Submitted:
06 November 2025
Posted:
06 November 2025
You are already at the latest version
Abstract
Thanks to the Next Generation Sequencing technologies, the diversity of Fungi can now be investigated through the analysis of their genome sequences. Naviculisporaceae is a family within the Sordariales, whose diversity is not well known, with only one genome sequence published for this family. Here, we report the isolation and cultivation of 20 new strains of Naviculisporaceae. Their genome sequences, as well as those of five commercially available strains, were determined, thus providing complete genome sequences for 25 new Naviculisporaceae strains. Species delimitation was conducted using a combination of (1) ITS + LSU phylogenetic analysis of the new isolates along with other known species of the family, (2) comparisons between DNA barcode sequences of the new strains with those of known species, and (3) average genome-wide nucleotide identity calculation. We built a phylogenomic tree and studied the organization of the mating-type locus. In vitro fruiting was obtained for 16 strains, enabling the definition of seven new species: Pseudorhypophila gallica, Pseudorhypophila guyanensis Rhypophila alpibus, Rhypophila brasiliensis, Rhypophila camarguensis, Rhypophila reunionensis and Rhypophila thailandica, as well as two new com-binations: Pseudorhypophila latipes and Pseudorhypophila oryzae. Eight strains for which in-vitro fruiting was not obtained may belong to additional new species. These results ex-pand the known diversity of the Naviculisporaceae and greatly enlarge the genomic data available for the family.
Keywords:
Sordariales
; Naviculisporaceae
; Phylogenomics
; new species
1. Introduction
Complete genome sequences, when combined with crossing assays and morphological analyses, enable accurate species. In particular, the Average Nucleotide Identity (ANI) can be very useful to delimit species using genome-wide sequences [1,2,3]. In most cases, a nucleotide divergence threshold of 1% allows clear species delimitation using the FungANI program: strains with ANI values above 99.5% consistently belong to the same species, whereas those with ANI values below 99% usually correspond to distinct species [2]. Genome analyses often reveal unexpected or cryptic diversity in fungi, uncovering distinct genetic lineages that remain indistinguishable based on morphology alone (see [4,5,6,7,8,9,10] for examples in the Sordariales). Beyond taxonomy, access to the full genome also provides insights into the mating-type locus structure, contributing to our understanding of the breeding system (e.g., homothallism versus heterothallism). This for example can help to explain fertility or sterility observed under culture conditions. Moreover, genome availability opens avenues for mining enzymes of interest, as well as regulatory and secondary metabolite pathways.
The Naviculisporaceae is a family within the Sordariales (Sordariomycetes, Ascomycota) that has been recently defined through molecular phylogenies using four DNA barcodes: the internal transcribed spacer region (ITS), and parts of the nuclear rDNA large subunit (LSU), the RNA polymerase II subunit 2 (RPB2) and the β-tubulin (TUB2) genes [11]. It currently contains four recognized genera, Areotheca, Naviculispora, Pseudorhypophila and Rhypophila, which collectively contain 11 recognized species: two species of Areotheca, one of Naviculispora, four of Pseudorhypophila and four of Rhypophila. Naviculisporaceae also includes three species of Arnium and one species of Gilmaniella, all needing taxonomic revision [11,12,13]. It is likely that additional species will be added to this family as more strains are analyzed and more genomes are sequenced. To date, only one Naviculisporaceae strain has been sequenced and Published: PSN293, isolated in 2017 from donkey dung collected in the Auvergne region of central France [14]. It was sequenced under the name Podospora decipiens (G. Winter ex Fuckel) Niessl [14], which is the old name for this species, now known as Rhypophila decipiens (G. Winter) Y. Marín, A.N. Mill. & Guarro [11].
Sordariales fungi occur in a wide range of biotopes and rank among the most wide-spread fungi present in soils [15]. They are also commonly found in the dung of herbivores [16]. Yet, their actual phylogenetic and morphological diversity remains largely unexplored [17]. In particular, genome sequences are unavailable for many species. To address this gap, we undertook a comprehensive sampling effort to characterize the diversity of Sordariales in both soil and dung environments. For soil, we developed a dedicated method to facilitate their recovery [18], whereas traditional moist chamber techniques were used to isolate species from dung. Using these approaches, we obtained Naviculisporaceae species that appeared to be new to science. Here, we report the genome sequences of 25 additional Naviculisporaceae strains, including five commercially available ones and twenty newly isolated ones. Morphological descriptions of the sexual reproductive structures are provided for the 16 strains that proved fertile in laboratory conditions. These analyses enabled us to identify seven new Naviculisporaceae species and to establish new combinations for Zopfiella latipes and Apodus oryzae, as Pseudorhypophila latipes comb. nov. and Pseudorhypophila oryzae comb. nov., respectively. Altogether, these findings significantly expand the known diversity of the family.
2. Materials and Methods
2.1. Sample Collection, Fungal Isolation and Availability
Strains CBS 256.59 and CBS 365.69 were purchased from the Westerdjik Institute (https://wi.knaw.nl/). Strains IMI229743, IMI229747 and IMI350600 were purchased from CABI (https://www.cabi.org/). All the others strains were newly isolated for this study. Soil and dung samples were collected and stored at 4°C, until they were processed according to previously described protocols [18,19], to recover potential Sordariales species. Newly obtained strains were isolated by germinating complete asci or batches of several ascospores to recover mat1-1/mat1-2 heterokaryons for heterothallic species. DNAs were extracted using the rapid method previously described in ref. [18] and were subjected to Internal Transcribed Spacer (ITS) sequencing. Taxonomic assignment to the Naviculisporaceae was performed by BLAST analysis of the ITS sequences at the National Center for Biotechnology Information (NCBI) querying the ITS Fungi RefSeq database (https://www.ncbi.nlm.nih.gov/bioproject/PRJNA177353/). Table 1 provides the origin and main characteristics of the strains used in this study.
The holotype specimens for the new species were deposited in the Herbarium of the “Museum National d'Histoire Naturelle” (MNHN, Paris, France). All the newly isolated strains, including the ex-holotype living cultures for the new species, were deposited in the “Centre International de Ressources Microbiennes - Champignons Filamenteux” (CIRM-CF, INRAE, France).
2.2. DNA Isolation for Genome Sequencing, Next Generation Sequencing and Genome Assembly
DNA for genome sequencing by Novogene (Cambridge, United Kingdom) and the Biomics facility (Institut Pasteur, Paris, France) was extracted using the NucleoSpin® Soil from Macherey Nagel (Düren, Germany) and libraries were prepared by the sequencing facilities. For genome sequencing by the JGI, DNA was extracted and libraries were prepared, following “Method 4” described in the supplementary materials of ref. [14].
For genome sequenced by Novogene (Cambridge, United Kingdom) and Biomics (project B16526), DNA was submitted to 2 × 150 bp Novaseq Illumina sequencing. Sequence reads were assembled with Unicycler (Galaxy Version 0.5.0+galaxy1) using the default parameters [20]. Although designed to assemble prokaryotic genomes, this program works well with fungal genomes, as demonstrated by the Busco score (Table S1 and see [2]).
For the genomes sequenced by JGI, Illumina 400 bp Regular Fragment libraries were sequenced using the NovaSeq S4 in 2 × 150 bp paired reads. All raw Illumina sequence data were filtered for artifacts/process contamination using BBTools version 38.79 kmercountexact.sh (http://sourceforge.net/projects/bbmap). Two million reads were subsampled to assemble mitochondria from the filtered reads using the GetOrganelle toolkit [21], and any organelle-matching reads were removed. The remaining reads were then k-mer-matched against the resulting contigs using BBTools version 38.79 with bbduk.sh [22]. An assembly of the target genome was generated using a 20 million read-pair subsample of the resulting non-organelle reads with SPAdes v3.15.2 [23].
For transcriptomes, stranded RNASeq libraries were sequenced using Illumina NovaSeq S4, filtered for artifacts and contaminants using BBDuk, trimmed for quality using the phred Q6, and assembled using Trinity v.2.11.0 [24]. Genomes were annotated using the JGI Annotation pipeline [25].
Raw data and genome assemblies are available under the GenBank Bioproject accession numbers listed in Table S1.
2.3. Phylogenetic Analyses
Table 2 provides the ITS and rDNA Large Subunit (LSU) GenBank accession numbers for the previously known sequences used in the combined ITS+LSU phylogenetic analysis. ITS+LSU sequences of the newly sequenced strains and that of PSN293 were manually extracted from the genome assemblies. Apodospora peruviana CBS 118394 was chosen as the outgroup for this analysis, as it was shown to be the closest relative of PSN293 [14]. The sequences were aligned using MAFFT V7 with the default parameters [26], and maximum likelihood phylogeny was computed by IQ-tree using the default parameters and 1000 ultrafast bootstraps [27].
2.4. Phylogenomic Analyses
We used sequences of Busco v.5.5.0 genes to infer phylogenetic relationships. We identified 2798 single-copy orthologs among BUSCO genes (Benchmarking Universal Single-Copy Orthologs). Sequences for each orthogroup were aligned at the codon level (i.e., keeping sequences in coding reading frame) with TranslatorX 1.1 [29], using MAFFT v7 [26] as the aligner and default parameters for Gblocks 0.91b [30]. Sequences were concatenated, and a maximum likelihood phylogeny was inferred using raxml-ng v.1.2.0 [31] with model GTR+G, ten parsimony starting trees, and 100 bootstrap replicates.
The ANI were calculated using FungANI with the default parameters [2].
2.5. Mating-Type Locus Annotation
The mating-type loci were manually annotated to ensure correct annotations.
3. Results
3.1. Fungal Isolates and Species Delimitation
Table 1 summarizes the dates of isolation, geographical origins and substrate of the strains purchased or isolated for this study. Table 1 also includes the main features of their genome assemblies (detailed statistics of the genome sequences used for phylogenomic analyses are presented in Table S1). As shown in Table S1, genome assemblies were generally of high quality, with BUSCO scores exceeding 97% in all cases, except for the previously published Diplogelasinospora grovesii CBS 340.73 genome, which was used as an outgroup in the phylogenomic analyses.
From most Naviculisporaceae genomes, we were able to retrieve the ITS + LSU region directly from the assemblies. When this region was absent, reads mapping to rRNA loci were extracted with Bowtie2 version 2.5.0 [32] and reassembled with Unicycler [20]. These sequences, combined with ITS + LSU data already available for Naviculisporaceae (Table 2), were used in a first step for species delimitation and assignment to known species. Apodospora peruviana CBS 118394 was used as an outgroup for this analysis, as this strain had previously been shown to be the closest relative of the sequenced PSN293 [14]. For this strain, the ITS and LSU sequences obtained from the genome assembly were identical to those deposited in GenBank (see GenBank accession number in Table 2). Finer species delimitation and assignment were then made by comparing with BLAST some genes of the investigated strains to orthologous ones from their closest relatives present in GenBank. These included the glyceraldehyde 3-phosphate dehydrogenase (gpd) gene, the RNA polymerase II subunit (RPB2) gene, the beta-tubulin (TUB2) gene and the sordarin secondary metabolite gene cluster. Finally, for fine comparison between genome sequences of closely related strains, ANI was calculated with FungANI [2].
Nine strains, PSN1062, PSN850, PSTH81, PSN1167, PSN1104, PSN637, PSN673, PSTH200 and PSN640, were distant from previously sequenced species in the ITS + LSU tree (Figure 1), indicating that they belonged to separate and possibly new species. We were able to obtain fruiting in vitro for five of them: PSN1062, PSTH81, PSN1167, PSN1104 and PSN673. Morphological analysis (Figure 2) showed that the four strains produced sexual structures with distinct and previously undescribed morphologies, making them four species new to science (see taxonomy section), while PSN1062 produced perithecia similar to Podospora araneosa (Cain) Cain [33,34].
PSN1062 ITS had 15 differences out of 334 aligned nucleotides with the ITS of P. araneosa F-116,361, which was used in previous phylogenetic analyses of the Naviculisporaceae (see accession number in Table 2). Accordingly, both strains branched at different positions in the ITS + LSU tree (Figure 1). PSN1062 also had four differences out of 362 aligned nucleotides in the ITS with P. araneosa TNM F17207 (GenBank accession number EF197073.1) and 36 differences in the gpd gene out of 492 aligned nucleotides (GenBank accession number EF197097.1). It did not contain the secondary metabolite gene cluster of Sordaria araneosa ATCC 36386 involved in sordarin biosynthesis (S. araneosa is the basionym of P. araneosa; GenBank accession number LC079035.1). However, it did contain a related cluster, i.e., containing similar genes, albeit with low sequence similarity between the two strains. Overall, these data showed that the P. araneosa strains previously investigated likely belong to at least two different species and that PSN1062 belongs to a third one. Moreover, PSN1062 perithecia were slightly larger than those of the original P. araneosa description (diameter = 935 + 86 µm instead of 550-700 x 400-460 µm) and the ascospores were slightly smaller (spore head = 19.8 + 1.2 × 13.9 + 0.5 µm instead of 24-26 × 16-17 µm; primary appendage = 9.5 + 1.6 × 4.4 + 0.9 µm instead of 11-12 × 6-8 µm), making it unclear whether it was a bona fide P. araneosa. This strain was thus labelled as Podospora aff. araneosa, pending the determination of DNA sequences for the type of P. araneosa TRTC 5211, to allow for the accurate delimitation of P. araneosa.
The four strains for which we could not get fruiting in vitro (PSN850, PSN637, PSTH200 and PSN640) are not formally described here, but may belong to four new species. PSN850 was related to N. terrestris Stchigel, Y. Marín, Cano & Guarro and likely belonged to the same genus; it was thus named Naviculispora sp. Similarly, PSN637 was related to R. myriaspora (P. Crouan & H. Crouan) Y. Marín, A.N. Mill. & Guarro and was thus named Rhypophila sp., PSTH200 was related to G. humicola G.L. Barron and was named Gilmaniella sp. and PSN640 was related to P. mangenotii (Arx & Hennebert) Y. Marín & Stchigel and was thus named Pseudorhypophila sp.
PSN1175 clustered with A. leporinum CBS 365.69 and A. japonense SANK10273 (Figure 1). ANI analysis indicated that PSN1175 and A. leporinum CBS 365.69 represented distinct species, as their genomes were only 93.01% similar and each strain had 8 to 9 % of specific sequences (Figure 3). PSN1175 displayed no difference with A. japonense SANK10273 across 1359 aligned nucleotides of the LSU sequence and five differences across 670 aligned nucleotides for TUB2, leaving its species status unresolved. As PSN1175 did not produce perithecia in vitro, its identification as A. japonense Furuya & Udagawa could not be verified. It is therefore provisionally designated A. aff. japonense.
Strains PSN658, PSN2212 and PSN2105 clustered in the ITS + LSU tree with the previously sequenced PSN293 as well as with R. decipiens CBS 258.69, R. pleiospora TNMF16889 and R. decipiens CBS 256.69 (Figure 1). As shown in Table 3, the ANI values of pairwise comparisons between the genomes of PSN658, PSN2212, PSN2105, CBS 256.69 and PSN293 were much lower than 99%, except for PSN2105 and CBS 256.69, indicating that the strains belonged to different species, except PSN2105 and CBS 256.69.
CBS 256.69 is labelled as R. decipiens in the Westerdjik Institute collection. However, PSN2105 had an ITS sequence identical to that of R. myriaspora CBS 115804 (GenBank accession number DQ166961.1) and produced 64-spored asci typical of R. myriaspora (P. Crouan & H. Crouan) Y. Marín, A.N. Mill. & Guarro (Figure 4). Unfortunately, CBS 256.69 did not fructify in vitro, making it impossible to confirm species assignment by morphological analysis; however, the genome sequence indicated that it is most likely a R. myriaspora strain, and it is designated as such hereafter.
PSN293 was closely related, in terms of nucleotide identity, to R. decipiens CBS 258.69 used in previous analyses of the Naviculisporaceae: we observed no difference in LSU across 1008 aligned nucleotides, one difference across 391 aligned nucleotides in the ITS sequence, three differences across 1198 aligned nucleotides in RPB2 (GenBank accession number AY780187.1) and no difference in TUB2 across 969 aligned nucleotides (GenBank accession number AY780130.1). Moreover, The ITS sequence of PSN293 was identical to that of R. decipiens CBS 113104 across 559 aligned nucleotides (GenBank accession number AY515359.1). It produced 8-spored asci typical of R. decipiens (G. Winter ex Fuckel) Y. Marín, A.N. Mill. & Guarro confirming that this strain actually belongs to this species (Figure 5).
Intriguingly, R. pleiospora TNM F16889, used in a previous phylogenetic analysis of the Naviculisporaceae, had four differences in the ITS across 544 aligned nucleotides with R. pleiospora CBS 113107 (GenBank accession number AY515364.1) and four differences across 550 aligned nucleotides with PSN658. These results suggested that these strains were unlikely to belong to the same species. Moreover, PSN658 had an ITS sequence strictly identical to that of R. pleiospora CBS 113107 across 556 aligned nucleotides. PSN658 also produced the 16-spored asci typical of R. pleiospora (G. Winter) Y. Marín, A.N. Mill. & Guarro (Figure 6). This suggested that the TNM F16889 strain used in previous phylogenetic analysis may not belong to R. pleiospora, but to another related species. Finally, PSN2212 produced 8-spored asci with ascospores that were larger than those of typical R. decipiens, making it a species new to science (see Taxonomy section).
Strains PSTH195, PSQ110, PSN2406, PSN540, PSN2022, PSN2407 and PSN2009 clustered with Zopfiella latipes IMI350600 and IFO9826, P. mangenotii IMI229747 and CBS419.67, P. pilifera CBS 413.73, P. marina IMI229743, CBS 155.77 and CBS 698.96, P. formosana NTUPPMCC 22-297, and Apodus oryzae CBS 376.74 (Figure 1). ANI analyses for these strains (Table 4) yielded values much lower than 99%, except for PSQ110 / IMI350600 and PSN540 / PSN 2022. In the latter case, the ANI value was lower than 99%, but this was due to the presence in the genome of a substantial proportion (approximately 4%) of highly divergent sequences with a similarity of 85% to 90% (Figure 7). The two strains shared mostly very similar sequences with a percentage of identity higher than 99.5% and contained few specific sequences (about 1% of their genome), indicating that they belonged to the same species (Figure 7).
Morphological analyses confirmed that IMI350600 and PSQ110 belong to Z. latipes (N. Lundq.) Malloch & Cain (Figure 8 and Figure S1), as expected for IMI350600 since it is the ex-type strain for this species. Since Z. latipes clearly branched with the known Pseudorhypophila species in the ITS and LSU tree (Figure 1), it was renamed as P. latipes (N. Lundq.) Tangthirasunun & Silar comb. nov. Note that IMI350600 and PSQ110 showed no differences across 559 aligned nucleotides in their ITS compared to Z. latipes IFO9826 previously used in a phylogenetic analysis of Zopfiella and allied genera [36]. There were two differences across 842 aligned nucleotides in their LSU and seven differences in TUB2 across 491 aligned nucleotides (accession number AY999146.1), suggesting it is unlikely that these strains belong to the same species. Especially, P. formosana, a recently identified species [13], branched in the ITS + LSU tree between IFO9826 and IMI350600 / PSQ110 (Figure 1).
PSN540, PSN1022, PSN2009 and PSN2407 branched with CBS 376.74, the ex-type strain of A. oryzae (Figure 1). PSN540, PSN1022, had three differences out of 550 aligned nucleotides in their ITS sequence with A. oryzae CBS 376.74, no differences out of 839 aligned nucleotides in their LSU region and three differences out of 492 aligned nucleotides in TUB2 (GenBank accession num AY681234.1). PSN2407 showed with A. oryzae CBS 376.74 no difference in the ITS across 550 aligned nucleotides, one difference across 839 aligned nucleotides in the LSU region and seven differences in TUB2 across 492 aligned nucleotides. PSN2009 had with A. oryzae CBS 376.74 zero differences in the ITS and LSU regions and a single difference in TUB2 out of 492 aligned nucleotides. In line with its close proximity to A. oryzae CBS 376.74 based on sequence comparisons, morphological analysis confirmed that PSN2009 exhibited a morphology typical of A. oryzae (Figure 9). Hence, PSN2009 was assigned to A. oryzae, while PSN540 and PSN2022 belonged to a closely related species (see Taxonomy section) and PSN2407 to another separate new species. Unfortunately, PSN2407 did not fructify in vitro, preventing us from assessing whether it was a species new to science or not. It was thus provisionally called Pseudorhypophila sp. A. oryzae was renamed as P. oryzae (Carolis & Arx) Tangthirasunun & Silar comb. nov., owing to its branching with other Pseudorypophila species in the ITS + LSU tree. Note that A. deciduus strain CBS 506.70, belonging to the type species for the Apodus genus, Malloch & Cain, is in the Schizotheciaceae [38], a position confirmed by whole genome sequence analysis (unpublished data).
Morphological analysis confirmed that IMI229747 and IMI229743 belonged to P. mangenotii and P. marina, respectively (Figure 10 and Figure 11). Consistent with this assignment, IMI229747 showed no difference compared to P. mangenotii CBS 419.67 across 891 aligned nucleotides in the LSU region, no difference across 484 aligned nucleotides in the ITS, no difference in TUB2 (GenBank accession num KP981571.1) across 618 aligned nucleotides and one difference across 1022 aligned nucleotides in RPB2 (GenBank accession num KP981627.1). Similarly, IMI229743 showed with P. marina CBS 155.77 no difference out of 1052 aligned nucleotides in its ITS+LSU region and no difference out of 849 aligned nucleotides in RPB2 (GenBank accession number MK876813.1); it also showed no difference out of 1052 aligned nucleotide in the ITS+LSU region with P. marina CBS 698.96 and one difference out of 831 aligned nucleotides in RPB2 (GenBank accession number MK876815.1; the species is labelled as Zopfiella submersa in GenBank).
Morphological analysis also showed that PSN2406 is a new species (see taxonomy section). Finally, PSTH195 did not fructify in vitro making it difficult to determine whether it was a species new to science. It was thus provisionally named Pseudorhypophila sp., as it appeared closely related to the other Pseudorhypophila species.
Overall, we generated complete genome sequences for:
- (1)
- six previously known species, two of which are newly placed in the Naviculisporaceae: R. myriaspora (PSN2105 and likely CBS 256.69), R. pleiospora (PSN658), P. latipes (formerly Z. latipes; IMI350600 and PSQ110), P. mangenotii (IMI229747), P. marina (IMI229743) and P. oryzae (formerly A. oryzae; PSN2009), adding to the previously sequenced R. decipiens (PSN293).
- (2)
- seven new species including five Rhypophila species: R. thailandica sp. nov. (PSTH81), R. reunionensis sp. nov. (PSN1167), R. brasiliensis sp. nov. (PSN1104), R. camarguensis sp. nov. (PSN673) and R. alpibus sp. nov. (PSN2212), as well as two Pseudorhypophila ones: P. guyanensis sp. nov (PSN2406), and P. gallica sp. nov. (PSN540 and PSN2022).
- (3)
- eight newly-isolated strains awaiting further characterization, some of which may belong to species new to science: P. aff. araneosa PSN1062, Naviculispora sp. PSN850, A. aff. japonense PSN1175, Rhypophila sp. PSN637, Gilmaniella sp. PSTH200, Pseudorhypophila sp. PSN640, Pseudorhypophila sp. PSN2407 and Pseudorhypophila sp. PSTH195.
- (4)
- one collection strain: A. leporinum: CBS 365.69.
3.2. Phylogenomic Analysis
As shown in Figure 1, statistical supports for many nodes in the ITS + LSU tree were very low. We thus computed a phylogenomic tree using using the concatenation of sequences of 2,798 BUSCO genes. We used as outgroups some representatives of major families of Sordariales (Podospora anserina for the Podosporaceae, Neurospora crassa for the Sordariaceae and Shizothecium tetrasporum for the Schizotheciaceae), as well as Diplogelasinospora grovesii. This later species belongs to the Diplogelasinosporaceae, the most divergent family of the Sordariales [14] and was used to root the tree (Figure 12A). All nodes received strong support, with 100% bootstrap values. As in the ITS + LSU tree, Pseudorhypophila and Rhypophila were each recovered as monophyletic genera. However, the relationships between the different species within each genus were distinct from those in the ITS + LSU tree. Additionally, Gilmaniella sp. PSTH200 branched between these two genera, closer to Pseudorhypophila, in both tree. Yet, the branch length in the phylogenomic tree is short, making it unclear whether it was a genus distinct from Pseudorhypophila. The two Arnium species clearly defined a separate genus, whose naming will await the molecular characterization of A. lanuginosum Nitschke, the type species for the genus. P. aff. araneoase PSN1062 and Naviculispora sp. PSN850 were sisters to all the other strains.
A noticeable difference between the Rhypophila and Pseudorhypophila species is their genome sizes: between 50 and 56 Mb for Pseudorhypophila, except for the most divergent strain of the genus (PSN640, with 47.5 Mb), and between 45 and 50 Mb for Rhypophila species (Table 1 and Table S1). Overall, all Naviculisporaceae species tended to have larger genome sizes exceeding 45 Mb, compared to the other Sordariales, which typically had genome sizes ranging from 35 to 40 Mb.
As shown in Table 1, the Rhypophila species were mostly recovered from dung, while the Pseudorhypophila ones were isolated from soils. The former differentiated ostiolate perithecia and the latter mostly non-ostiolate ones. This is consistent with the hypothesis that dispersing their ascospores over long distances from the growing substrate may be advantageous to dung-inhabiting fungi, while soil fungi ascospores might only be needed to persist within the soil. Therefore, they may not benefit from actively dispersing their ascospores, as this could be an energy-consuming process.
3.3. Mating-Type Locus Analysis
As shown in Figure 12B, some strains did not fructify in vitro under the tested conditions (i.e., at 27°C, with 24h of light and on six different media: M2, Oat Meal Agar, V8, M0+miscanthus and M0+hay). One possibility was that these strains were heterothallic and that the isolates we recovered carried a single mating type, although complete asci or batches of several ascospores were used to initiate cultures. To test this possibility, mating-type loci were manually annotated (Figure 12C), and putative breeding systems were inferred from the structures of the loci (Figure 12C). Three kinds of mating type-locus structures were observed.
Firstly, most strains (17 out of 26) contained both the genes for the mat1-1 idiomorph (mat1-1-1, mat1-1-2 and mat1-1-3) and the mat1-2-1 gene of the mat1-2 idiomorph, clustered together and flanked by the APN2 gene on one side and the SLA2 gene on the other (Figure 12C). Note that, in some cases, although the locus was split onto different contigs, the complete locus could nonetheless be reassembled because the mat genes were located at the edges of contigs. These contained thus the mating-type locus previously described for homothallic Sordariales such as Sordaria macrospora [42,43,44]. Most of these strains readily fructified in vitro. The exceptions were CBS 356.69, which had been kept in vitro for a long time, PSN2022, which fructified in an unreproducible manner, as well as PSN637 and PSN2407. These results altogether suggested that these strains were likely homothallic, but that the three, possibly four, last -mentioned ones required special conditions to fructify, not met in vitro.
Secondly, three strains (PSN850, CBS 365.69 and PSN1167) contained the two idiomorphs on different contigs, with a single one bordered by APN2 and SLA2 (Figure 12C). Such structure could be due to a usunusal homothallic mating-type locus structure in which the two idiomorphs would not be linked, or to heterothallism. Indeed, because we sequenced putative mat1-1/mat1-2 heterokaryons, one would expect the two idiomorphs to be on different contigs, each bordered by APN2 and SLA2. However, assembly programs do not perform well on such heterozygous loci and they may have yielded mating-type locus structures in which only one of the idiomorphs is bordered by APN2 and SLA2, the other one being located in a small independent contig. The two possiblities could in principle be differentiated by analyzing the fertility of single-spore isolates. Only PSN1167 permitted such analysis, as the other two strains were sterile. All the eight F1 single spore isolates of PSN1167 were fertile, indicating that the strain was homothallic. Moreover, sequence reads across APN2 (184 reads/kbp) and SLA2 (195 reads/kbp) did not display higher coverage than those for mat1-1-1 (196 reads/kbp), mat1-1-2 (208 reads/kbp), mat1-1-3 (224 reads/kbp) and mat1-2-1 (206 reads/kpb), indicating that all six genes were were present in the same number of copies, as expected if PSN1167 was homothallic. Indeed, one would expect in heterothallics APN2 and SLA2 to have a higher coverage of sequence reads than the mating-type genes, i.e., to have the coverage of the mat1-1 idiomorph added to the coverage of the mat1-2 idiomorph. Coverage calculation for the two sterile strains, PSN850 and CBS 365.69, also suggested that they could be homothallics, although we could not test this directly Indeed, the coverage calculations for PSN850 gave 202 reads/kbp for APN2, 226 reads/kbp for SLA2, 246 reads/kbp for mat1-1-1, 242 reads/kbp for mat1-1-2, 217reads/kbp for mat1-1-3, reads/kbp for 238 mat1-2-1, and for CBS 365.69 1241 reads/kbp for APN2, 1090 reads/kbp for SLA2, 1289 reads/kbp for mat1-1-1, 1289 reads/kbp for mat1-1-2, 1216 reads/kbp for mat1-1-3, and 1019 reads/kbp for mat1-2-1
Finally six strains (PSN1062, PSN1175, PSN1104, PSTH200, PSN640 and PSTH195) had a single idiomorph, either mat1-1 or mat1-2 bordered by APN2 and SLA2. Such structures could be due to heterothallism with loss of one mating type in the sequenced isolate. It could also correspond to a particular form of homothallism, as some rare fungal species can undergo single-mating-type mating [43]. This could only be tested for the fertile PSN1062 and PSN1104. Mycelia from all F1 single spores isolated from PSN1062 (n=5) produced progeny, showing that this strain underwent sexual reproduction with only the mat1-2 mating type. On the contrary, all mycelia from all single spore isolates from PSN1104 (n=8) were sterile; albeit, five out of eight differentiated small and barren perithecia. This showed that PSN1104 was heterothallic. Lack of the mat1-1 mating type in the sequence assembly of PSN1104 can be accounted for by a biased ratio of nuclei, with the mat1-1 muclei in too low amounts to contribute significantly to the extracted DNA, while even minute amount of mat1-1 nuclei can permit mating. For the remaining four strains, the best explanation for their sterility is that these strains are heterothallic and that the cultures we have obtained have lost one mating type.
3.4. Taxonomy
Pseudorhypophila gallica, Tangthirasunun & Silar sp. nov. Figure 13
Index Fungorum number: to be obtained
Etymology: from the location of first two isolations
Holotype: PC0820273, ex-type: PSN540 isolated from soil collected under bamboo in the public garden of Albert (Somme, Picardie), France in 2020; available under accession n° to be obtained at the “Centre International de Ressources Microbiennes - Champignons Filamenteux” (CIRM-CF, INRAE, France). Whole genome sequence available in GenBank under bioproject PRJNA1081049 for genomic data & PRJNA1339018 for transcriptomic data.
Other isolate: PSN2022 from St Maur sur le Loir (Eure et Loir), France– this strain did not reproducibly produce fruiting bodiest in vitro.
Description: Perithecia were readily differentiated on M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination; non-ostiolate, spherical, superficial or semi-immersed, 761 ± 116 µm diam (n=10); Their peridium was pale brown and semi-translucid, tissue type was textura angularis. Paraphyses absent. Asci were 8-spored, clavate with a tapering and narrow apex. Ascospores were biseriate and bicellular. Ascospore heads were 25.4 ± 2.5 × 11.8 ± 1.6 μm ((21.5-)30.7 × 9.2(-17.1) μm, n=30), ellipsoidal to oval, hyaline at first, then green and brown and dark brown when mature. Few ascospore with 1-septate (slightly curved or straight) toward the apical, first median cell from base 14.8 ± 2.5 µm and the second median cell 10.0 ± 2.0 μm (n=15).
Habitat & Distribution: soil; this species has been found twice in two different regions of France: Picardie and Centre-Val de Loire.
Breeding system: homothallic
In the ITS + LSU tree, the closest relative of P. gallica is P. oryzae CBS 376.74 and PSN2009 (Figure 1). In the phylogenomic tree, the clostest relative is R. oryzae PSN2009 (Figure 12). They differ by the fact that the ascospores are at first biseriate in P. gallica and uniseriate in P. oryzae. Also the ascospores of P. gallica are more elongated than those of P. oryzae: 25.4 ± 2.5 × 11.8 ± 1.6 μm versus 21.7 ± 2.2 × 13.2 ± 1.1 μm, respectively.
Pseudorhypophila guyanensis, Tangthirasunun & Silar sp. nov. Figure 14
Index Fungorum number: to be obtained
Etymology: from the location of first isolation
Holotype: PC0820276, ex-type: PSN2406 isolated from soil taken in 2024 under a tree in a disturbed rainforest in French Guyana; available under accession n° to be obtained at the “Centre International de Ressources Microbiennes - Champignons Filamenteux” (CIRM-CF, INRAE, France). ). Whole genome sequence available in GenBank under Bioproject PRJNA1345941.
Description: Perithecia were readily differentiated on M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination; ostiolate, pyriform, superficial or semi-immersed,499 ± 66 µm diam (n=10); their neck was black and slightly elongated. Their peridium was pale brown and semi-translucid, tissue type was textura angularis. Paraphyses absent. Asci were 8-spored, clavate with a tapering and narrow apex. Ascospores were biseriate and bicellular. They were at first vermiform, then the head enlarged and pigmented to yield Cercophora-like ascospores. Ascospore heads were 17.0 ± 1.0 × 9.6 ± 1.2 µm ((15.0-)18.6 × 6.4(-11.1) µm, n=30), ellipsoidal with a slightly flattened base, hyaline at first, then green and black when mature. The primary appendage was very long (often longer than the spore head,), flat, often curved, frequently collapsing, 23.4 ± 2.2 × 5.6 ± 0.9 µm ((18.0-)25.5 × 4.6(-7.2) µm, n=10), Secondary appendages were present at the apex of the spore head as a 1-whip structure and at the end of the primary appendage as a whip.
Habitat & Distribution: soil; this species has been found once from French Guyana.
Breeding system: homothallic
P. guyanensis can be easily differentiated from the other Pseudorhypophila spp. by the fact that it differentiates ostiolate perithecia, like P. mangenotii. However, unlike P. mangenotii, which differentiate triangular ascospores with a rounded primary appendage, P. guyanensis differentiates Cercophora-like ascospores.
Rhypophila alpibus, Tangthirasunun & Silar sp. nov. Figure 15
Index Fungorum number: to be obtained
Etymology: from the location of first isolation
Holotype: PC0820272, ex-type: PSN2212 isolated from cow dung collected in a mountain pasture in the Alps of Italy in 2024; available under accession n° to be obtained at the “Centre International de Ressources Microbiennes - Champignons Filamenteux” (CIRM-CF, INRAE, France). ). Whole genome sequence available in GenBank under bioproject PRJNA1345941.
Description: Perithecia were readily differentiated on M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination; they were ostiolate, pyriform, superficial or semi-immersed, covered with numerous hairs, 592 ± 100 µm diam (n=10); their neck was black and slightly elongated. Their peridium was pale brown and semi-translucid, tissue type was textura angularis. Paraphyses absent. Asci were 8-spored, clavate with a tapering and narrow apex,. Ascospores were biseriate and bicellular. Ascospore heads were 42.9 ± 2.2 × 21.8 ± 1.1 µm ((39.0-)47.1 × 19.5(-24.2) µm, n=30), ellipsoidal with a slightly flattened base, hyaline at first, then green and black when mature. The primary appendage was very long (often longer than the spore head) and flat, frequently collapsing, 50.5 ± 9.8 × 7.5 ± 1.8 µm ((36.4-)64.2 × 5.3(-10.8) µm, n=7), Secondary appendages were present at the apex of the spore head as a fan-shaped structure of juxtaposed fibrils and at the junction between the spore head and the primary appendage as fibrils often two in number.
Habitat & Distribution: dung; this species has been collected only once in the Alps mountains.
Breeding system: homothallic
Both in the ITS + LSU and phylogenomic tree, the closest relatives of R. alpibus are R. camarguensis, R. decipiens, R. pleiospora and R. myriaspora. Like R. decipiens and R. camarguensis, R. alpibus differentiated 8-spored asci, while R. pleiospora and R. myriaspora differentiated 16-spored and 64-spored asci, respectively. The ascospore heads of R. alpibus (42.9 ± 2.2 × 21.8 ± 1.1 µm) are larger than those of R. decipiens (35.5 ± 2.8 × 18.6 ± 1.2 µm) and R. camarguensis (36.0 ± 2.9 × 18.9 ± 1.3 µm).
Rhypophila brasiliensis, Tangthirasunun & Silar sp. nov. Figure 16
Index Fungorum number: to be obtained
Etymology: from the location of first isolation
Holotype: PC0820275, ex-type: PSN1104 isolated from capybara dung collected in the Parc Taquaral, Campinas, Brazil in 2021; available under accession n° to be obtained at the “Centre International de Ressources Microbiennes - Champignons Filamenteux” (CIRM-CF, INRAE, France). ). Whole genome sequence available in GenBank under bioproject PRJNA1345941.
Description: Perithecia were readily differentiated on M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination; they were ostiolate, pyriform, superficial or semi-immersed, and 498 ± 44 µm diam (n=10); their neck was black and slightly elongated. Their peridium was pale brown and semi-translucid, tissue type was textura angularis. Paraphyses absent. Asci were 8-spored, clavate with a tapering and narrow apex,. Ascospores were biseriate and bicellular. Ascospore heads were 33.2 ± 2.0 × 17.2 ± 1.6 µm ((28.1-)38.4 × 14.8(-22.5) µm, n=30), ellipsoidal with a slightly flattened base, hyaline at first, then green and black when mature. The primary appendage was very long (often longer than the spore head) and flat, frequently collapsing, 34.6 ± 4.1 × 7.3 ± 0.7 µm ((28.7-)41.1 × 5.9(-8.1) µm, n=10), Secondary appendages were present at the apex of the spore head as a whip-shaped or fibrillate or lamellate structure. No obvious appendage(s) at the base of the spore head.
Habitat & Distribution: dung, isolated once in Brazil.
Breeding system: heterothallic
In the ITS + LSU, the closest unequivocal relative of R. brasiliensis was R. cochleariformis. They differ by the number of ascospores per ascus (eight for R. brasiliensis and 128 for R. cochleariformis; [45]). In the phylogenomic tree, R. brasiliensis is related to R. reunionensis and R. thailandica. R. reunionsensis produced 64-spored asci and R. thailandica 8-spored asci. However the ascospore heads of R. brasiliensis are more elongated than those of R. thailandica: 33.2 ± 2.0 × 17.2 ± 1.6 µm versus 31.9 ± 3.3 × 19.8 ± 1.7 µm.
Rhypophila camarguensis, Tangthirasunun & Silar sp. nov. Figure 17
Index Fungorum number: to be obtained
Etymology: from the location of first isolation
Holotype: PC0820277, ex-type: PSN673 isolated from cow dung collected in 2021 in Camargue, France; available under accession n° to be obtained at the “Centre International de Ressources Microbiennes - Champignons Filamenteux” (CIRM-CF, INRAE, France). ). Whole genome sequence available in GenBank under bioproject PRJNA1345941.
Description: Perithecia were readily differentiated on M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination; they were ostiolate, pyriform, superficial or semi-immersed, covered with numerous hairs, 510 ± 51 µm diam (n=10); their neck was black and slightly elongated. Their peridium was pale brown and semi-translucid, tissue type was textura angularis. Paraphyses absent. Asci were 8-spored, clavate with a tapering and narrow apex,. Ascospores were biseriate and bicellular. Ascospore heads were 36.0 ± 2.9 × 18.9 ± 1.3 µm ((27.8-)39.7 × 16.5(-21.0) µm, n=30), ellipsoidal with a slightly flattened base, hyaline at first, then green and black when mature. The primary appendage was very long (often longer than the spore head) and flat, frequently collapsing, 54.0 ± 4.6 × 7.4 ± 0.8 ((46.0-)58.9 × 5.9(-8.5) µm, n=10), Secondary appendages were present at the apex of the spore head as a fan-shaped structure of juxtaposed fibrils and at the junction between the spore head and the primary appendage as fibrils often two in number.
Habitat & Distribution: dung; collected once in the South of France
Breeding system: Homothallic
In the ITS + LSU and phylogenomic trees, R. camarguensis is closely related to the other 8-spored species of Rhypophila, R. decipiens and R. alpibus. Its ascospore heads are smaller than those of R. alpibus: 36.0 ± 2.9 × 18.9 ± 1.3 µm versus 42.9 ± 2.2 × 21.8 ± 1.1 µm and are similar to those of R. decipiens, which are 35.5 ± 2.8 × 18.6 ± 1.2 µm. R. decipiens and R. camarguensis can easily be differentiated by their mycelium, which is darker on M2 medium for R. camarguensis and by their genome sequence. Especially, their ITSs differ by 15 differences.
Rhypophila reunionensis, Tangthirasunun & Silar sp. nov. Figure 18
Index Fungorum number: to be obtained
Etymology: from the location of first isolation
Holotype: PC0820274, ex-type: PSN1167 isolated from cow dung collected near Piton de la Fournaise in Ile de la Réunion, oversea France in the Indian Ocean; available under accession n° to be obtained at the “Centre International de Ressources Microbiennes - Champignons Filamenteux” (CIRM-CF, INRAE, France). ). Whole genome sequence available in GenBank under bioproject PRJNA1345941.
Description: Perithecia were readily differentiated on M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination; There were covered with hairs, ostiolate, pyriform, superficial or semi-immersed, 558 ± 63 µm diam (n=10); their neck was black and slightly elongated. Their peridium was pale brown and semi-translucid, tissue type was textura angularis. Paraphyses absent. Asci were 64-spored, clavate with a tapering and narrow apex. Ascospores were irregularly arranged., Ascospore heads were 19.6 ± 1.3 × 12.8 ± 0.8 µm ((16.9-)22.1 × 11.5(-14.6) µm, n=30), ellipsoidal with a slightly flattened base, hyaline at first, then green and black when mature. The primary appendage was very long (often longer than the spore head) and flat, frequently collapsing, 30.8 ± 5.7 × 4.9 ± 1.0 µm ((19.5-)40.6 × 2.9(-6.1) µm, n=10), Secondary appendages were present at the apex of the spore head as a rot-shaped, apical gelatinous caudae absent or present.
Habitat & Distribution: dung; collected only once in Ile de la Réunion in the Indian Ocean.
Breeding system: homothallic
R. reunionensis is related to R. myriaspora, the other 64-spored species known in the Rhypophila genus. It has smaller ascospore heads than R. myriaspora: 19.6 ± 1.3 × 12.8 ± 0.8 µm versus 27.3 ± 2.8 × 17.1 ± 1.2 µm.
Rhypophila thailandica, Tangthirasunun & Silar sp. nov. Figure 19
Index Fungorum number: to be obtained
Etymology: from the location of first isolation
Holotype: PC0820271, ex-type: PSTH81 isolated from elephant dung collected in Ayuthaya, Thailand, in 2023; available under accession n° to be obtained at the “Centre International de Ressources Microbiennes - Champignons Filamenteux” (CIRM-CF, INRAE, France). Whole genome sequence available in GenBank under bioproject PRJNA1345941.
Description: Perithecia were readily differentiated on M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination; they were ostiolate, pyriform and 527 ± 103 µm diam (n=10); their neck was black and slightly elongated. Their peridium was pale brown and semi-translucid, tissue type was textura angularis. Paraphyses were absent. Asci were 8-spored, clavate with a tapering and narrow apex. Ascospores were biseriate and bicellular. Ascospore heads were 31.9 ± 3.3 × 19.8 ± 1.7 µm ((25.2(-41.9) × 16.1-(22.9) µm, n=30), ellipsoidal with a slightly flattened base, hyaline at first, then green and black when mature. The primary appendage was very long (often longer than the spore head) and flat, frequently collapsing, 49.3 ± 3.3 × 7.2 ± 1.5 µm ((42.6(-54.0) × 5.4-10.3) µm, n=10), Secondary appendages were present at the apex of the spore head as a whip-shaped or fibrillate or lamellate structure.
Habitat & Distribution: dung, isolated once in Thailand.
Breeding system: homothallic
In the ITS + LSU tree, the closest relative of R. thailandica is R. cochleariformis (Figure 1). In the phylogenomic tree, the clostest relative is R. reunionensis (Figure 12). The three species differ by the number of ascospores per ascus eight for R. thailandica, 64 for R. reunionensis and 128 for R. cochleariformis.
Pseudorhypophila latipes (N. Lundq., ex Malloch & Cain) Tangthirasunun & Silar IF= To be obtained Figure 8
Basionym : Tripterospora latipes N. Lundq., IF=340467
Synonym : Zopfiella latipes (N. Lundq.) Malloch & Cain, IF=325683
Material examined: living cultures of IMI350600 and PSQ110, whose genome sequences are available in GenBank under bioproject PRJNA1345941.
Morphological analysis of both IMI350600 and PSQ110 is consistent with the original description of Tripterospora latipes Lundq. IMI350600 is the ex-type of the species. Phylogenetic analysis based on 2798 genes clearly places both strains in the Pseudorhypophila genus. This species is homothallic.
Pseudorhypophila oryzae (Carolis & Arx) Tangthirasunun & Silar IF= To be obtained Figure 9
Basionym : Apodus oryzae Carolis & Arx, IF=308867
Material examined: living cultures of PSN2009, whose genome sequence is available in GenBank under bioproject PRJNA1345941.
Morphological analysis of PSN2009 is consistent with the original description of Apodus oryzae Carolis & Arx. PSN2009 has with A. oryzae CBS 376.74 no difference in its ITS and LSU, and a single difference in TUB2. Phylogenetic analysis based on 2798 genes clearly places this strain in the Pseudorhypophila genus. This species is homothallic.
4. Discussion
Sordariales is an important order of fungi [17]. It hosts species producing enzymes and secondary metabolites that are of interest for industrial purposes [17], including some made by several thermophilic fungi [46,47,48]. It also hosts three important models used in genetical and biochemical studies: N. crassa, Sordaria macrospora and P. anserina, putting them among the best-known fungi [17]. Yet, although species of Sordariales are among the most frequent species present in soils [15] and dung [16], the overall diversity of the order is poorly known. Especially, several well-studied species were found to be complexes of cryptic species [4,5,6,7,8,9,10]. Here, we tackle through a combination of genome sequences and morphological analyses the diversity of Naviculisporaceae, one of the families hosted by Sordariales.
Firstly, by comparing the DNA barcodes (ITS, LSU, TUB2, RPB2, gpd…) extracted from our genome sequences to that of previously characterized strains, we found that many isolates attributed to the same species presented high levels of nucleotide differences. This might result from sequencing errors or be the results of misidentifications in the collections, as previously exemplified [4]. However, sequence differences could also be indicative of the presence of complexes of cryptic species. The availability of only limited barcode sequences is not enough to disentangle the different possibilities, especially when the isolates do not fruit or no living culture is available, so that the morphology of the strain cannot be compared with the original description. In the case of Sordariales, the task is rendered more complex by the fact that some species, which were described a long time ago, have missing holotypes, insufficient description in the protologue or holotypes of historical values or so poor condition that they cannot be used to extract DNA. Here, for example, we suggest that strain IFO 9826 may not belong to P. latipes and that the strains of P. araneosa previously analyzed (F-116,361, TNM F17207 and ATCC 36386) and our newly isolated strain PSN1062 may belong to different species. Availability of complete genome sequence can on the contrary settle some issues. Indeed, comparison using FungANI [2] of the genome sequence of CBS 256.69 (labelled as R. decipiens in the collection) to that of PSN2105, for which morphology has been determined, shows that both belong to the same species, R. myriaspora. The diminishing cost of Next Generation Sequencing and the ease with which raw data can be assembled using Unicycler that freely runs on Galaxy platforms make it possible for any mycologist with a limited knowledge of bioinformatics to obtain genome sequence useful for species delimitation. The determination of the barcodes that are presently recommended for fungal identification [49] is often time consuming, as it often requires several PCR amplifications in different conditions to get a significant set of barcodes (nowadays studies often determine four barcodes: ITS, LSU, TUB2 and RPB2). Then Sanger sequencing must be done. While the Sanger sequencing can be externalized, PCR amplifications usually cannot. Overall, with slightly higher cost and less intensive internal work, genome sequences can be used for species delimitation with little efforts and provide a wealth of additional data, including for example the possibility to determine structure of the mating type loci. We therefore advocate that complete genome becomes the gold standard for the description of new species.
Analyses of the mating-type loci showed that most strains had mating type loci similar to homothallic Sordariales such as S. macrospora [42], with the mat1-1 and mat1-2 idiomorphs located at the same position in the genome between the APN2 and SLA2 genes. However, some homothallic Naviculisporaceae had atypical mat loci with the two haplotypes at two different locations in the genome. This situation is not unprecedented and has also been observed in homothallic Chaetomiaceae, another Sordariales family [50]. Additionally, one Naviculisporaceae strains underwent sexual reproduction with only the mat1-2 mating type. Among the ten strains that would not fructify in vitro, four seemed to carry only one of the two idiomorphs of heterothallics, arguing that their sterility in vitro might primarily be due to lack of an appropriate mate to initiate sexual reproduction. However, the other sterile strains appeared to carry the whole complement of mat genes (i.e., mat1-1-1, mat1-1-2, mat1-1-3 and mat1-2-1), suggesting that they required special conditions to fructify that we were not able to provide. These may include the presence of special metabolite(s) or microorganism(s) present in their natural environment.
In addition to the genomes for the ex-types of the new species described here, our study provides genome references for well-known species such as R. myriaspora and P. mangenotii, which are the type species of Rhypophila and Pseudorhypophila, respectively. Phylogenomic analysis clearly confirms the separation of the two genera. It also suggests that additional data (i.e., the determination of the genomes and morphology of additional species) are required for Gimaniella to be definitively validated; so far, the two strains known for this genus do not undergo sexual reproduction in vitro. Additional strains are also needed to get a better idea of the diversity of the genus Navicula and to get some sequences for the genus Areotheca that we have not encountered during our strain isolation.
As shown in Figure 1, many species lack an identified type for which sequences are available. Many of these were described a long time ago. Their original description might be too scant to accurately identify them and/or a type is missing, is of historical value or in too poor condition to make a reliable identification (see for example refs [16,17]). Here, we introduce four 8-spored Rhypophila species that closely resemble R. decipiens: R. alpibus, R. brasiliensis, R. camarguensis and R. thailandica. All five species differentiate very similar fruiting bodies and asci containing biseriate ascospores having a very long primary appendage. They appear to differ by the sizes of the ascospore heads, shape and presence of the secondary appendages, and/or mycelium morphology. Similarly, we introduce R. reunionsensis, another 64-spored species closely resembling R. myriaspora, but with smaller ascospore heads and P. gallica an 8-spored species closely resembling P. oryzae having more elongated ascospores. All these species can be clearly differentiated by their genome sequences. The phylogenomic tree shows that the 64-spored species, along with the 16-spored R. pleiospora, are intermingled with the 8-spored ones. Indicating recurrent changes in the number of ascospores per ascus. Because a single strain for most species has been analyzed, we do not know the intraspecific variability of the size of the ascospores, which in other Sordariales species can be substantial [4,10]. More strains for each species must therefore be analyzed before fully accepting this character as discriminant. Moreover, the Sordariales species that have been thoroughly investigated turned out to actually be complexes of cryptic species, indistinguishable by looking at the morphology of the sexual reproductive structures [4,5,6,7,8,9,10]. So far, we have sequenced one or two strains for each Naviculisporaceae species, which is insufficient to detect complexes of closely-related cryptic species. We thus advocate that before epitypification, one awaits the sequences of several additional strains. We also advocate that more than four DNA barcodes should be determined for the types of some species, such as P. araneosa or A. japonense, as to unequivocally identify newly isolated strains, such as PSN1062 and PSN1175.
5. Conclusions
Through the isolation of new and collection strains, combined with genome sequencing and morphological analyses, our study improves the knowledge of the diversity of Naviculisporaceae. Genome sequences are provided for seven previously known species, some of which are commonly found on dung or in soil (e.g., R. myriaspora and P. mangenotii, the type species for the Rhypophila and Pseudorhypophila genera), in addition to that of the widespread and common R. decipiens that was previously published. Seven species new to science are described and eight strains, which may belong also to species new to science, await further characterization. A phylogenomic tree of the family with 100% statistical support for all nodes is provided.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Morphology of Pseudorhypophila latipes PSQ110; Table S1: Main features of the genomes used for phylogenomic analysis.
Author Contributions Conceptualization, N.T., P.G., T.G. & P.S..; investigation, N.T., C.L., V.G., P.S.; resources, R.H., S.C., R.R., J.P., A.L., V.N., I.V.G, C.L. & P.S..; data curation, P.S.; writing—original draft preparation, N.T. & P.S..; writing—review and editing, P.G. & T.G.; supervision, I.V.G, P.S.; funding acquisition, P.G., T.G. & P.S. All authors have read and agreed to the published version of the manuscript.
Funding
The work conducted at University Paris Cité was funded by intramural funding from Université Paris Cité. Narumon Tangthirasunun was funded by The Academic Melting Pot grant, which was supported by King Mongkut’s Institute of Technology Ladkrabang (grant number KREF206612). The work (proposal: 10.46936/10.25585/60001199) conducted by the U.S. Department of Energy Joint Genome Institute (https://ror.org/04xm1d337), a DOE Office of Science User Facility, is supported by the Office of Science of the U.S. Department of Energy operated under Contract No. DE-AC02-05CH11231.
Data Availability Statement
All newly isolated strains can be purchased at CIRM-CF (https://www.cirm-fungi.fr/). NGS Raw data are available in the GenBank SRA repository (see Table S1 for accession numbers) and/or the JGI MycoCosm website (https://mycocosm.jgi.doe.gov). Assemblies can be downloaded from a GitHub repository (https://github.com/podo-gec/fungi-public-data/tree/master/naviculisporaceae) or the JGI MycoCosm website (https://mycocosm.jgi.doe.gov).
Acknowledgments
We would like to thank Antonio Camara, Jérôme Letty, Violaine Llaurens, Jacqui Shykoff, Camille Silar, Greg Thorn and Catherine Villard for providing soil and dung samples and Aurélien Renault for his technical expertise. We would like to thank for genome sequencing Yakov Vitrenko and Laure Lemée, Biomics Platform, C2RT, Institut Pasteur, Paris, France, supported by France Génomique (ANR-10-INBS-09) and IBISA.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ANI | Average Nucleotide Identity |
| ITS | Internal Transcribed Spacer |
| kbp | Kilobase-pair |
| LSU | rDNA Large Subunit |
| RPB2 | RNA polymerase II subunit 2 |
| TUB2 | β-tubulin |
References
- Gostinčar, C. Towards Genomic Criteria for Delineating Fungal Species. Journal of fungi (Basel, Switzerland) 2020, 6. [Google Scholar] [CrossRef]
- Lalanne, C.; Silar, P. FungANI, a BLAST-based program for analyzing average nucleotide identity (ANI) between two fungal genomes, enables easy fungal species delimitation. Fungal Genet Biol 2025, 177, 103969. [Google Scholar] [CrossRef]
- Lin, P.; Kook, M.; Yi, T.H.; Yan, Z.F. Current Fungal Taxonomy and Developments in the Identification System. Curr Microbiol 2023, 80, 375. [Google Scholar] [CrossRef]
- De Filippo, E.; Gautier, V.; Lalanne, C.; Levert, E.; Chahine, E.; Hartmann, F.E.; Giraud, T.; Silar, P. Huge genetic diversity of Schizothecium tetrasporum (Wint.). N. Lundq.: delimitation of 18 species distributed into three complexes through genome sequencing. Mycosphere 2025, 16, 2936–2974. [Google Scholar] [CrossRef]
- Dettman, J.R.; Jacobson, D.J.; Taylor, J.W. A MULTILOCUS GENEALOGICAL APPROACH TO PHYLOGENETIC SPECIES RECOGNITION IN THE MODEL EUKARYOTE NEUROSPORA. Evolution 2003, 57, 2703–2720. [Google Scholar] [CrossRef] [PubMed]
- Dettman, J.R.; Jacobson, D.J.; Turner, E.; Pringle, A.; Taylor, J.W. REPRODUCTIVE ISOLATION AND PHYLOGENETIC DIVERGENCE IN NEUROSPORA: COMPARING METHODS OF SPECIES RECOGNITION IN A MODEL EUKARYOTE. Evolution 2003, 57, 2721–2741. [Google Scholar] [CrossRef] [PubMed]
- Dettman, J.R.; Jacobson, D.J.; Taylor, J.W. Multilocus sequence data reveal extensive phylogenetic species diversity within the Neurospora discreta complex. Mycologia 2006, 98, 436–446. [Google Scholar] [CrossRef]
- Svedberg, J.; Vogan, A.A.; Rhoades, N.A.; Sarmarajeewa, D.; Jacobson, D.J.; Lascoux, M.; Hammond, T.M.; Johannesson, H. An introgressed gene causes meiotic drive in Neurospora sitophila. Proceedings of the National Academy of Sciences of the United States of America 2021, 118. [Google Scholar] [CrossRef]
- Menkis, A.; Bastiaans, E.; Jacobson, D.J.; Johannesson, H. Phylogenetic and biological species diversity within the Neurospora tetrasperma complex. Journal of evolutionary biology 2009, 22, 1923–1936. [Google Scholar] [CrossRef]
- Boucher, C.; Nguyen, T.-S.; Silar, P. Species delimitation in the Podospora anserina/P. pauciseta/P. comata species complex (Sordariales). Crypt. Mycol. 2017, 38, 485–506. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Miller, A.N.; Cano-Lira, J.F.; Guarro, J.; García, D.; Stadler, M.; Huhndorf, S.M.; Stchigel, A.M. Re-Evaluation of the Order Sordariales: Delimitation of Lasiosphaeriaceae s. str., and Introduction of the New Families Diplogelasinosporaceae, Naviculisporaceae, and Schizotheciaceae. Microorganisms 2020, 8, 1430. [Google Scholar] [CrossRef]
- Harms, K.; Milic, A.; Stchigel, A.M.; Stadler, M.; Surup, F.; Marin-Felix, Y. Three New Derivatives of Zopfinol from Pseudorhypophila mangenotii gen. et comb. nov. Journal of Fungi 2021, 7, 181. [Google Scholar] [CrossRef]
- Cheng, K.W.; Yang, J.I.; Srimongkol, P.; Stadler, M.; Karnchanatat, A.; Ariyawansa, H.A. Fungal frontiers in toxic terrain: Revealing culturable fungal communities in Serpentine paddy fields of Taiwan. IMA Fungus 2025, 16, e155308. [Google Scholar] [CrossRef] [PubMed]
- Hensen, N.; Bonometti, L.; Westerberg, I.; Brännström, I.O.; Guillou, S.; Cros-Aarteil, S.; Calhoun, S.; Haridas, S.; Kuo, A.; Mondo, S.; et al. Genome-scale phylogeny and comparative genomics of the fungal order Sordariales. Molecular Phylogenetics and Evolution 2023, 189, 107938. [Google Scholar] [CrossRef] [PubMed]
- Egidi, E.; Delgado-Baquerizo, M.; Plett, J.M.; Wang, J.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A few Ascomycota taxa dominate soil fungal communities worldwide. Nature Communications 2019, 10, 2369. [Google Scholar] [CrossRef] [PubMed]
- Lundqvist, N. Nordic Sordariaceae S. Lat.; Acta Universitatis Upsaliensis: Uppsala, 1972; pp. 147–152. [Google Scholar]
- Thiyagaraja, V.; Hyde, K.; Piepenbring, M.; Davydov, E.; Dai, D.; Abdollahzadeh, J.; Bundhun, D.; Chethana, K.; Crous, P.; Gajanayake, A. Orders of Ascomycota. mycosphere 2025, 16. [Google Scholar] [CrossRef]
- Silar, P.; Gautier, V.; Lalanne, C.; Tangthirasunun, N.; Arthur, M.; Hartmann, F.E.; Giraud, T. The Podospora anserina species complex in metropolitan and overseas France with description of a new species, Podospora reunionensis sp. nov. Cryptogamie, Mycologie 2025, in the press.
- Silar, P. Podospora anserina. 2020. https://doi.org/.
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS computational biology 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Jin, J.-J.; Yu, W.-B.; Yang, J.-B.; Song, Y.; dePamphilis, C.W.; Yi, T.-S.; Li, D.-Z. GetOrganelle: a fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biology 2020, 21, 241. [Google Scholar] [CrossRef]
- Bushnell, B. BBMap: a fast, accurate, splice-aware aligner; Lawrence Berkeley National Laboratory, 2014.
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. Journal of computational biology : a journal of computational molecular cell biology 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nature biotechnology 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: gearing up for 1000 fungal genomes. Nucleic acids research 2014, 42, D699–704. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Briefings in Bioinformatics 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Molecular biology and evolution 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Crouan, P.L.; Crouan, H.M. Sordaria DNtrs. In Florule du Finistère; F. Klincksieck - J.B. & A. Lefournier: Paris & Brest, France, 1867.
- Abascal, F.; Zardoya, R.; Telford, M.J. TranslatorX: multiple alignment of nucleotide sequences guided by amino acid translations. Nucleic acids research 2010, 38, W7–13. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Molecular biology and evolution 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: a fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nature Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Cain, R.F. Studies of coprophilous Sphaeriales in Ontario. University of Toronto, Toronto, 1934.
- Mirza, J.H.; Cain, R.F. Revision of the genus Podospora. Canadian Journal of Botany 1969, 47, 1999–2048. [Google Scholar] [CrossRef]
- Doveri, F. Fungi Fimicoli Italici; Associazione Micologica Bresadola: Trento, Italy, 2004. [Google Scholar]
- Cai, L.; Jeewon, R.; Hyde, K.D. Molecular systematics of Zopfiella and allied genera: evidence from multi-gene sequence analyses. Mycological Research 2006, 110, 359–368. [Google Scholar] [CrossRef]
- Lundqvist, N. Tripterospora (Sordariaceae s. lat., Pyrenomycetes). Botaniska Notiser 1969.
- Huang, S.-K.; Hyde, K.D.; Mapook, A.; Maharachchikumbura, S.S.N.; Bhat, J.D.; McKenzie, E.H.C.; Jeewon, R.; Wen, T.-C. Taxonomic studies of some often over-looked Diaporthomycetidae and Sordariomycetidae. Fungal Diversity 2021. [Google Scholar] [CrossRef]
- Arx, v.J.A. On Thielavia and some similar genera of ascomycetes. Studies in Mycology 1975, 8, 1–29 + 22pl. [Google Scholar]
- Von Arx, J.; Hennebert, G. Triangularia mangenotii nov. sp. Bull. Soc. Mycol. France 1968, 84, 423–426. [Google Scholar]
- Furuya, K.; Udagawa, S. Two new species of cleistothecial Ascomycetes. Journal of Japanese botany 1975. [Google Scholar]
- Pöggeler, S.; Risch, S.; Kück, U.; Osiewacz, H.D. Mating-type genes from the homothallic fungus Sordaria macrospora are functionally expressed in a heterothallic ascomycete. Genetics 1997, 147, 567–580. [Google Scholar] [CrossRef]
- Bennett Richard, J.; Turgeon, B.G. Fungal Sex: The Ascomycota. Microbiology Spectrum 2016, 4, 10.1128/microbiolspec.funk-0005-2016. [Google Scholar] [CrossRef]
- Debuchy, R.; Berteaux-Lecellier, V.; Silar, P. Mating Systems and Sexual Morphogenesis in Ascomycetes. In Cellular and Molecular Biology of Filamentous Fungi; Borkovich, K., Ebbole, D., Eds.; ASM Press: Washington, DC, 2010; pp. 501–535. [Google Scholar]
- Cailleux, R. Champignons stercoraux de République Centrafricaine, III Podospora nouveaux. Cahier de la Maboké 1969, 7, 87–102. [Google Scholar]
- Berka, R.M.; Grigoriev, I.V.; Otillar, R.; Salamov, A.; Grimwood, J.; Reid, I.; Ishmael, N.; John, T.; Darmond, C.; Moisan, M.C.; et al. Comparative genomic analysis of the thermophilic biomass-degrading fungi Myceliophthora thermophila and Thielavia terrestris. Nat Biotechnol 2011, 29, 922–927. [Google Scholar] [CrossRef]
- Wang, X.W.; Han, P.J.; Bai, F.Y.; Luo, A.; Bensch, K.; Meijer, M.; Kraak, B.; Han, D.Y.; Sun, B.D.; Crous, P.W.; et al. Taxonomy, phylogeny and identification of Chaetomiaceae with emphasis on thermophilic species. Studies in Mycology 2022, 101, 121–243. [Google Scholar] [CrossRef]
- Steindorff, A.S.; Aguilar-Pontes, M.V.; Robinson, A.J.; Andreopoulos, B.; LaButti, K.; Kuo, A.; Mondo, S.; Riley, R.; Otillar, R.; Haridas, S.; et al. Comparative genomic analysis of thermophilic fungi reveals convergent evolutionary adaptations and gene losses. Communications biology 2024, 7, 1124. [Google Scholar] [CrossRef]
- Aime, M.C.; Miller, A.N.; Aoki, T.; Bensch, K.; Cai, L.; Crous, P.W.; Hawksworth, D.L.; Hyde, K.D.; Kirk, P.M.; Lücking, R.; et al. How to publish a new fungal species, or name, version 3.0. IMA Fungus 2021, 12, 11. [Google Scholar] [CrossRef]
- Hutchinson, M.I.; Powell, A.J.; Tsang, A.; O’Toole, N.; Berka, R.M.; Barry, K.; Grigoriev, I.V.; Natvig, D.O. Genetics of mating in members of the Chaetomiaceae as revealed by experimental and genomic characterization of reproduction in Myceliophthora heterothallica. Fungal Genetics and Biology 2016, 86, 9–19. [Google Scholar] [CrossRef]
Figure 1.
ITS + LSU Phylogenetic tree of the Naviculisporaceae. The tree was rooted with Apodospora. peruviana CBS 118394 The strains whose genome sequences were obtained in the present study are shown in red, and the strains whose genome sequences were previously published in blue [14]. On the left panel, Ts indicate ex-type strains. Insets show cladograms of Rhypophila and Pseudorhypophila displaying bootstrap values.
Figure 1.
ITS + LSU Phylogenetic tree of the Naviculisporaceae. The tree was rooted with Apodospora. peruviana CBS 118394 The strains whose genome sequences were obtained in the present study are shown in red, and the strains whose genome sequences were previously published in blue [14]. On the left panel, Ts indicate ex-type strains. Insets show cladograms of Rhypophila and Pseudorhypophila displaying bootstrap values.
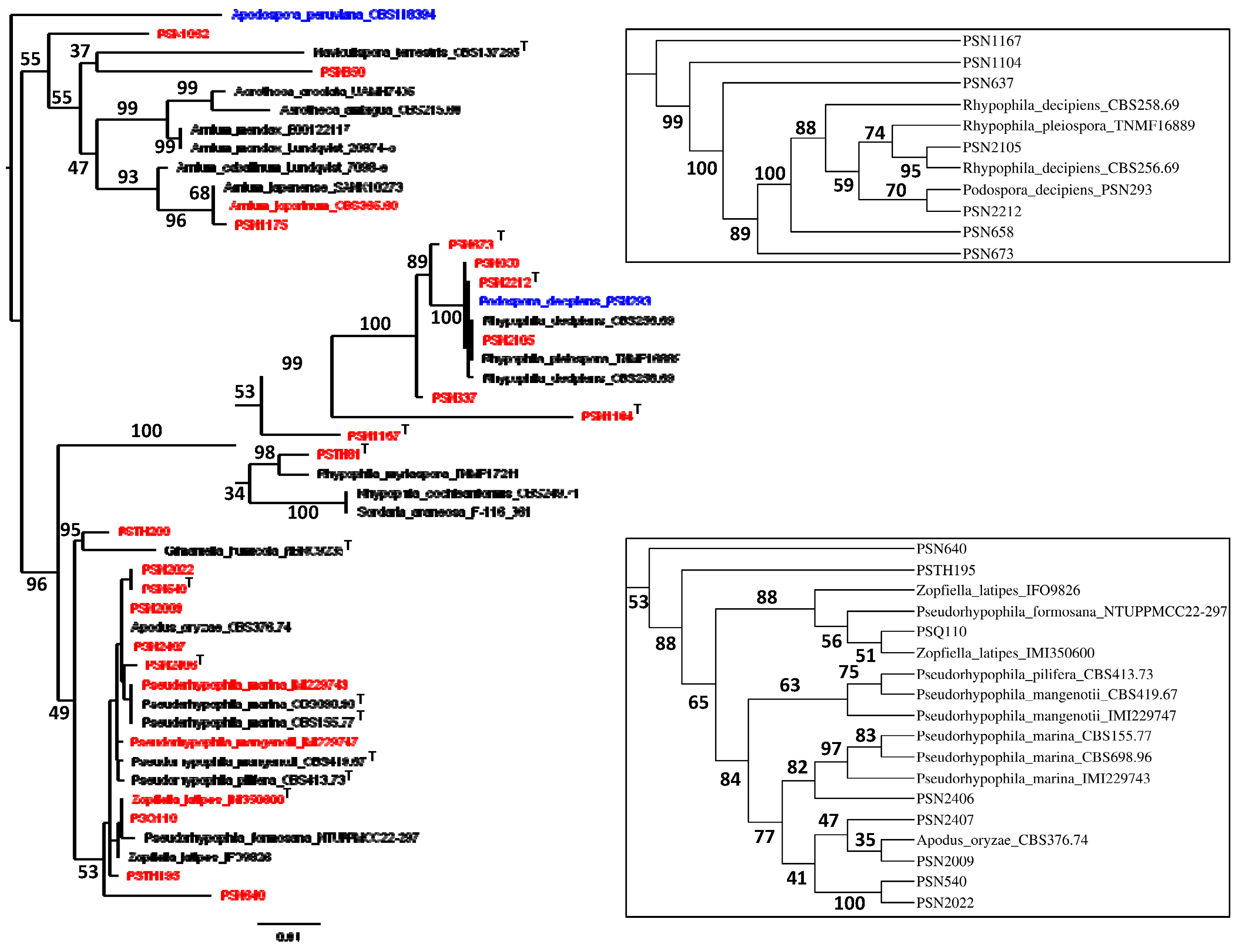
Figure 2.
Morphology of Podospora aff. araneosa PSN1062. Cultures on M2 (A) and M0 + hay (B); arrowheads point towards perithecia. Both plates were inoculated at the center and incubated for one month at 27°C and 24h illumination. Growth was very slow on M2 and much faster on M0 + hay. Perithecia differentiated only on M0 + hay after 2-4 weeks at room temperature (C). Their diameter measured 935 + 86 µm (n=10). Their peridium was light-brown (D) and densely-covered with long flexuous hair (C). They contained rosettes of clavate asci bearing around 256 ascospores (E). Young asci (F) were at first filled with ascospores only in the central part. Ascospores were bicellular (G) and multiseriate (E-F). Spore heads were 19.8 + 1.2 µm × 13.9 + 0.5 µm (n=30), olivaceous when young and dark brown when mature (G). The primary appendages were 9.5 + 1.6 µm × 4.4 + 0.9 µm (n=10), cylindrical and hyaline (G). Two thin and long secondary appendages of variable length were present, one at the apex of the ascospore and one at the end of the primary appendage (G). After ejection, the primary and secondary appendages were often lost. Except for the slightly larger sizes of the perithecia and slightly smaller size of the ascospores, this description fits well with that of P. araneosa (Cain) Cain. Scale bars: C = 1 mm, D and F = 100 µm, E = 200 µm and G = 10 µm.
Figure 2.
Morphology of Podospora aff. araneosa PSN1062. Cultures on M2 (A) and M0 + hay (B); arrowheads point towards perithecia. Both plates were inoculated at the center and incubated for one month at 27°C and 24h illumination. Growth was very slow on M2 and much faster on M0 + hay. Perithecia differentiated only on M0 + hay after 2-4 weeks at room temperature (C). Their diameter measured 935 + 86 µm (n=10). Their peridium was light-brown (D) and densely-covered with long flexuous hair (C). They contained rosettes of clavate asci bearing around 256 ascospores (E). Young asci (F) were at first filled with ascospores only in the central part. Ascospores were bicellular (G) and multiseriate (E-F). Spore heads were 19.8 + 1.2 µm × 13.9 + 0.5 µm (n=30), olivaceous when young and dark brown when mature (G). The primary appendages were 9.5 + 1.6 µm × 4.4 + 0.9 µm (n=10), cylindrical and hyaline (G). Two thin and long secondary appendages of variable length were present, one at the apex of the ascospore and one at the end of the primary appendage (G). After ejection, the primary and secondary appendages were often lost. Except for the slightly larger sizes of the perithecia and slightly smaller size of the ascospores, this description fits well with that of P. araneosa (Cain) Cain. Scale bars: C = 1 mm, D and F = 100 µm, E = 200 µm and G = 10 µm.
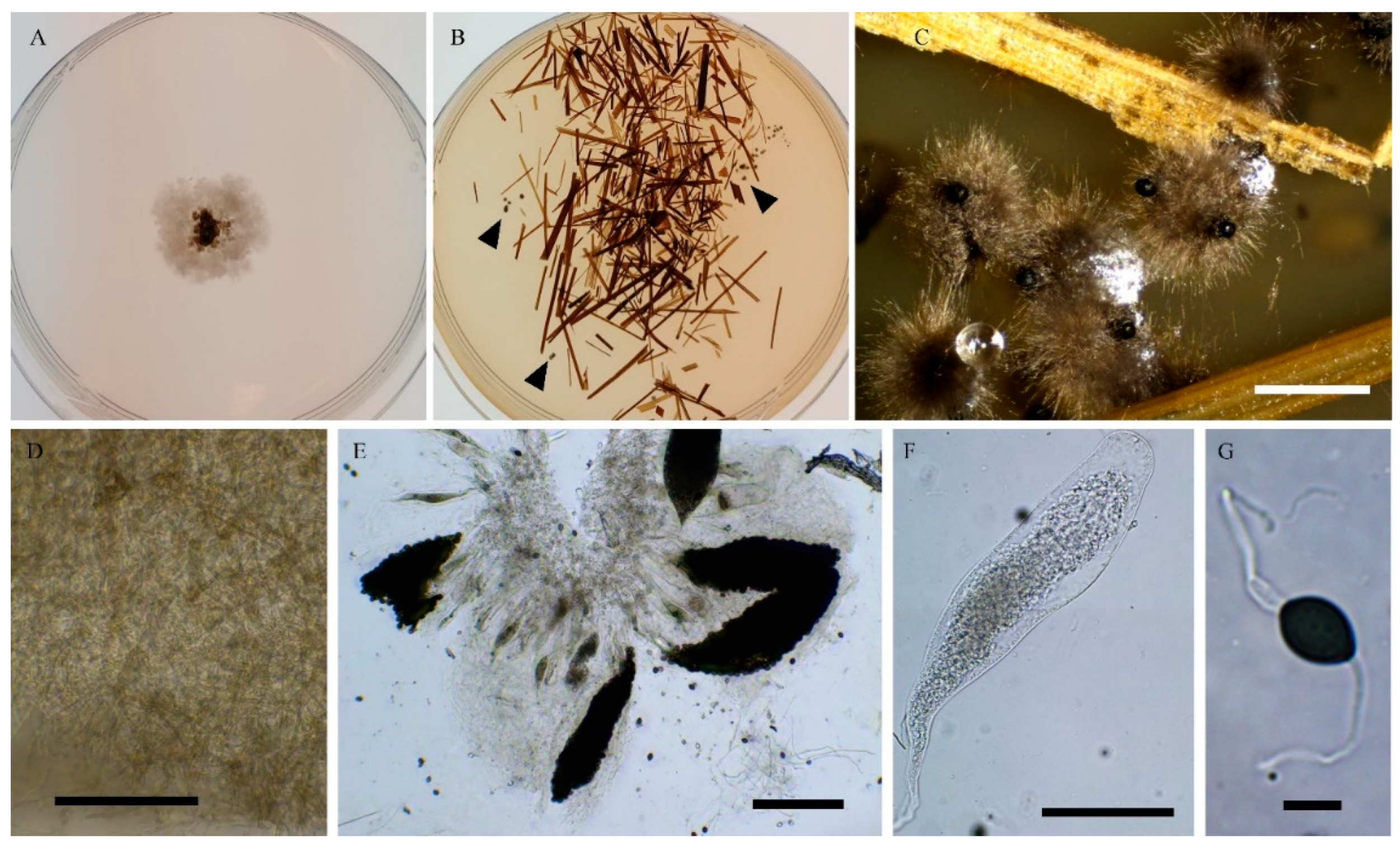
Figure 3.
Average Nucleotide Identity (ANI) analysis of PSN1175 and Arnium leporinum CBS 365.69. Horizontal bars quantify the number of BLAST results for each range of similarity percentages defined and color-coded at the top of the plot. The calculations were made in both directions, i.e., with on the left PSN1175 as query (test) and CBS 365.69 as subject (reference) and reciprocally on the right. The final ANI is the average of the two values. The numbers of BLAST obtained for each percentage range are boxed in the corresponding bars and the total number of BLAST made during the analysis is given at the top of the figure. The zero hits correspond to BLASTs where no similarity was found. For more information see ref. [2]. PSN1175 and CBS 365.69 displayed very few sequences with a similarity higher than 99.5%, corresponding to the pink and magenta bars at the top of the graphic. Moreover, each strain had about 8% to 9% of specific sequences.
Figure 3.
Average Nucleotide Identity (ANI) analysis of PSN1175 and Arnium leporinum CBS 365.69. Horizontal bars quantify the number of BLAST results for each range of similarity percentages defined and color-coded at the top of the plot. The calculations were made in both directions, i.e., with on the left PSN1175 as query (test) and CBS 365.69 as subject (reference) and reciprocally on the right. The final ANI is the average of the two values. The numbers of BLAST obtained for each percentage range are boxed in the corresponding bars and the total number of BLAST made during the analysis is given at the top of the figure. The zero hits correspond to BLASTs where no similarity was found. For more information see ref. [2]. PSN1175 and CBS 365.69 displayed very few sequences with a similarity higher than 99.5%, corresponding to the pink and magenta bars at the top of the graphic. Moreover, each strain had about 8% to 9% of specific sequences.

Figure 4.
Morphology of Rhypophila myriaspora PSN2105. Mycelium on M2 medium after two weeks of growth (A). Perithecia were readily differentiated on M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination; they were 542 ± 68 µm diam (n=10), pyriform and covered with numerous hairs (B); their neck was black and slightly elongated (B-D). Their peridium was pale brown and semi-translucid (C). Asci were 64-spored, clavate with a tapering and narrow apex (E-F). Ascospores were multiseriate and bicellular (F-I). Ascospore heads were 27.3 ± 2.8 × 17.1 ± 1.2 µm (n=30), ellipsoidal with a slightly flattened base, hyaline at first (E, G), then green and black when mature (F, H-I). The primary appendage was long (often longer than the spore head) and flat (G), frequently collapsing (G-I), 38.6 ±.5.2 × 6.3 ± 0.9 µm (n=10). Secondary appendages were present at the apex of the spore head as a fan-shaped structure of four fibrillar strands and at the junction between the spore head (I). Primary and secondary appendages were often lost upon ascospore ejection. This description fits with the ones made by Lundqvist for R. myriaspora (perithecium= 670-1150 × 400-530 µm, spore head= 23-34 × 14-19 µm, primary appendage = 18-45 × 14-19 µm; [16]) and Doveri (perithecium= 625-750 × 400- 500 µm, spore head= 27.3-34.6 × 17.8-19 µm, primary appendage = 30-37 × 5-7 µm; [35]). Scale bars: B-D = 200 µm, E-H = 20 µm, I = 10 µm.
Figure 4.
Morphology of Rhypophila myriaspora PSN2105. Mycelium on M2 medium after two weeks of growth (A). Perithecia were readily differentiated on M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination; they were 542 ± 68 µm diam (n=10), pyriform and covered with numerous hairs (B); their neck was black and slightly elongated (B-D). Their peridium was pale brown and semi-translucid (C). Asci were 64-spored, clavate with a tapering and narrow apex (E-F). Ascospores were multiseriate and bicellular (F-I). Ascospore heads were 27.3 ± 2.8 × 17.1 ± 1.2 µm (n=30), ellipsoidal with a slightly flattened base, hyaline at first (E, G), then green and black when mature (F, H-I). The primary appendage was long (often longer than the spore head) and flat (G), frequently collapsing (G-I), 38.6 ±.5.2 × 6.3 ± 0.9 µm (n=10). Secondary appendages were present at the apex of the spore head as a fan-shaped structure of four fibrillar strands and at the junction between the spore head (I). Primary and secondary appendages were often lost upon ascospore ejection. This description fits with the ones made by Lundqvist for R. myriaspora (perithecium= 670-1150 × 400-530 µm, spore head= 23-34 × 14-19 µm, primary appendage = 18-45 × 14-19 µm; [16]) and Doveri (perithecium= 625-750 × 400- 500 µm, spore head= 27.3-34.6 × 17.8-19 µm, primary appendage = 30-37 × 5-7 µm; [35]). Scale bars: B-D = 200 µm, E-H = 20 µm, I = 10 µm.

Figure 5.
Morphology of Rhypophila decipiens PSN293. Mycelium on M2 medium after two weeks of growth (A). Perithecia were readily differentiated on M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination; they had a diameter of 439 + 71 µm and were pyriform and covered with numerous hairs (B); their neck was black and slightly elongated (B-C). Their peridium was pale brown and semi-translucid; their textura was angularis (D-E). Asci were eight-spored, clavate with a tapering and narrow apex (F-G). Ascospores were biseriate and bicellular (G-L). Ascospore heads were 35.5 ± 2.8 × 18.6 ± 1.2 µm (n=30), ellipsoidal with a slightly flattened base, hyaline at first (F-G), then green (G) and black when mature (H-L). The primary appendage was very long (often longer than the spore head) and flat (J), frequently collapsing (L), 44.3 ± .7.3 × 6.5 ± 1.5 µm (n=10), Secondary appendages were present at the apex of the spore head as a fan-shaped structure of juxtaposed fibrils and at the junction between the spore head and the primary appendage as fibrils often two in number (L). This description fits with the ones made for R. decipiens by Mirza and Cain (perithecium= 600-800 × 350-450 µm, spore head= 35-46 × 19-23 µm, primary appendage = 40-60 × 7-8 µm ; [34]), Lundqvist (perithecium= 480-1050 × 290-530 µm, spore head= 36-42 × 20-22 µm, primary appendage = 55-73 × 7.2-8.5 µm; [16]) and Doveri (perithecium= 750-1050 × 620- 740 µm, spore head= 34.6-42 × 19-23 µm, primary appendage = 33-42 × 5.5-8 µm; [35]). Scale bars: B = 200 µm, C = 200 µm, D-K = 20 µm, L = 10 µm.
Figure 5.
Morphology of Rhypophila decipiens PSN293. Mycelium on M2 medium after two weeks of growth (A). Perithecia were readily differentiated on M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination; they had a diameter of 439 + 71 µm and were pyriform and covered with numerous hairs (B); their neck was black and slightly elongated (B-C). Their peridium was pale brown and semi-translucid; their textura was angularis (D-E). Asci were eight-spored, clavate with a tapering and narrow apex (F-G). Ascospores were biseriate and bicellular (G-L). Ascospore heads were 35.5 ± 2.8 × 18.6 ± 1.2 µm (n=30), ellipsoidal with a slightly flattened base, hyaline at first (F-G), then green (G) and black when mature (H-L). The primary appendage was very long (often longer than the spore head) and flat (J), frequently collapsing (L), 44.3 ± .7.3 × 6.5 ± 1.5 µm (n=10), Secondary appendages were present at the apex of the spore head as a fan-shaped structure of juxtaposed fibrils and at the junction between the spore head and the primary appendage as fibrils often two in number (L). This description fits with the ones made for R. decipiens by Mirza and Cain (perithecium= 600-800 × 350-450 µm, spore head= 35-46 × 19-23 µm, primary appendage = 40-60 × 7-8 µm ; [34]), Lundqvist (perithecium= 480-1050 × 290-530 µm, spore head= 36-42 × 20-22 µm, primary appendage = 55-73 × 7.2-8.5 µm; [16]) and Doveri (perithecium= 750-1050 × 620- 740 µm, spore head= 34.6-42 × 19-23 µm, primary appendage = 33-42 × 5.5-8 µm; [35]). Scale bars: B = 200 µm, C = 200 µm, D-K = 20 µm, L = 10 µm.
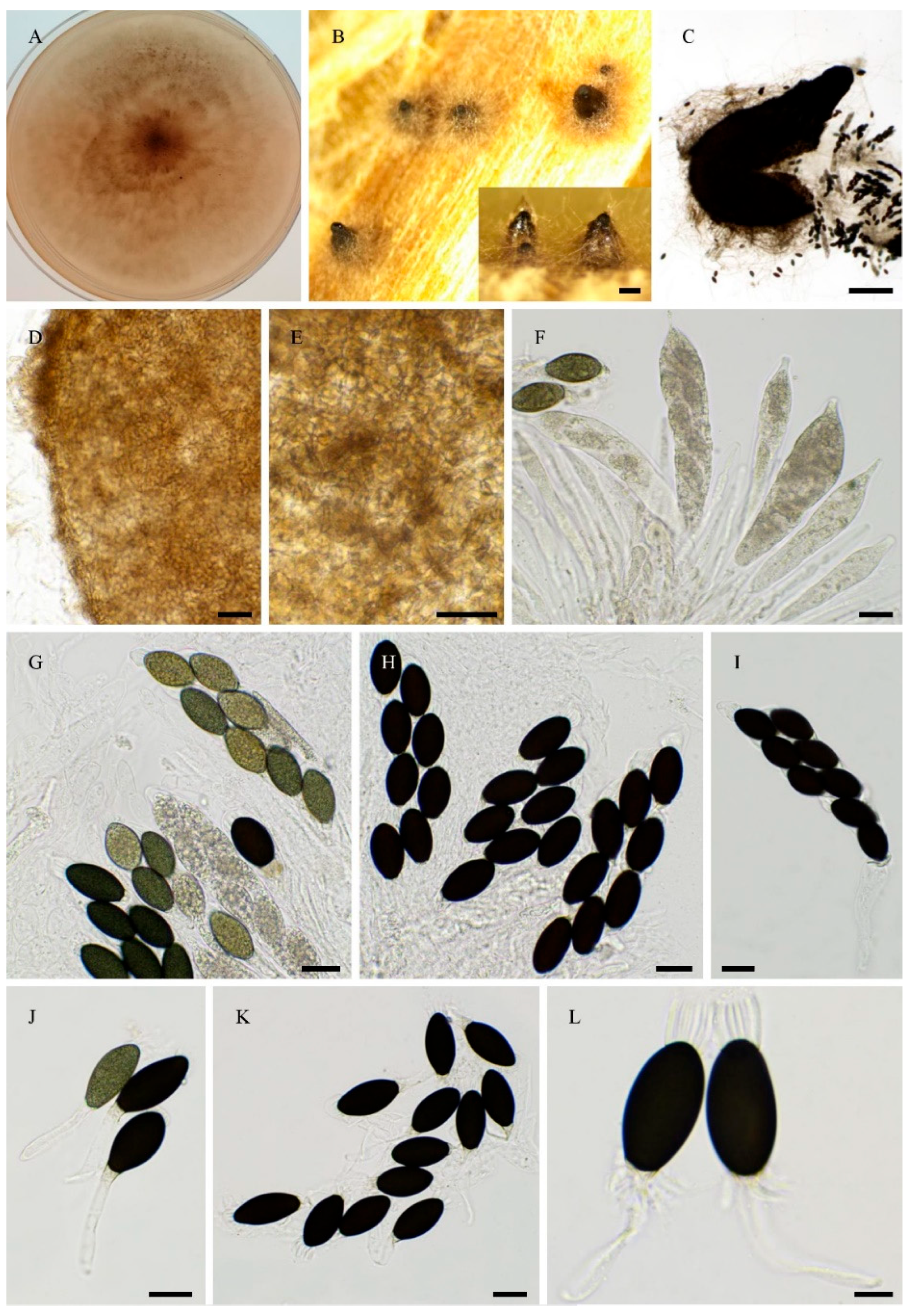
Figure 6.
Morphology of Rhypophila pleiospora PSN658. Mycelium on M2 medium after two weeks of growth (A). Perithecia were readily differentiated on M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination; they were 579 ± 71 µm diam (n=10), pyriform and covered with numerous hairs (B); their neck was black and short (B-C). Their peridium was pale brown and semi-translucid, their textura was angularis (D). Asci were sixteen-spored, clavate with a tapering and narrow apex (E). Ascospores were biseriate or triseriate and bicellular (E-I). Ascospore heads were 29.8 ± 2.3 × 17.0 ± 1.2 µm (n=30), ellipsoidal with a slightly flattened base, hyaline at first (E-F), then green (E) and black when mature (G-I). The primary appendage was long (often longer than the spore head) and flat (F), frequently collapsing (G-I), 38.7 ±.9.6 × 7.1 ± 1.5 µm (n=10). Secondary appendages were present at the apex of the spore head as a fan-shaped structure of juxtaposed fibrils and at the junction between the spore head and the primary appendage as fibrils often two in number (L). This description fits with the ones for R. pleiospora made by Mirza and Cain (perithecium= 600-1000 × 375- 550 µm, spore head= 31-36 × 19-24 µm for 16-spored asci, primary appendage = 25-50 × 5-8 µm; [34]), Lundqvist (perithecium= 530-1100 × 335-550 µm, spore head= 30-37 × 18-23 µm, primary appendage = 35-65 µm; [16]) and Doveri (perithecium= 1000-1200 × 500-650 µm, spore head= 31.5-36-7 × 17.8-21 µm, primary appendage = 22-45 × 5-7 µm; [35]). Scale bars: B-C = 200 µm, D-G = 20 µm, H-I = 10 µm.
Figure 6.
Morphology of Rhypophila pleiospora PSN658. Mycelium on M2 medium after two weeks of growth (A). Perithecia were readily differentiated on M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination; they were 579 ± 71 µm diam (n=10), pyriform and covered with numerous hairs (B); their neck was black and short (B-C). Their peridium was pale brown and semi-translucid, their textura was angularis (D). Asci were sixteen-spored, clavate with a tapering and narrow apex (E). Ascospores were biseriate or triseriate and bicellular (E-I). Ascospore heads were 29.8 ± 2.3 × 17.0 ± 1.2 µm (n=30), ellipsoidal with a slightly flattened base, hyaline at first (E-F), then green (E) and black when mature (G-I). The primary appendage was long (often longer than the spore head) and flat (F), frequently collapsing (G-I), 38.7 ±.9.6 × 7.1 ± 1.5 µm (n=10). Secondary appendages were present at the apex of the spore head as a fan-shaped structure of juxtaposed fibrils and at the junction between the spore head and the primary appendage as fibrils often two in number (L). This description fits with the ones for R. pleiospora made by Mirza and Cain (perithecium= 600-1000 × 375- 550 µm, spore head= 31-36 × 19-24 µm for 16-spored asci, primary appendage = 25-50 × 5-8 µm; [34]), Lundqvist (perithecium= 530-1100 × 335-550 µm, spore head= 30-37 × 18-23 µm, primary appendage = 35-65 µm; [16]) and Doveri (perithecium= 1000-1200 × 500-650 µm, spore head= 31.5-36-7 × 17.8-21 µm, primary appendage = 22-45 × 5-7 µm; [35]). Scale bars: B-C = 200 µm, D-G = 20 µm, H-I = 10 µm.

Figure 7.
ANI analysis of PSN540 and PSN2022. See legend of Figure 3 for a detailed explanation of the graphic. The analysis revealed that the two strains shared mostly highly similar sequences (e.g., with a similarity higher than 99.5%; pink and magenta bars). They have very few specific sequences (around 1%). The ANI is lowered to less than 99% due to the presence of about 4% of very dissimilar sequences with a similarity of around 85 and 90 % (bars around the yellow-orange ones at the bottom of the graphic).
Figure 7.
ANI analysis of PSN540 and PSN2022. See legend of Figure 3 for a detailed explanation of the graphic. The analysis revealed that the two strains shared mostly highly similar sequences (e.g., with a similarity higher than 99.5%; pink and magenta bars). They have very few specific sequences (around 1%). The ANI is lowered to less than 99% due to the presence of about 4% of very dissimilar sequences with a similarity of around 85 and 90 % (bars around the yellow-orange ones at the bottom of the graphic).
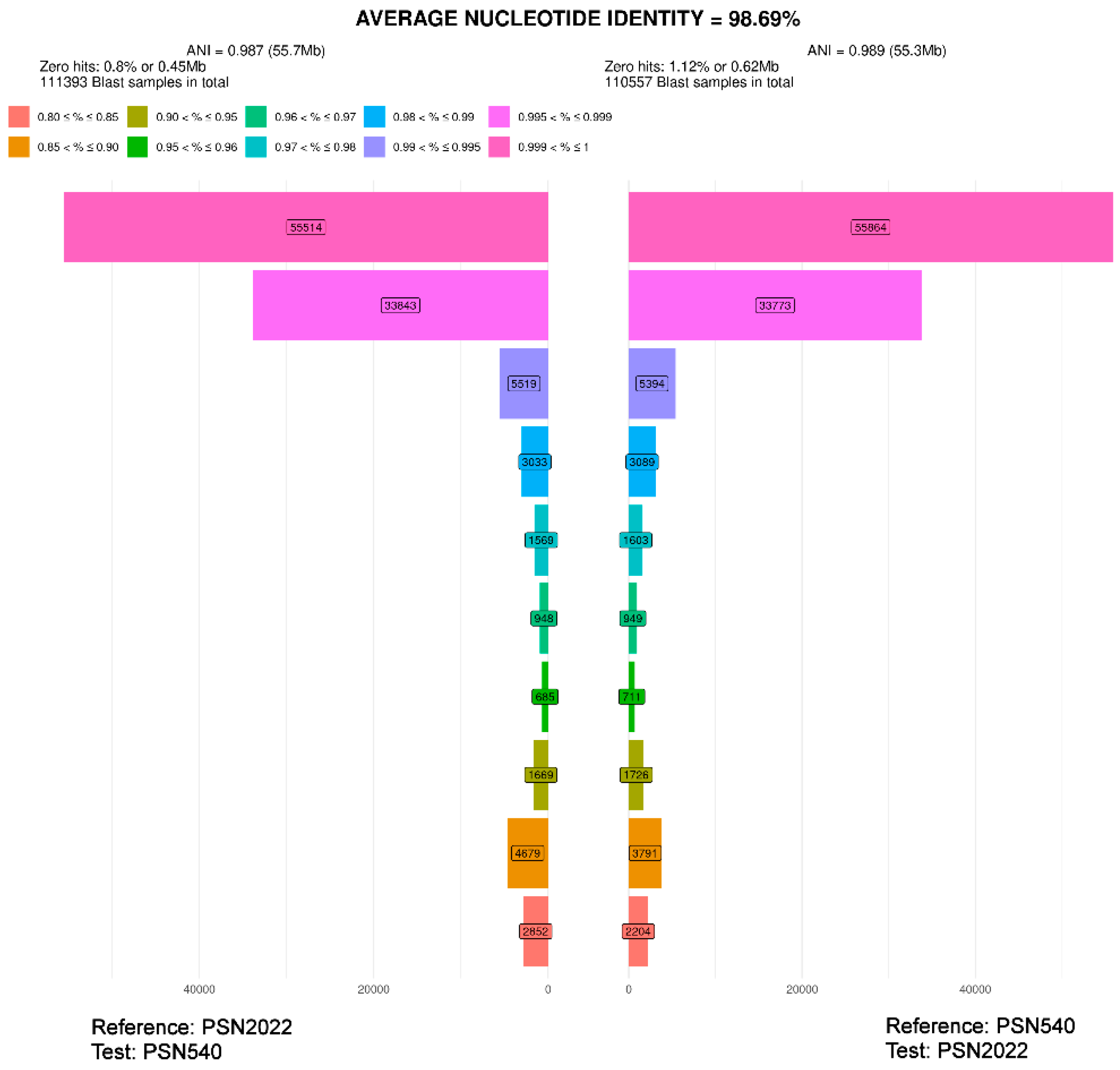
Figure 8.
Morphology of Pseudorhypophila latipes comb. nov. IMI350600. Mycelium on M2 medium after two weeks of growth (A). Perithecia, non-ostiolate, were readily differentiated on M2 and M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination, superficial or immersed, black, globose to subglobose, they were 449 ± 89 µm diam (n=10) (B-C); their peridium was pale brown and semi-translucid, tissue type was textura angularis (D). Asci were 8-spored, clavate with a tapering and narrow apex (F-G). Ascospores were biseriate and bicellular (G). Ascospore heads were 18.0 ± 0.8 × 10.6 ± 0.5 µm (n=30), ovoid with a slightly flattened base, hyaline at first (I), then green (J) and brown to dark brown when mature (H-I, K). The primary appendage with a rounded head and hyaline (J), frequently collapsing at maturity (K), they were 4.5 ± 0.6 × 6.4 ± 0.6 µm (n=10). This description fits the original description of Lundqvist (as Tripterospora latipes: perithecium= 290-700 µm diam, spore head= 15.5-20.5 × 10-13 µm, primary appendage= 6.3-7.7 × 5.5-7 µm; [37]). Scale bars: B = 200 µm, C = 100 µm, D-E = 20 µm, F-K = 10 µm.
Figure 8.
Morphology of Pseudorhypophila latipes comb. nov. IMI350600. Mycelium on M2 medium after two weeks of growth (A). Perithecia, non-ostiolate, were readily differentiated on M2 and M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination, superficial or immersed, black, globose to subglobose, they were 449 ± 89 µm diam (n=10) (B-C); their peridium was pale brown and semi-translucid, tissue type was textura angularis (D). Asci were 8-spored, clavate with a tapering and narrow apex (F-G). Ascospores were biseriate and bicellular (G). Ascospore heads were 18.0 ± 0.8 × 10.6 ± 0.5 µm (n=30), ovoid with a slightly flattened base, hyaline at first (I), then green (J) and brown to dark brown when mature (H-I, K). The primary appendage with a rounded head and hyaline (J), frequently collapsing at maturity (K), they were 4.5 ± 0.6 × 6.4 ± 0.6 µm (n=10). This description fits the original description of Lundqvist (as Tripterospora latipes: perithecium= 290-700 µm diam, spore head= 15.5-20.5 × 10-13 µm, primary appendage= 6.3-7.7 × 5.5-7 µm; [37]). Scale bars: B = 200 µm, C = 100 µm, D-E = 20 µm, F-K = 10 µm.
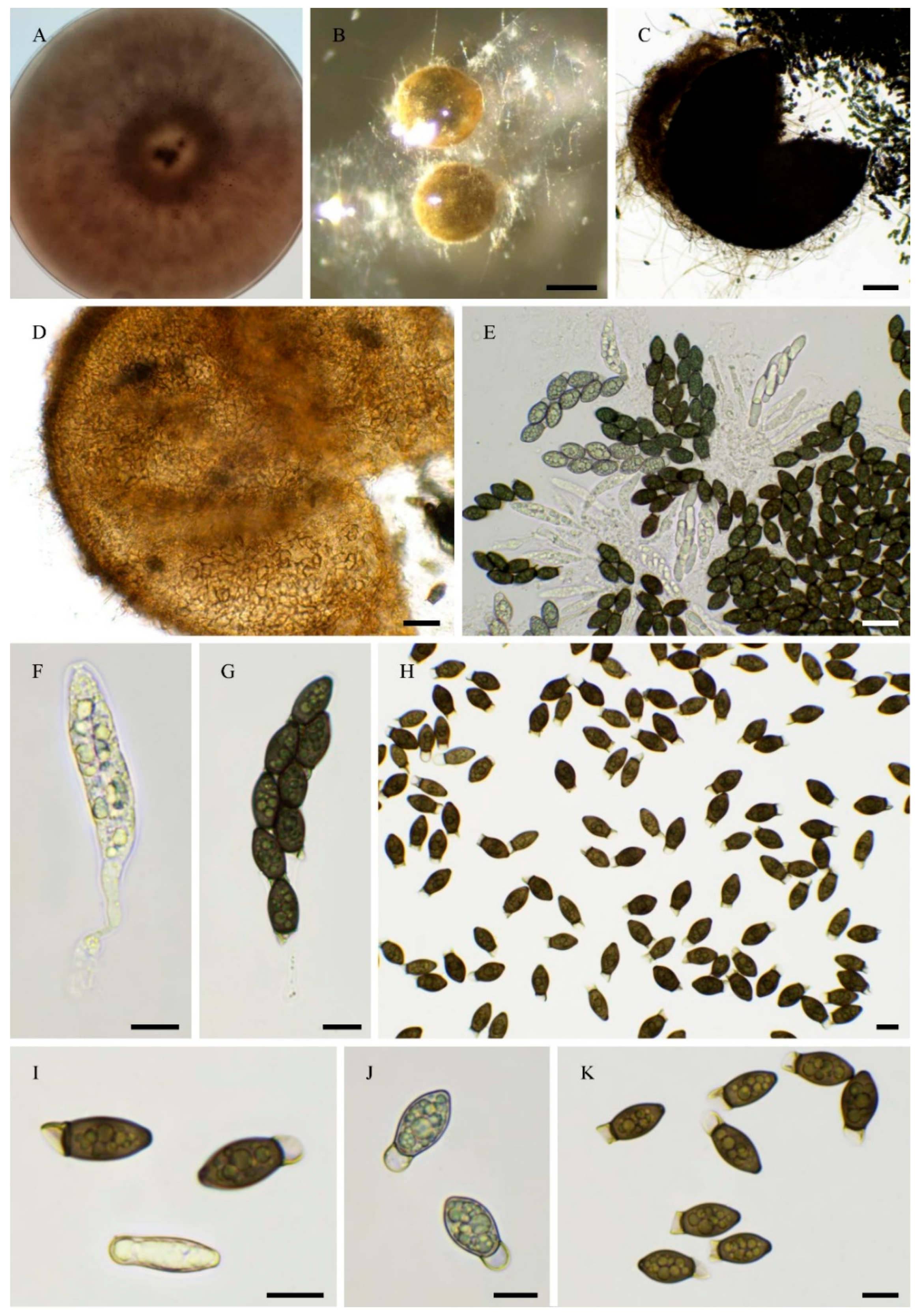
Figure 9.
Morphology of Pseudorhypophila oryzae PSN2009. Mycelium on M2 medium after two weeks of growth (A-B), Non ostiolate perithecia were readily differentiated on M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination; pyriform, superficial or semi-immersed, 415 ± 107 µm diam (n=10); (C-D), Their peridium was pale brown and semi-translucid, tissue type was textura angularis. (E-F). Paraphyses absent. Asci were 8-spored, clavate with a tapering and narrow apex (G). Ascospores were uniseriate in young asci and were disordered in older ones (G-H). Ascospore heads were 21.7 ± 2.2 × 13.2 ± 1.1 μm (n=30), ellipsoidal to oval, hyaline at first, then green and brown to dark brown when mature (I-K). Few ascospores (less than 10%) were 1-septate (septum slightly curved or straight toward the apex), one cell 12.4 ± 1.9 µm and the other 8.5 ± 1.8 μm (n=15) (I-K). This description fits with that of A. oryzae Carolis & Arx (non-ostiolate perithecium= 200-500 µm; ascospore= 21-27 × 10-14 µm, more than 90% of the ascospores remaining single-celled; [39]). Scale bars: C = 200 µm, D = 100 µm, E-H= 20 µm, I-K = 10 µm.
Figure 9.
Morphology of Pseudorhypophila oryzae PSN2009. Mycelium on M2 medium after two weeks of growth (A-B), Non ostiolate perithecia were readily differentiated on M0 + miscanthus after 2-3 weeks incubation at 27°C with constant illumination; pyriform, superficial or semi-immersed, 415 ± 107 µm diam (n=10); (C-D), Their peridium was pale brown and semi-translucid, tissue type was textura angularis. (E-F). Paraphyses absent. Asci were 8-spored, clavate with a tapering and narrow apex (G). Ascospores were uniseriate in young asci and were disordered in older ones (G-H). Ascospore heads were 21.7 ± 2.2 × 13.2 ± 1.1 μm (n=30), ellipsoidal to oval, hyaline at first, then green and brown to dark brown when mature (I-K). Few ascospores (less than 10%) were 1-septate (septum slightly curved or straight toward the apex), one cell 12.4 ± 1.9 µm and the other 8.5 ± 1.8 μm (n=15) (I-K). This description fits with that of A. oryzae Carolis & Arx (non-ostiolate perithecium= 200-500 µm; ascospore= 21-27 × 10-14 µm, more than 90% of the ascospores remaining single-celled; [39]). Scale bars: C = 200 µm, D = 100 µm, E-H= 20 µm, I-K = 10 µm.
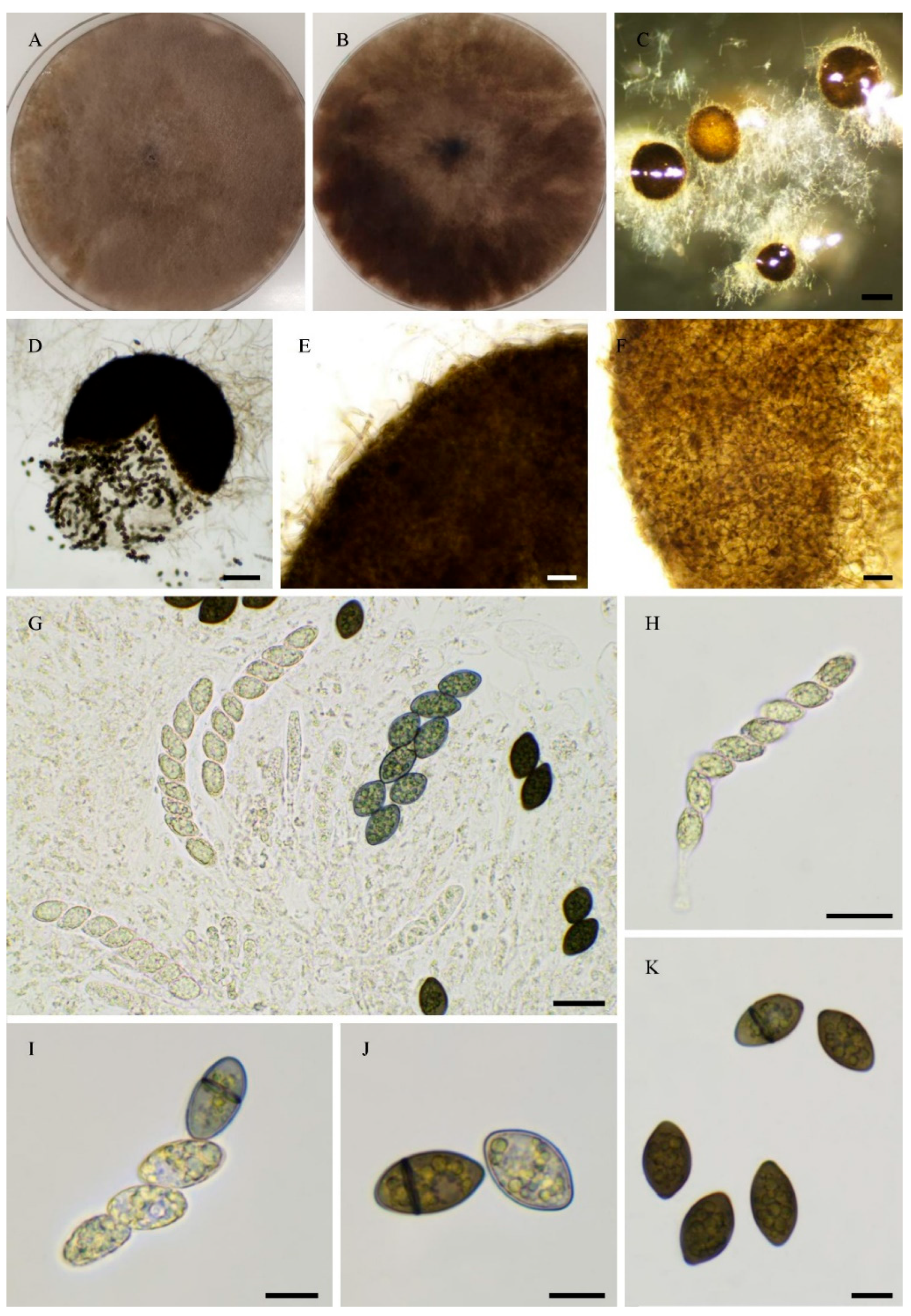
Figure 10.
Morphology of Pseudorhypophila mangenotii IMI229747 Mycelium on OA medium after two weeks of growth (A). Perithecia were readily differentiated on OA after 2-3 weeks incubation at 27°C with constant illumination, immersed or semi-immersed, black, they were 266 ± 47 µm diam (n=10) (B-C), with a clearly defined and narrow neck (B, F). Their peridium was pale brown and semi-translucid, tissue type was textura angularis (D-E). Asci were 8-spored, clavate with a tapering and narrow apex (G-H). Ascospores were biseriate and bicellular (G-I). Ascospore heads were 15.8 ± 0.9 × 8.0 ± 0.4 µm (height x base size; n=30), oval with a slightly flattened base, hyaline at first (G, I-J), then green (I) and black when mature (I-J). The primary appendage shape had a rounded end and was hyaline(I-J), they were 5.7 ± 0.5 × 7.9 ± 0.4 µm (n=10). This description fits the original one made by von Arx and Hennebert (as Triangularia mangenotii: perithecium = 170–260 µm diam., ascospore size= 21-24 × 7-9 µm, primary appendage= 7-9 µm diam.; [40]). Scale bars: B-C= 200 µm, D-I = 20 µm, J = 10 µm.
Figure 10.
Morphology of Pseudorhypophila mangenotii IMI229747 Mycelium on OA medium after two weeks of growth (A). Perithecia were readily differentiated on OA after 2-3 weeks incubation at 27°C with constant illumination, immersed or semi-immersed, black, they were 266 ± 47 µm diam (n=10) (B-C), with a clearly defined and narrow neck (B, F). Their peridium was pale brown and semi-translucid, tissue type was textura angularis (D-E). Asci were 8-spored, clavate with a tapering and narrow apex (G-H). Ascospores were biseriate and bicellular (G-I). Ascospore heads were 15.8 ± 0.9 × 8.0 ± 0.4 µm (height x base size; n=30), oval with a slightly flattened base, hyaline at first (G, I-J), then green (I) and black when mature (I-J). The primary appendage shape had a rounded end and was hyaline(I-J), they were 5.7 ± 0.5 × 7.9 ± 0.4 µm (n=10). This description fits the original one made by von Arx and Hennebert (as Triangularia mangenotii: perithecium = 170–260 µm diam., ascospore size= 21-24 × 7-9 µm, primary appendage= 7-9 µm diam.; [40]). Scale bars: B-C= 200 µm, D-I = 20 µm, J = 10 µm.

Figure 11.
Morphology of Pseudorhypophila marina IMI229743. Mycelium on medium after two weeks of growth (A-B). Perithecia, non-ostiolate, were readily differentiated on M2, M0 + miscanthus, OA, V8 and M0 + hay after 2-3 weeks incubation at 27°C with constant illumination, superficial or immersed, black, globose to subglobose, they were 287 ± 20 µm diam (n=10) (C-D). Their peridium was pale brown and semi-translucid, tissue type was textura angularis (E-F). Asci were 8-spored, clavate with a tapering and narrow apex (G-H). Ascospores were biseriate and bicellular (G-I). Ascospore heads were 17.7 ± 0.9 × 12.2 ± 0.5 µm (n=30), oval with a slightly flattened base or somewhat truncate base, hyaline at first (G-H & J), then green (J-K) and brown to dark brown when mature (J, L). The primary appendage was semi-long and flat, hyaline, frequently collapsing (J-L), they were 12.2 ± 4 × 5.2 ± 1.3 (n=10). IMI229743 (=NHL 2731) is the ex-type strain for P. marina and this description fits the original one made by Furuya and Udagawa (as Zopfiella marina: perithecium= 180-450 µm, ascospore head= (14-)15-20 × 10-13(-14) µm, primary appendage= 8-10 × 3-4 µm; [41]). Scale bars: C-D = 100 µm, E-H = 20 µm, I-L = 10 µm.
Figure 11.
Morphology of Pseudorhypophila marina IMI229743. Mycelium on medium after two weeks of growth (A-B). Perithecia, non-ostiolate, were readily differentiated on M2, M0 + miscanthus, OA, V8 and M0 + hay after 2-3 weeks incubation at 27°C with constant illumination, superficial or immersed, black, globose to subglobose, they were 287 ± 20 µm diam (n=10) (C-D). Their peridium was pale brown and semi-translucid, tissue type was textura angularis (E-F). Asci were 8-spored, clavate with a tapering and narrow apex (G-H). Ascospores were biseriate and bicellular (G-I). Ascospore heads were 17.7 ± 0.9 × 12.2 ± 0.5 µm (n=30), oval with a slightly flattened base or somewhat truncate base, hyaline at first (G-H & J), then green (J-K) and brown to dark brown when mature (J, L). The primary appendage was semi-long and flat, hyaline, frequently collapsing (J-L), they were 12.2 ± 4 × 5.2 ± 1.3 (n=10). IMI229743 (=NHL 2731) is the ex-type strain for P. marina and this description fits the original one made by Furuya and Udagawa (as Zopfiella marina: perithecium= 180-450 µm, ascospore head= (14-)15-20 × 10-13(-14) µm, primary appendage= 8-10 × 3-4 µm; [41]). Scale bars: C-D = 100 µm, E-H = 20 µm, I-L = 10 µm.
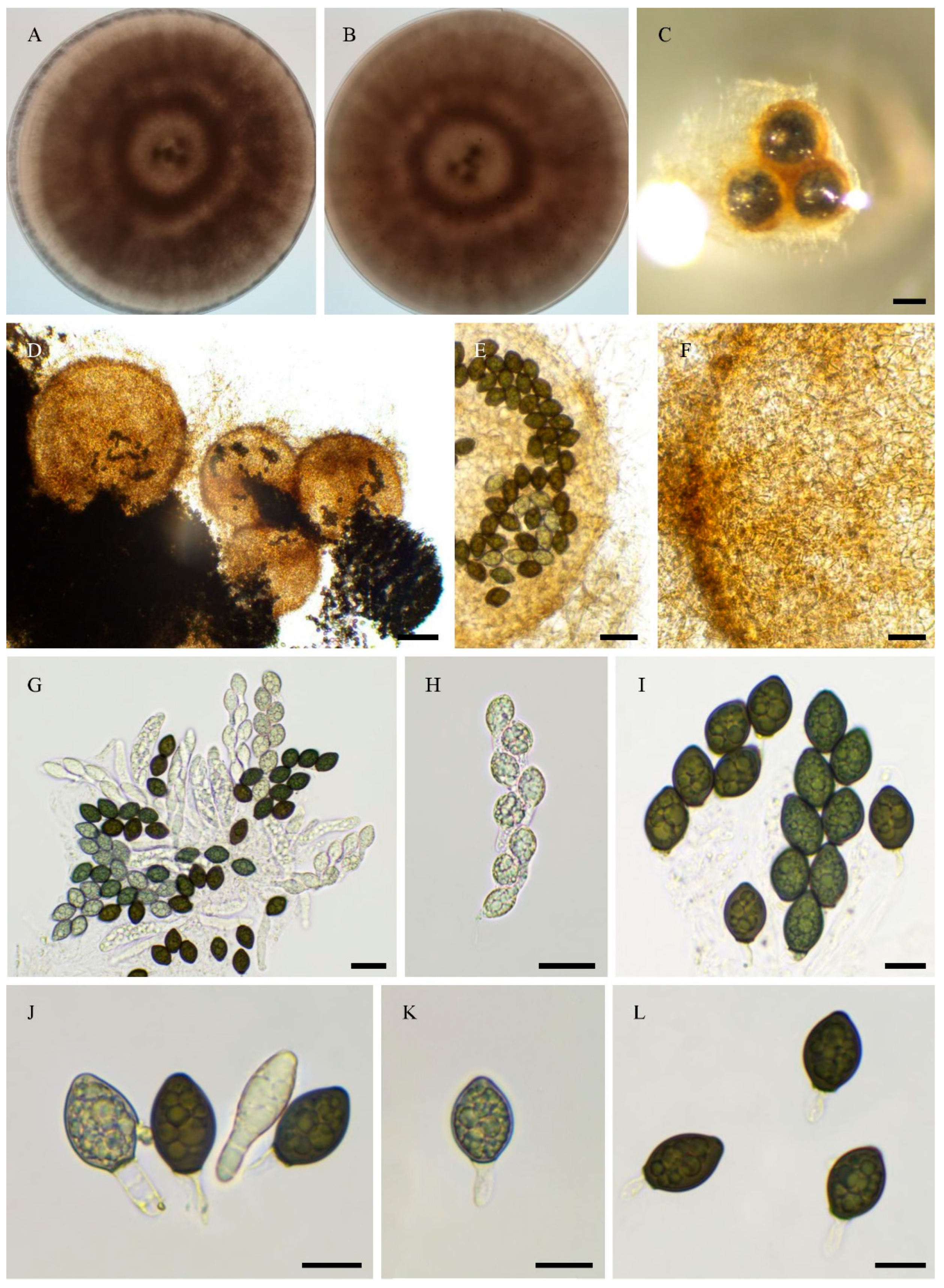
Figure 12.
Phylogenomic tree (A), in vitro fertility (B) and mating-type locus organizations (C) of the sequenced Naviculisporaceae. Presence of fruiting bodies is recorded as ostiolate perithecia (O) non-ostiolate perithecia (NO), red cross: no in vitro fruiting. For the schematic representations of the mating-type loci, the black vertical bar indicates the end of contigs, often due to the presence of repeated sequences. The red ψ denotes probable mat1-1-3 pseudogenes.
Figure 12.
Phylogenomic tree (A), in vitro fertility (B) and mating-type locus organizations (C) of the sequenced Naviculisporaceae. Presence of fruiting bodies is recorded as ostiolate perithecia (O) non-ostiolate perithecia (NO), red cross: no in vitro fruiting. For the schematic representations of the mating-type loci, the black vertical bar indicates the end of contigs, often due to the presence of repeated sequences. The red ψ denotes probable mat1-1-3 pseudogenes.

Figure 13.
Morphology of Pseudorhypophila gallica sp. nov. PSN540. Mycelium on M2 medium after two weeks of growth (A), Perithecia (B), Perithecium wall is textura angularis (C-D), Asci and ascospores (G-H), Ascospores (F-I). Scale bars: B = 400 µm, C-E = 20 µm, G-I = 10 µm.
Figure 13.
Morphology of Pseudorhypophila gallica sp. nov. PSN540. Mycelium on M2 medium after two weeks of growth (A), Perithecia (B), Perithecium wall is textura angularis (C-D), Asci and ascospores (G-H), Ascospores (F-I). Scale bars: B = 400 µm, C-E = 20 µm, G-I = 10 µm.
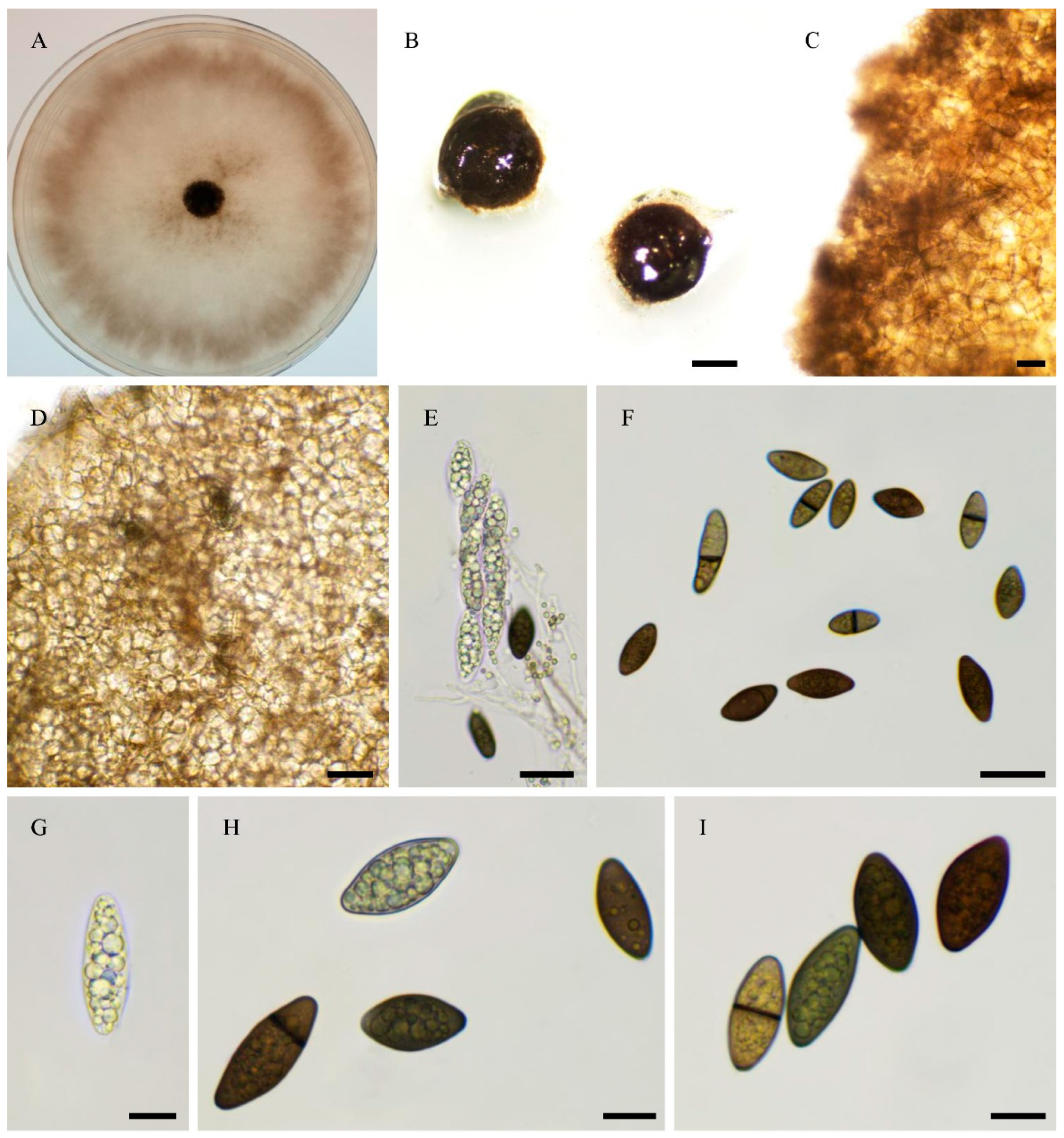
Figure 14.
Morphology of Pseudorhypophila guyanensis sp. nov. PSN2406. Mycelium on M2 medium after two weeks of growth (A), Perithecia (B), Peridium with textura angularis (C), Young asci (D), Mature asci with ascospores (E-F) Young ascospores (G-I), Mature ascospores (J). Scale bars: B = 200 µm, C-F, J = 20 µm, G-I = 10 µm.
Figure 14.
Morphology of Pseudorhypophila guyanensis sp. nov. PSN2406. Mycelium on M2 medium after two weeks of growth (A), Perithecia (B), Peridium with textura angularis (C), Young asci (D), Mature asci with ascospores (E-F) Young ascospores (G-I), Mature ascospores (J). Scale bars: B = 200 µm, C-F, J = 20 µm, G-I = 10 µm.

Figure 15.
Morphology of Rhypophila alpibus sp. nov. PSN2022. Mycelium on M2 medium after two weeks of growth (A-B), Perithecia (C-D), Perithecia wall of textura angularis (D-E), Asci and ascospores (F-G), Ascospores (H-J). Scale bars: C = 200 µm, F = 100 µm, D-E & G-J = 20 µm.
Figure 15.
Morphology of Rhypophila alpibus sp. nov. PSN2022. Mycelium on M2 medium after two weeks of growth (A-B), Perithecia (C-D), Perithecia wall of textura angularis (D-E), Asci and ascospores (F-G), Ascospores (H-J). Scale bars: C = 200 µm, F = 100 µm, D-E & G-J = 20 µm.

Figure 16.
Morphology of Rhypophila brasiliensis sp. nov. PSN1104. Mycelium on M2 medium after two weeks of growth (A-B), Perithecia (C-D), Perithecia wall was with textura angularis. (E), Asci and ascospores (F-H), Ascospores (I-L). Scale bars: D = 200 µm, E-F = 20 µm, = 100 µm, G-L = 10 µm.
Figure 16.
Morphology of Rhypophila brasiliensis sp. nov. PSN1104. Mycelium on M2 medium after two weeks of growth (A-B), Perithecia (C-D), Perithecia wall was with textura angularis. (E), Asci and ascospores (F-H), Ascospores (I-L). Scale bars: D = 200 µm, E-F = 20 µm, = 100 µm, G-L = 10 µm.
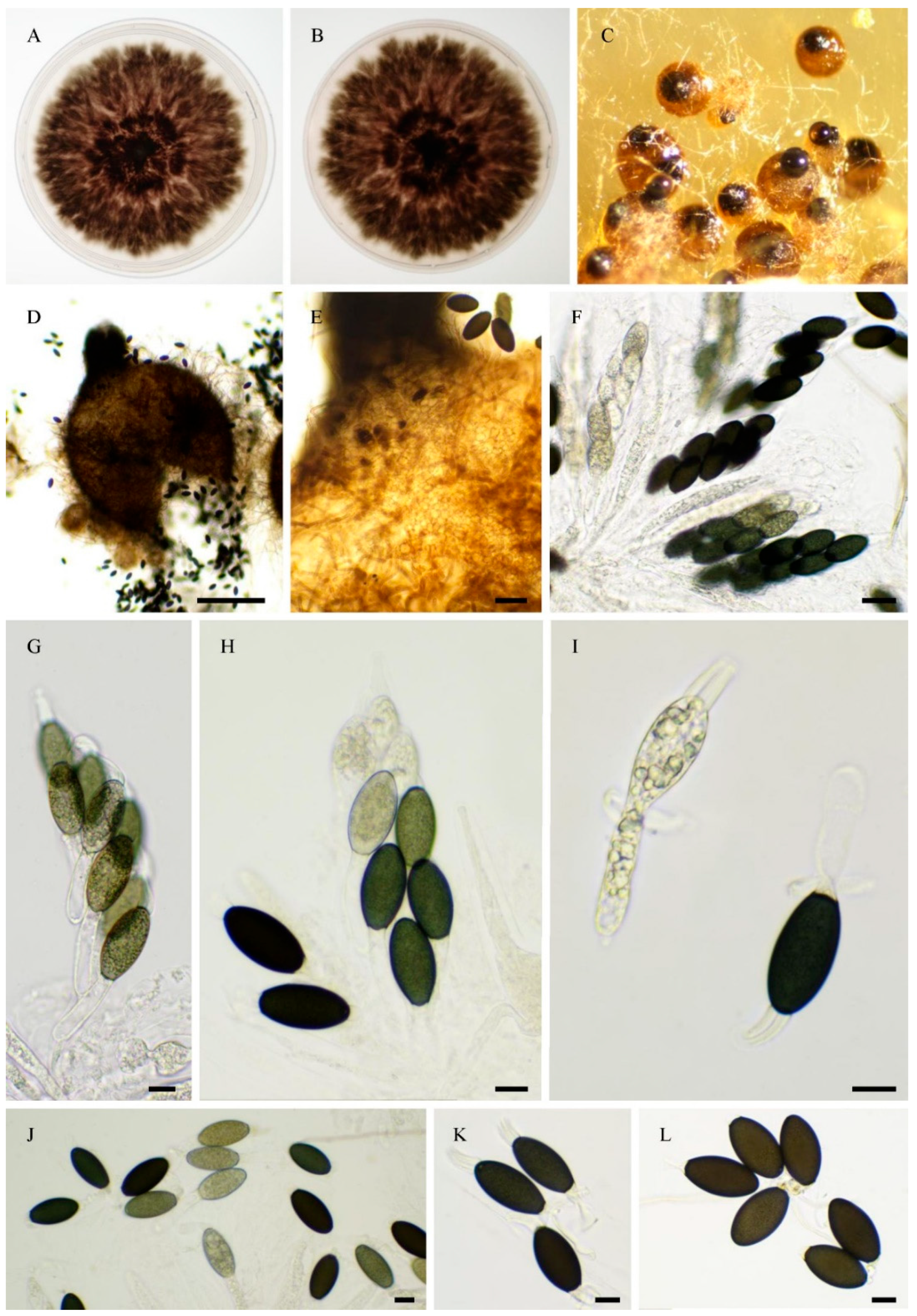
Figure 17.
Morphology of Rhypophila camarguensis sp. nov. PSN673. Mycelium on M2 medium after two weeks of growth (A-B), Perithecia (C-D), Perithecium wall with textura angularis (E), Asci and ascospores (F-G), Mature ascospores (H, J-K), Young ascospore (I). Scale Bars: C-D = 200 µm, E-I = 20 µm, J-K = 10 µm.
Figure 17.
Morphology of Rhypophila camarguensis sp. nov. PSN673. Mycelium on M2 medium after two weeks of growth (A-B), Perithecia (C-D), Perithecium wall with textura angularis (E), Asci and ascospores (F-G), Mature ascospores (H, J-K), Young ascospore (I). Scale Bars: C-D = 200 µm, E-I = 20 µm, J-K = 10 µm.
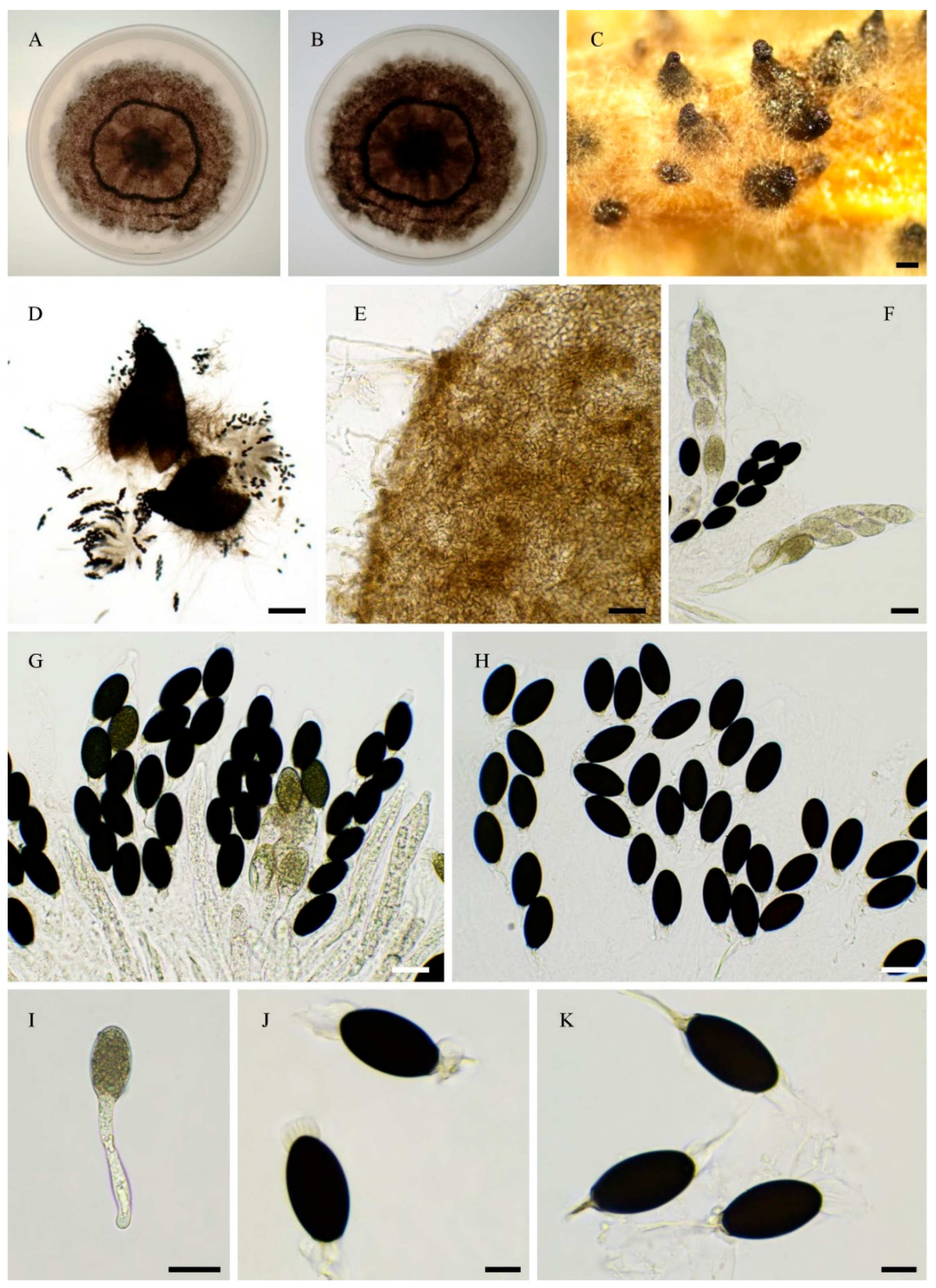
Figure 18.
Morphology of Rhypophila reunionensis sp. nov. PSN1167. Mycelium on M2 medium after two weeks of growth (A-B), Perithecia (C-D), Perithecia with neck (E), Peridium with textura angularis (F), Asci with ascospores (G-H), Yong ascospores (I-J), Mature ascospores (K-L). Scale bars: C-D = 200 µm, E = 100 µm, F-H= 20 µm, I-L = 10 µm.
Figure 18.
Morphology of Rhypophila reunionensis sp. nov. PSN1167. Mycelium on M2 medium after two weeks of growth (A-B), Perithecia (C-D), Perithecia with neck (E), Peridium with textura angularis (F), Asci with ascospores (G-H), Yong ascospores (I-J), Mature ascospores (K-L). Scale bars: C-D = 200 µm, E = 100 µm, F-H= 20 µm, I-L = 10 µm.
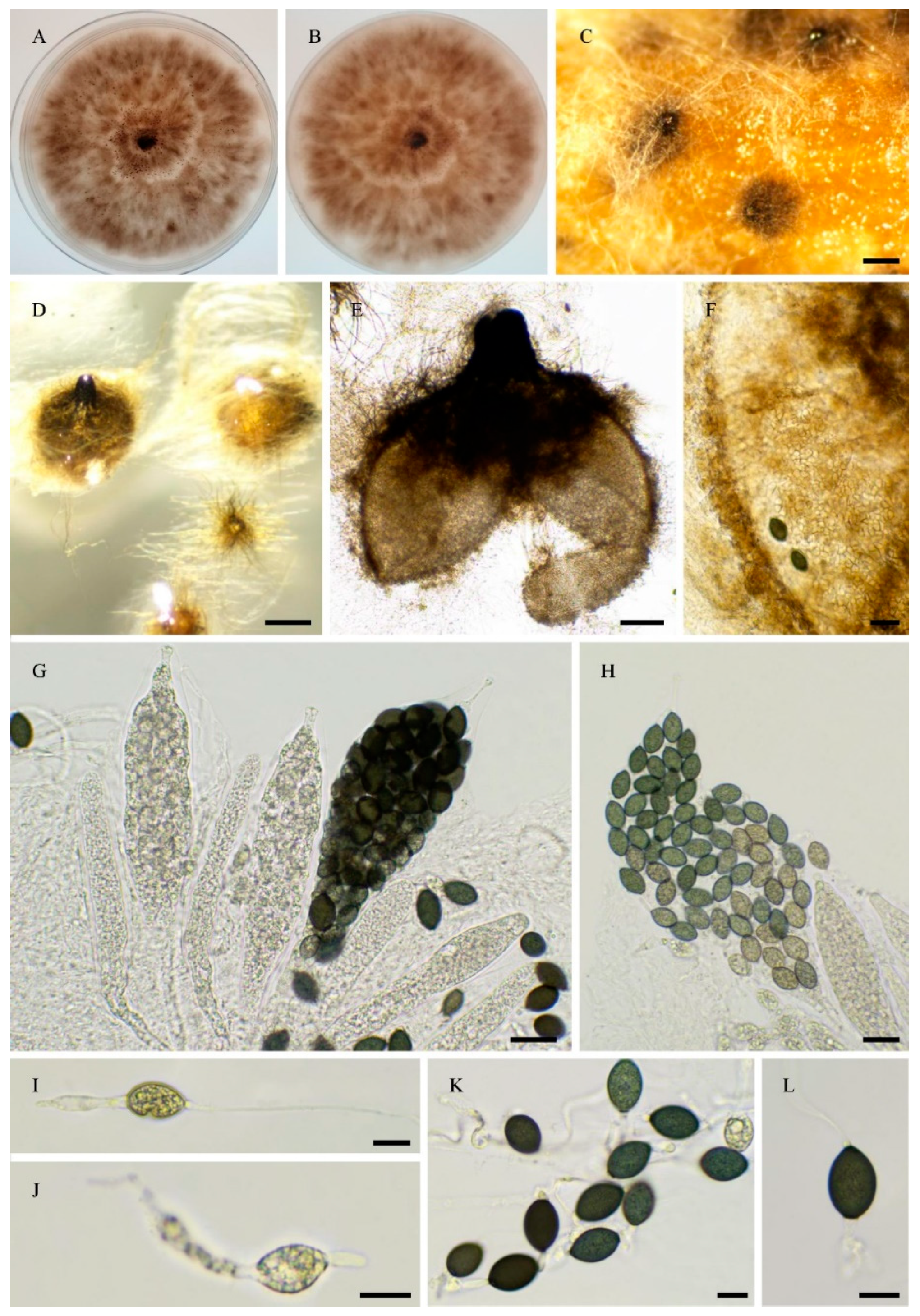
Figure 19.
Morphology of Rhypophila thailandica sp. nov. PSTH81. Mycelium on M2 medium at 6 days (A-B), Perithecia (C-D), Neck of perithecia with visible ostiole (E), Peridium with textura angularis (F), Young asci (G), Asci and ascospores (H-J), Ascospores (K-N). Scale bars: C-D = 200 µm, E, G = 100 µm, F = 20 µm, H-N = 10 µm.
Figure 19.
Morphology of Rhypophila thailandica sp. nov. PSTH81. Mycelium on M2 medium at 6 days (A-B), Perithecia (C-D), Neck of perithecia with visible ostiole (E), Peridium with textura angularis (F), Young asci (G), Asci and ascospores (H-J), Ascospores (K-N). Scale bars: C-D = 200 µm, E, G = 100 µm, F = 20 µm, H-N = 10 µm.
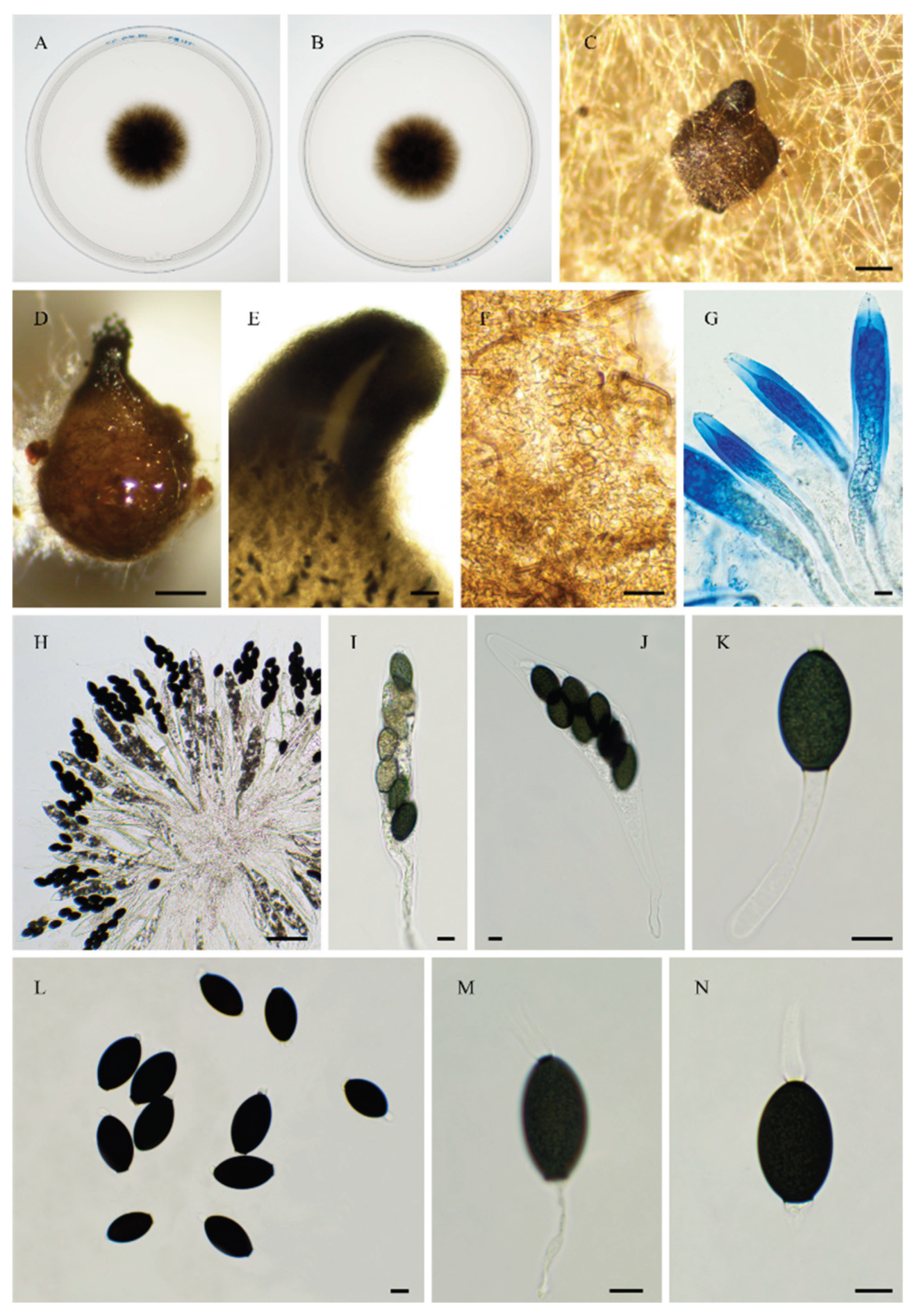

Table 1.
Strain origins and main characteristics of their genomes. Taxonomic novelties are indicated in bold.
Table 1.
Strain origins and main characteristics of their genomes. Taxonomic novelties are indicated in bold.
| Strain |
Species (other names*) |
Isolation year | Origin | Substrate | Sequencing palteform | Genome size (Mb) | Probable breeding system |
| PSN1062 | Podospora aff. araneosa | 2021 | Alps, France | Hare dung | Novogene | 40.3 | heterothallic |
| PSN850 | Naviculispora sp. | 2021 | Camargue, France | Horse dung | Novogene | 41.8 | heterothallic |
| CBS 365.69 | Arnium leporinum (Podospora leporina) | 1969 | The Netherlands | Rabbit dung | JGI | 48.5 | heterothallic |
| PSN1175 | Arnium aff. japonense | 2021 | Kobarld, Slovenia | Deer Dung | Biomics | 49.6 | heterothallic |
| PSTH81 | Rhypophila thailandica sp. nov. | 2023 | Ayuthaya, Thailand | Elephant dung | Novogene | 50.3 | homothallic |
| PSN1167 | Rhypophila reunionensis sp. nov. | 2022 | La Réunion Island, France | Cow dung | Novogene | 46.3 | heterothallic |
| PSN1104 | Rhypophila brasiliensis sp. nov. | 2021 | Campinas, Brazil | Capybara dung | Biomics | 45.9 | heterothallic |
| PSN637 | Rhypophila sp. | 2021 | Ontario, Canada | White rabbit dung | JGI | 46.9 | homothallic |
| PSN673 | Rhypophila camarguensis sp. nov. | 2021 | Camargue, France | Cow dung | Novogene | 45.9 | homothallic |
| PSN293$ |
Rhypophila decipiens (Podospora decipiens) |
2017 | Auvergne, France | Donkey dung | JGI | 47.1 | homothallic |
| PSN2105 | Rhypophila myriaspora | 2024 | Picardie, France | Horse dung | Novogene | 48.2 | homothallic |
| CBS 256.69 |
Rhypophila myriaspora (Rhypophila decipiens) |
1969 | The Netherlands | Rabbit dung | JGI | 48.3 | homothallic |
| PSN658 | Rhypophila pleiospora | 2021 | Ontario, Canada | Sylvillagus floridanus dung | JGI | 46.7 | homothallic |
| PSN2212 | Rhypophila alpibus sp. nov. | 2024 | Alps, Italy | Cow dung | Novogene | 47.9 | homothallic |
| PSTH200 | Gilmaniella sp. | 2023 | Chonburi, Thailand | Elephant dung | Novogene | 51.8 | heterothallic |
| PSN640 | Pseudorhypophila sp. | 2021 | Chad | Camel dung | JGI | 47.6 | heterothallic |
| PSTH195 | Pseudorhypophila sp. | 2023 | Chiang Rai, Thailand | Soil | Novogene | 50.6 | heterothallic |
| IMI229743 |
Pseudorhypophila marina (Zopfiella marina) |
1978 | Japan | Marine mud | JGI | 53.7 | homothallic |
| IMI350600 |
Pseudorhypophila latipes comb. nov. (Zopfiella latipes) |
1991 | U.K. | Not stated by the provider | Novogene | 52.3 | homothallic |
| PSQ110 | Pseudorhypophila latipescomb. nov. | 2019 | Québec, Canada | Soil | Novogene | 52.6 | homothallic |
| IMI229747 |
Pseudorhypophila mangenotii (Triangularia mangenotii) |
1978 | Japan | Soil | JGI | 54.3 | homothallic |
| PSN2406 | Pseudorhypophila guyanensis sp. nov. | 2025 | Guyane | Soil | Novogene | 52.6 | homothallic |
| PSN2407 | Pseudorhypophila sp.. | 2025 | Guyane | Soil | Novogene | 52.8 | homothallic |
| PSN2009 | Pseudorhypophila oryzae comb. nov. | 2024 | Centre-Val de Loire, France | Soil | Novogene | 56.2 | homothallic |
| PSN540 | Pseudorhypophila gallica sp. nov. | 2020 | Picardie, France | Soil | JGI | 55.4 | homothallic |
| PSN2022 | Pseudorhypophila gallica sp. nov. | 2024 | Centre-Val de Loire, France | Soil | Novogene | 54.6 | homothallic |
*: names under which the strain is commercially available or was sequenced. $: PSN293 genome sequence was previously reported [14].
Table 2.
ITS and LSU accession numbers of the strains used in the ITS + LSU phylogenetic analysis.
| Strain | Taxa | ITS | LSU |
| CBS 118394 | Apodospora peruviana | EU573703 | KF557665 |
| CBS 506.70T | Apodus deciduus | NR_145141.1 | NG_056953 |
| CBS 376.74 | Apodus oryzae | AY68120 | AY681166 |
| CBS 215.60 | Areotheca ambigua | AY999137 | AY999114 |
| UAMH 7495 | Areotheca areolata | AY587911 | AY587936 |
| Lundqvist 7098-e | Arnium caballinum | NA | KF557672 |
| SANK 10273 | Arnium japonense | NA | KF557680 |
| Lundqvist20874-c | Arnium mendax | NA | KF557687 |
| E00122117 | Arnium mendax | NA | KF557688 |
| NBRC 9235T | Gilmaniella humicola | available at https://www.nite.go.jp/nbrc/catalogue/ | |
| CBS 137295T | Naviculispora terrestris | MT784136 | KP981439 |
| NTUPPMCC 22-297T | Pseudorhypophila formosana | PV476805 | PV476867 |
| CBS 155.77T | Pseudorhypophila marina | MK926851 | MK926851 |
| CBS 698.96 | Pseudorhypophila marina | MK926853 | MK926853 |
| CBS 413.73T | Pseudorhypophila pilifera | MK926852 | MK926852 |
| CBS 419.67T | Pseudorhypophila mangenotii | MT784143 | KP981444 |
| CBS 249.71 | Rhypophila cochleariformis | AY999123 | AY999098 |
| CBS 258.69 | Rhypophila decipiens | KX171946 | AY780073 |
| TNM F17211 | Rhypophila myriaspora$ | EF197083 | NA |
| TNM F16889 | Rhypophila pleiospora | EF197084 | NA |
| F-116,361 | Sordaria araneosa | FJ175160 | NA |
| IFO9826 | Zopfiella latipes | AY999129 | AY999107 |
T: type. $: Marin-Felix et al. [11] labelled this strain as R. myriaspora in their phylogenetic tree, which is the name classically used, and R. myriospora in their diagnosis, which is the name now retained in several databanks. In the present paper, we use the traditional name of R. myriaspora, as the protolog described the species as Sordaria myriaspora Crn. Mscr. [28]. NA: not available.
Table 3.
Average nucleotide identity (ANI) between selected Rhypophila spp. strains. The ANI value in red identifies strains assigned to the same species.
Table 3.
Average nucleotide identity (ANI) between selected Rhypophila spp. strains. The ANI value in red identifies strains assigned to the same species.
| PSN2212 | PSN2105 | CBS 256.69 | PSN293 | |
| PSN658 | 94.99% | 95.07% | 95.08% | 95.17% |
| PSN2212 | 95.08% | 95.09% | 95.21% | |
| PSN2105 | 99.74% | 95.20% | ||
| CBS 256.69 | 95.20% |
Table 4.
Average nucleotide identity (ANI) between selected Pseudorhypophila spp. strains. The ANI value in red identifies strains assigned to the same species.
Table 4.
Average nucleotide identity (ANI) between selected Pseudorhypophila spp. strains. The ANI value in red identifies strains assigned to the same species.
| PSQ110 | IMI350600 | IMI229747 | PSN2406 | IMI229743 | PSN540 | PSN2022 | PSN2407 | PSN2009 | |
| PSTH195 | 87.90% | 88.47% | 88.49% | 87.75% | 89.14% | 87.86% | 87.86% | 87.85% | 87.87% |
| PSQ110 | 99.26% | 87.91% | 87.25% | 88.36% | 87.34% | 87.36% | 87.28% | 87.42% | |
| IMI350600 | 87.93% | 87.25% | 88.37% | 87.38% | 87.38% | 87.32% | 87.46% | ||
| IMI229747 | 88.25% | 89.02% | 88.35% | 88.36% | 88.35% | 88.35% | |||
| PSN2406 | 87.78% | 94.18% | 94.18 | 94.10% | 94.00% | ||||
| IMI229743 | 87.90% | 87.91% | 87.84% | 87.95% | |||||
| PSN540 | 98.69%* | 96.28% | 97.26% | ||||||
| PSN2022 | 96.43% | 97.30% | |||||||
| PSN2407 | 96.44% |
*Although ANI is lower than 99%, PSN540 and PSN2022 belong to the same species. Low ANI was due to the presence of about 4% of DNA regions diverging by 10%-15% (see Figure 7).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



