
Submitted:
11 November 2025
Posted:
12 November 2025
You are already at the latest version
Abstract
Natural products are sources of secondary metabolites with various biological activities. This review highlights the promising potential of the Annonaceae family, a large clade of flowering plants with 107 genera and over 2300 species. Known for their vast pharmacological activities, several genera and species within this family are considered excellent sources of bioactive molecules due to the diversity of their secondary metabolites. Chemical investigations have revealed the presence of alkaloids, mainly isoquinolinic alkaloids, phenolic compounds, terpenoids, lactones, and acetogenins. The Annonaceae family exhibits anti-inflammatory, insecticidal, antimicrobial, leishmanicidal, cytotoxic, antitumor, trypanocidal, antioxidant, gastroprotective, and antimalarial activities. However, most studies focus on plant extracts and essential oils, with few isolated molecules and mechanisms of action identified. Investigating the biological activity of isolated compounds is crucial for new drug discovery. This review also compiles important information for the pharmaceutical and agricultural industries.
Keywords:
annonaceae
; bioactivity
; natural products
; biological activities
1. Introduction
In addition to carrying out photosynthesis, plants also produce secondary metabolites, chemical molecules that can eventually play a biological role in living beings (Moghadamtousi et al. 2013; Ma et al. 2017). Thus, plants are among the main sources for new therapeutic drug candidates (Newman and Cragg 2016; Aminimoghadamfarouj et al. 2020).
Drugs produced from molecules obtained from plants frequently have important advantages over non-plant based products; they exhibit reduced side effects (as well as excellent efficacy and safety) and increased accessibility, which in turn decreases cost and contributes to more sustainable production process. All of this increases the incentive to search for new plant-based therapeutics (Pan et al. 2013; Chakraborty 2018; Sharifi-Rad et al. 2018; Tekuri et al. 2019; Mohammadi et al. 2020).
The Annonaceae family, first described by Antonie Laurent de Jussieu in 1789, stands out as one of the most anatomically and structurally uniform families (Cronquist 1981; Doyle et al. 2004; Cunha 2009; Silva and Domingues Neta 2011). It is one of the richest families of the Manoliophyta (flowering plant), having 107 genera and roughly 2400 species currently recognized (Guo et al. 2017). The Annonaceae has Pantropical distribution in the world, but requires specific characteristics of soil, altitude, temperature and humidity for growth, limiting its distribution to other regions (Popenoe 1921; Encina et al. 2014; Ferreira et al. 2019).
The Annonaceae is rich in fruit species, most of which are edible, making it the target of many studies (Ribeiro et al. 1999; Cunha 2009; Rabêlo 2014). Annonaceae are known for having many bioactive secondary metabolites with vast pharmacological activities. In addition to the classic secondary metabolite classes such as alkaloids, flavonoids, terpenes, etc., Annonaceae are characterized by the presence of isoquinoline alkaloids and acetogenins, a compound class exclusive to this family (Leboeuf et al. 1980; Bermejo et al. 2005; Aminimoghadamfarouj et al. 2011; Cortes et al. 2014). Thus, the Annonaceae family has been massively studied to explore potentially bioactive metabolites.
The objective of this review is to highlight the biological activities recorded by Annonaceae derived compounds and to emphasize the significance of these plants as a source of new therapeutics.
2. Methodology
Information was collected from the literature regarding the biological use of the Annonaceae plants as well as their extracts, essential oils, and secondary metabolites. The articles were collected from Web of Science, PubMed and ScienceDirect using the keywords “Annonaceae”, “Biological activity”, “Activity”, “Pharmacological activities”, and “Biological properties”. Articles were collected through April 2022, and those focusing on synthesis or semi-synthesis, even if inspired by compounds isolated from Annonaceae, were excluded.
3. Results and Discussion
Plants in the Annonaceae family have been widely used in traditional medicine and are well known in the tropical regions of the world. Some traditional uses include the treatment of arthritis, rheumatism, and neuralgia (Cercato et al. 2015), asthma (Auddy et al. 2003; Bhalke and Chavan 2011), wound healing (Tan et al. 2015), cancer (Cascaes et al. 2021), parasitic infections (Moghadamtousi et al. 2015b), as well as fever, diabetes, insomnia, and headaches (Attiq et al. 2017; Cascaes et al. 2021).
As mentioned earlier, the Annonaceae family comprises of approximately 2400 species. Our review includes analysis of about 177 species with proven biological activity, constituting roughly 7.37% of known Annonaceae species. These species are spread across 56 genera, representing 52.33% of all Annonaceae genera. These 56 genera encompass the four subfamilies of Annonaceae (Malmeoideae, Annonoideae, Anaxagoreoidae, and Ambabavioideae), with 8.92% belonging to the Ambavioideae and Anaxagoreoideae subfamilies, which lack tribes. The remaining 91.08% are distributed among seven tribes of Annonoideae and three tribes of Malmeoideae. These data are depicted in a Venn diagram in Figure 1, where the larger square represents the Annonaceae family with its four subfamilies, denoted as circles A, B, C, and D. Within A and B, the Annonoideae and Malmeoideae subfamilies respectively, there are tribes, illustrated by the smaller circles A1 and B1. C represents the Ambavioideae subfamily and D represents the Anaxagoreoideae subfamily. The species examined in the review article are highlighted in red.
The activities listed as “other” include: anti-convulsant activity (Okoye et al. 2013; Moghadamtousi et al. 2015a; Manoj Kumar et al. 2021), anti-Onchocerca, (Dikti Vildina et al. 2021), anti-hyperprolactinemic (Yakubu and Fayemo 2021), α-Glucosidase inhibitory (Suthiphasilp et al. 2021), anti-diabetic (Shirwaikar et al. 2004; Kaleem et al. 2008; Mohd et al. 2009; Qi et al. 2010; Basha and Subramanian 2011; Brindis et al. 2013; Ahalya et al. 2014; Florence et al. 2014; Sahu et al. 2016; Calzada et al. 2017, 2019; Coria-Téllez et al. 2018; Taha et al. 2018; Alsenosy et al. 2019; Mazumdar et al. 2021; Chowdhury et al. 2021; Martínez-Solís et al. 2021), antidiarrhea (Owusu et al. 2021), anti-SARS-CoV-2 (Prasad et al. 2021), anti-acetylcholinesterase (Leite et al. 2021), anthelmintic (Nwosu et al. 2022), antischistosoma (Matchi et al. 2022), anti-Platelet aggregation (Yang et al. 2002), apoptosis induction (Chen et al. 2004; Machana et al. 2012; Pumiputavon et al. 2017), testicular function (Abarikwu et al. 2017), anti-HIV (Piacente et al. 1994; Yang-Chang Wu et al. 1996; Chang et al. 1998; Wafo et al. 1999; Wu et al. 2003; Ding et al. 2010; Saepou et al. 2010; Hongthong et al. 2016; Silprakob et al. 2018; Yu et al. 2019), antiviral activity (Betancur-Galvis et al. 1999; Paredes et al. 2001; Kanokmedhakul et al. 2006; Paarakh et al. 2009; Gajalakshmi et al. 2012; Gavamukulya et al. 2014; Silva et al. 2016), anxiolytic-like activity (López-Rubalcava et al. 2006; Rejón-Orantes et al. 2011), antidepressant (Martínez-Vázquez et al. 2012), and anti-hypertensive (Nwokocha et al. 2012). Herein, we discuss the studies of the most common biological therapeutic aims of the previous studies.
Graph 1 represents the number of articles evaluated in this review grouped by years and in it, one can observe the growth in interest surrounding the Annonaceae family, particularly since 2006 (Menezes et al. 2021). Given the rich structural diversity, it is easy to understand this increase in the search for bioactive Annonaceae-based substances (Menezes et al. 2021).
Our bibliographic survey on biological activities of the Annonaceae resulted in many published bioactivities for a variety of disease states. Graph 2 depicts the number of articles collected for each disease, and due to their prevalence, the top 10 will be discussed in this review.
3.1. Anti-Inflammatory
Inflammation is a physiological response triggered by several factors, such as physical trauma, exposure to allergens, chemical stimuli, infections, etc (Wilson and Trumpp 2006; Guo et al. 2015; Attiq et al. 2017). The inflammatory process can also be initiated by tissue malfunctions or disruptions in homeostasis (Nathan and Ding 2010). In this context, inflammation serves as a protective immune response capable of restoring the body’s homeostasis. However, if unregulated, it can become harmful and lead to several diseases, such as autoimmune diseases (rheumatoid arthritis), hypertension , obesity, and cancer, among others (Jachak 2006; Huscher et al. 2009; Nathan and Ding 2010; Attiq et al. 2017).
Inflammation is didactically divided into two phases, the acute phase, and the chronic phase. The acute phase is characterized by the initial stages of inflammation, the emergence of the five cardinal signs of inflammation: pain, heat, redness, swelling, and loss of function (Sedgwick and Lees 1986; Serhan 2010). It also exhibits vasodilation, exudation of protein-rich plasma fluid, and migration of cells to the site of injury. When inflammation is persistent, the chronic phase begins, inducing changes in the composition of infiltrating leukocytes, that is, the replacement of neutrophils with high concentrations of lymphocytes and macrophages (Sherwood and Toliver-Kinsky 2004; Aller et al. 2007). The chronic phase has a prolonged duration, lasting anywhere from several months to years and is associated with blood vessel proliferation, fibrosis, and tissue necrosis. Inflammatory responses tend to present clinically in a progressive way, combining elements of the two phases (Sherwood and Toliver-Kinsky 2004; Fujiwara and Kobayashi 2005).
Several medications such as non-steroidal anti-inflammatory drugs (NSAIDs), corticosteroids, and disease-modifying anti-rheumatic drugs (DMARDs) are available for the treatment of painful and life-threatening inflammatory conditions (Hoes et al. 2010; Weber and Noels 2011; Attiq et al. 2017). Prolonged use of these drugs can induce toxicity delivering side effects such as uncontrolled hypertension, gastric ulcer, acute kidney failure, liver failure, heart failure, glaucoma, etc (Jachak 2006; Huscher et al. 2009; Attiq et al. 2017). This toxicity remains a major concern with these treatments.
Attiq, Jalil and Husain, 2017, carried out a literature review study highlighting the great potential of the Annonaceae family in fighting inflammation, pointing out isolated substances as great candidates for possible drugs. The authors further highlight terpenes as a promising secondary metabolite class in anti-inflammatory activity. However, there is still much to be studied in the Annonaceae family, especially in the elucidation of substances with biological potential, given the evidence for extracts with potent anti-inflammatory activity. Due to the abundance of studies discussing this anti-inflammatory potential, data from all these studies are summarized in Table 1, and some are discussed below to highlight the variety and complexity of the inflammatory pathways involved. Overall, these products and their derivatives are shown repeatedly to have significant anti-inflammatory activity, regularly displaying IC50 values below that of the therapeutic controls.
Xylopia genus
Xylopia aethiopica is a plant commonly used in African traditional medicine for the healing of wounds, inflammatory disorders, and the treatment of post-natal pain. Based in this traditional knowledge, in 2010 an oil extract from the fruits of Xylopia aethiopica was studied by Ezekwesili et al (2010). Specifically, the effects of the oils carbohydrates, glycosides, flavonoids, saponins, tannins and phytosterols on cell membrane stability and prostaglandin synthetase activity were evaluated (Ezekwesili et al. 2010).
The fruit of . aethiopica is recognized as a source of unsaturated fatty acid. This was demonstrated by HPLC studies of the lipid extract confirming the presence of palmitic acid (19.21%), palmitoleic acid (0.81%), stearic acid (4.54%), oleic acid (39.12%), linoleic acid (25.98%) and linolenic acid (1.10%). The X. aethiopica extract demonstrated the ability to stabilize the erythrocyte membrane, which the authors associated with the stabilization of the lysosomal membrane, a well-known marker of anti-inflammatory potential in drugs. This extract also preserved cellular membrane integrity and acted as a substrate for prostaglandin synthetase, thereby promoting prostaglandin biosynthesis (Ezekwesili et al. 2010).
Xylopic acid is the main compound isolated from the dried fruit of X. aethiopica. Anti-inflammatory effects of this acid have been observed specifically in H2S-induced paw edema models, and demonstrate its intervention along the arachidonic acid pathway (Osafo et al. 2016). The same authors found that xylopic acid was effective in suppressing experimentally induced ulcerative colitis by increasing superoxide dismutase (SOD), ascorbate peroxidase (APX), and catalase (CAT) activity (Osafo et al. 2019). These enzymes had been previously identified as prime biomarkers for colon damage, and treatment with xylopic acid treatment resulted in significantly increased expression in the three enzymes at all administered doses (10-100 mg kg−1). Additionally, rats showed reduced mucosal injury, granulomatous inflammation, and cellular proliferation. Finally, the levels of myeloperoxidase (MPO) and malondialdehyde (MDA), both reactive oxygen metabolites (ROMs), were reduced with all treatments of xylopic acid (Osafo et al. 2019). These two compounds are released by immune cells and play an important role in the physiopathology of ulcerative colitis, where upregulation is an indication of oxidative damage and free radical-induced lipid peroxidation (Liu and Wang 2011).
The anti-inflammatory effects of X. aethiopica leaves may involve phenolic compounds identified through HPLC-DAD, including cynaroside, rutin, quercitrin, astragalin, and nicotiflorin. O-caffeoylquinic acids and the luteolin monoglycoside cynaroside were also detected in the extract (Macedo et al. 2020).
The anti-inflammatory properties of essential oils obtained from eight batches of X. aethiopica fruits from Ghana and Nigeria were obtained by hydrodistillation and tested on RAW 264.7 macrophage cells. The results showed that the oils inhibit NO production in LPS-stimulated RAW264.7 cells concentration-dependently, and better than the control drug, dexamethasone. The oils from Ghana demonstrated better anti-inflammatory effects than those of Nigerian origin, and the authors suggested this was due to the synergistic effects of the 14 different metabolites commonly found in the extract (Alolga et al. 2019).
Other species of Xylopia, including Xylopia parviflora, Xylopia sericea, and Xylopia vielana, have also been investigated. The essential oil from striped African pepper (Xylopia parviflora) was studied at different concentrations in RAW 264.7 macrophages stimulated with LPS and shown to decrease NO production by 37% (Woguem et al. 2014). However, a chemical profile of this essential oil (finding mainly β-pinene (34.0%) and α-pinene (10.3%)), does not report any compounds previously identified as anti-inflammatory (Woguem et al. 2014).
An in-silico analysis assessed the anti-inflammatory potential of five guaiane-type sesquiterpene dimers, known as xylopidimers A-E, extracted from Xylopia vielana. Molecular docking was performed on the COX-2 protein as anti-inflammatory target (PDB: 1CX2). All the five evaluated dimers showed potent inhibitory activity against COX-2, with binding energy values among -10.51 Kcal/mol and -9.23 Kcal/mol, significantly lower than the controls Ibuprofen and Felbinac (Hassan et al. 2022). Molecular dynamics simulations suggested that these guaiane-type sesquiterpenes allow a ligand-protein stability. This in silico work is one of the few studies that report specific secondary metabolites together with the xylopic acid, since most anti-inflammatory assays for this genus are of the polar extracts.
Annona genus
In a bio-guided fractionation study, the anti-inflammatory activity of the hydromethanolic extracts of the leaves, pulp, and seeds of Annona cacans were evaluated in a paw edema model. These compounds inhibited the increase in MPO activity after 6 hrs when compared to both the dexamethasone and control groups (Volobuff et al. 2019).
Similarly, a polyphenol-enriched fraction of Annona crassiflora collected in Brazil was studied to evaluate its anti-inflammatory activity. The main components were identified as chlorogenic acid, epi-catechin, procyanidins B2 and C1, quercetin-glucoside, kaempferol, and caffeoyl-glucoside (de Moura et al. 2019). Through 1H NMR studies, kaempferol 3-O-β-glucoside and kaempferol 3-O-β-diglucoside were identified in the extract of Annona crassiflora and experimentally determined to inhibit paw edema, reduce myeloperoxidase activity, and reduce the total leukocyte count (Rocha et al. 2016). These results were complimentary to those observed by De Moura et al., 2019, (de Moura et al. 2019)using the polar extracts of this species.
The lyophilized fruit extract of another species of this genus, Annona muricata, which is found in different tropical regions and is commonly used in Africa, was studied in vivo and found to possesses analgesic and anti-inflammatory activities in various models (Ishola et al. 2014).
Several compounds from Annona squamosa L. and Annona reticulata L. barks were evaluated in vivo by Chavan et al., including a sesquiterpene fraction composed of copaene (35.40%), patchoulane (13.49%) and 1H-cycloprop(e)azulene (22.77%) (Chavan et al. 2012) as well as caryophyllene oxide (Chavan et al. 2010a), 18-acetoxy-ent-kaur-16-ene (Chavan et al. 2011) and saponified petroleum ether extract from the bark of Annona reticulata L. bark (Chavan et al. 2010b). All of these compounds exhibited anti-inflammatory activity, with many also demonstrating analgesic effects.
The anti-inflammatory properties of eight ent-kauranes were evaluated using fruits from Annona glabra, including three novel compounds: 7β,16α,17-trihydroxy-ent-kauran-19-oic acid, 7β,17-dihydroxy-16α-ent-kauran-19-oic acid 19-O-β-D-glucopyranoside ester, and 7β,17-dihydroxy-ent-kaur-15-en-19-oic acid 19-O-β-D-glucopyranoside ester. All tested ent-kauranes demonstrated greater inhibition of NO production compared to the control, dexamethasone (Nhiem et al. 2015).
Duguetia genus
The essential oil of Duguetia furfuracea, consisting of 24 volatile compounds, demonstrated significant inhibition of LPS-induced inducible nitric oxide synthase (iNOS) expression. It also reduced tumor necrosis factor alpha (TNF-α) production and inhibited the recruitment of polymorphonuclear leukocytes (Saldanha et al. 2019). Furthermore, extraction and enrichment of the phenylpropanoids further attenuated the disease states through the same pathways (Saldanha et al. 2020, 2021). Using leaves of this same species, a methanolic extract with high contents of phenols (624.37 mg/g), flavonoids (580.21 mg/g), and flavonols (254.44 mg/g) was obtained and showed significant decrease in inflammation in edema (do Santos et al. 2018).
Duguetia staudtii is a species found in Africa, mainly in a forest from Sierra Leone to Cameroon. The anti-inflammatory activity of eight compounds (one pachypolignan, one bisnorlignan, four flavonoids, one alkaloid, and one triterpenoid) isolated from Duguetia staudtii were evaluated in myeloperoxidase dependent (luminol/zymosan) and independent (lucigenin/PMA) oxidative burst assays. Inhibitory (IC50) values between 6.44 and 14.13 μg/mL were found for pachypodol, kumatakenin, and 5,4′-dihydroxy-3,7,3′,5′-tetramethoxyflavone in macrophages (Ngouonpe et al. 2019).
Polyalthia genus
An extensive review conducted by Yao et al. in 2019 on the Polyalthia genus revealed several compounds with potential anti-inflammatory properties. These compounds comprised 16-oxocleroda-3,13-dien-15,16-oic acid and 16-hydroxycleroda-3,13-dien-15,16-olide (clerodane diterpenoids), rutin and quercetin (flavonoids), spinasterol and α-spinasterol (phytosterols), as well as goniothalamin and (−)-5-hydroxygoniothalamin (6S-styrylpyrones). All tested compounds showed IC50 values comparable to the positive control. The primary anti-inflammatory mechanisms identified included the inhibition of nuclear factor kappa B (NF-κB), prostaglandins (PGs), pro-inflammatory cytokines, inducible nitric oxide synthase (iNOS), and reactive oxygen species (ROS) (Yao et al. 2019).
Recently, Chen et al., 2021, in a review study of the pharmacological activities of phytochemicals from Polyalthia, added new compounds with potential anti-inflammatory activity: polycerasoidol (prenylated benzopyran), 16-Hydroxycleroda-4(18),13-dien-15,16-olide, (-)-3α,16α-dihydroxycleroda-4(18),13(14)Z-dien-15,16-Olide, and (4→2)-abeo-16(R&S)-2,13Z-clerodadien-15,16-olide-3-al. These clerodane compounds, together with dehydrogoniothalamin and the two 6S-styrylpyrones mentioned earlier, were assessed collectively (Chen et al. 2021).
Clerodane diterpenoids were obtained from the methanolic extract of Polyalthia longifolia and tested against neutrophil fMLP/CB induced superoxide generation and phorbol 12-myristate 13-acetate (PMA)-induced action. The compound 16-oxocleroda-4(18),13-dien-15,16-olide obtained an IC50 value very close to that of the well-known NADPH oxidase inhibitor, diphenyleneiodonium (Chang et al. 2006).
The diterpenoid 16(R&S)-3,13-kolavadien-15,16-olide-2-one, at a concentration of 10μg/mL, showed the lowest IC50 value against phorbol 12-myristate 13-acetate (PMA)-induced action. The authors related this action with a critical structural feature, an E form double bond at C-13(14) of the 3-ene-clerodane skeleton (Chang et al. 2006). Recently, three of five compounds isolated from a methanolic extract of Polyalthia longifolia (16-hydroxy-cleroda-4(18),13-dien-16,15-olide, 3α,16α-dihydroxycleroda-4(18),13(14)Z-dien-15,16-olide, and 16α-hydroxy-cleroda-3,13(14)Z-dien-15,16-olide) exhibited potency as COX-1, COX-2, and 5-LOX inhibitors with IC50 values similar or lower to Indomethacin (COX-1 and COX-2) and diclofenac (5-LOX) controls (Nguyen et al. 2020).
Interestingly, these authors performed an in-silico study using the PDB structures of the three proteins: COX-1 (PDB ID: 2OYU), COX-2 (PDB ID: 4COX), and 5-LOX (PDB ID: 3V99), and found a high concurrence with the in vitro studies. Two structures, 3α,16α-dihydroxycleroda-4(18),13(14)Z-dien-15,16-olide and 16α-hydroxy-cleroda-3,13(14)Z-dien-15,16-olide, were reported as better inhibitors of COX-1, COX-2, and 5-LOX compared to their respective control drugs (Nguyen et al. 2020).
The anti-inflammatory activity of extracts from other two Polyalthia species, P. simiarum and P. suberosa were evaluated through carrageenan induced paw edema and Xylene-Induced ear edema models, respectively. All three extracts showed significant inhibition at all time points and doses, and acted comparably to the ear edema model control (Yasmen et al. 2018; Kabir et al. 2019).
Others Annonaceae species
Spectroscopic techniques led to the identification of seven compounds from the methanolic extract of Uvaria flexuosa leaves, including flexuvaroxepine A, flexuvarin A–D, and flexuvarol A–B, along with four known flavonoids: 6,7-di-O-methyl-baicalein, chrysin, negletein, and 6-hydroxy-5,7-dimethoxy-flavone. In a fMLP/CB model of human neutrophils, both Flexuvarol B and chrysin demonstrated greater anti-inflammatory activity compared to the positive control, genistein (Hsu et al. 2016).
In another study, (-) zeylenol, a polyoxygenated cyclohexene derivative, was isolated from the ethyl acetate extract of U. grandiflora stems and displayed anti-inflammatory activity in an EPP-induced rat ear edema model. The authors found that (-) zeylenol produced a significant inhibitory effect on the edema formation at all times tested, and the values were similar to those obtained by a dose of 1mg/ear of phenylbutazone, a positive control (Seangphakdee et al. 2013).
An isoquinoline alkaloid, dactyllactone A, was isolated from Dactylicapnos scandens and demonstrated a more potent, dose-dependent inhibition of LPS-treated RAW264.7 cells compared to the positive control, without showing cytotoxic effects (Wang et al. 2018).
Additionally, the same researchers isolated two novel aporphines from Dactylicapnos scandens, identified as dactylicapnosines A and B, which feature unique five-membered D carbon rings. In LPS-induced RAW 264.7 cells, dactylicapnosine A exhibited anti-inflammatory and analgesic properties by inhibiting the expression of TNF-α, IL-1β, and PGE2. Furthermore, an evaluation of its effect on xylene-induced inflammation in mice, through intraperitoneal injection of dactylicapnosine A (10.2 mg/kg), revealed superior swelling inhibition compared to the positive control Parecoxib (10 mg/kg) (Wang et al. 2020).
In another study, twenty-three chemical components of Phaeanthus vietnamensis were isolated, with three being novel. Anti-inflammatory properties of these molecules were evaluated through an assay looking at inhibitory NO production in BV-2 cells. Four compounds displayed significant inhibition, and the IC50 of spathulenol was lower than the control. Interestingly, none of the 12 tested compounds displayed cytotoxicity in a cell viability assay (treatment with 20 μM of each compound maintained viability above of 95%) (Nhiem et al. 2017).
The phenolic amide, Melodamide A, along with 12 known compounds was isolated from the leaves of Melodorum fruticosum. The inhibitory effects of these compounds were evaluated using a human neutrophil model with fMLP/CB. Melodamide A demonstrated the most potent inhibition of superoxide anion generation induced by fMLP/CB, without causing cytotoxicity (Chan et al. 2013).
To enhance the anti-inflammatory properties of Melodamide A, a series of analogues were synthesized and tested. Among the sixteen analogues, only three with modifications on the A-ring demonstrated inhibitory effects on superoxide anion formation, with IC50 values of 7.49 μM (2-Cl), 5.59 μM (3-F), and 5.19 μM (2-Br). However, none of the analogues exhibited elastase inhibitory activity, leading the authors to conclude that the substitutions on the A-ring did not significantly improve anti-inflammatory activity (Chan et al. 2013).

Table 1.
Summary of anti-inflammatory activity of species of the Annonaceae.
| Annonaceae Species | Used Material | Substances/Extracts | Methodology | Inhibition | Ref. |
|---|---|---|---|---|---|
|
Annona cacans |
Leaves Pulp Seeds |
Hydromethanolic extract (HME) Ethyl acetate fraction (EAF) |
Myeloperoxidase (MPO) activity carrageenan-induced paw oedema | After 6 h, 28% (300 mg/kg HME-Leaves) 53% (100 mg/kg HME-Pulp) 58% (300 mg/kg HME-Pulp) 43% (30 mg/kg EAF), 51% (100 mg /kg EAF) |
(Volobuff et al. 2019) |
| Annona crassiflora | Fruit peel Leaves Leaves Leaves Leaves |
Polyphenol-enriched fraction (PEF) Methanolic extract Methanolic extract Methanolic extract Methanolic extract |
Wound closure in C57 mice Carrageenan-induced edema MPO activity Total leukocytes Carrageenan-induced leukocyte migration |
75% (2% PEF topical) 84%(6% PEF topical) 53% (100mg/kg) 47% (300mg/kg) 60% (100mg/kg) 78% (100mg/kg) 90% (300mg/kg) 43% (300mg/kg) |
(de Moura et al. 2019) (Rocha et al. 2016) (Rocha et al. 2016) (Rocha et al. 2016) (Rocha et al. 2016) (Rocha et al. 2016) |
| Annona glabra | Fruits | 7β,16α,17-trihydroxy-ent-kauran-19-oic acid | NO production in LPS-stimulated RAW264.7 cells | IC50= 0.39 ± 0.12 μM | (Nhiem et al. 2015) |
| Fruits | 16β,17-dihydroxy-ent-kauran-19-al | NO production in LPS-stimulated RAW264.7 cells | IC50 = 0.32 ± 0.04 μM | (Nhiem et al. 2015) | |
|
Annona muricata |
Fruits | Lyophilized extract | Xylene-induced ear edema | 34.04% (50μg/mL) 63.83(100μg/mL) 80.85(200μg/mL) |
(Ishola et al. 2014) |
| Annona muricata | Fruits | Lyophilized extract | Cyclooxygenase (COX)-1 activity | 39.44% (100μg/mL) | (Ishola et al. 2014) |
| Fruits | Lyophilized extract | Cyclooxygenase (COX)-2 activity | 55.71% (100μg/mL) | (Ishola et al. 2014) | |
|
Annona nutans |
Fruits | Lyophilized extract | Cyclooxygenase (COX)-2 activity | 55.71% (100μg/mL) | (Ishola et al. 2014) |
| Annonaceae Species | Used Material | Substances/Extracts | Methodology | Inhibition | Ref. |
|
Annona Senegalensis |
Seeds | N-cerotoyltryptamine | ROS production in zymosan stimulated human whole blood phagocytes | IC50 = 2.7 ± 0.1 μg/mL | (Tamfu et al. 2021) |
| Seeds | Methanolic extract | ROS production in zymosan stimulated human whole blood phagocytes | IC50 =8.7 ± 10.2 μg/mL | (Tamfu et al. 2021) | |
| Seeds | Acetogenin | NO production in lipopolysaccharide (LPS) stimulated J774.2 mouse macrophages | IC50 = 3.9 ± 0.2 μg/mL | (Tamfu et al. 2021) | |
|
Annona squamosa |
Bark | Caryophyllene oxide | Carrageenan-induced paw edema | After 2 hours 45% (12.5mg/kg) 51% (25mg/kg) |
(Chavan et al. 2011) |
| Bark | Sesquiterpene fraction (copaene (35.40%), patchoulane (13.49%) and 1H-cycloprop(e)azulene (22.77%)) | Carrageenan-induced paw edema | After 2 hours 38% (12.5mg/kg) 34% (25mg/kg) |
(Chavan et al. 2012) | |
| Bark | 18-acetoxy-ent-kaur-16-ene | Carrageenan-induced paw edema | After 2 hours 51.6% (12.5mg/kg) 60.9% (25mg/kg) |
(Chavan et al. 2011) | |
| Annona vepretorum | Leaves | Ethanolic extract | leukocyte migration to the peritoneal cavity |
62%(25 mg/kg), 76% (50 mg/kg) 98% (100 mg/kg) |
(Silva et al. 2015) |
| Leaves | Ethanolic extract | Carrageenan-induced paw edema | After 2 hours 58%(25 mg/kg) 45% (50 mg/kg) 72% (100 mg/kg) |
(Silva et al. 2015) | |
| Annona vepretorum | Leaves | Ethanolic extract | Histamine-induced paw edema | After 1 hour >65% (100mg/kg) |
(Silva et al. 2015) |
| Cyathocalyx pruniferus | Leaves | Spathulenol Cyclopropa-azulene Polycarpol Koetjapic acid 2-Octaprenyl-benzoquinone 14-methylloctadec-1-ene 1-Docosene β-Sitosterol |
PGE2 | 71.4 (IC50 = 25.8) 8.6 (IC50 = -) 70.1 (IC50 = 24.7) 80.4 (IC50 = 13.1) 86.1 (IC50 = 11.2) 3.5 (IC50 = -) 5.8 (IC50 = -) 21.6 (IC50 = -) |
(Attiq et al. 2021) |
| Annonaceae Species | Used Material | Substances/Extracts | Methodology | Inhibition | Ref. |
| Cyathocalyx pruniferus | Leaves | α-Tocopherol 11-prenol Quercetin Epicatechin Chrysin Indomethacin (Positive control) |
PGE2 | 15.9 (IC50 = -) 2.1 (IC50 = -) 79.8 (IC50 = 15.4) 77.3 (IC50 =17.3) 73.8 (IC50 = 21.8 88.1 (IC50 = 11.8) |
(Attiq et al. 2021) |
| Cyathocalyx pruniferus | Leaves | Spathulenol Cyclopropa-azulene Polycarpol Koetjapic acid 2-Octaprenyl-benzoquinone 14-methylloctadec-1-ene 1-Docosene β-Sitosterol α-Tocopherol 11-prenol |
COX-2 |
21.1 (IC50 = -) 4.6 (IC50 = -) 29.6 (IC50 = -) 85.6 (IC50 = 8.1) 88.1 (IC50 = 6.6) 2.1 (IC50 = -) 2.4 (IC50 = -) 11.1 (IC50 = -) 10.5 (IC50 = -) |
(Attiq et al. 2021) |
| Cyathocalyx pruniferus | Leaves | Quercetin Epicatechin Chrysin Dexamethasone (Positive control) |
COX-2 | 3.3 (IC50 = -) 80.1 (IC50 = 10.3) 74.7 (IC50 =12.5) 70.5 (IC50 =15.7) 92.8 (IC50 = 5.1) |
(Attiq et al. 2021) |
|
Duguetia furfuracea |
stem bark | Essential oil | LPS-induced paw edema | After two hours 41.67% (3mg/kg) 86.11% (10mg/kg) |
(Saldanha et al. 2019) |
| stem bark | Essential oil | LPS-induced paw edema | After four hours 45.45% (1mg/kg) 63.64% (3mg/kg) 92.42% (10mg/kg) |
(Saldanha et al. 2019) | |
| Leaves | Methanolic extract | Carrageenan-induced paw edema | After two hours 39% (300mg/kg) 22% (100mg/kg) 17.5% (30mg/kg) |
(do Santos et al. 2018) | |
| Annonaceae Species | Used Material | Substances/Extracts | Methodology | Inhibition | Ref. |
| Duguetia furfuracea | Leaves | Methanolic extract | Carrageenan-induced paw edema | After four hours 40% (300mg/kg) 25% (100mg/kg) |
(do Santos et al. 2018) |
| Leaves | Dicentrinone | Carrageenan-induced paw edema | For 100 mg/kg 69.1% (2 hours) 50.4% (4 hours), |
(do Santos et al. 2018) | |
| Leaves | Methanolic extract | Zymosan-induced edema | 38.1% (300mg/kg) | (do Santos et al. 2018) | |
| Leaves | Dicentrinone | Zymosan-induced edema | 27.1% (300mg/kg) | (do Santos et al. 2018) | |
| Leaves | Enriched phenylpropanoid extract | LPS-induced paw edema | After two hours 90.91% (3mg/kg) 92.42% (10mg/kg) |
(Saldanha et al. 2021) | |
| Duguetia furfuracea | Leaves | Enriched phenylpropanoid extract | LPS-induced paw edema | After four hours 77.78% (1mg/kg) 77.78% (3mg/kg) 81.48% (10mg/kg) |
(Saldanha et al. 2021) |
| Leaves | α-asarone | LPS-induced paw edema | After two hours 62.12% (3mg/kg) 69.70% (10mg/kg) 69.70% (30mg/kg) |
(Saldanha et al. 2020) | |
| Leaves | α-asarone | LPS-induced paw edema | After four hours 72.22% (3mg/kg) 81.48% (10mg/kg) 81.48% (30mg/kg) |
(Saldanha et al. 2020) | |
|
Duguetia moricandiana |
Fruits | Discretamine | NO production in LPS-stimulated macrophages | Around 50%. (100 and 200 μg/mL) |
(Lemos et al. 2017) |
| Fruits | Discretamine | IL-6 production in LPS-stimulated macrophages | 74.1% (50μg/mL) 76.6% (100μg/mL) 75.1% (200 μg/mL) |
(Lemos et al. 2017) | |
| Fruits | Discretamine | IL1-b production in LPS-stimulated macrophages | 89.4% (50μg/mL) 87.4% (100μg/mL) 71.8% (200 μg/mL) |
(Lemos et al. 2017) | |
| Annonaceae Species | Used Material | Substances/Extracts | Methodology | Inhibition | Ref. |
|
Duguetia moricandiana |
Fruits | Discretamine | TNF-α production in LPS-stimulated macrophages | 61.0% (50μg/mL) 45.2% (100μg/mL) 52.6% (200 μg/mL) |
(Lemos et al. 2017) |
| Fruits | Discretamine | Carrageenan-induced paw edema | After one hour 42% (10mg/kg) 62% (20mg/kg) |
(Lemos et al. 2017) | |
| Fruits | Discretamine | Carrageenan-induced paw edema | After two hours 44% (10mg/kg) 67% (20mg/kg) |
(Lemos et al. 2017) | |
|
Duguetia moricandiana |
Fruits | Discretamine | Carrageenan-induced paw edema | After four hours 59% (5mg/kg) 49% (10mg/kg) 48% (20mg/kg) |
(Lemos et al. 2017) |
|
Duguetia staudtii |
Stem bark | Pachypodol | Myeloperoxidase dependent (luminol/zymosan) oxidative burst | IC50 = 8.32 μg/mL | (Ngouonpe et al. 2019) |
| Stem bark | Kumatakenin | Myeloperoxidase dependent (luminol/zymosan) oxidative burst | IC50 = 10.64 μg/mL | (Ngouonpe et al. 2019) | |
| Stem bark | 5,4′-dihydroxy-3,7,3′,5′-tetramethoxyflavone | Myeloperoxidase dependent (luminol/zymosan) oxidative burst | IC50 = 6.44 μg/mL | (Ngouonpe et al. 2019) | |
| Stem bark | Pachypodol | Myeloperoxidase independent (lucigenin/PMA) oxidative burst | IC50 = 11.04 μg/mL | (Ngouonpe et al. 2019) | |
| Stem bark | Kumatakenin | Myeloperoxidase independent (lucigenin/PMA) oxidative burst | IC50 = 14.13 μg/mL | (Ngouonpe et al. 2019) | |
| Stem bark | 5,4′-dihydroxy-3,7,3′,5′-tetramethoxyflavone | Myeloperoxidase independent (lucigenin/PMA) oxidative burst | IC50 = 8.55 μg/mL | (Ngouonpe et al. 2019) | |
| Enicosanthum membranifolium |
2β-methoxyhardwickiic acid (-)-Hardwicckiic acid 2β-acetoxyhardwickiic acid 2β-hidroxyhardwickiic acid 15-methozypatagonic acid Indomethacin (Positive control) |
NO production | IC50 μM 65.4 38.9 16.1 82.4 28.9 32.2 |
(Polbuppha et al. 2022) | |
| Annonaceae Species | Used Material | Substances/Extracts | Methodology | Inhibition | Ref. |
| Isolona dewevrei | Leaves | Essential oil Nordihydroguaiaretic (Positive control) |
Inhibit lipoxygenases (LOX) | 0.0020 (mg/mL) 0.013 (mg/ml) |
(Kambiré et al. 2021) |
| Melodorum fruticosum | Leaves | Melodamide A | Inhibition of superoxide anion generation | IC50 = 5.25 μM | (Chan et al. 2013) |
| Leaves | Melodamide A derivate (2-Cl) |
Inhibition of superoxide anion generation | IC50 = 7.49 μM | (Chan et al. 2013) | |
| Leaves | Melodamide A derivate (3-F) |
Inhibition of superoxide anion generation | IC50 = 5.59 μM | (Chan et al. 2013) | |
| Leaves | Melodamide A derivate (2-Br) |
Inhibition of superoxide anion generation | IC50 = 5.19 μM | (Chan et al. 2013) | |
|
Phaeanthus vietnamensis |
Leaves | spathulenol | NO production in LPS-stimulated BV2 cells | IC50 = 15.7 μM | (Nhiem et al. 2017) |
| Leaves | (8R,80R)-bishydrosyringenin | NO production in LPS-stimulated BV2 cells | IC50 = 25.3 μM | (Nhiem et al. 2017) | |
| Leaves | 1αH,5βH-aromandendrane-4α,10α-diol | NO production in LPS-stimulated BV2 cells | IC50 = 23.0 μM | (Nhiem et al. 2017) | |
| Leaves | 1βH,5βH-aromandendrane-4α,10β-diol | NO production in LPS-stimulated BV2 cells | IC50 = 22.6 μM | (Nhiem et al. 2017) | |
|
Polyalthia longifolia |
Seeds | 16-oxo-cleroda-3,13(14)E-dien-15-oic acid | COX-1, COX-2, and 5-LOX inhibitory activities | 62.85% (COX-2) 26.41% (LOX-5) |
(Nguyen et al. 2020) |
| Seeds | 16-hydroxy-cleroda-3,13-dien-15-oic acid | COX-1, COX-2, and 5-LOX inhibitory activities | 84.98% (COX-2) 30.51% (LOX-5) |
(Nguyen et al. 2020) | |
| Seeds | 16-hydroxy-cleroda-4(18),13-dien-16,15-olide | COX-1, COX-2, and 5-LOX inhibitory activities | 82.97% (COX-2) 12.73% (LOX-5) |
(Nguyen et al. 2020) | |
| Seeds | 3α,16α-dihydroxy-cleroda-4(18),13(14)Z-dien-15,16-olide | COX-1, COX-2, and 5-LOX inhibitory activities | 75.14% (COX-2) 14.38% (LOX-5) |
(Nguyen et al. 2020) | |
|
Polyalthia longifolia |
Seeds | 16α-hydroxy-cleroda-3,13(14)Z-dien-15,16-olide | COX-1, COX-2, and 5-LOX inhibitory activities | 92.94% (COX-1) 79.41% (COX-2) 16,94% (LOX-5) |
(Nguyen et al. 2020) |
| Bark |
16-oxocleroda-4(18),13-dien-15,16-olide |
fMLP/CB induced superoxide generation by neutrophils |
IC50 = 0.60 mg/mL |
(Chang et al. 2006) | |
| Annonaceae Species | Used Material | Substances/Extracts | Methodology | Inhibition | Ref. |
|
Diphenyleneiodonium (positive control) 16(R&S)-3,13-kolavadien-15,16-olide-2-one |
fMLP/CB induced superoxide generation by neutrophils Phorbol 12-myristate 13 acetate (PMA)-induced action |
IC50 = 0.11 mg/mL IC50 = 10μg/mL |
(Chang et al. 2006) | ||
| Stem bark | Ethyl acetate (EA) extract | Carrageenan-induced paw edema | After four hours 27.5%(50mg/kg) 39.1% (100mg/kg) |
(Kabir et al. 2019) | |
| Leaves | Diethyl ether extract | Xylene-induced ear edema | 42.70% (200mg/kg) 62.67% (400mg/kg) |
(Yasmen et al. 2018) | |
| Leaves | n-hexane extract | Xylene-induced ear edema | 48.54% (200mg/kg) 65.92% (400mg/kg) |
(Yasmen et al. 2018) | |
| Polyalthia viridis | Leaves Stem |
Leaf Essential oil Stem Essential oil Butein (Positive control) |
NO production in LPS stimulated BV2 cells | 80.8 (IC50 = 76.7 μg/mL) 87.2 (IC50 = 57.6 μg/mL) 91.8 (IC50 = 16.1 μg/mL) |
(Son et al. 2021) |
|
Uvaria flexuosa |
Leaves | Flexuvarol B | Superoxide anion generation assay | IC50 = 4.72 mM | (Hsu et al. 2016) |
| Leaves | Chrysin | Superoxide anion generation assay | IC50 = 2.25 mM | (Hsu et al. 2016) | |
| Leaves | Flexuvarol B | Elastase release assay | IC50 = 5.55 mM | (Hsu et al. 2016) | |
| Leaves | Chrysin | Elastase release assay | IC50 = 2.44 mM | (Hsu et al. 2016) | |
| Uvaria grandiflora | Stems | (-) zeylenol | EPP-induced rat ear edema | 90% (15min) 69% (30min) 52% (1 hour) 52% (2 hours) |
(Seangphakdee et al. 2013) |
|
Xylopia aethiopica |
Fruits | Methanolic extract | Prostaglandin synthetase activity | - | (Ezekwesili et al. 2010) |
| Fruits | Ethanolic extract | Mouse pinnal inflammation in carrageenan-induced paw oedema | 23% (30 μg/mL) 62% (300 μg/mL) |
(Obiri and Osafo 2013) | |
| Annonaceae Species | Used Material | Substances/Extracts | Methodology | Inhibition | Ref. |
| Xylopia aethiopica | Fruits | Extract 30 100 300 |
Superoxide dismutase (SOD), ascorbate peroxidase (APX), catalase (CAT), myeloperoxidase (MPO) and malondialdehyde (MDA) activity | 23.06 40.91 62.83 |
(Osafo et al. 2016) |
| Leaves | Hydroethanolic extract | TNF-α in (Lipopolysaccharide) LPS challenged THP-1-derived macrophages | >90% (500 mg/kg) | (Macedo et al. 2020) | |
| Leaves | Hydroethanolic extract | Inhibition of IL-6 production, in a LPS challenged THP-1-derived macrophages, | 84.6% (250 μg/mL) 96.3% (500 μg/mL) |
(Macedo et al. 2020) | |
| Leaves | Hydroethanolic extract | Interferences of 5-LOX in a LPS challenged THP-1-derived macrophages | IC50 = 85 μg/mL | (Macedo et al. 2020) | |
| Fruits | Essential oil | NO production in LPS-stimulated RAW264.7 cells | - | (Alolga et al. 2019) | |
|
Xylopia parviflora |
Fruits | Volatile oil | NO production in LPS-stimulated RAW264.7 cells | 37% (12μg/mL) | (Woguem et al. 2014) |
|
Xylopia sericea |
Leaves | Ethanolic extract | NO production in LPS-stimulated RAW264.7 cells | 76% | (Gomes et al. 2022) |
| Leaves | Ethanolic extract | IL-6 production in LPS-stimulated RAW264.7 cells | 85% | (Gomes et al. 2022) |
3.2. Insecticidal, Larvicidal and Pesticidal
Larvicidal, pesticidal and insecticidal activities are intricately linked, as they are commonly related to the external transmission of diseases or harm to the pestilent being. From now on, we will address the term “insecticide” to refer, in a global way, to larvicidal, insecticidal and pesticidal activities. Compounds classified as insecticides are those capable of killing, attracting and repelling insects (Klocke et al. 1991; Simões et al. 2003; Viegas 2003). However, an ideal insecticide must also be effective at low concentrations, non-toxic to mammals, people, or plants, easy to obtain, handle and apply, economically accessible, and non-cumulative in human and domestic animal adipose tissue (Klocke et al. 1991; Simões et al. 2003; Viegas 2003). All these properties describe a perfect insecticide candidate, which will rarely be found.
The use of plants and their isolated compounds for insecticidal activity has been used since ancient times (Viegas Júnior 2003; Krinski et al. 2014). They were widely used until the 1940s, when synthetic products began to gain space in the market. However, synthetic insecticides are shown to be extremely potent and, on the other hand, are quite unspecific, presenting several toxicity problems (Viegas 2003). In the search for insecticides that have high efficacy, safety, and selectivity, natural products have regained interest. There more than two thousand species of plants recorded in literature to have insecticidal properties. However, few of these species have been used commercially (Ndumu et al. 1999; Simões et al. 2003; Viegas 2003).
It has been found that the use of plant-based secondary metabolites are a low-cost alternative for pest control (Santos et al. 2010; Spletozer et al. 2021). Thus, the search for these compounds has intensified greatly in recent decades, with numerous publications and studies emphasizing the efficacy, along with the economic and ecological advantages of plant-based insecticides (Spletozer et al. 2021).
In this context, the Annonaceae family stands out for its structural variability of secondary metabolites, that are rich in alkaloids and acetogenins (Jossang et al. 1984; Spletozer et al. 2021). Acetogenins, a unique class of natural products found exclusively in Annonaceae, have demonstrated significant promise as prototypes for insecticidal agents (Viegas 2003; Castillo et al. 2010; Spletozer et al. 2021). Studies by Colom et al. (2008) and Alvarez et al. (2007) serve as notable examples of research evaluating the insecticidal potential of acetogenins derived from Annonaceae (Alvarez Colom et al. 2007; Colom et al. 2008).
The review article written by Krinski, Massaroli, and Machado, 2014, presents an analysis of the insecticidal potential of plants from the Annonaceae family, which are found across diverse tropical regions such as Central and South America, Asia and Africa. Despite the potential of Annonaceae as a source of natural insecticides, there is a lack of comprehensive studies on these plants for pest control, which can be partly attributed to the recent discovery of their biocidal properties against insects. However, growing concern about the adverse effects of synthetic insecticides on the environment and human health has prompted a resurgence in studies on botanical insecticides, aiming to obtain compounds that are less environmentally aggressive and avoid insect resistance.
The authors also highlight that Annonaceae metabolites are promising insecticidal substances, especially acetogenins. Approximately 42 species of Annonaceae have been studied for their insecticidal potential, primarily against the main orders of insects considered pests, such as Lepidoptera, Coleoptera, Hemiptera, Diptera, and Blattodea (Krinski et al. 2014).
Although there are promising studies demonstrating the potential of Annonaceae in pest control, further research is needed, especially under field conditions, to validate the results obtained in the laboratory. Additionally, it is essential to isolate the active compounds present in these plants to better understand their mechanisms of action and develop safer and more effective pest control methods (Krinski et al. 2014). Challenges related to the scarcity of plant resources and the lack of standardization and quality control of formulations aside, growing interest in organic products and the environmental impact of synthetic insecticides are driving this search for sustainable alternatives (Krinski et al. 2014).
Krinski, Massaroli, and Machado, 2014, in their review article, also created a table presenting information about the species of plants from the Annonaceae family studied regarding their insecticidal activity, the genera involved, the insect orders evaluated, and other relevant data for studying the insecticidal potential of these plants.
In this review, we updated the table from Krinski, Massaroli, and Machado, 2014, by adding information about activity values and methods used to evaluate the insecticidal, pesticidal, and larvicidal activity. Despite the plethora of promising results, most studies only assess essential oils, extracts, and fractions. Therefore, research is needed to isolate the secondary metabolites from the species already studied to evaluate them separately and confirm their insecticidal, larvicidal, pesticidal, and cytotoxic potential.
Annona genus
The Annona genus is the most extensively studied within the Annonaceae family for pesticide, larvicide, and insecticidal activity. Scientific reports cover 17 Annona species, providing evidence of insecticidal activity. Among these species, A. squamosa, A. muricata, A. coriaceae, and A. crassiflora emerge as the most investigated. The studies encompass a variety of approximately 41 species of insects, with about 14 research efforts specifically aimed at assessing activity against Aedes aegypti. These findings underscore the relevance of the genus Annona as a potential source of insecticidal and larvicidal compounds, especially concerning the control of disease-transmitting mosquitoes (Kawazu et al. 1989; Sinchaisri et al. 1991; Schmeda-Hirschmann and de Arias 1992; Epino and Chang 1993; Saxena et al. 1993; Monzon et al. 1994; Aku et al. 1998; Alali et al. 1998; Fontana et al. 1998; Guadaño et al. 2000; Kotkar et al. 2002; Leatemia and Isman 2004; LS et al. 2004; Morales et al. 2004; Pérez-Pacheco et al. 2004; Saito et al. 2004; Rao et al. 2005; Bobadilla et al. 2005; Khalequzzaman and Sultana 2006; Rodrigues et al. 2006, 2021; da Silva et al. 2007; de Omena et al. 2007; Henao et al. 2007; Alvarez Colom et al. 2007; Souza, E. M.; Cordeiro, J. R.; Pereira 2007; Coelho et al. 2007; Llanos et al. 2008; Colom et al. 2008; Dadang and Prijono 2009; Guarido 2009; Karunaratne and Arukwatta 2009; Magadula et al. 2009; Oliveira and Pereira 2009; Deshmukhe et al. 2010; Kumar et al. 2010; Araújo 2010; Begum et al. 2010; Toto Blessing et al. 2010; Carneiro 2010; de Cássia Seffrin et al. 2010; Cruz 2011; de Moraes et al. 2011; Kamaraj et al. 2011; Kempraj and Bhat 2011; Sharma et al. 2011; Costa et al. 2012; Dill et al. 2012; González-Esquinca et al. 2012; Kesetyaningsih 2012; Sreeletha and Geetha 2012; Costa, Marilza da Silva, Mônica Josene Barbosa Pereira, Simone Santos de Oliveira, Paulo Teixeira de Souza, Evandro Luiz Dall’oglio 2013; Cruz-Estrada et al. 2013; Allison et al. 2013; Massaroli et al. 2013; Ribeiro et al. 2013; Da Silva et al. 2013b; Krinski and Massaroli 2014).
Others Annonaceae species
Other Annonaceae genera have also been investigated for insecticidal, larvicidal, and pesticidal activity, such as the Artabotrys (Kabir 2010), Asimina (Mikolajczak et al. 1988), Cardiopetalum (Costa, Marilza da Silva, Mônica Josene Barbosa Pereira, Simone Santos de Oliveira, Paulo Teixeira de Souza, Evandro Luiz Dall’oglio 2013), Dennetia (Ewete et al. 1996; Okonkwo and Okoye 2001; Akinwumi et al. 2007; Umeotok et al. 2013), Duguetia (Rodrigues et al. 2006; Luciana et al. 2013), Guatteria (Aciole et al. 2011), Mikilua (Odalo et al. 2005), Oxandra (Rojano et al. 2007), Rollinia (Tolosa et al. 2012), Uvaria (Anza et al. 2021) and Xylopia genus (Ewete et al. 1996; Rodrigues et al. 2006; Zaridah et al. 2006; Aina et al. 2009). Approximately 11 species of insects have been studied for the insecticidal potential of these genera of Annonaceae.
Table 2.
Summary of insecticidal, pesticidal and larvicidal activity of species of the Annonaceae.
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
|---|---|---|---|---|---|---|---|
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% CL - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona atemoya | Seed | Seed extract | Trichoplusia ni (Lep.) | 197.67 | 301.30 | (de Cássia Seffrin et al. 2010) | |
| Annona squamosa | Seed | Seed extract | Trichoplusia ni (Lep.) | 382.37 | 167.48 | (de Cássia Seffrin et al. 2010) | |
|
Annona cherimola |
Seed |
Squamocin Molvizarin Almunequin Itrabin Deltamethrin (Positive control) |
Oncopeltus fasciatus |
0.16 0.34 11.23 14.91 7.4 |
(Colom et al. 2008) | ||
| Annona cherimola | Seed | Neoannonin Itrabin Almunequin Asimicin Squamocin Motrilin Cherimolin-1 Cherimolin-2 Tucumanin Control |
Spodoptera Frugiperda | 15.5 18.8 19.7 17.3 Instant death 18.0 14.0 17.7 14.7 12.1 |
10 30 30 30 100 20 0 10 20 10 |
0.90 0.59 1.10 0.77 0.16 1.19 0.91 0.97 0.81 1.02 |
(Alvarez Colom et al. 2007) |
| Annona coriaceae |
Seed |
Seed Extract |
Aedes aegypti (Dip.) |
0.01 |
- |
- |
(Costa et al. 2012) |
| Annona coriaceae | Seed | 100 ppm 50 ppm DMSO 0.1% Água (Control) |
Aedes aegypti (Dip.) | 0.50 0.40 0.30 0.20 |
3.00 6.00 - - |
3.5 5.5 - - |
(Dill et al. 2012) |
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods |
Ref. |
||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% CL - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona coriaceae | Seed | Seed Extract 0 ppm 50 ppm 100 ppm 200 ppm 300 ppm 400 ppm 500 ppm |
Aedes aegypti (Dip.) |
0.0 0.7 0.7 1.0 2.2 4.5 6.2 |
0.0 7.5 7.5 10.0 22.5 45.0 62.5 |
10.0 9.25 9.25 9.00 7.75 5.50 3.75 |
(de Moraes et al. 2011) |
| Annona coriaceae | Seed | Seed Extract Methanol (des. Hexane) Hexane Dichloromethane Methanol (des. DCM) |
Aedes aegypti (Dip.) |
0.1 0.1 0.1 0.1 |
100.0 100.0 58.75 0.0 |
0.007 0.007 0.805 0.0 |
(Costa, Marilza da Silva, Mônica Josene Barbosa Pereira, Simone Santos de Oliveira, Paulo Teixeira de Souza, Evandro Luiz Dall’oglio 2013) |
| Annona coriaceae | Seed | Diet | Anagasta kuehniella (Lep.) | 0.0 2.0 |
26.3 16.8 |
71.8 81.9 |
(Coelho et al. 2007) |
| Annona coriaceae | Seed | Diet | Corcyra Cephalonica (Lep.) | 0.0 2.0 |
23.16 32.25 |
69.7 49.3 |
(Coelho et al. 2007) |
| Annona coriaceae | Seed | Seed Extract Hexanic 8.0% 4.0% 2.0% 1.0% 0.5% Methanolic 8.0% 4.0% |
Dichelops melacanthus (Hem.) |
78.00 86.00 68.00 58.00 42.00 96.00 |
(Souza, E. M.; Cordeiro, J. R.; Pereira 2007) | ||
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% CL - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona coriaceae | Seed | Seed Extract Methanolic 2.0% 1.0% 0.5% Ethanolic 8.0% 4.0% 2.0% 1.0% 0.5% Distilled water (Positive Control) C 01 C 02 |
94.00 94.00 70.00 40.00 100.00 100.00 90.00 84.00 80.00 6.00 4.00 6.00 12.00 0.00 2.00 |
(Souza, E. M.; Cordeiro, J. R.; Pereira 2007) | |||
| Annona coriaceae | Seed | Seed Extract Preview 2 Days 5 Days 7 Days DMSO 20% (Positive Control 01) Water (Positive Control 02) |
Euschistus heros (Hem.) |
3.0 4.6 3.4 3.7 3.1, 5.0, 3.4 and 5.0 4.1, 4.4, 2.8 and 5.2 |
- 8.91 - 26.73 - - |
(Da Silva et al. 2013b) | |
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% CL - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona coriaceae | Seed | Seed Extract Methanolic 0.5 1.0 2.0 4.0 8.0 DMSO 20% Water |
Tuta absoluta (Lep.) |
- - - - - - - |
8.0 100 100 86.4 86.6 6.6 13.2 |
- - - - - - - |
(da Silva et al. 2007) |
| Annona cornifolia | Leaves | Leave extract 2.5 2.0 1.5 1.0 0.5 |
Anticarsia gemmantalis (Lep.) |
- - - - - |
0.41 0.38 -0.10 -0.25 0.01 |
- - - - - |
(Saito et al. 2004) |
| Annona cornifolia | Leaves | Leave extract 2.5 2.0 1.5 1.0 0.5 |
Spodoptera frugiperda (Lep.) |
- - - - - |
0.96 0.68 0.55 0.66 0.36 |
- - - - - |
(Saito et al. 2004) |
| Annona cornifolia | Leaves | Leave extract 2.5 2.0 1.5 1.0 0.5 |
Spodoptera frugiperda (Lep.) |
- - - - - |
0.96 0.68 0.55 0.66 0.36 |
- - - - - |
(Saito et al. 2004) |
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% CL - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona cornifolia | Leaves | Leave extract 2.5 2.0 1.5 1.0 0.5 |
Spodoptera frugiperda (Lep.) |
- - - - - |
0.96 0.68 0.55 0.66 0.36 |
- - - - - |
(Saito et al. 2004) |
| Annona Crassiflora | Fruits/ Twigs/ Roots | Extract Hexanic SB RW RB Ethanolic RW RB |
Aedes aegypti (Dip.) |
192.57 154.02 264.15 26.89 23.06 |
- - - - - |
- - - - - |
(Rodrigues et al. 2006) |
| Annona Crassiflora | Roots | Extract Root bark Root wood Stem |
Aedes aegypti (Dip.) |
0.71 8.94 16.1 |
- - - |
- - - |
(de Omena et al. 2007) |
| Annona Crassiflora | Seeds | Seeds Extract Methanol (Des. DCM) Hexanic Hydroalcoholic Fraction Ethyl Acetate |
Aedes aegypti (Dip.) |
1.0 1.0 1.0 1.0 |
0.0 91.25 0.0 0.0 |
- 0.433 - - |
(Costa, Marilza da Silva, Mônica Josene Barbosa Pereira, Simone Santos de Oliveira, Paulo Teixeira de Souza, Evandro Luiz Dall’oglio 2013) |
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% CL - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona Crassiflora | Seeds | Fraction Chloroform Crude Methanolic |
Aedes aegypti (Dip.) |
1.0 1.0 |
0.0 11.25 |
- 3.189 |
(Costa, Marilza da Silva, Mônica Josene Barbosa Pereira, Simone Santos de Oliveira, Paulo Teixeira de Souza, Evandro Luiz Dall’oglio 2013) |
| Annona Crassiflora | Seeds | Seeds Extract 1% 2% 4% DMSO 40% (Positive control) |
Euschistus heros (Hem.) |
481.50 542.00 372.00 683.00 |
1.25 1.50 2.00 1.25 |
353.00 396.00 306.00 309.20 |
(Oliveira and Pereira 2009) |
| Annona Crassiflora | Seeds | Seeds Extract Preview 2 Days 5 Days 7 Days DMSO 20% (Positive control 01) Water (Positive control 02) |
Euschistus heros (Hem.) |
2.4 4.1 3.0 4.2 3.1 5.0 3.4 5.0 4.1 4.4 2.8 5.2 |
17.82 13.04 16.83 - - - - - - - - |
(Da Silva et al. 2013b) |
|
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% CL - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona Crassiflora | Seeds | Seed Extract 24 h 8.0% 4.0% 2.0% 1.0% 0.5% 72 h 8.0% 4.0% 2.0% 1.0% 0.5% 120h 8.0% 4.0% 2.0% 1.0% 0.5% Water + Tween 80 (Positive Control 01) Water (Positive Control 02) |
Tibraca limbativentris (Hem.) |
70.0 64.0 44.0 20.0 6.0 78.0 72.0 54.0 30.0 10.0 81.0 76.0 58.0 34.0 10.0 2.0 4.0 9.0 0.0 2.0 6.0 |
4.46 3.30 |
4.34 3.16 |
(Krinski and Massaroli 2014) |
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% CL - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona dioica | Seeds | Seeds Extract Fraction Dichloromethane Methanol |
Aedes Aegypti (Dip.) |
1.0 1.0 |
10.00 3.75 |
2.447 5.196 |
(Costa, Marilza da Silva, Mônica Josene Barbosa Pereira, Simone Santos de Oliveira, Paulo Teixeira de Souza, Evandro Luiz Dall’oglio 2013) |
| Annona dioica | Seeds | Seed Extract Topical application method 25 50 100 200 Cantate application method 25 50 100 200 |
Rhodnius neglectus (Hem.) |
6.2 85 90 100 88.2 91.6 95.6 96.0 |
(Carneiro 2010) | ||
| Annona diversifolia | Leaves/ Branches | Extract Stem Aqueous Ethanolic Leaf Aqueous Ethanolic |
Anastrepa ludens (Dip.) |
588.685 409.139 >1000 52.0284 |
(González-Esquinca et al. 2012) | ||
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods |
Ref. |
||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% CL - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona foetida | Seeds | Seed Extract Methanolic 24h 48h |
Aedes aegypti (Dip.) |
76.15 62.28 |
(Guarido 2009) |
||
| Annona foetida | Seeds | Hexanic 24h 48h Dichloromethanic 24h 48h |
Aedes aegypti (Dip.) |
15.17 6.72 0.73 0.33 |
(Guarido 2009) |
||
| Annona glabra | Seeds | Seed Extract | Aedes aegypti (Dip.) | 0.06 | (de Omena et al. 2007) | ||
| Annona montana | Leaves/ Branches | Annonacin Cis-annonacin-10-one Densicomacin-1 Gigantetronenin Murihexocin-B Tucupentol Control |
Spodoptera frugiperda (Hem.) |
49.20 44.75 60.00 55.20 55.12 59.00 27.12 |
50 60 40 70 30 30 0.0 |
50 40 60 30 70 70 10 |
(Toto Blessing et al. 2010) |
| Annona mucosa | Fruits and branches | Ethanolic extract Rollinicin Rolliniastacin-1 |
Aedes aegypti |
2.60 0.78 0.43 |
- |
(Rodrigues et al. 2021) |
|
| Annona mucosa | Fruits and branches | Ethanolic extract Rollinicin Rolliniastacin-1 |
Aedes albopictus |
0.55 1.13 0.20 |
- |
(Rodrigues et al. 2021) |
|
| Annona mucosa | Seeds | Seed Extract 0.5 1.0 2.0 |
Chrysodeixis includens (Lep.) |
< 55% < 55% < 55% |
(Massaroli et al. 2013) |
||
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods |
Ref. |
||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% CL - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona mucosa | Seeds | Seed Extract 4.0 8.0 |
Chrysodeixis includens (Lep.) |
86.6% 93.3% |
(Massaroli et al. 2013) | ||
| Annona mucosa | Seeds/ Branches/ Leaves | Extract Seeds 300 mg kg 1500 mg kg Leaves 300 mg kg 1500 mg kg Branches 300 mg kg 1500 mg kg Control |
Sitophilus zeamais (Col.) |
0.80 0.00 36.80 5.60 34.70 39.10 36.90 37.00 |
98.00 100.00 0.50 61.50 1.50 0.00 0.00 0.00 |
7.84 1.12 81.97 57.90 83.71 64.97 81.94 81.74 |
(Ribeiro et al. 2013) |
| Annona mucosa | Seeds | Seed Extract 24h 8.0% 4.0% 2.0% 1.0% 0.5% 72h 8.0% 4.0% 2.0% |
Timbraca limbativentris (Hem.) |
100.0 92.0 90.0 76.0 28.0 100.0 96.0 96.0 |
1.59 1.18 |
0.49 -0.24 |
(Krinski and Massaroli 2014) |
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods |
Ref. |
||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona mucosa | Seeds | 1.0% 0.5% 120h 8.0% 4.0% 2.0% 1.0% 0.5% Water + Tween80 (Positive Control 01) Water (Positive Control 02) |
Timbraca limbativentris (Hem.) |
86.0 42.0 100.0 98.0 96.0 88.0 56.0 2.0 4.0 9.0 0.0 2.0 6.0 |
0.91 |
-0.76 |
(Krinski and Massaroli 2014) |
| Annona muricata | Seeds | Seed Extract | Aedes aegypti (Dip.) | 236.23 | 74.68 | - | (Morales et al. 2004) |
| Annona muricata | Seeds | Seed Extract 12h 24h 36h 48h |
Aedes aegypti (Dip.) |
0.18 0.06 0.04 0.02 |
0.10 0.05 0.03 0.01 |
(Bobadilla et al. 2005) |
|
| Annona muricata | Seeds | Seed Extract | Aedes aegypti (Dip.) | 900.0 | 380.0 | (Henao et al. 2007) | |
| Annona muricata | Leaves/ Branches | Extract Leaves Ethanolic Aqueous |
Anastrepha ludens (Dip.) |
831.445 >1000 |
2058.3 3852.6 |
(González-Esquinca et al. 2012) | |
| Annona muricata | Leaves/ Branches | Stems Ethanolic Aqueous |
Anastrepha ludens (Dip.) |
865.0 >1000 |
4539 3984.2 |
(González-Esquinca et al. 2012) |
|
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods |
Ref. |
||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona muricata | Seed | Seed extract | Anopheles albimanus (Dip.) | 16.20 | 0.82 | (Morales et al. 2004) | |
| Annona muricata | Seed | Food consumption (%) Low dose Medium dose Water (Positive Control 01) 10% Ethanol (Positive Control 02) |
Anticarsia gemmantallis (Lep.) |
10.0 30.0 0.0 0.0 |
25.0 29.3 19.9 19.3 |
(Fontana et al. 1998) |
|
| Annona muricata | Seed | Parviflorin Asimicin Sylvaticin Bullatalicin Annomontacin Gigantetrocin A Cypermethrin Chlorpyrifos Hydramethylnon Propoxur Bendiocarb |
Blatella germanica (Blat.) |
0.6 1.8 1.5 6.5 3.6 4.1 0.003 0.3 5.6 39.9 43.2 |
6 10 8 23 23 34 6 3 12 - - |
(Alali et al. 1998) |
|
| Annona muricata | Leaves | Leaves Extract |
Culex Quinquefascintus |
20.87 | 56.47 | (Magadula et al. 2009) | |
| Annona muricata | Seed | Compounds 1 2 Rotenone |
Leptinotarsa dercemlineata (Col.) |
92.18 29.68 100 |
(Guadaño et al. 2000) |
||
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods |
Ref. |
||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona muricata | Seeds | Seed Extract | Plutella xylostella (Lep.) | 43.0 | 60.0 | (Sinchaisri et al. 1991) | |
| Annona muricata | Seeds | Seed Extract Hexanic 24h 48h 72h Ethyl Acetathe 24h 48h 72h |
Sithophillus zeamais (Col.) |
11.447 - - - - - |
4.009 3.854 3.760 3.280 2.667 2.542 |
(Llanos et al. 2008) |
|
| Annona muricata | Seeds | Seeds Extract | Zabrotes subsfasciattus (Col.) | 46.0 | 39.1 | 36.4 | (Araújo 2010) |
| Annona reticulata | Seeds | Seed Extract (95% of Methanol) In two periods: 24h and 48h. g/l 2.5 g/l 5.0 g/l 7.5 g/l 10.0 g/l 15.0 g/l 20.0 g/l |
Epilachna vigintioctopunctata (Col.) |
(%) 24h and 48h 6.7 - 13.4 40.0 – 53.4 80.0 – 100 100 – 100 100 – 100 100 – 100 100 – 100 |
(Karunaratne and Arukwatta 2009) |
||
| Annona reticulata | Uninformed | Petroleum ether extract Ethanolic extract |
Rhodnius neglectus (Hem.) |
35.0 Not significant |
(Schmeda-Hirschmann and de Arias 1992) |
||
| Annona reticulata | Seeds | Methanolic extract | Spodoptera litura (Lep.) |
301.30 (259.15-326.33) |
50.0% |
167.48 (110.43-383.65) |
(de Cássia Seffrin et al. 2010) |
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods |
Ref. |
||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona salzmannii | Barks |
Hexanic Extract (8,68 g (0,48%) Methanolic Extract 143,29 g (7,96%) CHCl3 alkaloid fraction – FCA |
Aedes aegypti (Dip.) |
615.18 >700.00 163.53 |
(473.71 -981.44) (00.00-00.00) (107.90-238.82) (13.31-19.40) (130.00-218.00) (0.035-0.050) |
(Cruz 2011) |
|
| Annona salzmannii | Barks | Neutral CHCl3 fraction – FCN caryophyllene oxide Temephos |
Aedes aegypti (Dip.) |
15.92 167.00 0.042 |
(Cruz 2011) |
||
| Annona senegalensis | Root | Root Extract Control |
Callosobruchus maculatus (Col.) | 18.7 4.7 |
3.7 98.0 |
0.1 79.5 |
(Aku et al. 1998) |
| Annona senegalensis | Fruits | Ethanolic extract | Culex quinquefascintus (Dip.) | 0.67 | 23.42 | 29.78 |
(Magadula et al. 2009) |
| Annona senegalensis | Uninformed | Extract | Sitophilus zeamais (Col.) | 220.71 | 0.19 – 0.06 | (LS et al. 2004) | |
| Annona squamosa | Leaves | Aqueous extract (g/100 ml) 100.0 50.0 25.0 12.5 6.25 3.125 1.5625 Control |
Aedes aegypti (Dip.) |
100.0 100.0 83.3 90.0 70.0 76.6 73.3 5.7 |
(Monzon et al. 1994) | ||
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods |
Ref. |
||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona squamosa | Seeds | Extract (µg/ml) 1 |
Aedes albopictus (Dip.) |
388.3 |
397.4 |
486.6 |
(Kempraj and Bhat 2011) |
| Annona squamosa | Seeds | Extract (µg/ml) 2 4 6 8 10 20 |
Aedes albopictus (Dip.) |
231.3 162.3 114.0 64.9 45.6 0.0 |
240.3 163.8 112.0 72.4 47.8 0.0 |
268.5 185.4 121.1 84.9 56.3 0.0 |
(Kempraj and Bhat 2011) |
| Annona squamosa | Leaves | Ethanol extract (mg/ml) at 24h, 48h and 72h. 5 10 20 30 |
Anopheles gambiae (Dip.) |
3.33 (24h) 6.67 (48h) 23.33 (72h) 16.67 (24h) 33.33 (48h) 63.33 (72h) 40.0 (24h) 66.67 (48h) 76.67 (72h) 53.33 (24h) 73.33 (48h) 90.0 (72h) |
(Allison et al. 2013) |
||
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods |
Ref. |
||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona squamosa | Leaves | Ethanol extract (mg/ml) at 24h, 48h and 72h. 40 |
Anopheles gambiae (Dip.) |
70.0 (24h) 90.0 (48h) 1000 (72h) |
(Allison et al. 2013) | ||
| Annona squamosa | Whole plant | Extract (ppm) 50 100 150 200 |
Anopheles stephensi (Dip.) | (%) 58 60 70 74 |
(%) 4 6 16 18 |
(%) 52 76 86 92 |
(Saxena et al. 1993) |
| Annona squamosa | Leaves | Extract (mgl) 500 250 125 62.5 31.25 15.63 7.82 |
Anopheles subpictus (Dip.) | (%) 100 – 0.0 82.6 – 2.46 63.0 – 1.84 48.2 – 4.62 16.4 – 2.04 92.0 – 3.28 4.6 – 4.60 |
(Kamaraj et al. 2011) | ||
| Annona squamosa | Leaves | Ethanolic extract Aqueous extract |
Bemisia tabaci (Hem.) |
100 – 0.0 99.3 – 1.05 |
(Cruz-Estrada et al. 2013) | ||
| Annona squamosa | Seeds | Extract (mg/ml) 0.01 0.03 0.05 |
Callasobruchus chinensis (Col.) | (%) 9.66 9.66 41.00 |
(Kotkar et al. 2002) | ||
| Annona squamosa | Seeds | Extract (mg/ml) 0.07 0.09 |
Callasobruchus chinensis (Col.) | % 81.33 99.00 |
(Kotkar et al. 2002) | ||
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona squamosa | Seeds | Extract (mg/ml) 0.07 0.09 |
Callasobruchus chinensis (Col.) | % 81.33 99.00 |
(Kotkar et al. 2002) | ||
| Annona squamosa | Seeds | Extract (µg/cm2) Petroleo ether Ethanol Acethone Methanol |
Ceratitis capitata (Dip.) | (%) 0.031 0.632 0.591 4.038 |
(%) 198.57 614.26 1000.40 135.25 |
(Epino and Chang 1993) | |
| Annona squamosa | Seeds | Extract (%) 0.05 0.1 Deltamethrin |
Crocidiolomia pavonana (Lep.) |
10.4 ± 1.2 6.3 ± 2.7 11.0 ± 3.6 |
32.0 ± 6.9 69.0 ± 4.8 62.4 ± 5.2 |
(Dadang and Prijono 2009) | |
| Annona squamosa | Seeds |
Aqueous extract (25%) |
Culex quinquefasciatus (Dip.) |
33.6% |
(Pérez-Pacheco et al. 2004) | ||
| Annona squamosa | Leaves | Ethanol extract (mg/ml) at 24h, 48h and 72h. 5 10 |
Culex quinquefasciatus (Dip.) | (%) 0.0 (24h) 13.33 (48h) 46.67 (72h) 33.33 (24h) 56.67 (48h) 73.33 (72h) |
(Allison et al. 2013) | ||
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona squamosa | Leaves | Ethanol extract (mg/ml) at 24h, 48h and 72h. 20 30 40 |
Culex quinquefasciatus (Dip.) |
56.67 (24h) 70.0 (48h) 90.0 (72h) 80.0 (24h) 96.67 (48h) 100 (78h) 93.33 (24h) 100 (48h) 1000 (72h) |
(Allison et al. 2013) | ||
| Annona squamosa | Leaves | Aqueous extract (g/100 ml) 100.0 50.0 25.0 12.5 6.25 3.125 1.5625 |
Culex quinquefasciatus (Dip.) | (%) 100.0 60.0 50.0 36.7 26.7 33.3 10.0 |
(Monzon et al. 1994) | ||
| Annona squamosa | Leaves | Extract | Culex quinquefasciatus (Dip.) | 0.64 | 14.69 | (Magadula et al. 2009) | |
| Annona squamosa | Leaves | Methanol Extract (mg/l) 500 |
Culex tritaeniorhynchus (Dip.) | (%) 100.0 ± 00.0 |
(Kamaraj et al. 2011) | ||
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona squamosa | Leaves | Methanol Extract (mg/l) 250 125 62.5 31.25 15.63 7.82 |
Culex tritaeniorhynchus (Dip.) | (%) 88.4 ± 1.64 52.6 ± 4.63 34.2 ± 2.84 20.6 ± 1.67 12.4 ± 2.45 6.8 ± 1.87 |
(Kamaraj et al. 2011) | ||
| Annona squamosa | Seeds | Diet (µg/2g) 48h |
Drosophila melanogaster (Dip.) |
62.5 |
(Kawazu et al. 1989) | ||
| Annona squamosa | Seeds | Extract (g/l) 1.0 2.5 5.0 10.0 15.0 20.0 |
Epilachna vigintioctopunctata (Col.) | 24h and 48h (%) 6.7 and13.4 40.0 and 53.4 80.0 and 53.4 100 and 100 100 100 |
(%) 64.8 83.4 92.3 95.9 95.9 100.0 |
(Karunaratne and Arukwatta 2009) | |
| Annona squamosa | Branches | Extract (%) | Musca domestica (Dip.) | 41.00 | (Sharma et al. 2011) | ||
| Annona squamosa | Leaves | Extract (mg/l) 0 200 400 600 800 1000 |
Musca domestica (Dip.) |
(%) 100 80 65 50 30 0 |
(%) 100 62.5 53.85 40.0 33.33 0.0 |
(Begum et al. 2010) | |
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona squamosa | Leaves | Extract (% plant poder) 5 10 20 |
Oryctes rhinoceros (Col.) | (%) 10 30 50 |
(%) 10 20 20 |
(Sreeletha and Geetha 2012) | |
| Annona squamosa | Leaves | Extract (%) 100 75 50 25 10 5 0.1 |
Periplaneta americana (Blat.) | (%) 80 60 50 20 10 10 0 |
Average 4.00 ± 0.0 3.00 ± 0.0 2.5 ± 0.71 1.00 ± 0.0 0.5 ± 0.701 0.5 ± 0.701 0.0 ± 0.0 |
(Kesetyaningsih 2012) | |
| Annona squamosa | Seeds | Extract (mg/ml) 5 10 |
Plutella xylostella (Lep.) | (%) 46.7 70.0 |
(Sinchaisri et al. 1991) | ||
| Annona squamosa | Seeds | Aqueous extract Larval instar (Time h) 3rd: 24h 48h 72h 4th: 24 48 72 |
Plutella xylostella (Lep.) | (%) 5.2 (3.1-8.5); 1.7 (1.3-2.2); 0.9 (0.7-1.2). 8.7 (6.6-11.3); 4.2 (3.5-5.1); 2.0 (1.7-2.4). |
(%) 2.5 ± 1.4 10.0 ± 6.8 12.5 ± 6.0 0 1.3 ± 1.3 5.0 ± 2.0 |
(Leatemia and Isman 2004) | |
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods |
Ref. |
||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona squamosa | Seeds | Aqueous extract 1% 5% Control 1 (Acetone) Control 2 (Methanol) Control 3 (Without solvente) |
Sitophilus oryzae (Col.) | (LD 50 Min) 23.1 (22.1-23.9) 11.4 (10.7-12.2) 0.0 0.0 0.0 |
(% min) 39.6±1.4 14.5±1.1 - - - |
(Kumar et al. 2010) | |
| Annona squamosa | Seeds | Extract (%) 0.5 |
Spodoptera litura (Lep.) |
21.66±1.66 |
0.0 |
28.33±1.66 |
(Deshmukhe et al. 2010) |
| Annona squamosa | Seeds | Extract (%) 1 5 10 15 20 25 |
Spodoptera litura (Lep.) |
23.33±1.66 38.33±1.66 48.33±1.66 56.66±1.66 51.66±1.66 61.66±1.66 |
0.0 0.0 0.0 0.0 1.66±1.66 1.66±1.66 |
33.33±1.66 51.66±1.66 58.33±1.66 78.33±3.33 75.00±0.0 80.00±0.0 |
(Deshmukhe et al. 2010) |
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona squamosa | Seeds | Extract Petroleum ether EtOH Acetone Methanol |
Tribolium castaneum (Col.) | LD50 (µg/cm2) 0.031 0.632 0.591 4.038 |
95% (Lower and Upper) 0.006 and 0.150; 0.315 and 1.265; 0.285 and 1.224; 1.727 and 9.440. |
(Khalequzzaman and Sultana 2006) | |
| Annona squamosa | Seeds | Extracts using two methods of application: Topical (µg/larva) Oral (ppm fresh weight in diet. |
Trichoplusia ni (Lep.) |
301.30 (259.15-326.33) |
167.48 (110.43-383.65) |
(de Cássia Seffrin et al. 2010) | |
| Annona squamosa | Seeds | Extract (ppm) Hexane extract 250 500 750 1000 1250 1500 Ethyl Acetate extract 50 250 500 750 1000 1250 |
Trogoderma granarium (Dip.) | (%) 10th day and 15th day 0.0 and 11.13 4.47 and 11.13 11.13 and 17.8 17.8 and 28.87 48.9 and 53.33 75.33 and 82.2 4.47 and 20.00 24.5 and 33.33 26.7 and 55.53 51.13 and 57.8 64.47 and 80.0 84.5 and 91.13 |
(mg) 10th day and 15th day. 66.1 and 80.5 75.1 and 84.6 64.9 and 70.9 63.6 and 71.8 64.9 and 70.1 58.3 and 61.5 73.0 and 86.2 65.6 and 73 72.4 and 81.0 62.9 and 69.5 59.8 and 66.0 56.0 and 61.0 |
(Rao et al. 2005) | |
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Annona squamosa | Seeds | Extract (ppm) Methanol extract 500 750 1000 1250 1500 2000 Acetone control Control |
Trogoderma granarium (Dip.) | (%) 10th day and 15th day 6.67 and 8.87 13.34 and 17.8 22.2 and 24.47 20.0 and 26.67 48.87 and 57.7 66.67 and 77.7 0.0 and 0.0; 0.0 and 0.0. |
(mg) 10th day and 15th day. 78.0 and 96.6 74.1 and 93.1 75.2 and 98.2 74.6 and 92.4 58.0 and 85.0 53.4 and 62.5 100.9 and 150.6. 94.9 and 161.6. |
(Rao et al. 2005) | |
| Artabotrys odoratissimus | Bark | Larval instar and exposure periods (h) Second 12h 24h |
Culex quinquefascintus (Dip.) | LC50 52.92 42.03 |
95% (Lower and Upper) 33.59 and 83.87 26.18 and 67.47 |
(Kabir 2010) | |
| Artabotrys odoratissimus | Bark | Larval instar and exposure periods (h) Third 12h 24h Fouth 12h 24h |
Culex quinquefascintus (Dip.) |
110.03 99.13 170.12 110.41 |
72.51 and 166.9 60.2 and 163.29 137.6 and 210.26 89.6 and 135.95 |
(Kabir 2010) | |
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Asimina triloba | Roots | Ethanolic extract (Fraction nº.) 017 018 019 020 021 Asimicin |
Acalymma vittatum (Col.) | LC50 (p.p.m.) 7.56 >1000 1.67 0.04 715 0.03 |
(Mikolajczak et al. 1988) |
||
| Cardiopetalum calophyllum | Seeds |
Methanolic extract (1.0 mg/mL) |
Aedes aegypti (Dip.) | (%) 5.00 |
(mg/mL) 1.789 |
(Costa, Marilza da Silva, Mônica Josene Barbosa Pereira, Simone Santos de Oliveira, Paulo Teixeira de Souza, Evandro Luiz Dall’oglio 2013) | |
| Dennettia tripetala | Leaves and roots | Ethanol extract (5mL/100g) in 1, 3 and 7 days. 0.0 |
Dermestes maculatus (Col.) | (%) In 1, 3 and 7 days. 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0 |
(Akinwumi et al. 2007) | ||
| Dennettia tripetala | Leaves and roots | Ethanol extract (5mL/100g) in 1, 3 and 7 days. 2.50 5.00 |
Dermestes maculatus (Col.) | (%) In 1, 3 and 7 days. 26.67 ± 0.88 71.67 ± 0.88 100.0 ± 0.0 26.67 ± 0.67 75.0 ± 1.16 100.0 ± 0.0 |
(Akinwumi et al. 2007) | ||
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Dennettia tripetala | Leaves and roots | Ethanol extract (5mL/100g) in 1, 3 and 7 days. 7.50 10.0 |
Dermestes maculatus (Col.) | (%) In 1, 3 and 7 days. 48.33 ± 0.88 91.67 ± 0.33 100.0 ± 0.0 51.67 ± 0.66 98.33 ± 0.33 100.0 ± 0.0 |
(Akinwumi et al. 2007) | ||
| Dennettia tripetala | Seeds | Concentration of plant powder/25g fish Extract Untreated control |
Dermestes maculatus (Col.) | (%) 87.0 100 |
(g) 0.25 0.50 |
(Okonkwo and Okoye 2001) | |
| Dennettia tripetala | Uninformed | Extracts (ppm) 0 10 100 1000 |
Ostrinia nubialis (Lep.) |
5.79 3.13 3.87 3.84 |
(Ewete et al. 1996) | ||
| Dennettia tripetala | Seeds | Extract (ml/25g) 0.1 0.2 0.3 |
Necrobia rufipes (Col.) | (%) 100 100 100 |
(Okonkwo and Okoye 2001) | ||
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Dennettia tripetala | Leaves and roots | Extract (%) at 24h, 48h and 72h. 0% |
Sitophilus zeamais (Col.) | (%) at 24h, 48h and 72h. 1.83 1.09 1.75 |
(Umeotok et al. 2013) | ||
| Dennettia tripetala | Leaves and roots | Extract (%) at 24h, 48h and 72h. 1% 5% 10% |
Sitophilus zeamais (Col.) | (%) at 24h, 48h and 72h. 1.50 2.09 2.58 1.50 2.09 3.08 2.83 3.58 4.67 |
(Umeotok et al. 2013) | ||
| Duguetia furfuraceae | Leaves/barks and roots | Extract | Aedes aegypti (Dip.) | (µg/ml) 56.6 |
(Rodrigues et al. 2006) | ||
| Duguetia furfuraceae | Leaves | Extract | Sitophilus zeamais (Col.) | (Proportion of insects in the treated area (SDM)) 0.500 (0.113) |
(Luciana et al. 2013) | ||
| Guatteria blephrophylla | Leaves | Extract (ppm) | Aedes aegypti (Dip.) | 85.74 | (74.05 – 112.78) | 4.48±0.89 | (Aciole et al. 2011) |
| Guatteria friesiana | Leaves | Extract (ppm) | Aedes aegypti (Dip.) | 52.60 | (50.11 – 55.17) | 6.48±0.55 | (Aciole et al. 2011) |
| Guatteria hispida | Leaves | Extract (ppm) | Aedes aegypti (Dip.) | 85.74 | (74.05 – 112.78) | 4.48±0.89 | (Aciole et al. 2011) |
| Mikilua fragans | Aerial parts | Essential oil | Anopheles gambiae (Dip.) |
RC75 (x10-5 mgcm-2) 481 (11, 145) |
(Odalo et al. 2005) | ||
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Oxandra cf xylopioides | Leaves | Extract 24h 48h 72h |
Spodoptera frugiperda (Lep.) | ppm 319.61 311.47 294.13 |
(Rojano et al. 2007) | ||
| Rollinia occidentalis | Seeds | Extract (ppm) 50 100 250 |
Spodoptera frugiperda (Lep.) | (%) 5 35 50 |
(%) 30 45 50 |
(%) - 20 - |
(Tolosa et al. 2012) |
| Uvaria scheflerri |
Roots |
Essential oil 5% 1% 0.5% 0.1 |
Anisakis L3 |
- 5 - - |
(Anza et al. 2021) |
||
| Xylopia aethiopica | Leaves and Roots | Extract Ethanolic Water |
Anopheles gambiae (Dip.) | LC50 3.57 4.50 |
(%) 125 (34.72%) 106 (29.44%) |
(Aina et al. 2009) | |
| Xylopia aethiopica | Uninformed | Extract (ppm) 0 10 100 1000 |
Ostrinia nubilalis (Lep.) | (mg) 3.78 3.69 3.87 3.84 |
(Ewete et al. 1996) | ||
| Annonaceae species | Used Material | Substances / Extracts | Insect species (Order) | Methods | Ref. | ||
| Topical (LC50 µg/ larva) | Topical (LD50 – 95% LC - µg/nymph) | Oral (LC50 ppm fresh weight in diet) | |||||
| Xylopia aromatica | Leaves/Barks and Roots | Extract | Aedes aegypti (Dip.) | (µg/ml) 384.37 |
(Rodrigues et al. 2006) | ||
| Xylopia caudata | Leaves | Oil | Aedes aegypti (Dip.) | 29.83 (21.87-37.45) | 60.33 (48.04-82.47) | 4.19±0.69 | (Zaridah et al. 2006) |
| Xylopia ferruginea | Leaves | Oil | Aedes aegypti (Dip.) | 74.51 (68.39-86.52) | 106.45 (90.23-159.78) | 8.27±1.88 | (Zaridah et al. 2006) |
3.3. Antimicrobial
Microbial diseases are infections caused by invading microorganisms or imbalances in the individual’s microbiota due to low immunity, overpopulation of a certain microorganism, or the presence of bacteria outside its natural habitat. The microbiome of the human body is extremely important, coexisting in a mutual relationship where it protects the host from invading microorganisms. Microorganisms in the microbiota can produce important nutrients, such as the production of vitamin K and B, and contribute to the development of the immune system (Rabêlo et al. 2014; Mendes et al. 2020).
Pathogenic bacteria, those that can cause disease in humans, can be classified into three different groups on account of the composition of their cell wall: Gram-positive bacteria have a thick layer of peptideoglycan on their cell wall and after Gram stain it takes on a violet color; Gram-negative bacteria have a thin layer of peptideoglycan on their cell wall and an outer portion of lipopolysaccharide and lipoproteins so when stained by the Gram method they assume a pink color; and, finally, acid-resistant bacteria, BAARS (acid-resistant bacilli), whose cell wall has complex lipids (mycolic acids) that obtain the primary dye even after subjected to acid-acid discoloration in the Ziehl Neelsen method (Fernandes 2000; Moreira et al. 2015; Mendes 2019).
There are also fungi that are present in the human microbiota and those that can be inhaled, but they do not cause damage to the organism so long as the natural defense barriers are intact. In this way, fungi have almost no infectious power until heavy virulence (Sidrim and Rocha 2004; Gomes et al. 2010; Barros 2014).
The most common bacterial and fungal infections are induced by Staphylococcus aureus, Staphylococcus epidermides, Escherichia coli, Pseudomonas aeruginosa, Salmonella Typhimarium, Enterococcus faecalis, Klebisiella pneumonieae, Candida krusei, Candida albicans and Candida tropicalis (Fernandes 2000; Moreira et al. 2015).
Infections were commonly fatal until antibiotics were discovered. In addition to being extremely important for the treatment and control of infectious diseases in humans and animals, antibiotics have made it possible to perform complex surgeries, intensive therapy, organ transplants, etc (Acar and Moulin 2012; Andrade 2018).
In the late 1930s, when antibiotics were introduced into therapy, mortality from pneumococcal pneumonia decreased by 20-40%, mortality from pneumococcal bacteremia by 50-80%, and mortality from severe infections such as bacterial meningitis and endocarditis, by 60-75%. Antibiotics have sparked a major revolution in medicine and countless lives have been saved since their discovery (Lepper and Dowling 1951; Breiman et al. 1990; Tomasz 1997; Ortqvist et al. 2005; Davies and Davies 2010; IOM 2010; Andrade 2018).
However, microorganisms have begun to show resistance to antibiotics in the wake of the influx in use. This has increased the spread of antibiotic-resistant bacteria and fungi, as well as the number of deaths from infections (Andrade 2018). According to the World Health Organization (WHO), bacterial and fungal resistance is characterized as a worldwide public health problem, whose simple strategies of combat will not be enough to ameliorate the emergence and spread of resistant infectious organisms (World Health Organization (WHO) 2014; Andrade 2018). Antimicrobial resistance (AMR) is a natural process for microorganisms, but it can be accelerated through the misuse of antibiotics such as the selective, repeated, or incomplete use of some antimicrobial agents (Neill 2016; Andrade 2018; Mendes et al. 2020).
AMR has become a major threat to the health of living beings, making treatment more difficult as well as increasing morbidity and mortality. Bacteria and fungi such as E. coli, S. aureus, P. aeruginosa, and C. albicans are examples of pathogenic microorganisms that show frequent microbial resistance. In 2020, WHO alerted the world to the shortage of new antimicrobial agents effective against super-resistant bacteria (Barreiras et al. 2020; Mendes et al. 2020; World Health Organization (WHO) 2020). Clinical research focused on the discovery and implementation of new antimicrobial agents has been greatly reduced in recent years. Natural products are rich sources of compounds with antimicrobial potential that need to be explored. Thus, the search for new plant-based antimicrobial agents is increasingly frequent (Neill 2016; Andrade 2018; Mendes et al. 2020).
Some species of the Annonaceae family have been studied for their antimicrobial profile against bacteria and fungi pathogenic to humans and plants. Table 3 describes the species of Annonaceae that have been studied for their antimicrobial profile in strains of bacteria and fungi most prevalent in human pathologies: S. aureus, S. epidermides, E. coli, P. aeruginosa, E. faecalis, K. pneumonie, C. krusei, C. albicans e C. tropicalis.
Annona genus
Several species of Annona have been extensively studied for their potential antimicrobial effects. One such species, A. hypoglauca Mart., underwent evaluation on two distinct alkaloid fractions. These fractions were assessed for their effectiveness against S. aureus (ATCC 29213), E. coli (ATCC 25922), and E. faecalis (ATCC 299212). The study revealed that the FA5 fraction, characterized by a higher concentration of isoboldine alkaloid, displayed noteworthy activity against S. aureus and E. faecalis, demonstrating a remarkable MIC value of 70 µg/mL for both microorganisms. Conversely, the FA6 fraction, enriched with actinodaphine alkaloid, exhibited antimicrobial efficacy against all three microorganisms: S. aureus, E. coli, and E. faecalis, with MIC values of 70 µg/mL, 90 µg/mL, and 80 µg/mL, respectively (Rinaldi et al. 2017).
Moreover, the methanol-methylene chloride extract from the root bark of A. senegalensis demonstrated potential antimicrobial activity, particularly against S. aureus and P. aeruginosa, with MIC values of 8750 µg/mL and 1080 µg/mL, respectively. The F1 subfraction extracted from these samples was found to consist of a diverse range of compounds, including kaur-16-en-19-oic acid, 1-dodecanol, 1-naphthalenemethanol, 6,6-dimethyl-bicyclo[3.1.1]hept-2-ene-2-ethanol, 3,3-dimethyl-2-(3-methylbuta-1,3-dienyl)cyclohexane-1-methanol, and 3-hydroxyandrostan-17-carboxylic acid. Impressively, the F1 subfraction exhibited significant activity against P. aeruginosa, with an MIC value of 40 µg/mL (Akah et al. 2012).
The antimicrobial potential of various Annona species was investigated by assessing the activity of their leaves. Notably, the essential oil extracted from A. foetida exhibited significant antimicrobial properties against S. aureus (ATCC 6538) and C. albicans (ATCC 10231), demonstrating MIC values of 200 µg/mL and 60 µg/mL, respectively (Costa et al. 2009).
In a parallel study, different samples of dried A. muricata leaf powder, varying in granulation, were subjected to evaluation against a range of microorganisms, including S. aureus (ATCC 25923), E. coli (ATCC 25922), P. aeruginosa (ATCC 27853), and K. pneumoniae (ATCC 4352). Notably, samples AM3 (with size bands between 0.149 and 0.074) and AM4 (size bands < 0.074) demonstrated the most promising results, with an MIC of 780 µg/mL against S. aureus, E. coli, and K. pneumoniae (de Andrade et al. 2019).
Furthermore, extracts derived from A. vepretorum leaves using different organic solvents such as ethanol, hexane, and chloroform were examined for their antimicrobial potential against S. aureus (ATCC 25923), E. coli (ATCC 25922), E. faecalis (ATCC 19433), and K. pneumoniae (ATCC 13883). Among these extracts, the hexanic extract exhibited noteworthy activity against E. coli, displaying an MIC value of 390 µg/mL (Almeida et al. 2014).
Additionally, a combination of two substances, 11-hydroxy-16-hentriacontanone and 10-hydroxy-hentricontanone, along with the isolated substance palmitone from A. squamosa leaves, was assessed for antimicrobial efficacy against S. aureus (ATCC 96) and P. aeruginosa (ATCC 741). These compounds exhibited notable antimicrobial effects, with palmitone showing a minimum inhibitory concentration (MIC) of 12.5 µg/mL against S. aureus and 6.25 µg/mL against P. aeruginosa (Shanker et al. 2007).
Guatteria genus
Extensive research has been conducted on numerous species within the Guatteria genus to explore their antimicrobial properties against human pathogens. In particular, the essential oils from the seeds of three species—Guatteriopsis hispida, Guatteriopsis blephaphylla, and Guatteriopsis friesiana—were evaluated for their efficacy against S. aureus (ATCC 6538), S. epidermidis (ATCC 12228), E. coli (ATCC 11775), and P. aeruginosa (ATCC 133388). Among these, the essential oil from G. friesiana proved most effective, with MIC values of 125 µg/mL against S. aureus, 100 µg/mL against S. epidermidis, and 900 µg/mL against both E. coli and P. aeruginosa. Isolated compounds from G. blepharophyla essential oil, such as β-eudesmol, γ-eudesmol, and α-eudesmol, also showed significant activity. Notably, γ-eudesmol exhibited strong antimicrobial activity against P. aeruginosa with an MIC of 300 µg/mL, while α-eudesmol demonstrated potential against both S. aureus and P. aeruginosa, with MIC values of 250 µg/mL and 200 µg/mL, respectively (Costa et al. 2008).
In a separates studies, essential oils extracted from the leaves of G. blepharophylla (synonym Guatteria blepharophylla), G. costaricensis, G. diospyroid, and G. oliviformis were evaluated for their antimicrobial properties against S. aureus (ATCC 6538 (Costa et al. 2011c; Alcântara et al. 2017), ATCC 29213(Palazzo et al. 2009)), E. coli (ATCC 8739 (Costa et al. 2011c; Alcântara et al. 2017) and ATCC 259222 (Palazzo et al. 2009)), P. aeruginosa (ATCC 9027 (Costa et al. 2011c; Alcântara et al. 2017)), and E. faecalis (ATCC 29212 (Costa et al. 2011c; Alcântara et al. 2017)). Particularly, G. blepharophylla essential oil exhibited promising activity against S. aureus and E. faecalis, with an MIC of 50 µg/mL. Another significant discovery involved the efficacy of G. diospyroid essential oil against S. aureus, with an MIC of 312 µg/mL (Palazzo et al. 2009). Additionally, the isomoschatoline alkaloid, isolated from G. blepharophylla leaves, displayed significant activity against C. albicans (ATCC 10231) with an MIC of 50 µM L-1, surpassing the control compound nystatin, which had an MIC of 54 µM L-1 (Costa et al. 2011c; Alcântara et al. 2017).
Furthermore, the hydroalcoholic extract obtained from Guatteria citriodora leaves exhibited potential activity against S. aureus, with an MIC of 250 µg/mL, while the alkaloid fraction displayed promising activity against S. epidermidis (ATCC 4083) ( MIC of 125 µg/mL) (Rabelo et al. 2014). However, the alkaloid fraction derived from the stem bark did not exhibit activity against either S. aureus or S. epidermidis (Rabelo et al. 2014).
Finally, essential oils extracted from the aerial parts of G. selewiana, G. latifolia, G. ferruginea, G. australis, and G. punctata were tested against E. coli microorganisms (ATCC 11775 (Santos et al. 2017) and CDC-EDL 933-171-0157: H3 (Bay et al. 2019a)), S. aureus (ATCC 33591) (Bay et al. 2019a), and P. aeruginosa (ATCC 29336) (Bay et al. 2019a). Among these, G. selewiana, G. latifolia, and G. ferruginea exhibited activity against E. coli (ATCC 11775), with an MIC of 600 µg/mL (Santos et al. 2017). Conversely, G. australis and G. punctata did not demonstrate activity against these microorganisms (Santos et al. 2017; Bay et al. 2019a).
Polyalthia genus
The species P. longifolia stands out as the most extensively studied within its genus, with antimicrobial activity investigations conducted on extracts derived from its stem bark, roots, and leaves. In stem bark studies, the butanol fraction displayed noteworthy MIC activity levels of 320 µg/mL against S. aureus (ATCC 29213 and MRSA 512), as well as E. coli (ATCC 29212), and an MIC of 160 µg/mL against P. aeruginosa. A compound isolated from the stem bark, 3-o-methyl ellagic acid, exhibited MIC values of 80 µg/mL against S. aureus (ATCC 29213), E. coli, and P. aeruginosa, while it exhibited an MIC of 160 µg/mL against MRSA 512 strains (Jain et al. 2014).
Root-based studies yielded promising results, where three alkaloids—pendulamine A, pendulamine B, and penduline—were isolated, exhibiting intriguing antimicrobial activity against S. aureus, P. aeruginosa, S. typhimurium, and K. pneumoniae. Pendulamine A demonstrated an impressive MIC of 0.2 µg/mL against S. aureus, 2 µg/mL against P. aeruginosa, 0.02 µg/mL against S. typhimurium, and 2 µg/mL against K. pneumoniae. Likewise, Pendulamine B was effective against S. aureus, S. typhimurium, and K. pneumoniae, with corresponding MICs of 0.2 µg/mL for both S. aureus and S. typhimurium, and 2 µg/mL for K. pneumoniae. In contrast, the alkaloid penduline exhibited targeted activity solely against S. aureus, achieving an MIC of 12.5 µg/mL (Faizi et al. 2003).
Derived from P. longifolia leaves, the methanolic extract showcased substantial activity against S. aureus and P. aeruginosa, with MIC values of 125 µg/mL. Two diterpenes, namely 16(R and S)-hydroxy-cleroda-3,13(14)Z-dien-15,16-olide and 16-oxo-cleroda-3,13(14)E-dien-15-oic acid, were isolated from the leaves. Of these, the diterpene 16(R and S)-hydroxy-cleroda-3,13(14)Z-dien-15,16-olide exhibited the most promising antimicrobial activity against S. aureus, boasting an MIC of 7.8 µg/mL (Faizi et al. 2008).
Additionally, investigations into the antimicrobial potential of P. cinnamomea were conducted. However, the leaf extract did not reveal promising activity against S. aureus (ATCC 25923), S. epidermidis, E. coli (ATCC 10536), or S. typhimurium (ATCC 51812) (Mahmud et al. 2018).
Xylopia genus
Antimicrobial investigations have focused on three species within the Xylopia genus. X. staudtii bark hydroethanol extract revealed notable antimicrobial properties against E. coli (ATCC 25922), achieving an MIC of 83.33 µg/mL (Pouofo Nguiam et al. 2021). For both X. aromatica and X. sericea, the essential oils extracted from their leaves were subjected to analysis. X. aromatica’s essential oil displayed promising activity against E. faecalis (ATCC 29212), registering an MIC of 50 µg/mL (Alcântara et al. 2017). Meanwhile, the essential oil from X. sericea exhibited impressive antimicrobial potential against S. aureus (ATCC 6538), boasting an MIC of 7.8 µg/mL, and K. pneumoniae (ATCC 4552), with an MIC of 12.5 µg/mL (Mendes et al. 2017).
Uvaria genus
Furthermore, antimicrobial investigations extended to three species of Uvaria, yielding highly encouraging results. A particularly significant finding was the isolation of the flavonoid 5,7,8-trimethoxyflavone from U. schefflera leaves, which exhibited pronounced antimicrobial activity against E. coli (ATCC 10418), showing an MIC value of 125 µg/mL (Moshi et al. 2004). Additionally, a mixture of flavonoids, 2’,6’-dihydroxy-4’-methoxychalcone and 5,7-dihydroxyflavone, displayed notable antimicrobial activity against C. albicans, registering an MIC of 31.2 µg/mL (Moshi et al. 2004).
The methanolic extract from the root bark of U. tanzaniae exhibited remarkable antimicrobial activity against S. aureus (ATCC 25923), boasting an impressively low MIC of 1.25 µg/mL (Christopher et al. 2018). In a similar vein, the essential oil derived from the leaves of U. hamiltonii showcased substantial antimicrobial potential. It displayed noteworthy activities against S. aureus (ATCC 25923), P. aeruginosa (ATCC 27853), E. faecalis (ATCC 29212), and C. albicans (ATCC 10231), with MIC values of 20.34 µg/mL, 12.34 µg/mL, 7.99 µg/mL, and 32.57 µg/mL, respectively (Tsiang et al. 2022).
Other species of Annonaceae
Research conducted by Pereira et al., 2016, investigated the antimicrobial properties of Cleistochlamys kirkii (Benth) Oliv, a shrub commonly utilized in traditional medicine to treat infectious diseases. From the methanolic extract of C. kirkii, the authors isolated nine compounds and assessed their efficacy against six strains of S. aureus, as well as the bacterium E. faecalis. The compound dichamanetin was shown to be highly active against the tested strains and is often more potent than the control antibiotics used in the study. In addition to this substance, the compounds chamanetin, isochamanetin and cleistenolide also showed relevant antibacterial activity against the strains studied. Thus, the authors verified the antibacterial potential of C. kirkiii and identified active components as valuable compounds to continue with in antimicrobial studies (Pereira et al. 2016).
In a study conducted by Silva et al. (2015), the polycapol triterpene, a lanostane-type triterpene, was investigated. Polycarpol, which has been isolated from several Annonaceae species including Unonopsis duckei R.E. Fr., Unonopsis floribunda Diels, Unonopsis rufescens (Baill.) R. E. Fr., Unonopsis stipitata Diels, Onychopetalum amazonicum R.E. Fr., and Bocageopsis pleiosperma Maas, demonstrated significant antimicrobial activity. It exhibited MIC values ranging from 25 to 50 µg/mL against S. aureus, S. epidermidis, and E. coli (da Silva et al. 2015).
Numerous species belonging to the Annonaceae family have undergone comprehensive assessments of their essential oils against pathogenic microorganisms. These species include Bocageopsis multiflora (Alcântara et al. 2017; Bay et al. 2019a), Cananga odorata (Sacchetti et al. 2005), Cyathocalyx zeylanicus (Hisham et al. 2012), Dennetia tripetala (Oyemitan et al. 2019), Desmopsis bibacteata (Palazzo et al. 2009), Desmopsis macocarpa (Palazzo et al. 2009), Desmos chinensis (Hisham et al. 2012), Duguetia lanceolata (Sousa et al. 2012), Ephedranthus amazonicus (Alcântara et al. 2017), Fissistigma kwangsiensis (Tsiang et al. 2022), Fusaea longifolia (Bay et al. 2019a), Goniothalamus gracilipes (Trieu et al. 2021), and Unonopsis costaricensis (Palazzo et al. 2009). Notably, the study involving Fissistigma kwangsiensis stands out for its remarkable findings, as the essential oil displayed highly promising activity against P. aeruginosa (ATCC 27853), achieving an impressively low MIC of 3.45 µg/mL (Tsiang et al. 2022).
Furthermore, a range of species, including Goniothalamus longistipetes, Mitrephora celebica (Zgoda-Pols et al. 2002), Greenwayodendron suaveolens (Williams et al. 2010; Christopher et al. 2018), Enantia chlorantha (Etame et al. 2019), Cleistopholis patens (Hu et al. 2006), and Toussaintia orientalis Verdc (Samwel et al. 2011), underwent isolation of active compounds from their extracts, with subsequent evaluation of the antimicrobial profiles of these molecules. For example, the diterpene ent-trachyloban-19-oic acid, extracted from Mitrephora celebica, showed considerable effectiveness against S. aureus (ATCC 43300), with a remarkable MIC of 6.25 µg/mL (Zgoda-Pols et al. 2002).
In addition, the antimicrobial potential of extracts from species such as Annonidium mannii (Ngangoue et al. 2020), Monodora myristica (Mbosso et al. 2010), Mitrephora celebica (Zgoda-Pols et al. 2002), Greenwayodendron suaveolens (Williams et al. 2010; Christopher et al. 2018), and Uvariodendron usabarense (Christopher et al. 2018) has been evaluated. These collective efforts contribute valuable insights into the rich antimicrobial potential present within the Annonaceae family.
Table 3.
Summary of antimicrobial activity of species of the Annonaceae.
| Annonaceae species | Used Material | Substances/ Extracts |
Microorganisms / MIC (µg/mL) | Ref. | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Annona foetida |
Leaves |
Essential oil Chloramphenicol Nystatin (Positive control) |
ATCC 6538 200 20 - |
ATCC 10231 60 - 50 |
(Costa et al. 2009) |
|||||||||||||
|
Annona hypoglauca Mart. |
Bark |
Alkaloid phases FA5 (isoboldine) FA6 (actinodaphnine) |
ATCC 29213 70 70 |
ATCC 25922 - 90 |
ATCC 299212 70 80 |
(Rinaldi et al. 2017) |
||||||||||||
|
Annona muricata |
Leaves |
Dried leaf powder: AM0 (Size bands < 2000) AM1 (Size bands between 0.500 and 0.350) |
ATCC 25923 3120 1560 |
ATCC 25922 1560 1560 |
ATCC 27853 3120 3120 |
ATCC 4352 3120 3120 |
(de Andrade et al. 2019) |
|||||||||||
| Annonaceae species | Used Material |
Substances/ Extracts |
Microorganisms / MIC (µg/mL) | Ref. | ||||||||||||||
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Annona senegalensis |
Root bark |
methanol-methylene chloride extract kaur-16-en-19-oic acid F1 (Subfraction) Ciprofloxacin Gentamicin (Positive controls) |
8750 150 - 1.18 0.23 |
1080 - 40 3.6 0.79 |
(Akah et al. 2012) |
|||||||||||||
|
Annona squamosa |
Leaves |
11-hydroxy-16-hentriacontanona + 10-hydroxy-hentricontanona Palmitone Ciprofloxacin (positive control) |
MTCC 96 25 12.5 0.78 |
MTCC 741 25 6.25 0.78 |
(Shanker et al. 2007) |
|||||||||||||
|
Annona vepretorum |
Leaves |
Crude ethanolic extract Hexanic extract |
ATCC 25923 3120 780 |
ATCC 25922 390 390 |
ATCC 19433 3120 3120 |
ATCC 13883 3120 3120 |
(Almeida et al. 2014) |
|||||||||||
| Annonaceae species | Used Material |
Substances/ Extracts |
Microorganisms / MIC (µg/mL) | Ref. | ||||||||||||||
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Annona vepretorum |
Leaves |
Chloroform extract |
ATCC 25923 1560 |
ATCC 25922 780 |
ATCC 19433 12500 |
ATCC 13883 6250 |
(Almeida et al. 2014) |
|||||||||||
|
Annonidium mannii |
Root Bark |
Crude extract Ciproflocaxin (Positive control) |
ATCC 11296 64 2 |
(Ngangoue et al. 2020) |
||||||||||||||
|
Bocageopsis pleiosperma Maas |
Stem Bark |
Polycarpol Chloramphenicol (Positive control) Ketoconazole (Positive control) |
ATCC 6538 25 25 - |
ATCC 1228 50 50 - |
ATCC 10538 50 50 - |
ATCC 27853 - >500 - |
ATCC 10231 250 - 12.5 |
(da Silva et al. 2015) |
||||||||||
|
Bocageopsis multiflora (Mart.) R.E. Fr |
Leaves |
Essential oil |
ATCC 6538 190 |
ATCC 8739 1500 |
ATCC 9027 3000 |
ATCC 29212 90 |
(Alcântara et al. 2017) |
|||||||||||
| Annonaceae species | Used Material |
Substances/ Extracts |
Microorganisms / MIC (µg/mL) | Ref | ||||||||||||||
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Bocageopsis multiflora |
Aerial parts |
Essential oil TIENAM (Positive control) |
ATCC 33591 4.68 4.68 |
CDC-EDL 933-171-0157:H3 4.68 1.17 |
ATCC 29336 4.68 2.34 |
(Bay et al. 2019a) |
||||||||||||
|
Cananga odorata |
Essential oil Thymus vulgaris (reference oil) |
ATCC 48274 170 60 |
(Sacchetti et al. 2005) |
|||||||||||||||
|
Cleistopholis patens |
Leaves |
Cleistetroside-8 Cleistetroside-5 Cleistetroside-2 Vancomycin (positive control) |
ATCC 33591 8 8 0.5 1 |
(Hu et al. 2006) |
||||||||||||||
|
Cyathocalyx zeylanicus |
Leaves |
Essential oil Gentamycin Muconazol (Positive control) |
ATCC 25923 250 250 9.0 |
ATCC 25922 125 125 12.0 |
ATCC 37853 250 250 9.1 |
ATCC 27853 250 250 8.0 |
ATCC 90028 250 16 4.5 |
(Hisham et al. 2012) |
||||||||||
|
Annonaceae species |
Used Material |
Substances/ Extracts |
Microorganisms / MIC (µg/mL) |
Ref. |
||||||||||||||
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Cleistochlamys kirkii (Benth) Oliv. |
Root Bark |
Chamanetin Isochamanetin Dichamanetin Cleistenolide Acetylmelodorinol Amoxicillin Oxacillin Vancomycin (Positive control) |
S. aureus MSSA MRSA VISA ATCC ATCC ATCC FFHB ATCC CIP 6538 43866 9144 29593 700699 106760 7.5 15 30 - 125 15 62 125 250 30 62 62 2 2 1 4 2 2 30 30 7.5 15 30 30 >250 >250 125 >250 >250 >250 0.2 62 250 >250 250 250 0.2 125 125 250 250 250 0.2 0.4 0.4 0.8 2 4 |
VRE HB164 15 30 7.5 30 >250 - - 32 |
(Pereira et al. 2016) |
|||||||||||||
|
Dennetia tripetala Baker f. |
Seed |
Dried seeds essential oil Streptomycin Acriflavin (Positive control) |
NCIB 8586 3.13 - 0.13 |
NCIB 86 6.25 0.13 |
NCIB 950 25.0 1.0 |
NCCYC 6 6.25 - 2.0 |
(Oyemitan et al. 2019) |
|||||||||||
|
Desmopsis bibracteata Desmopsis macrocarpa |
Leaves Leaves |
Essential oil Essential oil |
ATCC 29213 625 1250 |
ATCC 259222 2500 1250 |
(Palazzo et al. 2009) |
|||||||||||||
| Annonaceae species | Used Material |
Substances/ Extracts |
Microorganisms / MIC (µg/mL) | Ref. | ||||||||||||||
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Desmos chinensis |
Leaves |
Essential oil Gentamycin Muconazol (Positive control) |
ATCC 25923 250 250 9.0 |
ATCC 25922 125 125 12.0 |
ATCC 37853 250 250 9.1 |
ATCC 27853 250 250 8.0 |
ATCC 90028 250 16 4.5 |
(Hisham et al. 2012) |
||||||||||
|
Duguetia lanceolata |
Stem bark |
Essential oil (T2) Essential oil (T4) Chloramphenicol (positive control) |
ATCC 6538 60 125 2 |
ATCC 10231 60 100 15 |
(Sousa et al. 2012) |
|||||||||||||
|
Enantia chlorantha |
Stem bark |
Palmitin Chloramphenicol (positive control) |
ATCC 25923 32 8 |
E.C 136 32 32 |
K.L 128 16 32 |
(Etame et al. 2019) |
||||||||||||
|
Ephedranthus amazonicus R.E. Fr. |
Leaves |
Essential oil Chlorhexidine digluconate (Positive controls) |
ATCC 6538 90 6 |
ATCC 8739 1500 6 |
ATCC 9027 3000 10 |
ATCC 29212 190 90 |
(Alcântara et al. 2017) |
|||||||||||
| Annonaceae species | Used Material |
Substances/ Extracts |
Microorganisms / MIC (µg/mL) | Ref. | ||||||||||||||
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Fissistigma kwangsiensis |
Leaves |
Essential oil Nystatine Streptomycin Cycloheximide (Positive controls) |
ATCC 25923 55.67 - 3.2 - |
ATCC 27853 3.45 8.0 - - |
ATCC 299212 33.62 2.07 - - |
ATCC 10231 16.45 - - 3.2 |
(Tsiang et al. 2022) |
|||||||||||
|
Fusaea longifolia |
Aerial parts |
Essential oil TIENAM (positive control) |
ATCC 33591 37.5 4.68 |
ATCC 29336 37.5 2.34 |
(Bay et al. 2019a) |
|||||||||||||
|
Greenwayodendron suaveolens subs. usambaricum |
Root Bark Roots |
Methanol extracts Pentacyclindole Polyalthenol Ciprofloxacin (Positive control) |
ATCC 25923 1000 4 4 2.5 |
(Christopher et al. 2018) (Williams et al. 2010) (Williams et al. 2010) (Christopher et al. 2018) |
||||||||||||||
|
Goniothalamus gracilipes |
Leaves |
Gracilipin C Streptomycin (Positive control) |
ATCC 25923 32 32 |
(Trieu et al. 2021) |
||||||||||||||
| Annonaceae species | Used Material |
Substances/ Extracts |
Microorganisms / MIC (µg/mL) | Ref. | ||||||||||||||
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Goniothalamus longistipetes |
Bark |
(+)-altholactone ((2S,3R,3aS,7aS)-3-hydroxy-2-phenyl-2,3,3a,7a-tetrahydrobenzo-5(4H)-5-one) (2S,3R,3aS,7aS)-3-hydroxy-2-phenyl-2,3,3a,7a-tetrahydrobenzo-5(4H)-5-one) 2,6-dimethoxyisonicotinaldehyde alkenyl-5-hydroxyl-phenyl benzoic acid |
eMRSA – 15 64 128 128 8-16 |
NCTC 10418 256 512 256 512 |
NCTC 10662 500 512 256 128 |
NCTC 10662 256 512 256 128 |
(Teo et al. 2020) |
|||||||||||
|
Guatteria blepharophylla |
Leaves |
Essential oil Isomoschatoline Chlorhexidine digluconate Nystatin (Positive control) |
ATCC 6538 50 - 6 |
ATCC 8739 1500 - 6 |
ATCC 9027 1500 - 10 |
ATCC 29212 50 - 90 |
ATCC 10231 - 50.81 µM L-1 54 µM L-1 |
(Costa et al. 2011c; Alcântara et al. 2017) |
||||||||||
| Annonaceae species | Used Material |
Substances/ Extracts |
Microorganisms / MIC (µg/mL) | Ref. | ||||||||||||||
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Guatteria costaricensis Guatteria diospyroides Guatteria oliviformis |
Leaves |
Essential oil Essential oil Essential oil |
ATCC 29213 1250 312 1250 |
ATC 25922 1250 1250 625 |
(Palazzo et al. 2009) |
|||||||||||||
|
Guatteria selowiana Guatteria latifolia Guatteria ferruginea Guatteria australis |
Aerial parts Aerial parts Aerial parts Aerial parts |
Essential oil Essential oil Essential oil Essential oil Chloramphenicol (Positive control) |
ATCC 11775 600 600 600 >1000 40 |
(Santos et al. 2017) |
||||||||||||||
| Annonaceae species | Used Material |
Substances/ Extracts |
Microorganisms / MIC (µg/mL) | Ref. | ||||||||||||||
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Guatteria citriodora |
Leaves and Stem bark |
Crude hydroalcoholic extract of leaves Alkaloid fraction of leaves Alkalidic fraction of stem bark Imipenem (Positive control) |
250 - - 15.6 |
ATCC 4083 - 125 - 62.5 |
(Rabelo et al. 2014) |
|||||||||||||
|
Guatteria punctata |
Aerial parts |
Essential oil TIENAM (positive control) |
ATCC 33591 - 4.68 |
CDC-EDL 933-171-0157:H3 - 1.17 |
ATCC 29336 - 2.34 |
(Bay et al. 2019a) |
||||||||||||
|
Guatteriopsis hispida |
Essential oil Oxide caryophyllene |
ATCC 6538 >1000 - |
ATCC 12228 >1000 - |
ATCC 11775 >1000 - |
ATCC 133388 >1000 - |
ATCC 10231 >1000 600 |
(Costa et al. 2008) |
|||||||||||
| Annonaceae species | Used Material |
Substances/ Extracts |
Microorganisms / MIC (µg/mL) | Ref. | ||||||||||||||
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Guatteriopsis blepharopylla Guatteriopsis friesiana |
seed |
Essential oil β-eudesmol Ƴ-eudesmol α-eudesmol Essential oil β - pinene α – pinene (E)- caryophyllene Chloramphenicol Nystatin (Positive control) |
ATCC 6538 1000 >1000 600 250 125 - - - 20 - |
ATCC 12228 >1000 600 700 700 100 - - - 40 - |
ATCC 11775 >1000 >1000 >1000 - 900 - - - 40 |
ATCC 133388 >1000 >1000 300 200 900 - - - 850 |
ATCC 10231 700 125 500 125 500 100 250 - 50 |
(Costa et al. 2008) |
||||||||||
|
Mitrephora celebica |
Leaves Stem bark Twigs |
Crude hydroalcoholic extract of leaves Crude hydroalcoholic extract Crude hydroalcoholic extract Ent-trachyloban-19-oic acid |
ATCC 43300 >100 12.5 >100 6.25 |
ATCC 27853 >100 >100 >100 >100 |
(Zgoda-Pols et al. 2002) |
|||||||||||||
| Annonaceae species | Used Material |
Substances/ Extracts |
Microorganisms / MIC (µg/mL) | Ref. | ||||||||||||||
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Mitrephora celebica |
ent-kaur-16-en-19-oic acid Gentamycin Oxacillin Vancomycin (Positive control) |
ATCC 43300 >100 >50 6.2-12.5 0.8 |
ATCC 27853 >100 0.8 - - |
(Zgoda-Pols et al. 2002) |
||||||||||||||
|
Monodora myristica |
Fruits |
Cyclohexane extract Ethyl acetate extract |
25 25 |
25 50 |
- 50 |
12.5 12.5 |
6.3 12.5 |
(Mbosso et al. 2010) |
||||||||||
|
Polyalthia cinnamomea |
Leaves |
Leaf extract Vancomycine Streptomicyne Kanamycine (Positive control) |
ATCC 25923 4000 - 30 - |
4000 30 - - |
ATCC 10536 1000 - - 30 |
(Mahmud et al. 2018) |
||||||||||||
|
Polyalthia longifolia |
Stem bark |
Butanol fraction 3-o-methyl ellagic |
ATCC 29213/ 512 (MRSA) 320/320 80/160 |
ATCC 29212 320 80 |
ATCC 27853 160 80 |
(Jain et al. 2014) |
||||||||||||
| Annonaceae species | Used Material |
Substances/ Extracts |
Microorganisms / MIC (µg/mL) | Ref. | ||||||||||||||
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Polyalthia longifolia |
Stem bark Roots Leaves |
Vancomycin Oxacillin Ciprofloxacin (Controle positive) Pendulamine A Pendulamine B Penduline Kanamycin sulfate (Control positive) Methanol extract 16(R and S)-hydroxy-cleroda-3,13(14)Z-dien-15,16-olide 16-oxo-cleroda-3,13(14)E-dien-15-oic acid |
ATCC 29213/ 512 (MRSA) 0.25/- -/8.0 - 0.2 0.2 12.5 0.31 125 7.8 500 |
ATCC 29212 - - 0.015 |
ATCC 27853 - - 0.25 2 - - 5 125 250 500 |
2 2 - 5 |
(Jain et al. 2014) (Jain et al. 2014) (Jain et al. 2014) (Faizi et al. 2003) (Faizi et al. 2003) (Faizi et al. 2003) (Faizi et al. 2003) (Faizi et al. 2008) (Faizi et al. 2008) (Faizi et al. 2008) |
|||||||||||
| Annonaceae species | Used Material |
Substances/ Extracts |
Microorganisms / MIC (µg/mL) | Ref. | ||||||||||||||
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Toussaintia orientalis Verdc. |
Seeds |
Toussintine A Toussintine B Toussintine C Toussintine D Toussintine E Ampicilin (Positive control) |
ATCC 25923 - 10 - 5 - 2.5 |
DSM 1103 10 20 10 - - 2.5 |
(Samwel et al. 2011) |
|||||||||||||
|
Unonopsis duckei R.E. Fr. Unonopsis floribunda Diels Unonopsis rufescens (Baill.) R.E. Fr. Unonopsis stipitala Diels Onychopetalum amazonicum R.E. Fr. |
Stem Bark |
Polycarpol Chloramphenicol (Positive control) Ketoconazole (Positive control) |
ATCC 6538 25 25 - |
ATCC 1228 50 50 - |
ATCC 10538 50 50 - |
ATCC 27853 - >500 - |
ATCC 10231 250 - 12.5 |
(da Silva et al. 2015) |
||||||||||
|
Unonopsis costaricensis |
Leaves |
Essential oil |
ATCC 29213 625 |
ATCC 25922 1250 |
(Palazzo et al. 2009) |
|||||||||||||
| Annonaceae species | Used Material |
Substances/ Extracts |
Microorganisms / MIC (µg/mL) | Ref. | ||||||||||||||
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Uvaria hamiltonii |
Leaves |
Essential oil Nystatine Streptomycin Cycloheximide (Positive controls) |
ATCC 25923 20.34 - 1.07 - |
ATCC 27853 12.34 8.0 - - |
ATCC 29212 7.99 0.48 - - |
ATCC 10231 32.57 - - 3.2 |
(Tsiang et al. 2022) |
|||||||||||
|
Uvaria schefflera |
Leaves |
5,7,8-trimetoxiflavona 2’,6’-dihydroxy-4’-methoxychalcone and 5,7-dihydroxyflavone (mixture) Ampicillin Ketoconazole (positive control) |
NCTC 6571 - 125 0.01 |
NCTC 10418 125 - 0.25 |
HG 392 - 31.2 0.125 |
(Moshi et al. 2004) |
||||||||||||
|
Uvaria tanzaniae |
Root Bark |
Methanol extracts Ciprofloxacin (Positive control) |
ATCC 25923 1.25 2.5 |
(Christopher et al. 2018) |
||||||||||||||
|
Annonaceae species |
Used Material |
Substances/ Extracts |
Microorganisms / MIC (µg/mL) |
Ref. | ||||||||||||||
| S. aureus | S. epidermidis | E. coli | P. aeruginosa | E. faecalis | K. pneumoniae | C. krusei | C. albicans | C. tropicalis | ||||||||||
|
Uvariodendron usambarense |
Leaves Stem Bark |
Methanol extracts Methanol extracts Ciprofloxacin (Positive control) |
ATCC 25923 8000 4000 2.5 |
ATCC 8740 500 - 0.63 |
(Christopher et al. 2018) |
|||||||||||||
|
Xylopia staudtii |
Bark |
Hydroethanolic extract Ciprofloxacin (Positive control) |
ATCC 25922 83.33 0.97 |
(Pouofo Nguiam et al. 2021) |
||||||||||||||
|
Xylopia aromatica (Lam.) Mart. |
Leaves |
Essential oil Chlorhexidine digluconate (Positive control) |
ATCC 6538 1200 6 |
ATCC 8739 3000 6 |
ATCC 9027 3000 10 |
ATCC 29212 50 90 |
(Alcântara et al. 2017) |
|||||||||||
|
Xylopia sericea |
Fruits |
Essential oil Chloramphenicol Ciprofloxacin (positive control) |
ATCC 6538 7.8 12.5 0.39 |
ATCC 10536 1000 6.25 1.56 |
ATCC 19433 1000 6.25 1.56 |
ATCC 4552 62.5 12.5 12.5 |
(Mendes et al. 2017) |
|||||||||||
3.4. Leishmanicidal
Leishmaniasis can be caused by any of the 20 species of hemoflagellate protozoa in the genus Leishmania. These protozoa are primarily transmitted to humans through the bites of female sandflies belonging to 30 species of Lutzomyia (associated with New World Leishmaniasis) and Phlebotomus (linked to Old World Leishmaniasis) (López et al. 2009; Brígido et al. 2020). This neglected disease is a public health problem that affects 98 tropical and subtropical countries with approximately 1.7 million cases each year (Lamidi et al. 2005; Ferreira et al. 2017). According to the WHO, it is considered the second most important protozoan public disease, with extensive morbidity and mortality in most developing countries (Lorenzo et al. 2016; Christopher et al. 2018).
Clinically, the disease can affect the skin (cutaneous leishmaniasis), mucosa (mucocutaneous leishmaniasis) and/or organs of the reticulo-endothelial system (visceral leishmaniasis). The forms of the disease are related to the type of parasite and differ in distribution, host and vector involved, incidence and mortality rate (López et al. 2009; Lorenzo et al. 2020).
Since the 1940s, pentavalent antimonial compounds (sodium stibogluconate and meglumine antimoniate) have been the first-line treatment for all forms of leishmaniasis but in cases of therapeutic resistance to these compounds, amphotericin B deoxycholate and liposomal amphotericin B (pentamidine and miltefosine) can be used (Lamidi et al. 2005). However, they are formulations that present high toxicity, high cost and development of parasitic resistance. Therefore, there is an urgent need for new therapies against leishmaniasis (Ferreira et al. 2017).
The lack of effective antiprotozoal drugs has renewed interest in the study of medicinal plants as sources of new therapeutic compounds with stronger activity and fewer side effects (Osorio et al. 2007; Christopher et al. 2018). The literature describes several classes of natural substances with proven leishmanicidal activity in in vitro assays on promastigotes and/or amastigotes of Leishmania such as terpenes, chalcones, acetogenins, alkaloids, diterpenic acids, quinones and phenolic derivatives (da Silva et al. 2012; Lorenzo et al. 2016, 2020; Ferreira et al. 2017).
The antiprotozoal activity generally reported for the Annonaceae family has been related to the traditional treatment of diseases such as malaria, Chagas disease, sleeping sickness and leishmaniasis (Vila-Nova et al. 2011; De Lima et al. 2012; Musuyu Muganza et al. 2015). A compilation of the leishmanicidal potential of essential oil, extracts, fractions and isolated compounds of species from the Annonaceae family is described in the following sections and in Table 4.
Annona genus
The ethanol extracts from the root barks, stem barks, and stem wood of A. crassiflora exhibited activity against L. donovani promastigotes, with effective concentrations ranging from 3.7 to 12.4 μg/mL. Additionally, the total alkaloids extracted from the leaves demonstrated activity against L. chagasi, with an IC50 value of 24.9 μg/mL (De Mesquita et al. 2005; Brígido et al. 2020). Furthermore, the essential oil derived from the leaves of A. coriacea showed anti-leishmanial effects against L. chagasi promastigotes (Siqueira et al. 2011).
Acetogenins such as annofolin and annotacin, isolated from A. cornifolia seeds, have shown activity against the amastigote forms of L. amazonensis with IC50 6.4 and 7.2 μM, respectively (Lima et al. 2014; Brígido et al. 2020). The essential oil, extracts, alkaloid fraction and some compounds (N-hydroxyanno-montine, O-methylmoschatolin, liriodenine and annomontine) isolated from bark and leaves of A. foetida, showed antileishmanial activity, giving the best results against promastigotes of L. braziliensis and L. guyanensis (Costa et al. 2009; Brígido et al. 2020).
The hydroalcoholic extract from the leaves of A. glabra exhibited activity against L. amazonensis promastigotes, with an IC50 of 37.8 μg/mL (Brígido et al. 2020). A study with Annonacin A and Goniothalamicin isolated from A. glauca seeds showed activity against the promastigote form of Leishmania spp. with IC50 16.75 and 8.37 μM, respectively (Waechter et al. 1998). From the hexanic extract of A. haematantha roots, argentilactone was isolated and exhibited in vitro activity against various strains of Leishmania ssp with IC50 value of 10.0 μg/ml (Waechter et al. 1997).
Various extracts from A. mucosa and the alkaloid liriodenine exhibited activity against L. amazonensis promastigotes (De Lima et al. 2012; Brígido et al. 2020). Efficacy against Leishmania species was also observed in extracts and fractions from the stems of A. muricata, which may be attributed to the presence of acetogenins, with increased activity noted following fractionation (Osorio et al. 2007; Vila-Nova et al. 2011; Brígido et al. 2020). Additionally, methanol and aqueous extracts from A. purpurea bark and seeds demonstrated significant activity against L. donovani promastigotes, while the fraction from the hydroalcoholic leaf extract was effective against L. panamensis, exhibiting an IC50 of 0.96 μg/mL (Malebo et al. 2013b).
Berbine alkaloids (pessione and spinosine) isolated from the bark and roots of A. spinescens were active against Leishmania ssp. Promastigotes (Emerson F. Queiroz et al. 1996). A benzylisoquinolinic alkaloid (O-methylarmepavine), and a C37 trihydroxy adjacent bistetrahydrofuran acetogenin isolated from leaves of A. squamosa, showed activities against promastigote and amastigote forms of L. chagasi (Vila-Nova et al. 2011). Ethanolic extracts from the leaves and stem of A. senegalensis exhibited activity against L. donovani promastigotes, showing IC50 values of 10.8 μM for the leaf extract and 27.8 μM for the stem extract (Ohashi et al. 2018; Brígido et al. 2020).
Guatteria genus
The alkaloids xylopine and nornuciferine (extracted from the leaves of G. amplifolia) and cryptodorine and nornantenine (from the leaves of G. dumetorum) demonstrated significant activity against L. mexicana and L. panamensis promastigotes (Lorenzo et al. 2016). Essential oil from the leaves of G. australis showed activity against L. infantum (IC50 = 30.71 μg/ml) (Lorenzo et al. 2016).
Bisbenzylisoquinoline alkaloids, including Puertogaline A and B, as well as the derivative Sepeerine, isolated from the stem bark of G. boliviana, exhibited moderate inhibition of Leishmania spp. promastigotes at a concentration of 100 μg/mL. This level of activity was comparable to that observed with the crude ethanolic extract in the screening assays (Lorenzo et al. 2016).
The branch extract of G. latifolia demonstrated notable activity against both L. amazonensis promastigotes and intracellular amastigotes and was subsequently fractionated based on in vitro assays. Among the fractions obtained, two alkaloid-rich fractions, GF1 and GF2, exhibited the highest activity against promastigotes, with IC50 values of 25.6 and 16 μg/mL, respectively (Ferreira et al. 2017).
Rollinia genus
The dichloromethane fraction of the stem bark of R. emarginata exhibited activity against various Leishmania spp. strains. Fractionation of this extract, guided by in vitro leishmanicidal activity, resulted in the isolation of five active compounds: four acetogenins (rolliniastatin-1, sylvaticin, squamocin, and rollidecin B) and one oxoaporphine (liriodenin)(Février et al. 1999). Additionally, extracts from the stem of R. exsucca and from the stem and leaves of R. pittieri demonstrated activity against Leishmania spp., with IC50 values below 25 μg/mL (Osorio et al. 2007).
Unonopsis genus
Three alkaloids isolated from the dichloromethane extract of the stem bark of U. buchtienii exhibited the highest activity against L. major and L. donovani. The oxoaporphine alkaloids, O-methylmoschatoline and lysicamine, had IC100 values of 50.0 and 25.0 mg/mL, respectively, and liriodenine, the most active alkaloid had an IC100 of 3.12 mg/mL (Waechter et al. 1999).
Alkaloidal fractions extracted from the twigs, bark, and leaves of U. guatterioides and U. duckei were evaluated for activity against promastigote forms of L. amazonensis. All fractions from U. guatterioides showed high activity, with IC50 values of 1.07, 1.90, and 2.79 mg/mL, respectively. In contrast, only the alkaloidal fraction from the twigs of U. duckei did not exhibit any activity (da Silva et al. 2012).
Xylopia genus
The methanol extract from the leaves of X. aromatica was active against Leishmania spp. Promastigotes, displaying an IC50 value of 20.8 μg/ml.(Osorio et al. 2007) The essential oil and eight extracts obtained from leaves and seed of X. discreta showed activity against L. panamensis promastigotes (López et al. 2009).
A new ent-kaurene diterpene glucoside, 7β-O-β-D-glucopyranoside-ent-kaur-16- ene, from the leaves of X. excellens, showed high in vitro antileishmanial activity (IC50 of 15.23 µg/mL) towards promastigote forms of L. amazonensis. The dichloromethane extract from roots of X. parviflora displayed high inhibitory effects on the growth of amastigote forms of L. donovani with IC50 value of 5.01 μg/ml (Bapela et al. 2017).
Others Annonaceae species
The azaphenanthrene alkaloids sampangine, imbiline 3, imbiline 1, and eupolaramine, extracted from the roots of Anaxagorea dolichocarpa, demonstrated significant activity against the promastigote forms of L. donovani, with IC50 values of ≤ 24.06 μM (Lorenzo et al. 2016). Furthermore, an aporphine alkaloid (lysicamine) and a bis-aporphine alkaloid (trivalvone) obtained from the leaves of Annickia kummeriae showed enhanced effectiveness against the amastigote form of L. donovani (Malebo et al. 2013b).
The Bocageopsis multiflora essential oil, richer in oxygenated sesquiterpenes, exhibited good activity against the L. amazonensis promastigote (IC50 =14.6 μg/mL) (Malebo et al. 2013b). The alkaloids duguetine β-N-oxide and dicentrinone isolated from the bark of Duguetia furfuracea, presented IC50 vlaues of 0.11 and 0.01 μM against the promastigote forms of L. braziliensis (da Silva et al. 2009).
The aporphine alkaloid glaucine, extracted from the leaves of Duguetia lanceolata, demonstrated effectiveness against L. infantum amastigotes. Additionally, an alkaloid-rich fraction containing isocorydine, norglaucine, and N-methyllaurotetanine exhibited activity against promastigotes (da Silva et al. 2009). Furthermore, the aqueous extract of Enantia chlorantha stem bark and the compound palmatine, isolated from the methanolic bark extract, also showed activity against L. infantum promastigotes (da Silva et al. 2009).
Extracts, fractions and isolated constituents (polycarpol, dihydropolycarpol and polyathenol) from fruits, leaves, root bark and stem bark of Greenwayodendron suaveolens showed in vitro activity against L. infantum promastigote (da Silva et al. 2009). The crude methanol extract of leaves of Isolona hexaloba and the two dichloromethane soluble fractions of the 80% ethanol extracts from root bark and stem bark showed good activity and selectivity against the L. infantum promastigote, with IC50 values equal to 6.35, 6.96 and 8.0 µg/ml, respectively (Musuyu Muganza et al. 2015).
A new diterpene, (4S,9R,10R) methyl 18-carboxy-labda-8,13(E)-diene-15-oate obtained from the stem barks of Polyalthia macropoda showed activity against the promastigote L. donovani.(Richomme et al. 1991) The methanolic extract of P. suaveolens stem bark demonstrated potent antiproliferative activity against L. infantum promastigotes (Lamidi et al. 2005).
Acetylene derivatives were isolated from seeds of Porcelia macrocarpa, where all isolated compounds demonstrated selectivity towards intracellular amastigotes of L. infantum, especially: 3-hydroxy-4-methylene-2-(eicos-11’-yn-19’-enyl)but-2-enolide, 3-hydroxy-4-methylene-2-(octadec-9’-yn-17’-enyl)but-2-enolide and 3-hydroxy-4-methylene-2-(hexadec-7’-yn-15’-enyl)but-2-enolide with IC50 values of 9.2, 10.4 and 11.0 µM, respectively and thus indicating superior activity over the positive control miltefosine (IC50 of 17.8 µM) (Brito et al. 2021).
An acetylene acetogenin (2S,3R,4R)-3-hydroxy-4-methyl-2-(n-eicos-11’-yn-19’-enyl)butanolide was obtained from the seeds of Porcelia macrocarpa and showed an IC50 value of 29.9 μM against the intracellular amastigote forms of L. infantum, whereas the similar compound (2S,3R,4R)-3-hydroxy-4-methyl-2-(n-eicos-11’-ynyl)butanolide was inactive. These results suggested that the terminal double bond plays an important role in the activity (Brito et al. 2022).
Three 6-substituted 5,6-dihydro-2H-pyran-2-ones were isolated from leaves of Raimondia monoica. All compounds showed activity against L. panamensis promastigotes at concentrations ranging between 0.4-10.0 μg/mL (Carmona et al. 2003). Bingervone, a β-triketone derivative, was isolated from the dichloromethane extract of the roots of Uvaria afzelii, and showed moderate antileishmanial activities against L. donovani and L. major promastigotes, with IC50 values of 38.9 and 44.4 µM, respectively (Okpekon et al. 2015).
Bioguided-fractionation of a dichloromethane extract of the stems of Uvaria klaineana led to isolation of a lactone (klaivanolide), that showed potent in vitro antileishmanial activity against both sensitive and amphotericin B-resistant promastigote forms of L. donovani (IC50 values of 1.75 and 3.12 µM, respectively) (Akendengue et al. 2002).
Some studies adopt the following criteria for the evaluation of in vitro antiprotozoal screening of extracts, fractions and isolated substances: IC50 ≤ 5 μg/mL: pronounced activity; 5 < IC50 ≤ 10 μg/mL: good activity; 10 < IC50 ≤ 20 μg/mL: moderate activity; 20 < IC50 ≤ 40 μg/mL: low activity; IC50 > 40 μg/mL: inactive (Musuyu Muganza et al. 2015; Muganza et al. 2016). This should be taken into consideration for the results compiled in Table 4, our summation of published anti-leishmaniasis screening of compounds isolated from Annonaceae plants.
Table 4.
Summary of leishmanicidal activity of species of the Annonaceae.
| Annonaceae species | Used Material | Substances/Extracts | Leishmania spp. | Parasite form | IC50 | Ref. | |
|---|---|---|---|---|---|---|---|
| µM | µg/ml | ||||||
| Anaxagorea dolichocarpa | root | Sampangine Imbiline 3 Imbiline 1 EupolaUramine |
L. donovani | Promastigote | 24.06 16.91 18.20 19.90 |
5.59 5.45 5.32 5.26 |
(Lorenzo et al. 2016) |
| Annickia kummeriae | leave | Methanolic extract Lysicamine Trivalvone Palmatine Jatrorrhizine Jatrorrhizinne/Columbamine Palmatine/Tetrahydro-palmetine |
L. donovani | Amastigote | 9.26 5.24 22.13 60.28 19.35 9.88 |
9.25 2.7 2.9 7.8 20.4 13.1 7.0 |
(Malebo et al. 2013b) |
| Annona crassiflora | Stem bark, stem wood, root bark and root wood | Ethanolic extract Stem bark Stem wood Root bark Root wood |
L. donovani | Promastigote |
12.4 8.3 3.7 8.7 |
(De Mesquita et al. 2005; Brígido et al. 2020) | |
| Annona coriaceae | leave | Essential oil | L. chagasi | Promastigote | 39.93 | (Siqueira et al. 2011) | |
| Annona cornifolia | seed | Annofolin Annotacin Extract |
L. amazonensis | Amastigote | 6.4 7.2 |
175.0 |
(Lima et al. 2014; Brígido et al. 2020) |
| Annona foetida | bark | Hexane extract Dichlorometahne extract Alkaloid fraction (Dichlorometane extract) Methanolic extract Alkaloid fraction (Methanolic extract) N-hydroxyanno-montine |
L. braziliensis and L. guyanensis | Promastigote |
911.3 and 1577.8 |
>160.0 and 42.7 23.0 and 2.7 23.0 and 2.7 40.4 and 23.6 24.3 and 9.1 252.7 and 437.5 |
(Costa et al. 2009; Brígido et al. 2020) |
| Annonaceae species | Used Material | Substances/Extracts | Leishmania spp. | Parasite form | IC50 | Ref. | |
| µM | µg/ml | ||||||
| Annona foetida | Bark Leave |
O-methylmoschatolin Liriodenine Annomontine Essential oil |
L. braziliensis and L. guyanensis L. amazonensis, L. braziliensis, L. chagasi and L. guyanensis |
Promastigote | 998.35 and 322.7 212.52 and 78.10 >2346.15 |
320.8 and 103.7 58.5 and 21.5 34.8 and >613.0 16.2, 9.9, 27.2 and 4.1 |
(Costa et al. 2009; Brígido et al. 2020) |
| Annona glabra | leave | Hydroalcoolic extrac | L. amazonensis | Promastigote | 37.8 | (Brígido et al. 2020) | |
| Annona glauca | seed | Dichloromethane extract Hexane extract Annonacin A Goniothalamicin Glaucanisin Rolliniastatin-2 Squamocin Glaucafilin Molvizarin Parviflorin Annonacin |
L. braziliensis, L. amazonensis and L. donovani |
Promastigote |
16.75 8.37 40.13 40.13 40.13 41.88 >168.09 >168.09 21.61 |
IC100 25.0 >100.0 10.0 5.0 25.0 25.0 25.0 25.0 >100.0 >100.0 12.5 |
(Waechter et al. 1998) |
| Annona haematantha | root | Argentilactone | L. donovani, L. major and L. amazonensis | Promastigote | 51.47 | 10.0 | (Waechter et al. 1997) |
| Annona mucosa | Leave Seed |
Hexane extract Dichloromethane extrac Methanol extract Hexane extract Methanol extract Liriodenine |
L. amazonensis and L. braziliensis |
Promastigote |
5.19 and 203.15 |
24.24 and 65.17 9.32 and 27.42 28.32 and 44.74 44.2 and 170.15 46.54 and 133.8 1.43 and 55.92 |
(De Lima et al. 2012; Brígido et al. 2020) |
| Annona muricata | leave | Ethil acetate extract | L. amazonensis, L. donovani | Promastigote | 25.0 | (Osorio et al. 2007; Vila-Nova et al. 2011; Brígido et al. 2020) | |
| Annonaceae species | Used Material | Substances/Extracts | Leishmania spp. | Parasite form | IC50 | Ref. | |
| µM | µg/ml | ||||||
| Annona muricata | Stem | Hexane extract Methanolic extrac Hexane extract Ethil acetate extract Methanolic extrac |
L. amazonensis, L. brasiliensis and L. donovani | Promastigote | >100.0 >100.0 98.6 63.2 98.6 |
(Osorio et al. 2007; Vila-Nova et al. 2011; Brígido et al. 2020) | |
| seeds | Annonacinone | L. chagasi | Promastigote and Amastigote | 63.20 and 22.69 | 37.6 and 13.5 | ||
| L. donovani, L. mexicana and L. major | Promastigote | 12.87, 13.44 and 11.29 | 7.66, 8.00 and 6.72 | ||||
| Corossolone | L. chagasi | Promastigote and Amastigote | 44.74 and 49.57 | 25.9 and 28.7 | |||
| L. donovani, L. mexicana and L. major | Promastigote | 32.35, 32.19 and 27.88 | 18.73, 18.64 and 16.14 | ||||
| Scoparone | L. donovani, L. mexicana and L. major | Promastigote | 133.42, 44.18 and 69.69 | 27.51, 9.11 and 14.37 | |||
| Annona purpurea | Bark Seed Leave |
Methanolic extract Aqueous extract Hydroalcoolic fraction |
L. donovani L. panamensis |
Promastigote | 113.24 28.57 289.0 0.961 |
(Brígido et al. 2020) | |
| Annona spinescens |
Bark Root |
Annonaine Liriodenine |
L. braziliensis L. amazonensis L. donovani L. braziliensis |
Promastigote |
188.45 94.22 376.91 363.29 |
IC100 50.0 25.0 100.0 100.0 |
(Emerson F. Queiroz et al. 1996) |
| Annona squamosa | leave | O-methylarmepavine C37 trihydroxy adjacent bistetrahydrofuran acetogenin |
L. chagasi | Promastigote and Amastigote | 71.16 and 77.58 42.44 and 40.67 |
23.3 and 25.4 26.4 and 25.3 |
(Vila-Nova et al. 2011) |
| Annona senegalensis | leave Stem |
Ethanolic extract | L. donovani | Promastigote | 10.8 27.8 |
(Ohashi et al. 2018; Brígido et al. 2020) | |
| Annonaceae species | Used Material | Substances/Extracts | Leishmania spp. | Parasite form | IC50 | Ref. | |
| µM | µg/ml | ||||||
| Bocageopsis multiflora | leave | Essential oil | L. amazonensis | Promastigote | 14.6 | (Oliveira et al. 2014) | |
| Duguetia furfuracea | bark | Alkaloid extract Duguetine Duguetine β-N-oxide Dicentrinone N-methyltetrahydropalmatine N-methylglaucine |
L. braziliensis | Promastigote | 16.32 4.32 0.11 0.01 17.03 4.88 |
(da Silva et al. 2009) | |
| Duguetia lanceolata | leave | Glaucine | L. infatum | Amastigote and Promastigote |
21.10 and >281.37 | 7.5 and >100.0 | (Dantas et al. 2020) |
| Enantia chlorantha | stem bark | Aqueous extract | L. infatum | Promastigote | 10.08 | (Olivier et al. 2015) | |
| Guatteria amplifolia | leave | Xylopine Nornuciferine |
L. mexicana and L. panamensis | Promastigote | 3.0 and 6.0 14.0 and 28.0 |
(Montenegro et al. 2003) | |
| Guatteria australis | leave | Essential oil | L. infatum | Promastigote | 30.71 | (Siqueira et al. 2015) | |
| Guatteria boliviana | bark | Ethanolic extract Puertogaline A Puertogaline B Sepeerine |
L. amazonensis, L. braziliensis and L. donovani |
Promastigote |
177.74 177.74 168.15 |
100.0 100.0 100.0 100.0 |
(Mahiou et al. 2000a) |
| Guatteria dumetorum | leave | Cryptodorine Normantenine |
L. mexicana and L. panamensis | Promastigote | 3.0 and 6.0 24.0 and 15.0 |
(Montenegro et al. 2003) | |
| Guatteria latifolia | branch | Crude extract Buthanolic fraction 1 Buthanolic fraction 2 |
L. amazonensis | Promastigote and Amastigote | 51.7 and 30.5 25.6 and 10.4 16.0 and 7.4 |
(Ferreira et al. 2017) | |
| Greenwayodendron suaveolens | Fruit, leave, root bark and stem bark | Dichlorometahne fraction rich in alkaloids Petroleum ether fraction rich in lipids and waxes Methanolic fraction rich in steroids and terpenes |
L. infatum | Promastigote | 24.05, 34.56, 0.63 and 20.32 24.05, 8.0, 27.27 and 5.04 40.32, 32.46, 7.51 and 6.82 |
(Muganza et al. 2016) | |
| Leave, root, stem bark | Crude ethanolic extract | 43.07, 8.11 and 24.05 | |||||
| Stem bark | Polycarpol Dihydropolycarpol Polyathenol |
3.2 8.0 8.1 |
|||||
| Annonaceae species | Used Material | Substances/Extracts | Leishmania spp. | Parasite form | IC50 | Ref. | |
| µM | µg/ml | ||||||
| Isolona hexaloba | Leave Root bark Stem bark |
Aqueous extract Methanolic extract Dichloromethane fraction Dichloromethane fraction |
L. infatum | Promastigote | 2.0 6.35 6.96 8.0 |
(Musuyu Muganza et al. 2015) | |
| Polyathia macropoda | Stem bark | (4S,9R,1OR) methyl 18- carboxy-labda-8, 1 3(E)-diene-15-oate |
L. donovani | Promastigote | 0.75 | (Richomme et al. 1991) | |
| Polyathia suaveolens | stem bark | Methanolic extract | L. infatum | Promastigote | 1.8 | (Lamidi et al. 2005) | |
| Porcelia macrocarpa | Seeds | Docos-13-yn-21-enoic acid 3-hydroxy-4-methylene-2-(eicos-11’-yn-19’-enyl)but-2-enolide 3-hydroxy-4-methylene-2-(octadec-9’-yn-17’-enyl)but-2-enolide 3-hydroxy-4-methylene-2-(hexadec-7’-yn-15’-enyl)but-2-enolide (2S,3R,4R)-3-hydroxy- 4-methyl-2-(eicos-11’-yn-19’-enyl)butanolide Miltefosine (positive control) |
L. infantum | Amastigotes | 48.5 9.2 10.4 11.0 29.9 17.8 |
(Brito et al. 2021) | |
| Porcelia macrocarpa | Seeds | (2S,3R,4R)-3-hydroxy- 4-methyl-2-(n-eicos-11’-yn-19’-enyl)butanolide (1) (2S,3R,4R)-3-hydroxy-4-methyl-2-(n-eicos-11’- ynyl)butanolide (2) Mixture of 1 and 2 2:1 1:1 1:2 |
L. infantum | Amastigotes |
29.9 Non active 8.4 13.6 19.4 |
(Brito et al. 2022) | |
| Annonaceae species | Used Material | Substances/Extracts | Leishmania spp. | Parasite form | IC50 | Ref. | |
| µM | µg/ml | ||||||
| Porcelia macrocarpa | Seeds | (2S,3R,4R)-3-hydroxy-4-methyl-2-(n-eicosyl)butanolide (3) | L. infantum | Amastigotes | Non active | (Brito et al. 2022) | |
| Raimondia monoica | leave | (–)-argentilactone (6S)-(50-oxohepten-10E,30Edienyl)- 5,6-dihydro-2H-pyran-2-one (6R)- (50-oxohepten-10Z,30E-dienyl)-5,6-dihydro-2H-pyran-2-one |
L. panamensis | Promastigote | 51.47 9.2 2.03 |
10.0 1.9 0.42 |
(Carmona et al. 2003) |
| Rollinia emarginata | stem bark |
Hexanic extract Dichloromethane extract Methanolic extract Rollidecin B Rolliniastatin-1 Lirioresinol B Squamocin Liriodenine Sylvaticin |
L. braziliensis, L. amazonensis and L. donovani |
Promastigote |
78.25 8.02 >239.0 8.02 18.16 15.65 |
IC100 >100.0 100.0 >100.0 50.0 5.0 >100.0 5.0 5.0 10.0 |
(Février et al. 1999) |
| Rollinia exsucca | stem | Hexane extract |
L. amazonensis, L. braziliensis and L. donovani |
Promastigote | 20.8 | (Osorio et al. 2007) | |
| Rollina pittieri | leave | Hexane extract Ethyl acetate extract Metanolic extract |
L. amazonensis, L. braziliensis and L. donovani |
Promastigote | 12.6, 10.7 and 10.7 20.8 19.7, 31.4 and 43.8 |
(Osorio et al. 2007) | |
| stem | Hexane extract Ethyl acetate extract |
13.5, 15.1 and 15.1 20.8, 25.0 and 19.7 |
|||||
| Unonopsis buchtienii | Stem bark |
Petroleum ether extract Dichloromethane extract O-methylmoschatoline Lysicamine Fraction containing Unonopsine |
L. major and L. donovani | Promastigote |
155.61 85.82 |
IC100 50.0 100.0 50.0 25.0 25.0 |
(Waechter et al. 1999) |
| Annonaceae species | Used Material | Substances/Extracts | Leishmania spp. | Parasite form | IC50 | Ref. | |
| µM | µg/ml | ||||||
| Unonopsis buchtienii | Stem bark | β-Sitosterol Stigmasterol Liriodenine |
L. major and L. donovani | Promastigote | >241.13 >242.30 11.33 |
>100.0 >100.0 3.12 |
(Waechter et al. 1999) |
| Unonopsis duckei | Twigs, barks and leaves | Alkaloidal fraction | L. amazonensis | Promastigote | 155.61, 32.16 and 4.0 | (da Silva et al. 2012) | |
| Unonopsis guatteriodes | Twigs, barks and leaves | Alkaloidal fraction | L. amazonensis | Promastigote | 1.07, 1.90 and 2.79 | (da Silva et al. 2012) | |
| Uvaria afzelii | root | Bigervone | L. donovani and L. major | Promastigote | 38.9 and 44.4 | (Okpekon et al. 2015) | |
| Uvaria klaineana | stem | Klaivanolide | L. donovani | Promastigote | 1.75 | (Akendengue et al. 2002) | |
| Xylopia aromatica | leave | Methanolic extract |
L. amazonensis, L. braziliensis and L. donovani |
Promastigote | 20.8 | (Osorio et al. 2007) | |
| Xylopia discreta | Leave | Ethanol extract Ether petroleum extract Acetate extract Methanol extract Essential oil |
L. panamensis Promastigote |
25.0 50.0 50.0 37.5 6.25 |
(López et al. 2009) | ||
| seed | Ethanol extract | 6.25 | |||||
| Xylopia excellens | leave | 7β-O-β-D-glucopyranoside-ent-kaur-16-ene | L. amazonensis | Promastigote | 33.74 | 15.23 | (Christopher et al. 2018) |
| Xylopia parviflora | roots | Dichloromethane extract | L. donovani | Amastigote | 5.01 | (Bapela et al. 2017) | |
3.5. Cytotoxic
Cytotoxicity is any form of fatal harm to cells (Costa et al. 2011b). It is said that a cell can be classified as cytotoxic when it has the ability to release harmful substances that culminate in the destruction of other cells (Alberts et al. 2010). Cytotoxic compounds can induce cell death through mechanisms such as necrosis, where there is loss of cell membrane integrity leading to cell death through lysis or rupture (Ferreira et al. 2004); as well as apoptosis, where cell growth is inhibited and there is uncontrolled disruption of cells (Riss and Moravec 2004; Horton and Mathew 2015).
Cytotoxicity tests consist of placing the biological material under study directly or indirectly in contact with a culture of mammalian cells, checking cellular changes by different mechanisms, including the incorporation of vital dyes or the inhibition of the formation of cell colonies (Otręba and Kośmider 2020). The most widely used marker to assess toxicity is cell viability, which can be analyzed with the help of vital dyes, such as neutral red, a water-soluble dye that passes through the cell membrane, settling on the lysosomes (Hadjichristou et al. 2020). The distinction of cell viability is made by measuring the color intensity of the cell culture (Rogero et al. 2003).
Annona genus
Several studies have investigated Annonaceae species in search of novel cytotoxic agents, with a focus on the Annona genus, which was the subject of seven studies. Fatope and collaborators, 1996, evaluated the cytotoxic activity of four ent-kaurenoid derivatives isolated from A. senegalensis stem bark. The brine shrimp lethality test (BST) was conducted according to standard protocols, with LC50 values determined for various fractions and isolated compounds obtained from methanolic bark extracts. These fractions were then tested against human solid tumor cell lines including lung carcinoma (A-549), breast carcinoma (MCF-7), colon adenocarcinoma (HT-29), kidney carcinoma (A-498), prostate adenocarcinoma (PC-3), and pancreatic carcinoma (PACA-2), with Adriamycin as a positive control. The 10% aqueous MeOH partitioned fraction from the stem bark chloroform extract exhibited significant lethality in the BST (LC50 <1.0 µg/mL) and cytotoxicity against multiple solid tumor cell lines (ED50 values of <10-2 µg/mL). ent-Kaur-16- en-19-oic acid displayed activity in the BST (LC50 16 µg/mL) and significant selectivity for MCF-7 cells (breast cancer, ED50 1.0 µg/mL) over the other lines tested. Therefore, ent-Kaur-16- en-19-oic acid demonstrated noteworthy cytotoxic selectivity, warranting further investigations into the structure-activity relationships of this compound class (Fatope et al. 1996).
Silva et al., 2016, evaluated the cytotoxic activity of select carotenoids, flavonoids, and tannins extracted from the leaves and fruits of A. squamosa L. Cytotoxic assays were conducted on Vero cell lines, using the MTT mitochondrial reduction assay to assess cytotoxicity. Additionally, cellular morphological changes were analyzed using May-Grünwald-Giemsa staining. The cytotoxic concentration for 50% of the cell culture (CC50) was calculated. The morphological findings confirmed the apoptotic effects of A. squamosa compounds on tumor cells, with signs of apoptosis observed in cells treated with hexane and aqueous extracts. The aqueous extract of the leaves exhibited higher toxicity than the aqueous extract of the seeds, consistent with its tannin content. Generally, hydrophobic extracts showed higher cytotoxicity than hydroalcoholic extracts (Silva et al. 2016).
Lima and colleagues, 2012, evaluated the cytotoxic potential of leaves and seeds of A. mucosa against Leishmania spp., as well as the oxoaporphine alkaloid liriodenine isolated from leaf dichloromethane extract. The study included in vitro evaluations against promastigote and amastigote forms of Leishmania spp., with the main compound, liriodenine, exhibiting significant cytotoxicity against peritoneal macrophages (De Lima et al. 2012).
Volobuff et al., 2019, conducted bio-guided fractionation of A. cacans Warm extracts to evaluate antioxidant, antiproliferative, and anti-inflammatory activities. Various assays were employed, including DPPH, ABTS, and β-carotene/linoleic acid methods for antioxidant activity, sulforhodamine B assay for cell proliferation, and measurement of myeloperoxidase (MPO) activity for anti-inflammatory activity. The study isolated four compounds and demonstrated significant antioxidant and antiproliferative activities in pulp extracts, with the ethyl acetate fraction exhibiting potent antiproliferative activity against ovarian cancer. Additionally, extracts showed significant inhibition of edema and MPO activity, highlighting the antioxidant and antiproliferative potential of A. cacans Warm compounds (Volobuff et al. 2019).
Two distinct studies examined the cytotoxic profile of A. vepretorum. In the first study, Silva et al., 2017, assessed cytotoxicity and acute toxicity of leaf extracts from the plant, identifying compounds that exhibited high cytotoxic activity against various tumor cell lines. For instance, the methanolic extract and chloroform extract displayed high cytotoxic activity against the HCT-116 cell line, with cell growth inhibitions of 98.16% and 74.28%, respectively, highlighting the significant cytotoxic potential of A. vepretorum extracts (Araújo et al. 2017).
In contrast, Dutra et al., 2014, examined the cytotoxic effects of Ent-kaurane-type diterpenes isolated from the stem bark of A. vepretorum. Among the isolated compounds, a novel derivative exhibited the highest tumor inhibition rate, showing the most potent cytotoxic effect on the K562 cell line, with an IC50 of 2.49 μg/mL, thus highlighting the notable cytotoxic potential of this Annona species (Dutra et al. 2014).
Peña-Hidalgo et al., 2021, assessed the cytotoxic activity of alkaloids from A. crassiflora leaf extract. The study identified compounds through NMR, IR, and Mass Spectrometry methods, with cytotoxic activity evaluated against HCT-116 and MCF-7 cell lines. However, the observed cytotoxic activity was limited for these cell lines (Peña-Hidalgo et al. 2021).
Xylopia genus
The cytotoxicity of Xylopia species has been the focus of several studies, revealing their potential as sources of anticancer agents. Menezes et al., 2016, isolated 19 alkaloids from X. laevigata stem extract, including lanuginosine, (+)-xylopine, and (+)-norglaucine, which exhibited potent cytotoxic activity against various tumor cell lines. Notably, lanuginosine and (+)-xylopine both triggered apoptosis, with (+)-xylopine specifically causing G2/M cell cycle arrest in HepG2 cells. These findings underscore X. laevigata as a promising reservoir of cytotoxic alkaloids (Menezes et al. 2016).
Anadozie et al., 2021, explored the aqueous extract of X. aethiopica fruits, synthesizing gold nanoparticles (AuNPs) and assessing their cytotoxic and antitumor activities. The nanoparticles displayed stability and antioxidant activity, while the extract demonstrated cytotoxicity against breast cancer (MCF-7) and colorectal cancer (MDA-MB and Caco-2 cells), without toxicity to non-cancerous human fibroblastic cells (KMST-6) up to 200 μg/mL. These findings suggest the potential of X. aethiopica as a therapeutic agent against breast and colorectal cancer (Anadozie et al. 2021).
In a study by Tavares et al., 2006, two new Ent-trachylobane-type diterpenes (3β,5β,16α-trihydroxyhalima-13(14)-en-15,16-olide (1) and (-)-8-oxopolyalthiaine(2)) from X. langsdorffiana stem extract exhibited cytotoxicity against V79 and K562 cells, with compound 1 displaying IC50 values of 224 µM and 200 µM, respectively. These results highlight the cytotoxic potential of compounds derived from X. langsdorffiana (Tavares et al. 2006).
Bakarnga-Via et al. (2014) evaluated the cytotoxicity of essential oils from various Xylopia species, including X. aethiopica and X. paviflora, as well as Monodora myristica. All the oils exhibited cytotoxic effects against both cancer (MCF-7) and normal epithelial (ARPE-19) cell lines. Notably, certain oils, such as X. paviflora from Chad and Cameroon, showed greater selectivity, demonstrating higher cytotoxicity against MCF-7 cells compared to ARPE-19 cells. These results highlight the potential of Xylopia essential oils as effective cytotoxic agents against cancer cell (Bakarnga-Via et al. 2014).
Polyalthia genus
Studies focusing on the cytotoxic activity of Polyalthia species have revealed their potential as sources of anticancer agents. Suedee et al., 2007, investigated P. jucunda, isolating four compounds from the stem bark extract. Among them, 24-methylenelanosta-7,9(11)-dien-3-β,15a-diol displayed significant growth inhibitory effects on various tumor cell lines, including breast adenocarcinoma (MCF-7), lung cancer (NCI-H460), and CNS cancer (SF-268), as well as non-tumor human fetal lung cells (MRC-5), indicating its potential for inducing apoptosis in cancer cells (Suedee et al. 2007).
Chen et al., 2000, explored P. longifolia var Pendula, identifying 20 compounds from the methanolic extract, including two novel compounds. Annonaine exhibited notable inhibitory potential against gastric, colon, and hepatoma cancer cell lines, highlighting its cytotoxic activity (Chen et al. 2000).
Similarly, Chang et al., 2006, investigated P. longifolia var Pendula bark extract, isolating 23 compounds, including novel clerodane diterpenes. These compounds demonstrated significant cytotoxicity against hepatoma cell lines (Hep G2 and Hep 3B), suggesting their potential as anticancer agents (Chang et al. 2006).
Tuchinda et al., 2006, evaluated P. crassa leaf and branch extracts, isolating 11 compounds, including novel stilide lactone derivatives. Among them, (+)-Crassalactone A exhibited potent cytotoxic activity against various cancer cell lines, including leukemia, epithelial carcinoma, and lung adenocarcinoma cells. Overall, these studies underscore the cytotoxic potential of Polyalthia species and their constituents, suggesting their promising role in cancer therapy (Tuchinda et al. 2006).
Guatteria and Desmopsis genus
Costa et al., 2020, and Palazzo et al., 2009, conducted studies focusing on the cytotoxic potential of essential oils from various species of the genera Guatteria and Desmopsis within the Annonaceae family. In Costa et al.’s study, the essential oil obtained from G. megalophylla Diels leaves demonstrated significant cytotoxic activity, especially against HL-60 human promyelocytic leukemia cells, with an IC50 of 12.51 μg/mL. This essential oil also demonstrated anti-leukemic activity in vivo, with inhibition rates of the tumor mass ranging from 16.6% to 48.8%. The major constituents identified in this essential oil included spathulenol, γ-muurolene, bicyclogermacrene, β-elemene, and δ-elemene. Similarly, Palazzo et al. evaluated the cytotoxicity of essential oils from species such as D. bibracteata, D. macrocarpa, G. costaricensis, G. diospyroides, G. oliviformis, and Unonopsis costariensis. These essential oils displayed high cytotoxicity against MDA-MB-231 breast adenocarcinoma cells, with ≥ 99% cell death observed at 100 µg/mL for D. bibracteata, G. diospyroides, G. oliviformis, and U. costariensis. Moreover, D. bibracteata essential oil exhibited 100% kill results against Hs 578T breast ductal carcinoma cells at the same concentration. Notably, germacrene D emerged as a predominant compound in the essential oils of G. oliviformis and U. costariensis, further emphasizing their cytotoxic potential. These studies underscore the significant cytotoxic activity of essential oils derived from Annonaceae species, particularly against cancer cell lines and highlight their potential as therapeutic agents in cancer treatment (Palazzo et al. 2009; Costa et al. 2020).
Others Annonaceae species
Other studies have viewed other species of Annonaceae that had smaller occurrences; these species include the Milliusa balanceae, Uvaria pandensis, Diclinanona calycina, Neouvaria acuminatissima, Dasymaschalon blumei, Asimina triloba, Artabotrys zeylanicus, Pseuduvaria trimera (Craib), Anaxagorea dolichocarpa and Duguetia chrysocarpa.
Huong et al., 2005, explored the cytotoxicity of a novel flavone named Miliufavol, along with four other known compounds isolated from the leaves and branches of Miliusa balansae. Among these compounds, paquipodol exhibited strong cytotoxic activity against KB and Hep-G2 cell lines, with IC50 values of 0.7 mg/mL and 0.55 mg/mL, respectively (Huong et al. 2005).
In another study by Maeda et al., 2022, the methanolic extract of Uvaria pandensis Verdc. leaves yielded 12 compounds, including three novel derivatives and two new flavonoids. However, only Pandensenol D and Pandesona A exhibited moderate cytotoxic effects against MCF-7 breast cancer cells, with EC50 values greater than 100 μM (Maeda et al. 2022).
Additionally, Costa et al., 2021, investigated the cytotoxic potential of benzylated dihydroflavones and isoquinoline-derived alkaloids from Diclinanona calycina bark extract. Dichamanetin and a mixture of uvarinol and isouvarinol demonstrated moderate cytotoxic activity against various cancer cell lines, with IC50 values ranging from 9.74 to 25.0 μg/mL. Importantly, they also exhibit low cytotoxicity against non-cancerous cell lines (Costa et al. 2021).
Furthermore, Ik-Soo-Lee et al., 1995, examined labdane-type diterpenes from Neouvaria acuminatissima stem bark, revealing broad cytotoxicity from Acuminolide and 17-O-Acetylacuminolide across several human cancer cell lines, with ED50 values ranging from 10 to 100 μg/mL. However, in vivo testing did not yield significant activity (Ik-Soo Lee et al. 1995).
Lastly, Chanakul et al., 2011, explored the cytotoxic potential of Dasymaschalon blumei extracts, isolating seven compounds with notable cytotoxic effects against various cancer cell lines. Among these compounds, oxodiscoguattine stood out, exhibiting low ED50 values (<19.11 μg/mL) against all tested cell lines (Chanakul et al. 2011).
Woo, Kim, and McLaughlin, 1999, explored the cytotoxicity of compounds from Asimina triloba seeds, isolating two novel acetogenins, asitrilobins A and B, which exhibited high potency against the MIA PaCa-2 pancreatic cancer cell line, with ED50 values of 3.99x10-5 µg/ml and 2.88x10-4 µg/ml, respectively, significantly outperforming the positive control Adriamycin (Woo et al. 1999).
Additionally, Wijeratne et al., 1995, examined the cytotoxic potential of constituents from Artabotrys zeylanicus, highlighting compound N-methoxinorcepharadione A (1) and atherospenine (2), with compound 1 displaying superior inhibitory activity across various cancer cell lines, including RS 322YK (rad52Y), RS 321N, and P-388 (Camptothecin resistant) (Kithsiri Wijeratne et al. 1995).
Sesang et al., 2014, investigated Pseuduvaria trimera, isolating two alkaloids, including a novel compound, 8-hydroxyartabonatin C, which demonstrated cytotoxic activity against HepG2 and MDA-MB231 cancer cells, albeit with lower potency compared to Doxorubicin (Sesang et al. 2014).
Lastly, Pinheiro et al., 2016, studied Anaxagorea dolichocarpa and Duguetia chrysocarpa stem bark extracts, revealing their cytotoxic effects against HCT-116 colon cancer cells, with the hexane extract of A. dolichocarpa showing the highest potency, inhibiting cell growth by 89.02% (Pinheiro et al. 2016).
The studies collectively highlight the diverse cytotoxic activities of compounds from Annonaceae species, suggesting their potential as therapeutic agents against various cancer types. Alkaloids, terpenoids, and flavonoids emerge as the most significant contributors to this cytotoxic bioactivity. These findings further emphasize the therapeutic potential of Annonaceae species in combating cancer, as summarized in Table 5, which lists the tested substances and their respective cytotoxic potential.
Table 5.
Summary of cytotoxic activity of species of the Annonaceae.
| Annonaceae species | Used material | Substances/ Extracts | Methodology | Cell Proliferation inhibition (%) | Ref. | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Anaxagorea dolichocarpa | Stem barks |
Extract Ad-EtOH (Ethanolic) Ad-Hex (Hexanic) Ad-CHCl3 (Chloroform) Ad-AcOEt (Ethyl acetate) |
Human tumor cell lines, including OVCAR-8 (ovarian), SF-295 (brain) and HCT-116 (colon). | OVCAR-8 | SF-295 | HCT-116 | (Pinheiro et al. 2016) | |||||||||||||||||||||||||||||||
| 53.94 44.31 52.51 −5.38 |
65.49 62.68 58.82 0.09 |
50.19 89.02 67.15 3.40 |
||||||||||||||||||||||||||||||||||||
|
Annona cacans Warm |
The fruits and leaves |
PHME-AC (pulp extract) PEAF-AC (ethyl acetate fraction) Acetogenin – PAC-1 Positive control Doxorubicin chloride (0.025–25 lg/mL) |
MCF-7 (breast) OVCAR-3 (ovarian) K-562 (leukemia) |
GI50 - lg/mL | (Volobuff et al. 2019) | |||||||||||||||||||||||||||||||||
| MCF-7 | OVCAR-3 | K-562 | ||||||||||||||||||||||||||||||||||||
| - 11.3 - |
8.8 5.68 6.4 |
6.10 7.84 - |
||||||||||||||||||||||||||||||||||||
| Annona crassiflora | Leaves |
Crassiflorine Xylopine Stephalagine Doxorubicin (Positive control) |
Cells line: HCT-116 – Colon carcinoma MCF-7 – Breast |
IC50 µM | (Peña-Hidalgo et al. 2021) | |||||||||||||||||||||||||||||||||
| HCT-116 | MCF-7 | |||||||||||||||||||||||||||||||||||||
|
143.4 30.2 48.5 0.07 |
Not determined 32.9 Not determined 0.15 |
|||||||||||||||||||||||||||||||||||||
| Annona Mucosa | Leaves and Fruits |
Fraction Hexane Extrat (L); Dichloromethane extract (L); Methanol extract (L); |
Cytotoxicity was studied after 96 h incubation of peritoneal macrophages with concentrations ranging from 6 to 100 µg.mL-1 of each extract and liriodenine. Results are expressed as 50% lethal concentrations (LC50) |
Cytotoxicity LC50 (µg.mL-1) + SEMc | LC50/ IC50 | (De Lima et al. 2012) | ||||||||||||||||||||||||||||||||
| PH8 | M2903 | |||||||||||||||||||||||||||||||||||||
| 62.63 ± 4.10 24.07 ± 4.02 29.41 ± 0.89 |
62.63 ± 4.10 24.07 ± 4.02 29.41 ± 0.89 |
2.58 2.58 1.03 |
||||||||||||||||||||||||||||||||||||
| Annonaceae species | Used material | Substances/ Extracts | Methodology | Cell Proliferation inhibition (%) | Ref. | |||||||||||||||||||||||||||||||||
| Annona Mucosa | Leaves and Fruits |
Fraction Hexane extrat (S); Methanol extrat (S); Compounds Liriodenine Pentamiddine |
Cytotoxicity was studied after 96 h incubation of peritoneal macrophages with concentrations ranging from 6 to 100 µg.mL-1 of each extract and liriodenine. Results are expressed as 50% lethal concentrations (LC50) |
Cytotoxicity LC50 (µg.mL-1) + SEMc | LC50/ IC50 | (De Lima et al. 2012) | ||||||||||||||||||||||||||||||||
| PH8 | M2903 | |||||||||||||||||||||||||||||||||||||
| 262.33 ± 5.81 139.00 ± 3.13 19.11 ± 1.6 51.99 ± 0.58 |
262.33 ± 5.81 139.00 ± 3.13 19.11 ± 1.6 51.99 ± 0.58 |
5.93 2.98 13.36 742.71 |
||||||||||||||||||||||||||||||||||||
| Annona senegalensis Pers. | The bark |
Fractions MeOH Extract Compounds Adriamicina |
BST (brine shrimp lethality test) Human solid tumor cell lines: A-549 (lung carcinoma) MCF-7 (breast carcinoma) HT-29 colon (adenocarcinoma) A-498 (kidney carcinoma) PC-3 (prostate adenocarcinoma) PACA-2 (pancreatic carcinoma) |
ED50 (µg/mL) | (Fatope et al. 1996) | |||||||||||||||||||||||||||||||||
| MeOH Extract | Adriamicina | |||||||||||||||||||||||||||||||||||||
| LC50 <1,0 <10-2 <10-2 1,0 <10-2 <10-2 <10-2 |
- - 1,85 × 10-1 4,37 × 10-2 1 × 10-2 2,23 × 10-2 2,05 ×10-2 |
|||||||||||||||||||||||||||||||||||||
| Anonna squamosa | Mature leaves and fruits |
Leaves aqueous extract (LAq) Seeds aqueous extract (SAq) |
Vero cell lines, (Vero, ATCC-CL 81) kindly provided by Instituto Butantan (São Paulo, Brazil), were used to perform cytotoxic. | Cytotoxicity Assays | (Silva et al. 2016) | |||||||||||||||||||||||||||||||||
| Tree Age | CC50 (mg.mL-1) | |||||||||||||||||||||||||||||||||||||
| 4 14 4 14 |
0.32 0.42 1.43 1.51 |
|||||||||||||||||||||||||||||||||||||
| Annonaceae species | Used material | Substances/ Extracts | Methodology | Cell Proliferation inhibition (%) | Ref. | |||||||||||||||||||||||||||||||||
| Anonna squamosa | Mature leaves and fruits |
Seeds hexane extract (SHex) |
Vero cell lines, (Vero, ATCC-CL 81) kindly provided by Instituto Butantan (São Paulo, Brazil), were used to perform cytotoxic. | Cytotoxicity Assays | (Silva et al. 2016) | |||||||||||||||||||||||||||||||||
| Tree Age | CC50 (mg.mL-1) | |||||||||||||||||||||||||||||||||||||
| 4 14 |
0.49 0.40 |
|||||||||||||||||||||||||||||||||||||
| Annona vepretorum | Stem bark |
Compounds ent-3β-hydroxy-kaur-16-en-19-al ent-3β,19-dihydroxy-kaur-16-eno ent-3β-hydroxy-kaur-16-eno ent-3β-acetoxy-kaur-16-eno ent-3β-hydroxy-kaurenoic acid Kaurenoic acid Positive Control Doxorubicin |
Cytotoxic activities towards tumor and non-tumor cells lines were investigated for compounds 1–6 with Tumor cells: B16-F10, Hep-G2, HL-60, K562 and non-tumor cells: PBMC | IC50 in µg/mL (µM) | (Dutra et al. 2014) | |||||||||||||||||||||||||||||||||
| B16-F10 | Hep-G2 | HL-60 | K562 | PBMC | ||||||||||||||||||||||||||||||||||
| 21.02 >25 19.12 >25 >25 16.56 2.30 |
15.50 >25 19.38 >25 >25 15.33 0.23 |
9.92 >25 9.86 >25 24.21 13.33 0.83 |
2.49 >25 2.94 >25 20.21 21.92 0.68 |
7.20 8.93 6.49 >25 >25 24.41 5.09 |
||||||||||||||||||||||||||||||||||
| Annona vepretorum | Leaves |
Extract Av-MeOH Av-HexC Av-Hex |
Human tumor cell lines were plated in 96-well plates: HCT-116, SF-295, HL-60, Sarcoma-180. The concentration that caused 50% cell growth inhibition (IC50) was determined from the concentration-response curves by non-linear regression with a confidence interval of 95%. |
Cell proliferation inhibition (%) | (Araújo et al. 2017) | |||||||||||||||||||||||||||||||||
| HCT-116 | SF-295 | HL-60 | Sarcoma-180 | IC50 (µM) | ||||||||||||||||||||||||||||||||||
| 98.16 ±0.92 29.96 ±1.60 56.04 ±21.0 |
63.98 ±4.84 86.54 ±3.31 65.43 ±6.52 |
82.23 ±4.84 17.11 ±7.34 55.85 ±3.56 |
82.34 ±1.36 79.43 ±4.39 86.24 ±1.09 |
2.81 ±0.41 4.87 ±0.83 45.82 ±9.07 |
||||||||||||||||||||||||||||||||||
| Annonaceae species | Used material | Substances/ Extracts | Methodology | Cell Proliferation inhibition (%) | Ref. | |||||||||||||||||||||||||||||||||
| Annona vepretorum | Leaves |
Av-CHCl3 AV-AcOEt Av-H2O |
Human tumor cell lines were plated in 96-well plates: HCT-116, SF-295, HL-60, Sarcoma-180. The concentration that caused 50% cell growth inhibition (IC50) was determined from the concentration-response curves by non-linear regression with a confidence interval of 95%. |
HCT-116 | SF-295 | HL-60 | Sarcoma-180 | IC50 (µM) | (Araújo et al. 2017) | |||||||||||||||||||||||||||||
| 74.28 ±0.25 17.8 7±6.45 9.52 ±11.68 |
82.05 ±24.67 27.98 ±5.16 6.72 ±1.25 |
29.79 ±1.82 68.72 ±38.28 -8.20 ±3.33 |
81.32 ±6.79 63.15 ±6.57 78.57 ±5.91 |
2.88 ±1.39 22.82 ±3.76 71.18 ±1.69 |
||||||||||||||||||||||||||||||||||
| Artabotrys zeylanicus | Not specified |
Compounds N-methoxynorcepharadione A (1) Atherospennidine (2) Positive control Camptothecin |
Cytotoxicity assessed by IC50 by assay with cells: RS 322YK (rad52Y) RS 321N RS 188N (rad+) P-388 (wild-type) P-388 (Camptothecin resistant) |
IC50 (µg/ml) | (Kithsiri Wijeratne et al. 1995) | |||||||||||||||||||||||||||||||||
| 1 | 2 | Camptothecin | ||||||||||||||||||||||||||||||||||||
| 2.16 1.20 >200 159 1.12 |
16 27 >50 Not tested Not tested |
0.6 - 100 0.012 >20 |
||||||||||||||||||||||||||||||||||||
| Asimina triloba | Seeds |
Acetogenins 1 2 Adramicyn |
Cytotoxicity tests against human tumor cell lines: A-549 (human lung carcinoma), MCF-7 (human breast carcinoma), HT-29 (human colon adenocarcinoma), A-498 (human kidney carcinoma), PC-3 (human prostate adenocarcinoma) and MIA PaCa-2 (human pancreatic carcinoma). | Human cancer cell line (ED50 µg/ml) | (Woo et al. 1999) | |||||||||||||||||||||||||||||||||
| BST LC50 | A-549 | MCF-7 | HT-29 | A-498 | PC-3 | MIA PaCa-2 | ||||||||||||||||||||||||||||||||
| 0.131 0.00429 Not tested |
0.00439 0.00165 0.0174 |
0.00211 0.00169 0.440 |
2.09 0.440 0.0116 |
2.78 2.19 0.0116 |
2.28 1.06 0.0461 |
0.000039 0.000028 0.00781 |
||||||||||||||||||||||||||||||||
|
Desmopsis bibracteata Desmopsis macrocarpa |
Leaves | Essential oil | Human MDA-MB-231 breast adenocarcinoma cells and Human Hs 578T breast ductal carcinoma cells. | % Kill at 100 µg/mL | (Palazzo et al. 2009) | |||||||||||||||||||||||||||||||||
| MDA-MB-231 | Hs 578T | |||||||||||||||||||||||||||||||||||||
| 99.3 (0.7) 53.0 (9.6) |
100 8.2 (14.0) |
|||||||||||||||||||||||||||||||||||||
| Annonaceae species | Used material | Substances/ Extracts | Methodology | Cell Proliferation inhibition (%) | Ref. | |||||||||||||||||||||||||||||||||
| Dasymaschalon blumei | The combined leaves and twigs Stems |
Extract: Acetate de ethyl extract Acetate de ethyl extract Compounds: 3,5-diydroxy-2,4-dimethoxyaristolactam Aristolactam BI Goniopedaline Griffithinam Oxodiscoguattine Dicentrinone Duguevalline Positive control Elipticine |
Cell culture: P-388 (mouse lymphoid neoplasm), KB (human epidermoid carcinoma in the mouth), Col-2 (human colon cancer), MCF-7 (human breast cancer), Lu-1 (human lung cancer), ASK (rat glioma), Hek 293 (noncancerous human embryonic kidney cell). The Hek 293 cell assay was used as a primary assay for assessing the specificity of an anticancer agent toward cancer cell lines in comparison with the normal mammalian cell. |
ED50 values (μg/ml) | (Chanakul et al. 2011) | |||||||||||||||||||||||||||||||||
| P-388 | KB | Col-2 | MCF-7 | Lu-1 | ASK | Hek 293 | ||||||||||||||||||||||||||||||||
| <4 <4 2.13 11.18 2.59 13.82 0.60 10.28 9.43 0.65 |
<4 4.12 2.97 - 1.98 - 2.30 4.56 - 0.62 |
17.38 - - - - - 0.91 7.34 - 0.65 |
- - - 3.60 9.45 - 2.74 9.05 19.11 0.69 |
16.46 14.04 - - - - 0.76 4.0 - 0.17 |
- - 3.04 - 12.76 - 2.11 10.20 - 0.61 |
- - 14.75 - 17.97 - 1.6 3.76 - 0.56 |
||||||||||||||||||||||||||||||||
|
Diclinanona calycina |
Barks |
Compounds Thalifoline (S)-(+)-Reticuline 1S,2R-Reticuline Nβ-oxide 1S,2S-Reticuline Nα-oxide Bisnorargemonine Isochamanetin Dichamanetin Uvarinol + Isouvarinol Doxorubicin (Positive control) |
Cancer cells HL-60 – Human promyelocytic leukemia MCF-7 – Breast adenocarcinoma HepG2 – hepatocellular carcinoma HCT116 – colon carcinoma Non-cancerous cell: MRC-5 – Human lung fibroblast |
µM | (Costa et al. 2021) | |||||||||||||||||||||||||||||||||
| HL-60 | MCF-7 | HCT116 | HepG2 | MRC-5 | ||||||||||||||||||||||||||||||||||
| Not determined Not determined Not determined Not determined Not determined Not determined 15.78 9.74 0.04 |
Not determined Not determined Not determined Not determined Not determined Not determined 23.59 >25 3.08 |
>25 >25 >25 >25 >25 >25 18.99 17.31 0.85 |
20.08 22.54 23.11 >25 >25 19.79 >25 >25 2.05 |
>25 >25 >25 >25 >25 24.69 >25 >25 3.19 |
||||||||||||||||||||||||||||||||||
| Annonaceae species | Used material | Substances/ Extracts | Methodology | Cell Proliferation inhibition (%) | Ref. | |||||||||||||||||||||||||||||||||
| Duguetia chrysocarpa | Stem barks |
Dc-EtOH (Ethanolic) Dc-Hex (Hexanic) Dc-CHCl3 (Chloroform) Dc-AcOEt (Ethyl acetate) |
Human tumor cell lines, including OVCAR-8 (ovarian), SF-295 (brain) and HCT-116 (colon). | OVCAR-8 | SF-295 | HCT-116 | (Pinheiro et al. 2016) | |||||||||||||||||||||||||||||||
| 49.65 18.39 42.24 13.01 |
43.95 34.84 63.17 30.99 |
60.16 15.61 59.35 9.76 |
||||||||||||||||||||||||||||||||||||
|
Guatteria costaricensis Guatteria diospyroides Guatteria oliviformis |
Leaves | Essential oil | Human MDA-MB-231 breast adenocarcinoma cells and Human Hs 578T breast ductal carcinoma cells. | % Kill at 100 µg/mL | (Palazzo et al. 2009) | |||||||||||||||||||||||||||||||||
| MDA-MB-231 | Hs 578T | |||||||||||||||||||||||||||||||||||||
| 54.6 (5.7) 98.8 (1.2) 100 |
0 21.1 (8.2) 35.6 (1.9) |
|||||||||||||||||||||||||||||||||||||
| Guatteria megalophylla Diels | Leaves |
Essential oil (EO) Positive Control Doxorubicin (DOX) 5-Fluorouracil (5-FU) |
Toxicity Assays with Human cancer cell lines: HL-60 Promyelocytic leukemia; MCF-7 Breast adenocarcinoma; Cal27 Oral squamous cell carcinoma; HSC-3 Oral squamous cell carcinoma; HepG2 Hepatocellular carcinoma; HCT116 Colon carcinoma and Human non-cancer cell line: MRC-5 Lung fibroblast. | IC50 (µg/mL) | (Costa et al. 2020) | |||||||||||||||||||||||||||||||||
| HL-60 | MCF-7 | CAL-27 | HSC-3 | Hep-g2 | HCT116 | MRC-5 | ||||||||||||||||||||||||||||||||
| 12.51 0.02 1.85 |
33.45 6.16 10.13 |
7.58 1.09 2.56 |
14.90 0.86 1.01 |
21.62 0.02 13.71 |
30.07 0.02 0.53 |
29.85 3.32 5.96 |
||||||||||||||||||||||||||||||||
| Miliusa balansae | Leaves and branches |
Flavonoids Ombuine Crysosplenol B Pachypodol Crysosplenol C Control Eliptin (Sigma) |
Cell culture: IC50 (µg/mL) - KB (Human Epidermoide Carcinom); - Hep-G2 (Hepatoma G2); - RD (Rhabdosarcoma). |
KB | Hep-G2 | RD | (Huong et al. 2005) | |||||||||||||||||||||||||||||||
|
˃ 5 4.6 0.7 4.3 0.002 |
1.5 0.93 0.55 0.57 0.001 |
˃ 5 ˃ 5 3.01 2.09 0.001 |
||||||||||||||||||||||||||||||||||||
| Annonaceae species | Used material | Substances/ Extracts | Methodology | Cell Proliferation inhibition (%) | Ref. | |||||||||||||||||||||||||||||||||
| Neouvaria acuminatissima | Stem bark |
Compounds Acuminolide (1) 17-O-Acetylacuminolide (2) Spiroacuminolide (3) Positive Control Doxorubicin |
Screened for cytotoxicity against a panel of human cancer cell lines and murine P388 cells, according to established protocols. 14 ED50 values of >4 ~tg/ml were regarded as negative. Among the cell lines represented, a human lung cancer cell line (Lul) was used to guide the fractionation against the HT-29 human colorectal and KB human epidermoid carcinoma models, Doxorubicin was run as a positive control. | ED50 Values: tg/ml | (Ik-Soo Lee et al. 1995) | |||||||||||||||||||||||||||||||||
| Compounds I and 2 were broadly cytotoxic, exhibiting ED50 values, ranging from 10 -~ to 10 ° ttg/ml in several cell lines. With the human cell lines, the most potent activity was observed with melanoma (Mel2) (ED50: 0.7 ttg/ml) and prostate (LNCaP) (ED50: 0.8 ~tg/ml) cells for compounds 1 and 2, respectively. Compound 3 was not significantly active for any of the cancer cell lines tested. Acuminolide (1) was inactive when tested in vivo against a HT-29 human colorectal xenograft model in nude mice at 40-60 mg/kg (maximum tolerated dose 70 mg/kg). 17-O-Acetylacuminolide (2) showed no significant activity when tested in vivo against a KB human epidermoid carcinoma murine model at 110 mg/kg. | ||||||||||||||||||||||||||||||||||||||
| Polyalthia crassa | Leaves and twigs |
Compounds (+)-Crassalactone A (+)-Crassalactone B (+)-Crassalactone C (+)-Crassalactone D Aristolactam AII (+)-tricinnamate Positive control Ellipticine |
Cytotoxicity assays of compounds 1-4, 10, and 11 were performed employing the colorimetric method. | Cell line (ED50 µg/mL) | (Tuchinda et al. 2006) | |||||||||||||||||||||||||||||||||
| P-388 | KB | Col-2 | BCA-1 | Lu-1 | ASK | |||||||||||||||||||||||||||||||||
| 0.18 3.8 >5 1.1 2.7 3.1 0.52 |
1.7 >5 >5 3.3 >5 >5 0.65 |
1.9 >5 >5 4.0 >5 >5 0.53 |
0.92 >5 >5 3.2 >5 >5 0.53 |
1.9 >5 >5 >5 >5 >5 0.56 |
1.6 >5 >5 3.1 >5 >5 0.60 |
|||||||||||||||||||||||||||||||||
| Polyalthia jucunda | Dried and powdered stem bark. |
Compound 4-Hydroxy-4,7-dimethyl-α-tetralone 4,5-Dihydroblu- menol A N-trans-feruloyltyramine 24–Methy- lenelanosta-7,9(11)-dien-3-β,15α-diol |
The effects of compounds on the growth of the human tumor and non–tumor cell lines were evaluated according to the procedure adopted by the National Cancer Institute (USA) for the in vitro anticancer drug discovery |
GI50 (µM) | (Suedee et al. 2007) | |||||||||||||||||||||||||||||||||
| MCF-7 | MDA-MB-231 | SF-268 | NCL-H460 | MRC-5 | ||||||||||||||||||||||||||||||||||
| >150 >150 >150 19.3±1.2 |
>150 >150 >150 18.8±2.0 |
>150 >150 >150 21.8±0.6 |
>150 >150 >150 23.0±1.7 |
>150 >150 >150 40.3±3.4 |
||||||||||||||||||||||||||||||||||
| Annonaceae species | Used material | Substances/ Extracts | Methodology | Cell Proliferation inhibition (%) | Ref. | |||||||||||||||||||||||||||||||||
| Polyalthia longifolia var. pendula | Leaves |
16α-hydroxycleroda3,13-dien-15,16-olide 5-hydroxy-6- Methoxyonychine (-)-anonnaine |
Tested against four human cancer cell lines: AGS (gastric cancer cells), DLD1 (colon cancer cells), HepG2 (hepatoma cells), and HA59T (hepatoma cells). | IC50 (µM) | (Chen et al. 2000) | |||||||||||||||||||||||||||||||||
| AGS | DLD | HA597 | HepG2 | |||||||||||||||||||||||||||||||||||
| 26.9 >30 8.6 |
>30 >30 28.9 |
23.6 21.7 16.4 |
>30 >30 20.8 |
|||||||||||||||||||||||||||||||||||
| Polyalthia longifólia var. pendula | The bark samples |
Compounds 16(R&S)- 3,13Z-kolavadien-15,16-olide-2-one 16-hydroxycleroda- 3,13-dien-15,16-olide 16-hydroxycleroda-4(18),13-dien- 15,16-olide 16-oxocleroda-3,13(14)E-dien-15-oic acid methyl ester Solidagonal acid (4→2)-abeo- 16(R&S)-2,13Z-clerodadien-15,16-olide-3-al labd-13E-en-8-ol-15-oic acid Polylongine Liriodenine Lysicamine (+)-Stepharine (−)-Stepholidine N-trans-feruloyltyramine N-trans-p-coumaroyltyramine Positive control Doxorubicin |
Tested against four human cancer cell lines: MCF-7, MDA-MB-231, Hep-G2, Hep 38. | IC50 (µM) | (Chang et al. 2006) | |||||||||||||||||||||||||||||||||
| MCF-7 | MDA-MB-231 | Hep-G2 | Hep38 | |||||||||||||||||||||||||||||||||||
| 18.28 14.42 10.43 14.34 18.12 11.89 - 10.41 4.46 8.94 9.40 16.56 25.53 17.35 0.04 |
4.50 8.29 3.22 13.22 14.67 11.65 - 9.94 10.28 16.75 9.90 - 25.54 - 0.32 |
2.88 4.42 3.35 - - 2.36 18.33 - - - - - 21.17 - 0.18 |
2.96 2.83 1.97 - - 8.94 15.40 - - - - - 24.98 - 0.23 |
|||||||||||||||||||||||||||||||||||
| Annonaceae species | Used material | Substances/ Extracts | Methodology | Cell Proliferation inhibition (%) | Ref. | |||||||||||||||||||||||||||||||||
| Pseuduvaria trimera (Craib) | Leaves And twigs |
8-hydroxyartabonatine C Ouregidione Positive control Doxorubicin |
The human hepatocellular carcinoma HepG2 and breast cancer MDA-MB231 cells. | IC50 | (Sesang et al. 2014) | |||||||||||||||||||||||||||||||||
| HepG2 | MDA-MB231 | |||||||||||||||||||||||||||||||||||||
| Mean | SD | Mean | SD | |||||||||||||||||||||||||||||||||||
| 26.36 12.88 2.21 |
±5.18 ±2.49 ±1.72 |
64.75 67.06 1.83 |
±4.45 ±3.5 ±0.09 |
|||||||||||||||||||||||||||||||||||
| Uvaria pandensis | Leaves |
Pandensenol D Pandensone A (8’α,9’β-dihydroxy)-3-farnesylindole (6’,7’-dihydro-8’α,9’β-dihydroxy)- 3-farnesylindole |
Cell culture: MCF-7 (breast cancer cells) |
EC50 (µM) | (Maeda et al. 2022) | |||||||||||||||||||||||||||||||||
|
>523.4 349.8 117.1 >563.4 | ||||||||||||||||||||||||||||||||||||||
| Unonopsis costariensis | Leaves |
Essential oil |
Human MDA-MB-231 breast adenocarcinoma cells and Human Hs 578T breast ductal carcinoma cells. |
% Kill at 100 µg/mL | (Palazzo et al. 2009) | |||||||||||||||||||||||||||||||||
| MDA-MB-231 | Hs 578T | |||||||||||||||||||||||||||||||||||||
| 100 | 17.3 (10.3) | |||||||||||||||||||||||||||||||||||||
| Xylopia aethiopica | Fruits |
Aqueous extract Aqueous extrat in gold nanoparticles Positive control Fulvestrant 5-Fluororacil |
Cell culture: IC50 (µg/mL) - MCF-7 (Bresat cancer); - MDA-MB-231; - Caco-2 cells. |
MCF-7 | MDA-MB-231 | Caco-2 | (Anadozie et al. 2021) | |||||||||||||||||||||||||||||||
| 171.3 >200 >120 nM - |
94.5 141.4 <120 nM - |
199.8 >200 - >100 |
||||||||||||||||||||||||||||||||||||
| Xylopia langsdorffiana | Stems |
Compound ent-7r-Acetoxytrachyloban-18-oic acid |
Cytotoxic activity of compound 1 was evaluated against V79 cells and rat hepatocytes using the MTT method. | IC50 (µM) | (Tavares et al. 2006) |
|||||||||||||||||||||||||||||||||
| V79 | K562 | |||||||||||||||||||||||||||||||||||||
| 224 and 231 µM | 200 µM | |||||||||||||||||||||||||||||||||||||
| Annonaceae species | Used material | Substances/ Extracts | Methodology | Cell Proliferation inhibition (%) | Ref. | |||||||||||||||||||||||||||||||||
|
Xylopia aethiopica (Dunal) A. Rich; Xylopia paviflora A. Rich (Benth.); |
Fruits |
Essential oil Composition: Monoterpenes hydrocarbons; Oxygenated monoterpenes; Sesquiterpene hydrocarbons; Oxygenated sesquiterpenes. |
The cytotoxic activity of all essential oils was evaluated on human breast cancer (MCF-7) and normal epithelial (ARPE-19) cell lines using the MTT assay based on cell viability. Cells were exposed to the oils at concentrations ranging from 0.1 to 2 μL/mL. | IC50 values μL/mL | (Bakarnga-Via et al. 2014) | |||||||||||||||||||||||||||||||||
| The six essential oils exerted cytotoxic activity against cancer (MCF-7) and normal cell lines (ARPE-19), with more pronounced effect on neoplastic cells in most cases. The highest selectivity was obtained with the essential oils of X. parviflora from Chad and Cameroon (5.87 and 5.54) which were more cytotoxic against MCF-7 than against normal cell line (ARPE-19) with IC50 values of 0.155 μL/mL and 0.166 μL/mL respectively | ||||||||||||||||||||||||||||||||||||||
| Xylopia laevigata | Leaves |
Compounds (-)-Roemerine (+)-Anonaine Lanuginosine (+)-Glaucine (+)-Xylopine Oxoglaucine (+)-Norglaucine (-)-Xylopinine (+)-Norpurpureine (+)-N-Methyllaurotetanine (+)-Norpredicentrine (+)-Discretine (+)-Calycinine (+)-Laurotetanine (+)-Reticuline (-)-Corytenchine (+)-Discretamine (+)-Flavinantine Positive Control Doxorubicin |
B16-F10, HepG2, K562 and HL-60 tumor cell lines were kindly donated by Hospital A.C. Camargo, São Paulo, Brazil. Cell viability was quantified using the Alamar Blue assay. | IC50 µg/mL (µM) | (Menezes et al. 2016) | |||||||||||||||||||||||||||||||||
| B16-F10 | HepG2 | HL60 | K562 | PBMC | ||||||||||||||||||||||||||||||||||
| NA 18.80 8.46 NA 3.77 19.14 8.48 NA 21.08 NA NA 16.15 22.17 NA NA NA 18.80 NA 0.08 |
NA 14.04 3.89 NA 1.87 NA 3.78 NA NA NA NA 7.89 NA NA 15.35 NA 14.04 NA 0.08 |
NA 10.09 7.81 NA 1.87 5.90 6.84 NA 10.11 NA NA 12.97 18.59 NA 23.81 NA 10.09 NA 0.09 |
NA 10.62 6.61 NA 3.12 12.48 7.84 NA 16.72 NA NA 14.85 NA NA NA NA 10.62 NA 0.15 |
NA NA 24.53 NA 4.08 10.25 6.70 NA 17.94 NA NA NA NA NA NA NA NA NA 2.47 |
||||||||||||||||||||||||||||||||||
3.6. NA - Not Active Instead (IC50 > 25µg/mL)Antitumor
Cancer is a heterogeneous disease caused by a series of “genetic” alterations selected clonally in tumor suppressor genes and oncogenes (Aguirre-Ghiso 2007). However, evidence accumulated in recent years indicates that the heterogeneity of tumor cells is partly due to the contributions of “epigenetic” changes in cancer cells (Ducasse and Brown 2006). Thus, it is believed that cancer is the manifestation of genetic and epigenetic changes (Esteller 2008; Ellis et al. 2009). Despite a few examples of genetic inheritance of tumorogenesis, it is believed that most cancers can result from changes that accumulate throughout life due to exposure to various endogenous factors such as nutrients, infections, physical activity, social behavior and other environmental factors (Link et al. 2010).
In Brazil, according to data from the National Cancer Institute (INCA, 2019) the disease is already emerging as the second leading cause of mortality in the country (Instituto Nacional de Câncer José Alencar Gomes da Silva (INCA) 2019). In this way, innumerable anticancer drugs have been introduced in therapy in the last decades, many of which were obtained by plant screening programs. Considering the high incidence of cancer and the toxic effects that antineoplastic drugs have on normal cells, it is important to highlight the role that natural products have played in the search for efficient alternatives for antineoplastic therapy (Zhang et al. 2019; Núcleo de estudos e Pesquisas de Produtos Naturais 2020; Twilley et al. 2020).
Thus, some authors conducted research on species of the Annonaceae family in search of new antitumor agents. The predominant species evaluated include Annona, Guatteria and Xylopia. Through these studies, the tumor inhibition potential of Annonaceae derived compounds is evident, where the top five studies addressed the action of acetogenins, terpenoids, and alkaloids. Table 6 shows the substances tested in each study as well as their respective antitumor potential.
Annona genus
Rinaldi et al., 2017, discovered that a crude extract of Annona hypoglauca Mart., commonly known in Brazil as “beribá,” exhibits cytotoxic activity against human cancer cells. Through Gas Chromatography–Mass Spectrometry (GC/MS) analysis, four aporphine alkaloids—actinodaphnine, anonaine, isoboldine, and nornuciferine—were identified as responsible for the antitumoral activity. In cytotoxicity assays, the crude extract demonstrated a lethal effect against breast and colon cancer cells (Rinaldi et al. 2017).
The total alkaloid fraction showed significant cytotoxicity against MCF-7 (−8.90% lethality) and showed cytotoxicity against SF-268 (26.7% growth inhibition), against NCIH461 (29.0% growth inhibition) and against KM-12 (67.6% growth inhibition). Additionally, both the fraction containing Isoboldine and the fraction containing Actinodaphnine displayed activity against the breast cancer cell line. In contrast, alkaloid-free fractions did not demonstrate significant activity against cancer cell lines (Rinaldi et al. 2017).
Chen and colleagues, 2011, examined the antitumor effects of acetogenins (ACGs) derived from the leaves of Annona squamosa Linn. Their study involved using S180 and HepS xenograft-bearing mice to test the antitumor activity. The results demonstrated that some ACGs effectively inhibited tumor growth in a dose-dependent manner, with a particular selectivity for HepS. Among the acetogenins evaluated, adjacent bis-THF ACGs showed greater activity compared to mono-THF and nonadjacent bis-THF ACGs against both HepS and S180. Conversely, nonadjacent bis-THF ACGs were more effective than mono-THF ACGs against S180, while mono-THF ACGs proved more potent than nonadjacent bis-THF ACGs against HepS (Chen et al. 2012).
Additionally, the cytotoxic activity of the fruit pericarp of the same species was investigated using in vitro cultures of Dalton’s lymphoma cells and HeLa cells. The results revealed that the chloroform extract of Annona squamosa pericarp exhibited cytotoxicity against the tested cell lines. The inhibitory concentration required for 50% cytotoxicity (IC50) was also determined. Through bioactivity-directed isolation, two diterpenoids, namely (-)-ent-kaur-16-en-19-oic acid and 16a,17-dihydroxy-ent-kauran-19-oic acid, were identified as the compounds responsible for the observed cytotoxic activity (Joy and Remani 2008).
In the volatile oil extracted from fresh leaves of Annona leptopetala, spathulenol was found to be the major component, comprising 12.56% of the total. This oil was utilized to assess its antitumor effects through both in vitro (using the sulforhodamine B assay) and in vivo (sarcoma 180 murine tumor model) experiments. In the in vitro study, the volatile oil demonstrated antitumor activity, particularly against the leukemia cell line (K-562), with a total growth inhibition (TGI) concentration of 0.64 µg/ml when tested up to 250 µg/ml. For the in vivo experiments, the 50% lethal dose in mice was approximately 447.2 mg/kg when administered intraperitoneally. Furthermore, the inhibition rates for Sarcoma 180 tumor growth were 59.29% and 58.77% at doses of 100 and 150 mg/kg intraperitoneally, respectively (Brito et al. 2018).
Leaves of Annona vepretorum Mart. were utilized to extract an essential oil, primarily composed of bicyclogermacrene (35.71%), spathulenol (18.89%), (E)-β-ocimene (12.46%), α-phellandrene (8.08%), o-cymene (6.24%), germacrene D (3.27%), and α-pinene (2.18%). The in vitro cytotoxicity of the essential oil and some of its major constituents was assessed in tumor cell lines representing different histotypes, using the alamar blue assay. Notably, both the essential oil and spathulenol demonstrated promising cytotoxic effects (Bomfim et al. 2016).
In vivo experiments revealed that the treatment with the essential oil led to a 34.46% inhibition of tumor growth. Prominently, when the essential oil was complexed with β-cyclodextrin in a microencapsulation, there was a significant increase in in vivo tumor growth inhibition, reaching 62.66% (Bomfim et al. 2016).
Among the other Annona species highlighted are Annona muricata and Annona crassiflora. Annona muricata, or graviola, boasts over 212 phytochemicals, including annonaceous acetogenins, alkaloids, flavonoids, and sterols. Extensively researched for therapeutic potential, graviola exhibits several biological activities including antitumoral properties. In an in vivo rodent study, Annona muricata leaf extract demonstrated a 59.8% inhibition of pancreatic cancer cell growth and metastasis induced by CD18/HPAF cells. The ethanolic extract selectively induced cytotoxicity in three tumor cell lines without affecting normal spleen cells. The extract inhibited EGFR overexpression, EGFR mRNA expression, induced G0/G1 phase cell cycle arrest, and activated caspase-3-mediated apoptosis. In athymic mice, it inhibited MDA-MB-468 tumor growth by 32%, reducing protein expression of EGFR, p-ERK, and p-EGFR. The 80% aqueous ethanol leaf extract suppressed tumor initiation and promotion even at lower dosages, showcasing its multifaceted potential in cancer treatment (Rady et al. 2018).
Using the methanolic extract of Annona crassiflora, the antimutagenic evaluation in the micronucleus test showed a damage reduction of 75.00 and 64.58% for the pre-treatment and simultaneous protocols, respectively. The post-treatment protocol enhanced the effects of cyclophosphamide by 45.83%. In contrast, pre-treatment with 15 mg/L of the extract resulted in a significant reduction in the mitotic index, decreasing it by 45.95%. This specific protocol did not effectively reduce MMS-induced toxicity. However, all other protocols and treatments, across various concentrations, led to an increased mitotic index. This implies that the Annona crassiflora methanolic extract has the potential to reverse the toxicity induced by MMS treatment in cultures (Rocha et al. 2016).
Xylopia genus
The leaf essential oil of Xylopia frutescens contains significant compounds, including (E)-caryophyllene (31.48%), bicyclogermacrene (15.13%), germacrene D (9.66%), δ-cadinene (5.44%), viridiflorene (5.09%), and α-copaene (4.35%), which were identified by GC/FID and GC/MS. In vitro cytotoxicity assays conducted on NCI-H358M and PC-3M tumor cells showed that the essential oil had IC50 values ranging from 24.6 μg/mL to 40.0 μg/mL. After, the in vivo evaluation using Sarcoma 180-bearing mice was performed. The essential oil (dosed at 50 and 100 mg/kg/day) inhibited tumor growth with rates by 31.0–37.5%. Notably, it demonstrated anticancer effects without significant toxicity, presenting a potential alternative for cancer therapy. The essential oil’s composition, coupled with its in vitro and in vivo outcomes, underscores its promising role in cancer treatment (Ferraz et al. 2013).
Cavalcanti et al., 2010, explored the genotoxic and mutagenic potential of kaurenoic acid, isolated from dried roots of Xylopia sericeae St. Hill, using diverse in vitro and in vivo methods. They investigated structure–activity relationships for two natural diterpenoids and three semi-synthetic derivatives of kaurenoic acid. The study unveiled genotoxic and mutagenic effects in human blood cells, yeast, and mice for some compounds, potentially attributed to DNA double-strand breaks or topoisomerase I inhibition. Intriguingly, certain compounds, including kaurenoic acid, exhibited no such effects, pointing to the exocyclic double bond (C16) as the active genotoxic moiety in kaurenoic acid derivatives (Cavalcanti et al. 2010).
The essential oil extracted from the leaves of Xylopia laevigata revealed compounds with inhibitory effects against tumor cells in culture, as well as on tumor growth in vivo. The essential oil exhibited consistent chemical composition, with major constituents identified as γ-muurolene, δ-cadinene, germacrene, α-copaene, germacrene D, bicyclogermacrene, and (E)-caryophyllene. In vitro tests showed that the essential oil has cytotoxicity across tested tumor cell lines, showing consistent profiles without hemolytic or genotoxic effects. After, the in vivo studies revealed tumor growth inhibition rates ranging from 37.3% to 42.5%. Essential oil treatment had no significant impact on body weight, organ macroscopy, or blood leukocyte counts (Quintans et al. 2013).
Guatteria genus
Santos et al., 2017, evaluated diverse essential oils extracted from the aerial parts of various Guatteria species, including G. australis, G. ferruginea, G. latifolia, and G. sellowiana. Following this, they investigated the antiproliferative activity of these extracts on a panel of tumor cell lines, measuring it as the concentration required for complete inhibition of cell growth, expressed as total growth inhibition (TGI) (Santos et al. 2017).
The results showed that while the essential oil from G. sellowiana demonstrated limited activity (TGI > 50 μg/ml) against most tumor cell lines, exceptions were observed for leukemia (K562, TGI = 1.1 μg/ml) and ovarian adenocarcinoma (OVCAR-03, TGI = 4.1 μg/ml) cell lines. The essential oils extracted from G. latifolia, G. ferruginea, and G. australis exhibited pronounced selective effects on OVCAR-03 cells, with TGIs of 1.1, 1.8, and 3.2 μg/ml, respectively. Additionally, they displayed intriguing effects on the multiresistant ovarian adenocarcinoma cell line (NCI-ADR/RES) with TGIs of 10.0, 34.6, and 15.2 μg/ml, respectively (Santos et al. 2017).
Guatteria elliptica, another species under examination for its antitumoral properties, underwent assessment using essential oils extracted from leaves sourced from distinct regions in Sao Paulo, Brazil. Notably, the essential oil from Paranapiacaba demonstrated significant antitumor activity against breast (IC50 = 7.01 μg/mL) and prostate (IC50 = 5.35 μg/mL) cancer cells. Importantly, it exhibited low cytotoxicity towards normal fibroblast cells (IC10 = 18.55 μg/mL). Spathulenol, the primary compound isolated from this essential oil, displayed notable efficacy against MCF-7 (5.38 μg/mL and PC-3 (2.25 μg/mL) (Rajca Ferreira et al. 2018).
Mitrephora genus
Bioassay-guided fractionation of Mitrephora thorelii led to the discovery of two clerodane-type diterpenes, 6α,16,18-trihydroxycleroda-3(4),13(14)-dien-15,16-olide and 16-hydroxycleroda-3(4),13(14)-dien-15,16-olide. Both compounds exhibited significant inhibitory activity against human hepatoma BEL-7402 cells in vitro. Compound 16-hydroxycleroda-3(4),13(14)-dien-15,16-olide demonstrated promising in vivo anti-tumor effects, inhibiting hepatoma H22 growth by 30.7% in mice. In vitro, two clerodane-types diterpenes inhibited BEL-7402 cell proliferation with IC50 values of 44.6 (6α,16,18-trihydroxycleroda-3(4),13(14)-dien-15,16-olide) and 20.1 μM (16-hydroxycleroda-3(4),13(14)-dien-15,16-olide). The compound 16-hydroxycleroda-3(4),13(14)-dien-15,16-olide, was well-tolerated in mice and represents a potential natural anti-tumor agent (Meng et al. 2007).
Mitrephora glabra stem bark underwent bioactivity-guided fractionation, yielding nine compounds, including three new ent-kaurenoids, five polyacetylenic acids/esters, and the alkaloid liriodenine. Evaluation against cancer cell lines and microorganisms revealed that ent-kaurane diterpenoids were inactive (IC50 >10 μM), contrasting with more potent ent-trachylobane diterpenoids from the same plant. Polyacetylenes exhibited varying IC50 values (10–40 μM), with the compound methyloropheate displaying no cytotoxicity, suggesting the methyl ester diminished its activity. Notably, the alkaloid liriodenine showed significant cytotoxicity (IC50 close to 5 μM) against all tested cell lines (Li et al. 2009).
Others Annonaceae species
Among the other genus of Annonaceae which have showed antitumoral activity are Asimina, Anaxagorea, Polyalthia, Duguetia and Miliusa. Kim and collaborators, 2005, evaluated the antitumor activity of acetogenins obtained from the leaves of Asimina triloba Possessing. The isolated compounds were tested on A-549 (human lung carcinoma), MCF-7 (human breast carcinoma), HT-29 (human colon adenocarcinoma), A-498 (human kidney carcinoma), PC-3 (human prostate adenocarcinoma), and MIA PaCa-2 (human pancreatic carcinoma) cells, with adriamycin as a positive control. The results revealed that asimitrin was selectively cytotoxic against prostate adenocarcinoma (PC3) and 104 times more potent than the control. Hydroxytrilobin was equally more active against colon adenocarcinoma (HT-29) than adriamycin. The acetogenins exert their in vivo antitumor effects, in part, by inhibiting complex I of the electron transport system in the mitochondria and by blocking the NADH oxidase enzyme particular to the plasma membranes of cancerous cells (Kim et al. 2005).
The antitumor activity of Annonaceae-derived alkaloids was also the objective of Suassuna et al., 2011. In this study, the authors evaluated the activity of alkaloids isolated from the stem bark of Anaxagorea dolichocarpa. The alkaloids eupolauramine and sampangine demonstrated strong antitumor activity against K569 cells. In addition, the anticancer potential of Polyalthia evecta was evaluated using leaves extracts (Machana et al. 2012). Cytotoxicity against HepG2 and apoptosis induction were systematically examined, unveiling heightened efficacy in 50% ethanol-water crude leaf extract compared to its fractions. Notably, a hexane extract exhibited significant effects, albeit surpassed by the ethanol-water crude leaf extract. The amalgamation of water and hexane extracts demonstrated augmented cytotoxicity and apoptosis induction, presenting a twofold increase in % apoptotic cells compared to the sole hexane extract. The authors suggest the indispensable role of the polar extract fraction in the anticancer activity of the non-polar extract fraction (Lúcio et al. 2011).
The compounds Duguetine and duguetine β-N-oxide displayed significant antitumoral effects across all cell lines when utilizing an alkaloid extract and five isolated alkaloids from the subterranean stem bark of Duguetia furfuracea. The alkaloid extract, containing the compounds duguetine and duguetine β-N-oxide, demonstrated notable cytotoxicity against cancer cell lines. These two compounds displayed low IC50 values, with duguetine β-N-oxide (IC50 7.27 μM) outperforming duguetine (IC50 12.39 μM) in MDA/MB-435 (da Silva et al. 2009).
Fissistigma cavaleriei (Levl) Rehd underwent a bioassay-guided investigation by Yang et al., 2012. Using the dried roots, they identified an isoindolin derivative displaying antiangiogenic properties, selective COX-2 inhibition, and in vitro cytotoxicity against tumor cells. In vivo, this compound effectively hindered S-180 cell growth in mice, synergistically enhancing doxorubicin’s antitumor efficacy. The authors propose its role as a multi-target molecule, urging further exploration for its potential as a lead in tumor treatment. Administered at 20 mg/kg, 36 mg/kg, and 64 mg/kg daily, the isoindolin derivative significantly reduced tumor weights in sarcoma 180-bearing mice to 2.33 ± 0.32 g, 1.87 ± 0.22 g, and 1.15 ± 0.11 g, respectively, after an 8-day regimen (Yang et al. 2012).
Finally, a novel compound named Miliufavol, along with known flavones such as ombuine, chrysosplenol B, pachypodol, and chrysosplenol C, was extracted and identified from the leaves and branches of Miliusa balansae. The extraction process involved multiple cycles with MeOH––H2O. The cytotoxic effects of the known compounds were assessed against cancer cell lines KB (human epidermoid carcinoma), Hep-G2, and RD (Rhabdomyosarcoma). Results indicated activity against all three tested cell lines for each of the four compounds. Particularly noteworthy was pachypodol, demonstrating significant potency against two cell lines (KB: 0.7 mg/ml, Hep-G2: 0.55 mg/ml). These findings underscore the remarkable cytotoxic potential of pachypodol and its counterparts (Huong et al. 2005).
Table 6.
Summary of antitumor activity of species of the Annonaceae.
| Annonaceae species | Used material | Substances/ Extracts |
Methodology | Inhibition | Ref. | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Annona crassiflora | The leaves | A. crassiflora (AC) | To evaluate the antimutagenic/ chemopreventive activity through the Allium cepa test, we used 5, 10, and 15 mg/L of extract, and for the micronucleus test in the peripheral blood, we used the dose of 15 mg/kg. | Chromosomal aberrations | (Rocha et al. 2016) | ||||||||||||||||
| Treatment | Damage Reduction Percentages of mitotic index |
Total (values compared with the control group) |
Damage Reduction Percentages | ||||||||||||||||||
| Mutagenicity AC – 5mg/L AC – 10mg/L AC – 15mg/L Control group (distilled water) Positive control MMS (Methyl methanesulfonate) Antimutagenicity AC – 5mg/L Pre-treatment Simple simultaneous Simultaneous with pre-incubation Post-treatment Continuous AC – 10mg/L Pre-treatment Simple simultaneous Simultaneous with pre-incubation |
−1.62 −11.35 −29.46 3.51 - −31.35 −44.86 −2.53 −58.38 −37.03 −64.05 −15.95 −35.68 |
31 27 34 238 40 - - - |
- - - - - −66.17 72.72 100.5 89.39 86.36 75.75 33.33 93.93 |
||||||||||||||||||
| Annonaceae species | Used material |
Substances/ Extracts |
Methodology | Inhibition | Ref. | ||||||||||||||||
| Annona crassiflora | The leaves | A. crassiflora (AC) | To evaluate the antimutagenic/ chemopreventive activity through the Allium cepa test, we used 5, 10, and 15 mg/L of extract, and for the micronucleus test in the peripheral blood, we used the dose of 15 mg/kg. | Chromosomal aberrations | (Rocha et al. 2016) | ||||||||||||||||
| Treatment | Damage Reduction Percentages of mitotic index |
Total (values compared with the control group) |
Damage Reduction Percentages | ||||||||||||||||||
| AC – 10mg/L Post-treatment Continuous AC – 10mg/L Pre-treatment Simple simultaneous Simultaneous with pre-incubation Post-treatment Continuous |
−21.35 −37.03 45.95 −23.78 −16.49 −36.76 −44.59 |
- |
79.79 81.31 69.19 22.22 102.52 84.34 93.43 |
||||||||||||||||||
| Annona hypoglauca | Stem |
Crude extract Fraction hexane Dichloromethane/Methanol Fraction ethyl acetate Fraction butanol Total alkaloid fraction |
Human tumor cell lines (MCF-7, breast adenocarcinoma; KM-12, colon adenocarcinoma; RPMI-8226, multiple myeloma; PC-3, prostate carcinoma; SF-268 glioblastoma and NCI-H460, non-small lung-cell carcinoma). | MCF-7 | PC-3 | NCI-H460 | KM-12 | SF-268 | RPMI-8226 | (Rinaldi et al. 2017) | |||||||||||
| −32.80 NI 2.90 Not tested 11.90 −8.90 |
11.4 77.4 59.6 67.1 46.6 17.04 |
1.3 48.1 44.9 38.6 65.6 71.7 |
−2.5 NI NI NI NI 32.4 |
58.5 NI 93.7 77.4 NI 73.3 |
17.3 53.5 53.4 NI 83.8 NI |
||||||||||||||||
| Annonaceae species | Used material |
Substances/ Extracts |
Methodology | Inhibition | Ref. | ||||||||||||||||
| Annona hypoglauca | Stem |
Fraction alkaloid 4.4 Fraction alkaloid 5 Fraction alkaloid 9 Positive control Doxorubicin |
Human tumor cell lines (MCF-7, breast adenocarcinoma; KM-12, colon adenocarcinoma; RPMI-8226, multiple myeloma; PC-3, prostate carcinoma; SF-268 glioblastoma and NCI-H460, non-small lung-cell carcinoma). | MCF-7 | PC-3 | NCI-H460 | KM-12 | SF-268 | RPMI-8226 | (Rinaldi et al. 2017) | |||||||||||
|
Not tested -11.60 -3.10 -16.31 |
-3.8 Not tested Not tested -50.0 |
-26.0 Not tested Not tested -46.2 |
11.7 -14.4 -8.4 -5.1 |
45.4 Not tested Not tested -34.0 |
29.7 Not tested Not tested -14.3 |
||||||||||||||||
| Annona leptopetala | Leaves | Essential oil Annona leptopetala leaves (ALO) Doxurubicin (DOX) Positive control |
The tumor cell lines used were: U251 – glioma, MCF-7 – breast, NCI/ADR-RES - multidrug-resistant ovarian, 786-0 – kidney, NCI-H460 – non-small cell lung cancer, PC-3 – prostate, OVCAR – ovarian, HT29 – colon and K562 – leukemia, and HaCaT human keratinocytes served as the normal cell line. Sarcoma 180 tumor cells were maintained in the peritoneal cavity of Swiss mice. | Total inhibition of cancer cells proliferation (µg/ml) | (Brito et al. 2018) | ||||||||||||||||
| Cell Lines | DOX | ALO | |||||||||||||||||||
| Glioma (U251) Breast (MCF-7) Ovary Multidrug Resistance Phenotype (NCI-ADR/RES) Kidney (786-O) Lung (NCI-H460) Prostate (PC-3) Ovary (OVCAR) Colon (HT-29) |
0.06 0.21 1.35 0.04 0.01 0.27 0.26 0.22 |
47.23 49.91 >250 101.52 75.53 45.12 >250 75.26 |
|||||||||||||||||||
| Annonaceae species | Used material |
Substances/ Extracts |
Methodology | Inhibition | Ref. | ||||||||||||||||
| Annona leptopetala | Leaves | Essential oil Annona leptopetala leaves (ALO) Doxurubicin (DOX) Positive control |
The tumor cell lines used were: U251 – glioma, MCF-7 – breast, NCI/ADR-RES - multidrug-resistant ovarian, 786-0 – kidney, NCI-H460 – non-small cell lung cancer, PC-3 – prostate, OVCAR – ovarian, HT29 – colon and K562 – leukemia, and HaCaT human keratinocytes served as the normal cell line. Sarcoma 180 tumor cells were maintained in the peritoneal cavity of Swiss mice. | Total inhibition of cancer cells proliferation (µg/ml) | (Brito et al. 2018) | ||||||||||||||||
| Cell Lines | DOX | ALO | |||||||||||||||||||
|
Leukemia (K-562) Skin (line of non-tumor cells) (HaCat) |
0.40 0.23 |
0.64 >250 |
|||||||||||||||||||
| Annona muricata | Leaf, Seed, Fruit, Pericarp, Twing, Root |
Acetogenins, extracts and fractions | This current review demonstrates A. muricata’s anticancer potential and other health-related benefits by providing insights into its bioactive chemical constituents as well as the in vitro and in vivo studies that have been carried out to elucidate the molecular mechanisms of action of these constituents. |
Review Article Acetogenins or other A. muricata-derived compounds could be tested as monotherapy or as sensitizers in combination with standard cancer treatments for cancer patients. |
(Rady et al. 2018) | ||||||||||||||||
| Annona squamosa | Seeds |
Compound Squamostatiin A |
The antitumor activities of 1–5 and standard control taxol against the growth of S180 and HepS in mice were measured by methods reported previously. | Treatment effects of annonaceous acetogenins in the Heps and S180 xenograft tumor model | (Chen et al. 2012) | ||||||||||||||||
| Treatment group | Dose (µg/kg) | HepS | S180 | ||||||||||||||||||
| Tumor weight (g: mean±SD) | Inhibition ratio (%) | Tumor weight (g: mean±SD) | Inhibition ratio (%) | ||||||||||||||||||
| Squamostatin A | 15 60 |
0.96±0.19 0.79±0.12 |
15.0 31.2 |
0.36±0.09 0.34±0.10 |
52.7 54.2 |
||||||||||||||||
| Annonaceae species | Used material |
Substances/ Extracts |
Methodology | Inhibition | Ref. | ||||||||||||||||
| Annona squamosa | Seeds |
Compounds Squamostatiin E 4-deoxyannoreticuin Desacetyluvaricin Bullatacin Taxol Positive control |
The antitumor activities of 1–5 and standard control taxol against the growth of S180 and HepS in mice were measured by methods reported previously. | Treatment effects of annonaceous acetogenins in the Heps and S180 xenograft tumor model | (Chen et al. 2012) | ||||||||||||||||
| Treatment group | Dose (µg/kg) | HepS | S180 | ||||||||||||||||||
| Tumor weight (g: mean±SD) | Inhibition ratio (%) | Tumor weight (g: mean±SD) | Inhibition ratio (%) | ||||||||||||||||||
| Control group Taxol Squamostatin E 4-deoxyannoreticuin Desacetyluvaricin Bullatacin |
40 15 60 15 60 15 60 15 60 |
1.13±0.46 0.43±0.15 0.75±0.16 0.55±0.18 0.43±0.12 0.37±0.14 0.77±0.18 0.33±0.22 0.38±0.18 - |
- 62.2 34.2 51.3 61.5 67.3 32.5 70.9 63.4 - |
0.75±0.16 0.38±0.12 0.55±0.10 0.37±0.08 0.42±0.10 0.39±0.11 0.43±0.12 0.27±0.08 0.26±0.06 - |
- 48.9 27.2 51.1 43.9 48.3 42.3 63.9 65.8 - |
||||||||||||||||
| Annona squamosa | Fruits |
Compounds (-)-entkaur-16-en-19-oic acid (1) 16-α,17- dihydroxy-ent-kauran-19-oic acid (2) |
Cytotoxic activity using in vitro cultures of Dalton’s lymphoma cells as well as HeLa cells. Cytotoxicity was detected by the Trypan blue exclusion test and induction of apoptosis was evaluated by [3-(4,5-Dimethylthiazol-2- yl)-2,5 diphenyltetrazolium bromide] assay (MTS assay) and DNA ladder assay. The inhibitory concentration required for 50% cytotoxicity (IC50) was also determined. | (%) of cytotoxicity |
(Joy and Remani 2008) |
||||||||||||||||
| There was an increase in the percentage cytotoxicity with increasing concentrations of the fraction containing the compounds. However a slight decrease in activity with increasing incubation time was noted, which implies that the compound has very high cytotoxicity even after 24 h of incubation. Even at a concentration of 1.65 lg/ml, the compounds exhibited more than 50% cytotoxicity after 24 h. The cytotoxic activity for different time intervals is compared. The highest activity was noted after 24 h, which implies that the efficacy of secondary metabolites as cytotoxic agents at a low dose and a short duration. | |||||||||||||||||||||
| Annonaceae species | Used material |
Substances/ Extracts |
Methodology | Inhibition | Ref. | ||||||||||||||||
|
Annona vepretorum |
Leaves |
Essential Oil Spathulenol α-Phellandrene σ- Cymene α- Pinene 5-Fluorouracil Positive control |
The in vivo antitumour effect was evaluated in C57BL/6 mice inoculated with B16-F10 melanoma. Tumour cells (2 9 106 cells per 500 lL) were implanted subcutaneously into the left hind groin of mice. Animals were euthanized by cervical dislocation, and tumours were excised and weighed. Drug effects are expressed as the per cent inhibition of control. | IC50 (µg/mL) | (Bomfim et al. 2016) | ||||||||||||||||
| B16-F10 | HepG2 | K562 | HL-60 | PBMCs | |||||||||||||||||
| 9.90 7.81-12.55 11.67 9.76-13.96 15.44 6.54-36.42 >25 11.46 5.46-24.04 0.68 0.21-1.45 |
10.60 8.55-13.26 11.19 9.58-13.07 17.30 13.89-21.55 >25 13.05 9.79-17.38 0.04 0.01-1.22 |
8.43 5.48-12.97 3.79 1.48-9.70 >25 >25 14.00 10.56-18.55 0.15 0.01-1.86 |
6.14 4.15-9.12 11.38 8.46-15.31 20.18 16.91-24.08 >25 14.96 12.25-18.26 0.29 0.21-0.38 |
22.82 19.18-27.15 15.59 13.12-18.53 >25 >25 >25 14.00 9.83-23.31 |
|||||||||||||||||
| Asimina triloba | Seeds | Compounds Asimitrin 4-Hydroxytrilobin Adriamicyn Positive control |
In vitro cytotoxicity tests against human tumor cell lines s for A-549 (human lung carcinoma),26 MCF-7 (human breast carcinoma),27 HT-29 (human colon adenocarcinoma),28 A-498 (human kidney carcinoma),26 PC-3 (human prostate adenocarcinoma),29 and MIA PaCa-2 (human pancreatic carcinoma) | Human cancer cell line ED50 (µg/mL) | (Kim et al. 2005) | ||||||||||||||||
| Cell lines | Asimitrin | 4-Hydroxytrilobin | Adriamicyn | ||||||||||||||||||
| BST A-549 MCF-7 HT-29 A-498 PC-3 MIA PaCa-2 |
2.07x10-2 1.19 2.12 1.19x10-4 7.50x10-1 1.72x10-6 2.11x10-4 |
7.00x10-2 1.54 3.79 1.54x10-6 3.62x10-2 2.01x10-4 2.01x10-4 |
NT 6.22x10-4 9.53x10-1 2.87x10-2 2.86x10-3 5.77x10-2 6.10x10-3 |
||||||||||||||||||
| Annonaceae species | Used material |
Substances/ Extracts |
Methodology | Inhibition | Ref. | ||||||||||||||||
| Anaxagorea dolichocarpa Sprague & Sandwith | The stem bark | Alkaloids Compounds Eupolauramine (1) Sampangine (2) |
The cytotoxity was evaluated through the MTT reduction assay, which determines the number of living cells able to reduce the yellow dye 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) to formazan. The determination of the 50% inhibition concentration for cell growth (IC50) of The human leukemic strain K562. | IC50 (µg/mL) | (Lúcio et al. 2011) | ||||||||||||||||
| The in vitro effects of eupolauramine (1) and sampangine (2) against the K562 cell line were determined in three experiments and in quadruplicate. Both compounds exhibited concentration-dependent inhibitory effect on the proliferation of K562 cells. The IC50 values were 18.97 (17.06–21.10) μg/mL and 10.95 (10.15–11.80) μg/mL respectively. | |||||||||||||||||||||
|
Duguetia furfuracea |
Bark |
Duguetine Duguetine β-N-oxide Dicentrinone N-methyltetrahydropalmatine N-methylglaucine Alkaloid Extract Doxorrubicin Positive control |
The cytotoxic potential of alkaloids was evaluated by the MTT assay. Against three human tumor cell lines: SF-295 (glyoblastoma), HCT-8 (colon cancer) and MDA/MB-435 (melanoma). | Percentage of inhibition (%) | (da Silva et al. 2009) | ||||||||||||||||
| HCT-8 (%) | SF-295 (%) | MDA/MB-435 (%) | |||||||||||||||||||
| 91.1±0.8 92.0±1.0 68.0±0.7 57.6±0.7 45.2±2.1 90.2±1.1 93.3±1.7 |
86.0±1.1 87.1±0.7 50.3±1.5 39.8±0.7 25.1±2.3 85.1±0.6 92.3±2.2 |
98.1±0.1 84.5±1.6 37.5±4.0 31.1±0.8 15.1±6.4 99.6±0.4 97.0±1.1 |
|||||||||||||||||||
| Annonaceae species | Used material |
Substances/ Extracts |
Methodology | Inhibition | Ref. | ||||||||||||||||
| Fissistigma cavaleriei | The dried roots |
5-methoxy-2-methylisoindolin-1-yl, 4-methoxyphenyl Doxorubicin Positive control |
Evaluation of the antiproliferative effect of compound 1 under study against cells: K562, S-180, A549. | IC50 (mol/L) | (Yang et al. 2012) | ||||||||||||||||
| K562 | S-180 | A549 | |||||||||||||||||||
|
2.54±0.22x10-5 1.16±0.14x10-6 |
7.26±0.24x10-5 1.61±0.11x10-7 |
8.76±0.18x10-5 8.12±0.12x10-7 |
|||||||||||||||||||
|
Guatteria autralis Guatteria ferruginea Guatteria latifolia Guatteria sellowiana |
The aerial parts | Essential Oils (EO) Doxorubicin Positive control |
The antiproliferative activity of the materials tested was evaluated using eight human tumor cell lines: U251 (central nervous system, CNS, glioma), MCF-7 (breast cancer) NCI-ADR/RES (ovarian tumor with multidrug resistance phenotype), 786-0 (kidney cancer), NCI-H460 (non-small-cell lung cancer), OVCAR-03 (ovarian carcinoma), HT-29 (colorectal cancer), and K562 (leukemia). Using the concentration-response curve for each cell line. | Total growth inhibition (TGI) [µg/ml] | (Santos et al. 2017) | ||||||||||||||||
| Cell lines | EO G. australis | EO G. ferruginea | EO G. latifolia | EO G. sellowiana | Doxorubicin | ||||||||||||||||
| U251 MCF-7 NCI-ADR/RES 786-0 NCI-H460 OVCAR-3 HT-29 K562 HaCat |
40.4 37.8 15.2 45.6 49.5 3.2 38.6 86.2 48.0 |
36.3 37.6 34.6 50.9 69.4 1.8 52.7 18.6 63.4 |
36.6 47.2 10.0 63.9 44.9 1.1 39.7 15.6 41.0 |
89.1 95.5 250 107.8 82.8 4.1 143.1 1.1 75.6 |
2.7 0.88 >25 3.1 >25 11.7 3.6 0.031 1.0 |
||||||||||||||||
| Annonaceae species | Used material |
Substances/ Extracts |
Methodology | Inhibition | Ref. | ||||||||||||||||
| Guatteria elliptica R. E. Fries | Leaves |
Essential oil Spathulenol |
The antitumor against the normal cell line derived from mouse fibroblasts (BALB/c 3T3, ATCC CCL163) was tested using the MTS method. The IC50 values were used to determine the median lethal dose (LD50) for cell lines MCF-7 (human breast cancer) and PC-3 (human prostate cancer. | IC50 ± SE (µg/mL) | (Rajca Ferreira et al. 2018) | ||||||||||||||||
| PC-3 | MCF-7 | ||||||||||||||||||||
| 5.32 ± 0.35 2.25 ± 0.28 |
7.01 ± 0.23 5,38 ± 0.20 |
||||||||||||||||||||
| Miliusa balansae | Leaves and branches |
Ombuine Crysosplenol B Pachypodol Crysosplenol C Eliptin Positive Control |
Cell culture: - KB (Human Epidermoide Carcinom); - Hep-G2 (Hepatoma G2); - RD (Rhabdosarcoma). |
IC50 (µg/mL) | (Huong et al. 2005) | ||||||||||||||||
| KB | Hep-G2 | RD | |||||||||||||||||||
|
˃ 5 4.6 0.7 4.3 0.002 |
1.5 0.93 0.55 0.57 0.001 |
˃ 5 ˃ 5 3.01 2.09 0.001 |
|||||||||||||||||||
| Mitrephora glabra | Stem Bark | 4-epi-kaurenoic acid (1) Mitrekaurenone (2) Methylmitrekaurenate (3) Oropheic acid (4) Methylloropheate (5) Octadeca-9,11,13-triynoic acid (6) Oropheolide (7) 9,10-Dehydrooropheolide (8) |
The cytotoxicity measurements against the KB human oral epidermoid carcinoma: MCF-7 human breast carcinoma, NCI-H460 human large cell lung carcinoma, and SF-268 human astrocytoma. | IC50 µM | (Li et al. 2009) | ||||||||||||||||
| The ent-kaurane diterpenoids (1-3), all were inactive (IC50 values >10 µM); the diterpenoids of the ent-trachylobane class, reported previously from the same plant, were more potent in this regard. The polyacetylenes (4 and 6-8) gave IC50 values ranging from 10 to 40 µM. However, compound 5 was completely inactive, suggesting that the methyl ester diminishes cytotoxicity. The known alkaloid liriodenine (9) was the most cytotoxic against all four of the cell lines in which it was evaluated (IC50 value ∼5 µM). | |||||||||||||||||||||
| Annonaceae species | Used material |
Substances/ Extracts |
Methodology | Inhibition | Ref. | ||||||||||||||||
| Mitrephora thorelii | The aerial parts | 6a,16,18-Trihydroxycleroda-3(4),13(14)-dien-15,16-olide (1) 16-Hydroxycleroda-3(4),13(14)-dien-15,16-olide (2) Cyclophosphamide Positive control |
In vivo evaluation female mice (5–6 weeks old) of Kunming strain were purchased from Shanghai SLAC Laboratory Animal Co. (Shanghai, China). Murine hepatoma H22 is maintained by serial intraperitoneal passage in Kunming mice. H22 cells were subcutaneously implanted into Kunming mice at Diterpenes from M. thorelii 683 1 £ 106 cells/mouse. | IC50 mM | (Meng et al. 2007) | ||||||||||||||||
| Compounds 1 and 2 inhibited the proliferation of BEL-7402 cells in vitro with the IC50 values 44.6 and 20.1 mM, respectively. In vivo anti-tumor effect of compound 2 was further evaluated in murine hepatoma H22 model. It was found that compound 2 significantly inhibited the growth of hepatoma H22 with the percentage inhibition of 30.7% (P, 0.05 vs control). The alkylation agent cyclophosphamide served as a positive control (63.8%, P, 0.01 vs control). The mice were well tolerated towards compound 2, and no significant loss of body weight was observed compared with control group (P. 0.05, data not shown). | |||||||||||||||||||||
| Polyalthia evecta | Leaves |
Hexane fraction (F1) 500 (µg/mL) Chloroform fraction (F2) 500 (µg/mL) Ethylacetate fraction (F3) 500 (µg/mL) Dichloromethane fraction (F4) 500 (µg/mL) Ethanol fraction (F5) 500 (µg/mL) |
Determination the cytotoxicity of the samples in the cell model, neutral red (NR) uptake assay was used for identification of vital cells. The cytotoxicity assays were performed with HepG2 and Vero cells. | % Cytotoxicity | % Apoptotic cells in HepG2 | (Machana et al. 2012) | |||||||||||||||
| HepG2 | Vero | ||||||||||||||||||||
|
74.6 ± 1.6 24.3 ± 9.3 29.6 ± 8.8 24.0 ± 7.4 22.7 ± 8.8 |
54.2 ± 15.4 2.2 ± 3.8 32.5 ± 8.1 25.5 ± 8.1 7.6 ± 8.3 |
46.4 ± 2.6 9.8 ± 4.7 3.2 ± 1.6 2.9 ± 1.1 2.4 ± 1.9 |
|||||||||||||||||||
| Annonaceae species | Used material |
Substances/ Extracts |
Methodology | Inhibition | Ref. | ||||||||||||||||
| Polyalthia evecta | Leaves |
Methanol fraction (F6) 500 (µg/mL) Water fraction (F7) 500 (µg/mL) Hexane: water (500:100) Hexane: water (500:250) Hexane: water (500:500) Hexane: methanol (500: 100) Hexane: methanol (500: 250) Hexane: methanol (500:500) P. evecta crude extract (EW-L) 140 (µg/mL) P. evecta crude extract (EW-L) 500 (µg/mL) |
Determination the cytotoxicity of the samples in the cell model, neutral red (NR) uptake assay was used for identification of vital cells. The cytotoxicity assays were performed with HepG2 and Vero cells. | % Cytotoxicity | % Apoptotic cells in HepG2 | (Machana et al. 2012) | |||||||||||||||
| HepG2 | Vero | ||||||||||||||||||||
|
9.9 ± 2.5 7.8 ± 2.5 98.9 ± 1.8 81.8 ± 10.6 100.0 ± 1.0 91.0 ± 6.2 100.0 ± 5.7 100.0 ± 8.9 60.0 ± 4.8 100.0 ± 4.1 |
12.3 ± 1.3 10.8 ± 0.8 4.0 ± 2.9 43.9 ± 3.3 45.1 ± 4.1 26.9 ± 5.6 36.1 ± 8.0 39.9 ± 3.2 30.2 ± 9.3 37.2 ± 4.2 |
4.6 ± 1.6 9.2 ± 2.1 n.d. n.d. 72.7 ± 13.6 n.d. n.d. 54.9 ± 10.8 46.4 ± 2.6 92.8 ± 10.8 |
|||||||||||||||||||
| Annonaceae species | Used material |
Substances/ Extracts |
Methodology | Inhibition | Ref. | ||||||||||||||||
| Polyalthia evecta | Leaves |
Melphalan 76 (µg/mL) |
Determination the cytotoxicity of the samples in the cell model, neutral red (NR) uptake assay was used for identification of vital cells. The cytotoxicity assays were performed with HepG2 and Vero cells. | % Cytotoxicity | % Apoptotic cells in HepG2 | (Machana et al. 2012) | |||||||||||||||
| HepG2 | Vero | ||||||||||||||||||||
| 67.2 ± 3.1 | 70.3 ± 3.1 | 41.6 ± 2.1 |
|||||||||||||||||||
| Polyalthia longifolia var. pendula | Stems | 15 compounds isolated were evaluated for cytotoxicity against MCF-7 (human breast carcinoma) and A549 (non-small cell lung cancer) cells with cell viabilities assessed using a MTT assay. | Cytotoxicity assessment was performed with Human breast carcinoma (MCF-7) cells and non-small cell lung cancer (A549) cells. | IC50 µM | (Lee et al. 2009) | ||||||||||||||||
| Compounds tested, only 16-oxo-cleroda3,13-dien-15-oic acid (8) was cytotoxic against both MCF-7 and A549 cell lines, with IC50 values of 3.7 (0.2 and 3.1 (0.3 µM, respectively. Under the same conditions, the IC50 values of the corresponding positive controls, paclitaxel, and doxorubicin, were 0.0020 (0.0001 and 0.837 (0.034 µM, respectively. | |||||||||||||||||||||
|
Xylopia frutescens Aubl. |
Leaves |
Essential oil Doxorubicin Positive Control |
The in vivo antitumor effect was evaluated using Sarcoma 180 ascites tumor cells. Ten-day-old Sarcoma 180 ascites tumor cells (2 106 cells per 500 ll) were implanted subcutaneously into the left hind groin of mice. | IC50 (µg/mL) | |||||||||||||||||
| OVCAR-8 | NCI-H358M | PC-3M |
(Ferraz et al. 2013) |
||||||||||||||||||
| 33.9 24.9-46.3 1.2 0.9-1.6 |
24.6 14.9-40.7 0.9 0.6-1.3 |
40.0 31.3-51.2 1.6 1.1-2.4 |
|||||||||||||||||||
|
Xylopia laevigata |
Leaves |
5% DMSO Essential oil 5-Fluororacil Positive control |
The in vivo antitumor effect was evaluated using sarcoma 180 ascites tumor cells. Ten-day-old sarcoma 180 ascites tumor cells were implanted subcutaneously into the left hind groin of mice. | Antitumor activity sarcoma 180 ascites tumor cells | (Quintans et al. 2013) | ||||||||||||||||
| Dose (mg/kg/day) | Tumor (g) | Inhibition (%) | |||||||||||||||||||
| - 50 100 25 |
1.97±0.14 1.24±0.09 1.13±0.27 0.63±0.16 |
- 37.3 42.5 67.8 |
|||||||||||||||||||
| Annonaceae species | Used material |
Substances/ Extracts |
Methodology | Inhibition | Ref. | ||||||||||||||||
| Xylopia sericea | Dried roots |
Compounds kauren-19-oic acid (KA); (−)-kauran-19-oic acid (KAH). |
The human cancer cell-lines used in this work were HL60 and K562 (leukemias), MDA-MB435 (melanoma) and SF295 (glioblastoma). The growth of tumour cells and PBLs was quantified by the ability of living cells to reduce the yellow dye 3-(4,5-dimethyl-2-thiozolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) to a purple formazan product. | Mitotic index, frequency of chromosomal aberrations and numeric changes in human lymphocytes in culture after kaurenoic acid (KA) and their hydrogenated derivative (KAH) treatments. | (Cavalcanti et al. 2010) | ||||||||||||||||
| Substance | Treatment | Mitotic index | Aberrant cells | ||||||||||||||||||
| % | Mean±S.E.M | % | Mean±S.E.M | ||||||||||||||||||
| MMS (Positive control) DMSO (Vehicle) KA KAH |
4x10-5 M 0.1% 2.5µg/mL 5.0 µg/mL 10.0 µg/mL 30.0 µg/mL 60.0 µg/mL 2.5 µg/mL |
2.8 2.1 2.5 4.1 4.0 3.5 4.3 4.1 4.3 4.0 3.8 4.2 3.3 3.0 3.1 2.3 1.8 2.6 1.3 1.7 1.1 4.4 4.0 4.0 |
2.46±0.3 3.86±0.3 4.23±0.1 4.0±0.2 3.13±0.1 2.23±0.4 1.36±0.3 4.13±0.2 |
5.3 8.3 5.0 0.7 0.0 0.7 0.0 0.0 0.0 0.0 0.0 0.0 0.7 0.0 0.7 5.3 4.7 5.3 4.3 5.0 6.2 0.0 0.0 0.0 |
6.2±1.8 0.46±0.4 0.0 0.23±0.4 0.46±0.4 5.1±0.3 5.3±1.2 0.0 |
||||||||||||||||
| Annonaceae species | Used material |
Substances/ Extracts |
Methodology | Inhibition | Ref. | ||||||||||||||||
|
Xylopia sericea |
Dried roots |
(−)-kauran-19-oic acid (KAH). |
The human cancer cell-lines used in this work were HL60 and K562 (leukemias), MDA-MB435 (melanoma) and SF295 (glioblastoma). The growth of tumour cells and PBLs was quantified by the ability of living cells to reduce the yellow dye 3-(4,5-dimethyl-2-thiozolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) to a purple formazan product. | Mitotic index, frequency of chromosomal aberrations and numeric changes in human lymphocytes in culture after kaurenoic acid (KA) and their hydrogenated derivative (KAH) treatments. | (Cavalcanti et al. 2010) | ||||||||||||||||
| Substance | Treatment | Mitotic index | Aberrant cells | ||||||||||||||||||
| % | Mean±S.E.M | % | Mean±S.E.M | ||||||||||||||||||
|
KAH |
5.0 µg/mL 10.0 µg/mL 30.0 µg/mL 60.0 µg/mL |
4.2 4.1 3.7 3.5 3.9 3.1 3.2 3.7 3.4 3.0 3.1 3.6 |
4.0±0.2 3.5±0.4 3.43±0.2 3.43±0.3 |
0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.7 0.0 0.0 0.7 0.7 |
0.23±0.4 0.0 0.23±0.4 0.43±0.4 |
||||||||||||||||
3.7. Trypanocidal
Chagas disease (CD) is categorized as a neglected tropical disease, affecting primarily tropical regions that often face insufficient financial support for research and the development of new treatments (Chatelain and Ioset 2011; Chatelain 2015; Armenio et al. 2020; Soerio 2022). The disease came to be known as Chagas disease in honor of Brazilian scientist Carlos Ribeiro Justina das Chagas, who in 1909 first discovered the entry of the monoflagellated protozoan into the bloodstream of human beings through the feces of triatomids. The protozoan was named Trypanosoma cruzi, honoring Oswaldo Cruz (Rester 2008; Song et al. 2009; Soerio 2022).
T. cruzi has a complex biological cycle where its intermediate host can be triatomids, insects, and vertebrate animals such as humans, armadillos, bats, paca, porcupine, monkeys, dogs, cats, etc. These hosts can then assume the role of definitive host, and complete the parasite’s evolutionary cycle (Chatelain 2015; Oliveira de Souza et al. 2017; Santos et al. 2020).
T. cruzi has different names and morphologies that vary according to the stage they are in. The infecting morphology of the disease is trypomastigotes. They are transmitted through the excreta of the triatomids, and when this hematophagous insect bites to feed, it generates a gateway for the disease-causing agent by defecating on the skin after feeding. The acute phase of CD is recognized when trypomastigotes are found in the host’s blood (Sales et al. 2017; Yeung et al. 2021; Santos et al. 2022; Soerio 2022). Amastigotes, characterized by their ability to multiply by binary fission, spherical shape, and absence of flagellum, are the morphological form found in the tissues of infected hosts. They are primarily the heart, colon, and esophagus, and signify the chronic form of CD (Armenio et al. 2020; Santos et al. 2020; Soerio 2022).
CD can also be transmitted through other routes such as blood transfusion or organ transplants, from mother to child during pregnancy or childbirth, contaminated drinking water, laboratory accidents, among others (Armenio et al. 2020; Santos et al. 2020; Soerio 2022). The symptoms of CD vary depending on the phase of the disease. The acute phase is brief and typically presents with few symptoms, while the chronic phase is prolonged and may lead to serious cardiac or digestive complications, potentially resulting in the patient’s death (Chatelain and Ioset 2011; Chatelain 2015; Armenio et al. 2020; Soerio 2022).
One of the main causes of heart disease in endemic areas is CD, and it is estimated that more than six million people are infected with T. cruzi. These infections occur mainly in the poorest regions of 21 Latin American countries, and it is estimated that 1 million women are infected at their reproductive age (Soerio 2022). According to the WHO 2022,(World Health Organization 2021) and the Drug for Neglected Diseases initiative (DNDi), 2022, it is estimated that more than 75 million people are at risk of acquiring CD, and that 173,000 new cases occur annually with more than 75,000 deaths per year, yet less than 10% of infected people are diagnosed and less than 1% are treated. Compared to other parasitic diseases, CD kills more people each year, and even still treatment represents a great challenge (World Health Organization 2021; DNDi 2022; Soerio 2022).
The treatment of CD is based on only two drugs that have been in clinical use for about 50 years, nifurtimox and benznidazole. Both drugs have low efficacy in the treatment of the chronic phase, are contraindicated during pregnancy, and cause serious side effects. This all leads to a high rate of treatment abandonment by patients, in addition to the fact that these two drugs are ineffective in the treatment of nitroderivative-resistant strains of parasites (Pecoul et al. 2016; Vannier-Santos et al. 2019; Bay et al. 2019b; Armenio et al. 2020; Soerio 2022).
These deficiencies justify the search for new therapeutic options, and once again the search for new molecules from natural products (Armenio et al. 2020; Newman and Cragg 2020; Soerio 2022). In this scenario, the Annonaceae family stands out as a promising source of active substances against T. cruzi. Several researchers have been exploring species belonging to this family in search of new compounds with trypanocidal properties.
Some of the most studied species of Annonaceae for trypanocidal activity belong to the Annona, Guatteria, Xylopia, and Duguetia genera. Studies also highlight classes of secondary metabolites such as alkaloids and acetogenins as promising substances for trypanocidal activity. Table 7 describes some species of Annonaceae that have been studied for their trypanocidal activity against different parasitic forms of T. cruzi (epimastigote, trypomastigote, and amastigote).
Annona genus
Eight species from the Annona genus were examined to assess their potential as trypanocidal agents. A. amazonica, A. cornifolia, and A. foetida were subjected to isolation and investigation of their substances for activity against T. cruzi.
Pinheiro et al., 2009, isolated the diterpene Acanthoic acid from A. amazonica and assessed its efficacy against the epimastigote form of T. cruzi, obtaining an IC50 of 59µM. Similarly, Costa et al., 2011, investigated the trypanocidal potential of an alkaloid from A. against the epimastigote and trypomastigote stages of T. cruzi. The alkaloids liriodenine, O-methylmoschatoline, and annomontine exhibited low activity against the epismastigote form (with IC50 values of 645.2, 286.3, and 757.8µM, respectively). However, these alkaloids demonstrated significant activity against the trypomastigote form with IC50 values of 14.3, 11.82, and 16.07µM, respectively, and surpassing the positive control, crystal violet (with an IC50 of 31.37µM) (Pinheiro et al. 2009; Costa et al. 2011a).
Silva et al., 2009, investigated A. cornifolia and isolated acetogenins for evaluation against the amastigote and trypomastigote forms of T. cruzi. Three pure acetogenins—squamocin M, annofolin, and annotacin—along with three acetogenin mixtures—4-deoxylongimicin B + folianin, Glaucanisin + parviflorin, and glaucanisin + glaucanetin—displayed potent activity against the analyzed forms of T. cruzi, with IC50 values ranging from 0.1 to 1.7µM, outperforming the positive control, benznidazole, which had an IC50 of 3.8µM. Despite the promising nature of acetogenins, they also exhibited toxicity, with a selectivity index (SI) of 1, whereas the control demonstrated an SI of 625 (Lima et al. 2014).
Three other species of Annona had their essential oils (EO) investigated for trypanocidal activity. The species A. coriaceae, examined by Siqueira et al., 2011, demonstrated the lowest EO activity against the trypomastigote form, with an IC50 of 168.5 µg/ml, compared to the benznidazole control which showed an IC50 of 45.02 mg/ml. In a study conducted by Meira et al., 2015, the EOs of the A. coriaceae and A. vepretorum species were evaluated against the trypomastigote and epimastigote forms. The EO of A. vepretorum proved to be the most active against the trypomastigote form of T. cruzi, with an IC50 of 11.2 µg/ml, while that of A. coriaceae recorded 12.7 µg/ml. The EO of A. coriaceae stood out as the most effective against the epimastigote form, with an IC50 of 14.9 µg/ml, followed by A. vepretorum, with an IC50 of 16.2 µg/ml. Benznidazole was used as a control, exhibiting an IC50 of 2.7 µg/ml (Siqueira et al. 2011; Meira et al. 2015).
Finally, extracts from two species of Annona were analyzed for activity against the epimastigote and amastigote forms of T. cruzi. The hexanic and ethyl acetate extracts from the leaves of A. muricata were studied by Valencia et al., 2011, with only the ethyl acetate extract being active against the epimastigote form, with an IC50 of 40.2 µg /ml (Valencia et al. 2011).
Osorio et al., 2007, investigated the trypanocidal profile of the hexanic and ethyl acetate extracts from the leaves of A. muricata, as well as the hexanic, ethyl acetate, and methanolic extracts from the branches of this species. The ethyl acetate extract demonstrated the highest potential, with an IC50 of 25 µg/mL for the leaf extract and 63.2 µg/mL for the branch extract (Osorio et al. 2007).
The ethanolic and hexanic extracts from the stem bark, stem wood, root bark, and root wood of A. crassiflora were investigated by Mesquita et al., 2005, against the amastigote form of T. cruzi. All extracts showed activity, with the ethanolic extract of root bark and the ethanolic extract of root wood being the most promising with IC50 values of 5.9 and 9.9 µg/ml, respectively. The results suggest that the roots contain substances with greater potential activity against T. cruzi amastigotes (De Mesquita et al. 2005).
Guatteria genus
The genus Guatteria has also been thoroughly explored for trypanocidal agents. Extracts from two species, G. tonduzii and G. elliptica, were examined for their effects on the epimastigote form of T. cruzi. The ethanolic extract of G. elliptica leaves showed an IC50 of 345.1 µg/ml,(Alves et al. 2012) while the hexanic extracts from the leaves and branches of G. tonduzii were more active, with IC50 values of 34 and 25.2 µg/ml, respectively (Valencia et al. 2011).
The essential oils of three Guatteria species were also studied. Meira et al., 2017, evaluated the trypanocidal potential of essential oils from the leaves of G. friesiana and G. pogonopus against the epimastigote and trypomastigote forms of T. cruzi. Both essential oils showed activity (IC50 values of 11.9 and 28 µg/ml, respectively) (Meira et al. 2017). Another study by Bay et al., 2019a, assessed the essential oil from the aerial parts of G. punctata against the trypomastigote and amastigote forms. The essential oil exhibited remarkable efficacy, with an IC50 of 0.029 µg/mL, in contrast to benznidazole, which had an IC50 of 1 µg/mL as a positive control. The pronounced activity of the G. punctata essential oil is credited to its major constituents, (E)-caryophyllene and germacrene D. Published studies indicate that essential oils containing high concentrations of these compounds possess antiprotozoal properties, particularly against T. cruzi (Da Silva et al. 2013a; Bay et al. 2019a, b).
Mahiou et al., 2000b, conducted a study in which bisbenzylisoquinoline alkaloids were isolated from the stem bark of G. boliviana, and their trypanocidal activity was subsequently evaluated. Eight out of nine isolated alkaloids showed activity against the trypomastigote form of T. cruzi, with funiferine being the most promising alkaloid, with an IC50 of 47.66 µM (Mahiou et al. 2000b).
Xylopia genus
Research on the Xylopia species has explored the trypanocidal properties of both extracts and essential oils. Three different studies examined extracts of X. aromatica against the epimastigote and amastigote forms of T. cruzi. Osorio et al., 2007, analyzed the hexane, ethyl acetate, and methanol extracts from the leaves and branches of X. aromatica against the epimastigote form. The leaf extracts demonstrated superior activity compared to the branch extracts, with the methanolic leaf extract showing the highest efficacy, exhibiting an IC50 of 26.1 µg/mL (Osorio et al. 2007). The ethanolic extract of the fruits, investigated by Alves et al., 2012, showed slight activity against the epimastigote form, with an IC50 of 253.1 µg/ml (Alves et al. 2012). The hexanic extracts of root wood and root bark of X. emarginata were analyzed by Mesquista et al., 2005, against the amastigote form, and both extracts showed activity with IC50 values of 21.6 and 23.5 µg/ml, respectively (De Mesquita et al. 2005).
Silva et al., 2013, investigated the leaf essential oils from X. frutescens and X. laevigata for their activity against the epimastigote and trypomastigote forms. The study revealed that both essential oils were effective against both forms, with a greater potency observed against the trypomastigote form, displaying IC50 values of 11.9 and 12.7 µg/mL, respectively (Da Silva et al. 2013a).
Duguetia genus
Mesquita et al.,2005, conducted a study investigating the trypanocidal potential of hexanic and ethanolic extracts derived from stem, root bark, and root wood of D. furfuracea. The authors found that these extracts exhibit considerable activity against the amastigote form, with the hexanic extract from the root bark displaying the most potency and boasting an IC50 of 6.6 µg/ml (De Mesquita et al. 2005).
In a separate study by Silva et al., 2009, the alkaloid extract and five isolated alkaloids from the bark of the underground stem of Duguetia furfuracea were examined for their biological potential against the trypomastigote form of T. cruzi. The authors also investigated the potential antitumor and leishmanicidal activity of these molecules. The five isolated alkaloids, duguetine, duguetine B-N-oxide, and dicentrinone, in addition to the alkaloid extract were shown to be active against the trypomastigote forms of T. cruzi, with duguetine being the most active (IC50 = 11.82 µM) and stronger than the positive control Gentian violet (IC50 = 31 µM) (da Silva et al. 2009).
Alkaloids isolated from D. lanceolata leaves were examined against both the amastigote and trypomastigote forms of T. cruzi. Dantas et al., 2020, observed that only glaucine showed activity against both forms, demonstrating greater efficacy against the amastigote form with an IC50 of 28.6 µg/ml. The oxoglaucine + liriodenine alkaloid mixture exhibited activity against trypomastigotes, with an IC50 of 83 µg/ml. The other alkaloid mixtures did not demonstrate activity against T. cruzi (Dantas et al. 2020).
Ethanol extracts from the leaves and branches of D. lanceolata were investigated by Alves et al., 2012, against the epimastigote form of T. cruzi. Both extracts showed activity, and these studies suggested promising activity of the substances present in D. lanceolata (Alves et al. 2012).
Bay et al., 2019, carried out a study to evaluate the chemical composition and trypanocidal and antimicrobial activity of four species of Annonaceae: Bocageopsis multiflora (MART.) REFR., Fusaea longifolia (AUBL.) SAFF., Duguetia quitarensis BENTH., and Guatteria punctata (AUBL.) RAHOWARD. The four essential oils showed trypanocidal activity, with IC50s of 0.46, 0.26, 0.3 and 0.029 µg/ml respectively. The synergistic action of the substances present in the essential oil should still be considered (Bay et al. 2019a).
Others Annonaceae species
Two separate studies explored the trypanocidal potential of various Colombian plants, including Desmos panamensis, Pseudomalmea boyacana, Rollinia exsucca, and Rollinia pittieri from the Annonaceae family. Hexanic, ethyl acetate, and methanolic extracts derived from both the leaves and branches were evaluated for their activity against the epimastigote stage of T. cruzi. Among them, the branch extracts of Desmos panamensis and Rollinia pittieri demonstrated the highest activity, with IC50 ranging from 20 to 14 µg/ml (Osorio et al. 2007; Valencia et al. 2011).
Some studies isolated substances and tested them against the trypomastigote form of T. cruzi. Armenio et al., 2020, isolated sesquiterpenes from the stem of Oxandra sessiflora. All sesquiterpenes showed activity against trypomastigotes, with 4β,10α-dihydroxy-guai-6-eno and 4β,6β,7β,10α-tetrahydroxy-guaiane being the most promising. With IC50 values of 16.3 and 17.6 µM, respectively, they performed similarly to the positive control benznidazole (IC50 of 16.4 µM) (Armenio et al. 2020). Ngantchou et al., 2009, assessed the efficacy of polycarpol isolated from Piptostigma preussi, revealing significant activity with an IC50 of 5.114 µM (Ngantchou et al. 2009). Studies by Londero et al., 2018, and Santos et al., 2015, investigated acetylene fatty acid derivatives from flowers and seeds of Porcelia macrocarpa. The authors observed that these compounds have activity against trypomastigotes with IC50 ranging between 27.6 µM and 59.9 µM. Only isanolic acid did not show activity (Santos et al. 2015; Londero et al. 2018).
The hexanic extract from the stem bark of Cardiopetalum calophyllum was analyzed for its potential activity against the amastigote form, which showed activity with an IC50 of 60.4 µg/ml (De Mesquita et al. 2005). Extracts from the fruits, leaves, and root bark of Greenwayodendron suaveolens were studied for potential activity against T. cruzi, and the authors observed that the extracts showed trypanocidal activity. The dichloromethane fraction rich in alkaloids from the root bark was the most active, with an IC50 of 0.25 µg/ml (Muganza et al. 2016). Another study investigated the trypanocidal potential of extracts from the leaves, root bark, and stem of Isolona hexaloba, and it was observed that most of the extracts showed some trypanocidal activity, with the 80% crude ethanol extract from the leaves displaying an IC50 of 8.33 µg/ml (Musuyu Muganza et al. 2015).
These findings collectively support the potential of compounds derived from Annonaceae in combating T. cruzi.
Table 7.
Summary of trypanocidal activity of species of the Annonaceae.
| Annonaceae species | Used material | Extracts/ Compounds | Parasite form | T. cruzi | Ref. | ||
|---|---|---|---|---|---|---|---|
| IC50 µM |
IC50 µg/ml |
SI | |||||
| Annona amazonica | Stem bark | Acanthoic acid Benznidazole (Positive control) |
Epimastigote | 59 7 |
5.9 | (Pinheiro et al. 2009) | |
| Annona cornifolia | Seed | 4-Desoxylongimicin B + Folianin A Squamocin M Annofolin Annotacin Glaucanisin + Parviflorin Glaucanisin + Glaucanetin Benznidazole (Positive Control) |
Amastigote and Trypomastigote | 0.12 0.1 0.11 1.7 0.13 1.7 3.8 |
1 1 1 1 1 1 625 |
(Lima et al. 2014) | |
| Annona coriaceae | Leaves | Essential oil Benznidazole (Positive control) |
Trypomastigotes | 168.5 45.02 |
(Siqueira et al. 2011) | ||
| Annona crassiflora | Stem bark Stem wood Root Bark |
Ethanolic extract Ethanolic extract Hexanic extract Ethanolic extract Hexanic extract |
Amastigote | 14.9 ±2.3 20.5 ±1.1 45.9 ±3.1 5.9 ±1.3 18.6 ±6.8 |
(De Mesquita et al. 2005) | ||
| Annonaceae species | Used material | Extracts/ Compounds | Parasite form | T. cruzi | Ref. | ||
|
IC50 µM |
IC50 µg/ml |
SI | |||||
| Annona crassiflora | Root wood | Ethanolic extract Benznidazole (Positive control) |
Amastigote | 9.9 ±0.5 1.0 ±0.1 |
(De Mesquita et al. 2005) | ||
| Annona foetida | Stem | Liriodenine O-methylmoschatoline Annomontine Benznidazole (Positive control) Crystal violet (Positive control) |
Epimastigote and Trypomastigote | 645.2 / 14.53 286.3 / 11.82 757.8 / 16.07 7.6 / - - / 31.37 |
(Costa et al. 2011a) | ||
| Annona muricataL. |
Leaves Stems |
Hexane extract Ethyl acetate extract Methanol extract Hexane extract Ethyl acetate extract Methanol extract Benznidazole (Positive control) |
Epimastigote | 100.0 25.0 40.2 ±11 >100.0 74.9 63.2 98.6 2.0 |
(Osorio et al. 2007) (Valencia et al. 2011) (Osorio et al. 2007) |
||
| Annona squamosa | Leaves | Essential oil Benznidazole (Positive control) |
Trypomastigote Epimastigote |
12.7 14.9 2.7 |
(Meira et al. 2015) | ||
| Annona vepretorum | Leaves | Essential oil Benznidazole (Positive control) |
Trypomastigote Epimastigote |
11.2 16.2 2.7 |
(Meira et al. 2015) | ||
| Annonaceae species | Used material | Extracts/ Compounds | Parasite form | T. cruzi | Ref. | ||
|
IC50 µM |
IC50 µg/ml |
SI | |||||
| Bocageopsis multiflora(MART.) REFR. | Aerial parts | Essential oil Benznidazole (Positive control) |
Trypomastigote and Amastigote | 0.46 ± 0.07 1 |
2.9 625 |
(Bay et al. 2019a) | |
| Cardiopetalum calophyllum | Stem bark | Hexanic extract Benznidazole (Positive control) |
Amastigote | 60.4 ±0.1 1.0 ±0.1 |
(De Mesquita et al. 2005) | ||
| Desmos panamensis(B.L. Rob) Saff. |
Leaves Stems |
Hexane extract Ethyl acetate extract Methanol extract Hexane extract Ethyl acetate extract Methanol extract Benznidazole (Positive control) |
Epimastigote |
61.4 10.7 ±3.5 85.6 26 ±13.6 98.6 11.9 ±5.2 98.6 17.7 ±6.4 >100.0 14.5 ±2.7 87.5 18.2 ±3.6 2.0 |
1.77 1.02 3.36 1.35 2.4 2.86 |
(Osorio et al. 2007) (Valencia et al. 2011) |
|
| Duguetia furfuracea | Stem Root bark Root wood |
Hexanic extract Ethanolic extract Hexanic extract Ethanolic extract Benznidazole (Positive control) |
Amastigote | 50.0 ±1.6 30.4 ±1.3 6.6 ±0.6 25.6 ±1.5 ±0.1 |
(De Mesquita et al. 2005) | ||
| Annonaceae species | Used material | Extracts/ Compounds | Parasite form | T. cruzi | Ref. | ||
|
IC50 µM |
IC50 µg/ml |
SI | |||||
| Duguetia furfuracea | Stem bark | Duguetine Duguetine β-N-oxide Dicentrinone N-methyltetrahydropalmatine N-methylglaucine Alkaloid extract Gentian violet (Positive control) |
Trypomastigote | 9.32 30.79 18.83 9072 4957 22.44 31 |
(da Silva et al. 2009) | ||
| Duguetia lanceolata | Branches Leaves |
Ethanolic extract Ethanolic extract Glaucine Oxoglaucine + liriodenine Oxoglaucine + lanuginosine + dehydroglaucine Norglaucine + Isocorydine + N-methyllaurotetanine Benznidazole (Positive control) |
Epimastigote Trypomastigote / Amastigote |
250.2 157.9 46.0/28.6 83/not active >100/not active >100/not active 4.6/1.3 11.77 (epimastigote) |
0.21 2.11 |
(Alves et al. 2012) (Dantas et al. 2020) |
|
| Duguetia quitarensisBENTH. | Aerial parts | Essential oil Benznidazole (Positive control) |
Trypomastigote and Amastigote | 0.26 ± 0.06 1 |
2.1 625 |
(Bay et al. 2019a) | |
| Fusaea longifolia(AUBL.) SAFF | Aerial parts | Essential oil Benznidazole (Positive control) |
Trypomastigote and Amastigote | 0.3 ± 0.11 1 |
3.1 625 |
(Bay et al. 2019a) | |
| Annonaceae species | Used material | Extracts/ Compounds | Parasite form | T. cruzi | Ref. | ||
|
IC50 µM |
IC50 µg/ml |
SI | |||||
|
Greenwayodendron suaveolens (Engl. & Diels) Verdc. (sin. Polyalthia suaveolens Enlg. & Diels) |
Fruits Leaves Root bark |
Crude ethanol extract Dichloromethane fraction rich in alkaloids Alkaline aqueous rich in salts e hydrophilic substances Petroleum ether fraction rich in lipids and waxes 90% methanol fraction rich in steroids and terpenes Crude ethanol extract Dichloromethane fraction rich in alkaloids Alkaline aqueous rich in salts e hydrophilic substances Petroleum ether fraction rich in lipids and waxes 90% methanol fraction rich in steroids and terpenes Crude ethanol extract Dichloromethane fraction rich in alkaloids Alkaline aqueous rich in salts e hydrophilic substances Petroleum ether fraction rich in lipids and waxes 90% methanol fraction rich in steroids and terpenes |
Not determined |
34.27 14.79 >64.0 7.49 3.58 27.86 31.17 >64.0 8.33 8.06 7.38 0.25 >64.0 7.88 2.11 |
>1.87 2.34 - 4.07 >17.88 >2.30 >2.05 - 3.84 4.62 1.04 >256.0 - 2.90 13.92 |
(Muganza et al. 2016) |
|
| Annonaceae species | Used material | Extracts/ Compounds | Parasite form | T. cruzi | Ref. | ||
|
IC50 µM |
IC50 µg/ml |
SI | |||||
|
Greenwayodendron suaveolens (Engl. & Diels) Verdc. (sin. Polyalthia suaveolens Enlg. & Diels) |
Stem bark | Crude ethanol extract Dichloromethane fraction rich in alkaloids Alkaline aqueous rich in salts e hydrophilic substances Petroleum ether fraction rich in lipids and waxes 90% methanol fraction rich in steroids and terpenes |
2.0 28.28 >64.0 2.0 2.05 |
17.94 >2.26 - 8.66 3.64 |
(Muganza et al. 2016) | ||
| Guatteria boliviana | Stem bark | Lanuginosine Pangkorimine Funiferine Tiliageine Antioquine Puertogaline A Puertogaline B Sepeerine Guatteboline |
Trypomastigote | >818.8 203.32 47.69 287.66 77.87 242.26 76.13 131.32 100.05 |
(Mahiou et al. 2000b) | ||
| Guatteria elliptica | Leaves | Ethanolic extract Benznidazole (Positive control) |
Epimastigote | 345.1 11.77 |
0.30 | (Alves et al. 2012) | |
| Guatteria friesiana | Leaves | Essential oil Benznidazole (Positive control) |
Epimastigote / Trypomastigote | 11.9 2.7 |
(Meira et al. 2017) | ||
| Guatteria pogonopus | Leaves | Essential oil Benznidazole (Positive control) |
Epimastigote / Trypomastigote | 28.0 2.7 |
(Meira et al. 2017) | ||
| Guatteria punctata(AUBL.) RAHOWARD. | Aerial parts | Essential oil Benznidazole (Positive control) |
Trypomastigote and Amastigote | 0.029 ± 0.014 1 |
32 625 |
(Bay et al. 2019a) | |
| Annonaceae species | Used material | Extracts/ Compounds | Parasite form | T. cruzi | Ref. | ||
|
IC50 µM |
IC50 µg/ml |
SI | |||||
| Guatteria xf. tonduzii | Leaves Stem |
Hexane extract Hexane extract |
Epimastigote | 34.0 ±17 25.2 ±5.2 |
0.77 1.09 |
(Valencia et al. 2011) | |
| Isolona hexaloba |
Leaves Root bark Stem bark |
Aqueous Extract Dried crude extract 80% crude ethanol extract Dichloromethane fraction rich in alkaloids Alkaline aqueous fraction rich in salts and hydrophilic substances Petroleum ether fraction rich in lipids and waxes 90% methanol fraction rich in steroids and terpenes Aqueous Extract Dried crude extract 80% crude ethanol extract Dichloromethane fraction rich in alkaloids Alkaline aqueous fraction rich in salts and hydrophilic substances Petroleum ether fraction rich in lipids and waxes 90% methanol fraction rich in steroids and terpenes Aqueous Extract Dried crude extract 80% crude ethanol extract Dichloromethane fraction rich in alkaloids Alkaline aqueous fraction rich in salts and hydrophilic substances |
Not determined | 30.05 33.71 8.33 32.79 >64.00 8.50 16.54 33.07 21.30 34.56 10.34 >64.00 21.71 20.71 - 15.06 34.27 17.55 >64.00 |
>2.13 >1.90 3.92 >1.95 - 2.52 >3.87 >1.94 >3.00 >1.85 4.11 - 1.59 >3.09 - >4.25 >1.87 >3.65 - |
(Musuyu Muganza et al. 2015) | |
| Annonaceae species | Used material | Extracts/ Compounds | Parasite form | T. cruzi | Ref. | ||
|
IC50 µM |
IC50 µg/ml |
SI | |||||
| Isolona hexaloba | Stem bark |
Petroleum ether fraction rich in lipids and waxes 90% methanol fraction rich in steroids and terpenes Benznidazole (Positive control) Suramine (Positive control) |
Not determined | >64.00 45.02 3.19 - |
- >1.42 |
(Musuyu Muganza et al. 2015) | |
| Oxandra sessiflora | Stem | 4α,10β-aromadendronediol 4β,10α -aromadendronediol 4α,10α-aromadendranediol 1β,6α-dihydroxy-4(15)-eudesmeno 4β,10α-dihydroxy-guai-6-eno 4β,6β,7β,10α-tetrahydroxy-guaiane Benznidazole (Positive control) |
Trypomastigote | 23.7 ±3.8 31.7 ±6.9 29.8 ±8.3 47.5 ±5.8 16.3 ±4.7 17.6 ±2.3 16.4 ±0.8 |
(Armenio et al. 2020) | ||
| Piptostigma preussi | - | Polycarpol | Trypomastigote | 5.114 | (Ngantchou et al. 2009) | ||
| Porcelia macrocarpa |
Flours Seed |
Stearolic acid Santalbic acid 8-hydroxyoctadec-9,11-diynoic Isanolic acid 12,14-octadecadiynoic acid/macrocarpic acid Benznidazole (Positive control) |
Trypomastigote | 27.6 59.9 57.3 Not active 38,77 16.4 534.2 |
>7.2 >3.3 >3.5 - 4.1 >12.2 0.9 |
(Londero et al. 2018) (Santos et al. 2015) |
|
| Annonaceae species | Used material | Extracts/ Compounds | Parasite form | T. cruzi | Ref. | ||
|
IC50 µM |
IC50 µg/ml |
SI | |||||
| Pseudomalmea boyacana(J.F. Macbr.) L. W. Chatrou. |
Leaves Stems |
Hexane extract Ethyl acetate extract Methanol extract Hexane extract Ethyl acetate extract Methanol extract Benznidazole (Positive control) |
Epimastigote |
74.0 89.2 100.0 15.2 ±3.6 50.4 100.0 87.5 12.8 ±1.3 2.0 |
1.19 |
(Osorio et al. 2007) (Valencia et al. 2011) (Osorio et al. 2007) (Valencia et al. 2011) |
|
|
Rollinia exsucca (DC. Ex Dunal) A. DC. |
Leaves Stems |
Hexane extract Ethyl acetate extract Methanol extract Hexane extract Ethyl acetate extract Methanol extract Benznidazole (Positive control) |
Epimastigote |
74.4 98.6 61.4 26.1 18.1 ±7.2 58.3 41.7 ±12 >100.0 2.0 |
(Osorio et al. 2007) (Valencia et al. 2011) (Osorio et al. 2007) (Valencia et al. 2011) (Osorio et al. 2007) |
||
|
Rollinia pittieri Saff |
Leaves Stems |
Hexane extract Ethyl acetate extract Methanol extract Hexane extract Ethyl acetate extract Methanol extract Benznidazole (Positive control) |
Epimastigote |
16.5 46.4 ±3.1 20.8 39.8 16.4 20.8 >100.0 2.0 |
(Osorio et al. 2007) (Valencia et al. 2011) (Osorio et al. 2007) |
||
| Annonaceae species | Used material | Extracts/ Compounds | Parasite form | T. cruzi | Ref. | ||
|
IC50 µM |
IC50 µg/ml |
SI | |||||
| Xylopia aromatica(Lam.) Mart. |
Leaves Stems |
Hexane extract Ethyl acetate extract Methanol extract Hexane extract Ethyl acetate extract Methanol extract Benznidazole (Positive control) |
Epimastigote |
99.2 66.0 26.1 >100.0 58.3 >100.0 2.0 |
(Osorio et al. 2007) |
||
| Xylopia aromatica | Fruits Root wood Root bark |
Ethanolic extract Benznidazole (Positive control) Hexanic extract Hexanic extract Benznidazole (Positive control) |
Epimastigote Amastigote |
253.1 11.77 21.6 ±6.0 23.5 ±4.7 1.0 ±0.1 |
0.39 | (Alves et al. 2012) (De Mesquita et al. 2005) |
|
| Xylopia emarginata | Leaves | Hexanic extract Benznidazole (Positive control) |
Amastigote | 57.6 ±2.4 1.0 ±0.1 |
(De Mesquita et al. 2005) | ||
| Xylopia frutescens | Leaves | Essential oil Benznidazole (Positive control) |
Epimastigote / Trypomastigote | 20.2 / 11.9 2.8 / 2.8 |
(Da Silva et al. 2013a) | ||
| Xylopia laevigata | Leaves | Essential oil Benznidazole (Positive control) |
Epimastigote / Trypomastigote | 22.2 / 12.7 2.8 / 2.8 |
(Da Silva et al. 2013a) | ||
3.8. Antioxidant
Antioxidant compounds have the ability to interfere with oxidation chain reactions that result in the production of toxic compounds, both in the initiation or propagation of these reactions, slowing, neutralizing or preventing actions of oxidizable substrates, and through the inhibition of lipid peroxidation and lipoxygenase (Huber and Rodriguez-Amaya 2008; Silva et al. 2009; Silva 2015).
Antioxidant compounds exert their effects through various mechanisms, including free radical inhibition and metal ion complexation (Pietta 2000; Duarte-Almeida et al. 2006). Free radicals can be acquired exogenously and are also produced endogenously. When in excess the radicals can cause oxidative stress which culminate in the appearance of cardiovascular and neurological diseases, some cancers, cataracts, diabetes, rheumatism, aging of the body, etc (Núñez-Sellés 2005; Prado 2009; Sousa et al. 2011; Silva et al. 2018). Thus, antioxidants bring great benefits to human health by protecting the body from damage caused by free radicals (Oliveira 2015).
Antioxidants can be synthetic or natural. The synthetic ones have greater stability and good efficiency when compared to the natural ones, but the natural ones have less toxicity. In addition to having a greater effect on the oxidation processes, natural antioxidants are generally more sought after than the synthetic ones (Duarte-Almeida et al. 2006; Prado 2009; Silva 2015). For the same reasons, researchers have analyzed the antioxidant potential of Annonaceae species.
Xylopia genus
Several studies have explored the antioxidant properties of Xylopia species, yielding encouraging results. In one such study, Karioti et al., 2004, analyzed the essential oils from different parts of X. aethiopica to evaluate their antioxidant activity and determine their main constituents. The findings revealed that the essential oil derived from the leaves displayed the strongest ability to neutralize free radicals (Karioti et al. 2004).
Similarly, Konan et al., 2009, examined the antioxidant activity and chemical composition of essential oils extracted from dried fruits of X. aethiopica leaves. Surprisingly, they discovered that the nut oil displayed slightly higher antioxidant activity compared to the essential oil extracted from the leaves (Konan et al. 2009). This contradicted the findings of Karioti et al., 2004, highlighting the variability in antioxidant activity among different parts of the plant.
In another study by Silva et al., 2009, the antioxidant potential of X. langsdorffiana was investigated. The authors isolated seven compounds from the leaves and stem bark of the plant, including alkaloids, diterpenes, phaephorbide, and flavonoids. Among these compounds, discretamine, an alkaloid, exhibited exceptional antioxidant activity, with a percentage of activity greater than 90% in the 1,2-diphenyl-2-picryl-hydrazyl (DPPH) free radical scavenging assay (Da Silva et al. 2009).
These studies collectively demonstrate the significant antioxidant potential of Xylopia species, highlighting the importance of further research into their therapeutic applications in combating oxidative stress-related disorders.
Annona genus
Silva et al., 2019, explored the antioxidant potential of A. nutans, a species within the Annonaceae family. They conducted comprehensive analyses, including the evaluation of total phenolic content, flavonoids, and tannins, along with quantification of main metabolites using LC-MS. The methanolic fraction of the extract exhibited significant anti-inflammatory effects, reducing paw edema, and inhibiting acute inflammation even six hours after carrageenan injection. Intraperitoneal treatment with the methanolic extract also showed efficacy in reducing inflammation and elicited central antinociceptive effects. Furthermore, the hydromethanolic fraction of A. nutans extract demonstrated dose-dependent antioxidant activity comparable to commercial antioxidant BHT. Metabolite analysis identified several compounds contributing to its antioxidant properties, including quercetin derivatives and chlorogenic acid (Silva et al. 2019).
In another study conducted by Leite et al., 2021, the antioxidant potential of extracts from the pulp and seeds of A. squamosa L. was evaluated. Chemical profiling revealed the presence of anthocyanidins, flavones, flavonols, and alkaloids. Phenol analysis indicated higher levels in the seed extract, while vitamin C content was higher in the pulp extract. Antioxidant activity was assessed using various methods, with the ABTS assay showing the most potent activity for both seed and pulp extracts, with IC50 values of 0.14 ± 0.02 and 0.38 ± 0.02 µg/mL, respectively. Additionally, the pulp extract exhibited superior acetylcholinesterase inhibitory activity compared to the seed extract, suggesting potential cognitive benefits. A. squamosa emerges as a promising source of antioxidants, further emphasizing its value as a nutritious food source (Leite et al. 2021).
Together, these studies underscore the significant antioxidant potential of Annona species, highlighting their possible therapeutic applications in combating oxidative stress-related conditions and promoting overall health and well-being.
Others Annonaceae species
Several studies have investigated the antioxidant activity of various species within the Annonaceae family, shedding light on their potential therapeutic applications. Jain et al., 2014, investigated the chemical profile of the butanolic fraction derived from the hydroalcoholic extract of P. longifolia stem bark. They discovered that 3-O-methyl ellagic acid exhibited antioxidant activity with an IC50 of 24.28 μg/mL, while the butanolic fraction had an IC50 of 266.59 μg/mL in the DPPH assay (Jain et al. 2014).
Sacchetti et al., 2004, evaluated eleven essential oils, including Cananga odorata, to assess their functional ingredients and potential as synthetic preservatives. Among these oils, Cananga odorata oil showed remarkable antioxidant activity, with an activity percentage greater than 75% in the DPPH test (Sacchetti et al. 2005).
Xavier et al., 2016, characterized the chemical composition of the essential oil from the leaves of Cardiopetalum calophyllum and evaluated its antioxidant potential. The study demonstrated significant antioxidant activity, with an IC50 of 9.66 μg/ml in the DPPH method, validating the antioxidant potential of C. calophyllum (Xavier et al. 2016).
Santos et al., 2018, analyzed the methanolic extract of Duguetia furfuracea and its fractions for antioxidant and anti-inflammatory potential. The methanolic extract exhibited significant antioxidant activity with an IC50 of 22.46 µg/mL in the DPPH assay. Additionally, it showed inhibitory effects on carrageenan-induced edema and leukocyte migration, suggesting its potential in treating inflammatory conditions (do Santos et al. 2018).
A recent study by Nghi et al., 2022, highlighted the antioxidant activity of Rumdul fruit (Sphaerocoryne affinis) extract. The aqueous extract showed strong antioxidant capacity with an IC50 value of 85.62 ± 1.05 μg/mL in the DPPH assay. The extract also exhibited potential in improving locomotor deficiencies and preventing degeneration of dopaminergic neurons, indicating its promising role in treating Parkinson’s Disease (Nghi et al. 2022).
Overall, these studies underscore the significant antioxidant potential of Annonaceae species, suggesting their value as sources of natural antioxidants with potential therapeutic benefits. Table 8 provides a summary of the results discussed in this section.
Table 8.
Antioxidant activity of substances and extracts from species of the Annonaceae.
| Annonaceae species | Used material | Substances/ Extracts |
IC50 | DPPH | Ref. | |||
|---|---|---|---|---|---|---|---|---|
| %Interaction | %Interaction 20 min |
%Interaction 60 min |
%Free radical scavenging | |||||
|
Annona nutans |
Leaves |
Hydromthanolic fraction |
4.89 µg/mL |
- | - | - |
50.0 |
(Silva et al. 2019) |
|
Annona squamosa |
Seed Pulp |
Extract Extract Vitamin C (positive control) |
0.36 µg/mL 0.83 µg/mL 0.011 µg/mL |
- | - | - | - | (Leite et al. 2021) |
| Cananga Odorata | Essential oil | Essential oil | 75.50 | - | - | 63.80 | (Sacchetti et al. 2005) | |
| Cardiopetalum calophyllum | Leaf |
Essential oil Quercetin (standard compound) |
9.66 µg/mL 3.13 µg/mL |
- - |
- - |
- - |
- - |
(Xavier et al. 2016) |
| Duguetia furfuracea | Leaves | Methanolic extract Chloroform fraction Ethyl acetate fraction Hydromethanol fraction |
22.46 µg/mL 176.88 µg/mL 60.56 µg/mL 28.21 µg/mL |
50.0 50.0 50.0 50.0 |
- | - | - | (do Santos et al. 2018) |
| Polyalthia longifolia | Stem bark | 3-O-methyl Ellagic Acid | 2.5 µg/mL 5 µg/mL 10 µg/mL 20 µg/mL 40 µg/mL 24.28 μg/ml |
- | - | - | 10.36 28.17 34.83 45.80 69.81 50.00 |
(Jain et al. 2014) |
| Annonaceae species | Used material |
Substances/ Extracts |
IC50 | DPPH | Ref. | |||
| %Interaction |
%Interaction 20 min |
%Interaction 60 min |
%Free radical scavenging | |||||
| Sphaerocoryne affinis | Fruit |
Rumdul fruit water extract |
85.62 µg/mL | - | - | - | (Nghi et al. 2022) | |
| Xylopia aethiopica | Essential oil of leaf, stem bark, roots, and fruits | Leaf oil Leaf oil Root bark oil Stem bark oil Fresh fruit oil Dried fruit oil Dried fruit oil |
0.048 g/mL 4.9 mg/mL 0.033 g/mL 0.037 g/mL 0.045 g/mL 0.042 g/mL 4.1 mg/mL |
- 50.0 - - - - 50.0 |
43.8 - 36.5 32.4 85.6 21.2 - |
75.9 - 43.2 40.3 85.8 54.8 - |
86.8 - 68.4 73.9 66.5 57.2 - |
(Karioti et al. 2004) (Konan et al. 2009) (Karioti et al. 2004) (Karioti et al. 2004) (Karioti et al. 2004) (Karioti et al. 2004) (Konan et al. 2009) |
|
Xylopia langsdorffiana |
Leaf and stem bark | Discretamine | Concentrations Tested 240 µg/mL 120 µg/mL 60 µg/mL 30 µg/mL |
94.25 93.93 91.51 90.05 |
- | - | - | (Da Silva et al. 2009) |
3.9. Antimalarials
Malaria is an endemic disease in tropical and subtropical areas, occurring mainly in Southeast Asia, the Amazon and Africa (Nascimento et al. 2019; Hammami et al. 2020). Despite numerous efforts to control and eliminate the disease, it is estimated that there were 228 million cases in 2018, with a global mortality rate of up to 30% in severe cases in 2019. Therefore, malaria is considered a public health problem (Talapko et al. 2019; World Health Organization (WHO) 2019; Figueroa-Miranda et al. 2020; Hammami et al. 2020).
The etiological agent of malaria is a Plasmodium protozoan that infects humans (White et al. 2014; Walker et al. 2017; Talapko et al. 2019). This protozoan is an intracellular parasite that can infect red blood cells and some tissues, depending on the species. There are five species that can infect humans, P. falciparum, P. vivax, P. ovale, P. malarie e P. knowlesi (Vuk et al. 2008; Talapko et al. 2019; World Health Organization (WHO) 2019; Hammami et al. 2020).
The vector, mosquitoes, and parasite have developed resistance to some drugs used for their control and eradication. On top of administration problems, the resistance has made it increasingly difficult to eradicate this disease globally (Asase et al. 2010; Talapko et al. 2019). Thus, research for new drugs and insecticides is still important, as well as studies with the vaccine.
Plants are always an excellent source of new drugs, so the Annonaceae, due to its chemical diversity, can be interesting in the study of new antimalarial drugs. In this sense, several researchers have already analyzed the antimalarial potential of several species of Annonaceae, and the results are described below.
Asase et al., 2010, studied 30 species of plants, belonging to 28 genera and 20 families, used to treat malaria by indigenous communities in the western district of Dangme, Ghana. Five species were described for the first time in the treatment of malaria, including the Annonaceae species Greenwayodendron sp. The authors reported that the leaves were boiled, and one cup of this tea was administered three times a day for adults (half a cup for children) until complete recovery (Asase et al. 2010).
Polyathia genus
The dichloromethane extract and two dimeric aporphinoid alkaloids, named bidebiline C and D, obtained from roots of P. debilis showed activity against P. falciparum (Somdej Kanokmedhakul et al. 2003). The indolosesquiterpene alkaloids, N-acetyl-8α-polyveolinone and N-acetyl-polyveoline, isolated from the stem bark of P. oliveri, exhibited moderate antiplasmodial effects against the blood stages of the chloroquine-sensitive P. falciparum NF54 strain. (Gbedema et al. 2015).
Fractionation of the ethanolic extract from the stem bark of P. longifolia led to the isolation of several compounds through bioassay-guided techniques. This process led to the identification of three clerodane diterpenes (16-hydroxycleroda-3,13-dien-16,15-olide, 16-oxocleroda-3,13E-dien-15-oic acid, and 3,16-dihydroxycleroda-4(18),13(14)Z-dien-15,16-olide), a steroid (β-stigmasterol), and two alkaloids (darienine and stepholidine) with significant antiplasmodial activity. The clerodane diterpenes, in particular, exhibited strong activity against multi-resistant P. falciparum K1 strains, with IC50 values between 3 and 6 μg/mL. (Gbedema et al. 2015).
Others Annonaceae species
Methanolic extract of the leaves of Annikia kummeriae showed strong activity against the multi-resistant strain of P. falciparum (K1), with an IC50 value of 0.12 μg/mL. Through a biomonitored study four pure alkaloids, lysicamine, trivalvone, palmatine and jatorrizine, were isolated and identified for having strong to moderate activity against P. falciparum (IC50 ranging from 0.08-2.4 μg/mL) (Malebo et al. 2013a).
Essential oils from the stem bark and leaves of Cleistopholis patens and Uvariastrum pierreanum, were found to be moderately active against P. falciparum. These results indicate that essential oils may offer a promising alternative for the development of new antimalarials (Boyom et al. 2011).
Four essential oils extracted from steam bark ofXylopia phloiodora, Xylopia aethiopica, Pachypodanthium confine and Hexalobus crispiflorus were evaluated regarding their anti-plasmodial activity against the W2 strain of P. falciparum, all oils were active, the most effective was the oil of H. crispiflorus, with an IC50 of 2 μg/mL (Boyom et al. 2011).
The phytochemical analysis of methanol extracts from the leaves and twigs of Mitrephora tomentosa led to the isolation of six polyacetylenic ester-neolignan derivatives, designated mitrephentosins A-F. Among these, mitrephentosins C, E, and F exhibited moderate antimalarial activity, with IC50 values ranging from 13.3 to 24.6 μM, against the P. falciparum strains TM4/8.2 and K1CB1 (Boyom et al. 2011).
Significant antiplasmodial activity was noted in the ethyl acetate extracts from the stems of Rollinia pittieri and Pseudomalmea boyacana. From R. pittieri, researchers isolated one oxoaporphinic alkaloid (O-methylmoschatolin) and one 7,7-dimethylaporphinic alkaloid (melosmin). Meanwhile, P. boyacana yielded three oxoaporphinic alkaloids (aislamient, liriodenine, and atherospermidin). All isolated alkaloids demonstrated activity, with liriodenine showing the strongest effect against both chloroquine-sensitive and chloroquine-resistant strains of P. falciparum (IC50 = 8.0-10.0 μg/mL)(Boyom et al. 2011).
The results presented so far reinforce the antiplasmodial activity of substances and extracts of the Annonaceae species, and a table of those tested are elaborated in Table 9.
Table 9.
Antiplasmodial activity of substances and extracts from species of the Annonaceae.
| Annonaceae species | Used material | Substances/Extracts |
P. falciparum (strain) (IC50) |
Ifβ-h (%) 2mg/ml | Ref. | |||
|---|---|---|---|---|---|---|---|---|
| F32 (µg/ml) | W2 (µg/ml) | K1 (µg/ml) |
NF54 (µM) |
|||||
|
Annickia kummeriae |
Leaves |
Extract metanolic Lysicamine Trivalvone Palmatine Jatrorrhizine |
- |
- |
0.12 2.4 1.6 0.08 0.24 |
- |
- |
(Malebo et al. 2013a) |
| Cleistopholis patens | Steam bark Leaves | Essential oil | 9.19 15.19 |
(Boyom et al. 2011) | ||||
| Hexalobus crispiflorus | Stem bark | Essential oil | 2.0 | (Fekam Boyom et al. 2003) | ||||
| Mitrephora tomentosa | Leaves and twigs | (-)-(7R,8S)-mitrephentosin C (-)-(7R,8S)-mitrephentosin F Cyclogunil (Positive control) |
13.3 µM 18.7 µM 5.87 µM |
(Wongsomboon et al. 2021) | ||||
| Pachypodanthium confine | Steam bark | Essential oil | 16.6 | (Fekam Boyom et al. 2003) | ||||
| Polyalthia debilis | Roots | dichloromethane extract Bidebiline C Bidebiline D |
1.35 5.4 4.1 |
(Kanokmedhakul et al. 2003) | ||||
| Polyalthia longifólia | Steam bark | 16-hydroxycleroda-3,13-dien-16,15-olide Acid 16-oxocleroda-3,13E-dien-15-óico |
5.33 3.05 |
(Gbedema et al. 2015) | ||||
| Annonaceae species | Used material | Substances/Extracts |
P. falciparum (strain) (IC50) |
Ifβ-h (%) 2mg/ml | Ref. | |||
| F32 (µg/ml) | W2 (µg/ml) |
K1 (µg/ml) |
NF54 (µM) |
|||||
| Polyalthia longifolia | Steam bark | 3,16-dihydroxycleroda-4 (18),13 (14) Z-dien-15,16-olida β-Stigmasterol Darienine L-Stepholidine |
6.15 63.3 22.05 104.33 |
(Gbedema et al. 2015) | ||||
|
Polyalthia oliveri |
Steam bark |
N-acetil-8α-polivolinona N-acetil-polveolina Chloroquine (Positive Control) |
7.6 29.1 0.006 |
(Kouam et al. 2014) | ||||
| Pseudomalmea boyacana | Liriodenina Atherospermidina Isomoschatolina Chloroquine (Positive control) |
8 – 10 0.01 |
8.0 0.90 |
11.4 5.12 2.40 97.0 |
(Osorio D et al. 2006) | |||
| Rollinia pittieri | O-metilmoschatolina Melosmina Chloroquine (Positive control |
98.3 96.4 97.0 |
(Osorio D et al. 2006) | |||||
| Uvariastrum pierreanum | Steam bark Leaves | Essential oil | 6.08 13.96 |
(Boyom et al. 2011) | ||||
|
Xylopia phloiodara Xylopia aethiopica |
Steam bark | Essential oil | 17.9 17.8 |
(Fekam Boyom et al. 2003) | ||||
K1 - multidrug resistant strain, F32 - chloroquine sensitive strain, W2 - chloroquine resistant strain, Ifβ-h - inhibition of β-hematin formation.
3.10. Gastroprotective
Gastric ulcer (GU) is a very common disease of the digestive system that has a high morbidity and thus is a major public health concern (2016 2017; Zhou et al. 2020). The cause of a GU can be related to several factors, such as for example, the indiscriminate use of non-steroidal anti-inflammatory drugs. This disrupts the protective mechanisms of the gastric mucosa, particularly those associated with prostaglandin levels and antioxidant activity. As a result, the defensive barrier of the gastric mucosa is compromised, leading to the development of gastric ulcers (Franke et al. 2005; Woolf and Rose 2020; Zhou et al. 2020). Oxidative stress induced by ethanol, which generates highly cytotoxic free radicals, has also been linked to GU developement (X et al. 2018; Aziz et al. 2019; Simões et al. 2019; Zhou et al. 2020). Other factors that cause GU are stress and infection by Helicobacter pylori bacteria (Tulassay and Herszényi 2010; Montenegro et al. 2014).
Although there are many medications on the market for the treatment of GU, such as antacids, proton pump inhibitors, anticholinergics, cytoprotective agents, etc., the majority of which induce side effects such as hypersensitivity, arrhythmia , impotence, gynecomastia, hematopoietic changes, and hyper gastrinemia, with chronic use (Sheen and Triadafilopoulos 2011; Montenegro et al. 2014; 2016 2017).
Thus, it is necessary to search for new drugs that are less toxic and more effective in the treatment of GU. In this context, plants are an excellent source of new drugs and the Annonaceae stands out for having numerous pharmacological activities, as previously mentioned. The Annonaceae genus which were evaluated include Annona, Goniothalamus, Polyalthia and Xylopia (Table 10).
Annona genus
In a study conducted by Moghadamtousi et al. (2014) the gastroprotective effects of the ethyl acetate extract from Annona muricata leaves (EEAM) were analyzed using the ethanol-induced gastric lesion model in rats. The results indicated a significant reduction in the ulcer injury rate among rats pretreated with oral administration of EEAM, which was comparable to the effect observed with omeprazole in the control group. The authors further concluded that the potential and promising gastroprotective effect of EEAM could be attributed to its suppressive impact on oxidative damage and its conservative effects on gastric wall mucus (Moghadamtousi et al. 2014).
Goniothalamus sp.
The gastroprotective activity of the racemic mixture of goniothalamine (GNT), a lactone naturally occurring in enantiomeric form in plants of the genus Goniothalamus (Annonaceae), was evaluated using an ethanol-induced gastric lesion model. The study demonstrated a strong gastroprotective effect of GNT at concentrations of 30 and 60 mg/kg, with an ED50 of 18 mg/kg. This activity was found to rely on the generation of sulfhydryl and prostaglandin compounds but was independent of nitric oxide (NO), gastric secretion, and mucus production. The authors proposed that GNT functions as a mild irritant, promoting the production of sulfhydryl and prostaglandin compounds, a process known as adaptive cytoprotection. This suggested mechanism was reinforced by the observation that Michael acceptors, including GNT, are effective inducers of antioxidant responses through mild oxidative stress. The gastroprotective effect of GNT was diminished when pretreatment with N-ethylmaleimide and a NSAID was used, underscoring the critical role of sulfhydryl and prostaglandin compounds in GNT’s activity (Vendramini-Costa et al. 2014).
Polyalthia genus
In this sense, some authors have carried out research on some species of the Polyalthia genus looking for new gastroprotective agents. Olate et al. (2012), evaluated the gastroprotective potential of 11 amides derived from the diterpene (4S, 9R, 10R) methyl 18-carboxy-labda-8,13 (E) -dien-15-oate (PMD) and its 8 (9) -en isomer [ PMD 8 (9) -en], isolated in the species Polyalthia macropoda. The gastric lesion model in mice was induced using ethanol/HCl. When administered as a single oral dose of 0.1 mg/kg, compounds 1, 10, and 11 exhibited significant gastroprotective effects, comparable to the reference lansoprazole at 1 mg/kg, with reductions in gastric lesions of 76.7%, 67.7%, and 77.2%, respectively. Compounds 10 and 11 are particularly promising as they demonstrated strong gastroprotective effects without any cytotoxicity (Olate et al. 2012).
Xylopia genus
Montenegro et al. (2014) investigated the gastroprotective properties of X. langsdorffiana A. St.-Hil. & Tul. by analyzing the ethanolic extract (EtOHE) and hexane phase (HexPh) derived from its leaves. The study aimed to assess the gastroprotective efficacy of these extracts and determine their underlying mechanisms of action. They found that both the XL-EtOHE extract and the XL-HexPh phase significantly reduced ulcer formation compared to the negative controls, with the most effective doses being 500 mg/kg for the EtOHE and 250 mg/kg for the HexPh. The authors concluded that the gastroprotective activity is likely due to the involvement of sulfhydryl and nitric oxide (NO) groups, rather than anti-secretory effects or increased mucus production. Additionally, phytochemical analysis of the extracts revealed four major diterpenes, which may contribute to the observed gastroprotective effects of X. langsdorffiana. (Montenegro et al. 2014).
Table 10.
Summary of the gastroprotective activity of Annonaceae species.
| Annonaceae species | Used material | Substances/Extracts | Methodology | Dose | Ulcer lesion index | %Lesion reduction (gastroprotection) |
Ref. |
|---|---|---|---|---|---|---|---|
|
Annona muricata |
Leaves |
EEAM Control Omeprazole |
Ethanol induced ulcer model |
200 mg/kg 400 mg/kg 20 mg/kg |
39.0 23.0 9.0 |
- |
(Moghadamtousi et al. 2014) |
|
Goniothalamus sp. |
- |
Rac-GNT Control Carbenoxolone 200 |
Ethanol induced ulcer model |
15 mg/kg 30 mg/kg 60 mg/kg 18 mg/kg (ED50) 200mg/kg |
- |
41.7 70.5 86.7 50.0 93.5 |
(Vendramini-Costa et al. 2014) |
|
Polyalthia macropoda |
Stem bark |
PMD Amides from PMD 1 10 11 Control Lansoprazole |
Ethanol induced ulcer model |
1 mg/kg 5 mg/kg 10 mg/kg 0.1 mg/kg 0.1 mg/kg 0.1 mg/kg 1 mg/kg |
20.4 17.7 9.4 10.4 20.8 14.7 13.8 |
68.3 72.5 85.4 76.7 67.7 77.2 78.6 |
(Olate et al. 2012) |
|
Xylopia langsdorffiana |
Leaves |
EtOHE HexPh Control Carbenoxolone |
Ethanol induced ulcer model |
500 mg/kg 250 mg/kg 500 mg/kg 100 mg/kg |
- |
83 81 84 40 |
(Montenegro et al. 2014) |
4. Perspectives
Studies related to natural products are extremely important because they are sources of secondary metabolites that have different biological activities and applications for humans. In this review, the exciting biological potential of the Annonaceae family was highlighted and is extremely promising. These plants continue to be identified as potential therapeutics in many disease states but herein we discussed the most promising to date, the anti-inflammatory, insecticidal, antimicrobial, leishmanicidal, cytotoxic, antitumor, trypanocidal, antioxidant, gastroprotective, and antimalarial activities. Unfortunately, most studies focus on the activities of different plant extracts and essential oils, thus be limited by a lack of isolated molecules and mechanisms of action. The identification of biological activity in isolated compounds is of fundamental importance for the identification of new drugs and pharmaceutical and agricultural production. Where isolated in vitro data is available, we encourage researchers to delve deeper into in vivo and clinical experiments, for the production and commercialization of new drugs. This review also compiles vital information for the pharmaceutical and agricultural industry.
Funding
This research was funded by National Council for Scientific and Technological Development (CNPq) and Coordination of Superior Level Sta Improvement (CAPES).
Acknowledgments
We would like to thank the CNPq and Capes for financial Support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 2016 G (2017) GBD 2016 Disease Injury and Incidence and Prevalence Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990–2016: a systemic analysis for the Global Burden. Lancet 390:1211–1259.
- Abarikwu SO, Ogunlaja A, Otuechere CA, Gideon OB (2017) Effect of Ethanolic Extract from Seeds or Pods of Xylopia Aethiopica (Dunal) A. Rich (Annonaceae) on the Testicular Function of Adult Male Rats. Indian J Clin Biochem 32:420–428. [CrossRef]
- Acar JF, Moulin G (2012) Antimicrobial resistance: a complex issue Pre-existence of antimicrobial resistance determinants Origin of antimicrobial resistance determinants. Rev Sci Tech (Internationail Off Epizoot 31:23–31.
- Aciole SDG, Piccoli CF, COSTA E V, et al (2011) Insecticidal activity of three species of Guatteria (Annonaceae) against Aedes aegypti (Diptera: Culicidae). Rev Colomb Entomol 37:262–268.
- Aguirre-Ghiso JA (2007) Models, mechanisms and clinical evidence for cancer dormancy. Nat Rev Cancer 7:834–846.
- Ahalya B, Shankar Ravi K, Kiranmayi GVN (2014) Exploration of anti-hyperglycemic and hypolipidemic activities of ethanolic extract of Annona muricata bark in alloxan induced diabetic rats. Int J Pharm Sci Res 25:21–27.
- Aina SA, Banjo AD, Lawal OA, Jonathan K (2009) Efficacy of some plant extracts on Anopheles gambiae mosquito larvae. Acad J Entomol 2:31–35.
- Akah PA, Odoh UE, Ezike AC, et al (2012) Antimicrobial Effects of a Lipophilic Fraction and Kaurenoic Acid Isolated from the Root Bark Extracts of Annona senegalensis . Evidence-Based Complement. Altern. Med. 2012:1–10.
- Akendengue B, Roblot F, Loiseau PM, et al (2002) Klaivanolide, an antiprotozoal lactone from Uvaria klaineana. Phytochemistry 59:885–888. [CrossRef]
- Akinwumi FO, Fasakin EA, Adedire CO (2007) Toxic and repellence activities of four plant extracts to Dermestes maculatus Degeer on smoked african mud catfish, Clarias gariepinus Burchell. J Entomol 4:149–154.
- Aku AA, Ogunwolu EO, Attah JA (1998) Annona senegalensis L.(Annonaceae): Performance as a botanical insecticide for controlling cowpea seed bruchid, Callosobruchus maculatus (F.)(Coleoptera: Bruchidae) in Nigeria/Annona senegalensis L.(Annonaceae): Wirksamkeit als pflanzliches Insektizid zur. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz/Journal Plant Dis Prot 513–519.
- Alali FQ, Kaakeh W, Bennett GW, McLaughlin JL (1998) Annonaceous Acetogenins as Natural Pesticides: Potent Toxicity Against Insecticide-Susceptible and -Resistant German Cockroaches (Dictyoptera: Blattellidae). J Econ Entomol 91:641–649. [CrossRef]
- Alberts B, Johnson A, Lewis J, et al (2010) Biologia molecular da célula. Artmed Editora.
- Alcântara JM, De Lucena JMVM, Facanali R, et al (2017) Chemical composition and bactericidal activity of the essential oils of four species of annonaceae growing in brazilian amazon. Nat Prod Commun 12:619–622. [CrossRef]
- Aller MA, Arias JL, Arias JI, et al (2007) The inflammatory response recapitulates phylogeny through trophic mechanisms to the injured tissue. Med Hypotheses 68:202–209. [CrossRef]
- Allison LN, Dike KS, Opara FN, et al (2013) Evaluation of larvicidal efficacy and phytochemical potential of some selected indigenous plant against Anopheles gambiense and Culex quinquefasciatus. Adv Biosci Biotechnol 2013:.
- Almeida JRG da S, Araújo C de S, Pessoa C do Ó, et al (2014) Antioxidant, cytotoxic and antimicrobial activity of Annona vepretorum MART. (Annonaceae). Rev Bras Frutic 36:258–264.
- Alolga RN, Chávez León MASC, Osei-Adjei G, Onoja V (2019) GC-MS-based metabolomics, antibacterial and anti-inflammatory investigations to characterize the quality of essential oil obtained from dried Xylopia aethiopica fruits from Ghana and Nigeria. J Pharm Pharmacol 71:1544–1552. [CrossRef]
- Alsenosy AA, El-Far AH, Sadek KM, et al (2019) Graviola (Annona muricata) attenuates behavioural alterations and testicular oxidative stress induced by streptozotocin in diabetic rats. PLoS One 14:e0222410. [CrossRef]
- Alvarez Colom O, Neske A, Popich S, Bardón A (2007) Toxic effects of annonaceous acetogenins from Annona cherimolia (Magnoliales: Annonaceae) on Spodoptera frugiperda (Lepidoptera: Noctuidae). J Pest Sci (2004) 80:63–67. [CrossRef]
- Alves RT, Regasini LO, Funari CS, et al (2012) Trypanocidal activity of Brazilian plants against epimastigote forms from Y and Bolivia strains of Trypanosoma cruzi. Rev Bras Farmacogn 22:528–533. [CrossRef]
- Aminimoghadamfarouj N, Nematollahi A, Wiart C, et al (2020) Secondary metabolismin in Annonaceae: potencial source of drugs. Rev Bras Frutic 36:120554. [CrossRef]
- Aminimoghadamfarouj N, Nematollahi A, Wiart C (2011) Annonaceae: Bio-resource for tomorrow’s drug discovery. J Asian Nat Prod Res 13:465–476. [CrossRef]
- Anadozie SO, Adewale OB, Meyer M, et al (2021) In vitro anti-oxidant and cytotoxic activities of gold nanoparticles synthesized from an aqueous extract of the Xylopia aethiopica fruit. Nanotechnology 32:. [CrossRef]
- Andrade AL (2018) Atividade Antibacteriana e Antibiofilme de Cmplexos de Rutênio do Tipo cis-[RuCl2(dppb)(NN-R)] 2+ Sobre Bactérias Resistentes e o Efeito destes Combinados aos Antibióticos Ampicilina e Tetraciclina. Universidade Federal do Ceará.
- Anza M, Endale M, Cardona L, et al (2021) Chemical composition, cytotoxicity and larvicidal activity of essential oils of three medicinal plants of Ethiopian flora against anisakis L3 larvae. Res J Pharmacogn 8:21–30. [CrossRef]
- Araújo C de S, Silva JC, Mendes RL, et al (2017) Phytochemical screening, cytotoxicity and acute toxicity of Annona vepretorum Mart (Annonaceae) leaf extracts. Trop J Pharm Res 16:597. [CrossRef]
- Araújo AMN de (2010) Bioatividade de espécies vegetais em relação a Zabrotes subfasciatus (Boheman, 1833)(Coleoptera: Chrysomelidae: Bruchinae) em feijão (Phaseolus vulharis L. 1753).
- Armenio AA, de Sousa EA, Alves Veras MD, et al (2020) In vitro anti-Trypanosoma cruzi evaluation of sesquiterpenes from the branches of Oxandra sessiliflora. Phytochem Lett 37:59–62. [CrossRef]
- Asase A, Akwetey GA, Achel DG (2010) Ethnopharmacological use of herbal remedies for the treatment of malaria in the Dangme West District of Ghana. J Ethnopharmacol 129:367–376. [CrossRef]
- Attiq A, Jalil J, Husain K (2017) Annonaceae: Breaking the wall of inflammation. Front Pharmacol 8:. [CrossRef]
- Attiq A, Jalil J, Husain K, et al (2021) A new prenylated benzoquinone from Cyathocalyx pruniferus abrogates LPS-induced inflammatory responses associated with PGE2, COX-2 and cytokines biosynthesis in human plasma. Inflammopharmacology 29:841–854. [CrossRef]
- Auddy B, Ferreira M, Blasina F, et al (2003) Screening of antioxidant activity of three Indian medicinal plants, traditionally used for the management of neurodegenerative diseases. J Ethnopharmacol 84:131–138. [CrossRef]
- Aziz RS, Siddiqua A, Shahzad M, et al (2019) Oxyresveratrol ameliorates ethanol-induced gastric ulcer via downregulation of IL-6, TNF-α NF-ĸB, and COX-2 levels, and upregulation of TFF-2 levels. Biomed Pharmacother 110:554–560. [CrossRef]
- Bakarnga-Via I, Hzounda JB, Fokou PVT, et al (2014) Composition and cytotoxic activity of essential oils from Xylopia aethiopica (Dunal) A. Rich, Xylopia parviflora (A. Rich) Benth.) and Monodora myristica (Gaertn) growing in Chad and Cameroon. BMC Complement Altern Med 14:1–8. [CrossRef]
- Bapela MJ, Kaiser M, Meyer JJM (2017) Antileishmanial activity of selected South African plant species. South African J Bot 108:342–345. [CrossRef]
- Barreiras DG, Ruiz FM, Gomes JEG, Souza BMS (2020) Eficácia da ação antimicrobiana do extrato de própolis de abelha jataí (Tetragonisca angustula) em bactérias Gram-positivas e Gram-negativas. Cad Ciências Agrárias 12:1–5. [CrossRef]
- Barros RPC (2014) Identificação de linhagens de Candida sp. e perfil de sensibilidade a antifúngicos convencionais e de origem vegetal. Universidade Estadual da paraíba.
- Basha SKH, Subramanian S (2011) Biochemical evaluation of antidiabetic and antioxidant potentials of Annona squamosa leaves extracts studied in STZ-induced diabetic rats. Int J Pharm Sci Res 2:643–655.
- Bay M, Souza de Oliveira JV, Sales Junior PA, et al (2019a) In Vitro Trypanocidal and Antibacterial Activities of Essential Oils from Four Species of the Family Annonaceae. Chem Biodivers 16:. [CrossRef]
- Bay M, Souza de Oliveira JV, Sales Junior PA, et al (2019b) In Vitro Trypanocidal and Antibacterial Activities of Essential Oils from Four Species of the Family Annonaceae. Chem Biodivers 16:. [CrossRef]
- Begum N, Sharma B, Pandey RS (2010) Evaluation of insecticidal efficacy of Calotropis procera and Annona squamosa ethanol extracts against Musca domestica. J Biofertil Biopestici 1:1–6.
- Bermejo A, Figadère B, Zafra-Polo MC, et al (2005) Acetogenins from annonaceae: Recent progress in isolation, synthesis and mechanisms of action. Nat. Prod. Rep. 22:269–303.
- Betancur-Galvis LA, Saez J, Granados H, et al (1999) Antitumor and Antiviral Activity of Colombian Medicinal Plant Extracts. Mem Inst Oswaldo Cruz 94:531–535. [CrossRef]
- Bhalke RD, Chavan MJ (2011) Analgesic and CNS depressant activities of extracts of Annona reticulata Linn. bark. Phytopharmacology 1:160–165.
- Bobadilla M, Zavala F, Sisniegas M, et al (2005) Evaluación larvicida de suspensiones acuosas de Annona muricata Linnaeus “guanábana” sobre Aedes aegypti Linnaeus (Diptera, Culicidae). Rev Peru Biol 12:145–152. [CrossRef]
- Bomfim LM, Menezes LRA, Rodrigues ACBC, et al (2016) Antitumour Activity of the Microencapsulation of Annona vepretorum Essential Oil. Basic Clin Pharmacol Toxicol 118:208–213. [CrossRef]
- Boyom FF, Ngouana V, Kemgne EAM, et al (2011) Antiplasmodial volatile extracts from Cleistopholis patens Engler & Diels and Uvariastrum pierreanum Engl. (Engl. & Diels) (Annonaceae) growing in Cameroon. Parasitol Res 108:1211–1217. [CrossRef]
- Breiman A, Hand L, Mashall JE, et al (1990) Pneumococcal bacteremia in Charleston County, South Carolina. A decade later. Arch Intern Med 150:1401–05.
- Brígido HPC, Correa-Barbosa J, da Silva-Silva JV, et al (2020) Antileishmanial activity of Annona species (Annonaceae). SN Appl Sci 2:1–8. [CrossRef]
- Brindis F, González-Trujano ME, González-Andrade M, et al (2013) Aqueous Extract of Annona macroprophyllata : A Potential α -Glucosidase Inhibitor. Biomed Res Int 2013:1–6. [CrossRef]
- Brito IA, Oliveira EA, Chaves MH, et al (2021) Antileishmanial acetylene fatty acid and acetogenins from seeds of porcelia macrocarpa. J Braz Chem Soc 32:447–453. [CrossRef]
- Brito IA, Thevenard F, Costa-Silva TA, et al (2022) Antileishmanial Effects of Acetylene Acetogenins from Seeds of Porcelia macrocarpa (Warm.) R.E. Fries (Annonaceae) and Semisynthetic Derivatives. Molecules 27:893. [CrossRef]
- Brito MT, Ferreira RC, Beltrão DM, et al (2018) Antitumor activity and toxicity of volatile oil from the leaves of Annona leptopetala. Rev Bras Farmacogn 28:602–609. [CrossRef]
- Calzada F, Solares-Pascasio J, Ordoñez-Razo R, et al (2017) Antihyperglycemic activity of the leaves from Annona cherimola miller and rutin on alloxan-induced diabetic rats. Pharmacognosy Res 9:1. [CrossRef]
- Calzada F, Valdes M, Garcia-Hernandez N, et al (2019) Antihyperglycemic activity of the leaves from annona diversifolia Safford. and farnesol on normal and alloxan-induced diabetic mice*. Pharmacogn Mag 15:5. [CrossRef]
- Carmona D, Sáez J, Granados H, et al (2003) Antiprotozoal 6-substituted-5,6-dihydro-α-pyrones from Raimondia cf. Monoica. Nat Prod Res 17:275–280. [CrossRef]
- Carneiro ÂP (2010) Atividade biocida de Annona coriacea Mart 1841 sobre o vetor Rhodnius neglectus Lent 1954 (Hemiptera: Reduviidae). Universidade do Estado de Mato Grosso.
- Cascaes MM, Carneiro ODS, Do Nascimento LD, et al (2021) Essential oils from annonaceae species from brazil: A systematic review of their phytochemistry, and biological activities. Int J Mol Sci 22:. [CrossRef]
- Castillo L, Castillo LE, Jimenez JJ, Delgado MA (2010) Secondary metabolites of the Annonaceae, Solanaceae and Meliaceae families used as biological control of insects. Trop Subtrop Agroecosystems 12:445–462.
- Cavalcanti BC, Ferreira JRO, Moura DJ, et al (2010) Structure-mutagenicity relationship of kaurenoic acid from Xylopia sericeae (Annonaceae). Mutat Res - Genet Toxicol Environ Mutagen 701:153–163. [CrossRef]
- Cercato LM, White PAS, Nampo FK, et al (2015) A systematic review of medicinal plants used for weight loss in Brazil: Is there potential for obesity treatment? J Ethnopharmacol 176:286–296. [CrossRef]
- Chakraborty P (2018) Herbal genomics as tools for dissecting new metabolic pathways of unexplored medicinal plants and drug discovery. Biochim. Open 6:9–16.
- Chan H-H, Hwang T-L, Thang T, et al (2013) Isolation and Synthesis of Melodamide A, a New Anti-inflammatory Phenolic Amide from the Leaves of Melodorum fruticosum. Planta Med 79:288–294. [CrossRef]
- Chanakul W, Tuchinda P, Anantachoke N, et al (2011) Cytotoxic alkaloids from stems, leaves and twigs of Dasymaschalon blumei. Fitoterapia 82:964–968. [CrossRef]
- Chang FR, Hwang TL, Yang YL, et al (2006) Anti-inflammatory and cytotoxic diterpenes from formosan Polyalthia longifolia var. pendula. Planta Med 72:1344–1347. [CrossRef]
- Chang FR, Yang P-Y, Lin J-Y, et al (1998) Bioactive Kaurane diterpenoids from Annona glabra. J Nat Prod 61:437–439. [CrossRef]
- Chatelain E (2015) Chagas Disease drug discovery: toward a new era. J Biomol Screen 20:.
- Chatelain E, Ioset JR (2011) Drug discovery and development for neglected diseases: the DNDi model. Drug Des Devel Ther 5:175. [CrossRef]
- Chavan MJ, Wakte PS, Shinde DB (2011) Analgesic and anti-inflammatory activities of 18-acetoxy-ent-kaur-16-ene from Annona squamosa L. bark. Inflammopharmacology 19:111–115. [CrossRef]
- Chavan MJ, Wakte PS, Shinde DB (2010a) Analgesic and anti-inflammatory activity of Caryophyllene oxide from Annona squamosa L. bark. Phytomedicine 17:149–151. [CrossRef]
- Chavan MJ, Wakte PS, Shinde DB (2012) Analgesic and anti-inflammatory activities of the sesquiterpene fraction from Annona reticulata L. bark. Nat Prod Res 26:1515–1518. [CrossRef]
- Chavan MJ, Wakte PS, Shinde DB (2010b) Analgesic and anti-inflammatory activities of saponified fraction from Annona reticulata L. Bark. Lat Am J Pharm 29:1246–1249.
- Chen CH, Hsieh TJ, Liu TZ, et al (2004) Annoglabayin, a novel dimeric kaurane diterpenoid, and apoptosis in Hep G2 cells of annomontacin from the fruits of Annona glabra. J Nat Prod 67:1942–1946. [CrossRef]
- Chen CY, Chang FR, Shih YC, et al (2000) Cytotoxic constituents of Polyalthia longifolia var. pendula. J Nat Prod 63:1475–1478. [CrossRef]
- Chen Y-C, Chia Y-C, Huang B-M (2021) Phytochemicals from Polyalthia Species: Potential and Implication on Anti-Oxidant, Anti-Inflammatory, Anti-Cancer, and Chemoprevention Activities. Molecules 26:5369. [CrossRef]
- Chen Y, Chen JW, Xu SS, et al (2012) Antitumor activity of annonaceous acetogenins in HepS and S180 xenografts bearing mice. Bioorganic Med Chem Lett 22:2717–2719. [CrossRef]
- Chowdhury SS, Tareq AM, Tareq SM, et al (2021) Screening of antidiabetic and antioxidant potential along with phytochemicals of Annona genus: a review. Futur J Pharm Sci 7:. [CrossRef]
- Christopher R, Mgani Q, Nyandoro S, et al (2018) Antitrypanosomal, antiplasmodial, and antibacterial activities of extracts from selected Diospyros and Annonaceae species. J Complement Med Res 7:161. [CrossRef]
- Coelho MB, Marangoni S, Macedo MLR (2007) Insecticidal action of Annona coriacea lectin against the flour moth Anagasta kuehniella and the rice moth Corcyra cephalonica (Lepidoptera: Pyralidae). Comp Biochem Physiol - C Toxicol Pharmacol 146:406–414. [CrossRef]
- Colom OA, Barrachina I, Mingol IA, et al (2008) Toxic effects of annonaceous acetogenins on Oncopeltus fasciatus. J Pest Sci (2004) 81:85–89. [CrossRef]
- Coria-Téllez A V., Montalvo-Gónzalez E, Yahia EM, Obledo-Vázquez EN (2018) Annona muricata: A comprehensive review on its traditional medicinal uses, phytochemicals, pharmacological activities, mechanisms of action and toxicity. Arab J Chem 11:662–691. [CrossRef]
- Cortes D, Moreno L, Párraga J, et al (2014) Nuevos fármacos inspirados en annonáceas. Rev Bras Frutic 36:22–31. [CrossRef]
- Costa, Marilza da Silva, Mônica Josene Barbosa Pereira, Simone Santos de Oliveira, Paulo Teixeira de Souza, Evandro Luiz Dall’oglio TCA (2013) ARTIGO Anonáceas provocam mortalidade em larvas de Aedes aegypti ( Linnaeus , 1762 ) ( Diptera : Culicidae ). Rev Bras Biociências v. 11, n 2:184–190.
- Costa EV, Pinheiro MLB, De Souza ADL, et al (2011a) Trypanocidal activity of oxoaporphine and pyrimidine-β-carboline alkaloids from the branches of annona foetida mart. (annonaceae). Molecules 16:9714–9720. [CrossRef]
- Costa EV, Pinheiro MLB, Silva JRDA, et al (2009) Antimicrobial and antileishmanial activity of essential oil from the leaves of Annona foetida (Annonaceae). Quim Nova 32:78–81. [CrossRef]
- Costa MS, Pinheiro DO, Serrão JE, Pereira MJB (2012) Morphological Changes in the Midgut of Aedes aegypti L. (Diptera: Culicidae) Larvae Following Exposure to an Annona coriacea (Magnoliales: Annonaceae) Extract. Neotrop Entomol 41:311–314. [CrossRef]
- Costa RAP, Romagna CD, Pereira JL, Souza-Pinto NC (2011b) The role of mitochondrial DNA damage in the citotoxicity of reactive oxygen species. J Bioenerg Biomembr 43:25–29.
- Costa RGA, Anunciação TA da, Araujo M de S, et al (2020) In vitro and in vivo growth inhibition of human acute promyelocytic leukemia HL-60 cells by Guatteria megalophylla Diels (Annonaceae) leaf essential oil. Biomed Pharmacother 122:109713. [CrossRef]
- Costa E V., Marques F de A, Pinheiro MLB, et al (2011c) Chemical constituents isolated from the bark of Guatteria blepharophylla (Annonaceae) and their antiproliferative and antimicrobial activities. J Braz Chem Soc 22:1111–1117. [CrossRef]
- Costa E V., Soares LDN, Chaar J da S, et al (2021) Benzylated dihydroflavones and isoquinoline-derived alkaloids from the bark of diclinanona calycina (Annonaceae) and their cytotoxicities. Molecules 26:1–13. [CrossRef]
- Costa E V., Teixeira SD, Marques FA, et al (2008) Chemical composition and antimicrobial activity of the essential oils of the Amazon Guatteriopsis species. Phytochemistry 69:1895–1899. [CrossRef]
- Cronquist A (1981) An integrated system of classification of flowering plants. Columbia University Press.
- Cruz-Estrada A, Gamboa-Angulo M, Borges-Argáez R, Ruiz-Sánchez E (2013) Insecticidal effects of plant extracts on immature whitefly Bemisia tabaci Genn.(Hemiptera: Aleyroideae). Electron J Biotechnol 16:6.
- Cruz PEO da (2011) Estudo fitoquímico e investigação das atividades antioxidante, antimicrobiana e larvicida das cascas de Annona salzmannii A. DC.(Annonaceae).
- Cunha LMA (2009) Estudo Fitoquímico e Biológico de Duguetia riparia ( Annonaceae ). Univerdidade Federal do Amazonas.
- da Silva APT, Pereira MJB, Bento LF (2007) Extrato metanólico da semente de Annona coriacea (MART.) sobre a mortalidade da traça-do-tomateiro. Rev Bras Agroecol 2:1150–1153.
- da Silva DB, Tulli ECO, Militão GCG, et al (2009) The antitumoral, trypanocidal and antileishmanial activities of extract and alkaloids isolated from Duguetia furfuracea. Phytomedicine 16:1059–1063. [CrossRef]
- da Silva FMA, de Lima BR, Soares ER, et al (2015) Polycarpol in unonopsis, Bocageopsis and Onychopetalum Amazonian species: Chemosystematical implications and antimicrobial evaluation. Rev Bras Farmacogn 25:11–15. [CrossRef]
- da Silva FMA, Koolen HHF, de Lima JPS, et al (2012) Leishmanicidal activity of fractions rich in aporphine alkaloids from Amazonian Unonopsis species. Rev Bras Farmacogn 22:1368–1371. [CrossRef]
- Da Silva MS, Tavares JF, Queiroga KF, et al (2009) Alcaloides e outros constituintes de Xylopia langsdorffiana (Annonaceae). Quim Nova 32:1566–1570.
- Da Silva TB, Menezes LRA, Sampaio MFC, et al (2013a) Chemical composition and anti-trypanosoma cruzi activity of essential oils obtained from leaves of Xylopia frutescens and X. laevigata (Annonaceae). Nat Prod Commun 8:403–406. [CrossRef]
- Da Silva VP, Pereira MJB, Turchen LM (2013b) Efeito de extratos vegetais no controle de Euschistus heros (F.) (Hemiptera: Pentatomidae) em lavoura de soja na região sudeste do estado de Mato Grosso. BRAZILIAN J Agric - Rev Agric 88:185. [CrossRef]
- Dadang EDF, Prijono D (2009) Effectiveness of two botanical insecticide formulations to two major cabbage insect pests on field application. J ISSAAS 15:42–51.
- Dantas EP, Monteiro J, Medeiros LS de, et al (2020) Dereplication of Aporphine Alkaloids by UHPLC-HR-ESI-MS/MS and NMR from. 31:1908–1916.
- Davies J, Davies D (2010) Origins and Evolution of Antibiotic resistance. Microbiol Mol Biol Rev 74:417–433.
- de Andrade FHD, de Araújo Batista RS, Lins TB, et al (2019) Thermal characterization and microbiology assay of Annona muricata L. leaves. J Therm Anal Calorim 138:3737–3745. [CrossRef]
- de Cássia Seffrin R, Shikano I, Akhtar Y, Isman MB (2010) Effects of crude seed extracts of Annona atemoya and Annona squamosa L. against the cabbage looper, Trichoplusia ni in the laboratory and greenhouse. Crop Prot 29:20–24. [CrossRef]
- De Lima JPS, Pinheiro MLB, Antonio AM, et al (2012) In Vitro Atileishmanial and Cytotoxic Activities of Annona mucosa (Annonaceae). Rev Virtual Quim 4:692–702. [CrossRef]
- De Mesquita ML, Desrivot J, Bories C, et al (2005) Antileishmanial and trypanocidal activity of Brazilian Cerrado plants. Mem Inst Oswaldo Cruz 100:783–787. [CrossRef]
- de Moraes JM, Pereira MJB, da Silva Costa M, et al (2011) Avaliação da atividade de Annona coriacea (Annonaceae) sobre pupas e adultos de Aedes aegypti (Diptera: Culicidae) em laboratório. BRAZILIAN J Agric Agric 86:115–121.
- de Moura F, Justino A, Ferreira B, et al (2019) Pro-Fibrogenic and Anti-Inflammatory Potential of a Polyphenol-Enriched Fraction from Annona crassiflora in Skin Repair. Planta Med 85:570–577. [CrossRef]
- de Omena MC, Navarro DMAF, de Paula JE, et al (2007) Larvicidal activities against Aedes aegypti of some Brazilian medicinal plants. Bioresour Technol 98:2549–2556. [CrossRef]
- Deshmukhe P V, Hooli AA, Holihosur SN (2010) Bioefficacy of cold ethyl alcohol extract of Annona squamosa against Spodoptera litura Fabricius. J Biopestic 3:271.
- Dikti Vildina J, Ndjonka D, Schmidt TJ, Liebau E (2021) Identification of anti-Caenorhabditis and anti-Onchocerca constituents from leaves of Annona senegalensis Pers. (Annonaceae). South African J Bot 138:84–93. [CrossRef]
- Dill EM, Pereira MJB, Costa MS (2012) Efeito residual do extrato de Annona coriacea sobre Aedes aegypti. Arq Inst Biol (Sao Paulo) 79:595–602. [CrossRef]
- Ding L, Münch J, Goerls H, et al (2010) Xiamycin, a pentacyclic indolosesquiterpene with selective anti-HIV activity from a bacterial mangrove endophyte. Bioorganic Med Chem Lett 20:6685–6687. [CrossRef]
- DNDi D for ND initiative (2022) Chagas Disease.
- do Santos RC, de Souza AV, Andrade-Silva M, et al (2018) Antioxidant, anti-rheumatic and anti-inflammatory investigation of extract and dicentrinone from Duguetia furfuracea (A. St.-Hil.) Benth. & Hook. f. J Ethnopharmacol 211:9–16. [CrossRef]
- Doyle JA, Sauquet H, Scharaschkin T, Le Thomas A (2004) Phylogeny, molecular and fossil dating, and biogeographic history of annonaceae and myristicaceae (Magnoliales). In: International Journal of Plant Sciences.
- Duarte-Almeida JM, Santos RJ dos, Genovese MI, Lajolo FM (2006) Avaliação da atividade antioxidante utilizando sistema beta-caroteno/ácido linoléico e método de seqüestro de radicais DPPH. Ciência e Tecnol Aliment 26:446–452. [CrossRef]
- Ducasse M, Brown MA (2006) Epigenetic aberrations and cancer. Mol Cancer 5:60.
- Dutra LM, Bomfim LM, Rocha SLA, et al (2014) Bioorganic & Medicinal Chemistry Letters ent -Kaurane diterpenes from the stem bark of Annona vepretorum ( Annonaceae ) and cytotoxic evaluation. 24:3315–3320. [CrossRef]
- Ellis L, Atadja PW, Johnstone RW (2009) Epigenetics in cancer: targeting chromatin modifications. Mol Cancer Ther 8:1409–1420.
- Emerson F. Queiroz, François Roblot and, Cavé* A, et al (1996) Pessoine and Spinosine, Two Catecholic Berbines from Annona spinescens1. [CrossRef]
- Encina CL, Martin EC, Lopez AA, Padilla IMG (2014) Biotechnology applied to Annona species: a review. Rev Bras Frutic 36:17–21. [CrossRef]
- Epino PB, Chang F (1993) Insecticidal activity of Annona squamosa L. seed extracts against the Mediterranean fruit fly, Ceratitis capitata (Wiedemann)(Diptera: Tephritidae). Philipp Entomol.
- Esteller M (2008) Epigenetics in cancer. N Engl J Med 358:1148–1159.
- Etame RME, Mouokeu RS, Poundeu FSM, et al (2019) Effect of fractioning on antibacterial activity of n-butanol fraction from Enantia chlorantha stem bark methanol extract. BMC Complement Altern Med 19:1–7. [CrossRef]
- Ewete FK, Arnason JT, Larson J, Philogene BJR (1996) Biological activities of extracts from traditionally used Nigerian plants against the European corn borer, Ostrinia nubilalis. Entomol Exp Appl 80:531–537.
- Ezekwesili CN, Nwodo OFC, Eneh FU, Ogbunugafor HA (2010) Investigation of the chemical composition and biological activity of Xylopia aethiopica Dunal (Annonaceae). African J Biotechnol 9:7352–7356.
- Faizi S, Khan RA, Azher S, et al (2003) New Antimicrobial Alkaloids from the Roots of Polyalthia longifolia var. pendula. Planta Med 69:350–355. [CrossRef]
- Faizi S, Khan RA, Mughal NR, et al (2008) Antimicrobial activity of various parts of Polyalthia longifolia var. pendula: Isolation of active principles from the leaves and the berries. Phyther Res 22:907–912. [CrossRef]
- Fatope MO, Audu OT, Takeda Y, et al (1996) Bioactive ent-kaurene diterpenoids from Annona senegalensis. J Nat Prod 59:301–303. [CrossRef]
- Fekam Boyom F, Ngouana V, Amvam Zollo PH, et al (2003) Composition and anti-plasmodial activities of essential oils from some Cameroonian medicinal plants. Phytochemistry 64:1269–1275. [CrossRef]
- Fernandes AT (2000) Infecção Hospitalar e suas interfaces na área da saúde, 1st editio. São Paulo, Brasil.
- Ferraz RPC, Cardoso GMB, Da Silva TB, et al (2013) Antitumour properties of the leaf essential oil of Xylopia frutescens Aubl. (Annonaceae). Food Chem 141:542–547. [CrossRef]
- Ferreira C, Passos CLA, Soares DC, et al (2017) Leishmanicidal activity of the alkaloid-rich fraction from Guatteria latifolia. Exp Parasitol 172:51–60. [CrossRef]
- Ferreira G, De-La-Cruz-Chacón I, Fernandes Boaro CS, et al (2019) Propagation of Annonaceous plants. Rev Bras Frutic 41:1–14. [CrossRef]
- Ferreira J, Kurachi C, Melo CAS, et al (2004) Necrosis Characteristics of Photodynamics Therapy. Laser Phys 14:209–212.
- Février A, Ferreira ME, Fournet A, et al (1999) Acetogenins and other compounds from Rollinia emarginata and their antiprotozoal activities. Planta Med 65:47–49. [CrossRef]
- Figueroa-Miranda G, Wu C, Zhang Y, et al (2020) Polyethylene glycol-mediated blocking and monolayer morphology of anelectrochemical aptasensor for malaria biomarker detection in humanserum. Bioelectrochemistry 136:1–11.
- Florence NT, Benoit MZ, Jonas K, et al (2014) Antidiabetic and antioxidant effects of Annona muricata (Annonaceae), aqueous extract on streptozotocin-induced diabetic rats. J Ethnopharmacol 151:784–790. [CrossRef]
- Fontana JD, Passos M, Baron M, et al (1998) Selective polarity-and adsorption-guided extraction/purification of Annona sp. polar acetogenins and biological assay against agricultural pests. In: Biotechnology for Fuels and Chemicals. Springer, pp 67–76.
- Franke A, Teyssen S, Singer M V. (2005) Alcohol-Related Diseases of the Esophagus and Stomach. Dig Dis 23:204–213. [CrossRef]
- Fujiwara N, Kobayashi K (2005) Macrophages in Inflammation. Curr Drug Target -Inflammation Allergy 4:281–286. [CrossRef]
- Gajalakshmi S, Vijayalakshmi S, Davi Rajeswari V (2012) Phytochemical and pharmacological properties of Annona muricata: a review. Int J Pharm Sci Res 4:3–6.
- Gavamukulya Y, Abou-Elella F, Wamunyokoli F, AEl-Shemy H (2014) Phytochemical screening, anti-oxidant activity and in vitro anticancer potential of ethanolic and water leaves extracts of Annona muricata (Graviola). Asian Pac J Trop Med 7:S355–S363. [CrossRef]
- Gbedema SY, Bayor MT, Annan K, Wright CW (2015) Clerodane diterpenes from Polyalthia longifolia (Sonn) Thw. var. pendula: Potential antimalarial agents for drug resistant Plasmodium falciparum infection. J Ethnopharmacol 169:176–182. [CrossRef]
- Gomes CL, Cavalcante JE, Cunha FA, et al (2010) Identificação e perfil de sensibilidade de Candida spp. isoladas de urina de pacientes com candidúria em Iguatu-Ceará. Rev Bras Análises Cl;inicas 42:223–225.
- Gomes JVD, Borges AS, Athaydes BR, et al (2022) Anti-Helicobacter pylori potential, antioxidant capacity, and anti-inflammatory activity of Xylopia sericea A. St.-Hil. (Annonaceae) leaves. Phytomedicine Plus 2:100214. [CrossRef]
- González-Esquinca AR, Luna-Cazáres LM, Schlie Guzmán MA, et al (2012) In Vitro Larvicidal Evaluation of Annona muricata L., A. diversifolia Saff. and A. lutescens Saff. Extracts Against Anastrepha ludens Larvae (Diptera, Tephritidae). Interciencia 37:284–289.
- Guadaño A, Gutiérrez C, de la Peña E, et al (2000) Insecticidal and mutagenic evaluation of two annonaceous acetogenins. J Nat Prod 63:773–776. [CrossRef]
- Guarido MM (2009) Atividade inseticida de extratos de Annona foetida Mart.(Annonaceae) sobre imaturos de Aedes aegypti (Linnaeus, 1762)(Diptera: Culicidae).
- Guo H, Callaway JB, Ting JP-Y (2015) Inflammasomes: mechanism of action, role in disease, and therapeutics. Nat Med 21:677–687. [CrossRef]
- Guo X, Tang CC, Thomas DC, et al (2017) A mega-phylogeny of the Annonaceae: Taxonomic placement of five enigmatic genera and support for a new tribe, Phoenicantheae. Sci Rep 7:1–11. [CrossRef]
- Hadjichristou C, Koidis P, Bakopoulou A (2020) Advanced in Vitro Experimental Models for Tissue Engineering-based Reconstruction of a 3D Dentin/pulp Complex: a Literature Review. Stem Cell Rev Reports 1–18.
- Hammami F, Koubaa M, Chaari M, et al (2020) Haemophagocytic lymphohistiocytosis secondary to Plasmodium falciparum malaria: Case report and review of the literature. Asian Pac J Trop Med 13:467–471. [CrossRef]
- Hassan SS ul, Zhang W-D, Jin H, et al (2022) In-silico anti-inflammatory potential of guaiane dimers from Xylopia vielana targeting COX-2. J Biomol Struct Dyn 40:484–498. [CrossRef]
- Henao GJP, Pajón CMG, Torres JMC (2007) Actividad insecticida de extractos vegetales sobre Aedes aegypti (Diptera: Culicidae) vector del dengue en Colombia. Ces Med 21:.
- Hisham A, Rameshkumar KB, Sherwani N, et al (2012) The composition and antimicrobial activities of Cyperus conglomeratus, Desmos chinensis var. lawii and Cyathocalyx zeylanicus essential oils. Nat Prod Commun 7:663–666. [CrossRef]
- Hoes JN, Jacobs JWG, Buttgereit F, Bijlsma JWJ (2010) Current view of glucocorticoid co-therapy with DMARDs in rheumatoid arthritis. Nat Rev Rheumatol 6:693–702. [CrossRef]
- Hongthong S, Kuhakarn C, Jaipetch T, et al (2016) A new neolignan, and the cytotoxic and anti-HIV-1 activities of constituents from the roots of Dasymaschalon sootepense. Nat Prod Commun 11:809–813. [CrossRef]
- Horton NC, Mathew PA (2015) NKp44 and natural cytotoxicity receptors as damage-associated molecular pattern recognition receptors. Front Immunol 6:31.
- Hsu Y-M, Wu T-Y, Du Y-C, et al (2016) 3-Methyl-4,5-dihydro-oxepine, polyoxygenated seco-cyclohexenes and cyclohexenes from Uvaria flexuosa and their anti-inflammatory activity. Phytochemistry 122:184–192. [CrossRef]
- Hu JF, Garo E, Hough GW, et al (2006) Antibacterial, partially acetylated oligorhamnosides from Cleistopholis patens. J Nat Prod 69:585–590. [CrossRef]
- Huber LS, Rodriguez-Amaya DB (2008) Flavonóis e flavonas: fontes brasileiras e fatores que influenciam a composição em alimentos. Aliment e Nutr 18:97–108.
- Huong DT, Luong D V., Thao TTP, Sung T Van (2005) A new flavone and cytotoxic activity of flavonoid constituents isolated from Miliusa balansae (Annonaceae). Pharmazie 60:627–629. [CrossRef]
- Huscher D, Thiele K, Gromnica-Ihle E, et al (2009) Dose-related patterns of glucocorticoid-induced side effects. Ann Rheum Dis 68:1119–1124. [CrossRef]
- Ik-Soo Lee, Xianjian Ma, Hee-Byung Chai, et al (1995) Novel cytotoxic labdane diterpenoids from Neouvaria acuminatissima. Tetrahedron 51:21–28. [CrossRef]
- Instituto Nacional de Câncer José Alencar Gomes da Silva (INCA) (2019) Estimativa 2020: incidência de Cancer no Brasil. p 120p.
- IOM I of M (2010) Antibiotic resistance: implications for global health and novel intervention strategies: workshop summary. Washington, DC: The National Academies Press.
- Ishola IO, Awodele O, Olusayero AM, Ochieng CO (2014) Mechanisms of Analgesic and Anti-Inflammatory Properties of Annona muricata Linn. (Annonaceae) Fruit Extract in Rodents. J Med Food 17:1375–1382. [CrossRef]
- Jachak S (2006) Cyclooxygenase Inhibitory Natural Products: Current Status. Curr Med Chem 13:659–678. [CrossRef]
- Jain P, Patra A, Jain S, et al (2014) Antibacterial and Antioxidant Activities of 3-O-methyl Ellagic Acid 4’-rhamnoside from Stem Bark of Polyalthia longifolia Thw. 45:b031. [CrossRef]
- Jossang A, Leboeuf M, Cavé A, et al (1984) Alcaloïdes des Annonacees, L: Alcaloïdes De Polyalthia cauliflora. J Nat Prod 47:504–513. [CrossRef]
- Joy B, Remani P (2008) Antitumor constituents from Annona squamosa fruit pericarp. Med Chem Res 17:345–355. [CrossRef]
- Kabir KE (2010) Larvicidal effect of an alkaloidal fraction of Artabotrys odoratissimus (Annonaceae) bark against the filarial mosquito Culex quinquefasciatus (Diptera: Culicidae). Int J Trop Insect Sci 30:167–169.
- Kabir S, Zahan R, Chowdhury AMS, et al (2019) Evaluation of Analgesic and Anti-inflammatory Activities of Polyalthia simiarum (Hook. F. &Thomson). J Natl Inst Neurosci Bangladesh 5:18–23. [CrossRef]
- Kaleem M, Medha P, Ahmed QU, et al (2008) Beneficial effects of Annona squamosa extract in streptozotocin-induced diabetic rats. Singapore Med J 49:800–4.
- Kamaraj C, Bagavan A, Elango G, et al (2011) Larvicidal activity of medicinal plant extracts against Anopheles subpictus & Culex tritaeniorhynchus. Indian J Med Res 134:101.
- Kambiré DA, Boti JB, Kablan ACL, et al (2021) Chemical Variability and In Vitro Anti-Inflammatory Activity of Leaf Essential Oil from Ivorian Isolona dewevrei (De Wild. & T. Durand) Engl. & Diels. Molecules 26:6228. [CrossRef]
- Kanokmedhakul S, Kanokmedhakul K, Kantikeaw I, Phonkerd N (2006) 2-Substituted furans from the roots of Polyalthia evecta. J Nat Prod 69:68–72. [CrossRef]
- Kanokmedhakul S, Kanokmedhakul K, Yodbuddee D, Phonkerd N (2003) New antimalarial bis-dehydroaporphine alkaloids from Polyalthia debilis. J Nat Prod 66:616–619. [CrossRef]
- Karioti A, Hadjipavlou-Litina D, Mensah MLK, et al (2004) Composition and antioxidant activity of the essential oils of Xylopia aethiopica (Dun) A. Rich. (Annonaceae) leaves, stem bark, root bark, and fresh and dried fruits, growing in Ghana. J Agric Food Chem 52:8094–8098. [CrossRef]
- Karunaratne M, Arukwatta A (2009) Efficacy of three plant species on the mortality and food consumption of Epilachna vigintioctopunctata.
- Kawazu K, Alcantara JP, Kobayashi A (1989) Isolation and structure of neoannonin, a novel insecticidal compound from the seeds of Annona squamosa. Agric Biol Chem 53:2719–2722.
- Kempraj V, Bhat SK (2011) Acute and reproductive toxicity of Annona squamosa to Aedes albopictus. Pestic Biochem Physiol 100:82–86.
- Kesetyaningsih TW (2012) Efficacy of Annona squamosa leaf extract as an insecticide against Cockroach (Periplaneta americana).
- Khalequzzaman M, Sultana S (2006) Insecticidal activity of Annona squamosa L. seed extracts against the red flour beetle, Tribolium castaneum (Herbst). J Bio-Science 14:107–112.
- Kim EJ, Suh KM, Kim DH, et al (2005) Asimitrin and 4-hydroxytrilobin, new bioactive annonaceous acetogenins from the seeds of Asimina triloba possessing a bis-tetrahydrofuran ring. J Nat Prod 68:194–197. [CrossRef]
- Kithsiri Wijeratne EM, Leslie Gunatilaka AA, Kingston DGI, et al (1995) Artabotrine: A novel bioactive alkaloid from Artabotrys zeylanicus. Tetrahedron 51:7877–7882. [CrossRef]
- Klocke JA, Mei-Ying H, Shin-Foon C, Kubo I (1991) Grayanoid diterpene insect antifeedants and insecticides from Rhododendron molle. Phytochemistry 30:1797–1800. [CrossRef]
- Konan N, Kouame BA, Mamyrbekova-Bekro JA, et al (2009) Chemical composition and antioxidant activities of essential oils of xylopia aethiopica (dunal) a. rich. Eur J Sci Res 37:311–318.
- Kotkar HM, Mendki PS, Sadan SVGS, et al (2002) Antimicrobial and pesticidal activity of partially purified flavonoids of Annona squamosa. Pest Manag Sci Former Pestic Sci 58:33–37.
- Kouam SF, Ngouonpe AW, Lamshöft M, et al (2014) Indolosesquiterpene alkaloids from the Cameroonian medicinal plant Polyalthia oliveri (Annonaceae). Phytochemistry 105:52–59. [CrossRef]
- Krinski D, Massaroli A (2014) Nymphicidal effect of vegetal extracts of Annona mucosa and Anonna crassiflora (Magnoliales, Annonaceae) against rice stalk stink bug, Tibraca limbativentris (Hemiptera, Pentatomidae). Rev Bras Frutic 36:217–224. [CrossRef]
- Krinski D, Massaroli A, Machado M (2014) Potencial inseticida de plantas da família Annonaceae. Rev Bras Frutic 36:225–242. [CrossRef]
- Kumar JA, Rekha T, Devi SS, et al (2010) Insecticidal activity of ethanolic extract of leaves of Annona squamosa. J Chem Pharm Res 2:177–180.
- Lamidi M, DiGiorgio C, Delmas F, et al (2005) In vitro cytotoxic, antileishmanial and antifungal activities of ethnopharmacologically selected Gabonese plants. J Ethnopharmacol 102:185–190. [CrossRef]
- Leatemia JA, Isman MB (2004) Efficacy of crude seed extracts of Annona squamosa against diamondback moth, Plutella xylostella L. in the greenhouse. Int J Pest Manag 50:129–133.
- Leboeuf M, Cavé A, Bhaumik PK, et al (1980) The phytochemistry of the annonaceae. Phytochemistry 21:2783–2813. [CrossRef]
- Lee TH, Wang MJ, Chen PY, et al (2009) Constituents of Polyalthia longifolia var. pendula. J Nat Prod 72:1960–1963. [CrossRef]
- Leite DOD, Camilo CJ, Nonato C de FA, et al (2021) Chemical profile and evaluation of the antioxidant and anti-acetylcholinesterase activities of annona squamosa l. (annonaceae) extracts. Foods 10:. [CrossRef]
- Lemos DECV, Cavalcante-Silva LHA, de Almeida Lima É, et al (2017) Anti-inflammatory Effect of Discretamine, a Protoberberine Alkaloid Isolated from Duguetia moricandiana. Nat Prod Commun 12:1934578X1701201. [CrossRef]
- Lepper MH, Dowling HF (1951) Treatment of pneumococcic meningitis with penicillin compared with penicillin plus aureomycin: Studies including observations on an apparent antagonism between penicillin and aureomycin. AMA Arch Intern Med 88:489–494.
- Li C, Lee D, Graf TN, et al (2009) Bioactive Constituents of the Stem Bark of Mitrephora glabra. J Nat Prod 72:1949–1953. [CrossRef]
- Lima LAR dos S, Alves TMA, Zani CL, et al (2014) In vitro cytotoxic, antifungal, trypanocidal and leishmanicidal activities of acetogenins isolated from Annona cornifolia A. St. -Hil. (Annonaceae). An Acad Bras Cienc 86:829–839. [CrossRef]
- Link A, Balaguer F, Goel A (2010) Cancer chemoprevention by dietary polyphenols: promising role for epigenetics. Biochem Pharmacol 80:1771–1792.
- Liu X, Wang J (2011) Anti-inflammatory effects of iridoid glycosides fraction of Folium syringae leaves on TNBS-induced colitis in rats. J Ethnopharmacol 133:780–787. [CrossRef]
- Llanos CAH, Arango DL, Giraldo MC (2008) Actividad insecticida de extractos de semilla de Annona muricata (Anonaceae) sobre Sitophilus zeamais (Coleoptera: Curculionidae). Rev Colomb Entomol 34:76–82.
- Londero VS, da Costa-Silva TA, Gomes KS, et al (2018) Acetylenic fatty acids from Porcelia macrocarpa (Annonaceae) against trypomastigotes of Trypanosoma cruzi: Effect of octadec-9-ynoic acid in plasma membrane electric potential. Bioorg Chem 78:307–311. [CrossRef]
- López-Rubalcava C, Piña-Medina B, Estrada-Reyes R, et al (2006) Anxiolytic-like actions of the hexane extract from leaves of Annona cherimolia in two anxiety paradigms: Possible involvement of the GABA/benzodiazepine receptor complex. Life Sci 78:730–737. [CrossRef]
- López R, Cuca LE, Delgado G (2009) Antileishmanial and immunomodulatory activity of Xylopia discreta. Parasite Immunol 31:623–630. [CrossRef]
- Lorenzo V, Lúcio A, Scotti L, et al (2016) Structure- and Ligand-Based Approaches to Evaluate Aporphynic Alkaloids from Annonaceae as Multi-Target Agent Against Leishmania donovani. Curr Pharm Des 22:5196–5203. [CrossRef]
- Lorenzo VP, Scotti L, da Silva Almeida JRG, Scotti MT (2020) Annonaceae Family Alkaloids as Agents Against Leishmaniasis: A Review and Molecular Docking Evaluation. Curr Drug Metab 21:482–492. [CrossRef]
- LS NT, Goudoum A, Ngassoum MB, et al (2004) Persistance of the insecticidal activity of five essential oils on the maize weevil Sitophilus zeamais (Motsch.)(Coleoptera: Curculionidae). Commun Agric Appl Biol Sci 69:145–147.
- Luciana BS, Jorlan C da S, Bruno EP, et al (2013) Insecticide irritability of plant extracts against Sitophilus zeamais. African J Agric Res 8:978–983.
- Lúcio ASSC, Da Silva Almeida JRG, Barbosa-Filho JM, et al (2011) Azaphenanthrene alkaloids with antitumoral activity from anaxagorea dolichocarpa sprague and sandwith (annonaceae). Molecules 16:7125–7131. [CrossRef]
- Ma C, Chen Y, Chen J, et al (2017) A Review on Annona squamosa L.: Phytochemicals and Biological Activities . Am J Chin Med 45:933–964. [CrossRef]
- Macedo T, Ribeiro V, Oliveira AP, et al (2020) Anti-inflammatory properties of Xylopia aethiopica leaves: Interference with pro-inflammatory cytokines in THP-1-derived macrophages and flavonoid profiling. J Ethnopharmacol 248:112312. [CrossRef]
- Machana S, Weerapreeyakul N, Barusrux S, et al (2012) Synergistic anticancer effect of the extracts from Polyalthia evecta caused apoptosis in human hepatoma (HepG2) cells. Asian Pac J Trop Biomed 2:589–596. [CrossRef]
- Maeda G, Gilissen PJ, Bourgard C, et al (2022) Polyoxygenated cyclohexene derivatives and flavonoids from the leaves of Uvaria pandensis. Fitoterapia 158:. [CrossRef]
- Magadula JJ, Innocent E, Otieno JN (2009) Mosquito larvicidal and cytotoxic activities of 3 Annona species and isolation of active principles. J Med Plants Res 3:674–680.
- Mahiou V, Roblot F, Fournet A, Hocquemiller R (2000a) Bisbenzylisoquinoline alkaloids from Guatteria boliviana (Annonaceae). Phytochemistry 54:709–716. [CrossRef]
- Mahiou V, Roblot F, Fournet A, Hocquemiller R (2000b) Bisbenzylisoquinoline alkaloids from Guatteria boliviana (Annonaceae). Phytochemistry 54:709–716. [CrossRef]
- Mahmud PIAM, Yaacob WA, Ibrahim N, Bakar MA (2018) Antibacterial activity and major constituents of polyalthia cinnamomea basic fraction. Sains Malaysiana 47:2063–2071. [CrossRef]
- Malebo HM, Wenzler T, Cal M, et al (2013a) Anti-protozoal activity of aporphine and protoberberine alkaloids from Annickia kummeriae (Engl. & Diels) Setten & Maas (Annonaceae). BMC Complement Altern Med 13:1–10. [CrossRef]
- Malebo HM, Wenzler T, Cal M, et al (2013b) Anti-protozoal activity of aporphine and protoberberine alkaloids from Annickia kummeriae. BMC Complement Altern Med 13:1–10.
- Manoj Kumar S, Azamthulla M, Saravanan KS (2021) Pharmacognostical evaluation and anti-convulsant property of Annona reticulata Linn. (Annonaceae) root. Futur J Pharm Sci 7:. [CrossRef]
- Martínez-Solís J, Calzada F, Barbosa E, Valdés M (2021) Antihyperglycemic and Antilipidemic Properties of a Tea Infusion of the Leaves from Annona cherimola Miller on Streptozocin-Induced Type 2 Diabetic Mice. Molecules 26:2408. [CrossRef]
- Martínez-Vázquez M, Estrada-Reyes R, Araujo Escalona AG, et al (2012) Antidepressant-like effects of an alkaloid extract of the aerial parts of Annona cherimolia in mice. J Ethnopharmacol 139:164–170. [CrossRef]
- Massaroli A, Pereira MJB, Foerster LA (2013) Annona mucosa como fitoinseticida para o controle de Chrysodeixis includens (Lepidoptera: Noctuidae). SIMPÓSIO Control BIOLÓGICO 13:.
- Matchi JLT, Noungoue DT, Chaubet G, et al (2022) Antischistosomal evaluation of stem bark’s extract and chemiacl constituents from Anonidium mannii against schistosoma mansoni. Pharmacogn Mag 17:752–758.
- Mazumdar S, Ghosh AK, Dinda M, et al (2021) Evaluation of wound healing activity of ethanol extract of Annona reticulata L. leaf both in vitro and in diabetic mice model. J Tradit Complement Med 11:27–37. [CrossRef]
- Mbosso EJT, Ngouela S, Nguedia JCA, et al (2010) In vitro antimicrobial activity of extracts and compounds of some selected medicinal plants from Cameroon. J Ethnopharmacol 128:476–481. [CrossRef]
- Meira CS, Guimarães ET, MacEdo TS, et al (2015) Chemical composition of essential oils from Annona vepretorum Mart. and Annona squamosa L. (Annonaceae) leaves and their antimalarial and trypanocidal activities. J Essent Oil Res 27:160–168. [CrossRef]
- Meira CS, Menezes LRA, dos Santos TB, et al (2017) Chemical composition and antiparasitic activity of essential oils from leaves of Guatteria friesiana and Guatteria pogonopus (Annonaceae). J Essent Oil Res 29:156–162. [CrossRef]
- Mendes FLR (2019) Avaliação da atividade antibacteriana de extratos de Eugenia brasiliensis. Fundação Oswaldo Cruz.
- Mendes FLR, Carvalho EM de, Abrantes JA, Nogueira JM da R (2020) Buscando novos antimicrobianos: avaliação da atividade antibacteriana de extratos de Eugenia brasiliensis. Rev Bras Análises Clínicas 52:. [CrossRef]
- Mendes R de F, Pinto N de CC, da Silva JM, et al (2017) The essential oil from the fruits of the Brazilian spice Xylopia sericea A. St.-Hil. presents expressive in-vitro antibacterial and antioxidant activity. J Pharm Pharmacol 69:341–348. [CrossRef]
- Menezes L, Costa C, Rodrigues A, et al (2016) Cytotoxic Alkaloids from the Stem of Xylopia laevigata. Molecules 21:890. [CrossRef]
- Menezes R, Sessions Z, Muratov E, et al (2021) Secondary metabolites extracted from annonaceae and chemotaxonomy study of terpenoids. J Braz Chem Soc 00:1–10. [CrossRef]
- Meng DH, Xu YP, Chen WL, et al (2007) Anti-tumour clerodane-type diterpenes from Mitrephora thorelii. J Asian Nat Prod Res 9:679–684. [CrossRef]
- Mikolajczak KL, McLaughlin JL, Rupprecht JK (1988) Control of pests with annonaceous acetogenins.
- Moghadamtousi S, Fadaeinasab M, Nikzad S, et al (2015a) Annona muricata (Annonaceae): A Review of Its Traditional Uses, Isolated Acetogenins and Biological Activities. Int J Mol Sci 16:15625–15658. [CrossRef]
- Moghadamtousi SZ, Fadaeinasab M, Nikzad S, et al (2015b) Annona muricata (Annonaceae): A review of its traditional uses, isolated acetogenins and biological activities. Int J Mol Sci 16:15625–15658. [CrossRef]
- Moghadamtousi SZ, Goh BH, Chan CK, et al (2013) Biological activities and phytochemicals of Swietenia macrophylla king. Molecules 18:10465–10483. [CrossRef]
- Moghadamtousi SZ, Rouhollahi E, Karimian H, et al (2014) Gastroprotective activity of Annona muricata leaves against ethanol-induced gastric injury in rats via Hsp70/Bax involvement. Drug Des Devel Ther 8:2099–2111. [CrossRef]
- Mohammadi S, Jafari B, Asgharian P, et al (2020) Medicinal plants used in the treatment of Malaria: A key emphasis to Artemisia, Cinchona, Cryptolepis, and Tabebuia genera. Phyther Res 1–14. [CrossRef]
- Mohd M, Alam KS, Mohd A, et al (2009) Antidaiabetic activity of the aqueous extract o Annona squamosa in streptozotocin induced-hyperglycemic rats. Pharma Res 2:59–63.
- Montenegro C de A, Lima GR de M, Gomes IF, et al (2014) Gastroprotective effect of Xylopia langsdorffiana A. St.-Hil. & Tul. (Annonaceae): Involvement of endogenous sulfhydryls compounds and nitric oxide. Rec Nat Prod 8:165–183.
- Montenegro H, Gutiørrez M, Romero LI, et al (2003) - G. amplifolia e G. dumetorum (2003) - Montenegro - Alcaloides - atividade antileishmania. 677–679.
- Monzon RB, Alvior JP, Luczon LL, et al (1994) Larvicidal potential of five Philippine plants against Aedes aegypti (Linnaeus) and Culex quinquefasciatus (Say). Southeast Asian J Trop Med Public Health 25:755–759.
- Morales CA, González R, Aragón R (2004) Evaluación de la actividad larvicida de extractos polares y no polares de acetogeninas de annona muricata sobre larvas de Aedes aegypti y Anopheles albimanus (Diptera: Culicidae). Rev Colomb Entomol 30:187–192.
- Moreira JLB, Carvalho CBM de, Frota cristiane C (2015) Visualização bacteriana e colorações. Fortaleza.
- Moshi MJ, Joseph CC, Innocent E, Nkunya MHH (2004) In vitro antibacterial and antifungal activities of extracts and compounds from Uvaria scheffleri. Pharm Biol 42:269–273. [CrossRef]
- Muganza DM, Fruth B, Nzunzu JL, et al (2016) In vitro antiprotozoal activity and cytotoxicity of extracts and isolated constituents from Greenwayodendron suaveolens. J Ethnopharmacol 193:510–516. [CrossRef]
- Musuyu Muganza D, Fruth BI, Nzunzu Lami J, et al (2015) In vitro antiprotozoal activity and cytotoxicity of extracts and fractions from the leaves, root bark and stem bark of Isolona hexaloba. J Ethnopharmacol 174:187–194. [CrossRef]
- Nascimento TL do, Vasconcelos SP, Peres Y, et al (2019) Prevalence of malaria relapse: Systematic review with meta-analysis. Rev Lat Am Enferm 18:e3111.
- Nathan C, Ding A (2010) Nonresolving Inflammation. Cell 140:871–882. [CrossRef]
- Ndumu PA, George JBD, Choudhury MK (1999) Toxicity of neem seed oil (Azadiracta indica) against the larvae ofAmblyomma variegatum a three-host tick in cattle. Phyther Res 13:532–534. [CrossRef]
- Neill JO (2016) Nature reviews. Drug Discov 15:526.
- Newman DJ, Cragg GM (2016) Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 79:629–661.
- Newman DJ, Cragg GM (2020) Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J Nat Prod 83:770–803. [CrossRef]
- Ngangoue MO, Ngameni B, Ambassa P, et al (2020) A phenanthridin-6(5H)-one derivative and a lanostane-type triterpene with antibacterial properties from Anonidium mannii (Oliv). Engl. & Diels (Annonaceae). Nat Prod Res 0:1–10. [CrossRef]
- Ngantchou I, Nkwengoua E, Nganso Y, et al (2009) Antitrypanosomal activity of polycarpol from Piptostigma preussi (Annonaceae). Fitoterapia 80:188–191. [CrossRef]
- Nghi NBT, Uyen TT, Anh HM, et al (2022) Rumdul (Sphaerocoryne affinis) Antioxidant Activity and Its Potential for Parkinson’s Disease Treatment. Oxid Med Cell Longev 2022:. [CrossRef]
- Ngouonpe AW, Mbobda ASW, Happi GM, et al (2019) Natural products from the medicinal plant Duguetia staudtii (Annonaceae). Biochem Syst Ecol 83:22–25. [CrossRef]
- Nguyen HT, Vu T-Y, Chandi V, et al (2020) Dual COX and 5-LOX inhibition by clerodane diterpenes from seeds of Polyalthia longifolia (Sonn.) Thwaites. Sci Rep 10:15965. [CrossRef]
- Nhiem NX, Hien NTT, Tai BH, et al (2015) New ent-kauranes from the fruits of Annona glabra and their inhibitory nitric oxide production in LPS-stimulated RAW264.7 macrophages. Bioorganic Med Chem Lett 25:254–258. [CrossRef]
- Nhiem NX, Tuong NT, Ky PT, et al (2017) Chemical Components from Phaeanthus vietnamensis and Their Inhibitory NO Production in BV2 Cells. Chem Biodivers 14:1–7. [CrossRef]
- Núcleo de estudos e Pesquisas de Produtos Naturais (2020) Atividade Antitumoral. Univ Fed Sergipe - UFS.
- Núñez-Sellés AJ (2005) Antioxidant therapy: Myth or reality? J Braz Chem Soc 16:699–710. [CrossRef]
- Nwokocha CR, Owu DU, Gordon A, et al (2012) Possible mechanisms of action of the hypotensive effect of Annona muricata (soursop) in normotensive Sprague–Dawley rats. Pharm Biol 50:1436–1441. [CrossRef]
- Nwosu RA, Suleiman MM, Makun HJ, et al (2022) Anthelmintic activity of methanol extract of Dennettia tripetala G. Baker (Annonaceae) fruits against Haemonchus contortus in red Sokoto goats. Trop Anim Health Prod 54:. [CrossRef]
- Obiri DD, Osafo N (2013) Aqueous ethanol extract of the fruit of Xylopia aethiopica (Annonaceae) exhibits anti-anaphylactic and anti-inflammatory actions in mice. J Ethnopharmacol 148:940–945. [CrossRef]
- Odalo JO, Omolo MO, Malebo H, et al (2005) Repellency of essential oils of some plants from the Kenyan coast against Anopheles gambiae. Acta Trop 95:210–218. [CrossRef]
- Ohashi M, Amoa-Bosompem M, Kwofie KD, et al (2018) In vitro antiprotozoan activity and mechanisms of action of selected G hanaian medicinal plants against Trypanosoma , Leishmania , and Plasmodium parasites. Phyther Res 32:1617–1630. [CrossRef]
- Okonkwo EU, Okoye WI (2001) Insecticidal activity of Dennettia tripetaia Baker f. and Piper guineense Schum. and Thonn. against Dermestes maculatus Degeer (Coleoptera: Dermestidae) and Necrobia rufipes Degeer (Coleoptera: Cleridae) on dried fish. Niger J Entomol 18:. [CrossRef]
- Okoye TC, Akah PA, Omeje EO, et al (2013) Anticonvulsant effect of kaurenoic acid isolated from the root bark of Annona senegalensis. Pharmacol Biochem Behav 109:38–43. [CrossRef]
- Okpekon TA, Dade JME, Say M V, et al (2015) Bingervone, an antiprotozoal beta-triketone derivative from the roots of Uvaria afzelii (Annonaceae). Int J Pharm Sci Res 6:4210–4215. [CrossRef]
- Olate VR, Pertino MW, Theoduloz C, et al (2012) New Gastroprotective Labdeneamides from. Planta Med 13:362–367.
- Oliveira ÂC de, Pereira MJB (2009) Efeito Antialimentar do Extrato Metanólico de Annona crassiflora Mart . sobre o Percevejo Marrom Euschistus heros ( Fabr . 1798 ) ( Heteroptera : Pentatomidae ). Rev Bras Agroecol 4:2633–2636.
- Oliveira de Souza LI, Bezzera-Silva PC, do Amaral Ferraz Navarro DM, et al (2017) The chemical composition and trypanocidal activity of volatile oils from Brazilian Caatinga plants. Biomed Pharmacother 96:1055–1064. [CrossRef]
- Oliveira ESC, Amaral ACF, Lima ES, Jefferson JR (2014) Chemical composition and biological activities of Bocageopsis multiflora essential oil. J Essent Oil Res 26:161–165. [CrossRef]
- Oliveira GLS (2015) Determinação da capacidade antioxidante de produtos naturais in vitro pelo método do dpph•: Estudo de revisão. Rev Bras Plantas Med 17:36–44. [CrossRef]
- Olivier DK, Van Vuuren SF, Moteetee AN (2015) Annickia affinis and A. chlorantha (Enantia chlorantha) - A review of two closely related medicinal plants from tropical Africa. J Ethnopharmacol 176:438–462. [CrossRef]
- Ortqvist A, Hedlund J, Kalin M (2005) Streptocuccus pneumonie: epidemiology, risk factors, and clinical features. Semin Respir Crit Care Med 26:563–574.
- Osafo N, Biney RP, Obiri DD (2016) Aqueous Ethanol Fruit Extract of Xylopia aethiopica and Xylopic Acid Exhibit Anti-inflammatory Activity Through Inhibition of the Arachidonic Acid Pathway. Pharm Biosci J 49–55. [CrossRef]
- Osafo N, Obiri DD, Danquah KO, et al (2019) Potential effects of xylopic acid on acetic acid-induced ulcerative colitis in rats. Turkish J Gastroenterol 30:732–744. [CrossRef]
- Osorio D EJ, Montoya P. GL, Muñoz D. K, Arango A. GJ (2006) Antiplasmodial Activity of Aporphinic Alkaloids from Rollinia pittieri and Pseudomalmea boyacana (Annonaceae). Vitae 13:49–54.
- Osorio E, Arango GJ, Jiménez N, et al (2007) Antiprotozoal and cytotoxic activities in vitro of Colombian Annonaceae. J Ethnopharmacol 111:630–635. [CrossRef]
- Otręba M, Kośmider L (2020) In vitro anticancer activity of fluphenazine, perphenazine and prochlorperazine. A review. J Appl Toxicol.
- Owusu FWA, Boakye-Gyasi M El, Entsie P, et al (2021) Formulation and Pharmaceutical Assessment of Annona muricata Oral Capsules and Suspension as Antidiarrhea Dosage Forms. J Chem 2021:6–11. [CrossRef]
- Oyemitan IA, Elusiyan CA, Akinkunmi EO, et al (2019) Memory enhancing, anticholinesterase and antimicrobial activities of β-phenylnitroethane and essential oil of Dennettia tripetala Baker f. J Ethnopharmacol 229:256–261. [CrossRef]
- Paarakh PM, Chansouria JPN, Khosa RL (2009) Wound healing activity of Annona muricata extract. J Pharm Res 2:404–406.
- Palazzo MC, Wright HL, Agius BR, et al (2009) Chemical Compositions and Biological Activities of Leaf Essential Oils of Six Species of Annonaceae from Monteverde, Costa Rica. Rec Nat Prod 3:153–160.
- Pan S-Y, Zhou S-Z, Gao S-H, et al (2013) New Perspectives on How to Discover Drugs from Herbal Medicines: CAM’s Outstanding Contribution to Modern Therapeutics. Evidence-Based Complement Altern Med 2013:1–25. [CrossRef]
- Paredes A, Hasegawa M, Prieto F, et al (2001) Biological activity of Guatteria cardoniana fractions. J Ethnopharmacol 78:129–132. [CrossRef]
- Pecoul B, Batista C, Stobbaerts E, et al (2016) The BENEFIT Trial: Where Do We Go from Here? PLoS Negl Trop Dis 10:e0004343. [CrossRef]
- Peña-Hidalgo M, Furtado LC, Costa-Lotufo L V., et al (2021) Alkaloids from the Leaves of Annona crassiflora and Their Cytotoxic Activity. Rev Bras Farmacogn 31:244–248. [CrossRef]
- Pereira F, Madureira AM, Sancha S, et al (2016) Cleistochlamys kirkii chemical constituents: Antibacterial activity and synergistic effects against resistant Staphylococcus aureus strains. J Ethnopharmacol 178:180–187. [CrossRef]
- Pérez-Pacheco R, Rodríguez Hernández C, Lara-Reyna J, et al (2004) Toxicidad de aceites, esencias y extractos vegetales en larvas de mosquito Culex quinquefasciatus Say (Diptera: Culicidae). Acta zoológica Mex 20:141–152.
- Piacente S, Aquino R, Tommasi N, et al (1994) Constituents of Weneria ciliolata and their in vitro anti-HIV activity. Phytochemistry 36:4–9.
- Pietta PG (2000) Flavonoids as antioxidants. J Nat Prod 63:1035–1042.
- Pinheiro MLB, Xavier CM, Souza ADL De, et al (2009) Acanthoic Acid and other Constituents from the Stem of. J Braz Chem Soc 20:1095–1102.
- Pinheiro R da S, Rabelo SV, de Oliveira AP, et al (2016) Phytochemical screening and evaluation of cytotoxicity of stem bark extracts of Anaxagorea dolichocarpa and Duguetia chrysocarpa (Annonaceae). Trop J Pharm Res 15:793–798. [CrossRef]
- Polbuppha I, Suthiphasilp V, Maneerat T, et al (2022) Nitric oxide production inhibitory activity of clerodane diterpenes from Monoon membranifolium. Nat Prod Res 36:2513–2517. [CrossRef]
- Popenoe W (1921) The native home of the cherimoya. J Hered 12:331–336.
- Pouofo Nguiam M, Njonte Wouamba SC, Longo F, et al (2021) Antibacterial and antishigellosis activity of Xylopia staudtii (engl. & diels), Annonaceae. J Ethnopharmacol 280:114406. [CrossRef]
- Prado A (2009) Composição fenólica e atividade antioxidante de frutas tropicais. Universidade de São Paulo.
- Prasad SK, Pradeep S, Shimavallu C, et al (2021) Evaluation of Annona muricata Acetogenins as Potential Anti-SARS-CoV-2 Agents Through Computational Approaches. Front Chem 8:1–7. [CrossRef]
- Pumiputavon K, Chaowasku T, Saenjum C, et al (2017) Cell cycle arrest and apoptosis induction by methanolic leaves extracts of four Annonaceae plants. BMC Complement Altern Med 17:1–11. [CrossRef]
- Qi L-W, Liu E-H, Chu C, et al (2010) Anti-Diabetic Agents from Natural Products — An Update from 2004 to 2009. Curr Top Med Chem 10:434–457. [CrossRef]
- Quintans JSS, Soares B, Ferraz RC, et al (2013) Chemical constituents and anticancer effects of the essential oil from leaves of xylopia laevigata. Planta Med 79:123–130. [CrossRef]
- Rabelo D de M, Pinheiro MLB, Barison A, et al (2014) Isoquinoline alkaloids and investigation of the antibacterial and antiplasmodial activities of Guatteria citriodora (Annonaceae). Quim Nova 37:1453–1458. [CrossRef]
- Rabêlo SV (2014) Revisão de alcalóides do gênero Annona, estudo fitoquímico e avaliação da atividade biológica de Atemoia (Annona cherimola x Annona squamosa ). Universidade Federal do Vale do São Francisco.
- Rabêlo SV, Da Costa MM, Libório RC, Almeida JRGDS (2014) Antioxidant and antimicrobial activity of extracts from atemoia (Annona cherimola Mill. x A. squamosa L.). Rev Bras Frutic 36:265–271. [CrossRef]
- Rady I, Bloch MB, Chamcheu RCN, et al (2018) Anticancer Properties of Graviola (Annona muricata): A Comprehensive Mechanistic Review. Oxid Med Cell Longev 2018:. [CrossRef]
- Rajca Ferreira AK, Lourenço FR, Young MCM, et al (2018) Chemical composition and biological activities of Guatteria elliptica R. E. Fries (Annonaceae) essential oils. J Essent Oil Res 30:69–76. [CrossRef]
- Rao NS, Sharma K, Sharma RK (2005) Anti-feedant and growth inhibitory effects of seed extracts of custard apple, Annona squamosa against Khapra Beetle, Trogoderma granarium. J Agric Technol 1:43–54.
- Rejón-Orantes J, González-Esquinca A, de la Mora M, et al (2011) Annomontine, an Alkaloid Isolated from Annona purpurea , Has Anxiolytic-Like Effects in the Elevated Plus-Maze. Planta Med 77:322–327. [CrossRef]
- Rester U (2008) Fom virtually to reality - Virtual screening in lead discovery and lead optimization: a medicinal chemistry perspective. Curr Opin Drug Discov Devel 11:559–568.
- Ribeiro JEL da S, Hopkins MJG, Vicentini A, et al (1999) Flora da reserva Ducke - Guia de Identificação das plantas vasculares de uma floresta de terra-firme na Amazónia Central. INPA-DFID.
- Ribeiro L do P, Vendramim JD, Bicalho KU, et al (2013) Annona mucosa Jacq. (Annonaceae): A promising source of bioactive compounds against Sitophilus zeamais Mots. (Coleoptera: Curculionidae). J Stored Prod Res 55:6–14. [CrossRef]
- Richomme P, Godet MC, Foussard F, et al (1991) A novel leishmanicidal labdane from Polyalthia macropoda. Planta Med 57:552–554. [CrossRef]
- Rinaldi MVN, Díaz IEC, Suffredini IB, Moreno PRH (2017) Alkaloids and biological activity of beribá (Annona hypoglauca). Rev Bras Farmacogn 27:77–83. [CrossRef]
- Riss TL, Moravec RA (2004) Use of multiple assay endpoints to investigate the effects of incubation time, dose of toxin, and plating density in cell-based cytotoxicity assays. Assay Drug Dev Technol 2:51–62.
- Rocha RS, Kassuya CAL, Formagio ASN, et al (2016) Analysis of the anti-inflammatory and chemopreventive potential and description of the antimutagenic mode of action of the Annona crassiflora methanolic extract. Pharm Biol 54:35–47. [CrossRef]
- Rodrigues AM, da Silva AA, de Freitas JCC, et al (2021) Larvicidal activity of Annona mucosa Jacq. extract and main constituents rolliniastatin 1 and rollinicin against Aedes aegypti and Aedes albopictus. Ind Crops Prod 169:. [CrossRef]
- Rodrigues AMS, De Paula JE, Degallier N, et al (2006) Larvicidal activity of some Cerrado plant extracts against Aedes aegypti. J Am Mosq Control Assoc 22:314–317. [CrossRef]
- Rogero SO, Lugão AB, Ikeda TI, Cruz ÁS (2003) Teste in vitro de citotoxicidade: estudo comparativo entre duas metodologias. Mater Res 6:317–320. [CrossRef]
- Rojano BA, GAVIRIA M CA, SÁEZ V JA, et al (2007) Berenjenol isolated from Oxandra cf xylopioides (Annonaceae) as insecticide. Vitae 14:95–100.
- Sacchetti G, Maietti S, Muzzoli M, et al (2005) Comparative evaluation of 11 essential oils of different origin as functional antioxidants, antiradicals and antimicrobials in foods. Food Chem 91:621–632. [CrossRef]
- Saepou S, Pohmakotr M, Reutrakul V, et al (2010) Anti-HIV1 Diterpenoids from Leaves and Twigs of Polyalthia sclerophylla. Planta Med 76:721–725. [CrossRef]
- Sahu M, Sahoo NK, Alagarsamy V, Rath BP (2016) Comparative Evaluation of Antidiabetic and Antioxidant Activities of Aqueous Fruit Peel and Leaf Extracts of Annona Squamosa on High Fat Diet and Multiple Low Dose Streptozotocin Mouse Model of Diabetes. Austin J Pharmacol Ther 4:1081.
- Saito ML, Pott A, Ferraz JMG, Nascimento R dos S (2004) Avaliação de plantas com atividade deterrente alimentar em Spodoptera frugiperda (J.E.SMITH) E Anticarsia gemmatalis HUBNER. Pestic Rev Ecotoxicologia e Meio Ambient 14:1–10. [CrossRef]
- Saldanha AA, Vieira L, de Oliveira FM, et al (2020) Anti-inflammatory and central and peripheral anti-nociceptive activities of α-asarone through the inhibition of TNF-α production, leukocyte recruitment and iNOS expression, and participation of the adenosinergic and opioidergic systems. Inflammopharmacology 28:1039–1052. [CrossRef]
- Saldanha AA, Vieira L, Maia DS da S, et al (2021) Anti-inflammatory and antinociceptive activities of a phenylpropanoid-enriched fraction of Duguetia furfuracea. Inflammopharmacology 29:409–422. [CrossRef]
- Saldanha AA, Vieira L, Ribeiro RIM de A, et al (2019) Chemical composition and evaluation of the anti-inflammatory and antinociceptive activities of Duguetia furfuracea essential oil: Effect on edema, leukocyte recruitment, tumor necrosis factor alpha production, iNOS expression, and adenosinergic and opioid. J Ethnopharmacol 231:325–336. [CrossRef]
- Sales PA, Molina I, Murta SMF, et al (2017) Experimental and Clinical Treatment of Chagas Disease: A Review. Am J Trop Med Hyg 97:1289–1303. [CrossRef]
- Samwel S, Odalo JO, Nkunya MHH, et al (2011) Toussaintines A-E: Antimicrobial indolidinoids, a cinnamoylhydrobenzofuranoid and a cinnamoylcyclohexenoid from Toussaintia orientalis leaves. Phytochemistry 72:1826–1832. [CrossRef]
- Santos AR, Benghi TGS, Nepel A, et al (2017) In vitro Antiproliferative and Antibacterial Activities of Essential Oils from Four Species of Guatteria. Chem Biodivers 14:. [CrossRef]
- Santos F, Magalhães-Junior JT, Carneiro I de O, et al (2022) Eco-epidemiology of vectorial Trypanosoma cruzi transmission in a region of northeast Brazil. Acta Trop 225:106184. [CrossRef]
- Santos L de Á, Cavalheiro AJ, Tempone AG, et al (2015) Antitrypanosomal acetylene fatty acid derivatives from the seeds of Porcelia macrocarpa (Annonaceae). Molecules 20:8168–8180. [CrossRef]
- Santos MRA, Lima RA, Silva AG, et al (2010) Composição química e atividade inseticida do extrato acetônico de Piper alatabaccum Trel&Yuncker (Piperaceae) sobre Hypothenemus hampei Ferrari. Rev Bras Plantas Med 15:332–336.
- Santos SS, Araújo RV, Giarolla J, et al (2020) Searching for drugs for Chagas disease, leishmaniasis and schistosomiasis: a review. Int J Antimicrob Angents 55:105906. [CrossRef]
- Saxena RC, Harshan V, Saxena A, et al (1993) Larvicidal and chemosterilant activity of Annona squamosa alkaloids against Anopheles stephensi. Journal-American Mosq Control Assoc 9:84.
- Schmeda-Hirschmann G, de Arias AR (1992) A screening method for natural products on triatomine bugs. Phyther Res 6:68–73.
- Seangphakdee P, Pompimon W, Meepowpan P, et al (2013) Anti-inflammatory and anticancer activities of (−)-zeylenol from stems of Uvaria grandiflora. ScienceAsia 39:610. [CrossRef]
- Sedgwick AD, Lees P (1986) A comparison of air pouch, sponge and pleurisy models of acute carrageenan inflammation in the rat. Agents Actions 18:439–446. [CrossRef]
- Serhan CN (2010) Novel Lipid Mediators and Resolution Mechanisms in Acute Inflammation. Am J Pathol 177:1576–1591. [CrossRef]
- Sesang W, Punyanitya S, Pitchuanchom S, et al (2014) Cytotoxic Aporphine Alkaloids from Leaves and Twigs of Pseuduvaria trimera (Craib). Molecules 19:8762–8772. [CrossRef]
- Shanker KS, Kanjilal S, Rao BVSK, et al (2007) Isolation and antimicrobial evaluation of isomeric hydroxy ketones in leaf cuticular waxes of annona squamosa. Phytochem Anal 18:7–12. [CrossRef]
- Sharifi-Rad M, Roberts TH, Matthews KR, et al (2018) Ethnobotany of the genus Taraxacum—Phytochemicals and antimicrobial activity. Phyther. Res. 32:2131–2145.
- Sharma PP, Pardeshi AB, Vijigiri D (2011) Bioactivity of some medicinal plant extracts against Musca domestica L. J Ecobiotechnology.
- Sheen E, Triadafilopoulos G (2011) Adverse Effects of Long-Term Proton Pump Inhibitor Therapy. Dig Dis Sci 56:931–950. [CrossRef]
- Sherwood ER, Toliver-Kinsky T (2004) Mechanisms of the inflammatory response. Best Pract Res Clin Anaesthesiol 18:385–405. [CrossRef]
- Shirwaikar A, Rajendran K, Dinesh Kumar C, Bodla R (2004) Antidiabetic activity of aqueous leaf extract of Annona squamosa in streptozotocin–nicotinamide type 2 diabetic rats. J Ethnopharmacol 91:171–175. [CrossRef]
- Sidrim JJC, Rocha MFG (2004) Aspectos gerais de fungos filamentosos e dimórficos na apresentação filamentosa. In: Editora Guanabara Koogan (ed) Micologia médica à luz de autores com=ntemporâneos, 1st editio. Rio de Janeiro - RJ, pp 8–88.
- Silprakob W, Sukhamsri N, Kuhakarn C, et al (2018) A new oxoaporphine alkaloid from the root of dasymaschalon glaucum. Nat Prod Commun 13:1471–1474. [CrossRef]
- Silva AG, Rocha LC, Canniatti Brazaca SG (2009) Caracterização físico-química, digestibilidade proteica e atividade antioxidante de feijão (Phaseolus vulgaris L.). Aliment e Nutr 20:591–598.
- Silva CA, Domingues Neta AM (2011) Aspectos reprodutivos e visitantes florais de Duguetia marcgraviana Mart. (Annonaceae) na região sudoeste de Mato Grosso. Biotemas 23:69–76. [CrossRef]
- Silva LA da, Raposo JDA, Campos LPG, et al (2018) Atividade antioxidante do óleo essencial de Myrcia sylvatica (G. Mey.) DC. por diferentes métodos de análises antioxidantes (ABTS, DPPH, FRAP, β-caroteno/ácido linoleico). Rev Fitos 12:117–126. [CrossRef]
- Silva ITSS, Fernandes MJB, Oliveira RA, et al (2016) Annona squamosa L. (Annonaceae): Chemical bioprospection and biological activity in two phenological stages. Appl Ecol Environ Res 14:133–147. [CrossRef]
- Silva JC, Araújo C de S, de Lima-Saraiva SRG, et al (2015) Antinociceptive and anti-inflammatory activities of the ethanolic extract of Annona vepretorum Mart. (Annonaceae) in rodents. BMC Complement Altern Med 15:197. [CrossRef]
- Silva MO (2015) Atividade antioxidante e composição de oligossacarídeos em subproduto obtido do processamento industrial da goiaba (Psidium guajava). Universidade Estadual de Campinas.
- Silva NL, Saldanha AA, Silva DB, et al (2019) Anti-inflammatory, antinociceptive and antioxidant activities of the hydromethanolic fraction from Annona nutans leaves. Biosci J 35:1599–1613. [CrossRef]
- Simões CM de O, Schenkel EP, Gosmann G, et al (2003) Farmacognocia: da planta ao medicamento, Fifth edit. UFRGS editora.
- Simões S, Lopes R, Campos MCD, et al (2019) Animal models of acute gastric mucosal injury: Macroscopic and microscopic evaluation. Anim Model Exp Med 2:ame2.12060. [CrossRef]
- Sinchaisri N, Roongsook D, Chungsamarnyart N (1991) Insecticidal Activity of plant crude-extracts on Diamondback Moth Larvae. Kasetsart J. (Nat. Sci. Suppl.) 25:106–110.
- Siqueira CAT, Oliani J, Sartoratto A, et al (2011) Chemical constituents of the volatile oil from leaves of Annona coriacea and in vitro antiprotozoal activity. Rev Bras Farmacogn 21:33–40. [CrossRef]
- Siqueira CAT, Serain AF, Pascoal ACRF, et al (2015) Bioactivity and chemical composition of the essential oil from the leaves of Guatteria australis A.St.-Hil. Nat Prod Res 29:1966–1969. [CrossRef]
- Soerio M de NC (2022) Perspectives for a new drug candidate for Chagas disease therapy. Mem Inst Oswaldo Cruz 117:e220004. [CrossRef]
- Somdej Kanokmedhakul *, Kwanjai Kanokmedhakul, Daungrudee Yodbuddee and, Phonkerd N (2003) New Antimalarial Bis-dehydroaporphine Alkaloids from Polyalthia debilis†. [CrossRef]
- Son NT, Le TA, Thuy DTT, et al (2021) Essential Oils of the Leaf and Stem of Polyalthia viridis Craib and Their Biological Activities. Nat. Prod. Commun. 16.
- Song CM, Lim SJ, Tong JC (2009) Recnt advances in computer-aided drug design. Brief Bioinform 10:579–591.
- Sousa MSB, VIeira LM, Lima A (2011) Fenólicos totais e capacidade antioxidante in vitro de resíduos de polpas de frutas tropicais. Brazilian J Food Technol 14:202–210.
- Sousa O V., Del-Vechio-Vieira G, Alves MS, et al (2012) Chemical composition and biological activities of the essential oils from Duguetia lanceolata St. Hil. barks. Molecules 17:11056–11066. [CrossRef]
- Souza, E. M.; Cordeiro, J. R.; Pereira MJB (2007) Avaliação da atividade inseticida dos diferentes extratos das sementes de Annona coriaceae sobre Dichelops melacanthus (Dallas, 1851). In: Resumos do V CBA - Manejo de agroecossistemas sustentáveis. Rev Bras Agroecol 2:1107–1110.
- Spletozer AG, Santos CR dos, Sanches LA, Garlet J (2021) Plantas com potencial inseticida: enfoque em espécies amazônicas. Ciência Florest 31:974–997. [CrossRef]
- Sreeletha C, Geetha PR (2012) Pesticidal effects of Annona squamosa L. on male Oryctes rhinoceros Linn.(Coleoptera: Scarabaeidae) in relation to reproduction. Curr Biot 6:8–21.
- Suedee A, Mondranondra IO, Kijjoa A, et al (2007) Constituents of Polyalthia jucunda and their cytotoxic effect on human cancer cell lines. Pharm Biol 45:575–579. [CrossRef]
- Suthiphasilp V, Maneerat T, Andersen RJ, et al (2021) α-Glucosidase inhibitory activity of compounds isolated from the twig and leaf extracts of Desmos dumosus. Heliyon 7:e06180. [CrossRef]
- Taha H, Arya A, Khan AK, et al (2018) Effect of Pseuduvaria macrophylla in attenuating hyperglycemia mediated oxidative stress and inflammatory response in STZ-nicotinamide induced diabetic rats by upregulating insulin secretion and glucose transporter-1, 2 and 4 proteins expression. J Appl Biomed 16:263–273. [CrossRef]
- Talapko J, Škrlec I, Alebić T, et al (2019) Malaria: The past and the present. Microorganisms 7:. [CrossRef]
- Tamfu AN, Tagatsing Fotsing M, Talla E, et al (2021) Bioactive constituents from seeds of Annona Senegalensis Persoon (Annonaceae). Nat Prod Res 35:1746–1751. [CrossRef]
- Tan LTH, Lee LH, Yin WF, et al (2015) Traditional Uses, Phytochemistry, and Bioactivities of Cananga odorata (Ylang-Ylang). Evidence-Based Complement Altern Med 2015:1–30. [CrossRef]
- Tavares JF, Queiroga KF, Silva MVB, et al (2006) ent-trachylobane diterpenoids from Xylopia langsdorffiana. J Nat Prod 69:960–962. [CrossRef]
- Tekuri S, Pasupuleti S, Konidala K, Pabbaraju N (2019) Pharmacological Effects of Polyalthia cerasoides (Roxb.) Bedd.: a brief Review. J Complement Med Res 10:38. [CrossRef]
- Teo SP, Bhakta S, Stapleton P, Gibbons S (2020) Bioactive compounds from the bornean endemic plant goniothalamus longistipetes. Antibiotics 9:1–11. [CrossRef]
- Tolosa D, Colom OA, Bardón A, Neske A (2012) Insecticidal effects of acetogenins from Rollinia occidentalis seed extract. Nat Prod Commun 7:1934578X1200701226.
- Tomasz A (1997) Antibiotic resistance in Streptococcus pneumonie. Clin Infect Dis 24:S85-88.
- Toto Blessing L Di, Colom OÁ, Popich S, et al (2010) Antifeedant and toxic effects of acetogenins from Annona montana on Spodoptera frugiperda. J Pest Sci (2004) 83:307–310. [CrossRef]
- Trieu QH, Pham VC, Retailleau P, et al (2021) Rare flavonoids and sesquiterpenoids isolated from the leaves of Goniothalamus gracilipes. Fitoterapia 155:105034. [CrossRef]
- Tsiang F, Li PT, Huong LT, et al (2022) Annonaceae Essential Oils : Antimicrobial and Compositions of the , Nguyen Thanh Chung Do Ngoc Dai. 4:387–392.
- Tuchinda P, Munyoo B, Pohmakotr M, et al (2006) Cytotoxic styryl-lactones from the leaves and twigs of Polyalthia crassa. J Nat Prod 69:1728–1733. [CrossRef]
- Tulassay Z, Herszényi L (2010) Gastric mucosal defense and cytoprotection. Best Pract Res Clin Gastroenterol 24:99–108. [CrossRef]
- Twilley D, Rademan S, Lall N (2020) A review on traditionally used South African medicinal plants, their secondary metabolites and their potential development into anticancer agents. J Ethnopharmacol 113101.
- Umeotok SBA, Isah MD, Ukeh JA, Udo IA (2013) Evaluation of the insecticidal and deterrence properties of pepper fruit, Dennettia tripetala (G. Baker) and Ginger Zingiber officinale Roscoe against maize weevil Sitophilus zeamais (Motsch.). J Biol Agric Healthc 3:76–82.
- Valencia L, Muñoz DL, Robledo SM, et al (2011) Trypanocidal and cytotoxic activity of extracts of Colombian plants. Biomedica 31:552–559. [CrossRef]
- Vannier-Santos MA, Brunoro G V., Soeiro M de NC, et al (2019) Parasite, Compartments, and Molecules: Trick versus Treatment on Chagas Disease. In: Biology of Trypanosoma cruzi , 1st edition. IntechOpen.
- Vendramini-Costa DB, Monteiro KM, Iwamoto LH, et al (2014) Gastroprotective effects of goniothalamin against ethanol and indomethacin-induced gastric lesions in rats: Role of prostaglandins, nitric oxide and sulfhydryl compounds. Chem Biol Interact 224:206–212. [CrossRef]
- Viegas C (2003) Terpenes with insecticidal activity: An alternative to chemical control of insects. Quim Nova 26:390–400. [CrossRef]
- Viegas Júnior C (2003) Terpenos com atividade inseticida: uma alternativa para o controle químico de insetos. Quim Nova 26:390–400.
- Vila-Nova NS, de Morais SM, Falcão MJC, et al (2011) Atividade leishmanicida e citotoxicidade de constituintes quimicos de duas especies de annonaceae cultivadas no Nordeste do Brasil. Rev Soc Bras Med Trop 44:567–571. [CrossRef]
- Volobuff CRF, Pederiva MMC, Benites RSR, et al (2019) Bioguided Fractionation, and Antioxidant, Antiproliferative, and Anti-Inflammatory Activity of Annona cacans Warm. J Med Food 22:1078–1086. [CrossRef]
- Vuk I, Rajic Z, Zorc B (2008) Malaria and antimalarial drugs. Farm Glas 64:51–60.
- Waechter AI, Cavé A, Hocquemiller R, et al (1999) Antiprotozoal activity of aporphine alkaloids isolated from Unonopsis buchtienii (Annonaceae). Phyther Res 13:175–177. [CrossRef]
- Waechter AI, Ferreira ME, Fournet A, et al (1997) Experimental treatment of cutaneous leishsmaniasis with argentilactone isolated from annona haematantha. Planta Med 63:433–435. [CrossRef]
- Waechter AI, Yaluff G, Inchausti A, et al (1998) Leishmanicidal and trypanocidal activities of acetogenins isolated from Annona glauca. Phyther Res 12:541–544. [CrossRef]
- Wafo P, Nyasse B, Fontaine C (1999) A 7,8-dihydro-8-hydroxypalmatine from Enantia chlorantha. Phytochemistry 50:279–281. [CrossRef]
- Walker N, Nadjm B, Whitty C (2017) Malaria. Medicine (Baltimore) 42:52–58.
- Wang B, Yang Z-F, Zhao Y-L, et al (2018) Anti-Inflammatory Isoquinoline with Bis- seco -aporphine Skeleton from Dactylicapnos scandens. Org Lett 20:1647–1650. [CrossRef]
- Wang B, Zhao Y-J, Zhao Y-L, et al (2020) Exploring Aporphine as Anti-inflammatory and Analgesic Lead from Dactylicapnos scandens. Org Lett 22:257–260. [CrossRef]
- Weber C, Noels H (2011) Atherosclerosis: current pathogenesis and therapeutic options. Nat Med 17:1410–1422. [CrossRef]
- White NJN, Pukrittayakamee S, Hien TTT, et al (2014) Malaria. Lancet 383:723–735.
- Williams RB, Hu JF, Olson KM, et al (2010) Antibiotic indole sesquiterpene alkaloid from Greenwayodendron suaveolens with a new natural product framework. J Nat Prod 73:1008–1011. [CrossRef]
- Wilson A, Trumpp A (2006) Bone-marrow haematopoietic-stem-cell niches. Nat Rev Immunol 6:93–106. [CrossRef]
- Woguem V, Fogang HPD, Maggi F, et al (2014) Volatile oil from striped African pepper (Xylopia parviflora, Annonaceae) possesses notable chemopreventive, anti-inflammatory and antimicrobial potential. Food Chem 149:183–189. [CrossRef]
- Wongsomboon P, Rattanajak R, Kamchonwongpaisan S, et al (2021) Unique polyacetylenic ester-neolignan derivatives from Mitrephora tomentosa and their antimalarial activities. Phytochemistry 183:112615. [CrossRef]
- Woo MH, Kim DH, McLaughlin JL (1999) Asitrilobins A and B: Cytotoxic mono-THF annonaceous acetogenins from the seeds of Asimina triloba. Phytochemistry 50:1033–1040. [CrossRef]
- Woolf A, Rose R (2020) Gastric Ulcer - StatPearls - NCBI Bookshelf. In: StatPearls. https://www.ncbi.nlm.nih.gov/books/NBK537128/. Accessed 22 Jul 2020.
- World Health Organization (2021) Chagas disease (also known as American trypanosomiasis). https://www.who.int/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis). Accessed 16 Feb 2022.
- World Health Organization (WHO) (2014) Antimicrobial resistance. Global Report on Surveillance. In: Bulletin of the World Health Organization. pp 383–384.
- World Health Organization (WHO) (2020) Lack of new antibiotics threatens global efforts to contain drug-resistant infections. Geneva.
- World Health Organization (WHO) (2019) World malaria report 2019.
- Wu JH, Wang XH, Yi YH, Lee KH (2003) Anti-AIDS agents 54. A potent anti-HIV chalcone and flavonoids from genus Desmos. Bioorganic Med Chem Lett 13:1813–1815. [CrossRef]
- X W, Q H, N X, et al (2018) Antioxidative and Anti-Inflammatory Effects of Water Extract of Acrostichum aureum Linn. against Ethanol-Induced Gastric Ulcer in Rats. Evid Based Complement Alternat Med 2018:. [CrossRef]
- Xavier MN, Alves CCF, Cazal C de M, Santos NH (2016) Chemical composition of the volatile oil of Cardiopetalum calophyllum collected in the Cerrado area. Ciência Rural 46:937–942. [CrossRef]
- Yakubu MT, Fayemo HT (2021) Anti-hyperprolactinemic activities of aqueous extract of Uvaria chamae (P. Beauv) roots and associated biochemical changes in chlorpromazine-induced hyperprolactinemic female Wistar rats. J Ethnopharmacol 271:113863. [CrossRef]
- Yang-Chang Wu *,†, Yu-Chun Hung †, Fang-Rong Chang †, et al (1996) Identification of ent-16β,17-Dihydroxykauran-19-oic Acid as an Anti-HIV Principle and Isolation of the New Diterpenoids Annosquamosins A and B from Annona squamosa. [CrossRef]
- Yang YL, Chang FR, Wu CC, et al (2002) New ent-kaurane diterpenoids with anti-platelet aggregation activity from Annona squamosa. J Nat Prod 65:1462–1467. [CrossRef]
- Yang Z, Lu W, Ma X, Song D (2012) Bioassay-guided isolation of an alkaloid with antiangiogenic and antitumor activities from the extract of Fissistigma cavaleriei root. Phytomedicine 19:301–305. [CrossRef]
- Yao LJ, Jalil J, Attiq A, et al (2019) The medicinal uses, toxicities and anti-inflammatory activity of Polyalthia species (Annonaceae). J Ethnopharmacol 229:303–325. [CrossRef]
- Yasmen N, Aziz MA, Tajmim A, et al (2018) Analgesic and Anti-Inflammatory Activities of Diethyl Ether and n-Hexane Extract of Polyalthia suberosa Leaves. Evidence-Based Complement Altern Med 2018:1–8. [CrossRef]
- Yeung C, Mendoza I, Echeverria LE, Baranchuk A (2021) Chagas’ cardiomyopathy and Lyme carditis: Lessons learned from two infectious diseases affecting the heart. Trends Cardiovasc Med 31:233–239. [CrossRef]
- Yu Z, Han C, Song X, et al (2019) Bioactive aporphine alkaloids from the stems of Dasymaschalon rostratum. Bioorg Chem 90:103069. [CrossRef]
- Zaridah MZ, Azah MAN, Rohani A (2006) Mosquitocidal activities of Malaysian plants. J Trop For Sci 74–80.
- Zgoda-Pols JR, Freyer AJ, Killmer LB, Porter JR (2002) Antimicrobial diterpenes from the stem bark of Mitrephora celebica. Fitoterapia 73:434–438. [CrossRef]
- Zhang Y, Li H, Zhang J, et al (2019) The combinatory effects of natural products and chemotherapy drugs and their mechanisms in breast cancer treatment. Phytochem Rev 1–19.
- Zhou D, Yang Q, Tian T, et al (2020) Gastroprotective effect of gallic acid against ethanol-induced gastric ulcer in rats: Involvement of the Nrf2/HO-1 signaling and anti-apoptosis role. Biomed Pharmacother 126:110075. [CrossRef]
Figure 1.
Venn diagram of the subfamilies of species with biological activity reported by the studies covered in this review.
Figure 1.
Venn diagram of the subfamilies of species with biological activity reported by the studies covered in this review.
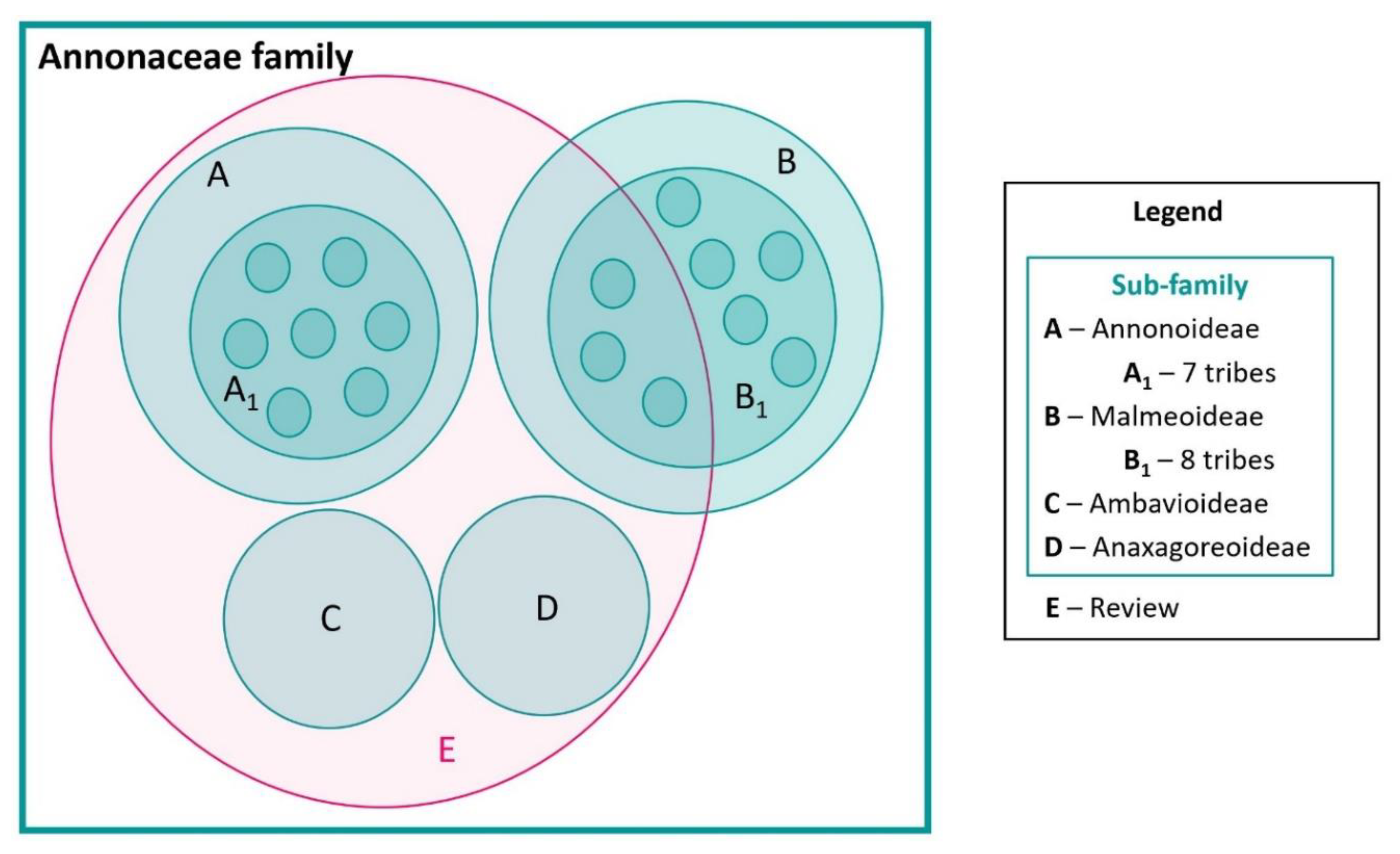
Graph 1.
Distribution of papers analyzed in this review over time, based on their year of publication.
Graph 1.
Distribution of papers analyzed in this review over time, based on their year of publication.
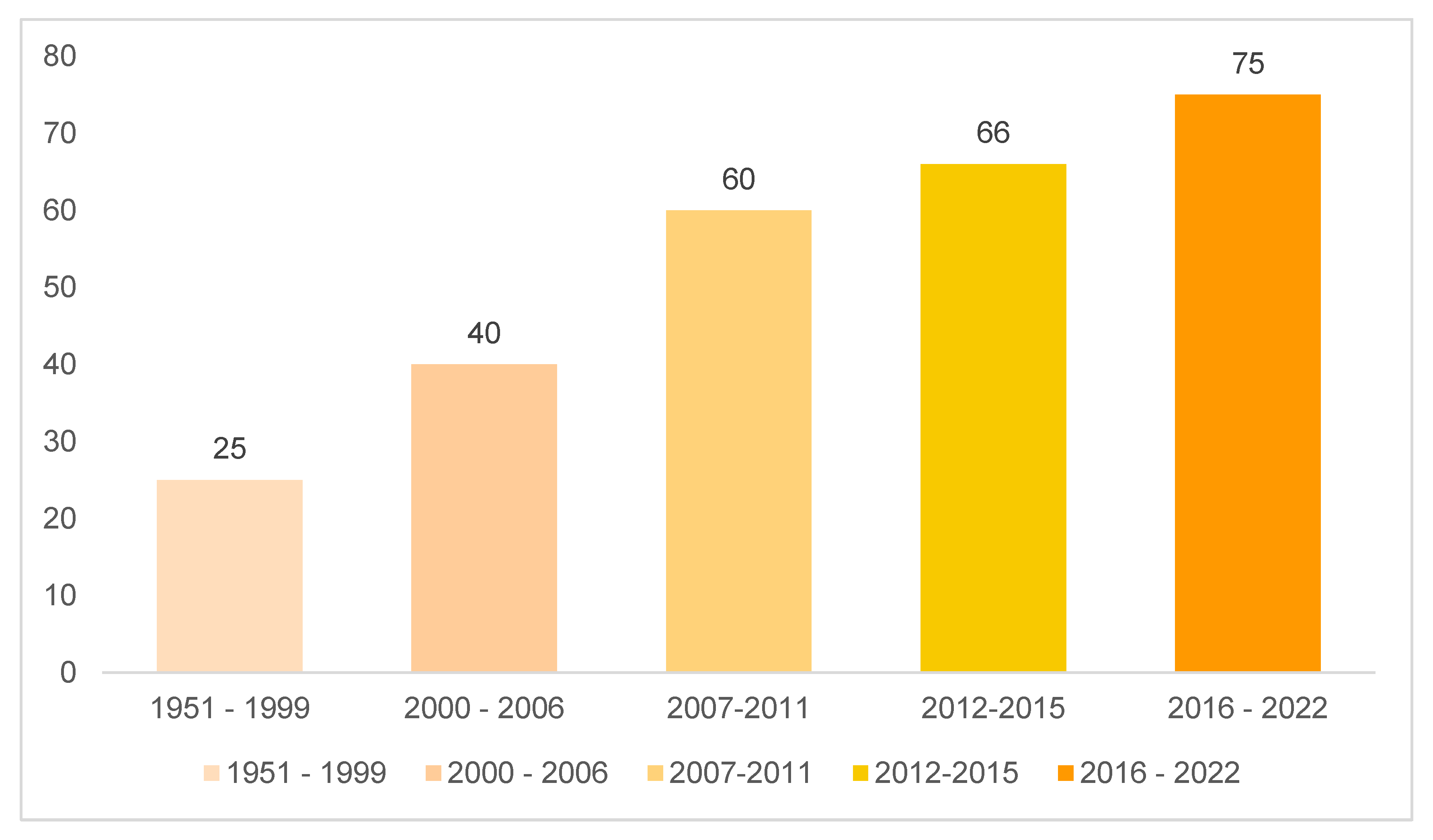
Graph 2.
Number of articles collected for each biological activity already studied for the Annonaceae.
Graph 2.
Number of articles collected for each biological activity already studied for the Annonaceae.
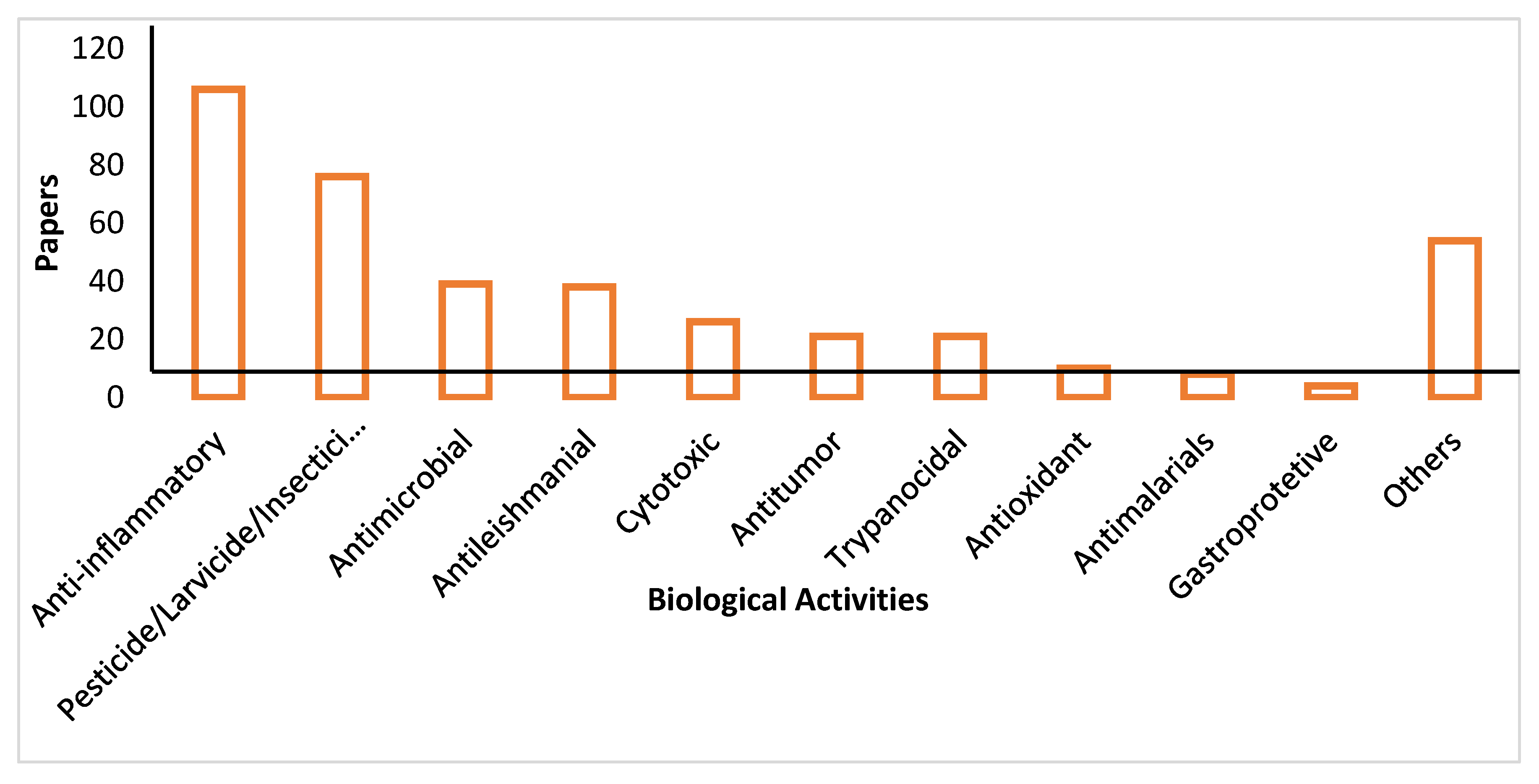
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



