Submitted:
17 November 2025
Posted:
18 November 2025
You are already at the latest version
Abstract
Melatonin, an ancient and conserved indolamine, has always attracted attention for its multifunctional roles in redox balance. More recently, it has been studied in relation to immune regulation and cancer biology. Beyond its well-known circadian function, melatonin modulates oxidative stress by directly scavenging reactive oxygen and nitrogen species and by upregulating antioxidant enzymes such as superoxide dismutase, catalase, and glutathione peroxidase. At the same time, it exerts wide-ranging immunomodulatory functions by influencing both the innate and adaptive immune responses. All these actions converge in the tumor microenvironment, where oxidative stress and immune suppression drive cancer progression. Notably, melatonin influences macrophage polarization, favoring antitumor M1 phenotypes over pro-tumoral M2 states, while attenuating chronic inflammation and restoring mitochondrial function. This review summarizes current knowledge on melatonin’s antioxidant and immunoregulatory mechanisms, highlighting its impact on the tumor immune microenvironment and its potential use as an adjuvant in cancer prevention and treatment.
Keywords:
melatonin
; oxidative stress
; antioxidants
; immune modulation
; macrophages
; tumor microenvironment
; cancer therapy
1. Introduction
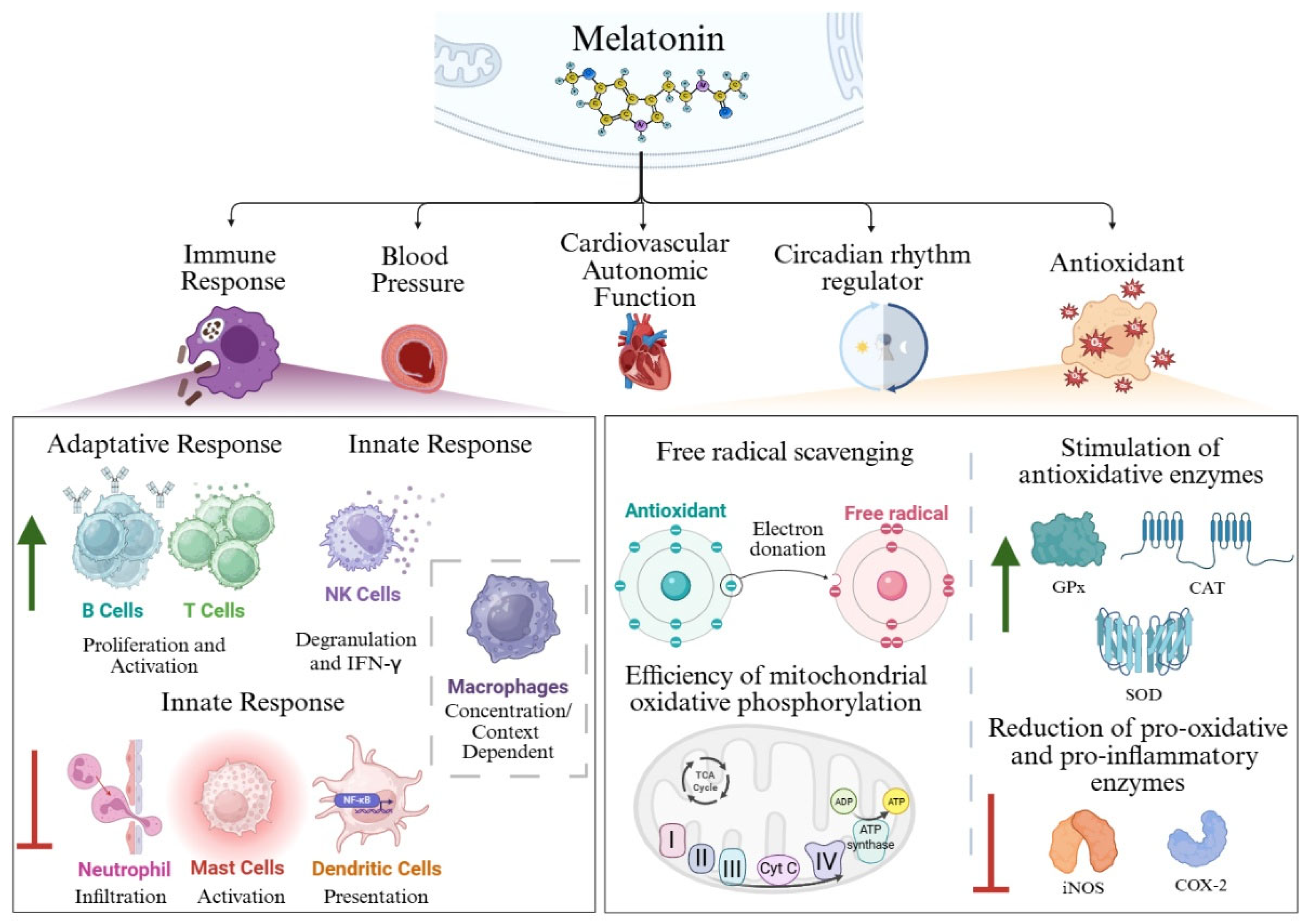
Melatonin, or N-acetyl-5-methoxytryptamine, is a small indolamine with many physiological and biological roles [1]. Aaron Lerner and colleagues isolated melatonin from cow pineal glands in 1958 for the first time [2,3,4]. They nhamed it melatonin because those pineal gland extracts contained a molecule able to light frog and fish melanocytes [5]. Melatonin production is found across all domains of life, including organisms without a pineal gland such as archaea, bacteria, unicellular eukaryotes, invertebrates, and even vascular plants [6]. Several studies suggest that mitochondria serve as the primary site of melatonin synthesis, supporting the production by species so phylogenetically apart [5,7]. In plants, additional evidence indicates that chloroplasts also contribute hto melatonin biosynthesis. The precise synthetic pathways in microorganisms remain largely unknown, although some recent studies have shed some light in this area [8,9,10]. In humans, melatonin is synthesized in almost all cell types, tissues, and organs. The pineal gland is the most well-known site of production, but melatonin production is very similar in other parts of the body such as the enterochromaffin cells in the gastrointestinal tract and the skin cells [5]. Melatonin derives from tryptophan through a process of hydroxylation by the enzyme tryptophan hydroxylase [1,11,12]. The resulting product, 5-hydroxytryptophan, is decarboxylated by the enzyme aromatic amino acid decarboxylase resulting in serotonin. Serotonin will be transformed into melatonin by the sequential action of the enzymes N-acetyltransferase and N-acetylserotonin O-methyltransferase [11,12]. One synthesized, this hormone is released into the systemic circulation to reach all tissues [11]. This molecule is associated with a broad spectrum of physiological processes, including regulation of blood pressure, cardiovascular autonomic function, and immune response (Figure 1). Furthermore, it plays key roles as a circadian rhythm regulator, including the modulation of sleep-wake cycles and antioxidant [1,5,13,14]. Melatonin can exert its effect through endocrine, autocrine and paracrine mechanisms. Over the course of evolution, melatonin has acquired additional functions, spotlighting as a signaling molecule involved in the regulation of photoperiodic responses through endocrine pathways [5,18]. Its amphiphilicity allows the molecule to enter any cell, compartment or body fluid [15]. Melatonin mainly exerts its functions through binding to G protein-coupled membrane receptors (Melatonin Receptor 1 (MT1), MT2 and MT3), orphan nuclear receptors (such as orphan receptor retinoid Z receptor beta (RZR-β), retinoic acid-related orphan receptor alpha (ROR-α)), as well as through its interaction with intracellular proteins such as Calmodulin [16]. MT1 receptors contribute to the regulation of circadian rhythms and contraction of cardiac vessels, mainly. MT2 receptors are located especially in the retina and brain, exerting functions related to retinal physiology and to the inflammatory response. They are also involved in circadian rhythm and cardiac vessel dilation (Figure 1). Finally, the MT3, that was renamed from Mel2 receptors, plays a similar role to MT2 [16,17].
2. Antioxidant Function of Melatonin
Melatonin initial and most conserved function across diverse organisms is as an antioxidant and free radicals’ scavenger, vital functions for cellular homeostasis [5,18]. Melatonin acts as a unique antioxidant, neutralizing more than one molecule, moving across physiological barriers, stimulating other endogenous antioxidant enzymes and improving the efficacy of other antioxidants and protecting mitochondria. Thus, it is significantly better than classical antioxidants [5,19]. Melatonin's functions as an antioxidant include (Figure 1):
These functions are carried out by melatonin, but also by the three main metabolites of melatonin: cyclic 3-hydroxymelatonin (3OHM), N-acetyl-N-formyl-5-methoxyquinuramine (AFMK) and N-acetyl-5-methoxyquinuramine (AMK) [29]. AFMK can be formed both by enzymatic or pseudoenzymatic and nonenzymatic metabolic pathways [29,30]. AMK is also formed by deformylation of AFMK [29].
2.1. Free radical Scavenging
The use of oxygen represented an important cost to aerobic organisms. It produces free radicals: reactive oxygen species (ROS) and reactive nitrogen species (RNS). Their accumulation creates an imbalance of homeostasis that is detrimental to cell survival [31]. Both melatonin, and its metabolites, can initiate a cascade to neutralize free radicals such as superoxide anions (O2-), hydroxyl radicals(·OH), nitric oxide(·NO) and peroxynitrite anions (ONOO-) [32,33]. A study carried out by Pieri et al. (1996) showed that melatonin exhibits antioxidant activity about three times greater than reduced glutathione (GSH) and about twice greater than vitamins C and E [13]. Furthermore, melatonin´s metabolite AMK is a potent scavenger of singlet oxygen (¹O₂) and NO [30], whereas the metabolite 3OHM effectively scavenges ·OH, 2, 2′-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid) (ABST), and peroxide radicals [29,30].
2.2. Stimulation of Antioxidative Enzymes
In addition to its free radical scavenging capacity, melatonin stimulates Sirtuin 3 (SIRT3), which deacetylates FoxO3a activating an antioxidant response by modulating superoxide dismutase (SOD) and catalase (CAT) enzymes [23,34]. It also upregulates the expression of antioxidant enzyme genes through MT1/MT2 receptors, such as glutathione peroxidase (GPx) [14,23,24,34]. This effect is mediated by the rise of γ-glutamylcysteine synthase activity, which is the rate-limiting enzyme of the GSH synthesis pathway[14,24,35]. As an indirect antioxidant it can also downregulate prooxidative and pro-inflammatory enzymes, such us cyclooxygenase 2 and inducible nitric oxide synthase (iNOS) [31]. (Figure 1)
2.3. Efficiency of Mitochondrial Oxidative Phosphorylation
Melatonin synthesis in the mitochondria is well conserved and its presence in this organelle can be further increased by entering through the oligopeptide transporters 1 (PEPT-1) and PEPT2. This indicates the relevance of melatonin neutralizing ROS produced naturally by the electron respiratory chain complexes [26]. Studies by Okatani et al. (2003) showed that melatonin restored the respiratory physiology of aged mouse hepatocytes, specifically at the level of complexes I and IV [36]. Besides, melatonin has been observed to stimulate oxidative phosphorylation and promote ATP production in neuronal and hepatocyte mitochondria [26]. In addition, melatonin increases the activity of uncoupling proteins (UCP) by direct action or by upregulating their gene expression [28]. These proteins transport intermembrane protons back to the mitochondrial matrix, reducing the membrane potential (Δψ), thus accelerating electron transport through the electron transport chain. This phenomenon drastically reduces the loss of electrons and, consequently, the formation of ROS [28]. Another protective function of melatonin through the mitochondria is related to apoptosis. When melatonin binds to the mitochondrial MT1 receptor, it activates Gαi, which blocks adenylate cyclase activity, and inhibits stress-induced cytochrome c release, thereby dampening caspase activation [27].
Figure 1.
Schematic representation of the biological functions of melatonin. Melatonin exerts multiple physiological roles. Within the immune system, melatonin enhances the proliferation and activation of T and B lymphocytes. In mast cells, it promotes cellular degranulation, IFN-γ release, neutrophil infiltration, and antigen presentation by dendritic cells. In macrophages, however, the effects of melatonin appear to be context- and concentration-dependent. Regarding its antioxidant capacity, melatonin functions as a potent free radical scavenger, stimulates antioxidant enzymes, downregulates pro-oxidant and pro-inflammatory enzymes, and improves the efficiency of mitochondrial oxidative phosphorylation. Created in BioRender. Garcia-Macia, M. (2025) https://BioRender.com/v72fvv9.
Figure 1.
Schematic representation of the biological functions of melatonin. Melatonin exerts multiple physiological roles. Within the immune system, melatonin enhances the proliferation and activation of T and B lymphocytes. In mast cells, it promotes cellular degranulation, IFN-γ release, neutrophil infiltration, and antigen presentation by dendritic cells. In macrophages, however, the effects of melatonin appear to be context- and concentration-dependent. Regarding its antioxidant capacity, melatonin functions as a potent free radical scavenger, stimulates antioxidant enzymes, downregulates pro-oxidant and pro-inflammatory enzymes, and improves the efficiency of mitochondrial oxidative phosphorylation. Created in BioRender. Garcia-Macia, M. (2025) https://BioRender.com/v72fvv9.
3. Immunomodulatory and Anti-inflammatory Actions of Melatonin
The first line of defense (after the skin) against pathogens and diseases is the immune system. A complex biological network formed by different cell types, tissues, and organs that releases specific proteins and molecules, and provides protection [37]–[39]. There are two main types of immunity: the adaptative, or specific response; and the innate, or non-specific response [37]–[39].
The adaptive immune response is more specific, and its magnitude increases with successive exposures to a specific insult. This type of response is slower since it requires cellular activation [37]–[39]. The cell types involved in this type of response are T and B lymphocytes, along with various cytokines and antibodies (humoral immunity) [37]–[39].
The innate response includes defense mechanisms that are present before infection occurs, facilitating a rapid response. The main cellular components are macrophages, neutrophils, basophils, eosinophils and natural killer (NK) cells. These cells release various soluble factors, such as cytokines: tumor necrosis factor alpha (TNF-α), (IL)-1β, IL-6 and IL-8.
Interleukins and cytokines produced in the immune response modulate the synthesis and release of melatonin. For instance, an increase in the inflammatory cytokine IL-1β stimulates the signals of the endocrine system (melatonin and cortisol) to counteract the secretion of IL-1β [40]. This reveals an intimate relationship between the immune system and the endocrine system. Many immune system cells, such as lymphoid cells, express melatonin receptors and a bidirectional communication circuit among both systems (immune and melatonin) is established [40,41]. As an example of the role of melatonin in the immune system, Maestroni et al. (1986) described in mice that inhibition of melatonin synthesis reduces the adaptive, cellular and humoral response, which can be reversed by melatonin administration [42]. Besides, circulating leukocytes and monocytes are a well-established source of extra pineal melatonin [40,43].
3.1. Adaptative Immune Response
3.1.1. T Cells
T cells develop from bone marrow (BM)-derived thymocyte progenitors and mature in the thymus. Broadly speaking, this cell population is composed of CD4+ and CD8+ T lymphocytes [39,44]. They activate and clonally expand by the interaction of the T cell receptor (TCR) with the processed antigen present on the major histocompatibility complex (MHC) molecules of antigen presenting cells (APCs) (signal 1) [39,44]. In addition, they can interact with co-stimulatory molecules (signal 2) and cytokines (signal 3) [39,44]. Upon activation, these cells release cytokines which initiate signals that trigger cell death in the target cell, mainly through direct interaction [39,44]. Depending on the cytokine-mediated activation signal, these T cells can be activated toward different programs [45]. IFN-γ and IL-12 drive the differentiation of Th1 cells responsible for protection against intracellular viruses and bacteria. IL-13 and IL-4 cytokines induce differentiation towards a Th2 phenotype acting against helminth infections [45]. This phenotype also facilitates tissue repair and can contribute to chronic inflammation, typically in allergies and asthma. While IL-6, IL-21 and IL-23 stabilize the Th17 lineage, characterized by its response against extracellular pathogens [45]. The Th9 program, activated by the cytokines IL-4 and TGF-β, plays an essential role in infectious diseases, cancer, and autoimmune diseases [45]. There is another subpopulation of CD4+ T lymphocytes called TReg, induced by TGF-β, which seem very relevant for damage resolution. TReg are focused on suppressing immune system cells and preventing exacerbated reactions [44].
In murine T cells melatonin has different roles depending on the situation. Exogenous melatonin promotes both the activation of T cells and their proliferation [41,46]. This effect occurs through the reduction of IL-10, and the increased the level of the T cell activation marker CD69 [41,46]. Additionally, there is a notable increase in the abundance of proliferation markers such as Ki67 and Bcl2 [46]. The underlying mechanism might be related to MT1 receptors since its antagonists’ limit T cell proliferation [46]. Melatonin might not only control proliferation. Stimulation of ovalbumin-activated T cells with melatonin increased IL-4 levels contributing to the clonal expansion of Th2 cells in vivo. However, melatonin did not increase the production of this interleukin in naïve T cells not activated with ovalbumin, suggesting that the effect of melatonin only occurs in antigen-activated T cells [47]. In myasthenia gravis, an autoimmune disorder, melatonin decreases the Th1 and Th17 response, as well as the production of pro-inflammatory cytokines, which have benefit effects due to the base of the disease [48]. Besides, in a model of AIDS infection it has been observed that melatonin administration was associated with an increase in IL-2 and IFN-γ levels; and a decrease in Th2 cytokine production. This increase in the proliferation of T-cell as well as increase of Th1 cytokine secretion have protective effect against retrovirus infection [49].
Following cell activation, most T cells enter apoptosis. But a small population persists in the form of memory T cells (TM) [50]. These TM can differentiate into central memory T cells (TCM), effector memory T cells (TEM), memory stem T cells (TSCM), resident memory T cells (TRM), and follicular memory T cells (TFH) [50]. This classification is based on cell surface expression of homing and selectin molecules, production of effector cytokines, and location [50]. Studies conducted by Álvarez-Sánchez et al. (2015) showed that melatonin increased the population of TCM cells in the central nervous system in an Experimental Autoimmune Encephalomyelitis (EAE) model [51]. These T cells are the most frequent population of immune cells in the cerebrospinal fluid of healthy humans [51]. Melatonin also decreased the population of TEM cells, which are more characteristic in EAE patients [51]. These studies showed that melatonin reduces the level of CD44, a marker of effector cells, and contributes to the increase of TReg cells, which reduces the immune response, alleviating the symptoms of the disease[51].
Taken together, these results suggest that melatonin has a dual environment-dependent role. It acts as a negative modulator of the effector immune response in the presence of exacerbated immune responses, but as a stimulator of the immune response under conditions of immunosuppression.
3.1.2. B Cells
B cells recognize and respond specifically to each antigenic epitope without the need to interact with APCs [52]. They originate from hematopoietic stem cells in the bone marrow [52]. They mature within secondary lymphoid organs and transform into plasmacytes after activation by direct interaction between antigen and B-cell receptor (BCR) [52]. These cells also present co-receptors and co-stimulatory molecules, like T lymphocytes, which can be regulated by the BCR [52]. Plasma cells, or plasmacytes, have the capacity to synthesize antibodies (AB), both natural ABs, which are generated without external antigenic stimulation, and antigen-specific ABs produced in response to pathogens such as bacteria, viruses, fungi, and parasites [53]. There are different populations of B-cells. Memory B cells, which are long-lived and have two subpopulations depending on the origin and expression of markers. Then, regulatory B cells (BReg), which can be short-lived cells, although not necessarily, and restrict excessive inflammatory responses. These BReg cells ensure tolerance through the release of cytokines and interleukins, such as IL-10, that occur in all inflammatory reactions. This release of cytokines is exacerbated during autoimmune diseases or unresolved infections [53].
There are just few studies about the melatonin biological effects on B lymphocytes [17]. Melatonin specifically binds to the nuclear receptor RZR/RORα on human B cells and inhibits the expression of the 5-lipoxygenase gene. Which encodes for a key enzyme in the biosynthesis of leukotrienes, which are involved in inflammatory and allergic processes. This may suggest a melatonin role in these immunological reactions through B lymphocytes [17]. In vivo studies in chicken have shown that exogenous administration of melatonin stimulates the proliferation of B lymphocytes [17]. Melatonin promotes B cells proliferation also in mice models [17]. It has been suggested that melatonin may be a checkpoint regulator in early B cell development as it prevented apoptosis of bone marrow B cells during early differentiation when it was administrated orally to mice [17,54]. Studies conducted by Luo et al. (2020) suggested that the role of melatonin in B cell activation depends on concentration and timing of the administration [55]. For instance, melatonin treatment for two weeks inhibited B cells, while same treatment for four weeks caused the opposite effect [55].
3.2. Innate Immune Response.
In addition to its well-documented effects on adaptive immunity, melatonin has also been shown to modulate cells of the innate immune system. For instance, in vivo experiments by Liang et al. (2024) demonstrated that treatment of aged mice with melatonin for 1–2 weeks enhanced the proliferation of innate immune cells in the spleen, liver, and bone marrow [56].
Natural killer (NK) cells are innate immune lymphocytes whose activity is regulated by a balance of inhibitory and activating receptors [57]. Unlike T cells, NK cells do not require antigen presentation by APCs because they can recognize their target cells directly [57]. They discriminate between healthy and abnormal cells through recognition of major histocompatibility complex (MHC) class I molecules, which engage inhibitory receptors [58]. Upon activation, NK cells secrete a variety of cytokines, such as interferon-γ (IFN-γ), as well as growth factors including granulocyte–macrophage colony-stimulating factor (GM-CSF) and diverse chemokines [57]. NK cells kill target cells by releasing perforin and granzymes or through contact-dependent mechanisms mediated by Fas–FasL and TRAIL, collectively known as death-ligand pathways. In addition, NK cells mediate antibody-dependent cellular cytotoxicity (ADCC) [57,58]. Melatonin promotes NK cell degranulation and IFN-γ secretion via activation of the JAK3/STAT5 and T-bet signaling pathways [56]. Notably, these effects were absent in young mice, suggesting that melatonin’s restorative action on NK cell function is context dependent.
Mast cells are hematopoietic-derived immune cells. Under normal conditions mature mast cells do not circulate in the bloodstream. Instead, mast cell progenitors migrate into peripheral tissues, where they differentiate under the influence of stem cell factors and various cytokines [59]. These cells contain large cytoplasmic granules, like those in neutrophils, which store inflammatory mediators such as histamine and heparin [59]. Their primary mechanism of action involves the binding of immunoglobulin E (IgE) during allergic reactions. They can also recognize harmful antigens and release inflammatory mediators that contribute to the elimination of pathogen [59]. Evidence from Caiyun et al. (2023) highlights the role of melatonin in innate immunity, showing that mice with aberrant melatonin production exhibited increased mortality following H1N1 viral infection [60]. This heightened mortality was attributed to melatonin-mediated suppression of mast cell activation, which in turn reduced the inflammatory response through regulation of cytokine secretion [60].
Dendritic cells (DCs) are among the most professional antigen-presenting cell types [61]. In the conventional presentation pathway, exogenous antigens are displayed on major histocompatibility complex (MHC) class II molecules to CD4⁺ T cells, whereas endogenous antigens are presented on MHC class I molecules to CD8⁺ T cells [61]. DCs are also highly efficient at presenting exogenous antigens on MHC class I molecules through an unconventional pathway known as cross-presentation (Liu, 2015). Huang et al. (2024) reported that melatonin influences T cell differentiation by suppressing NF-κB activation in DCs, thereby indirectly shaping adaptive responses through modulation of innate immune cell activity [62].
Neutrophils are the most abundant leukocyte in humans and are essential components of the innate immune response. They can rapidly kill invading pathogens by phagocytosis [63]. Although these cells are not classic cytokine producers and do not function as APCs, they play a central role in inflammation and release a variety of granule proteins, such as calreticulin [63]. More recently, Tai et al. (2025) described an additional neuroimmune facet of melatonin’s action. In a murine model of transient focal cerebral ischemia, melatonin administration reduced neutrophil infiltration and attenuated microglial and macrophage activation in the brain [64].
Macrophages represent another type of professional antigen-presenting cell, and their principal function is phagocytosis [65]. Macrophages exhibit binary M1/M2 polarization states in response to specific stimuli, playing a central role in immune regulation [65]. Classically activated M1 macrophages, induced by stimuli such as lipopolysaccharide (LPS) and IFN-γ, adopt a predominantly aerobic glycolytic metabolism and display a tumor-suppressive profile characterized by releasing of IL-6, IL-12, TNF and iNOS [66,67]. Alternatively activated macrophages (M2) promote the resolution of inflammation, healing and phagocytosis. Their metabolism depends on oxidative phosphorylation, and they show the activation of autophagy. This latter phenotype has a pro-tumoral profile. Membrane melatonin receptors (MT1/MT2) were identified on murine peritoneal macrophages as early as 1999, and pharmacological blockade with the non-selective MT1/MT2 antagonist luzindole diminished phagocytic activity in RAW264.7 macrophages [68,69]. Melatonin modulates macrophage polarization in an environment-dependent manner via pathways such as NF-κB [70]. It negatively regulates iNOS mRNA expression through NF-κB inhibition, thereby reducing ROS generation that can impair electron-transport chain function [69]. Yet the effects are context-specific: classic studies reported that human monocytes exposed to LPS and then treated with melatonin differentiated toward a pro-inflammatory, cytotoxic macrophage phenotype, whereas monocytes pre-activated with melatonin and subsequently challenged with LPS exhibited pro-oxidant and pro-inflammatory profiles [71]. In RAW264.7 cells, danger signals that activate NF-κB, have been shown to induce melatonin synthesis, a mechanism involved in both the initiation and resolution of the innate immune response [72]. In addition, Wang et al. (2023) reported that 0.3 mM melatonin induced apoptosis in RAW264.7 cells through Bmal1 and the MAPK-p38 pathway [73]. This effect was associated with upregulation of Bmal1, reduced levels of phosphorylated MAPK-p38, and a subsequent increase in ROS [73].
4. Melatonin in Cancer: Mechanistic Insights and Therapeutic Potential
Cancer remains one of the leading causes of mortality worldwide [74]. Cancer is a multifactorial disease, a collection of disorders arising primarily from the body’s own cells, which acquire abnormal growth potential and the capacity to invade and metastasize beyond their tissue of origin [75]. These changes in cancer cells are due to genetic alterations in key cellular pathways such as DNA repair, proliferation control, and defense mechanisms [75]. The hallmark of cancer is cellular immortalization, wherein tumor cells acquire the ability to proliferate independently of external growth signals while evading apoptosis [76,77,78]. They also undergo profound metabolic reprogramming through the Warburg effect, shifting energy production from mitochondrial oxidative phosphorylation towards aerobic glycolysis in the cytosol [79].
Due to its anti-inflammatory role and its ability to modulate the immune system, melatonin has been proposed as a novel antitumor agent, targeting key events of cancer initiation, progression, and metastasis. Experimental evidence highlights the multifaceted anticancer actions of melatonin, ranging from direct modulation of tumor cell survival to TEM regulation and metabolic reprogramming. Laborda-Illanes et al. (2023) demonstrated that vascular endothelial growth factor (VEGF) and MCF-7 breast cancer cells induce angiogenic sprouting in three-dimensional spheroids of human umbilical vein endothelial cells (HUVECs) [80]. In this setting, 1 mM melatonin effectively suppressed neovascular outgrowth by inhibiting angiogenesis (Figure 2). Complementary studies by Zeppa et al. (2024) revealed that melatonin reduced the viability of pancreatic ductal adenocarcinoma cell lines PANC-1 and MiaPaCa-2 through both apoptotic and necroptotic mechanisms [81] (Figure 2). Combined treatments with antitumoral agents (cannabidiol or sorafenib) and melatonin inhibits proliferation and enhanced cell death indicators[81]. Melatonin metabolites also display anticancer potential, as they inhibited proliferation in SKMEL-188 human melanoma cells without affecting melanogenesis [82]. Pinealectomy studies highlight melatonin’s protective role. The removal of the pineal gland in rats led to larger and thicker oral squamous cell carcinomas, potentially due to loss of melatonin’s capacity to inactivate ROS-dependent Akt signaling and reduced cyclin D1, PCNA, and Bcl-2 expression [83].
Immune checkpoint regulation has emerged as a significant anticancer mechanism [84]. Programmed death-ligand 1 (PD-L1) is key for immune evasion. After PD-1 binding to this receptor on T cells, proliferation, and activation is suppressed in these immune cells [84]. Melatonin reduces PD-L1 expression in hepatocellular carcinoma through hypoxia-inducible factor-1α (HIF-1α) and MAPK-dependent pathways [85] (Figure 2). An effect replicated in a model of non-small-cell lung cancer [86] and head-and-neck squamous cell carcinoma (HNSCC) [86]. Melatonin treatment also blocked epithelial-to-mesenchymal transition (EMT) in HNSCC [87] (Figure 2). This effect, tested in vitro and in vivo, was mediated through the inhibition of ERK1/2–FOSL1 signaling, thereby enhancing CD8⁺ T-cell infiltration and potentiating antitumoral immunity [87].
Tumors are hypoxic and show an elevated HIF-1α levels. This promotes expression of pyruvate dehydrogenase kinase 1 (PDK1), which inhibits the pyruvate dehydrogenase complex (PDC) and thereby prevents pyruvate conversion to acetyl-CoA. This metabolic switch limits mitochondrial synthesis of melatonin via aralkylamine N-acetyltransferase (AANAT), fostering a Warburg-type glycolytic phenotype [88,89]. Melatonin in ovarian cancer counteracts this effect by inhibiting PDK activity, restoring pyruvate flux into acetyl-CoA, promoting mitochondrial oxidative metabolism, and reversing the Warburg effect [89] (Figure 2).
Clinical applications have been explored using melatonin as an adjuvant with traditional treatments [93]. Oral melatonin was administrated along with the radiofrequency ablation (RFA) in early-stage non-small-cell lung cancer patients with multiple ground-glass nodules [93]. This therapy yielded significantly fewer or smaller untreated nodules compared with conventional surgery [93]. The beneficial effects were mediated by a combined induction of and RFA of the NK-cell antitumor responses. This treatment reprogrammed residual tumor metabolism toward oxidative phosphorylation, and reduced hypoxia in the tumor microenvironment, consistent with Warburg effect reversal [93]. Melatonin has been also investigated for mitigating cancer-related fatigue (CRF), a common adverse effect of conventional therapies [94]. In patients with invasive breast carcinoma receiving chemotherapy and radiotherapy, 18 mg/day oral melatonin reduced CRF and improved quality of life [94]. However, other randomized controlled trials in early-stage breast cancer or ductal carcinoma in situ reported no significant benefit [95].
5. Modulation of the Tumor Immune Microenvironment
- Elimination. Innate and adaptive immune components act to eradicate emerging tumor cells.
- Equilibrium. Tumor cells with low immunogenicity survive immune pressure and continue unchecked proliferation.
- Escape. Tumor cells downregulate major histocompatibility complex class I (MHC-I) expression, impairing immune recognition and culminating in the emergence of clinically detectable tumors.
Tumor cells can evade immune surveillance by increasing the production of proteins that activate immune regulatory pathways, thereby suppressing the immune system [76,77,78]. Treatment challenges arise from both types of immune evasion (II and III) and the cellular heterogeneity of the tumor microenvironment (TME) [98,99]. The term TME was first introduced in the 19th century by Paget, who proposed that bidirectional communication between tumors and their surrounding milieu was critical to understanding relapses, metastasis, and drug resistance [100,101]. The TME contains non-malignant cells, including immune cells, cancer-associated fibroblasts (CAF), endothelial cells (EC), pericytes, and tissue-specific cell types such as adipocytes or neurons, whose composition varies depending on tumor location [98,99,101]. Once considered passive bystanders, the TME cells and their secreted factors are now recognized as active participants in tumorigenesis and as potential therapeutic targets [99,101,102]. From these TME cells, the immune infiltrate within tumors, which can be broadly categorized as tumor-antagonizing: effector T lymphocytes (CD8⁺ cytotoxic and CD4⁺ helper subsets), NK cells, dendritic cells (DC), and M1 macrophages; or tumor-promoting: TReg, tumor associated macrophages (TAMs) and myeloid-derived suppressor cells (MDSCs) [98]. Tumor cells actively shape the TME by recruiting and reprogramming host cells, remodeling the vasculature, and reorganizing the extracellular matrix (ECM). This process depends on complex heterotypic interactions between malignant and non-malignant components [99].
Within the TME, adaptive immune cells (principally T and B lymphocytes) play pivotal roles in tumor control and immune evasion. This is mainly to it is called T cell “exhaustion”. This term refers to the diminished cytokine production and increased levels of inhibitory receptors of lymphocytes, which impair their activation [103]. In solid tumors, CD8⁺ T lymphocytes mediate direct tumor cell elimination, whereas TReg represent the predominant immunosuppressive CD4⁺ subset [102]. The presence of effector CD4⁺ T cell subsets in both circulation and the TME has been linked to favorable prognosis across multiple cancer types [104,105]. CD4⁺ T cells exert dual and context-dependent effects: Th1 cells promote tumor senescence and drive tumor MHC-II expression (anti-tumoral), but they may also facilitate tumor maintenance via pro-angiogenic activity (pro-tumoral); Th2 cells can recruit cytotoxic eosinophils (anti-tumoral) but also favor the accumulation of immunosuppressive M2 macrophages (pro-tumoral) [104,105]. Melatonin increases Th1 cells in mice with tumors [106] and reduced the proliferation of TReg, which have a pro-tumoral role [107]. Furthermore, metabolic constraints within the TME further compromise T cell function [108]. Elevated lactate concentrations and extracellular acidity inhibit cytokine production by both CD8⁺ and CD4⁺ T cells, markedly reducing their cytotoxic potential [108]. B lymphocytes contribute to tumor control primarily through the production of antigen-specific antibodies, which promote antibody-dependent cellular cytotoxicity (ADCC) by NK cells [109]. In addition, direct antigen recognition via the B cell receptor (BCR) can trigger the release of proteolytic enzymes such as granzyme B (GzmB), inducing apoptosis in tumor cells [109,110].
Among innate immune effectors, NK cells recognize tumor targets independently of MHC-I expression, complementing CD8⁺ T cell surveillance [111]. NK-mediated cytotoxicity involves perforin-dependent pore formation and granzyme-induced apoptosis, as well as death receptor engagement via FasL and TRAIL [112]. They also secrete IFN-γ and TNF-α, which can suppress tumor proliferation and survival. Nevertheless, NK cells often exhibit functional impairment within the TME [102]. Tumor cells modulate ligands for NK cell receptors, which activate the Natural Killer group 2, member D (NKG2D) and NKp30. Besides, tumor cells can inhibit killer immunoglobulin-like receptors (KIR) [111,113]. It is described that melatonin amplifies lytic activity of NK cells [107]. Also, melatonin induces the release of cytokines, such as Il-6, Il-12 and IFN-γ, related to their proliferation [107].This will be useful to reduce the TME. Neutrophils, including tumor-associated neutrophils (TANs), exhibit polarized phenotypes: antitumorigenic N1 and pro-tumoral N2 subsets [102]. Their canonical antimicrobial role, phagocytosis, and ROS generation could exert anti-tumoral functions, but their ROS in the TME induces mutagenesis and supports tumor progression [114]. Karadas et al. in 2021 indicate that melatonin reduced the number of neutrophils in peripheral blood and decrease TME infiltration in mice model of breast cancer [115]. Myeloid-derived suppressor cells (MDSCs), expand in cancer patients, further suppressing T cell activity, enhancing PD-L1 expression, secreting ROS and RNS such as nitric oxide, which downregulates genes required for T cell extravasation and tumor infiltration, and promoting angiogenesis and pre-metastatic niche formation [116]. Melatonin reduces the accumulation of MDSCs by decreasing TGF-β secretion by tumor cells [117]. TAMs, Macrophages that infiltrate TME, exemplify the functional plasticity of myeloid cells [118]. However, due to macrophages relevance in the TME a whole section about the melatonin actions in the TAMs is next to this one.
In addition to these hematopoietic components, non-hematopoietic stromal cells such as cancer-associated fibroblasts (CAFs) orchestrate multiple tumor-supportive mechanisms [125]. CAFs remodel the extracellular matrix, secrete pro-angiogenic factors (VEGFA, PDGFC, CXCL12, osteopontin), and recruit endothelial and myeloid cells, thereby facilitating neovascularization [125]. They also produce TGF-β and CXCL12, fostering TReg and M2 macrophage differentiation while impairing T cell recruitment and activation. Melatonin prevents the conversion of fibroblast into CAFs by reducing the production of metalloproteinase 2 and 9 in gastric cancer cells [126]. This helps to disrupt cancer progression.
The immune system exerts a determinant influence on cancer evolution. Thus, the modulation of the immune cells through different approaches, pharmacological or genetical ones, has been proposed as therapeutic targets for cancer [1,127]. Here melatonin is a potential adjuvant for cancer prevention and therapy [128].
6. Macrophages as a Therapeutic Target of Melatonin in Cancer
Macrophages, particularly TAMs, account for half of the cells of the TEM and are key for both tumor progression and regression [118]. TAMs show different metabolic and functional states in response to microenvironmental perturbations, from pro-inflammatory, classically activated M1-like programs to anti-inflammatory, alternatively activated M2-like programs [118]. M1 effector mechanisms include ROS and RNS production with direct, albeit relatively slow, cytotoxicity toward tumor cells, and rapid antibody-dependent cellular cytotoxicity (ADCC). M1 activity is also associated with increased numbers of activated NK cells with death-receptor ligands, thereby amplifying tumor cell killing [66,67]. M2-like TAMs support tumor progression through well-documented mechanisms [67]. In different malignancies, the prevalence of M2-polarized TAMs correlates with reduced patient survival, increased metastatic dissemination, and enhanced tumor proliferation [119]–[121]. M2 phenotypes are associated with expression of proangiogenic factors (VEGF, basic fibroblast growth factor (bFGF) and others), secretion of matrix-remodeling enzymes (matrix metalloproteinases, serine proteases, cathepsins) that degrade collagens and extracellular-matrix components, promotion of endothelial proliferation and neovascularization, and facilitation of stromal and tumor cell migration [67]. M2 macrophages can also suppress CD8⁺ T cell proliferation via L-arginine metabolism [67]. Tumor-derived metabolites further modulate phenotype towards M2. For example, extracellular lactate produced by tumor metabolism promotes an M2-like activation through histone lactylation and related metabolic–epigenetic crosstalk [122,123]. Melatonin usually prevents M2 induction [124].
Given the central role of the immune system in cancer control and the capacity of melatonin to enhance immune function, the modulation of macrophage phenotype by melatonin has emerged as a promising therapeutic avenue [118]. This could be mediated by multiple mechanisms: suppression of senescence, enhancement of antigen presentation, interruption of tumor-promoting inflammatory circuits, metabolic reprogramming, and modulation of immune checkpoints [118].
A selective clearance of senescent macrophages could reverse the immunosuppressive TME in theory. This would create an immunostimulatory TME and would disrupt tumoral vascular networks. Thereby reducing nutrient and oxygen supply to cancer cells [119]. Recent mechanistic work by Zhang et al. (2025) demonstrated that melatonin-deficient macrophages secrete high levels of senescence-associated secretory phenotype (SASP) factors, driving inflammation, macrophage senescence, and immune dysfunction, which contribute to chemotherapy resistance [129]. Melatonin counteracts these effects by upregulating Trim26, which in turn blocks activation of the cyclic GMP–AMP synthase–stimulator of interferon genes (cGAS–STING) pathway, thereby attenuating macrophage senescence, promoting a protumoral response [129] (Figure 3). However, chronic activation of cGAS–STING is known to sustain pro-tumorigenic inflammation and foster an immunosuppressive TME [130]. Melatonin also enhances macrophage-mediated antigen presentation by upregulating MHC-II expression, amplifying both specific and non-specific T-cell responses [131] (Figure 3). Melatonin treatment interrupts tumor invasion and migration by reducing MIF (macrophage migration inhibitory factor) and IL-1β levels in THP-1 cells, thereby limiting the crosstalk between TAMs and cancer cells, reducing the inflammation in oral squamous cell carcinoma [132] (Figure 3).
Metabolic reprogramming of TAMs is another relevant therapy [133]. In colostrum-derived macrophages co-cultured with breast cancer cell lines, exposure to melatonin increases the M1/M2 ratio. Thus, reducing both pro-tumoral and immunosuppressive phenotypes [134]. In lymphoma models, melatonin inhibited arginase activity, leading to increased nitric oxide (NO) production and decreased urea levels within the TME, culminating in tumor cell death [133] (Figure 3). In relation to this, melatonin alters intercellular communication by modulating cancer cell–derived exosomes [135,136]. In gastric cancer, melatonin-treated macrophages displayed reduced PD-L1 levels, enhancing their antitumor activity. This effect extended to tumor cells themselves and was associated with altered exosomal microRNA content capable of suppressing PD-L1 expression [135,136] (Figure 3). In lung cancer, melatonin combined with LPS reduced M2 macrophage polarization and induced M2 apoptosis via activation of JAK–STAT, Toll-like receptor, and chemokine signaling pathways [137]. Prolonged LPS exposure, however, induces tolerance and favors M2 polarization, underscoring the importance of temporal dynamics in therapeutic strategies [137]. Inhibition of the JAK2/STAT3 pathway in M2 macrophages diminished PD-L1 production, further relieving T-cell suppression [137]. Thus, melatonin can bias macrophage behavior in divergent directions depending on prior activation state and microenvironmental cues, a property that could be harnessed to reprogram TAMs toward tumor-suppressive phenotypes.
5. Conclusions
Melatonin emerges as a potent regulator of the cancer microenvironment, which accounts for a very relevant effector of tumoral growth. We have found interesting literature on how melatonin modulates TEM cells and activates some immune cells to fight against tumoral cells, for instance, T-cells and macrophages. The latter stand up as they can account for half of the TEM. Melatonin modulation of macrophage function in cancer will offer relevant translational opportunities for combinatorial immunotherapeutic approaches.
Author Contributions
Conceptualization, E.L.F., A.S.L. and M.G.M..; writing—original draft preparation, E.L.F, C.P.M. and M.R.G.; writing—review and editing, A.S.L. and M.G.M.; supervision, A.S.L. and M.G.M. All authors have read and agreed to the published version of the manuscript.
Funding
E.L.F. has an FPI contract (PRE2023-138478) funded by the grant PID2022-138478OA-100. C.P.M. has a contract from the Plan Especial Grado Medicina (USAL), A.S.L research is funded by by the Lanzadera TCUE and C2 program (Universidad de Salamanca); and M.G.M. is a Ramón y Cajal researcher (grant RYC2021-033684-I, funded by MICIU/AEI/10.13039/501100011033 and by European Union NextGenerationEU/PRTR), and her research is funded by the PID2022-138478OA-100, funded by MICIU/AEI/10.13039/501100011033 and by FEDER, UE.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
No new data were created or analysed in this study. Data sharing is not applicable to this article.
Acknowledgments
We are members of and thank Women in Autophagy (WIA), ALBA Network, Sociedad Española de Bioquímica y Biología Molecular (SEBBM), Sociedad Española de Autofagia (SEFAGIA), Sociedad Anatómica Española (SAE), Asociación Española de Investigación sobre el Cáncer (ASEICA), and European Society of Cardiology (ESC).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AANAT | Aralkylamine N-Acetyltransferase |
| ABST | 2,2′-Azino-Bis(3-Ethylbenzthiazoline-6-Sulfonic Acid) |
| ADCC | Antibody-Dependent Cellular Cytotoxicity |
| AFMK | N-Acetyl-N-Formyl-5-Methoxyquinuramine |
| AMK | N-Acetyl-5-Methoxyquinuramine |
| Arg1 | Arginase 1 |
| ATP | Adenosine Triphosphate |
| Bcl-2 | B-Cell Lymphoma 2 |
| BCR | B Cell Receptor |
| bFGF | Basic Fibroblast Growth Factor |
| BMAL1 | Brain And Muscle ARNT-Like Protein 1 |
| BReg | Regulatory B Cells |
| CAFs | Cancer-Associated Fibroblasts |
| CAT | Catalase |
| cGAMP | Cyclic GMP–AMP |
| cGAS | Cyclic GMP–AMP Synthase |
| CD8 | Cluster Of Differentiation 8 (Cytotoxic T Lymphocyte Marker) |
| COX-2 | Cyclooxygenase-2 |
| CRF | Cancer-Related Fatigue |
| CXCL12 | C-X-C Motif Chemokine Ligand 12 |
| Cyt C | Cytochrome C |
| DC | Dendritic Cells |
| EAE | Experimental Autoimmune Encephalomyelitis |
| EC | Endothelial Cells |
| ECM | Extracellular Matrix |
| EMT | Epithelial–Mesenchymal Transition |
| ERK1/2–FOSL1 | Extracellular Signal-Regulated Kinases 1 And 2 – Fos-Like Antigen 1 |
| FasL | Fas Ligand (CD95 Ligand) |
| GLUT | Glucose Transporter |
| GLUT1 | Glucose Transporter 1 |
| GM-CSF | Granulocyte–Macrophage Colony-Stimulating Factor |
| GPx | Glutathione Peroxidase |
| GR | Glutathione Reductase |
| GSH | Reduced Glutathione |
| GSSG | Oxidized Glutathione |
| GTP | Guanosine Triphosphate |
| GzmB | Granzyme B |
| HIF | Hypoxia-Inducible Factor |
| HNSCC | Head And Neck Squamous Cell Carcinoma |
| HUVECs | Human Umbilical Vein Endothelial Cells |
| iNOS | Inducible Nitric Oxide Synthase |
| IL-1β | Interleukin-1 Beta |
| JAK-STAT | Janus Kinase And Signal Transducer And Activator Of Transcription |
| Ki67 | Proliferation Marker Protein Ki-67 |
| KIR | Killer Immunoglobulin-Like Receptors |
| MAPK | Mitogen-Activated Protein Kinase |
| MCF-7 | Michigan Cancer Foundation-7 |
| MDSCs | Myeloid-Derived Suppressor Cells |
| MHC-I/II | Major Histocompatibility Complex Class I/II |
| MiaPaCa-2 | Human Pancreatic Cancer |
| MIF | Macrophage Migration Inhibitory Factor |
| MT1R | Melatonin Receptor Type 1 |
| NKG2D | Natural Killer Group 2, Member D |
| NK | Natural Killer |
| NKp30 | Natural Killer Cell P30-Related Protein (NCR3) |
| NLRP3 | NOD-Like Receptor Family Pyrin Domain-Containing 3 |
| ·NO | Nitric Oxide Radical |
| ·NO/NO | Nitric Oxide |
| O2- | Superoxide Anions |
| ·OH | Hydroxyl Radicals |
| ONOO- | Peroxynitrite Anions |
| PANC-1 | Human Pancreatic Carcinoma Cell Line |
| PCNA | Proliferating Cell Nuclear Antigen |
| PDC | Pyruvate Dehydrogenase Complex |
| PD-1 | Programmed Cell Death Protein 1 |
| PDCA | Programmed Cell Death Activator |
| PDGFC | Platelet Derived Growth Factor C |
| PD-L1 | Programmed Death-Ligand 1 |
| PD-L1/2 | Programmed Death-Ligand 1/2 |
| PEPT1/2 | Peptide Transporter 1/2 |
| RFA | Radiofrequency Ablation |
| RNS | Reactive Nitrogen Species |
| ROR-α | Retinoic Acid-Related Orphan Receptor Alpha |
| ROS | Reactive Oxygen Species |
| RZR-β | Orphan Receptor Retinoid Z Receptor Beta |
| SAPS | Serum Amyloid P Component |
| SIRT3 | Stimulates Sirtuin 3 |
| siRNA | Small Interfering RNA |
| SOD | Superoxide Dismutase |
| STRING | Search Tool For The Retrieval Of Interacting Genes/Proteins |
| TAM | Tumor-Associated Macrophage |
| TANs | Tumor-Associated Neutrophils |
| TCA Cycle | Tricarboxylic Acid Cycle (Krebs Cycle) |
| TCR | T-Cell Receptor |
| TGF-β | Transforming Growth Factor Beta |
| THP-1 Cells | Human Monocytic Cell Line Derived From A Patient With Acute Monocytic Leukemia |
| TNF-α | Tumor Necrosis Factor Alpha |
| TRAIL | TNF-Related Apoptosis-Inducing Ligand |
| TRIM26 | Tripartite Motif-Containing Protein 26 |
| Treg | Regulatory T Cells |
| TSCM | Memory Stem T Cells |
| TFH | Follicular Memory T Cells |
| TEM | Tumor Microenvironment |
| TEM | Effector Memory T Cells |
| TM | Memory T Cells |
| TRM | Resident Memory T Cells |
| UCP | Uncoupling Proteins |
| VEGF | Vascular Endothelial Growth Factor |
| VEGFA | Vascular Endothelial Growth Factor A |
| Δψ | Membrane Potential |
| ¹O₂ | Singlet Oxygen |
References
- Minich, D. M.; Henning, M.; Darley, C.; Fahoum, M.; Schuler, C. B.; Frame, J. Is Melatonin the “Next Vitamin D”?: A Review of Emerging Science, Clinical Uses, Safety, and Dietary Supplements. Nutrients. MDPI October 2022. [CrossRef] [PubMed]
- Lerner, A. B.; Case, J. D.; Takahashi, Y.; Lee, T. H.; Mori, W. ISOLATION OF MELATONIN, THE PINEAL GLAND FACTOR THAT LIGHTENS MELANOCYTES1. J Am Chem Soc 2002. 80 (10), 2587. [Google Scholar] [CrossRef]
- LERNER, A. B.; CASE, J. D.; TAKAHASHI, Y. Isolation of Melatonin and 5-Methoxyindole-3-Acetic Acid from Bovine Pineal Glands. Journal of Biological Chemistry 1960, 235 (7), 1992–1997. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A. B.; Case, J. D.; Mori, W.; Wright, M. R. Melatonin in Peripheral Nerve. Nature 1959, 183 (4678). [Google Scholar] [CrossRef]
- Tan, D. X.; Manchester, L. C.; Esteban-Zubero, E.; Zhou, Z.; Reiter, R. J. Melatonin as a Potent and Inducible Endogenous Antioxidant: Synthesis and Metabolism. Molecules, MDPI AG October 2015, pp 18886–18906. [CrossRef]
- Reiter, R. J.; Mayo, J. C.; Tan, D. X.; Sainz, R. M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an Antioxidant: Under Promises but over Delivers. J Pineal Res 2016, 61, 253–278. [Google Scholar] [CrossRef]
- Reiter, R. J.; Sharma, R. N.; Chuffa, L. G. de A.; Silva, D. G. H. da; Rosales-Corral, S. Intrinsically Synthesized Melatonin in Mitochondria and Factors Controlling Its Production. Histology and Histopathology, Histology and Histopathology March 2025, pp 271–282. [CrossRef]
- Arnao, M. B.; Giraldo-Acosta, M.; Castejón-Castillejo, A.; Losada-Lorán, M.; Sánchez-Herrerías, P.; El Mihyaoui, A.; Cano, A.; Hernández-Ruiz, J. Melatonin from Microorganisms, Algae, and Plants as Possible Alternatives to Synthetic Melatonin. Metabolites 2023, 13. [Google Scholar] [CrossRef]
- Lee, K.; Choi, G. H.; Back, K. Functional Characterization of Serotonin N-Acetyltransferase in Archaeon Thermoplasma Volcanium. Antioxidants 2022, 11(3), 596. [Google Scholar] [CrossRef]
- Zimmermann, P.; Kurth, S.; Pugin, B.; Bokulich, N. A. Microbial Melatonin Metabolism in the Human Intestine as a Therapeutic Target for Dysbiosis and Rhythm Disorders. NPJ Biofilms Microbiomes 2024, 10, 1–17. [Google Scholar] [CrossRef]
- Tordjman, S.; Chokron, S.; Delorme, R.; Charrier, A.; Bellissant, E.; Jaafari, N.; Fougerou, C. Melatonin: Pharmacology, Functions and Therapeutic Benefits. Curr Neuropharmacol 2017, 15, 434–443. [Google Scholar] [CrossRef]
- Zhao, D.; Yu, Y.; Shen, Y.; Liu, Q.; Zhao, Z.; Sharma, R.; Reiter, R. J. Melatonin Synthesis and Function: Evolutionary History in Animals and Plants. Front Endocrinol (Lausanne) 2019, 10, 249. [Google Scholar] [CrossRef]
- Pieri, C.; Marra, M.; Moroni, F.; Recchioni, R.; Marcheselli, F. Melatonin: A Peroxyl Radical Scavenger More Effective than Vitamin E. Life Sci 1994, 55. [Google Scholar] [CrossRef]
- Reiter, R. J.; Carneiro, R. C.; Oh, C. S. Melatonin in Relation to Cellular Antioxidative Defense Mechanisms. Hormone and Metabolic Research, Georg Thieme Verlag 1997, pp 363–372. [CrossRef]
- Poeggeler, B.; Thuermann, S.; Dose, A.; Schoenke, M.; Burkhardt, S.; Hardeland, R. Melatonin’s Unique Radical Scavenging Properties – Roles of Its Functional Substituents as Revealed by a Comparison with Its Structural Analogs. J Pineal Res 2002, 33, 20–30. [Google Scholar] [CrossRef]
- Ahmad, S. B.; Ali, A.; Bilal, M.; Rashid, S. M.; Wani, A. B.; Bhat, R. R.; Rehman, M. U. Melatonin and Health: Insights of Melatonin Action, Biological Functions, and Associated Disorders. Cellular and Molecular Neurobiology, Springer August 2023, pp 2437–2458. [CrossRef]
- Calvo, J. R.; Maldonado, M. D. Immunoregulatory Properties of Melatonin in the Humoral Immune System: A Narrative Review. Immunol Lett 2024, 269, 106901. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, V. S.; Shukla, M. R.; Sherif, S. M.; Murch, S. J.; Saxena, P. K. Role of Melatonin in Alleviating Cold Stress in Arabidopsis Thaliana. J Pineal Res 2014, 56, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Chitimus, D. M.; Popescu, M. R.; Voiculescu, S. E.; Panaitescu, A. M.; Pavel, B.; Zagrean, L.; Zagrean, A. M. Melatonin’s Impact on Antioxidative and Anti-Inflammatory Reprogramming in Homeostasis and Disease. Biomolecules 2020, 10, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Liu, J.; Liu, X.; Xin, Y.; Hou, Y.; Xiang, X.; Deng, Y.; Yang, B.; Yu, W. Melatonin-Derived Carbon Dots with Free Radical Scavenging Property for Effective Periodontitis Treatment via the Nrf2/HO-1 Pathway. ACS Nano 2024, 18, 8307–8324. [Google Scholar] [CrossRef]
- Tan, D.; Reiter, R.; Manchester, L.; Yan, M.; El-Sawi, M.; Sainz, R.; Mayo, J.; Kohen, R.; Allegra, M.; Hardelan, R. Chemical and Physical Properties and Potential Mechanisms: Melatonin as a Broad Spectrum Antioxidant and Free Radical Scavenger. Curr Top Med Chem 2005, 2, 181–197. [Google Scholar] [CrossRef]
- Beyer, C. E.; Steketee, J. D.; Saphier, D. Antioxidant Properties of Melatonin—an Emerging Mystery. Biochem Pharmacol 1998, 56, 1265–1272. [Google Scholar] [CrossRef]
- Bocheva, G.; Bakalov, D.; Iliev, P.; Tafradjiiska-Hadjiolova, R. The Vital Role of Melatonin and Its Metabolites in the Neuroprotection and Retardation of Brain Aging. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Sharafati-Chaleshtori, R.; Shirzad, H.; Rafieian-Kopaei, M.; Soltani, A. Melatonin and Human Mitochondrial Diseases. J Res Med Sci 2017, 22, 2. [Google Scholar] [CrossRef]
- Urata, Y.; Honma, S.; Goto, S.; Todoroki, S.; Iida, T.; Cho, S.; Honma, K.; Kondo, T. Melatonin Induces γ-Glutamylcysteine Synthetase Mediated by Activator Protein-1 in Human Vascular Endothelial Cells. Free Radic Biol Med. 1999, 27 (7–8), 838–847. [CrossRef]
- Reiter, R. J.; Rosales-Corral, S.; Tan, D. X.; Jou, M. J.; Galano, A.; Xu, B. Melatonin as a Mitochondria-Targeted Antioxidant: One of Evolution’s Best Ideas. Cellular and Molecular Life Sciences, Birkhauser Verlag AG November 2017, pp 3863–3881. [CrossRef]
- Suofu, Y.; Li, W.; Jean-Alphonse, F. G.; Jia, J.; Khattar, N. K.; Li, J.; Baranov, S. V.; Leronni, D.; Mihalik, A. C.; He, Y.; Cecon, E.; Wehbi, V. L.; Kim, J. H.; Heath, B. E.; Baranova, O. V.; Wang, X.; Gable, M. J.; Kretz, E. S.; Di Benedetto, G.; Lezon, T. R.; Ferrando, L. M.; Larkin, T. M.; Sullivan, M.; Yablonska, S.; Wang, J.; Minnigh, M. B.; Guillaumet, G.; Suzenet, F.; Richardson, R. M.; Poloyac, S. M.; Stolz, D. B.; Jockers, R.; Witt-Enderby, P. A.; Carlisle, D. L.; Vilardaga, J. P.; Friedlander, R. M. Dual Role of Mitochondria in Producing Melatonin and Driving GPCR Signaling to Block Cytochrome c Release. Proc Natl Acad Sci U S A 2017, 114, E7997–E8006. [Google Scholar] [CrossRef]
- Tan, D. X.; Manchester, L. C.; Qin, L.; Reiter, R. J. Melatonin: A Mitochondrial Targeting Molecule Involving Mitochondrial Protection and Dynamics. International Journal of Molecular Sciences. MDPI AG December 2016. [CrossRef]
- Galano, A.; Medina, M. E.; Tan, D. X.; Reiter, R. J. Melatonin and Its Metabolites as Copper Chelating Agents and Their Role in Inhibiting Oxidative Stress: A Physicochemical Analysis. J Pineal Res 2015, 58, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Tan, D. X.; Reiter, R. J. On the Free Radical Scavenging Activities of Melatonin’s Metabolites, AFMK and AMK. J Pineal Res 2013, 54, 245–257. [Google Scholar] [CrossRef]
- Zhang, H. M.; Zhang, Y. Melatonin: A Well-Documented Antioxidant with Conditional pro-Oxidant Actions. Journal of Pineal Research, 2014, pp 131–146. [CrossRef]
- POEGGELER, B.; SAARELA, S.; REITER, R. J.; TAN, D. -X; CHEN, L. -D; MANCHESTER, L. C.; BARLOW-WALDEN, L. R. Melatonin--a Highly Potent Endogenous Radical Scavenger and Electron Donor: New Aspects of the Oxidation Chemistry of This Indole Accessed in Vitro. Ann N Y Acad Sci 1994, 738, 419–420. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, K. K. A. C.; Shiroma, M. E.; Damous, L. L.; Simões, M. de J.; Simões, R. dos S.; Cipolla-Neto, J.; Baracat, E. C.; Soares-Jr, J. M. Antioxidant Actions of Melatonin: A Systematic Review of Animal Studies. Antioxidants 2024, Vol. 13, Page 439 2024, 13. (4), 439. [Google Scholar] [CrossRef]
- Singh, C. K.; Chhabra, G.; Ndiaye, M. A.; Garcia-Peterson, L. M.; MacK, N. J.; Ahmad, N. The Role of Sirtuins in Antioxidant and Redox Signaling. Antioxidants and Redox Signaling, Mary Ann Liebert Inc. March 2018, pp 643–661. [CrossRef]
- Urata, Y.; Honma, S.; Goto, S.; Todoroki, S.; Iida, T.; Cho, S.; Honma, K.; Kondo, T. Melatonin Induces γ-Glutamylcysteine Synthetase Mediated by Activator Protein-1 in Human Vascular Endothelial Cells. Free Radic Biol Med 1999, 27, (7–8). [Google Scholar] [CrossRef] [PubMed]
- Okatani, Y.; Wakatsuki, A.; Reiter, R. J.; Enzan, H.; Miyahara, Y. Protective Effect of Melatonin against Mitochondrial Injury Induced by Ischemia and Reperfusion of Rat Liver. Eur J Pharmacol 2003, 469, 145–152. [Google Scholar] [CrossRef]
- Abbott, M.; Ustoyev, Y. Cancer and the Immune System: The History and Background of Immunotherapy. Semin Oncol Nurs 2019, 35. (5), 150923. [Google Scholar] [CrossRef]
- Hillion, S.; Arleevskaya, M. I.; Blanco, P.; Bordron, A.; Brooks, W. H.; Cesbron, J. Y.; Kaveri, S.; Vivier, E.; Renaudineau, Y. The Innate Part of the Adaptive Immune System. Clinical Reviews in Allergy and Immunology, Springer April 2020, pp 151–154. [CrossRef]
- Ruterbusch, M.; Pruner, K. B.; Shehata, L.; Pepper, M. In Vivo CD4+ T Cell Differentiation and Function: Revisiting the Th1/Th2 Paradigm. Annual Review of Immunology, Annual Reviews Inc. April 2020, pp 705–725. [CrossRef]
- Zefferino, R.; Di Gioia, S.; Conese, M. Molecular Links between Endocrine, Nervous and Immune System during Chronic Stress. Brain and Behavior. John Wiley and Sons Ltd February 2021. [CrossRef] [PubMed]
- Kühlwein, E.; Irwin, M. Melatonin Modulation of Lymphocyte Proliferation and Th1/Th2 Cytokine Expression. J Neuroimmunol 2001, 117, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Maestroni, G. J. M.; Conti, A.; Pierpaoli, W. Role of the Pineal Gland in Immunity. Circadian Synthesis and Release of Melatonin Modulates the Antibody Response and Antagonizes the Immunosuppressive Effect of Corticosterone. J Neuroimmunol 1986, 13, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Stelzer, I. A.; Arck, P. C. Immunity and the Endocrine System. In Encyclopedia of Immunobiology; Elsevier Inc., 2016; Vol. 5, pp 73–85. [CrossRef]
- Sun, L.; Su, Y.; Jiao, A.; Wang, X.; Zhang, B. T Cells in Health and Disease. Signal Transduction and Targeted Therapy. Springer Nature December 2023. [CrossRef]
- Carrillo-Vico, A.; Lardone, P. J.; Álvarez-Śnchez, N.; Rodrĩguez-Rodrĩguez, A.; Guerrero, J. M. Melatonin: Buffering the Immune System. Int J Mol Sci 2013, 14. (4), 8638. [Google Scholar] [CrossRef]
- Ren, W.; Liu, G.; Chen, S.; Yin, J.; Wang, J.; Tan, B.; Wu, G.; Bazer, F. W.; Peng, Y.; Li, T.; Reiter, R. J.; Yin, Y. Melatonin Signaling in T Cells: Functions and Applications. J Pineal Res 2017, 62. (3), e12394. [Google Scholar] [CrossRef]
- Shaji, A. V.; Kulkarni, S. K.; Agrewala, J. N. Regulation of Secretion of IL-4 and IgG1 Isotype by Melatonin-Stimulated Ovalbumin-Specific T Cells. Clin Exp Immunol 1998, 111, 181–185. [Google Scholar] [CrossRef]
- Chang, T.; Niu, C.; Sun, C.; Ma, Y.; Guo, R.; Ruan, Z.; Gao, Y.; Lu, X.; Li, H.; Lin, Y.; Lin, J.; Li, Z. Melatonin Exerts Immunoregulatory Effects by Balancing Peripheral Effector and Regulatory T Helper Cells in Myasthenia Gravis. Aging (Albany NY) 2020, 12. (21), 21147. [Google Scholar] [CrossRef]
- Zhang, Z.; Araghi-Niknam, M.; Liang, B.; Inserra, P.; Ardestani, S. K.; Jiang, S.; Chow, S.; Watson, R. R. Prevention of Immune Dysfunction and Vitamin E Loss by Dehydroepiandrosterone and Melatonin Supplementation during Murine Retrovirus Infection. Immunology 1999, 96, 291–297. [Google Scholar] [CrossRef]
- Künzli, M.; Masopust, D. CD4+ T Cell Memory. Nat Immunol 2023, 24. (6), 903. [Google Scholar] [CrossRef]
- Álvarez-Sánchez, N.; Cruz-Chamorro, I.; López-González, A.; Utrilla, J. C.; Fernández-Santos, J. M.; Martínez-López, A.; Lardone, P. J.; Guerrero, J. M.; Carrillo-Vico, A. Melatonin Controls Experimental Autoimmune Encephalomyelitis by Altering the T Effector/Regulatory Balance. Brain Behav Immun 2015, 50, 101–114. [Google Scholar] [CrossRef]
- Cano, R. L. E.; Lopera, H. D. E. Introduction to T and B Lymphocytes. 2013.
- Wang, Y.; Liu, J.; Burrows, P. D.; Wang, J. Y. B Cell Development and Maturation. In Advances in Experimental Medicine and Biology; Springer, 2020; Vol. 1254, pp 1–22. [CrossRef]
- Yu, Q.; Miller, S. C.; Osmond, D. G. Melatonin Inhibits Apoptosis during Early B-Cell Development in Mouse Bone Marrow. J Pineal Res 2000, 29, 86–93. [Google Scholar] [CrossRef]
- Luo, J.; Zhang, Z.; Sun, H.; Song, J.; Chen, X.; Huang, J.; Lin, X.; Zhou, R. Effect of Melatonin on T/B Cell Activation and Immune Regulation in Pinealectomy Mice. Life Sci 2020, 242, 117191. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Song, R.; Zhang, J.; Yao, J.; Guan, Z.; Zeng, X. Melatonin Enhances NK Cell Function in Aged Mice by Increasing T-Bet Expression via the JAK3-STAT5 Signaling Pathway. Immunity and Ageing 2024, 21. [Google Scholar] [CrossRef]
- Mace, E. M. Human Natural Killer Cells: Form, Function, and Development. J Allergy Clin Immunol 2022, 151. [Google Scholar] [CrossRef] [PubMed]
- Cantoni, C.; Falco, M.; Vitale, M.; Pietra, G.; Munari, E.; Pende, D.; Mingari, M. C.; Sivori, S.; Moretta, L. Human NK Cells and Cancer. Oncoimmunology 2024, 13. (1), 2378520. [Google Scholar] [CrossRef]
- Krystel-Whittemore, M.; Dileepan, K. N.; Wood, J. G. Mast Cell: A Multi-Functional Master Cell. Front Immunol 2016, 6. (JAN), 165675. [Google Scholar] [CrossRef] [PubMed]
- Huo, C.; Tang, Y.; Li, X.; Han, D.; Gu, Q.; Su, R.; Liu, Y.; Reiter, R. J.; Liu, G.; Hu, Y.; Yang, H. Melatonin Alleviates Lung Injury in H1N1-Infected Mice by Mast Cell Inactivation and Cytokine Storm Suppression. PLoS Pathog 2023, 19. [Google Scholar] [CrossRef]
- Liu, K. Dendritic Cells. Encyclopedia of Cell Biology 2015, 3, 741. [Google Scholar] [CrossRef]
- Huang, S. H.; Hong, Z. J.; Chen, M. F.; Tsai, M. W.; Chen, S. J.; Cheng, C. P.; Sytwu, H. K.; Lin, G. J. Melatonin Inhibits the Formation of Chemically Induced Experimental Encapsulating Peritoneal Sclerosis through Modulation of T Cell Differentiation by Suppressing of NF-ΚB Activation in Dendritic Cells. Int Immunopharmacol 2024, 126, 111300. [Google Scholar] [CrossRef]
- Rungelrath, V.; Kobayashi, S. D.; DeLeo, F. R. Neutrophils in Innate Immunity and Systems Biology-Level Approaches: An Update. Wiley Interdiscip Rev Syst Biol Med 2019, 12. (1), e1458. [Google Scholar] [CrossRef]
- Tai, S. H.; Hsu, H. H.; Huang, S. Y.; Chen, Y. N.; Chen, L. Y.; Lee, A. H.; Lee, A. C.; Lee, E. J. Melatonin Promotes B-Cell Maturation and Attenuates Post-Ischemic Immunodeficiency in a Murine Model of Transient Focal Cerebral Ischemia. J Neuroimmune Pharmacol 2025, 20. [Google Scholar] [CrossRef]
- Chen, S.; Saeed, A. F. U. H.; Liu, Q.; Jiang, Q.; Xu, H.; Xiao, G. G.; Rao, L.; Duo, Y. Macrophages in Immunoregulation and Therapeutics. Signal Transduct Target Ther 2023, 8, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Yunna, C.; Mengru, H.; Lei, W.; Weidong, C. Macrophage M1/M2 Polarization. European Journal of Pharmacology. Elsevier B.V. June 2020. [CrossRef]
- Pan, Y.; Yu, Y.; Wang, X.; Zhang, T. Tumor-Associated Macrophages in Tumor Immunity. Front Immunol 2020, 11, 583084. [Google Scholar] [CrossRef] [PubMed]
- García-Pergañeda, A.; Guerrero, J. M.; Rafii-El-Idrissi, M.; Paz Romero, M.; Pozo, D.; Calvo, J. R. Characterization of Membrane Melatonin Receptor in Mouse Peritoneal Macrophages: Inhibition of Adenylyl Cyclase by a Pertussis Toxin-Sensitive G Protein. J Neuroimmunol 1999, 95, 85–94. [Google Scholar] [CrossRef]
- Xia, Y.; Chen, S.; Zeng, S.; Zhao, Y.; Zhu, C.; Deng, B.; Zhu, G.; Yin, Y.; Wang, W.; Hardeland, R.; Ren, W. Melatonin in Macrophage Biology: Current Understanding and Future Perspectives. J Pineal Res 2019, 66. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, C.; Choi, W. S. Use of Melatonin in Cancer Treatment: Where Are We? MDPI April. International Journal of Molecular Sciences. MDPI April 2022. [CrossRef]
- Morrey, K. M.; McLachlan, J. A.; Serkin, C. D.; Bakouche, O. Activation of Human Monocytes by the Pineal Hormone Melatonin. J Immunol 1994, 153, 2671–2680. [Google Scholar] [CrossRef]
- Muxel, S. M.; Pires-Lapa, M. A.; Monteiro, A. W. A.; Cecon, E.; Tamura, E. K.; Floeter-Winter, L. M.; Markus, R. P. NF-ΚB Drives the Synthesis of Melatonin in RAW 264.7 Macrophages by Inducing the Transcription of the Arylalkylamine-N-Acetyltransferase (AA-NAT) Gene. PLoS One 2012, 7. (12), e52010. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, W.; Pan, K.; Tao, L.; Zhu, Y. Melatonin Induces RAW264.7 Cell Apoptosis via the BMAL1/ROS/MAPK-P38 Pathway to Improve Postmenopausal Osteoporosis. Bone Joint Res 2023, 12, 677–690. [Google Scholar] [CrossRef]
- Cancer Today. https://gco.iarc.who.int/today/en/dataviz/bars?types=0_1&mode=population (accessed 2025-09-30).
- Schwartz, S. M. Epidemiology of Cancer. Clin Chem 2024, 70, 140–149. [Google Scholar] [CrossRef]
- Hassanpour, S. H.; Dehghani, M. Review of Cancer from Perspective of Molecular. Journal of Cancer Research and Practice 2017, 4, 127–129. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R. A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Vaupel, P.; Multhoff, G.; Bennet, L.; Chan, J.; Vaupel, P.; Multhoff, G.; Physiol, J. Revisiting the Warburg Effect: Historical Dogma versus Current Understanding. J Physiol 2021, 599, 1745–1757. [Google Scholar] [CrossRef] [PubMed]
- Laborda-Illanes, A.; Sánchez-Alcoholado, L.; Castellano-Castillo, D.; Boutriq, S.; Plaza-Andrades, I.; Aranega-Martín, L.; Peralta-Linero, J.; Alba, E.; González-González, A.; Queipo-Ortuño, M. I. Development of in Vitro and in Vivo Tools to Evaluate the Antiangiogenic Potential of Melatonin to Neutralize the Angiogenic Effects of VEGF and Breast Cancer Cells: CAM Assay and 3D Endothelial Cell Spheroids. Biomedicine & Pharmacotherapy 2023, 157, 114041. [Google Scholar] [CrossRef]
- Zeppa, L.; Aguzzi, C.; Morelli, M. B.; Marinelli, O.; Amantini, C.; Giangrossi, M.; Santoni, G.; Fanelli, A.; Luongo, M.; Nabissi, M. In Vitro and In Vivo Effects of Melatonin-Containing Combinations in Human Pancreatic Ductal Adenocarcinoma. J Pineal Res 2024, 76. [Google Scholar] [CrossRef]
- Kim, T. K.; Lin, Z.; Tidwell, W. J.; Li, W.; Slominski, A. T. Melatonin and Its Metabolites Accumulate in the Human Epidermis in Vivo and Inhibit Proliferation and Tyrosinase Activity in Epidermal Melanocytes in Vitro. Mol Cell Endocrinol 2014, 404, 1. [Google Scholar] [CrossRef]
- Kayahara, G. M.; Valente, V. B.; Pereira, R. B.; Lopes, F. Y. K.; Crivelini, M. M.; Miyahara, G. I.; Biasoli, É. R.; Oliveira, S. H. P.; Bernabé, D. G. Pineal Gland Protects against Chemically Induced Oral Carcinogenesis and Inhibits Tumor Progression in Rats. Oncotarget 2020, 11, 1816–1831. [Google Scholar] [CrossRef]
- Gou, Q.; Dong, C.; Xu, H.; Khan, B.; Jin, J.; Liu, Q.; Shi, J.; Hou, Y. PD-L1 Degradation Pathway and Immunotherapy for Cancer. Cell Death Dis 2020, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Rao, P. G.; Liao, B. Z.; Luo, X.; Yang, W. W.; Lei, X. H.; Ye, J. M. Melatonin Suppresses PD-L1 Expression and Exerts Antitumor Activity in Hepatocellular Carcinoma. Sci Rep 2025, 15. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y. C.; Lee, K. Y.; Wu, S. M.; Kuo, D. Y.; Shueng, P. W.; Lin, C. W. Melatonin Downregulates Pd-L1 Expression and Modulates Tumor Immunity in Kras-Mutant Non-Small Cell Lung Cancer. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Chen, Y.; Tang, H.; Wang, H.; Jiang, E.; Shao, Z.; Liu, K.; Zhou, X.; Shang, Z. Melatonin Inhibits EMT and PD-L1 Expression through the ERK1/2/FOSL1 Pathway and Regulates Anti-Tumor Immunity in HNSCC. Cancer Sci 2022, 113, 2232–2245. [Google Scholar] [CrossRef]
- Wei, X.; Hou, Y.; Long, M.; Jiang, L.; Du, Y. Molecular Mechanisms Underlying the Role of Hypoxia-Inducible Factor-1 α in Metabolic Reprogramming in Renal Fibrosis. Front Endocrinol (Lausanne) 2022, 13. [Google Scholar] [CrossRef]
- Cucielo, M. S.; Cesário, R. C.; Silveira, H. S.; Gaiotte, L. B.; Dos Santos, S. A. A.; de Campos Zuccari, D. A. P.; Seiva, F. R. F.; Reiter, R. J.; de Almeida Chuffa, L. G. Melatonin Reverses the Warburg-Type Metabolism and Reduces Mitochondrial Membrane Potential of Ovarian Cancer Cells Independent of MT1 Receptor Activation. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Cucielo, M. S.; Freire, P. P.; Emílio-Silva, M. T.; Romagnoli, G. G.; Carvalho, R. F.; Kaneno, R.; Hiruma-Lima, C. A.; Delella, F. K.; Reiter, R. J.; Chuffa, L. G. de A. Melatonin Enhances Cell Death and Suppresses the Metastatic Capacity of Ovarian Cancer Cells by Attenuating the Signaling of Multiple Kinases. Pathol Res Pract 2023, 248, 154637. [Google Scholar] [CrossRef]
- Cardenas-Romero, S.; Saderi, N.; Ramirez-Plascencia, O. D.; Baez-Ruiz, A.; Flores-Sandoval, O.; Briones, C. E.; Salgado-Delgado, R. C. Melatonin Prevents Tumor Growth: The Role of Genes Controlling the Circadian Clock, the Cell Cycle, and Angiogenesis. J Pineal Res 2025, 77. (4), e70064. [Google Scholar] [CrossRef]
- Ozturk, I.; Elbe, H.; Bicer, Y.; Karayakali, M.; Onal, M. O.; Altinoz, E. Therapeutic Role of Melatonin on Acrylamide-Induced Hepatotoxicity in Pinealectomized Rats: Effects on Oxidative Stress, NF-ΚB Signaling Pathway, and Hepatocellular Proliferation. Food and Chemical Toxicology 2023, 174, 113658. [Google Scholar] [CrossRef]
- Li, M.; Hao, B.; Zhang, M.; Reiter, R. J.; Lin, S.; Zheng, T.; Chen, X.; Ren, Y.; Yue, L.; Abay, B.; Chen, G.; Xu, X.; Shi, Y.; Fan, L. Melatonin Enhances Radiofrequency-Induced NK Antitumor Immunity, Causing Cancer Metabolism Reprogramming and Inhibition of Multiple Pulmonary Tumor Development. Signal Transduct Target Ther 2021, 6. (1), 330. [Google Scholar] [CrossRef]
- Sedighi Pashaki, A.; Sheida, F.; Moaddab Shoar, L.; Hashem, T.; Fazilat-Panah, D.; Nemati Motehaver, A.; Ghanbari Motlagh, A.; Nikzad, S.; Bakhtiari, M.; Tapak, L.; Keshtpour Amlashi, Z.; Javadinia, S. A.; Keshtpour Amlashi, Z. A Randomized, Controlled, Parallel-Group, Trial on the Long-Term Effects of Melatonin on Fatigue Associated With Breast Cancer and Its Adjuvant Treatments. Integr Cancer Ther 2023, 22. [Google Scholar] [CrossRef]
- Mukhopadhyay, N. D.; Khorasanchi, A.; Pandey, S.; Nemani, S.; Parker, G.; Deng, X.; Arthur, D. W.; Urdaneta, A.; Fabbro, E. Del. Melatonin Supplementation for Cancer-Related Fatigue in Patients With Early Stage Breast Cancer Receiving Radiotherapy: A Double-Blind Placebo-Controlled Trial. Oncologist 2024, 29, E206–E212. [Google Scholar] [CrossRef] [PubMed]
- Mittal, D.; Gubin, M. M.; Schreiber, R. D.; Smyth, M. J. New Insights into Cancer Immunoediting and Its Three Component Phases — Elimination, Equilibrium and Escape. Curr Opin Immunol 2014, 27. (1), 16. [Google Scholar] [CrossRef] [PubMed]
- Dunn, G. P.; Old, L. J.; Schreiber, R. D. The Immunobiology of Cancer Immunosurveillance and Immunoediting. Immunity 2004, 21, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Lei, Y.; Li, J.-K.; Du, W.-X.; Li, R.-G.; Yang, J.; Li, J.; Li, F.; Tan, H.-B. Immune Cells within the Tumor Microenvironment: Biological Functions and Roles in Cancer Immunotherapy. Cancer Lett 2020, 470, 126–133. [Google Scholar] [CrossRef]
- de Visser, K. E.; Joyce, J. A. The Evolving Tumor Microenvironment: From Cancer Initiation to Metastatic Outgrowth. Cancer Cell 2023, 41, 374–403. [Google Scholar] [CrossRef]
- Paget, S. THE DISTRIBUTION OF SECONDARY GROWTHS IN CANCER OF THE BREAST. The Lancet 1889, 133, 571–573. [Google Scholar] [CrossRef]
- Gallo, G.; Vescio, G.; De Paola, G.; Sammarco, G. Therapeutic Targets and Tumor Microenvironment in Colorectal Cancer. J Clin Med 2021, 10. [Google Scholar] [CrossRef]
- Liu, Y.; Li, C.; Lu, Y.; Liu, C.; Yang, W. Tumor Microenvironment-Mediated Immune Tolerance in Development and Treatment of Gastric Cancer. Front Immunol 2022, 13. [Google Scholar] [CrossRef]
- Blank, C. U.; Haining, W. N.; Held, W.; Hogan, P. G.; Kallies, A.; Lugli, E.; Lynn, R. C.; Philip, M.; Rao, A.; Restifo, N. P.; Schietinger, A.; Schumacher, T. N.; Schwartzberg, P. L.; Sharpe, A. H.; Speiser, D. E.; Wherry, E. J.; Youngblood, B. A.; Zehn, D. Defining ‘T Cell Exhaustion. ’ Nat Rev Immunol 2019, 19, 665–674. [Google Scholar] [CrossRef]
- Montauti, E.; Oh, D. Y.; Fong, L. CD4+ T Cells in Antitumor Immunity. Trends Cancer 2024, 10, 969–985. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D. B.; Nixon, M. J.; Wang, Y.; Wang, D. Y.; Castellanos, E.; Estrada, M. V.; Ericsson-Gonzalez, P. I.; Cote, C. H.; Salgado, R.; Sanchez, V.; Dean, P. T.; Opalenik, S. R.; Schreeder, D. M.; Rimm, D. L.; Kim, J. Y.; Bordeaux, J.; Loi, S.; Horn, L.; Sanders, M. E.; Ferrell, P. B.; Xu, Y.; Sosman, J. A.; Davis, R. S.; Balko, J. M. Tumor-Specific MHC-II Expression Drives a Unique Pattern of Resistance to Immunotherapy via LAG-3/FCRL6 Engagement. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Talib, W. H.; Alsayed, A. R.; Abuawad, A.; Daoud, S.; Mahmod, A. I. Melatonin in Cancer Treatment: Current Knowledge and Future Opportunities. Molecules 2021, 26. [Google Scholar] [CrossRef] [PubMed]
- Mortezaee, K.; Potes, Y.; Mirtavoos-Mahyari, H.; Motevaseli, E.; Shabeeb, D.; Musa, A. E.; Najafi, M.; Farhood, B. Boosting Immune System against Cancer by Melatonin: A Mechanistic Viewpoint. Life Sci 2019, 238, 116960. [Google Scholar] [CrossRef]
- Navarro, F.; Casares, N.; Martín-Otal, C.; Lasarte-Cía, A.; Gorraiz, M.; Sarrión, P.; Llopiz, D.; Reparaz, D.; Varo, N.; Rodriguez-Madoz, J. R.; Prosper, F.; Hervás-Stubbs, S.; Lozano, T.; Lasarte, J. J. Overcoming T Cell Dysfunction in Acidic PH to Enhance Adoptive T Cell Transfer Immunotherapy. Oncoimmunology 2022, 11. [Google Scholar] [CrossRef]
- Downs-Canner, S. M.; Meier, J.; Vincent, B. G.; Serody, J. S. B Cell Function in the Tumor Microenvironment. Annual Review of Immunology Downloaded from www.annualreviews.org. Guest 2025, 44, 53. [Google Scholar] [CrossRef]
- I, R.; E, K. Granzyme B-Induced Apoptosis in Cancer Cells and Its Regulation (Review). Int J Oncol 2010, 37. [Google Scholar] [CrossRef]
- Terrén, I.; Orrantia, A.; Vitallé, J.; Zenarruzabeitia, O.; Borrego, F. NK Cell Metabolism and Tumor Microenvironment. Front Immunol 2019, 10. (SEP), 2278. [Google Scholar] [CrossRef]
- Guillerey, C. NK Cells in the Tumor Microenvironment. Adv Exp Med Biol 2020, 1273, 69–90. [Google Scholar] [CrossRef]
- Curti, A.; Ruggeri, L.; D’Addio, A.; Bontadini, A.; Dan, E.; Motta, M. R.; Trabanelli, S.; Giudice, V.; Urbani, E.; Martinelli, G.; Paolini, S.; Fruet, F.; Isidori, A.; Parisi, S.; Bandini, G.; Baccarani, M.; Velardi, A.; Lemoli, R. M. Successful Transfer of Alloreactive Haploidentical KIR Ligand-Mismatched Natural Killer Cells after Infusion in Elderly High Risk Acute Myeloid Leukemia Patients. Blood 2011, 118, 3273–3279. [Google Scholar] [CrossRef]
- Wu, L.; Saxena, S.; Singh, R. K. Neutrophils in the Tumor Microenvironment. Adv Exp Med Biol 2020, 1224, 1. [Google Scholar] [CrossRef]
- Karadas, A. K.; Dilmac, S.; Aytac, G.; Tanriover, G. Melatonin Decreases Metastasis, Primary Tumor Growth and Angiogenesis in a Mice Model of Breast Cancer. Hum Exp Toxicol 2021, 40, 1545–1557. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Shi, H.; Zhang, B.; Ou, X.; Ma, Q.; Chen, Y.; Shu, P.; Li, D.; Wang, Y. Myeloid-Derived Suppressor Cells as Immunosuppressive Regulators and Therapeutic Targets in Cancer. Signal Transduction and Targeted Therapy 2021 6:1 2021, 6, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhang, H.; Chen, X.; Li, C.; Chen, J. Melatonin: A Natural Guardian in Cancer Treatment. Front Pharmacol 2025, 16, 1617508. [Google Scholar] [CrossRef] [PubMed]
- Vitale, I.; Manic, G.; Coussens, L. M.; Kroemer, G.; Galluzzi, L. Macrophages and Metabolism in the Tumor Microenvironment. Cell Metab 2019, 30, 36–50. [Google Scholar] [CrossRef]
- Haston, S.; Gonzalez-Gualda, E.; Morsli, S.; Ge, J.; Reen, V.; Calderwood, A.; Moutsopoulos, I.; Panousopoulos, L.; Deletic, P.; Carreno, G.; Guiho, R.; Manshaei, S.; Gonzalez-Meljem, J. M.; Lim, H. Y.; Simpson, D. J.; Birch, J.; Pallikonda, H. A.; Chandra, T.; Macias, D.; Doherty, G. J.; Rassl, D. M.; Rintoul, R. C.; Signore, M.; Mohorianu, I.; Akbar, A. N.; Gil, J.; Muñoz-Espín, D.; Martinez-Barbera, J. P. Clearance of Senescent Macrophages Ameliorates Tumorigenesis in KRAS-Driven Lung Cancer. Cancer Cell 2023, 41, 1242–1260e6. [Google Scholar] [CrossRef]
- Sumitomo, R.; Hirai, T.; Fujita, M.; Murakami, H.; Otake, Y.; Huang, C.-L. M2 Tumor-Associated Macrophages Promote Tumor Progression in Non-Small-Cell Lung Cancer. Exp Ther Med 2019, 18. [Google Scholar] [CrossRef]
- Wang, S.; Wang, J.; Chen, Z.; Luo, J.; Guo, W.; Sun, L.; Lin, L. Targeting M2-like Tumor-Associated Macrophages Is a Potential Therapeutic Approach to Overcome Antitumor Drug Resistance. NPJ Precis Oncol 2024, 8, 1–19. [Google Scholar] [CrossRef]
- Zhang, D.; Tang, Z.; Huang, H.; Zhou, G.; Cui, C.; Weng, Y.; Liu, W.; Kim, S.; Lee, S.; Perez-Neut, M.; Ding, J.; Czyz, D.; Hu, R.; Ye, Z.; He, M.; Zheng, Y. G.; Shuman, H. A.; Dai, L.; Ren, B.; Roeder, R. G.; Becker, L.; Zhao, Y. Metabolic Regulation of Gene Expression by Histone Lactylation. Nature 2019, 574, 575–580. [Google Scholar] [CrossRef]
- Chen, D.; Zhang, X.; Li, Z.; Zhu, B. Metabolic Regulatory Crosstalk between Tumor Microenvironment and Tumor-Associated Macrophages. Theranostics 2021, 11. [Google Scholar] [CrossRef]
- Hekmatirad, S.; Moloudizargari, M.; Fallah, M.; Rahimi, A.; Poortahmasebi, V.; Asghari, M. H. Cancer-Associated Immune Cells and Their Modulation by Melatonin. Immunopharmacol Immunotoxicol 2023, 45, 788–801. [Google Scholar] [CrossRef]
- Zhang, H.; Yue, X.; Chen, Z.; Liu, C.; Wu, W.; Zhang, N.; Liu, Z.; Yang, L.; Jiang, Q.; Cheng, Q.; Luo, P.; Liu, G. Define Cancer-Associated Fibroblasts (CAFs) in the Tumor Microenvironment: New Opportunities in Cancer Immunotherapy and Advances in Clinical Trials. Mol Cancer 2023, 22. [Google Scholar] [CrossRef]
- Liu, D.; Shi, K.; Fu, M.; Chen, F. Melatonin Indirectly Decreases Gastric Cancer Cell Proliferation and Invasion via Effects on Cancer-Associated Fibroblasts. Life Sci 2021, 277. [Google Scholar] [CrossRef] [PubMed]
- Talib, W. H.; Alsayed, A. R.; Abuawad, A.; Daoud, S.; Mahmod, A. I. Melatonin in Cancer Treatment: Current Knowledge and Future Opportunities. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Li, Y.; Li, S.; Zhou, Y.; Meng, X.; Zhang, J.-J.; Xu, D.-P.; Li, H.-B. Melatonin for the Prevention and Treatment of Cancer. Oncotarget 2017, 8, 39896–39921. [Google Scholar] [CrossRef]
- Zhang, X.; Zhuang, M.; Zhang, H.; Zhu, Y.; Yang, J.; Wu, X.; Yu, X.; Tao, J.; Liu, X. Melatonin-Mediated CGAS-STING Signal in Senescent Macrophages Promote TNBC Chemotherapy Resistance and Drive the SASP. J Biol Chem 2025, 301. [Google Scholar] [CrossRef] [PubMed]
- Won, J. K.; Bakhoum, S. F. The Cytosolic DNA-Sensing CGAS-STING Pathway in Cancer. Cancer Discov 2020, 10, 26–39. [Google Scholar] [CrossRef]
- Pioli, C.; Caroleo, M. C.; Nistico’, G.; Doriac, G. Melatonin Increases Antigen Presentation and Amplifies Specific and Non Specific Signals for T-Cell Proliferation. Int J Immunopharmacol 1993, 15, 463–468. [Google Scholar] [CrossRef]
- Wang, L.; Wang, C.; Tao, Z.; Zhu, W.; Su, Y.; Choi, W. S. Tumor-Associated Macrophages Facilitate Oral Squamous Cell Carcinomas Migration and Invasion by MIF/NLRP3/IL-1β Circuit: A Crosstalk Interrupted by Melatonin. Biochim Biophys Acta Mol Basis Dis 2023, 1869. [Google Scholar] [CrossRef]
- Kumari, A.; Syeda, S.; Rawat, K.; Kumari, R.; Shrivastava, A. Melatonin Modulates L-Arginine Metabolism in Tumor-Associated Macrophages by Targeting Arginase 1 in Lymphoma. Naunyn Schmiedebergs Arch Pharmacol 2024, 397, 1163–1179. [Google Scholar] [CrossRef]
- Silva, K. M. R.; França, D. C. H.; de Queiroz, A. A.; Fagundes-Triches, D. L. G.; de Marchi, P. G. F.; Morais, T. C.; Honorio-França, A. C.; França, E. L. Polarization of Melatonin-Modulated Colostrum Macrophages in the Presence of Breast Tumor Cell Lines. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Wang, K.; Cai, R.; Fei, S.; Chen, X.; Feng, S.; Zhang, L.; Liu, H.; Zhang, Z.; Song, J.; Zhou, R. Melatonin Enhances Anti-Tumor Immunity by Targeting Macrophages PD-L1 via Exosomes Derived from Gastric Cancer Cells. Mol Cell Endocrinol 2023, 568–569, 111917. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Qureshi, M. Z.; Tahir, F.; Yilmaz, S.; Romero, M. A.; Attar, R.; Farooqi, A. A. Role of Melatonin in Carcinogenesis and Metastasis: From Mechanistic Insights to Intermeshed Networks of Noncoding RNAs. Cell Biochem Funct 2024, 42. (4), e3995. [Google Scholar] [CrossRef] [PubMed]
- Fan Gao; Lin, Y. ; Zhang, M.; Niu, Y.; Sun, L.; Li, W.; Xia, H.; Lin, H.; Guo, Z.; Du, G. The Combination of LPS and Melatonin Induces M2 Macrophage Apoptosis to Prevent Lung Cancer. http://www.xiahepublishing.com/ 2022, 7, 201–216. [Google Scholar] [CrossRef]
Figure 2.
Schematic representation of the antitumor mechanisms of melatonin across different types of cancer. In HNSCC, it enhances the infiltration of cytotoxic CD8⁺ T lymphocytes and suppresses EMT. In ovarian cancer, melatonin attenuates the Warburg effect by restoring metabolic flux toward acetyl-CoA and enhancing TCA cycle activity, improving cellular bioenergetic efficiency. In the pancreatic cancer cell line PANC-1, it increases cell death markers. In hepatocellular carcinoma, melatonin downregulates PD-L1 expression in tumor cells, preventing T-cell inhibition mediated by the PD-1/PD-L1 pathway. In breast cancer, its anti-angiogenic effect restricts nutrient and oxygen supply to the tumor microenvironment, strengthening its overall antineoplastic potential. Created in BioRender. Garcia-Macia, M. (2025) https://BioRender.com/v72fvv9.
Figure 2.
Schematic representation of the antitumor mechanisms of melatonin across different types of cancer. In HNSCC, it enhances the infiltration of cytotoxic CD8⁺ T lymphocytes and suppresses EMT. In ovarian cancer, melatonin attenuates the Warburg effect by restoring metabolic flux toward acetyl-CoA and enhancing TCA cycle activity, improving cellular bioenergetic efficiency. In the pancreatic cancer cell line PANC-1, it increases cell death markers. In hepatocellular carcinoma, melatonin downregulates PD-L1 expression in tumor cells, preventing T-cell inhibition mediated by the PD-1/PD-L1 pathway. In breast cancer, its anti-angiogenic effect restricts nutrient and oxygen supply to the tumor microenvironment, strengthening its overall antineoplastic potential. Created in BioRender. Garcia-Macia, M. (2025) https://BioRender.com/v72fvv9.
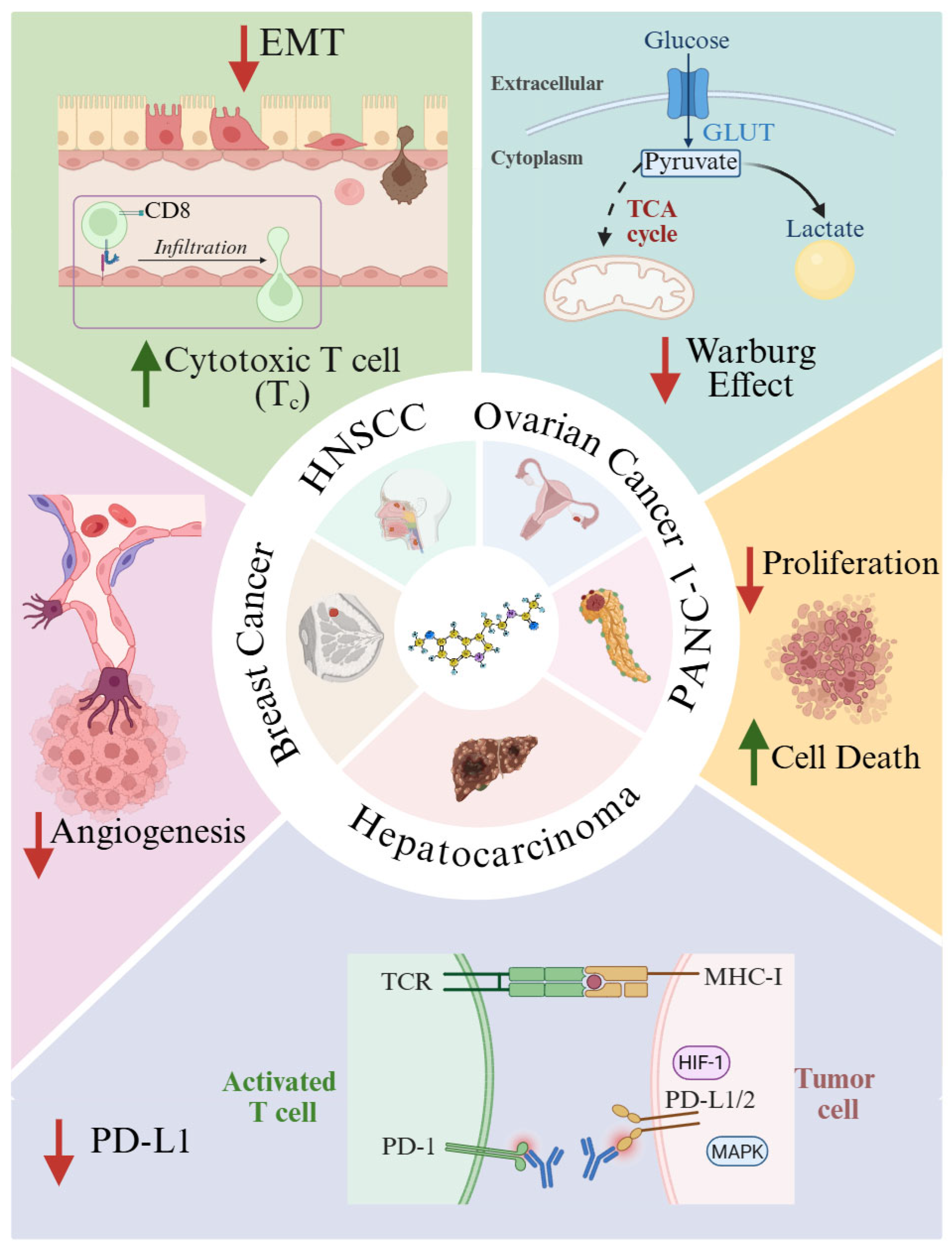
Figure 3.
Melatonin impact on macrophages. Melatonin upregulates Trim26 that contributes to macrophage senescence. It also enhances MHC-II expression, thereby increasing antigen-presenting capacity and amplifying immune activation. Melatonin suppress the MIF/NLRP3/IL-1β signaling cascade, reducing the crosstalk between macrophages and tumor cells. Melatonin inhibits arginase activity increasing NO production. It also modifies the microRNA content of tumor-derived exosomes, leading to decreased PD-L1 expression and attenuated immune suppression. Lack of pineal melatonin, in rats, increases TAM infiltration and promotes tumor progression. Melatonin combined with LPS increases M2 macrophage apoptosis through JAK-STAT and TLR pathways. Created in BioRender. Garcia-Macia, M. (2025) https://BioRender.com/v72fvv9.
Figure 3.
Melatonin impact on macrophages. Melatonin upregulates Trim26 that contributes to macrophage senescence. It also enhances MHC-II expression, thereby increasing antigen-presenting capacity and amplifying immune activation. Melatonin suppress the MIF/NLRP3/IL-1β signaling cascade, reducing the crosstalk between macrophages and tumor cells. Melatonin inhibits arginase activity increasing NO production. It also modifies the microRNA content of tumor-derived exosomes, leading to decreased PD-L1 expression and attenuated immune suppression. Lack of pineal melatonin, in rats, increases TAM infiltration and promotes tumor progression. Melatonin combined with LPS increases M2 macrophage apoptosis through JAK-STAT and TLR pathways. Created in BioRender. Garcia-Macia, M. (2025) https://BioRender.com/v72fvv9.
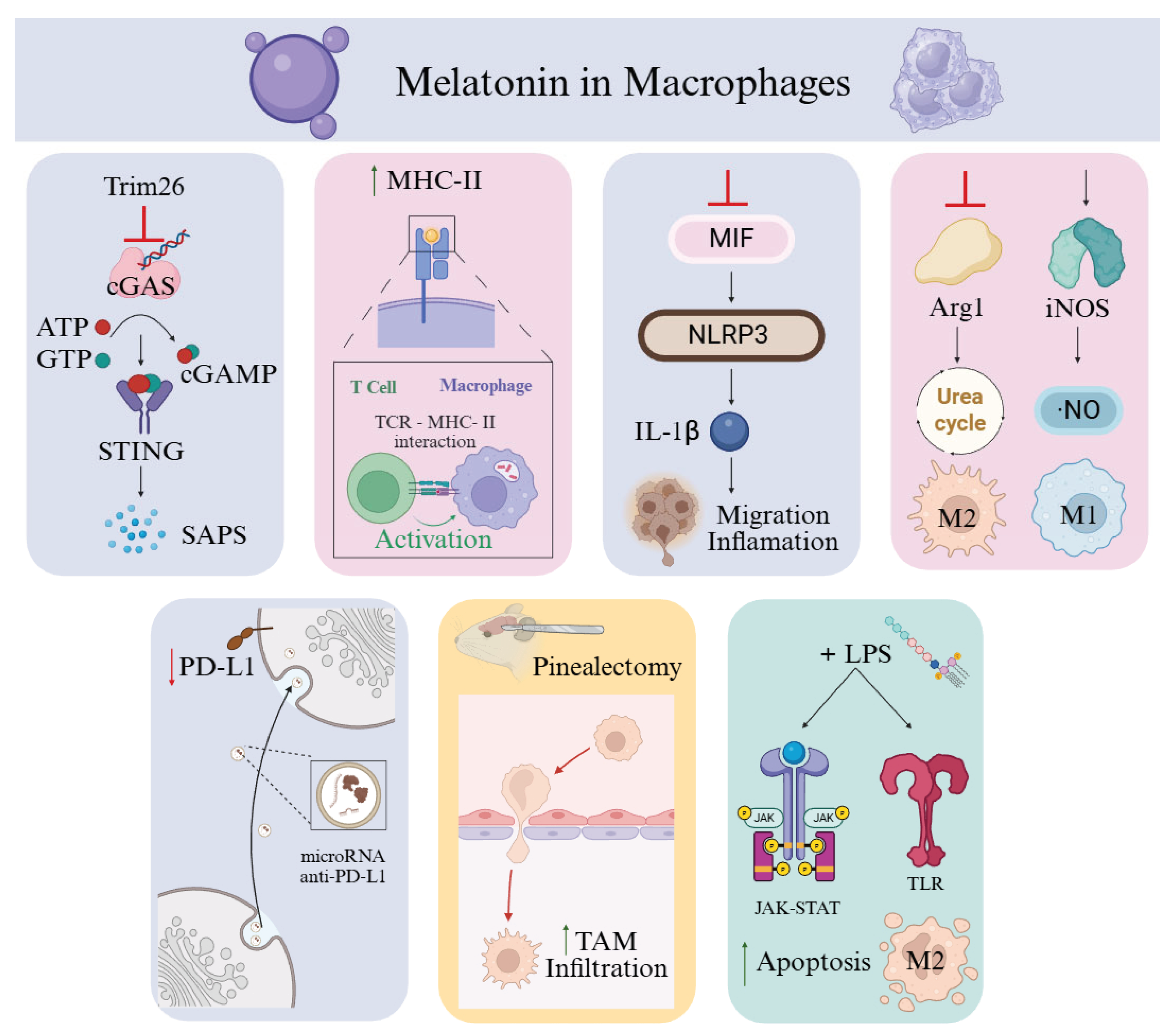
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



