Submitted:
15 November 2025
Posted:
18 November 2025
You are already at the latest version
Abstract
Insect pollination, a critical ecological process, pre-dates the emergence of angiosperms by nearly 300 million years, with fossil evidence indicating pollination interactions between insects and non-angiosperm seed plants during the Late Paleozoic. This re-view examines the symbiotic relationships between insects and gymnosperms in pre-angiosperm ecosystems, highlighting the complexity of these interactions. Fossil records suggest that the mutualistic relationships between insects and gymnosperms, which facilitated plant reproduction, were as intricate and diverse as the modern in-teractions between angiosperms and their pollinators, particularly bees. These early pollination systems likely involved specialized behaviors and plant adaptations, re-flecting a sophisticated evolutionary dynamic long before the advent of flowering plants.
The Anthropocene presents a dichotomy: while climate change and anthropogenic pressures threaten insect biodiversity and risk disrupting angiosperm reproduction, such upheaval may simultaneously generate opportunities for novel plant–insect in-teractions as ecological niches are vacated. Understanding the deep evolutionary his-tory of pollination offers critical insight into the mechanisms underlying the resilience and adaptability of these mutualisms. The evolutionary trajectory of bees—originating from predatory wasps, diversifying alongside angiosperms, and reorganizing after mass extinctions—exemplifies this dynamic, demonstrating how pollination networks persist and reorganize under environmental stress and underscoring the enduring health, resilience, and adaptability of these essential ecological systems.
Keywords:
pollination evolution
; pre-angiosperm pollination
; insect-gymnosperm pollination
; insect-angiosperm mutualism
; pollinator resilience
; bee evolution
; Anthropocene
; plant–pollinator networks
; pollinator health
1. Introduction
One of the main prejudices still rooted in the understanding of insect pollination is the belief that it began with the appearance and expansion of angiosperms in the late Lower Cretaceous. This process has long been attributed to a co-evolutionary relationship between flowering plants and pollinating insects, which was thought to have dramatically accelerated their biodiversity [1]. However, this view overlooks the extensive history of insect–plant mutual interactions that preceded angiosperms, with insect pollination predating the emergence of flowering plants (angiosperms) by nearly 200 million years.
Although the origin of insects is dated to the Early Ordovician [2], the first fossil evidence of insect taxa appears in the Early Devonian, somewhat later than the colonization of land by vascular plants. Even at this stage, clues suggest that spore consumption—i.e., feeding on the reproductive cells of early plants and fungi—was among the earliest feeding strategies of ancient hexapods [3]. Throughout their long evolutionary history, insects and plants have undergone multiple radiations, developing both mutualistic and antagonistic strategies. Interactions with plants increasingly involved reproductive organs, which insects exploited for valuable nutrients, inadvertently favoring plant outcrossing. Fossil evidence demonstrates such relationships as far back as the Upper Carboniferous, when early gymnosperms dominated terrestrial ecosystems alongside cryptogams [3,4,5,6].
These interactions further evolved during the Mesozoic Era, often referred to as the “Age of Gymnosperms.” During this time, gymnosperms exhibited remarkable diversity—including shrubs, lianas, mangroves, succulents, fast-growing herbaceous plants, and palm- or dicot-like forms [7,8,9,10,11]. Insects and gymnosperms formed complex associations that were ecologically comparable to modern pollination systems, though involving now-extinct plant lineages and insect groups that today play little or no role as pollinators. These fossil interactions reveal a sophisticated evolutionary prelude to modern angiosperm pollination [9,12].
From the mid-Cretaceous onward (around 120 million years ago), gymnosperms faced increasing competition from emerging angiosperms, which progressively displaced them from many ecosystems [13]. In the modern era, gymnosperms persist as a “relictual” group, with several lineages occupying specialized ecological niches such as boreal forests (dominated by conifers) or tropical highlands (e.g., Cycadales). Figure 1 outlines the evolutionary history of plants and insects with reference to entomogamy in plants and anthophily in insects, both of which appeared independentlyacross multiple lineages over time.
The long-term history of insect pollination has been marked by major ecological disruptions and evolutionary turnovers. The Permian–Triassic and Cretaceous–Paleogene mass extinction events, rather than collapsing pollination systems, triggered extensive restructuring of insect–plant associations. These transitions included the replacement of declining Paleozoic paleopteran pollinators by neopterans—a process already underway before the Permian–Triassic crisis—and the reorganization of pollination networks during the Albian–Aptian angiosperm radiation. During this latter phase, specialized Mesozoic holometabolous pollinators declined in prominence as modern groups diversified, and some pollinators shifted from gymnosperms to angiosperms roughly 60 million years before the Cretaceous–Paleogene extinction [14,15].
Despite this evidence, the notion that complex mutualistic interactions between entomophilous plants and specialized pollinating insects predate the emergence of flowering plants remains underappreciated. This limited awareness reinforces the misconception that the ongoing ecological crisis of the Anthropocene will inevitably lead to the collapse of pollination—a mechanism that has sustained biodiversity for more than 300 million years and persisted through multiple episodes of profound environmental change.
Bees represent a paradigmatic example of such evolutionary turnover and adaptation. Originating from carnivorous, wasp-like ancestors in the Early Cretaceous, they underwent a profound ecological transition to pollen and nectar feeding. They subsequently diversified alongside flowering plants, reshaping pollination networks and establishing many of the interactions that underpin modern ecosystems [1,16,17]. Their trajectory highlights how pollinator lineages can rapidly radiate in response to plant innovations, reinforcing the long-term adaptability of plant–insect relationships [15,18].
Accordingly, this review provides a framework for understanding the evolutionary history of insect pollination, focusing on mutual relationships in pre-angiosperm ecosystems and the key steps leading to modern plant–pollinator systems. The angiosperm radiation marked a critical turning point, driving the diversification of pollinator lineages and promoting the coevolution of traits that sustain modern interactions—bees standing as the most prominent and emblematic example of this evolutionary partnership. Finally, we emphasize how this deep-time history demonstrates the inherent resilience of pollination systems, offering valuable insights into how they may respond to the unprecedented challenges of the Anthropocene.
2. Fossil Evidence of Host-Plant and Insect Interactions and the Evolutionary Phases of Insect Pollination
Past interactions between host plants and insects are well represented in the fossil record, as evidenced by various feeding modes on plant tissues [19]. Fossil evidence of entomophily reveals several factors comparable to those observed in modern pollination systems, such as the interplay between insect structures—especially mouthparts, but also other body parts like pollen baskets on the legs—adapted for collecting pollen and/or nectar, and the corresponding plant reproductive structures. Additional indicators include the presence of attractive floral features (e.g., nectar glands, showy organs), the morphology of pollen grains, and the occurrence of pollen associated with fossilized insect bodies or preserved within their coprolites [3,6,7,9,18,20,21,22,23,24,25,26,27]. The habit of pollinator in a fossil insect could be also supposed if it belongs to a crown group of monophyletic lineages of extant taxa known as pollinators (e.g. Corbiculata bees [28,29]. Finally, palaeoecological evidence of environmental context of the possible plant-insect association is important for revealing feeding types and pollination modes that may be extinct or have survived to the present [6,20,30]. Actually, there is general acceptance that the application of these criteria in fossil records show interactions between seed plants and insect pollinators preluding early angiosperm pollination since more than 100 million years before flower plants radiation, with associations ecologically similar to modern ones in complexity, especially during the Mesozoic [3,7,9,12,15,24,31]. An example of such associations can still be seen today in the obligate mutualistic relationship between Cycadales, the most ancient seed-plants living today, and their pollinating insects, certainly as complex as other species-specific relationships existing in various angiosperms [32,33].
Fossils tell a 420 m.y. long story of vascular-plant hosts, their insect herbivores, and associated functional feeding groups, including palynivory and nectarivory, therefore involved also in plant fecundation. A long story which can be sorted spatiotemporally into four major herbivore expansions [19], discussed in detail in the subsequent paragraphs and summarized in Figure 2.
- 1. A Late Silurian to Late Devonian phase (about 60 million years) characterized by herbivores arthropods, including apterygote hexapods (Enthognathate and perhaps early Ectognathate) , feeding on several clades of primitive vascular-plant hosts.
- 2. A Late Mississippian to end-Permian phase (85 million years) involving principally apterygotes, palaeopterans, non-holometabolan and (later) basal holometabolan neopterans, feeding on pteridophyte, basal and more advanced gymnosperm plant hosts.
- 3. A Middle Triassic to Middle Cretaceous phase (ca. 130 million years) dominated by polyneopterans, paraneopterans and holometabolan, feeding mostly on gymnosperm plant hosts.
- 4. A mid-Early Cretaceous to Recent phase (115 million years) featuring modern hemimetabolous and holometabolous, feeding principally on angiosperm plant hosts. This phase also witnessed the emergence of bees, establishing one of the most important modern pollinator lineages.
While the underlying drivers of these four major associations are still debated, they likely reflect broad paleoclimatic and atmospheric dynamics, including greenhouse–icehouse cycles and fluctuations in O2 and CO2 concentrations. Notably, although the specific plant and arthropod taxa involved have changed through time, the fundamental feeding strategies have remained remarkably conserved across much of this evolutionary continuum.
2.1. Early Mandibles on Early Spores: Silurian-Devonian First Evidence of Palynivory
The first radiation of primordial vascular plants on land during the Silurian-Devonian period, alongside nonvascular plants and prototaxites fungi, led to early associations with ancient terrestrial arthropods, whose presence is documented in fossil deposits from that era [3,34,35,36]. Late Silurian and Early Devonian coprolites, i.e. fossilized faecal pellets, provide evidence of consumption by herbivores arthropods of various organic materials, including spores [36,37]. Faecal pellets were likely produced by different groups of arthropods, potentially including early Entognathate hexapods such as the collembola species Rhyniella praecursor Hirst & Maulik, 1926, one of the oldest known hexapods dating back to the Early Devonian period, and possibly basal Ectognathate wingless insects [3,9]. According to Labandeira [3,5,7,19] and Labandeira et al. [9], these Late Silurian-Early Devonian interactions represent the earliest phase in the development of palynivory (i.e., the consumption of spores and pollen) and related feeding strategies, though at this stage, only spore consumption occurred. At that time, wind and water were the primary vectors for the dispersal of undifferentiated spores from these early tracheophytes, similar to the mechanisms used by extant ferns and horsetails, with no evidence of animal involvement in their reproduction or dispersal [13].
2.2. Seed Plants and the Second Phase of Plants/Insect Associations: Late Paleozoic Pollination
The Late Paleozoic marks the second major phase of insect exploitation of plant reproductive structures and likely the origin of plant–pollinator interactions, laying the groundwork for the complexity developed in the Mesozoic [7,9].
Critical innovations began in the Middle Devonian. Some Progymnospermopsida already with gymnosperm-like anatomy but still reproducing with undifferentiated spores, evolved distinct male microspores and female megaspores, a crucial step toward the specialized heterospory of seed plants [11,38,39,40]. Early gymnosperms advanced further with the evolution of the ovule (Figure 3), in which a retained megaspore was enclosed by integuments, leaving a micropylar opening through which a nucellar exudate secreted a pollination drop that captured airborne pollen [8,9,41]. Fossil evidence of such a mechanism is well documented in a Late Carboniferous Callistophytales, where a prepollen-filled exudate was preserved protruding from the micropyle, demonstrating a pollination-drop system likely ancestral in seed plants—first serving wind pollination and later co-opted for insect mediation [9,25,42]. Gymnosperms diversified during the Carboniferous, with early seed plants such as Medullosales and Callistophytales together with more advanced lineages like Cordaitales, radiating into trees, shrubs, and climbers [11,43,44]. While early spermatophytes were probably wind-pollinated, evidence from Upper Carboniferous medullosaceans suggests the earliest insect involvement: their large, heavy prepollen, enclosed in structures with glandular trichomes and fleshy tissues (Figure 3), likely offered nutritive rewards to large arthropod pollinators such as palaeodictyopterans [9].
In the Permian, increasingly arid climates favored xeromorphic gymnosperms, and probable insect pollination is indicated in groups like Peltaspermales and Glossopteridales, the latter bearing flower-like female organs and producing abundant pollen and seeds suggestive of entomophilous adaptations [43,45,46].
These botanical shifts coincided with the first massive radiation of pterygote insects during the Carboniferous and Permian, likely driven by plant evolution [3,7,47,48].
Archaic palaeopterans were the first herbivore insects which developed complex modified mouthparts (Figure 4) and most likely played a crucial role in the reproduction of Carboniferous seed plants [7,9,14,31,47,49], together with early neopterans, represented by Polyneoptera and stem Paraneoptera.
The Permian saw the decline of palaeopterans [50,51] and the rise of neopterans such as early bugs, thrips, and early holometabolans (beetles, lacewings and stem mecopteroids), while Orthoptera and Grylloblattodea diversified among Polyneoptera [3,44,52]. These groups also evolved increasingly specialized mouthparts from basic chewing mouthparts (Figure 5): Acercaria developed piercing–sucking styles, and holometabolans produced the earliest siphonate, non-piercing proboscises, establishing the major mouthpart classes that persist today [3,52,53].
Exceptional insight comes from the Early Permian Chekarda Lagerstätte in Russia, which preserves over 290 insect species across 25 orders alongside diverse gymnosperms [7,54,55]. Fossilized gut contents show monospecific or mixed accumulations of pollen from Peltaspermales, Glossopteridales, and conifers, providing clear evidence of specialized palynophagy and intense insect–plant interactions [5,25]. Among the most remarkable insects is Marimerobius splendens (Protomeropidae), the earliest holometabolan with a siphon-like proboscis adapted for fluid feeding, probably on pollination drops of Peltaspermales (Figure 6) [24,25].
Protomeropidae, early mecopteroids perhaps stem-Amphiesmenoptera (superorder which includes caddisflies, moths and butterflies), represent the first documented radiation of non-piercing long-proboscid nectar feeders, precursors to later specialised Mesozoic guilds, and likely went extinct with the climatic upheavals of Pangea [24,56].
By the Late Permian, insect faunas remained diverse, and analyses indicate no catastrophic family-level extinction at the Permian–Triassic boundary. Instead, the Early Triassic retained about two-thirds of Late Permian families, with diversity loss attributed more to slowed diversification than to wholesale extinction [51,57]. Even so, the transition marked a profound restructuring of insect assemblages, as dominant Paleozoic groups waned and a more modern, “post-Paleozoic” fauna began to take shape [50].
2.3. The Mesozoic Third Phase: Advanced Gymnosperms and Pollinator Guilds
The Mesozoic represents a fundamental restructuring of plant-insect relationships, driven by the Permian-Triassic extinction. It was characterized not by simple recovery, but by the rise of advanced gymnosperms and the evolution of sophisticated, often specialized, pollination mutualisms with insects—a complex ecological world of pollinators that flourished for over 100 million years before the dominance of angiosperms.
This evolutionary phase is defined by three major developments, described in the subsequent paragraphs: (1) the P-T extinction as a catalyst wiping out Paleozoic ecosystems and creating the ecological vacuum and evolutionary opportunity that allowed for a new phase to begin; (2) the rise of gymnosperms as advanced plants with complex reproductive structures and rewards (fleshy tissues, sugary pollination drops) to attract insects; and consequently (3) a pre-angiosperm pollination revolution, i.e. a spectacular radiation of pollinator guilds that engaged in specialized, often obligate, relationships with gymnosperms.
2.3.1. The Permian-Triassic Crisis and Its Aftermath
The Permian–Triassic (P–T) crisis, the most severe extinction in Earth’s history, eliminated ~95% of species and about half of marine and terrestrial families [58,59]. On land, forests collapsed, producing a global “coal gap” lasting nearly 10 million years, with palynological assemblages dominated by pioneer pteridophytes [60,61,62,63,64]. Some authors, however, argue that floral losses may be overstated due to taphonomic biases and localized refugia [65,66,67].
Insects show a similar pattern: diversity dips in Early Triassic deposits may reflect faunal replacement or preservational bias rather than true extinction [12,51,68,69]. Nonetheless, the P–T transition marks a major evolutionary turnover, with the extinction of Paleozoic orders such as Palaeodictyopteroidea and the rise of holometabolous lineages, including the appearance of new orders, such as Diptera, Hymenoptera, and Lepidoptera [1,5,48,70]. The Triassic records a progressive recovery, from Early Triassic faunas dominated by survivors from end of Permian to Late Triassic assemblages comparable to those of the Early Jurassic [50,71]. Although direct evidence such as pollen-filled gut contents is absent, indirect data indicate the establishment of new insect–gymnosperm pollination mutualisms, which radiated in the Early Jurassic and underpin the diversification of pollination systems still evident today [5,6,69].
2.3.2. Floristic Recovery and the Rise of Mesozoic Gymnosperms
Starting in the early Middle Triassic, global vegetation began to recover [64]. This recovery was not a simple return to the Paleozoic status quo but a fundamental restructuring of plant communities. Early gymnosperms, such as Lyginopteridales, Medullosales, and Callistophytales, were already extinct in Early Permian. Others, like Cordaitales, Glossopteridales, Peltaspermales and Lebachiales, did not cross or only briefly survived the P-T boundary [11,72,73].
In their place, a new flora emerged. Late Paleozoic survivors like Cycadales, Ginkgoales, Pinales, and Gnetales became fundamental components of Mesozoic ecosystems, alongside groups that originated in the Late Permian, such as Bennettitales and Czekanowskiales [11,74]. They were joined by new groups that radiated during the Triassic and Early Jurassic, including Voltziales, cupressoid and Cheirolepidiaceae conifers, Pentoxylales, and Caytoniales [11,72].
This Mesozoic flora, in terms of classes, represented the peak historical diversity of insect-pollinated seed plant lineages. These included Cycadopsida, Pinopsida, Bennettitopsida, Gnetopsida, possibly Ginkgoopsida (if Czekanowskiales are included) and Angiospermopsida, compared to only three extant classes (Cycadopsida, Gnetopsida, and Angiospermopsida) with major insect-pollinated members (Figure 1). This vast diversity of gymnosperms began to decline in the mid-Cretaceous with the rise of angiosperms. Today, only Cycadales, Pinales (though excluding many Mesozoic families like Cheirolepidiaceae), Ginkgoales (a single species), and Gnetales remain as survivors [6,9,11,72].
Pollination systems of mid-Mesozoic gymnosperms were diverse but can be broadly grouped into two categories based on the reward offered and the corresponding insect pollinators: (a) the pollen and tissue reward systems; and (b) the pollination drop and nectar reward systems.
(a) The pollen and tissue reward systems primarily included plants with compact cones that produced abundant pollen and offered fleshy tissues as a reward. These structures were typically visited by small, mandibulate (e.g., beetles) or with piercing-sucking mouthparts (e.g., thrips), insects that lived in close association with the reproductive organs, often as larvae feeding within the cone tissues [6,9,75,76].
Evidence of this association, in the form of tunneling damage and coprolites in strobili likely caused by beetle larvae, dates back to the Late Triassic and is known throughout the Mesozoic in plants like cycads, Bennettitales, and Pentoxylales [23,32,76,77]. Compelling evidence comes from a well-preserved nitidulid beetle larva discovered inside the fleshy fructifications of a Cretaceous Pentoxylales, confirming beetle consumption of seed plant reproductive structures prior to angiosperm dominance [78].
Cycadales are the most ancient seed plants living today, although their modern diversity is the result of a recent radiation in the Miocene-Pliocene (12–5 Ma) or even later ([79,80,81,82]. Actually, a mutualism involves beetles (mainly weevils and cucujids) that consume pollen and cone tissues while acting as pollen vectors, involving highly specific and complex interactions [32,79,80]. Coleoptera and Cycadales have both a very long evolutionary history [44] and their mutualist association is thought as an Early Mesozoic heritage [7,9,76,81,82]. However, although there is evidence of associations between cycads and beetles dating back to Mesozoic [23,44,83], cycad-specific lineages of modern weevils – currently the most important and diverse host-specific pollinators of extant cycads – are relatively young, and likely derived from a shifting from angiosperm hosts to these plants [33,84]. Another modern guild involves primitive thrips (Aeolothripidae) restricted to pollinating the Australian genus Macrozamia [85,86,87]. While basal Thysanoptera likely played a broader role in Mesozoic gymnosperm pollination [6,81,86,87,88], the specific Macrozamia–thrips mutualism is also relatively young, probably less than 10 million years old [33,89].
Bennettitales (Figure 7) provide another quintessential example of this pollination mode. They produced highly complex cones, often with abundant bracts and, uniquely among gymnosperms, bisexual reproductive structures in several species [11,72]. Despite superficial similarity, they were not closely related to cycads [72,90]. Fossil evidence from both families (Cycadeoideaceae and Williamsoniaceae) shows tunneling damage at the interface of ovulate and pollen organs, likely caused by beetle larvae [9,75,91]. Attractive structures like resin bodies together with robust woody tissues may have been adaptations to lure Coleoptera while limiting destructive feeding [92]. Cycadeoideaceae cones were hermaphroditic and remained closed at maturity, suggesting self-pollination, potentially aided by boring beetles [9,75,91,93]. Williamsonaceae bore open, flower-like structures that may have secreted sugary substances or volatiles to attract pollinators like beetles and true flies [94,95]. Features like long, arching bracts may have restricted access to ovules to insects with long proboscises [13,24]. Decoy mechanisms common in modern cycads (thermogenesis, volatiles) were likely present in Bennettitales and other Mesozoic gymnosperms [32,33,96] and likely mimicked, in Early Cretaceous, by certain basic angiosperms with large, showy flowers, similarly pollinated [6,93,97,98,99].
(b) The pollination drop and nectar reward systems involved a modification of the basal pollination drop, introducing higher levels of nutritional substances to produce an energy-rich reward for highly mobile, fluid-feeding insects [6,24,41].
Extant Gnetales are the only group that has preserved this mechanism (Figure 8) secreting sugary drops (including on sterile ovules in male strobili) with nutritional levels comparable to angiosperm nectar, which lure visitors and ensure pollen collection; many Mesozoic lineages likely used similar tactics [9,24,41,100,101,102,103,104]. Unlike pollen/tissue-reward plants, this second category of gymnosperms often have loosely arranged unisexual organs, with notable exceptions such as Cheirolepidiaceae and other presumed insect-pollinated Mesozoic conifers bearing compact cones; funneled cone scales and abundant Classopollis pollen—frequently associated with insect fossils—strongly indicate entomophily within a largely wind-pollinated clade [6,9,72,105,106].
2.3.3. Pre-Angiosperm Complex Mutualist Balances
This syndrome chiefly engaged pollinators with non-piercing haustellate mouthparts that evolved repeatedly in Holometabola [6,24,107,108,109,110,111]. Functionally, proboscides derive from maxillary/labial modifications and range from short, mobile sponges for superficial fluids (e.g., fly labellum) to long, tubular siphons for concealed rewards (e.g., butterflies, several bees, bombyliids flies), with associated flight and sensory adaptations [1,3,6,24,112,113,114,115,116,117,118]. A classic “Darwin’s race”—progressive concealment of rewards vs. proboscis elongation—promoted pollen contact [24,119,120].
Fossil records recognize two major waves of Mesozoic long-proboscis nectarivores: a mid-Mesozoic phase (Middle Triassic–Early Cretaceous) dominated by specialized Mecoptera, Neuroptera, and Diptera, and a mid-Early Cretaceous–Recent phase coincident with angiosperm radiation which led to the pollinator guilds that we know today; Early Cretaceous overlap implies early interactions with flowering plants [6,22].
Before angiosperms, mid-Mesozoic fossils already show long-proboscid nectarivores matched to tubular gymnosperm organs and insect-type pollen, implying complex insect pollination predated flowering-plant dominance [6,7,9,20,22,23,24,30,88,106,121,122]. Cheirolepidiaceae exemplify this: funneled female scales guided small or long-proboscid insects to nectary-like secretions (Figure 9), and their Classopollis pollen is ubiquitous in Jurassic–Cretaceous deposits, often found in association with herbivorous insect fossils [6,9,72,105,106]. Extant Gnetales attract diverse short-proboscid or non-proboscid visitors (flies, midges, wasps, bees, beetles, thrips); Mesozoic gnetaleans likely ranged from extant-like forms to species with exceptionally long micropyles and bracteate/hairy tufts—potential adaptations to long, thin proboscides [6,10,101,102,104].
From the Upper Triassic to Upper Cretaceous, non-angiosperm mutualisms were in part already represented by groups that are now familiar as anthophilous (Tysanoptera, Coleoptera, Diptera). However, on the other side, they were dominated by groups that today have no or little importance as pollinators. Mecoptera (scorpionflies) and Neuroptera (lacewings) were much more diverse in the Mesozoic, developing very successful nectarivore lineages with long proboscis, some of which became popular for their incredible convergence with current butterflies (Figure 10) [6,9,20,24,30,123,124,125,126].
Large mandibulate Polyneoptera formed a now-vanished palynivory guild; Late Jurassic gut contents show Ensifera consuming Cheirolepidiaceae pollen, and gymnosperm pollen appears in some Phasmatodea and Embioptera, though regularity is uncertain [6,9,18,106,127,128,129]. Cretaceous ambers reveal anthophilous blattodeans with gymnosperm or early angiosperm pollen and floral parts [130]. Actually, Orthoptera and Blattodea are minor pollinators, whereas Phasmatodea and Embioptera are not known to consume pollen [6,131,132,133]. Within Acercaria, thrips and the archaic Permopsocida (with a mouth-cone intermediate between chewing and piercing–sucking) fed on pollen/nectar; Permopsocida vanished by mid-Cretaceous, likely displaced by angiosperm-associated pollinators [2,134,135,136]. An enigmatic order, Tarachoptera (Amphiesmenoptera), known from Burmese amber only also failed to cross the Cretaceous [122,137,138].
Hymenoptera and Lepidoptera were less diversified and marginal until the mid-Cretaceous [6,9,22,24]. The oldest Hymenoptera are Middle-Triassic xyelid sawflies and the earliest unambiguous Lepidoptera are Early-Jurassic micropterigid-like moths with mandibulate adults and primitive scales [1,68,109,139,140,141,142,143,144]. Through the Mesozoic, Hymenoptera diversified modestly until the mid-Cretaceous appearance of most modern groups, including the first bees [1,13,16]; apoditrysian Lepidoptera are absent before the Early Eocene, though basal Glossata occur earlier [24,145].
Amid these shifts, Diptera remained central from the Triassic onward and today are second only to bees in pollination importance; both Nematocera and Brachycera include long-proboscid anthophiles with flight/sensory traits suited to deep corollas [1,9,24,113,118,136,146,147,148,149,150,151,152,153,154]. The findings of Upper Jurassic flies with modern-like adaptations to anthophily, were initially interpreted as evidence of the existence of angiosperms in that time [155]. However, since no remains of angiosperms in deposits older than Early Cretaceous have never been found, and Classopollis-type pollen were often found attached to the hairs of their bodies, it is more logic to deduce that nectarivore flies were already important pollinators in a world still devoid of flowering plants [6,7,9,118,156,157,158,159].
Finally, Mesozoic sees the great radiation of beetles; larval feeding on gymnosperm sporophylls is known since Middle Triassic and persists in modern cycad mutualisms. Cretaceous ambers record a surge of plant-associated polyphagans, newly emerged groups related to the new flora, extinctions of gymnosperm-tied pollinators that failed to shift hosts, and at least one gymnosperm-angiosperm transition documented [22,23,26,44,70,76,98,121,160,161,162,163,164,165,166,167].
2.4. The Cretaceous Terrestrial Revolution: Angiosperm Radiation and the Evolution of Bees
Angiosperms (flowering plants) currently represent approximately 90% of all land plants in terms of species diversity and biomass [168]. They dominate nearly all terrestrial ecosystems and constitute a fundamental component of the modern food chain for numerous organisms, including humans. Their evolutionary success is often attributed to innovations in reproductive strategy compared to gymnosperms, most notably the enclosure of seeds within fruits that develop from the ovary after fertilization [15,90,169,170].
The fossil record provides robust evidence of an intense diversification and radiation of angiosperms during the late-Early to early-Late Cretaceous, beginning with the first unambiguous tricolpate pollens from the Barremian–Aptian (~121 Ma) and floral assemblages from the Aptian (~115 Ma), followed by an explosive increase in diversity during the mid- and late-Cretaceous (Aptian–Turonian, ~115–90 Ma) [15,158,171,172]. Despite this evidence, the timing and mechanisms underlying the origin and early radiation of angiosperms remain a subject of vigorous debate—the “abominable mystery” of Darwin (1879)—owing to conflicting interpretations of macrofossil versus microfossil data and discrepancies between paleontological and molecular datasets [169,173,174]. Molecular clock estimates often suggest pre-Cretaceous origins, ranging from the Late Carboniferous to the Middle–Late Jurassic, implying a long cryptic evolutionary history unrepresented in the fossil record [15,172,173,175]. However, such estimates may be inflated if they fail to account for dramatic accelerations in diversification rates during the angiosperm radiation [172].
What is clear is that angiosperms rose to ecological dominance during the Late Cretaceous, catalyzing the Cretaceous Terrestrial Revolution (KTR), a profound restructuring of trophic networks that set the stage for modern terrestrial ecosystems [16,176,177]. A hallmark of this ecological shift was the co-diversification of flowering plants and their pollinators, most prominently bees. Although bees are today the most diverse lineage of pollinivorous insects and a vital component of terrestrial biomes worldwide [17,178,179,180,181], their evolutionary origins trace back to the mid-Cretaceous in close association with angiosperm expansion [1,5,13,15,16].
Bees are thought to have originated from wasp-like ancestors in western Gondwana (modern Africa and South America) in arid habitats similar to present-day biodiversity hotspots for these insects [17,178,181]. This evolutionary transition marked a profound ecological innovation, shifting from carnivorous brood provisioning with insect prey to a strictly herbivorous, anthophilous diet of pollen and nectar, a change tightly coupled to the concurrent rise and ecological expansion of angiosperms during the Cretaceous (~130–66 Ma) [1,13,182,183]. A central morphological adaptation was the evolution of branched, plumose body hairs. Unlike the simple setae of their wasp ancestors, these hairs dramatically increased the surface area for pollen adherence and could attract airborne particles through electrostatic forces, making bees unparalleled pollen vectors [184,185,186]. Over time, these hairs were further specialized into compact pollen-transporting structures (scopae) on the hind legs or abdomen, culminating in the evolution of corbiculae (pollen baskets) in the corbiculate bee clade [187].
The fossil record, though sparse, corroborates this early diversification. The controversial Melittosphex burmensis from Burmese amber (~100 Ma) exhibits a mosaic of bee-like traits (e.g., branched hairs) and wasp-like features, leaving its status as a stem bee or close apoid relative uncertain [188,189]. This, alongside other mid-Cretaceous transitional fossils and possible halictid nests, points to ongoing evolution. The oldest uncontested bee fossil, Cretotrigona prisca from New Jersey amber (~65 Ma; Figure 11), already belongs to the crown-group Meliponini (Apidae), implying a much earlier, hidden diversification [190,191].
Phylogenetic and molecular data support an Early Cretaceous origin of Apoidea (~120 Ma), a process likely facilitated by the fragmentation of Gondwana beginning ~175 Ma, which promoted geographic isolation and early lineage divergence [16,17]. By the end of the Cretaceous, all extant bee families were present.
This early diversification was further structured by major geological and climatic events. The continued separation of landmasses promoted endemism, while transient Tertiary land bridges facilitated dispersal into the Northern Hemisphere, where bees adapted to temperate floras [16,17]. Mass extinctions also played a formative role; the K–Pg event (~66 Ma) and Eocene-Oligocene transition eliminated numerous taxa but created ecological opportunities for surviving clades to undergo adaptive radiations in tandem with their eudicot hosts [190,192]. Morphologically, early bees were likely short-tongued, a condition retained in basal families (Andrenidae, Colletidae, Halictidae). The increasing floral complexity of angiosperms later drove the evolution of long-tongued lineages (e.g., Apidae, Megachilidae), firmly anchoring bees in their mutualistic role and reshaping plant reproductive strategies [16]. Collectively, these geological, climatic, and biological factors forged bees into one of the most specialized and ecologically significant pollinator groups, whose coupled diversification with angiosperms during and after the Cretaceous Terrestrial Revolution fundamentally reshaped terrestrial ecosystems.
3. The Resilience of Plant–Pollinator Interactions: Lessons from Deep Time for the Anthropocene
Deep-time evidence suggests that pollination systems have demonstrated resilience through past environmental crises. Even profound environmental crises, such as the Cretaceous–Paleogene mass extinction, did not cause the collapse of entomophilous pollination but instead reshaped ecological networks. Generalist species often persisted and temporarily dominated, while specialists were disproportionately vulnerable [30,190,192,193,194]. This dynamic of disruption, survival, and reassembly illustrates the adaptive flexibility of plant–pollinator interactions—a theme that continues to resonate in the Anthropocene. Today, however, pollination systems face unprecedented pressures from climate change and human-driven environmental change. Urbanization, deforestation, agricultural intensification, and pollution are driving alarming declines in insect biodiversity and pollinator populations, with cascading consequences for ecosystem services and plant reproduction [195,196,197,198,199,200,201,202,203,204,205,206]. These stressors disrupt pollination dynamics and increase the risk of decline or extinction in plants dependent on specific pollinators. Yet, as in the deep past, such disruptions may also create opportunities for new associations. Empirical evidence demonstrates the capacity of networks to reorganize: in a century-scale study of a temperate forest understory community, 76% of original plant–pollinator interactions were lost and nearly half of historical bee species were extirpated, but extraordinary, novel interactions emerged, indicating a surprising degree of resilience and adaptability [207] (Burkle et al., 2013).
Adaptive responses are evident on both sides of the interaction. Among pollinators, foraging plasticity is a critical buffer against environmental stress. Generalist pollinators such as Bombus terrestris and syrphid flies expand their floral ranges when preferred resources decline, maintaining ecosystem function despite habitat fragmentation and phenological mismatches [195,196,197,208,209]. Urban studies similarly show bees incorporating non-native plants into their diets, allowing persistence in modified habitats [199,210]. However, specialists—such as oligolectic bees dependent on single plant taxa—remain highly vulnerable, heightening the risk of local extinctions and potential network collapse [196,211]. Plants, in turn, exhibit their own forms of plasticity, altering floral morphology, nectar production, scent, or phenology to attract alternative pollinators under stress [198,202,207,212,213].
Taken together, the evolutionary history of angiosperm–insect interactions illustrates both the creative power and the resilience of pollination systems. The Cretaceous angiosperm radiation was a critical turning point, driving the diversification of pollinators and promoting the coevolutionary traits that sustain modern ecosystems, with bees as their most prominent and emblematic representatives. At the same time, the persistence and reorganization of pollination through deep-time crises underscores the resilience and adaptive capacity of these systems. This long-term perspective offers crucial insights for the Anthropocene: while pollination networks are dynamic and capable of reassembly, their survival in the face of current unprecedented pressures will depend on conservation strategies that safeguard both pollinator diversity and the ecological conditions that enable such resilience.
4. Conclusions
The history of pollination in the Paleozoic and Mesozoic reveals a dynamic and evolving set of interactions between plants and insects. From their earliest co-evolutionary stages, both groups developed adaptations that enabled intimate ecological relationships, laying the foundations for the complex pollination systems of today. These ancient syndromes, though less conspicuous than modern ones, were no less intricate or fundamental to the history of life on Earth.
Deep-time evidence shows that major ecological transitions—such as the rise of Mesophytic floras in the Permian and the Cretaceous Terrestrial Revolution with angiosperm expansion—drove more profound changes in insect diversity than the mass extinctions of the last 300 million years. These transitions opened new ecological opportunities, fueling adaptive radiations in both plants and pollinators. While this precedent suggests that current pollinator declines may also foster novel ecological arrangements, the unprecedented pace of human-driven environmental change may exceed the adaptive capacity of many species [195,196,211].
The Anthropocene thus presents both a challenge and an opportunity. Generalist pollinators and opportunistic plants may persist, but specialists risk functional extinction, creating gaps in pollination networks that may not be easily replaced [207,210]. Conservation strategies must therefore focus on safeguarding habitat diversity, genetic resources, and irreplaceable ecological roles, while research should track how networks reorganize in the face of loss.
Integrating deep-time insights with present-day challenges is essential for developing adaptive conservation strategies that foster both ecological stability and evolutionary resilience. Such an approach also offers a broader framework for understanding pollinator health and guiding sustainable management practices under accelerating environmental change.
Funding
This work received no external funding.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
We acknowledge the essential contributions of paleontologists, botanists, entomologists, and zoologists, whose research forms the basis of this review. Our aim has been to outline selected aspects and distinctive features of plant–pollinator co-evolution through deep time, without attempting to cover the full breadth of paleontological evidence. Reconstructions of fossil plants and insects are included to aid visualization of these ancient organisms and their interactions, while readers are referred to the cited literature for primary fossil data. This work also seeks to draw attention to the evolutionary significance of pre-angiosperm pollination, an aspect often overlooked yet fundamental to understanding the origins and long-term dynamics of plant–insect relationships.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Morris, S.C. D. Grimaldi & M. S. Engel 2005. Evolution of the Insects. Xv + 755 Pp. Cambridge, New York, Melbourne: Cambridge University Press. Price £45.00, US $75.00 (Hard Covers). ISBN 0 521 82149 5. Geological Magazine 2007, 144, 1035–1036. [Google Scholar] [CrossRef]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics Resolves the Timing and Pattern of Insect Evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Labandeira, C.C. The Fossil Record of Insect Mouthparts: Innovation, Functional Convergence, and Associations with Other Organisms. Zoological Monographs 2019, 567–671. [Google Scholar] [CrossRef]
- Labandeira, C.C. Chapter 2 The Historyofassociations between Plants and Animals.
- Labandeira, C. Silurian to Triassic Plant and Hexapod Clades and Their Associations: New Data, a Review, and Interpretations. Arthropod Systematics & Phylogeny 2006, 64, 53–94. [Google Scholar] [CrossRef]
- Labandeira, C.C. The Pollination of Mid Mesozoic Seed Plants and the Early History of Long-Proboscid Insects1,2,3. Annals of the Missouri Botanical Garden 2010, 97, 469–513. [Google Scholar] [CrossRef]
- Labandeira, C.C. The Paleobiology of Pollination and Its Precursors. The Paleontological Society Papers 2000, 6, 233–270. [Google Scholar] [CrossRef]
- Rothwell, G.W.; Grauvogel-Stamm, L.; Mapes, G. An Herbaceous Fossil Conifer: Gymnospermous Ruderals in the Evolution of Mesozoic Vegetation. Palaeogeography, Palaeoclimatology, Palaeoecology 2000, 156, 139–145. [Google Scholar] [CrossRef]
- Labandeira, C.C.; Kvaček, J.; Mostovski, M.B. Pollination Drops, Pollen, and Insect Pollination of Mesozoic Gymnosperms. TAXON 2007, 56, 663–695. [Google Scholar] [CrossRef]
- Krassilov, V.A. Diversity of Mesozoic Gnetophytes and the First Angiosperms. Paleontological Journal 2009, 43, 1272–1280. [Google Scholar] [CrossRef]
- McLoughlin, S. Gymnosperms. Encyclopedia of Geology 2021, 476–500. [Google Scholar] [CrossRef]
- Labandeira, C.C.; Sepkoski, J.J. Insect Diversity in the Fossil Record. Science 1993, 261, 310–315. [Google Scholar] [CrossRef]
- Friis, E.M.; Crane, P.R.; Pedersen, K.R. Early Flowers and Angiosperm Evolution, 1st ed.; Cambridge University Press, 2011; ISBN 978-0-521-59283-3. [Google Scholar]
- Schachat, S.R.; Labandeira, C.C. Are Insects Heading Toward Their First Mass Extinction? Distinguishing Turnover From Crises in Their Fossil Record. Annals of the Entomological Society of America 2020, 114, 99–118. [Google Scholar] [CrossRef]
- Peris, D.; Condamine, F.L. The Angiosperm Radiation Played a Dual Role in the Diversification of Insects and Insect Pollinators. Nature Communications 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Cardinal, S.; Danforth, B.N. Bees Diversified in the Age of Eudicots. Proceedings of the Royal Society B: Biological Sciences 2013, 280, 20122686. [Google Scholar] [CrossRef] [PubMed]
- Almeida, E.A.B.; Bossert, S.; Danforth, B.N.; Porto, D.S.; Freitas, F.V.; Davis, C.C.; Murray, E.A.; Blaimer, B.B.; Spasojevic, T.; Ströher, P.R.; et al. The Evolutionary History of Bees in Time and Space. Current Biology 2023, 33, 3409–3422.e6. [Google Scholar] [CrossRef]
- Labandeira, C. Why Did Terrestrial Insect Diversity Not Increase During the Angiosperm Radiation? Mid-Mesozoic, Plant-Associated Insect Lineages Harbor Clues. Evolutionary Biology: Genome Evolution, Speciation, Coevolution and Origin of Life 2014, 261–299. [Google Scholar] [CrossRef]
- LABANDEIRA, C. The Four Phases of Plant-Arthropod Associations in Deep Time. Geologica Acta 2006, 4, 409–438. [Google Scholar] [CrossRef]
- Ren, D.; Labandeira, C.C.; Santiago-Blay, J.A.; Rasnitsyn, A.; Shih, C.; Bashkuev, A.; Logan, M.A.V.; Hotton, C.L.; Dilcher, D. A Probable Pollination Mode Before Angiosperms: Eurasian, Long-Proboscid Scorpionflies. Science 2009, 326, 840–847. [Google Scholar] [CrossRef]
- Hu, S.; Dilcher, D.L.; Taylor, D.W. Pollen Evidence for the Pollination Biology of Early Flowering Plants. Evolution of Plant-Pollinator Relationships 2011, 165–236. [Google Scholar] [CrossRef]
- Peris, D.; Labandeira, C.C.; Peñalver, E.; Delclòs, X.; Barrón, E.; Pérez-de la Fuente, R. The Case of Darwinylus Marcosi (Insecta: Coleoptera: Oedemeridae): A Cretaceous Shift from a Gymnosperm to an Angiosperm Pollinator Mutualism. Communicative & Integrative Biology 2017, 10, e1325048. [Google Scholar] [CrossRef]
- Cai, C.; Escalona, H.E.; Li, L.; Yin, Z.; Huang, D.; Engel, M.S. Beetle Pollination of Cycads in the Mesozoic. Current Biology 2018, 28, 2806–2812.e1. [Google Scholar] [CrossRef]
- Khramov, A.V.; Bashkuev, A.S.; Lukashevich, E.D. The Fossil Record of Long-Proboscid Nectarivorous Insects. Entomological Review 2020, 100, 881–968. [Google Scholar] [CrossRef]
- Khramov, A.V.; Naugolnykh, S.V.; Węgierek, P. Possible Long-Proboscid Insect Pollinators from the Early Permian of Russia. Current Biology 2022, 32, 3815–3820.e2. [Google Scholar] [CrossRef]
- Tihelka, E.; Li, L.; Fu, Y.; Su, Y.; Huang, D.; Cai, C. Angiosperm Pollinivory in a Cretaceous Beetle. Nature Plants 2021, 7, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Peña-Kairath, C.; Delclòs, X.; Álvarez-Parra, S.; Peñalver, E.; Engel, M.S.; Ollerton, J.; Peris, D. Insect Pollination in Deep Time. Trends in Ecology & Evolution 2023, 38, 749–759. [Google Scholar] [CrossRef]
- ENGEL, M.S. A MONOGRAPH OF THE BALTIC AMBER BEES AND EVOLUTION OF THE APOIDEA (HYMENOPTERA). Bulletin of the American Museum of Natural History 2001, 259, 1–192. [Google Scholar] [CrossRef]
- Vajda, V.; McLoughlin, S.; Slater, S.M.; Gustafsson, O.; Rasmusson, A.G. The ‘Seed-Fern’ Lepidopteris Mass-Produced the Abnormal Pollen Ricciisporites during the End-Triassic Biotic Crisis. Palaeogeography, Palaeoclimatology, Palaeoecology 2023, 627, 111723. [Google Scholar] [CrossRef]
- Labandeira, C.C.; Yang, Q.; Santiago-Blay, J.A.; Hotton, C.L.; Monteiro, A.; Wang, Y.-J.; Goreva, Y.; Shih, C.; Siljeström, S.; Rose, T.R.; et al. The Evolutionary Convergence of Mid-Mesozoic Lacewings and Cenozoic Butterflies. Proceedings of the Royal Society B: Biological Sciences 2016, 283, 20152893. [Google Scholar] [CrossRef]
- GORELICK, R. Did Insect Pollination Cause Increased Seed Plant Diversity? Biological Journal of the Linnean Society 2001, 74, 407–427. [Google Scholar] [CrossRef]
- Toon, A.; Terry, L.I.; Tang, W.; Walter, G.H.; Cook, L.G. Insect Pollination of Cycads. Austral Ecology 2020, 45, 1033–1058. [Google Scholar] [CrossRef]
- Hsiao, Y.; Oberprieler, R.G.; Zwick, A.; Zhou, Y.-L.; Ślipiński, A. Museomics Unveil Systematics, Diversity and Evolution of Australian Cycad-Pollinating Weevils. Proceedings of the Royal Society B: Biological Sciences 2023, 290. [Google Scholar] [CrossRef]
- Pirozynski, K.A.; Malloch, D.W. The Origin of Land Plants: A Matter of Mycotrophism. Biosystems 1975, 6, 153–164. [Google Scholar] [CrossRef]
- Taylor, T.N.; Osborn, J.M. The Importance of Fungi in Shaping the Paleoecosystem. Review of Palaeobotany and Palynology 1996, 90, 249–262. [Google Scholar] [CrossRef]
- Dunlop, J.A.; Garwood, R.J. Terrestrial Invertebrates in the Rhynie Chert Ecosystem. Philosophical Transactions of the Royal Society B: Biological Sciences 2017, 373, 20160493. [Google Scholar] [CrossRef]
- Edwards, D.; Selden, P.A.; Richardson, J.B.; Axe, L. Coprolites as Evidence for Plant–Animal Interaction in Siluro–Devonian Terrestrial Ecosystems. Nature 1995, 377, 329–331. [Google Scholar] [CrossRef]
- Beck, C.B. ON THE ORIGIN OF GYMNOSPERMS. TAXON 1966, 15, 337–339. [Google Scholar] [CrossRef]
- BATEMAN, R.M.; DiMICHELE, W.A. HETEROSPORY: THE MOST ITERATIVE KEY INNOVATION IN THE EVOLUTIONARY HISTORY OF THE PLANT KINGDOM. Biological Reviews 1994, 69, 345–417. [Google Scholar] [CrossRef]
- Bonacorsi, N.K.; Leslie, A.B. Sporangium Position, Branching Architecture, and the Evolution of Reproductive Morphology in Devonian Plants. International Journal of Plant Sciences 2019, 180, 493–503. [Google Scholar] [CrossRef]
- von Aderkas, P.; Prior, N.A.; Little, S.A. The Evolution of Sexual Fluids in Gymnosperms From Pollination Drops to Nectar. Frontiers in Plant Science 2018, 9. [Google Scholar] [CrossRef]
- Rothwell, G.W. Evidence for a Pollination-Drop Mechanism in Paleozoic Pteridosperms. Science 1977, 198, 1251–1252. [Google Scholar] [CrossRef] [PubMed]
- DiMichele, W.A.; Pfefferkorn, H.W.; Gastaldo, R.A. Response of Late Carboniferous and Early Permian Plant Communities to Climate Change. Annual Review of Earth and Planetary Sciences 2001, 29, 461–487. [Google Scholar] [CrossRef]
- Beutel, R.G.; Xu, C.; Jarzembowski, E.; Kundrata, R.; Boudinot, B.E.; McKenna, D.D.; Goczał, J. The Evolutionary History of Coleoptera ( Insecta ) in the Late Palaeozoic and the Mesozoic. Systematic Entomology 2024, 49, 355–388. [Google Scholar] [CrossRef]
- DOS SANTOS, T.B.; DE SOUZA PINHEIRO, E.R.; IANNUZZI, R. FIRST EVIDENCE OF SEED PREDATION BY ARTHROPODS FROM GONDWANA AND ITS EARLY PALEOZOIC HISTORY (RIO BONITO FORMATION, PARANÁ BASIN, BRAZIL). PALAIOS 2020, 35, 292–301. [Google Scholar] [CrossRef]
- McLoughlin, S.; Prevec, R. The Reproductive Biology of Glossopterid Gymnosperms—A Review. Review of Palaeobotany and Palynology 2021, 295, 104527. [Google Scholar] [CrossRef]
- Retallack, G.J.; Dilcher, D.L. Reconstructions of Selected Seed Ferns. Annals of the Missouri Botanical Garden 1988, 75, 1010. [Google Scholar] [CrossRef]
- Jarzembowski, E.A.; Ross, A.J. Insect Origination and Extinction in the Phanerozoic. Geological Society, London, Special Publications 1996, 102, 65–78. [Google Scholar] [CrossRef]
- Prokop, J.; Engel, M.S. Palaeodictyopterida. Current Biology 2019, 29, R306–R309. [Google Scholar] [CrossRef] [PubMed]
- Shcherbakov, D.E. On Permian and Triassic Insect Faunas in Relation to Biogeography and the Permian-Triassic Crisis. Paleontological Journal 2008, 42, 15–31. [Google Scholar] [CrossRef]
- Aristov, D.S.; Bashkuev, A.S.; Golubev, V.K.; Gorochov, A.V.; Karasev, E.V.; Kopylov, D.S.; Ponomarenko, A.G.; Rasnitsyn, A.P.; Rasnitsyn, D.A.; Sinitshenkova, N.D.; et al. Fossil Insects of the Middle and Upper Permian of European Russia. Paleontological Journal 2013, 47, 641–832. [Google Scholar] [CrossRef]
- Prokop, J.; Nel, A.; Engel, M.S. Diversity, Form, and Postembryonic Development of Paleozoic Insects. Annual Review of Entomology 2023, 68, 401–429. [Google Scholar] [CrossRef]
- Wootton, R.J. Palaeozoic Insects. Annual Review of Entomology 1981, 26, 319–344. [Google Scholar] [CrossRef]
- Жyжгoвa, Л.B. [and three others].; Zhuzhgova, L.V. Чeкapдa -- Mecтoнaxoждeниe Пepмcкиx Иcкoпaeмыx Haceкoмыx и Pacтeний: Moнoгpaφия Пo Гeoлoгии, Пaлeнтoэнтoмoлoгии и Пaлeoбoтaникe Чeкapды = Chekarda Is a Location of Fossil Insects and Plants from the Permian Period : Monograph on the Geology, Paleobotany and Paleoentomology of Chekard, Ed.; Издaтeльcкий цeнтp Пepмcкoгo гocyдapcтвeннoгo нaциoнaльнoгo иccлeдoвaтeльcкoгo yнивepcитeтa: Permʹ, 2015; ISBN 978-5-7944-2545-1. [Google Scholar]
- Novokshonov, V.G. New Insects (Insecta: Hypoperlida, Mischopterida, Jurinida) from the Lower Permian of the Middle Urals. Paleontological Journal. 1998, 32, 46–53. [Google Scholar]
- POSCHMANN, M.J.; NEL, A. The First Permian Scorpionfly from Germany (Insecta, Panorpida: Protomeropidae). Palaeoentomology 2021, 4. [Google Scholar] [CrossRef]
- Dmitriev, V.Yu.; Aristov, D.S.; Bashkuev, A.S.; Vasilenko, D.V.; Vřsanský, P.; Gorochov, A.V.; Lukashevitch, E.D.; Mostovski, M.B.; Ponomarenko, A.G.; Popov, Yu.A.; et al. Insect Diversity from the Carboniferous to Recent. Paleontological Journal 2018, 52, 610–619. [Google Scholar] [CrossRef]
- Erwin, D.H. The End-Permian Mass Extinction: What Really Happened and Did It Matter? Trends in Ecology & Evolution 1989, 4, 225–229. [Google Scholar] [CrossRef]
- Benton, M.J.; Twitchett, R.J. How to Kill (Almost) All Life: The End-Permian Extinction Event. Trends in Ecology & Evolution 2003, 18, 358–365. [Google Scholar] [CrossRef]
- Looy, C.V.; Brugman, W.A.; Dilcher, D.L.; Visscher, H. The Delayed Resurgence of Equatorial Forests after the Permian–Triassic Ecologic Crisis. Proceedings of the National Academy of Sciences 1999, 96, 13857–13862. [Google Scholar] [CrossRef] [PubMed]
- Galfetti, T.; Hochuli, P.A.; Brayard, A.; Bucher, H.; Weissert, H.; Vigran, J.O. Smithian-Spathian Boundary Event: Evidence for Global Climatic Change in the Wake of the End-Permian Biotic Crisis. Geology 2007, 35, 291. [Google Scholar] [CrossRef]
- McElwain, J.C.; Punyasena, S.W. Mass Extinction Events and the Plant Fossil Record. Trends in Ecology & Evolution 2007, 22, 548–557. [Google Scholar] [CrossRef]
- Vajda, V.; McLoughlin, S.; Mays, C.; Frank, T.D.; Fielding, C.R.; Tevyaw, A.; Lehsten, V.; Bocking, M.; Nicoll, R.S. End-Permian (252 Mya) Deforestation, Wildfires and Flooding—An Ancient Biotic Crisis with Lessons for the Present. Earth and Planetary Science Letters 2020, 529, 115875. [Google Scholar] [CrossRef]
- Bodnar, J.; Coturel, E.P.; Falco, J.I.; Beltrán, M. An Updated Scenario for the End-Permian Crisis and the Recovery of Triassic Land Flora in Argentina. Historical Biology 2021, 33, 3654–3672. [Google Scholar] [CrossRef]
- Peng, H.; Yang, W.; Wan, M.; Liu, J.; Liu, F. Refugium amidst Ruins: Unearthing the Lost Flora That Escaped the End-Permian Mass Extinction. Sci. Adv. 2025, 11, eads5614. [Google Scholar] [CrossRef]
- Nowak, H.; Schneebeli-Hermann, E.; Kustatscher, E. No Mass Extinction for Land Plants at the Permian–Triassic Transition. Nature Communications 2019, 10. [Google Scholar] [CrossRef]
- Nowak, H.; Vérard, C.; Kustatscher, E. Palaeophytogeographical Patterns Across the Permian–Triassic Boundary. Frontiers in Earth Science 2020, 8. [Google Scholar] [CrossRef]
- Montagna, M.; Tong, K.J.; Magoga, G.; Strada, L.; Tintori, A.; Ho, S.Y.W.; Lo, N. Recalibration of the Insect Evolutionary Time Scale Using Monte San Giorgio Fossils Suggests Survival of Key Lineages through the End-Permian Extinction. Proceedings of the Royal Society B: Biological Sciences 2019, 286, 20191854. [Google Scholar] [CrossRef]
- Montagna, M.; Magoga, G.; Stockar, R.; Magnani, F. The Contribution of the Middle Triassic Fossil Assemblage of Monte San Giorgio to Insect Evolution. Communications Biology 2024, 7. [Google Scholar] [CrossRef]
- Zhang, S.-Q.; Che, L.-H.; Li, Y.; Dan Liang; Pang, H. ; Ślipiński, A.; Zhang, P. Evolutionary History of Coleoptera Revealed by Extensive Sampling of Genes and Species. Nature Communications 2018, 9. [Google Scholar] [CrossRef]
- Shcherbakov, D.E. Insect Recovery after the Permian/Triassic Crisis.
- Taylor, T.N. Paleobotany: The Biology and Evolution of Fossil Plants; 2nd ed.; Elsevier Science & Technology: Oxford, 2009; ISBN 978-0-08-055783-0. [Google Scholar]
- Yi, Z.; Shaolin, Z.; Singh, K.J.; Yongdong, W.; Shanzhen, Z.; Saxena, A. Glossopterids Survived End-Permian Mass Extinction in North Hemisphere.
- Kustatscher, E.; Visscher, H.; Van Konijnenburg-van Cittert, J.H.A. Did the Czekanowskiales Already Exist in the Late Permian? PalZ 2019, 93, 465–477. [Google Scholar] [CrossRef]
- Crepet, W.L. INVESTIGATIONS OF NORTH AMERICAN CYCADEOIDS: POLLINATION MECHANISMS IN CYCADEOIDEA. American Journal of Botany 1972, 59, 1048–1056. [Google Scholar] [CrossRef]
- Klavins, S.D. Coprolites in a Middle Triassic Cycad Pollen Cone: Evidence for Insect Pollination in Early Cycads? Evolutionary Ecology Research 2005, 7, 479–488. [Google Scholar]
- Procheş, Ş.; Johnson, S.D. Beetle Pollination of the Fruit-scented Cones of the South African Cycad Stangeria Eriopus. American Journal of Botany 2009, 96, 1722–1730. [Google Scholar] [CrossRef]
- Nishida, H.; Hayashi, N. Cretaceous Coleopteran Larva Fed on a Female Fructification of Extinct Gymnosperm. Journal of Plant Research 1996, 109, 327–330. [Google Scholar] [CrossRef]
- Terry, I.; Walter, G.H.; Moore, C.; Roemer, R.; Hull, C. Odor-Mediated Push-Pull Pollination in Cycads. Science 2007, 318, 70–70. [Google Scholar] [CrossRef]
- Terry, I.; Tang, W.; Taylor Blake, A.S.; Donaldson, J.S.; Singh, R.; Vovides, A.P.; Cibrián Jaramillo, A. An Overview of Cycad Pollination Studies.; The New York Botanical Garden Press, 2012; pp. 352–394. 12 October.
- Schneider, D.; Wink, M.; Sporer, F.; Lounibos, P. Cycads: Their Evolution, Toxins, Herbivores and Insect Pollinators. Naturwissenschaften 2002, 89, 281–294. [Google Scholar] [CrossRef]
- Oberprieler, R.G. “Evil Weevils” – the Key to Cycad Survival and Diversification? Pp. (170–194).; Lindstrom, A.J., 2004; pp. 170–194.
- Liu, Z.; Ślipiński, A.; Lawrence, J.F.; Ren, D.; Pang, H. Palaeoboganium Gen. Nov. from the Middle Jurassic of China (Coleoptera: Cucujoidea: Boganiidae): The First Cycad Pollinators? Journal of Systematic Palaeontology 2017, 16, 351–360. [Google Scholar] [CrossRef]
- Downie, D.A.; Donaldson, J.S.; Oberprieler, R.G. Molecular Systematics and Evolution in an African Cycad-Weevil Interaction: Amorphocerini (Coleoptera: Curculionidae: Molytinae) Weevils on Encephalartos. Molecular Phylogenetics and Evolution 2008, 47, 102–116. [Google Scholar] [CrossRef]
- Mound, L. The First Thrips Species (Insecta, Thysanoptera) from Cycad Male Cones, and Its Family Level Significance. Journal of Natural History 1991, 25, 647–652. [Google Scholar] [CrossRef]
- Mound, L.A.; Terry, I. Thrips Pollination of the Central Australian Cycad, Macrozamia Macdonnellii (Cycadales). International Journal of Plant Sciences 2001, 162, 147–154. [Google Scholar] [CrossRef]
- GRIMALDI, D.; SHMAKOV, A.; FRASER, N. MESOZOIC THRIPS AND EARLY EVOLUTION OF THE ORDER THYSANOPTERA (INSECTA). Journal of Paleontology 2004, 78, 941–952. [Google Scholar] [CrossRef]
- Peñalver, E.; Labandeira, C.C.; Barrón, E.; Delclòs, X.; Nel, P.; Nel, A.; Tafforeau, P.; Soriano, C. Thrips Pollination of Mesozoic Gymnosperms. Proceedings of the National Academy of Sciences 2012, 109, 8623–8628. [Google Scholar] [CrossRef]
- Brookes, D.R.; Hereward, J.P.; Terry, L.I.; Walter, G.H. Evolutionary Dynamics of a Cycad Obligate Pollination Mutualism – Pattern and Process in Extant Macrozamia Cycads and Their Specialist Thrips Pollinators. Molecular Phylogenetics and Evolution 2015, 93, 83–93. [Google Scholar] [CrossRef]
- Crane, P.R.; Friis, E.M.; Pedersen, K.R. The Origin and Early Diversification of Angiosperms. Nature 1995, 374, 27–33. [Google Scholar] [CrossRef]
- Crepet, W.L. Investigations of North American Cycadeoids: The Reproductive Biology of Cycadeoidea. Palaeontographica Abteilung B 1974, 148, 144–169. [Google Scholar]
- Popa, M.E. Review of the Bennettitalean Genus Weltrichia. Journal of Palaeogeography 2019, 8. [Google Scholar] [CrossRef]
- Gottsberger, G. THE REPRODUCTIVE BIOLOGY OF PRIMITIVE ANGIOSPERMS. TAXON 1988, 37, 630–643. [Google Scholar] [CrossRef]
- Watson, J.; Henderson, C.M.B.; Sincock, C.A. Bennettitales of the English Wealden. Monographs of the Palaeontographical Society 1991, 145, 2–224. [Google Scholar] [CrossRef]
- Pott, C. A Revision of Wielandiella Angustifolia, a Shrub-Sized Bennettite from the Rhaetian-Hettangian of Scania, Sweden, and Jameson Land, Greenland. International Journal of Plant Sciences 2014, 175, 467–499. [Google Scholar] [CrossRef]
- Roemer, R.; Terry, I.; Chockley, C.; Jacobsen, J. Experimental Evaluation and Thermo-Physical Analysis of Thermogenesis in Male and Female Cycad Cones. Oecologia 2005, 144, 88–97. [Google Scholar] [CrossRef]
- Dieringer, G.; Cabrera R., L.; Lara, M.; Loya, L.; Reyes-Castillo, P. Beetle Pollination and Floral Thermogenicity inMagnoliaTamaulipana(Magnoliaceae). International Journal of Plant Sciences 1999, 160, 64–71. [Google Scholar] [CrossRef]
- Bernhardt, P. Convergent Evolution and Adaptive Radiation of Beetle-Pollinated Angiosperms. Plant Systematics and Evolution 2000, 222, 293–320. [Google Scholar] [CrossRef]
- SEYMOUR, R.S.; MATTHEWS, P.G.D. The Role of Thermogenesis in the Pollination Biology of the Amazon Waterlily Victoria Amazonica. Annals of Botany 2006, 98, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Kubitzki, K. Welwitschiaceae. Pteridophytes and Gymnosperms 1990, 387–391. [Google Scholar] [CrossRef]
- Kato, M.; Inoue, T. Origin of Insect Pollination. Nature 1994, 368, 195–195. [Google Scholar] [CrossRef]
- Kato, M.; Inoue, T.; Nagamitsu, T. Pollination Biology of Gnetum (Gnetaceae) in a LOWLAND MIXED DIPTEROCARP Forest in Sarawak. American Journal of Botany 1995, 82, 862–868. [Google Scholar] [CrossRef]
- Gong, Y.; Yang, M.; Vamosi, J.C.; Yang, H.; Mu, W.; Li, J.; Wan, T. Wind or Insect Pollination? Ambophily in a Subtropical Gymnosperm Gnetum Parvifolium (Gnetales). Plant Species Biology 2015, 31, 272–279. [Google Scholar] [CrossRef]
- Rydin, C.; Bolinder, K. Moonlight Pollination in the gymnospermEphedra(Gnetales). Biology Letters 2015, 11, 20140993. [Google Scholar] [CrossRef]
- Balme, B.E. Fossil in Situ Spores and Pollen Grains: An Annotated Catalogue. Review of Palaeobotany and Palynology 1995, 87, 81–323. [Google Scholar] [CrossRef]
- Krassilov, V.A.; Rasnitsyn, A.P.; Afonin, S.A. Pollen Eaters and Pollen Morphology : Co-Evolution through the Permian and Mesozoic.
- Chaudonneret, J. Les Piéces Buccales Des Insectes: Théme et Variations. Éditions hors série du Bulletin scientifique de Bourgogne, Dijon, 1990. [Google Scholar]
- Labandeira, C.C. Insect Mouthparts: Ascertaining the Paleobiology of Insect Feeding Strategies. Annual Review of Ecology and Systematics 1997, 28, 153–193. [Google Scholar] [CrossRef]
- Krenn, H.W. Feeding Mechanisms of Adult Lepidoptera: Structure, Function, and Evolution of the Mouthparts. Annual Review of Entomology 2010, 55, 307–327. [Google Scholar] [CrossRef]
- Krenn, H.W. Form and Function of Insect Mouthparts. Zoological Monographs 2019, 9–46. [Google Scholar] [CrossRef]
- Krenn, H.W. Fluid-Feeding Mouthparts. Zoological Monographs 2019, 47–99. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Daniel, T.L. Mechanics of Food Handling by Fluid-Feeding Insects. Regulatory Mechanisms in Insect Feeding 1995, 32–73. [Google Scholar] [CrossRef]
- Grimaldi, D.A. Basal Cyclorrhapha in Amber from the Cretaceous and Tertiary(Insecta: Diptera), and Their Relationships: Brachycera in Cretaceous Amber Part IX. Bulletin of the American Museum of Natural History 2018, 423, 1–97. [Google Scholar] [CrossRef]
- Gillung, J.P.; Winterton, S.L. Evolution of Fossil and Living Spider Flies Based on Morphological and Molecular Data (Diptera, Acroceridae). Systematic Entomology 2019, 44, 820–841. [Google Scholar] [CrossRef]
- Szucsich, N.U.; Krenn, H.W. Morphology and Function of the Proboscis in Bombyliidae (Diptera, Brachycera) and Implications for Proboscis Evolution in Brachycera. Zoomorphology 2000, 120, 79–90. [Google Scholar] [CrossRef]
- Krenn, H.W.; Mauss, V.; Plant, J. Evolution of the Suctorial Proboscis in Pollen Wasps (Masarinae, Vespidae). Arthropod Structure & Development 2002, 31, 103–120. [Google Scholar] [CrossRef]
- Krenn, H.W.; Plant, J.D.; Szucsich, N.U. Mouthparts of Flower-Visiting Insects. Arthropod Structure & Development 2005, 34, 1–40. [Google Scholar] [CrossRef]
- Labandeira, C.C. Fossil History and Evolutionary Ecology of Diptera and Their Associations with Plants. In The Evolutionary Biology of Flies; Yeates, D.K. & Wiegmann, B.M. (eds.) Columbia Univ. Press: New York, NY, 2005; pp. 217–272. [Google Scholar]
- Nilsson, L.A. The Evolution of Flowers with Deep Corolla Tubes. Nature 1988, 334, 147–149. [Google Scholar] [CrossRef]
- Pauw, A.; Stofberg, J.; Waterman, R.J. FLIES AND FLOWERS IN DARWIN’S RACE. Evolution 2009, 63, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Peris, D.; Pérez-de la Fuente, R.; Peñalver, E.; Delclòs, X.; Barrón, E.; Labandeira, C.C. False Blister Beetles and the Expansion of Gymnosperm-Insect Pollination Modes before Angiosperm Dominance. Current Biology 2017, 27, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Xu, C.; Jarzembowski, E.A. Ecological Radiations of Insects in the Mesozoic. Trends in Ecology & Evolution 2022, 37, 529–540. [Google Scholar] [CrossRef]
- Bicha, W.J. Biodiversity of Mecoptera. Insect Biodiversity 2018, 705–720. [Google Scholar] [CrossRef]
- Liu, Q.; Lu, X.; Zhang, Q.; Chen, J.; Zheng, X.; Zhang, W.; Liu, X.; Wang, B. High Niche Diversity in Mesozoic Pollinating Lacewings. Nature Communications 2018, 9. [Google Scholar] [CrossRef]
- Lin, X.; Labandeira, C.C.; Shih, C.; Hotton, C.L.; Ren, D. Life Habits and Evolutionary Biology of New Two-Winged Long-Proboscid Scorpionflies from Mid-Cretaceous Myanmar Amber. Nature Communications 2019, 10. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, B.; Bashkuev, A.S.; Aria, C.; Zhang, Q.; Zhang, H.; Tang, W.; Engel, M.S. Mouthpart Homologies and Life Habits of Mesozoic Long-Proboscid Scorpionflies. Science Advances 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Krassilov, V.A. & Rasnitsyn, A.P. Plant Remains from the Guts of Fossil Insects: Evolutionary and Paleoecological Inferences.; Vršanský, P., 1999; pp. 65–72.
- Gu, J.-J.; Qiao, G.-X.; Ren, D. Revision and New Taxa of Fossil Prophalangopsidae (Orthoptera: Ensifera). Journal of Orthoptera Research 2010, 19, 41–56. [Google Scholar] [CrossRef]
- Rasnitsyn, A.P.; Krassilov, V.A. The First Documented Occurrence of Phyllophagy in Pre-Cretaceous Insects: Leaf Tissues in the Gut of Upper Jurassic Insects from Southern Kazakhstan.
- Sendi, H.; Hinkelman, J.; Vršanská, L.; Kúdelová, T.; Kúdela, M.; Zuber, M.; van de Kamp, T.; Vršanský, P. Roach Nectarivory, Gymnosperm and Earliest Flower Pollination Evidence from Cretaceous Ambers. Biologia 2020, 75, 1613–1630. [Google Scholar] [CrossRef]
- Vlasáková, B.; Kalinová, B.; Gustafsson, M.H.G.; Teichert, H. Cockroaches as Pollinators of Clusia Aff. Sellowiana (Clusiaceae) on Inselbergs in French Guiana. Annals of Botany 2008, 102, 295–304. [Google Scholar] [CrossRef]
- Pérez-Gómez, Á.; León-Osper, M.; Pareja, D.; Robla, J. Flower Visits of Cockroaches (Insecta: Blattodea) in the Iberian Peninsula: Are They Neglected Pollinators? Journal of Applied Entomology 2023, 147, 565–576. [Google Scholar] [CrossRef]
- The Plecopteroid, Blattoid, and Orthopteroid Orders. In Entomology; Springer Netherlands: Dordrecht, 2005; pp. 147–197. ISBN 978-1-4020-3184-7.
- Huang, D.-Y.; Bechly, G.; Nel, P.; Engel, M.S.; Prokop, J.; Azar, D.; Cai, C.-Y.; van de Kamp, T.; Staniczek, A.H.; Garrouste, R.; et al. New Fossil Insect Order Permopsocida Elucidates Major Radiation and Evolution of Suction Feeding in Hemimetabolous Insects (Hexapoda: Acercaria). Scientific Reports 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- YOSHIZAWA, K.; LIENHARD, C. Bridging the Gap between Chewing and Sucking in the Hemipteroid Insects: <br/>new Insights from Cretaceous Amber. Zootaxa 2016, 4079. [Google Scholar] [CrossRef]
- Grimaldi, D. The Co-Radiations of Pollinating Insects and Angiosperms in the Cretaceous. Annals of the Missouri Botanical Garden 1999, 86, 373. [Google Scholar] [CrossRef]
- Mey, W.; Wichard, W.; Müller, P.; Wang, B. The Blueprint of the Amphiesmenoptera – Tarachoptera, a New Order of Insects from Burmese Amber (Insecta, Amphiesmenoptera). Fossil Record 2017, 20, 129–145. [Google Scholar] [CrossRef]
- Mey, W.; Wichard, W. Figure 10 from: Mey W, Wichard W (2023) Tarachoptera: The Extinct and Enigmatic Cousins of Trichoptera and Lepidoptera, with Descriptions of Two New Species. Contributions to Entomology 73(2): 137-146. Pensoft Publishers, 2023; [CrossRef]
- Rasnitsyn, A.P. New Triassic Hymenoptera from Central Asia. Paleontological Journal (Paleontologicheskii Zhurnal) 1964, 1, 88–96. [Google Scholar]
- Lara, M.B.; Rasnitsyn, A.P.; Zavattieri, A.M. Potrerilloxyela Menendezi Gen. et Sp. Nov. from the Late Triassic of Argentina: The Oldest Representative of Xyelidae (Hymenoptera: Symphyta) for Americas. Paleontological Journal 2014, 48, 182–190. [Google Scholar] [CrossRef]
- Denisova, E.A.; Kopylov, D.S.; Rasnitsyn, A.P. New Archexyelinae (Hymenoptera: Xyelidae) from the Triassic Madygen Formation of Kyrgyzstan. PalZ 2023, 98, 95–104. [Google Scholar] [CrossRef]
- Zhang, W.; Shih, C.; Labandeira, C.C.; Sohn, J.-C.; Davis, D.R.; Santiago-Blay, J.A.; Flint, O.; Ren, D. New Fossil Lepidoptera (Insecta: Amphiesmenoptera) from the Middle Jurassic Jiulongshan Formation of Northeastern China. PLoS ONE 2013, 8, e79500. [Google Scholar] [CrossRef]
- Mitter, C.; Davis, D.R.; Cummings, M.P. Phylogeny and Evolution of Lepidoptera. Annual Review of Entomology 2017, 62, 265–283. [Google Scholar] [CrossRef]
- van Eldijk, T.J.B.; Wappler, T.; Strother, P.K.; van der Weijst, C.M.H.; Rajaei, H.; Visscher, H.; van de Schootbrugge, B. A Triassic-Jurassic Window into the Evolution of Lepidoptera. Science Advances 2018, 4. [Google Scholar] [CrossRef]
- Sohn, J.-C.; Labandeira, C.C.; Davis, D.R. The Fossil Record and Taphonomy of Butterflies and Moths (Insecta, Lepidoptera): Implications for Evolutionary Diversity and Divergence-Time Estimates. BMC Evolutionary Biology 2015, 15. [Google Scholar] [CrossRef]
- Kalugina, N.S. & Kovalev, V.G. Dvukrylye Nasekomye Yury Sibiri (Diptera of the Jurassic of Siberia); Nauka, Moscow.: Moscow, 1985. [Google Scholar]
- Gilbert, F.; Jervis, M. Functional, Evolutionary and Ecological Aspects of Feeding-Related Mouthpart Specializations in Parasitoid Flies. Biological Journal of the Linnean Society 1998, 63, 495–535. [Google Scholar] [CrossRef]
- Krzemiński, W. & Krzemińska, E. Triassic Diptera: Descriptions, Revisions and Phylogenetic Relations. Acta Zoologica Cracoviensia 2003, 46, 153–184. [Google Scholar]
- ARILLO, A.; PEÑALVER, E.; PÉREZ-DE LA FUENTE, R.; DELCLÒS, X.; CRISCIONE, J.; BARDEN, P.M.; RICCIO, M.L.; GRIMALDI, D.A. Long-proboscid Brachyceran Flies in C Retaceous Amber ( D Iptera: S Tratiomyomorpha: Z Hangsolvidae). Systematic Entomology 2015, 40, 242–267. [Google Scholar] [CrossRef]
- Peñalver, E.; Arillo, A.; Pérez-de la Fuente, R.; Riccio, M.L.; Delclòs, X.; Barrón, E.; Grimaldi, D.A. Long-Proboscid Flies as Pollinators of Cretaceous Gymnosperms. Current Biology 2015, 25, 1917–1923. [Google Scholar] [CrossRef]
- Lukashevich, E.D.; Ribeiro, G.C. Mesozoic Fossils and the Phylogeny of Tipulomorpha (Insecta: Diptera). Journal of Systematic Palaeontology 2018, 17, 635–652. [Google Scholar] [CrossRef]
- Ollerton, J. Pollinator Diversity: Distribution, Ecological Function, and Conservation. Annual Review of Ecology, Evolution, and Systematics 2017, 48, 353–376. [Google Scholar] [CrossRef]
- Willmer, P.G.; Cunnold, H.; Ballantyne, G. Insights from Measuring Pollen Deposition: Quantifying the Pre-Eminence of Bees as Flower Visitors and Effective Pollinators. Arthropod-Plant Interactions 2017, 11, 411–425. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, B. Evolution of Lower Brachyceran Flies (Diptera) and Their Adaptive Radiation with Angiosperms. Front. Plant Sci. 2017, 8, 631. [Google Scholar] [CrossRef]
- Ren, D. Flower-Associated Brachycera Flies as Fossil Evidence for Jurassic Angiosperm Origins. Science 1998, 280, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Labandeira, C.C. How Old Is the Flower and the Fly? Science 1998, 280, 57–59. [Google Scholar] [CrossRef]
- Goldblatt, P.; Manning, J.C. The Long-Proboscid Fly Pollination System in Southern Africa. Annals of the Missouri Botanical Garden 2000, 87, 146. [Google Scholar] [CrossRef]
- Herendeen, P.S.; Friis, E.M.; Pedersen, K.R.; Crane, P.R. Palaeobotanical Redux: Revisiting the Age of the Angiosperms. Nature Plants 2017, 3. [Google Scholar] [CrossRef]
- Mostovski, M.B. A Revision of the Nemestrinid Flies (Diptera, Nemestrinidae) Described by Rohdendorf, and a Deascription of New Taxa of the Nemestrinidae from the Upper Jurassic of Kazakhstan. Paleontological Journal (Paleontologicheskii Zhurnal) 1998, 32, 369–375. [Google Scholar]
- Arnol’di, L.V. , Zherikin, V.V., Nikritin, L.M. & Ponomarenko, A.G. Mesozoic Coleoptera.; New Delhi and Calcutta: Oxonian Press Pvt Ltd, 1992. [Google Scholar]
- Ponomarenko, A.G. The Geological History of Beetles. In Biology, phylogeny, and classification of Coleoptera: papers celebrating the 80th Birthday of Roy A. Crowson.; Pakaluk, J. & Slipinski, S.A., 1995.
- Ponomarenko, A.G. Ecological Evolution of Beetles (Insecta: Coleoptera). Acta Zoologica Cracoviensia 2003, 46, 319–328. [Google Scholar]
- McKenna, D.D.; Sequeira, A.S.; Marvaldi, A.E.; Farrell, B.D. Temporal Lags and Overlap in the Diversification of Weevils and Flowering Plants. Proceedings of the National Academy of Sciences 2009, 106, 7083–7088. [Google Scholar] [CrossRef]
- Peris, D.; Kundrata, R.; Delclòs, X.; Mähler, B.; Ivie, M.A.; Rust, J.; Labandeira, C.C. Unlocking the Mystery of the Mid-Cretaceous Mysteriomorphidae (Coleoptera: Elateroidea) and Modalities in Transiting from Gymnosperms to Angiosperms. Scientific Reports 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Peris, D.; Labandeira, C.C.; Barrón, E.; Delclòs, X.; Rust, J.; Wang, B. Generalist Pollen-Feeding Beetles during the Mid-Cretaceous. iScience 2020, 23, 100913. [Google Scholar] [CrossRef] [PubMed]
- Peris, D.; Rust, J. Cretaceous Beetles (Insecta: Coleoptera) in Amber: The Palaeoecology of This Most Diverse Group of Insects. Zoological Journal of the Linnean Society 2019, 189, 1085–1104. [Google Scholar] [CrossRef]
- Bao, T.; Wang, B.; Li, J.; Dilcher, D. Pollination of Cretaceous Flowers. Proceedings of the National Academy of Sciences 2019, 116, 24707–24711. [Google Scholar] [CrossRef]
- CHRISTENHUSZ, M.J.M.; BYNG, J.W. The Number of Known Plants Species in the World and Its Annual Increase. Phytotaxa 2016, 261, 201. [Google Scholar] [CrossRef]
- Sauquet, H.; Ramírez-Barahona, S.; Magallón, S. What Is the Age of Flowering Plants? Journal of Experimental Botany 2022, 73, 3840–3853. [Google Scholar] [CrossRef]
- Crane, P.R.; Herendeen, P.; Friis, E.M. Fossils and Plant Phylogeny. American Journal of Botany 2004, 91, 1683–1699. [Google Scholar] [CrossRef]
- Clarke, J.T.; Warnock, R.C.M.; Donoghue, P.C.J. Establishing a Time-scale for Plant Evolution. New Phytologist 2011, 192, 266–301. [Google Scholar] [CrossRef] [PubMed]
- Barba-Montoya, J.; dos Reis, M.; Schneider, H.; Donoghue, P.C.J.; Yang, Z. Constraining Uncertainty in the Timescale of Angiosperm Evolution and the Veracity of a Cretaceous Terrestrial Revolution. New Phytologist 2018, 218, 819–834. [Google Scholar] [CrossRef]
- Silvestro, D.; Cascales-Miñana, B.; Bacon, C.D.; Antonelli, A. Revisiting the Origin and Diversification of Vascular Plants through a Comprehensive Bayesian Analysis of the Fossil Record. New Phytologist 2015, 207, 425–436. [Google Scholar] [CrossRef] [PubMed]
- van der Kooi, C.J.; Ollerton, J. The Origins of Flowering Plants and Pollinators. Science 2020, 368, 1306–1308. [Google Scholar] [CrossRef]
- Coiro, M.; Doyle, J.A.; Hilton, J. How Deep Is the Conflict between Molecular and Fossil Evidence on the Age of Angiosperms? New Phytologist 2019, 223, 83–99. [Google Scholar] [CrossRef] [PubMed]
- Meredith, R.W.; Janečka, J.E.; Gatesy, J.; Ryder, O.A.; Fisher, C.A.; Teeling, E.C.; Goodbla, A.; Eizirik, E.; Simão, T.L.L.; Stadler, T.; et al. Impacts of the Cretaceous Terrestrial Revolution and KPg Extinction on Mammal Diversification. Science 2011, 334, 521–524. [Google Scholar] [CrossRef]
- Augusto, L.; Davies, T.J.; Delzon, S.; De Schrijver, A. The Enigma of the Rise of Angiosperms: Can We Untie the Knot? Ecology Letters 2014, 17, 1326–1338. [Google Scholar] [CrossRef]
- Michener, C.D. Biogeography of the Bees. Annals of the Missouri Botanical Garden 1979, 66, 277. [Google Scholar] [CrossRef]
- The Bees of the World; Michener, C., Ed.; Johns Hopkins University Press, 2007; ISBN 978-0-8018-8573-0.
- Ollerton, J.; Winfree, R.; Tarrant, S. How Many Flowering Plants Are Pollinated by Animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Orr, M.C.; Hughes, A.C.; Chesters, D.; Pickering, J.; Zhu, C.-D.; Ascher, J.S. Global Patterns and Drivers of Bee Distribution. Current Biology 2021, 31, 451–458.e4. [Google Scholar] [CrossRef]
- Sann, M.; Niehuis, O.; Peters, R.S.; Mayer, C.; Kozlov, A.; Podsiadlowski, L.; Bank, S.; Meusemann, K.; Misof, B.; Bleidorn, C.; et al. Phylogenomic Analysis of Apoidea Sheds New Light on the Sister Group of Bees. BMC Evolutionary Biology 2018, 18. [Google Scholar] [CrossRef]
- Sann, M.; Meusemann, K.; Niehuis, O.; Escalona, H.E.; Mokrousov, M.; Ohl, M.; Pauli, T.; Schmid-Egger, C. Reanalysis of the Apoid Wasp Phylogeny with Additional Taxa and Sequence Data Confirms the Placement of Ammoplanidae as Sister to Bees. Systematic Entomology 2021, 46, 558–569. [Google Scholar] [CrossRef]
- Negri, I.; Mavris, C.; Di Prisco, G.; Caprio, E.; Pellecchia, M. Honey Bees (Apis Mellifera, L.) as Active Samplers of Airborne Particulate Matter. PLOS ONE 2015, 10, e0132491. [Google Scholar] [CrossRef]
- Pellecchia, M.; Papa, G.; Barbato, M.; Capitani, G.; Negri, I. Origin of Non-Exhaust PM in Cities by Individual Analysis of Particles Collected by Honey Bees (Apis Mellifera). Environmental Pollution 2023, 331, 121885. [Google Scholar] [CrossRef]
- Capitani, G.; Papa, G.; Pellecchia, M.; Negri, I. Disentangling Multiple PM Emission Sources in the Po Valley (Italy) Using Honey Bees. Heliyon 2021, 7, e06194. [Google Scholar] [CrossRef]
- Martins, A.C.; Melo, G.A.R.; Renner, S.S. The Corbiculate Bees Arose from New World Oil-Collecting Bees: Implications for the Origin of Pollen Baskets. Molecular Phylogenetics and Evolution 2014, 80, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Poinar, G.O.; Danforth, B.N. A Fossil Bee from Early Cretaceous Burmese Amber. Science 2006, 314, 614–614. [Google Scholar] [CrossRef]
- Danforth, B.N.; Poinar, G.O. Morphology, Classification, and Antiquity of Melittosphex Burmensis (Apoidea: Melittosphecidae) and Implications for Early Bee Evolution. Journal of Paleontology 2011, 85, 882–891. [Google Scholar] [CrossRef]
- ENGEL, M.S. A New Interpretation of the Oldest Fossil Bee (Hymenoptera: Apidae). American Museum Novitates 2000, 3296, 1–11. [Google Scholar] [CrossRef]
- Michener, C. D. & Grimaldi, D. The Oldest Fossil Bee: Apoidhistory, Evolutionary Stasis, and Antiquity of Social Behavior. Proceedings of the National Academy of Sciences of the United States ofAmerica 1988, 85, 6424–6426. [Google Scholar]
- Rehan, S.M.; Leys, R.; Schwarz, M.P. First Evidence for a Massive Extinction Event Affecting Bees Close to the K-T Boundary. PLoS ONE 2013, 8, e76683. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.S. Monophyly and Extensive Extinction of Advanced Eusocial Bees: Insights from an Unexpected Eocene Diversity. Proceedings of the National Academy of Sciences 2001, 98, 1661–1664. [Google Scholar] [CrossRef]
- Rust, J.; Singh, H.; Rana, R.S.; McCann, T.; Singh, L.; Anderson, K.; Sarkar, N.; Nascimbene, P.C.; Stebner, F.; Thomas, J.C.; et al. Biogeographic and Evolutionary Implications of a Diverse Paleobiota in Amber from the Early Eocene of India. Proceedings of the National Academy of Sciences 2010, 107, 18360–18365. [Google Scholar] [CrossRef]
- Winfree, R.; Aguilar, R.; Vázquez, D.P.; LeBuhn, G.; Aizen, M.A. A Meta-analysis of Bees’ Responses to Anthropogenic Disturbance. Ecology 2009, 90, 2068–2076. [Google Scholar] [CrossRef]
- Vanbergen, A.J.; Woodcock, B.A.; Gray, A.; Grant, F.; Telford, A.; Lambdon, P.; Chapman, D.S.; Pywell, R.F.; Heard, M.S.; Cavers, S. Grazing Alters Insect Visitation Networks and Plant Mating Systems. Functional Ecology 2013, 28, 178–189. [Google Scholar] [CrossRef]
- Kovács-Hostyánszki, A.; Espíndola, A.; Vanbergen, A.J.; Settele, J.; Kremen, C.; Dicks, L.V. Ecological Intensification to Mitigate Impacts of Conventional Intensive Land Use on Pollinators and Pollination. Ecology Letters 2017, 20, 673–689. [Google Scholar] [CrossRef]
- Yang, P.; Peng, Y.; Zhao, R.; Yang, D. Biological Characteristics, Threat Factors and Conservation Strategies for the Giant Honey Bee Apis Dorsata. Biodiversity Science 2018, 26, 476–485. [Google Scholar] [CrossRef]
- LeBuhn, G.; Vargas Luna, J. Pollinator Decline: What Do We Know about the Drivers of Solitary Bee Declines? Current Opinion in Insect Science 2021, 46, 106–111. [Google Scholar] [CrossRef]
- Papa, G.; Di Prisco, G.; Spini, G.; Puglisi, E.; Negri, I. Acute and Chronic Effects of Titanium Dioxide (TiO2) PM1 on Honey Bee Gut Microbiota under Laboratory Conditions. Sci Rep 2021, 11, 5946. [Google Scholar] [CrossRef]
- Plutino, M.; Bianchetto, E.; Durazzo, A.; Lucarini, M.; Lucini, L.; Negri, I. Rethinking the Connections between Ecosystem Services, Pollinators, Pollution, and Health: Focus on Air Pollution and Its Impacts. IJERPH 2022, 19, 2997. [Google Scholar] [CrossRef] [PubMed]
- TOLEDO-HERNÁNDEZ, E.; PEÑA-CHORA, G.; HERNÁNDEZ-VELÁZQUEZ, V.M.; LORMENDEZ, C.C.; TORIBIO-JIMÉNEZ, J.; ROMERO-RAMÍREZ, Y.; LEÓN-RODRÍGUEZ, R. The Stingless Bees (Hymenoptera: Apidae: Meliponini): A Review of the Current Threats to Their Survival. Apidologie 2022, 53. [Google Scholar] [CrossRef]
- Forister, M.L.; Dyer, L.A.; Gompert, Z.; Smilanich, A.M. Editorial Overview: Global Change Biology (2023) — Novel Perspectives on Futures, Mechanisms, and the Human Element of Insect Conservation in the Anthropocene. Current Opinion in Insect Science 2024, 62, 101175. [Google Scholar] [CrossRef]
- López-Vázquez, K.; Lara, C.; Corcuera, P.; Castillo-Guevara, C.; Cuautle, M. The Human Touch: A Meta-Analysis of Anthropogenic Effects on Plant-Pollinator Interaction Networks. PeerJ 2024, 12, e17647. [Google Scholar] [CrossRef]
- Margaoan, R.; Papa, G.; Nicolescu, A.; Cornea-Cipcigan, M.; Kösoğlu, M.; Topal, E.; Negri, I. Environmental Pollution Effect on Honey Bees and Their Derived Products: A Comprehensive Analysis. Environmental Science and Pollution Research 2024. [Google Scholar] [CrossRef]
- Saunders, M.E.; Lees, A.C.; Grames, E.M. Understanding and Counteracting the Denial of Insect Biodiversity Loss. Current Opinion in Insect Science 2025, 68, 101338. [Google Scholar] [CrossRef] [PubMed]
- Burkle, L.A.; Marlin, J.C.; Knight, T.M. Plant-Pollinator Interactions over 120 Years: Loss of Species, Co-Occurrence, and Function. Science 2013, 339, 1611–1615. [Google Scholar] [CrossRef]
- Tylianakis, J.M.; Didham, R.K.; Bascompte, J.; Wardle, D.A. Global Change and Species Interactions in Terrestrial Ecosystems. Ecology Letters 2008, 11, 1351–1363. [Google Scholar] [CrossRef]
- González-Varo, J.P.; Biesmeijer, J.C.; Bommarco, R.; Potts, S.G.; Schweiger, O.; Smith, H.G.; Steffan-Dewenter, I.; Szentgyörgyi, H.; Woyciechowski, M.; Vilà, M. Combined Effects of Global Change Pressures on Animal-Mediated Pollination. Trends in Ecology & Evolution 2013, 28, 524–530. [Google Scholar] [CrossRef]
- Geslin, B.; Le Féon, V.; Folschweiller, M.; Flacher, F.; Carmignac, D.; Motard, E.; Perret, S.; Dajoz, I. The Proportion of Impervious Surfaces at the Landscape Scale Structures Wild Bee Assemblages in a Densely Populated Region. Ecology and Evolution 2016, 6, 6599–6615. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel Declines in Pollinators and Insect-Pollinated Plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Hegland, S.J.; Nielsen, A.; Lázaro, A.; Bjerknes, A.; Totland, Ø. How Does Climate Warming Affect Plant-pollinator Interactions? Ecology Letters 2009, 12, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Thomson, J.D. Flowering Phenology, Fruiting Success and Progressive Deterioration of Pollination in an Early-Flowering Geophyte. Philosophical Transactions of the Royal Society B: Biological Sciences 2010, 365, 3187–3199. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of the evolutionary history of insects (top) and seed plants (bottom). Lineages that include taxa known to have engaged in mutualistic pollination relationships—both in the fossil record and in modern ecosystems—are highlighted. Modified and expanded from Labandeira et al. (2007).
Figure 1.
Schematic representation of the evolutionary history of insects (top) and seed plants (bottom). Lineages that include taxa known to have engaged in mutualistic pollination relationships—both in the fossil record and in modern ecosystems—are highlighted. Modified and expanded from Labandeira et al. (2007).
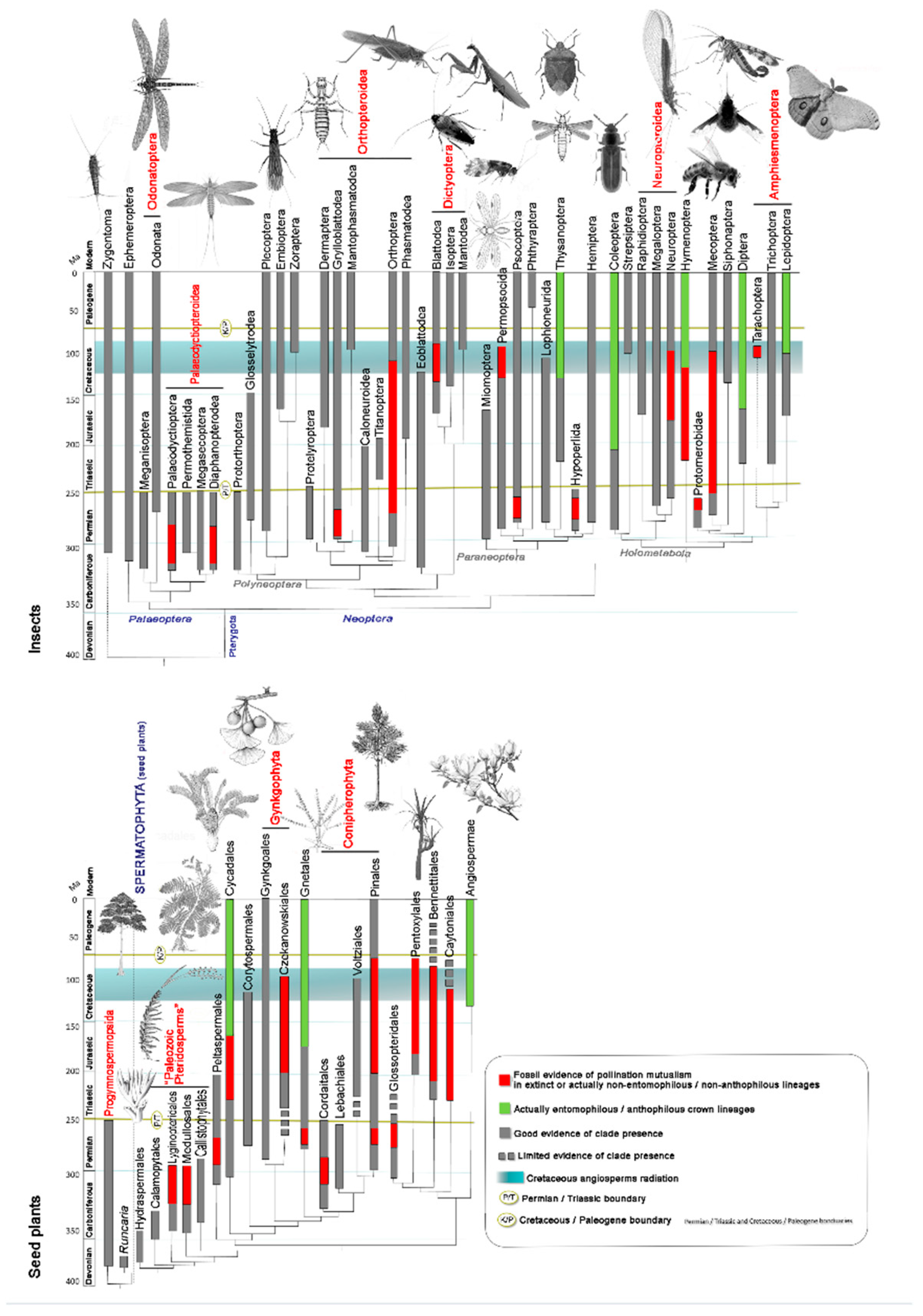
Figure 2.
Schematic representation of the four major phases of plant and herbivorous insect expansion over the past 420 million years—from early arthropods associated with primitive vascular plants, through the rise of gymnosperms and their pollinators, to the establishment of modern pollination networks in angiosperms. Each phase is distinguished by a different background color.
Figure 2.
Schematic representation of the four major phases of plant and herbivorous insect expansion over the past 420 million years—from early arthropods associated with primitive vascular plants, through the rise of gymnosperms and their pollinators, to the establishment of modern pollination networks in angiosperms. Each phase is distinguished by a different background color.
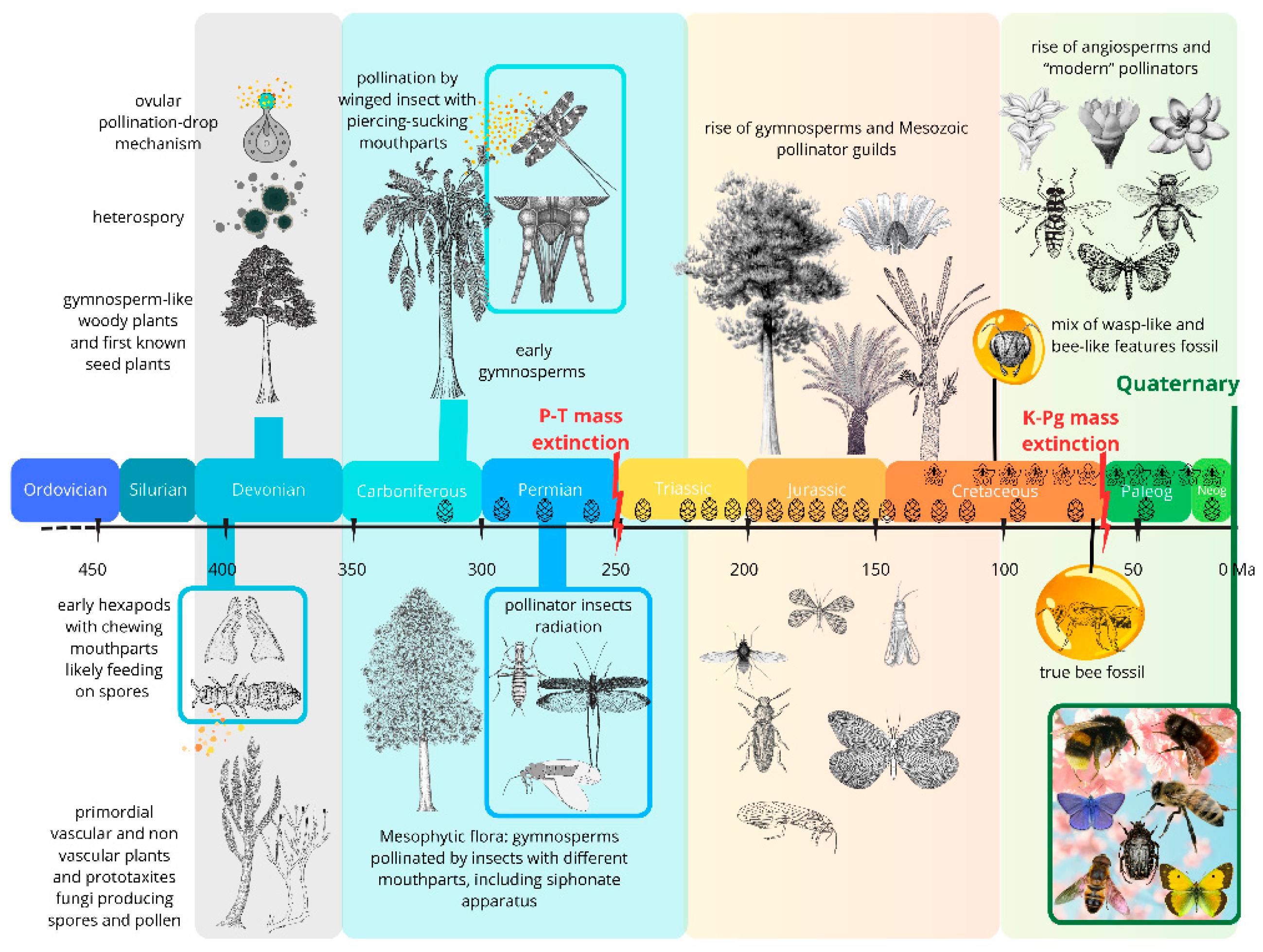
Figure 3.
Paleozoic early seed plants. (A) restoration of a Peltaspermales brunch with foliage and male reproductive organs (modified from Vajda et al., 2023). (B-C) Medullosales reproductive organs: (B) Ovule of Pachytesta attached to Alethopteris foliage; (C) pollen organs (Whittleseya) attached to Neuropteris foliage and, below, the structure of Dolerotheca showing complex system of pollen sacs (modified from: https://ucmp.berkeley.edu).
Figure 3.
Paleozoic early seed plants. (A) restoration of a Peltaspermales brunch with foliage and male reproductive organs (modified from Vajda et al., 2023). (B-C) Medullosales reproductive organs: (B) Ovule of Pachytesta attached to Alethopteris foliage; (C) pollen organs (Whittleseya) attached to Neuropteris foliage and, below, the structure of Dolerotheca showing complex system of pollen sacs (modified from: https://ucmp.berkeley.edu).
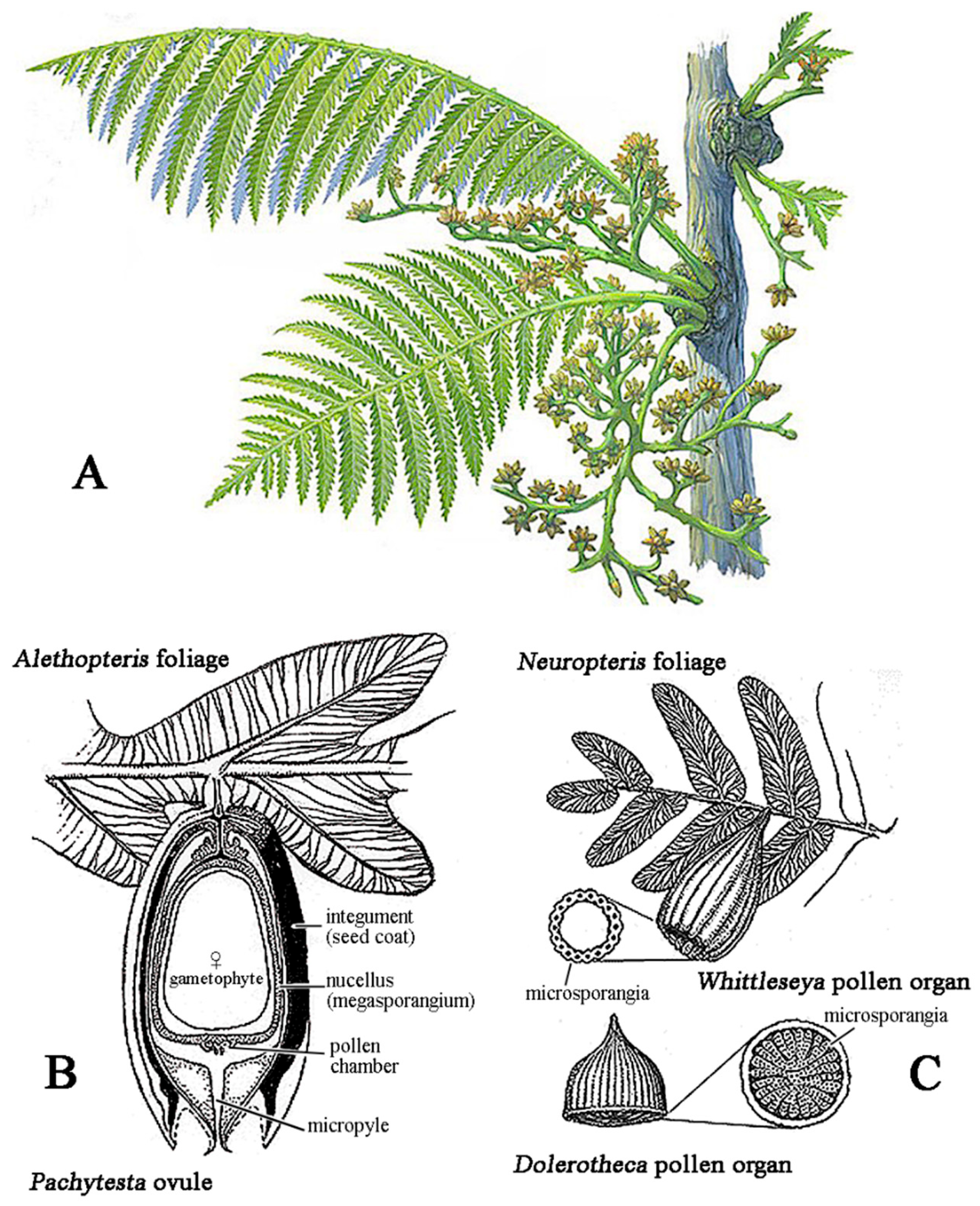
Figure 4.
The Permian saw the decline of palaeopterans (Shcherbakov et al. 2009; Aristov et al. 2013) and the rise of neopterans such as early bugs, thrips, and early holometabolans (beetles, lacewings and stem mecopteroids), while Orthoptera and Grylloblattodea diversified among Polyneoptera (Labandeira 2019; Prokop et al. 2023; Beutel et al. 2024). These groups also evolved increasingly specialized mouthparts from basic chewing mouthparts (Figure 5): Acercaria developed piercing–sucking styles, and holometabolans produced the earliest siphonate, non-piercing proboscises, establishing the major mouthpart classes that persist today (Wootton 1981; Labandeira 2019; Prokop et al. 2023).
Figure 4.
The Permian saw the decline of palaeopterans (Shcherbakov et al. 2009; Aristov et al. 2013) and the rise of neopterans such as early bugs, thrips, and early holometabolans (beetles, lacewings and stem mecopteroids), while Orthoptera and Grylloblattodea diversified among Polyneoptera (Labandeira 2019; Prokop et al. 2023; Beutel et al. 2024). These groups also evolved increasingly specialized mouthparts from basic chewing mouthparts (Figure 5): Acercaria developed piercing–sucking styles, and holometabolans produced the earliest siphonate, non-piercing proboscises, establishing the major mouthpart classes that persist today (Wootton 1981; Labandeira 2019; Prokop et al. 2023).
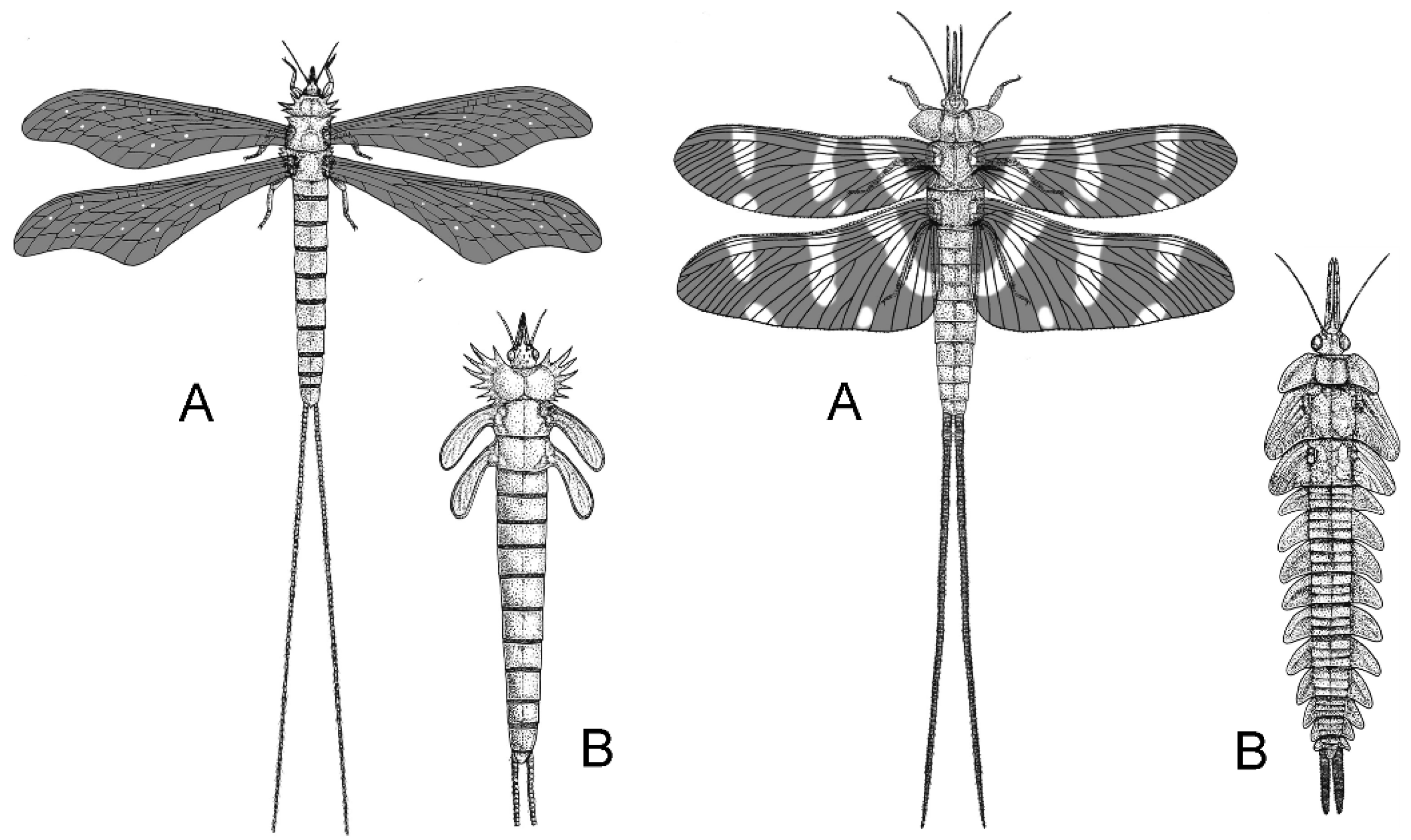
Figure 5.
Schematic representation of head and mouthpart morphology in various Acercaria, illustrating the hypothesized evolutionary transition from basal chewing to advanced piercing–sucking mouthparts. From left to right: psocid with basal mandibulate mouthparts; permopsocid with elongate mandibles, intermediate between chewing and piercing–sucking; thripidan ground pattern with piercing–sucking mouthparts; and hemipteran with highly specialized piercing–sucking structures. Color code: mandible—green; maxilla—yellow; anterior part of gena (mandibular lobe)—grey; posterior part of gena (maxillary lobe?)—blue. Modified from Huang et al. (2016).
Figure 5.
Schematic representation of head and mouthpart morphology in various Acercaria, illustrating the hypothesized evolutionary transition from basal chewing to advanced piercing–sucking mouthparts. From left to right: psocid with basal mandibulate mouthparts; permopsocid with elongate mandibles, intermediate between chewing and piercing–sucking; thripidan ground pattern with piercing–sucking mouthparts; and hemipteran with highly specialized piercing–sucking structures. Color code: mandible—green; maxilla—yellow; anterior part of gena (mandibular lobe)—grey; posterior part of gena (maxillary lobe?)—blue. Modified from Huang et al. (2016).
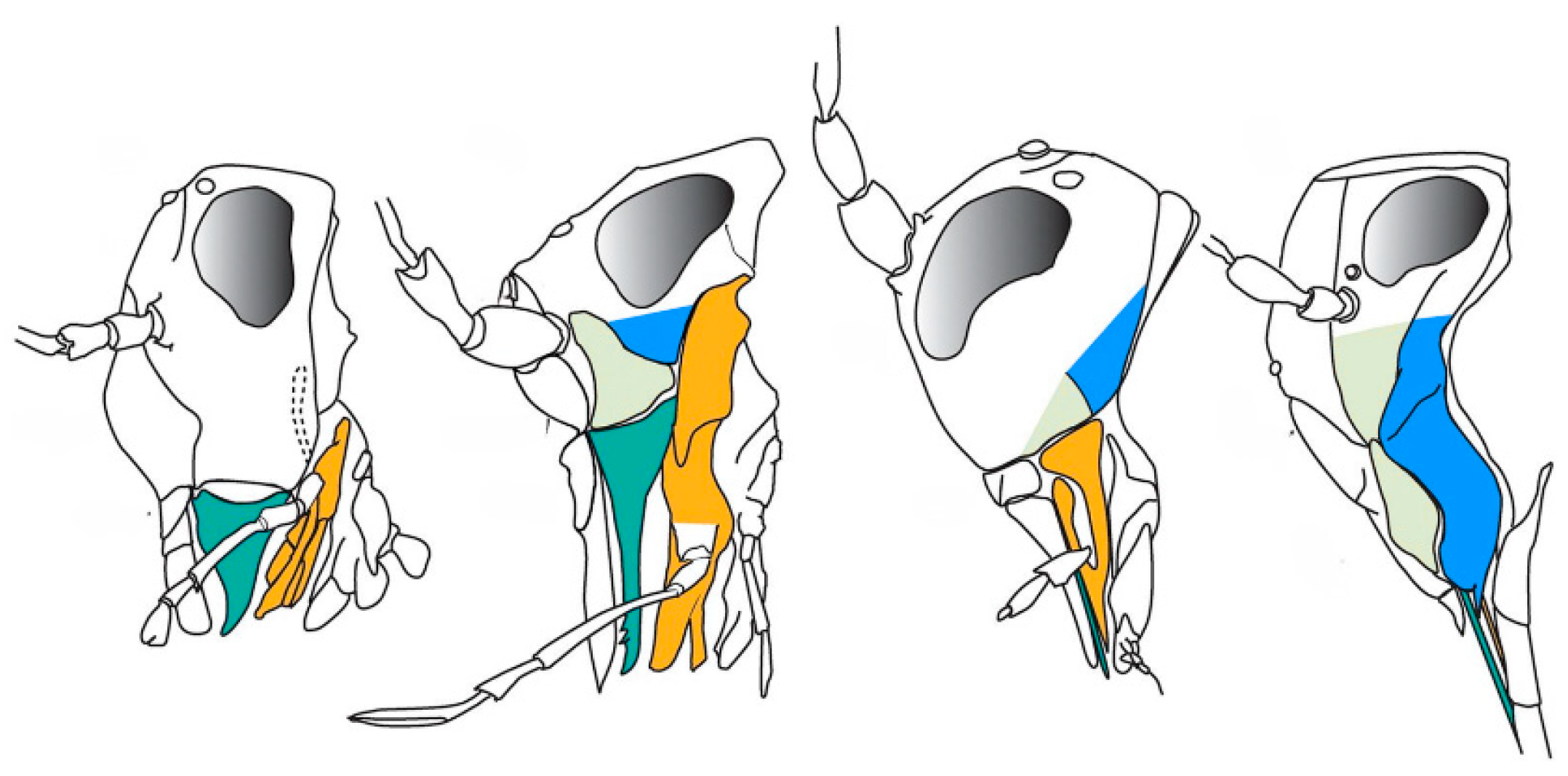
Figure 6.
Marimerobius splendens (Protomeropidae) feeding through its proboscis on a Peltaspermales cupula (shown dissected to reveal an ovule with pollination drops). This species represents the earliest known example of non-piercing haustellate mouthparts in holometabolous insects, likely adapted for absorbing sugary fluids. Multiple fossils of M. splendens have been found in association with Peltaspermales pollen, suggesting the involvement of these insects in early gymnosperm pollination during the Lower Permian. Modified from Khramov et al. (2022).
Figure 6.
Marimerobius splendens (Protomeropidae) feeding through its proboscis on a Peltaspermales cupula (shown dissected to reveal an ovule with pollination drops). This species represents the earliest known example of non-piercing haustellate mouthparts in holometabolous insects, likely adapted for absorbing sugary fluids. Multiple fossils of M. splendens have been found in association with Peltaspermales pollen, suggesting the involvement of these insects in early gymnosperm pollination during the Lower Permian. Modified from Khramov et al. (2022).
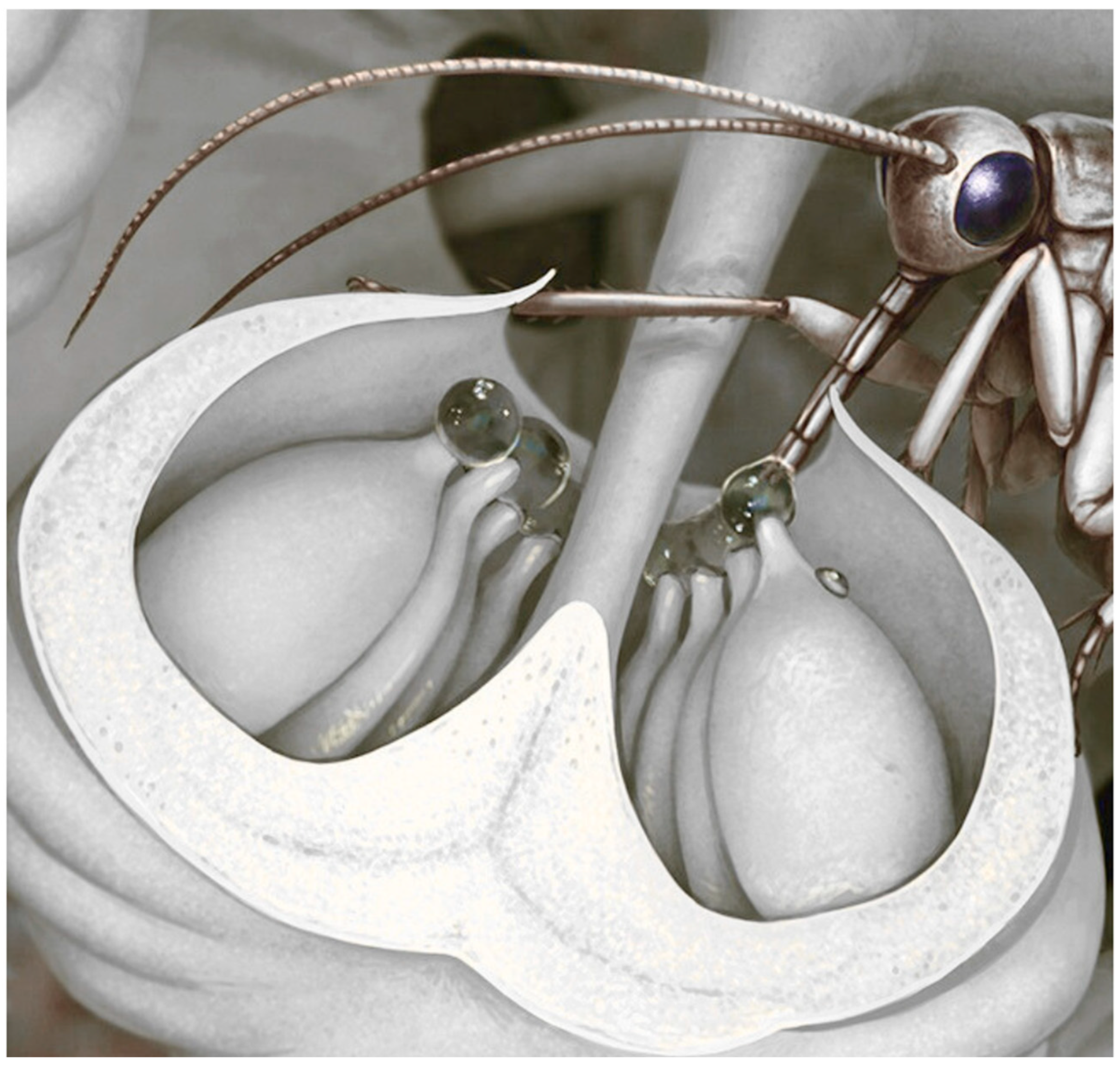
Figure 7.
Bennettitales, Williamsoniaceae. Top: restoration of the shrub-sized Wielandiella, showing dichotomous branching and female cones at various stages of maturation and senescence (from Pott, 2014). Bottom: schematic reconstruction of female (left) and male (right) “flower-like” strobili (modified from Rothwell et al., 2009). Abbreviations: m, micropyles; o, ovules; ps, pollination sacs.
Figure 7.
Bennettitales, Williamsoniaceae. Top: restoration of the shrub-sized Wielandiella, showing dichotomous branching and female cones at various stages of maturation and senescence (from Pott, 2014). Bottom: schematic reconstruction of female (left) and male (right) “flower-like” strobili (modified from Rothwell et al., 2009). Abbreviations: m, micropyles; o, ovules; ps, pollination sacs.
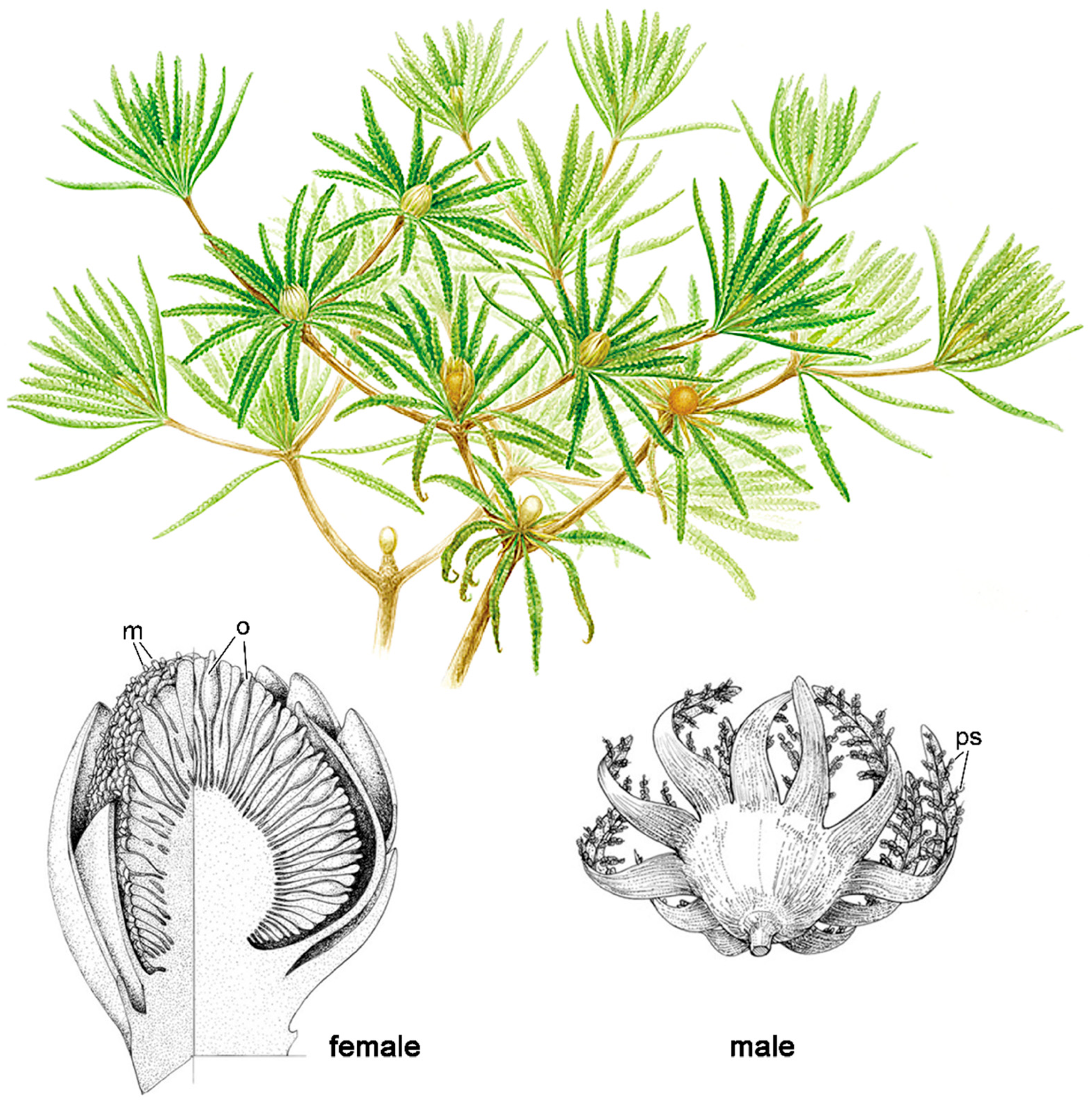
Figure 8.
Ephedra foeminea (Gnetales, Epherdaceae): male cone with pollination drops produced by sterile ovules at the distal centre of the cone (left) and (right) female cones with pollination drops exposed at the micropylar openings (modified from Rydin & Bolinder 2015).
Figure 8.
Ephedra foeminea (Gnetales, Epherdaceae): male cone with pollination drops produced by sterile ovules at the distal centre of the cone (left) and (right) female cones with pollination drops exposed at the micropylar openings (modified from Rydin & Bolinder 2015).

Figure 9.
Reconstruction of a Lower Cretaceous association between an entomophilous female cone of a cheirolepidiaceous conifer (Alvinia) and a group of meropsychid, specialized anthophilous scorpionflies (Vitimopsyche), which acted as vectors of Classopollis pollen. The insects were likely attracted by the bright coloration of the funnel orifice and its sugary secretions (modified from Labandeira, 2010).
Figure 9.
Reconstruction of a Lower Cretaceous association between an entomophilous female cone of a cheirolepidiaceous conifer (Alvinia) and a group of meropsychid, specialized anthophilous scorpionflies (Vitimopsyche), which acted as vectors of Classopollis pollen. The insects were likely attracted by the bright coloration of the funnel orifice and its sugary secretions (modified from Labandeira, 2010).
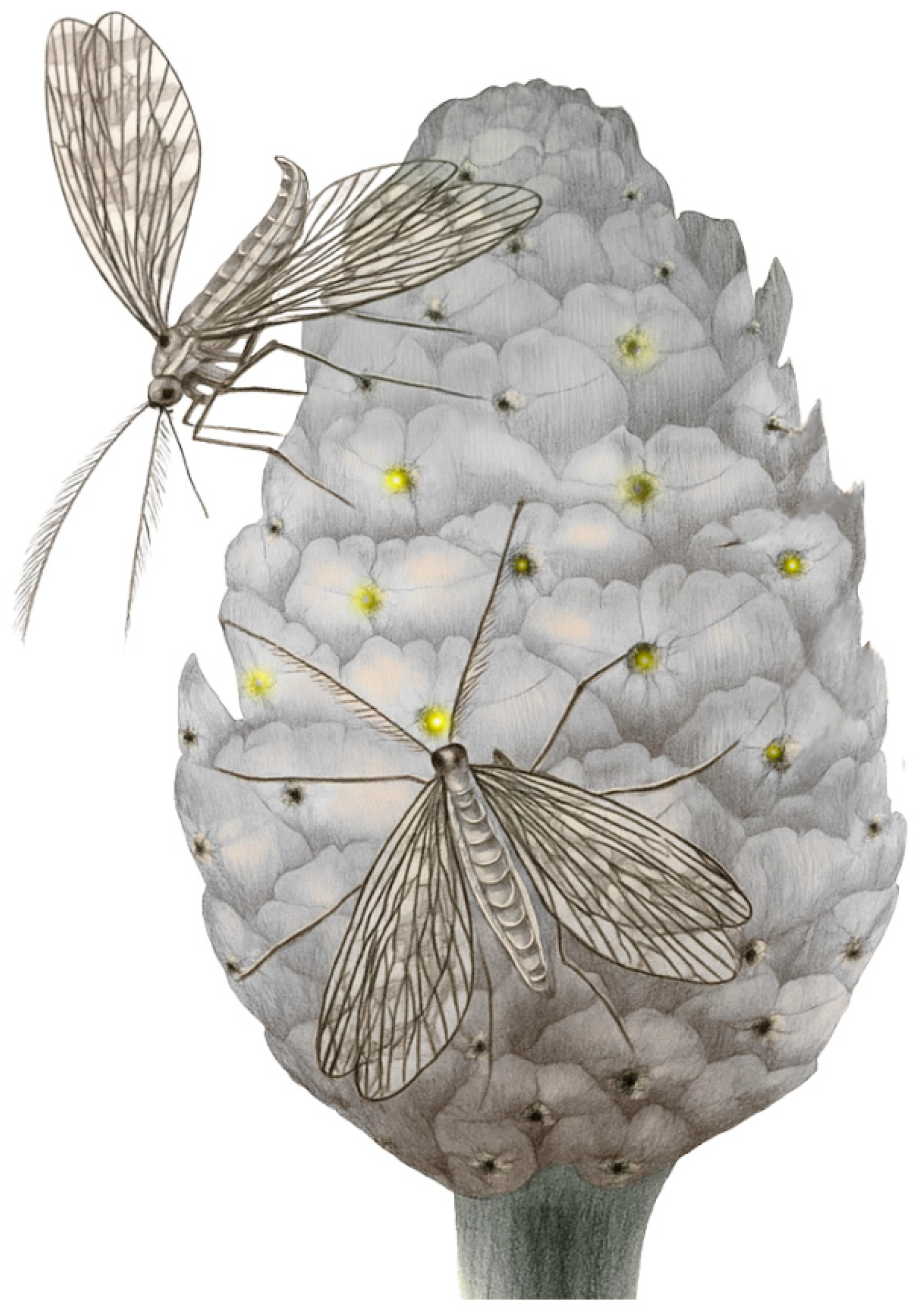
Figure 10.
Kalligrammatidae (Neuroptera) — often referred to as the “Mesozoic butterflies” — represent a remarkable example of convergent evolution with modern Papilionoidea, which appeared more than 50 million years after their extinction. Their resemblance to modern butterflies was striking: large, colorful wings bearing eyespots and covered with scale-like structures, though anatomically distinct from those of Lepidoptera. Their elongated proboscises enabled them to feed on sugary secretions from the cones of entomophilous gymnosperms such as Bennettitales and Cheirolepidiaceae. The decline of these plant groups, coinciding with the expansion of angiosperms, likely contributed to the extinction of these extraordinary insects. (A) Fossil of the kalligrammatid Oregramma compared with (B) a modern butterfly (modified from https://www.enn.com). (C) Schematic restoration of Oregramma morphology, showing the right side with left wings removed (modified from https://palaeoflora.blogspot.com).
Figure 10.
Kalligrammatidae (Neuroptera) — often referred to as the “Mesozoic butterflies” — represent a remarkable example of convergent evolution with modern Papilionoidea, which appeared more than 50 million years after their extinction. Their resemblance to modern butterflies was striking: large, colorful wings bearing eyespots and covered with scale-like structures, though anatomically distinct from those of Lepidoptera. Their elongated proboscises enabled them to feed on sugary secretions from the cones of entomophilous gymnosperms such as Bennettitales and Cheirolepidiaceae. The decline of these plant groups, coinciding with the expansion of angiosperms, likely contributed to the extinction of these extraordinary insects. (A) Fossil of the kalligrammatid Oregramma compared with (B) a modern butterfly (modified from https://www.enn.com). (C) Schematic restoration of Oregramma morphology, showing the right side with left wings removed (modified from https://palaeoflora.blogspot.com).
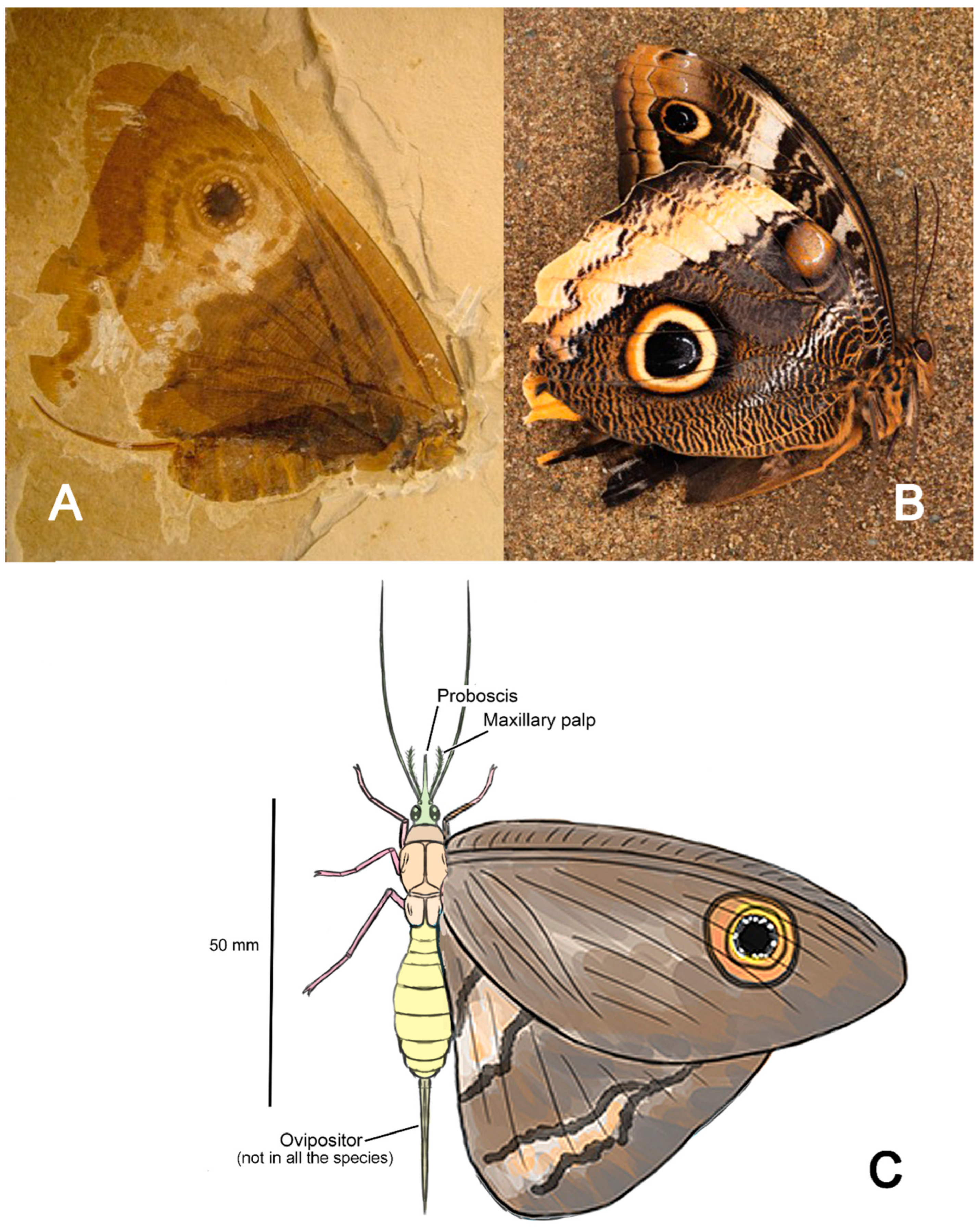
Figure 11.
Cretotrigona prisca, the earliest known fossil bee, discovered in Cretaceous (presumed Maastrichtian) amber from New Jersey, USA (modified from Friis et al 2011).
Figure 11.
Cretotrigona prisca, the earliest known fossil bee, discovered in Cretaceous (presumed Maastrichtian) amber from New Jersey, USA (modified from Friis et al 2011).
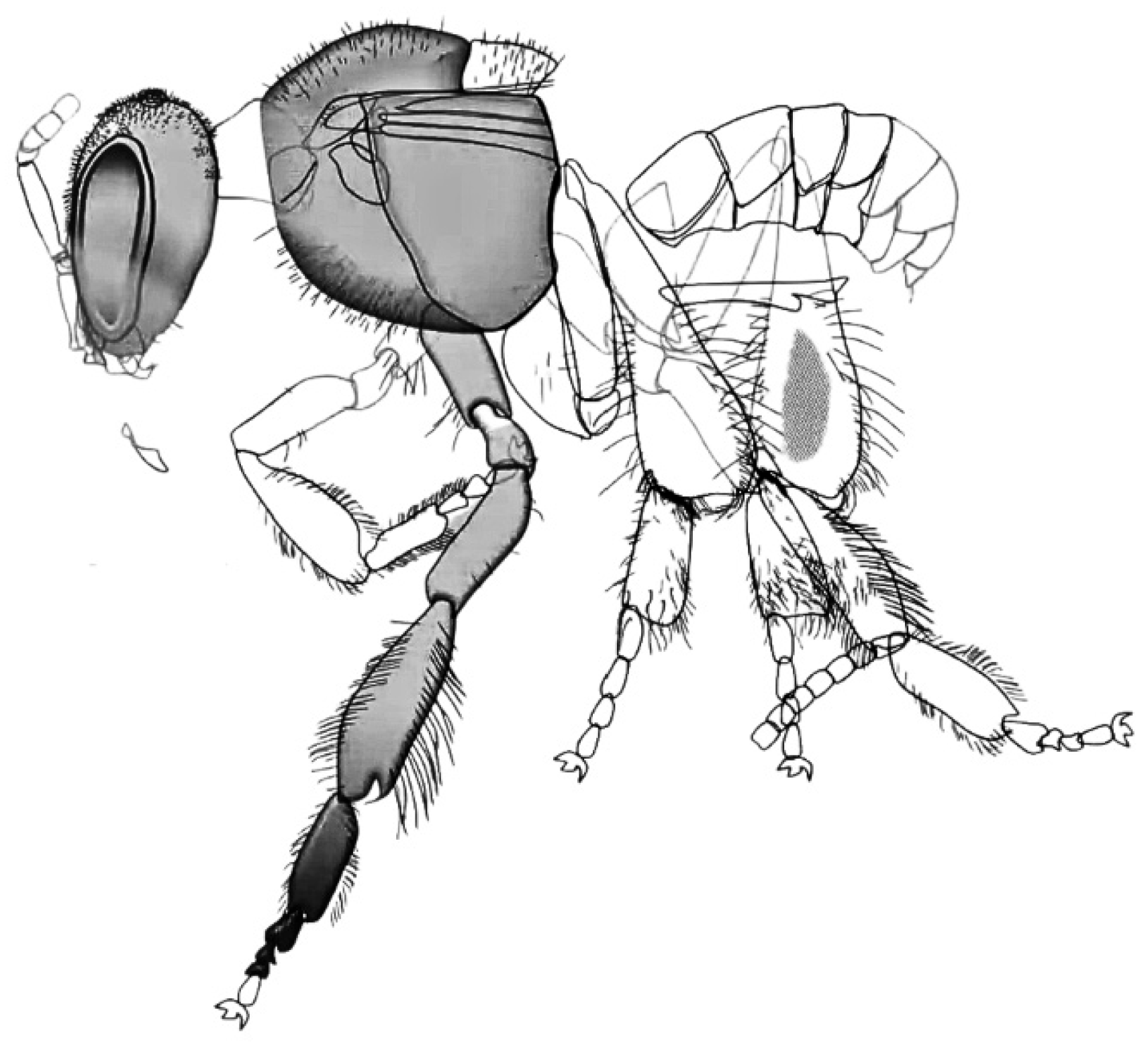
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



