
Aquatic organisms, and fish in particular, are affected by a variety of stressors caused by anthropogenic influences that lead to changes in environmental parameters. These in turn elicit stress responses of the organism in the sense that the affected organisms show reactions outside their normal range[48]. The impacts of multiple stressors in freshwater biota at spatial scales and ecosystems, point out that anthropic actions can interfere in water quality dynamics, these include chemical pollutants, nutrients, flow velocity, pH, dissolved oxygen, disturbances in light and temperature regimes, and many other physical-chemical variables that can be significantly altered[49].
Metabolic disorders and oxidative stress can be caused by increasing temperature, affecting the synthesis of release and actions of stress hormones[50]. Acute or prolonged exposure to high temperatures affects the functions of stress axes and responses to other stressors, compromising the long-term coping capacity of each organism[51].
The impacts of climate change on the functional diversity of freshwater fish, influenced the change in the distribution of fish species, with some species being able to lose up to one hundred percent of the areas to which they are climatically adapted in the Paraná-Paraguay River basin[31]. To this end, these impacts make it difficult for the hosts to interact with their respective ecological niches, reducing the community, either by death or abandonment of the place[51].
The declining biodiversity could also bring food problems. Climate change could lead to disruptions in ocean currents, which will stop bringing nutrients to Antarctica's shores, hampering the growth of krill, which is the base of all the oceans' food chain. This could lead to a decrease in marine biota and a low supply of food to communities traditionally consuming fish and/or seafood[52].
A study about current knowledge in climate change-related eco-immunology in teleosts fishes, focusing on temperature, hypoxia, salinity, and acidification showed that acute and chronic changes in temperature and dissolved oxygen compromise the immunity of fish, increasing susceptibility to disease[53]. The review highlights the need for further studies on acidification and multiple stressor interactions. In addition, it explores the acclimatization potential of the fish's immune response, although comprehensive conclusions cannot yet be drawn due to the limited number of epigenetic studies.
The formation of reactive oxygen species (ROS) responsible for oxidative stress is a common aspect generated by stress. The term oxidative stress is used for the state of imbalance between the generation of oxidants (free radicals and ROS) and the availability of endogenous antioxidants to eliminate ROS[54]. Many studies show that ROS play a central role in the pathogenesis and progression of many infectious diseases and inflammatory disorders[55,56].
Inflammation is a host defense mechanism, generating immune response to physical stimuli, external chemicals or pathogen presence that plays a crucial role in the removal of the pathogens in the hosts, involving an increased generation of ROS by target cells and the immune system[57]. As part of the inflammatory response, ROS are produced that facilitate the elimination of invasive tissue pathogens, however when produced for prolonged periods promote oxidative stress and problems related to chronic inflammation[56]. Immune system cells such as mast cells and leukocytes are recruited during inflammation, they migrate to the site of infection, leading to a “respiratory burst” due to increased oxygen uptake and therefore greater release and accumulation of ROS at the site of infection that stimulate pathways that lead to inflammation activation[55].
The immune system of vertebrates comprises cells, tissues and organs that serve to protect a host organism from infections by bacteria, viruses or parasites. Innate immunity is the first line of defense against invading pathogens. This type of immunity involves physical barriers such as skin and mucus, cellular components and soluble components, including cytokines and complement[58,59]. Adaptive immune response is slower but more efficient, and includes humoral components, B lymphocytes, and cell-mediated components such as T lymphocytes. Adaptive immunity is very specific in its targets and has a memory component that allows a faster response to a reinfection[60].
Fish have both innate and adaptive immunity. In the innate immune response, pathogens are detected by recognition receptors, which induce specific responses to various molecular patterns associated with pathogens (PAMPs) in the infected animal[61]. Water temperature can affect the immune system of fish. Acute and chronic changes in temperature also have different impacts on animals, with short-term episodes being offset by processes such as heat-shock protein response (HSPs), while chronic temperature variations are less likely to be resolved by such responses and can still impact the physiology of the organism[62,63].
In a study on the projection of climate change in Colossoma macropomum (Characiforme) in the Amazon, a greater leukocyte immune response was found in the proportions of lymphocytes and granulocytes. The test determined a change in climate for the year 2100, where the increase in temperature closely influenced the immune response of the host in relation to the degree of monogeneans parasites in the gills[35].
There is a limitation between the correlation of evidence that climate change is related to fish diseases[35]. Host-parasite relationships are affected by several factors, especially by environmental changes. For fish, changes in water temperature can lead to an increase in the intensity and virulence of the pathogen, decrease in immune resistance and increased frequency of disease outbreaks in the host. However, little attention has been paid to clarifying how dependence on fish temperature can directly affect parasites[64].
4. Climate Change and Fish Production
Aquaculture is defined as the cultivation of aquatic organisms under controlled conditions, with the objective of producing a marketable product in the most efficient and economical way possible. Aquaculture is among the most important sectors for food production, providing high quality animal protein and income generation and employment[35,64,65]. Aquaculture plays an important role in food production, in addition to a large international presence with an impact on the global economy[66].
Aquaculture can contribute to compensating for the negative impacts that climate change can have on fisheries[67]. However, mobile organisms, such as fish, may be tolerant of some environmental changes or may escape these changes by moving from one place to another, but aquaculture fish raised in cages or ponds in fixed places cannot move to avoid these changes [67,68].
Climate changes can be considered stressful and promote the emergence of diseases in the growing environment of aquaculture[39,62], since temperature is an environmental factor that affects productivity in fish farming, it can affect the growth, reproduction and behavior of fish in nature and aquaculture, changing individual and population dynamics[1]. The increase in infectious diseases in aquaculture is associated with large economic losses. These diseases are controlled by the treatment of fish with chemicals and medications such as antibiotics. This seemingly stable state, in which the effective transmission of a disease is controlled by drugs, is highly delicate and can easily lose balance if environmental conditions change. One of these parameters is water temperature, which promotes the occurrence of diseases in nature and also in fish farms, since they obtain water from natural sources and the water temperature in the facilities accompanies environmental change [31,32,69]. Some studies have addressed the relationship between temperature modulation and fish susceptibility to pathogens[70,71].
Research on the impacts of climate change on aquaculture and fisheries improves understanding of environmental effects and their consequences for aquatic ecosystems. Highlighting the socioeconomic implications, providing essential data for public policies. The promotion of sustainable adaptation strategies and international cooperation to mitigate these impacts. In addition to investment in research and innovation to address climate challenges and ensure global food security[72].
The impact of climate change on aquaculture can vary in relation to both types (e.g., water temperature, saline water intrusion, ocean acidification) and extent of climate change, depending on climate zones (temperate, arid, tropical or Mediterranean), geographical areas (inland, sea or coastal), types of aquaculture production systems and aquatic species that are grown. In the Northern Hemisphere, rising temperatures can result in mild weather and positive effects on animal growth, but it can also introduce new diseases. In the Southern Hemisphere, the increase in temperature can result in heat stress, floods, droughts and extreme weather conditions[73]. In addition, ocean productivity decreases due to climate change and raw materials for fishmeal and fish oil may be affected[73,74]. In this circumstance, climate change indirectly puts more pressure on improving characteristics related to food efficiency, especially in carnivorous species.
The protozoan Ichthyophthirius multifiliis is one of the main parasites that causes damage to fish farms. Due to lesions proven by these agents in intense infections combined with the enormous reproductive capacity of the protozoa, these can cause high mortality rates even in populations of native species[75]. This fact confirms the potential for dissemination of parasitological agents. In addition, these can act as vectors, bringing new pathogens from different regions and further intensifying the parasite load when it comes to introduced in farmed fish. The intensity of infection was higher in polluted and eutrophicated lakes, demonstrating the importance of environmental characteristics in the proliferation of these agents. Thus, the food supply on farmed fish contributes significantly to the eutrophication process of the environment. Associating this issue with rising temperatures, these environments are always more susceptible to new pathogens and will never be completely free of parasites[76].
The interactions between hosts-parasites can be influence in multiples variables in function of the temperature. The host and parasite thermal preferences curves, the prevalence, abundance, intensity and modes of transmission, the distribution and availability of the hosts are factors can be positively or negatively affected by temperature[24,28].
Whenever the increase in temperature does not exceed the tolerance curve of the parasite or the host, it favours the intensity of parasitism and its transmission. In a generalized scenario where the host suffers from oxygen stress, the metabolism of the parasites increases[24].
As an example, fish species with great commercial importance may suffer more from the emergence of diseases and parasitic infections due to changes in water temperature in the marine environment. The myxosporid Kudoa musculoliquefaciens responsible for muscle myoliquefaction in swordfish (Xiphias gladius), increased its infection power with a variable from 2,500 to 290,000 spores/g of muscle directly in the marine environment at warmer temperatures between 26 ⁰C-28 ⁰C[33]. While the ectoparasite Lepeophtheirus salmonis, which can cause a decrease in the production of Atlantic salmon (Salmo salar) by spoliative action on the host, had its potential for proliferation and infection at temperatures of up to 22 ⁰C[40].
Given the requirement for the control of environmental parameters within aquaculture, recirculation systems provide total control of water temperature. Nevertheless, their economic feasibility is still uncertain. Conversely, cage aquaculture is integrating monitoring technologies to gather valuable data regarding the impact of climate change on fish and pathogens. Smart aquaculture, employing sensors, the internet of things, and data analytics, is increasingly becoming a reality for numerous commercial fish farms[77].
5. Climate Change and Parasites
The increase in temperature affects system transmission, with direct increase in the metabolism of the parasite, increasing the feeding or replication of the parasite in the host, increasing the damage and resulting in the production of a greater number of stages of transmission, as well as a faster spread of the disease in a single outbreak[15,69]. The increase in temperature can extend the duration of transmission windows, resulting in a general spread of the disease in the host population. Both mechanisms, alone or in combination, would result in a higher prevalence of the disease. However, some diseases could also show opposite effects with the increase in temperature if their ideal temperature for growth and transmission were lower[69,78].
The fact that climate change mainly affects fish communities, whether in cold or hot waters with the determining factors: ocean acidification, increased temperature, salinity, disposal of pollutants, among others, has shown significant changes in the disposition of host species. These factors are also influencing the reproduction, fertility and feeding of fish species. Parasites can be acquired by their hosts in two ways: phylogenetic and ecological. By the phylogenetic route, the parasites are inherited from species or groups of ancestral species, and by the ecological route, the parasites are acquired from the environment or from other host species[79].
In a study about climatic variations in the Neolithic period in Europe and their impacts on lake communities and their intestinal parasites in humans, 14 taxa were found, of which 4 taxa were attributed to fishes (Diphyllobothrium, Opistorchis, Echinostoma and Dioctophyme). It was found that in the periods where there was greater rainfall influence or melting associated with the rise in the water level of the lakes, mainly favored species of the genus Diphyllobothrium associated with increased fishing activity and the consumption of infected raw fish. While in the drier periods, with the decrease in the water level of the lakes, it favoured species of the genus Trichuris, closely related to agropastoral activities[42]. These changes reflect dietary and livelihood adaptations to climatic conditions.
Parasites with monoxenous development/life cycle, there may be dispersion or disappearance of the definitive host from a certain area where the parasite is present, which may disadvantage the entire parasite community in that region or ecological niche, with an increase in parasitic intensity in non-habitual hosts or definitive disappearance of the parasitic species. Parasites with heteroxenous development/life cycle, climate change could be even more severe. These parasitic species need one or more intermediate hosts to be able to complete their biological cycle. If these hosts are more susceptible to environmental variations, they may decrease or disappear from the environment, reflecting negatively on the parasitic biological cycle. These parasites necessarily need the intermediate host to evolve and be able to parasitize the definitive host, such as digenetic parasites[80,81].
Wood and collaborators analyzed 699 fish species from North America from 1880 to 2020. A total intensity of 17,702 parasites was found and divided into 85 taxa (n= 17,259 parasite specimens). Of all parasite species found, 52% require three or more obligate intermediate hosts, in these species there was a 10.9% decline in parasite abundance for each decade. Also, for these parasitic species, a 38% decline in parasite abundance was detected for every 1⁰C increase in sea temperature. For parasite species requiring one or two obligate intermediate hosts, no decline in parasite abundance was observed. The authors concluded that other factors may also contribute to the decline in the abundance of these parasites, despite the strong effect of temperature[45].
Parasites may respond to increasing temperatures more strongly than their hosts. Trematodes digenetic are among the most common parasites of vertebrates. They require a molluscan intermediate host in which infective cercariae are asexually produced and released into the water in search of the next host in the life cycle. In a review of temperature effects on cercarial shedding, it was shown that temperature differentially affected cercarial production, with increases in cercarial emergence from the first intermediate host, after an increase of few degrees in temperature.[82].
However, the increase in temperature can benefit parasites with an heteroxenous cycle, up to the point of host death, reflecting negatively on the parasite's biological cycle[83]. Fluctuations as low as 0 – 1 ⁰C can reflect on the dynamics of parasite distribution, from exponential growth to parasite extinction[84].
The environmental conditions, as the temperature affect directly Free-living stages of parasites [85]. The effects of temperature on free-living stages have been analysed in different groups of parasites. The best studies were with trematodes [86].
The average increase across trematode species was a 200-fold increase with a 10°C rise in temperature, which is much higher than the two-to-three-fold elevation predicted by standard metabolic principles. Small increases in temperature should promote the proliferation and emergence of cercariae, with little or no reduction in transmission efficiency. Even short-term spikes in cercarial emergence are sufficient to cause mortality in the next host, with consequences that may cascade through the ecosystem [82]. Furthermore, there may be serious consequences for human populations at risk of trematode diseases, such as schistosomiasis, where the cercariae infect the man directly, if temperatures increase[24]. In contrast, Morley and Lewis (2013)[87], in another meta-analysis of low and midlatitude species parasites showed a declining at higher temperatures, while cercarial emergence was not affected between 20 and 25 C.
The impact of climate change on infection by the trematode parasite Clinostomum complanatum in Trichogaster fasciatus forage fish in the Aligarh region, India, over seven years has been observed. The results show that infection depends on the temperature of the air at the surface, highlighting the biological relevance of climate change in the ecology of this parasite. At temperatures above 40 0C, it favoured the stabilization of the infection in the intermediate host. The model also showed that the greater the evaporation of the waters, the smaller the area of the water depth, favouring the success of infection of the first intermediate host, positively interfering in the biological cycle of the parasite. This is the first study to implicate air temperature as an environmental variable that may contribute to parasitism in fish hosts[38].
Carlson et al. (2017), analyzed the variations in parasite habitat loss influenced by differences in general circulation models (GCMs). In the most optimistic scenario of Representative Greenhouse Gas Concentration (RCP 2.6), the average loss of native habitat of parasites was 20.2%, while in the most pessimistic scenario (RCP 8.5), the loss was 37.4%. No parasitic species has completely lost its suitable habitat in all climate scenarios, on average, species have lost 29.0% of their total habitat without dispersal. 86 species have lost more than 50% of their habitat, and 8 species have lost more than 80%. Even accounting for dispersal, 202 parasitic species still lost habitat by 2070, and 32 species lost more than half of their adequate global habitat. Despite these losses, parasitic species gained an average of 16.2% of suitable habitat, with 29 species doubling their area and 7 species tripling it, and none of these 7 species has zoonotic potential or records of human infection (Table 2)[81].
Monogeneans (Platyhelminths) are the most common and most diverse ectoparasites of fish, usually live on external surfaces, such as the skin, fins, head, gills and eyes, oral and branchial cavities[88]. Haptor is the main organ for fixing these parasites to their hosts and is the main morphological characteristic used to distinguish these from cestode and digenea parasites[89]. They have monoxenous life cycles and stages of rapid transmission, feed on mucus and epithelial cells, which results in damage to host tissue, such as deep wounds in the epidermis that can reach the dermis. The presence of these ectoparasites is correlated with increased susceptibility to secondary infections such as bacterial infections and disturbances in the host immune system[90]. They are widespread in aquaculture, including closed, semi-closed and open systems, and are responsible for economic losses. Therefore, the fight against infectious diseases, which is already considered one of the main current challenges of fish farming, can become an important issue in climate change scenarios[69,91].
In a senary of climate change until 2100, with the increase in temperature, a higher intensity of monogeneans Notozothecium janauachense, Anacanthorus spathulatus and Mymarothecium boegeri was observed in the host C. macropomum, with a 200% increase in mean intensity and a prevalence of 100% in the hosts. The authors concluded that the environment favours activity and increase in parasitic metabolism[92].
Several factors influence the responses of species to temperature rises, both intrinsic to each organism and extrinsic linked to the ecological niches in which they are inserted, which are directly subject to the environmental conditions existing in each geographic region and how these conditions are and will be affected or not by climate change[5,22].
Melo et al. (2024) analyzed 1,973 fish of the species Hoplias malabaricus in South America, found 120 parasites, two species of Monogeneans (ectoparasites): 17 Urocleidoides cuiabai and 44 U. eremitus; three species of Digenea (endoparasites): 15 Clinostomatopsis sorbens; 30 C. dimorphum and 14 Pseudosellacotyla lutzi. In a climate change scenario until 2050 using an atmosphere-ocean general circulation model, where the main variables analyzed were: temperature, precipitation, carbon and pH, where SSP (Shared Socioeconomic Path) of 4.5 for an optimistic scenario and SSP 8.5 for a pessimistic scenario was used. In an optimistic scenario, there was a loss of 93% of the niches suitable for all parasites, with a 100% loss for species C. dimorphum. In a pessimistic scenario, there was a loss of 97% of the suitable areas, leaving 3% of the niches for parasitic species. The increase in temperature can mainly influence the biological cycle of endoparasite species, as they require one or more intermediate hosts. While the host H. malabaricus showed adaptability to climate change[41].
Certain parasites may have a higher incidence with increasing temperatures, leading to a decrease in host species or even their extinction. In the literature there are reports of massive deaths among lions, frogs, birds, wild dogs, snails, mussels, storks, eagles, corals and various types of plants, all caused by pathogens. Fungi and viruses are especially sensitive to climate change and can quickly grow when the temperature rises, especially if this elevation is accompanied by increased humidity. Fungi and insects have their activity increased and can be responsible for the elimination of entire species of trees. Moreover, parasitosis in both cattle and wild animals are also greatly increased with higher temperatures[93].
The same impact of climate change on the diversity of animals and plants is predicted for parasitic diversity. Parasites, such as helminths, are known as modulators of the immune system, can have a synergistic effect on the effects of climate change on hosts. One suggests between host and parasites interaction is that the reactivity of the immune system of host vertebrates depends on exposure to parasites, which are important for the host to develop an adequate functional immune response. Parasitic-host interactions are the main force in the evolution of the immune system that is designed to maintain optimal basal immunity, minimizing the risk of developing potentially pathological reactions [94,95].
Modifications in environmental elements such as water bodies, soil moisture, vegetation cover, among others, interfere in parasites, vectors and their hosts, as well as in the interactions between them[96]. In the case of vectors of parasitic diseases, changes in temperature influence the life cycles of parasites. After all, a portion of them have part of the life cycle in the environment[10].
Climate change has brought about significant alterations in ecosystems, intricately affecting the interactions between parasites and their hosts. The response to these environmental changes is not uniform, varying considerably even among phylogenetically related species of both parasites and hosts [97]. This variability stems from physiological, ecological, and behavioural differences that influence each organism’s sensitivity to environmental shifts[98].
Moreover, many parasites possess complex life cycles involving multiple stages that may occur in different hosts or environments[86]. Each stage may respond differently to variations in temperature, humidity, and other climatic factors, directly impacting parasite survival, development, and transmission rates[86,98]. Studies have shown that these thermal responses are often non-linear, with infection rates increasing up to an optimal point and then declining under extreme temperatures[98].
Therefore, understanding the effects of climate change on host–parasite systems require an integrative approach that considers the ecological and physiological specificities of each species involved, as well as the interactions among the various components of the system across time and space[86,97].
Climate change may favor the emergence of infectious and parasitic diseases in fish in nature or in intensive, semi-intensive and open production systems. The increase in temperature can increase parasite metabolism, the adaptation of parasitic species to new hosts, the intensification of the most abundant parasitic species to search for new hosts, contributing to greater economic loss due to parasitic lesions in these hosts, greater parasitic contamination in fish production leading to a higher dietary risk for consumers. In addition to directly affecting the physiological and immunological dynamics of the hosts. However, future studies need to be carried out to be able to understand more deeply how climate changes can interfere with host-parasite adaptations and interactions.
Acknowledgments
This research was supported by national funds through FCT - Foundation for Science and Technology within the scope of UIDB/04423/2020 and UIDP/04423/2020, and under the support FCT-Tenure 2023.15937.TENURE.005.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Brander, K.; Cochrane, K.; Barange, M.; Soto, D. Climate change implications for fisheries and aquaculture. Climate change impacts on fisheries aquaculture: A global analysis 2017, 1, 45–62. [Google Scholar] [CrossRef]
- Pachauri, R. K.; Allen, M. R.; Barros, V. R.; Broome, J.; Cramer, W.; Christ, R.; Church, J. A.; Clarke, L.; Dahe, Q.; Dasgupta, P. Climate change 2014: synthesis report. Contribution of Working Groups I, II and III to the fifth assessment report of the Intergovernmental Panel on Climate Change; Ipcc, 2014. Available in: https://www.ipcc.ch/site/assets/uploads/2018/05/SYR_AR5_FINAL_full_wcover.pdf. Accessed in: Jan 20, 2025.
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecology letters 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Ojima, R.; Marandola Jr, E. Mudanças climáticas e as cidades: novos e antigos debates na busca da sustentabilidade urbana e social; Editora Blucher, 2013.
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Patz, J. A.; Graczyk, T. K.; Geller, N.; Vittor, A. Y. Effects of environmental change on emerging parasitic diseases. International journal for parasitology 2000, 30, 1395–1405. [Google Scholar] [CrossRef]
- Charron, D. F.; Thomas, M. K.; Waltner-Toews, D.; Aramini, J. J.; Edge, T.; Kent, R. A.; Maarouf, A. R.; Wilson, J. Vulnerability of waterborne diseases to climate change in Canada: a review. Journal of Toxicology Environmental Health, Part A 2004, 67, 1667–1677. [Google Scholar] [CrossRef]
- Confalonieri, U. E.; Margonari, C.; Quintão, A. F. Environmental change and the dynamics of parasitic diseases in the Amazon. Acta tropica 2014, 129, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F. Climate change, biodiversity, ticks and tick-borne diseases: the butterfly effect. International Journal for Parasitology: parasites wildlife 2015, 4, 452–461. [Google Scholar] [CrossRef]
- Haines, A.; Kovats, R. S.; Campbell-Lendrum, D.; Corvalán, C. Climate change and human health: impacts, vulnerability and public health. Public health 2006, 120, 585–596. [Google Scholar] [CrossRef]
- Lafferty, K. D. Environmental parasitology: what can parasites tell us about human impacts on the environment? Parasitology today 1997, 13, 251–255. [Google Scholar] [CrossRef]
- McMichael, A. J.; Woodruff, R. E.; Hales, S. Climate change and human health: present and future risks. The Lancet 2006, 367, 859–869. [Google Scholar] [CrossRef]
- Pascual, M.; Dobson, A. Seasonal patterns of infectious diseases. PLoS Medicine 2005, 2, e5. [Google Scholar] [CrossRef]
- Reeves, W. C.; Hardy, J. L.; Reisen, W. K.; Milby, M. M. Potential effect of global warming on mosquito-borne arboviruses. Journal of medical entomology 1994, 31, 323–332. [Google Scholar] [CrossRef]
- Byers, J. E. Marine parasites and disease in the era of global climate change. Annual Review of Marine Science 2021, 13, 397–420. [Google Scholar] [CrossRef]
- Araujo, A.; Rangel, A.; Ferreira, L. F. R. Climatic change in northeastern Brazil: paleoparasitological data. Memórias do Instituto Oswaldo Cruz 1993, 88, 577–579. [Google Scholar] [CrossRef]
- Dunn, R. R.; Davies, T. J.; Harris, N. C.; Gavin, M. C. Global drivers of human pathogen richness and prevalence. Proceedings of the Royal Society B: Biological Sciences 2010, 277, 2587–2595. [Google Scholar] [CrossRef] [PubMed]
- Nava, A.; Shimabukuro, J. S.; Chmura, A. A.; Luz, S. L. B. The impact of global environmental changes on infectious disease emergence with a focus on risks for Brazil. ILAR journal 2017, 58, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, A. R. S. d.; Motta-Veiga, M. Análise dos impactos sociais e à saúde de grandes empreendimentos hidrelétricos: lições para uma gestão energética sustentável. Ciência Saúde Coletiva 2012, 17, 1387–1398. [Google Scholar] [CrossRef]
- Rosado-García, F. M.; Guerrero-Flórez, M.; Karanis, G.; Hinojosa, M. D. C.; Karanis, P. Water-borne protozoa parasites: the Latin American perspective. International Journal of Hygiene Environmental Health 2017, 220, 783–798. [Google Scholar] [CrossRef]
- Short, E. E.; Caminade, C.; Thomas, B. N. Climate change contribution to the emergence or re-emergence of parasitic diseases. Infectious Diseases: Research Treatment 2017, 10, 1178633617732296. [Google Scholar] [CrossRef]
- Fecchio, A.; Wells, K.; Bell, J. A.; Tkach, V. V.; Lutz, H. L.; Weckstein, J. D.; Clegg, S. M.; Clark, N. J. Climate variation influences host specificity in avian malaria parasites. Ecology Letters 2019, 22, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Kadlec, D.; Šimková, A.; Jarkovský, J.; Gelnar, M. Parasite communities of freshwater fish under flood conditions. Parasitology Research 2003, 89, 272–283. [Google Scholar] [CrossRef]
- Marcogliese, D. J. The impact of climate change on the parasites and infectious diseases of aquatic animals. Rev. sci. tech. Off. int. Epiz 2008, 27, 467–484. [Google Scholar] [CrossRef] [PubMed]
- Rohr, J. R.; Dobson, A. P.; Johnson, P. T. J.; Kilpatrick, A. M.; Paull, S. H.; Raffel, T. R.; Ruiz-Moreno, D.; Thomas, M. B. Frontiers in climate change–disease research. Trends in ecology evolution 2011, 26, 270–277. [Google Scholar] [CrossRef]
- Godfrey, G. H. Effect of temperature and moisture on nematode root knot. JOURNAL OF AGRICULTURAL RESEARCH 1926, 33, 0223–0254. [Google Scholar]
- Lesage, V.; Hammill, M. O. The status of the Grey Seal, Halichoerus grypus, in yhe Northwest Atlantic. THE CANADIAN FIELD-NATURALIST 2001, 115, 653–662. [Google Scholar] [CrossRef]
- Byers, J. E. Marine Parasites and Disease in the Era of Global Climate Change. Annual Review ofMarine Science 2021, 397, 420. [Google Scholar] [CrossRef] [PubMed]
- Lauringson, M.; Nousiainen, I.; Kahar, S.; Burimski, O.; Gross, R.; Kaart, T.; Vasemägi, A. Climate change-driven disease in sympatric hosts: Temporal dynamics of parasite burden and proliferative kidney disease in wild brown trout and Atlantic salmon. Journal Fish Diseases 2021, 44, 689–699. [Google Scholar] [CrossRef]
- Ros, A.; Baer, J.; Basen, T.; Chucholl, C.; Schneider, E.; Teschner, R.; Brinker, A. Current and projected impacts of the parasite Tetracapsuloides bryosalmonae (causative to proliferative kidney disease) on Central European salmonid populations under predicted climate change. Freshwater Biology 2021, 66, 1182–1199. [Google Scholar] [CrossRef]
- Borgwardt, F.; Unfer, G.; Auer, S.; Waldner, K.; El-Matbouli, M.; Bechter, T. Direct and indirect climate change impacts on brown trout in central Europe: How thermal regimes reinforce physiological stress and support the emergence of diseases. Frontiers in Environmental Science 2020, 59. [Google Scholar] [CrossRef]
- Bruneaux, M.; Visse, M.; Gross, R.; Pukk, L.; Saks, L.; Vasemägi, A. Parasite infection and decreased thermal tolerance: impact of proliferative kidney disease on a wild salmonid fish in the context of climate change. Functional Ecology 2017, 31, 216–226. [Google Scholar] [CrossRef]
- Bolin, J. A.; Evans, K.; Schoeman, D. S.; Spillman, C. M.; Moore, T. S.; Hartog, J. R.; Cummins, S. F.; Scales, K. L. A warming western boundary current increases the prevalenceof commercially disruptive parasites in broadbill swordfish. Fisheries Oceanography 2024, 33. [Google Scholar] [CrossRef]
- da Costa, J. C.; de Souza, S. S.; Castro, J. C.; Amanajás, R. D.; Val, A. L. Climate change affects the parasitism rate and impairs the regulation of genes related to oxidative stress and ionoregulation of Colossoma macropomum. Scientific Reports 2021, 11. [Google Scholar] [CrossRef]
- da Costa, J. C.; Val, A. L. Extreme climate scenario and parasitism affect the Amazonian fish Colossoma macropomum. Science of the Total Environment 2020, 726. [Google Scholar] [CrossRef]
- Marcotegui, P.; Merlo, M.; Irigoitia, M. M.; Gutiérrez, M. P.; Buratti, C.; Pon, J. P. S.; Parietti, M.; Timi, J. T. Local extinction of a parasite of Magellanic penguins? The effect of a warming hotspot on a ‘cold’ trematode. Parasitology 2025, 152, 275–283. [Google Scholar] [CrossRef]
- Klemme, I.; Hyvärinen, P.; Karvonen, A. Cold water reduces the severity of parasite-inflicted damage: support for wintertime recuperation in aquatic hosts. Oecologia 2021, 195, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, A.; Chaudhary, R.; Haider, M.; Naseem, I. How climate change affects parasites: the case of trematode parasite Clinostomum complanatum and its fish host Trichogaster fasiatus. Journal Parasite Disease 2020, 44, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Scharsack, J. P.; Wieczorek, B.; Schmidt-Drewello, A.; Büscher, J.; Franke, F.; Moore, A.; Branca, B.; Witten, A.; Stoll, M.; Bornberg-Bauer, E.; et al. Climate change facilitates a parasite’s host exploitation via temperature-mediated immunometabolic processes. Global Change Biology 2020, 27, 94–107. [Google Scholar] [CrossRef] [PubMed]
- Godwin, S. C.; Fast, M. D.; Kuparinen, A.; Medcalf, K. E.; Hutchings, J. A. Increasing temperatures accentuate negative fitness consequences of a marine parasite. Scientific Reports 2020, 10. [Google Scholar] [CrossRef]
- de Melo H. P, S; Takemoto, R. M.; Gonçalves, G. S. R.; Frederico, R. G.; Virgilio, L. R. Effects of climate change on the distribution of Hoplias malabaricus and its ecto- and endoparasite species in South America. Aquatic ecology 2024, 58, 999–1011. [Google Scholar] [CrossRef]
- Maicher, C.; Le Bailly, M. The impact of climate change upon intestinal parasites in central Europe during the 4th millennium BCE. Quaternary Science Reviews 2024, 338. [Google Scholar] [CrossRef]
- Lanfranchi, A. L.; Canel, D.; Alarcos, A. J.; Levy, E.; Braicovich, P. E.; Marcotegui, P.; Timi, J. T. Parasite assemblages as indicators of stability in stock structure of Cynoscion guatucupa (Sciaenidae) after a quarter of century of exploitation in a marine warming hotspot. Rev Fish Biol Fisheries 2024, 34, 1149–1166. [Google Scholar] [CrossRef]
- Porter, M.; Barton, B. P.; Williams, J.; Randall, J.; Ovaskainen, O.; Crook, D. A.; Shamsi, S. Influence of body size and environmental conditions on parasite assemblages of the black-spotted croaker (Protonibea diacanthus) (Teleostei: Sciaenidae) in northern Australia. Parasitology 2024, 151, 864–874. [Google Scholar] [CrossRef]
- Wood, C. L. W., R. L.; Preisser, W. C.; Leslie, K. L.; Mastick, N.; Greene, C.; Maslenikov, K. P.; ; Tornabene, L. J. M. K., J. M.; Essington, T. E. A reconstruction of parasite burden reveals one century of climate-associated parasite decline. PNAS Ecology 2023, 120. [Google Scholar] [CrossRef]
- Burraco, P.; Orizaola, G.; Monaghan, P.; Metcalfe, N. B. Climate change and ageing in ectotherms. Global Change Biology 2020, 26, 5371–5381. [Google Scholar] [CrossRef]
- Pörtner, H. O. Climate-dependent evolution of Antarctic ectotherms: an integrative analysis. Deep Sea Research Part II: Topical Studies in Oceanography 2006, 53, 1071–1104. [Google Scholar] [CrossRef]
- Sures, B.; Nachev, M.; Selbach, C.; Marcogliese, D. J. Parasite responses to pollution: what we know and where we go in ‘Environmental Parasitology’. Aquatic Toxicology Vectors 2017, 10, 1–19. [Google Scholar] [CrossRef]
- Birk, S.; Chapman, D.; Carvalho, L.; Spears, B. M.; Andersen, H. E.; Argillier, C.; Auer, S.; Baattrup-Pedersen, A.; Banin, L.; Beklioğlu, M. Impacts of multiple stressors on freshwater biota across spatial scales and ecosystems. Nature Ecology Evolution 2020, 4, 1060–1068. [Google Scholar] [CrossRef]
- Topal, A.; Özdemir, S.; Arslan, H.; Çomaklı, S. How does elevated water temperature affect fish brain?(A neurophysiological and experimental study: Assessment of brain derived neurotrophic factor, cFOS, apoptotic genes, heat shock genes, ER-stress genes and oxidative stress genes). Fish Shellfish Immunology 2021, 115, 198–204. [Google Scholar] [CrossRef]
- Alfonso, S.; Gesto, M.; Sadoul, B. Temperature increase and its effects on fish stress physiology in the context of global warming. Journal of Fish Biology 2021, 98, 1496–1508. [Google Scholar] [CrossRef]
- Novacek, M. J., Cleland, E. E. The current biodiversity extinction event: scenarios for mitigation and recovery, in: National Academy of Sciences Colloquium The Future of Evolution. Irvine, California 2001. [CrossRef]
- Franke, A.; Beemelmanns, A.; Miest, J. J. Are fish immunocompetent enough to face climate change? Biology Latters 2024, 20. [Google Scholar] [CrossRef]
- Feidantsis, K.; Georgoulis, I.; Zachariou, A.; Campaz, B.; Christoforou, M.; Pörtner, H. O.; Michaelidis, B. Energetic, antioxidant, inflammatory and cell death responses in the red muscle of thermally stressed Sparus aurata. Journal of Comparative Physiology B 2020, 190, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S. Oxidative stress, inflammation, and disease. In Oxidative stress and biomaterials, Elsevier, 2016; pp 35-58. DOI: B978-0-12-803269-5.00002-4.
- Reuter, S.; Gupta, S. C.; Chaturvedi, M. M.; Aggarwal, B. B. Oxidative stress, inflammation, and cancer: how are they linked? Free radical biology medicine 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.-Y.; Cho, S.-H.; Kwon, S.-H.; Eom, C.-Y.; Jeong, M. S.; Lee, W.; Kim, S.-Y.; Heo, S.-J.; Ahn, G.; Lee, K. P. The roles of NF-κB and ROS in regulation of pro-inflammatory mediators of inflammation induction in LPS-stimulated zebrafish embryos. Fish shellfish immunology 2017, 68, 525–529. [Google Scholar] [CrossRef]
- Poynter, S.; Dixon, B. Bony Fish Immunology. Elsevier Ltd 2017, 1–7. [Google Scholar] [CrossRef]
- Press, C. M.; Evensen, Ø. The morphology of the immune system in teleost fishes. Fish shellfish immunology 1999, 9, 309–318. [Google Scholar] [CrossRef]
- Alvarez-Pellitero, P. Fish immunity and parasite infections: from innate immunity to immunoprophylactic prospects. Veterinary immunology imunopathology 2008, 126, 171–198. [Google Scholar] [CrossRef]
- Tort, L.; Balasch, J. C.; Mackenzie, S. Fish immune system. A crossroads between innate and adaptive responses. Inmunología 2003, Imunologia. 22, 277-286.
- Filipe, J. F.; Herrera, V.; Curone, G.; Vigo, D.; Riva, F. Floods, hurricanes, and other catastrophes: a challenge for the immune system of livestock and other animals. Frontiers in veterinary science 2020, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Makrinos, D. L.; Bowden, T. J. Natural environmental impacts on teleost immune function. Fish Shellfish Immunology 2016, 53, 50–57. [Google Scholar] [CrossRef]
- Stankus, A. State of world aquaculture 2020 and regional reviews: FAO webinar series. FAO Aquaculture Newsletter 2021, (63), 17-18. Available in: https://www.fao.org/fishery/en/openasfa/403fc2bd-ab68-4549-b8fc-6f7b6fff9336. Accessed in: May 30, 2025.
- Tacon, A. G. J. Trends in global aquaculture and aquafeed production: 2000–2017. Reviews in Fisheries Science Aquaculture 2020, 28, 43–56. [Google Scholar] [CrossRef]
- Valladão, G. M. R.; Gallani, S. U.; Pilarski, F. South American fish for continental aquaculture. Reviews in Aquaculture 2018, 10, 351–369. [Google Scholar] [CrossRef]
- Methling, C.; Aluru, N.; Vijayan, M. M.; Steffensen, J. F. Effect of moderate hypoxia at three acclimation temperatures on stress responses in Atlantic cod with different haemoglobin types. Comparative Biochemistry Physiology Part A: Molecular Integrative Physiology 2010, 156, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Jesus, T. F.; Moreno, J. M.; Repolho, T.; Athanasiadis, A.; Rosa, R.; Almeida-Val, V. M. F.; Coelho, M. M. Protein analysis and gene expression indicate differential vulnerability of Iberian fish species under a climate change scenario. PLoS One 2017, 12, e0181325. [Google Scholar] [CrossRef] [PubMed]
- Karvonen, A.; Rintamäki, P.; Jokela, J.; Valtonen, E. T. Increasing water temperature and disease risks in aquatic systems: climate change increases the risk of some, but not all, diseases. International journal for parasitology 2010, 40, 1483–1488. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J. Y.; Markkandan, K.; Han, K.; Kwon, M. G.; Seo, J. S.; Yoo, S.; Hwang, S. D.; Ji, B. Y.; Son, M.; Park, J. Temperature-dependent immune response of olive flounder (Paralichthys olivaceus) infected with viral hemorrhagic septicemia virus (VHSV). Genes genomics 2018, 40, 315–320. [Google Scholar] [CrossRef]
- Kayansamruaj, P.; Pirarat, N.; Hirono, I.; Rodkhum, C. Increasing of temperature induces pathogenicity of Streptococcus agalactiae and the up-regulation of inflammatory related genes in infected Nile tilapia (Oreochromis niloticus). Veterinary microbiology 2014, 172, (1–2). [Google Scholar] [CrossRef]
- Bhanumathi, K.; Sasirekhamani, M. Sustaining Aquatic Resources: Climatic Changes Impacts and Adaptation Strategies in Aquaculture and Fisheries. Paradigm Shift: Multidisciplinary Research for a Chanching World 2024, 2, 77–84. [Google Scholar]
- De Silva, S. S.; Soto, D. Climate change and aquaculture: potential impacts, adaptation and mitigation. Climate change implications for fisheries aquaculture: overview of current scientific knowledge. FAO Fisheries Aquaculture Technical Paper 2009, 530, 151–212. [Google Scholar]
- Schmittner, A. Decline of the marine ecosystem caused by a reduction in the Atlantic overturning circulation. Nature 2005, 434, 628–633. [Google Scholar] [CrossRef]
- Eiras, J. C. Elementos da Ictioparasitologia 1994. Porto : Fundação Eng. António de Almeida.
- Koskivaara, M. V., E. T.; Prost, M. Dactylogyrids on the gills of roach in central Finland: features of infection and species composition. International Journal of Parasitology 1991, 21, 565–572. [Google Scholar] [CrossRef]
- Cascarano, M. C.; Stavrakidis-Zachou, O.; Mladineo, I.; Thompson, K. D.; Papandroulakis, N.; Katharios, P. Mediterranean Aquaculture in a Changing Climate: Temperature Effects on Pathogens and Diseases of Three Farmed Fish Species. Pathogens 2021, 10. [Google Scholar] [CrossRef]
- Pounds, J. A.; Bustamante, M. R.; Coloma, L. A.; Consuegra, J. A.; Fogden, M. P. L.; Foster, P. N.; La Marca, E.; Masters, K. L.; Merino-Viteri, A.; Puschendorf, R.; et al. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 2006, 439, 161–167. [Google Scholar] [CrossRef]
- Chame, M.; Batouli-Santos, A. L.; Brandão, M. L. As migrações humanas e animais ea introdução de parasitas exóticos invasores que afetam a saúde humana no Brasil. FUMDHAMentos 2008, 7, 47–62. [Google Scholar]
- Lafferty, K. D.; Shaw, J. C. Comparing mechanisms of host manipulation across host and parasite taxa. Journal of Experimental Biology 2013, 216(1), 56–66. [Google Scholar] [CrossRef]
- Carlson, C. J.; Burgio, K. R.; Dougherty, E. R.; Phillips, A. J.; Bueno, V. M.; Clements, C. F.; Castaldo, G.; Dallas, T. A.; Cizauskas, C. A.; Cumming, G. S. Parasite biodiversity faces extinction and redistribution in a changing climate. Science advances 2017, 3, e1602422. [Google Scholar] [CrossRef]
- Poulin, R. Global warming and temperaturemediated increases in cercarial emergence in trematode parasites. Parasitology 2006, 132, 143–151. [Google Scholar] [CrossRef]
- Studer, A.; Thieltges, D. W.; Poulin, R. Parasites and global warming: net effects of temperature on an intertidal host–parasite system. Marine Ecology Progress Series 2010, 415, 11–22. [Google Scholar] [CrossRef]
- Lõhmus, M.; Björklund, M. Climate change: what will it do to fish—parasite interactions? Biological Journal of the Linnean Society 2015, 116, 397–411. [Google Scholar] [CrossRef]
- Pietrock, M.; Marcogliese, D. J. Free-living endohelminth stages: at the mercy of environmental conditions. Trends Parasitol 2023, 19, 293–299. [Google Scholar] [CrossRef]
- Marcogliese, D. J. Climate Change and Parasitism in Aquatic Ecosystems; Smit, N. J., Sures, B., Eds.; 2025. [CrossRef]
- Morley, N. J.; Lewis, J. W. Thermodynamics of cercarial development and emergence in trematodes. Parasitology 2013, 140, 1211–1224. [Google Scholar] [CrossRef]
- Buchmann, K.; Lindenstrøm, T. Interactions between monogenean parasites and their fish hosts. International journal for parasitology 2002, 32, 309–319. [Google Scholar] [CrossRef]
- Ogawa, K. Diseases of cultured marine fishes caused by Platyhelminthes (Monogenea, Digenea, Cestoda). Parasitology 2015, 142, 178–195. [Google Scholar] [CrossRef]
- WOO, P. T.; GREGORY, D. W. B. Diseases and disorders of finfish in cage culture. CABI 2014. [Google Scholar]
- Macnab, V.; Barber, I. Some (worms) like it hot: fish parasites grow faster in warmer water, and alter host thermal preferences. Global Change Biology 2012, 18, 1540–1548. [Google Scholar] [CrossRef]
- Costa, J. C. V., A. L. Extreme climate scenario and parasitism affect the Amazonian fish Colossoma macropomum. Science of the Total Environment 2020, 726. [Google Scholar] [CrossRef]
- Harvell, C. D. , Mitchell, C. E., Ward, J. R., Altizer, S., Dobson, A. P., Ostfeld, R. S., Samuel, M. D. Climate Warming and Disease Risks for Terrestrial and Marine Biota. Science’s Compass Review 2002, 296. [Google Scholar] [CrossRef]
- Peuß, R.; Box, A. C.; Chen, S.; Wang, Y.; Tsuchiya, D.; Persons, J. L.; Kenzior, A.; Maldonado, E.; Krishnan, J.; Scharsack, J. P. Adaptation to low parasite abundance affects immune investment and immunopathological responses of cavefish. Nature ecology evolution 2020, 4, 1416–1430. [Google Scholar] [CrossRef] [PubMed]
- Scharsack, J. P.; Wieczorek, B.; Schmidt-Drewello, A.; Büscher, J.; Franke, F.; Moore, A.; Branca, A.; Witten, A.; Stoll, M.; Bornberg-Bauer, E. Climate change facilitates a parasite's host exploitation via temperature-mediated immunometabolic processes. Global Change Biology 2020, 27, 94–107. [Google Scholar] [CrossRef] [PubMed]
- Haines, A. P., J. A. Health Effects of Climate Change. Journal of the American Medical Association 2004, 291, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Ismail, N.; Seppälä, O.; Valtonen, E. T. Temperature and intraspecific variation affect host–parasite interactions. Oecologia 2023, 202, 567–578. [Google Scholar] [CrossRef]
- Wells, K.; Flynn, R. Managing host-parasite interactions in humans and wildlife in times of global change. Parasitology Research 2022, 121, 3361–3375. [Google Scholar] [CrossRef]
Figure 1.
Distribution of the total number of articles published until June 2025 on climate change in the most varied areas of science, according to the platform Web of Science.
Figure 1.
Distribution of the total number of articles published until June 2025 on climate change in the most varied areas of science, according to the platform Web of Science.
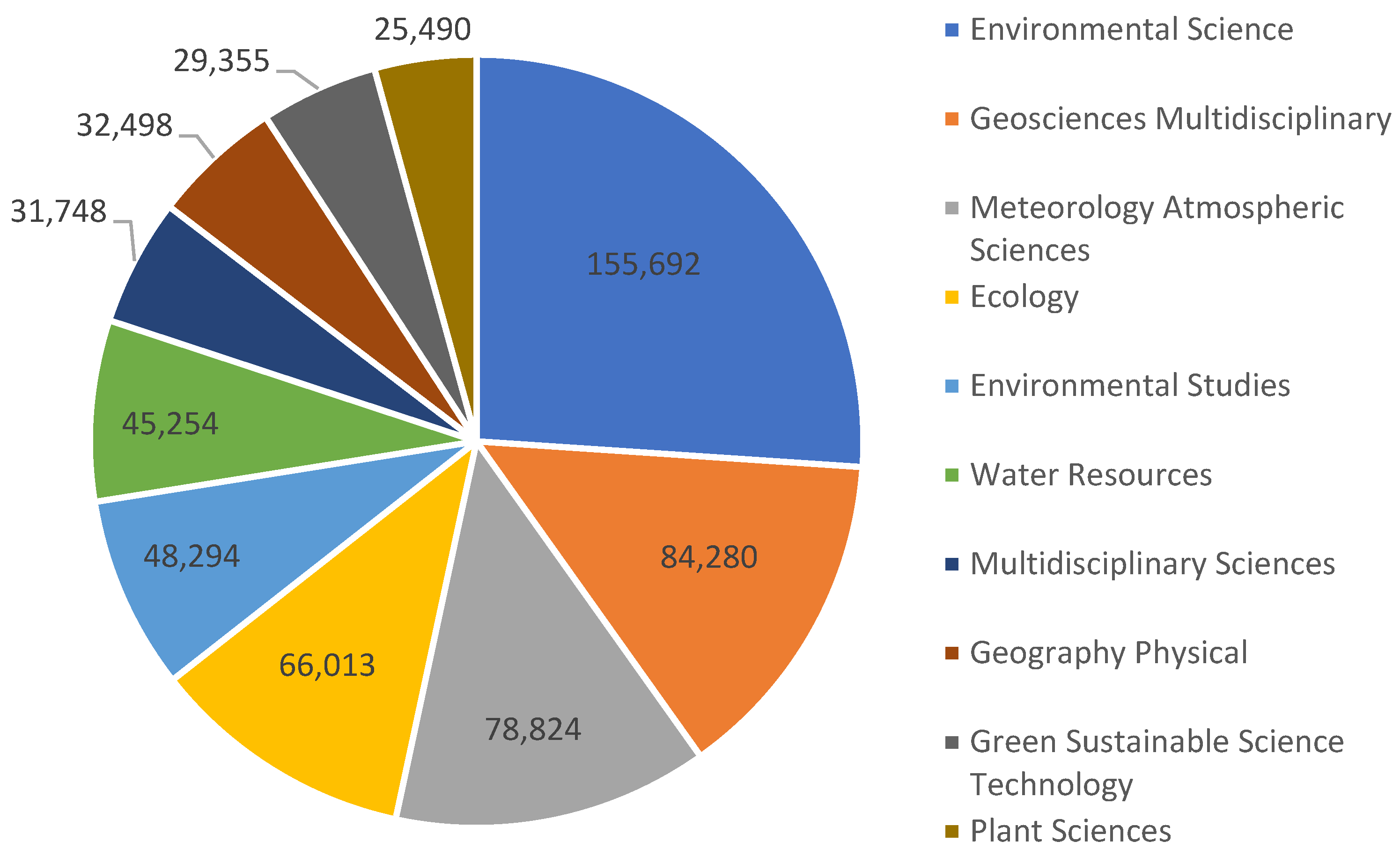
Figure 2.
Annual distribution (2001 – 2025) of the number of publications that related climate changes, parasites and fish in freshwater or marine environment, according to the platform of the Web of Science.
Figure 2.
Annual distribution (2001 – 2025) of the number of publications that related climate changes, parasites and fish in freshwater or marine environment, according to the platform of the Web of Science.
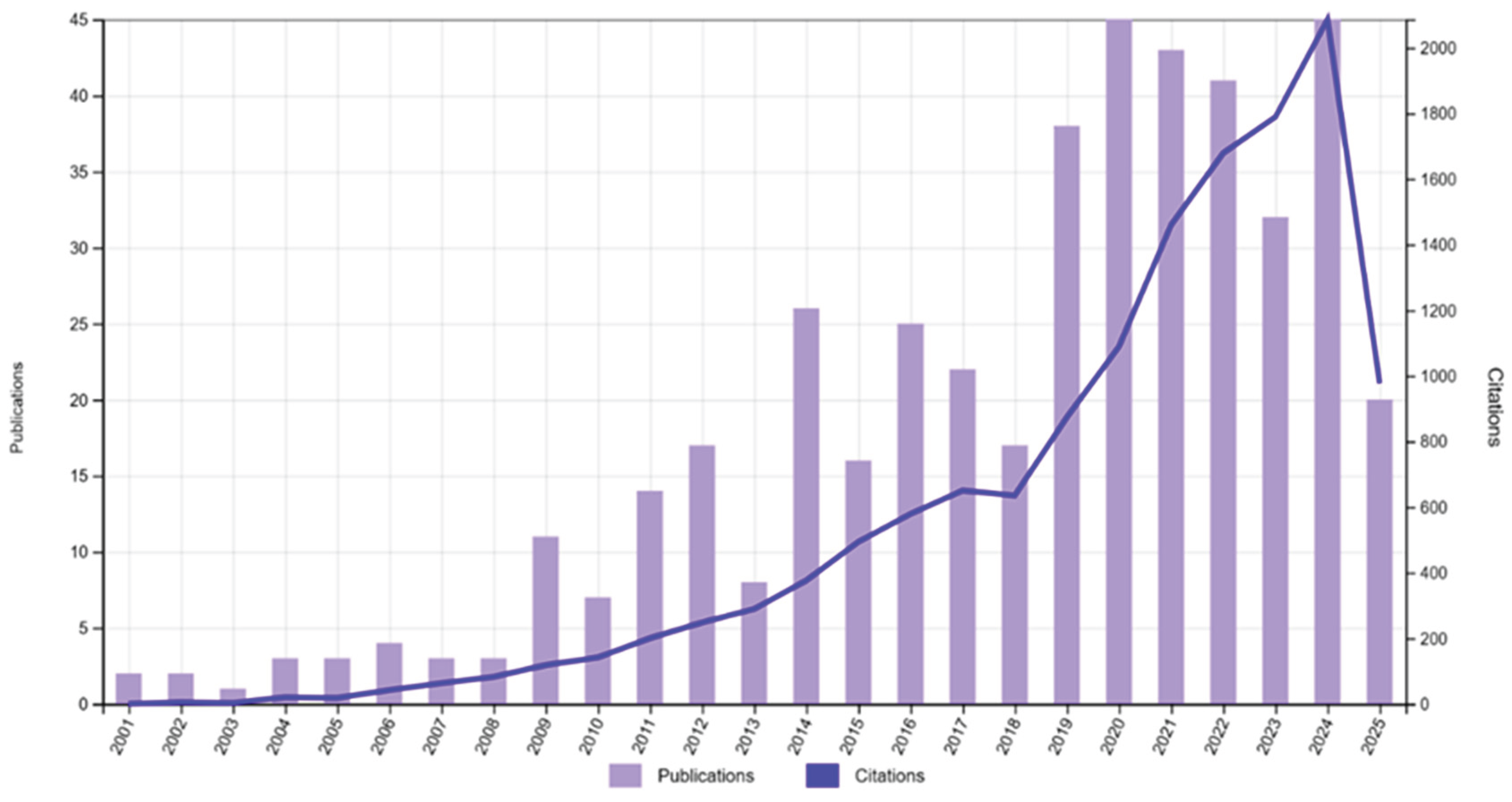

Table 1.
Articles that present the contributions in the literature that relate climate change as the main interference in the parasite-host interaction.
Table 1.
Articles that present the contributions in the literature that relate climate change as the main interference in the parasite-host interaction.
| Parasite Group/ Genus or Specie | Host | Major goals |
Major finding | Reference |
|---|---|---|---|---|
| Dinoflagelatta: Syndiniophyceae | ||||
|
Hematodinium sp. H. perezi |
Homarus americanus; Chionoecete sopilio; Callinectes sapidus | To examine how climate change affects ecological processes and interactions, including parasitism, especially in marine systems. |
|
[28] |
| Cnidaria Malacosporea | ||||
| Tetracapsuloides bryosalmonae |
Salmo salar; Salmo trutta |
To investigate how climate change influences the temporal dynamics of parasite load and clinical signs of proliferative kidney disease (PKD) in two species of sympatric salmonid fish: brown trout and Atlantic salmon. |
|
[29] |
| Tetracapsuloides bryosalmonae |
Salmo salar; Salmo trutta |
To investigate the impacts of proliferative kidney disease (PKD), caused by the parasite Tetracapsuloides bryosalmonae, on salmonid populations in Central Europe, considering the effects of climate change. |
|
[30] |
| Tetracapsuloides bryosalmonae | Salmo trutta | To assess how climate change, through increased water temperature, directly and indirectly affects brown trout in Austria, with a focus on proliferative kidney disease (PKD) emergence and physiological heat stress. |
|
[31] |
| Tetracapsuloides bryosalmonae | Salmo trutta | To investigate how infection by the parasite that causes proliferative kidney disease (PKD) affects the physiology of fry brown trout in a natural environment, focusing on thermal tolerance and aerobic performance, in the context of climate change. |
|
[32] |
| ||||
|
Kudoa musculoliquefaciens |
Xiphias gladius |
To investigate the environmental factors that influence the prevalence and intensity of infection by the parasite Kudoa musculoliquefaciens in swordfish caught at the Eastern Tuna and Billfish Fisheries (ETBF), focusing on how the warming of the Australian Eastern Current may increase the risk of myoliquefaction. |
|
[33] |
| ||||
| Monogenea (2 species) |
Colossoma macropomum |
To investigate whether different periods of exposure to climate change scenarios (7 and 30 days) and two degrees of parasitism (low and high) would affect the environment-parasite-host interaction and the host antioxidant and ion regulatory responses. |
|
[34] |
| Monogenea (3 species) |
Colossoma macropomum | To investigate the combined effects of an extreme climate scenario (predicted for 2100) and different levels of parasitism by monogenea on the hematological and immunological response of the host, a keystone species in Amazonian aquaculture. |
|
[35] |
| ||||
| Cardiocephaloides physalis | Engraulis anchoita | To corroborate the specific identity of metacercariae from anchovies as C. physalis and to assess if changes in parasitism did occur in both fish and seabird hosts and the congruence between them, analysing the possible causes for such changes in a scenario of a rapid regional tropicalization. |
|
[36] |
|
Diplostomum pseudospathaceum |
Salmo salar Salmo trutta |
To investigate how water temperature affects parasite damage (parasite virulence) in trematode-infected salmonid fish, focusing on the possibility of host recovery during periods of cold water. |
|
[37] |
|
Clinostomum complanatum |
Trichogaster fasciatus |
To investigate the influence of climatic variables, especially air temperature, on the prevalence of the trematode parasite in its intermediate host, fish, over seven years (2007–2013) in the Aligarh region, India. |
|
[38] |
| ||||
|
Schistocephalus solidus |
Gasterosteus aculeatus |
To investigate how the increase in temperature, associated with climate change, affects the interaction between the ectothermic three-spiny spiny fish and the parasite, focusing on the immunometabolic effects on the host. |
|
[39] |
| ||||
|
Lepeophtheirus salmonis |
Solmo salar | To investigate how the increase in water temperature influences the impacts of the marine parasite (salmon lice) on the growth, body condition and survival of Atlantic salmon, a fish of ecological and economic importance. |
|
[40] |
| ||||
| Monogenea (2 species) Digenea (3 species) |
Hoplias malabaricus | To develop tools to assist in monitoring, conservation, and actions that contribute to the biodiversity of parasites in South America |
|
[41] |
| Digenea (2 species) Cestoda (1 species) Nematoda (1 specie) |
Fishes | To know about the dispersion of zoonoses via food and to know the relationships between zoonotic cycles, anthropization and climate. |
|
[42] |
| Monogenea (2 species) Digenea (5 species) Cestoda (4 specie) Nematoda (5 species) Acanthocephala (2 species) Copepoda (2 species) |
Cynoscion guatucupa |
To evaluate the possible changes in the structure of parasitic communities of the fish host over approximately 25 years in the northern Argentine Sea, using parasites as biological markers ("tags") to investigate the stability of the stock structure of this species in a marine region characterized by accelerated warming. |
|
[43] |
| Monogenea (1 specie) Digenea (5 species) Nematoda (3 species) Copepoda (2 species) |
Protonibea diacanthus | This study showed as parasites respond the environmental changes and their impacts on hosts as prevalence, abundance a distribution parasite. Also was consider dissolved oxygen, temperature, salinity and ammonia into parameters. |
|
[44] |
| Monogenea; Trematoda; Cestoda; Acanthocephala; Nematoda; Hirudinea and Copepoda |
Clupea pallasii; Embiotoca lateralis; Hydrolagus colliei; Hypomesus pretiosus; Merluccius productus; Parophrys vetulus; Sebastes caurinus and Gabus chalcogrammus | To determine how parasite populations have changed over the past century and whether these changes are linked to environmental factors, particularly climate change. |
|
[45] |
Table 2.
Habitat loss and projected extinction risk by dispersal scenario and great group and values are averaged across all general circulation models (GCMs) and RCP scenarios, and the percentage of species committed to extinction. Adapted from Carlson et al. 2017 76.
Table 2.
Habitat loss and projected extinction risk by dispersal scenario and great group and values are averaged across all general circulation models (GCMs) and RCP scenarios, and the percentage of species committed to extinction. Adapted from Carlson et al. 2017 76.
| Great Group | Number of the species | Habitat loss (mean) | Habitat loss range (5-95t%) | % Committed to extinction |
|---|---|---|---|---|
| 0% dispersal | ||||
| Acanthocephala | 14 | −16.6% | (−50.6%, −0.2%) | 3.8%/4.4%/4.9% |
| Astigmata | 18 | −19.0% | (−43.6%, −4.0%) | 4.4%/5.1%/5.3% |
| Cestoda | 25 | −13.6% | (−29.1%, −2.9%,) | 4.0%/3.6%/3.7% |
| Ixodida | 141 | −31.9% | (−57.0%, −1.9%) | 8.1%/9.2%/9.8% |
| Nematoda | 147 | −28.0% | (−74.4%, −2.6%) | 5.4%/7.9%/9.3% |
| Phthiraptera | 5 | −55.8% | (−71.5%, −34.4%) | 10.5%/18.5%/19.3% |
| Siphonaptera | 67 | −40.6% | (−69.5%, −11.0%) | 10.0%/12.2%/12.9% |
| Trematoda | 40 | −17.8% | (−47.4%, −0.4%) | 3.8%/4.8%/6.0% |
| 100% dispersal | ||||
| Acanthocephala | 14 | 48.80% | (−10.4%, +129.0%) | 0.21%/0.54%/0.60% |
| Astigmata | 18 | 13.80% | (−41.2%, +64.4%) | 1.3%/2.0%/2.3% |
| Cestoda | 25 | 57.10% | (+3.7%, +131.1%) | 0.07%/0.07%/0.07% |
| Ixodida | 141 | −8.6% | (−54.1%, +67.7%) | 4.9%/5.7%/6.4% |
| Nematoda | 147 | 18.70% | (−53.6%, +87.6%) | 1.3%/2.5%/3.3% |
| Phthiraptera | 5 | 110.50% | (−57.7%, +514.8%) | 4.6%/6.2%/7.4% |
| Siphonaptera | 67 | −5.0% | (−50.0%, +43.8%) | 1.9%/4.1%/4.6% |
| Trematoda | 40 | 82.20% | (−30.4%, +242.4%) | 0.11%/1.0%/1.2% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



