
Submitted:
06 November 2025
Posted:
07 November 2025
You are already at the latest version
Abstract
Antimicrobial resistance remains one of the most urgent challenges in modern medicine, demanding innovative research tools for understanding and combating bacterial adaptation. Microfluidic technologies enable precise control over experimental conditions, single-cell resolution and high-throughput analysis, offering unique advantages over traditional microbiological methods. This review summarizes recent (2020–2025) developments in the application of microfluidics to antibiotic resistance research, emphasizing approaches used in fundamental studies rather than diagnostic implementations. The discussed technologies are grouped according to their primary research focus: (i) microfluidic cultivation and screening of antibiotic-producing microorganisms; (ii) tools for antibiotic screening and mechanistic studies and (iii) models for studying microbial stress responses and resistance development. Collectively, these approaches provide unprecedented insight into antibiotic action, resistance evolution and microbial physiology. Continued development and integration of microfluidics with complementary analytical tools will further accelerate the discovery of novel antimicrobials and the rational design of combination therapies, ultimately bridging the gap between fundamental microbiology and translational applications in antimicrobial resistance research.
Keywords:
antibiotic resistance
; microfluidics
; drug screening
; single-cell imaging
1. Introduction
Microfluidic technologies have an extremely wide range of applications and have revolutionized many fields: organic [1,2] and nanoparticle [3,4] synthesis, drug discovery [5,6,7] and delivery [8,9], production of RNA- and DNA-loaded lipid nanoparticles for gene delivery [10,11,12,13], reproductive technologies [14,15], live-cell delivery [16], CAR-T cell immunotherapy [17], cell separation [18] and other optical sorting [19], environmental analysis [20], aging and rejuvenation studies [21], biomarker diagnostics [22], food quality analysis [23,24] and biosensing [25,26], organ-on-chip [27,28] and 3D cell culture [29,30] models, single-cell analysis [31,32,33] and cell nanoencapsulation [34], protein screening [35], protein interaction studies [36] and many others.
Antibiotic resistance is recognized as an increasing global threat for public health [37,38,39,40,41]. Naturally, this huge problem increasingly attracting power of various areas of microfluidic technologies, especially a breakthrough droplet ones. The related areas of microfluidics have been extensively reviewed previously. Specific review articles in past five years are devoted to droplet microfluidics [31,32,42,43,44,45,46,47,48] and single-cell analysis [49,50,51,52], specific applications, including antimicrobial susceptibility testing and resistance development studies [53,54], bacterial patterning [55], synthetic microbial communities construction [56,57,58,59,60], biofilm investigation [61], environmental studies [62,63,64].
The objective of this review is to provide a concise yet comprehensive overview of recent (2020-2025) applications of microfluidic technologies that contribute to addressing the pressing issue of antimicrobial resistance. By summarizing key experimental designs across different application areas, this work aims to serve as a valuable foundation for future research in the field. The review focuses primarily on microfluidic approaches used in fundamental studies of antibiotic resistance. Newly developed technologies, as well as successful examples of the adaptation or refinement of existing methods, are organized into three major sections according to their principal research focus. The first section highlights studies on microbial strains that produce antimicrobial compounds, emphasizing bioactive molecules and antibiotics themselves. The second section summarizes microfluidic approaches for antibiotic screening and mechanistic investigations. Finally, as overcoming resistance and identifying novel targets depends on a detailed understanding of microbial physiology, the third section reviews methods for studying the emergence of resistance and the stress responses of model microorganisms.
The widespread use of antibiotics in the food and agricultural industries significantly contributes to the spread of antibiotic resistance, leading to the accumulation of antibiotics in the environment and creating evolutionary pressure for the selection of resistant isolates. Microfluidic technologies are used to detect antibiotics in food products and other complex matrices [65,66,67], but this area is only indirectly related to antibiotic resistance and is therefore outside the scope of this research.
Specific microfluidic models simulating infection conditions are also being developed to study infectious agents in clinical practice. For example, Microfluidic Silicon Membrane Canalicular Arrays (μSiM-CA) [68] were developed to investigate the colonization of the osteocyte lacuno-canalicular network of cortical bone by Staphylococcus aureus. Despite the importance of these approaches to medicine, they are not directly related to the problem of antibiotic resistance and are therefore also not discussed in this review.
2. Producing Strains: Discovery and Cultivation
Many ecological processes, including the production of secondary metabolites, arise from single-cell behaviors and interaction-dependent regulation. At present, natural microbial consortia are mainly explored through cultivation-based approaches, which limits the number of strains that can be screened and often provides a biased view of microbial diversity [32]. Microfluidic technologies—particularly droplet-based systems—have emerged as powerful tools for single-cell studies [32,42], including the discovery and development of biocontrol agents and antibiotic-producing microorganisms.
One of the most notable examples of a microfluidic platform for biodiversity screening involves single-cell cultivation of microorganisms in double water-in-oil-in-water (W/O/W) emulsions, followed by fluorescence-activated cell sorting (FACS), as described previously [69] (Figure 1A). This approach was recently enhanced through the integration of live biosensors, allowing for precise monitoring of antimicrobial activity [70]. This platform has been applied to the functional profiling of diverse microbial communities, enabling the identification of amicoumacin-producing strains [71], screening of wild-animal microbiota for potential biocontrol agents [72] and isolation of a novel bacteriocin active against methicillin-resistant S. aureus (MRSA) [73]. A similar concept underlies the GrowMIDE platform, developed for studying human gut microbiota, which demonstrated an ability to recover of rare microbial species [74]. The key advantage of such double-emulsion systems lies in their compatibility with conventional flow cytometry equipment, enabling effective enrichment of bacterial populations containing bioactive isolates through fluorescence-gated sorting. Gel microbeads share this advantage with W/O/W emulsions; a recent proof-of-concept study demonstrated their application for high-throughput screening of antibiotic-producing strains using FACS-based sorting [75].
Selective isolation of individual colonies of interest for further cultivation and analysis remains particularly challenging in most microfluidic systems. A droplet-based microscale cultivation platform was developed to enable high-throughput screening of antimicrobial activity prior to strain isolation [76]. In this approach, the microbial community is encapsulated into water-in-oil (W/O) droplets, which are incubated in bulk and subsequently injected into a 100 μm capillary for deposition onto an agar surface (Figure 1B). This results in spatially separated microcolonies derived from individual droplets. Although the current method requires specialized equipment, related techniques for automated microdroplet dispensing with single-colony resolution are under active development [77].
An alternative on-chip cultivation approach, termed the digital plating (DP) platform, was proposed to simplify single-cell isolation [78]. The DP workflow involves discretizing the sample into a high-density picoliter microwell array (PicoArray device), followed by coverage with a thin agar medium sheet for incubation and subsequent microscopic examination (Figure 1C). Although the throughput of this method is relatively limited, it provides a straightforward route for isolating individual microcolonies at the single-cell level.
A large fraction of natural microbial biodiversity—often referred to as “microbial dark matter”—remains uncultivable under standard laboratory conditions but represents a vast, untapped reservoir of novel secondary metabolites. Exploration of this “dark microbiome” through in situ cultivation has proven to be an effective strategy for discovering unique metabolites, exemplified by the isolation of teixobactin [79]. To increase the throughput of in situ approaches, a microfluidic adaptation known as the microbe domestication pod (MD Pod) was recently introduced [80]. In this system, single cells are encapsulated into agarose microbeads—generated using standard microfluidic protocols—and subsequently incubated within a modified growth chamber (the MD Pod) in their natural environment (Figure 1D). The device was later miniaturized (μMD Pod) to enable its use in small habitats, such as marine invertebrates [81]. The in situ cultivation strategy shows great promise for isolating rare antibiotic producers. A recent study on taxonomic profiling of soil microbiota cultivated in droplets containing soil suspensions enriched with key natural metabolites demonstrated the preservation of low-abundance taxa and recovery of otherwise inaccessible microorganisms [82], emphasizing the critical role of natural environmental factors in microbial cultivability.
A distinct and practically significant challenge in antimicrobial research is the development of industrial microorganisms capable of high antibiotic production. Integrating of whole-cell biosensors, W/O emulsion and fluorescence-activated droplet sorting (FADS) led to the establishment of a co-cultivation-based microfluidic screening platform for actinomycetes, termed WELCOME (Whole-cell biosensor and producer co-cultivation-based microfluidic platform for screening actinomycetes) [83]. Subsequent variations of this platform introduced fine-tuning of regulatory element expression through ribosome-binding site engineering of an MphR-based Escherichia coli biosensor [84], as well as the development of dual-color biosensors that provide normalized fluorescence outputs [85]. The WELCOME system was successfully applied to select Saccharopolyspora erythraea strains hyperproducing erythromycin.
Screening libraries of chemically induced mutant strains also represents a promising route for diversifying natural metabolite profiles. The metabolic patterns of mutant strains selected using a droplet-based microfluidic platform were shown to be comparable to those obtained via classical screening procedures [86]. In this approach, chemically mutagenized libraries of actinomycetes and bacteria were encapsulated into W/O droplets and processed using an integrated microfluidic device featuring single-droplet trapping sites equipped with pneumatic microvalves. Fluorescence-based detection enabled selective extraction of mutant strains for subsequent mass-spectrometry profiling (Figure 2B).
Functional screening of microbial consortia is challenging, because their study requires not only detection and sorting of single cells, but also recovery of the selected strains for further cultivation. Current microfluidic capabilities for functionality screens offer the flexibility needed for diverse assay designs in microbial ecology and have so far been applied predominantly for enzyme evolution, with only scarce reports suggesting that droplet isolation tends to improve recovery of natural biodiversity [32]. A lot of microfluidic cultivation approaches are being developed for specific tasks [32,42]. Therefore, the transfer of these technologies is an emerging field for microbiome screening.
3. Antibiotics: Screening and Mechanism Elucidation
For both mechanism of action studies and screening studies and alternative approaches to determining inhibitory concentrations, a critical step is creating a gradient of one or more components. Most of the existing solutions are based on either on-chip fluid manipulation (Figure 3A-D) or generation, merging and diluting droplets (Figure 3E-F) [87].
It is worth noting that microfluidic technologies are constantly evolving, including the development of new approaches to creating gradients of chemical compounds. For example, to test combinations of anticancer drugs on single cells, an alternative tree-shaped gradient generation architecture was developed [88]. Another recently designed compound mixer is 3D microchannel network design for multiple concentration gradient formations and mixed solution combinations [89]. Development of novel systems for clinical AST includes introduction of novel on-chip (e.g. MVM2 platform [90]) and off-chip (e.g. microfluidic mixer [91]) gradient generators. In this rapidly developing field transfer of technologies and approaches from related fields holds promise for effective development of novel platforms, recently cross-disciplinary gradient devices were comprehensively summarized [92].
3.1. Advances in Screening Approaches
Screening natural or synthetic compounds for potential drug candidates remains significantly constrained by the low throughput and labor-intensive operations required for bioactivity testing. Microfluidic technologies have emerged as powerful tools for analyzing microbial susceptibility to diverse agents and their combinations. The most rapidly advancing area within this field is antimicrobial susceptibility testing (AST), aimed at facilitating rapid therapeutic decision-making in clinical settings. These methods are designed for the fast evaluation of clinical isolate resistance to antibiotics, requiring minimal instrumentation and providing short turnaround times. While numerous recent reviews summarize microfluidic AST approaches [93,94,95,96,97,98,99,100,101,102,103,104,105,106], the present section focuses on methods developed for (or adaptable to) in vitro drug discovery and combinatorial drug development.
3.1.1. Screening of Chemicals and Their Combinations
Microfluidic approaches to drug screening are predominantly droplet-based. The first challenge in this workflow is controlled generation of multicomponent droplets. Once droplets containing bacteria and varying drug concentrations are generated, bacterial growth within individual droplets must be analyzed in relation to the corresponding antibiotic dose. The first approach to control of droplet interior is co-encapsulation with array of fluorescent dyes. Fluorescent droplet barcoding is typically combined with off-chip bulk incubation followed by aliquot sampling and microscopy imaging [107,108,109] (Figure 4).
Droplet-based microfluidic systems enable precise control of experimental parameters for determining the minimum inhibitory concentration (MIC) and related metrics. For example, color-coded control of cell density improved the accuracy of inoculum-effect evaluation [110]. In this study, antibiotics were co-encapsulated with yellow fluorescent protein (YFP)-expressing E. coli; initial cell counts were barcoded using two additional fluorescent dyes. Subsequent fluorescence-activated droplet sorting (FADS) with three detection channels enabled quantification of the MIC (defined as growth inhibition in 50% of droplets) and a more accurate parameter—the minimum inhibitory amount (MIA), representing the antibiotic dose per bacterium, which remains constant regardless of inoculum density (Figure 4A).
Efficient high-throughput droplet screening also requires gradient generation, implying precise droplet manipulation steps. Several recent studies have explored improved or alternative droplet control strategies for screening assays. The simplest method involves generating separate droplet populations containing different antibiotic concentrations [111], though its throughput is limited. To advance the droplet-merging technique (Figure 3), an integrated chip, allowing re-flow of generated droplets and synchronized reagent addition via droplet cleaving, was introduced (Figure 4B) [107]. Another approach achieves droplet multiplexing and tracking through combinatorial sample preparation [108]: the droplet generator receives automatically dosed and merged sample plugs, each coded by fluorescent dyes, with merging achieved via oscillatory flow (Figure 4C). An interesting alternative approach for droplet generation is based on sequential spraying of fluorophore-encoded components (Figure 4D). Although this equipment-free method yields polydisperse droplets, fluorescence staining enables quantitative determination of compound concentrations via imaging [109].
It is worth noting, that droplet-based assays can suffer from chemical escape from droplets. Limited cross-talk may be advantageous in microbiome cultivation studies, allowing inter-droplet chemical signaling within isolated microenvironments. In susceptibility assays, by contrast, maintaining consistent antibiotic and dye concentrations across droplets is critical. This limitation should be taken into account in the design of droplet experiments, for example, by means of prediction of antibiotic retention in emulsion [112]. A recent alternative approach addresses permeation control by using double-emulsion-templated giant unilamellar vesicles (GUVs) as microincubators for bacterial cultivation [113]. In this system, permeability of the polydimethylsiloxane (PDMS)-based block copolymer membrane is controlled by introducing biopores formed by the amphiphilic peptide melittin (Figure 4E).
A fundamental challenge of fluorescent barcoding lies in the differing physicochemical properties of dyes and tested compounds, which can cause variable leakage and inconsistencies in measured results [112]. Microvalve-based droplet generation supports alternative detection method − spatially encoded reagent gradients.
Microvalve-based droplet systems remain limited by the complexity of microdevice design and fabrication. However, related techniques continue to evolve toward greater precision and simplicity. The recently developed Tubing Eliminated Sample Loading Interface (TESLI) utilizes vacuum-based actuation to streamline sample handling [114]. A recent innovation - Artificial intelligence-accelerated high-throughput combinatorial drug evaluation system (AI-HTCDES) – achieves effective combinatorial dosing on the basis of readily available commercial multiway valves and programmable syringe pumps (Figure 5A) [115], addressing some of these limitations.
Typically, automated droplet production is followed by ordered transfer into tubing (Figure 5B) or S-shaped collection chip (Figure 5A) for incubation and subsequent ordered ejection through a detection channel for sequential detection. This method was successfully used for combinatorial antibiotic screening [115,116]. Imaging of spatially-ordered droplets generates large datasets that demand automated analysis. To address this challenge, deep learning–based image processing was incorporated into the AI-HTCDES platform [115]. In this assay, sequential bright-field imaging of over 7,000 droplets was efficiently analyzed using machine learning algorithms to identify bacterial growth automatically.
Sequential detection constrains throughput and scalability, so the detection approach was upgraded in the Robotic-Printed Combinatorial Droplet (RoboDrop) [117] system by the addition of a robotic arm after tubing, which automatically prints the ejected droplets into an array, enabling simultaneous imaging of thousands droplets using a standard camera (Figure 5C).
Straightforward observation of growth inhibition in conventional microfluidic channels has proven unsuitable for detecting antimicrobial compounds within complex natural product matrices [118]. SlipChip devices present an attractive alternative to standard microfluidic protocols, offering comparable precision in fluid manipulation without the need for external instrumentation. A detailed review of SlipChip design principles and their applications in digital bioassays was recently published [119].
In a SlipChip droplets are generated through the relative movement of two closely aligned microstructured glass plates, each patterned with microwells and microducts on their contacting surfaces. This configuration was adapted for screening natural products with antimicrobial activity in the nanoliter matrix SlipChip (nm-SlipChip) [120]. In this assay ten antimicrobial candidates (either pure compounds or extract fractions) are simultaneously tested against ten bacterial cultures within microwell-generated droplets (Figure 6). The compounds or fractions (and potentially different drug concentrations) are loaded into the X-axis channels of the chip, while bacterial suspensions are introduced into the Y-axis channels. A controlled sliding motion merges the two arrays, initiating on-chip incubation. Following incubation, droplet growth inhibition is assessed microscopically. Compared with classical bioactivity testing methods, this approach not only minimizes sample consumption but also reduces analysis time to approximately three hours. Aside for fabrication of two glass plates, comprising the nm-SlipChip, by etching method, this approach does not require any specific equipment. Importantly, it integrates droplet generation and spatial control of droplet contents within a single device, and it is compatible with the recovery of droplets of interest for downstream analysis or alternative detection methods.
3.1.2. Screening of DNA-Encoded Antimicrobials
Specific microfluidic screening strategies can be applied to evaluate libraries of DNA-encoded antimicrobial peptides (AMPs). The genetically encoded sequences of AMPs enable the generation of libraries containing unnatural modifications through directed evolution techniques [121]. AMP libraries can be produced either by chemical synthesis, heterologous recombinant expression systems, or cell-free translation platforms. In all cases, high-throughput screening of target biological activities is essential for hit identification, making microfluidic methods particularly suitable for this area of research. For example, droplet based screening in double W/O/W emulsion was applied for evaluation of protegrin-1 production in Pichia pastoris yeast, establishing a platform for screening of recombinant AMP activity against gram negative bacteria [122] (Figure 7A).
Since most AMPs act through membrane permeabilization and frequently lack selectivity, often displaying high hemolytic activity, strategies that assess their membrane specificity are critical. microfluidic system enabling simultaneous cell-free production and screening of AMP membrane specificity was developed to address this need [123]. In this approach, AMPs are produced within W/O/W double emulsion droplets, co-encapsulated with sensor large unilamellar vesicles are generated that generate a membrane-specific fluorescence signal. Two distinct populations of sensor liposomes—with bacterial-like or mammalian-like lipid compositions—are preloaded with self-quenched fluorescent dyes. Dye leakage in droplets can be observed by droplet trapping of FACS and is proportional to the preference of the AMP for the specific membrane composition. (Figure 7B). Sensor liposomes-based approach was further shown to be applicable to detect and evaluate membrane-active antimicrobial peptides in non-ribosomal peptide synthase mutant libraries, exemplified by gramicidin S heterologous production in E. coli [124].
An interesting alternative approach allowed to assess AMP selectivity on whole cells on the basis of microfluidic impedance cytometry [125]. In this approach, bacterial cells and human red blood cells are treated with AMP off-chip, and the resulting individual or mixed samples are analyzed using an impedance cytometer. The system records electrical signatures of single cells, reflecting AMP-induced changes in membrane integrity. The key advantage of this approach lies in its rapid, label-free, and co-culture-compatible evaluation of AMP selectivity, providing physiologically relevant insights into peptide activity.
3.1.3. Bacteriophage Evaluation and Development
Bacteriophages are increasingly recognized as a promising alternative strategy for combating antibiotic-resistant bacteria. However, conventional phage isolation methods remain labor-intensive and may introduce significant bias [126]. Microfluidic technologies offer new opportunities for phage research by providing precise control over environmental parameters and enabling high-throughput experimentation. Despite this potential, reported applications of microfluidics in phage biology are still relatively limited.
The first demonstration of a droplet-based approach for isolating infectious phages involved the co-encapsulation of phages and E. coli host cells in W/O droplets [127]. he fluorescent nucleic acid dye YOYO-1 was used to label phage DNA; due to its cell-impermeant nature, it selectively stained viral particles without affecting host cells. Droplets were analyzed and sorted using an On-chip Droplet Selector, allowing the isolation of target droplets into individual wells of a 96-well plate (Figure 8A). This proof-of-concept study established the feasibility of droplet microfluidics for phage discovery and has since inspired further developments, as exemplified by two recent preprints expanding this approach [128,129].
Aforementioned technology for on-chip droplet generation was also adapted for phage quantification in digital phage SlipChip (dp-SlipChip), relying on bright-field microscopy for host bacterial cells growth detection [130] (Figure 8B). An alternative on-chip cultivation strategy utilized a commercially available microfluidic platform (Mimetas Organoplates), originally developed for eukaryotic cell culture, and repurposed it for phage isolation and quantification [131]. Each chip consists of two parallel microchannels converging into a shared interaction chamber separated by a narrow phaseguide. In this setup, one channel is filled with bacteria immobilized in agarose gel, while the adjacent channel contains phage suspensions in liquid medium (Figure 8C).
3.1.4. Antibiofilm Activity Testing
The development of biofilms plays a pivotal role in microbial resistance to treatment in clinic. Consequently, compounds that can either prevent biofilm formation or disrupt mature biofilms are of significant therapeutic interest. Microfluidic platforms have proven particularly valuable in biofilm research, as they provide precisely controlled, dynamic environments that closely mimic natural growth conditions
Well-established open biofilm models, available commercially, e.g. BioFlux [132] and similar microfluidic chambers remain widely used for biofilm studies and screening applications. In these devices, biofilms are formed within a microchannel attached to a glass slide under continuous flow conditions (Figure 9A). Even this relatively simple experimental configuration yields substantially more physiologically relevant data compared with traditional static models. For instance, the BioFlux system enabled the identification of the antibiofilm activity of the peptide Temporin L against P. fluorescens, demonstrating effects beyond its known antimicrobial properties [133]. A similar setup was employed to reveal the antibiofilm efficacy of murepavadin against P. aeruginosa [134] and to study modulation of intestinal bacterial adhesion by naringenin derivatives [135].
Further applications of BioFlux-based models include investigations of ultrashort antifungal lipopeptides against Candida biofilms [136] marine invertebrate-derived AMPs active against opportunistic pathogens Aurelia aurita and Mnemiopsis leidyi [137]. These studies demonstrated that microfluidic biofilm systems can generate reproducible and biologically meaningful results relevant to complex natural and clinical environments
A three-port modification of the BioFlux design was later introduced, allowing the application of spatially separated flows of distinct media within a single device. This configuration was validated for antibiofilm screening [138] (Figure 9B). To further improve screening capacity, a high-throughput microfluidic perfusion biofilm reactor (HT-μPBR) was recently developed specifically for antibiofilm testing [139] (Figure 9C). Compared with the standard BioFlux system, the HT-μPBR offers enhanced shear control and full compatibility with standard microtiter plates and incubator setups.
Research on the formation and eradication of mixed-species biofilms is critically important, as many infections in the human host are caused by polymicrobial biofilms. Inoculation of BioFlux device with mixed S. aureus and Pseudomonas aeruginosa culture was applied to achieve diabetic foot environment model for antibiotic screening [140]. A more advanced platform incorporating a herringbone micromixer for combining separate microbial flows, coupled with optofluidic imaging for real-time monitoring, was applied to study S.aureus + C. albicans mixed biofilms [141] (Figure 9D). Recently, an even more sophisticated system for modeling biofilms—including polymicrobial and clinically derived communities—was introduced [142]. The BiofilmChip design (Figure 9E) features a prechamber and three growth chambers with optimized geometry, supporting both confocal microscopy and impedance measurements. Validation using clinical samples demonstrated that BiofilmChip effectively supports the growth and characterization of polymicrobial biofilms.
Bacterial colonization on solid surfaces and subsequent biofilm formation pose particular challenges in the context of implanted devices application. The development of antimicrobial coatings and materials is therefore an active research area, though their reliable screening and evaluation remain difficult. Microfluidic technologies have provided efficient solutions to this problem. For instance, a custom microfluidic device was used to investigate the attachment behavior of P. aeruginosa and S. aureus species on nanostructured surface under fluid wall shear stress [143]. Fluorescent staining and microscopic visualization enabled detailed quantification of bacterial adhesion (Figure 9F). Another study employed a standard biofilm model to assess the antimicrobial performance of AMP-functionalized surfaces [144]. In this work, continuous monitoring of E. coli growth revealed crucial factors to aid AMP-grafted coatings design.
3.2. Studies on Antibiotics Mode of Action
Identification of a hit compound or effective drug combination represents only the first stage in antimicrobial drug development. The next critical step is elucidating the molecular mode of action (MoA) underlying the observed biological effects. In contrast to screening-oriented platforms, microfluidic systems designed for mechanistic studies typically shift from endpoint growth detection toward time-resolved single-cell imaging, requiring modified experimental setups and device architectures.
One of the most established microfluidic designs for single-cell is a configuration nicknamed the “mother machine” (MM, [145]). The MM consists of a large central flow channel intersected by numerous perpendicular, single-cell-wide dead-end side trenches. During inoculation, individual bacterial cells become trapped in these narrow side channels, while their progeny are continuously flushed away by a constant flow of fresh medium in the main channel (Figure 10A). Time-lapse microscopy in this configuration enables the observation of single-cell lineages over hundreds of generations [146]. The MM remains widely applied in recent antibiotic MoA studies (Table 1).
Building upon this principle, a modified design termed the isolated mother machine (iMM) was developed for the capture of extracellular vesicles [147]. In this system, individual cells are retained in narrow side channels similar to the MM, but all daughter cells are immediately removed by continuous flow. The winding microchannels are selectively coated with poly-l-lysine, forming a positively charged surface that enables electrostatic trapping of negatively charged extracellular vesicles (Figure 10B).
Another widely adopted strategy for single-cell imaging involves trapping bacteria in monolayers to track microcolony dynamics. Various devices, including commercially available (e.g. CellASIC® ONIX B04A-03 bacteral plates), have been employed for detailed investigation of bacteria–drug interactions (Table 1). Working principle includes loading and trapping of single cells in one-cell high monolayer with subsequent time-resolved imaging of microcolonies development (Figure 11A). Device with variable chamber height have been applied to selectively trap normal bacterial cells or cell-wall-deficient L-forms (Figure 11B) [148]. An alternative design employs hydropneumatic chambers to form bacterial monolayers (Figure 11C). Such systems have been utilized in studies of Mycobacterium smegmatis studies, including the μDeSCRiPTor platform – a microfluidic dynamic single-cell screening for pheno-tuning compounds, used for mechanism-informed development of anti-tubercular adjuvant therapies [149].
A different mechanistic approach involves on-chip generation of immobilized droplets, enabling real-time observation of bacterial growth and lysis dynamics (Figure 11D). This setup has been applied to study E. coli challenged with phages [150] and antibiotics [151]. Immobilized droplet arrays provide stable and isolated microenvironments for time-resolved imaging of individual bacterial responses.

Table 1.
Summary of single-cell imaging-based studies of modes of antimicrobial action.
| Compound | Label | Device | Detection method | Test strain | Observed effect | Ref |
|---|---|---|---|---|---|---|
| Ultrashort peptides | Live/Dead stain |
Figure 10 MM |
Fluorescent imaging | Е. Coli | Variable single cell killing kinetics | [152] |
| Roxithromycin | Labelled antibiotic |
Figure 10 MM |
Fluorescent imaging | E.coli, expressing phage secretin f1pIV | Drug uptake | [153] |
| Ofloxacin | Antibiotic natural fluorescence |
Figure 10 MM |
Epifluorescent imaging | Е. Coli | Drug uptake | [154] |
| Polymyxin | GFP, membrane stain |
Figure 10B iMM |
Fluorescent imaging | Е. Coli | Extracellular vesicles secretion | [147] |
| protein capsids | - |
Figure 10A MM |
Bright-field microscopy | Е. Coli | Variable single cell killing kinetics | [155] |
| Proof-of principle study | Labelled vancomycin |
Figure 10A MM |
Fluorescent imaging | Е. Coli | Outer membrane damage by probe uptake | [156] |
| Polymixin | - | Figure 10B | Bright-field microscopy | Е. Coli | L-forms morphology and proliferation | [148] |
| Aminoglycosides and fluoruquinolones | GFP, mCherry | Figure 11A | Fluorescent imaging | Е. Coli with genetic compartment markers | Hyperosmotic shock, cytoplasmic condesation | [157] |
| Meropenem, berberine | Live/Dead stain | Figure 11A | Bright-field and fluorescent microscopy | A. baumannii | Single-cell growth kinetic under combination therapy | [158] |
| Listeriolysin S | GFP, Sytox blue | Figure 11A | Fluorescent imaging | Listeria monocytogenes | Contact-killing by producing strain | [159] |
| Moxifloxacin | GFP, PI | Figure 11C | Bright-field microscopy | M. smegmatis | Single-cell dose response | [160] |
| M06 pheno-tuning compound | mCherry, GFP | Figure 11C | Fluorescent imaging | M. smegmatis | Single-cell phenotypes | [149] |
| Phages | GFP | Figure 11D | Fluorescent imaging | Е. Coli | Growth and lysis kinetics | [150] |
| Ciprofloxacin | mRFP | Figure 11D | Fluorescent imaging | Е. Coli | Growth kinetic | [151] |
To emulate drug bioavailability in intracellular infections, a specialized model based on droplet interface bilayers (DIBs) was developed [161]. In such infections, the host-cell membrane forms a barrier between the pathogen and antimicrobial agents. The DIB model reconstitutes this condition by forming lipid bilayers at the interfaces of lipid-coated water-in-oil (W/O) droplets. Arrays of droplets are generated on a chip equipped with a movable chamber, following previously established methods [162]. Controlled contact between antibiotic-containing droplets and bacteria-loaded droplets allows monitoring of drug diffusion across the DIB, mimicking intracellular drug exposure (Figure 12A).
Another approach to study drug–membrane interactions utilizes freestanding planar lipid bilayers, though their formation and buffer exchange have traditionally been difficult to control. A recently introduced on-chip platform allows arrayed planar bilayer formation and was successfully applied to study the effects of azithromycin on the mechanical properties of POPC lipid bilayers (Figure 12B) [163].
Model membranes permeabilization under antibiotic treatment can be also examined using GUVs. A label-free, impedance-based on-chip GUV detection system integrating microelectrodes within the microfluidic channel was developed and validated using norfloxacin as a model compound [164] (Figure 12C). Notably, this system could distinguish between antibiotic permeation through the lipid bilayer and transport mediated by inserted porin channels, offering detailed mechanistic insights into drug–membrane interactions.
4. Pathogens: Stress Responses and Resistance Development
Systematic studies of how bacterial populations respond to antibiotic stress and how resistant subpopulations emerge provide the mechanistic and quantitative foundation for rational antimicrobial therapy. Microfluidic technologies have become indispensable in this context, offering precise environmental control, high temporal resolution, and single-cell-level insight into the complex dynamics of resistance acquisition.
4.1. Genetic Resistance
Adaptive laboratory evolution (ALE) is a key method for investigating the emergence of antibiotic resistance and elucidating its underlying mechanisms. Microfluidic systems have transformed ALE workflows by accelerating mutant generation, improving environmental control, and enabling high-throughput selection of desired traits [54]. Beyond mechanistic insights, these approaches facilitate detailed preclinical resistance prediction, potentially extending the clinical lifespan of new antimicrobial agents [53]. Recent review summarizes current approaches to the assessment of resistance development, focusing on technologies of laboratory evolution on resistance mutants on hit-to-lead stage of drug development [53].
Several microfluidic systems have been developed recently to enhance ALE efficiency (Figure 13). Standard ALE protocols are time-consuming, but a centrifugal microfluidic system was designed to accelerate resistance evolution [165]. In this device, a PDMS chip pre-loaded with antibiotic and bonded to glass is filled with bacterial suspension. On-chip centrifugation generates a condensed bacterial matrix, allowing survival under antibiotic concentrations several times higher than the MIC and promoting rapid resistance development (Figure 13A).
Conventional batch ALE methods are also limited by competitive exclusion, where slow-growing but strongly resistant variants are outcompeted, leading to loss of diversity. Emulsion-based cultivation provides spatial isolation and fine control over environmental conditions, enabling the detection of distinct evolutionary trajectories—as shown for P. aeruginosa evolving resistance to colistin [166] and E. coli adaptation to doxycycline [167] (Figure 13B).
A related approach employs spatially organized stress gradients to promote adaptation toward complex phenotypes (Figure 13C). This method enabled the identification of previously unrecognized mutations in E. coli conferring resistance to nalidixic acid [168]. Further studies on E. coli mutants, resistant to ciprofloxacin, generated in on-chip spatial gradient, revealed that microfluidic environment itself can induce mutations beneficial for the subsequent evolution of highly resistant mutants [169].
In addition to identification of genetic determinants of resistance, microfluidic protocols can be applied to phenotyping of mutant strains and detalization of resistance genes dissemination processes by single cell imaging (Table 2). For example, single-cell imaging of growth rate was applied to show fitness cost of gene amplification-mediated resistance, demonstrating rapid loss of amplification in the absence of selection for clinical isolates of E. coli and Salmonella enterica [170]. Moreover, observaton of single colonies in monolayer can be utilized for detection of individual events of horizontal and vertical gene transfer [171,172]. Physiological role of known resistance-associated mutation can be elucidated on the basis of single-cell phenotyping. For instance, previously described monolayer trap (Figure 11C) for M. smegmatis was utilized to compare persistent phenotypes in wild type cells and the the msm2570::Tn transposon mutant that has an insertion in themsm2570 gene encoding a putative xanthine/uracil permease.
4.2. Phenotypic Resistance and Bacterial Stress Responses
Antibiotic therapy is never applied to a homogeneous, static target. Even within a single clonal culture, bacterial populations are dynamic, spatially structured, and phenotypically diverse. Studies that track how bacterial cultures respond to antibiotic exposure and how resistant subpopulations arise are therefore essential to understanding treatment success and failure. Single-cell resolution is extremely informative in a realm of non-heritable, phenotypic resistance studies, recently applied experimental settings are summarized in Table 3.
Classical colony-forming unit (CFU)-based killing assays can misrepresent persister dynamics due to post-exposure killing, as demonstrated for Salmonella [174]. Tracking of pre- and post-exposure history of persisters formation was also successfully achieved for E. coli using membrane-covered microchamber array (MCMA) [175].
Many recent studies aimed to characterize subpopulations of bacteria, surviving drug exposure, and elucidate key features, underlying resistant phenotype. Phenotypic resistance is typically associated with growth-arrested persisters. Indeed, several studies revealed persister subpopulations, exhibiting dormant state of reduced growth rates. For example, viable but non-cultural state was induced in B. subtilis prior to antibiotic exposure by osmotic stress, leading to antibiotic tolerance [176]. Fluorescent reporter for in vivo ATP measurments revealed that a subpopulations of E. coli with a low level of ATP survive ampicillin exposure, suggesting a “low-energy” mechanism of persistence [177].
Suprisingly, subpopulation of cells, surviving the antibiotic exposure, were found to exhibit diverse growth rates. Under ampicillin treatment, E. coli cells with ~50% reduced growth rate demonstrated lower antibiotic susceptibility, compared with both normal and growth-arrested cells [178]. Moreover, fast growing resistant to macrolides subpopulation with high expression of ribosomal promoters was detected under investigation of cell-to-cell differences of drugs uptake. Presumably drug-free ribosomes facilitated essential cellular processes including efflux of macrolides, leading to fast-growing resistant phenotype [179]. Comparative studies across antibiotics revealed distinct stress-response mechanisms. Among nine tested antibiotics, treatment with rifamycin and nitrofurantoin, differing in both structure and mode of action, led to surviving sub-populations [180]. Phenotypically resistant strains, particularly those with high growth rate, are significantly increasing risk of genetically fixed resistance development.
Phenotypic resistance has diverse and elusive mechanisms, depending on vast variety of factors. For example, studies of E. coli phenotypic resistance showed that multicopy plasmids can serve as drivers of bacterial adaptation, enabling rapid modulation of gene copy numbers [181]. Moreover, E. coli demonstrated physiological adaptation to antibiotic treatment after deletion of resistance genes [182]. Phenotypic resistance to AMP can be mediated by efflux activity and was shown to be reverted by efflux pump inhibitor [183]. Similar results were obtained for S. aureus, escaping β-lactam treatment by AbcA transporter overexpression [184]. Visualization of plasmid-encoded fluorescent pH reporter showed that E. coli persisters, resistant to ampicillin, have lower intracellular pH than viable but non-culturable and susceptible cells [185], highlighting the manipulation of the intracellular pH can be a strategy for targeting persisters.
All the aforementioned studies applied fluorescent or optical imaging to monitor stressed populations. As a non-destructive and label-free alternative to imaging, Raman spectroscopy combined with acoustic trapping enabled monitoring of M. smegmatis response to antibiotic treatment [186]. Despite challenges in interpretation, Raman fingerprints monitoring is promising approach to access metabolic changes in live cells. The most informative approach to assess stress-induced metabolomics changes is mass-spectrometry. On-chip lysis of antibiotic-treated bacteria with subsequent ESI-MS monitoring revealed up-regulation of cyclic dinucleotide, c-di-GMP, in ESBL-producing E. coli. Treatment with chemical inhibitor of c-di-GMP biosynthesis indeed reverted resistance to beta-lactams, verifying the identified self-saving response as target for adjuvant development [187]. Despite obvious advantages, coupling of microfluidic approaches with MS-detection is very challenging.
Table 3.
Summary of experimental approaches to study heteroresistance.
| Antibiotic | Device | Detection method | Studied strains | Observed effect | Ref |
|---|---|---|---|---|---|
| Ampicillin Ciprofloxacin |
MCMA | Microscopy | E. coli | Pre- and post-exposure imaging of individual cells | [175] |
| Ampicillin |
Figure 10A MM |
Microscopy | E.coli, expressing pHluorin | Intracellular pH in individual cells | [185] |
| Ampicillin |
Figure 10A MM |
Microscopy | Е. Coli, expressing iATPSnFr1.0 | ATP levels in individual cells | [177] |
| Trimetoprim Linezolid Ciprofloxacin Roxithromycin Vancomycin Polymyxin Octapeptin Tachyplesin |
Figure 10A MM |
Microscopy |
E. coli P. aeruginosa Burkholderia cenocepacia S. aureus |
Accumulation of fluorescent antibiotic derivative | [179] |
| Chloramphenicol |
Figure 10A MM |
Microscopy | E.coli | Growth kinetics after resistance gene deletion | [182] |
| Chloroamphenicol Gentamycin Spectinomycin Tetracyclin Rifampicin Ciprofloxacin Nirtofurantoin Carbenicillin Ceftriaxone |
Figure 10A MM |
Microscopy | E.coli | Growth response under antibiotic treatment | [180] |
| Nafcillin Oxacillin |
Figure 10A MM |
Microscopy | E.coli with AbcA transporter overexpression | Individual cells growth rate | [184] |
| Tachyplesin (AMP) |
Figure 10A MM |
Microscopy |
E.coli P. aeruginosa |
Fluorescent antibiotic uptake | [183] |
| Ampicillin |
Figure 10A MM |
Microscopy | E.coli | Growth kinetic under antibiotic treatment | [178] |
| Flucloxacillin | Figure 11A | Microscopy |
Salmonella S. aureus |
Single-cell growth and regrowth kinetic | [174] |
| Kanamycin | Figure 11A | Microscopy | B. subtilis | Single-cell growth kinetic, fluorescent antibiotic uptake | [176] |
| Cefotaxime | Droplet (W/O) |
Microscopy | E.coli, expressing β-lactamases | Susceptibility distribution | [188] |
| Isoniazid | Acoustic trap |
Raman spectroscopy | M. smegmatis | Single-cell metabolic response by Raman fingerprint | [186] |
| Ampicillin | Figure 11A | Microscopy | E. coli | Indirect monitoring of plasmid copy number | [181] |
| Ceftriaxone | HV-coupled channel |
Mass-spectrometry | E. coli | Metabolic response to antibiotic treatment | [187] |
Emerging research highlights the role of biomolecular condensates in bacterial stress responses. Increasing evidence indicates that key cell cycle proteins form phase-separated condensates under stress, serving as adaptive organizational hubs and potential drug targets [189]. Droplet microfluidics has been used to model such condensates, providing mechanistic insight into E. coli division machinery [190].
Recovery of specific phenotypes in microfluidic assays often requires active sorting (e.g., FACS or FADS). Recently, passive sorting techniques have gained attention as scalable, low-cost alternatives [18]. One of the approaches to passive sorting is vascoelasting microfluidics, it was recently successfully adapted for shape-based separation of β-lactam-treated E. coli cells [191] The device (Figure 14) integrates two inlets for viscoelastic and Newtonian fluids and seven outlets; spherical bacteria migrate toward the channel center, while elongated cells are directed to the sides, enabling label-free morphological separation.
4.3. Chemotaxis and Bacterial Motility
Microfluidic models can reproduce spatial gradients of antibiotics and nutrients, reflecting environmental and host conditions (Figure 15). For example, studies of P. aeruginosa in long gradient channels revealed that flow-shaped gradients significantly alter antibiotic susceptibility [192]. Several approaches to study bacterial chemotaxis were applied recently, including imaging of E. coli migration in single bacterial layer [193] (Figure 15A), Shewanella oneidensis chemotaxis in microfluidic gradient chamber [194] and E. coli growth in spatially controlled population under tetracycline treatment (Figure 15B), suicidal chemotaxis of P. aeruginosa towards high antibiotic concentration [195,196]. The latter effect was studied in detail using reconfigurable microfluidic device, allowing to isolate subpopulations after time-lapse imaging by building fluid walls around migrating cells (Figure 15C).
A complementary “static” strategy uses hydrodynamic trapping of individual bacteria followed by high-speed video microscopy (Figure 15D). As demonstrated on E. coli MG1655 and Salmonella typhimurium, reduced single-cell motility under kanamycin exposure correlates with growth inhibition and can serve as a proxy for heteroresistance [197].
4.4. Biofilms Antibiotic Susceptibility
As discussed in Section 3, biofilms represent highly organized microbial communities that are inherently more resistant to antibiotics than planktonic cells. Moreover, they act as hotspots for horizontal gene transfer and resistance dissemination. Microfluidic platforms, including flow chambers and the BioFlux system (Figure 9A) are widely used to study biofilm-associated resistance. For instance, flow-chamber models were employed to identify P. aeruginosa biofilm resistance genes [198] and to investigate the role of the flagellar hook in biofilm structure [199]. The BioFlux platform, combined with fluorescence microscopy, revealed a strong correlation between biofilm formation and clarithromycin resistance in Helicobacter pylori clinical isolates [200].
Fluid shear stress is another key factor influencing biofilm morphology and antibiotic susceptibility. For example, Klebsiella pneumoniae clinical isolates exposed to shear stress in microfluidic channels showed increased viability under antibiotic treatment and enhanced proliferation within macrophages [201]. To simultaneously evaluate shear stress and antibiotic exposure, a double-layer microfluidic biofilm chip (2PAB) was developed, featuring a concentration gradient generator (tree-shaped, Figure 3A) and expanding flow chambers (Figure 16A) [202]. Studies using this system demonstrated species-specific differences in E. coli and P. aeruginosa biofilm eradication, emphasizing the need for controlled hydrodynamic environments.
Geometric design can strongly modulate biofilm behavior. The wavy channel geometry can create heterogeneous flow conditions, promoting localized regions of low shear stress [203] (Figure 16E). Introduction of an isolated pillar in a straight channel, located at its half-width, induced reproducible formation of biofilm streamers – filaments, suspended in flow [204] (Figure 16B). Using this platform, antibiotic-induced genotoxic stress and SOS-response was shown to cause explosive cell lysis and stimulate streamer formation in the opportunistic pathogen Burkholderia cenocepacia [205].
To overcome limitations of standard flow-chamber models—such as random adhesion, clogging, or limited harvesting—new systems have been developed (Table 4). A spatially controlled seeding chip with a pressure-based seeding gap (Figure 16C) allows reproducible biofilm formation [206]. A microfluidic chip called Brimor was developed to sudy the dynamics of enrichment of antibiotic-resistant bacteria in biofilms using confocal microscopy [207]. Brimor chip is manufactured with 3D-printed mold and includes two channels for simultaneous observation of experimental and control biofilms (Figure 16D). Modular chips integrating electrodes have been introduced for monitoring biofilm migration and regrowth dynamics (Figure 16E) [203]. Finally, a dual-chamber microfluidic device was designed to model P. aeruginosa biofilm formation at the air–liquid interface, with electrochemical detection of pyocyanin serving as a proxy for viable cell density [208] (Figure 16F).
5. Conclusions and Future Directions
Microfluidic technologies have reshaped the landscape of antibiotic resistance research by enabling precise, high-resolution observation of microbial dynamics under controlled conditions. The versatility of these systems allows them to bridge the gap between molecular-scale mechanisms and population-level phenomena, providing insights that are often inaccessible through traditional methods.
Recent advances demonstrate that microfluidics can be applied across the full spectrum of antimicrobial studies—from the discovery of new antibiotic producers and the screening of natural or synthetic compounds to mechanistic dissection of drug action and quantitative analysis of stress responses and resistance evolution. Integrated approaches combining microfluidic assays with imaging, spectroscopy and mass spectrometry are rapidly expanding the analytical capabilities of these platforms. Moreover, the rise of AI-driven data processing, automated droplet manipulation and organism-specific model systems is accelerating the throughput and enhancing reproducibility.
Despite these achievements, several challenges remain. Limited standardization of device designs and protocols continues to hinder data comparability and large-scale implementation. Many promising systems remain at the proof-of-concept stage, with limited transition to industrial or clinical workflows. Future progress will depend on the development of modular, user-friendly and scalable microfluidics platforms compatible with existing laboratory infrastructure and high-content data analysis pipelines.
Microfluidic technologies have found applications in a wide variety of research areas and continue to evolve rapidly. Each study addressing a specific scientific problem often introduces technical innovations with potential utility in other, even distant, disciplines. Furthermore, many works focus on improving encapsulation and imaging methods without direct application to particular biological problems. For instance, recent studies have proposed enhanced image analysis algorithms for MM experiments [209]. Although such developments hold great promise, identifying their relevance to specialized research domains can be challenging, emphasizing the importance of comprehensive literature analyses and reviews summarizing emerging applications of microfluidic technologies
The most rapidly advancing fields of microfluidic research aimed at combating antibiotic resistance remain pathogen detection, antimicrobial susceptibility testing, and rapid drug selection in clinical settings. Studies in this area focus on simplifying device architecture, reducing analysis time and enabling measurements at low bacterial concentrations within complex biological matrices. Nevertheless, many of these developments also hold significant potential for fundamental microbiological research. For example, the application of SlipChip technology for antimicrobial susceptibility testing (AST) [210,211,212] has evolved into dedicated platforms for novel antibiotic [120] and phage discovery [130]. Also, alternative methods for bacterial growth detection are continuously emerging and being refined, including those based on optofluidics [213,214], light scattering [215], Raman spectroscopy [216,217], impedance sensing [218], THz detection [219] and fluorescence measurments [220].
As microfluidics technologies continue to mature, they tend to become indispensable tools in antimicrobial research—supporting natural compound discovery, rational drug design, phenotypic screening and real-time resistance monitoring. Ultimately, the convergence of microfluidics with systems biology, synthetic biology and computational modeling holds the potential to redefine our understanding of microbial adaptation and guide the next generation of strategies to overcome antibiotic resistance.
Author Contributions
Conceptualization, V.A.A. and V.A.K.; writing—original draft preparation, V.A.A., M.Y.Z.; writing—review and editing, V.A.A., V.A.K.; visualization, V.A.A., M.Y.Z.; supervision, V.A.K.; funding acquisition, V.A.A. All authors have read and agreed to the published version of the manuscript.
Funding
The work was supported by the Russian Science Foundation project no. 25-14-00281, https://rscf.ru/en/project/25-14-00281/.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zafar, A.; Takeda, C.; Manzoor, A.; Tanaka, D.; Kobayashi, M.; Wadayama, Y.; Nakane, D.; Majeed, A.; Iqbal, M.A.; Akitsu, T. Towards Industrially Important Applications of Enhanced Organic Reactions by Microfluidic Systems. Molecules 2024, 29, 398. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Min, K.-I.; Inoue, K.; Im, D.J.; Kim, D.-P.; Yoshida, J. Submillisecond Organic Synthesis: Outpacing Fries Rearrangement through Microfluidic Rapid Mixing. Science 2016, 352, 691–694. [Google Scholar] [CrossRef]
- Gimondi, S.; Ferreira, H.; Reis, R.L.; Neves, N.M. Microfluidic Devices: A Tool for Nanoparticle Synthesis and Performance Evaluation. ACS Nano 2023, 17, 14205–14228. [Google Scholar] [CrossRef]
- Saeed, M.M.; Carthy, E.; Dunne, N.; Kinahan, D. Advances in Nanoparticle Synthesis Assisted by Microfluidics. Lab Chip 2025, 25, 3060–3093. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, L.; Zhang, H.; Shang, L.; Zhao, Y. Microfluidics for Drug Development: From Synthesis to Evaluation. Chem. Rev. 2021, 121, 7468–7529. [Google Scholar] [CrossRef] [PubMed]
- Elvira, K.S. Microfluidic Technologies for Drug Discovery and Development: Friend or Foe? Trends in Pharmacological Sciences 2021, 42, 518–526. [Google Scholar] [CrossRef]
- Liu, J.; Du, H.; Huang, L.; Xie, W.; Liu, K.; Zhang, X.; Chen, S.; Zhang, Y.; Li, D.; Pan, H. AI-Powered Microfluidics: Shaping the Future of Phenotypic Drug Discovery. ACS Appl. Mater. Interfaces 2024, 16, 38832–38851. [Google Scholar] [CrossRef]
- Alavi, S.E.; Alharthi, S.; Alavi, S.F.; Alavi, S.Z.; Zahra, G.E.; Raza, A.; Ebrahimi Shahmabadi, H. Microfluidics for Personalized Drug Delivery. Drug Discovery Today 2024, 29, 103936. [Google Scholar] [CrossRef]
- Fan, R.; Wu, J.; Duan, S.; Jin, L.; Zhang, H.; Zhang, C.; Zheng, A. Droplet-Based Microfluidics for Drug Delivery Applications. International Journal of Pharmaceutics 2024, 663, 124551. [Google Scholar] [CrossRef]
- Ali, M.S.; Hooshmand, N.; El-Sayed, M.; Labouta, H.I. Microfluidics for Development of Lipid Nanoparticles: Paving the Way for Nucleic Acids to the Clinic. ACS Appl. Bio Mater. 2023, 6, 3566–3576. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Oh, D.; Cheng, M.; Chintapula, U.; Liu, S.; Reynolds, D.; Zhang, X.; Zhou, Y.; Xu, X.; Ko, J. Enhancing Chimeric Antigen Receptor T-Cell Generation via Microfluidic Mechanoporation and Lipid Nanoparticles. Small 2025, 21, 2410975. [Google Scholar] [CrossRef]
- Yu, M.; Liu, D.; Shah, P.; Qiu, B.; Mathew, A.; Yao, L.; Guan, T.; Cong, H.; Zhang, N. Optimizing Microfluidic Channel Design with Tilted Rectangular Baffles for Enhanced mRNA-Lipid Nanoparticle Preparation. ACS Biomater. Sci. Eng. 2025, 11, 3762–3772. [Google Scholar] [CrossRef]
- Fardoost, A.; Karimi, K.; Govindaraju, H.; Jamali, P.; Javanmard, M. Applications of Microfluidics in mRNA Vaccine Development: A Review. Biomicrofluidics 2024, 18, 061502. [Google Scholar] [CrossRef]
- Bhat, G.R.; Lone, F.A.; Dalal, J. Microfluidics—A Novel Technique for High-quality Sperm Selection for Greater ART Outcomes. FASEB BioAdvances 2024, 6, 406–423. [Google Scholar] [CrossRef]
- Bouloorchi Tabalvandani, M.; Saeidpour, Z.; Habibi, Z.; Javadizadeh, S.; Firoozabadi, S.A.; Badieirostami, M. Microfluidics as an Emerging Paradigm for Assisted Reproductive Technology: A Sperm Separation Perspective. Biomed Microdevices 2024, 26, 23. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Yang, X.; Wang, C.; Shang, L. Microfluidics-Based Microcarriers for Live-Cell Delivery. Advanced Science 2025, 12, 2414410. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, S.; Lim, H.; Chung, A.J. Expanding CAR-T Cell Immunotherapy Horizons through Microfluidics. Lab Chip 2024, 24, 1088–1120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Di Carlo, D.; Lim, C.T.; Zhou, T.; Tian, G.; Tang, T.; Shen, A.Q.; Li, W.; Li, M.; Yang, Y.; et al. Passive Microfluidic Devices for Cell Separation. Biotechnology Advances 2024, 71, 108317. [Google Scholar] [CrossRef]
- Yang, M.; Shi, Y.; Song, Q.; Wei, Z.; Dun, X.; Wang, Z.; Wang, Z.; Qiu, C.-W.; Zhang, H.; Cheng, X. Optical Sorting: Past, Present and Future. Light Sci Appl 2025, 14, 103. [Google Scholar] [CrossRef]
- Aryal, P.; Hefner, C.; Martinez, B.; Henry, C.S. Microfluidics in Environmental Analysis: Advancements, Challenges, and Future Prospects for Rapid and Efficient Monitoring. Lab Chip 2024, 24, 1175–1206. [Google Scholar] [CrossRef]
- Zwi-Dantsis, L.; Jayarajan, V.; Church, G.M.; Kamm, R.D.; De Magalhães, J.P.; Moeendarbary, E. Aging on Chip: Harnessing the Potential of Microfluidic Technologies in Aging and Rejuvenation Research. Adv Healthcare Materials 2025, 14, 2500217. [Google Scholar] [CrossRef]
- Vafadar, A.; Takallu, S.; Alashti, S.K.; Rashidi, S.; Bahrani, S.; Tajbakhsh, A.; Mirzaei, E.; Savardashtaki, A. Advancements in Microfluidic Platforms for Rapid Biomarker Diagnostics of Infectious Diseases. Microchemical Journal 2025, 208, 112296. [Google Scholar] [CrossRef]
- Srivastava, S.; Pandey, V.K.; Singh, A.; Dash, K.K.; Dar, A.H.; Rustagi, S. Recent Insights on Microfluidics Applications for Food Quality and Safety Analysis: A Comprehensive Review. Food Control 2025, 168, 110869. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.; Zhang, Y.; Yang, X.; Guo, L.; Man, C.; Jiang, Y.; Zhang, W.; Zhang, X. Emerging Biosensors Integrated with Microfluidic Devices: A Promising Analytical Tool for on-Site Detection of Mycotoxins. npj Sci Food 2025, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- Sekhwama, M.; Mpofu, K.; Sivarasu, S.; Mthunzi-Kufa, P. Applications of Microfluidics in Biosensing. Discov Appl Sci 2024, 6, 303. [Google Scholar] [CrossRef]
- Milić, L.; Zambry, N.S.; Ibrahim, F.B.; Petrović, B.; Kojić, S.; Thiha, A.; Joseph, K.; Jamaluddin, N.F.; Stojanović, G.M. Advances in Textile-Based Microfluidics for Biomolecule Sensing. Biomicrofluidics 2024, 18, 051502. [Google Scholar] [CrossRef]
- Alonso-Roman, R.; Mosig, A.S.; Figge, M.T.; Papenfort, K.; Eggeling, C.; Schacher, F.H.; Hube, B.; Gresnigt, M.S. Organ-on-Chip Models for Infectious Disease Research. Nat Microbiol 2024, 9, 891–904. [Google Scholar] [CrossRef]
- Xue, X.; Kim, Y.S.; Ponce-Arias, A.-I.; O’Laughlin, R.; Yan, R.Z.; Kobayashi, N.; Tshuva, R.Y.; Tsai, Y.-H.; Sun, S.; Zheng, Y.; et al. A Patterned Human Neural Tube Model Using Microfluidic Gradients. Nature 2024, 628, 391–399. [Google Scholar] [CrossRef]
- Jiang, L.; Guo, K.; Chen, Y.; Xiang, N. Droplet Microfluidics for Current Cancer Research: From Single-Cell Analysis to 3D Cell Culture. ACS Biomater. Sci. Eng. 2024, 10, 1335–1354. [Google Scholar] [CrossRef]
- Abuwatfa, W.H.; Pitt, W.G.; Husseini, G.A. Scaffold-Based 3D Cell Culture Models in Cancer Research. J Biomed Sci 2024, 31, 7. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Xu, X. Droplet Microfluidics for Advanced Single-Cell Analysis. Smart Medicine 2025, 4, e70002. [Google Scholar] [CrossRef]
- Nauwynck, W.; Faust, K.; Boon, N. Droplet Microfluidics for Single-Cell Studies: A Frontier in Ecological Understanding of Microbiomes. FEMS Microbiology Reviews 2025, 49, fuaf032. [Google Scholar] [CrossRef]
- Carneiro, A.; Aranda Palomer, M.; Esteves, M.; Rodrigues, C.; Fernandes, J.M.; Oliveira, F.; Teixeira, A.; Honrado, C.; Dieguez, L.; Abalde-Cela, S.; et al. Advanced Microfluidics for Single Cell-Based Cancer Research. Advanced Science 2025, e00975. [Google Scholar] [CrossRef]
- Rheem, H.B.; Kim, N.; Nguyen, D.T.; Baskoro, G.A.; Roh, J.H.; Lee, J.K.; Kim, B.J.; Choi, I.S. Single-Cell Nanoencapsulation: Chemical Synthesis of Artificial Cell-in-Shell Spores. Chem. Rev. 2025, 125, 6366–6396. [Google Scholar] [CrossRef]
- Chen, A.; Xu, W.; Zhang, X.D.; Lao, J.; Zhao, X.; Milcic, K.; Weitz, D.A. Ultrahigh-Throughput Multiplexed Screening of Purified Protein from Cell-Free Expression Using Droplet Microfluidics. J. Am. Chem. Soc. 2025, 147, 28758–28772. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.M.; Knowles, T.P.J.; Keller, S.; Krainer, G. Microfluidics for Protein Interaction Studies: Current Methods, Challenges, and Future Perspectives. Eur Biophys J 2025. [Google Scholar] [CrossRef]
- Larsson, D.G.J.; Flach, C.-F. Antibiotic Resistance in the Environment. Nat Rev Microbiol 2022, 20, 257–269. [Google Scholar] [CrossRef]
- Darby, E.M.; Trampari, E.; Siasat, P.; Gaya, M.S.; Alav, I.; Webber, M.A.; Blair, J.M.A. Molecular Mechanisms of Antibiotic Resistance Revisited. Nat Rev Microbiol 2023, 21, 280–295. [Google Scholar] [CrossRef]
- Abbas, A.; Barkhouse, A.; Hackenberger, D.; Wright, G.D. Antibiotic Resistance: A Key Microbial Survival Mechanism That Threatens Public Health. Cell Host & Microbe 2024, 32, 837–851. [Google Scholar] [CrossRef] [PubMed]
- Brüssow, H. The Antibiotic Resistance Crisis and the Development of New Antibiotics. Microbial Biotechnology 2024, 17, e14510. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, L.; Huang, Y.; Xu, X.; Liu, Z.; Li, S.; Zhu, L.; Hu, B.; Zhang, T. Global Soil Antibiotic Resistance Genes Are Associated with Increasing Risk and Connectivity to Human Resistome. Nat Commun 2025, 16, 7141. [Google Scholar] [CrossRef]
- Potenza, L.; Krzak, J.; Andrzejewski, M.; Pyzik, A.; Kaminski, T.S. Ultra-High Throughput Droplet Microfluidics for Cultivation and Functional Screening of Environmental Microbial Strains and Consortia. 2025.
- Hinojosa-Ventura, G.; Acosta-Cuevas, J.M.; Velázquez-Carriles, C.A.; Navarro-López, D.E.; López-Alvarez, M.Á.; Ortega-de La Rosa, N.D.; Silva-Jara, J.M. From Basic to Breakthroughs: The Journey of Microfluidic Devices in Hydrogel Droplet Generation. Gels 2025, 11, 309. [Google Scholar] [CrossRef] [PubMed]
- Trinh, T.N.D.; Do, H.D.K.; Nam, N.N.; Dan, T.T.; Trinh, K.T.L.; Lee, N.Y. Droplet-Based Microfluidics: Applications in Pharmaceuticals. Pharmaceuticals 2023, 16, 937. [Google Scholar] [CrossRef]
- Moragues, T.; Arguijo, D.; Beneyton, T.; Modavi, C.; Simutis, K.; Abate, A.R.; Baret, J.-C.; deMello, A.J.; Densmore, D.; Griffiths, A.D. Droplet-Based Microfluidics. Nat Rev Methods Primers 2023, 3, 32. [Google Scholar] [CrossRef]
- Ma, H.; Zhang, Y.; Shen, R.; Jia, Y. Droplet-Based Microfluidics in Single-Bacterium Analysis: Advancements in Cultivation, Detection, and Application. Biosensors 2025, 15, 535. [Google Scholar] [CrossRef]
- Fergola, A.; Ballesio, A.; Frascella, F.; Napione, L.; Cocuzza, M.; Marasso, S.L. Droplet Generation and Manipulation in Microfluidics: A Comprehensive Overview of Passive and Active Strategies. Biosensors 2025, 15, 345. [Google Scholar] [CrossRef]
- Sadasivan, S.; Pradeep, S.; Ramachandran, J.C.; Narayan, J.; Gęca, M.J. Advances in Droplet Microfluidics: A Comprehensive Review of Innovations, Morphology, Dynamics, and Applications. Microfluid Nanofluid 2025, 29, 17. [Google Scholar] [CrossRef]
- Song, Y.; Yin, J.; Huang, W.E.; Li, B.; Yin, H. Emerging Single-Cell Microfluidic Technology for Microbiology. TrAC Trends in Analytical Chemistry 2024, 170, 117444. [Google Scholar] [CrossRef]
- Ripandelli, R.A.A.; Van Oijen, A.M.; Robinson, A. Single-Cell Microfluidics: A Primer for Microbiologists. J. Phys. Chem. B 2024, 128, 10311–10328. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Shen, Q.; Chen, Z.; He, Z.; Yan, X. Harnessing Microfluidic Technology for Bacterial Single-Cell Analysis in Mammals. TrAC Trends in Analytical Chemistry 2023, 166, 117168. [Google Scholar] [CrossRef]
- Zhang, D.; Qiao, L. Microfluidics Coupled Mass Spectrometry for Single Cell Multi-Omics. Small Methods 2024, 8, 2301179. [Google Scholar] [CrossRef]
- Krajewska, J.; Tyski, S.; Laudy, A.E. In Vitro Resistance-Predicting Studies and In Vitro Resistance-Related Parameters—A Hit-to-Lead Perspective. Pharmaceuticals 2024, 17, 1068. [Google Scholar] [CrossRef]
- Zoheir, A.E.; Stolle, C.; Rabe, K.S. Microfluidics for Adaptation of Microorganisms to Stress: Design and Application. Appl Microbiol Biotechnol 2024, 108, 162. [Google Scholar] [CrossRef]
- Xiao, M.; Lv, S.; Zhu, C. Bacterial Patterning: A Promising Biofabrication Technique. ACS Appl. Bio Mater. 2024, 7, 8008–8018. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Lin, H.; Liu, J.; Xin, F.; Chen, M.; Dong, W.; Qian, X.; Jiang, M. Insights into Constructing a Stable and Efficient Microbial Consortium System. Chinese Journal of Chemical Engineering 2024, 76, 95–104. [Google Scholar] [CrossRef]
- Nikoloudaki, O.; Aheto, F.; Di Cagno, R.; Gobbetti, M. Synthetic Microbial Communities: A Gateway to Understanding Resistance, Resilience, and Functionality in Spontaneously Fermented Food Microbiomes. Food Research International 2024, 192, 114780. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Navarro, O.; Aguilar-Salinas, B.; Rocha, J.; Olmedo-Álvarez, G. Higher-Order Interactions and Emergent Properties of Microbial Communities: The Power of Synthetic Ecology. Heliyon 2024, 10, e33896. [Google Scholar] [CrossRef] [PubMed]
- Lyu, X.; Nuhu, M.; Candry, P.; Wolfanger, J.; Betenbaugh, M.; Saldivar, A.; Zuniga, C.; Wang, Y.; Shrestha, S. Top-down and Bottom-up Microbiome Engineering Approaches to Enable Biomanufacturing from Waste Biomass. Journal of Industrial Microbiology and Biotechnology 2024, 51, kuae025. [Google Scholar] [CrossRef]
- Song, X.; Ju, Y.; Chen, L.; Zhang, W. Strategies and Tools to Construct Stable and Efficient Artificial Coculture Systems as Biosynthetic Platforms for Biomass Conversion. Biotechnol Biofuels 2024, 17, 148. [Google Scholar] [CrossRef]
- Abouhagger, A.; Celiešiūtė-Germanienė, R.; Bakute, N.; Stirke, A.; Melo, W.C.M.A. Electrochemical Biosensors on Microfluidic Chips as Promising Tools to Study Microbial Biofilms: A Review. Front. Cell. Infect. Microbiol. 2024, 14, 1419570. [Google Scholar] [CrossRef]
- Dou, J.; Yang, Z.; Singh, B.; Ma, B.; Lu, Z.; Xu, J.; He, Y. Discussion: Embracing Microfluidics to Advance Environmental Science and Technology. Science of The Total Environment 2024, 937, 173597. [Google Scholar] [CrossRef] [PubMed]
- Ugolini, G.S.; Wang, M.; Secchi, E.; Pioli, R.; Ackermann, M.; Stocker, R. Microfluidic Approaches in Microbial Ecology. Lab Chip 2024, 24, 1394–1418. [Google Scholar] [CrossRef]
- Kelliher, J.M.; Johnson, L.Y.D.; Robinson, A.J.; Longley, R.; Hanson, B.T.; Cailleau, G.; Bindschedler, S.; Junier, P.; Chain, P.S.G. Fabricated Devices for Performing Bacterial-Fungal Interaction Experiments across Scales. Front. Microbiol. 2024, 15, 1380199. [Google Scholar] [CrossRef] [PubMed]
- Diep Trinh, T.N.; Trinh, K.T.L.; Lee, N.Y. Microfluidic Advances in Food Safety Control. Food Research International 2024, 176, 113799. [Google Scholar] [CrossRef]
- Ranjbaran, M.; Verma, M.S. Microfluidics at the Interface of Bacteria and Fresh Produce. Trends in Food Science & Technology 2022, 128, 102–117. [Google Scholar] [CrossRef]
- Jin, Y.; Dou, M.; Zhuo, S.; Li, Q.; Wang, F.; Li, J. Advances in Microfluidic Analysis of Residual Antibiotics in Food. Food Control 2022, 136, 108885. [Google Scholar] [CrossRef]
- Masters, E.A.; De Mesy Bentley, K.L.; Gill, A.L.; Hao, S.P.; Galloway, C.A.; Salminen, A.T.; Guy, D.R.; McGrath, J.L.; Awad, H.A.; Gill, S.R.; et al. Identification of Penicillin Binding Protein 4 (PBP4) as a Critical Factor for Staphylococcus Aureus Bone Invasion during Osteomyelitis in Mice. PLoS Pathog 2020, 16, e1008988. [Google Scholar] [CrossRef]
- Terekhov, S.S.; Smirnov, I.V.; Stepanova, A.V.; Bobik, T.V.; Mokrushina, Y.A.; Ponomarenko, N.A.; Belogurov, A.A.; Rubtsova, M.P.; Kartseva, O.V.; Gomzikova, M.O.; et al. Microfluidic Droplet Platform for Ultrahigh-Throughput Single-Cell Screening of Biodiversity. Proc. Natl. Acad. Sci. U.S.A. 2017, 114, 2550–2555. [Google Scholar] [CrossRef]
- Baranova, M.N.; Babikova, P.A.; Kudzhaev, A.M.; Mokrushina, Y.A.; Belozerova, O.A.; Yunin, M.A.; Kovalchuk, S.; Gabibov, A.G.; Smirnov, I.V.; Terekhov, S.S. Live Biosensors for Ultrahigh-Throughput Screening of Antimicrobial Activity against Gram-Negative Bacteria. Antibiotics 2021, 10, 1161. [Google Scholar] [CrossRef]
- Terekhov, S.S.; Nazarov, A.S.; Mokrushina, Y.A.; Baranova, M.N.; Potapova, N.A.; Malakhova, M.V.; Ilina, E.N.; Smirnov, I.V.; Gabibov, A.G. Deep Functional Profiling Facilitates the Evaluation of the Antibacterial Potential of the Antibiotic Amicoumacin. Antibiotics 2020, 9, 157. [Google Scholar] [CrossRef] [PubMed]
- Baranova, M.N.; Kudzhaev, A.M.; Mokrushina, Y.A.; Babenko, V.V.; Kornienko, M.A.; Malakhova, M.V.; Yudin, V.G.; Rubtsova, M.P.; Zalevsky, A.; Belozerova, O.A.; et al. Deep Functional Profiling of Wild Animal Microbiomes Reveals Probiotic Bacillus Pumilus Strains with a Common Biosynthetic Fingerprint. IJMS 2022, 23, 1168. [Google Scholar] [CrossRef]
- Baranova, M.N.; Soboleva, E.A.; Kornienko, M.A.; Malakhova, M.V.; Mokrushina, Yu.A.; Gabibov, A.G.; Terekhov, S.S.; Smirnov, I.V. Bacteriocin from the Raccoon Dog Oral Microbiota Inhibits the Growth of Pathogenic Methicillin-Resistant Staphylococcus Aureus. Acta Naturae 2024, 16, 105–108. [Google Scholar] [CrossRef]
- McCully, A.L.; Loop Yao, M.; Brower, K.K.; Fordyce, P.M.; Spormann, A.M. Double Emulsions as a High-Throughput Enrichment and Isolation Platform for Slower-Growing Microbes. ISME Communications 2023, 3, 47. [Google Scholar] [CrossRef]
- Ochoa, A.; Gastélum, G.; Rocha, J.; Olguin, L.F. High-Throughput Bacterial Co-Encapsulation in Microfluidic Gel Beads for Discovery of Antibiotic-Producing Strains. Analyst 2023, 148, 5762–5774. [Google Scholar] [CrossRef] [PubMed]
- Mahler, L.; Niehs, S.P.; Martin, K.; Weber, T.; Scherlach, K.; Hertweck, C.; Roth, M.; Rosenbaum, M.A. Highly Parallelized Droplet Cultivation and Prioritization of Antibiotic Producers from Natural Microbial Communities. eLife 2021, 10. [Google Scholar] [CrossRef]
- Jung, H.; Zhang, H.; Hooper, J.; Huang, C.; Gupte, R.; Guzman, A.; Han, J.J.; Han, A. Size-Independent and Automated Single-Colony-Resolution Microdroplet Dispensing. Lab Chip 2025. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Han, X.; Wu, L.; Sun, B.; Li, G. A Digital Plating Platform for Robust and Versatile Microbial Detection and Analysis. Sci Rep 2025, 15, 25301. [Google Scholar] [CrossRef]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.P.; Mueller, A.; Schaberle, T.F.; Hughes, D.E.; Epstein, S.; et al. A New Antibiotic Kills Pathogens without Detectable Resistance. Nature 2015, 517, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Alkayyali, T.; Pope, E.; Wheatley, S.K.; Cartmell, C.; Haltli, B.; Kerr, R.G.; Ahmadi, A. Development of a Microbe Domestication Pod (MD Pod) for in Situ Cultivation of Micro-encapsulated Marine Bacteria. Biotech & Bioengineering 2021, 118, 1166–1176. [Google Scholar] [CrossRef]
- Wheatley, S.K.; Cartmell, C.; Madadian, E.; Badr, S.; Haltli, B.A.; Kerr, R.G.; Ahmadi, A. Microfabrication of a Micron-Scale Microbial-Domestication Pod for in Situ Cultivation of Marine Bacteria. RSC Adv. 2022, 12, 28123–28127. [Google Scholar] [CrossRef]
- Dai, J.; Ouyang, Y.; Gupte, R.; Liu, X.J.A.; Li, Y.; Yang, F.; Chen, S.; Provin, T.; Van Schaik, E.; Samuel, J.E.; et al. Microfluidic Droplets with Amended Culture Media Cultivate a Greater Diversity of Soil Microorganisms. Appl Environ Microbiol 2025, 91, e01794–24. [Google Scholar] [CrossRef]
- Hua, E.; Zhang, Y.; Yun, K.; Pan, W.; Liu, Y.; Li, S.; Wang, Y.; Tu, R.; Wang, M. Whole-Cell Biosensor and Producer Co-Cultivation-Based Microfludic Platform for Screening Saccharopolyspora Erythraea with Hyper Erythromycin Production. ACS Synth. Biol. 2022, 11, 2697–2708. [Google Scholar] [CrossRef]
- Wang, Y.; Li, S.; Xue, N.; Wang, L.; Zhang, X.; Zhao, L.; Guo, Y.; Zhang, Y.; Wang, M. Modulating Sensitivity of an Erythromycin Biosensor for Precise High-Throughput Screening of Strains with Different Characteristics. ACS Synth. Biol. 2023, 12, 1761–1771. [Google Scholar] [CrossRef]
- Zhao, L.; Li, S.; Yang, Y.; Qi, L.; Zhu, Q.; Zhao, Y.; Qi, H.; Liao, X.; Zhang, Y.; Wang, M. Biosensor-Based Dual-Color Droplet Microfluidic Platform for Precise High-Throughput Screening of Erythromycin Hyperproducers. Biosensors and Bioelectronics 2025, 278, 117376. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Li, G.; Zhou, L.; Guo, Y.; Tu, Q.; Wang, J. On-Chip Screening of Chemically Induced Mutant Strains and Assay of Their Metabolites by Liquid Chromatography Tandem Mass Spectrometry. Sensors and Actuators B: Chemical 2021, 348, 130655. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Z.; Pang, Y. Concentration Gradient Generation Methods Based on Microfluidic Systems. RSC Adv. 2017, 7, 29966–29984. [Google Scholar] [CrossRef]
- Shen, S.; Zhang, F.; Zhang, Y.; Li, Y.; Niu, Y.; Pang, L.; Wang, J. Construction of Multiple Concentration Gradients for Single-Cell Level Drug Screening. Microsyst Nanoeng 2023, 9, 46. [Google Scholar] [CrossRef]
- Zhao, M.; Yang, J.; Li, Z.; Zeng, Y.; Tao, C.; Dai, B.; Zhang, D.; Yamaguchi, Y. High-Throughput 3D Microfluidic Chip for Generation of Concentration Gradients and Mixture Combinations. Lab Chip 2024, 24, 2280–2286. [Google Scholar] [CrossRef]
- Azizi, M.; Davaji, B.; Nguyen, A.V.; Mokhtare, A.; Zhang, S.; Dogan, B.; Gibney, P.A.; Simpson, K.W.; Abbaspourrad, A. Biological Small-Molecule Assays Using Gradient-Based Microfluidics. Biosensors and Bioelectronics 2021, 178, 113038. [Google Scholar] [CrossRef]
- Sweet, E.; Yang, B.; Chen, J.; Vickerman, R.; Lin, Y.; Long, A.; Jacobs, E.; Wu, T.; Mercier, C.; Jew, R.; et al. 3D Microfluidic Gradient Generator for Combination Antimicrobial Susceptibility Testing. Microsyst Nanoeng 2020, 6, 92. [Google Scholar] [CrossRef]
- Wang, J.; Gong, G.; Hu, W.; Lv, H.; Li, Q.; Li, X.; Sun, X. Recent Advances of Concentration Gradient Microfluidic Chips for Drug Screening. BioChip J 2025, 19, 496–544. [Google Scholar] [CrossRef]
- Zhang, K.; Qin, S.; Wu, S.; Liang, Y.; Li, J. Microfluidic Systems for Rapid Antibiotic Susceptibility Tests (ASTs) at the Single-Cell Level. Chem. Sci. 2020, 11, 6352–6361. [Google Scholar] [CrossRef]
- Dietvorst, J.; Vilaplana, L.; Uria, N.; Marco, M.-P.; Muñoz-Berbel, X. Current and Near-Future Technologies for Antibiotic Susceptibility Testing and Resistant Bacteria Detection. TrAC Trends in Analytical Chemistry 2020, 127, 115891. [Google Scholar] [CrossRef]
- Trinh, T.N.D.; Lee, N.Y. Nucleic Acid Amplification-Based Microfluidic Approaches for Antimicrobial Susceptibility Testing. Analyst 2021, 146, 3101–3113. [Google Scholar] [CrossRef]
- Hsieh, K.; Mach, K.E.; Zhang, P.; Liao, J.C.; Wang, T.-H. Combating Antimicrobial Resistance via Single-Cell Diagnostic Technologies Powered by Droplet Microfluidics. Acc. Chem. Res. 2022, 55, 123–133. [Google Scholar] [CrossRef]
- Ruszczak, A.; Bartkova, S.; Zapotoczna, M.; Scheler, O.; Garstecki, P. Droplet-Based Methods for Tackling Antimicrobial Resistance. Current Opinion in Biotechnology 2022, 76, 102755. [Google Scholar] [CrossRef]
- Postek, W.; Pacocha, N.; Garstecki, P. Microfluidics for Antibiotic Susceptibility Testing. Lab Chip 2022, 22, 3637–3662. [Google Scholar] [CrossRef] [PubMed]
- Aubry, G.; Lee, H.J.; Lu, H. Advances in Microfluidics: Technical Innovations and Applications in Diagnostics and Therapeutics. Anal. Chem. 2023, 95, 444–467. [Google Scholar] [CrossRef] [PubMed]
- Ardila, C.M.; Jiménez-Arbeláez, G.A.; Vivares-Builes, A.M. The Potential Clinical Applications of a Microfluidic Lab-on-a-Chip for the Identification and Antibiotic Susceptibility Testing of Enterococcus Faecalis-Associated Endodontic Infections: A Systematic Review. Dentistry Journal 2023, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Lehnert, T.; Gijs, M.A.M. Microfluidic Systems for Infectious Disease Diagnostics. Lab Chip 2024, 24, 1441–1493. [Google Scholar] [CrossRef]
- Zhang, J.; Fu, Y.; Fong, C.Y.; Hua, H.; Li, W.; Khoo, B.L. Advancements in Microfluidic Technology for Rapid Bacterial Detection and Inflammation-Driven Diseases. Lab Chip 2025, 25, 3348–3373. [Google Scholar] [CrossRef]
- Yabbarov, N.G.; Nikolskaya, E.D.; Bibikov, S.B.; Maltsev, A.A.; Chirkina, M.V.; Mollaeva, M.R.; Sokol, M.B.; Epova, E.Yu.; Aliev, R.O.; Kurochkin, I.N. Methods for Rapid Evaluation of Microbial Antibiotics Resistance. Biochemistry Moscow 2025, 90, S312–S341. [Google Scholar] [CrossRef] [PubMed]
- Ahmed Nawaz Qureshi, Y.Z.; Li, M.; Chang, H.; Song, Y. Microfluidic Chip Systems for Color-Based Antimicrobial Susceptibility Test a Review. Biosensors and Bioelectronics 2025, 273, 117160. [Google Scholar] [CrossRef]
- Rastmanesh, S.; Zeinaly, I.; Alivirdiloo, V.; Mobed, A.; Darvishi, M. Biosensing for Rapid Detection of MDR, XDR and PDR Bacteria. Clinica Chimica Acta 2025, 567, 120121. [Google Scholar] [CrossRef] [PubMed]
- Duan, B.; Zeng, X.; Peng, J. Advances in Genotypic Antimicrobialresistance Testing: A Comprehensive Review. Sci. China Life Sci. 2025, 68, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Guzman, A.R.; Wippold, J.A.; Li, Y.; Dai, J.; Huang, C.; Han, A. An Ultra High-Efficiency Droplet Microfluidics Platform Using Automatically Synchronized Droplet Pairing and Merging. Lab Chip 2020, 20, 3948–3959. [Google Scholar] [CrossRef]
- Samimi, A.; Hengoju, S.; Rosenbaum, M.A. Combinatorial Sample Preparation Platform for Droplet-Based Applications in Microbiology. Sensors and Actuators B: Chemical 2024, 417, 136162. [Google Scholar] [CrossRef]
- Fukuda, R.; Cira, N.J. High-Throughput, Combinatorial Droplet Generation by Sequential Spraying. Lab Chip 2025, 25, 1502–1511. [Google Scholar] [CrossRef]
- Scheler, O.; Makuch, K.; Debski, P.R.; Horka, M.; Ruszczak, A.; Pacocha, N.; Sozański, K.; Smolander, O.-P.; Postek, W.; Garstecki, P. Droplet-Based Digital Antibiotic Susceptibility Screen Reveals Single-Cell Clonal Heteroresistance in an Isogenic Bacterial Population. Sci Rep 2020, 10, 3282. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, J.; Kim, J.-S.; Kim, W.; Lee, C.-S. Label-Free Single-Cell Antimicrobial Susceptibility Testing in Droplets with Concentration Gradient Generation. Lab Chip 2024, 24, 5274–5289. [Google Scholar] [CrossRef]
- Ruszczak, A.; Jankowski, P.; Vasantham, S.K.; Scheler, O.; Garstecki, P. Physicochemical Properties Predict Retention of Antibiotics in Water-in-Oil Droplets. Anal. Chem. 2023. [Google Scholar] [CrossRef] [PubMed]
- Heuberger, L.; Messmer, D.; Dos Santos, E.C.; Scherrer, D.; Lörtscher, E.; Schoenenberger, C.; Palivan, C.G. Microfluidic Giant Polymer Vesicles Equipped with Biopores for High-Throughput Screening of Bacteria. Advanced Science 2024, 11, 2307103. [Google Scholar] [CrossRef]
- Shao, F.; Hsieh, K.; Zhang, P.; Kaushik, A.M.; Wang, T.-H. Facile and Scalable Tubing-Free Sample Loading for Droplet Microfluidics. Sci Rep 2022, 12, 13340. [Google Scholar] [CrossRef]
- Yang, D.; Yu, Z.; Zheng, M.; Yang, W.; Liu, Z.; Zhou, J.; Huang, L. Artificial Intelligence-Accelerated High-Throughput Screening of Antibiotic Combinations on a Microfluidic Combinatorial Droplet System. Lab Chip 2023, 23, 3961–3977. [Google Scholar] [CrossRef]
- Li, H.; Zhang, P.; Hsieh, K.; Wang, T.-H. Combinatorial Nanodroplet Platform for Screening Antibiotic Combinations. Lab Chip 2022, 22, 621–631. [Google Scholar] [CrossRef]
- Shao, F.; Li, H.; Hsieh, K.; Zhang, P.; Li, S.; Wang, T.-H. Automated and Miniaturized Screening of Antibiotic Combinations via Robotic-Printed Combinatorial Droplet Platform. Acta Pharmaceutica Sinica B 2024, 14, 1801–1813. [Google Scholar] [CrossRef]
- Parsley, N.C.; Smythers, A.L.; Hicks, L.M. Implementation of Microfluidics for Antimicrobial Susceptibility Assays: Issues and Optimization Requirements. Front. Cell. Infect. Microbiol. 2020, 10, 547177. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Yu, Z.; Lyu, W.; Luo, Y.; Xu, L.; Shen, F. Digital Bioassays on the Slipchip Microfluidic Devices. Advanced Sensor Research 2025, 4, e00030. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, M.; Lyu, W.; Li, X.; Xu, L.; Qin, Y.; Ren, Y.; Deng, Z.; Tao, M.; Xiao, W.; et al. Rapid High-Throughput Discovery of Molecules With Antimicrobial Activity From Natural Products Enabled by a Nanoliter Matrix SlipChip. Small Methods 2025, 9, 2402045. [Google Scholar] [CrossRef] [PubMed]
- Pipiya, S.O.; Terekhov, S.S.; Mokrushina, Yu.A.; Knorre, V.D.; Smirnov, I.V.; Gabibov, A.G. Engineering Artificial Biodiversity of Lantibiotics to Expand Chemical Space of DNA-Encoded Antibiotics. Biochemistry Moscow 2020, 85, 1319–1334. [Google Scholar] [CrossRef]
- Pipiya, S.O.; Mirzoeva, N.Z.; Baranova, M.N.; Eliseev, I.E.; Mokrushina, Y.A.; Shamova, O.V.; Gabibov, A.G.; Smirnov, I.V.; Terekhov, S.S. Creation of Recombinant Biocontrol Agents by Genetic Programming of Yeast. Acta Naturae 2023, 15, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Nuti, N.; Rottmann, P.; Stucki, A.; Koch, P.; Panke, S.; Dittrich, P.S. A Multiplexed Cell-Free Assay to Screen for Antimicrobial Peptides in Double Emulsion Droplets. Angew Chem Int Ed 2022, 61, e202114632. [Google Scholar] [CrossRef] [PubMed]
- Pourmasoumi, F.; Hengoju, S.; Beck, K.; Stephan, P.; Klopfleisch, L.; Hoernke, M.; Rosenbaum, M.A.; Kries, H. Analysing Megasynthetase Mutants at High Throughput Using Droplet Microfluidics**. ChemBioChem 2023, 24, e202300680. [Google Scholar] [CrossRef]
- Troiano, C.; De Ninno, A.; Casciaro, B.; Riccitelli, F.; Park, Y.; Businaro, L.; Massoud, R.; Mangoni, M.L.; Bisegna, P.; Stella, L.; et al. Rapid Assessment of Susceptibility of Bacteria and Erythrocytes to Antimicrobial Peptides by Single-Cell Impedance Cytometry. ACS Sens. 2023, 8, 2572–2582. [Google Scholar] [CrossRef]
- Panteleev, V.; Kulbachinskiy, A.; Gelfenbein, D. Evaluating Phage Lytic Activity: From Plaque Assays to Single-Cell Technologies. Front. Microbiol. 2025, 16, 1659093. [Google Scholar] [CrossRef]
- Hoshino, M.; Ota, Y.; Suyama, T.; Morishita, Y.; Tsuneda, S.; Noda, N. Water-in-Oil Droplet-Mediated Method for Detecting and Isolating Infectious Bacteriophage Particles via Fluorescent Staining. Front. Microbiol. 2023, 14, 1282372. [Google Scholar] [CrossRef]
- Zhang, H.; Boeckman, J.; Gupte, R.; Prejean, A.; Miller, J.; Rodier, A.; De Figueiredo, P.; Liu, M.; Gill, J.; Han, A. PRISM: A Platform for Illuminating Viral Dark Matter 2025.
- Givelet, L.; Von Schönberg, S.; Katzmeier, F.; Simmel, F.C. Digital Phage Biology in Droplets 2025.
- Li, X.; Hu, Q.; Liu, X.; Liu, J.; Wu, N.; Shen, F. Rapid Bacteriophage Quantification by Digital Biosensing on a SlipChip Microfluidic Device. Anal. Chem. 2023, 95, 8632–8639. [Google Scholar] [CrossRef]
- Sidi Mabrouk, A.; Ongenae, V.; Claessen, D.; Brenzinger, S.; Briegel, A. A Flexible and Efficient Microfluidics Platform for the Characterization and Isolation of Novel Bacteriophages. Appl Environ Microbiol 2023, 89, e01596-22. [Google Scholar] [CrossRef] [PubMed]
- Benoit, M.R.; Conant, C.G.; Ionescu-Zanetti, C.; Schwartz, M.; Matin, A. New Device for High-Throughput Viability Screening of Flow Biofilms. Appl Environ Microbiol 2010, 76, 4136–4142. [Google Scholar] [CrossRef]
- Di Somma, A.; Recupido, F.; Cirillo, A.; Romano, A.; Romanelli, A.; Caserta, S.; Guido, S.; Duilio, A. Antibiofilm Properties of Temporin-L on Pseudomonas Fluorescens in Static and In-Flow Conditions. IJMS 2020, 21, 8526. [Google Scholar] [CrossRef]
- Díez-Aguilar, M.; Ekkelenkamp, M.; Morosini, M.-I.; Huertas, N.; Del Campo, R.; Zamora, J.; Fluit, A.C.; Tunney, M.M.; Obrecht, D.; Bernardini, F.; et al. Anti-Biofilm Activity of Murepavadin against Cystic Fibrosis Pseudomonas Aeruginosa Isolates. Journal of Antimicrobial Chemotherapy 2021, 76, 2578–2585. [Google Scholar] [CrossRef]
- Duda-Madej, A.; Kozłowska, J.; Baczyńska, D.; Krzyżek, P. Ether Derivatives of Naringenin and Their Oximes as Factors Modulating Bacterial Adhesion. Antibiotics 2023, 12, 1076. [Google Scholar] [CrossRef]
- Czechowicz, P.; Nowicka, J.; Neubauer, D.; Chodaczek, G.; Krzyżek, P.; Gościniak, G. Activity of Novel Ultrashort Cyclic Lipopeptides against Biofilm of Candida Albicans Isolated from VVC in the Ex Vivo Animal Vaginal Model and BioFlux Biofilm Model—A Pilot Study. IJMS 2022, 23, 14453. [Google Scholar] [CrossRef]
- Ladewig, L.; Gloy, L.; Langfeldt, D.; Pinnow, N.; Weiland-Bräuer, N.; Schmitz, R.A. Antimicrobial Peptides Originating from Expression Libraries of Aurelia Aurita and Mnemiopsis Leidyi Prevent Biofilm Formation of Opportunistic Pathogens. Microorganisms 2023, 11, 2184. [Google Scholar] [CrossRef]
- Straub, H.; Eberl, L.; Zinn, M.; Rossi, R.M.; Maniura-Weber, K.; Ren, Q. A Microfluidic Platform for in Situ Investigation of Biofilm Formation and Its Treatment under Controlled Conditions. J Nanobiotechnol 2020, 18, 166. [Google Scholar] [CrossRef]
- McLeod, D.; Wei, L.; Li, Z. A Standard 96-Well Based High Throughput Microfluidic Perfusion Biofilm Reactor for in Situ Optical Analysis. Biomed Microdevices 2023, 25, 26. [Google Scholar] [CrossRef] [PubMed]
- Pouget, C.; Pantel, A.; Dunyach-Remy, C.; Magnan, C.; Sotto, A.; Lavigne, J.-P. Antimicrobial Activity of Antibiotics on Biofilm Formed by Staphylococcus Aureus and Pseudomonas Aeruginosa in an Open Microfluidic Model Mimicking the Diabetic Foot Environment. Journal of Antimicrobial Chemotherapy 2023, 78, 540–545. [Google Scholar] [CrossRef]
- Tran, V.N.; Khan, F.; Han, W.; Luluil, M.; Truong, V.G.; Yun, H.G.; Choi, S.; Kim, Y.-M.; Shin, J.H.; Kang, H.W. Real-Time Monitoring of Mono- and Dual-Species Biofilm Formation and Eradication Using Microfluidic Platform. Sci Rep 2022, 12, 9678. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Cabra, N.; López-Martínez, M.J.; Arévalo-Jaimes, B.V.; Martin-Gómez, M.T.; Samitier, J.; Torrents, E. A New BiofilmChip Device for Testing Biofilm Formation and Antibiotic Susceptibility. npj Biofilms Microbiomes 2021, 7, 62. [Google Scholar] [CrossRef] [PubMed]
- Senevirathne, S.W.M.A.I.; Mathew, A.; Toh, Y.-C.; Yarlagadda, P.K.D.V. Bactericidal Efficacy of Nanostructured Surfaces Increases under Flow Conditions. ACS Omega 2022, 7, 41711–41722. [Google Scholar] [CrossRef]
- Hansson, A.; Karlsen, E.A.; Stensen, W.; Svendsen, J.S.M.; Berglin, M.; Lundgren, A. Preventing E. Coli Biofilm Formation with Antimicrobial Peptide-Functionalized Surface Coatings: Recognizing the Dependence on the Bacterial Binding Mode Using Live-Cell Microscopy. ACS Appl. Mater. Interfaces 2024, 16, 6799–6812. [Google Scholar] [CrossRef]
- Wang, P.; Robert, L.; Pelletier, J.; Dang, W.L.; Taddei, F.; Wright, A.; Jun, S. Robust Growth of Escherichia Coli. Current Biology 2010, 20, 1099–1103. [Google Scholar] [CrossRef]
- Allard, P.; Papazotos, F.; Potvin-Trottier, L. Microfluidics for Long-Term Single-Cell Time-Lapse Microscopy: Advances and Applications. Front. Bioeng. Biotechnol. 2022, 10, 968342. [Google Scholar] [CrossRef]
- Yokoyama, F.; Kling, A.; Dittrich, P.S. Capturing of Extracellular Vesicles Derived from Single Cells of Escherichia Coli. Lab Chip 2024, 24, 2049–2057. [Google Scholar] [CrossRef] [PubMed]
- Chikada, T.; Kanai, T.; Hayashi, M.; Kasai, T.; Oshima, T.; Shiomi, D. Direct Observation of Conversion From Walled Cells to Wall-Deficient L-Form and Vice Versa in Escherichia Coli Indicates the Essentiality of the Outer Membrane for Proliferation of L-Form Cells. Front. Microbiol. 2021, 12, 645965. [Google Scholar] [CrossRef] [PubMed]
- Mistretta, M.; Cimino, M.; Campagne, P.; Volant, S.; Kornobis, E.; Hebert, O.; Rochais, C.; Dallemagne, P.; Lecoutey, C.; Tisnerat, C.; et al. Dynamic Microfluidic Single-Cell Screening Identifies Pheno-Tuning Compounds to Potentiate Tuberculosis Therapy. Nat Commun 2024, 15, 4175. [Google Scholar] [CrossRef] [PubMed]
- Nikolic, N.; Anagnostidis, V.; Tiwari, A.; Chait, R.; Gielen, F. Droplet-Based Methodology for Investigating Bacterial Population Dynamics in Response to Phage Exposure. Front. Microbiol. 2023, 14, 1260196. [Google Scholar] [CrossRef] [PubMed]
- Le Quellec, L.; Aristov, A.; Gutiérrez Ramos, S.; Amselem, G.; Bos, J.; Baharoglu, Z.; Mazel, D.; Baroud, C.N. Measuring Single-Cell Susceptibility to Antibiotics within Monoclonal Bacterial Populations. PLoS ONE 2024, 19, e0303630. [Google Scholar] [CrossRef]
- Cama, J.; Al Nahas, K.; Fletcher, M.; Hammond, K.; Ryadnov, M.G.; Keyser, U.F.; Pagliara, S. An Ultrasensitive Microfluidic Approach Reveals Correlations between the Physico-Chemical and Biological Activity of Experimental Peptide Antibiotics. Sci Rep 2022, 12, 4005. [Google Scholar] [CrossRef]
- Conners, R.; McLaren, M.; Łapińska, U.; Sanders, K.; Stone, M.R.L.; Blaskovich, M.A.T.; Pagliara, S.; Daum, B.; Rakonjac, J.; Gold, V.A.M. CryoEM Structure of the Outer Membrane Secretin Channel pIV from the F1 Filamentous Bacteriophage. Nat Commun 2021, 12, 6316. [Google Scholar] [CrossRef]
- Cama, J.; Voliotis, M.; Metz, J.; Smith, A.; Iannucci, J.; Keyser, U.F.; Tsaneva-Atanasova, K.; Pagliara, S. Single-Cell Microfluidics Facilitates the Rapid Quantification of Antibiotic Accumulation in Gram-Negative Bacteria. Lab Chip 2020, 20, 2765–2775. [Google Scholar] [CrossRef]
- Zhang, Y.; Kepiro, I.; Ryadnov, M.G.; Pagliara, S. Single Cell Killing Kinetics Differentiate Phenotypic Bacterial Responses to Different Antibacterial Classes. Microbiol Spectr 2023, 11, e03667-22. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Phetsang, W.; Stone, M.R.L.; Kc, S.; Butler, M.S.; Cooper, M.A.; Elliott, A.G.; Łapińska, U.; Voliotis, M.; Tsaneva-Atanasova, K.; et al. Synthesis of Vancomycin Fluorescent Probes That Retain Antimicrobial Activity, Identify Gram-Positive Bacteria, and Detect Gram-Negative Outer Membrane Damage. Commun Biol 2023, 6, 409. [Google Scholar] [CrossRef]
- Wong, F.; Stokes, J.M.; Cervantes, B.; Penkov, S.; Friedrichs, J.; Renner, L.D.; Collins, J.J. Cytoplasmic Condensation Induced by Membrane Damage Is Associated with Antibiotic Lethality. Nat Commun 2021, 12, 2321. [Google Scholar] [CrossRef]
- Li, X.; Song, Y.; Chen, X.; Yin, J.; Wang, P.; Huang, H.; Yin, H. Single-Cell Microfluidics Enabled Dynamic Evaluation of Drug Combinations on Antibiotic Resistance Bacteria. Talanta 2023, 265, 124814. [Google Scholar] [CrossRef]
- Meza-Torres, J.; Lelek, M.; Quereda, J.J.; Sachse, M.; Manina, G.; Ershov, D.; Tinevez, J.-Y.; Radoshevich, L.; Maudet, C.; Chaze, T.; et al. Listeriolysin S: A Bacteriocin from Listeria Monocytogenes That Induces Membrane Permeabilization in a Contact-Dependent Manner. Proc. Natl. Acad. Sci. U.S.A. 2021, 118, e2108155118. [Google Scholar] [CrossRef]
- Mistretta, M.; Gangneux, N.; Manina, G. Microfluidic Dose–Response Platform to Track the Dynamics of Drug Response in Single Mycobacterial Cells. Sci Rep 2022, 12, 19578. [Google Scholar] [CrossRef]
- Strutt, R.; Jusková, P.; Berlanda, S.F.; Krämer, S.D.; Dittrich, P.S. Engineering a Biohybrid System to Link Antibiotic Efficacy to Membrane Depth in Bacterial Infections. Small 2025, 21, 2412399. [Google Scholar] [CrossRef]
- Barlow, N.E.; Bolognesi, G.; Flemming, A.J.; Brooks, N.J.; Barter, L.M.C.; Ces, O. Multiplexed Droplet Interface Bilayer Formation. Lab Chip 2016, 16, 4653–4657. [Google Scholar] [CrossRef] [PubMed]
- Yahyazadeh Shourabi, A.; Iacona, M.; Aubin-Tam, M.-E. Microfluidic System for Efficient Molecular Delivery to Artificial Cell Membranes. Lab Chip 2025, 25, 1842–1853. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Ghorbanpoor, H.; Öztürk, Y.; Kaygusuz, Ö.; Avcı, H.; Darcan, C.; Trabzon, L.; Güzel, F.D. On-chip Label-free Impedance-based Detection of Antibiotic Permeation. IET Nanobiotechnology 2021, 15, 100–106. [Google Scholar] [CrossRef]
- Xu, T.; Dai, Y.; Ge, A.; Chen, X.; Gong, Y.; Lam, T.H.; Lee, K.; Han, X.; Ji, Y.; Shen, W.; et al. Ultrafast Evolution of Bacterial Antimicrobial Resistance by Picoliter-Scale Centrifugal Microfluidics. Anal. Chem. 2024, 96, 18842–18851. [Google Scholar] [CrossRef]
- Disney-McKeethen, S.; Seo, S.; Mehta, H.; Ghosh, K.; Shamoo, Y. Experimental Evolution of Pseudomonas Aeruginosa to Colistin in Spatially Confined Microdroplets Identifies Evolutionary Trajectories Consistent with Adaptation in Microaerobic Lung Environments. mBio 2023, 14, e01506-23. [Google Scholar] [CrossRef]
- Seo, S.; Disney-McKeethen, S.; Prabhakar, R.G.; Song, X.; Mehta, H.H.; Shamoo, Y. Identification of Evolutionary Trajectories Associated with Antimicrobial Resistance Using Microfluidics. ACS Infect. Dis. 2022, 8, 242–254. [Google Scholar] [CrossRef]
- Zoheir, A.E.; Späth, G.P.; Niemeyer, C.M.; Rabe, K.S. Microfluidic Evolution-On-A-Chip Reveals New Mutations That Cause Antibiotic Resistance. Small 2021, 17, 2007166. [Google Scholar] [CrossRef]
- Nagy, K.; Dukic, B.; Hodula, O.; Ábrahám, Á.; Csákvári, E.; Dér, L.; Wetherington, M.T.; Noorlag, J.; Keymer, J.E.; Galajda, P. Emergence of Resistant Escherichia Coli Mutants in Microfluidic On-Chip Antibiotic Gradients. Front. Microbiol. 2022, 13, 820738. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Larsson, J.; Hjort, K.; Elf, J.; Andersson, D.I. The Highly Dynamic Nature of Bacterial Heteroresistance Impairs Its Clinical Detection. Commun Biol 2021, 4, 521. [Google Scholar] [CrossRef]
- Daniel, S.; Goldlust, K.; Quebre, V.; Shen, M.; Lesterlin, C.; Bouet, J.-Y.; Yamaichi, Y. Vertical and Horizontal Transmission of ESBL Plasmid from Escherichia Coli O104:H4. Genes 2020, 11, 1207. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, Q.; Li, M.; Li, R.; He, Z.; Dechesne, A.; Smets, B.F.; Sheng, G. Single-Cell Analysis Reveals Antibiotic Affects Conjugative Transfer by Modulating Bacterial Growth Rather than Conjugation Efficiency. Environment International 2025, 198, 109385. [Google Scholar] [CrossRef] [PubMed]
- Elitas, M.; Dhar, N.; McKinney, J.D. Revealing Antibiotic Tolerance of the Mycobacterium Smegmatis Xanthine/Uracil Permease Mutant Using Microfluidics and Single-Cell Analysis. Antibiotics 2021, 10, 794. [Google Scholar] [CrossRef] [PubMed]
- Fanous, J.; Claudi, B.; Tripathi, V.; Li, J.; Goormaghtigh, F.; Bumann, D. Limited Impact of Salmonella Stress and Persisters on Antibiotic Clearance. Nature 2025, 639, 181–189. [Google Scholar] [CrossRef]
- Umetani, M.; Fujisawa, M.; Okura, R.; Nozoe, T.; Suenaga, S.; Nakaoka, H.; Kussell, E.; Wakamoto, Y. Observation of Persister Cell Histories Reveals Diverse Modes of Survival in Antibiotic Persistence. eLife 2025, 14, e79517. [Google Scholar] [CrossRef]
- Morawska, L.P.; Kuipers, O.P. Antibiotic Tolerance in Environmentally Stressed Bacillus Subtilis : Physical Barriers and Induction of a Viable but Nonculturable State. microLife 2022, 3, uqac010. [Google Scholar] [CrossRef]
- Manuse, S.; Shan, Y.; Canas-Duarte, S.J.; Bakshi, S.; Sun, W.-S.; Mori, H.; Paulsson, J.; Lewis, K. Bacterial Persisters Are a Stochastically Formed Subpopulation of Low-Energy Cells. PLoS Biol 2021, 19, e3001194. [Google Scholar] [CrossRef]
- Kollerová, S.; Jouvet, L.; Smelková, J.; Zunk-Parras, S.; Rodríguez-Rojas, A.; Steiner, U.K. Phenotypic Resistant Single-Cell Characteristics under Recurring Ampicillin Antibiotic Exposure in Escherichia Coli. mSystems 2024, 9, e00256-24. [Google Scholar] [CrossRef]
- Łapińska, U.; Voliotis, M.; Lee, K.K.; Campey, A.; Stone, M.R.L.; Tuck, B.; Phetsang, W.; Zhang, B.; Tsaneva-Atanasova, K.; Blaskovich, M.A.; et al. Fast Bacterial Growth Reduces Antibiotic Accumulation and Efficacy. eLife 2022, 11, e74062. [Google Scholar] [CrossRef]
- Brandis, G.; Larsson, J.; Elf, J. Antibiotic Perseverance Increases the Risk of Resistance Development. Proc. Natl. Acad. Sci. U.S.A. 2023, 120, e2216216120. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Beltran, J.C.R.; Rodríguez-Beltrán, J.; Aguilar-Luviano, O.B.; Velez-Santiago, J.; Mondragón-Palomino, O.; MacLean, R.C.; Fuentes-Hernández, A.; San Millán, A.; Peña-Miller, R. Plasmid-Mediated Phenotypic Noise Leads to Transient Antibiotic Resistance in Bacteria. Nat Commun 2024, 15, 2610. [Google Scholar] [CrossRef]
- Koganezawa, Y.; Umetani, M.; Sato, M.; Wakamoto, Y. History-Dependent Physiological Adaptation to Lethal Genetic Modification under Antibiotic Exposure. eLife 2022, 11, e74486. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.K.; Łapińska, U.; Tolle, G.; Micaletto, M.; Zhang, B.; Phetsang, W.; Verderosa, A.D.; Invergo, B.M.; Westley, J.; Bebes, A.; et al. Heterogeneous Efflux Pump Expression Underpins Phenotypic Resistance to Antimicrobial Peptides. eLife 2025, 13, RP99752. [Google Scholar] [CrossRef] [PubMed]
- Truong-Bolduc, Q.C.; Wang, Y.; Ferrer-Espada, R.; Reedy, J.L.; Martens, A.T.; Goulev, Y.; Paulsson, J.; Vyas, J.M.; Hooper, D.C. Staphylococcus Aureus AbcA Transporter Enhances Persister Formation under β-Lactam Exposure. Antimicrob Agents Chemother 2024, 68, e01340-23. [Google Scholar] [CrossRef]
- Goode, O.; Smith, A.; Zarkan, A.; Cama, J.; Invergo, B.M.; Belgami, D.; Caño-Muñiz, S.; Metz, J.; O’Neill, P.; Jeffries, A.; et al. Persister Escherichia Coli Cells Have a Lower Intracellular pH than Susceptible Cells but Maintain Their pH in Response to Antibiotic Treatment. mBio 2021, 12, e00909-21. [Google Scholar] [CrossRef] [PubMed]
- Baron, V.O.; Chen, M.; Hammarstrom, B.; Hammond, R.J.H.; Glynne-Jones, P.; Gillespie, S.H.; Dholakia, K. Real-Time Monitoring of Live Mycobacteria with a Microfluidic Acoustic-Raman Platform. Commun Biol 2020, 3, 236. [Google Scholar] [CrossRef]
- Zhang, D.; Yin, F.; Qin, Q.; Qiao, L. Molecular Responses during Bacterial Filamentation Reveal Inhibition Methods of Drug-Resistant Bacteria. Proc. Natl. Acad. Sci. U.S.A. 2023, 120, e2301170120. [Google Scholar] [CrossRef]
- Shahryari, S.; Ahmad, S.; Foik, I.P.; Jankowski, P.; Samborski, A.; Równicki, M.; Vasantham, S.K.; Garstecki, P. Bacterial Strain Type and TEM-1 Enzyme Allele Impact Antibiotic Susceptibility Distribution in Monoclonal Populations: A Single-Cell Droplet Approach. R. Soc. Open Sci. 2025, 12, 242143. [Google Scholar] [CrossRef]
- Barros-Medina, I.; Robles-Ramos, M.Á.; Sobrinos-Sanguino, M.; Luque-Ortega, J.R.; Alfonso, C.; Margolin, W.; Rivas, G.; Monterroso, B.; Zorrilla, S. Evidence for Biomolecular Condensates Formed by the Escherichia Coli MatP Protein in Spatiotemporal Regulation of the Bacterial Cell Division Cycle. International Journal of Biological Macromolecules 2025, 309, 142691. [Google Scholar] [CrossRef]
- Monterroso, B.; Robles-Ramos, M.Á.; Sobrinos-Sanguino, M.; Luque-Ortega, J.R.; Alfonso, C.; Margolin, W.; Rivas, G.; Zorrilla, S. Bacterial Division Ring Stabilizing ZapA versus Destabilizing SlmA Modulate FtsZ Switching between Biomolecular Condensates and Polymers. Open Biol. 2023, 13, 220324. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, H.; Okano, K.; Tang, T.; Inoue, K.; Yamazaki, Y.; Kamikubo, H.; Cain, A.K.; Tanaka, Y.; Inglis, D.W.; et al. Shape-Based Separation of Drug-Treated Escherichia Coli Using Viscoelastic Microfluidics. Lab Chip 2022, 22, 2801–2809. [Google Scholar] [CrossRef]
- Shuppara, A.M.; Padron, G.C.; Sharma, A.; Modi, Z.; Koch, M.D.; Sanfilippo, J.E. Shear Flow Patterns Antimicrobial Gradients across Bacterial Populations. Sci. Adv. 2025, 11, eads5005. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lehnert, T.; Gijs, M.A.M. Effect of Inoculum Size and Antibiotics on Bacterial Traveling Bands in a Thin Microchannel Defined by Optical Adhesive. Microsyst Nanoeng 2021, 7, 86. [Google Scholar] [CrossRef] [PubMed]
- Alcalde, R.E.; Dundas, C.M.; Dong, Y.; Sanford, R.A.; Keitz, B.K.; Fouke, B.W.; Werth, C.J. The Role of Chemotaxis and Efflux Pumps on Nitrate Reduction in the Toxic Regions of a Ciprofloxacin Concentration Gradient. The ISME Journal 2021, 15, 2920–2932. [Google Scholar] [CrossRef]
- Oliveira, N.M.; Wheeler, J.H.R.; Deroy, C.; Booth, S.C.; Walsh, E.J.; Durham, W.M.; Foster, K.R. Suicidal Chemotaxis in Bacteria. Nat Commun 2022, 13, 7608. [Google Scholar] [CrossRef]
- Deroy, C.; Wheeler, J.H.R.; Rumianek, A.N.; Cook, P.R.; Durham, W.M.; Foster, K.R.; Walsh, E.J. Reconfigurable Microfluidic Circuits for Isolating and Retrieving Cells of Interest. ACS Appl. Mater. Interfaces 2022, 14, 25209–25219. [Google Scholar] [CrossRef]
- Pitruzzello, G.; Baumann, C.G.; Johnson, S.; Krauss, T.F. Single-Cell Motility Rapidly Quantifying Heteroresistance in Populations of Escherichia Coli and Salmonella Typhimurium. Small Science 2022, 2, 2100123. [Google Scholar] [CrossRef] [PubMed]
- Varadarajan, A.R.; Allan, R.N.; Valentin, J.D.P.; Castañeda Ocampo, O.E.; Somerville, V.; Pietsch, F.; Buhmann, M.T.; West, J.; Skipp, P.J.; Van Der Mei, H.C.; et al. An Integrated Model System to Gain Mechanistic Insights into Biofilm-Associated Antimicrobial Resistance in Pseudomonas Aeruginosa MPAO1. npj Biofilms Microbiomes 2020, 6, 46. [Google Scholar] [CrossRef] [PubMed]
- Valentin, J.D.P.; Straub, H.; Pietsch, F.; Lemare, M.; Ahrens, C.H.; Schreiber, F.; Webb, J.S.; Van Der Mei, H.C.; Ren, Q. Role of the Flagellar Hook in the Structural Development and Antibiotic Tolerance of Pseudomonas Aeruginosa Biofilms. The ISME Journal 2022, 16, 1176–1186. [Google Scholar] [CrossRef]
- Krzyżek, P.; Migdał, P.; Grande, R.; Gościniak, G. Biofilm Formation of Helicobacter Pylori in Both Static and Microfluidic Conditions Is Associated With Resistance to Clarithromycin. Front. Cell. Infect. Microbiol. 2022, 12, 868905. [Google Scholar] [CrossRef]
- Jain, S.; Singh, A.; Tiwari, N.; Naik, A.; Chatterjee, R.; Chakravortty, D.; Basu, S. Observations on Phenomenological Changes in Klebsiella Pneumoniae under Fluidic Stresses. Soft Matter 2023, 19, 9239–9253. [Google Scholar] [CrossRef]
- Nguyen, A.V.; Shourabi, A.Y.; Yaghoobi, M.; Zhang, S.; Simpson, K.W.; Abbaspourrad, A. A High-Throughput Integrated Biofilm-on-a-Chip Platform for the Investigation of Combinatory Physicochemical Responses to Chemical and Fluid Shear Stress. PLoS ONE 2022, 17, e0272294. [Google Scholar] [CrossRef] [PubMed]
- Islayem, D.; Arya, S.; Yasmeen, N.; Hallfors, N.; Ngoc, H.; Alkhatib, S.; Mela, I.; Anwar, S.; Truong, V.K.; Pappa, A.M. A Modular, Lego-Like Microfluidic Platform for Multimodal Analysis of Biofilm Formation and Antimicrobial Response. Adv Materials Inter 2025, e00303. [Google Scholar] [CrossRef]
- Savorana, G.; Słomka, J.; Stocker, R.; Rusconi, R.; Secchi, E. A Microfluidic Platform for Characterizing the Structure and Rheology of Biofilm Streamers. Soft Matter 2022, 18, 3878–3890. [Google Scholar] [CrossRef]
- Heredia-Ponce, Z.; Secchi, E.; Toyofuku, M.; Marinova, G.; Savorana, G.; Eberl, L. Genotoxic Stress Stimulates eDNA Release via Explosive Cell Lysis and Thereby Promotes Streamer Formation of Burkholderia Cenocepacia H111 Cultured in a Microfluidic Device. npj Biofilms Microbiomes 2023, 9, 96. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cai, Y.; Zeng, L.; Liu, P.; Ma, L.Z.; Liu, J. A Microfluidic Approach for Quantitative Study of Spatial Heterogeneity in Bacterial Biofilms. Small Science 2022, 2, 2200047. [Google Scholar] [CrossRef] [PubMed]
- Tang, P.-C.; Eriksson, O.; Sjögren, J.; Fatsis-Kavalopoulos, N.; Kreuger, J.; Andersson, D.I. A Microfluidic Chip for Studies of the Dynamics of Antibiotic Resistance Selection in Bacterial Biofilms. Front. Cell. Infect. Microbiol. 2022, 12, 896149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gholizadeh, H.; Young, P.; Traini, D.; Li, M.; Ong, H.X.; Cheng, S. Real-time In-situ Electrochemical Monitoring of Pseudomonas Aeruginosa Biofilms Grown on Air–Liquid Interface and Its Antibiotic Susceptibility Using a Novel Dual-chamber Microfluidic Device. Biotech & Bioengineering 2023, 120, 702–714. [Google Scholar] [CrossRef]
- Thiermann, R.; Sandler, M.; Ahir, G.; Sauls, J.T.; Schroeder, J.; Brown, S.; Le Treut, G.; Si, F.; Li, D.; Wang, J.D.; et al. Tools and Methods for High-Throughput Single-Cell Imaging with the Mother Machine. eLife 2024, 12, RP88463. [Google Scholar] [CrossRef]
- Liu, X.; Li, X.; Wu, N.; Luo, Y.; Zhang, J.; Yu, Z.; Shen, F. Formation and Parallel Manipulation of Gradient Droplets on a Self-Partitioning SlipChip for Phenotypic Antimicrobial Susceptibility Testing. ACS Sens. 2022, 7, 1977–1984. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, X.; Yu, Z.; Luo, Y.; Hu, Q.; Xu, Z.; Dai, J.; Wu, N.; Shen, F. Combinatorial Screening SlipChip for Rapid Phenotypic Antimicrobial Susceptibility Testing. Lab Chip 2022, 22, 3952–3960. [Google Scholar] [CrossRef]
- Wang, Q.; Li, X.; Ren, Y.; Hu, Q.; Xu, L.; Chen, W.; Liu, J.; Wu, N.; Tao, M.; Sun, J.; et al. Rapid and Precise Treatment Selection for Antimicrobial-Resistant Infection Enabled by a Nano-Dilution SlipChip. Biosensors and Bioelectronics 2025, 271, 117084. [Google Scholar] [CrossRef]
- Hengoju, S.; Wohlfeil, S.; Munser, A.S.; Boehme, S.; Beckert, E.; Shvydkiv, O.; Tovar, M.; Roth, M.; Rosenbaum, M.A. Optofluidic Detection Setup for Multi-Parametric Analysis of Microbiological Samples in Droplets. Biomicrofluidics 2020, 14, 024109. [Google Scholar] [CrossRef]
- Samimi, A.; Hengoju, S.; Martin, K.; Rosenbaum, M.A. Advancing Droplet-Based Microbiological Assays: Optofluidic Detection Meets Multiplexed Droplet Generation. Analyst 2025, 150, 3137–3146. [Google Scholar] [CrossRef]
- Pacocha, N.; Bogusławski, J.; Horka, M.; Makuch, K.; Liżewski, K.; Wojtkowski, M.; Garstecki, P. High-Throughput Monitoring of Bacterial Cell Density in Nanoliter Droplets: Label-Free Detection of Unmodified Gram-Positive and Gram-Negative Bacteria. Anal. Chem. 2021, 93, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-J.; Chao, P.-H.; Cheng, H.-W.; Wang, J.-K.; Wang, Y.-L.; Han, Y.-Y.; Huang, N.-T. An Antibiotic Concentration Gradient Microfluidic Device Integrating Surface-Enhanced Raman Spectroscopy for Multiplex Antimicrobial Susceptibility Testing. Lab Chip 2022, 22, 1805–1814. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-K.; Cheng, H.-W.; Liao, C.-C.; Lin, S.-J.; Chen, Y.-Z.; Wang, J.-K.; Wang, Y.-L.; Huang, N.-T. Bacteria Encapsulation and Rapid Antibiotic Susceptibility Test Using a Microfluidic Microwell Device Integrating Surface-Enhanced Raman Scattering. Lab Chip 2020, 20, 2520–2528. [Google Scholar] [CrossRef] [PubMed]
- Spencer, D.; Li, Y.; Zhu, Y.; Sutton, J.M.; Morgan, H. Electrical Broth Micro-Dilution for Rapid Antibiotic Resistance Testing. ACS Sens. 2023, 8, 1101–1108. [Google Scholar] [CrossRef]
- Zhao, Y.; Hou, Z.; Yan, B.; Cao, X.; Su, B.; Lv, M.; Cui, H.; Zhang, C. Research on Drug Efficacy Using a Terahertz Metasurface Microfluidic Biosensor Based on Fano Resonance Effect. ACS Appl. Mater. Interfaces 2024, 16, 52092–52103. [Google Scholar] [CrossRef]
- Nakamura, A.; Honma, N.; Tanaka, Y.; Suzuki, Y.; Shida, Y.; Tsuda, Y.; Hidaka, K.; Ogasawara, W. 7-Aminocoumarin-4-Acetic Acid as a Fluorescent Probe for Detecting Bacterial Dipeptidyl Peptidase Activities in Water-in-Oil Droplets and in Bulk. Anal. Chem. 2022, 94, 2416–2424. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of recently applied microfluidic approaches to natural biodiversity profiling for antibiotic producers.
Figure 1.
Schematic representation of recently applied microfluidic approaches to natural biodiversity profiling for antibiotic producers.
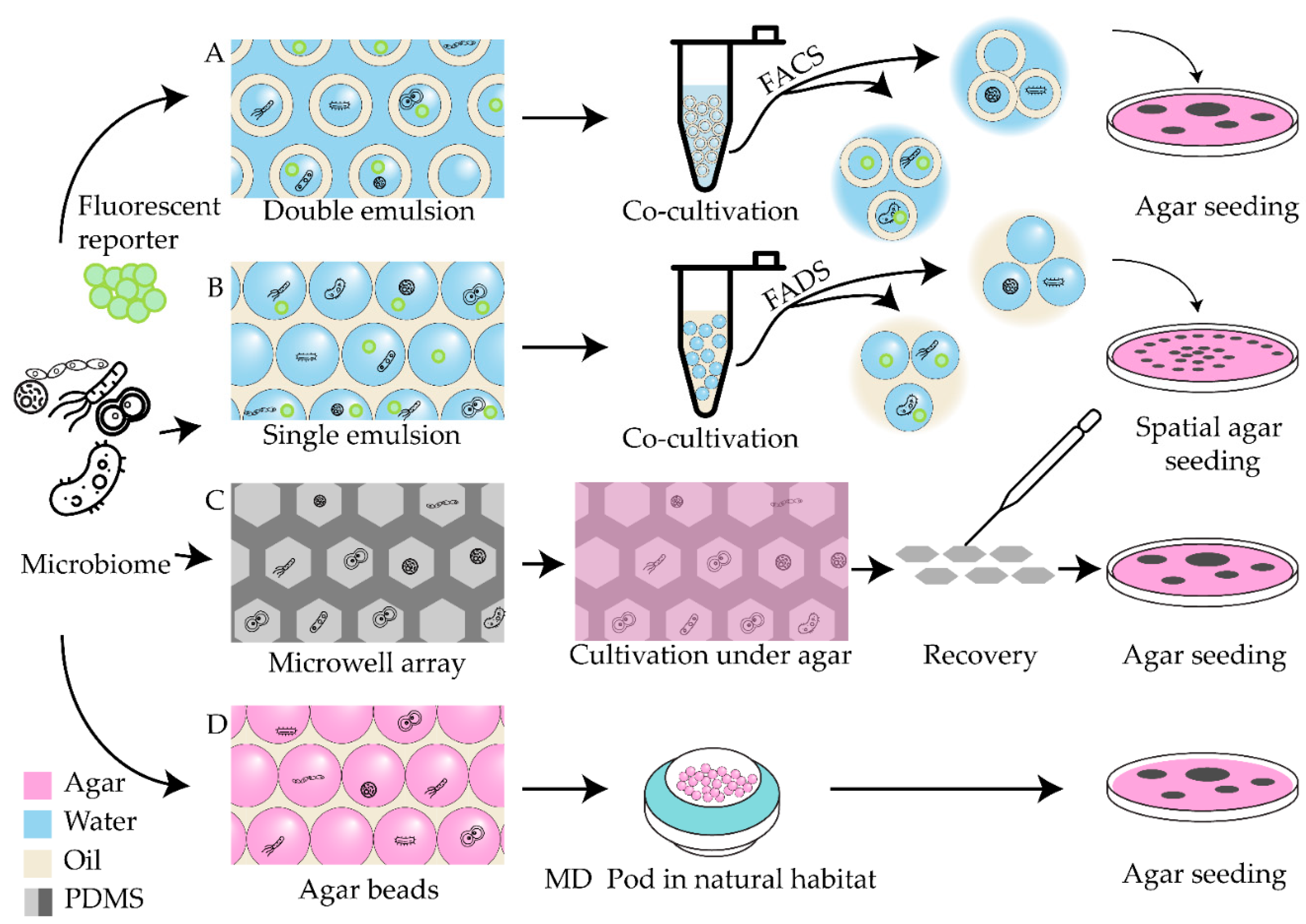
Figure 2.
Schematic representation of mutant library screening strategies.
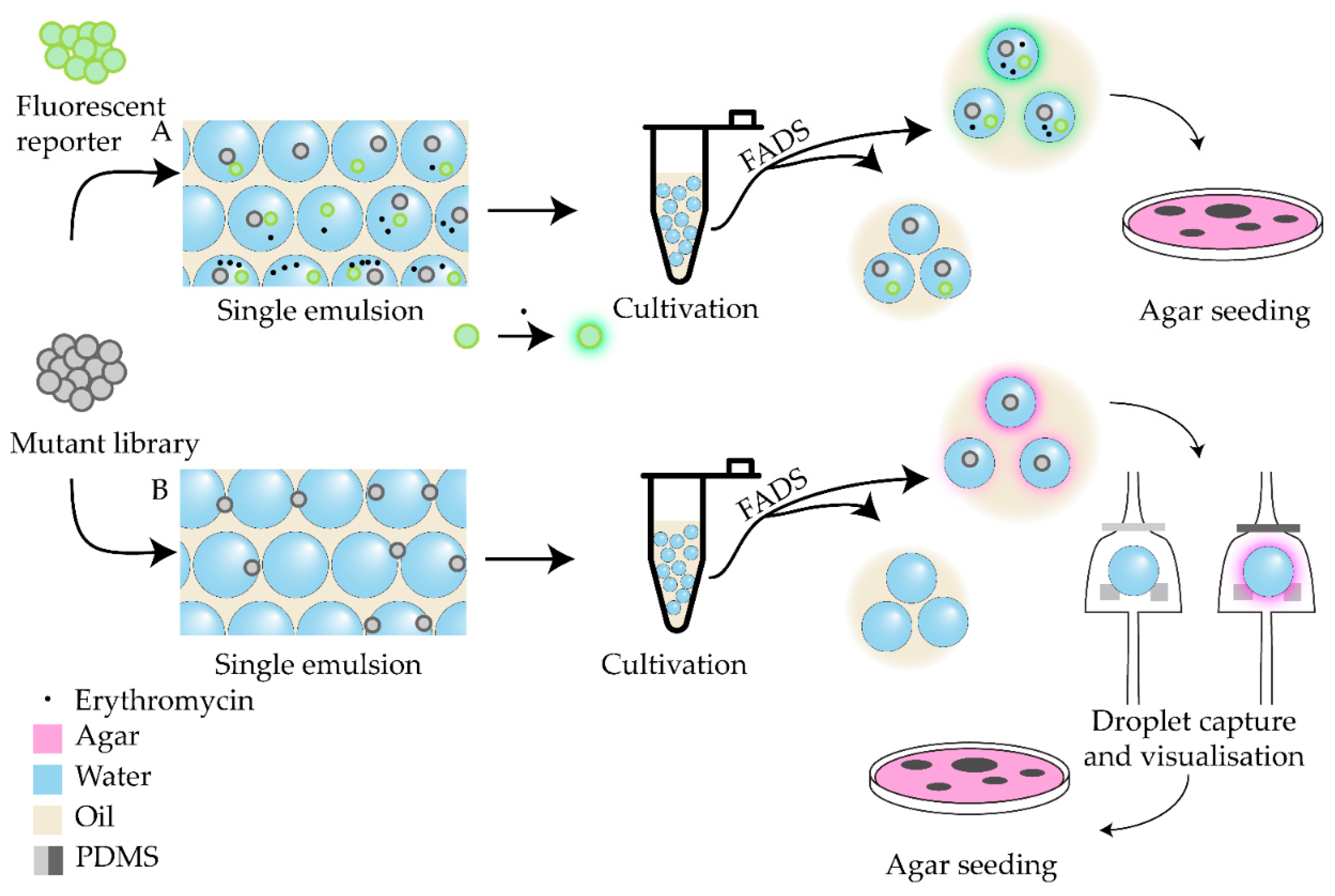
Figure 3.
Gradient generators for microfluidic applications. (A-D) On-chip gradient generators; (E-G) concentration variation in droplets.
Figure 3.
Gradient generators for microfluidic applications. (A-D) On-chip gradient generators; (E-G) concentration variation in droplets.
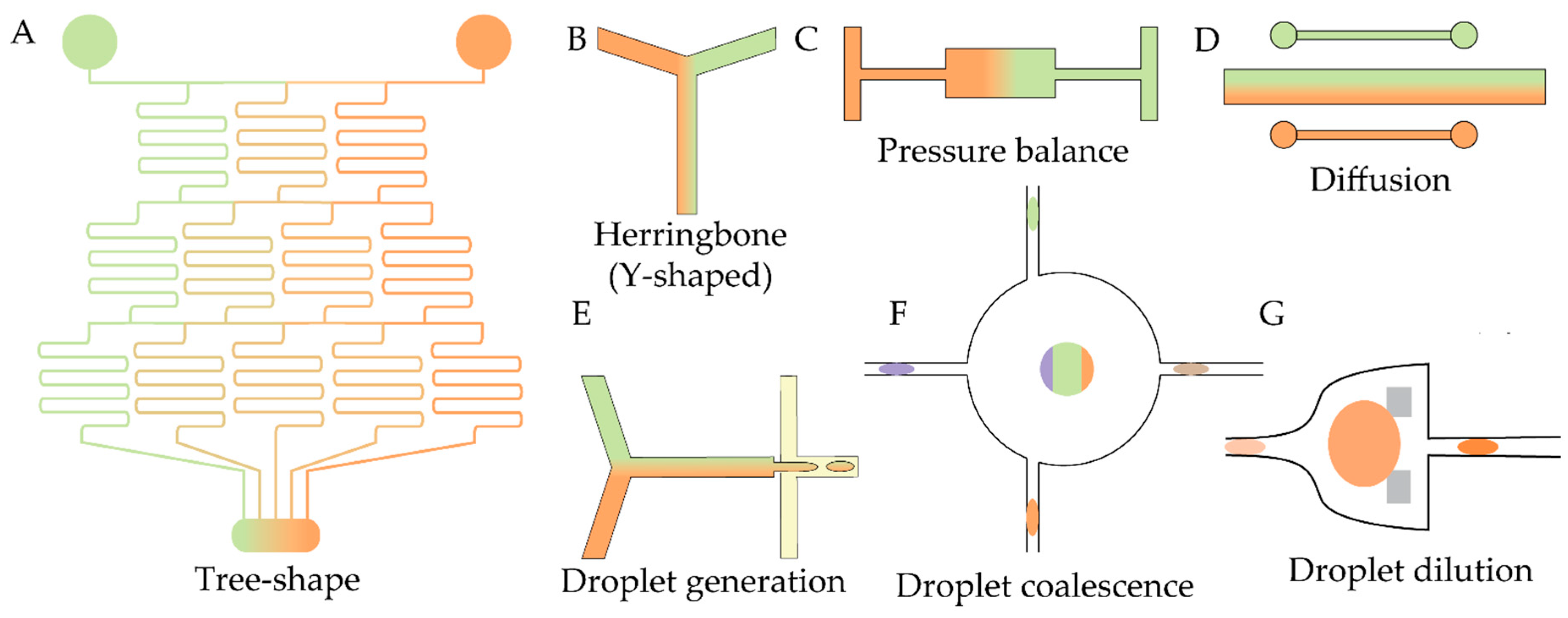
Figure 4.
Droplet-based screening approaches with imaging-based concentration control.
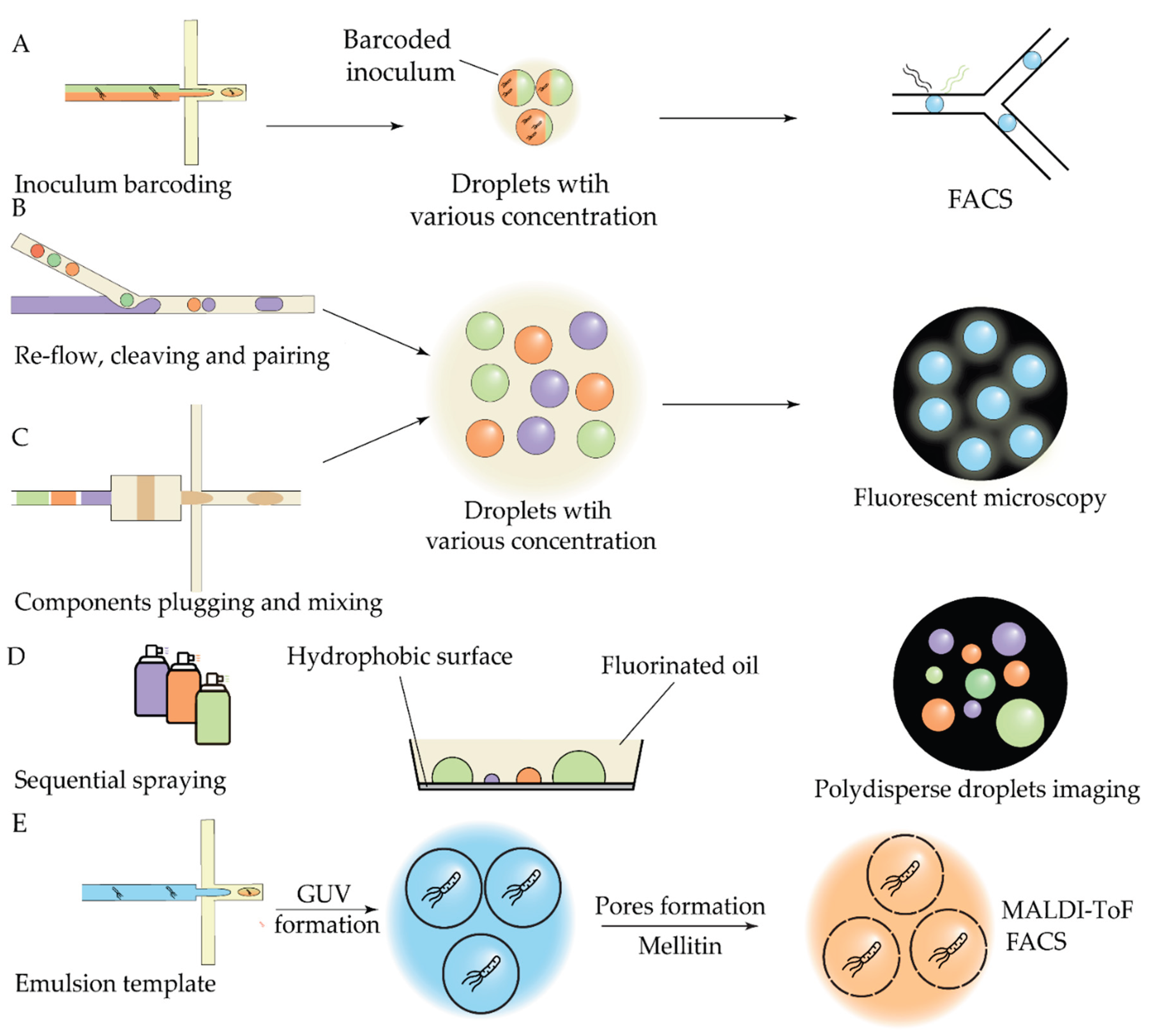
Figure 5.
Microfluidic screening approaches with spatially controlled droplets.
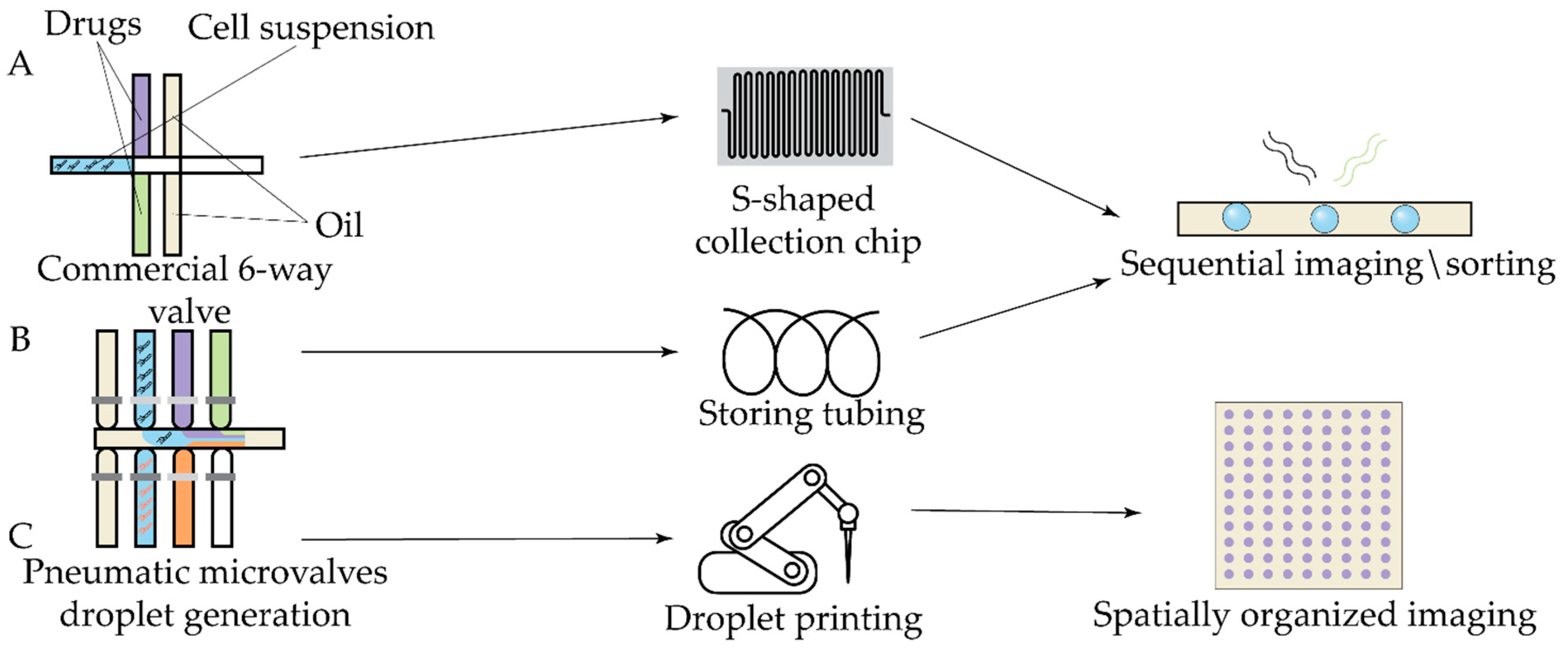
Figure 6.
Schematic representation of nm-SlipChip.
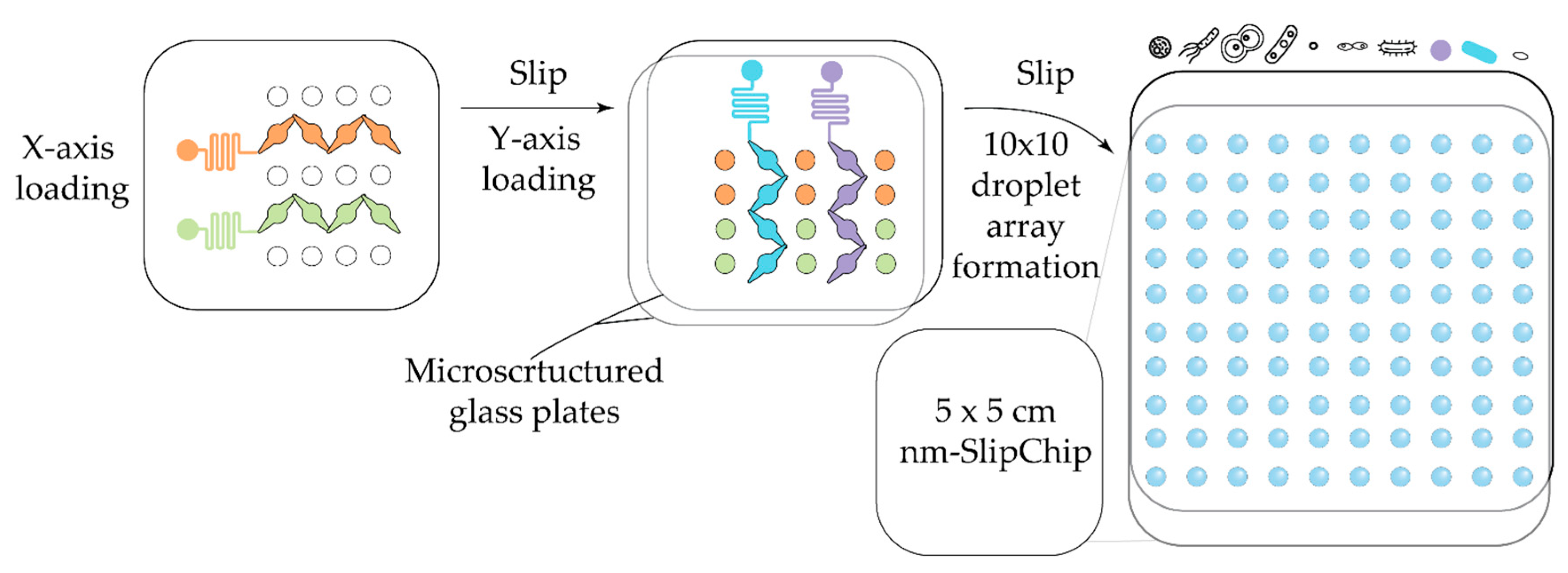
Figure 7.
Approaches to peptide libraries screening.
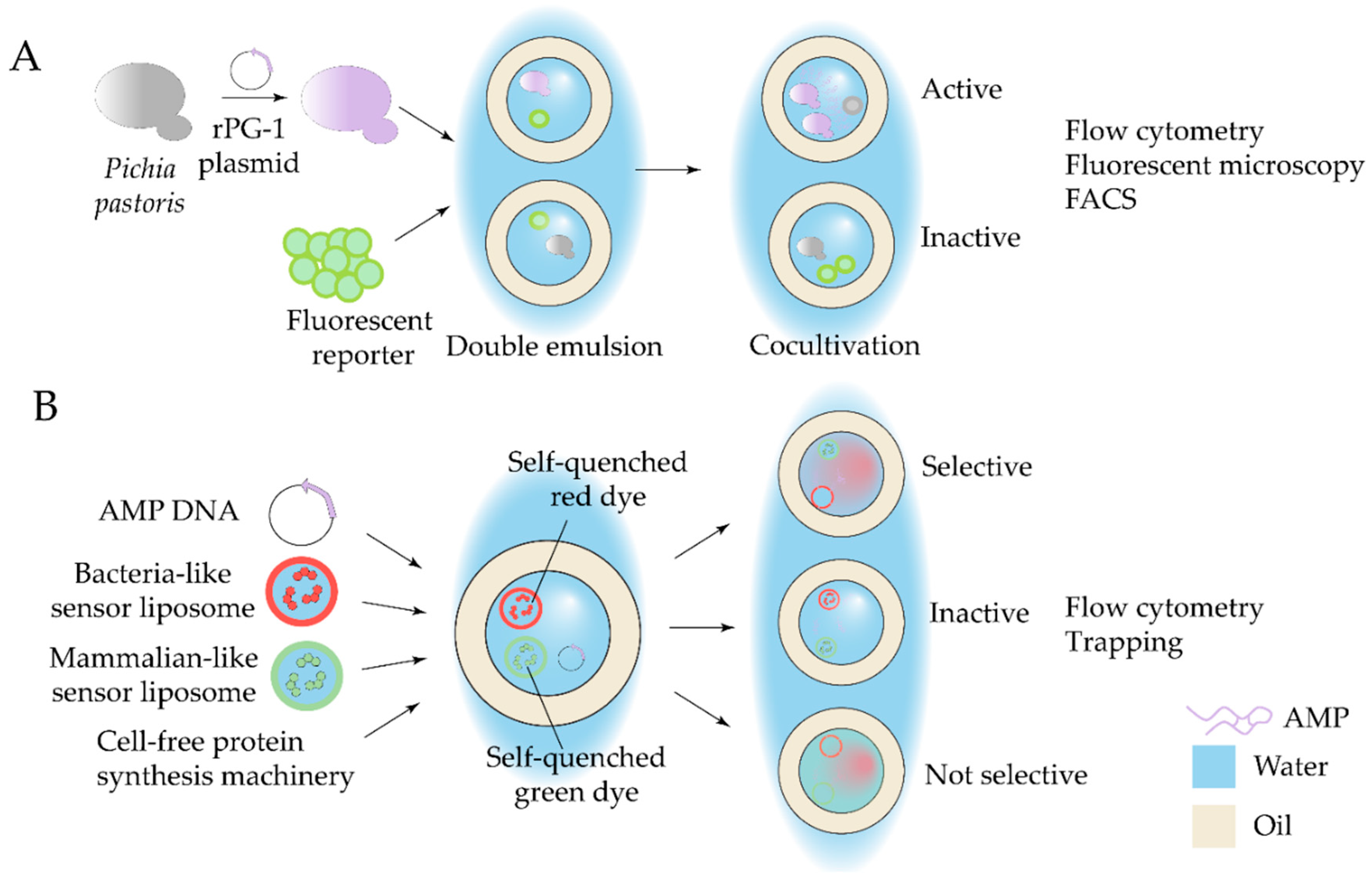
Figure 8.
Microfluidic-based phage discovery pipelines.
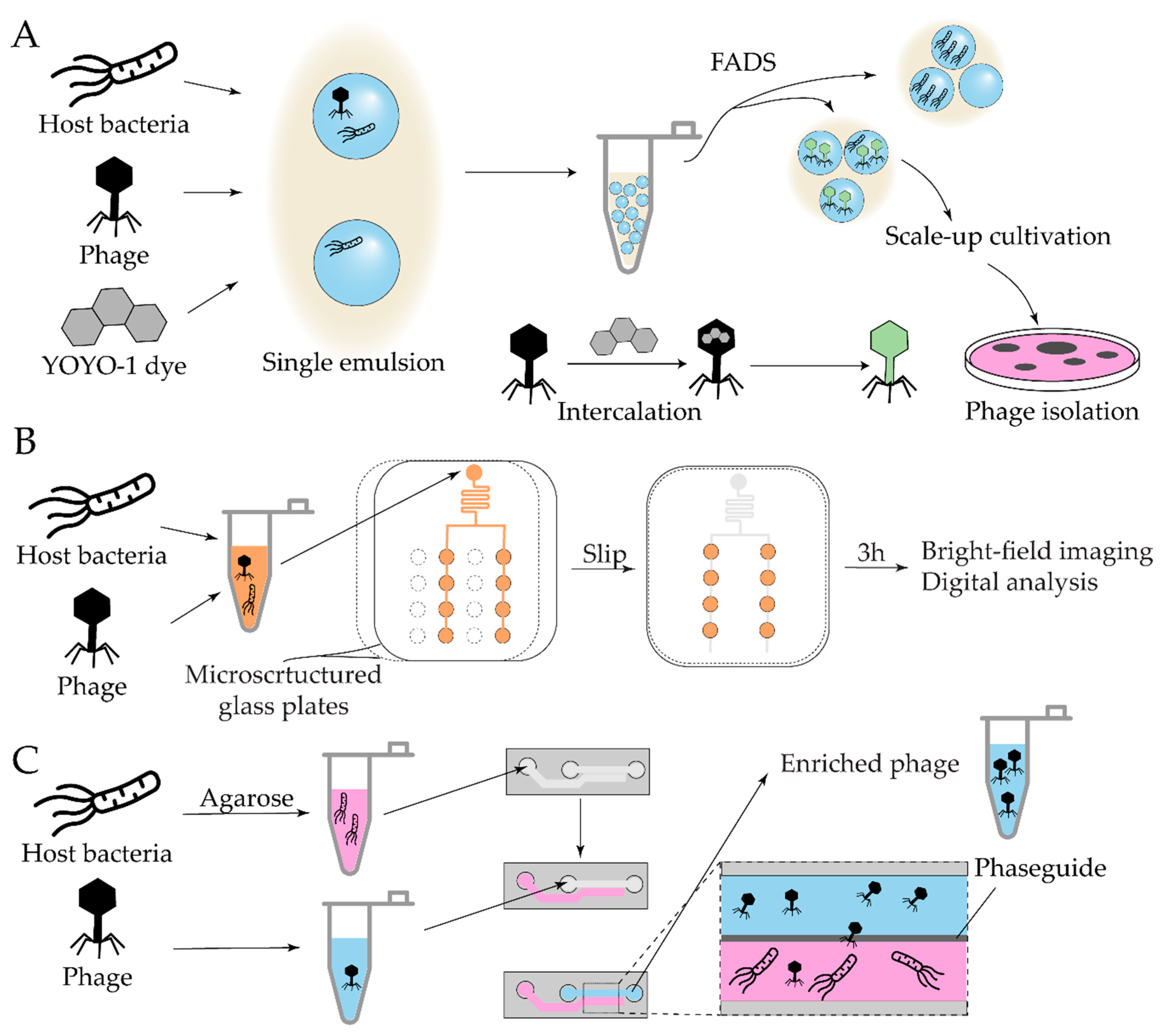
Figure 9.
Examples of microfluidic biofilm models for activity screening.
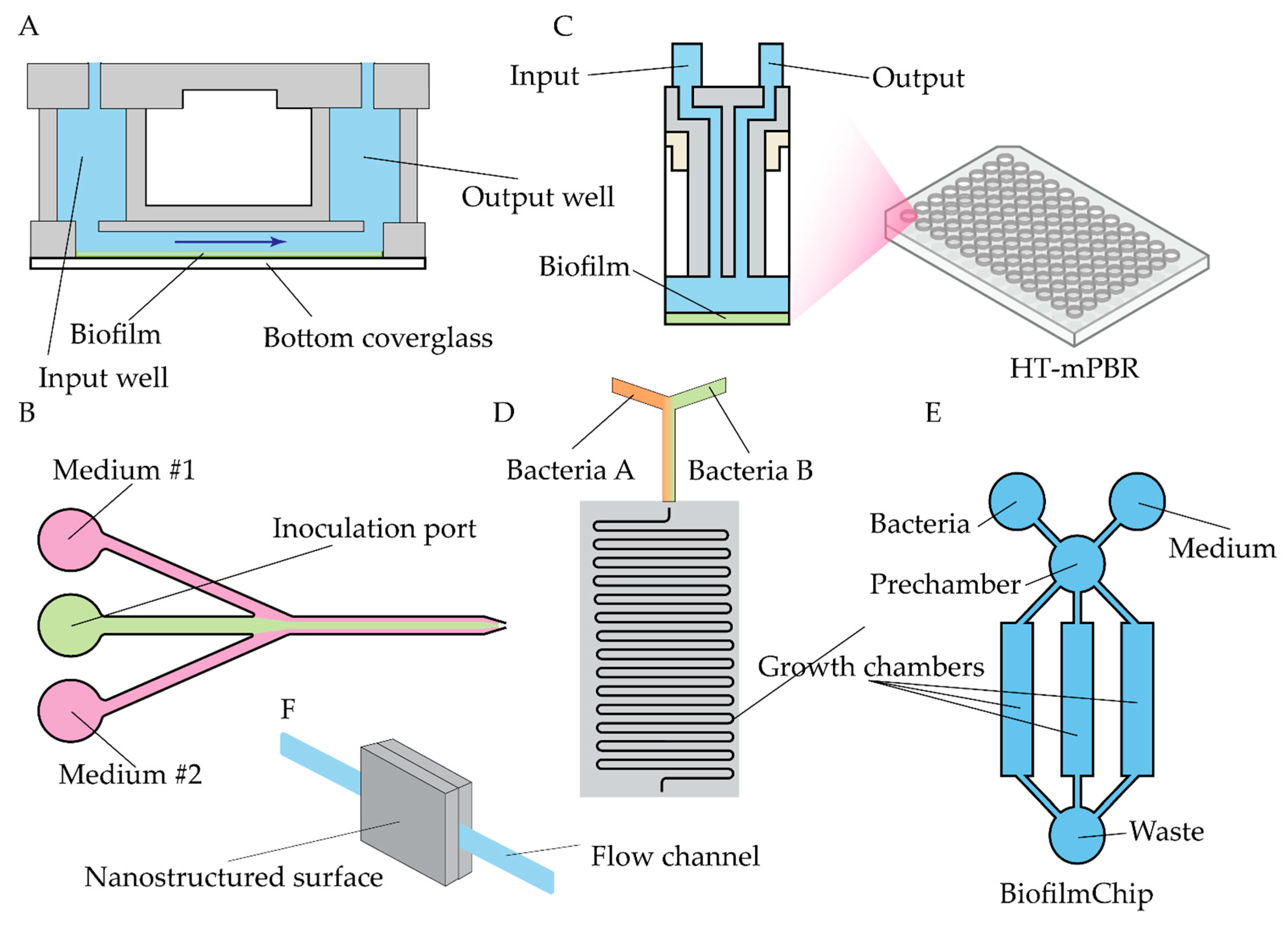
Figure 10.
Schematic representation of (A) mother machine (MM) and (B) similar design (iMM).
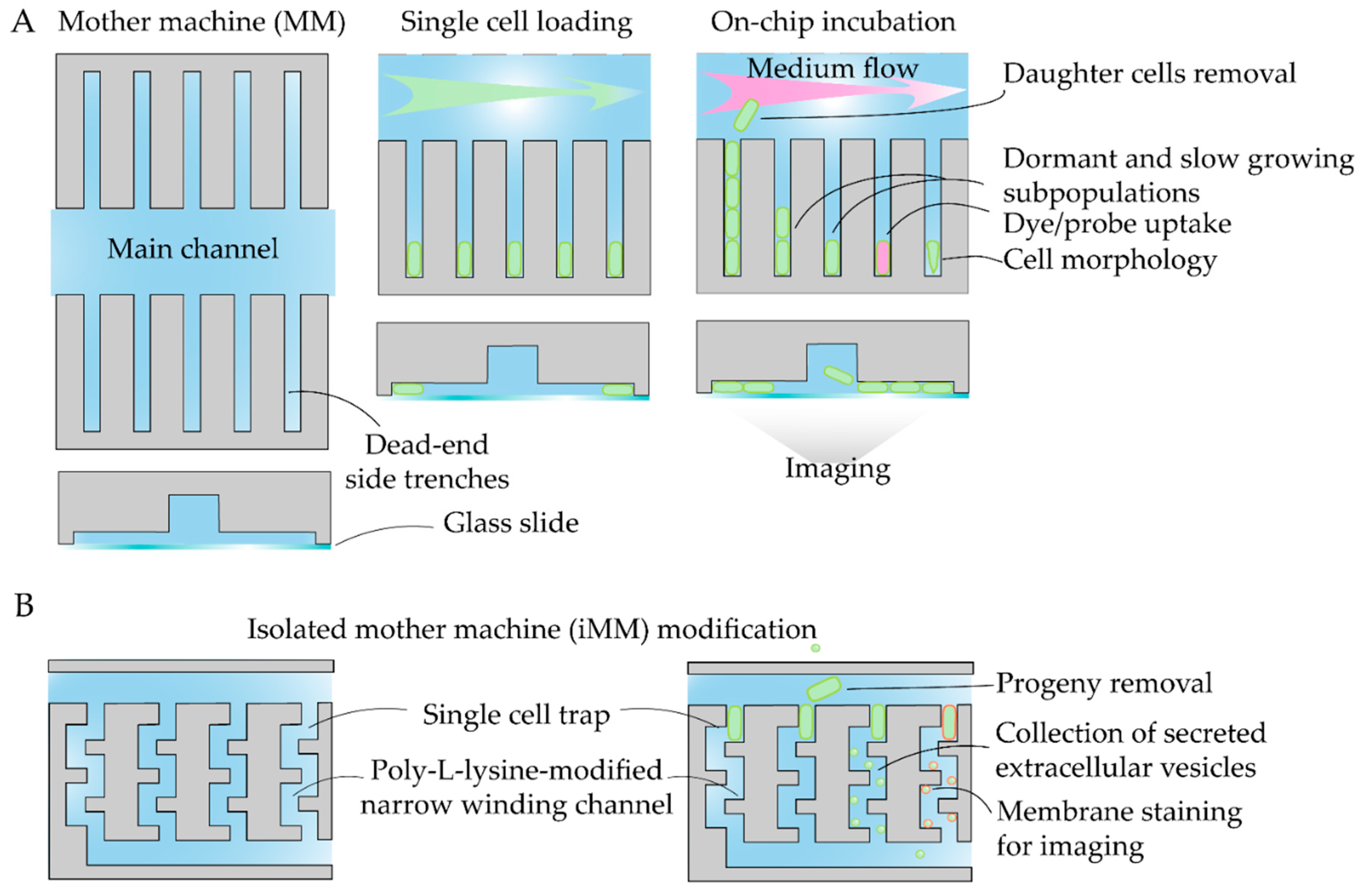
Figure 11.
Microfluidic experimental setups for on-chip cell imaging.
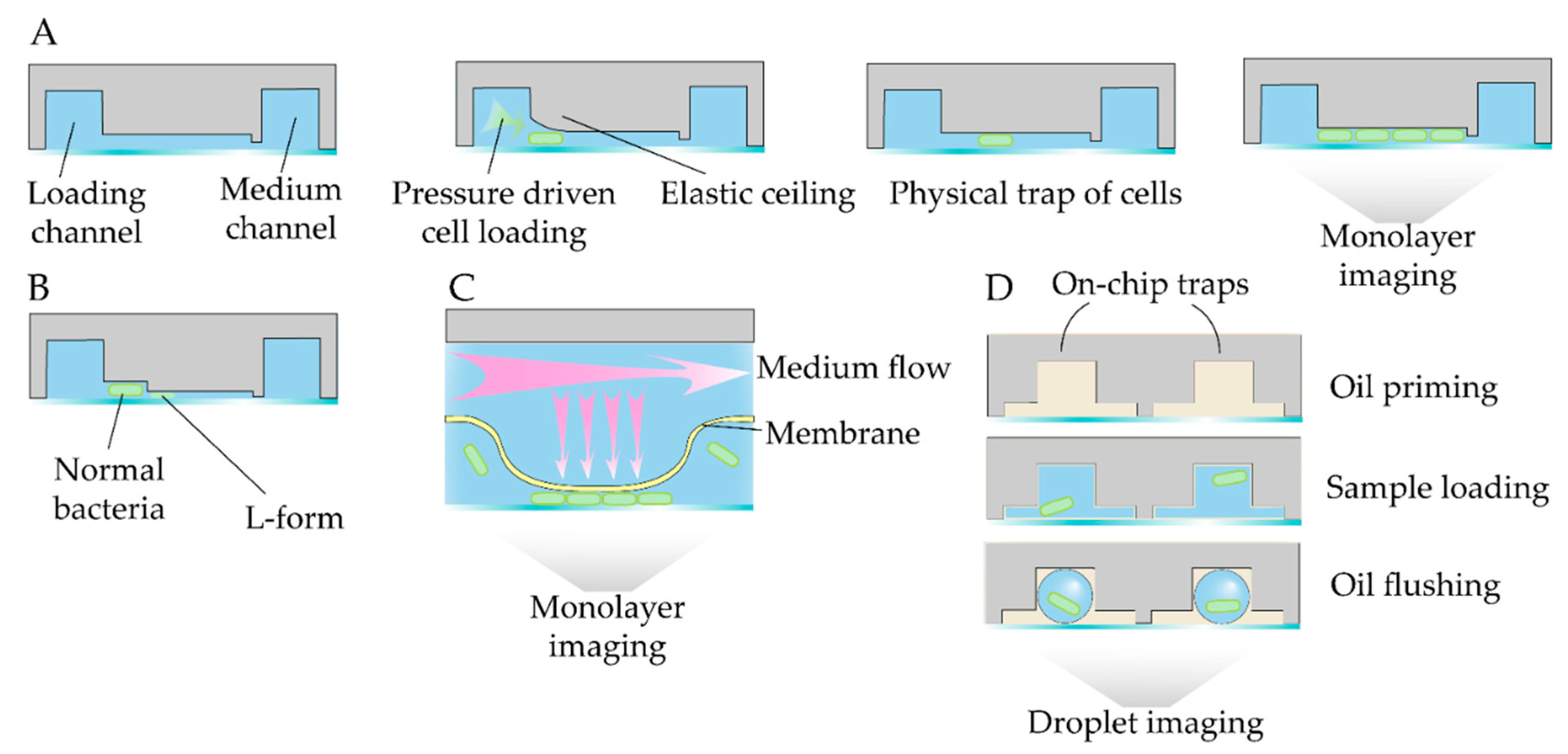
Figure 12.
Specific designs of mechanistic studies.
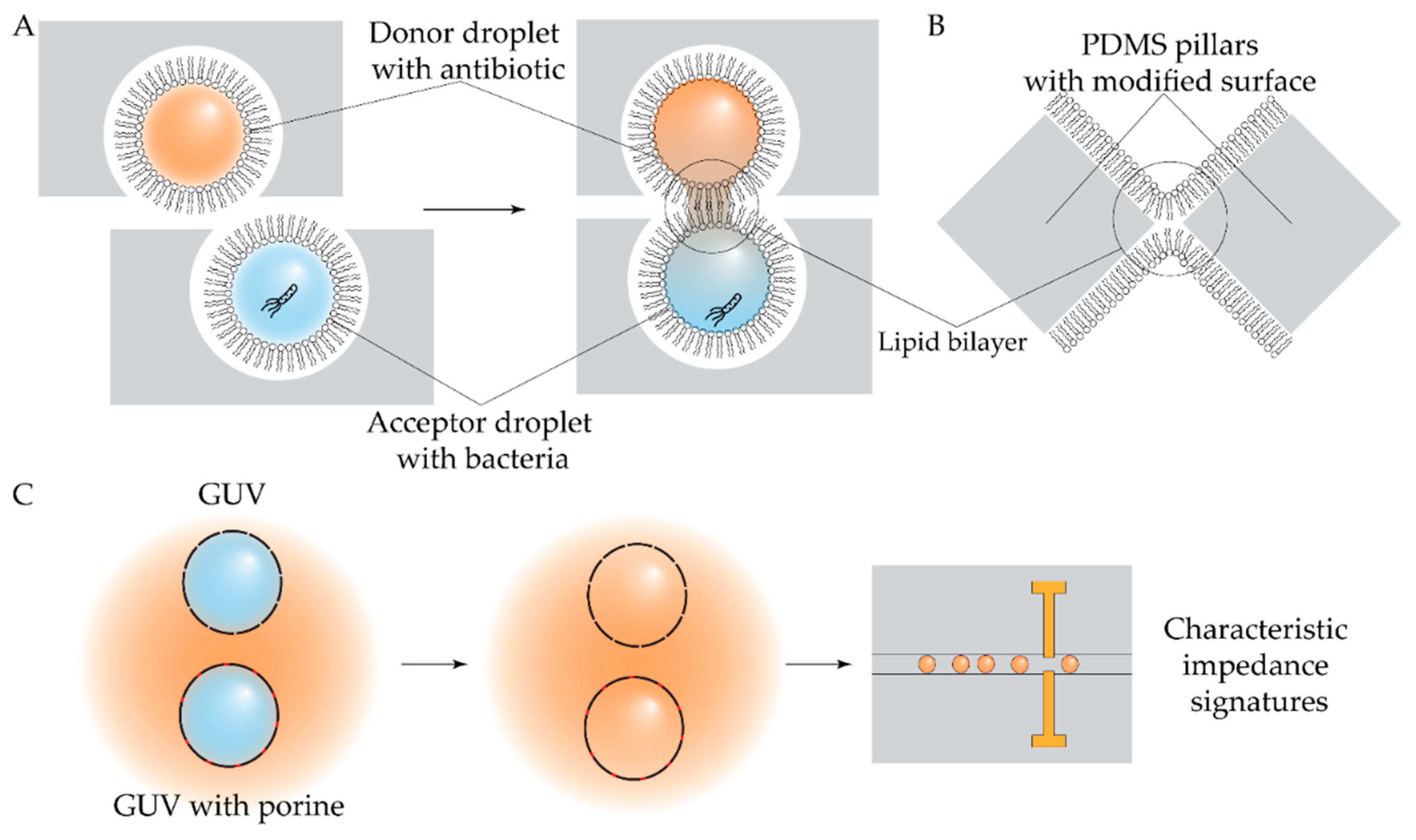
Figure 13.
Experimental workflows to perform microfluidic-driven ALE.
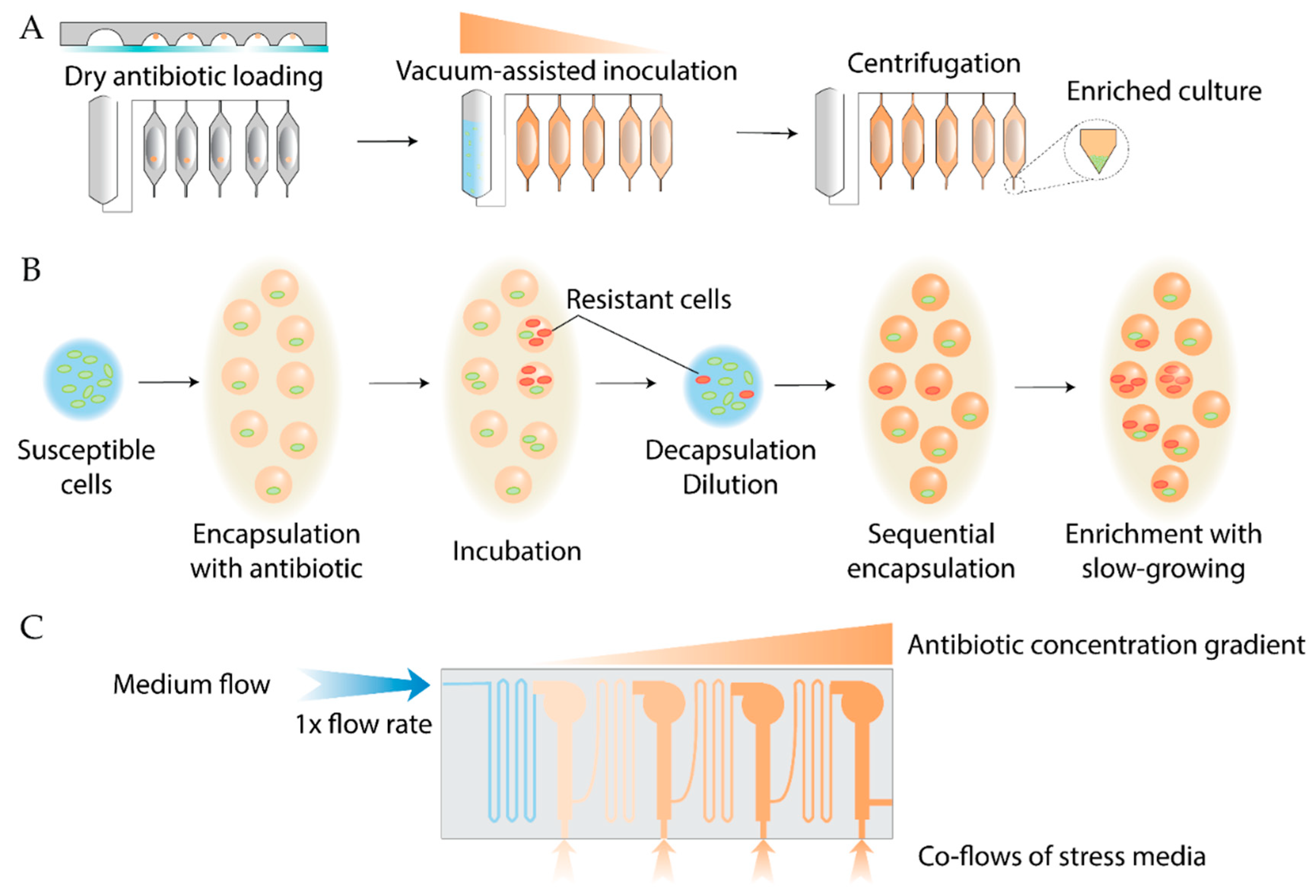
Figure 14.
Viscoelastic shape-based separation of E.coli subpopulations.
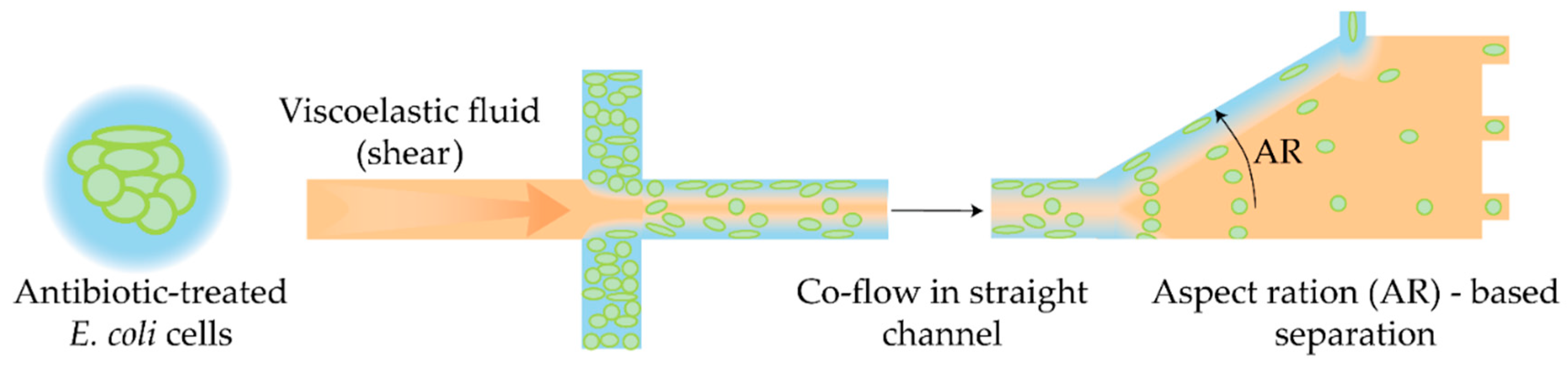
Figure 15.
Microfluidic models to study spatial gradients and bacterial motility.
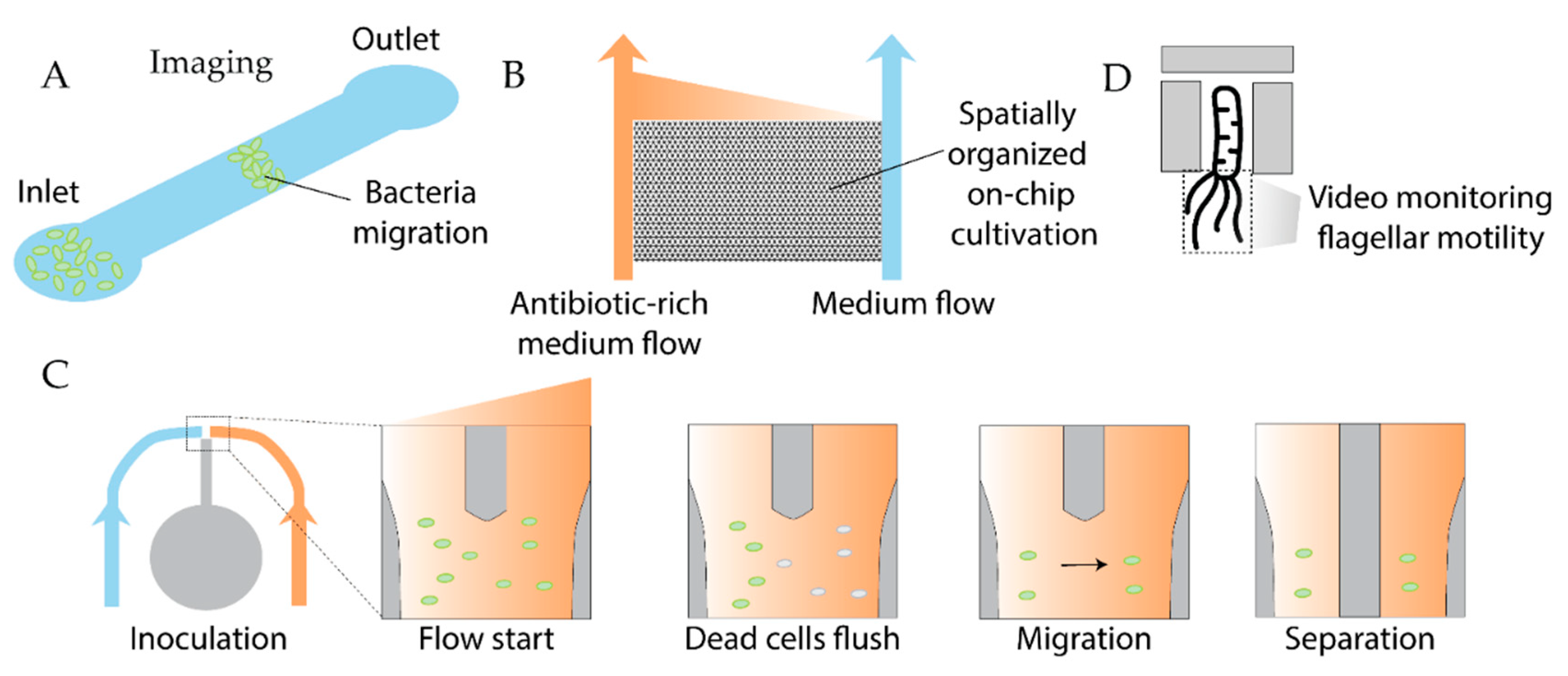
Figure 16.
Microfluidic biofilm models for precise monitoring of stress responses.
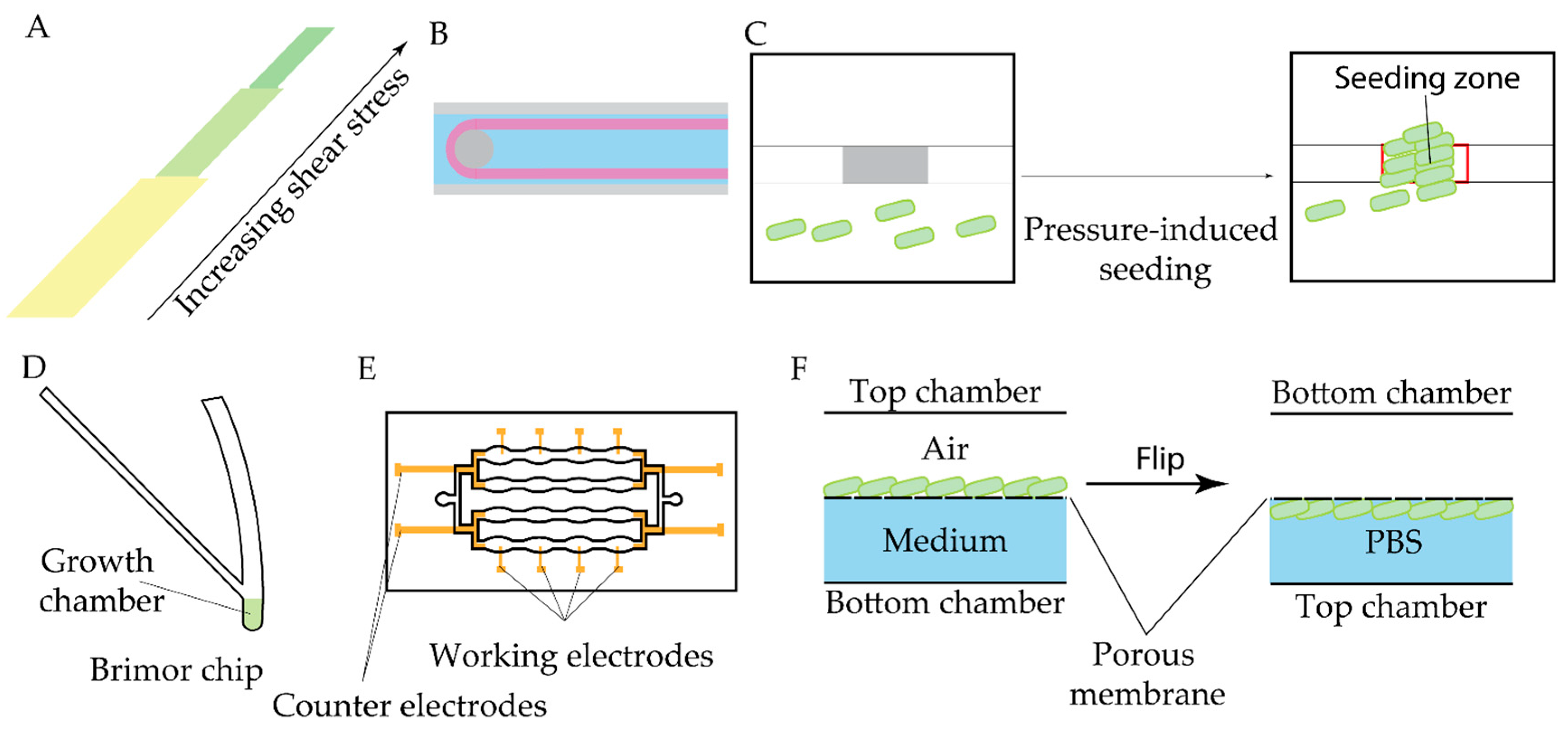
Table 2.
Recent single cell studies on genetic bacterial resistance.
| Antibiotic | Device | Detection method | Studied strains | Observed effect | Ref |
|---|---|---|---|---|---|
| MDR strains |
Figure 10A MM |
Microscopy |
Е. Coli Salmonella enterica |
Single cell growth rate | [170] |
| β-lactams | Figure 10A | Microscopy | E.coli | Visualization of conjugational and vertical transfer events | [171] |
| Isoniazid | Figure 11C | Microscopy |
M. smegmatis msm2570::Tn mutant |
Individual cells growth and lysis kinetics | [173] |
Table 4.
Summary of recently introduced biofilm eradication microfluidic models.
| Antibiotics | Device | Detection method | Tested biofilms | Key features | Ref |
|---|---|---|---|---|---|
| Gentamicin Streptomycin |
2PAB Figure 16A |
Microscopy |
E. coli P. aeruginosa |
Simultaneous control of antibiotic treatment and shear stress | [202] |
| Berberine | Figure 16C | Microscopy |
E. coli P. aeruginosa S. typimurium K. pneumonia B. subtilis S. aureus E. faecium M. smegmatis |
Spatially controlled seeding Epoxy-resin sealing to block oxygen penetration Suitable for broad range of bacteria |
[206] |
| Ciprofloxacin | Brimor Figure 16D |
Microscopy | E. coli | Long-term cultivation Simple manufacturing procedure |
[207] |
| Ciprofloxacin | Figure 16F | Pyocyanin detection | P. aeruginosa | Air-liquid interface biofilms Electochemical detection of biomarker |
[208] |
| Tetracycline Chloramphenicol Amikacin Coatings and nanoparticles |
Figure 16E | Microscopy, electrical impedance | P. aeruginosa | Label-free monitoring Modular structure to assess migration and regrowth Localized shear stress variations |
[203] |
| Mitomycin C Ciprofloxacin |
Figure 16B | Microscopy |
P. aeruginosa Burkholderia cenocepacia |
Visualization of streamers | [204,205] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



