
Submitted:
15 November 2025
Posted:
18 November 2025
You are already at the latest version
Abstract
Presence of pathogenic viruses in wastewater pose a potential threat to public health. Conventional treatment methods often yield moderate viral reduction and toxic byproducts, whereas advanced technologies are underutilised due to their high cost and energy demands. Antiviral phytoremediation emerges as an affordable, eco-friendly and sustainable approach for removing viruses. However, recent bibliometric analysis on wastewater treatment methods from 1976–2025 revealed that only ~0.4% of total literature (~23,000) was related to antiviral phytoremediation suggesting critical knowledge gaps persist. This critical review provides insights into viral removal mechanisms, recent advancements, practical applications, and challenges and opportunities. Antiviral phytoremediation offers a promising multilayer of viral removal mechanisms (i.e., sorption/filtration, rhizosphere-mediated inactivation, internalization, and intracellular degradation mechanisms). Hybrid systems integrating constructed wetlands (CWs) with complementary technologies could achieve high removal efficiencies (i.e., ∼3.0–7 log₁₀ reductions) compared to standalone CWs (i.e., ∼1–3 log₁₀). Although phytoremediation efficiency is moderate for viruses (i.e., ∼45–84%) relative to heavy metal removal (i.e., ∼70–100%), emerging technologies (i.e., CRISPR gene editing, engineered microconsortia, and biosensors) offer promise for enhancement, which is still at proof-of-concept levels. Hybrid antiviral phytoremediation approaches provide sustainable infrastructure supporting public health, climate adaptation, and pandemic preparedness.
Keywords:
phytoremediation
; viral removal
; wastewater treatment
; hybrid systems
; sustainable remediation
1. Introduction
Human pathogenic viruses, including enteroviruses, noroviruses, and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), persist in wastewater collection systems and are distributed through multiple environmental pathways (Figure 1), substantially threatening public health [1,2]. Millions of viral particles from infected individuals enter municipal sewer systems daily, where they survive for extended periods ranging from hours to months, depending on the viral strain, temperature, and organic content. These viruses can contaminate receiving water bodies, agricultural soils, and seafood production systems [3,4].
Gastrointestinal viruses, particularly norovirus, adenovirus, and hepatitis A virus, are highly resistant to traditional disinfectants (e.g., chlorination), ultraviolet (UV) irradiation, and selected advanced treatment processes [5,6]. Advanced technologies, such as membrane bioreactors (MBRs) and advanced oxidation processes (AOPs), yield relatively higher viral inactivation than traditional technologies, but the levels of treatment are still virus specific. The efficiency of MBRs for the removal of different viral pathogens depends on log₁₀ removals of ~1–7 along with published log₁₀ removals of ~4–6 for adenovirus and ~5–6 for norovirus genogroup II [7,8]. The removal efficiency translates into ~97–100% removal efficiency which indicates significant but not complete viral removal.
The widespread deployment of advanced treatment systems in resource-constrained regions is severely limited by (1) high capital expenditure, (2) substantial energy requirements (~0.4–5.0 kWh/m³ of treated wastewater), and (3) technical complexity, which demands skilled operators and sophisticated tracking infrastructure [9,10]. These barriers are acute in low-income regions, where wastewater infrastructure development lags population growth and climate variability exacerbates water stress.
Documented disease outbreaks linked to wastewater reuse disproportionately affect farmworkers, vulnerable consumers, and communities near disposal sites, with children and immunocompromised populations experiencing elevated morbidity and mortality [11].
Detection of SARS-CoV-2 ribonucleic acid (RNA) in wastewater samples from diverse geographic regions has shown the utility of wastewater-based epidemiology for disease surveillance [12,13,14] and has provided compelling evidence for integrating antiviral phytoremediation into post-treatment polishing steps [15].
Nature-based approaches, notably constructed wetland systems (CWs), are highly effective at removing conventional pollutants (i.e., nutrients and pathogens) and emerging micropollutants (pharmaceuticals and microplastics) across all locations and climates [16,17]. Chemical disinfection methods, particularly chlorination, produce potentially carcinogenic water chlorination products including trihalomethanes (THM) and haloacetic acids (HAA) that might persist in treated effluents and receiving waters, thus posing threats to human health, aquatic organisms and ecosystem integrity [18].
Plants are the functional units of constructed wetlands, and have various complementary removal pathways through phytoremediation [19,20]. These pathways encompass (1) rhizofiltration, that is oxidation and immobilisation by adsorbing biofilms of contaminants on the root surfaces, (2) phytoextraction, or selective uptake of metals and transport into harvestable biomass [21], (3) phytodegradation, involving enzymatic breakdown of contaminants within plant tissues, (4) rhizodegradation, where the rhizospheric microbial consortia degrade contaminants stimulated by plant root exudates [22,23,24]. Model macrophyte species, which include Lemna minor and Phragmites australis, have shown contaminant removal efficiencies of ~51–100% for heavy metals (concentration-dependent), ~70–99% for excess nutrients (nitrogen and phosphorus) and ~70–100% for culturable microbial pathogens in the possible configurations of constructed wetlands [25,26,27]. Elevated removal efficiencies are attributed to bioactive secondary metabolites produced, especially polyphenols, terpenoids, and organic acids that (a) have direct antimicrobial properties, or (b) stimulate the increase in beneficial rhizospheric bacteria capable of antagonising pathogenic microorganisms [28,29].
Plants have innate viral defence mechanisms such as non-host resistance pathways and salicylic acid [SA]-dependent inducible defences that inhibit the replication and movement of the virus within plant tissues [30,31,32]. Preliminary studies indicate that specific macrophytes have enhanced antiviral properties. Pennisetum purpureum, for example, effectively removes excess nutrients in constructed wetlands [33], whilst Perilla frutescens produces bioactive phytochemicals which include caffeic acid derivatives and polyphenols that target viral capsid proteins that at the same time modulate host immune signaling and show efficacy against SARS-CoV-2 in vitro [34,35].
To contextualize this review both within and among the existing literature, we performed a comprehensive bibliometric analysis of ~23,000 peer-reviewed publications on wastewater treatment systems within the 1976 to 2025 time-frame. Bibliometric analysis was performed through the online Dimensions platform [36]. Dimensions was chosen for this bibliometric evaluation based on the authenticity and comprehensive nature by which manuscripts, grants, patents, and policy documents are indexed across Google Scholar, Web of Science, and Scopus [37,38]. The insights revealed by this bibliometric survey unveiled a striking gap in knowledge: of the ~23,000 articles identified, only about 81 dealt with plant-based viral removal mechanisms, which reflects a representation of a little less than about 0.4% of the total articles. The phytoremediation literature base is primarily focused on heavy metal juggling (52% of the articles) or bacterial pathogen manipulation (34%), while antiviral phytoremediation is apparently underdeveloped considering the increasing evidence of viral contamination of wastewaters throughout the world gained through surveillance mechanisms (Figure 2). The asymmetrical representation of the forms of phytoremediation is alarming as it points to major voids existing regarding the mechanistic understandings of plant-viral interaction mechanisms among the enveloped and non-enveloped classes of viruses while highlighting the critical need for field-scale testing of hybrid antiviral platforms. Among the key issues of research that have arisen from this literature gap for these forms of phytoremediation are: 1) identifying the plant exudate-rhizospheric microbiota co-metabolic interaction mechanisms, 2) systematically comparing co-metabolic interactions of differing species under different standardized conditions, 3) harmonization of the method for assays of viral measurements among the research teams, 4) field-scale evaluations of the hybrid treatment configurations, 5) targeted genetic engineering of plant genetic traits to develop preferred antiviral phenotypes, and 7) techno-economic and life cycle assessment analyses of treatments used regarding their application to low-income settings. This critical review provides the insights gained from the researches representative of (1) the mechanistic pathways underlying antiviral phytoremediation applied to the molecular and ecosystem scales, (2) the strategies for system designs of phytoremedial platforms with reference to integrating disparities with the complementary technologies for vaunted viral removal, (3) the applications that have been implemented and the documented field-scale successes reached that indicate their operational feasibility, and (4) the ceaseless barriers that remain and the confirmed, evidence-based research directions that should be sought to globalize the use of antiviral phytoremediation.
By integrating perspectives from plant biology, environmental microbiology, wastewater engineering, and systems ecology, this review positions antiviral phytoremediation as a technically feasible, economically scalable, and environmentally resilient part of nature-based infrastructure supporting fair public health preparedness, climate adaptation, and environmental justice across diverse socioeconomic contexts.
2. Mechanistic Basis of Antiviral Phytoremediation
2.1. Phytoremediation Mechanisms
Phytoremediation in planted treatment systems runs through multiple, well-established pathways that function concurrently within the plant–soil–microbe continuum: rhizofiltration (physical interception and interfacial partitioning), phytostabilization (immobilisation within the rhizosphere), rhizodegradation/phytodegradation (microbially mediated and plant-enzymatic transformation), and phytoextraction/phytovolatilization (uptake, sequestration, and release of certain volatile species). The relative contributions of these mechanisms are influenced primarily by hydraulics, redox conditions, and microbial community composition, rather than only by the pathogen or contaminant class [39,40,41].
Rhizofiltration
Root mats, periphytic biofilms, and porous media act as tortuous filters that remove suspended and colloidal matter through straining, attachment, and gravitational sedimentation. Concurrently, dissolved and nano-colloidal species partition to organic and mineral surfaces via electrostatic, hydrophobic, and cation-bridging interactions [39,40]. The efficiency of contact is governed hydraulically: multi-port dosing, baffling, and media resistant to compaction maintain adequate home time and prevent short-circuiting; alternating day dosing in vertical subsurface-flow (VSSF) beds re-oxygenates pores and mitigates clogging, thus sustaining interception stability over time [41,42]. Plant traits reinforce these barriers. A high fine-root surface area and vigorous periphyton growth expand the active surface area, but root exudates modulate the near-root charge and pH, enhancing the sorption and co-precipitation of particulate and dissolved species [43,44].
Phytostabilization
Phytostabilization reduces contaminant mobility and bioavailability through sorption to roots and media, precipitation reactions (e.g., phosphate- or carbonate-mediated), and organic ligand complexation within the rhizosphere (Pilon-Smits 2005; Ruttens et al. 2011). Plants with deep or fibrous root systems create extensive sorptive matrices and microzones characterised by distinct redox potentials and pH conditions that favour the immobilisation of heavy metals. Root cell walls and vacuoles also serve as sinks for ions and polar organic compounds [45]. Phytostabilization efficacy is enhanced by soil amendments such as phosphate and biochar, and, where appropriate, microbial inoculants that reinforce root-associated microbial consortia. Routine tracking is essential to ensure the stability of immobilised fractions under seasonal wetting–drying cycles [46,47].
Rhizodegradation and phytodegradation
The rhizosphere is a chemically reactive and microbially rich zone where plant exudates, including organic acids, amino acids, and phenolics, serve as substrates for heterotrophic microorganisms and modulate the local redox conditions. This environment fosters division-of-labour microbial consortia that can transform dissolved organic compounds and co-metabolise diverse contaminants (rhizodegradation) [23,48,49]. Biofilms on roots and media secrete enzymes, such as oxidases, peroxidases, and hydrolases, which accelerate the depolymerisation and humification of organic matter. Oxygen release via aerenchyma creates mixed redox mosaics that support coupled nitrification–denitrification and sulfur/iron cycling [39,40]. Within plant tissues, endogenous enzymes, including peroxidases, dehalogenases, and nitrilases (via phytodegradation), further transform the absorbed organic compounds. Some transformation products are sequestered in cell walls or vacuoles, completing partial mineralisation across plant and microbial compartments [20,44]. Where coloured dissolved organic carbon (DOC) or recalcitrant micropollutants limit biodegradation, staging a photochemically active free-water cell upstream can pre-oxidise chromophores, thus enhancing downstream biodegradability without compromising rhizosphere function [50].
Phytoextraction and phytovolatilization
Phytoextraction involves the removal of dissolved ions and certain polar organic compounds via root uptake, followed by intracellular binding to organic acids and thiol-rich peptides, and compartmentalisation within vacuoles or deposition in cell walls [20,41,51]. Periodic harvesting exports sequestered contaminants and promotes belowground turnover and exudation. Species showing high growth rates, extensive aerenchyma, and dense fine-root systems sustain greater contaminant fluxes without excessive clogging [40,44]. Phytovolatilization contributes to a limited set of volatile elements and compounds, notably mercury (Hg) and selenium (Se) species, through biochemical reduction or transformation processes that generate volatile forms released via transpiration [52]. The desirability of phytovolatilization depends on the specific compounds involved and the regulatory policies in place. Most engineered systems prioritise retention and in situ transformation over atmospheric transfer [20,39].
2.2. Antiviral-Specific Mechanisms
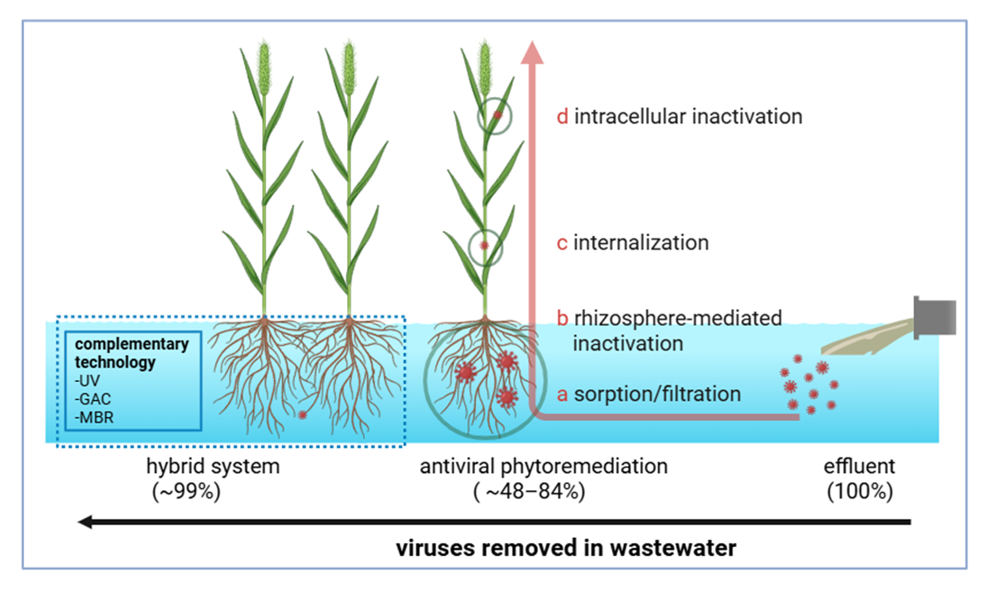
Antiviral phytoremediation functions as a multi-barrier system in which plants and their associated microbiomes attenuate viral contaminants through four interconnected pathways: sorption–filtration, rhizosphere-mediated inactivation, viral internalisation, and intracellular degradation (Figure 3). This framework extends classical phytoremediation ideas to encompass viruses by adapting plant-driven mass transfer, surface interactions, and immune-like responses to reduce the number of infective particles in complex wastewater matrices [23,53,54].
Sorption–filtration at plant, biofilm, and substrate interfaces
Sorption–filtration is the initial barrier, intercepting virions on high-surface-area matrices, including roots, biofilms, and porous substrates [55,56]. Hydrophobic interactions, electrostatic forces, and cation bridging immobilise viral particles, but size exclusion within tortuous pore networks physically retards their transport [39]. In constructed wetlands, plant-supported biofilms and mineral surfaces provide dominant specific surface areas similar to the mechanisms of immobilising dissolved metals via sorption and complexation [57,58]. The performance depends on the hydraulic retention time and the interfacial surface area. Field and pilot studies have consistently shown longer retention times and denser rootbiofilm matrices correlate with enhanced pathogen attenuation. But clogging and hydraulic short-circuiting reduce contact efficiency, helping with viral breakthrough [59]. Across systems, viral reductions attributable only to physical barriers typically range from ~1–3 log₁₀ and vary with virion surface chemistry, ionic strength, and organic matter content, highlighting the necessity of downstream biological mechanisms [60].
Rhizosphere-mediated inactivation
The rhizosphere acts as a chemically reactive zone where plant exudates and microbial metabolites, such as reactive oxygen species and lytic enzymes, can damage viral capsids and genomes. Root-derived phenolics and flavonoids could directly inhibit viral attachment and fusion, and modulate microbial consortia, providing a biochemical complement to physical capture [23,28,61,62]. Engineered and naturally assembled plant–microbe consortia enhance these effects through division-of-labour metabolism and enzyme secretion, amplifying viral inactivation within the root boundary layer [49,63]. Non-enveloped viruses, characterised by rigid icosahedral capsids and exceptional environmental stability, generally require stronger rhizosphere chemistry and prolonged contact times to achieve attenuation comparable to that of enveloped viruses [8,64,65].
Viral internalization (cell entry and early destabilization)
Following surface capture, some virions could internalised into root tissues via endocytic and non-endocytic pathways [66,67]. Enveloped viruses display greater susceptibility to plant-mediated internalisation and destabilisation, largely due to their fragile lipid envelopes, but non-enveloped viruses, protected by strong icosahedral capsids, exhibit greater persistence and resistance to internalisation and early chemical degradation [8,64,68]. Plant chitinases and β-1,3/1,6-glucanases (glycoside hydrolase family 19 [GH19]), along with lectins that recognise N-acetyl-glucosamine motifs, can target glycoprotein domains on enveloped viruses during or after entry, helping with particle disassembly [23,66]. Heterologous expression of antiviral membrane proteins (e.g., IFITM3 [Interferon-Induced Transmembrane Protein 3]) may enhance uptake-linked destabilisation by modulating late endosomal lipid dynamics in model plants, suggesting the potential for engineered improvements to this barrier [69,70].
Intracellular inactivation (genome silencing and proteolysis)
Once internalised, plant antiviral responses operate on multiple fronts. Small RNA pathways (RNA interference; RNAi) mediate post-transcriptional silencing of viral genomes, while the ubiquitin–proteasome system and macroautophagy (autophagy-related atg-dependent) help to degrade viral proteins and particles [71,72,73]. Phytochemicals, such as the bioflavonoid rutin from Ocimum basilicum (via phytoinactivation), bind to structural proteins across diverse viruses with favourable binding energies (−7 to −10 kcal·mol⁻¹), inhibiting attachment, fusion, and genome release [74]. Peroxidases and proteases also contribute to capsid degradation, particularly under oxidative conditions [23,49]. Owing to the pH and temperature tolerance of non-enveloped viruses and their resistance to lipid-targeting phytochemicals, achieving ~1–3 log₁₀ removal typically requires the concerted action of all four barriers combined with enough retention time, highlighting the importance of designing antiviral phytoremediation as a sequential coupled system [75,76]. From a systems perspective, intracellular pathways establish the ceiling for true viral inactivation (loss of infectivity), but upstream barriers regulate the encounter frequency and loading of the cellular machinery. Designs incorporating high-surface-area capture, chemically active rhizospheres, and enough retention time before intracellular processes could yield the most reliable viral removal performance across virus classes and seasonal variations [39,77].
2.3. Comparative Analysis of Viral Particles and Heavy Metals
Removal mechanisms
Plants and plant–microbe systems remove heavy metals and viruses through superficially similar stages: interception at interfaces, transformation in the rhizosphere, and organism-level processing. However, the underlying physicochemical and biochemical mechanisms differ significantly, with important implications for system design (Table 1). Heavy metals, primarily existing as ions or complexes, are predominantly removed via sorption and complexation to roots, biofilms, and mineral media [78]. Subsequent uptake, chelation (e.g. by organic acids and phytochelatins), and vacuolar sequestration stabilise metals within plant tissues, culminating in phytostabilization and phytoextraction [79,80]. Viruses (nanoscale colloids composed of proteinaceous capsids with or without lipid envelopes) are first immobilised through electrostatic and hydrophobic interactions, followed by inactivation mediated by exudate chemistry and microbial enzymes [81,82]. A subset undergoes internalisation and intracellular degradation [83].
These differences arise from particle identity: metals are elemental and cannot be degraded, only transformed in speciation and sequestered [84], but viral particles are supramolecular assemblies whose infectivity can be neutralised through capsid or envelope damage and genome cleavage [82]. Achieving “true removal” of metals focuses on preventing remobilization (e.g., via pH stability), but viral removal emphasises the loss of infectivity in addition to physical capture [85,86]. Structurally, the strong icosahedral capsids of many non-enveloped viruses confer resistance to desiccation, pH fluctuations, and moderate oxidants, but lipid envelopes are more labile [87]. This contributes to the generally higher susceptibility of enveloped viruses to rhizosphere chemistry and intracellular defences compared to their non-enveloped counterparts [77]. At the interface scale, the adsorption parameters also differ. For model bacteriophages and coronaviruses, the Freundlich adsorption coefficients range from ~2×10³–2.7×10⁵ mL·g⁻¹ and vary with ionic strength, organic matter content, and temperature [88]. Divalent metal cations show more predictable surface complexation trends, primarily governed by pH and competing ligands [89]. Therefore, design strategies emphasise charge neutralisation and aggregation aids (e.g. biofilm matrices and upstream destabilisation) for viruses, but pH and redox control, alongside ligand management, predominate for metals.
Factors influencing removal efficiency
Pilot- and field-scale macrophyte systems commonly report ~1–2 log₁₀ reductions in viral load attributable to physical capture alone, with higher total reductions achieved when rhizosphere inactivation and intracellular degradation pathways are activated (Table 1) [90]. Plant traits that increase the active surface area, such as fine-root density and healthy periphyton growth, combined with hydraulics that prevent short-circuiting, enhance encounter rates and viral attenuation [91]. Temperature fluctuations influence virion partitioning and adsorption efficiency, contributing to seasonality in virus removal, a phenomenon that is less pronounced in metals [92]. For heavy metals, removal efficiencies are often higher and more consistent under comparable hydraulic retention times, often reaching ~70–100% in well-maintained wetlands or rhizofiltration units if metal speciation favours sorption and that plant uptake and compartmentation capacities are not saturated [93]. Virus removal is more sensitive to virion architecture (enveloped vs. non-enveloped), dissolved organic carbon (DOC), and competing colloids. For heavy metals, removal efficiencies are often higher and more consistent under comparable hydraulic retention times, often reaching ~70–100% in well-maintained wetlands or rhizofiltration units if metal speciation favours sorption and that plant uptake and compartmentation capacities are not saturated [93]. Virus removal is more sensitive to virion architecture (enveloped vs. non-enveloped), dissolved organic carbon (DOC), and competing colloids. Chemical amendments can help to narrow this performance gap. Polyphenols, such as rutin, bind to viral surface proteins with favourable free energies (−7 to −11 kcal·mol⁻¹), promoting virion aggregation and enhancing capture efficiency. Upstream charge destabilisation methods, including electrocoagulation and optimised flocculation, further increase early-stage retention within root-associated matrices [83,94,95]. Maintaining young, metabolically active plant stands with a balanced nutrient status and moderate redox heterogeneity sustains both antiviral metabolite flux and metal-binding capacity [96,97,98].
Design considerations for combined virus and metal removal
When wastewater has both viruses and dissolved metals, the system design must integrate complementary removal barriers while minimising interference (Table 1). Organic acids and flavonoids that help with viral inactivation also chelate metals and influence speciation; generally, this enhances metal immobilisation but may increase competitive adsorption with natural organic matter, requiring careful control of influent chemistry and retention time [99,100]. Similarly, dense biofilms that capture virions provide ligands for metal binding. However, operational challenges include avoiding clogging while preserving a high surface area [80,101]. Operational tracking should reflect these differences. Viral removal requires infectivity assays or confirmed surrogates to confirm true inactivation, but metal removal tracking focuses on dissolved and particulate speciation and plant tissue burdens to assess stability against re-mobilisation [75,102]. For example, macrophyte beds based on Pistia stratiotes achieve ~0.5–1.0 log₁₀ reduction of bacteriophage surrogates per pass while simultaneously lowering the dissolved metal concentrations [83,103,104,105]. Coupling with polishing steps such as ultraviolet (UV) irradiation, ozone, or ferrate increases viral removal to ~3–7 log₁₀, with metal removal maintained or enhanced [106,107,108]. Cost and reliability considerations often favour modular hybrid systems. Downstream activated carbon or nanofiltration units provide resilience during peaks in organic load or viral spikes and control trace organics that may interfere with adsorption and enzymatic activities. These technologies have also been established for treating metal-bearing effluents [109,110]. Adaptive tracking and rule-based operational control, such as flow equalisation, intermittent aeration, and targeted coagulant dosing, support sustained performance under seasonal stressors [111,112,113,114].
Emerging innovations for enhanced removal performance
Three promising innovation streams address the differential removal challenges posed by viruses and heavy metals. First, engineered plant–microbe consortia that partition antiviral and metal-binding functions can amplify rhizosphere chemistry without compromising stability. Examples include Streptomyces and Bacillus species, which provide complementary lytic enzymes alongside exopolysaccharide-rich periphyton that enhance metal capture and complexation [115,116]. Second, targeted host enhancement through the choice of genotypes showing high fine-root areas and the introduction of traits that bolster antioxidant and proteostasis pathways can improve intracellular viral degradation while maintaining metal tolerance [117,118].

Table 1.
Virus and heavy metal removal mechanisms in plant-microbe rhizosphere system.
|
Mechanism/Parameter |
Virus (Antiviral Mechanism) |
Heavy Metal (co-Removal) |
Performance Metrics |
Critical Variables |
Example Systems |
Ref. |
|---|---|---|---|---|---|---|
| Primary removal | electrostatic/hydrophobic adsorption; aggregation; enzymatic inactivation | ion exchange; surface complexation; chelation; precipitation | virus: ~1–2 log₁₀ capture, up to 7 log₁₀ with polishing; metal: ~70–100% removal | charge density, DOC, pH, root potential, ionic strength | Pistia stratiotes, Typha latifolia, Phragmites australis beds | [80,81,90] |
|
Rhizosphere biochemistry |
exudate oxidation, proteolysis; polyphenol virion destabilization (ΔG ≈ -10 kcal/mol) | organic acid complexation; phytochelatin synthesis; redox cycling | inactivation rate: k = ~0.02–0.07 h⁻¹ (25°C); infectivity loss: 65 ± 12% (48 h) | root activity, flavonoid flux (~0.8–1.5 mg g⁻¹ DW), microbial profile, T-sensitivity | natural/ changed wetlands | [83,96,119] |
| Particle stability | capsid/envelope disruption, genome cleavage; enveloped viruses removed ~2–5× better | speciation-dependent stability; vacuolar sequestration post-uptake | enveloped removal: >90%; RNA decay: ~65–84% (~48–72 h) | temperature, pH, oxidative potential, virion charge | macrophyte–biofilm systems | [83,87] |
|
Adsorption/ partitioning |
Freundlich Kₙ = ~2×10³–2.7×10⁵ mL g⁻¹; mean capture: 58 ± 20% | surface complexation log K = ~4–8 (pH-driven) | capture efficiency: ~58 ± 20% (n=16); K = ~10³–10⁴ mL g⁻¹ for bacteriophages | ionic strength, DOC competition, surface pKa, hydrophobicity | rhizofiltration, periphyton-root systems | [88,89] |
| HRT requirement (d) | ~3–6 d for viral attenuation; ideal ~5–10 d | ~2–4 d for metal sorption equilibrium | virus: ~1–2 log₁₀ per stage; metal: ~70–100% removal | flow uniformity, aeration regime, recirculation, temperature effects | hybrid wetland + UV, VSSF units | [93,106] |
| Chemical aids | polyphenols (ΔG); electrocoagulation (EC) → ~2–3× capture boost | biochar, zeolite, Fe(OH)₃, molecular imprinted polymers composites | virus capture: +1–1.5 log₁₀ gain with EC; metal removal: +15–30% with media | coagulant dose (FeCl₃ ~5–20 mg/L), pH, oxidation reduction potential (ORP) | modular wetland-filter hybrids |
[109,120,121] |
| Microbial contribution | lytic enzymes, quorum-regulated proteases (Bacillus); ROS generation | extracellular polymeric substance matrix, siderophore secretion, biosorption | +0.5–1.0 log₁₀ increment; peroxidase activity ↑25–60%. | microbial diversity, nutrient ratio (C:N:P ≈ 100:10:1), rhizosphere age | engineered consortia | [41,116,122] |
| Seasonal sensitivity | strong T-dependence (−0.3 log₁₀ per 10°C drop); dissolved organic carbon (DOC) competition | moderate; resilient under redox/pH shift | winter: retains ~70–85% of summer rate with thermal buffering | temperature, DOC level, biofilm maturity, flow fluctuation | aerated/intermittent-flow constructed wetland systems | [91,92,123] |
| AI control | adaptive flow/dosing for dual targeting; real-time viral prediction (~12–18 h lead time) | dynamic ligand control via real-time speciation | ±10% variance reduction under fluctuating loads | pH/ORP sensors, metabolite biosensors, AI feedback | smart AI-integrated wetlands | [114,124] |
Third, upstream physicochemical aids tailored to viral colloids, such as mild electrocoagulation for charge neutralisation, reduce biological stage loading and stabilise performance under cold or high-DOC conditions while remaining compatible with metal removal pathways [125,126,127]. Looking forward, artificial intelligence (AI)-guided tracking and adaptive control of flow, aeration, and dosing parameters can optimise coupled system performance across seasonal and influent variability, enhancing both viral inactivation and metal immobilisation [124]. Collectively, these approaches translate mechanistic contrasts into complementary barriers that achieve high and stable removal efficiencies across various classes of contaminants.
3. Recent Advancements in Antiviral Phytoremediation
3.1. Plant Selection and Optimization
Strategic choice of antiviral plants via mechanisms to traits
Selecting plants for antiviral phytoremediation should be grounded in the four-barrier framework outlined in previous Section 2 (i.e., capture, rhizosphere inactivation, internalisation, and intracellular degradation) and mapped to specific plant traits (Table 2). These include a high fine-root specific surface area to enhance sorption–filtration, metabolically active rhizospheres that release phenolics and flavonoids and support dense periphyton to promote chemical and enzymatic inactivation, tissues exhibiting strong endocytosis and proteostasis for efficient internalisation and intracellular degradation, and shoot–root architectures compatible with target hydraulics and redox zoning [56,83,128]. Systems maintaining young, rapidly growing stands with vigorous root turnover and periphyton renewal provide more stable viral attenuation across seasons than overaged canopies with diminished belowground activity [40,129]. Seasonal variability and influent chemistry further refine the plant-selection process. Species that sustain exudation and peroxidase/oxidase activity at lower temperatures (°C) and tolerate dissolved organic carbon (DOC) fluctuations without impairing biofilm health mitigate performance decline during winter and high DOC shocks [130,131]. Matching species to hydraulic regimes is essential; for example, VSSF wetlands benefit from species that maintain porosity and resist clogging under intermittent loading, but free-water surface beds prioritise emergent canopies that enhance light and oxygen inputs for oxidative chemistry [132,133,134].
High-performance antiviral plant species
Evidence from bench, pilot, and field studies has identified macrophytes that combine extensive, cleanable surface areas with chemically active rhizospheres as high-performing (Table 2). P. australis, Typha spp., Cyperus spp., and selected aromatic or medicinal species, such as Ocimum basilicum and Strobilanthes cusia, showed superior antiviral phytoremediation potential [135,136]. P. australis consistently supports diverse, functionally rich rhizobacterial communities linked to pathogen degradation while preserving hydraulic conductivity in VSSF beds. Other macrophytes, such as Typha and Cyperus, provide comparable periphyton support and tolerate nutrient and DOC variability typical of municipal influents [137,138]. Certain medicinal plants confer distinct antiviral advantages through exudates or tissue-bound polyphenols and alkaloids. For example, rutin from O. basilicum exhibited favourable binding free energies (−9.7 to −10.9 kcal·mol⁻¹) against multiple viral surface proteins, promoting virion aggregation and helping with capture within root and biofilm matrices [74]. S. cusia produces tryptanthrin, which inhibits the coronavirus NL63 protease at low micromolar concentrations [139,140]. Interplanting these species with structural macrophytes, such as Phragmites, could results in a division of roles, with large surface areas for virus capture and enhanced rhizosphere chemistry, resulting in improved and consistent virus removal [138,141].
Selection criteria and screening workflow
A strong plant selection workflow integrates (i) desk-based pre-screening, (ii) bench assays, and (iii) pilot verification of the selected plant. Pre-screening filters candidate species based on local availability, invasiveness risk, hydraulic compatibility (root porosity, aerenchyma), and evidence of antiviral metabolites or vigorous rhizosphere metabolism [142,143]. Bench assays measure barrier-aligned metrics, including short-term sorption coefficients (mL·g⁻¹) on roots and periphyton, exudate-driven loss of infectivity for enveloped versus non-enveloped viral surrogates, early internalisation markers such as viral RNA decay within root tissues over ~24–72 h, and indicators of intracellular degradation, including peroxidase, protease, and RNase activities, alongside stress-response gene expression under realistic ionic strength and DOC levels [66,144,145]. Pilot verification in the intended hydraulic configuration should confirm (a) the stability of log reductions across seasons and loading rates, (b) maintenance requirements, such as harvesting frequency and clogging control, and (c) ecological safeguards to ensure no pathogen amplification or spread of non-native taxa. Comparative tests among Phragmites–Typha–Cyperus mixtures typically reveal complementary hydraulic and rhizosphere chemistries. Including a minority fraction (~10–30% stem density) of antiviral-rich aromatic species can enhance inactivation without compromising hydraulic conveyance [143].
Optimization strategies and pretreatment integration
Three consistent optimisation strategies can improve plant-mediated antiviral performance. First, maintaining young, metabolically active stands through staged planting and periodic harvesting preserves a high root-specific surface area and fresh periphyton [146]. Second, stabilising rhizosphere chemistry by ensuring a balanced nutrient supply and moderate redox heterogeneity, such as intermittent aeration in VSSF wetlands, sustains enzyme production and reactive oxygen species critical for capsid and envelope damage [147,148,149]. Third, pairing plants with gentle pretreatment methods that enhance early viral capture without damaging roots or biofilms, such as mild electrocoagulation or optimised flocculation to neutralise charge and aggregate virions before root zone contact, further improves performance [150,151,152]. In systems experiencing frequent viral loads and DOC spikes, interplanting antiviral-rich species (i.e., O. basilicum and S. cusia) with structural macrophytes (Table 2) and incorporating upstream destabilisation steps (iron-based coagulants or low-dose electrocoagulation) can increase first-barrier retention and reduce the intracellular processing burden [150,151,152]. Maintaining system flexibility for seasonal rebalancing through transient aeration and selective harvesting keeps the plant–microbe assembly near ideal performance [130,153].
3.2. System Optimization
Hydraulics and contact optimization
Virus attenuation in planted treatment systems is limited by contact efficiency; thus, optimising the flow architecture to maximise the uniform interaction between influents and active interfaces including roots, biofilms, and media is critical (Table 3). In VSSF beds, the influent should be evenly distributed via multiple ports or perforated headers, with hydraulic loading rates (HLR) matched to bed conductivity and maintained porosity to prevent short circuiting. In free-water surface (FWS) cells, the installation of baffles and staged islands can straighten flow paths and reduce wind-driven recirculation, which dilutes root zone contact [59,154,155].
Table 2.
Plant traits and viral removal in constructed wetland systems.
| Scientific Name (Common Name) |
Functional Traits (Key Mechanisms) |
Optimal Configurations |
Viral Removal Performance (log₁₀ Reduction) |
Co-Removal Benefits |
Critical Constraints |
Ref. |
|---|---|---|---|---|---|---|
| Monoculture systems | ||||||
| Phragmites australis (Common reed) | fine root area (>300 cm² g⁻¹); dense periphyton; strong O₂ transfer; high porosity (~25–35%) | VSSF (intermittent loading); baffled FWS | 1.2 ± 0.3 log₁₀ (capture-dominant); field stability ~60–75% | high N/P removal (~70–99%); stable heavy metal uptake (~70–90% Zn/Cu) | seasonal dormancy (winter); requires periodic harvest (~2–4× yr⁻¹); establishment time 4–6 wks | [40,138,156] |
| Ocimum basilicum (Sweet basil) | high phenolic/flavonoid exudates (0.8–1.5 mg g⁻¹ DW); elevated oxidase/peroxidase activity | horizontal/free-water flow with aeration; mixed beds (2:1 ratio) | 2.3 ± 0.4 log₁₀ (chemical inactivation); +40% for enveloped viruses | volatile oil antimicrobial effects; phenolic anti-biofilm agents; biomass valorization potential | high T-sensitivity (~20–30°C ideal); short lifespan (requires replacement ~2–3× yr⁻¹) | [83,139,140] |
| Strobilanthes cusia (Assam indigo) | indole alkaloid production (Tryptanthrin 10–50 µM IC₅₀); elevated RNase/protease activity | floating macrophytes; warm shallow beds (~20–28°C) | +0.7 ± 0.2 log₁₀ gain over baseline (intracellular enzymatic defense) | medicinal/commercial value co-product potential; strong nucleic acid hydrolysis capability | tropical requirement (dies <10°C); limited geographic deployment; alkaloid bioaccumulation risk | [104,139,140] |
|
Pistia stratiotes (Water lettuce) |
extensive adventitious root system; rapid biomass production; high transpiration | floating-bed systems; rhizofiltration units | ~0.5–1.0 log₁₀ per pass; ~3–5 log₁₀ in CWs–UV hybrid (high sorption capacity, K = ~10³–10⁴ mL g⁻¹) | high heavy metal uptake (~70–85%); scalable for rapid deployment | Invasive potential (requires containment); sensitive to low DOC/high shear; capture-dominant mechanism | [83,157,158] |
| Optimized polyculture systems | ||||||
| Phragmites + Typha + Ocimum (triculture) | trait complementarity: max surface area + diverse exudate chemistry + functional redundancy | coupled VSSF–free-surface system | 2.8 ± 0.5 log₁₀ reduction; 85 ± 10% infectivity loss (capture–inactivation synergy) | superior stability; buffering seasonal/load variations; showed performance over 3+ yrs | higher complexity in operation and maintenance (O&M); longer initial establishment (~8 wks); requires strict nutrient control | [138,143,159] |
Design heuristics recommend targeting a municipal-strength wastewater HRT that secures physical barrier removal of ~1–2 log₁₀ in the initial stage, followed by an additional ~0.5–1.0 bed volume for rhizosphere chemical and intracellular viral attenuation [128,160,161]. When the influent organic matter is elevated or the temperatures are low, incorporating a ~20–40% HRT safety margin or a compact pre-treatment stage (see Pretreatment, polishing, and seasonal adaptation) stabilises upstream capture and mitigates desorption risk [123,162]. Because viral removal scales with the interfacial area, selecting plant species and media that maintain a high specific surface area, such as fine roots and cleanable media, and avoiding compaction are paramount [163]. Operational strategies should maintain a high “contact efficiency” throughout the system lifespan. Alternating-day dosing in VSSF systems re-aerates pores and supports biofilm recovery, rotating inlet zones minimise localised clogging, and annual tracer tests reassess effective HRT. If the effective HRT falls below about ~70% of the design or home time variance goes up, remedial actions such as hydro-flushing, media scarification, or partial media replacement should be undertaken before an irreversible performance decline occurs [164].
Rhizosphere chemistry and redox management
Sustaining a metabolically active and chemically reactive rhizosphere is the second key optimisation lever, as it helps with capsid and envelope damage and accelerates the intracellular antiviral response (Table 3). Intermittent aeration in VSSF systems using duty cycles that maintain dissolved oxygen (DO) levels above 2 mg·L⁻¹ near inlets while preserving anoxic zones deeper in the bed could support oxidative enzymes and reactive oxygen species generation without inhibiting denitrification or causing biofilm overoxidation [48,165,166]. In FWS cells, shallow shelves combined with emergent canopies enhance light and oxygen penetration, promoting peroxidase activity and exudation, and providing a habitat for periphyton development [167]. Nutrient balance critically influences antiviral metabolite flux and microbial consortia dynamics. Maintaining carbon:nitrogen:phosphorus (C:N:P) ratios prevents carbon starvation, which suppresses enzyme production and avoids eutrophic blooms that can smother roots. Temperature-responsive regulation in beneficial Pseudomonas spp. and quorum-sensing mechanisms in biofilms further modulate in situ metabolite release [168,169,170,171]. Plant immune signalling pathways, such as salicylic acid/NPR1 and redox cues, potentiate rhizosphere responses under stress, linking systemic acquired resistance with localised oxidative bursts and enzymatic defences [172,173,174,175]. During cold seasons, stabilising metabolism by increasing water levels (thermal buffering) and reducing HLR to preserve contact time compensates for slowed reaction rates [123,176]. Tracking should prioritise functional indicators over surrogate parameters. Routine physicochemical profiling (DO, oxidation-reduction potential [ORP], pH, and conductivity) should be complemented by periodic assays of peroxidase and protease activities or proxies for exudate production. Declines in enzymatic activity justify stand rejuvenation via selective harvesting and short-term aeration changes to prevent downstream performance loss [177].
Biomass and media maintenance
Optimized phytoremediation systems maintain an actively renewing surface characterized by abundant fine roots, refreshed periphyton, and unclogged pore networks (Table 3). Staggered harvesting, which involves removing ~20–40% of the aboveground biomass per event two to four times annually depending on the growth rate, promotes root turnover and exudation while preventing thatch accumulation [178,179]. Rapid removal of senescent litter is essential, as its decomposition elevates dissolved organic carbon (DOC) levels and fosters anoxic mats that impair viral capture and enzymatic activity [180,181]. Media stewardship is also important. Use well-graded aggregates resistant to compaction that provide tortuous flow paths and minimise fines (<1–2 mm), except in thin reactive layers [182]. When the head loss goes up, back-flushing with clean water or air-pulse scouring during offline periods may restore the permeability. Persistent clogging near inlets requires dosing line rotation, raising or flipping near-surface media, or replacing the top ~10–15 cm of media to restore hydraulic conductivity and root penetration [183,184]. Excessive biofilm accumulation may slough during load shocks, releasing colloids and viruses. Moderate shear stress via pulsed dosing and avoidance of abrupt changes in the hydraulic loading rate mitigate the risk of sloughing. Recurrent sloughing events require shortening of the dosing pulses, increasing rest intervals, and assessing nutrient balance. Sometimes, a small upstream roughing filter captures sloughed material before it enters the planted beds [183,184].
Pretreatment, polishing, and seasonal adaptation
Gentle pretreatment stabilises the initial viral capture barrier without damaging the plants or biofilms (Table 3). Practical options include (i) low-dose coagulant addition (e.g. iron salts optimised by zeta potential) to neutralise the charge and promote virion aggregation and (ii) mild electrocoagulation in compact tanks operated at conservative current densities. Both approaches could enhance early retention within the root and biofilm matrices, reducing the intracellular processing burdens, particularly under high DOC or low-temperature conditions [121,185,186,187]. For stringent compliance, especially when non-enveloped viral surrogates dominate or during peak viral loads, macrophyte stages should be paired with compact polishing units such as ultraviolet (UV) irradiation (with confirmed fluence), ferrate or ozone micro-dosing, or short granular activated carbon (GAC) contactors. Hybrid treatment trains show additive viral inactivation relative to individual barriers [188,189]. Oxidant placement is critical, and oxidants should be kept downstream of plants to avoid phytotoxicity. If upstream stabilisation is necessary, low doses with short contact times and rapid quenching should be used [190,191]. Explicit seasonal operating envelopes can enhance the resilience of the system. In summer, the HLR within capacity should be increased, biomass removal intervals should be shortened, and brief daytime aeration should be added to suppress anoxic mats [192,193]. In cold-temperate climates, coupling wetlands with low-enthalpy geothermal systems supports year-round function and reduces the energy penalties associated with greenhouse enclosures [194,195].
3.3. Hybrid Systems and Technology Integration
Rationale and design principles
Hybrid systems may integrate plant antiviral mechanisms (i.e., sorption/filtration, rhizosphere inactivation, viral internalisation, and intracellular degradation) with on-site biophysiochemical barriers and/or other treatment methods to enhance log removal, stabilise performance under cold or high DOC conditions, and reduce spatial footprints for urban applications. Typically, vertical or subsurface constructed wetlands are preceded by charge-destabilisation units such as electrocoagulation or optimised coagulant dosing, and followed by ultraviolet (UV) irradiation or granular activated carbon (GAC) polishing. Membrane bioreactors (MBRs) serve as alternative polishing steps when stringent reuse standards are applied [7,196]. This design logic aligns with mechanistic complementarity: pretreatment enhances early virion capture, planted-vegetation beds help with biochemical inactivation, and polishing secures complete loss of infectivity, especially in non-enveloped viruses [75,197]. Across pilot studies, hybrid plant-based treatment systems have consistently outperformed single-stage planted systems under varying influent qualities.
Table 3.
Design and operational drivers of viral attenuation in constructed wetlands.
| Factor/Strategy | Target Parameters | Key Action/Specification | Performance Metric | Mechanistic Rationale | Ref. |
|---|---|---|---|---|---|
| Hydraulic loading rate (HLR) | HLR & distribution uniformity | VSSF: 0.05–0.15 m³ m⁻² d⁻¹; perforated manifold dosing | ~1–2 log₁₀ removal; HRT = ~4–15 d | maximizes root–water contact and filtration efficiency; prevents short-circuiting | [59,154,198] |
| Flow configuration | flow pattern & dead-zone control | baffled FWS/staged islands; dispersion index >0.7 | HRT efficiency ~70–95%; channeling causes up to −30% loss | promotes plug flow (extended home time); increases uniform virion–biofilm interaction | [161,199] |
| Hydraulic retention time (HRT) | retention stability & redundancy | design: ~4–10 d (+20–40% safety margin for low T) | stable up to 3 log₁₀ removal | sustains contact time for adsorption/inactivation kinetics; reduces desorption risk | [123,162] |
| Rhizosphere aeration | intermittent air cycles & DO | ON/OFF ~1–2 h cycles; DO > 2 mg L⁻¹ at inlet. | enzyme gain ~20–45%; redox maintained (+50 to +200 mV) | boosts oxidative/enzymatic antiviral activity (peroxidases, ROS); prevents anoxic clogging | [165,200] |
| Redox / nutrient balance | C:N:P Ratio & ORP | C:N:P ≈ 100:10:1; ORP target: +100–+250 mV | infectivity loss 2.5 ± 0.4 log₁₀ | optimizes synthesis of antiviral exudates and enzymatic function; stabilizes microbial consortia | [169,171] |
| Temperature buffering | seasonal heat retention | raise water depth ~10–20% (winter); optional geothermal loop (<10°C differential) | keeps ~70–85% of summer rate; viral loss: –0.3 log₁₀ per 10°C drop | counteracts T-dependent reduction in enzymatic/adsorption kinetics; ensures year-round stability | [123,176] |
| Biomass management | harvest fraction & frequency | remove ~20–40% biomass ~2–4× yr⁻¹ | +0.5–0.8 log₁₀ improvement post-harvest | renews roots and exudation capacity (young plants are more active); prevents DOC release from senescence | [178,179] |
| Pretreatment (chemical) | charge neutralization & aggregation | FeCl₃ 5–20 mg L⁻¹ or EC ~1–2 mA cm⁻² | +1–1.5 log₁₀ viral gain (primary capture); +20% DOC tolerance. | strengthens primary capture by neutralizing negative virion charge; flocculation enhances settling/adsorption | [121,185] |
| Polishing/disinfection | secondary oxidation | UV ~30–60 mJ cm⁻²; ferrate ~0.5–1 mg L⁻¹; ozone ~0.2–0.5 mg L⁻¹ | ~5–7 log₁₀ total removal; low phytotoxicity | eliminates residual, recalcitrant infectivity (non-enveloped viruses); ensures safety for reuse standards | [188,191] |
For example, macrophyte–UV sequences convert stable ~2–4 log₁₀ reductions in the planted stages into ~3–7 log₁₀ overall removal, while reducing land requirements relative to plant-only configurations [56,193]. These benefits are maximised when upstream mixing and contact are well-engineered, and downstream polishing units are sized to residual pathogen loads rather than influent peaks [201,202].
UV and advanced oxidation process (AOP)-enhanced macrophyte systems
UV is a compact and energy-efficient polishing step that inactivates RNA and DNA viruses primarily via nucleic acid photodamage. Performing these systems depends on the delivered fluence, water transmittance, and shielding effects of particulates and coloured dissolved organic matter [203,204]. Typical UVC doses ranging from ~10–48 mJ·cm⁻² achieve complete inactivation of many enveloped viral surrogates and large reductions in more resistant non-enveloped viruses. For example, SARS-CoV-2 exhibits ~50% reduction within 1.4 min at laboratory irradiance levels [197,198,199]. When combined with planted beds, UV treatment addresses residual virions that persist following rhizosphere-mediated inactivation, effectively ensuring thorough removal [56,205]. Advanced oxidation processes (AOPs), applied pre- or post-UV, further improve performance by degrading UV-absorbing chromophores and reducing DOC, which competes for adsorption sites on roots and periphyton. Constructed wetland–ultraviolet (UV)–hydroxyl radical (HO·) pretreatment shows the concurrent removal of pesticides and total organic carbon (TOC), illustrating the enhancement of mass transfer and enzymatic efficacy downstream [206]. Designers must consider light-absorbing humic substances and melanoidins that reduce UV penetration and photochemical quenching in specific matrices. In such cases, modest coagulation or roughing filtration upstream of UV reactors can restore the transmittance [199,200].
Genome editing and synthetic biology for enhancing plant–microbe defense
Targeted genome editing and microbiome engineering offer promising routes to strengthen biological antiviral barriers, specifically rhizosphere inactivation and intracellular degradation, without compromising the hydraulic performance. CRISPR-Cas (Clustered Regularly Interspaced Short Palindromic Repeats [CRISPR]-associated [Cas] nuclease)-mediated editing in aquatic and wetland plants has enhanced antioxidant capacities, ion transport, and exudation profiles, traits associated with improved contaminant removal and stress tolerance, thus bolstering antiviral efficacy under cold or high DOC conditions [207,208]. Concurrent advances in synthetic biology have enabled the design of engineered rhizobacterial consortia that partition functions, such as lytic enzyme secretion, reactive oxygen species (ROS) generation, and extracellular polymeric substance production, to augment virion capture and capsid damage at the root interface [209,210]. At the system scale, these enhancements can reduce the reliance on UV/GAC polishing or decrease bed volume, thus improving the feasibility of space-constrained urban catchments [209,211]. Field deployment must navigate biosafety and regulatory challenges, including genetically modified organism (GMO) approval and environmental release tracking. Emerging risk mitigation strategies include pilot studies within containment and engineered microbial kill-switch circuits [212,213].
Digital/artificial intelligence (AI) tracking and scaling feasibility
Digital tracking technologies, including optical and DOC sensors coupled with rule-based control systems, have stabilised the hydraulics and redox conditions in planted beds. Early AI-assisted operational frameworks promise predictive control of flow equalisation, aeration duty cycles, and coagulant dosing; however, successful implementation requires attention to accessibility, local operator skill sets, and cyber-physical system reliability [214]. Scaling analyses have identified major non-technical barriers, such as land scarcity, fragmented governance, and financing mechanisms. Modularization and decentralisation of wastewater treatment (DEWATS) hybrid systems address footprint constraints and enable phased implementation, but community engagement and capacity building foster public acceptance and sustainable operation [215,216]. Fair deployment requires coupling technology choices with community co-design and long-term operation and maintenance (O&M) support to prevent critical parts such as UV lamps, sensors, and control software from becoming single points of failure “minimum viable hybridization” configurations (e.g., coagulation + planted bed + compact UV reactor) often offer ideal reliability-to-cost ratios in resource-limited settings, with advanced oxidation or membrane steps reserved for applications demanding stringent water reuse standards [215,216].
4. Practical Applications and Implementation
4.1. Constructed Wetland Systems (CWs)
Wetland configurations and hydraulic design
Constructed wetlands (CWs) are engineered ecosystems that integrate capture, rhizosphere inactivation, viral internalisation, and intracellular degradation under controlled flow regimes (Table 4). FWS cells combine light exposure, oxygenation, and photochemical pathways with plant–biofilm interfaces, but subsurface-flow (SSF) beds prioritise filtration and intimate water–root contact [217]. Unit-process wetlands show phototransformation and microbial degradation can be arranged sequentially to address the co-location of pathogens and trace organics in municipal effluents [218,219]. However, densely vegetated FWS systems may suffer from canopy shading and recirculation, which suppress photolysis and reduce first barrier contact, potentially resulting in less than 1 log₁₀ removal without hydraulic correction [202]. Horizontal SSF (HSSF) wetlands provide steady filtration but are prone to anoxic conditions. Column and sand-bed studies have reported diminished adsorption and slower inactivation under anaerobic conditions, which follows weaker electrostatic fixation and reduced oxidative chemistry [81,202,220]. VSSF designs, intermittently dosed via perforated manifolds, maintain aerobic microzones, a high specific surface area, and short diffusion distances, which help prevent surface clogging and sustain enzymatic activity [221]. Consequently, field practice combines well-distributed dosing, baffling, and staged cells to maintain an effective hydraulic retention time (HRT) near the design values and stabilise contact during fluctuations in organic matter or temperature [113,202]. Design heuristics recommend sizing the initial planted stage to achieve ~1–2 log₁₀ viral reduction via capture/filtration, followed by an additional ~0.5–1.0 bed volume to support rhizosphere and intracellular processes beyond threshold HRTs and efficiency plateaus, owing to pore saturation and biofilm maturation [67,202,222]. Cold-season operation benefits from modest HRT goes up and equalisation to counteract reduced reaction rates [223].
Substrate, vegetation, and rhizosphere dynamics
Substrate selection governs the hydraulics and microbiology of the system (Table 4). High-porosity, biofilm-supportive media increase the interfacial area and microbial colonisation, but appropriate hydraulic loading rates limit short-circuiting [224]. Steel-slag derivatives, particularly basic oxygen furnace slag, contribute to pathogen reduction via transient alkalinity (pH ≈ 10.6–11.4) and enhanced phosphorus capture; however, pH management and safety assessment are essential [225,226]. Alternate-day loading in VSSF wetlands promotes re-oxygenation and nitrification–denitrification cycling; during rest periods, redox rebounds and substrate re-oxygenation maintain enzyme activity supportive of antiviral chemistry [221].
Table 4.
System configurations and innovations for antiviral hybrid wetlands.
| Components | Configuration Description | Primary Antiviral Mechanisms | Demonstrated Performance | Innovation Value/Application | Ref. |
|---|---|---|---|---|---|
| FWS wetland | shallow vegetated channels (~0.3–0.6 m); open photic zone | photolysis, oxidation, biofilm sorption (low shear) | ~0.5–1.0 log₁₀ baseline; up to 2.0 with baffling | simple, low-cost system; sensitive to temperature and climate variability | [202,217] |
| HSSF wetland | saturated porous bed; laminar flow | filtration and anoxic biofilm degradation (stable pH) | 1.0 ± 0.3 log₁₀ (n=15); high stability across pH changes | filtration-dominant removal; good hydraulic control; limited oxidative capacity | [81,220] |
| VSSF wetland | intermittent dosing (alt-day); aerated percolation | adsorptive capture and oxidative decay on roots/media (high O₂) | ~2–3 log₁₀ at HRT ~5–10 d | high efficiency (~2–3× HSSF); reduces land area; requires mechanical dosing/aeration | [59,227] |
| Multistage hybrid CWs | sequential VSSF–FWS or VSSF–UV trains (multi-barrier approach) | combined filtration, oxidation, photolysis, enzymatic action | ~3–7 log₁₀ total removal (highest efficacy) | meets stringent reuse standards; functional redundancy buffers system failures | [113,161] |
| Substrate innovation | gravel, slag, zeolite, biochar, ferric media | enhanced adsorption; pH ~10–11 microzones; ROS generation | +10–30% extra removal from reactive layers | increases specific surface area; biochar adds catalytic/adsorptive properties; controls metal mobility | [224,225,226] |
| Vegetation selection | Phragmites, Typha, Ocimum, Strobilanthes (targeted functional traits). | O₂ release, enzyme induction, antiviral metabolite exudation | ~2–4 log₁₀ (field mean); 85–95% infectivity loss | shifts CWs from simple filtration to biochemically active reactors; cost-effective performance boost | [138,228] |
| Digital monitoring/AI | IoT sensors (DO, ORP, metabolites, microbial activity) | predictive control and early alerting (machine learning integration) | ~12–18 h lead time before viral breakthrough prediction | improves reliability/uptime; enables adaptive dosing/flow control; important for fluctuating loads. | [229,230] |
|
Synthetic biology integration |
engineered microbial consortia & biosensors (PGPR, lytic strains) | self-regulated enzymatic capture loops; enhanced proteolysis | +1–2 log₁₀ added potential (proof-of-concept) | high potential for targeted virus/pathogen removal; highly specific mechanism; requires regulatory acceptance | [137,231,232] |
Plant selection must balance hydraulic integrity and biochemical potency. Phragmites australis and Typha spp. provide aerenchyma-driven oxygen transfer, strong root mats, and stable periphyton, but aromatic and medicinal taxa enrich the rhizosphere with polyphenols and other antiviral metabolites [138,233]. Root-associated biofilms structured by exudates (i.e., organic acids, amino acids, and fatty acids) enhance nutrient cycling and contaminant attenuation, and plant growth-promoting rhizobacteria (PGPR) further coordinate division-of-labour metabolism, including lytic enzymes relevant to viral inactivation [41,224]. Polyculture wetlands leverage complementarity and functional redundancy by mixing structural macrophytes with antiviral-rich species, thus widening seasonal operating windows and buffering influent variability (see previous Table 2) [142,234]. Cold-climate adaptations include greenhouse enclosures, choice of plant species tolerant to low temperatures, and experimental use of CRISPR-edited macrophytes with enhanced stress tolerance. However, genome-edited plants require a biosafety review before open system deployment [223,228].
Operational management and digital tracking
Operational management should maintain an actively renewing surface state characterised by staged harvesting (~20–40% of aboveground biomass per event), removal of senescent litter, and periodic media maintenance to prevent pore blockage and anoxic mats [67,235]. Routine functional tracking, including dissolved oxygen, oxidation–reduction potential, pH, conductivity, and head loss, should be complemented by periodic enzyme activity assays (peroxidase and protease) or exudate markers to detect declines in rhizosphere reactivity before performance degradation [221,224]. Machine learning (ML) models trained on multi-seasonal plant, hydraulic, and chemical data have shown high predictive accuracy (R² ≈ 0.85–0.95) for nitrogen and chemical oxygen demand (COD) and benefit from virtual-sample augmentation to improve predictions under sparse field data conditions [229]. Root exudate biomarkers integrated with artificial intelligence (AI) have been proposed as early warning indicators for viral breakthrough (lead times of ~12–18 h), and field-deployable CRISPR–Cas12a assays can detect viral RNA within ~30 min to support event-driven operational controls [229,230]. These digital layers enable rule-based changes, including flow equalisation, aeration duty cycle modulation, and coagulant dosing, to have system performance despite influent variability [113,236]. Seasonal operational playbooks may enhance reliability. Winter mode reduces hydraulic loading rates by ~10–30%, increases water depth for thermal buffering, and lengthens rest periods, but summer mode tightens harvesting intervals and may incorporate brief daytime aeration to suppress anoxic mats [113].
Implementation and governance
Pilot projects in resource-limited contexts have shown the feasibility and community benefits of CWs, including livelihood co-benefits and local stewardship. Participatory projects in the Philippines illustrate how co-design and community operation and maintenance (O&M) improve system longevity and compliance [237]. Persistent adoption gaps come from regulatory exclusion, limited long-term pathogen tracking and fragmented governance. Updated guidance emphasising adaptive management and performance verification would accelerate mainstreaming [202,238]. Looking forward, synthetic biology offers tools for augmenting CW robustness. Engineered microbial consortia partition lytic enzyme production, reactive oxygen species generation, and extracellular polymer synthesis to enhance capture and inactivation, but biosensors and gene circuit sentinels provide online diagnostics [231,232]. Root exudate manipulation and microbiome engineering represent active frontiers with the potential to improve antiviral performance without compromising hydraulic performance [137]. A pragmatic implementation pathway involves minimum-viable hybridization such as low-dose coagulation upstream of planted beds combined with compact UV downstream, while concurrently developing data and governance frameworks that certify viral infectivity loss alongside conventional water quality metrics [137,138,202]. Collectively, these strategies translate mechanistic insights into deployable, resilient antiviral constructed wetlands characterised by hydraulically disciplined, biochemically active, digitally supervised, and socially embedded systems capable of delivering ~3–7 log₁₀ virus reduction when paired with polishing, where required [113,161,202].
4.2. Modular and Scalable Designs
Modular treatment architectures
Modular treatment enables the assembly of plant-based units integrated with compact physicochemical steps into site-specific modules that can be scaled, duplicated, or reconfigured as load demands and reuse targets evolve [239,240]. This approach aligns with decentralised wastewater (DEWATS) strategies applicable in dense urban and dispersed peri-urban contexts, facilitating phased implementation and plug-in upgrades that reduce capital risk and accelerate service time [107]. Governance analyses highlight the role of modularity in lowering institutional barriers by enabling performance-verified subunits, such as a planted bed combined with UV as a minimum viable hybrid which communities can adopt and expand as financing and operational capacity mature [107].
The design follows the antiviral barrier framework: an initial compact charge destabilisation or roughing step enhances early virion capture; planted units provide sorption–filtration and rhizosphere inactivation; and a downstream UV or granular activated carbon (GAC) module ensures the loss of infectivity for recalcitrant, non-enveloped viruses [241,242,243]. Standardised hydraulic and electrical interfaces between modules, including header geometry, bypass loops, and 24 V control systems, help with field assembly and maintenance by local operators [244,245]. Module sizing targets stable partial reductions of ~2–4 log₁₀ during the planting stage, with enough headroom for seasonal performance dips. Polishing units are sized to treat residual pathogen loads rather than influent peaks, optimising energy use and footprint [209,246].
Compact wetland designs for urban areas
Compact constructed wetlands, including intensified vertical subsurface flow (VSSF) beds, baffled FWS cells, and unit-process wetlands, maximise the specific interfacial area and narrow home time distributions, thus enhancing viral contact efficiency within limited land areas [99,209]. Urban pilot studies have shown using high-porosity media, multi-port influent dosing, and short flow paths maintains effective first-barrier capture and rhizosphere chemistry under varying organic matter loads. These effects are amplified when combined with downstream UV polishing [244,245,247]. Rooftop and podium-deck wetlands leverage gravity-fed pulsed dosing and solar exposure to support periphyton productivity, but green corridors along drainage channels and rights-of-way convert linear hydraulics into staged contact zones with minimal land-use conflicts [248,249]. To mitigate performance declines during cold seasons or high dissolved organic carbon (DOC) events, modular pre-treatment stages, such as coagulation or electrocoagulation cartridges, stabilise virion capture upstream of compact beds, but UV or GAC skids ensure regulatory compliance during peak viral loads [250]. The operations remained straightforward, with alternating day dosing preserving media porosity and oxygen supply. Quarterly tracer tests have verified that the effective hydraulic retention time (HRT) remains near the design value despite biofilm growth [251,252].
Mobile and adaptive treatment units
Trailer-mounted or containerised modular units provide rapid deployment capabilities suitable for outbreak response, festivals, or disaster relief, with flexibility for redeployment as demand shifts [244,253]. To operationalise mobile modules as decision-ready assets, rapid pathogen analytics, such as Reverse Transcription Recombinase Polymerase Amplification (RT-RPA)–CRISPR–Cas12a assays, enable viral RNA detection within ~30 min, integrating with low-power field readers for event-driven operational control [254,255,256]. These data streams support rule-based changes to flow equalisation, aeration duty cycles, and coagulant dosing, thus maintaining antiviral treatment efficacy despite episodic influent variability [216,257]. Given the variable influent chemistry in mobile deployments, interchangeable upstream destabilisation cartridges (i.e., coagulant or electrocoagulation) and downstream UV modules preserve reliability when non-enveloped viral surrogates predominate or when coloured dissolved organics attenuate UV transmittance [169,258].
Advanced materials and bio-digital rhizosphere enhancements
Additive manufacturing (3D printing) helps to produce root-zone scaffolds and advanced media with high, cleanable surface areas and engineered pore networks that resist compaction and clogging under pulsed hydraulic regimes, thus improving initial viral capture and periphyton stability within a compact footprint [259]. Incorporating redox-active fillers or slow-release micronutrients into these scaffolds sustains enzymatic activity and reactive oxygen species generation, which are critical for capsid and envelope damage without phytotoxic effects [108,260]. The synthetic biology and biosensing layers further enhance antiviral capacity and enable closed-loop control. Engineered plant–microbe consortia partition lytic enzyme secretion, extracellular polymeric substance production, and reactive oxygen species generation, while gene circuit-based biosensors provide real-time reporting of stress or viral surrogate signals [261,262]. In decentralised contexts, these biosignals can tell simple control systems that autonomously adjust dosing or aeration, complementing modular governance frameworks [263,264,265]. A pragmatic implementation pathway involves “minimum-viable hybridization” constructed from standardised modules: upstream destabilisation via cartridge coagulation or mild electrocoagulation, a compact planted bed (i.e., intensified VSSF or baffled FWS), and a small ultraviolet (UV) skid. This configuration is equity-friendly, scalable, and capable of achieving ~3–7 log₁₀ overall viral reductions when tuned to residual pathogen loads rather than influent peaks [75,205,266].
4.3. Hybrid Treatment Systems and Economic Considerations
Cost–performance and governance alignment
Hybrid treatment integrates plant-based barriers with compact physicochemical processes to convert stable partial reductions achieved in planted units, such as viral capture and rhizosphere inactivation, into compliance-grade removals while minimising land use and operational complexity [108,266]. Specifying treatment targets based on established reuse and effluent standards explains the critical final stage of polishing work by hybrid systems particularly in agricultural reuse or urban unpotable application [267,268]. In resource constrained environments, technical limitations and fragmented governance often impede the implementation of one step wastewater treatment systems. Modular hybridisation reduces these risks and shares performance requirements amongst smaller, verifiable units [267,268]. Economically, hybrid systems affect the cost curve favourably by diminishing the treatment footprint (and attending land costs) and normalising seasonal performance, negating the need for over-sizing to accommodate winter lows. In land constrained settings compact pretreatment combined with vertical subsurface flow (VSSF) constructed wetlands and ultraviolet (UV) disinfection usually minimises total lifecycle costs for a given viral log-reduction goal [205,269,270]. Governance mechanisms, for example clear reuse standards, performance-based procurement and community-based operation and maintenance (O&M) contracts enhance the feasibility still further by aligning the incentives between regulatory agencies and operators [270,271].
Plant-based and physicochemical hybrid systems
Field and pilot-scale studies have shown macrophyte treatment stages constructed with Typha spp. and Phragmites spp. could achieve ~2.0–3.5 log₁₀ reductions in viral surrogates under optimised hydraulic conditions. Subsequent UV disinfection typically provides an additional ~1–3 log₁₀ reduction, depending on the UV fluence and water transmittance [75,272]. Floating wetland configurations employing Eichhornia crassipes combined with compact UV units have shown further ~1.5–2.1 log₁₀ reductions for norovirus surrogates while maintaining low capital expenditure (CAPEX) and enabling biomass valorization [273,274,275]. Where influent turbidity and coloured dissolved organic carbon (DOC) reduce UV effectiveness, upstream light-touch charge destabilisation via optimised coagulant dosing or mild electrocoagulation enhances early viral capture and restores downstream UV efficiency [276,277]. For applications requiring higher treatment assurance, electrochemically enhanced membrane bioreactors (e-MBRs) have been reported to achieve ~2.8–5.0 log₁₀ reductions in SARS-CoV-2 through combined filtration and in situ oxidation. When deployed as polishing units following the planting stages, e-MBRs can be downsized to treat residual viral loads, thus preserving energy efficiency [7,196]. The design sequence leverages mechanistic complementarity: upstream destabilisation for viral capture, planted beds for biochemical inactivation, and UV, advanced oxidation processes (AOP), or membrane bioreactors for losing infectivity [278].
Financing, operation and maintenance (O&M) economics, and scale-up
Hybridisation helps with stepwise financing approaches, beginning with a “minimum-viable hybrid” (e.g., coagulation cartridge, planted bed, and small UV unit) and allowing the incremental addition of modules as reuse standards tighten or influent flows go up [279]. Blended financing mechanisms, including municipal budgets combined with green bonds, microfinance for community O&M, and public-private partnerships (PPP), help bridge CAPEX gaps while securing maintenance commitments [280]. Lifecycle cost assessments should incorporate avoided land purchase costs, reductions in winter over-sizing, and revenues or cost offsets from biomass valorization products such as compost or biochar, thus improving the net present value of plant-centric treatment systems [7,196]. System reliability depends on lean O&M practices: periodic harvesting, inlet rotation, cartridge replacement for pretreatment units, UV sleeve cleaning, lamp maintenance on monthly or quarterly schedules, and simple sensor-based feedback loops (dissolved oxygen [DO], oxidation-reduction potential [ORP], head loss) to trigger rule-based operational changes [113,281,282,283]. Incorporating digital tracking layers can further reduce operational risks by forecasting influent perturbations and optimising set points if these tools remain accessible to local operators to prevent the introduction of new failure modes [113,284].
Equity, biosafety, and technological innovation
The fair deployment of hybrid treatment systems requires modular governance frameworks coupled with context-sensitive technology selection, including appropriately sized UV units, affordable consumables, and avoidance of dependence on reagents difficult to obtain [285,286,287]. Emerging synthetic biology innovations, such as engineered plant–microbe consortia that partition lytic enzymes, reactive oxygen species (ROS) generation, and extracellular polymer production, have the potential to enhance planted-stage viral removal performance and reduce polishing requirements. However, these approaches require rigorous biosafety assessments, continuous tracking, and, where appropriate, incorporating genetic kill switches [285,288,289]. Integrating biotechnological advances with pragmatic financial and operational strategies, including operator training, spare part logistics, and community stewardship, yields resilient and socially embedded wastewater treatment systems [290,291,292]. Collectively, these pathways enable antiviral hybrid treatment systems that are hydraulically disciplined, biochemically active, and economically viable across diverse income settings, achieving ~3–7 log10 overall viral reductions when sequenced and sized to treat residual loads rather than peak influent concentrations [278,293].
5. Challenges and Prospects
5.1. Current Challenges and Persistent Gaps
Variability of rhizosphere antiviral activity
Antiviral phytoremediation is limited by the intrinsic stability of many non-enveloped viruses and the variable nature of plant–microbe antiviral interactions. Enveloped virions are generally susceptible to oxidative chemistry and enzymatic degradation in the rhizosphere, but non-enveloped particles with strong icosahedral capsids often persist, resulting in only ~1–3 log₁₀ reduction without supplementary treatment steps [294,295,296]. This structural asymmetry implies that systems relying only on physical capture mechanisms, such as roots, biofilms, and porous media, are vulnerable to desorption or downstream breakthrough under fluctuating hydraulic conditions [297,298,299]. The antiviral activity of the rhizosphere varies constantly and depends on environmental conditions such as temperature, nutrient supply and redox conditions. These environmental variables influence the synthesis of antiviral metabolites and lytic enzymes, resulting in seasonal plateaux in viral inactivity despite adequate contact time [300,301]. For example, in the colder seasons and in influent with high dissolved organic carbon (DOC) there will be reduced exudation rates and enzyme turnover, reducing the strength of the intracellular degradation routes which are extra to those providing physical capture [302,303,304]. The diversity of viruses complicates predictive modelling and system design. The kinetics of persistence and inactivation is different between strains of virus (e.g., rotavirus vs. enterovirus) such that plant–microbe assemblages designed for one surrogate virus may perform poorly against others unless the systems are made to incorporate together multilayer effects [226,305]. These biological constraints show that hybrid systems and modes of operation should be used in winter and seasonal work rather than in single barrier effects [278,306,307].
Engineering and operational constraints
Constructed antiviral phytoremediation systems are limited by effective contact between viruses and reactive surfaces. Even minor dead zones or hydraulic short-circuiting can reduce effective hydraulic retention time (HRT), reducing first-barrier viral removal from less than ~1–2 log₁₀, despite favourable plant traits [278,308,309]. Clogging phenomena, including inlet obstruction, pore occlusion by fines, and biofilm aging, reduce media porosity and interfacial area, thus lowering virus capture and suppressing rhizosphere activity. Remediation of hydraulic performance typically requires disruptive media maintenance, which is rarely incorporated into routine operational planning or budgeting [40,238]. Material selection involves critical trade-offs between various factors. Alkaline industrial by-products and reactive substrates may transiently enhance pathogen reduction, but require precise pH control to avoid phytotoxic effects and regulatory non-compliance downstream [310,311]. But biologically favourable media with a high specific surface area can accelerate clogging if the dosing and harvesting protocols are not optimised to maintain a “young active surface” [113,312]. Operational discipline is paramount, and strategies such as alternating day dosing in vertical subsurface-flow beds, seasonal changes in hydraulic loading rates (HLR), selective harvesting, and tracer-based verification of HRT are rarely standardised beyond research pilots. This disconnect between laboratory-grade performance and long-term field reliability hampers confidence in antiviral phytoremediation for water reuse applications [113,313].
Tracking, standards, and knowledge gaps
Many demonstration studies have reported reductions in viral molecular signals (RNA/DNA) but have not assessed infectivity, which is the definitive measure of viral removal efficacy [314,315]. Standardised protocols that integrate surrogate virus selection, dose–response relationships, and side-by-side infectivity assays remain limited in long-term field deployments, complicating cross-study comparisons and meta-analyses [316]. Economic and governance data are also limited. Few investigations provide life-cycle cost analyses encompassing seasonal operational modes, routine media maintenance, and analytical tracking, thus obscuring budgeting and impeding adoption in decentralised systems, where modular hybrid designs are most effective [293,317]. The absence of widely accepted verification frameworks, such as minimum HRT testing, enzyme activity proxy panels, and event-driven ultraviolet (UV) backstops, further widens the gap between laboratory validation and field application [318,319]. Pathways for translating promising enhancements, such as biochar amendments that stabilise redox conditions and adsorptive capacity, or synthetic biology approaches that reinforce rhizosphere and intracellular antiviral barriers, into permitting and biosafety frameworks for open systems remain undefined [320,321]. Addressing these gaps through the development of infectivity-based tracking protocols, operational playbooks, and regulatory guidance for novel materials and engineered microbial consortia is critical for the mainstream adoption of antiviral phytoremediation technologies.
5.2. Prospects and Emerging Innovations
Genetic engineering to enhanced antiviral activities
Genetic engineering offers targeted strategies to enhance the two primary biological barriers in antiviral phytoremediation, rhizosphere inactivation, and intracellular degradation, without compromising hydraulic performance. Building on foundational phytoremediation principles, CRISPR–Cas genome editing can modulate the antioxidant capacity, exudation profiles, and proteostasis pathways, which are critical for capsid damage and viral genome cleavage [322]. Plants engineered for increased late-endosomal restriction, reactive oxygen species (ROS) generation, and protease/ribonuclease (RNase) activity are poised to reduce the contact time required for enveloped viruses, with potential benefits extending to more resilient non-enveloped viruses. Complementary approaches include the expression of antiviral decoy receptors or lectin-like proteins that bind viral surface motifs extracellularly, promoting aggregation and capture within the root or periphyton matrix before internalisation [323,324]. The localisation of these proteins in root exudates or apoplastic spaces may minimise growth penalties while intercepting virions at critical interfaces in planted treatment systems. Translational research should prioritise bench-to-pilot pipelines that link genetic modifications to measurable barrier-specific endpoints, such as enzyme activity proxies and infectivity loss assays for both enveloped and non-enveloped surrogate viruses, under environmentally relevant conditions of ionic strength, dissolved organic carbon (DOC), and temperature [325].
Biosafety and biocontainment strategies
Genetic enhancement programs must integrate strong and effective biocontainment strategies. These include conditional containment systems, sterile triploid or polyploid cultivars, and chloroplast-targeted transgenes, which minimise gene flow without compromising plant vigour or remediation efficacy [326,327]. Spatial confinement via biofilm-centric cultivation and physical barriers can further restrict dispersal while supporting dense periphyton communities that enhance antiviral activity [328]. Risk assessment frameworks should incorporate off-target effect evaluations, ecological interaction studies, staged field releases, environmental DNA tracking, and rollback plans, treating engineered consortia as time-limited interventions subject to periodic re-licencing [329]. Ethical oversight and community consent models developed for nature-based solutions provide valuable frameworks for transparent decision-making and shared stewardship during pilot deployments [330,331]. Governance toolkits that combine technical safeguards (e.g., sterility and genetic kill-switches), operational controls (e.g., harvest scheduling and inlet rotation), and social contracts (e.g., community operation and maintenance agreements) will help with responsible scaling from greenhouse experiments to neighbourhood-scale implementations [332,333].
Advanced materials and autonomous systems
Next-generation media and root-zone scaffolds, fabricated via additive manufacturing to provide tunable porosity gradients and high, cleanable surface areas, can enhance early viral capture while resisting compaction and clogging under variable hydraulic loading [334]. Incorporating redox-active fillers or micronutrient dopants sustains enzyme production and ROS generation, thus boosting rhizosphere antiviral activity without inducing phytotoxicity [335,336]. Biomimetic self-cleaning surfaces inspired by lotus leaf morphology show promise for fouling resistance in compact pretreatment and polishing cartridges positioned upstream or downstream of the planted units [337,338]. Artificial intelligence (AI)-enabled tracking is emerging as a critical control layer for modular and decentralised treatment systems. Edge-computing sensor networks tracking dissolved oxygen (DO), oxidation-reduction potential (ORP), head loss, and proxies for exudate or enzyme activity can feed predictive models to expect clogging and viral breakthrough events, enabling condition-based maintenance that reduces downtime by ~30–60% [338,339]. Techno-economic forecasting and adaptive operational dispatch—adjusting ultraviolet (UV) or coagulant dosing based on residual rather than influent peaks—can lower operational spending while maintaining regulatory compliance during seasonal performance lows [278,340]. In land-constrained settings, intensified VSSF units, baffled free-water surface cells, and minimum-viable hybrid systems (combining destabilisation, planted beds, and small UV units) give scalable and reliable building blocks simplified failure modes, suitable for peri-urban corridors and informal settlements [99].
Policy, climate resilience and circular value
Policy alignment is important for the widespread adoption of antiviral phytoremediation techniques. Harmonised verification protocols, including minimum hydraulic retention time (HRT) testing, infectivity-based endpoints, and enzyme activity proxy panels, integrated into state manuals and international guidance documents, would bridge the gap between laboratory validation and field application [1,341]. Climate resilience requires both biological and infrastructural adaptation. Drought-tolerant species, such as Vetiveria zizanoides and Pennisetum purpureum, sustain fine-root areas and exudation during water scarcity, but salt-tolerant taxa and hybrid desalination–phytoremediation systems address the challenges of coastal intrusion [342]. Circular economy strategies, including biochar production from macrophyte biomass (i.e., surface area ~200–400 m²/g), nutrient recovery, and the development of local soil amendment markets, enhance the lifecycle value and stabilise redox and adsorption properties when applied judiciously as media amendments [343,344]. Integrated roadmaps combining technology, finance, and policy—starting with minimum-viable hybrid systems and scaling via modular blocks under community operation and maintenance (O&M) and transparent performance verification offer the most credible pathway toward fair, climate-resilient antiviral phytoremediation deployment [345].
6. Conclusions
Antiviral phytoremediation is a promising method for removing viruses in wastewater owing to its complex series of multi-defense mechanisms (i.e., sorption-filtration, rhizosphere-mediated inactivation, viral internalisation, and intracellular degradation. Collectively, these mechanisms could convert hydraulic contact into loss of viral infectivity. Enveloped virions are more susceptible to rhizosphere chemicals and intracellular defence mechanisms but non-enveloped viruses with strong capsids present problems. Planted treatment measures lead to ~2–4 log₁₀ reduction in viral numbers, while their use with UV irradiation or GAC enables ~3–7 log₁₀ reduction without unacceptable growth of treatment footprint. Efficient system design depends on even flow distribution through the bed crown, verified HRT, and active interfacial surface area for initial ~1–2 logs of viral removal. Stabilising treatment rhizosphere chemistry requires balanced C:N:P ratios, periodic aeration, and seasonal operation modes to maintain enzyme activity. Optimising the polishing step, based on residual viral load, involves calibrated fluence level UV or compact sorptive units to reduce non-enveloped viruses. For decentralised applications, a system based on minimum viable hybrid modules would use cartridge coagulation, planted beds, UV units to simplify O&M and enable incremental adoption through standard interfaces. Future goals include viral infectivity tracking protocols, establishing operational basis for equipment maintenance through seasonal variations, and integrating new materials and autonomous systems in regulatory frameworks for reliable antiviral phytoremediation. This would create a system with satisfactory laboratory and field reliability, enabling worldwide water reuse and pestilential control implementation.
Author Contributions
DZ contributed to conceptualization, investigation, analysis, visualization, writing—original draft, and writing—review & editing. M-H.S, AM & EM contributed to writing—review & editing. AD contributed to supervision and writing—review & editing. DH-W.K contributed to supervision, visualization, writing—review & editing, resources, project administration, and funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Ministry of Science and Technology, Taiwan (MOST 105-2221-E-029-002 and 106-2221-E-029-003).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
This study was conducted at the Department of Environmental Science and Engineering, Tunghai University, with gratitude to the School of Forestry, PNG University of Technology, for providing space and resources for data analysis and writing.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AOPs | Advanced Oxidation Processes |
| AI | Artificial Intelligence |
| CAPEX | Capital Expenditure |
| C:N:P | Carbon:Nitrogen:Phosphorus Ratio |
| COD | Chemical Oxygen Demand |
| CRISPR | Clustered Regularly Interspaced Short Palindromic Repeats |
| CRISPR-Cas | Clustered Regularly Interspaced Short Palindromic Repeats [CRISPR]-associated [Cas] nuclease |
| CWs | Constructed Wetland Systems |
| DEWATS | Decentralisation of Wastewater Treatment |
| DO | Dissolved Oxygen |
| DOC | Dissolved Organic Carbon |
| EC | Electrocoagulation |
| e-MBRs | Electrochemically Enhanced Membrane Bioreactors |
| EPS | Extracellular Polymeric Substance |
| FWS | Free-Water Surface (or Free-Water Surface cells/systems) |
| GAC | Granular Activated Carbon |
| GH19 | Glycoside Hydrolase Family 19 |
| GMO | Genetically Modified Organism |
| HAA | Haloacetic Acids |
| HLR | Hydraulic Loading Rate |
| HO· | Hydroxyl Radical |
| HRT | Hydraulic Retention Time |
| HSSF | Horizontal Subsurface-Flow |
| IC₅₀ | Inhibitory Concentration 50% |
| IFITM3 | Interferon-Induced Transmembrane Protein 3 |
| MBRs | Membrane Bioreactors |
| ML | Machine Learning |
| O&M | Operation and Maintenance |
| ORP | Oxidation-Reduction Potential |
| PGPR | Plant Growth-Promoting Rhizobacteria |
| PPP | Public-Private Partnerships |
| RNA | Ribonucleic Acid |
| RNAi | RNA Interference |
| RNase | Ribonuclease |
| ROS | Reactive Oxygen Species |
| RT-RPA | Reverse Transcription Recombinase Polymerase Amplification |
| SA | Salicylic Acid |
| SARS-CoV-2 | Severe Acute Respiratory Syndrome Coronavirus 2 |
| SSF | Subsurface-Flow |
| THM | Trihalomethanes |
| TOC | Total Organic Carbon |
| UV | Ultraviolet |
| UVC | Ultraviolet C |
| VSSF | Vertical Subsurface-Flow |
References
- Carmo Dos Santos, M.; Cerqueira Silva, A.C.; Dos Reis Teixeira, C.; Pinheiro Macedo Prazeres, F.; Fernandes Dos Santos, R.; de Araújo Rolo, C.; de Souza Santos, E.; Santos da Fonseca, M.; Oliveira Valente, C.; Saraiva Hodel, K.V.; et al. Wastewater Surveillance for Viral Pathogens: A Tool for Public Health. Heliyon 2024, 10, e33873. [Google Scholar] [CrossRef]
- Ahmad, J.; Ahmad, M.; Usman, A.R.A.; Al-Wabel, M.I. Prevalence of Human Pathogenic Viruses in Wastewater: A Potential Transmission Risk as Well as an Effective Tool for Early Outbreak Detection for COVID-19. J. Environ. Manage. 2021, 298, 113486. [Google Scholar] [CrossRef]
- Bivins, A.; Greaves, J.; Fischer, R.; Yinda, K.C.; Ahmed, W.; Kitajima, M.; Munster, V.J.; Bibby, K. Persistence of SARS-CoV-2 in Water and Wastewater. Environ. Sci. Technol. Lett. 2020, 7, 937–942. [Google Scholar] [CrossRef]
- Girón-Guzmán, I.; Falcó, I.; Cuevas-Ferrando, E.; Ballesteros, S.; Barranquero, R.; Sánchez, G. Survival of Viruses in Water Microcosms. Science of The Total Environment 2025, 963, 178416. [Google Scholar] [CrossRef]
- Kahler, A.M.; Cromeans, T.L.; Roberts, J.M.; Hill, V.R. Effects of Source Water Quality on Chlorine Inactivation of Adenovirus, Coxsackievirus, Echovirus, and Murine Norovirus. Appl. Environ. Microbiol. 2010, 76, 5159–5164. [Google Scholar] [CrossRef]
- Lanrewaju, A.A.; Enitan-Folami, A.M.; Sabiu, S.; Swalaha, F.M. A Review on Disinfection Methods for Inactivation of Waterborne Viruses. Frontiers in Microbiology 2022, 13. [Google Scholar] [CrossRef]
- Chaudhry, R.M.; Nelson, K.L.; Drewes, J.E. Mechanisms of Pathogenic Virus Removal in a Full-Scale Membrane Bioreactor. Environ. Sci. Technol. 2015, 49, 2815–2822. [Google Scholar] [CrossRef]
- Chen, C.; Guo, L.; Yang, Y.; Oguma, K.; Hou, L. Comparative Effectiveness of Membrane Technologies and Disinfection Methods for Virus Elimination in Water: A Review. Science of The Total Environment 2021, 801, 149678–149678. [Google Scholar] [CrossRef]
- Krzeminski, P.; van der Graaf, J.H.J.M.; van Lier, J.B. Specific Energy Consumption of Membrane Bioreactor (MBR) for Sewage Treatment. Water Sci. Technol. 2012, 65, 380–392. [Google Scholar] [CrossRef]
- Skouteris, G.; Arnot, T.C.; Jraou, M.; Feki, F.; Sayadi, S. Modeling Energy Consumption in Membrane Bioreactors for Wastewater Treatment in North Africa. Water Environment Research 2014, 86, 232–244. [Google Scholar] [CrossRef]
- Adegoke, A.A.; Amoah, I.D.; Stenström, T.A.; Verbyla, M.E.; Mihelcic, J.R. Epidemiological Evidence and Health Risks Associated With Agricultural Reuse of Partially Treated and Untreated Wastewater: A Review. Front. Public Health 2018, 6, 337. [Google Scholar] [CrossRef]
- Haramoto, E.; Malla, B.; Thakali, O.; Kitajima, M. First Environmental Surveillance for the Presence of SARS-CoV-2 RNA in Wastewater and River Water in Japan. Science of The Total Environment 2020, 737, 140405. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, V.; Kapoor, D.; Dhanjal, D.S.; Bhatia, D.S.; Jan, S.; Singh, N.; Romero, R.; Ramamurthy, P.C.; Singh, J. Detection and Disinfection of COVID-19 Virus in Wastewater. Environmental Chemistry Letters 2021, 19, 1917–1933. [Google Scholar] [CrossRef]
- Tandukar, S.; Sthapit, N.; Thakali, O.; Malla, B.; Sherchan, S.P.; Shakya, B.M.; Shrestha, L.P.; Sherchand, J.B.; Joshi, D.R.; Lama, B.; et al. Detection of SARS-CoV-2 RNA in Wastewater, River Water, and Hospital Wastewater of Nepal. Sci. Total Environ. 2022, 824, 153816. [Google Scholar] [CrossRef]
- Shingare, R.P.; Nanekar, S.V.; Thawale, P.R.; Karthik, R.; Juwarkar, A.A. Comparative Study on Removal of Enteric Pathogens From Domestic Wastewater Using Typha Latifolia and Cyperus Rotundus Along With Different Substrates. 2017, 19, 899–908. [CrossRef]
- Rosendo, J.C.M.; da Paz, G.M.; Rosendo, A. Constructed Wetlands Applied on Domestic Wastewater for Decetralized Systems: Concepts, Processes, Modalities, Combinations and Enhancements; A Review. 2022, 21, 371–397. [CrossRef]
- Wang, L.; Ma, L.; Wang, J.; Zhao, X.; Jing, Y.; Liu, C.; Xiao, Y.; Li, C.; Jiao, C.; Xu, M. Research Progress on the Removal of Contaminants from Wastewater by Constructed Wetland Substrate: A Review. Water 2024, 16, 1848. [Google Scholar] [CrossRef]
- Li, X.-F.; Mitch, W.A. Drinking Water Disinfection Byproducts (DBPs) and Human Health Effects: Multidisciplinary Challenges and Opportunities. Environ. Sci. Technol. 2018, 52, 1681–1689. [Google Scholar] [CrossRef]
- Salt, D.E.; Smith, R.D.; Raskin, I. PHYTOREMEDIATION. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1998, 49, 643–668. [Google Scholar] [CrossRef]
- Pilon-Smits, E. PHYTOREMEDIATION. Annu. Rev. Plant Biol. 2005, 56, 15–39. [Google Scholar] [CrossRef]
- Fletcher, J.; Willby, N.; Oliver, D.M.; Quilliam, R.S. Phytoremediation Using Aquatic Plants. In Phytoremediation; Shmaefsky, B.R., Ed.; Concepts and Strategies in Plant Sciences; Springer International Publishing: Cham, 2020; pp. 205–260. ISBN 978-3-030-00098-1. [Google Scholar]
- Williams, J.B. Phytoremediation in Wetland Ecosystems: Progress, Problems, and Potential. Critical Reviews in Plant Sciences 2002, 21, 607–635. [Google Scholar] [CrossRef]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The Role of Root Exudates in Rhizosphere Interactions with Plants and Other Organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef]
- Sourav Kumar, P.; Surajit, D. Potential of Plant Growth-Promoting Microbes for Improving Plant and Soil Health for Biotic and Abiotic Stress Management in Mangrove Vegetation. Rev. Environ. Sci. Biotechnol. 2024, 23, 801–837. [Google Scholar] [CrossRef]
- Daud, M.K.; Ali, S.; Abbas, Z.; Zaheer, I.E.; Riaz, M.A.; Malik, A.; Hussain, A.; Rizwan, M.; Zia-ur-Rehman, M.; Zhu, S.J. Potential of Duckweed ( Lemna Minor ) for the Phytoremediation of Landfill Leachate. Journal of Chemistry 2018, 2018, 1–9. [Google Scholar] [CrossRef]
- Khan, A.U.; Khan, A.N.; Waris, A.; Ilyas, M.; Zamel, D. Phytoremediation of Pollutants from Wastewater: A Concise Review. Open Life Sciences 2022, 17, 488–496. [Google Scholar] [CrossRef]
- Aryal, M. Phytoremediation Strategies for Mitigating Environmental Toxicants. Heliyon 2024, 10, e38683. [Google Scholar] [CrossRef]
- Patra, A.K. An Overview of Antimicrobial Properties of Different Classes of Phytochemicals. In Dietary Phytochemicals and Microbes; Springer Netherlands: Dordrecht, 2012; pp. 1–32. ISBN 978-94-007-3925-3. [Google Scholar]
- Othman, L.; Sleiman, A.; Abdel-Massih, R.M. Antimicrobial Activity of Polyphenols and Alkaloids in Middle Eastern Plants. Front. Microbiol. 2019, 10, 911. [Google Scholar] [CrossRef]
- Dangl, J.L.; Jones, J.D. Plant Pathogens and Integrated Defence Responses to Infection. Nature 2001, 411, 826–833. [Google Scholar] [CrossRef]
- Nürnberger, T.; Lipka, V. Non-Host Resistance in Plants: New Insights into an Old Phenomenon. Mol. Plant Pathol. 2005, 6, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.A.; Lee, H.Y.; Seo, E.; Lee, J.; Kim, S.B.; Oh, S.; Choi, E.; Choi, E.; Lee, S.E.; Choi, D. Current Understandings of Plant Nonhost Resistance. Molecular Plant-Microbe Interactions 2017, 30, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Hunag, Z.; Wang, X.; Cui, L. Pennisetum Sinese Roxb and Pennisetum Purpureum Schum. as Vertical-Flow Constructed Wetland Vegetation for Removal of N and P from Domestic Sewage. Ecological Engineering 2015, 83, 120–124. [Google Scholar] [CrossRef]
- Tang, W.-F.; Tsai, H.-P.; Chang, Y.-H.; Chang, T.-Y.; Hsieh, C.-F.; Lin, C.-Y.; Lin, G.-H.; Chen, Y.-L.; Jheng, J.-R.; Liu, P.-C.; et al. Perilla (Perilla Frutescens) Leaf Extract Inhibits SARS-CoV-2 via Direct Virus Inactivation. Biomedical Journal 2021, 44, 293–303. [Google Scholar] [CrossRef]
- Chen, J.; Zhao, Y.; Cheng, J.; Wang, H.; Pan, S.; Liu, Y. The Antiviral Potential of Perilla Frutescens: Advances and Perspectives. Molecules 2024, 29, 3328. [Google Scholar] [CrossRef]
- Hook, D.W.; Porter, S.J.; Herzog, C. Dimensions: Building Context for Search and Evaluation. Front. Res. Metr. Anal. 2018, 3, 23. [Google Scholar] [CrossRef]
- Guerrero-Bote, V.P.; Chinchilla-Rodríguez, Z.; Mendoza, A.; de Moya-Anegón, F. Comparative Analysis of the Bibliographic Data Sources Dimensions and Scopus: An Approach at the Country and Institutional Levels. Front. Res. Metr. Anal. 2020, 5, 593494. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, P.; Karmakar, M.; Leta, J.; Mayr, P. The Journal Coverage of Web of Science, Scopus and Dimensions: A Comparative Analysis. Scientometrics 2021, 126, 5113–5142. [Google Scholar] [CrossRef]
- Kadlec, R.H.; Wallace, S. Treatment Wetlands; 0 ed.; CRC Press, 2008. ISBN 978-0-429-13795-2.
- Vymazal, J. Constructed Wetlands for Wastewater Treatment: Five Decades of Experience. 2010, 45, 61–69. [CrossRef]
- Saeed, T.; Sun, G. A Review on Nitrogen and Organics Removal Mechanisms in Subsurface Flow Constructed Wetlands: Dependency on Environmental Parameters, Operating Conditions and Supporting Media. 2012, 112, 429–448. [CrossRef]
- Tang, P.; Yu, B.; Zhou, Y.; Zhang, Y.; Li, J. Clogging Development and Hydraulic Performance of the Horizontal Subsurface Flow Stormwater Constructed Wetlands: A Laboratory Study. Environ Sci Pollut Res 2017, 24, 9210–9219. [Google Scholar] [CrossRef]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. THE ROLE OF ROOT EXUDATES IN RHIZOSPHERE INTERACTIONS WITH PLANTS AND OTHER ORGANISMS. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef]
- García-Ávila, F. Treatment of Municipal Wastewater by Vertical Subsurface Flow Constructed Wetland: Data Collection on Removal Efficiency Using Phragmites Australis and Cyperus Papyrus. Data in Brief 2020, 30, 105584. [Google Scholar] [CrossRef]
- García-Ávila, F. Treatment of Municipal Wastewater by Vertical Subsurface Flow Constructed Wetland: Data Collection on Removal Efficiency Using Phragmites Australis and Cyperus Papyrus. Data in Brief 2020, 30. [Google Scholar] [CrossRef]
- Kadlec, R.H.; Wallace, S. Treatment Wetlands. 2008. [CrossRef]
- Das, S.K.; Ghosh, G.K.; Avasthe, R. Biochar Application for Environmental Management and Toxic Pollutant Remediation. Biomass Convers. Biorefin. 2023, 13, 555–566. [Google Scholar] [CrossRef]
- Liu, H.; Hu, Z.; Zhang, J.; Ngo, H.H.; Guo, W.; Liang, S.; Fan, J.; Lu, S.; Wu, H. Optimizations on Supply and Distribution of Dissolved Oxygen in Constructed Wetlands: A Review. Bioresource Technology 2016, 214, 797–805. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Srivastava, A.K.; Rajput, V.D.; Chauhan, P.K.; Bhojiya, A.A.; Jain, D.; Chaubey, G.; Dwivedi, P.; Sharma, B.; Minkina, T. Root Exudates: Mechanistic Insight of Plant Growth Promoting Rhizobacteria for Sustainable Crop Production. Front. Microbiol. 2022, 13, 916488. [Google Scholar] [CrossRef]
- López-Vinent, N.; Cruz-Alcalde, A.; Santacruz, A.P.; Sans, C. Green Approach for Micropollutants Removal: Study of Constructed Wetlands as Pretreatment of Solar Photo-Fenton Catalyzed by Organic Fertilizers. Catal. Today 2024, 430, 114540. [Google Scholar] [CrossRef]
- Ruttens, A.; Boulet, J.; Weyens, N.; Smeets, K.; Adriaensen, K.; Meers, E.; Van Slycken, S.; Tack, F.; Meiresonne, L.; Thewys, T.; et al. Short Rotation Coppice Culture of Willows and Poplars as Energy Crops on Metal Contaminated Agricultural Soils. International Journal of Phytoremediation 2011, 13, 194–207. [Google Scholar] [CrossRef]
- Tiodar, E.D.; Văcar, C.L.; Podar, D. Phytoremediation and Microorganisms-Assisted Phytoremediation of Mercury-Contaminated Soils: Challenges and Perspectives. IJERPH 2021, 18, 2435. [Google Scholar] [CrossRef]
- Manorama Thampatti, K.C.; Beena, V.I.; Meera, A.V.; Ajayan, A.S. Phytoremediation of Metals by Aquatic Macrophytes. In Phytoremediation; Shmaefsky, B.R., Ed.; Concepts and Strategies in Plant Sciences; Springer International Publishing: Cham, 2020; pp. 153–204. ISBN 978-3-030-00098-1. [Google Scholar]
- Gersberg, R.M.; Gearheart, R.A.; Ives, M. Pathogen Removal in Constructed Wetlands. In Constructed Wetlands for Wastewater Treatment; Hammer, D.A., Ed.; CRC Press, 2020; pp. 431–445. ISBN 978-1-003-06985-0.
- Gersberg, R.M.; Lyon, S.R.; Brenner, R.; Elkins, B.V. Fate of Viruses in Artificial Wetlands. Appl Environ Microbiol 1987, 53, 731–736. [Google Scholar] [CrossRef]
- Jasper, J.T.; Nguyen, M.T.; Jones, Z.L.; Ismail, N.S.; Sedlak, D.L.; Sharp, J.O.; Luthy, R.G.; Horne, A.J.; Nelson, K.L. Unit Process Wetlands for Removal of Trace Organic Contaminants and Pathogens from Municipal Wastewater Effluents. Environ. Eng. Sci. 2013, 30, 421–436. [Google Scholar] [CrossRef]
- Yu, G.; Wang, G.; Chi, T.; Du, C.; Wang, J.; Li, P.; Zhang, Y.; Wang, S.; Yang, K.; Long, Y.; et al. Enhanced Removal of Heavy Metals and Metalloids by Constructed Wetlands: A Review of Approaches and Mechanisms. Science of The Total Environment 2022, 821, 153516. [Google Scholar] [CrossRef]
- Wang, J.; Long, Y.; Yu, G.; Wang, G.; Zhou, Z.; Li, P.; Zhang, Y.; Yang, K.; Wang, S. A Review on Microorganisms in Constructed Wetlands for Typical Pollutant Removal: Species, Function, and Diversity. Frontiers in Microbiology 2022, 13. [Google Scholar] [CrossRef]
- Liu, W.; Li, S.; Yang, K.; Zeng, X.; Li, Y.; Zou, M.; Deng, Y.; Qian, W. CFD-Based Study of Flow Field Characteristics and Clogging in Horizontal Flow Constructed Wetlands. Front. Environ. Sci. 2025, 12. [Google Scholar] [CrossRef]
- Ghermandi, A.; Bixio, D.; Traverso, P.; Cersosimo, I.; Thoeye, C. The Removal of Pathogens in Surface-Flow Constructed Wetlands and Its Implications for Water Reuse. Water Sci. Technol. 2007, 56, 207–216. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and Function of Root Exudates. Plant Cell & Environment 2009, 32, 666–681. [Google Scholar] [CrossRef]
- Qu, P.; Wang, B.; Qi, M.; Lin, R.; Chen, H.; Xie, C.; Zhang, Z.; Qiu, J.; Du, H.; Ge, Y. Medicinal Plant Root Exudate Metabolites Shape the Rhizosphere Microbiota. IJMS 2024, 25, 7786. [Google Scholar] [CrossRef]
- Chojnacka, K.; Moustakas, K.; Mikulewicz, M. The Combined Rhizoremediation by a Triad: Plant-Microorganism-Functional Materials. Environ Sci Pollut Res 2023, 30, 90500–90521. [Google Scholar] [CrossRef]
- Azad, K.; Banerjee, M. Structural Dynamics of Nonenveloped Virus Disassembly Intermediates. J Virol 2019, 93, e01115-19. [Google Scholar] [CrossRef]
- Moresco, V.; Charatzidou, A.; Oliver, D.M.; Weidmann, M.; Matallana-Surget, S.; Quilliam, R.S. Binding, Recovery, and Infectiousness of Enveloped and Non-Enveloped Viruses Associated with Plastic Pollution in Surface Water. Environ. Pollut. 2022, 308, 119594. [Google Scholar] [CrossRef]
- DiCaprio, E.; Culbertson, D.; Li, J. Evidence of the Internalization of Animal Caliciviruses via the Roots of Growing Strawberry Plants and Dissemination to the Fruit. Appl. Environ. Microbiol. 2015, 81, 2727–2734. [Google Scholar] [CrossRef]
- Khan, M.R.; Adam, V.; Rizvi, T.F.; Zhang, B.; Ahamad, F.; Jośko, I.; Zhu, Y.; Yang, M.; Mao, C. Nanoparticle-Plant Interactions: Two-Way Traffic. Small 2019, 15, e1901794. [Google Scholar] [CrossRef]
- Dimitrov, D.S. Virus Entry: Molecular Mechanisms and Biomedical Applications. Nat. Rev. Microbiol. 2004, 2, 109–122. [Google Scholar] [CrossRef]
- Bernaudat, F.; Frelet-Barrand, A.; Pochon, N.; Dementin, S.; Hivin, P.; Boutigny, S.; Rioux, J.-B.; Salvi, D.; Seigneurin-Berny, D.; Richaud, P.; et al. Heterologous Expression of Membrane Proteins: Choosing the Appropriate Host. PLoS One 2011, 6, e29191. [Google Scholar] [CrossRef]
- Kulshreshtha, A.; Sharma, S.; Padilla, C.S.; Mandadi, K.K. Plant-Based Expression Platforms to Produce High-Value Metabolites and Proteins. Front. Plant Sci. 2022, 13, 1043478. [Google Scholar] [CrossRef]
- Baulcombe, D. RNA Silencing in Plants. Biochem. (Lond.) 2015, 37, 10–13. [Google Scholar] [CrossRef]
- Sorel, M.; Mooney, B.; De Marchi, R.; Graciet, E. Ubiquitin/Proteasome System in Plant Pathogen Responses. In Annual Plant Reviews online; Roberts, J.A., Ed.; Wiley, 2019; pp. 65–116. ISBN 978-1-119-31299-4.
- Huang, X.; Chen, S.; Yang, X.; Yang, X.; Zhang, T.; Zhou, G. Friend or Enemy: A Dual Role of Autophagy in Plant Virus Infection. Front. Microbiol. 2020, 11, 736. [Google Scholar] [CrossRef]
- Zure, D.; Sung, M.-H.; Rahim, A.; Kuo, H.-W. In Silico Assessment of Chemical Disinfectants on Surface Proteins Unveiled Dissimilarity in Antiviral Efficacy and Suitability towards Pathogenic Viruses. IJMS 2024, 25, 6009. [Google Scholar] [CrossRef]
- Rachmadi, A.T.; Kitajima, M.; Pepper, I.L.; Gerba, C.P. Enteric and Indicator Virus Removal by Surface Flow Wetlands. Science of The Total Environment 2016, 542, 976–982. [Google Scholar] [CrossRef]
- Alexandros, S.I.; Akratos, C.S. Removal of Pathogenic Bacteria in Constructed Wetlands: Mechanisms and Efficiency. In Phytoremediation; Ansari, A.A., Gill, S.S., Gill, R., Lanza, G.R., Newman, L., Eds.; Springer International Publishing: Cham, 2016; pp. 327–346. ISBN 978-3-319-41810-0. [Google Scholar]
- Plaza-Garrido, A.; Limaico, M.; Villamar-Ayala, C.A. Influence of Wastewater Treatment Technologies on Virus Removal under a Bibliometric-Statistical Analysis. Journal of Water Process Engineering 2022, 47, 102642. [Google Scholar] [CrossRef]
- Shahid, M.; Khalid, S.; Abbas, G.; Shahid, N.; Nadeem, M.; Sabir, M.; Aslam, M.; Dumat, C. Heavy Metal Stress and Crop Productivity. In Crop Production and Global Environmental Issues; Hakeem, K.R., Ed.; Springer International Publishing: Cham, 2015; pp. 1–25. ISBN 978-3-319-23161-7. [Google Scholar]
- Vangronsveld, J.; Herzig, R.; Weyens, N.; Boulet, J.; Adriaensen, K.; Ruttens, A.; Thewys, T.; Vassilev, A.; Meers, E.; Nehnevajova, E.; et al. Phytoremediation of Contaminated Soils and Groundwater: Lessons from the Field. Environmental Science and Pollution Research 2009 16:7 2009, 16, 765–794. [Google Scholar] [CrossRef]
- Kataki, S.; Chatterjee, S.; Vairale, M.G.; Dwivedi, S.K.; Gupta, D.K. Constructed Wetland, an Eco-Technology for Wastewater Treatment: A Review on Types of Wastewater Treated and Components of the Technology (Macrophyte, Biolfilm and Substrate). 2021, 283, 111986–111986. [CrossRef]
- Armanious, A.; Aeppli, M.; Jacak, R.; Refardt, D.; Sigstam, T.; Kohn, T.; Sander, M. Viruses at Solid-Water Interfaces: A Systematic Assessment of Interactions Driving Adsorption. Environ. Sci. Technol. 2016, 50, 732–743. [Google Scholar] [CrossRef]
- Romanenko, A.; Peter, H.; Meibom, J.; Borchardt, M.A.; Kohn, T. Diversity of Lake Bacteria Promotes Human Echovirus Inactivation. Appl Environ Microbiol 2025, 91, e02366-24. [Google Scholar] [CrossRef]
- Zure, D.; David Kuo, H.-W.; Drizo, A. Insights of Phytoremediation Mechanisms for Viruses Based on In-Vitro, in-Vivo and in-Silico Assessments of Selected Herbal Plants. Chemosphere 2024, 351, 141101. [Google Scholar] [CrossRef]
- Han, Q.; Fu, G.; Liu, K.; Adnan, M.; Liu, S.; Wang, M.; Jiang, F.; Wang, M. Chemical Speciation and Transformation of Heavy Metals in Soil and Groundwater: Implications for Ecotoxicology and Remediation. J. Environ. Chem. Eng. 2025, 13, 119858. [Google Scholar] [CrossRef]
- Torrey, J.; Von Gunten, U.; Kohn, T. Differences in Viral Disinfection Mechanisms as Revealed by Quantitative Transfection of Echovirus 11 Genomes. Appl Environ Microbiol 2019, 85, e00961-19. [Google Scholar] [CrossRef]
- Nie, X.; Huang, X.; Li, M.; Lu, Z.; Ling, X. Advances in Soil Amendments for Remediation of Heavy Metal-Contaminated Soils: Mechanisms, Impact, and Future Prospects. Toxics 2024, 12, 872. [Google Scholar] [CrossRef]
- Firquet, S.; Beaujard, S.; Lobert, P.-E.; Sané, F.; Caloone, D.; Izard, D.; Hober, D. Survival of Enveloped and Non-Enveloped Viruses on Inanimate Surfaces. Microbes and environments 2015, 30, 140–144. [Google Scholar] [CrossRef]
- Roldan-Hernandez, L.; Boehm, A.B. Adsorption of Respiratory Syncytial Virus (RSV), Rhinovirus, SARS-CoV-2, and F+ Bacteriophage MS2 RNA onto Wastewater Solids from Raw Wastewater 2023.
- Zhang, C.; Liu, X.; Lu, X.; He, M.; Jan Meijer, E.; Wang, R. Surface Complexation of Heavy Metal Cations on Clay Edges: Insights from First Principles Molecular Dynamics Simulation of Ni(II). Geochimica et Cosmochimica Acta 2017, 203, 54–68. [Google Scholar] [CrossRef]
- Paraskevopoulos, S.; Smeets, P. Quantifying the Log Reduction of Pathogenic Microorganisms by Constructed Wetlands: A Review. In Proceedings of the The 4th International Electronic Conference on Water Sciences, MDPI, November 12 2019; p. 9. [Google Scholar]
- Shingare, R.P.; Thawale, P.R.; Raghunathan, K.; Mishra, A.; Kumar, S. Constructed Wetland for Wastewater Reuse: Role and Efficiency in Removing Enteric Pathogens. Journal of Environmental Management 2019, 246, 444–461. [Google Scholar] [CrossRef]
- Roldan-Hernandez, L.; Van Oost, C.; Boehm, A.B. Solid-Liquid Partitioning of Dengue, West Nile, Zika, Hepatitis A, Influenza A, and SARS-CoV-2 Viruses in Wastewater from across the United States 2024.
- Schück, M.; Greger, M. Screening the Capacity of 34 Wetland Plant Species to Remove Heavy Metals from Water. IJERPH 2020, 17, 4623. [Google Scholar] [CrossRef]
- Urase, T.; Yamamoto, K.; Ohgaki, S. Effect of Pore Structure of Membranes and Module Configuration on Virus Retention. Journal of Membrane Science 1996, 115, 21–29. [Google Scholar] [CrossRef]
- Agrawal, P.K.; Agrawal, C.; Blunden, G. Rutin: A Potential Antiviral for Repurposing as a SARS-CoV-2 Main Protease (Mpro ) Inhibitor. Natural Product Communications 2021, 16, 1934578X21991723. [Google Scholar] [CrossRef]
- Gargallo-Garriga, A.; Preece, C.; Sardans, J.; Oravec, M.; Urban, O.; Peñuelas, J. Root Exudate Metabolomes Change under Drought and Show Limited Capacity for Recovery. Sci. Rep. 2018, 8, 12696. [Google Scholar] [CrossRef]
- Jiang, T.; Hao, T.; Chen, W.; Li, C.; Pang, S.; Fu, C.; Cheng, J.; Zhang, C.; Ghorbanpour, M.; Miao, S. Reprogrammed Plant Metabolism During Viral Infections: Mechanisms, Pathways and Implications. Molecular Plant Pathology 2025, 26, e70066. [Google Scholar] [CrossRef]
- Ma, H.; Zhao, M.; Yang, N.; Feng, L.; Wang, L.; Jiang, C.; Jiang, M.; Guo, J.; Li, T. Regulated the Electrokinetic Application of Different Plant Growth Stages and Parameters Enhance the Economic Extraction of Soil Heavy Metals. Front. Plant Sci. 2025, 16, 1557261. [Google Scholar] [CrossRef]
- Porras-Socias, P.; Tomasino, M.P.; Fernandes, J.P.; De Menezes, A.B.; Fernández, B.; Collins, G.; Alves, M.J.; Castro, R.; Gomes, C.R.; Almeida, C.M.R.; et al. Removal of Metals and Emergent Contaminants from Liquid Digestates in Constructed Wetlands for Agricultural Reuse. Front. Microbiol. 2024, 15, 1388895. [Google Scholar] [CrossRef]
- Camargos, C.H.M.; Yang, L.; Jackson, J.C.; Tanganini, I.C.; Francisco, K.R.; Ceccato-Antonini, S.R.; Rezende, C.A.; Faria, A.F. Lignin and Nanolignin: Next-Generation Sustainable Materials for Water Treatment. ACS Appl. Bio Mater. 2025, 8, 2632–2673. [Google Scholar] [CrossRef]
- Zhou, Y.; Luo, S.; Yu, B.; Zhang, T.; Li, J.; Zhang, Y. A Comparative Analysis for the Development and Recovery Processes of Different Types of Clogging in Lab-Scale Vertical Flow Constructed Wetlands. Environ Sci Pollut Res 2018, 25, 24073–24083. [Google Scholar] [CrossRef]
- Dai, L.; Ren, J.; Ling, T.; Wei, B.; Wang, G. Chemical Speciation and Phytoavailability of Cr, Ni, Zn and Cu in Loess Amended with Attapulgite-Stabilized Sewage Sludge. Environmental Pollutants and Bioavailability 2019, 31, 112–119. [Google Scholar] [CrossRef]
- Di Luca, G.A.; Hadad, H.R.; Mufarrege, M.M.; Maine, M.A.; Sánchez, G.C. Improvement of Cr Phytoremediation by Pistia Stratiotes in Presence of Nutrients. Int. J. Phytoremediation 2014, 16, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Dhir, B. Effective Control of Waterborne Pathogens by Aquatic Plants. In Waterborne Pathogens; Elsevier, 2020; pp. 339–361. ISBN 978-0-12-818783-8.
- Nguyen Vo Chau, N.; Huynh Van, T.; Nguyen Cong, T.; Kim, L.; Pham, D.V. Water Lettuce ( Pistia Stratiotes L.) Increases Biogas Effluent Pollutant Removal Efficacy and Proves a Positive Substrate for Renewable Energy Production. PeerJ 2023, 11, e15879. [Google Scholar] [CrossRef]
- Sharrer, M.J.; Summerfelt, S.T. Ozonation Followed by Ultraviolet Irradiation Provides Effective Bacteria Inactivation in a Freshwater Recirculating System. Aquacultural Engineering 2007, 37, 180–191. [Google Scholar] [CrossRef]
- Sharma, V.K.; Zboril, R.; Varma, R.S. Ferrates: Greener Oxidants with Multimodal Action in Water Treatment Technologies. Acc. Chem. Res. 2015, 48, 182–191. [Google Scholar] [CrossRef]
- Kokkinos, P.; Venieri, D.; Mantzavinos, D. Advanced Oxidation Processes for Water and Wastewater Viral Disinfection. A Systematic Review. Food Environ. Virol. 2021, 13, 283–302. [Google Scholar] [CrossRef]
- Stoquart, C.; Servais, P.; Bérubé, P.R.; Barbeau, B. Hybrid Membrane Processes Using Activated Carbon Treatment for Drinking Water: A Review. Journal of Membrane Science 2012, 411–412, 1–12. [Google Scholar] [CrossRef]
- Stiegler, A.N.; Cecchetti, A.R.; Scholes, R.C.; Sedlak, D.L. Persistent Trace Organic Contaminants Are Transformed Rapidly under Sulfate- and Fe(III)-Reducing Conditions in a Nature-Based Subsurface Water Treatment System. Environ. Sci. Technol. 2023, 57, 16616–16627. [Google Scholar] [CrossRef]
- Uggetti, E.; Hughes-Riley, T.; Morris, R.H.; Newton, M.I.; Trabi, C.L.; Hawes, P.; Puigagut, J.; García, J. Intermittent Aeration to Improve Wastewater Treatment Efficiency in Pilot-Scale Constructed Wetland. Science of The Total Environment 2016, 559, 212–217. [Google Scholar] [CrossRef]
- Hargreaves, A.J.; Constantino, C.; Dotro, G.; Cartmell, E.; Campo, P. Fate and Removal of Metals in Municipal Wastewater Treatment: A Review. Environ. Technol. Rev. 2018, 7, 1–18. [Google Scholar] [CrossRef]
- Rahman, M.E.; Bin Halmi, M.I.E.; Bin Abd Samad, M.Y.; Uddin, M.K.; Mahmud, K.; Abd Shukor, M.Y.; Sheikh Abdullah, S.R.; Shamsuzzaman, S.M. Design, Operation and Optimization of Constructed Wetland for Removal of Pollutant. 2020, 17, 8339–8339. [CrossRef]
- Dykes, C.; Pearson, J.; Bending, G.; Abolfathi, S. Impact of Seasonal Climate Variability on Constructed Wetland Treatment Efficiency. J. Water Proc.engineering 2025, 72, 107350. [Google Scholar] [CrossRef]
- Cao, R.; Zhang, Y.; Ju, Y.; Wang, W.; Zhao, Y.; Liu, N.; Zhang, G.; Wang, X.; Xie, X.; Dai, C.; et al. Exopolysaccharide-Producing Bacteria Enhanced Pb Immobilization and Influenced the Microbiome Composition in Rhizosphere Soil of Pakchoi (Brassica Chinensis L.). Front. Microbiol. 2023, 14, 1117312. [Google Scholar] [CrossRef] [PubMed]
- Vojnovic, S.; Aleksic, I.; Ilic-Tomic, T.; Stevanovic, M.; Nikodinovic-Runic, J. Bacillus and Streptomyces Spp. as Hosts for Production of Industrially Relevant Enzymes. Appl Microbiol Biotechnol 2024, 108, 185. [Google Scholar] [CrossRef]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy Metal Tolerance in Plants: Role of Transcriptomics, Proteomics, Metabolomics, and Ionomics. Front. Plant Sci. 2016, 6. [Google Scholar] [CrossRef]
- Sun, H.; Jing, X.; Wang, C.; Wang, P.; Huang, Z.; Sun, B.; Li, P.; Li, H.; Zhang, C. The Great Game between Plants and Viruses: A Focus on Protein Homeostasis. IJMS 2023, 24, 12582. [Google Scholar] [CrossRef]
- Jiang, T.; Hao, T.; Chen, W.; Li, C.; Pang, S.; Fu, C.; Cheng, J.; Zhang, C.; Ghorbanpour, M.; Miao, S. Reprogrammed Plant Metabolism during Viral Infections: Mechanisms, Pathways and Implications. Mol. Plant Pathol. 2025, 26, e70066. [Google Scholar] [CrossRef]
- Agrawal, P.K.; Agrawal, C.; Blunden, G. Rutin: A Potential Antiviral for Repurposing as a SARS-CoV-2 Main Protease (Mpro) Inhibitor. Nat. Prod. Commun. 2021, 16, 1934578X2199172. [Google Scholar] [CrossRef]
- Yang, B.; Tian, L.; Zhou, P.; Babakhani, P.; Gregory, J.; Graham, N.; Elimelech, M.; Yu, W. Nanonet Trapping for Effective Removal of Nanoplastics by Iron Coagulation. Nat Commun 2025, 16, 5998. [Google Scholar] [CrossRef]
- Cao, R.; Zhang, Y.; Ju, Y.; Wang, W.; Zhao, Y.; Liu, N.; Zhang, G.; Wang, X.; Xie, X.; Dai, C.; et al. Exopolysaccharide-Producing Bacteria Enhanced Pb Immobilization and Influenced the Microbiome Composition in Rhizosphere Soil of Pakchoi (Brassica Chinensis L.). Front. Microbiol. 2023, 14, 1117312. [Google Scholar] [CrossRef]
- Varma, M.; Gupta, A.K.; Ghosal, P.S.; Majumder, A. A Review on Performance of Constructed Wetlands in Tropical and Cold Climate: Insights of Mechanism, Role of Influencing Factors, and System Modification in Low Temperature. 2021, 755, 142540–142540. [CrossRef]
- Liu, Y.; Mancuso, G.; Petrotto, L.; Lavrnić, S.; Dong, Z.; Tian, Y.; Zhang, J.; Toscano, A. AI-Driven Solutions in Wastewater Treatment and Agricultural Reuse Systems: A Comprehensive Review. J. Environ. Manage. 2025, 393, 127008. [Google Scholar] [CrossRef]
- Tanneru, C.T.; Chellam, S. Mechanisms of Virus Control during Iron Electrocoagulation – Microfiltration of Surface Water. Water Research 2012, 46, 2111–2120. [Google Scholar] [CrossRef]
- Kim, K.; Narayanan, J.; Sen, A.; Chellam, S. Virus Removal and Inactivation Mechanisms during Iron Electrocoagulation: Capsid and Genome Damages and Electro-Fenton Reactions. Environ. Sci. Technol. 2021, acs.est.0c04438. [CrossRef]
- Bicudo, B.; Medema, G.; van Halem, D. Inactivation of Escherichia Coli and Somatic Coliphage ΦX174 by Oxidation of Electrochemically Produced Fe2+. J. Water Proc.engineering 2022, 47, 102683. [Google Scholar] [CrossRef]
- Wang, J.; Man, Y.; Ruan, W.; Tam, N.F.; Tao, R.; Yin, L.; Yang, Y.; Dai, Y.; Tai, Y. The Effect of Rhizosphere and the Plant Species on the Degradation of Sulfonamides in Model Constructed Wetlands Treating Synthetic Domestic Wastewater. Chemosphere 2022, 288, 132487. [Google Scholar] [CrossRef]
- Hdidou, M.; Necibi, M.C.; Labille, J.; El Hajjaji, S.; Dhiba, D.; Chehbouni, A.; Roche, N. Potential Use of Constructed Wetland Systems for Rural Sanitation and Wastewater Reuse in Agriculture in the Moroccan Context. Energies 2021, 15, 156. [Google Scholar] [CrossRef]
- Wang, Q.; Xie, H.; Zhang, J.; Liang, S.; Ngo, H.H.; Guo, W.; Liu, C.; Zhao, C.; Li, H. Effect of Plant Harvesting on the Performance of Constructed Wetlands during Winter: Radial Oxygen Loss and Microbial Characteristics. Environ Sci Pollut Res 2015, 22, 7476–7484. [Google Scholar] [CrossRef]
- Nourredine, Y.; Naima, A.; Dalila, H.; Habib, S.; Karim, S.; Zohra, F.-L. Changes of Peroxidase Activities under Cold Stress in Annuals Populations of Medicago. Mol. Plant Breed. 2015. [Google Scholar] [CrossRef]
- Knowles, P.; Dotro, G.; Nivala, J.; García, J. Clogging in Subsurface-Flow Treatment Wetlands: Occurrence and Contributing Factors. Ecol. Eng. 2011, 37, 99–112. [Google Scholar] [CrossRef]
- Brix, H. Functions of Macrophytes in Constructed Wetlands. Water Sci. Technol. 1994, 29, 71–78. [Google Scholar] [CrossRef]
- Suhaib, K.H.; Bhunia, P. Dynamics of Clogging in Subsurface Flow Constructed Wetlands. J. Hazard. Toxic Radioact. Waste 2022, 26, 03121004. [Google Scholar] [CrossRef]
- Chiang, L.-C.; Ng, L.-T.; Cheng, P.-W.; Chiang, W.; Lin, C.-C. Antiviral Activities of Extracts and Selected Pure Constituents of Ocimum Basilicum. Clinical and Experimental Pharmacology and Physiology 2005, 32, 811–816. [Google Scholar] [CrossRef]
- Ebrahimi, A.; Taheri, E.; Ehrampoush, M.H.; Nasiri, S.; Jalali, F.; Soltani, R.; Fatehizadeh, A. Efficiency of Constructed Wetland Vegetated with Cyperus Alternifolius Applied for Municipal Wastewater Treatment. J. Environ. Public Health 2013, 2013, 815962. [Google Scholar] [CrossRef]
- Cangioli, L.; Salobehaj, M.; Del Duca, S.; Fagorzi, C.; Berardi, C.; Coppini, E.; Fibbi, D.; Fani, R.; Vassallo, A. Effect of Wastewater on the Composition of Bacterial Microbiota of Phragmites Australis Used in Constructed Wetlands for Phytodepuration. Plants 2022, 11, 3210. [Google Scholar] [CrossRef]
- Riva, V.; Vergani, L.; Rashed, A.A.; El Saadi, A.; Sabatino, R.; Di Cesare, A.; Crotti, E.; Mapelli, F.; Borin, S. Plant Species Influences the Composition of Root System Microbiome and Its Antibiotic Resistance Profile in a Constructed Wetland Receiving Primary Treated Wastewater. Front. Microbiol. 2024, 15, 1436122. [Google Scholar] [CrossRef]
- Tsai, Y.-C.; Lee, C.-L.; Yen, H.-R.; Chang, Y.-S.; Lin, Y.-P.; Huang, S.-H.; Lin, C.-W. Antiviral Action of Tryptanthrin Isolated from Strobilanthes Cusia Leaf against Human Coronavirus NL63. Biomolecules 2020, 10, 366. [Google Scholar] [CrossRef]
- Pantanam, A.; Mana, N.; Semkum, P.; Lueangaramkul, V.; Phecharat, N.; Lekcharoensuk, P.; Theerawatanasirikul, S. Dual Effects of Ipecac Alkaloids with Potent Antiviral Activity against Foot-and-Mouth Disease Virus as Replicase Inhibitors and Direct Virucides. Int. J. Vet. Sci. Med. 2024, 12, 134–147. [Google Scholar] [CrossRef]
- Bhatia, M.; Goyal, D. Analyzing Remediation Potential of Wastewater through Wetland Plants: A Review. Environ. Prog. Sustain. Energy 2014, 33, 9–27. [Google Scholar] [CrossRef]
- Kochi, L.Y.; Freitas, P.L.; Maranho, L.T.; Juneau, P.; Gomes, M.P. Aquatic Macrophytes in Constructed Wetlands: A Fight against Water Pollution. Sustainability 2020, 12, 9202. [Google Scholar] [CrossRef]
- Maranho, L.T.; Gomes, M.P. Morphophysiological Adaptations of Aquatic Macrophytes in Wetland-Based Sewage Treatment Systems: Strategies for Resilience and Efficiency under Environmental Stress. Plants 2024, 13, 2870. [Google Scholar] [CrossRef]
- Markland, S.M.; Bais, H.; Kniel, K.E. Human Norovirus and Its Surrogates Induce Plant Immune Response in Arabidopsis Thaliana and Lactuca Sativa. Foodborne Pathogens and Disease 2017, 14, 432–439. [Google Scholar] [CrossRef]
- Esseili, M.A.; Meulia, T.; Saif, L.J.; Wang, Q. Tissue Distribution and Visualization of Internalized Human Norovirus in Leafy Greens. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef]
- Muerdter, C.P.; LeFevre, G.H.; Cwiertny, D.; Just, C.; Lehmler, H.-J.; Schnoor, J. Vegetation-Facilitated Removal Kinetics and Transformation of Organic Biocides. Doctor of Philosophy, University of Iowa: Iowa City, IA, United States, 2021.
- Li, J.; Zheng, L.; Ye, C.; Ni, B.; Wang, X.; Liu, H. Evaluation of an Intermittent-Aeration Constructed Wetland for Removing Residual Organics and Nutrients from Secondary Effluent: Performance and Microbial Analysis. Bioresour. Technol. 2021, 329, 124897. [Google Scholar] [CrossRef]
- Yu, G.; Chen, J.; Wang, G.; Chen, H.; Huang, J.; Li, Y.; Wang, W.; Song, F.; Ma, Y.; Wang, Q.; et al. Recent Advances in Constructed Wetlands Methane Reduction: Mechanisms and Methods. Front. Microbiol. 2023, 14, 1106332. [Google Scholar] [CrossRef]
- Pandey, D.; Singh, S.V.; Savio, N.; Bhutto, J.K.; Srivastava, R.K.; Yadav, K.K.; Sharma, R.; Nandipamu, T.M.K.; Sarkar, B. Biochar Application in Constructed Wetlands for Wastewater Treatment: A Critical Review. J. Water Proc.engineering 2025, 69, 106713. [Google Scholar] [CrossRef]
- Pu, J.; Liu, Y.; Zhang, J.; An, B.; Li, Y.; Wang, X.; Din, K.; Qin, C.; Li, K.; Cui, M.; et al. Virus Disinfection from Environmental Water Sources Using Living Engineered Biofilm Materials. Advanced Science 2020, 7, 1903558. [Google Scholar] [CrossRef]
- Ndjomgoue-Yossa, A.C.; Nanseu-Njiki, C.P.; Ngameni, E. Effect of pH on Escherichia Coli Removal by Electrocoagulation and Elimination Kinetics after Treatment. J. Chem. 2022, 2022, 1–8. [Google Scholar] [CrossRef]
- Al-Hazmi, H.E.; Shokrani, H.; Shokrani, A.; Jabbour, K.; Abida, O.; Mousavi Khadem, S.S.; Habibzadeh, S.; Sonawane, S.H.; Saeb, M.R.; Bonilla-Petriciolet, A.; et al. Recent Advances in Aqueous Virus Removal Technologies. Chemosphere 2022, 305, 135441. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.; Zhang, Y.; Liu, J.; Huang, Y.; Li, J. Seasonal Effects of Constructed Wetlands on Water Quality Characteristics in Jinshan Lake: A Gate Dam Lake (Zhenjiang City, China). Biology (Basel) 2024, 13, 593. [Google Scholar] [CrossRef]
- Cui, L.; Feng, J.; Ouyang, Y.; Deng, P. Removal of Nutrients from Septic Effluent with Re-Circulated Hybrid Tidal Flow Constructed Wetland. Ecol. Eng. 2012, 46, 112–115. [Google Scholar] [CrossRef]
- Cui, L.; Feng, J.; Ouyang, Y.; Deng, P. Removal of Nutrients from Septic Effluent with Re-Circulated Hybrid Tidal Flow Constructed Wetland. Ecological Engineering 2012, 46, 112–115. [Google Scholar] [CrossRef]
- Cangioli, L.; Salobehaj, M.; Del Duca, S.; Fagorzi, C.; Berardi, C.; Coppini, E.; Fibbi, D.; Fani, R.; Vassallo, A. Effect of Wastewater on the Composition of Bacterial Microbiota of Phragmites Australis Used in Constructed Wetlands for Phytodepuration. Plants 2022, 11, 3210. [Google Scholar] [CrossRef]
- Ali, S.; Abbas, Z.; Rizwan, M.; Zaheer, I.; Yavaş, İ.; Ünay, A.; Abdel-DAIM, M.; Bin-Jumah, M.; Hasanuzzaman, M.; Kalderis, D. Application of Floating Aquatic Plants in Phytoremediation of Heavy Metals Polluted Water: A Review. Sustainability 2020, 12, 1927. [Google Scholar] [CrossRef]
- Zhilkina, T.; Gerasimova, I.; Babich, T.; Kanapatskiy, T.; Sokolova, D.; Kadnikov, V.; Kamionskaya, A. Evaluation of the Phytoremediation Potential of Aquatic Plants and Associated Microorganisms for the Cleaning of Aquatic Ecosystems from Oil Products. Sustainability 2024, 16, 9288. [Google Scholar] [CrossRef]
- Liang, K.; Dai, Y.; Wang, F.; Liang, W. Seasonal Variation of Microbial Community for the Treatment of Tail Water in Constructed Wetland. Water Science and Technology 2017, 75, 2434–2442. [Google Scholar] [CrossRef]
- Richter, A.Y.; Weaver, R.W. Ultraviolet Disinfection of Effluent from Subsurface Flow Constructed Wetlands. Environmental Technology 2003, 24, 1175–1182. [Google Scholar] [CrossRef]
- Arden, S.; Anderson, M.; Blue, J.; Ma, X. (Cissy); Jahne, M.; Garland, J. Towards the Definition of Treatment Wetland Pathogen Log Reduction Credits. Science of The Total Environment 2024, 957, 177613. [Google Scholar] [CrossRef]
- Shruthi, R.; Shivashankara, G.P. Effect of HRT and Seasons on the Performance of Pilot-Scale Horizontal Subsurface Flow Constructed Wetland to Treat Rural Wastewater. Water Practice and Technology 2022, 17, 445–455. [Google Scholar] [CrossRef]
- GARCÍA, J.; ROUSSEAU, D.P.L.; MORATÓ, J.; LESAGE, E.L.S.; MATAMOROS, V.; BAYONA, J.M. Contaminant Removal Processes in Subsurface-Flow Constructed Wetlands: A Review. 2010, 40, 561–661. [CrossRef]
- Wang, D.; Bai, Y.; Qu, J. The Phragmites Root-Inhabiting Microbiome: A Critical Review on Its Composition and Environmental Application. Engineering 2022, 9, 42–50. [Google Scholar] [CrossRef]
- Foladori, P.; Ruaben, J.; Ortigara, A.R.C.; Andreottola, G. Batch Feed and Intermittent Recirculation to Increase Removed Loads in a Vertical Subsurface Flow Filter. Ecol. Eng. 2014, 70, 124–132. [Google Scholar] [CrossRef]
- Shirdashtzadeh, M.; Chua, L.H.C.; Brau, L. Microbial Communities and Nitrogen Transformation in Constructed Wetlands Treating Stormwater Runoff. Front. Water 2022, 3, 751830. [Google Scholar] [CrossRef]
- Wu, H.; Wang, X.; He, X.; Zhang, S.; Liang, R.; Shen, J. Effects of Root Exudates on Denitrifier Gene Abundance, Community Structure and Activity in a Micro-Polluted Constructed Wetland. Science of The Total Environment 2017, 598, 697–703. [Google Scholar] [CrossRef]
- Wang, C.; Chen, W.; Xia, A.; Zhang, R.; Huang, Y.; Yang, S.; Ni, L.; Jin, F. Carbon Starvation Induces the Expression of PprB-Regulated Genes in Pseudomonas Aeruginosa. Appl Environ Microbiol 2019, 85, e01705-19. [Google Scholar] [CrossRef]
- Gao, Z.; Chen, H.; Wang, Y.; Lv, Y. Advances in AHLs-Mediated Quorum Sensing System in Wastewater Biological Nitrogen Removal: Mechanism, Function, and Application. Water Sci. Technol. 2022, 86, 1927–1943. [Google Scholar] [CrossRef]
- Mehmood, N.; Saeed, M.; Zafarullah, S.; Hyder, S.; Rizvi, Z.F.; Gondal, A.S.; Jamil, N.; Iqbal, R.; Ali, B.; Ercisli, S.; et al. Multifaceted Impacts of Plant-Beneficial Pseudomonas Spp. In Managing Various Plant Diseases and Crop Yield Improvement. ACS Omega 2023, 8, 22296–22315. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, R.; Li, M.; Guo, X.; Lu, S.; Li, M.; Wu, H. Intensified Performance and Mechanism of Nitrogen Removal in Constructed Wetland Incorporating Algal Pond for Treating Low Carbon Nitrogen Ratio Wastewater. Bioresource Technology 2025, 429, 132488. [Google Scholar] [CrossRef]
- Mou, Z.; Fan, W.; Dong, X. Inducers of Plant Systemic Acquired Resistance Regulate NPR1 Function through Redox Changes. Cell 2003, 113, 935–944. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, S.; Ding, P.; Wang, D.; Cheng, Y.T.; He, J.; Gao, M.; Xu, F.; Li, Y.; Zhu, Z.; et al. Control of Salicylic Acid Synthesis and Systemic Acquired Resistance by Two Members of a Plant-Specific Family of Transcription Factors. Proc. Natl. Acad. Sci. U.S.A. 2010, 107, 18220–18225. [Google Scholar] [CrossRef]
- Shine, M.B.; Xiao, X.; Kachroo, P.; Kachroo, A. Signaling Mechanisms Underlying Systemic Acquired Resistance to Microbial Pathogens. Plant Science 2019, 279, 81–86. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Q.; Mou, Z. Redox Signaling and Oxidative Stress in Systemic Acquired Resistance. J. Exp. Bot. 2024, 75, 4535–4548. [Google Scholar] [CrossRef]
- Idris, N.N.; Chua, L.H.C.; Mustaffa, Z.; Das, S.; Takaijudin, H. A Review Study on the Association between Hydraulic Performance and Treatment Effectiveness in Free Surface Flow Constructed Wetlands. Ecol. Eng. 2024, 203, 107258. [Google Scholar] [CrossRef]
- Tang, P.; Xiang, Z.; Zhou, Y.; Zhang, Y. Enzyme Treatment Improves the Performance of Laboratory-Scale Vertical Flow Constructed Wetland. Bioresource Technology 2018, 268, 665–671. [Google Scholar] [CrossRef]
- Verhofstad, M.J.J.M.; Poelen, M.D.M.; Van Kempen, M.M.L.; Bakker, E.S.; Smolders, A.J.P. Finding the Harvesting Frequency to Maximize Nutrient Removal in a Constructed Wetland Dominated by Submerged Aquatic Plants. Ecological Engineering 2017, 106, 423–430. [Google Scholar] [CrossRef]
- Datta, A.; Singh, H.O.; Raja, S.K.; Dixit, S. Constructed Wetland for Improved Wastewater Management and Increased Water Use Efficiency in Resource Scarce SAT Villages: A Case Study from Kothapally Village, in India. Int. J. Phytoremediation 2021, 23, 1067–1076. [Google Scholar] [CrossRef]
- Hernes, P.J.; Miller, R.L.; Dyda, R.Y.; Bergamaschi, B.A. Vegetation vs. Anoxic Controls on Degradation of Plant Litter in a Restored Wetland. Front. Environ. Sci. 2020, 8. [Google Scholar] [CrossRef]
- Lv, S.; Zhang, S.; Zhang, M.; Liu, F.; Cheng, L. Effects of Multi-Plant Harvesting on Nitrogen Removal and Recovery in Constructed Wetlands. Chemosphere 2024, 353, 141550. [Google Scholar] [CrossRef]
- Bai, S.-Y.; Song, Z.-X.; Ding, Y.-L.; You, S.-H.; He, S. Correlation of substrate structure and hydraulic characteristics in subsurface flow constructed wetlands. Huan Jing Ke Xue 2014, 35, 592–596. [Google Scholar]
- Pedescoll, A.; Corzo, A.; Alvarez, E.; García, J.; Puigagut, J. The Effect of Primary Treatment and Flow Regime on Clogging Development in Horizontal Subsurface Flow Constructed Wetlands: An Experimental Evaluation. Water Res. 2011, 45, 3579–3589. [Google Scholar] [CrossRef]
- Nivala, J.; Headley, T.; Wallace, S.; Bernhard, K.; Brix, H.; Van Afferden, M.; Müller, R.A. Comparative Analysis of Constructed Wetlands: The Design and Construction of the Ecotechnology Research Facility in Langenreichenbach, Germany. Ecological Engineering 2013, 61, 527–543. [Google Scholar] [CrossRef]
- Hakizimana, J.N.; Gourich, B.; Chafi, M.; Stiriba, Y.; Vial, C.; Drogui, P.; Naja, J. Electrocoagulation Process in Water Treatment: A Review of Electrocoagulation Modeling Approaches. Desalination 2017, 404, 1–21. [Google Scholar] [CrossRef]
- Kim, K.; Jothikumar, N.; Sen, A.; Murphy, J.L.; Chellam, S. Removal and Inactivation of an Enveloped Virus Surrogate by Iron Conventional Coagulation and Electrocoagulation. Environ. Sci. Technol. 2021, 55, 2674–2683. [Google Scholar] [CrossRef]
- Bicudo, B.; van der Werff, B.-J.; Medema, G.; van Halem, D. Disinfection during Iron Electrocoagulation: Differentiating between Inactivation and Floc Entrapment for Escherichia Coli and Somatic Coliphage ØX174. ACS ES T Water 2022, 2, 1707–1714. [Google Scholar] [CrossRef]
- Guo, B.; Pasco, E.V.; Xagoraraki, I.; Tarabara, V.V. Virus Removal and Inactivation in a Hybrid Microfiltration–UV Process with a Photocatalytic Membrane. Sep. Purif. Technol. 2015, 149, 245–254. [Google Scholar] [CrossRef]
- Marques dos Santos, M.; Li, C.; Jemain, M.H.; Yuen, J.W.; Snyder, S.A. Removal of Human Coronavirus OC43 (HCoV-OC43) in Simulated Drinking Water Treatment Processes. ACS ES T Water 2024. [Google Scholar] [CrossRef]
- Barocsi, A.; Csintalan, Z.; Kocsanyi, L.; Dushenkov, S.; Kuperberg, J.M.; Kucharski, R.; Richter, P.I. Optimizing Phytoremediation of Heavy Metal-Contaminated Soil by Exploiting Plants’ Stress Adaptation. Int. J. Phytoremediation 2003, 5, 13–23. [Google Scholar] [CrossRef]
- Mansoor, S.; Ali Wani, O.; Lone, J.K.; Manhas, S.; Kour, N.; Alam, P.; Ahmad, A.; Ahmad, P. Reactive Oxygen Species in Plants: From Source to Sink. Antioxidants 2022, 11, 225. [Google Scholar] [CrossRef]
- Liang, K.; Dai, Y.; Wang, F.; Liang, W. Seasonal Variation of Microbial Community for the Treatment of Tail Water in Constructed Wetland. Water Sci. Technol. 2017, 75, 2434–2442. [Google Scholar] [CrossRef]
- Allen, D.J.; Farrell, M.; Huang, J.; Reynolds, C.; Rupasinghe, M.; Mosley, L.M. Long-Term Water Quality Response to Increased Hydraulic Loadings in a Field-Scale Free Water Surface Constructed Wetland Treating Domestic Effluent. J. Environ. Manage. 2022, 311, 114858. [Google Scholar] [CrossRef]
- Liang, M.-Y.; Han, Y.-C.; Easa, S.M.; Chu, P.-P.; Wang, Y.-L.; Zhou, X.-Y. New Solution to Build Constructed Wetland in Cold Climatic Region. Sci. Total Environ. 2020, 719, 137124. [Google Scholar] [CrossRef]
- Abir Ahsan, T.M.; Rahman, M.S.; Ahamed, M.S. Geothermal Energy Application for Greenhouse Microclimate Management: A Review. Geothermics 2025, 127, 103209. [Google Scholar] [CrossRef]
- Wang, R.; Alamin, Md.; Tsuji, S.; Hara-Yamamura, H.; Hata, A.; Zhao, B.; Ihara, M.; Honda, R. Removal Performance of SARS-CoV-2 in Wastewater Treatment by Membrane Bioreactor, Anaerobic-Anoxic-Oxic, and Conventional Activated Sludge Processes. Science of The Total Environment 2022, 851, 158310. [Google Scholar] [CrossRef]
- Espinosa, M.F.; Verbyla, M.E.; Vassalle, L.; Rosa-Machado, A.T.; Zhao, F.; Gaunin, A.; Mota, C.R. Reduction and Partitioning of Viral and Bacterial Indicators in a UASB Reactor Followed by High Rate Algal Ponds Treating Domestic Sewage. Sci. Total Environ. 2021, 760, 144309. [Google Scholar] [CrossRef]
- Verbyla, M.E.; Mihelcic, J.R. A Review of Virus Removal in Wastewater Treatment Pond Systems. Water Research 2015, 71, 107–124. [Google Scholar] [CrossRef]
- Arden, S.; Ma, X. Constructed Wetlands for Greywater Recycle and Reuse: A Review. Science of the Total Environment 2018. [Google Scholar] [CrossRef]
- Liu, H.; Hu, Z.; Zhang, J.; Ngo, H.H.; Guo, W.; Liang, S.; Fan, J.; Lu, S.; Wu, H. Optimizations on Supply and Distribution of Dissolved Oxygen in Constructed Wetlands: A Review. Bioresour. Technol. 2016, 214, 797–805. [Google Scholar] [CrossRef]
- Sossalla, N.A.; Nivala, J.; Escher, B.I.; Reemtsma, T.; Schlichting, R.; Van Afferden, M.; Müller, R.A. Resilience of Micropollutant and Biological Effect Removal in an Aerated Horizontal Flow Treatment Wetland. Water 2020, 12, 3050. [Google Scholar] [CrossRef]
- Biswal, B.K.; Balasubramanian, R. Constructed Wetlands for Reclamation and Reuse of Wastewater and Urban Stormwater: A Review. Frontiers in Environmental Science 2022, 10, 836289–836289. [Google Scholar] [CrossRef]
- Augsburger, N.; Rachmadi, A.T.; Zaouri, N.; Lee, Y.; Hong, P.-Y. Recent Update on UV Disinfection to Fulfill the Disinfection Credit Value for Enteric Viruses in Water. Environ. Sci. Technol. 2021, 55, 16283–16298. [Google Scholar] [CrossRef]
- Yu, Z.; Xie, Y.; Li, X.; Liu, W.; Han, J.-L.; Zheng, C.; Zheng, Q.; Zhao, X.; Wang, A. Why the Disinfection Efficiency of Ultraviolet Radiation May Become Unsatisfactory at Low Suspended Solid Concentrations: The Mechanism of Extracellular Polymeric Substances Secretion Induced by Different Particles. Water Research 2025, 274, 123122. [Google Scholar] [CrossRef]
- Azaizeh, H.; Linden, K.G.; Barstow, C.; Kalbouneh, S.; Tellawi, A.; Albalawneh, A.; Gerchman, Y. Constructed Wetlands Combined with UV Disinfection Systems for Removal of Enteric Pathogens and Wastewater Contaminants. Water Science and Technology 2013, 67. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Rijnaarts, H.; Langenhoff, A. Mesocosm Constructed Wetlands to Remove Micropollutants from Wastewater Treatment Plant Effluent: Effect of Matrices and Pre-Treatments. Chemosphere 2022, 305, 135306. [Google Scholar] [CrossRef]
- Demirer, G.S.; Silva, T.N.; Jackson, C.T.; Thomas, J.B.; W Ehrhardt, D.; Rhee, S.Y.; Mortimer, J.C.; Landry, M.P. Nanotechnology to Advance CRISPR-Cas Genetic Engineering of Plants. Nat. Nanotechnol. 2021, 16, 243–250. [Google Scholar] [CrossRef]
- Wang, Y.; Zafar, N.; Ali, Q.; Manghwar, H.; Wang, G.; Yu, L.; Ding, X.; Ding, F.; Hong, N.; Wang, G.; et al. CRISPR/Cas Genome Editing Technologies for Plant Improvement against Biotic and Abiotic Stresses: Advances, Limitations, and Future Perspectives. Cells 2022, 11, 3928. [Google Scholar] [CrossRef]
- Singh, A.; Yadav, V.K.; Chundawat, R.S.; Soltane, R.; Awwad, N.S.; Ibrahium, H.A.; Yadav, K.K.; Vicas, S.I. Enhancing Plant Growth Promoting Rhizobacterial Activities through Consortium Exposure: A Review. Front. Bioeng. Biotechnol. 2023, 11, 1099999. [Google Scholar] [CrossRef]
- Ragland, C.J.; Shih, K.Y.; Dinneny, J.R. Choreographing Root Architecture and Rhizosphere Interactions through Synthetic Biology. Nat Commun 2024, 15, 1370. [Google Scholar] [CrossRef] [PubMed]
- Haskett, T.L.; Tkacz, A.; Poole, P.S. Engineering Rhizobacteria for Sustainable Agriculture. ISME J. 2021, 15, 949–964. [Google Scholar] [CrossRef]
- Chemla, Y.; Sweeney, C.J.; Wozniak, C.A.; Voigt, C.A. Design and Regulation of Engineered Bacteria for Environmental Release. Nat. Microbiol. 2025, 10, 281–300. [Google Scholar] [CrossRef] [PubMed]
- Lea-Smith, D.J.; Hassard, F.; Coulon, F.; Partridge, N.; Horsfall, L.; Parker, K.D.J.; Smith, R.D.J.; McCarthy, R.R.; McKew, B.; Gutierrez, T.; et al. Engineering Biology Applications for Environmental Solutions: Potential and Challenges. Nat. Commun. 2025, 16, 3538. [Google Scholar] [CrossRef]
- Mabrouki, J.; Fattah, G.; Kherraf, S.; Abrouki, Y.; Azrour, M.; Hajjaji, S.E. Artificial Intelligence System for Intelligent Monitoring and Management of Water Treatment Plants. In Emerging Real-World Applications of Internet of Things; CRC Press: Boca Raton, 2022; pp. 69–87. ISBN 978-1-003-30420-3. [Google Scholar]
- Reymond, P.; Chandragiri, R.; Ulrich, L. Governance Arrangements for the Scaling Up of Small-Scale Wastewater Treatment and Reuse Systems – Lessons From India. Front. Environ. Sci. 2020, 8, 72. [Google Scholar] [CrossRef]
- Garrido-Baserba, M.; Sedlak, D.L.; Molinos-Senante, M.; Barnosell, I.; Schraa, O.; Rosso, D.; Verdaguer, M.; Poch, M. Using Water and Wastewater Decentralization to Enhance the Resilience and Sustainability of Cities. Nat. Water 2024, 2, 953–974. [Google Scholar] [CrossRef]
- Chen, H. Surface-Flow Constructed Treatment Wetlands for Pollutant Removal: Applications and Perspectives. Wetlands (Wilmington) 2011, 31, 805–814. [Google Scholar] [CrossRef]
- Prasse, C.; Wenk, J.; Jasper, J.T.; Ternes, T.A.; Sedlak, D.L. Co-Occurrence of Photochemical and Microbiological Transformation Processes in Open-Water Unit Process Wetlands. Environ. Sci. Technol. 2015, 49, 14136–14145. [Google Scholar] [CrossRef]
- Thakur, T.; Barya, M.; Dutta, J.; Mukherjee, P.; Thakur, A.; Swamy, S.; Anderson, J. Integrated Phytobial Remediation of Dissolved Pollutants from Domestic Wastewater through Constructed Wetlands: An Interactive Macrophyte-Microbe-Based Green and Low-Cost Decontamination Technology with Prospective Resource Recovery. Water 2023, 15, 3877. [Google Scholar] [CrossRef]
- Tao, W.; Sauba, K.; Fattah, K.P.; Smith, J.R. Designing Constructed Wetlands for Reclamation of Pretreated Wastewater and Stormwater. Rev Environ Sci Biotechnol 2017, 16, 37–57. [Google Scholar] [CrossRef]
- Ni, Q.; Wang, T.; Liao, J.; Shi, W.; Huang, Z.; Miao, H.; Wu, P.; Ruan, W. Operational Performances and Enzymatic Activities for Eutrophic Water Treatment by Vertical-Flow and Horizontal-Flow Constructed Wetlands. Water (Basel) 2020, 12, 2007. [Google Scholar] [CrossRef]
- Merino-Solís, M.; Villegas, E.; De Anda, J.; López-López, A. The Effect of the Hydraulic Retention Time on the Performance of an Ecological Wastewater Treatment System: An Anaerobic Filter with a Constructed Wetland. Water (Basel) 2015, 7, 1149–1163. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, D.Q.; Dong, J.W.; Tan, S.K. Constructed Wetlands for Wastewater Treatment in Cold Climate — A Review. Journal of Environmental Sciences 2017, 57, 293–311. [Google Scholar] [CrossRef]
- Rani, A.; Chauhan, M.; Kumar Sharma, P.; Kumari, M.; Mitra, D.; Joshi, S. Microbiological Dimensions and Functions in Constructed Wetlands: A Review. Current Research in Microbial Sciences 2024, 7, 100311. [Google Scholar] [CrossRef]
- Barca, C.; Troesch, S.; Meyer, D.; Drissen, P.; Andrès, Y.; Chazarenc, F. Steel Slag Filters to Upgrade Phosphorus Removal in Constructed Wetlands: Two Years of Field Experiments. Environ. Sci. Technol. 2013, 47, 549–556. [Google Scholar] [CrossRef]
- Zhang, J.; Zou, Y.; Yu, X.; Ding, S.; Yan, J.; Min, Y. Vegetated Steel Slag Substrate Constructed Wetlands Can Achieve High Efficiency Simultaneous Nitrogen and Phosphorus Removal. Front. Environ. Sci. 2022, 10, 947783. [Google Scholar] [CrossRef]
- Ni, Q.; Wang, T.; Liao, J.; Shi, W.; Huang, Z.; Miao, H.; Wu, P.; Ruan, W. Operational Performances and Enzymatic Activities for Eutrophic Water Treatment by Vertical-Flow and Horizontal-Flow Constructed Wetlands. Water 2020, 12, 2007. [Google Scholar] [CrossRef]
- Erdoğan, İ.; Cevher-Keskin, B.; Bilir, Ö.; Hong, Y.; Tör, M. Recent Developments in CRISPR/Cas9 Genome-Editing Technology Related to Plant Disease Resistance and Abiotic Stress Tolerance. Biology (Basel) 2023, 12, 1037. [Google Scholar] [CrossRef]
- Dong, Q.; Bai, S.; Wang, Z.; Zhao, X.; Yang, S.; Ren, N. Virtual Sample Generation Empowers Machine Learning-Based Effluent Prediction in Constructed Wetlands. J. Environ. Manage. 2023, 346, 118961. [Google Scholar] [CrossRef] [PubMed]
- Aman, R.; Mahas, A.; Marsic, T.; Hassan, N.; Mahfouz, M.M. Efficient, Rapid, and Sensitive Detection of Plant RNA Viruses with One-Pot RT-RPA-CRISPR/Cas12a Assay. Front. Microbiol. 2020, 11, 610872. [Google Scholar] [CrossRef]
- Wan, X.; Saltepe, B.; Yu, L.; Wang, B. Programming Living Sensors for Environment, Health and Biomanufacturing. Microbial Biotechnology 2021, 14, 2334–2342. [Google Scholar] [CrossRef]
- Duncker, K.E.; Holmes, Z.A.; You, L. Engineered Microbial Consortia: Strategies and Applications. Microb. Cell Fact. 2021, 20, 211. [Google Scholar] [CrossRef]
- Dong, C.; Huang, Y.-H. The Effect of Key Physiological Features on Roots Oxygen Release of Five Wetland Vegetations. Appl. Ecol. Environ. Res. 2023, 21, 2241–2252. [Google Scholar] [CrossRef]
- Licata, M.; Gennaro, M.C.; Tuttolomondo, T.; Leto, C.; La Bella, S. Research Focusing on Plant Performance in Constructed Wetlands and Agronomic Application of Treated Wastewater - A Set of Experimental Studies in Sicily (Italy). PLoS One 2019, 14, e0219445. [Google Scholar] [CrossRef]
- Vymazal, J. Emergent Plants Used in Free Water Surface Constructed Wetlands: A Review. 2013, 61, 582–592. [CrossRef]
- Suquet, J.; Godo-Pla, L.; Valentí, M.; Verdaguer, M.; Martin, M.J.; Poch, M.; Monclús, H. Development of an Environmental Decision Support System for Enhanced Coagulation in Drinking Water Production. Water 2020, 12, 2115. [Google Scholar] [CrossRef]
- Agaton, C.B.; Guila, P.M.C. Success Factors and Challenges: Implications of Real Options Valuation of Constructed Wetlands as Nature-Based Solutions for Wastewater Treatment. Resources 2024, 13, 11. [Google Scholar] [CrossRef]
- Gorgoglione, A.; Torretta, V. Sustainable Management and Successful Application of Constructed Wetlands: A Critical Review. Sustainability 2018, 10, 3910. [Google Scholar] [CrossRef]
- Li, L.; Zhang, Z.; Shen, Y.; He, B.; Fu, Y.; Kou, S.; Gao, J. Optimized Design of Modular Constructed Wetland for Treating Rural Black–Odorous Water. Water (Basel) 2024, 16, 2492. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, J.; Han, R.; Luo, H.; Chen, L.; Liu, M.; He, B. Modular Constructed Wetlands for Treatment of Rural Domestic Wastewater: Laboratory Performance and Field Application. Sustainability 2025, 17, 4427. [Google Scholar] [CrossRef]
- Hayes, E.K.; Stoddart, A.K.; Gagnon, G.A. Adsorption of SARS-CoV-2 onto Granular Activated Carbon (GAC) in Wastewater: Implications for Improvements in Passive Sampling. Sci. Total Environ. 2022, 847, 157548. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhu, D.; Yao, Z.; Zhu, D.Z. Virus-Prokaryote Interactions Assist Pollutant Removal in Constructed Wetlands. Bioresource Technology 2025, 416, 131791. [Google Scholar] [CrossRef] [PubMed]
- Gomes, J.; Domingues, E.; Frasson, D.; Martins, R.C.; Matos, A.M. Virus Removal from Real Wastewater as an Environmental Management Approach. Molecules 2024, 29, 5601. [Google Scholar] [CrossRef]
- Waly, M.M.; Ahmed, T.; Abunada, Z.; Mickovski, S.B.; Thomson, C. Constructed Wetland for Sustainable and Low-Cost Wastewater Treatment: Review Article. Land 2022, 11, 1388. [Google Scholar] [CrossRef]
- Fernández Ramírez, L.E.; Zamora-Castro, S.A.; Sandoval-Herazo, L.C.; Herrera-May, A.L.; Salgado-Estrada, R.; De La Cruz-Dessavre, D.A. Technological Innovations in the Application of Constructed Wetlands: A Review. Processes (Basel) 2023, 11, 3334. [Google Scholar] [CrossRef]
- Zhao, P.; Wang, R.; Li, B.; Zhang, G.; Chen, Z.; Wang, S.; Hu, X.; Hao, R. Study on the Optimization of Hydrodynamic Characteristics and Pollutant Removal Efficiency in Integrated Vertical Flow Constructed Wetlands. Sci Rep 2025, 15, 6017. [Google Scholar] [CrossRef]
- Rossi, L.; Queloz, P.; Brovelli, A.; Margot, J.; Barry, D.A. Enhancement of Micropollutant Degradation at the Outlet of Small Wastewater Treatment Plants. PLoS ONE 2013, 8, e58864. [Google Scholar] [CrossRef]
- Sattler, S.; Zluwa, I.; Österreicher, D. The “PV Rooftop Garden”: Providing Recreational Green Roofs and Renewable Energy as a Multifunctional System within One Surface Area. Applied Sciences 2020, 10, 1791. [Google Scholar] [CrossRef]
- Cook, L.M.; Larsen, T.A. Towards a Performance-Based Approach for Multifunctional Green Roofs: An Interdisciplinary Review. Build. Environ. 2021, 188, 107489. [Google Scholar] [CrossRef]
- Uludag-Demirer, S.; Olson, N.; Ives, R.; Nshimyimana, J.P.; Rusinek, C.A.; Rose, J.B.; Liao, W. Techno-Economic Analysis of Electrocoagulation on Water Reclamation and Bacterial/Viral Indicator Reductions of a High-Strength Organic Wastewater—Anaerobic Digestion Effluent. Sustainability 2020, 12, 2697. [Google Scholar] [CrossRef]
- Torrens, A.; Folch, M.; Salgot, M. Design and Performance of an Innovative Hybrid Constructed Wetland for Sustainable Pig Slurry Treatment in Small Farms. Front. Environ. Sci. 2021, 8, 577186. [Google Scholar] [CrossRef]
- Li, J.; Hu, Z.; Li, F.; Fan, J.; Zhang, J.; Li, F.; Hu, H. Effect of Oxygen Supply Strategy on Nitrogen Removal of Biochar-Based Vertical Subsurface Flow Constructed Wetland: Intermittent Aeration and Tidal Flow. Chemosphere 2019, 223, 366–374. [Google Scholar] [CrossRef]
- Sallam, M.; Yaseen, A.; Refai, L.; El-Qanni, A. Conceptual Feasibility of a Portable Containerized and Decentralized Wastewater Treatment System for Rural Regions: A Case Study in Palestine. An-Najah University Journal for Research - A 2024, 38, 42–47. [Google Scholar] [CrossRef]
- Kabay, G.; DeCastro, J.; Altay, A.; Smith, K.; Lu, H.-W.; Capossela, A.M.; Moarefian, M.; Aran, K.; Dincer, C. Emerging Biosensing Technologies for the Diagnostics of Viral Infectious Diseases. Adv. Mater. 2022, 34, e2201085. [Google Scholar] [CrossRef]
- Xiao, M.; Tian, F.; Liu, X.; Zhou, Q.; Pan, J.; Luo, Z.; Yang, M.; Yi, C. Virus Detection: From State-of-the-Art Laboratories to Smartphone-Based Point-of-Care Testing. Advanced Science 2022, 9, 2105904. [Google Scholar] [CrossRef]
- Ji, T.; Fang, X.; Gao, Y.; Yu, K.; Gao, X. Research Progress on the Application of RPA-CRISPR/Cas12a in the Rapid Visual Detection of Pathogenic Microorganisms. Front. Cell. Infect. Microbiol. 2025, 15, 1640938. [Google Scholar] [CrossRef]
- Ventura, J.-R.S.; Tulipan, J.U.; Banawa, A.; Umali, K.D.C.; Villanueva, J.A.L. Advancements and Challenges in Decentralized Wastewater Treatment: A Comprehensive Review. Desalination and Water Treatment 2024, 320, 100830. [Google Scholar] [CrossRef]
- Badawi, A.K.; Salama, R.S.; Mostafa, M.M.M. Natural-Based Coagulants/Flocculants as Sustainable Market-Valued Products for Industrial Wastewater Treatment: A Review of Recent Developments. RSC Adv. 2023, 13, 19335–19355. [Google Scholar] [CrossRef]
- Proano-Pena, G.; Carrano, A.L.; Blersch, D.M. Analysis of Very-High Surface Area 3D-Printed Media in a Moving Bed Biofilm Reactor for Wastewater Treatment. PLoS ONE 2020, 15, e0238386. [Google Scholar] [CrossRef]
- Roy Barman, S.; Gavit, P.; Chowdhury, S.; Chatterjee, K.; Nain, A. 3D-Printed Materials for Wastewater Treatment. JACS Au 2023, 3, 2930–2947. [Google Scholar] [CrossRef]
- Del Valle, I.; Fulk, E.M.; Kalvapalle, P.; Silberg, J.J.; Masiello, C.A.; Stadler, L.B. Translating New Synthetic Biology Advances for Biosensing into the Earth and Environmental Sciences. Front. Microbiol. 2020, 11, 618373. [Google Scholar] [CrossRef]
- Khan, A.; Ostaku, J.; Aras, E.; Safak Seker, U.O. Combating Infectious Diseases with Synthetic Biology. ACS Synth. Biol. 2022, 11, 528–537. [Google Scholar] [CrossRef]
- Wölle, J.; Steinmetz, H.; Hansen, J.; Einsfeld, K.; Ebert, A. An Intelligent Visualisation and Decision Support System for Decentralised Wastewater Treatment Plants. Water Science and Technology 2007, 56, 183–191. [Google Scholar] [CrossRef]
- Pasternak, G.; Greenman, J.; Ieropoulos, I. Self-Powered, Autonomous Biological Oxygen Demand Biosensor for Online Water Quality Monitoring. Sens. Actuators B Chem. 2017, 244, 815–822. [Google Scholar] [CrossRef]
- McLamore, E.S.; Datta, S.P.A. A Connected World: System-Level Support through Biosensors. Annu. Rev. Anal. Chem. (Palo Alto Calif.) 2023, 16, 285–309. [Google Scholar] [CrossRef]
- Mohamed, A.Y.A.; Siggins, A.; Healy, M.G.; Ó hUallacháin, D.; Fenton, O.; Tuohy, P. A Novel Hybrid Coagulation-Constructed Wetland System for the Treatment of Dairy Wastewater. Sci. Total Environ. 2022, 847, 157567. [Google Scholar] [CrossRef]
- Crovella, T.; Paiano, A.; Falciglia, P.P.; Lagioia, G.; Ingrao, C. Wastewater Recovery for Sustainable Agricultural Systems in the Circular Economy - A Systematic Literature Review of Life Cycle Assessments. Sci. Total Environ. 2024, 912, 169310. [Google Scholar] [CrossRef]
- Christou, A.; Beretsou, V.G.; Iakovides, I.C.; Karaolia, P.; Michael, C.; Benmarhnia, T.; Chefetz, B.; Donner, E.; Gawlik, B.M.; Lee, Y.; et al. Sustainable Wastewater Reuse for Agriculture. Nat. Rev. Earth Environ. 2024, 5, 504–521. [Google Scholar] [CrossRef]
- Martinez-Guerra, E.; Castillo-Valenzuela, J.; Gude, V.G. Wetlands for Wastewater Treatment. Water Environ. Res. 2018, 90, 1537–1562. [Google Scholar] [CrossRef]
- Ghimire, U.; Nandimandalam, H.; Martinez-Guerra, E.; Gude, V.G. Wetlands for Wastewater Treatment. Water Environ. Res. 2019, 91, 1378–1389. [Google Scholar] [CrossRef]
- Gruyer, N.; Dorais, M.; Zagury, G.J.; Alsanius, B.W. Removal of Plant Pathogens from Recycled Greenhouse Wastewater Using Constructed Wetlands. Agric. Water Manag. 2013, 117, 153–158. [Google Scholar] [CrossRef]
- Younis, B.A.; Mahoney, L.E.; Yao, S. Field Evaluation of a Novel UV Water Disinfection System for Use in Underserved Rural Communities. Water Environment Research 2019, 91, 75–82. [Google Scholar] [CrossRef]
- Kamarasu, P.; Hsu, H.-Y.; Moore, M.D. Research Progress in Viral Inactivation Utilizing Human Norovirus Surrogates. Front. Sustain. Food Syst. 2018, 2. [Google Scholar] [CrossRef]
- Pal, D.B.; Tiwari, A.K.; Srivastava, N.; Ahmad, I.; Abohashrh, M.; Gupta, V.K. Biomass Valorization of Eichhornia Crassipes Root Using Thermogravimetric Analysis. Environ. Res. 2022, 214, 114046. [Google Scholar] [CrossRef]
- Bi, X.; Liu, D.; Wang, L.; Huo, Y.; Fu, M.-L.; Sun, W.; Yuan, B. Inactivation and Genome Damage of Rotavirus and a Human Norovirus Surrogate by Monochloramine Treatment and Sequential Application with UV. Water Res. 2022, 226, 119309. [Google Scholar] [CrossRef]
- Lau, M.; Monis, P.T.; King, B.J. The Efficacy of Current Treatment Processes to Remove, Inactivate, or Reduce Environmental Bloom-Forming Escherichia Coli. Microbiol. Spectr. 2024, 12, e0085624. [Google Scholar] [CrossRef]
- Gao, R.; Gao, S.-H.; Li, J.; Su, Y.; Huang, F.; Liang, B.; Fan, L.; Guo, J.; Wang, A. Emerging Technologies for the Control of Biological Contaminants in Water Treatment: A Critical Review. Engineering (Beijing) 2025, 48, 185–204. [Google Scholar] [CrossRef]
- Al-Hazmi, H.E.; Shokrani, H.; Shokrani, A.; Jabbour, K.; Abida, O.; Mousavi Khadem, S.S.; Habibzadeh, S.; Sonawane, S.H.; Saeb, M.R.; Bonilla-Petriciolet, A.; et al. Recent Advances in Aqueous Virus Removal Technologies. Chemosphere 2022, 305, 135441. [Google Scholar] [CrossRef]
- Gupta, A.K.; Majumder, A.; Ghosal, P.S. Introduction to Modular Wastewater Treatment System and Its Significance. In Modular Treatment Approach for Drinking Water and Wastewater; Elsevier, 2022; pp. 81–106. ISBN 978-0-323-85421-4.
- Wen, X.; Meng, F.; Li, S. Analyzing the Effect of Public Private Partnership Mode on Sewage Treatment in China. Sci Rep 2024, 14, 9531. [Google Scholar] [CrossRef]
- Luca, A.-V.; Simon-Várhelyi, M.; Mihály, N.-B.; Cristea, V.-M. Data Driven Detection of Different Dissolved Oxygen Sensor Faults for Improving Operation of the WWTP Control System. Processes (Basel) 2021, 9, 1633. [Google Scholar] [CrossRef]
- Ntalaperas, D.; Christophoridis, C.; Angelidis, I.; Iossifidis, D.; Touloupi, M.-F.; Vergeti, D.; Politi, E. Intelligent Tools to Monitor, Control and Predict Wastewater Reclamation and Reuse. Sensors (Basel) 2022, 22, 3068. [Google Scholar] [CrossRef]
- Reynaert, E.; Nagappa, D.; Sigrist, J.A.; Morgenroth, E. Ensuring Microbial Water Quality for On-Site Water Reuse: Importance of Online Sensors for Reliable Operation. Water Research X 2024, 22, 100215. [Google Scholar] [CrossRef]
- D, N.S.; C, M.; R. Kshirsagar, P.; Tirth, V.; Islam, S.; Qaiyum, S.; B, S.; Al Duhayyim, M.; Waji, Y.A. IOT Based Smart Wastewater Treatment Model for Industry 4.0 Using Artificial Intelligence. Scientific Programming 2022, 2022, 1–11. [CrossRef]
- Srivastava, R.R.; Singh, P.K. Reuse-Focused Selection of Appropriate Technologies for Municipal Wastewater Treatment: A Multi-Criteria Approach. Int. J. Environ. Sci. Technol. 2022, 19, 12505–12522. [Google Scholar] [CrossRef]
- Eseoglu, G.; Yapsakli, K.; Tozan, H.; Vayvay, O. A Novel Fuzzy Framework for Technology Selection of Sustainable Wastewater Treatment Plants Based on TODIM Methodology in Developing Urban Areas. Sci. Rep. 2022, 12, 8800. [Google Scholar] [CrossRef]
- Zagklis, D.P.; Bampos, G. Tertiary Wastewater Treatment Technologies: A Review of Technical, Economic, and Life Cycle Aspects. Processes 2022, 10, 2304. [Google Scholar] [CrossRef]
- Rottinghaus, A.G.; Ferreiro, A.; Fishbein, S.R.S.; Dantas, G.; Moon, T.S. Genetically Stable CRISPR-Based Kill Switches for Engineered Microbes. Nat Commun 2022, 13, 672. [Google Scholar] [CrossRef]
- Joshi, D.; Kaushik, A.; Kumar, R.; Arya, A.; Santoyo, G.; Singh, V.K.; Kashyap, N.; Solanki, M.K.; Kumari, M.; Bhardwaj, N.; et al. Improving Plant Performance through Microbiome Manipulation: The Potential Role of Current Bioengineering Approaches. Bacteria 2025, 4, 12. [Google Scholar] [CrossRef]
- Puyol, D.; Batstone, D.J.; Hülsen, T.; Astals, S.; Peces, M.; Krömer, J.O. Resource Recovery from Wastewater by Biological Technologies: Opportunities, Challenges, and Prospects. Front. Microbiol. 2017, 7. [Google Scholar] [CrossRef]
- Mehta, N.; Shah, K.J.; Lin, Y.-I.; Sun, Y.; Pan, S.-Y. Advances in Circular Bioeconomy Technologies: From Agricultural Wastewater to Value-Added Resources. Environments 2021, 8, 20. [Google Scholar] [CrossRef]
- Sharma, M.; Agarwal, S.; Agarwal Malik, R.; Kumar, G.; Pal, D.B.; Mandal, M.; Sarkar, A.; Bantun, F.; Haque, S.; Singh, P.; et al. Recent Advances in Microbial Engineering Approaches for Wastewater Treatment: A Review. Bioengineered 2023, 14, 2184518. [Google Scholar] [CrossRef]
- López-Serrano, M.J.; Lakho, F.H.; Van Hulle, S.W.H.; Batlles-delaFuente, A. Life Cycle Cost Assessment and Economic Analysis of a Decentralized Wastewater Treatment to Achieve Water Sustainability within the Framework of Circular Economy. Oecon. Copernic. 2023, 14, 103–133. [Google Scholar] [CrossRef]
- Bhatt, A.; Arora, P.; Prajapati, S.K. Occurrence, Fates and Potential Treatment Approaches for Removal of Viruses from Wastewater: A Review with Emphasis on SARS-CoV-2. J. Environ. Chem. Eng. 2020, 8, 104429. [Google Scholar] [CrossRef]
- Mousazadeh, M.; Kabdaşlı, I.; Khademi, S.; Sandoval, M.A.; Moussavi, S.P.; Malekdar, F.; Gilhotra, V.; Hashemi, M.; Dehghani, M.H. A Critical Review on the Existing Wastewater Treatment Methods in the COVID-19 Era: What Is the Potential of Advanced Oxidation Processes in Combatting Viral Especially SARS-CoV-2? Journal of Water Process Engineering 2022, 49. [Google Scholar] [CrossRef]
- Bleotu, C.; Matei, L.; Dragu, L.D.; Necula, L.G.; Pitica, I.M.; Chivu-Economescu, M.; Diaconu, C.C. Viruses in Wastewater-A Concern for Public Health and the Environment. Microorganisms 2024, 12, 1430. [Google Scholar] [CrossRef]
- Scheidweiler, D.; Peter, H.; Pramateftaki, P.; De Anna, P.; Battin, T.J. Unraveling the Biophysical Underpinnings to the Success of Multispecies Biofilms in Porous Environments. The ISME Journal 2019, 13, 1700–1710. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Tabasi, Z.A.; Liu, J.; Zhang, B.H.; Zhao, Y. Recent Advances in Functional Materials for Wastewater Treatment: From Materials to Technological Innovations. J. Mar. Sci. Eng. 2022, 10, 534. [Google Scholar] [CrossRef]
- Samari-Kermani, M.; De Vries, E.T.; Schijven, J.F.; Raoof, A. A Closer Look at the Role of Biofilms in Water Filtration: Bridging Microscopic Insights with System Performance. Journal of Water Process Engineering 2024, 67, 106104. [Google Scholar] [CrossRef]
- Checconi, P.; De Angelis, M.; Marcocci, M.E.; Fraternale, A.; Magnani, M.; Palamara, A.T.; Nencioni, L. Redox-Modulating Agents in the Treatment of Viral Infections. Int. J. Mol. Sci. 2020, 21, 4084. [Google Scholar] [CrossRef]
- Manjunatha, L.; Rajashekara, H.; Uppala, L.S.; Ambika, D.S.; Patil, B.; Shankarappa, K.S.; Nath, V.S.; Kavitha, T.R.; Mishra, A.K. Mechanisms of Microbial Plant Protection and Control of Plant Viruses. Plants 2022, 11, 3449. [Google Scholar] [CrossRef]
- Wen, H.; Perdrial, J.; Abbott, B.W.; Bernal, S.; Dupas, R.; Godsey, S.E.; Harpold, A.; Rizzo, D.; Underwood, K.; Adler, T.; et al. Temperature Controls Production but Hydrology Regulates Export of Dissolved Organic Carbon at the Catchment Scale. Hydrol. Earth Syst. Sci. 2020, 24, 945–966. [Google Scholar] [CrossRef]
- Baetge, N.; Behrenfeld, M.J.; Fox, J.; Halsey, K.H.; Mojica, K.D.A.; Novoa, A.; Stephens, B.M.; Carlson, C.A. The Seasonal Flux and Fate of Dissolved Organic Carbon through Bacterioplankton in the Western North Atlantic. Front. Microbiol. 2021, 12, 669883. [Google Scholar] [CrossRef]
- Berggren, M.; Guillemette, F.; Bieroza, M.; Buffam, I.; Deininger, A.; Hawkes, J.A.; Kothawala, D.N.; LaBrie, R.; Lapierre, J.-F.; Murphy, K.R.; et al. Unified Understanding of Intrinsic and Extrinsic Controls of Dissolved Organic Carbon Reactivity in Aquatic Ecosystems. Ecology 2022, 103, e3763. [Google Scholar] [CrossRef]
- Xagoraraki, I.; Yin, Z.; Svambayev, Z. Fate of Viruses in Water Systems. J. Environ. Eng. 2014, 140, 04014020. [Google Scholar] [CrossRef]
- Nasir, A.M.; Adam, M.R.; Mohamad Kamal, S.N.E.A.; Jaafar, J.; Othman, M.H.D.; Ismail, A.F.; Aziz, F.; Yusof, N.; Bilad, M.R.; Mohamud, R.; et al. A Review of the Potential of Conventional and Advanced Membrane Technology in the Removal of Pathogens from Wastewater. Sep. Purif. Technol. 2022, 286, 120454. [Google Scholar] [CrossRef] [PubMed]
- Asheghmoalla, M.; Mehrvar, M. Integrated and Hybrid Processes for the Treatment of Actual Wastewaters Containing Micropollutants: A Review on Recent Advances. Processes (Basel) 2024, 12, 339. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Han, Q.; Ju, K.; Wei, D.; Sun, Y.; Wan, Q. Effects of Hydraulic Retention Time (HRT)and Packing Height on the Performanceof Homemade Ceramsite-Soil Constructed Wetlandfor Rural Domestic Wastewater Treatment. Pol. J. Environ. Stud. 2021, 30, 4845–4854. [Google Scholar] [CrossRef]
- Miyazaki, C.K.; Morvannou, A.; Petitjean, A.; Nivala, J.; Molle, P. Hydraulic Characterization of a Hybrid Aerated Vertical and Horizontal Treatment Wetland. Ecol. Eng. 2024, 206, 107301. [Google Scholar] [CrossRef]
- Kulshreshtha, N.M.; Kumar, A.; Bisht, G.; Pasha, S.; Kumar, R. Usefulness of Organic Acid Produced by Exiguobacterium Sp. 12/1 on Neutralization of Alkaline Wastewater. ScientificWorldJournal 2012, 2012, 345101. [Google Scholar] [CrossRef]
- Monje, V.; Nobel, P.; Junicke, H.; Kjellberg, K.; Gernaey, K.V.; Flores-Alsina, X. Assessment of Alkaline Stabilization Processes in Industrial Waste Streams Using a Model-Based Approach. J. Environ. Manage. 2021, 293, 112806. [Google Scholar] [CrossRef]
- Shaikh, I.N.; Ahammed, M.M. Granular Media Filtration for On-Site Treatment of Greywater: A Review. Water Science and Technology 2022, 86, 992–1016. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Upadhyay, S.; Rani, A.; Sharma, P.K.; Rawat, J.M.; Rawat, B.; Prashant; Bhattacharya, P. Assessment of Pathogen Removal Efficiency of Vertical Flow Constructed Wetland Treating Septage. Sci Rep 2023, 13, 18703. [CrossRef]
- Rimoldi, S.G.; Stefani, F.; Gigantiello, A.; Polesello, S.; Comandatore, F.; Mileto, D.; Maresca, M.; Longobardi, C.; Mancon, A.; Romeri, F.; et al. Presence and Infectivity of SARS-CoV-2 Virus in Wastewaters and Rivers. Science of The Total Environment 2020, 744, 140911. [Google Scholar] [CrossRef]
- Serra-Compte, A.; González, S.; Arnaldos, M.; Berlendis, S.; Courtois, S.; Loret, J.F.; Schlosser, O.; Yáñez, A.M.; Soria-Soria, E.; Fittipaldi, M.; et al. Elimination of SARS-CoV-2 along Wastewater and Sludge Treatment Processes. Water Research 2021, 202, 117435. [Google Scholar] [CrossRef]
- Alamin, M.; Tsuji, S.; Hata, A.; Hara-Yamamura, H.; Honda, R. Selection of Surrogate Viruses for Process Control in Detection of SARS-CoV-2 in Wastewater. Sci. Total Environ. 2022, 823, 153737. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Cashman, S.; Gaglione, A.; Mosley, J.; Weiss, L.; Ma, X.C.; Cashdollar, J.; Garland, J. Holistic Analysis of Urban Water Systems in the Greater Cincinnati Region: (1) Life Cycle Assessment and Cost Implications. Water Research X 2019, 2, 100015. [Google Scholar] [CrossRef] [PubMed]
- Whitby, G.E.; Lawal, O.; Ropic, P.; Shima, S.; Ferran, B.; Dussert, B. Uniform Protocol for Wastewater UV Validation Applications. Water Practice and Technology 2013, 8, 83–94. [Google Scholar] [CrossRef]
- Liyanage, S.; Lay, M.; Glasgow, G.; Tanner, C.; Craggs, R.; Northcott, G. Nature Based Solutions for Removal of Steroid Estrogens in Wastewater. Front. Microbiol. 2024, 15, 1437795. [Google Scholar] [CrossRef]
- Muoghalu, C.C.; Owusu, P.A.; Lebu, S.; Nakagiri, A.; Semiyaga, S.; Iorhemen, O.T.; Manga, M. Biochar as a Novel Technology for Treatment of Onsite Domestic Wastewater: A Critical Review. Front. Environ. Sci. 2023, 11. [Google Scholar] [CrossRef]
- Hama Aziz, K.H.; Fatah, N.M.; Muhammad, K.T. Advancements in Application of Modified Biochar as a Green and Low-Cost Adsorbent for Wastewater Remediation from Organic Dyes. R. Soc. Open Sci. 2024, 11, 232033. [Google Scholar] [CrossRef]
- Chen, F.; Chen, L.; Yan, Z.; Xu, J.; Feng, L.; He, N.; Guo, M.; Zhao, J.; Chen, Z.; Chen, H.; et al. Recent Advances of CRISPR-Based Genome Editing for Enhancing Staple Crops. Front. Plant Sci. 2024, 15, 1478398. [Google Scholar] [CrossRef]
- Jovanović, I.; Frantová, N.; Zouhar, J. A Sword or a Buffet: Plant Endomembrane System in Viral Infections. Front. Plant Sci. 2023, 14, 1226498. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.; Hu, T.; Wang, Y.; Lozano-Durán, R.; Yang, X.; Zhou, X. Two Viral Proteins Translated from One Open Reading Frame Target Different Layers of Plant Defense. Plant Commun. 2024, 5, 100788. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Takaki, Y.; Yoshida-Takashima, Y.; Hiraoka, S.; Kurosawa, K.; Nunoura, T.; Takai, K. A Sequential One-Pot Approach for Rapid and Convenient Characterization of Putative Restriction-Modification Systems. mSystems 2023, 8, e00817-23. [Google Scholar] [CrossRef]
- Hoffmann, S.A.; Diggans, J.; Densmore, D.; Dai, J.; Knight, T.; Leproust, E.; Boeke, J.D.; Wheeler, N.; Cai, Y. Safety by Design: Biosafety and Biosecurity in the Age of Synthetic Genomics. iScience 2023, 26, 106165. [Google Scholar] [CrossRef]
- George, D.R.; Danciu, M.; Davenport, P.W.; Lakin, M.R.; Chappell, J.; Frow, E.K. A Bumpy Road Ahead for Genetic Biocontainment. Nat. Commun. 2024, 15, 650. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Wu, Y.; Sørensen, S.J.; Zhang, M.; Dai, K.; Gao, C.; Qu, C.; Huang, Q.; Cai, P. The Mitigation of Spatial Constraint in Porous Environments Enhances Biofilm Phylogenetic and Functional Diversity. Microbiome 2025, 13, 84. [Google Scholar] [CrossRef] [PubMed]
- Eckerstorfer, M.F.; Dolezel, M.; Engelhard, M.; Giovannelli, V.; Grabowski, M.; Heissenberger, A.; Lener, M.; Reichenbecher, W.; Simon, S.; Staiano, G.; et al. Recommendations for the Assessment of Potential Environmental Effects of Genome-Editing Applications in Plants in the EU. Plants 2023, 12, 1764. [Google Scholar] [CrossRef]
- Seddon, N.; Smith, A.; Smith, P.; Key, I.; Chausson, A.; Girardin, C.; House, J.; Srivastava, S.; Turner, B. Getting the Message Right on Nature-based Solutions to Climate Change. Global Change Biology 2021, 27, 1518–1546. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, J.; Pereira, A.; Jefferies, L.; Kemp, A.H.; Isham, A. Nature-Based Interventions for Individual, Collective and Planetary Wellbeing: A Protocol for a Scoping Review. PLoS One 2025, 20, e0314591. [Google Scholar] [CrossRef] [PubMed]
- Butler, D.; Ward, S.; Sweetapple, C.; Astaraie-Imani, M.; Diao, K.; Farmani, R.; Fu, G. Reliable, Resilient and Sustainable Water Management: The Safe & SuRe Approach. Global Chall. 2017, 1, 63–77. [Google Scholar] [CrossRef]
- Gerullis, M.; Pieruschka, R.; Fahrner, S.; Hartl, L.; Schurr, U.; Heckelei, T. From Genes to Policy: Mission-Oriented Governance of Plant-Breeding Research and Technologies. Front. Plant Sci. 2023, 14, 1235175. [Google Scholar] [CrossRef]
- Satyam, S.; Patra, S. Innovations and Challenges in Adsorption-Based Wastewater Remediation: A Comprehensive Review. Heliyon 2024, 10, e29573. [Google Scholar] [CrossRef]
- Liu, P.; Wu, X.; Gong, B.; Lü, G.; Li, J.; Gao, H. Review of the Mechanisms by Which Transcription Factors and Exogenous Substances Regulate ROS Metabolism under Abiotic Stress. Antioxidants (Basel) 2022, 11, 2106. [Google Scholar] [CrossRef]
- Bhandari, G.; Gangola, S.; Bhatt, P.; Rafatullah, M. Editorial: Potential of the Plant Rhizomicrobiome for Bioremediation of Contaminants in Agroecosystems. Front. Plant Sci. 2024, 15, 1397360. [Google Scholar] [CrossRef]
- Xu, Q.; Zhang, W.; Dong, C.; Sreeprasad, T.S.; Xia, Z. Biomimetic Self-Cleaning Surfaces: Synthesis, Mechanism and Applications. J. R. Soc. Interface. 2016, 13, 20160300. [Google Scholar] [CrossRef]
- Perumanath, S.; Pillai, R.; Borg, M.K. Contaminant Removal from Nature’s Self-Cleaning Surfaces. Nano Lett. 2023, 23, 4234–4241. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, M. Prediction of River Dissolved Oxygen (DO) Based on Multi-Source Data and Various Machine Learning Coupling Models. PLoS ONE 2025, 20, e0319256. [Google Scholar] [CrossRef]
- Duarte, M.S.; Martins, G.; Oliveira, P.; Fernandes, B.; Ferreira, E.C.; Alves, M.M.; Lopes, F.; Pereira, M.A.; Novais, P. A Review of Computational Modeling in Wastewater Treatment Processes. ACS ES T Water 2024, 4, 784–804. [Google Scholar] [CrossRef]
- McClary-Gutierrez, J.S.; Aanderud, Z.T.; Al-Faliti, M.; Duvallet, C.; Gonzalez, R.; Guzman, J.; Holm, R.H.; Jahne, M.A.; Kantor, R.S.; Katsivelis, P.; et al. Standardizing Data Reporting in the Research Community to Enhance the Utility of Open Data for SARS-CoV-2 Wastewater Surveillance. Environ. Sci. (Camb.) 2021, 9. [Google Scholar] [CrossRef]
- Chen, Y.; Yao, Z.; Sun, Y.; Wang, E.; Tian, C.; Sun, Y.; Liu, J.; Sun, C.; Tian, L. Current Studies of the Effects of Drought Stress on Root Exudates and Rhizosphere Microbiomes of Crop Plant Species. Int. J. Mol. Sci. 2022, 23, 2374. [Google Scholar] [CrossRef]
- Murtaza, G.; Ahmed, Z.; Eldin, S.M.; Ali, B.; Bawazeer, S.; Usman, M.; Iqbal, R.; Neupane, D.; Ullah, A.; Khan, A.; et al. Biochar-Soil-Plant Interactions: A Cross Talk for Sustainable Agriculture under Changing Climate. Front. Environ. Sci. 2023, 11, 1059449. [Google Scholar] [CrossRef]
- Nepal, J.; Ahmad, W.; Munsif, F.; Khan, A.; Zou, Z. Advances and Prospects of Biochar in Improving Soil Fertility, Biochemical Quality, and Environmental Applications. Front. Environ. Sci. 2023, 11. [Google Scholar] [CrossRef]
- Paredes Méndez, D.; Pérez-Sánchez, M.; Sánchez-Romero, F.J.; Coronado-Hernández, O.E. Assessment of the Effectiveness of Green Infrastructure Interventions to Enhance the Ecosystem Services in Developing Countries. Urban Sci. 2025, 9, 85. [Google Scholar] [CrossRef]
Figure 1.
Transmission pathway of pathogenic enteric viruses. Viruses transmit from infected to susceptible individuals through environmental routes. Transmission occurs when bodily fluids (sweat, droplets, tears, blood, feces, saliva) spread via different pathways (air, water, solids, vectors). Susceptible individuals may contract infections through multiple entry points (eyes, nose, mouth, blood, skin).
Figure 1.
Transmission pathway of pathogenic enteric viruses. Viruses transmit from infected to susceptible individuals through environmental routes. Transmission occurs when bodily fluids (sweat, droplets, tears, blood, feces, saliva) spread via different pathways (air, water, solids, vectors). Susceptible individuals may contract infections through multiple entry points (eyes, nose, mouth, blood, skin).
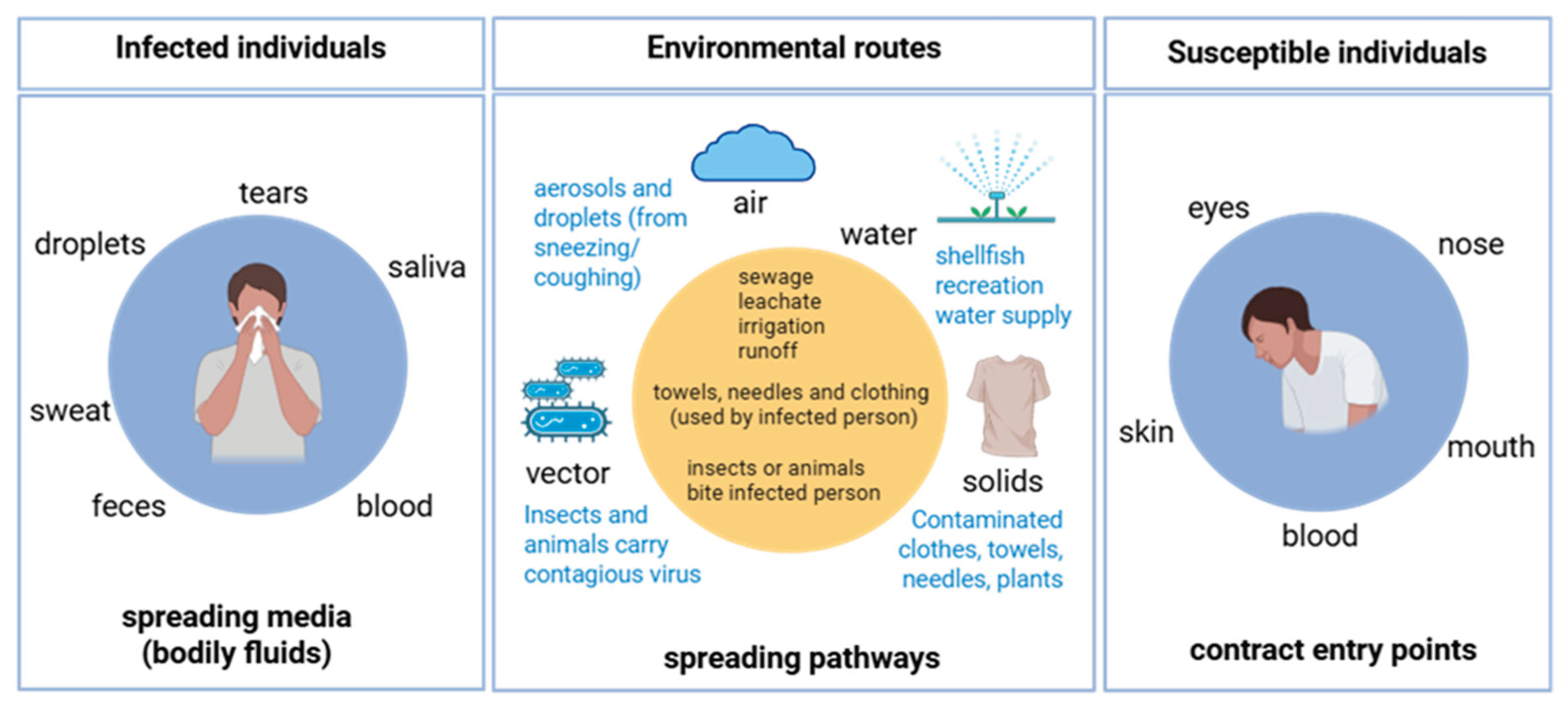
Figure 2.
Studies on the phytoremediation of viruses and other pathogens from wastewater over five decades (1976–2025). Conventional and advanced wastewater studies have evolved and become interconnected, leading to overlaps between treatment domains. Most studies (>90%) identified microbial contaminants as waterborne pathogens and emphasized their combined treatments, resulting in overlaps when categorizing them (i.e., bacteria: 25, viruses: 24, parasitic protozoa: 2, and helminths: 17, totaling 25). Data from the Dimensions Research Database (https://app.dimensions.ai) as of 31/03/2025 showed an evolution combining conventional and advanced methods, recognizing hybrid treatment as promising for microbial contaminants.
Figure 2.
Studies on the phytoremediation of viruses and other pathogens from wastewater over five decades (1976–2025). Conventional and advanced wastewater studies have evolved and become interconnected, leading to overlaps between treatment domains. Most studies (>90%) identified microbial contaminants as waterborne pathogens and emphasized their combined treatments, resulting in overlaps when categorizing them (i.e., bacteria: 25, viruses: 24, parasitic protozoa: 2, and helminths: 17, totaling 25). Data from the Dimensions Research Database (https://app.dimensions.ai) as of 31/03/2025 showed an evolution combining conventional and advanced methods, recognizing hybrid treatment as promising for microbial contaminants.
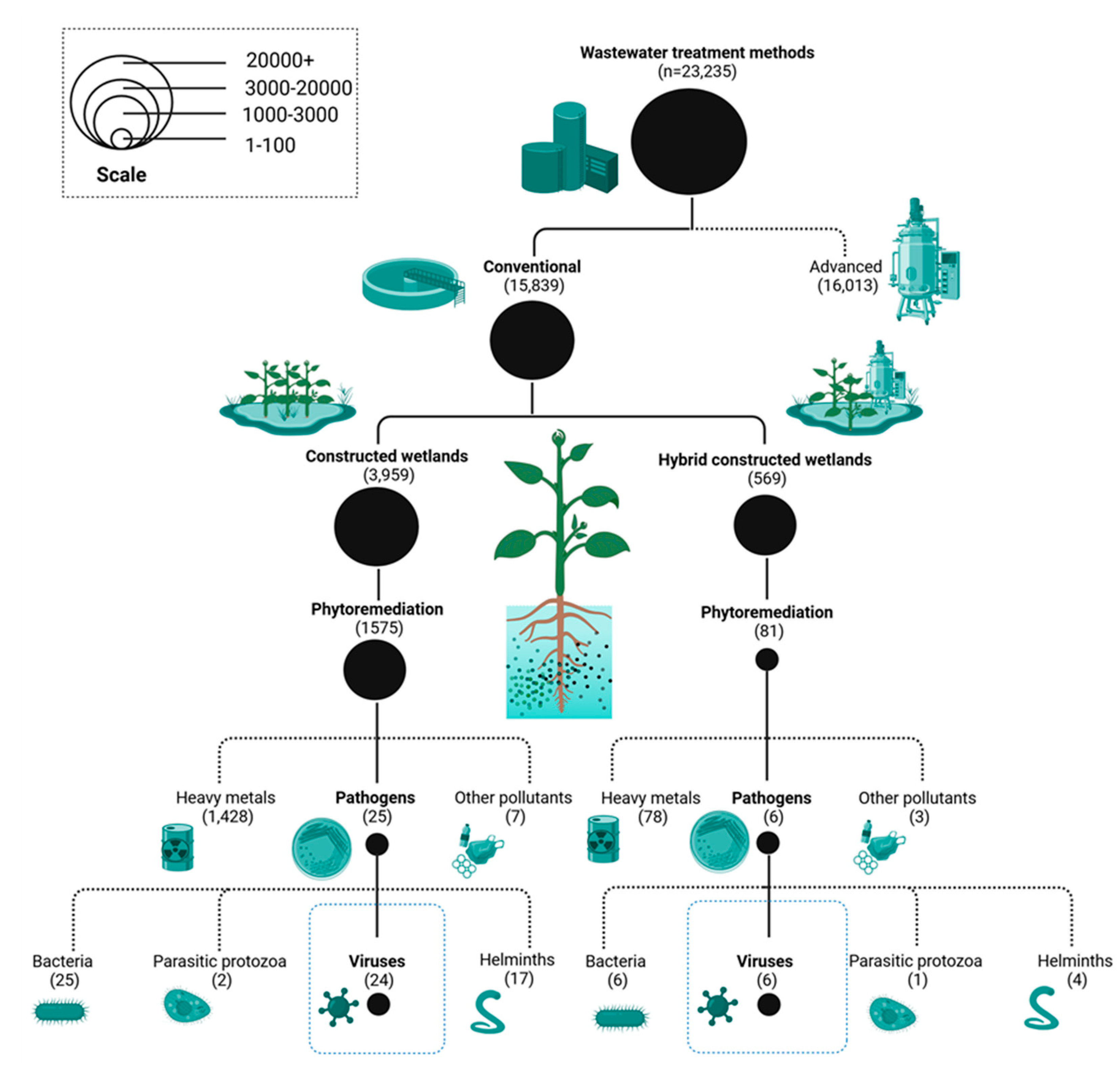
Figure 3.
Conceptual antiviral phytoremediation of viral pathogens in wastewater. Effluents with infectious viral particles could be rendered inactivated/degraded via a series of complex removal mechanisms. Active/partially active viruses initially encounter sorption/filtration (phytostabilization/rhizofiltration) followed by subsequent reduction involving rhizosphere mediated inactivation (microbial consortia/root exudes trigger rhizodegradation), internalization (viral particle uptake via endocytosis/membrane fusion), and intracellular inactivation (phytoinactivation and phytodegradation).
Figure 3.
Conceptual antiviral phytoremediation of viral pathogens in wastewater. Effluents with infectious viral particles could be rendered inactivated/degraded via a series of complex removal mechanisms. Active/partially active viruses initially encounter sorption/filtration (phytostabilization/rhizofiltration) followed by subsequent reduction involving rhizosphere mediated inactivation (microbial consortia/root exudes trigger rhizodegradation), internalization (viral particle uptake via endocytosis/membrane fusion), and intracellular inactivation (phytoinactivation and phytodegradation).
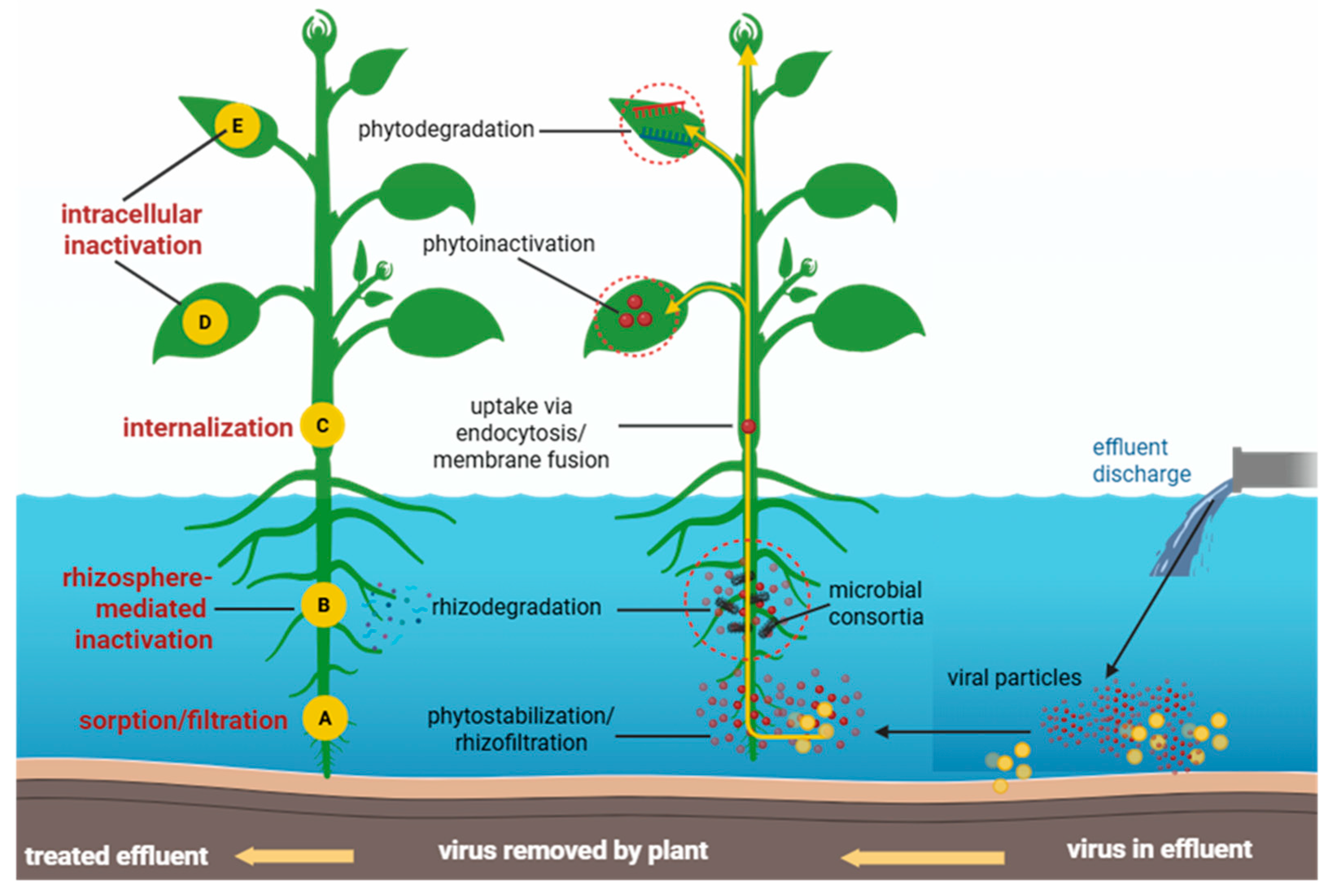
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



