
Submitted:
28 October 2025
Posted:
29 October 2025
You are already at the latest version
Abstract
The atmosphere plays a pivotal role in modulating the interactions between microorganisms and their surrounding environments, influencing ecological cycles, heritage conservation, and providing opportunities for novel applications. Recent studies have highlighted the role of microbial responses to atmospheric conditions as indicators of environmental change. This study highlights the role of climate change, particularly rising temperatures, on the growth of cyanobacteria and, consequently, the impact of this on the conservation of cultural heritage, as in the case study of the rock paintings of the Majella Massif (Lama dei Peligni - Abruzzo Region, central Italy). The region’s rock art, characterized by red and black schematic motifs, is increasingly impacted by microbial colonization, driven by climate-induced temperature variations. These impacts are consistent with broader research demonstrating the link between microbial growth patterns and climatic factors. Laboratory analyses were carried out on cyanobacteria samples collected near the rock paintings at the study site in the Majella National Park. Results revealed a significant increase in growth rates at the higher temperature, demonstrating their sensitivity to climatic shifts. These findings underscore the dynamic role of atmospheric factors in shaping microbial survival and propagation. Consequently, certain atmospheric parameters appear to play a crucial role in the deterioration of fragile cultural assets. Indeed, the enhanced growth of cyanobacteria due to rising temperatures also poses a challenge: their proliferation can degrade cultural heritage sites, threatening their preservation. This research advocates for interdisciplinary approaches that integrate atmospheric sciences, microbial ecology, and heritage studies to explore the role of temperature in affecting cyanobacteria growth and the conservation of a peculiar cultural heritage in the Majella Massif. By leveraging their biological traits, cyanobacteria can provide valuable insights into climate dynamics while emphasizing the urgency for proactive strategies to mitigate environmental impacts on vulnerable ecosystems and heritage sites.
Keywords:
climate change
; temperature
; cyanobacteria
; rock paintings
; cultural heritage
; atmosphere
1. Introduction
In the territories of the Maiella National Park there is the highest concentration of examples of rock art of the entire Italian Apennines Mountain chain [1]. This local heritage has increased over the last decades as a result of important discoveries. The largest catalogue [2] on the rock art of central Italy provides a first chronological and typological picture of over 30 sites documented in Abruzzo Region (Italy), placing it in the ambit of the post-paleolithic schematic art of the Mediterranean. Numerous testimonies have been found in a rather limited location. In fact, thanks to the investigations carried out since 2006, the panorama of prehistoric rock art sites in the valleys of Sangro and Aventino river (Chieti province) has greatly expanded. This extraordinary concentration of sites that have similar characteristics but also important differences, has allowed to start a systematic methodological approach for the interpretation of these sites, their environmental contexts and the recurring themes of their representations [3]. The dating and frequenting of the places can be traced back to the late Bronze Age and/or early Iron Age [4]. The black and red paintings (and engravings) include anthropomorphic motifs but mainly geometric and irregular patterns. The three sites of Civitalupparella and the site of Fallo (Chieti) have similarities also under the iconographic profile being united by a recurring symbol of extreme significance: the crusader circle, a stylized representation of the sun. All of these sites are located in inaccessible locations; the small space available suggests, in fact, that they were designed for initiatory rites for a few people. In some cases, the natural characteristics of the places and the proximity of the sites, suggest the presence of ceremonial landscapes (a kind of ritual path) as their possible interpretation. Specifically, in this study we focused on the difficult-to-reach site of “Riparo La Pineta” (La Pineta shelter) at Lama dei Peligni () (Figure 1).
In this site, already included in the census by Mattioli in 2007 [2,5] but not investigated, there is evidence of red anthropomorphic motifs paintings (Figure 2a). These red paintings are covered in some areas by a black biofilm which alters the appearance of surfaces (Figure 2b). The growth of peculiar biological organisms on a specific stone surface is determined, on the one hand, by environmental factors such as temperature, relative humidity, light conditions, levels of atmospheric pollution, wind and rainfall and, on the other hand, by the nature and properties of the stone itself (mineral composition, pH, relative percentage of various minerals, salinity, moisture content and consistency).
Therefore, the response of organisms to a potentially colonisable surface depends on the ecological and physiological requirements of the biological species involved [6,7,8]. There is a wide range of literature from recent decades on the fundamental role that biological agents play in the deterioration of stones. By now it is well established that the microbiota on stone represents a complex ecosystem which develops in various ways, depending on environmental conditions and the physicochemical properties of the material in question. The complex physical and chemical interactions of atmospheric pollutants with the mineral material accelerate the colonization of stones [9,10,11,12,13,14,15,16,17,18]. The biodeterioration of exposed stone depends mainly on the availability of light, water and nutrients. Microorganisms are able to obtain several elements that they need for their metabolism (e.g., calcium, aluminium, silicon, iron and potassium) from the limestone by biosolubilization. This biosolubilization process generally involves the production of various organic and inorganic acids by micro-organisms [6,12]. Algae and cyanobacteria are usually the first colonisers of stone surfaces due to their photosynthetic nature, causing both chemical and mechanical deterioration. The microclimate determines the degree of colonization, the type of community and its specific composition [19]. Their presence can be detected through the patinas or crusts they form [6,13]. The cyanobacteria can grow in the form of subaerial biofilms, of different colours (white, red, blue and black), complex communities that adhere to a surface and are exposed to the atmosphere [20,21]. They play a fundamental role in the ecology of many different habitats [22,23]. The characteristics that influence the colonisation of surfaces by cyanobacteria, determining the bioreceptivity, i.e. the ability of a material to be colonised by living organisms, are varied [24,25]. Among these characteristics are surface roughness and stone lithotype [19,24,26], angle of inclination, surface geometry and degree of shading [19,27,28,29]. The cyanobacteria adapt to the quality of light (spectral composition) by modifying the composition of their pigments and/or photosynthetic apparatus. Light therefore represents a determining factor in niche differentiation in natural environments [30,31]. Furthermore, these subaerial biofilm can serve as a substrate for the growth of other deteriogens. Both these and the cyanobacteria themselves can cause in association aesthetic, chemical, and physical decay affecting the durability of the material [11]. As highlighted by Macedo et al. 2009, reviewing studies conducted since the 1990s, the most widespread and commonly reported taxa in the stone cultural heritage of the Mediterranean Basin (Mediterranean climate) are, among cyanobacteria, Gloeocapsa, Phormidium and Chroococcus and, among chlorophytes, Chlorella, Stichococcus and Chlorococcum. These genera have been found associated with all lithotypes (siliceous or calcareous). The preference of cyanobacteria and chlorophytes for a specific stone substrate is not easy to correlate immediately. The data examined suggest that green algae and cyanobacteria can colonise a wide variety of substrates. In the complex set of factors mentioned above, it is often difficult to determine the influence of each of them individually; assessing their combined effects, synergy and dynamics is complex, and all factors are likely to be relevant. These biofilms can have a dual role on stone, oscillating between bioprotection and biodeterioration, depending on the surrounding environment. On a case-by-case basis, biodeterioration and bioprotection must be assessed comprehensively and quantitatively, taking into account the active and dominant species, the specific lithic matrix, environmental conditions, etc. [32]. Besides, different perceptions of issues related to biodeterioration generally are affected by the type of surface of the heritage concerned (a statue, a fountain, a building, a façade, an archaeological site, etc.) and on local cultural tradition [33]. In general, however, any covering caused by micro-organisms detracts from the original appearance of the heritage site. Therefore, conservators of outdoor stone heritage, particularly in the Latin cultural area, consider it a priority to keep all stone heritage surfaces clean (i.e. free of biological patina) and manage conservation plans accordingly [33]. Devitalisation and mechanical removal of lichen thalli and microbial biofilms are therefore regularly included in restoration interventions [34]. However, the widespread use of synthetic chemicals such as biocides, practised for decades, is now increasingly considered environmentally unsustainable, and new alternative products and/or chemical-free approaches to controlling biological growth are continually being sought [35,36]. Recent studies have highlighted the role of microbial responses to atmospheric conditions as indicators of environmental change [37,38]. Although it has been demonstrated that local climatic conditions influence the microbial population and community on stone, comparative studies on the microbiome of stone in different climatic conditions are very limited for a better understanding of this research topic [39,40,41,42,43]. The impact of climate change in this area (Abruzzo Region, Italy) have shown clear variations in terms of the atmospheric mean temperature. In this study, the effect of this parameter on the growth of bacteria responsible for the black patina has been evaluated, using the Majella Massif region of Central Italy as a case study. These impacts align with broader research showing the link between microbial growth patterns and climatic factors [38,44,45,46,47].
2. Materials and Methods
2.1. Research Design
In the territories of the Majella National Park (Figure 3) there is the highest concentration of examples of rock art of the entire Italian Apennines Mountain chain. This great Cultural Heritage is well documented by large catalogues (see Section 1), placing Abruzzo Region in the context of the post-paleolithic schematic art of the Mediterranean. In this framework, at the Lama dei Peligni (Abruzzo) site, black stripes are affecting and covering the listed red rock paintings. In order to assess their origin and any actions that could mitigate their effects on the paintings, two samples (May 21st, 2024 and July 10th, 2024) were taken (Figure 3) with the goal of conducting dedicated laboratory analyses (see Section 2.3), which revealed the presence of organic (i.e. cyanobacteria [48]) and inorganic substance (see Section 3).
Tests related to the average atmospheric temperature (see Section 2.2) were performed. Laboratory analysis, confirming the presence of cyanobacteria biofilms near the rock paintings, pave the way to growth tests, which were conducted in relation to the average atmospheric temperature. These tests aim to show the role of increasing temperature in the growth of cyanobacteria (see Section 3).
2.2. Atmospheric Analysis
Meteorological data are provided by the Regional Hydrographic Office. We accessed temperature data spanning from 1930 to 2023 for the Palena and Lama dei Peligni site stations (Figure 4). In this research, we applied a quality check that evaluated potential anomalies such as missing data or spikes. Two temperatures were selected for the tests: an average of the maximum temperatures for the period 1930-1970 at the Palena site (14.3 °C), the closest station (≈ 7.5 km) to the Lama dei Peligni site for which data were available, and an average of the maximum temperatures for the year 2023 at the Lama dei Peligni site (18.6 °C).
2.3. Laboratory Tests
Laboratory tests were performed to classify the different components of the biofilm samples and identify the cyanobacteria, with the aim of assessing their growth in relation to temperature variations linked to climate change in the area of interest. After the classification, as reported below, the collected samples have been analysed at two selected temperatures: 14.3 °C and 18.6 °C (see Section 2.2). The culture medium used was BG11 plus 1.2% of agar bacteriological to isolate the bacterial colonies. Plates were incubated at the two temperatures for four weeks and exposed at natural light/darkness cycles.
2.3.1. Cytometry Analysis
Bacterial samples collected by sterile swabs were analyzed by flow cytometry. Briefly, 1×10⁶ cells per sample were stained with SYTO9 at a final concentration of 5µM (Invitrogen, ThermoFisher Scientific, Waltham, MA, USA) and incubated for 30 minutes at room temperature in the dark. After incubation, the samples were washed, and at least 20,000 events per sample were acquired using either a FACSVerse™ flow cytometer or a FACSAria™ III cell sorter (both from BD Biosciences, San Jose, CA, USA). A trigger threshold was used on the FITC channel (SYTO9 emission) to exclude all non-nucleated events. Fluorescence Minus One (FMO) controls were used to identify positive populations. The Cytometer Setup & Tracking (CS&T) system was employed for daily instrument quality control. Furthermore, given that cyanobacteria exhibit an emission peak around 650–660 nm, cells ( events) emitting in the Alexa Fluor 647 channel () were isolated by cell sorting using a nozzle. Those cells were further cultured.
2.3.2. Genetics Analysis
Starting from the environmental samples, bacterial DNA was extracted using the MagPurix instrument and the Bacterial DNA Extraction Kit (Zinexts Life Science Corp., Taipei, Taiwan-CatZP02006) according to the manufacturer’s protocol. To ensure the accuracy and reliability of the metagenomic sequencing results, we included both negative and positive controls in our analysis. Negative controls, such as blank swabs and extraction blanks, were used to detect any potential contamination introduced during the sample processing and sequencing stages. Positive controls, consisting of known microbial communities, were included to verify the efficacy and sensitivity of the sequencing process. These controls allowed us to validate the presence of the detected microbial taxa and ensure that the results were not artifacts of contamination. The bacterial DNA was quantified with a Qubit 3.0 fluorometer (ThermoFisher, Waltham, MA, USA). NGS was performed with the Ion 16STM Metagenomics Kit (ThermoFisher, Waltham, MA, USA), designed for the analyses of mixed microbial populations using the Ion TorrentTM semiconductor sequencing workflow. The kit permits PCR amplification of the hypervariable regions of the 16S rDNA gene from bacteria. The kit includes two primer sets that selectively amplify the corresponding hypervariable regions of the 16S region in bacteria: Primer set V2-4-8; Primer set V3-6, 7-9. After the quantification of libraries with the Real-Time Step One PCR System (Thermo Fisher Scientific, Waltham, MA, USA), the amplified fragments can then be sequenced using the Ion S5TM (ThermoFisher, Waltham, MA, USA) platform, loaded onto an Ion 520TM chip and analyzed using the Ion 16STM metagenomics analyses module within the Ion ReporterTM software 5.14 (ThermoFisher, Waltham, MA, USA), enabling a rapid and semi-quantitative assessment of complex microbial samples. These comprehensive primer sets allow for accurate detection and identification of a broad range of bacteria down to the genus or species level. The primer sets are paired with Environmental Master Mix v2.0 that is optimized to tolerate high levels of PCR inhibitors and amplify targets from complex samples such as environmental, food, tissue, and other challenging samples. The combination of the two primer pools allows for the sequence-based identification of a broad range of bacteria within a mixed population. The bioinformatics analysis was performed with the Krona Software, to describe all the phylogenetic classes, starting from the major phyla.
2.3.3. Inductively Coupled Plasma—Mass Spectrometry (ICP-MS) Analysis
Samples of moss, blackened moss, blackened soil and rock surface were digested using acidic digestion by adding 69% () at a sample-to-acid ratio of , heating at 80 ∘C for 3 hours. After that digested samples were diluted with high-purity water (18 resistivity) to have a final concentration of 7% () . The same concentration of was used for in solution standard curve (levels from to ) using the multielement mixture (Ag, Ba, Be, Cd, Co, Cr, Cu, Fe, Mn, Ni, Pb, Rb, Se, Sr, Tl, U, V, and Zn) at 10 (Agilent Technologies, Santa Clara, CA, USA). Samples and calibrators were analyzed in triplicate by Inductively Coupled Plasma Mass Spectrometry (ICP-MS) with the Agilent 7900 ICP-MS quadrupolar mass spectrometer (Agilent Technologies, Santa Clara, CA, USA). An internal standard correction was performed by online addition of an internal standard solution of Y, Bi and In at 200 ) in a T-piece. In particular, some elements (Cr, Co, Se, Cu, Ni, Al, As, Cd, Pb, Zn and Tl) were acquired in helium (He) mode to reduce spectral interference and noise effects; other elements (V, Mn and Ag) were measured in no-gas mode due to the lack of interference. All specifications for ICP-MS acquisition in terms of its optimization are reported in Chiaudani et al. [49]. Whereas detailed instrumental parameters and operating ICP-MS conditions were detailed in Table 1.
2.3.4. Shotgun Proteomics Analysis
Shotgun proteomics was performed to assess the protein composition of sample containing 1 million cyanobacteria isolated by instrumental sorting from samples collected in the Lama dei Peligni area. The biological sample was concentrated with 30 KDa filters (Amicon®Ultra,Merck Millipore) and was lysed by sonication on ice (Sonicator U200S control, IKA Labortechnik, Staufen, Germany) at cycle 0.4 and 40% amplitude in a lysis buffer (urea 6 M in 100 mM Tris/HCl, pH = 7.5). Protein concentration was determined using the Bradford assay (Bio-Rad, Hercules, CA, USA) with Bovine Serum Albumin (BSA, Sigma-Aldrich, St. Louis, MI, USA) as the standard for the calibration curve, allowing digestion of 50 g of protein of sample. Filter Aided Sample Preparation (FASP) protocol was used for overnight tryptic digestion at 37 °C. The sample was analyzed in triplicate by injecting 3 l of digested proteins by nano LC-MS/MS through Ultimate™ 3000 UPLC (Thermo Fisher Scientific, Milan, Italy) chromatographic system coupled to the Orbitrap Fusion™ Tribrid™ (Thermo Fisher Scientific, Milan, Italy) mass spectrometer, using the column Easyspray PEPMAP RSLC C18 2 m, 25 cm x 75 µm (Thermo Fisher Scientific, Milan, Italy). The sample was acquired with a flow rate set at 300 nL/min and a total run time of 65 minutes, employing a chromatographic gradient increasing from 5% to 90% of phase B (mobile phase A: 0.1% formic acid in water; mobile phase B: 0.1% formic acid in acetonitrile). The MS parameters and acquisitions were previously described in two works [50,51] used in mass spectrometry employ the high-energy collision-induced dissociation (HCD) technique, which enables the fragmentation of selected molecular ions during acquisition. The MS/MS raw data were processed using Proteome Discoverer 2.5 software (Thermo Fisher Scientific, Milan, Italy), through the “All Entries” reviewed database (25,252 entries, released in 2021, UniProtKB/SwissProt) and Cyanobacteriota/Melainabacteria group (id: 1798711) (3,578 entries, released in 2024, UniProtKB/SwissProt) with a false positive rate (FDR) of 1% at protein level for both search engines. For the protein identification, the following processing filters were applied: all reported proteins were identified as “master proteins” and, additionally, a high confidence threshold was set, including in the dataset only proteins that were present with high confidence in at least 2 out of 3 analytical replicates. Functional enrichment and pathway analysis were performed using STRING (Search Tool for the Retrieval of Interacting Genes/Proteins) software, a bioinformatics tool used to explore protein-protein interactions based on functional relationships. p-values of functional networks were corrected for multiple testing within each category using the Benjamini–Hochberg procedure.
3. Results
Microorganism, obtained by resuspending the material collected from the rock paintings in 1X PBS, were analyzed by flow cytometry. As shown in Figure 5, a population of cells was identified in the side scatter height (SSC-H) versus forward scatter height (FSC-H) dot plot (“Morph” gate, Figure 5a), using a 1X PBS sample as a negative control (Figure 5d-Figure 5e). This population, containing a discrete amount of DNA (Figure 5b-Figure 5c), as indicated by the histogram shown in Figure 5b, where the blue peak represents the sample stained with SYTO9, while the red peak corresponds to its respective fluorescence minus one (FMO) control. Such a population of cells, obtained from an area of the painting covered by the biofilm, exhibited fluorescence in the Alexa Fluor 647 channel (Figure 5e), consistent with cyanobacterial pigment emission. A sample obtained from a painting control area (not covered by the biofilm) was used as a control (Figure 5d).
The sequencing performed with the 16S Metagenomics Kit revealed an important result. Using the Ion Reporter Software with the Krona Software, we analyzed all the phylogenetic classes, starting from the major phyla. In particular, in the sample 1 (Figure 6) the phylum cyanobacteria revealed 18% of the entire microbial population, with 32271 total reads, while in the sample 2 (Figure 7) cyanobacteria was found only with 3% with 5948 total reads.
The multi-elemental concentrations detected in the different samples were listed in Table 2. The main differences between moss and blackened moss shed light on probable presence by heavy metals (Pb, Cd, Zn and Mn) especially of the blackened samples. These data may confirm the colonization of cyanobacteria in areas where the moss appears blackened, possibly due to heavy metal stressors acting as a selective pressure.
As the Majella Massif hosts prehistoric rock art now threatened by microbial colonization, we performed proteomics analysis on cyanobacteria isolated by FACS. Comparing the searches through two databases, 22 proteins were robustly identified (9.44%) respect to “All entries” database reclassified as Cyanobacteriota phylum, as shown in Venn diagram of Figure 8. When research was reduced to only Cyanobacteriota phylum, 33 proteins were reclassified in Cyanobacteriota/Melainabacteria group. Of them, 22 proteins were the same of the previous identification in “All entries” database. Most of the identified proteins were reclassified to the species Nostoc punctiforme, a filamentous cyanobacterium characterized as one of the most active inteins, as demonstrated by the number of peptides with which they have been identified because it is greater than the proteins reclassified with other species of cyanobacteria. We studied PPI of these 33 proteins in order to visualize functional protein clusters, as shown in Figure 9a and Figure 9b, demonstrating that they are involved in photosynthesis and ATP metabolism. The whole protein list was reported in supplementary materials.
Following laboratory analyses aimed at characterising the sampled compound, and once the actual presence of cyanobacteria had been verified, we carried out a growth test on the colonies collected in situ. Tests at the two temperatures (see Section 2.2) showed two different growth rates: 40cfu(colony-forming units)/ml at 18.6°C and 35cfu/ml at 14.3°C). The results show how the effect of climate change, in particular temperature variations, has an impact on the growth of bacterial colonies and consequently on heritage conservation, highlighting the key role of the study of environmental changes.
4. Discussion
This study investigated the role of climate-driven temperature variability on the growth of cyanobacteria colonizing the open-air rock paintings of Lama dei Peligni (Majella Massif, Abruzzo, Italy). By combining atmospheric analysis with microbiological, chemical and proteomic data, we provide converging evidence that higher mean temperatures, consistent with recent regional warming, promote cyanobacterial proliferation on lithic substrates, with direct implications for the conservation of prehistoric rock art.
Flow cytometry and metagenomic profiling confirmed the presence of cyanobacteria as a substantial fraction of the microbiome on blackened surfaces (up to 18% of total reads in the most colonized sample), while proteomics identified proteins predominantly assigned to Nostoc punctiforme. The detected functional signatures associated with photosynthesis and ATP metabolism indicate metabolically active populations capable of sustaining growth under fluctuating microclimatic conditions. These observations align with prior work in Mediterranean settings showing that temperature, moisture and light regime jointly modulate lithobiont colonization and biofilm development on stone [19,38,45].
Growth assays performed at two representative temperatures, 14.3 ∘C (historical average of maximum temperatures, 1930–1970) and 18.6 ∘C (average of maximum temperatures for 2023), showed higher cfu at the warmer condition (40 vs. 35 cfu/mL). Although modest in absolute terms, this difference is directionally consistent with a temperature-driven acceleration of microbial proliferation, and mirrors expectations from metabolic theory and empirical reports on biofilm expansion under warmer climates on exposed heritage [38,45]. In situ, even small shifts in growth rates can amplify over seasonal cycles, tipping the balance towards thicker and more persistent subaerial biofilms that obscure pigment layers, alter surface optics, and contribute to chemical and mechanical decay via hydration cycles, pigment deposition and acid-mediated mineral dissolution [11,12].
Elemental analysis further revealed higher concentrations of Pb, Cd, Zn and Mn in blackened moss relative to unblackened moss, suggesting that heavy metals may act as selective pressures favouring tolerant cyanobacterial taxa and/or mixed biofilms with enhanced stress resilience. Such metal enrichment has been associated with shifts in lithic microbiomes and with black crust formation patterns used as markers of environmental change on heritage limestones [16,40]. While causality cannot be inferred from our cross-sectional data, the co-occurrence of higher temperature signals and metal loadings points to multi-stressor scenarios in which warming and pollutant availability jointly facilitate colonization and persistence.
From a conservation standpoint, these findings underscore the need to integrate climate indicators into routine diagnostic and preventative strategies for open-air rock art. First, site-based microclimate monitoring (temperature, RH, wetting/drying frequency, irradiance) should be coupled with periodic, minimally invasive microbiological assessments (e.g., qPCR or metabarcoding panels targeting cyanobacteria) to detect early deviations towards higher bioreceptivity states [19]. Second, given the environmental and regulatory concerns around repeated use of broad-spectrum biocides, the evaluation of lower-impact control options (e.g., essential oils or other nature-based antimicrobials, photodynamic or photocatalytic treatments, and cleaning protocols tuned to local bioreceptivity) is warranted [35,36]. These interventions should be framed within adaptive management plans that account for seasonal windows of highest biofilm activity and the specific geometry and shading of decorated shelters, known determinants of colonization intensity [19,28].
This work has limitations. Laboratory growth was assessed at two discrete temperatures without full replication across a gradient, and no formal hypothesis testing was applied to the cfu differences. Moreover, other key drivers (light spectrum/intensity, surface moisture dynamics, nutrient availability, and wind-driven rain) were not manipulated experimentally and may modulate the temperature response in situ [46,47]. In summary, our interdisciplinary evidence indicates that recent warming conditions are consistent with enhanced cyanobacterial activity on the Lama dei Peligni rock paintings, reinforcing concerns that climate change can accelerate biodeterioration in Mediterranean open-air heritage. Translating these insights into practice will require climate-informed monitoring, targeted and sustainable biocontrol trials, and site-specific risk models that jointly consider climate trends, pollutant loads and substrate bioreceptivity.
5. Conclusions
This study provides converging evidence that recent warming consistent with regional climate change can enhance cyanobacterial proliferation on lithic substrates at the Lama dei Peligni rock art site (Majella Massif, Italy). Integrating atmospheric records with microbiological, chemical and proteomic analyses, we showed that higher temperatures are associated with increased cyanobacterial growth, metabolically active biofilms (with proteins largely assigned to Nostoc punctiforme), and co-occurring elemental patterns suggestive of metal-related selective pressures. Even modest laboratory differences in growth at 18.6 ∘C versus 14.3 ∘C may translate, in situ, into thicker and more persistent subaerial biofilms that obscure pigments and accelerate chemical and mechanical decay of painted surfaces.
Our findings support the urgent incorporation of climate indicators into conservation diagnostics and preventative management for open-air rock art in Mediterranean settings. We recommend (i) site-specific microclimate monitoring linked to periodic, minimally invasive microbiome assessments; (ii) evaluation of lower-impact biocontrol options as alternatives or complements to synthetic biocides; and (iii) scheduling interventions around seasonal windows of peak bioreceptivity.
Future work should expand experimental designs to multi-factor gradients (temperature, irradiance, wetting/drying) and couple proteogenomic surveys with high-resolution imaging of paint layers to quantify aesthetic and material outcomes through time. Such climate-informed, interdisciplinary approaches are essential to mitigate the escalating biodeterioration risks posed by cyanobacterial colonization to vulnerable cultural heritage sites.
Author Contributions
Conceptualization, P.C., F.P., E.M.S., E.A., S.D., P.D.C., and A.M.; methodology, P.C., F.P., E.M.S., E.A., S.D., P.D.C., and A.M.; formal analysis, P.C., P.S., P.L., S.D.L., M.D.G., S.G., P.D.B., G.S., M.C.C., V.G., F.A. and A.M.; data curation, P.C., P.S., P.L., S.D.L., M.D.G., S.G., P.D.B., G.S., M.C.C., V.G., F.A. and A.M.; writing—original draft preparation, P.C., F.P., E.M.S., E.A., P.S., P.L., S.D.L., M.D.G., S.G., P.D.B., M.C.C., V.G., F.A., S.D., P.D.C. and A.M.; writing—review and editing, P.C., F.P., E.M.S., E.A., C.A., S.D., P.D.C. and A.M.; supervision, P.C., F.P., C.A., S.D., P.D.C. and A.M.; All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
Meteorological data belong to “Centro Funzionale e Ufficio Idrologia, Idrografico, Mareografico - Agenzia di Protezione Civile della Regione Abruzzo”. Data relating to laboratory tests can be requested directly from the authors.
Acknowledgments
The authors would like to acknowledge the cooperation and availability of the Maiella National Park President, Prof. Lucio Zazzera, the Maiella National Park Director, Dott. Luciano Di Martino, and President of the Castel di Sangro Section of the Alpine Club, Franco D’Ambrosio. Alessandra Mascitelli, Piero Chiacchiaretta, Eleonora Aruffo and Piero Di Carlo acknowledge financial support funded by the European Union NextGenerationEU, under the National Recovery and Resilience Plan (NRRP), Mission 4 Component 2-M4C2, Investment 1.5 Call for tender No. 3277 of 30 December 2021, Italian Ministry of University, Award Number: ECS00000041, Project Title: “Innovation, digitalization, and sustainability for the diffused economy in Central Italy”, Concession Degree No. 1057 of 23 June 2022 adopted by the Italian Ministry of University. CUP: D73C22000840006.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Palmerini, G. L’arte rupestre nel Parco Nazionale della Maiella. Ricerche storiche, indagini in corso e nuove prospettive; Ente Parco Nazionale della Maiella, 2023.
- Mattioli, T. L’arte rupestre in Italia centrale: Umbria, Lazio, Abruzzo; Vol. 4, Tommaso Mattioli, 2007.
- Palmerini, G.; Beck, L.; Di Martino, L.; Gallet, X.; Lebon, M.; Manzi, A.; Nicoud, E.; Mariano, A.; Villa, V. # MaiellaRockArtProject: Nuove ricerche sull’arte rupestre dell’Appennino abruzzese. In Proceedings of the XXVIII Valcamonica Symposium: ROCK-ART, A HUMAN HERITAGE. Centro Camuno di Studi Preistorici, 2021.
- Di Fraia, T. Le nuove scoperte di arte rupestre in Abruzzo: verso un’interpretazione sistemica. L’RTE RUPESTRE, 2015. [Google Scholar]
- Mattioli, T. Landscape analysis of a sample of rock-art sites in Central Italy. In Proceedings of the Layers of perception: Proceedings of the 35th International Conference on Computer Applications and Quantitative Methods in Archaeology (CAA), 2008, pp. 342–43.
- Nuhoglu, Y.; Oguz, E.; Uslu, H.; Ozbek, A.; Ipekoglu, B.; Ocak, I.; Hasenekoglu, I. The accelerating effects of the microorganisms on biodeterioration of stone monuments under air pollution and continental-cold climatic conditions in Erzurum, Turkey. Science of the total environment 2006, 364, 272–283. [Google Scholar] [CrossRef]
- Kumar, R.; Kumar, A.V. Biodeterioration of stone in tropical environments. An overview. The Getty Conservation Institute, Los Angeles, Calif, 1999. [Google Scholar]
- Ortega-Morales, B.O.; Gaylarde, C.C.; Englert, G.E.; Gaylarde, P.M. Analysis of salt-containing biofilms on limestone buildings of the Mayan culture at Edzna, Mexico. Geomicrobiology Journal 2005, 22, 261–268. [Google Scholar] [CrossRef]
- Tomaselli, L.; Lamenti, G.; Bosco, M.; Tiano, P. Biodiversity of photosynthetic micro-organisms dwelling on stone monuments. International Biodeterioration & Biodegradation 2000, 46, 251–258. [Google Scholar] [CrossRef]
- Gaylarde, P.M.; Gaylarde, C.C. Algae and cyanobacteria on painted buildings in Latin America. International Biodeterioration & Biodegradation 2000, 46, 93–97. [Google Scholar] [CrossRef]
- Crispim, C.A.; Gaylarde, C. Cyanobacteria and biodeterioration of cultural heritage: a review. Microbial ecology 2005, 49, 1–9. [Google Scholar] [CrossRef]
- Zanardini, E.; Abbruscato, P.; Ghedini, N.; Realini, M.; Sorlini, C. Influence of atmospheric pollutants on the biodeterioration of stone. International biodeterioration & biodegradation 2000, 45, 35–42. [Google Scholar]
- Warscheid, T.; Braams, J. Biodeterioration of stone: a review. International Biodeterioration & Biodegradation 2000, 46, 343–368. [Google Scholar] [CrossRef]
- Scheerer, S.; Ortega-Morales, O.; Gaylarde, C. Microbial deterioration of stone monuments—an updated overview. Advances in applied microbiology 2009, 66, 97–139. [Google Scholar] [PubMed]
- Ortega-Morales, O.; Montero-Muñoz, J.L.; Neto, J.A.B.; Beech, I.B.; Sunner, J.; Gaylarde, C. Deterioration and microbial colonization of cultural heritage stone buildings in polluted and unpolluted tropical and subtropical climates: A meta-analysis. International Biodeterioration & Biodegradation 2019, 143, 104734. [Google Scholar] [CrossRef]
- Louati, M.; Ennis, N.J.; Ghodhbane-Gtari, F.; Hezbri, K.; Sevigny, J.L.; Fahnestock, M.F.; Cherif-Silini, H.; Bryce, J.G.; Tisa, L.S.; Gtari, M. Elucidating the ecological networks in stone-dwelling microbiomes. Environmental microbiology 2020, 22, 1467–1480. [Google Scholar] [CrossRef]
- Sesana, E.; Gagnon, A.S.; Ciantelli, C.; Cassar, J.; Hughes, J.J. Climate change impacts on cultural heritage: A literature review. Wiley Interdisciplinary Reviews: Climate Change 2021, 12, e710. [Google Scholar] [CrossRef]
- Gu, J.D.; Katayama, Y. Microbiota and biochemical processes involved in biodeterioration of cultural heritage and protection. Microorganisms in the deterioration and preservation of Cultural Heritage 2021, 37, 37–58. [Google Scholar]
- Macedo, M.F.; Miller, A.Z.; Dionísio, A.; Saiz-Jimenez, C. Biodiversity of cyanobacteria and green algae on monuments in the Mediterranean Basin: an overview. Microbiology 2009, 155, 3476–3490. [Google Scholar] [CrossRef]
- Urzì, C.; Realini, M. Colour changes of Notos calcareous sandstone as related to its colonisation by microorganisms. International Biodeterioration & Biodegradation 1998, 42, 45–54. [Google Scholar] [CrossRef]
- Gambino, M.; Sanmartín, P.; Longoni, M.; Villa, F.; Mitchell, R.; Cappitelli, F. Surface colour: An overlooked aspect in the study of cyanobacterial biofilm formation. Science of the Total Environment 2019, 659, 342–353. [Google Scholar] [CrossRef]
- Ortega-Morales, O.; Guezennec, J.; Hernandez-Duque, G.; Gaylarde, C.C.; Gaylarde, P.M. Phototrophic biofilms on ancient Mayan buildings in Yucatan, Mexico. Current Microbiology 2000, 40, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Gorbushina, A.A. Life on the rocks. Environmental microbiology 2007, 9, 1613–1631. [Google Scholar] [CrossRef] [PubMed]
- Guillitte, O. Bioreceptivity: a new concept for building ecology studies. Science of the total environment 1995, 167, 215–220. [Google Scholar] [CrossRef]
- Sanmartín, P.; Miller, A.; Prieto, B.; Viles, H.A. Revisiting and reanalysing the concept of bioreceptivity 25 years on. Science of the total environment 2021, 770, 145314. [Google Scholar] [CrossRef]
- Miller, A.; Sanmartín, P.; Pereira-Pardo, L.; Dionísio, A.; Sáiz-Jiménez, C.; Macedo, M.; Prieto, B. Bioreceptivity of building stones: A review. Science of the total environment 2012, 426, 1–12. [Google Scholar] [CrossRef]
- Viles, H.; Ahmad, H. Architectural controls on the bioreceptivity of sandstone to green algal colonization. ECBSM2016. In Proceedings of the European Conference on Biodeterioration of Stone Monuments-Second Edition. Cergy-Pontoise, France. November 17–18, 2016, 2016.
- Cattò, C.; Mu, A.; Moreau, J.W.; Wang, N.; Cappitelli, F.; Strugnell, R. Biofilm colonization of stone materials from an Australian outdoor sculpture: importance of geometry and exposure. Journal of Environmental Management 2023, 339, 117948. [Google Scholar] [CrossRef]
- Trovão, J.; Portugal, A. The impact of stone position and location on the microbiome of a marble statue. The Microbe 2024, 2, 100040. [Google Scholar] [CrossRef]
- Ohkubo, S.; Miyashita, H. A niche for cyanobacteria producing chlorophyll f within a microbial mat. The ISME Journal 2017, 11, 2368–2378. [Google Scholar] [CrossRef]
- Involvement of Cyanobacterial Phytochromes in Growth Under Different Light Qualitities and Quantities¶.
- Liu, X.; Qian, Y.; Wu, F.; Wang, Y.; Wang, W.; Gu, J.D. Biofilms on stone monuments: biodeterioration or bioprotection? Trends in Microbiology 2022, 30, 816–819. [Google Scholar] [CrossRef]
- Favero-Longo, S.E.; Viles, H.A. A review of the nature, role and control of lithobionts on stone cultural heritage: weighing-up and managing biodeterioration and bioprotection. World Journal of Microbiology and Biotechnology 2020, 36, 100. [Google Scholar] [CrossRef]
- Pinna, D. Coping with biological growth on stone heritage objects: methods, products, applications, and perspectives; Apple academic press, 2017.
- Cappitelli, F.; Cattò, C.; Villa, F. The control of cultural heritage microbial deterioration. Microorganisms 2020, 8, 1542. [Google Scholar] [CrossRef]
- Macchia, A.; Aureli, H.; Prestileo, F.; Ortenzi, F.; Sellathurai, S.; Docci, A.; Cerafogli, E.; Colasanti, I.A.; Ricca, M.; La Russa, M.F. In-situ comparative study of eucalyptus, basil, cloves, thyme, pine tree, and tea tree essential oil biocide efficacy. Methods and Protocols 2022, 5, 37. [Google Scholar] [CrossRef]
- Decho, A. W. a. Microbial indicators of environmental change, 2010.
- C. Gaylarde, C. Influence of environment on microbial colonization of historic stone buildings with emphasis on cyanobacteria. Heritage 2020, 3, 1469–1482. [Google Scholar] [CrossRef]
- May, E.; Papida, S.; Abdulla, H.; Tayler, S.; Dewedar, A. Comparative studies of microbial communities on stone monuments in temperate and semi-arid climates. In Of microbes and art: the role of microbial communities in the degradation and protection of cultural heritage; Springer, 2000; pp. 49–62.
- Perez-Monserrat, E.M.; Varas-Muriel, M.J.; Alvarez De Buergo, M.; Fort, R. Black layers of decay and color patterns on heritage limestone as markers of environmental change. Geosciences 2016, 6, 4. [Google Scholar] [CrossRef]
- Chen, X.; Bai, F.; Huang, J.; Lu, Y.; Wu, Y.; Yu, J.; Bai, S. The organisms on rock cultural heritages: growth and weathering. Geoheritage 2021, 13, 56. [Google Scholar] [CrossRef]
- Ding, X.; Lan, W.; Yan, A.; Li, Y.; Katayama, Y.; Gu, J.D. Microbiome characteristics and the key biochemical reactions identified on stone world cultural heritage under different climate conditions. Journal of Environmental Management 2022, 302, 114041. [Google Scholar] [CrossRef]
- Wang, C.; Wang, L.; Bai, C.; Wang, M.; Ma, T.; Ma, H.; Zhang, G.; Wang, W.; Guo, Z.; Sun, Y.; et al. Identification of the key factors influencing biodeterioration of open-air cultural heritage in the temperate climate zone of China. International Biodeterioration & Biodegradation 2025, 196, 105954. [Google Scholar]
- Pointing, S. B. a. Microbial growth patterns linked to climatic factors, 2009.
- Viles, H.A.; Cutler, N.A. Global environmental change and the biology of heritage structures. Global Change Biology 2012, 18, 2406–2418. [Google Scholar] [CrossRef]
- Traversetti, L.; Bartoli, F.; Caneva, G. Wind-driven rain as a bioclimatic factor affecting the biological colonization at the archaeological site of Pompeii, Italy. International biodeterioration & biodegradation 2018, 134, 31–38. [Google Scholar]
- Orr, S.A.; Cassar, M. Exposure indices of extreme wind-driven rain events for built heritage. Atmosphere 2020, 11, 163. [Google Scholar] [CrossRef]
- Albertano, P. Cyanobacterial biofilms in monuments and caves. In Ecology of cyanobacteria II: their diversity in space and time; Springer, 2012; pp. 317–343.
- Chiaudani, A.; Flamminii, F.; Consalvo, A.; Bellocci, M.; Pizzi, A.; Passamonti, C.; Cichelli, A. Rare Earth Element Variability in Italian Extra Virgin Olive Oils from Abruzzo Region. Foods 2024, 13. [Google Scholar] [CrossRef]
- Potenza, F.; Cufaro, M.C.; Di Biase, L.; Panella, V.; Di Campli, A.; Ruggieri, A.G.; Dufrusine, B.; Restelli, E.; Pietrangelo, L.; Protasi, F.; et al. Proteomic analysis of marinesco–sjogren syndrome fibroblasts indicates pro-survival metabolic adaptation to SIL1 loss. International Journal of Molecular Sciences 2021, 22, 12449. [Google Scholar] [CrossRef]
- Ucci, A.; Giacchi, L.; Cufaro, M.C.; Puri, C.; Ciocca, M.; Di Ferdinando, F.; Del Boccio, P.; Cappariello, A.; Rucci, N. Human osteosarcoma cell secretome impairs neonatal mouse calvarial osteogenic cells functions and modifies the nanoparticles-derived protein profile. Life Sciences 2025, p. 123837.
Figure 1.
(a) Site location in Lama dei Peligni, Abruzzo Region - Central Italy (b) Site location (two red markers) in relation to the town center of Lama dei Peligni (red circle marker) and route to reach the site of interest (black line).
Figure 1.
(a) Site location in Lama dei Peligni, Abruzzo Region - Central Italy (b) Site location (two red markers) in relation to the town center of Lama dei Peligni (red circle marker) and route to reach the site of interest (black line).
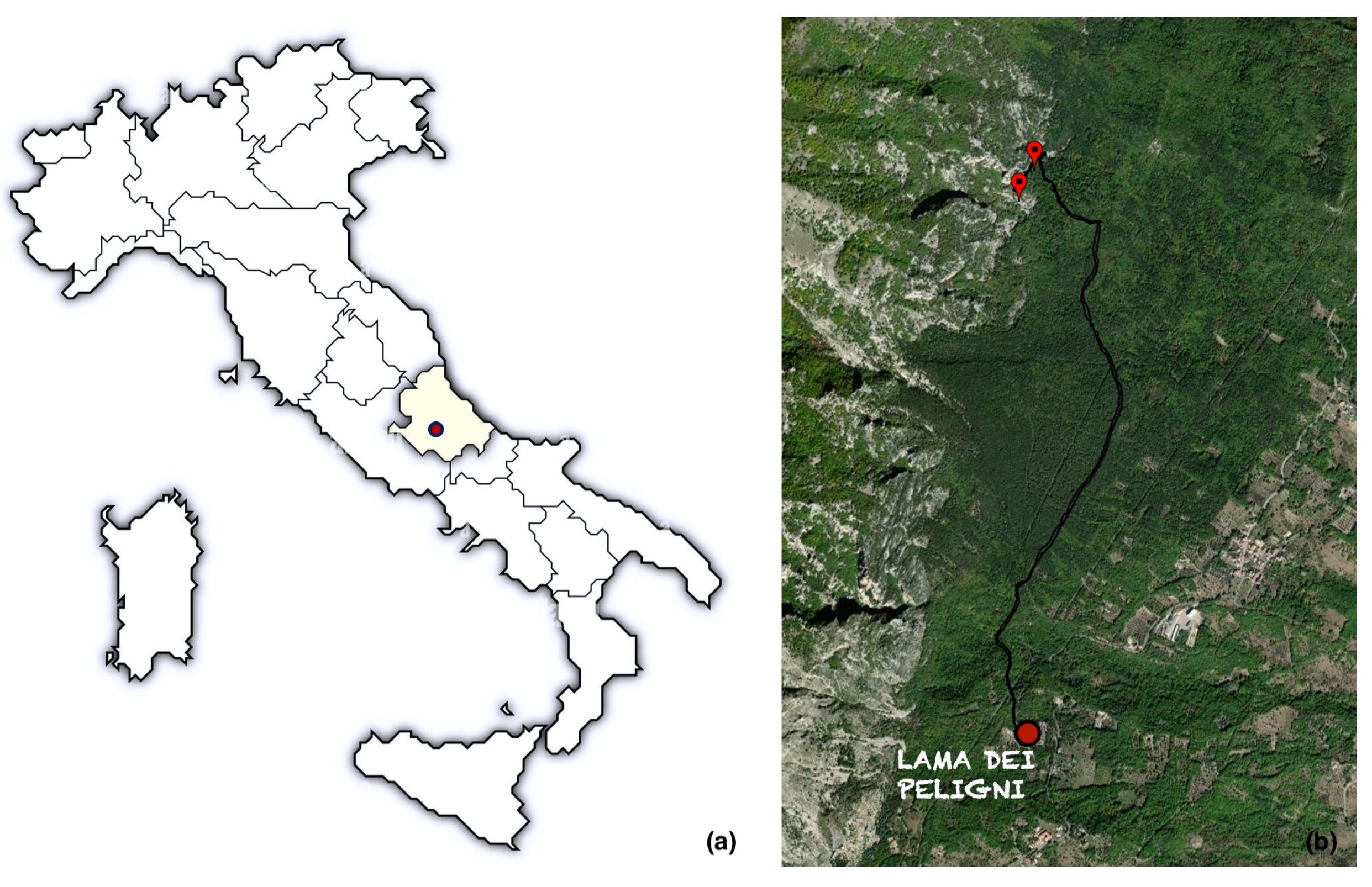
Figure 2.
(a) One of red anthropomorphic motifs paintings located in the site of interest (b) Black biofilm which alters the appearance of surfaces in some areas (photos by A. Mascitelli).
Figure 2.
(a) One of red anthropomorphic motifs paintings located in the site of interest (b) Black biofilm which alters the appearance of surfaces in some areas (photos by A. Mascitelli).

Figure 3.
(a) One of red anthropomorphic motifs paintings close to a black stripe (b) Sample collection (c-d-e) Views of the area (photos by A. Mascitelli).
Figure 3.
(a) One of red anthropomorphic motifs paintings close to a black stripe (b) Sample collection (c-d-e) Views of the area (photos by A. Mascitelli).

Figure 4.
Palena and Lama dei Peligni site stations(light-blue markers) and the distance between them (yellow line ≈ 7.5 km).
Figure 4.
Palena and Lama dei Peligni site stations(light-blue markers) and the distance between them (yellow line ≈ 7.5 km).
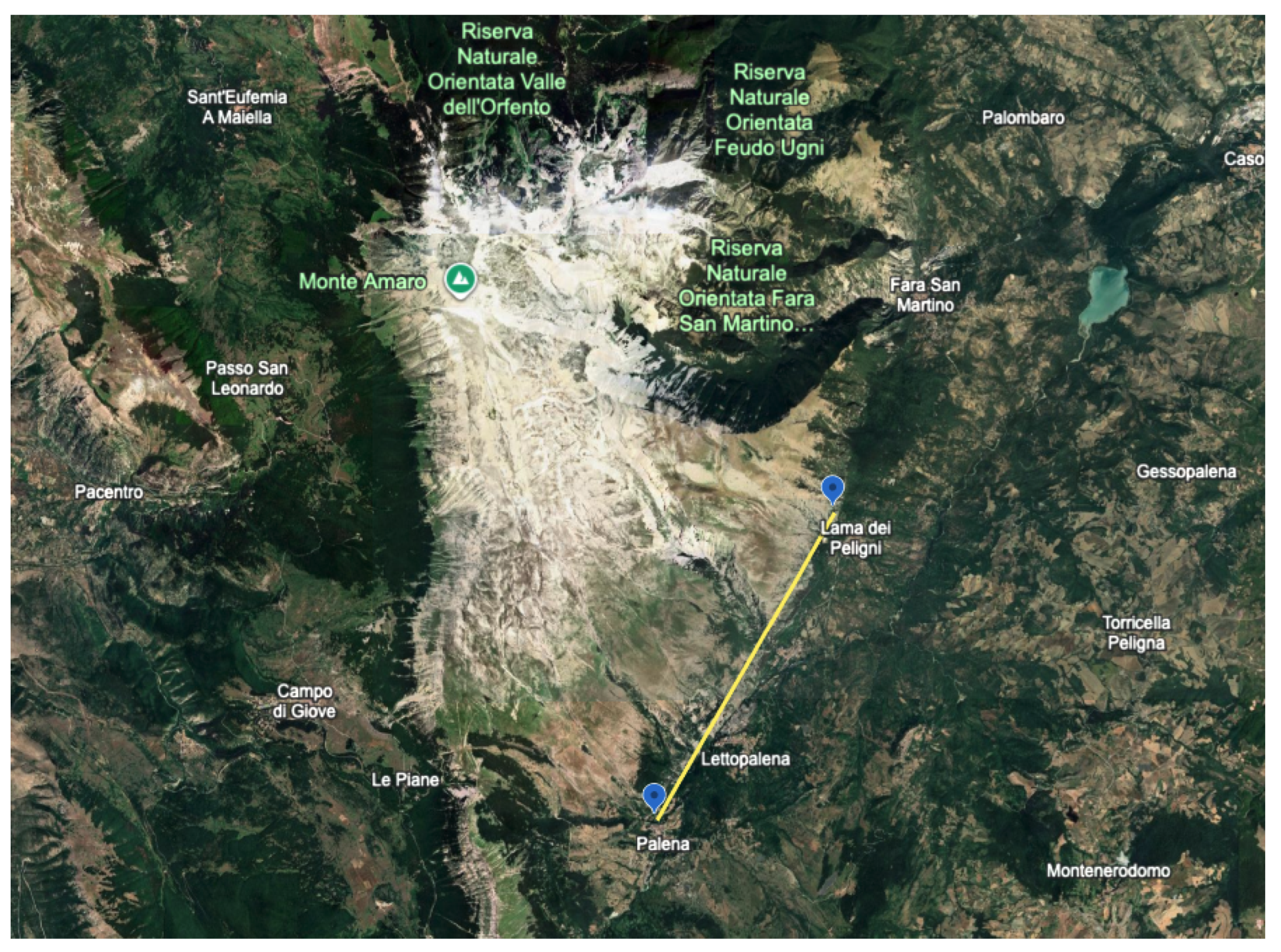
Figure 5.
Identification of microbial populations from rock painting samples by flow cytometry. Microorganisms resuspended in 1X PBS from rock painting swabs were analyzed by flow cytometry. (a) A distinct population was identified in a side scatter height (SSC-H) vs a forward scatter height (FSC-H) dot plot. (b) The whole identified population was analyzed for SYTO9 staining. The blue peak represents the SYTO9-stained population, while the red peak corresponds to the related fluorescence minus one (FMO) control. (c) The identified SYTO9+ population of cells was represented on a SSC-H vs Alexa Fluor 647-H contour plot. The Alexa Fluor 647 channel (660/20 nm) is the best suited for detecting cyanobacteria-specific fluorescence (emission peak around 650–660 nm). The same representation is shown bot for was analyzed for both a sample collected from a control area of the painting (not covered by biofilm), used as a negative control, and (d) the population of SYTO9+ cells obtained from a biofilm-covered region of the painting. (e) 1X PBS alone was analyzed as an additional negative control to exclude background on scatter signals. Data are representative of three independent experiments.
Figure 5.
Identification of microbial populations from rock painting samples by flow cytometry. Microorganisms resuspended in 1X PBS from rock painting swabs were analyzed by flow cytometry. (a) A distinct population was identified in a side scatter height (SSC-H) vs a forward scatter height (FSC-H) dot plot. (b) The whole identified population was analyzed for SYTO9 staining. The blue peak represents the SYTO9-stained population, while the red peak corresponds to the related fluorescence minus one (FMO) control. (c) The identified SYTO9+ population of cells was represented on a SSC-H vs Alexa Fluor 647-H contour plot. The Alexa Fluor 647 channel (660/20 nm) is the best suited for detecting cyanobacteria-specific fluorescence (emission peak around 650–660 nm). The same representation is shown bot for was analyzed for both a sample collected from a control area of the painting (not covered by biofilm), used as a negative control, and (d) the population of SYTO9+ cells obtained from a biofilm-covered region of the painting. (e) 1X PBS alone was analyzed as an additional negative control to exclude background on scatter signals. Data are representative of three independent experiments.
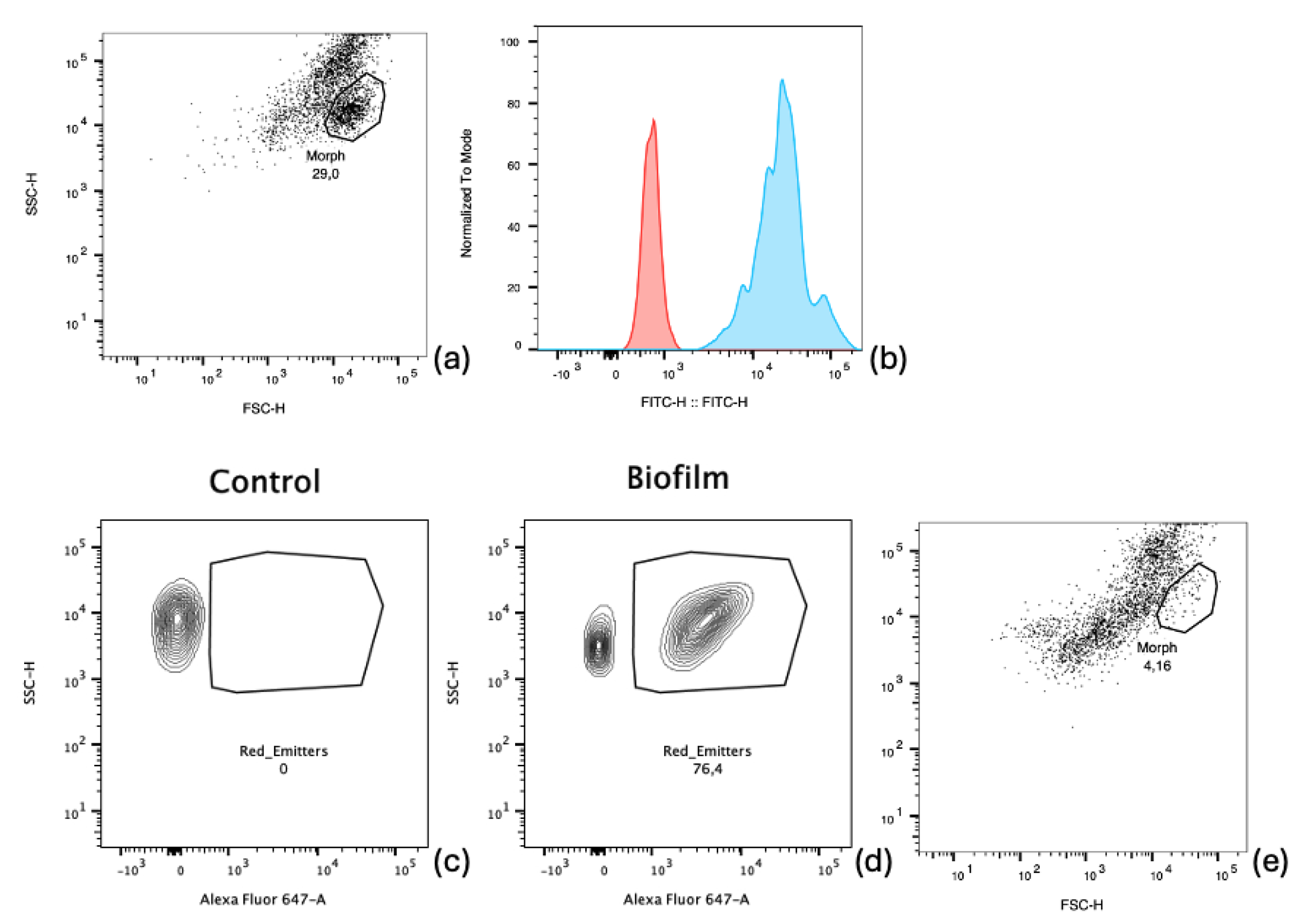
Figure 6.
Output referred to sample 1.
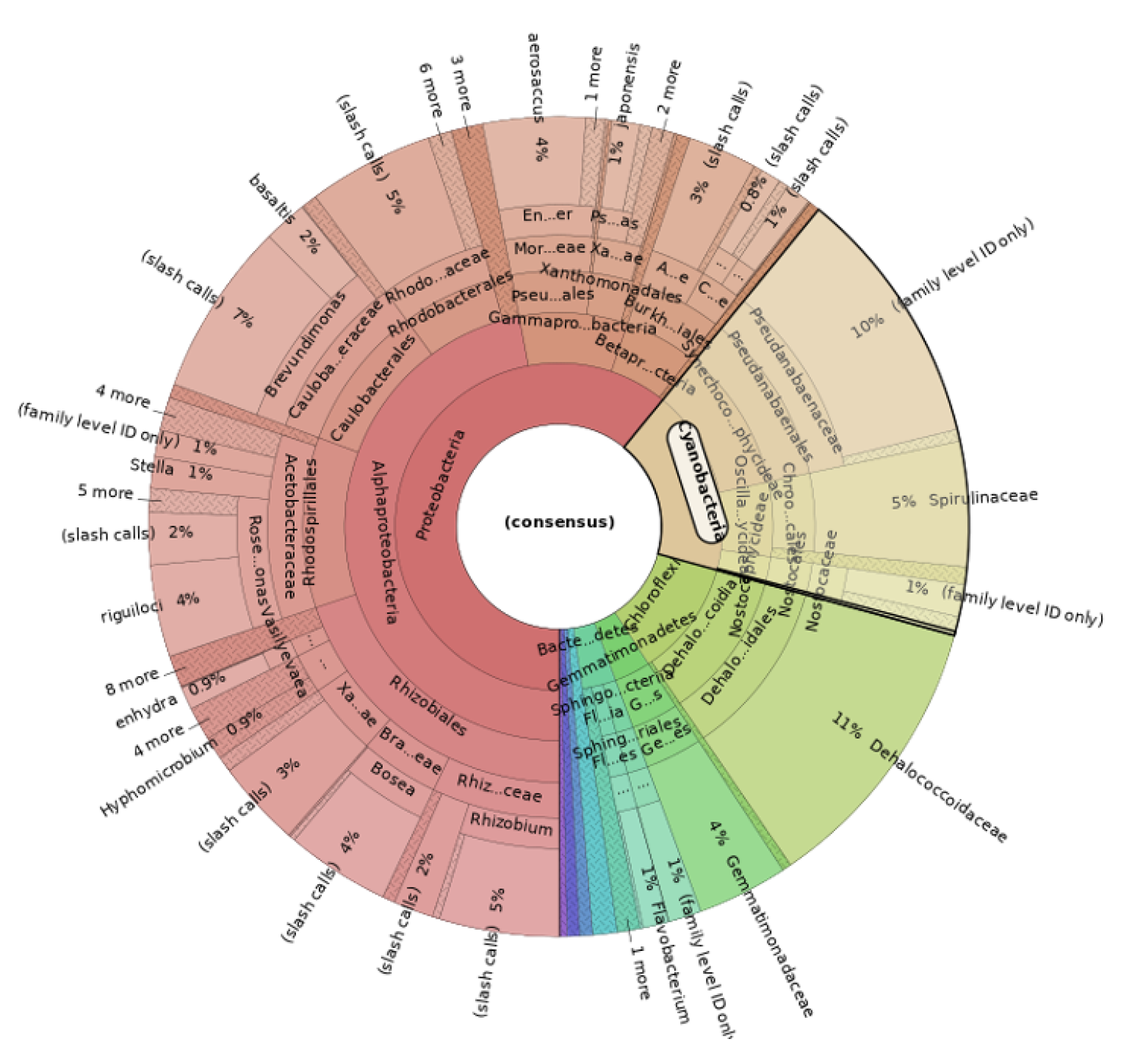
Figure 7.
Output referred to sample 2.
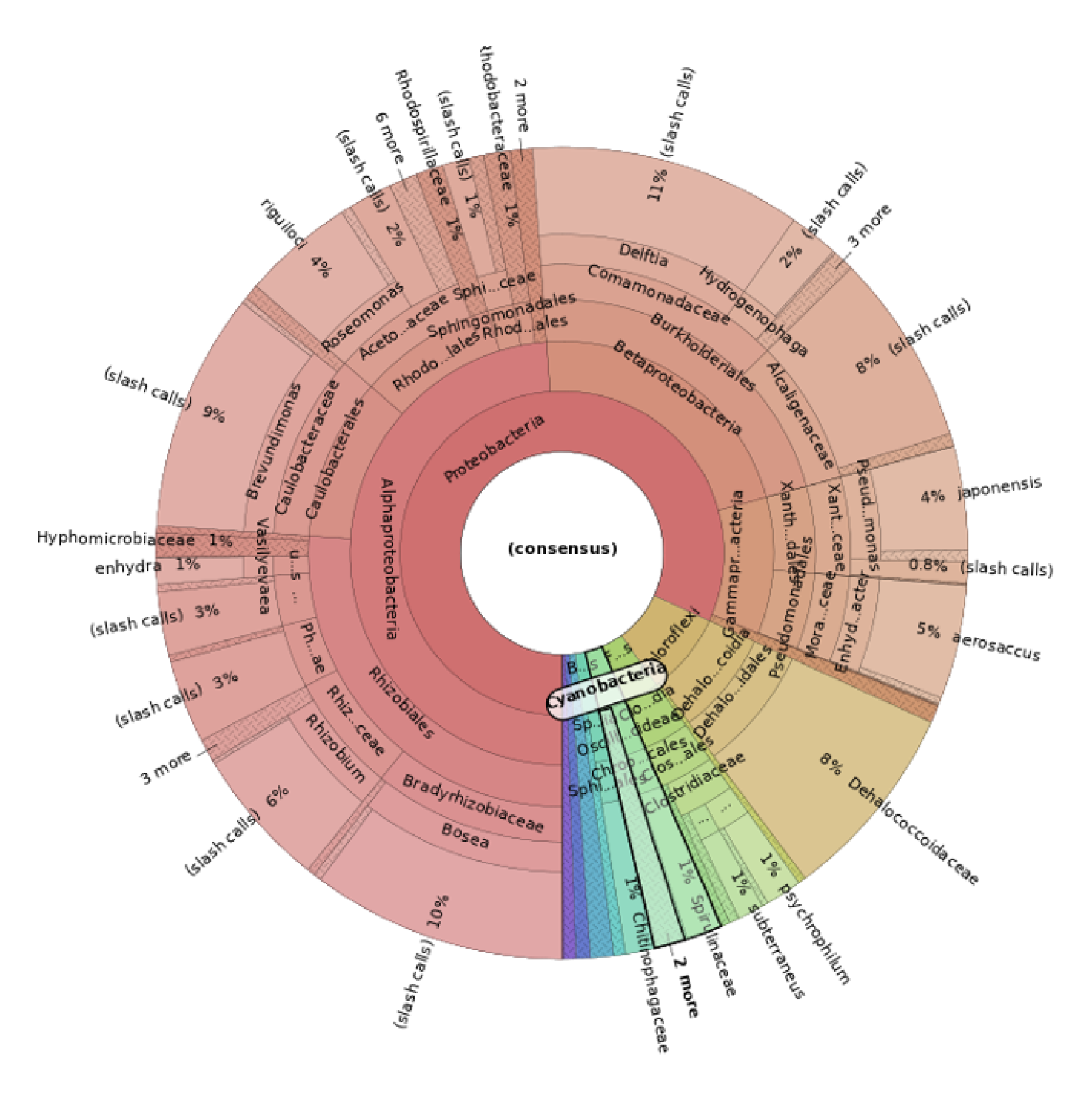
Figure 8.
BioVenn diagram showing the percentage of protein identified through Cyanobacteriota/Melainabacteria group (in green) related to the various species derived from the ’All Entries’ database of UniProtKB/SwissProt.
Figure 8.
BioVenn diagram showing the percentage of protein identified through Cyanobacteriota/Melainabacteria group (in green) related to the various species derived from the ’All Entries’ database of UniProtKB/SwissProt.
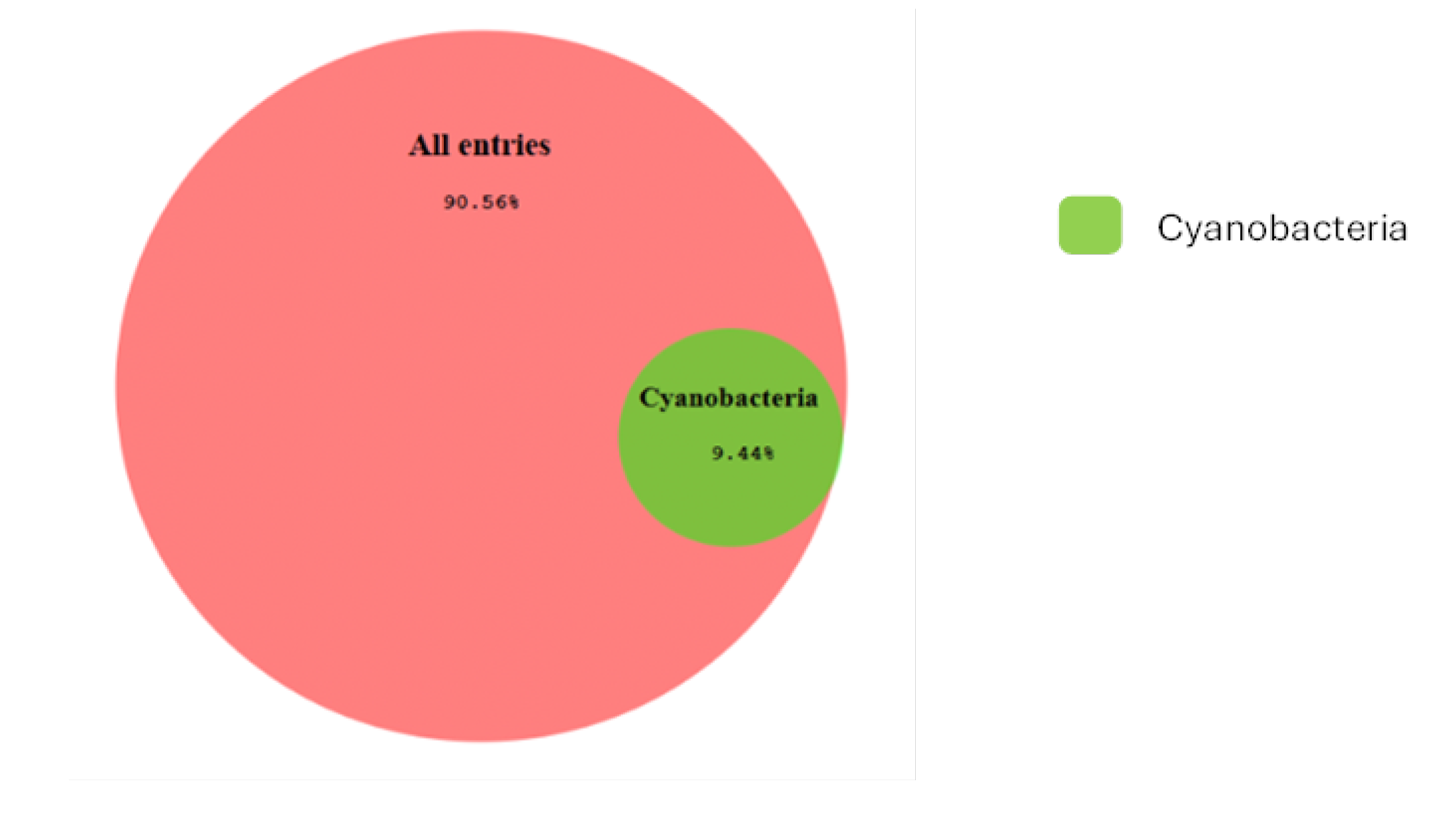
Figure 9.
(a) Network GO:0046933 of “Proton-transporting ATP synthase activity, rotational mechanism” (FDR=8.22x10−11) (b) Network GO:0015979 of “Photosynthesis” network (FDR=1.2x10−4).
Figure 9.
(a) Network GO:0046933 of “Proton-transporting ATP synthase activity, rotational mechanism” (FDR=8.22x10−11) (b) Network GO:0015979 of “Photosynthesis” network (FDR=1.2x10−4).
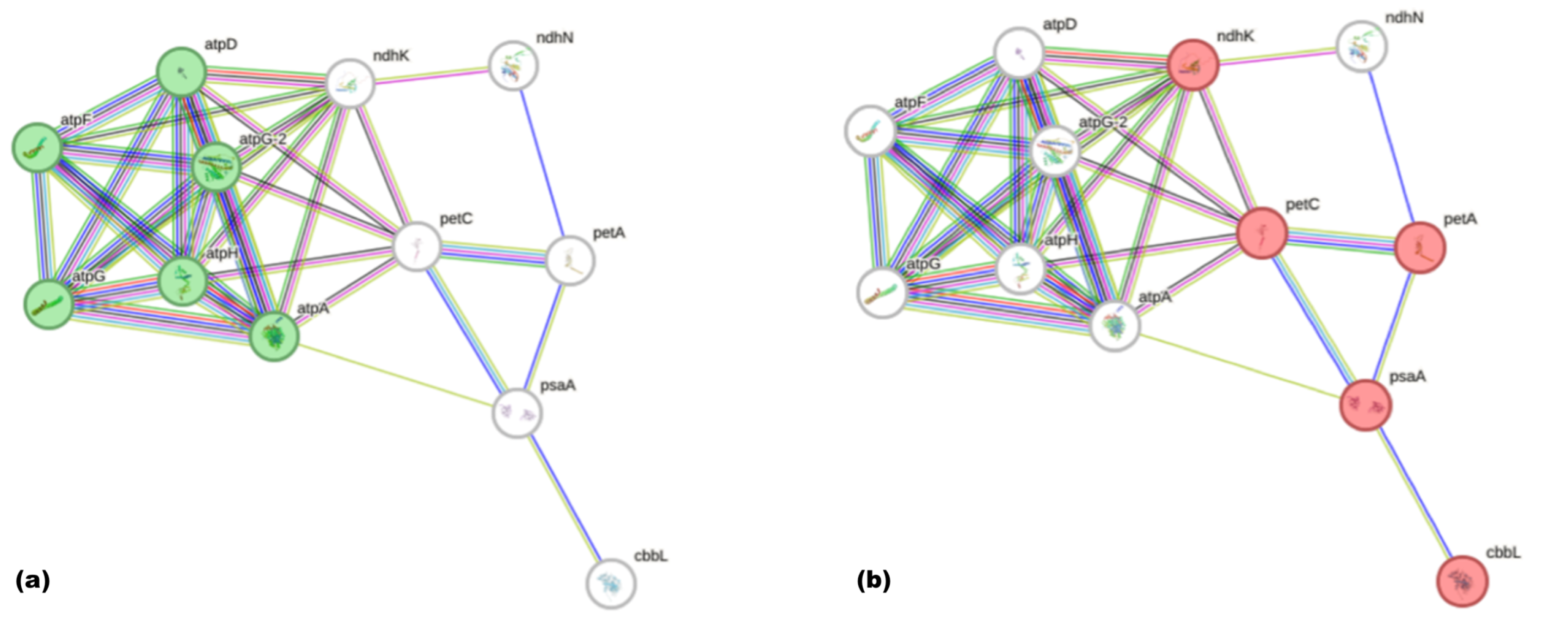

Table 1.
List of ICP-MS instrumentation parameters.
| Instrument parameters | |
|---|---|
| Nebulizer | Babington |
| Torch | Quartz glass torch |
| Spray chamber | Scott double-pass type at 2 ∘C |
| Sample cone Nickel | 1.00 mm aperture |
| Skimmer cone Nickel | 0.40 mm aperture |
| Plasma mode | Normal plasma |
| RF power (W) | 1550 |
| RF matching (V) | 1.8 |
| Sample depth (mm) | 10 |
| Nebulizer gas (L ) | 1.03 |
| Nebulizer pump (rps) | 0.1 |
| Plasma gas (L ) | 15 |
| Sampling period (s) | 0.3 |
| Repetitions 3 | 3 |
| Sample uptake rate (mL ) | 0.4 |
| Integration time (s) | 0.1 |
Table 2.
Elemental composition of the samples (ng mL-1 ± relative standard deviation (n = 3). In the table the main differences between blackened moss and only moss are highlighted in bold).* Values fall outside the calibration curve used. ** Values are below the limit of quantification.
Table 2.
Elemental composition of the samples (ng mL-1 ± relative standard deviation (n = 3). In the table the main differences between blackened moss and only moss are highlighted in bold).* Values fall outside the calibration curve used. ** Values are below the limit of quantification.
| MOSS | BLACKENED MOSS | SOIL | ROCK SURFACE |
|---|---|---|---|
| Co = 6.6 ± 0.2 | Co = 2.9 ± 1 | Co = 12.9 ± 5.7 | Co = 3.8 ± 2.7 |
| Cr = 2.2 ± 2.9 | Cr = 5.6 ± 2.2 | Cr = 1.5 ± 6.5 | Cr = 11.7 ± 4.7 |
| Al = 713.3*± 8.4 | Al = 3329.3*± 0.7 | Al = 435.2*± 1.7 | Al = 1532.3*± 2.4 |
| Ag = N/A** | Ag = 0.23 ± 4.7 | Ag = N/A** | Ag = N/A** |
| As = 4.4 ± 5.1 | As = 2.01 ± 1.9 | As = 4.8 ± 0.6 | As = 12.6 ± 4.2 |
| Cd = 0.13 ± 3.4 | Cd = 5.4 ± 1.7 | Cd = 0.18 ± 10.9 | Cd = 624.2* ± 1.3 |
| Cu = 16.9 ± 6.7 | Cu = 19.3 ± 2.3 | Cu = 16.2 ± 1 | Cu = 42.2 ± 3.1 |
| Mn = 25.2 ± 7.2 | Mn = 115.5* ± 0.6 | Mn = 17.7 ± 1.7 | Mn = 188.4* ± 2.7 |
| Ni = 56.3 ± 0.1 | Ni = 12.9 ± 0.9 | Ni = 107 ± 6.5 | Ni = 41.4 ± 8.7 |
| Pb = 1.9 ± 8.6 | Pb = 255.6 ± 4.9 | Pb = 1.4 ± 1 | Pb = 24.8 ± 6.9 |
| Se = 1.8 ± 13.4 | Se = 11.5 ± 7.2 | Se = 3.2 ± 16 | Se = 18 ± 6.9 |
| V = 11.7 ± 7 | V = 6.9 ± 0.8 | V = 12.1 ± 0.5 | V = 23.31 ± 1.6 |
| Zn = 17.9 ± 7.1 | Zn = 54.7 ± 1.7 | Zn = 22.2 ± 1.1 | Zn = 82.9 ± 3.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



