Submitted:
13 November 2025
Posted:
14 November 2025
You are already at the latest version
Abstract
As a primary macronutrient, nitrogen is integral to plant growth and developmental reg-ulation; ammonium transporters (AMTs) mediate its absorption and involvement in ni-trogen metabolism. In this study, nine BcAMT genes were identified from flowering Chi-nese cabbage (Brassica campestris), which were systematically categorized into two sub-families. Their evolutionary relationships, conserved motifs, chromosomal distribution, cis-regulatory elements, and expression profiling were systematically characterized. RNA-sequencing and RT-PCR analyses demonstrated that BcAMT1.1 was abundantly expressed in roots, leaves, and stems of flowering Chinese cabbage, and was markedly up-regulated under nitrogen deficiency or low-nitrogen conditions. Subcellular location using GFP-fusion demonstrated that BcAMT1.1 localized to the plasma membrane. Func-tional assays through heterologous expression in the yeast mutant strain 31019b and transgenic Arabidopsis validated that BcAMT1.1 acted as a functional ammonium trans-porter. Compared with wildtype, overexpressing BcAMT1.1 promoted seedling growth, enhanced NH4+ influxes and NO3- effluxes under low nitrogen conditions, and signifi-cantly increased the transcription levels of key nitrogen assimilation genes (i.e., AtGLN1.1, AtGLN2, AtGDH2). Collectively, our findings enhance the fundamental understanding of BcAMT gene functions, and highlighting of BcAMT1.1 as a crucial component in nitrogen uptake and assimilation under low nitrogen conditions, and providing valuable genetic resources for improving nitrogen efficiency in vegetable crops.
Keywords:
ammonium transporter
; flowering Chinese cabbage
; low nitrogen conditions
; genomewide analysis
1. Introduction
Nitrogen, as a critical macronutrient, is indispensable for plant physiological functioning and is responsible for up to 50% of biomass accumulation [1]. Due to its significant added value, nitrogen fertilizers are often applied in excess of actual crop demands as a low-cost insurance against potential yield losses [2,3]. In vegetable crop production, over-application, combined with low nitrogen recovery and intensive irrigation, markedly reduces nitrogen use efficiency (NUE) and generates dual environmental and health risks, including water pollution, greenhouse gas emissions, and excessive nitrate accumulation in edible plant organs, particularly in leafy vegetables [2]. Thus, improving NUE is essential for sustainable agricultural production and relies on a thorough understanding of the mechanisms governing nitrogen absorption, transport, and metabolism.
Ammonium (NH4+) and nitrate (NO3-) are the main inorganic nitrogen sources utilized by plants [4]. In soils, NH4+ concentrations (20 to 200 μmol·L-1) are lower than those of NO3- (100 μmol·L-1 to 70 mmol·L-1) [5,6]. Although NO3- is generally the predominant nitrogen form for most plants, NH4+ may be preferred under nitrogen deficiency or elevated atmosphere carbon dioxide, because it requires less energy for assimilation via the glutamine synthetase/NADH-dependent glutamate synthase (GS/GOGAT) pathway [7,8]. Nevertheless, excessive NH4+ can be toxic to plants [6,9]. Therefore, NH4+ uptake and transportation require to be tightly regulated. NH4+ uptake in plants is mediated by two distinct systems: the high-affinity transport system (HATS) and the low-affinity transport system (LATS). HATS operates predominantly at low NH4+ levels, ensuring efficient NH4+ absorption, whereas LATS becomes more active at higher NH4+ concentrations [10,11]. NH4+ uptake at low NH4+ level (< 1 mmol·L-1) is predominantly regulated by HATS [12]. Ammonium transporters (AMTs), members of AMT/methylammonium permease (MEP)/rhesus (Rh) protein family, are key mediators of NH4+ uptake and transport in plants [11,13]. AMT proteins, mainly found in plants, are grouped into two major subfamilies: the AMT1 subfamily, comprising only AMT1 cluster, and the AMT2 subfamily, including AMT2, AMT3, and AMT4 clusters [14]. The AMT2 shows high homology to yeast MEP and Escherichia coli AmtB, but less homologous to AMT1 proteins [15].
Following the identification of the first AMT in Arabidopsis [16], the members of AMT have subsequently been reported in various species, including rice [17,18], rapeseed [19], soybean [20], and tobacco [21]. In Arabidopsis, six AMTs have been isolated and characterized, including five members of AMT1 subfamily and a single AMT2 isoform [22,23]. Among them, AtAMT1.1 ~ AtAMT1.3, and AtAMT1.5 are primarily expressed in roots and strongly induced by nitrogen deficiency or low nitrogen conditions, with AtAMT1.1 ~ AtAMT1.3 account for nearly 90% of high-affinity NH4+ absorption [22]. AtAMT1.4 is a pollen-specific AMT member and mediates NH4+ absorption in pollen [24]. In rice, 12 AMTs are isolated, OsAMT1.1 ~ OsAMT1.3 account for up to 95% of high-affinity NH4+ uptake [18]; the knockout of OsAMT1.1 reduces NH4+ uptake by 25~30% [25]. In rapeseed, 20 BnaAMT genes are identified, with 14 members from the AMT1 subfamily and six from the AMT2 subfamily, most of which are highly responsive to external nitrogen conditions [19].
Flowering Chinese cabbage (Brassica campestris L. ssp. chinensis var. utilis Tsen et Lee), is typical stalk vegetable originated from South China, possessing significant health and economic values [26,27]. Flowering Chinese cabbage is now cultivated globally, particularly in Asian countries [26,28], and requires substantial nitrogen input to achieve optimal yield; hence, excessive NO3- often accumulates in edible organs, especially in stalks, posing potential health risks [29,30]. Understanding the molecular mechanism of nitrogen uptake and transportation in flowering Chinese cabbage could inform strategies to optimize fertilization practices and improve NUE. Here, we present a genome-wide survey and detailed analysis of the AMT gene family in flowering Chinese cabbage. We examined their phylogenetic relationships, conserved motifs, gene structures, chromosomal localization, syntenic patterns, cis-regulatory elements, and expression patterns. Among these, BcAMT1.1 showed strong transcriptional induction under low nitrogen concentration. Functional assays of BcAMT1.1 in yeast mutant strain 31019b confirmed its functional NH4+ transport activity, and overexpression in Arabidopsis enhanced seedling growth, NH4+ uptake, and nitrogen assimilation-related genes transcription under low-nitrogen conditions. This study underscores the pivotal role of AMTs in plant growth and nitrogen metabolism, with BcAMT1.1 being critical for nitrogen acquisition and assimilation under low nitrogen conditions, and provide genetic resources for improving NUE in vegetable crops.
2. Results
2.1. Identification and Chromosomal Localization of BcAMT Gene Family in Flowering Chinese Cabbage
Potential AMT members were identified from flowering Chinese cabbage genome through BLASTP searches using Arabidopsis AMT protein sequences, obtained from the BRAD database. Candidate sequences were confirmed to contain the Pfam domain of ammonium transporter family (PF00909). In total, nine BcAMT members were identified and designated BcAMT1.1 to BcAMT2.1-like, according to their homology with Arabidopsis AMTs. The physiochemical properties of BcAMTs were assessed using the EXPASY ProtParam, including molecular weight (MW), isoelectric point (pI), amino acids number, and grand average of hydropathy (GRAVY). Amino acid number ranged from 476 (BcAMT1.3-like) to 512 aa (BcAMT1.2), with the predicted MW ranging from 50.79 (BcAMT1.3-like) to 54.88 kDa (BcAMT1.2). The pI values varied from 5.45 (BcAMT1.4-like) to 7.73 (BcAMT1.2), indicating that most members were weakly acidic. All instability indices were below 40, and the GRAVY value was higher than zero, suggesting that these BcAMT proteins were stable and hydrophobic. Subcellar localization analysis revealed that all BcAMT proteins are positioned in the plasma membrane, with 9 to 11 transmembrane domains (Table 1), consistent with membrane-associated transport functions of AMT. Chromosomal mapping showed that nine BcAMTs were dispersed across six chromosomes (Chr) of flowering Chinese cabbage genome (Table 1; Figure 1), BcAMT1.3-like and BcAMT1.4 were located on Chr1; BcAMT1.4-like and BcAMT1.5 on Chr3; BcAMT1.1 and BcAMT2.1 on Chr5; BcAMT2.1-like, BcAMT1.3, and BcAMT1.2 on Chr4, Chr7, and Chr9.
2.2. Phylogenetic Tree and Conserved Domains Analyses of BcAMT Gene Family
To evaluate the evolutionary relationships of AMT proteins, a maximum likelihood (ML) phylogenetic tree was generated in MEGA 7.0, incorporating AMTs from flowering Chinese cabbage, Arabidopsis thaliana, Brassica napus, Solanum lycopersicum, Nicotiana tabacum, Oryza sativa, and Populus trichocarpa (Table S1). The phylogenetic tree resolved into two dominant clades (Figure 2A): seven BcAMTs grouped within the AMT1 subfamily and two within AMT2. Both two AMT2 members were found in Cluster II, and no flowering Chinese cabbage or other Cruciferae AMTs were placed in cluster III or IV. Homology analysis showed that BcAMTs shared high homology with Arabidopsis AtAMTs and rapeseed BnaAMTs, with orthologous counterparts in flowering Chinese cabbage (Figure 2A). Furthermore, several paralogous gene pairs were identified, including BcAMT1.3/BcAMT1.3-like, BcAMT1.4/ BcAMT1.4-like, and BcAMT2.1/BcAMT2.1-like, each gene pair clustering with the corresponding AMT members in Arabidopsis and rapeseed, suggesting possible functional redundancy. Signature motif analysis, performed in Jalview v2.11.2.7, revealed that all AMT1 subfamily members shared the conserved sequence “DFAGSGVVHMVGGIAGLWGALIEGPR”, except BnaAMT1.3c in rapeseed; while all AMT2 members universally contained the conserved sequence “DYSGGYIHLSSGVAGFTAAYW WGPR”, except OsAMT4.1 in O. sativa (Figure 2B,C). The signatures motifs strongly support the phylogenetic classification.
2.3. Conserved Motifs and Gene Structure of BcAMT Gene Family
Motif composition and gene structure were analyzed in relation to evolutionary classification. Among all BcAMT members, ten consensus motifs were identified, which were present in seven BcAMT1 subfamily, whereas BcAMT2.1 and BcAMT2.1-like contained seven and six motifs, respectively. Six conversed motifs (motif 3, 4, 5, 6, 8 and 10) were conserved across all BcAMT members (Figure 3A). Domain prediction showed that all BcAMT proteins contained the characteristic ammonium transporter domain (amt) (Figure 3B). Gene structure analysis revealed marked differences between subfamilies: BcAMT2 members harbored four introns, while most BcAMT1 members were intronless except BcAMT1.2 and BcAMT1.3-like (Figure 3C). These differences in gene structure may underpin functional divergence between BcAMT1 and BcAMT2 subfamilies.
2.4. Gene Duplication and Synteny Analyses of the BcAMT Gene Family
To understand the evolutionary expansion of the BcAMT gene family, genome-wide collinearity analysis was performed using the MCScan X module in TBtools-II. Five segmentally gene pairs were identified: BcAMT1.3/BcAMT1.3-like, BcAMT1.4/BcAMT1.4-like, BcAMT1.3/ BcAMT1.5, BcAMT1.5/BcAMT1.3-like, and BcAMT2.1/BcAMT2.1-like (Figure 4A; Table 2). The segmental duplications likely played a vital role in the expansion of BcAMT family in flowering Chinese cabbage. Based on the rates of synonymous (Ks) and nonsynonymous (Ka) substitution, and assuming a neutral divergence rate of 1.5×10-8 substitutions per site per year, divergence times of these paralogous pairs were estimated to range from 10.5503 to 24.7684 million years ago (MYA), averaging ~16.9060 MYA (Table 2). The ratios of Ka/Ks ranged from 0.0962 and 0.1383, remaining well below 1.0, implying that these duplicates of BcAMTs have experienced intense purifying selection during evolution.
Comparative synteny mapping revealed the extensive collinear relationships between BcAMT genes and AMTs in Arabidopsis and B. napus, while no syntenic associations with O. sativa. Seven BcAMT genes exhibited syntenic correlation with Arabidopsis AMTs and nine with B. napus AMTs (Figure 4B; Table S2). Specific relationships included BcAMT1.3, BcAMT1.3-like, BcAMT1.5 with AtAMT1.1 and AtAMT1.5; BcAMT1.4 and BcAMT1.4-like with AtAMT1.4; BcAMT2.1 and BcAMT2.1-like with AtAMT2.1 (Figure 4; Table S2). In B. napus, there were many syntenic relationships, BcAMT1.1 with BnaAMT1.1b, BcAMT1.2 with BnaAMT1.2a and BnaAMT1.2b; BcAMT1.3, BcAMT1.3-like and BcAMT1.5 with BnaAMT1.3a and BnaAMT1.5a; BcAMT1.4 and BcAMT1.4-like with BnaAMT1.4a, BnaAMT1.4b and BnaAMT1.4c; BcAMT2.1 and BcAMT2.1-like with BnaAMT2.1a, BnaAMT2.1c, and BnaAMT2.2c (Figure 4; Table S2). These syntenic relationships highlight a conserved genomic context of BcAMTs within Brassica species and the divergence from monocot AMTs.
2.5. Analysis of Cis-Acting Elements in BcAMT Promoter Regions
To clarify transcriptional regulation of BcAMT, 2,000 bp upstream promoter regions were used to analyze cis-acting elements using PlantCARE database. 42 types of elements were detected and grouped into four functional categories. (1) Light-responsive elements: 14 light-related regulatory motifs, including G-Box, GT1-motif, and TCT-motif, were detected in most BcAMT promoters. (2) Growth- and development-related elements: AAGAA-motif, O2-site, and CAT-box motifs were present in BcAMT promoters, while circadian element was identified exclusively in the promoters of BcAMT1.1 and BcAMT1.4. (3) Hormone-response elements: Promoters contained binding sites responsive to gibberellin (GARE-motif, TATC-box, P-box), auxin (AuxRR-core, TGA-element), abscisic acid (ABRE), salicylic acid (TCA-element), methyl jasmonate (CGTCA-motif), and ethylene (ERE). Notably, ABRE, ERE, and CGTCA-motif were widely distributed among promoters. (4) Stress-related elements: 13 types of stress-related motifs were detected, including stress-responsiveness element (STRE), anaerobic induction element (ARE), salicylic acid or pathogen-induced signaling (as-1), low-temperature responsiveness (LTR), wound-responsive element (WUN-motif), and several MYB/ MYC- binding sites (Figure 5). These findings indicate that BcAMTs transcription may be regulated by various environmental and endogenous factors, including light, hormonal signaling, growth and development, and abiotic or biotic stresses.
2.6. Expression Patterns of BcAMTs Across Tissues of Flowering Chinese Cabbage and Under Different Nitrogen Forms
2.6.1. Tissue-Specific Expression
The data of RNA-sequencing (RNA-seq) from the cultivar ‘49 Caixin’(SRP427920) revealed distinct spatial expression patterns for BcAMTs in seven tissues (roots, stems, flowers, seedpods, and young/mature/senescent leaves). BcAMT1.1, BcAMT2.1, and BcAMT2.1-like were broadly expressed in tissues, with BcAMT1.1 showing high transcript abundance in leaves, stems, and roots; BcAMT2.1 in young/mature leaves, seedpods, stems and roots; and BcAMT2.1-like in young/mature leaves, and stems (Figure 6A). BcAMT1.2 was strongly expressed in roots and stems, BcAMT1.3 and BAMT1.5 were specifically high expressed in roots, BcAMT1.4 and BcAMT1.4-like were specifically expressed in flowers, while BcAMT1.3-like exhibited low expression in most tissues (Figure 6A). The results indicate that BcAMTs may play distinct roles in regulating plant growth and development of flowering Chinese cabbage.
2.6.2. BcAMTs Response to Nitrogen Forms
Expression changes were evaluated following treatments with different nitrogen sources: 1 mmol·L-1 NH4+, 0.5 mmol·L-1 NH4+ and 0.5 mmol·L-1 NO3-, and 1 mmol·L-1 NO3- for 4 d (PRJCA021671). The transcripts of BcAMT1.1 were abundant in both leaves and roots in response to different nitrogen sources. In leaves, BcAMT1.1 transcription was significantly up-regulated under both NH4+ and the mixed nitrogen nutrition compared with NO3- treatment; in roots, its expression was reduced by NH4+ but increased under the mixed treatment (Figure 6B). The transcription of BcAMT1.3 and BcAMT1.5 in leaves was unaffected by nitrogen forms; while in roots, the ones were significantly decreased by NH4+ and only slightly altered by the mixed nitrogen treatment (Figure 6B). In contrast, the transcripts of BcAMT2.1 and BcAMT2.1-like were strongly induced by NH4+ and the mixed nitrogen- nutrition in both roots or leaves, compared with the ones of NO3- (Figure 6B). These results show that the BcAMT gene exhibited a dynamic response to various nitrogen treatments, suggesting that it may play a key role in nitrogen uptake and metabolism.
2.6.3. Response to Nitrogen Deficiency and Different NH4+ Concentration
The qRT-PCR analysis revealed that BcAMT1.1 transcript was markedly induced under nitrogen deficiency. After 72 h of nitrogen starvation, BcAMT1.1 expression increased 6.78-fold in roots and 2.01-fold in leaves compared with the control (Figure 7A,B). When exposed to different NH4+ concentrations, BcAMT1.1 transcripts were significantly up-regulated by lower NH4+ levels, reaching 1.95 ~ 4.10 times that of the control. In contrast, the ones were obviously down-regulated by higher NH4+ concentration, only being 1.25 ~ 1.73 times of the control (Figure 7C,D). Considering the established role of AMT1 subfamily members in high-affinity NH4+ uptake, their inducibility under nitrogen deficiency or low nitrogen conditions, BcAMT1.1 was selected for subsequent functional characterization.
2.7. Subcellular Localization and NH4+ Transport Activity of BcAMT1.1
Transient expression assays indicated that BcAMT1.1 was localized in plasma membrane of onion epidermal cells (Figure 8A). NH4+ transport function of BcAMT1.1 was further examined using the yeast mutant 31019b. Transformants harboring pYES2-BcAMT1.1 grew normally on medium supplied with 2 mmol·L-1 NH4+ (Figure 8B). These findings indicate that BcAMT1.1 can complement the growth defect of the mutant, confirming its potential involvement in ammonium transport and utilization in yeast.
2.8. Overexpressing BcAMT1.1 Promotes NH4+ Uptake and Accelerates Plant Growth of Arabidopsis Under Low NH4+ Concentration
To evaluate the potential role of BcAMT1.1, three independent T4 homozygous Arabidopsis lines were selected to analyze (Figure S1). After pre-cultivation on medium with 4 mmol·L-1 NO₃⁻ for 4 d, seedlings were subsequently transferred into vertical agar plates with 0.25 mmol·L-1 NH₄⁺ for 10 d. Compared with wildtype (WT), overexpressing BcAMT1.1 obviously promoted the growth of Arabidopsis seedlings, with shoots and roots fresh weight increased by 1.32 ~ 1.34 times and 2.04 ~ 2.36 times, respectively. Primary root length was increased by 1.10 ~ 1.16 folds (Figure 9A-C). NH₄⁺ content in transgenic lines was 1.03 ~ 1.13 times higher than that of WT (Figure 9D). Net NH₄⁺ influx rate was measured in the OE-2 line, was increased by 20% relative of WT (Figure 9E).
The physiological relevance of this trait was confirmed by sensitivity testing with methylammonium (MeA), a toxic analog of NH₄⁺. On medium with 20 mmol·L-1 MeA, overexpression lines showed severe growth inhibition, with fresh weight reduced by 35 ~ 48% of WT, accompanied by chlorosis and shorter primary roots (Figure S2). Collectively, these results demonstrate that overexpressing BcAMT1.1 enhances NH₄⁺ uptake capacity under low NH4+ conditions, and promotes plant growth.
2.9. Overexpressing BcAMT1.1 Alters Nitrogen Ion Fluxes and the Expression of Nitrogen Assimilation-Related Genes in Arabidopsis
To further explore the function of BcAMT1.1, the OE-2 line was used to detect the function under a nitrogen mixed nutrition (0.0625 mmol·L-1 NH4+ and 0.1875 mmol·L-1 NO3-). Overexpressing BcAMT1.1 lines exhibited better growth potential than WT, characterized by significantly increased biomass, elongated primary root length, increased lateral roots number and density (Figure 10A-D). Net NH4+ influx in OE-2 was 1.74-fold higher than in WT, whereas NO3- flux shifted from net influx in WT to net efflux in OE-2 (Figure 10E). NO3- content was significantly decreased in transgenic seedlings, whereas NH4+ content remained unchanged (Figure 10F).
Furthermore, we detected the transcript level of key nitrogen assimilation-related genes, such as GLN, GLT, and GDH, encoding GS, GOGAT, and glutamate dehydrogenase. Transcript analysis of nitrogen assimilation-related genes revealed that overexpressing BcAMT1.1 significantly up-regulated AtGLN1;2 transcription, being about 2.60 folds of WT, and significantly down-regulated the expression of AtGLN2 in roots; while having no obvious influences on AtGLN1.1, AtGDH2, and AtGLT1 (Figure 10G). In shoots, the transcription of AtGLN1.1, AtGLN2, and AtGDH2 in overexpression lines were significantly improved by 2.08, 2.16, and 7.31 times, respectively, compared to those of WT (Figure 10H).
2.10. Protein-Protein Interaction (PPI) Network of BcAMT1.1
To further elucidate the potential molecular mechanism of BcAMT1.1, a PPI network was inferred from orthologous genes in Arabidopsis using the STRING database. The analysis identified 46 putative interaction pairs (Figure 11; Table S3). In this network, BcAMT1.1 was predicted to interact with AMT1.3, nitrate transporter (NPF6.3/NRT1.1, NRT2.1 and NRT2.4), GLT1.1, GLN (GLN1.3, GLN1.4, and GLN2), and GLB1, which is a PII protein involved in nitrogen-sensing signal transduction pathway [31]. In addition, BcAMT1.1 may interact with CBL-interacting protein kinase 23 (CIPK23), which forms a complex with the calcineurin B-like protein (CBL), who play a prominent role in activation on plant nutrient transporter [32]. Notably, CIPK23 was also predicted to interact with AMT1.3, NRT2.1, NRT2.4, and NPF6.3. The observations suggest that AMT1.1 may participate in nitrogen absorption and assimilation through interactions with CIPK23 and other key regulators within nitrogen metabolism network.
3. Discussion
AMT proteins are key mediators of NH4+ absorption and transport from rhizosphere to intercellular space, thereby maintaining cellular NH4+ homeostasis [14,33]. Although the AMT gene families have been well characterized in a variety of species, such as Arabidopsis, O. sativa, and B. napus, a comprehensive and systematic identification in flowering Chinese cabbage remains lacking. In this study, nine BcAMT genes were systematically identified and categorized into two phylogenetic subfamilies: AMT1 (7 members) and AMT2 (2 members), consistent with the classification of AtAMTs in Arabidopsis and BnaAMTs in B. napus (Figure 2). Interestingly, while each Arabidopsis AMT has a single homologous copy, most B. napus AMTs occur as 2 ~ 3 copies. In flowering Chinese cabbage, BcAMT1.1, BcAMT1.2 and BcAMT1.5 existed as a single copy, BcAMT1.3, BcAMT1.4 and BcAMT2.1 contained two copies. These homologous genes clustered into the corresponding branches with AMT proteins from other species (Figure 2), consistent with the higher number of duplicated orthologs in Brassica species than Arabidopsis [34], supporting previous evidence for higher levels of duplicated orthologs in Brassica compared with Arabidopsis. Previous study has reported that three polyploidization events in B. rapa: γ triplication (135 MYA), β duplication (90 ~ 100 MYA), and α duplication (24 ~ 40 MYA) [35]. In our study, five of nine BcAMT genes (55.56%) were products of segmental duplication (Figure 4; Table 2), suggesting that the expansion of BcAMTs in flowering Chinese cabbage was primarily driven by segmental duplication.
Polyploidization often leads to the diversification in both structural features and functional domains of genes. In this study, motif analysis revealed that BcAMT1 subfamily members in flowering Chinese cabbage possessed 10 uniform motifs, whereas BcAMT2 members lacked motif 1, motif 2, and motif 9, with BcAMT2.1-like further lacking motif 7 (Figure 3). Similar reductions of conserved motif number in AMT2 subfamily compared with AMT1 have been reported in B. napus [19]. Previous studies have reported that most AMT1 genes lack introns, with exception including LjAMT1.1 in Lotus japonicus [36], SlAMT1.2 in Solanum lycopersicum [37], and MeAMT1.2 in Manihot esculenta [38]. In this study, both BcAMT1.2 and BcAMT1.3-like contained one intron, with the intron length of BcAMT1.3-like exceeding 4500 bp (Figure 3). A similarly long intron structure is found in rapeseed BnaAMT1.3a [19]. All BcAMT2 genes contained four introns and five exons, being consistent with those reported in Arabidopsis [33], B. napus [19], and soybean [20]. It indicates that these AMT2 genes may undergo more complex regulation, i.e., chromatin assembly, mRNA transport, and alternative splicing [39].
In plants, the members of AMT1 subfamily are widely recognized as high-affinity ammonium transporters [11,22], whereas AMT2 members are primarily associated with low-affinity transport [33]. In flowering Chinese cabbage, BcAMTs exhibits distinct expression profiles. BcAMT1.5 is predominantly expressed in roots [40]. In this study, RNA-seq analysis revealed that BcAMT1.1, BcAMT2.1, and BcAMT2.1-like were mainly expressed in roots, stems, and leaves. Notably, BcAMT1.1 was strongly induced under nitrogen-deficient or low nitrogen availability, particularly in roots (Figure 7). It is consistent with previous studies that AtAMT1.1 in Arabidopsis [41], OsAMT1.1 in O. sativa [18], and PsAMT1.1 in Populus simonii [42] are induced under limited nitrogen availability and function as high-affinity ammonium transporters.
In the present study, BcAMT1.1 was located at the plasma membrane and was able to complement the growth defect of yeast mutant 31019b on medium containing 2 mmol·L-1 NH4+. In Arabidopsis, overexpressing BcAMT1.1 significantly promoted plant growth in low nitrogen conditions, being increased both net NH4+ influx and NH4+ content in comparison with the ones of WT. It indicates that BcAMT1.1 encodes a high-affinity ammonium transporter in flowering Chinese cabbage. It is consistent with previous reports for AMT1.1 in Arabidopsis [22,41], Oryza sativa [18,43], and P. simonii [42], as well as with the results of BcAMT1.2 and BcAMT1.5 in B. campestris, which indicate that AMT1 members primarily function in NH4+ absorption and their overexpression lines can enhance NH4+ absorption at low NH4+ conditions. Under the mixed nitrogen nutrition, BcAMT1.1-overexpressing lines enhanced transcription of AtGLN1.1, AtGLN1.2, AtGLN2, AtGDH2, the key genes of GS/GOGAT or GDH pathway, leading to increasing net NH4+ influx without significant changes in NH4+ content in comparison to WT. This is in accordance with our earlier observations, which showed that overexpressing BcAMT1.2 or BcAMT1.5 significantly enhanced the transcription of most nitrogen assimilation-related genes [40,44]. Previous study has further indicated that potential NH4+ toxicity can be mitigated if nitrogen assimilation rates keep pace with the ones of NH4+ uptake [45].
Beyond their role in NH4+ absorption, AMTs may also affect NO3- uptake, owing to that the acquisition of these two nitrogen sources can be synergistically regulated [4]. Our results showed that in BcAMT1.1-overexpressing lines, net NO3- flux shifted from influx in WT to efflux, leading to a marked decrease in NO3- concentration. It aligns with the observations for BcAMT1.2 in flowering Chinese cabbage [44], while contrasts with the results for PsAMT1.1 in P. simonii [42] and BcAMT1.5 in flowering Chinese cabbage [40]. It indicates that AMT members may have divergent functions in modulating the interaction between NH4+ and NO3- uptake [44].
AMTs activities are tightly regulated to adapt to external nitrogen status, with control exerted at both transcriptional and post-transcriptional levels [46,47,48]. Plant nutrient uptake, including both NO3- and NH4+, is often modulated by protein kinase-mediated phosphorylation at post-transcriptional levels [49]. For instance, CIPK23 regulates NO3- uptake by phosphorylating of a threonine in NPF6.3/NRT1.1 [32,50,51]. Similarly, conserved threonine residues in AtAMT1.1 and AtAMT1.2 are targeted for phosphorylation by the CBL1-CIPK23 complex [32,47,49]. In Arabidopsis, phosphorylation of threonine residues T464 and T494 in AtAMT1.3 serves as a key regulatory switch, enabling plant to fine-tune its response to varying nitrogen forms. [48]. In the present study, PPI network indicated that BcAMT1.1 might interact with CIPK23, as well as with other proteins involved in nitrogen uptake and metabolism, including AMT, NRT, GLT, GLN, and GLB proteins (Figure 11). These findings suggest that CIPK23 might contribute to regulating nitrogen uptake and assimilation by modifying phosphorylation residues on AMT, NRT, or other interacting proteins. Further identifying novel upstream regulators of AMT1s will be crucial for elucidating the molecular mechanisms of nitrogen transport [48].
4. Materials and Methods
4.1. Geonmoe-Wide Identification of AMT Genes in Flowering Chinese Cabbage and Chromosome Location
The genome database of flowering Chinese cabbage was obtained from the China National Gene Bank (CNGB) released by Li et al. [34]. Sequences of AtAMT in Arabidopsis were acquired from the Brassicaceae Database (BRAD) (http://brassicadb.cn/, accessed on 12 July 2024), and utilized as query inputs for BLASTP analysis against flowering Chinese cabbage genome (E-value < 1×10-5). To confirm domain composition, candidate sequences were analyzed in the Pfam database (http://pfam.sanger.ac.uk/search, accessed on 12 July 2024) for the presence of the of ammonium transporter family domain (Pfam ID: PF00909). Phyicochemical properties of predicted AMT proteins were predicted using the ExPASY ProtParam (https://web.expasy.org/protparam/, accessed on 12 July 2024), and conserved domains were examined usng the NCBI Batch CD-Search tool (https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi, accessed on 12 July 2024). Nine AMT proteins in flowering Chinese cabbage were finally confirmed as AMT family members. Predictions of subcellular localization were conducted with Deeploc-2.0 (https://services.healthtech.dtu.dk/cgi-bin/webface2.cgi, accessed on 12 July 2024), while transmembrane helices were identified with TMHMM v2.0 (https://services.healthtech.dtu.dk/services/TMHMM-2.0/, accessed on 12 July 2024). Chromosomal localization of AMT genes were analyzed and visualized using TBtools-II v2.345 [52].
4.2. Phylogenetic Tree, Conserved Motifs, Domains, and Gene Structure Analyses of AMT Members
Homologous AMT protein sequences from A. thaliana, B. napus, S. lycopersicum, N. tabacum, O. sativa, and P. trichocarpa were obtained from public database (BRAD, GenBank, UniProt, Genoscope) and literature [18,19,21,22]. A maximum-likelihood (ML) phylogenetic tree was generated based on the multiple sequence alignment of these seven species with 1000 bootstrap replicates, and visualized using Figtree v1.4.4. Sequences alignments were visualized and subfamily-specific conserved signature sequences were identified using Jalview v2.11.2.7 [53]. Conserved motifs of the flowering Chinese cabbage AMTs were performed using MEME Suite (https://meme-suite.org/meme/, assessed on 20 July, 2024), followed by analysis and visualization of domain organization and gene structures with TBtools-II.
4.3. Gene Duplication and Geneome-Wide Synteny Analysis of AMTs
Gene duplication events were analyzed and Ks and Ka parameters were calculated utilizing MCScan X module in TBtools-II. The diveragence rate (λ) was set to 1.5 × 10-8 for B. rapa [54], divergence time (T) was estimated as T = Ks/2λ, and the Ka/Ks ratio was calculated. Comparative synteny between flowering Chinese cabbage, A. thaliana, B. napus, and O. sativa was evaluated and visualized in TBtools-II.
4.4. Identification of Cis-Acting Elements in AMT Promoter Regions
The 2000 bp upstream sequences of AMT genes from flowering Chinese cabbage genome were obtained via TBtools-II, and cis-acting regulatory elements were identified using the PlantCARE database (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/, assessed on 23 August, 2024). Elements were classified according to functional categories, their distribution patterns were visualized using TBtools-II.
4.5. Expression Profiling of BcAMTs
Expression levels of BcAMTs in different tissues (roots, stems, young leaves, mature leaves, old leaves, flowers, and seedpods) were retrieved from RNA-seq data (SRP427920) of flowering Chinese cabbage cultivar ‘49 Caixin’. Expression patterns under different nitrogen forms were obtained from unpublished RNA-seq datasets (PRJCA021671) of cultivar ‘Youlv 501’, After 4 d of nitrogen starvation, seedlings were treated with 1 mmol·L-1 NH4+, 0.5 mmol·L-1 NH4+ and 0.5 mmol·L-1 NO3-, and 1 mmol·L-1 NO3- for 4 d, the roots and leaves of which were collected for RNA sequencing. The values of fragments per kilobase of exon model per million mapped reads (FPKM) were utilized to generate heatmaps with TBtools-II.
Seedlings of flowering Chinese cabbage cultivar ‘Youlv 501’ at three-leaf stage were grown in a modified Hoagland solution with 4 mmol·L-1 NO3- for two weeks. Seedlings were rinsed thoroughly with deionized water and transplanted to a nitrogen-free modified Hoagland solution for nitrogen starvation. Leaves and roots were collected at 0, 24, 48, and 72 h after the initiation of nitrogen deprivation. The remaining seedlings were subsequently exposed to NH4+ at concentrations of 0.1, 1, 4, and 8 mmol·L-1 for 2 h, after which roots and leaves were harvested. Total RNA extraction, first-strand cDNA synthesis, and quantitative real-time PCR (RT-PCR) were conducted according to the method described by Zhu et al. [40].
4.6. Subcellular Localization of BcAMT1.1
Using primers listed in Table S4, the coding sequence (CDS) of BcAMT1.1 without the termination codon was amplified and cloned into the pBI121-GFP vector after the vector was linearized by Sma I and Xba I. The BcAMT1.1-GFP plasmid was transformed into onion epidermal cells via Agrobacterium tumefaciens EHA105. GFP signals were observed using a Zeiss Axio Imager D2 fluorescence microscope.
4.7. Functional Complementation Analysis of BcAMT1.1 in Yeast
Using the ClonExpress II OneStep Cloning Kit (Vazyme Biotech, Nanjing, China), the BcAMT1.1 CDS was inserted into the pYES2 vector, digested by EcoR I and Xba I. Recombinant plasmid and empty vector controls were transformed into yeast mutant strain 31019b (Δmep1, Δmep2, Δmep3, and ura3) via the method of lithium acetate. This strain is unable to grow on medium containing NH4+ concentration below 5 mmol·L-1 as the sole nitrogen source [22]. Transformants were cultured at 30 °C for 3 d on yeast nitrogen base medium (2% galactose) containing 2 mmol·L-1 arginine or NH4+ as the sole nitrogen source.
4.8. Overexpression of BcAMT1.1 in Arabidopsis
The BcAMT1.1 CDS was cloned into pCAMBIA3301 and transformed into Arabidopsis via floral dip using Agrobacterium tumefaciens GV1301 [55]. Transgenic seeds were screened on phosphinothricin and analyzed by qRT-PCR. The T4 generation homozygous lines were used to analyze plant phenotype and physicochemical indices.
Sterilized seeds were germinated on 1/2 Murashige and Skong (MS) agar medium containing 4 mmol·L-1 NO3- for 4 d, then transferred to 1/2 MS plates with 0.25 mmol·L-1 NH4+ for 10 d. Fresh weight, primary root length, and NH4+ content and NH4+ ion flux were measured as described by Zhu et al. [44].
For mixed nitrogen treatments, Arabidopsis seedlings were pre-cultured on 1/2 MS medium supplemented with 4 mmol·L-1 NO3- for 7 d, followed by transfer to 1/2 MS medium with 0.0625 mmol·L-1 NH4+ and 0.1875 mmol·L-1 NO3- for another 7 d. NH4+ and NO3- contents were determined following Ivančič and Degobbis [56]. Gene expression were analyzed via qRT-PCR using TB Green® Premix Ex TaqTM II (TaKaRa Bio, Shiga, Japan), with GADPH as the internal reference. Primers sequences were listed in Table S4. Relative transcript levels were evaluated using the 2-ΔΔCT method [57].
4.9. Prediction of Protein-Protein Interaction Network of BcAMT1.1
A potential protein interaction partners of BcAMT1.1 was predicted using STRING database (https://cn.string-db.org, accessed on 25 August, 2024), with Arabidopsis orthologs used as reference.
4.10. Statistial Analysis
Statistical analyses were conducted using SPSS v21.0 (IBM, Armonk, NY, USA). Differenes among treatments were assessed by Duncan’s multiple range test, with significance defined at P < 0.05 or P < 0.01. In the firgures, distinct lowercase letters or asterisks denote statistically significant differences among treatments.
5. Conclusions
This study presents a comprehensive characterization of the ammonium transporter (AMT) gene family in flowering Chinese cabbage, with emphasis on BcAMT1.1, a plasm membrane-localized isoform. Nine BcAMT genes were identified and categorized into two subgroups according to phylogenetic relationships and conserved domains. Expression profiles revealed that BcAMT1.1 was preferentially expressed in roots, leaves, and stems, and displayed differential transcriptional responses to different nitrogen sources. Notably, BcAMT1.1 was strongly induced under nitrogen starvation or low NH4+ levels, but markedly suppressed at higher NH4+ level. Functional assays demonstrated that overexpressing BcAMT1.1 in Arabidopsis accelerated plant growth under low-nitrogen conditions, increased NH4+ influx, and content, and altered the expression of nitrogen assimilation-related genes compared with WT. Drawing upon these results, we propose a mechanistic model outlining how BcAMT1.1 regulates nitrogen uptake and assimilation (Figure 12). Collectively, these findings advance our understanding of AMT functions in flowering Chinese cabbage, and provide molecular insights for AMT-mediated regulation of nitrogen uptake and assimilation.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Table S1. The sequence information for each AMT protein used for the phylogenetic analysis; Table S2. All gene pairs of three different genomes (Brassica campestris vs Arabidopsis thaliana, and Brassica campestris vs Brassica napus); Table S3: Protein-protein interaction network prediction of BcAMT1.1; Table S4: The primers were used in this study. Figure S1: Identification of qRT-PCR in different BcAMT1.1-overexpressing lines; Figure S2: Growth phenotype and fresh weight of overexpressing BcAMT1.1 on NH4+ toxic analog methylammoium (MeA).
Author Contributions
R.C. and S. S. conceived and designed the project; Y.Z. and L.Z. performed the experiments; Y.Z., Q.Z. and X.H. analyzed the data, Y.Z. and L.Z. wrote the manuscript; A.A., W.S., and S.S. reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32202465), the Natural Science Foundation of Guangdong Province, China (2023A1515012687), the Guangdong Provincial Special Fund for Modern Agriculture Industry Technology Innovation Teams (2024CXTD01), and the China Agriculture Research System (CARS-23-B11).
Data Availability Statement
All datasets supporting this work are included within the article and Supplementary Materials.
Acknowledgments
We thank Bruno André from the Université Libre de Bruxelles for kindly providing the yeast mutant strain 31019b. We also thank the Guangxi First-class Disciplines (Agricultural Resources and Environment) for suggestions of this paper.
Conflicts of Interest
The authors declared no conflict of interest.
References
- Kindred, D.R.; Verhoeven, T.M.O.; Weightman, R.M.; Swanston, J.S.; Agu, R.C.; Brosnan, J.M.; Sylvester-Bradley, R. Effects of variety and fertiliser nitrogen on alcohol yield, grain yield, starch and protein content, and protein composition of winter wheat. J. Cereal. Sci. 2008, 48, 46–57. [Google Scholar] [CrossRef]
- Tei, F.; De Neve, S.; de Haan, J.; Kristensen, H.L. Nitrogen management of vegetable crops. Agr. Water Manag. 2020, 240, 106316. [Google Scholar] [CrossRef]
- Valenzuela, H. Optimizing the nitrogen use efficiency in vegetable crops. Nitrogen 2024, 5, 106–143. [Google Scholar] [CrossRef]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef]
- Dechorgnat, J.; Nguyen, C.T.; Armengaud, P.; Jossier, M.; Diatloff, E.; Filleur, S.; Daniel-Vedele, F. From the soil to the seeds: the long journey of nitrate in plants. J. Exp. Bot. 2011, 62, 1349–1359. [Google Scholar] [CrossRef]
- Li, S.; Yan, L.; Zhang, W.; Yi, C.; Haider, S.; Wang, C.; Liu, Y.; Shi, L.; Ding, G. Nitrate alleviates ammonium toxicity in Brassica napus by coordinating rhizosphere and cell pH and ammonium assimilation. Plant J. 2024, 117, 786–804. [Google Scholar] [CrossRef]
- Bloom, A.J.; Burger, M.; Asensio, J.S.R.; Cousins, A.B. Carbon dioxide enrichment inhibits nitrate assimilation in wheat and Arabidopsis. Science 2010, 328, 899–903. [Google Scholar] [CrossRef]
- Liu, X.; Hu, B.; Chu, C. Nitrogen assimilation in plants: current status and future prospects. J. Genet. Genomics. 2022, 49, 394–404. [Google Scholar] [CrossRef]
- Omari Alzahrani, F. Ammonium transporter 1 (AMT1) gene family in pomegranate: genome-wide analysis and expression profiles in response to salt stress. Curr. Issues. Mol. Biol. 2025, 47, 59. [Google Scholar] [CrossRef]
- Couturier, J.; Montanini, B.; Martin, F.; Brun, A.; Blaudez, D.; Chalot, M. The expanded family of ammonium transporters in the perennial poplar plant. New. Phytol. 2007, 174, 137–150. [Google Scholar] [CrossRef]
- Williamson, G.; Harris, T.; Bizior, A.; Hoskisson, P.A.; Pritchard, L.; Javelle, A. Biological ammonium transporters: evolution and diversification. FEBS J. 2024, 291, 3786–3810. [Google Scholar] [CrossRef]
- Glass, A.D.M.; Britto, D.T.; Kaiser, B.N.; Kinghorn, J.R.; Kronzucker, H.J.; Kumar, A.; Okamoto, M.; Rawat, S.; Siddiqi, M.Y.; Unkles, S.E.; Vidmar, J.J. The regulation of nitrate and ammonium transport systems in plants. J. Exp. Bot. 2002, 53, 855–864. [Google Scholar] [CrossRef]
- McDonald, T.R.; Ward, J.M. Evolution of electrogenic ammonium transporters (AMTs). Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Ludewig, U.; Neuhäuser, B.; Dynowski, M. Molecular mechanisms of ammonium transport and accumulation in plants. Febs. Lett. 2007, 581, 2301–2308. [Google Scholar] [CrossRef]
- Sohlenkamp, C.; Shelden, M.; Howitt, S.; Udvardi, M. Characterization of Arabidopsis AtAMT2, a novel ammonium transporter in plants. Febs. Lett. 2000, 467, 273–278. [Google Scholar] [CrossRef]
- Ninnemann, O.; Jauniaux, J.C.; Frommer, W.B. Identification of a high affinity NH4+ transporter from plants. EMBO J. 1994, 13, 3464–3471. [Google Scholar] [CrossRef]
- Li, B.; Merrick, M.; Li, S.; Li, H.; Zhu, S.; Shi, W.; Su, Y. Molecular basis and regulation of ammonium transporter in rice. Rice Sci. 2009, 16, 314–322. [Google Scholar] [CrossRef]
- Konishi, N.; Ma, J.F. Three polarly localized ammonium transporter 1 members are cooperatively responsible for ammonium uptake in rice under low ammonium condition. New Phytol. 2021, 232, 1778–1792. [Google Scholar] [CrossRef]
- Dai, J.; Han, P.; Walk, T.C.; Yang, L.; Chen, L.; Li, Y.; Gu, C.; Liao, X.; Qin, L. Genome-wide identification and characterization of ammonium transporter (AMT) genes in rapeseed (Brassica napus L.). Genes 2023, 14, 658. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Dong, X.; Yuan, Z.; Li, X.; Wang, Y. Genome-wide identification and expression analysis of the ammonium transporter family genes in soybean. Int. J. Mol. Sci. 2023, 24, 3991. [Google Scholar] [CrossRef]
- Liu, L.; Fan, T.; Shi, D.; Li, C.; He, M.; Chen, Y.; Zhang, L.; Yang, C.; Cheng, X.; Chen, X.; Li, D.; Sun, Y. Coding-sequence identification and transcriptional profiling of nine AMTs and four NRTs from tobacco revealed their differential regulation by developmental stages, nitrogen nutrition, and photoperiod. Front. Plant Sci. 2018, 9, 210. [Google Scholar] [CrossRef]
- Yuan, L.; Loqué, D.; Kojima, S.; Rauch, S.; Ishiyama, K.; Inoue, E.; Takahashi, H.; von Wirén, N. The organization of high-affinity ammonium uptake in Arabidopsis roots depends on the spatial arrangement and biochemical properties of AMT1-type transporters. Plant Cell 2007, 19, 2636–2652. [Google Scholar] [CrossRef]
- Sohlenkamp, C.; Wood, C.C.; Roeb, G.W.; Udvardi, M.K. Characterization of Arabidopsis AtAMT2, a high-affinity ammonium transporter of the plasma membrane. Plant Physiol. 2002, 130, 1788–1796. [Google Scholar] [CrossRef]
- Yuan, L.; Graff, L.; Loqué, D.; Kojima, S.; Tsuchiya, Y.N.; Takahashi, H.; von Wirén, N. AtAMT1;4, a Pollen-specific high-affinity ammonium transporter of the plasma membrane in Arabidopsis. Plant Cell Physiol. 2009, 50, 13–25. [Google Scholar] [CrossRef]
- Li, C.; Tang, Z.; Wei, J.; et al. The OsAMT1.1 gene functions in ammonium uptake and ammonium-potassium homeostasis over low and high ammonium concentration ranges. J. Genet. Genomics 2016, 43, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, G.; Zhu, Z.; Hu, M.; Chen, M.; Wang, J.; Zhang, K.; Zheng, Y.; Liao, Y.; Chen, C. Genomic selection and genetic architecture of agronomic traits during modern flowering Chinese cabbage breeding. Hortic Res. 2025, 12, uhae299. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; He, L.; Chen, Y.; Liu, Y.; Liu, J.; Chen, R.; Zhu, Y.; Song, S.; He, Q. Genome-wide identification of SWEET gene family and functional analysis of BcSWEET1-2 associated with flowering in flowering Chinese cabbage (Brassica campestris). BMC Genomics 2025, 26, 605. [Google Scholar] [CrossRef]
- Zhang, S.; Li, G.; Wang, Y.; Anwar, A.; He, B.; Zhang, J.; Chen, C.; Hao, Y.; Chen, R.; Song, S. Genome-wide identification of BcGRF genes in flowering Chinese cabbage and preliminary functional analysis of BcGRF8 in nitrogen metabolism. Front. Plant Sci. 2023, 14, 1144748. [Google Scholar] [CrossRef]
- Zhu, Y.; Qi, B.; Hao, Y.; Liu, H.; Sun, G.; Chen, R.; Song, S. Appropriate NH4+/NO3– ratio triggers plant growth and nutrient uptake of flowering Chinese cabbage by optimizing the pH value of nutrient solution. Front. Plant Sci. 2021, 12, 656144. [Google Scholar] [CrossRef]
- Song, S.; Yi, L.; Zhu, Y.; Liu, H.; Sun, G.; Chen, R. Effects of ammonium and nitrate ratios on plant growth, nitrate concentration and nutrient uptake in flowering Chinese cabbage. Bangl. J. Bot. 2017, 46, 1259–1267. [Google Scholar]
- Hsieh, M.H.; Lam, H.M.; van de Loo, F.J.; Coruzzi, G. A PII-like protein in Arabidopsis: putative role in nitrogen sensing. PNAS 1998, 95, 13965–13970. [Google Scholar] [CrossRef]
- Ródenas, R.; Vert, G. Regulation of root nutrient transporters by CIPK23: ‘One kinase to rule them all’. Plant Cell Physiol. 2021, 62, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Giehl, R.; Laginha, A.M.; Duan, F.; Rentsch, D.; Yuan, L.; von Wirén, N. A critical role of AMT2;1 in root-to-shoot translocation of ammonium in Arabidopsis. Mol. Plant. 2017, 10, 1449–1460. [Google Scholar] [CrossRef]
- Li, G.; Jiang, D.; Wang, J.; Liao, Y.; Zhang, T.; Zhang, H.; Dai, X.; Ren, H.; Chen, C.; Zheng, Y. A high-continuity genome assembly of Chinese flowering cabbage (Brassica rapa var. parachinensis) provides new insights into Brassica genome structure evolution. Plants 2023, 12, 2498. [Google Scholar] [CrossRef]
- Wang, X.W.; Kole, C. The Brassica rapa genome. In Compendium of Plant Genomes; Chittaranjan, K., Mohanpur, W.B., Eds.; Springer-Verlag GmbH Berlin Heidelberg: Berlin, Germany, 2015; pp. 1–165. [Google Scholar] [CrossRef]
- Salvemini, F.; Marini, A.; Riccio, A.; Patriarca, E.J.; Chiurazzi, M. Functional characterization of an ammonium transporter gene from Lotus japonicus. Gene 2001, 270, 237–243. [Google Scholar] [CrossRef]
- Filiz, E.; Akbudak, M.A. Ammonium transporter 1 (AMT1) gene family in tomato (Solanum lycopersicum L.) bioinformatics, physiological and expression analyses under drought and salt stresses. Genomics 2020, 112, 3773–3782. [Google Scholar] [CrossRef]
- Xia, Y.; Liu, Y.; Zhang, T.; Wang, Y.; Jiang, X.; Zhou, Y. Genome-wide identification and expression analysis of ammonium transporter 1 (AMT1) gene family in cassava (Manihot esculenta Crantz) and functional analysis of MeAMT1;1 in transgenic Arabidopsis. 3 Biotech 2022, 12, 4. [Google Scholar] [CrossRef]
- Jo, B.; Choi, S.S. Introns: The Functional Benefits of Introns in Genomes. Genomics Inform. 2015, 13, 112. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhong, L.; Huang, X.; Su, W.; Liu, H.; Sun, G.; Song, S.; Chen, R. BcAMT1;5 mediates nitrogen uptake and assimilation in flowering Chinese cabbage and improves plant growth when overexpressed in Arabidopsis. Horticulturae 2023, 9, 43. [Google Scholar] [CrossRef]
- Gazzarrini, S.; Lejay, L.; Gojon, A.; Ninnemann, O.; Frommer, W.B.; von Wirén, N. Three functional transporters for constitutive, diurnally regulated, and starvation-induced uptake of ammonium into Arabidopsis roots. Plant Cell 1999, 11, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, J.; Zhuang, S.; Feng, Z.; Fan, J. Functional characterization of PsAMT1.1 from Populus simonii in ammonium transport and its role in nitrogen uptake and metabolism. Environ. Exp. Bot. 2023, 208, 105255. [Google Scholar] [CrossRef]
- Ranathunge, K.; El-kereamy, A.; Gidda, S.; Bi, Y.; Rothstein, S.J. AMT1;1 transgenic rice plants with enhanced NH4+ permeability show superior growth and higher yield under optimal and suboptimal NH4+ conditions. J. Exp. Bot. 2014, 65, 965–979. [Google Scholar] [CrossRef]
- Zhu, Y.; Huang, X.; Hao, Y.; Su, W.; Liu, H.; Sun, G.; Chen, R.; Song, S. Ammonium transporter (BcAMT1.2 mediates the interaction of ammonium and nitrate in Brassica campestris. Front. Plant Sci. 2020, 10, 1776. [Google Scholar] [CrossRef]
- The, S.V.; Snyder, R.; Tegeder, M. Targeting nitrogen metabolism and transport processes to improve plant nitrogen use efficiency. Front. Plant Sci. 2021, 11, 628366. [Google Scholar] [CrossRef]
- Yuan, L.; Loqué, D.; Ye, F.; Frommer, W.B.; von Wirén, N. Nitrogen-dependent posttranscriptional regulation of the ammonium transporter AtAMT1;1. Plant Physiol. 2007, 143, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Straub, T.; Ludewig, U.; Neuhäuser, B. The kinase CIPK23 inhibits ammonium transport in Arabidopsis thaliana. Plant Cell 2017, 29, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, T.; Zhang, Y.; Duan, F.; Neuhäuser, B.; Ludewig, U.; Schulze, W.X.; Yuan, L. Ammonium and nitrate regulate NH4+ uptake activity of Arabidopsis ammonium transporter AtAMT1;3 via phosphorylation at multiple C-terminal sites. J. Exp. Bot. 2019, 70, 4919–4930. [Google Scholar] [CrossRef]
- Wang, T.; Chen, X.; Ju, C.; Wang, C. Calcium signaling in plant mineral nutrition: From uptake to transport. Plant Commun. 2023, 4, 100678. [Google Scholar] [CrossRef]
- Wang, W.; Hu, B.; Li, A.; Chu, C. NRT1.1s in plants: functions beyond nitrate transport. J. Exp. Bot. 2020, 71, 4373–4379. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.; Lin, S.; Hu, H.; Tsay, Y. CHL1 Functions as a Nitrate Sensor in Plants. Cell 2009, 138, 1184–1194. [Google Scholar] [CrossRef]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y.; Xia, R. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Procter, J.B.; Carstairs, G.M.; Soares, B.; Mourão, K.; Ofoegbu, T.C.; Barton, D.; Lui, L.; Menard, A.; Sherstnev, N.; Roldan-Martinez, D.; Duce, S.; Martin, D.M.A.; Barton, G.J. Alignment of Biological Sequences with Jalview. In Methods in Molecular Biology; Katoh, K., Ed.; Humana: New York, NY, USA, 2021; Volume 2231, pp. 203–224. [Google Scholar] [CrossRef]
- Yin, X.; Wang, Q.; Chen, Q.; Xiang, N.; Yang, Y.; Yang, Y. Genome-wide identification and functional analysis of the calcineurin B-like protein and calcineurin B-like protein-interacting protein kinase gene families in turnip (Brassica rapa var. rapa). Front. Plant Sci. 2017, 8, 1191. [Google Scholar] [CrossRef]
- Wiktorek-Smagur, A.; Hnatuszko-Konka, K.; Kononowicz, A.K. Flower bud dipping or vacuum infiltration—two methods of Arabidopsis thaliana transformation. Russ. J. Plant Physiol. 2009, 56, 560–568. [Google Scholar] [CrossRef]
- Ivančič, I.; Degobbis, D. An optimal manual procedure for ammonia analysis in natural waters by the indophenol blue method. Water Res. 1984, 18, 1143–1147. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chromosomal location of BcAMT genes in flowering Chinese cabbage. Bars represent individual chromosomes, with color gradients indicating gene density along each chromosome. Physical gene positions are marked, and scale bar denotes relative chromosome length in megabase (Mb) units.
Figure 1.
Chromosomal location of BcAMT genes in flowering Chinese cabbage. Bars represent individual chromosomes, with color gradients indicating gene density along each chromosome. Physical gene positions are marked, and scale bar denotes relative chromosome length in megabase (Mb) units.
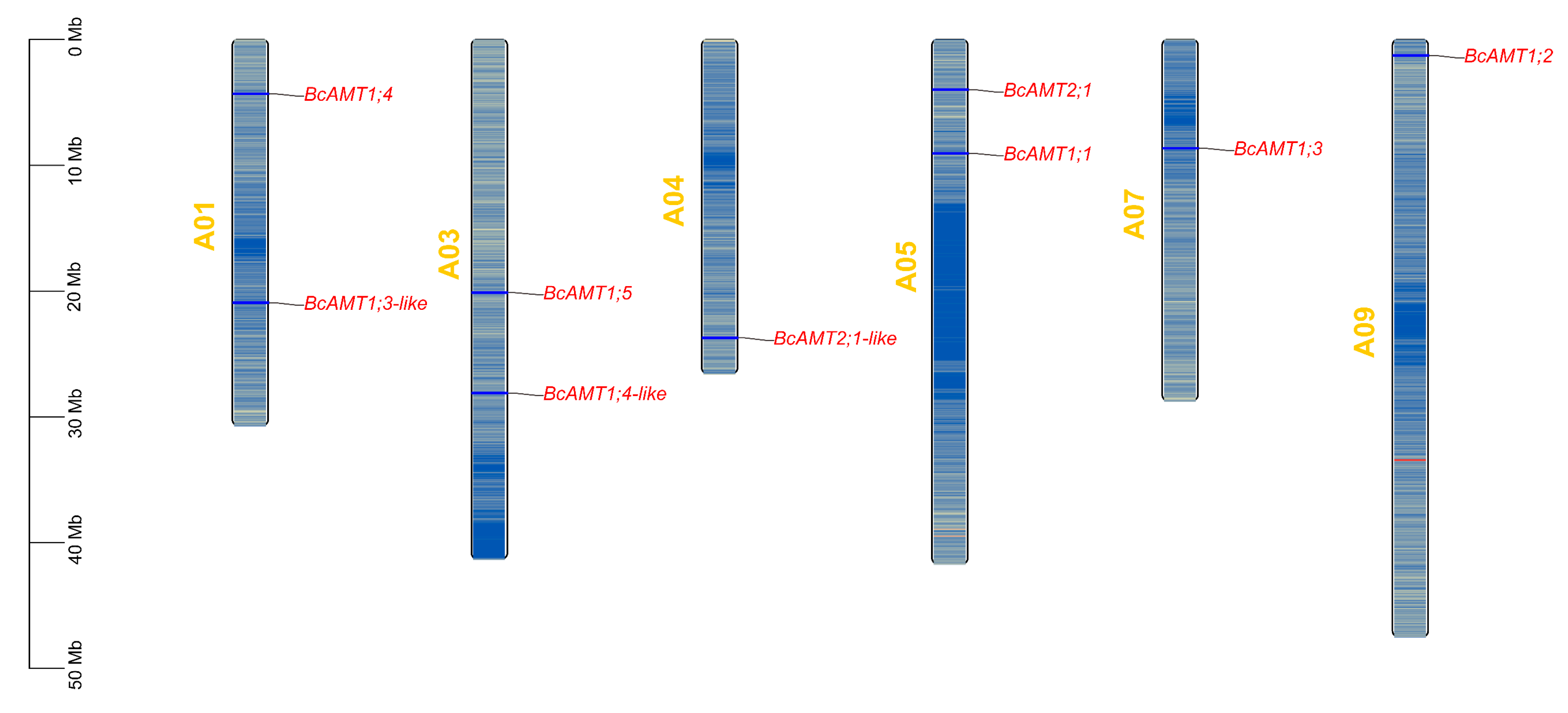
Figure 2.
Phylogenetic relationships and conserved signature sequences of AMTs. (A) Phylogenetic tree of AMTs was constructed using MEGA 7.0, with bootstrap values calculated from 1000 replications. At: Arabidopsis thaliana; Sl: Solanum lycopersicum; Nt: Nicotiana tabacum; Os: Oryza sativa; Ptr: Populus trichocarpa; Bn: Brassica napus; Bc: Brassica campestris. (B) Conserved signature sequences of the AMT1 subfamily. (C) Conserved signature sequences of the AMT2 subfamily.
Figure 2.
Phylogenetic relationships and conserved signature sequences of AMTs. (A) Phylogenetic tree of AMTs was constructed using MEGA 7.0, with bootstrap values calculated from 1000 replications. At: Arabidopsis thaliana; Sl: Solanum lycopersicum; Nt: Nicotiana tabacum; Os: Oryza sativa; Ptr: Populus trichocarpa; Bn: Brassica napus; Bc: Brassica campestris. (B) Conserved signature sequences of the AMT1 subfamily. (C) Conserved signature sequences of the AMT2 subfamily.
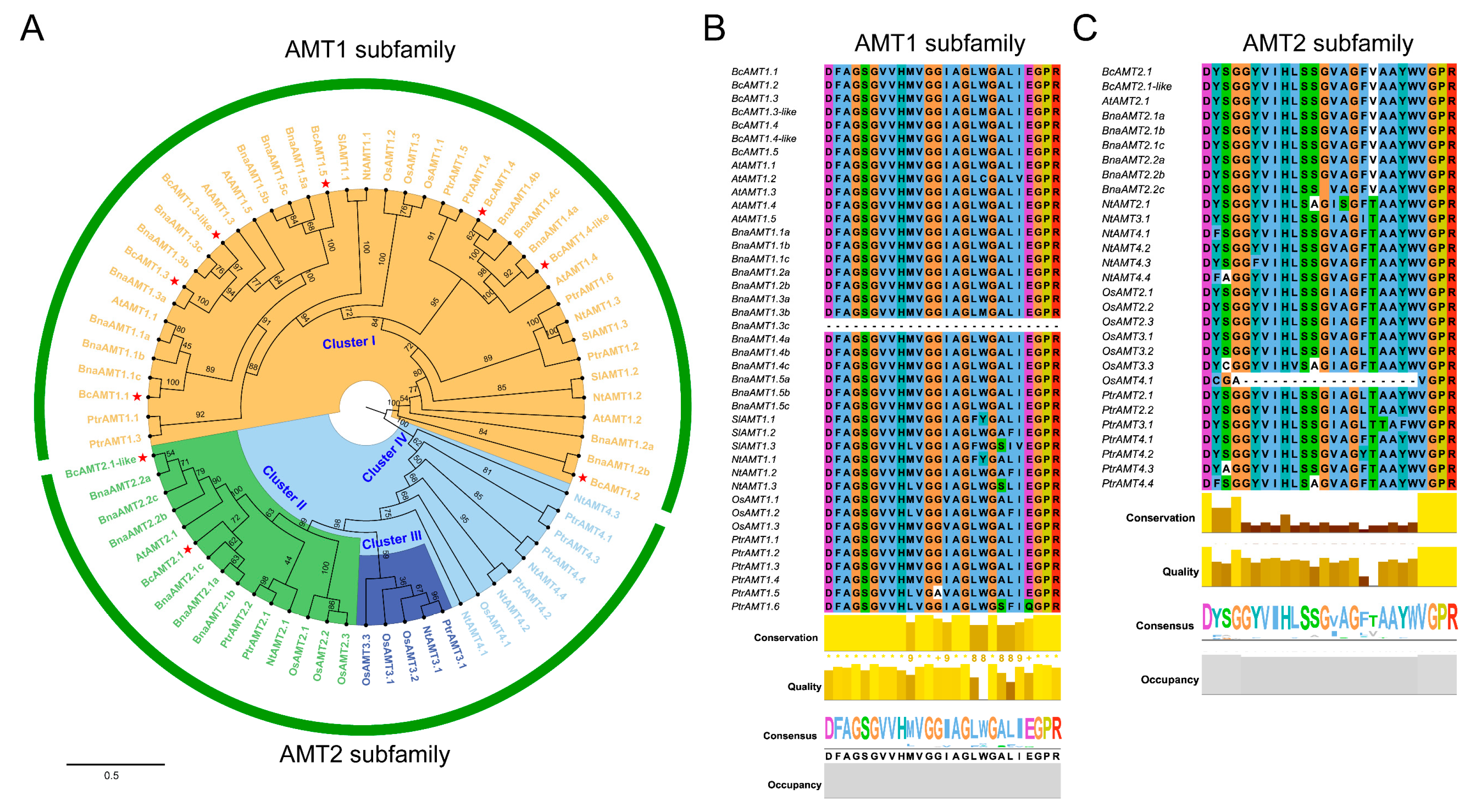
Figure 3.
Conserved motif composition, domain architecture, and gene structure of BcAMTs. (A) Phylogenetic tree and motifs distribution. (B) Conserved domain analysis. (C) Gene structure analysis.
Figure 3.
Conserved motif composition, domain architecture, and gene structure of BcAMTs. (A) Phylogenetic tree and motifs distribution. (B) Conserved domain analysis. (C) Gene structure analysis.
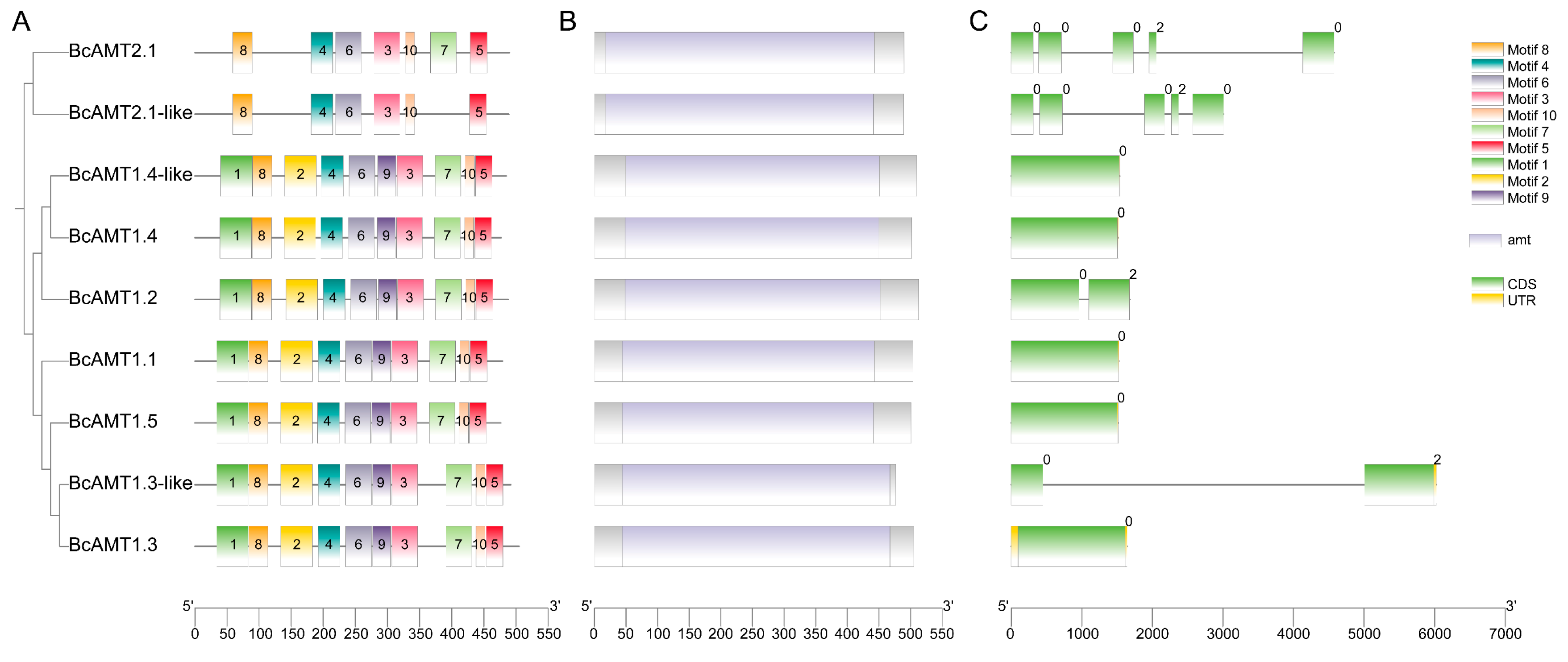
Figure 4.
Collinearity patterns of the AMTs gene family. (A) Collinearity relationships of AMTs in flowering Chinese cabbage. (B) Comparative synteny among A. thaliana, B. napus, and flowering Chinese cabbage.
Figure 4.
Collinearity patterns of the AMTs gene family. (A) Collinearity relationships of AMTs in flowering Chinese cabbage. (B) Comparative synteny among A. thaliana, B. napus, and flowering Chinese cabbage.
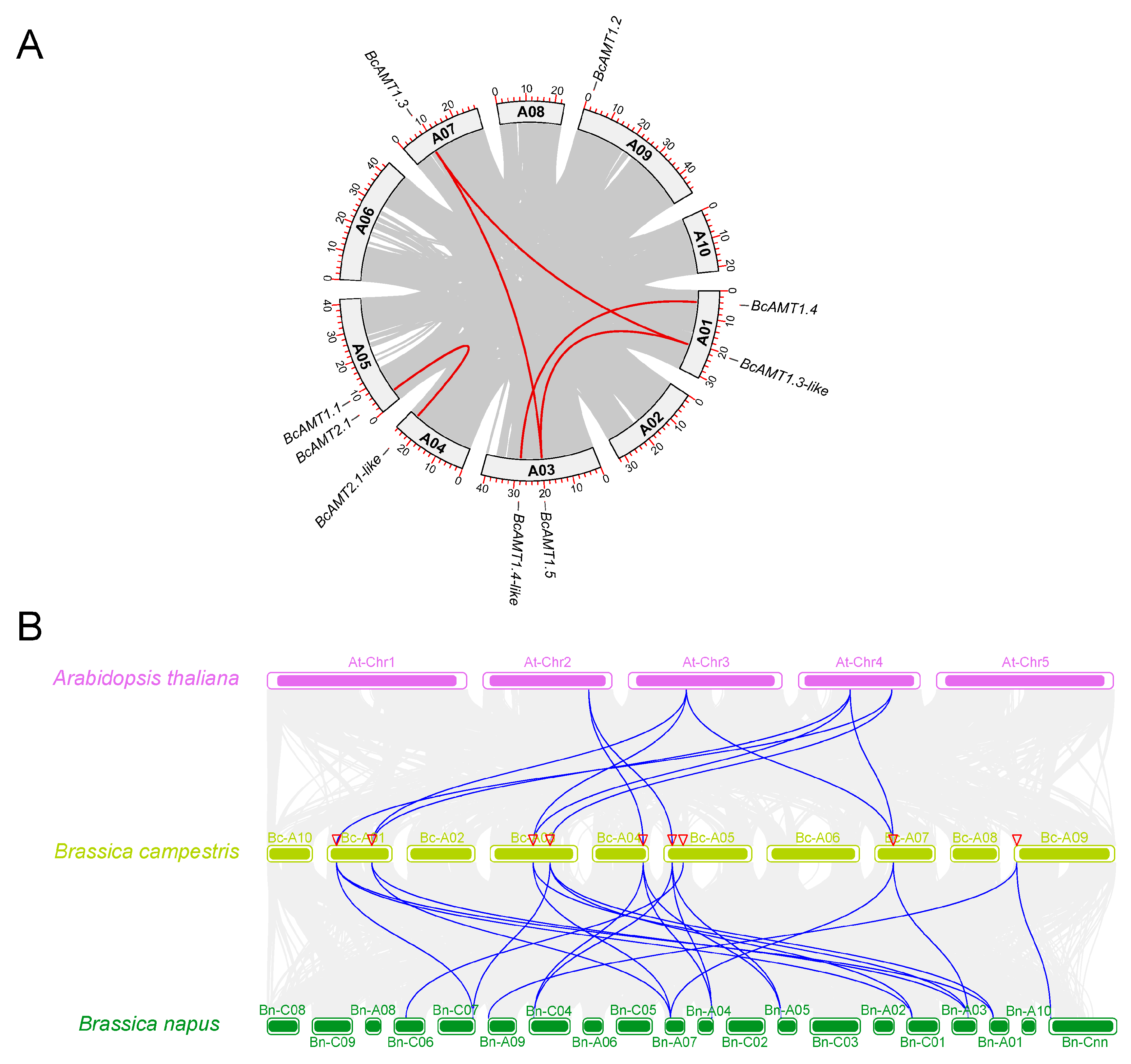
Figure 5.
Analysis of cis-acting elements in BcAMTs promoters. (A) Distribution of cis-acting elements. (B) Number of each cis-acting element type.
Figure 5.
Analysis of cis-acting elements in BcAMTs promoters. (A) Distribution of cis-acting elements. (B) Number of each cis-acting element type.
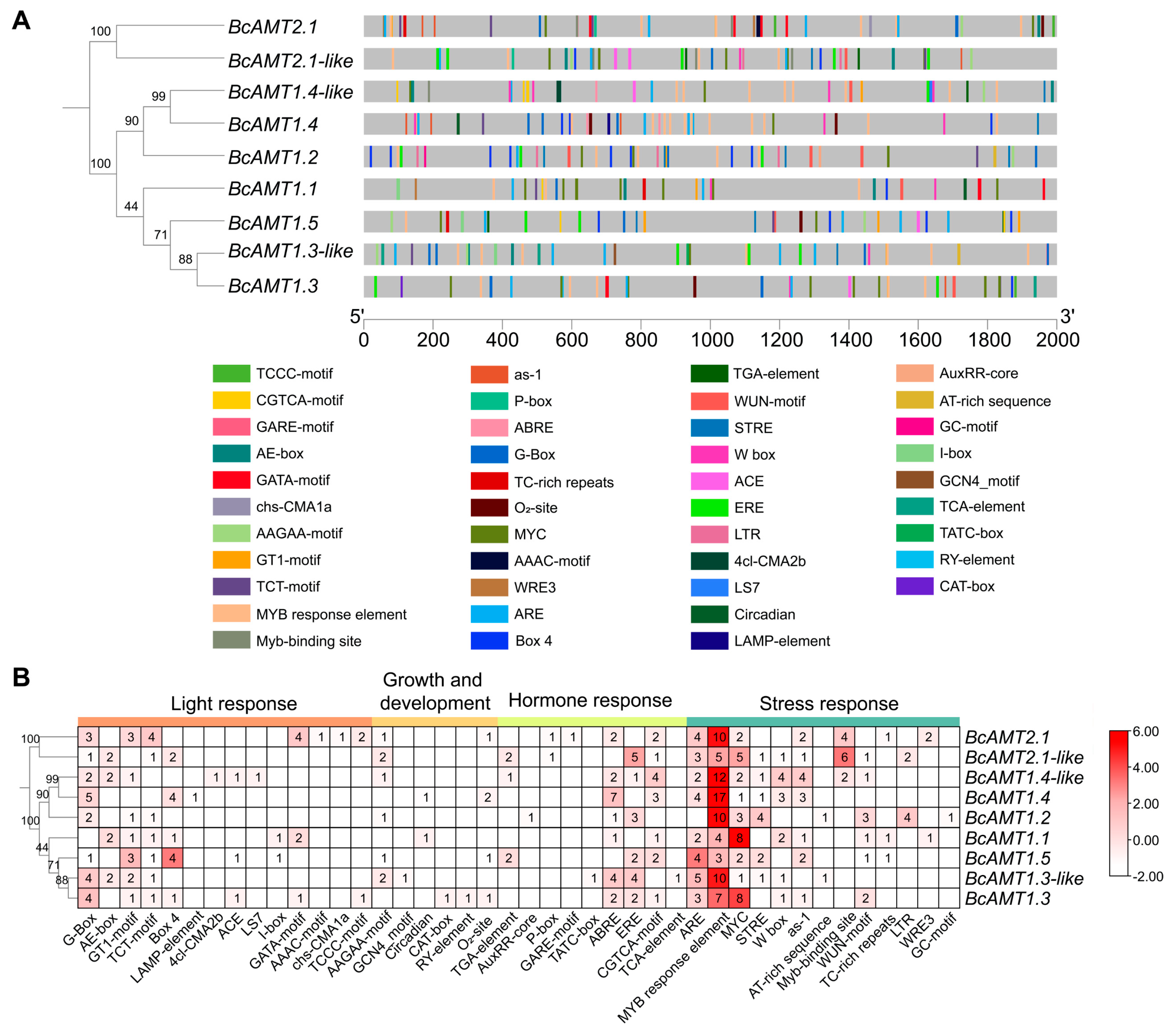
Figure 6.
Expression characteristics of BcAMTs in flowering Chinese cabbage. (A) Expression in different tissues. (B) Expression under different nitrogen regimes. AL and AR: leaf and root in 1 mmol·L-1 NH4+; NAL and NAR: leaf and root in 0.5 mmol·L-1 NH4+ and 0.5 mmol·L-1 NO3-; NL and NR: leaf and root in 1 mmol·L-1 NO3-. Heatmap represents normalized FPKM and values in rectangle represent raw data.
Figure 6.
Expression characteristics of BcAMTs in flowering Chinese cabbage. (A) Expression in different tissues. (B) Expression under different nitrogen regimes. AL and AR: leaf and root in 1 mmol·L-1 NH4+; NAL and NAR: leaf and root in 0.5 mmol·L-1 NH4+ and 0.5 mmol·L-1 NO3-; NL and NR: leaf and root in 1 mmol·L-1 NO3-. Heatmap represents normalized FPKM and values in rectangle represent raw data.
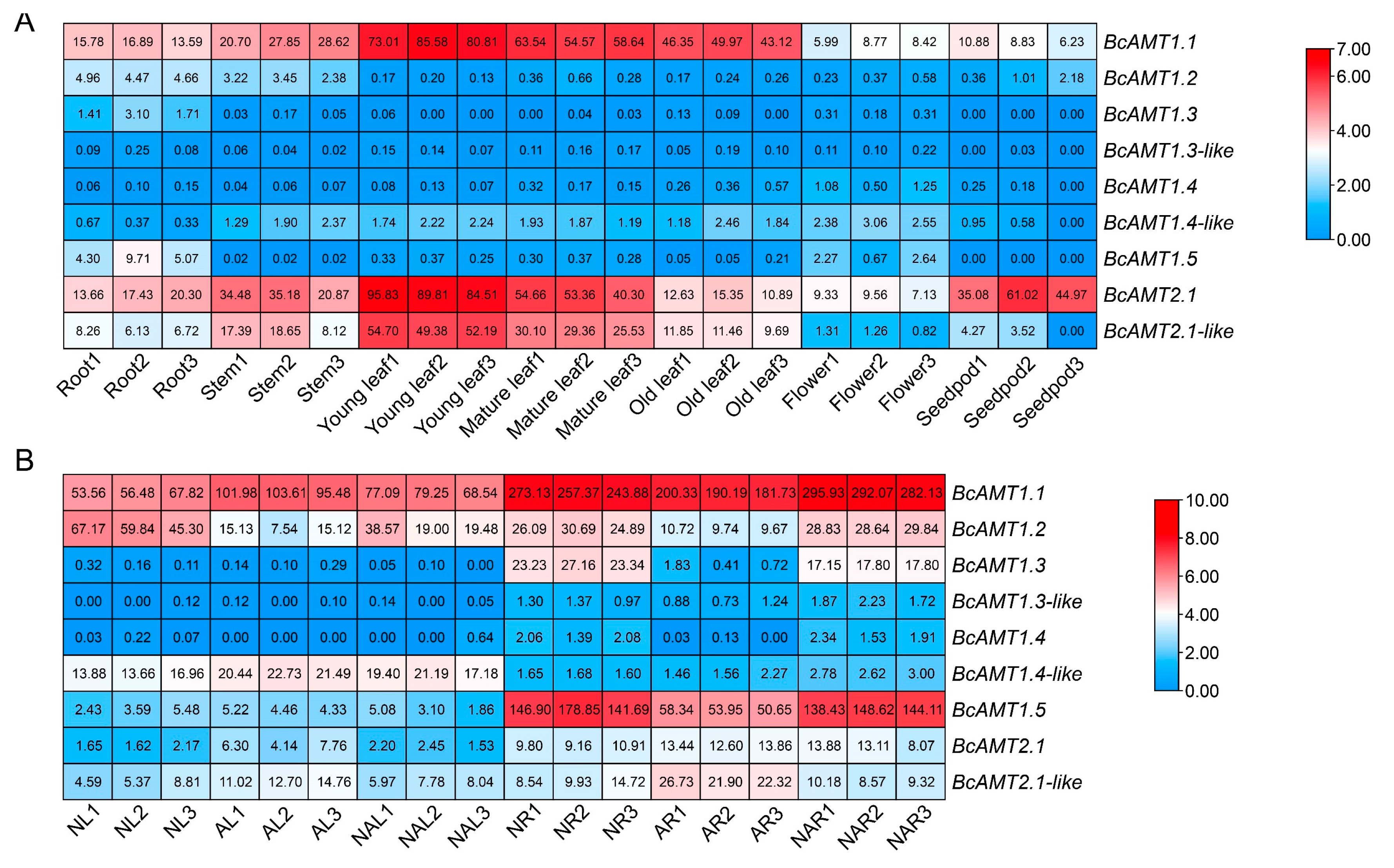
Figure 7.
BcAMT1.1 expression patterns in flowering Chinese cabbage determined by qRT-PCR under different nitrogen regimes. After cultivation for two weeks at 4 mmol·L-1 NO3-, seedlings of flowering Chinese cabbage were subjected to different nitrogen treatments. (A, B) BcAMT1.1 expression in roots and leaves under nitrogen deficiency for 0, 24, 48, and 72 h. (C, D) BcAMT1.1 expression in roots and leaves subjected to different NH4+ concentrations for 2 h. Different lowercase letters present the differences at the level of 0.05.
Figure 7.
BcAMT1.1 expression patterns in flowering Chinese cabbage determined by qRT-PCR under different nitrogen regimes. After cultivation for two weeks at 4 mmol·L-1 NO3-, seedlings of flowering Chinese cabbage were subjected to different nitrogen treatments. (A, B) BcAMT1.1 expression in roots and leaves under nitrogen deficiency for 0, 24, 48, and 72 h. (C, D) BcAMT1.1 expression in roots and leaves subjected to different NH4+ concentrations for 2 h. Different lowercase letters present the differences at the level of 0.05.
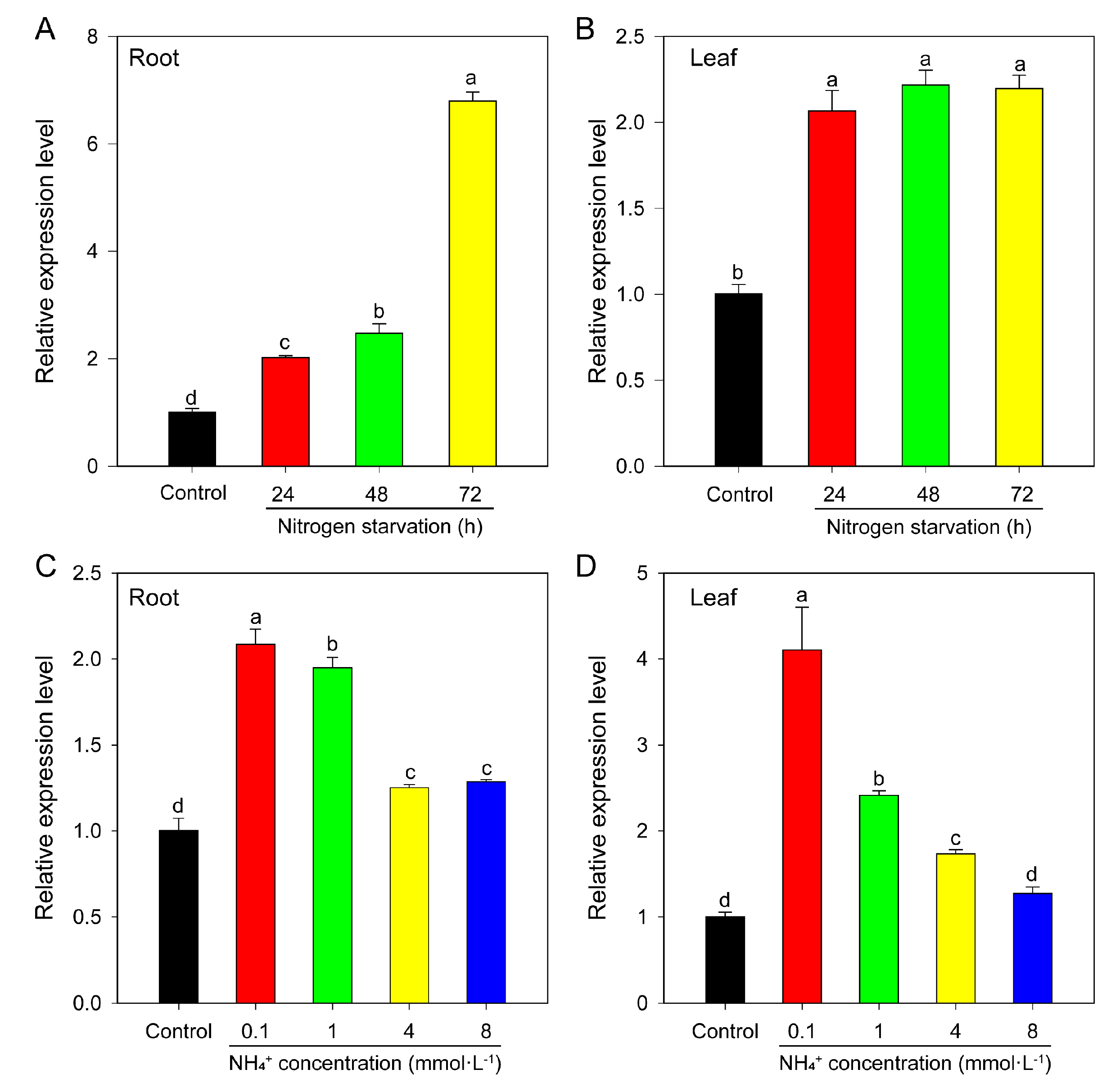
Figure 8.
Subcellular localization and NH4+ transport activity of BcAMT1.1. (A) Subcellular localization of BcAMT1.1. Scale bar is 50 μm. (B) Yeast complementation assay of BcAMT1.1 in mutant strain 31019b.
Figure 8.
Subcellular localization and NH4+ transport activity of BcAMT1.1. (A) Subcellular localization of BcAMT1.1. Scale bar is 50 μm. (B) Yeast complementation assay of BcAMT1.1 in mutant strain 31019b.
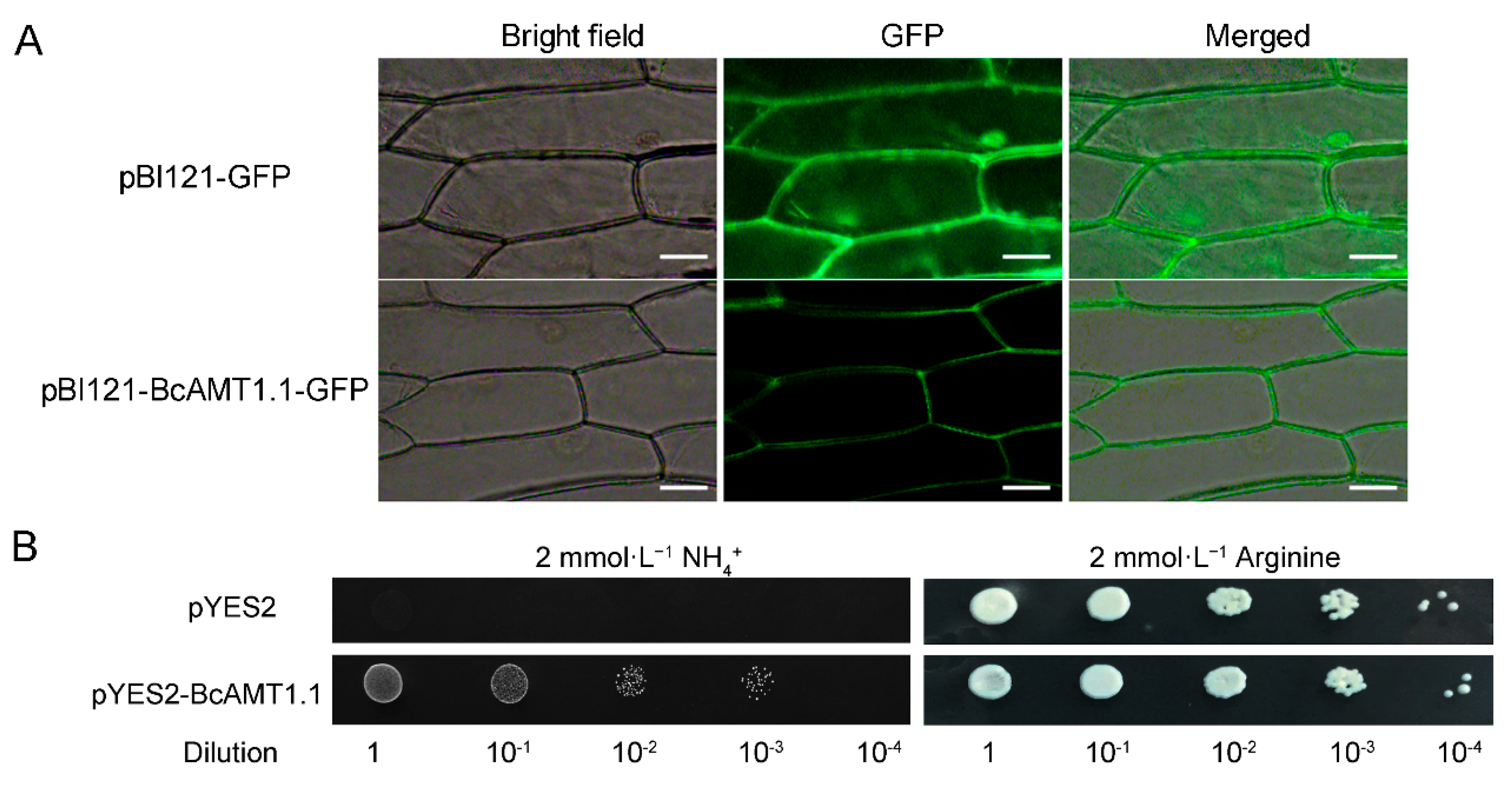
Figure 9.
Phenotype and physiological traits of BcAMT1.1-overexpressing Arabidopsis under 0.25 mmol·L-1 NH4+. (A) Growth phenotype of wild type (WT), vector control and overexpressing BcAMT1.1 lines. After precultured on solid medium contained 4 mmol·L-1 NO3- for 4 d, seedlings were transferred to 0.25 mmol·L-1 NH4+ for 10 d. (B) Fresh weight of whole pants. (C) Primary root length. (D) NH4+ content of whole plants. (E) NH4+ ion flux in root surface. WT: wild type; vector: overexpressing pCAMBIA 3301 vector; OE-2, OE-3, an OE-6: overexpression lines 2, 3, and 6. Line chart shows the real-time change of NH4+ ion at root surface during 5 min; small histogram presents net NH4+ change. Lowercase letters or asterisks denote significant difference (P < 0.05).
Figure 9.
Phenotype and physiological traits of BcAMT1.1-overexpressing Arabidopsis under 0.25 mmol·L-1 NH4+. (A) Growth phenotype of wild type (WT), vector control and overexpressing BcAMT1.1 lines. After precultured on solid medium contained 4 mmol·L-1 NO3- for 4 d, seedlings were transferred to 0.25 mmol·L-1 NH4+ for 10 d. (B) Fresh weight of whole pants. (C) Primary root length. (D) NH4+ content of whole plants. (E) NH4+ ion flux in root surface. WT: wild type; vector: overexpressing pCAMBIA 3301 vector; OE-2, OE-3, an OE-6: overexpression lines 2, 3, and 6. Line chart shows the real-time change of NH4+ ion at root surface during 5 min; small histogram presents net NH4+ change. Lowercase letters or asterisks denote significant difference (P < 0.05).
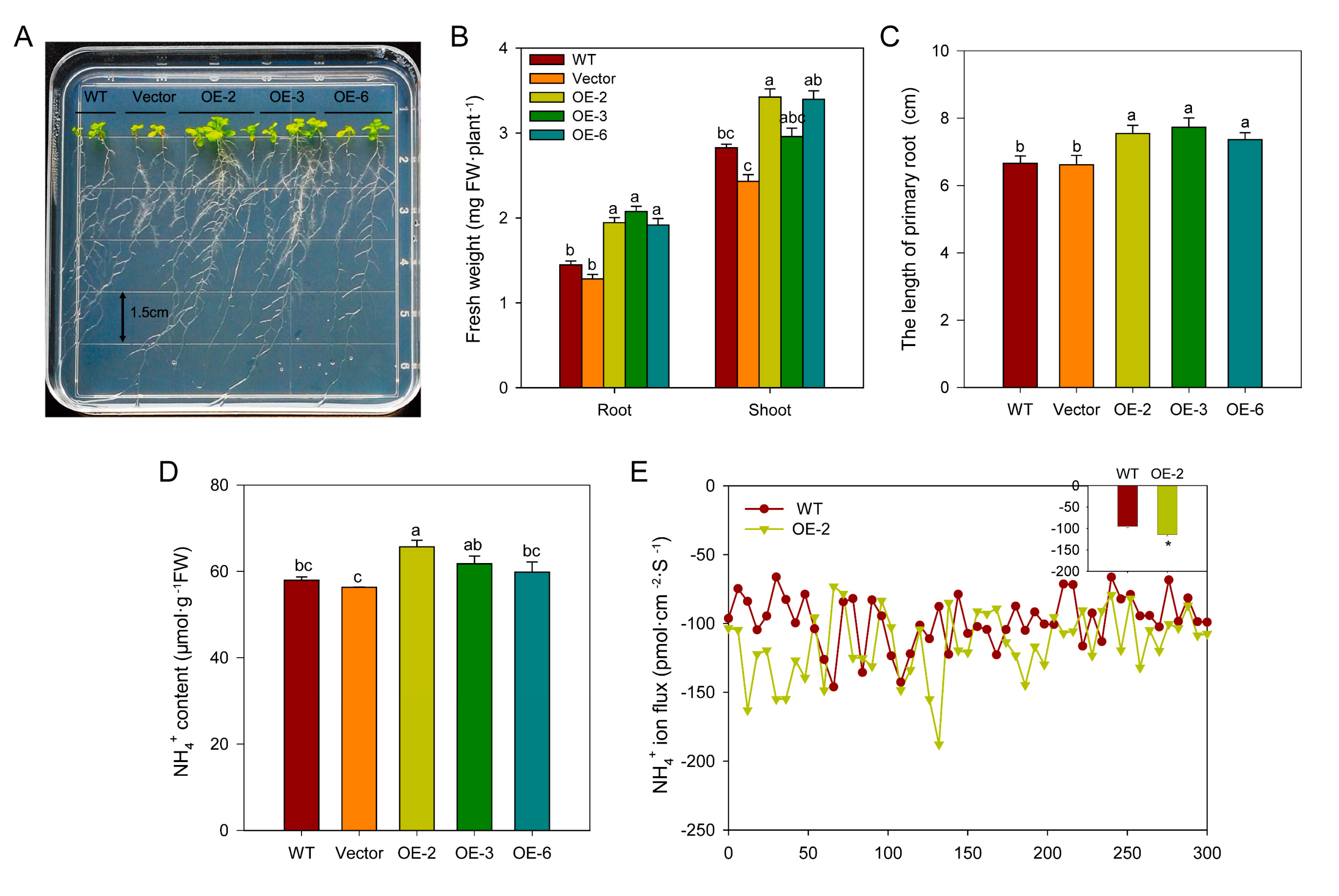
Figure 10.
Effect of Overexpressing BcAMT1.1 on plant growth, nitrogen uptake, and assimilation under mixed NH4+ and NO3- nutrition. (A) Plant phenotype. (B) Fresh weight of roots and shoots. (C) Primary root length and lateral roots numbers. (D) Lateral roots density. (E) NH4+ and NO3- ion fluxes. (F) NH4+ and NO3- content. (G, H) Expression of nitrogen assimilation-related genes in roots and shoots. WT: wild type; OE-2: overexpression line 2. ns: no significant difference; * or ** represents significant differences at P < 0.05 or P < 0.01, respectively.
Figure 10.
Effect of Overexpressing BcAMT1.1 on plant growth, nitrogen uptake, and assimilation under mixed NH4+ and NO3- nutrition. (A) Plant phenotype. (B) Fresh weight of roots and shoots. (C) Primary root length and lateral roots numbers. (D) Lateral roots density. (E) NH4+ and NO3- ion fluxes. (F) NH4+ and NO3- content. (G, H) Expression of nitrogen assimilation-related genes in roots and shoots. WT: wild type; OE-2: overexpression line 2. ns: no significant difference; * or ** represents significant differences at P < 0.05 or P < 0.01, respectively.
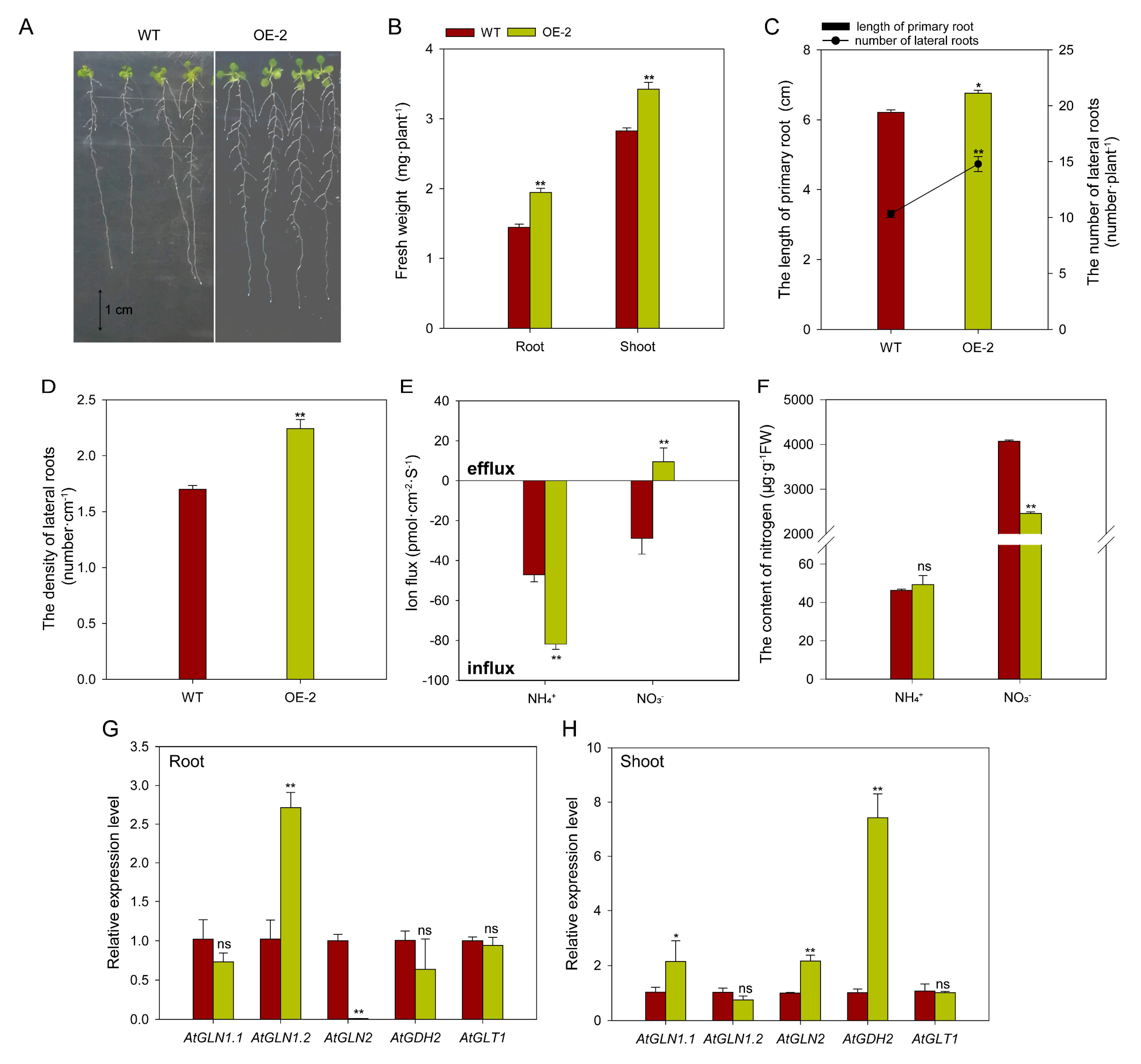
Figure 11.
Predicted protein-protein interaction network of BcAMT1.1.
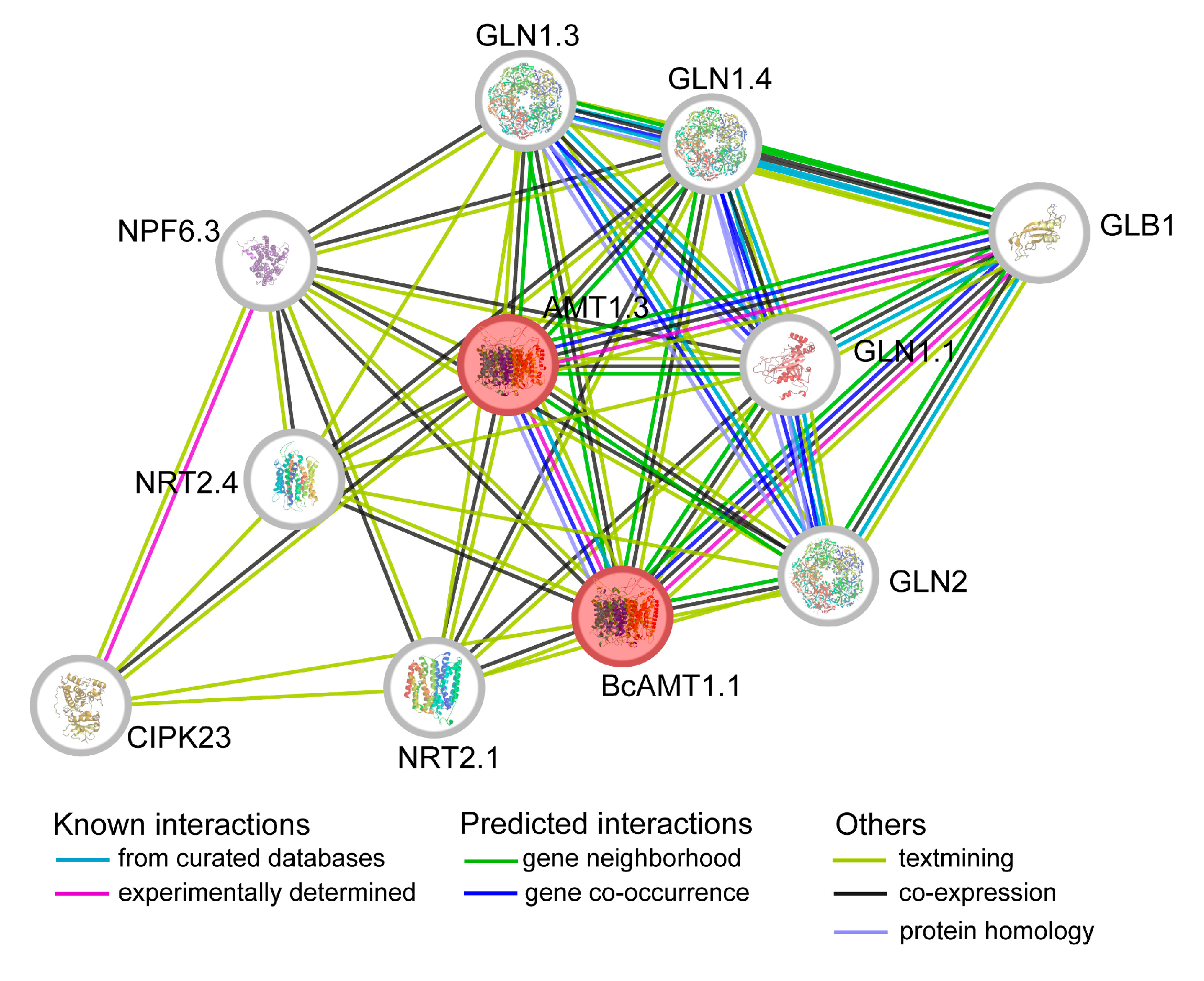
Figure 12.
A proposed model for BcAMT1.1 regulates nitrogen uptake and assimilation under low NH4+ conditions.
Figure 12.
A proposed model for BcAMT1.1 regulates nitrogen uptake and assimilation under low NH4+ conditions.
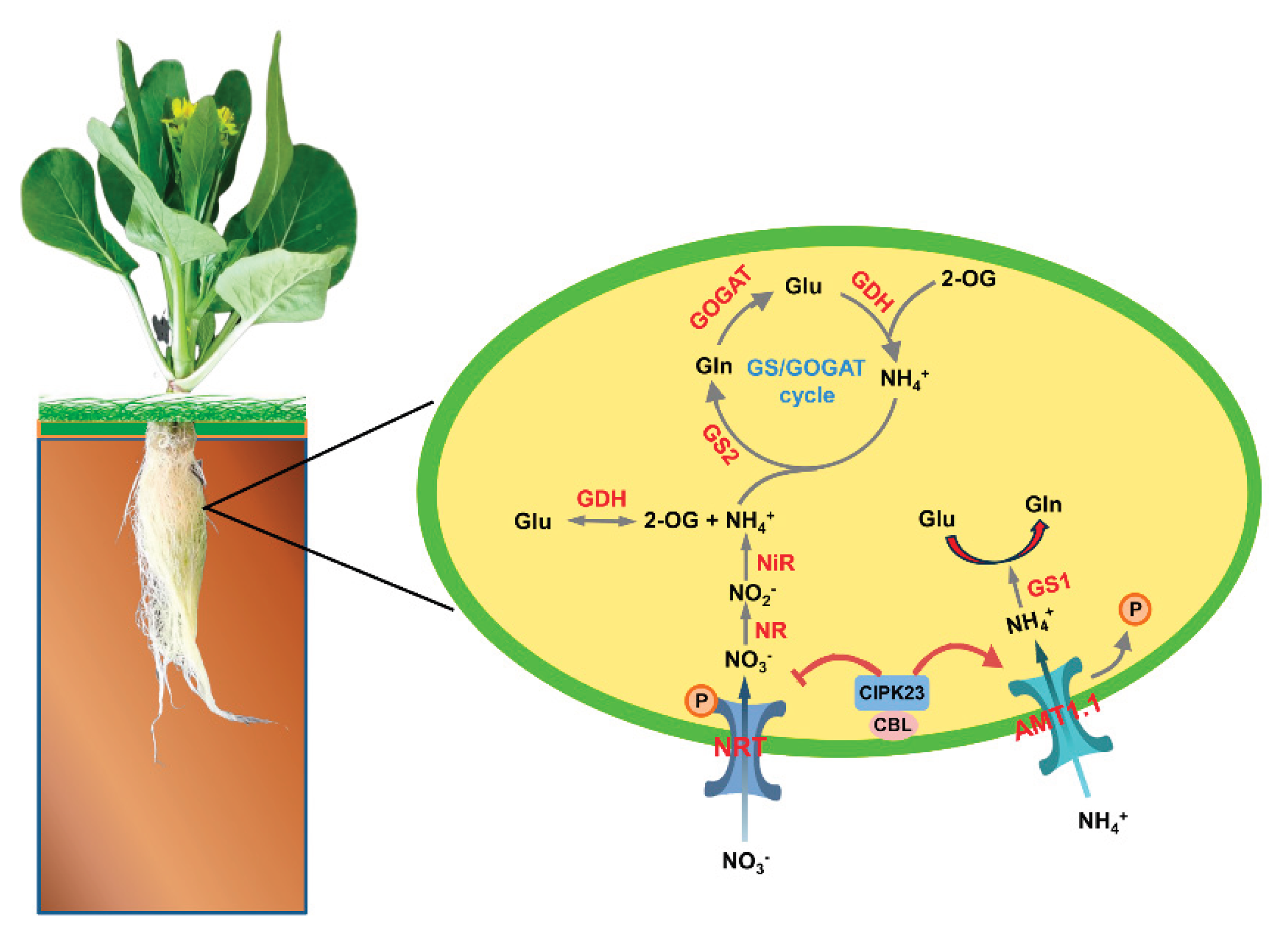

Table 1.
Characteristics of BcAMT gene family members in flowering Chinese cabbage.
| Gene ID | Gene Name | Chr | Start | End | MW (kDa) |
pI | AA (aa) |
Instability index | GRAVY | TM | Subcellular localization |
Category |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bra_cxA05g029310.1 | BcAMT1.1 | A05 | 9051433 | 9052957 | 53.62 | 7.13 | 503 | 25.33 | 0.38 | 9 | Cell membrane | AMT1 |
| Bra_cxA09g068650.1 | BcAMT1.2 | A09 | 1262097 | 1263778 | 54.88 | 7.73 | 512 | 24.10 | 0.35 | 10 | Cell membrane | AMT1 |
| Bra_cxA07g035530.1 | BcAMT1.3 | A07 | 8638823 | 8640466 | 54.11 | 6.74 | 504 | 28.46 | 0.35 | 9 | Cell membrane | AMT1 |
| Bra_cxA01g016480.1 | BcAMT1.3-like | A01 | 20920458 | 20926484 | 50.79 | 5.87 | 476 | 26.46 | 0.39 | 10 | Cell membrane | AMT1 |
| Bra_cxA01g038520.1 | BcAMT1.4 | A01 | 4314030 | 4315548 | 53.66 | 5.7 | 501 | 26.06 | 0.43 | 10 | Cell membrane | AMT1 |
| Bra_cxA03g011620.1 | BcAMT1.4-like | A03 | 28109231 | 28110768 | 54.41 | 5.45 | 509 | 27.01 | 0.45 | 10 | Cell membrane | AMT1 |
| Bra_cxA03g025790.1 | BcAMT1.5 | A03 | 20119522 | 20121038 | 53.17 | 5.96 | 500 | 25.32 | 0.43 | 10 | Cell membrane | AMT1 |
| Bra_cxA05g037880.1 | BcAMT2.1 | A05 | 3982245 | 3986822 | 52.63 | 6.32 | 489 | 28.53 | 0.45 | 11 | Cell membrane | AMT2 |
| Bra_cxA04g005660.1 | BcAMT2.1-like | A04 | 23713465 | 23716475 | 52.52 | 7.28 | 488 | 25.86 | 0.45 | 11 | Cell membrane | AMT2 |
AA: amino acids; Chr: chromosome; GRAVY: grand average of hydropathy; MW: molecular weight; pI: isoelectric point; TM: transmembrane.
Table 2.
Divergence time estimation for paralogous gene pairs of BcAMTs.
| Seq_1 | Seq_2 | Identity (%) | Ka | Ks | Ka/Ks | T/ (MYA) |
|---|---|---|---|---|---|---|
| BcAMT1.3 | BcAMT1.3-like | 87.30 | 0.0394 | 0.4100 | 0.0962 | 13.6678 |
| BcAMT1.5 | BcAMT1.3 | 84.33 | 0.0903 | 0.6636 | 0.1360 | 22.1207 |
| BcAMT1.5 | BcAMT1.3-like | 81.04 | 0.0782 | 0.7431 | 0.1053 | 24.7684 |
| BcAMT1.4 | BcAMT1.4-like | 90.57 | 0.0454 | 0.4027 | 0.1129 | 13.4226 |
| BcAMT2.1 | BcAMT2.1-like | 92.84 | 0.0438 | 0.3165 | 0.1383 | 10.5503 |
Ks: synonymous substitution rate, Ka: nonsynonymous substitution rate, T: divergence time.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



