
Submitted:
25 October 2025
Posted:
27 October 2025
You are already at the latest version
Abstract
This study aims to elucidate the physiological and biochemical alterations induced by parasitic Cuscuta sp. (dodder) in lucerne (Medicago sativa L.), a key forage crop. Comparative analyses between infected and healthy plants revealed that significant reductions in chlorophyll a, b and total chlorophyll, and protein levels in the leaf and stem tissues of Cuscuta-infested plants were evident. The parasitic infection led to increased activities in antioxidant enzymes such as catalase (CAT) and peroxidase (POX) in stems, but not in leaves. Phenolic compounds were significantly lower both in leaves and stems of dodder-infected lucerne plants. No statistically significant changes were detected in jasmonic acid (JA) and salicylic acid (SA) levels in both plant parts, suggesting that classical defense signaling pathways may not be predominantly activated under Cuscuta-mediated stress. Possibly, host defense might be impaired. Histological examinations demonstrated active structural defense responses including localized tissue remodeling and the formation of callose-like structures at haustorial penetration sites. DNA fragmentations showed that Cuscuta-infected M. sativa plants exhibited slightly higher instability. Collectively, these findings provide novel insights into the molecular and biochemical basis of the Cuscuta-lucerne interactions and highlight the need for further investigation into host defense mechanisms. The results also offer a valuable foundation for the development of Cuscuta-resistant lucerne cultivars and support the design of integrated, sustainable weed management strategies to mitigate the detrimental effects of parasitic plants on forage production systems.
Keywords:
Cuscuta sp.
; Medicago sativa
; Parasitic weed
; Oxidative stress
; host-parasite interaction
1. Introduction
Lucerne (Medicago sativa L.), a perennial leguminous forage crop of the Fabaceae family, is recognized as one of the most widely cultivated and nutritionally valuable plant species in global agriculture. Its high dry matter yield, rich protein content, and superior digestibility make it a cornerstone in ruminant nutrition. Beyond its forage quality, lucerne contributes significantly to agroecosystem sustainability through its extensive root system, which enhances soil structure by improving aeration, reducing compaction, and limiting erosion. Additionally, it can fix atmospheric nitrogen via symbiosis with Rhizobium spp., and it enriches soil fertility and reduces the need for synthetic fertilizers [1]. Despite these agronomic and ecological advantages, lucerne cultivation is highly susceptible to a range of abiotic (e.g. drought, salinity, temperature extremes) and biotic (e.g. pathogens, pests, parasitic plants) stress factors. Among biotic threats, species of Cuscuta sp. (dodder) have recently gained attention due to their aggressive parasitic nature and destructive impact on plant health and productivity [2,3]. Members of the dodder genus, belonging to the Convolvulaceae family, are obligatory holoparasitic plants devoid of chlorophyll and photosynthetic pigments. They are entirely reliant on host plants for water, nutrients, and organic metabolites, and they absorb materials through specialized invasive structures known as haustoria [4]. The economic loss caused by holophyte parasitic weeds on crop plants could be expressed in hundreds of millions of US Dollars around the globe, and this figure is enormously increasing every year [5].
The establishment of haustorial connections by Cuscuta sp. leads to severe physiological and biochemical disruptions in the host plant. These include the depletion of vital resources, suppression of photosynthesis, and ultimately, significant reductions in biomass. Parasitic plants use a special organ called haustorium to attach to host plants, and extract water and nutrients from the host plants. Cuscuta spp. are rootless and leafless holoparasitic plants on crop plants. They have vine-like stems which twine around the host stem and selectively take nutrients and genes from the host. Infection strategies of parasitic plants are quite similar to microbial plant pathogens. Therefore, they must interact closely with host plants to extract nutrients and suppress defense responses. Although plants possess large arsenals of immune receptors capable of recognizing all pathogen classes, however, invading pathogens, including holoparasites, suppress or evade immune perception and drive nutrients from host tissues.
Dodder infection markedly lowers chlorophyll a and b levels, reduces total protein content, and alters antioxidant defense pathways by increasing the activity of key enzymes such as catalase (CAT) and peroxidase (POX) as well as inducing phenolic compound biosynthesis [6,7]. These responses are believed to reflect the defense capacity of host plants against the oxidative stress triggered by parasitic invasion. In addition to biochemical responses, structural defense mechanisms have also been documented, including localized tissue remodeling and the formation of callose-like structures at infection sites—suggesting a targeted response to haustorial penetration [8]. The cumulative effects of such infections lead to stunted growth, reduced forage quality, and diminished yield potential, posing a significant threat to the economic viability of lucerne-based forage systems [9,10]. If Cuscuta sp. damage reaches DNA molecule, the recovery of host plants might be extremely difficult since all metabolic activities are coded there. Therefore, any damage and instability in DNA molecules affect the whole metabolism.
Given the increasing prevalence of dodder infestations and their substantial agronomic impact, there is a critical need to better understand the dodder–lucerne interactions at molecular, physiological, and biochemical levels. In this study, comparative analyses of healthy and infected lucerne plants were conducted to evaluate the physiological and biochemical consequences of parasitic stress. Insights gained from such studies are essential for the development of resistant lucerne cultivars and will play a vital role in designing innovative and sustainable crop management strategies in the face of parasitic weed challenges.
2. Materials and Methods
2.1. Plant Materials and Sampling Procedure
Lucerne (Medicago sativa L. cv. Alsancak) seeds were sown in an experimental field plot (3 m x 3 m) in Harran University, Şanlıurfa, Türkiye. Cuscuta sp. seeds were artificially inoculated within the roots of lucerne by mixing with soil. Within weeks, stem colonization of lucerne by dodder was observed. The lucerne plant samples were first collected at the 90th day, and followed by 15-day intervals, for the growth parameters from the dodder-infested and the control plots. All samples were carefully labeled in detail, including sampling location, date, and observed symptoms and the leaf and stem samples were stored at −20°C until analysis. Height, fresh and dry weights of the plants were recorded at each sampling time. For the biochemical analysis, the samples were collected at the final harvest stage.
2.2. Yield and Growth Measurements
Height measurements (cm) of lucerne plants were made on dodder-infested and non-infested plots by measuring the height from the soil surface to the plant apex. To determine green forage yield, plants were harvested from 10 different locations, and their fresh weights (Fwt) were recorded. The same samples were then oven-dried at 70°C for 72 hours, and dry forage yield (Dwt) was determined thereafter. Yield values from infested and non-infested plots were recorded and converted to yield da-1.
2.3. Biochemical Analysis
Biochemical analyses were performed on Cuscuta sp.-infected and non-infected plants. To minimize standard errors arising from biological variability among samples, a single representative plant was selected from each experimental group. This approach not only enhanced the accuracy of the biochemical data obtained from both infected and healthy groups but also ensured their comparability.
Determination of Protein Content
To quantify protein content, approximately 0.5 g of fresh leaf tissue was homogenized in 5 mL of 50 mmol L-1 sodium phosphate buffer (pH 7.0). From the resulting plant extract, 100 μL was mixed with 5 mL of Coomassie Brilliant Blue G-250 dye reagent. The absorbance of the mixture was measured spectrophotometrically at 595 nm. Protein concentration was quantified by constructing a standard curve using Bovine Serum Albumin Fraction V (Sigma, St. Louis, MO, USA) with concentrations ranging from 10 to 100 μg mL-1.
Determination of Chlorophyll Content
Chlorophyll contents in lucerne leaves were determined following the method of Arnon [11] with minor modifications as described by Karakaş et al. [12]. Approximately 0.5 g of fresh leaf tissue was homogenized in 5 mL of 80% acetone solution (acetone: water, 80:20 v/v). The resulting suspension was filtered through filter paper to separate the liquid phase, which was then transferred into light-proof tubes. Chlorophyll a and b concentrations were measured spectrophotometrically at wavelengths of 663.5 nm and 645 nm, respectively, against a blank solution prepared with 80% acetone (Epoch-BioTek spectrophotometer, BioTek Instruments, Winooski, VT, USA). Total chlorophyll content was calculated in mg fresh weight leaf tissue with the following equations.
Total chlorophlyll (mg L-1)= 20.2 A625+8.02 A663.5
Chlorophlyll a (mg L-1)= 12.7 A663.5-2.69 A645
Chlorophlyll b (mg L-1)= 22.9 A645-4.68 A663.5
Determination of Total Phenolic Content
Total phenolic content was measured using a modified version of the method described by Shetty et al. [13]. Approximately 0.5 g of lucerne leaf tissue was homogenized in 80% methanol and incubated at 95°C for 30 minutes, followed by centrifugation at 10,000 × g for 10 minutes. For the phenolic assay, 300 μL of the extract was mixed with 1.5 mL of diluted (1:10) Folin–Ciocalteu reagent. After 5 minutes, 1.2 mL of 20% sodium carbonate (Na₂CO₃) was added, and the mixture was incubated in the dark at 40°C for 30 minutes. The absorbance of the resulting solution was measured spectrophotometrically at 760 nm, and total phenolic content was calculated using a gallic acid standard calibration curve.
Peroxidase (POX, E.C. 1.11.1.7) Activity Assay
Peroxidase (POX) activity was determined using the method of Cvıkorová et al. [14] with minor modifications [15]. Approximately 0.5 g of leaf tissue was homogenized in 50 mmol L-1 phosphate buffer (pH 7.0). Subsequently, 100 μL of the extract was added to a 3 mL reaction mixture containing 13 mmol L-1 guaiacol, 5 mmol L-1 H₂O₂, and 50 mmol L-1 sodium phosphate buffer (pH 6.5). The enzymatic reaction was initiated by the addition of H₂O₂, and the increase in absorbance was recorded spectrophotometrically at 470 nm every minute for three consecutive measurements at 25°C. POX activity was expressed as the change in absorbance (ΔA470) of 0.1 per minute and reported as units per mg of protein.
Catalase (CAT, E.C. 1.11.1.6) Activity Assay
Catalase (CAT) activity was measured following the method described by Milosevic and Slusarenko [16] with minor modifications introduced by Karakaş [17]. Approximately 0.5 g of leaf tissue was homogenized in phosphate buffer at pH 7.0. From the resulting supernatant, 50 μL was mixed with a 2.95 mL reaction mixture containing 10 mmol L-1 H₂O₂, 50 mmol L-1 phosphate buffer (pH 7.0), and 4 mmol L-1 Na₂EDTA. The decrease in absorbance was recorded spectrophotometrically at 240 nm for 30 seconds at 25°C. Enzyme activity was defined as the amount of enzyme decomposing 1 μmol of H₂O₂ per minute [18] and expressed as units per mg of protein.
Determination of Jasmonic acid (JA) content
Determination of jasmonic acid (JA) content was carried out according to the method described by Annigeri et al. [19]. Initially, 1 g of fresh leaf tissue was extracted with 10 mL of absolute ethanol and incubated in the dark at room temperature for 12 hours. At the end of the extraction period, the mixture was filtered through Whatman No. 1 filter paper, and 1 mL of the filtrate was used to measure absorbance at 323 nm using a spectrophotometer (Epoch-BioTek). The JA concentration in the samples was calculated based on a standard curve prepared from known concentrations of JA dissolved in absolute ethanol.
Determination of Salicylic Acid (SA) Content
Salicylic acid (SA) content was determined with slight modifications based on the method of Rainsford [20]. One gram of leaf sample was incubated in 10 mL of ethanol in the dark at room temperature for 12 hours, then centrifuged at 10,000 × g for 10 minutes. A 100 μL aliquot of the supernatant was mixed with 1% ferric chloride solution, and the volume was adjusted to 3 mL. The violet Fe³⁺-SA complex formed was measured spectrophotometrically at 540 nm. SA concentrations were calculated using a standard curve prepared within the range of 0–100 ppm. The reliability of the method is supported by the high solubility of SA and its ability to react readily with various solvents.
2.4. Histological Analysis of Cuscuta sp. Infection on Medicago sativa L.
Histological analysis was made in lucerne plants from the infection point of Cuscuta sp. followed by 20 days of infection. The cut-stem section around the infection point was fixed using formalin: acetic acid: ethanol (1:1:1, volume) solution. To enhance visualization of cellular structures, the prepared sections were stained with Safranin-O/Fast Green or Toluidine Blue. When necessary, histochemical stains such as Periodic Acid-Schiff (PAS) for polysaccharide accumulation and Aniline Blue for detecting callose deposition were applied [21].
2.5. Determination of DNA fragmentation in Medicago sativa and Cuscuta sp.
The pathogenic effect of Cuscuta sp on Medicago sativa L. plants was evaluated to find out if DNA integrity was disturbed. DNA fragments were also evaluated in non-infected lucerne and Cuscuta sp. plants. The degree of DNA damage or disruption of DNA integrity can provide clues as to whether the plant can compensate for the loss caused by stress, i.e. it can firmly say whether the stressed plant can recover or not. For this purpose, plant genomic DNA was isolated using methods developed by Ahrens and Seemuller [23] and Surapu et al. [24] with minor modifications. Fresh wheat leaves (1 g) were ground in a mortar with 4 mL CTAB buffer (2% w/v cetyltrimethylammonium bromide, 1.4 mol L⁻¹ NaCl, 0.2% β-mercaptoethanol, 20 mmol L⁻¹ EDTA, 100 mmol L⁻¹ Tris-HCl, 2% polyvinylpyrrolidone, pH 8.0). Followed by the protocol, the resulting DNA solution was dissolved in 80-100 μL Tris-EDTA (TE) buffer (10 mmol L-1 Tris-HCl, 1 mmol L-1 EDTA, pH 8). The DNA fragments were separated by electrophoresis in a 1% agarose gel (Sigma, Aldrich) prepared in 1xTris Acetate EDTA (TAE) at 85 V for 75 minutes, which was proved that the yellow colour in the loading buffer solution reached the opposite end of the gel tank (25). (Sigma, Aldrich) prepared in 1xTris Acetate EDTA (TAE) (Table 3.7) at 85 V for 75 minutes. Post-electrophoresis DNA images were obtained by staining with ethidium bromide (1 μg mL⁻¹). Since DNA is a molecule visible under UV fluorescent light, the fragmented DNA bands became visible and exhibit ladder-like patterns using the imaging system.
2.6. Statistical Analysis
Biochemical data obtained from dodder-infected and non-infected plants were analyzed using Minitab 20.1 software (Minitab Inc., State College, PA, USA) [26]. Results are presented as mean ± standard error (n=3). Statistical comparisons and relevant tests were performed to assess significant differences between treatment groups.
3. Results
3.1. Yield and Growth Measurements
The initial plant height measurement was conducted on the 90th day after sowing; the subsequent measurements were made at 15-day intervals on the 105th, 120th, and 135th. Based on the data, the shoot lengths of lucerne (Medicago sativa L. cv. Alsancak) plants infected with Cuscuta sp. were found to be significantly lower when compared to non-infected plants (Figure 1) (p < 0.05).
The harvest commenced on the 90th day post-sowing, with a total of four harvests conducted at 15-day intervals (on 105th days, 120th days, and 135th days). Analysis of data revealed that both fresh and dry forage yields of dodder-infested lucerne plants were significantly lower than those of non-infested plants (p < 0.05) (Figure 2 and Figure 3).
The results regarding fresh and dry weight were converted to kg yield per da area. When conversion was made, the average green herbage yield in the dodder-infected area was measured at 847.7 kg/da, while the dry herbage yield was 324.64 kg/da. In contrast, the corresponding values for the uninfected control plants were recorded as 1153.43 kg/da and 402.22 kg/da, respectively.
3.2. Biochemical Analysis
3.2.1. Chlorophyll and Protein Contents
Leaves and stems of dodder-infected plants were analyzed separately. According to the results, total chlorophyll and chlorophyll a and b contents in infected lucerne leaves and stems were significantly lower when compared to those of healthy plants. Although protein contents were low in infected plants, no statistical differences were observed between the infected and healthy leaves (Figure 4, Figure 5 and Figure 6). In contrast, a statistically significant decrease in protein content was detected when infected and healthy stems were compared (p < 0.05) (Figure 7).
3.2.2. Quantitative Evaluation of Phenolic Compounds
In order to determine the effects of dodder infection on the host plant, the leaf and stem tissues of infected plants were evaluated separately. The analyses revealed that the phenol content in both tissues of infected lucerne plants was significantly lower when compared to that of healthy plants (p < 0.05) (Figure 8).
3.2.3. Quantitative Assessment of Antioxidant Enzyme Activities
The activities of catalase (CAT) and peroxidase (POX) enzymes, jasmonic acid (JA) and salicylic acid (SA) were independently assessed in the leaf and stem tissues of dodder-infected and healthy plants. The analyses revealed that CAT and POX activities in the leaf tissues of infected plants did not differ significantly from those of healthy controls. However, CAT and POX activities in the stem tissues of infected plants were significantly different when compared to healthy controls. On the other hand, it was determined that the levels of JA and SA in both leaf and stem tissues did not show a statistically significant difference from those of control plants (Figure 11 and Figure 12).
Figure 9.
Catalyse (CAT) activity was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
Figure 9.
Catalyse (CAT) activity was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
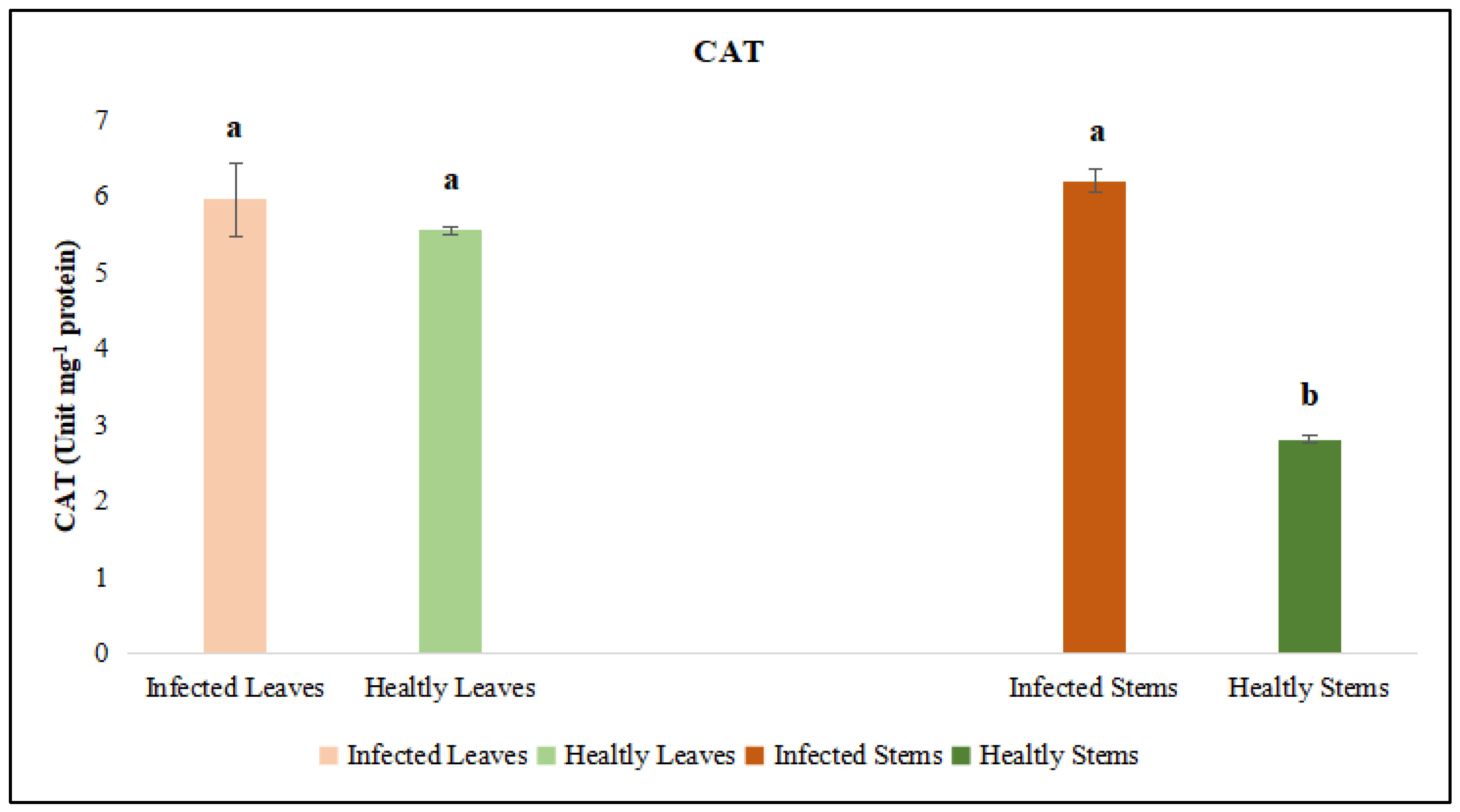
Figure 10.
Peroxidase (POX) activity was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
Figure 10.
Peroxidase (POX) activity was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
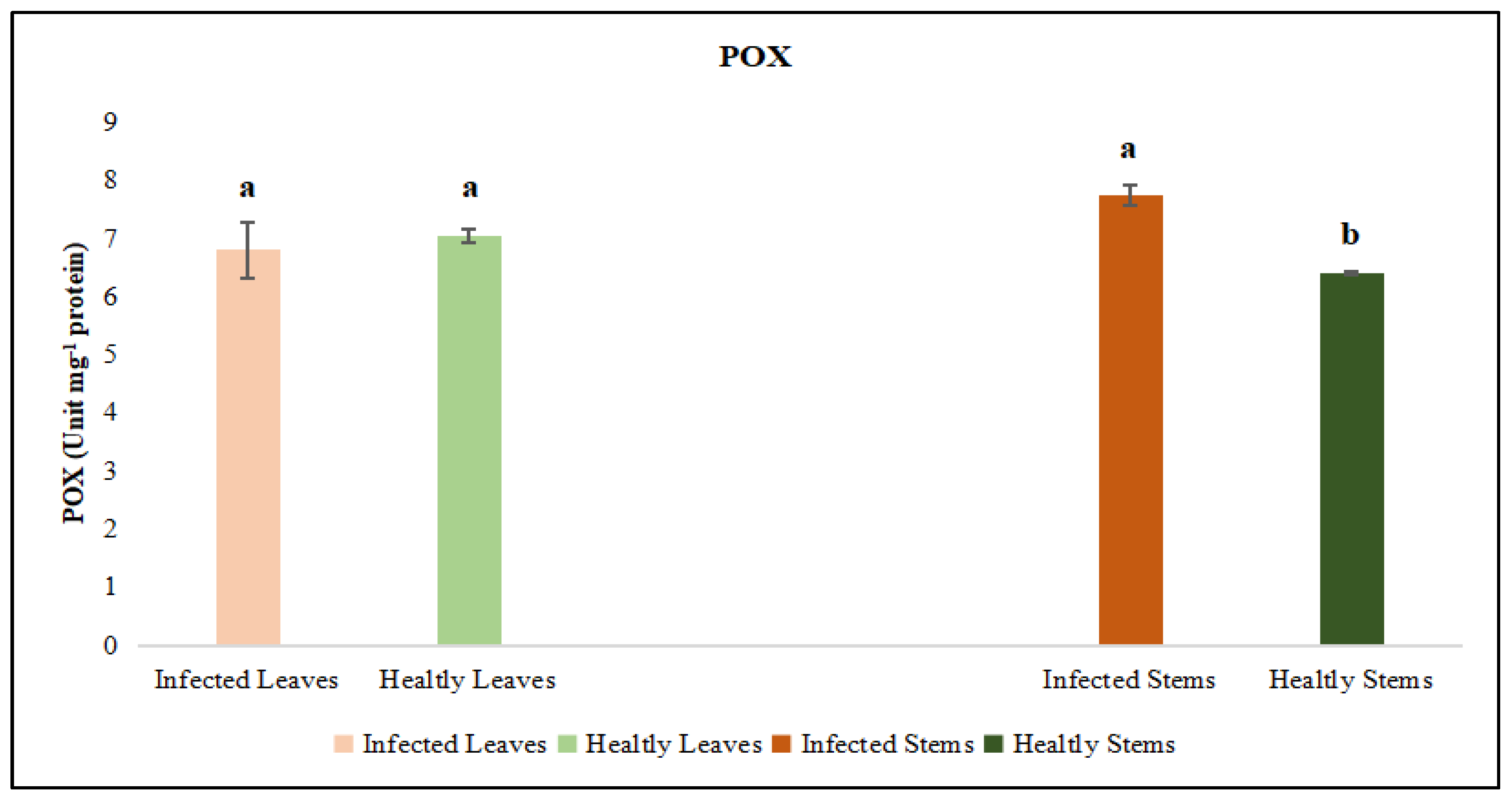
Figure 11.
Jasmonic acid (JA) activity was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
Figure 11.
Jasmonic acid (JA) activity was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
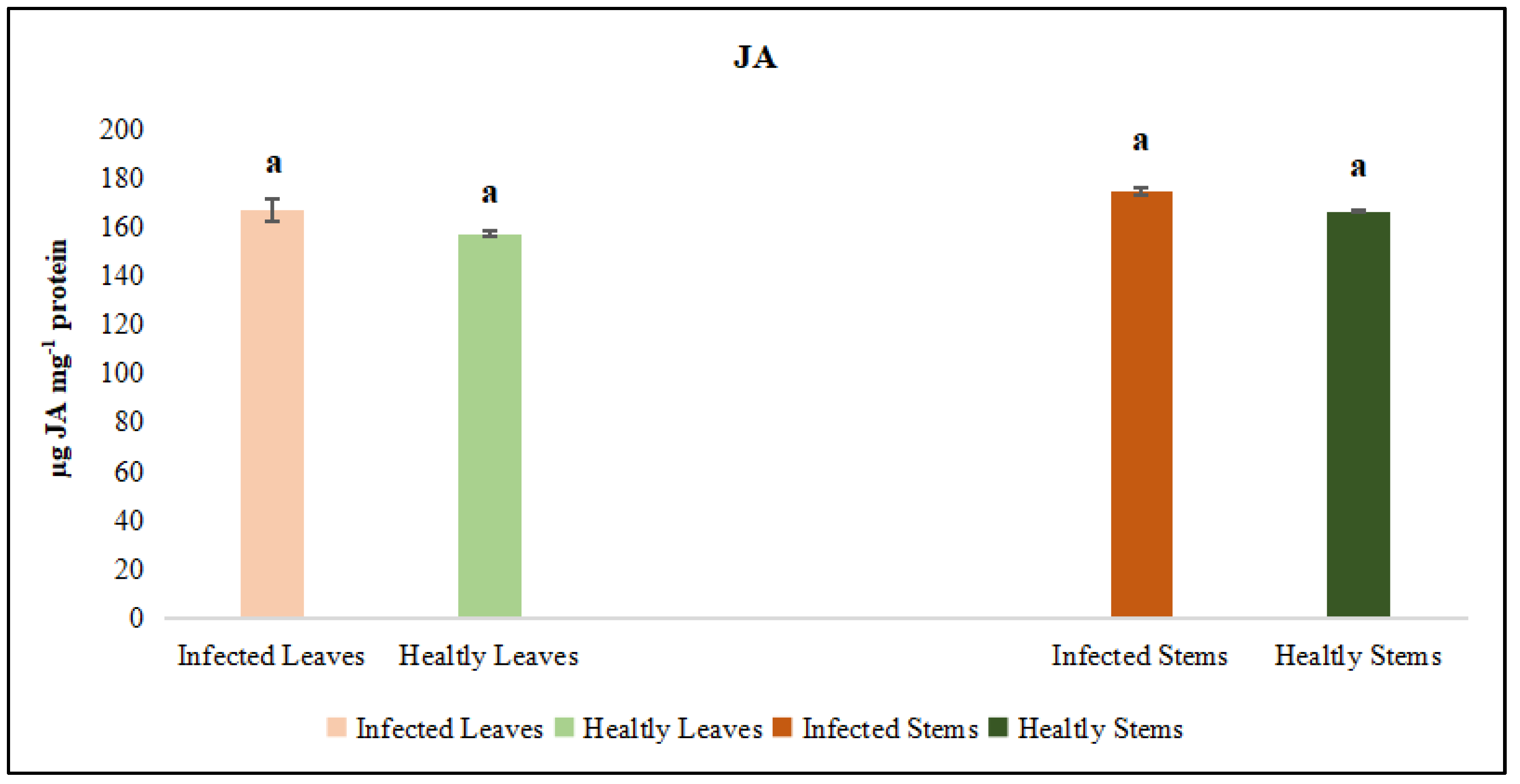
Figure 12.
Salicylic acid (SA) activity was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
Figure 12.
Salicylic acid (SA) activity was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
3.2.4. Evaluation of Callose Responses Induced by Parasitic Infection in Plants
Samples were fixed in formalin: acetic acid: ethanol solution and subsequently dehydrated through a graded ethanol series. The tissues were then embedded in paraffin, and 7–10 µm-thick sections were obtained. The sections were stained with Safranin-O/Fast Green or Toluidine Blue, examined under a light microscope, and images were captured using a digital camera. Microscopic analysis clearly revealed prominent callose formation at the sites where Cuscuta sp. penetrated the host plant.
Figure 13.
Attachment of Cuscuta sp to the stem of lucerne (Medicago sativa L. cv. Alsancak) via haustoria and the formation of feeding sites at the points of contact (×400 magnification).
Figure 13.
Attachment of Cuscuta sp to the stem of lucerne (Medicago sativa L. cv. Alsancak) via haustoria and the formation of feeding sites at the points of contact (×400 magnification).
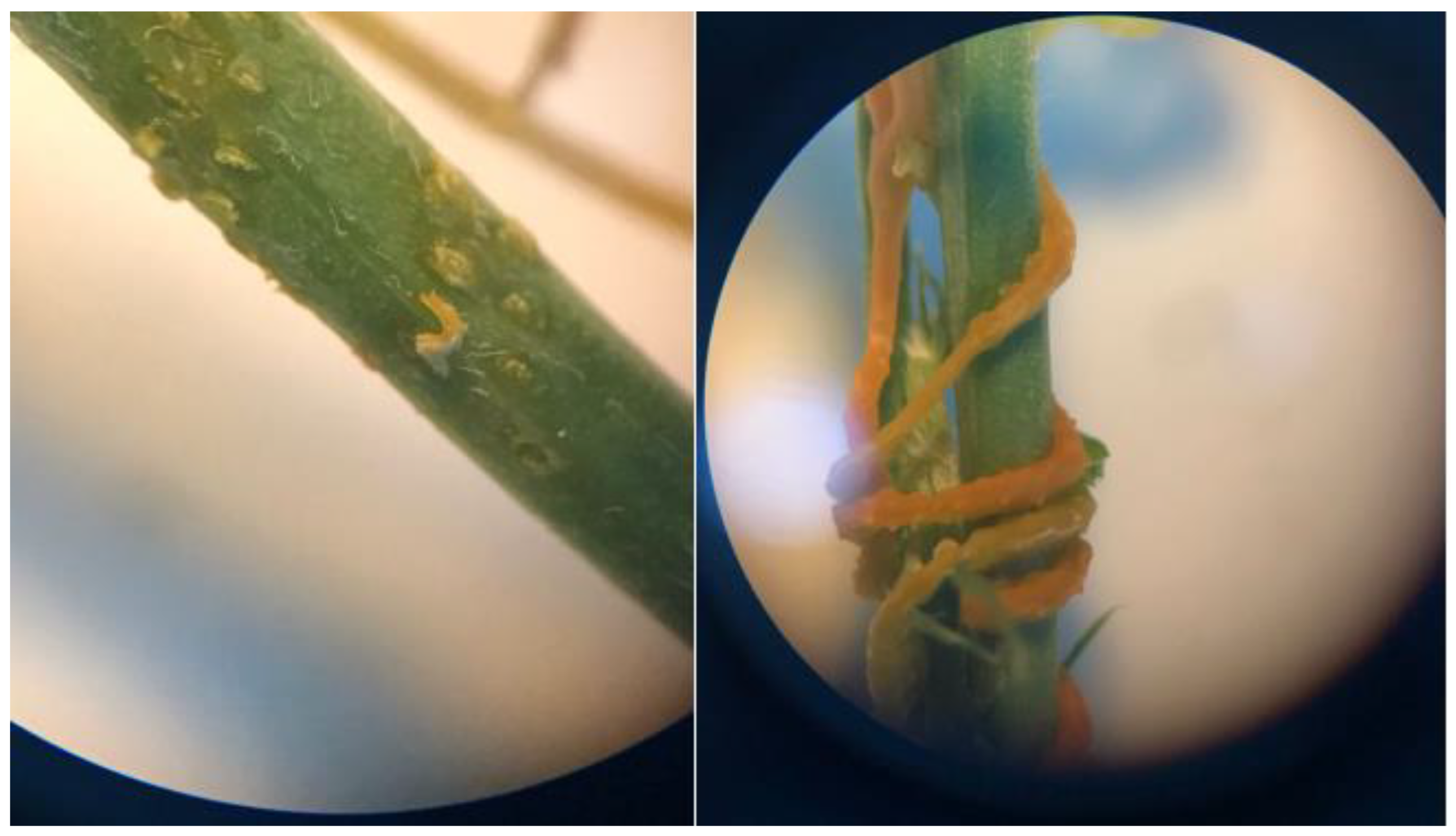
Figure 14.
Structural appearance and internal positioning of Cuscuta sp. haustoria penetrating the lucerne (Medicago sativa L. cv. Alsancak) stem (×400 magnification).
Figure 14.
Structural appearance and internal positioning of Cuscuta sp. haustoria penetrating the lucerne (Medicago sativa L. cv. Alsancak) stem (×400 magnification).
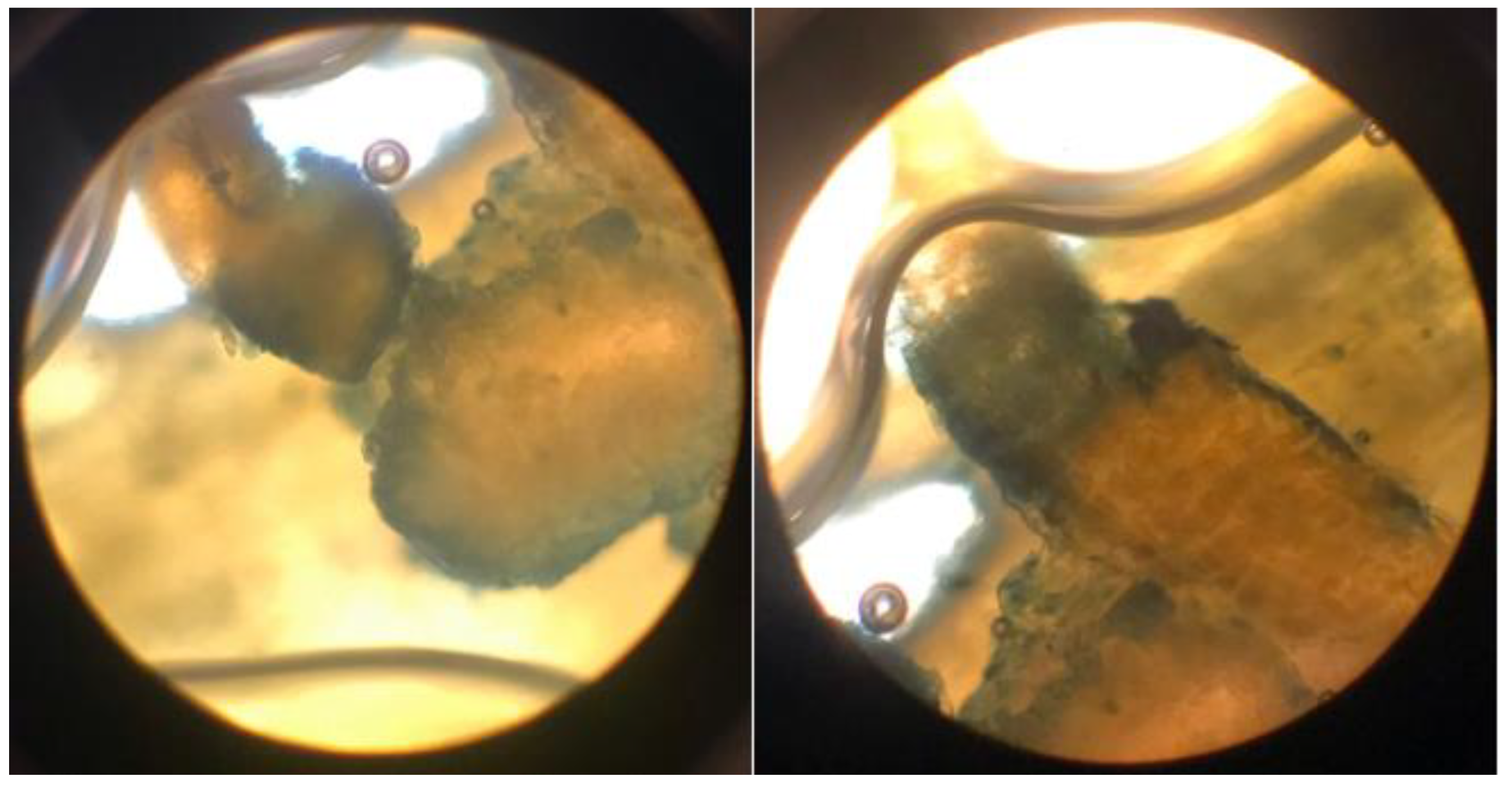
Figure 15.
Light microscopy image of the callose tissue developed in the host tissue following the penetration of Cuscuta sp. into the (Medicago sativa L. cv. Alsancak) stem (×400 magnification).
Figure 15.
Light microscopy image of the callose tissue developed in the host tissue following the penetration of Cuscuta sp. into the (Medicago sativa L. cv. Alsancak) stem (×400 magnification).

3.2.5. Evaluation of DNA damage in Medicago sativa infected with Cuscuta sp.
Lucerne plants followed by the infection of Cuscuta sp. Showed significant responses in terms of physiological and biochemical means, most of these symptoms have been differentially expressed in leaves and stems. We evaluated if DNA was damaged or distrupted upon infection by dodders. We found that the damage caused by Cuscuta sp. was not severe but evident in infected lucerne plants, Figure 16.
A-DNA fragmentation images, B-Infected and control lucerne plants with Cuscuta sp.
Although not a very big difference was observed between inoculated control plants, the infected lucerne showed slightly more damage than the control plants in terms of DNA fragmentation. Cuscuta sp. made this infection with very little DNA contents. It is important to note that lucerne plants carried out their metabolic functions with very little damage, however, this analysis was made at the end of the harvest period in which the control plants might have shown the sign of ageing
4. Discussion
Cuscuta sp. (dodder) is a holoparasitic plant, which extracts water and nutrients directly from its host, severely compromising the host’s growth and development. This parasitism is particularly detrimental to economically important forage crops such as lucerne (Medicago sativa L.) and other crop plants and vegetables, leading to substantial yield losses. Attachments of the parasite to the host vascular system via haustoria facilitates the transfer of essential resources, including water, carbohydrates, and minerals [27]. Consistent with previous research, this study confirms that dodder infection markedly affects the anatomical and physiological parameters of lucerne. “Dooders” is the common name for a group of holoparasitic plants of the angiosperm genus Cuscuta spp. Since every infection starts with the parasite coiling around the host stem and glueing itself onto its host surface with a sticky mucilage, the infection becomes more severe when compared to those of any fungal pathogens due to the single infection point being the main route to pathogenic infection. Therefore, suppression of host defense created by holoparasite plants could be much easier when compared to those of other infection strategies. Here, the haustoria break through the host surface and grow until they have reached the vascular tissue, and spread laterally and apically via finger-like scavenging cells called “feeding hyphae”. During penetration, the haustoria soften and decompose the middle lamellae of the host to suppress defense response. For a successful infection, Cuscuta has to inhibit signaling mechanisms of the host. Dodders can also form phloem and/or xylem fusion in the host vascular systems. In this way, dodders can take up minerals, water and organic compounds at the same time from the host. It could nourish itself with organic compounds while decreasing the defense mechanisms of the host via transferring the secondary metabolites, messenger RNAs, micro RNAs, amino acids, proteins, and noncoding RNAs to suppress the signaling molecules in the host [28]. Yang et al. [29] reported that horizontal gene transfer (HGT), the movement and genomic integration of DNA across species generally associated with bacteria and other fungal organisms, was also observed between Cuscuta and its hosts. It is known that transfer of genetic material between bacteria and their hosts forms the basis of antibiotic resistance. The authors stated that 108 genes transferred to the dodder’s genome were involved in parasitism, haustorial structure and amino acid metabolism, and microRNAs (miRNAs) produced in the cell metabolism of dodders were sent back to the host plant to suppress defense through gene silencing. Although major pathways have been revealed followed by dodder infection, however, how dodders find host plants and locate the infection points and carry out pathogenicity without significant defense responses remains to elusive. Recent studies showed that the mechanism of defense suppression is more complex than previously thought. For example, Shahid et al. [30] showed that Cuscuta campestris accumulated high levels of many novel miRNAs while paraziting Arabidopsis thaliana through haustoria. Many of these miRNAs decreased mRNA accumulation in A. thaliana. The authors suggested that these miRNAs acted as virulence factors during parasitism. For example, Förste et al. [31] concluded that the uptake of minerals was selectively regulated. The authors stated that calcium and manganese were depleted in the host. As a result of the depletion of Ca in host plants led to a reduction in pectin content, which resulted in further penetration of Cuscuta. This could partly explain why defense mechanisms are interrupted and suppressed. Similar issues were made with those involved in plant and fungal pathogen interactions. Tian et al [32] reported that obligate plant pathogens could secrete effector proteins to suppress the plant defense response to make successful colonization. They stated that Puccinia striiformis f.sp. tritici (Pst), one of the most devastating wheat pathogens, secreted effector proteins to suppress plant defense mechanisms. So, it is important to note that here, obligate parasites have the ability to secrete effectors to suppress the host defense. Unlike facultative pathogens, obligate plant pathogens or obligate holoparasitic plants want to control the activities of the host; therefore, they can suppress the host defense depending on the genetic background of the host. Similarly, Xie et al. [33] reported the same mechanism with that of Meloidogyne incognita on the suppression of plant defense mechanism via Misp 12 gene involved in effector biosynthesis. We can enlarge our examples of plant defense suppression. Blaazer et al. [34] reported that several species of herbivorous mites, such as Tetranychus urticae and T. evansi maximized their performance by suppressing plant defenses. The more complex issue is underlined by Hettenhausen et al. [35] who stated that metabolites, proteins, and mRNAs are known to be transferred from hosts to Cuscuta, and systemic herbivory signals are transmitted from attacked plants to unattacked plants via Cuscuta bridges. Some authors suggested that Cuscuta spp. induced ethylene in susceptible crop cultivars, including vegetables such as tomato and tobacco plants [36]. This pathway may have been involved in suppression of the defense responses of M. sativa. However, more pathways are needed other than hormone signaling to reveal the mechanisms underlying defense suppression.
Saric-Krsmanovic et al. [37] reported significant anatomical alterations and physiological impairments in infected plants. The findings are further supported by field studies that show dodder infestation weakens host plants, resulting in decreased growth and reproductive capacity [38]. Notably, dodder parasitism has been associated with reduced dry matter accumulation, thereby negatively impacting yield and biomass production.
At the molecular level, transcriptomic analyses revealed that dodder infection downregulated photosynthesis-related gene expression and disrupted protein synthesis in the host [39]. This downregulation is reflected physiologically by a significant decrease in photosynthetic capacity in lucerne during infection [7,40]. The parasite also induced activation of host defense-related genes and signaling pathways, which further suppressed photosynthetic activity [41]. Consequently, the reduction in photosynthesis adversely affects the protein synthesis, which correlates with decreased chlorophyll content—including total chlorophyll and chlorophyll a and b—in infected tissues [42,43]. Energy spent for the defense purposes will cost the quality parameters, as shown in the work of Baran et al. [25] Moreover, dodder infection significantly disrupts photosystem II activity and other chlorophyll fluorescence parameters, indicating compromised light-harvesting efficiency and photosynthetic electron transport [44]. This reduction in photosystem II function is likely linked to diminished chlorophyll levels and disruption of the light-harvesting chlorophyll-protein complex II [42,45]. The suppression of photosynthetic protein gene expression coupled with the activation of defense responses highlights a complex host-parasite interaction, where energy production is sacrificed to mount an effective defense.
Lucerne serves as a major source of animal fodder; the observed decline in protein synthesis and photosynthetic efficiency has important implications for both crop yield and nutritional quality. Protein reduction in parasitized leaves adversely affects forage quality, directly impacting livestock nutrition and productivity. Farah and Al-Abdulsalam [46] reported yield losses exceeding 50% in heavily infected fields, underscoring the significant economic threat posed by dodder. Overall, these findings highlight the multifaceted impact of Cuscuta sp. on lucerne, involving anatomical damage, physiological dysfunction, and molecular disruptions that collectively reduce host vitality and productivity. Future research should focus on elucidating the molecular mechanisms underpinning host resistance and parasite virulence. Such insights could facilitate the development of resistant lucerne cultivars or innovative management strategies to mitigate the detrimental effects of dodder parasitism.
Phenolic compounds are secondary metabolites that play a crucial role in plant defense mechanisms against various biotic stress factors, including parasitic infections. These compounds function as direct antimicrobial agents against pathogens, participate in signal transduction processes, and contribute to the reinforcement of cell walls through lignin biosynthesis, thereby forming a physical barrier [47]. However, several studies have reported that Cuscuta sp. infections suppress phenolic metabolism or reduce the levels of these compounds in host plants. For instance, Runyon et al. demonstrated that phenolic accumulation was quite low followed by the suppression of pheylpropanoid pathway in tomato plants infected by dodder pentagona [48]. Similarly, Albert et al. [49] reported that dodder species can actively suppress host defense responses, particularly phenolic compound production, to ensure successful parasitism. In the present study, a significant reduction in phenol content was observed in both leaf and stem tissues of infected lucerne plants, suggesting that dodder remarkably weakened the host defense systems. This suppression may be mediated through the disruption of crosstalk between phytohormones, especially ethylene, SA and JA, which play a key role in the biosynthesis of phenolic compounds [50]. Moreover, parasitic plants such as dodder have been reported to alter phenolic accumulation and inhibitory activity in host tissues, thereby compromising the host’s defense capacity [51].
Parasitic plant infections disrupt the oxidative balance in host plants by inducing metabolic stress and promoting the accumulation of reactive oxygen species (ROS). Under such biotic stress conditions, plants activate tissue-specific antioxidant defense mechanisms to maintain cellular homeostasis. The findings of this study demonstrate that enzymatic responses to dodder infection vary depending on the tissue type, reflecting differential regulation of stress signaling across plant organs. A notable increase in CAT activity observed specifically in stem tissues suggests a critical role for this enzyme in ROS detoxification. Previous studies have reported that CAT activity increases during certain developmental stages in plant–pathogen interactions to transiently regulate intracellular H₂O₂ levels [52]. The elevation of CAT activity in stems of dodder-infected plants may represent a localized defense response aimed at preserving tissue integrity, especially at the site of haustorial invasion.
Similarly, the significant increase in POX activity, again limited to stem tissues, highlights another key component of the host defense system. POX enzymes are involved in lignin biosynthesis and cell wall strengthening, processes that contribute to the formation of structural barriers against invading pathogens [53]. The increased POX activity observed in infected regions has been linked to activation of the phenylpropanoid pathway and suggests a defence-induced remodeling of cell walls [54]. However, in leaves of lucerne plants, increases in antioxidant enzymes ceased, indicating that defense responses were not carried out further. These tissue-specific increases in CAT and POX activities imply that the host plant develops a localised defence strategy in response to dodder infection. Given that the stem is the primary site of haustorial penetration and resource extraction, the enhanced activation of antioxidant enzymes in this region appears to be a biologically adaptive response. In contrast, the absence of significant enzymatic changes in the leaf tissue may indicate that the defense response is concentrated at the infection site and that systemic responses are either limited or delayed.
Jasmonic acid (JA) and its derivatives, along with salicylic acid (SA), are key regulators of plant defense responses against diseases and environmental stress factors [55]. The effects of changes in JA levels on plant defense responses are quite complex and multifaceted [56]. In particular, increases in JA levels enhance the expression of defence-related genes, thereby promoting the synthesis of defense metabolites in plants [57,58]. Plants regulate their defense responses against environmental stress and pathogen attacks not only through JA but also via other hormonal signaling pathways, such as SA, ethylene, and abscisic acid are involved. Therefore, even if a significant increase in JA levels is not observed, effective defense responses can still be activated through these alternative pathways. Moreover, JA responses are typically rapid and transient; JA levels may rise shortly after stress exposure and quickly return to baseline [59]. The synthesis and degradation of JA are tightly controlled by negative feedback mechanisms, preventing unnecessary accumulation. As a result, stable JA levels may be detected at the time of measurement. This dynamic regulation ensures efficient use of the plant’s energy resources and maintains hormonal balance. On the other hand, SA is a phenolic phytohormone involved in key physiological processes such as photosynthesis, transpiration, and the uptake and transport of ions [60,61]. It also plays a crucial regulatory role in shaping leaf anatomy and maintaining the structural organization of chloroplasts [62,63]. The synthesis, accumulation, and metabolism of SA can rapidly fluctuate depending on environmental conditions [64]. Derivatized forms of SA produced during these processes may not always be detectable, which can result in the absence of a noticeable change in total SA levels [65]. Moreover, plants can activate their defense responses not only through SA but also through other phytohormones, defense proteins, low concentrations of ROS, and phytoalexins can take significant roles [66,67]. In certain plant–pathogen interactions, SA signaling may be suppressed or activated with a delay. Our results indicate that dodder infection elicited tissue-specific oxidative stress responses in the host plant. The observed enzymatic changes supported the hypothesis that host defense involved both the regulation of ROS levels and structural changes in infected tissues to counteract parasitic invasion. Parasitic infections induced localized anatomical changes in the host and partially suppressed the defense mechanisms. Prominent callose formation was observed at the sites where Cuscuta sp. attached to the host plant. This suggests that the plant attempts to limit parasite progression by creating a physical barrier through localized cell proliferation at the infection site. The literature frequently emphasizes that callose tissue develops as a mechanical and chemical defense response against parasitic plants [68,69]. The intense callose formation in stem tissues particularly highlights the critical role of localized defence, given that dodder haustoria obtain nutrients through this region. In conclusion, callose development represents an effective tissue-level defense strategy employed by the host plant against dodder infection.
When DNA fragementaion assay was performed, we observed that Cuscuta-infected M. sativa accumulated slightly more low molecular weight DNA molecules when compared to control groups. Although Cuscuta sp. did not cause significant DNA damages, low level of DNA damages could distrupt the whole metabolism. We are about to evaluate the progress of DNA damages through timescale experiment if recovery takes place. Distruption in DNA molecules could be very costly for the host plant. For example, Turan et al. [70] reported that DNA fragmentation in spermatozoa affected the fertility in men. Similar issues were made by Kaya et al. [71] who stated that high-dose vanadium resulted in DNA fragmentaion in Allium plants and signifcantly disturbed the metabolism. Again, Plitta-Michalak et al. [72] showed that the loss of DNA integrity distrupted the whole metabolism and either delayed or prevented the recovery of Acer pseudoplatanus L.
5. Conclusions
Understanding the biochemical and molecular pathways involved in the pathogenicity of holoparasites is essential to the control of the parasitic weeds. The majority of the research has been conducted between parasitic plants and their hosts; however, stress-tolerant parasitic plants may threat to agricultural production, followed by climate changes that could modify irrigation regimes, and cause drought, salinity, and other environmental stresses. Under these circumstances, tolerance or resistance levels of host plants break, and defense suppression could be easier for the stress-tolerant holoparasitic plants. To better understand the defense suppression of host plants followed by holoparasitic infection, this study formed a strong database to evaluate the behaviour of holoparasites under stress conditions. Demirbas et al. [73] reported that A. thaliana seedlings exposed to salinity were more sensitive to Phelipanche ramosa when compared to the uninoculated plants, indicating that there was a positive correlation between salt sensitivity and the ability of P. ramosa to infect A. thaliana plants. Defense barrier was further suppressed. Therefore, resistant plants could not defend themselves when exposed to multiple stressors. Our findings demonstrated that dodder parasitism in lucerne triggered significant declines in chlorophyll content, photosynthetic efficiency, and protein synthesis, alongside notable structural modifications in host tissues. The observed suppression of phenolic compounds, which are key components of plant defense suggests that dodder strategically impairs the host’s protective metabolic pathways—particularly those associated with the phenylpropanoid biosynthetic route and phytohormonal crosstalk involving SA and JA. Concurrently, a tissue-specific oxidative stress response was recorded with elevated CAT and POX activities in stem tissues and decreased or unchanged activities in leaves, indicating a localized antioxidant defense strategy.
Histological analysis further revealed prominent callose formation at the penetration sites. This localized cell proliferation likely serves as a physical barrier against parasitic invasion and represents a structural manifestation of host defense. The intensity of callose development in stem tissues underscores the biological importance of localized defense responses in regions directly targeted by the parasite.
As DNA fragmentation was examined, the infected plants showed signs of DNA damage. The observation regarding DNA damage was obtained at the very end of the trial, a sort of recovery might have been occured. In our coming studies, we would observe DNA integrity at earlier times with more frequent time intervals to elucidate the threshold level of tolerance of the plant against Cuscuta infection.
Collectively, the data suggest that dodder infection elicits a complex interplay of localized and systemic responses in lucerne involving both the suppression and activation of defense pathways. Understanding these intricate host–parasite interactions is critical for developing resistant lucerne cultivars and designing effective management strategies to mitigate the impact of dodder infestation in agricultural systems.
Host defense response suppression should be evaluated further what genes and metabolites are involved. Under the combined stress, suppression of host defense should be studied extensively if further suppression is carried out in terms of downregulation of defence-related genes. If defence-related genes could show an increase or stability, we could follow that pathway to generate or breed multiple stress-tolerant plants, including those resistant to holoparasitic plants; if not, we could come back to square one and start generating multiple stress-tolerant crop plants after revealing all pathways. If tolerance- or resistance-related defense genes and metabolites are downregulated, the only way to generate more toleran or resistant crop plants should follow revealing whole plant metabolism. Therefore, breeding tasks would be more difficult as the stress get more complex.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, Hümeyra Ayvacı and Murat Dikilitas.; methodology, Hümeyra Ayvacı and Murat Dikilitas; software, Hümeyra Ayvacı ; validation, Hümeyra Ayvacı, Ertuğrul Mehmet Güldür and Murat Dikilitas; formal analysis, Hümeyra Ayvacı; investigation, Hümeyra Ayvacı; resources, Hümeyra Ayvacı and Murat Dikilitas; data curation, Hümeyra Ayvacı and Murat Dikilitas; writing—original draft preparation, Hümeyra Ayvacı and Murat Dikilitas; writing—review and editing, Murat Dikilitas; visualization, Hümeyra Ayvacı; supervision, Murat Dikilitas; project administration, Hümeyra Ayvacı, Ertuğrul Mehmet Güldür and Murat Dikilitas. All authors have read and agreed to the published version of the manuscript.”
Funding
This research received no external funding.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
The authors would like to thank Harran University for providing access to its laboratory facilities, which greatly contributed to the completion of this study. The authors have reviewed and revised the final version of the manuscript and take full responsibility for its content.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| CAT | Catalase |
| POX | Peroxidase |
| JA | Jasmonic acid |
| SA | Salicylic acid |
| Fwt | Fresh |
| Dwt | Dry |
| HGT | Horizontal Gene Transfer |
References
- Gaweł, E.; Grzelak, M. Protein from lucerne in animals supplement diet. Journal of Food Agriculture and Environment. 2014, 12, 314–319. [Google Scholar]
- Dechassa, N.; Regassa, B. Current Status, Economic Importance and Management of Dodders (Cuscuta spp.) of Important Crops. Advances in Life Science and Technology. 2021, 87, 16–21. [Google Scholar] [CrossRef]
- Masanga, J.; Mwangi, B.N.; Kibet, W.; Sagero, P.; Wamalwa, M.; Oduor, R.; Ngugi, M.; Alakonya, A.; Ojola, P.; Bellis, ES.; Runo, S. Physiological and ecological warnings that dodders pose an exigent threat to farmlands in Eastern Africa. Plant Physiology. 2021, 185, 1457–1467. [Google Scholar] [CrossRef] [PubMed]
- Kokla, A.; Melnyk, CW. Developing a thief: Haustoria formation in parasitic plants. Developmental biology. 2018, 442, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Shen, G.; Wu, J. Jasmonic acid and salicylic acid transcriptionally regulate CuRe1 in cultivated tomato to activate resistance to parasitization by dodder Cuscuta australis. Plant Diversity. 2025, 47, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Moradkhani, S.; Jabbari, H. The effects of foliar spray of chitosan nanoparticles on tomato resistance against of Cuscuta campestris yunck. The Open Medicinal Chemistry Journal. 2023, 17. [Google Scholar] [CrossRef]
- Saric-Krsmanovic, M.; Bozic, D.; Radivojevic, L.; Gajic Umiljendic, J.; Vrbnicanin, S. Response of alfalfa and sugar beet to field dodder (Cuscuta campestris Yunck.) parasitism: a physiological and anatomical approach. Canadian Journal of Plant Science. 2018, 99, 199–209. [Google Scholar] [CrossRef]
- Hegenauer, V.; Slaby, P.; Körner, M.; Bruckmüller, J.A.; Burggraf, R.; Albert, I.; Kaiser, B.; Löffelhardt, B.; Droste-Borel. I.; Sklenar, J.; Menke, FLH.; Maček, B.; Ranjan, A.; Sinha, N.; Nürnberger, T.; Felix, G.; Krause, K.; Stahl, M.; Albert, M. The tomato receptor CuRe1 senses a cell wall protein to identify Cuscuta as a pathogen. Nature communications. 2020, 11, 5299. [Google Scholar] [CrossRef]
- Meighani, F.; Mamnoei, E.; Hatami, S.; Samadi-Kalkhoran, E.; Khalil-Tahmasebi, B.; Korres, NE.; Bajwa, AA. Chemical control of the field dodder (Cuscuta campestris) in new-seeded alfalfa (Medicago sativa). Agronomy. 2024, 14, 1643. [Google Scholar] [CrossRef]
- Sarić-Krsmanović, M.; Božić, D.; Radivojević, L.; Gajić-Umiljendić, J.; Vrbničanin, S. Impact of field dodder (Cuscuta campestris Yunk.) on physiological and anatomical changes in untreated and herbicide-treated alfalfa plants. Pesticidi i fitomedicina. 2016, 31, 115–120. [Google Scholar] [CrossRef]
- Arnon, DL. Copper enzyme is isolated chloroplast: polyphenol oxidase in Beta vulgaries. Plant Physiol. 1949, 24, 15. [Google Scholar] [CrossRef]
- Karakas, S.; Dikilitas, M.; Almaca, A.; Tipirdamaz, R. Physiological and biochemical responses of (Aptenia cordifolia) to salt stress and its remediative effect on saline soils. Appl. Ecol Environ. Res. 2020, 18, 1329–1345. [Google Scholar] [CrossRef]
- Shetty, K.; Curtis, OF.; Levin, RE.; Wikowsky, R.; Ang, W. Prevention of verification associated with in vitro shoot culture of oregano (Origanum vulgare) by Pseudomonas spp. J. Plant Physiol. 1995, 147, 447–451. [Google Scholar] [CrossRef]
- Cvıkorová, M.; Hrubcová, M.; Vägner, M.; Macháčková, I.; Eder, J. Phenolic acids and peroxidase activity in alfalfa (Medicago sativa) embryogenic cultures after ethephon treatment. Physiol. Plant. 1994, 91, 226–233. [Google Scholar] [CrossRef]
- Karakas, S.; Dikilitaş, M.; Tıpırdamaz, R. Biochemical and molecular tolerance of Carpobrotus acinaciformis L. halophyte plants exposed to high level of NaCl stress. Harran, J. Agric. Food Sci. 2019, 23, 99–107. [Google Scholar] [CrossRef]
- Milosevic, N.; Slusarenko, A J. Active oxygen metabolism and lignification in the hypersensitive response in bean. Physiol. Mol. Plant Pathol. 1996, 49, 143–158. [Google Scholar] [CrossRef]
- Karakas, SD. Development of tomato growing ın soils differing in salt levels and effects of companion plants on some physiological parameters and soil remediation. PhD thesis, Harran University, Şanlıurfa-Turkey, 2013.
- Wang, X.S.; Han, J.G. Changes of proline content, activity, and active isoforms of antioxidative enzymes in two alfalfa cultivars under salt stress. Agric. Sci Chin. 2009, 8, 431–440. [Google Scholar] [CrossRef]
- Annigeri, S.; Pankaj Shakil, NA.; Kumar, J.; . Singh, K. Effect of jasmonate (jasmonic acid) foliar spray on resistance in tomato infected with root-knot nematode, Meloidogyne incognita. Ann. Plant Prot Sci. 2011, 19, 446–450. [Google Scholar]
- Rainsford, KD. Aspirin and related drugs, 1rd ed.; Taylor & Francis: London, UK, 2004; pp. 1–23. [Google Scholar]
- Bojko, M.; Kędra, M.; Adamska, A.; Jakubowska, Z.; Tuleja, M.; Myśliwa-Kurdziel, B. Induction and characteristics of callus cultures of the medicinal plant Tussilago farfara L. Plants. 2024, 13, 3080. [Google Scholar] [CrossRef]
- Pan, H.; Liao, R.; Zhang, Y.; Arif, M.; Zhang, Y.; Zhang, S.; Zhao, YWP. ; Wang, Z.; Han. B.; Song, C. Establishment of callus induction and plantlet regeneration systems of Peucedanum Praeruptorum dunn based on the tissue culture method. Plant Methods. 2024, 20, 174. [Google Scholar] [CrossRef]
- Ahrens, U.; Seemüller, E. Detection of DNA of plant pathogenic mycoplasmalike organisms by a polymerase chain reaction that amplifies a sequence of the 16 S rRNA gene. Phytopathology. 1992, 82, 828–832. [Google Scholar] [CrossRef]
- Surapu, V.; Ediga, A.; Meriga, B. Salicylic Acid Alleviates Aluminum Toxicity in Tomato Seedlings (Lycopersicum esculentum Mill.) through Activation of Antioxidant Defense System and Proline Biosynthesis. Advances in Bioscience and Biotechnology. 2014, 5, 777–789. [Google Scholar] [CrossRef]
- Baran, B.; Ölmez, F.; Çapa, B.; Dikilitas, M. Defense Pathways of Wheat Plants Inoculated with Zymoseptoria tritici under NaCl Stress Conditions: An Overview. Life. 2024, 14, 648. [Google Scholar] [CrossRef] [PubMed]
- Minitab. https://www.minitab.com/en-us/products/minitab-solution-center/free-trial/ (3.10. 2025.
- Kaiser, B.; Vogg, G.; Fürst, UB.; Albert, M. Parasitic plants of the genus Cuscuta and their interaction with susceptible and resistant host plants. Frontiers in plant science. 2015, 6, 45. [Google Scholar] [CrossRef]
- Martinez, G.; Krause, K. The parasitic plant haustorium: a trojan horse releasing microRNAs that take control of the defense responses of the host. Non-coding RNA Investigation. 2018, 2. [CrossRef]
- Yang, Z.; Wafula, EK.; Kim, G.; Shahid, S.; McNeal, JR.; Ralph, PE.; Timilsena, PR.; Yu, W.; Kelly, EA.; Zhang, H.; Person, TN.; Altman, NS.; Axtell, MJ.; Westwood, JH.; dePamphilis, CW. Convergent horizontal gene transfer and cross-talk of mobile nucleic acids in parasitic plants. Nature Plants. 2019, 5, 991–1001. [Google Scholar] [CrossRef]
- Shahid, S.; Kim, G.; Johnson, NR.; Wafula, E.; Wang, F.; Coruh, C.; Bernal-Galeano, V.; Phifer, T.; dePamphilis. CW.; Westwood, HT.; Axtell, MJ. MicroRNAs from the parasitic plant Cuscuta campestris target host messenger RNAs. Nature. 2018, 553, 82–85. [Google Scholar] [CrossRef]
- Förste, F.; Mantouvalou, I.; Kanngießer, B.; Stosnach, H.; Lachner, LAM. ; Fischer, K.; Krause, K. Selective mineral transport barriers at Cuscuta-host infection sites. Physiologia Plantarum. 2020, 168, 934–947. [Google Scholar] [CrossRef]
- Tian, M.; Zhang, Z.; Bi, X.; Xue, Y.; Zhou, J.; Yuan, B.; Feng, Z.; Wang, J. A Putative effector Pst-18220, from Puccinia striiformis f. sp. tritici, participates in rust pathogenicity and plant defense suppression. Biomolecules. 2024, 14, 1092. [Google Scholar] [CrossRef]
- Xie, J.; Li, S.; Mo, C.; Wang, G.; Xiao, X.; Xiao, Y. A novel Meloidogyne incognita effector Misp12 suppresses plant defense response at latter stages of nematode parasitism. Frontiers in Plant Science. 2016, 7, 964. [Google Scholar] [CrossRef]
- Blaazer, CJH. ; Villacis-Perez, EA.; Chafi, R.; Van Leeuwen, T.; Kant, MR.; Schimmel, BC. Why do herbivorous mites suppress plant defenses? Frontiers in plant science, 2018, 9, 1057. [Google Scholar] [CrossRef]
- Hettenhausen, C.; Li, J.; Zhuang, H.; Sun, H.; Xu, Y.; Qi, J.; Sun, G.; Wang, L.; Baldwin, lanT. ; Wu, J. Stem parasitic plant Cuscuta australis (dodder) transfers herbivory-induced signals among plants. Proceedings of the National Academy of Sciences. 2017, 114, E6703–E6709. [Google Scholar] [CrossRef]
- Hegenauer, V.; Fürst, U.; Kaiser, B.; Smoker, M.; Zipfel, C.; Felix, G.; Albert, M. Detection of the plant parasite Cuscuta reflexa by a tomato cell surface receptor. Science. 2016, 353, 478–481. [Google Scholar] [CrossRef]
- Saric-Krsmanovic, M.; Bozic, D.; Radivojevic, L.; Gajic Umiljendic, J.; Vrbnicanin, S. Response of alfalfa and sugar beet to field dodder (Cuscuta campestris Yunck.) parasitism: a physiological and anatomical approach. Canadian Journal of Plant Science. 2018, 99, 199–209. [Google Scholar] [CrossRef]
- Wolswinkel, P. Complete inhibition of setting and growth of fruits of Vicia faba L. resulting from the draining of the phloem system by Cuscuta species. Acta botanica neerlandica. 1974, 23, 48–60. [Google Scholar] [CrossRef]
- Ranjan, A.; Ichihashi, Y.; Farhi, M.; Zumstein, K.; Townsley, B.; David-Schwartz, R.; Sinha, NR. De novo assembly and characterization of the transcriptome of the parasitic weed dodder identifies genes associated with plant parasitism. Plant physiology. 2014, 166, 1186–1199. [Google Scholar] [CrossRef]
- Zagorchev, L.; Du, Z.; Shi, Y.; Teofanova, D.; Li, J. Cuscuta australis Parasitism-Induced Changes in the Proteome and Photosynthetic Parameters of Arabidopsis thaliana. Plants. 2022, 11, 2904. [Google Scholar] [CrossRef]
- Zhou, L.; Zawaira, A.; Lu, Q.; Yang, B.; Li, J. Transcriptome analysis reveals defense-related genes and pathways during dodder (Cuscuta australis) parasitism on white clover (Trifolium repens). Frontiers in Genetics. 2023, 14, 1106936. [Google Scholar] [CrossRef] [PubMed]
- Boex-Fontvieille, E.; Daventure, M.; Jossier, M.; Zivy, M.; Hodges, M.; Tcherkez, G. Photosynthetic control of Arabidopsis leaf cytoplasmic translation initiation by protein phosphorylation. PLoS one. 2013, 8, e70692. [Google Scholar] [CrossRef] [PubMed]
- Ayvacı, H.; Güldür, ME.; Dikilitas, M. Physiological and biochemical changes in lucerne (Medicago sativa) plants infected with ‘Candidatus Phytoplasma australasia’-related strain (16SrII-D Subgroup). The Plant Pathology Journal. 2022, 38, 146–10.5423. [Google Scholar] [CrossRef] [PubMed]
- Vrbničanin, SP.; Sarić-Krsmanović, MM.; Božić, DM. The effect of field dodder (Cuscuta campestris Yunck.) on morphological and fluorescence parameters of giant ragweed (Ambrosia trifida L.). Pesticides and Phytomedicine/Pesticidi i fitomedicina, 2013,1(1).
- Arce-Leal, Á.P.; Bautista, R.; Rodríguez-Negrete, E.A.; Manzanilla-Ramírez, M.Á.; Velázquez-Monreal, J.J.; Santos-Cervantes, M.E.; Méndez-Lozano, J.; Beuzón, C.R.; Bejarano, E.R.; Castillo, A.G.; et al. Gene Expression Profile of Mexican Lime (Citrus aurantifolia) Trees in Response to Huanglongbing Disease caused by Candidatus Liberibacter asiaticus. Microorganisms 2020, 8, 528. [Google Scholar] [CrossRef]
- Farah, A.F.; Al-Abdulsalam, MA. Effect of field dodder (Cuscuta campestris Yuncker) on some legume crops. Scientific Journal of King Faisal University (Basic and Applied Sciences). 2004, 5, 103–113. [Google Scholar]
- Heldt, HW.; Piechulla, B. Phenylpropanoids comprise a multitude of plant secondary metabolites and cell wall components. Plant biochemistry. 2011, 4, 431–449. [Google Scholar] [CrossRef]
- Runyon, J.B.; Mescher, M.C. , De Moraes, C. M. Plant defenses against parasitic plants show similarities to those induced by herbivores and pathogens. Plant signaling & behavior. 2010, 5, 929–931. [Google Scholar] [CrossRef]
- Albert, M.; Belastegui-Macadam, XM.; Bleischwitz, M.; Kaldenhoff, R. Cuscuta spp:“Parasitic plants in the spotlight of plant physiology, economy and ecology. In Progress in botany; Springer: Berlin, The Heidelberg, 2008; pp. 267–277. ISSN 978-3-540-72954-9. [Google Scholar]
- Koornneef, A.; Leon-Reyes, A.; Ritsema, T.; Verhage, A.; Den Otter, F. C.; Van Loon, LC.; Pieterse, CM. Kinetics of salicylate-mediated suppression of jasmonate signaling reveal a role for redox modulation. Plant physiology. 2008, 147, 1358–1368. [Google Scholar] [CrossRef] [PubMed]
- Assanga, SBI. ; Luján, LML.; Ruiz, JCG.; McCarty, MF.; Cota-Arce, JM.; Espinoza, CLL.; Salido, AAG.; Ángulo, DF. Comparative analysis of phenolic content and antioxidant power between parasitic Phoradendron californicum (toji) and their hosts from Sonoran Desert. Results in Chemistry. 2020, 2, 100079. [Google Scholar] [CrossRef]
- Guan, LM.; Scandalios, JG. Catalase gene expression in response to auxin-mediated developmental signals. Physiologia Plantarum. 2002, 114, 288–295. [Google Scholar] [CrossRef]
- Hiraga, S.; Sasaki, K.; Ito, H.; Ohashi, Y.; Matsui, H. A large family of class III plant peroxidases. Plant and Cell Physiology. 2001, 42, 462–468. [Google Scholar] [CrossRef]
- Yadav, V.; Wang, Z.; Wei, C.; Amo, A.; Ahmed, B.; Yang, X.; Zhang, X. Phenylpropanoid pathway engineering: An emerging approach towards plant defense. Pathogens. 2020, 9, 312. [Google Scholar] [CrossRef]
- Zhang, P.İ.; Jackson, E.; Li, X.; Zhang, Y. Salicylic acid and jasmonic acid in plant immunity. Horticulture Research. 2025, 12, uhaf082. [Google Scholar] [CrossRef]
- Li, C.; Xu, M.; Cai, X.; Han, Z.; Si, J.; Chen, D. Jasmonate signaling pathway modulates plant defense, growth, and their trade-offs. International Journal of Molecular Sciences. 2022, 23, 3945. [Google Scholar] [CrossRef]
- Chen, X.; Wang, DD.; Fang, X.; Chen, XY.; Mao, YB. Plant specialized metabolism regulated by jasmonate signaling. Plant and Cell Physiology. 2019, 60, 2638–2647. [Google Scholar] [CrossRef] [PubMed]
- Ghorbel, M.; Brini, F.; Sharma, A.; Landi, M. Role of jasmonic acid in plants: the molecular point of view. Plant Cell Reports. 2021, 40, 1471–1494. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Seomun, S.; Yoon, Y.; Jang, G. Jasmonic acid in plant abiotic stress tolerance and interaction with abscisic acid. Agronomy. 2021, 11, 1886. [Google Scholar] [CrossRef]
- Aziz, A.; Kapoor, D. Salicylic acid: it's physiological role and Interactions. Research Journal of Pharmacy and Technology. 2018, 11, 3171–3177. [Google Scholar] [CrossRef]
- de Freitas, PAF. , de Carvalho, HH.; Costa, JH.; de Souza Miranda, R.; da Cruz Saraiva, KD.; de Oliveira, FDB.; Gomes Filho, D.; Prisco, JT.; Gomes-Filho, E. Salt acclimation in sorghum plants by exogenous proline: physiological and biochemical changes and regulation of proline metabolism. Plant Cell Rep. 2019, 38, 403–416. [Google Scholar] [CrossRef]
- Warrier, RR. , Paul, M.; Vineetha, MV. Estimation of salicylic acid in Eucalyptus leaves using spectrophotometric methods. Genet Plant Physiol. 2013, 3, 90–97. [Google Scholar]
- Poór, P.; Borbély, P.; Bódi, N.; Bagyánszki, M.; Tari, I. Effects of salicylic acid on photosynthetic activity and chloroplast morphology under light and prolonged darkness. Photosynthetica. 2019, 57, 367–376. [Google Scholar] [CrossRef]
- Rossi, CA.; Marchetta, EJ.; Kim, JH.; Castroverde, CDM. Molecular regulation of the salicylic acid hormone pathway in plants under changing environmental conditions. Trends in Biochemical Sciences. 2023, 48, 699–712. [Google Scholar] [CrossRef]
- Berim, A.; Gang, DR. Accumulation of salicylic acid and related metabolites in Selaginella moellendorffii. Plants. 2022, 11, 461. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, PP. Plant hormone-mediated regulation of stress responses. BMC plant biology. 2016, 16, 86. [Google Scholar] [CrossRef]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, LJ. Signaling crosstalk between salicylic acid and ethylene/jasmonate in plant defense: do we understand what they are whispering? International journal of molecular sciences. 2019, 20, 671. [Google Scholar] [CrossRef]
- Albert, M.; Axtell, MJ.; Timko, MP. Mechanisms of resistance and virulence in parasitic plant–host interactions. Plant physiology 2021, 185, 1282–1291. [Google Scholar] [CrossRef]
- Takagawa, M.; Yokoyama, R. Current understanding of the role of the cell wall in Cuscuta parasitism. Plant Biology. 2025. [CrossRef] [PubMed]
- Turan, P.; Şentürk, G.E.; Ercan, F. Cryopreservation triggers DNA fragmentation and ultrastructural damage in spermatozoa of oligoasthenoteratozoospermic men. Marmara Medical Journal. 2017, 30, 63–72. [Google Scholar] [CrossRef]
- Kaya, M.; Çavuşoğlu, K.; Yalçin, E.; Acar, A. DNA fragmentation and multifaceted toxicity induced by high-dose vanadium exposure determined by the bioindicator Allium test. Scientific Reports. 2023, 13, 8493. [Google Scholar] [CrossRef] [PubMed]
- Plitta-Michalak, B.P.; Ramos, A.; Stępień, D.; Trusiak, M.; Michalak, M. PERSPECTIVE: The comet assay as a method for assessing DNA damage in cryopreserved samples. CryoLetters. 2024, 45, 1–15. [Google Scholar] [CrossRef]
- Demirbas, S.; Vlachonasios, KE.; Acar, O. , & Kaldis, A. The effect of salt stress on Arabidopsis thaliana and Phelipanche ramosa interaction. Weed Research. 2013, 53, 452–460. [Google Scholar] [CrossRef]
Figure 1.
Evaluation of stem length measurements of Cuscuta sp.-infected lucerne (Medicago sativa L. cv. Alsancak) plants at 90th, 105th, 120th, and 135th days post-sowing. Different letters above the bars indicate statistically significant differences from the control plants based on the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
Figure 1.
Evaluation of stem length measurements of Cuscuta sp.-infected lucerne (Medicago sativa L. cv. Alsancak) plants at 90th, 105th, 120th, and 135th days post-sowing. Different letters above the bars indicate statistically significant differences from the control plants based on the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
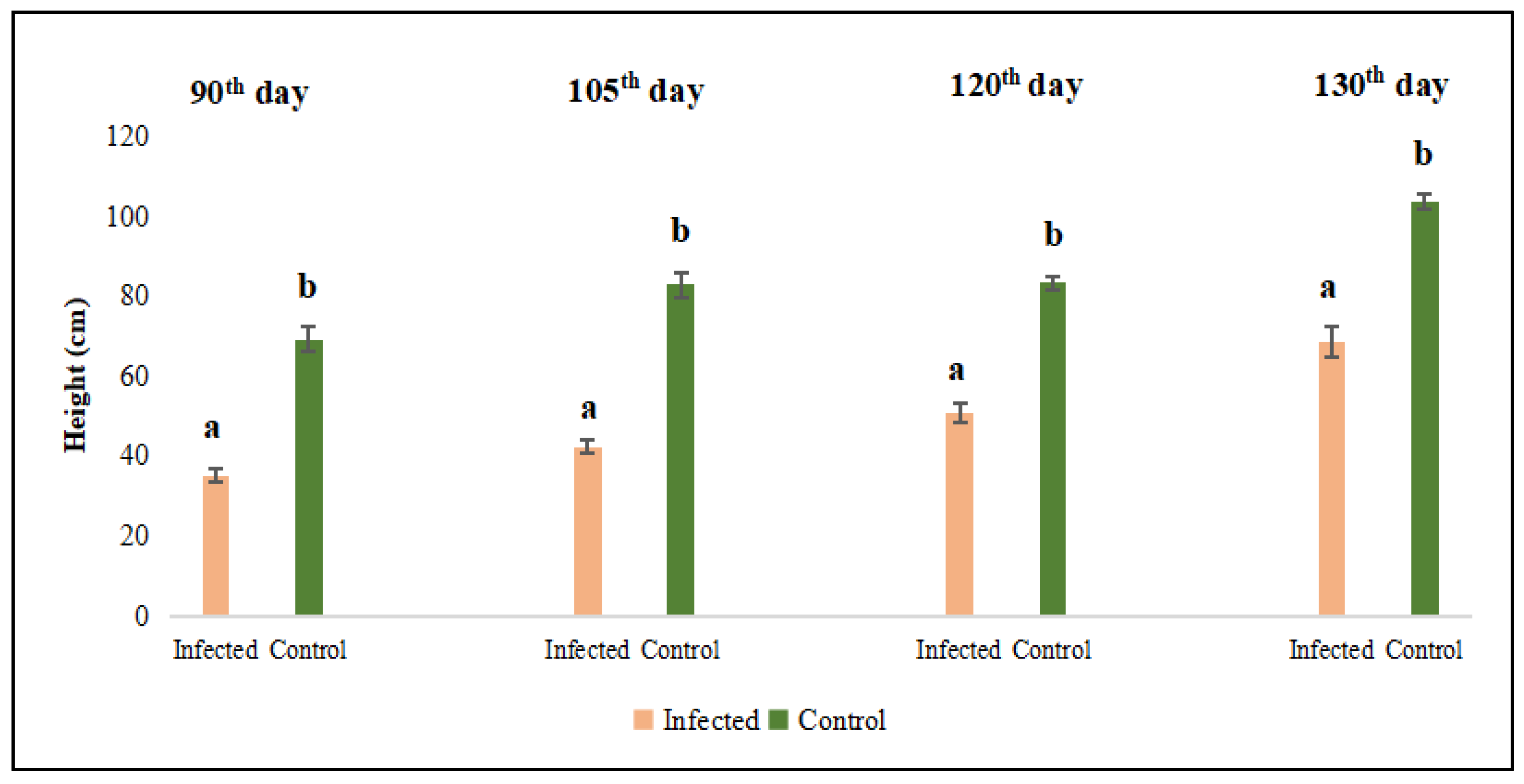
Figure 2.
Evaluation of fresh weight of lucerne (Medicago sativa L. cv. Alsancak) plants infected with Cuscuta sp. at 90th , 105th, 120th, and 135th days after sowing. Different letters above the bars indicate statistically significant differences from the control plants according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
Figure 2.
Evaluation of fresh weight of lucerne (Medicago sativa L. cv. Alsancak) plants infected with Cuscuta sp. at 90th , 105th, 120th, and 135th days after sowing. Different letters above the bars indicate statistically significant differences from the control plants according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
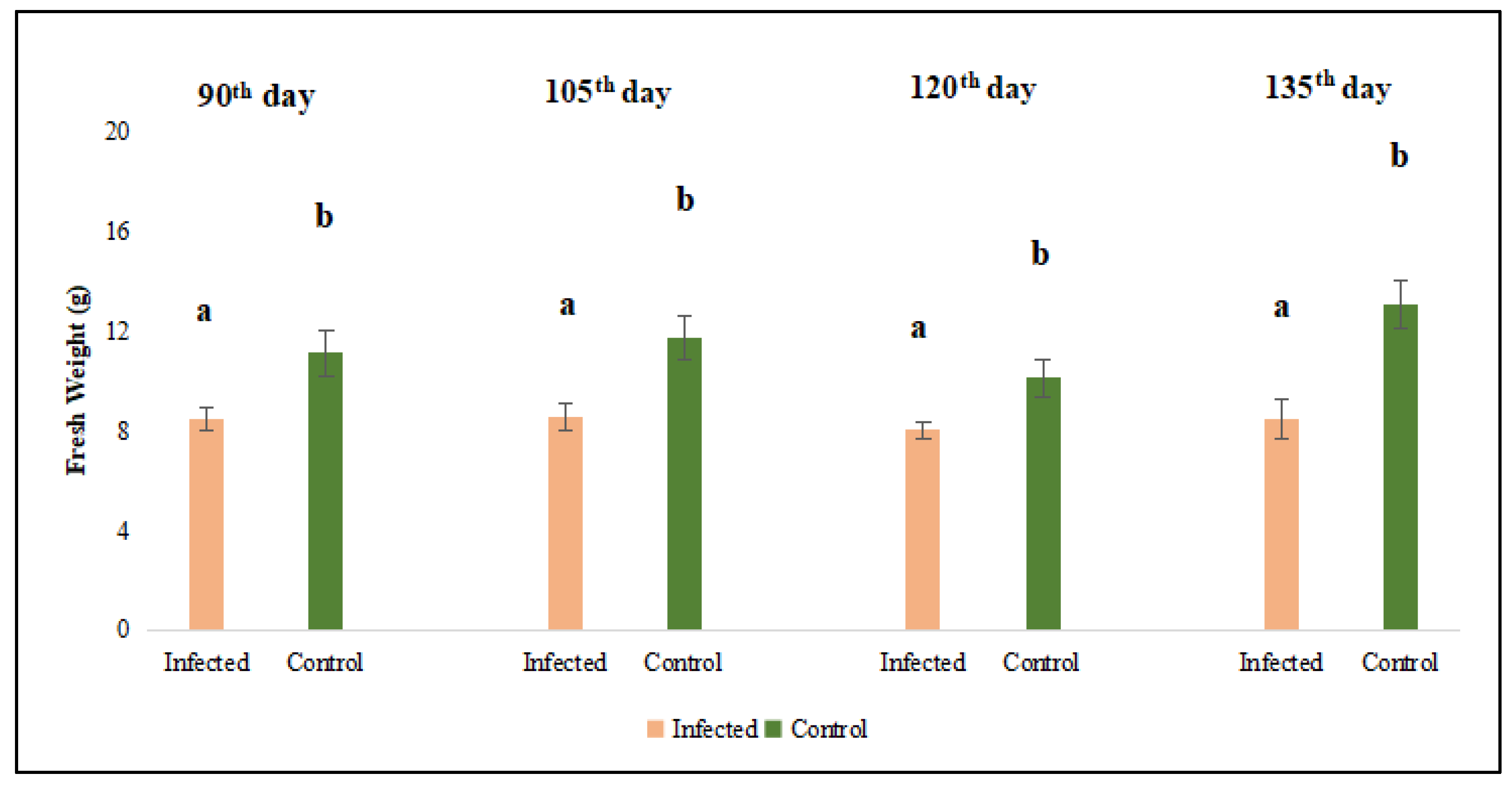
Figure 3.
Evaluation of dry weight of lucerne (Medicago sativa L. cv. Alsancak) plants infected with Cuscuta sp. at 93th, 108th, 123th, and 138th days after sowing. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
Figure 3.
Evaluation of dry weight of lucerne (Medicago sativa L. cv. Alsancak) plants infected with Cuscuta sp. at 93th, 108th, 123th, and 138th days after sowing. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
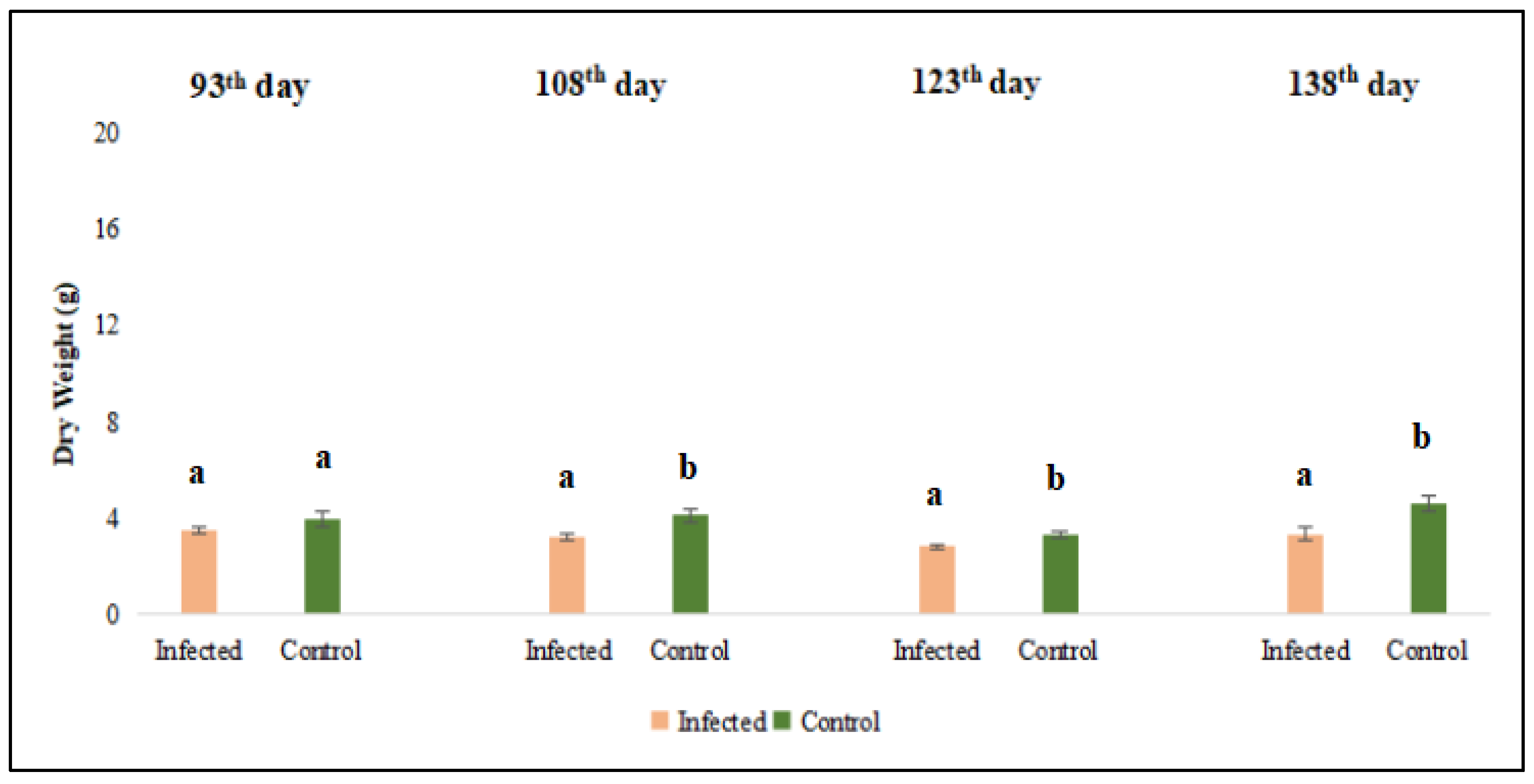
Figure 4.
Total Chlorophyll was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
Figure 4.
Total Chlorophyll was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
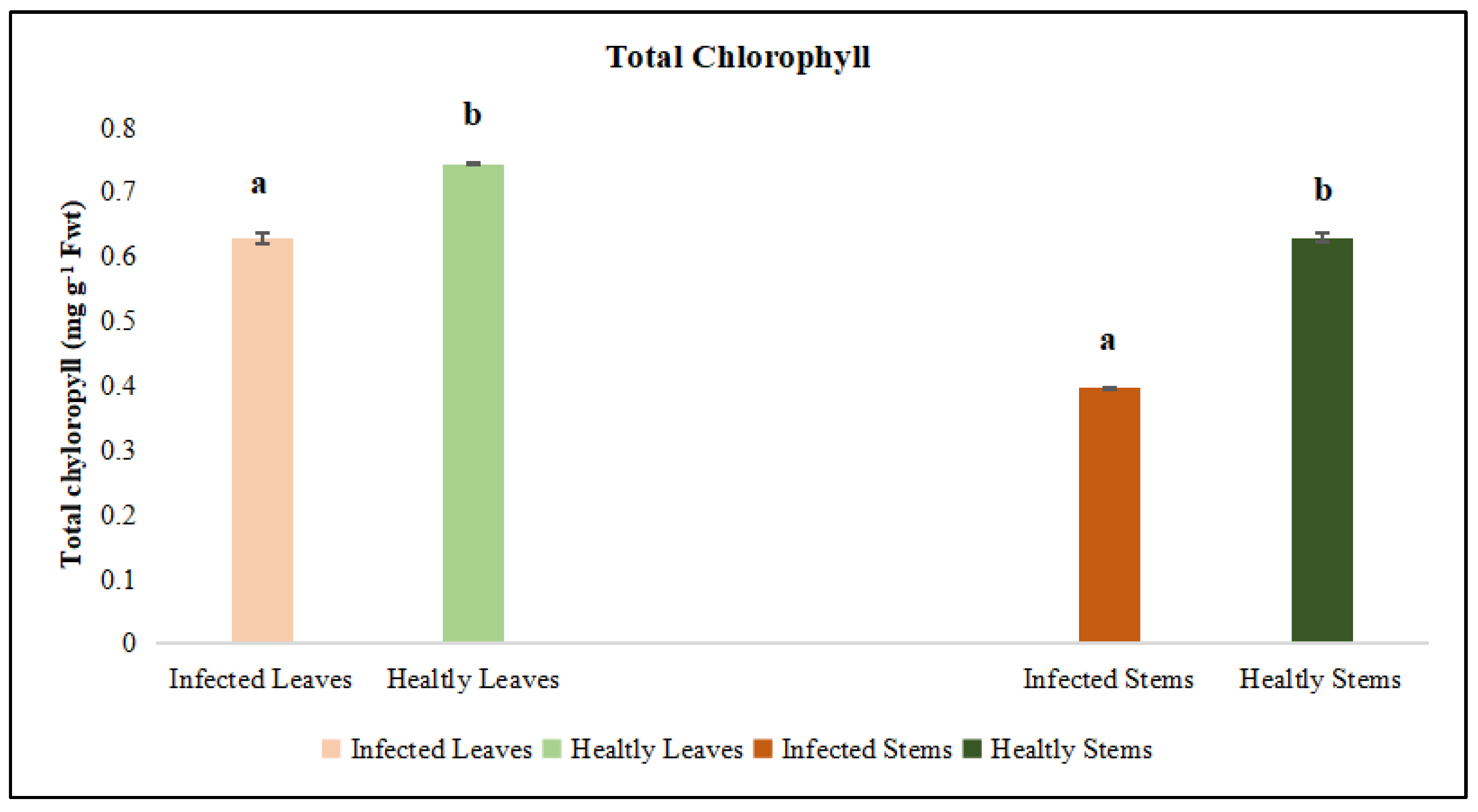
Figure 5.
Chlorophyll a was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
Figure 5.
Chlorophyll a was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
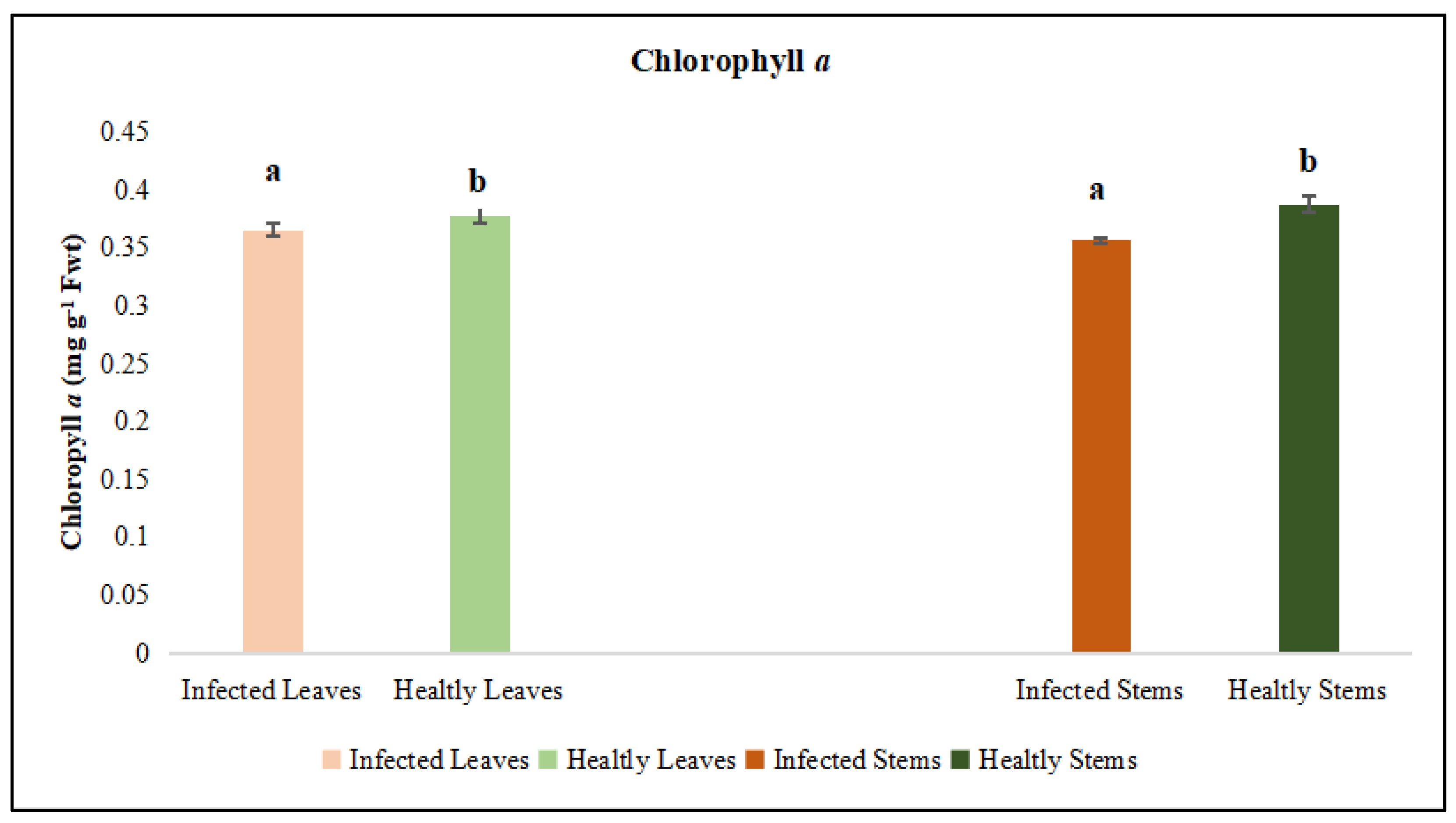
Figure 6.
Chlorophyll b was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
Figure 6.
Chlorophyll b was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
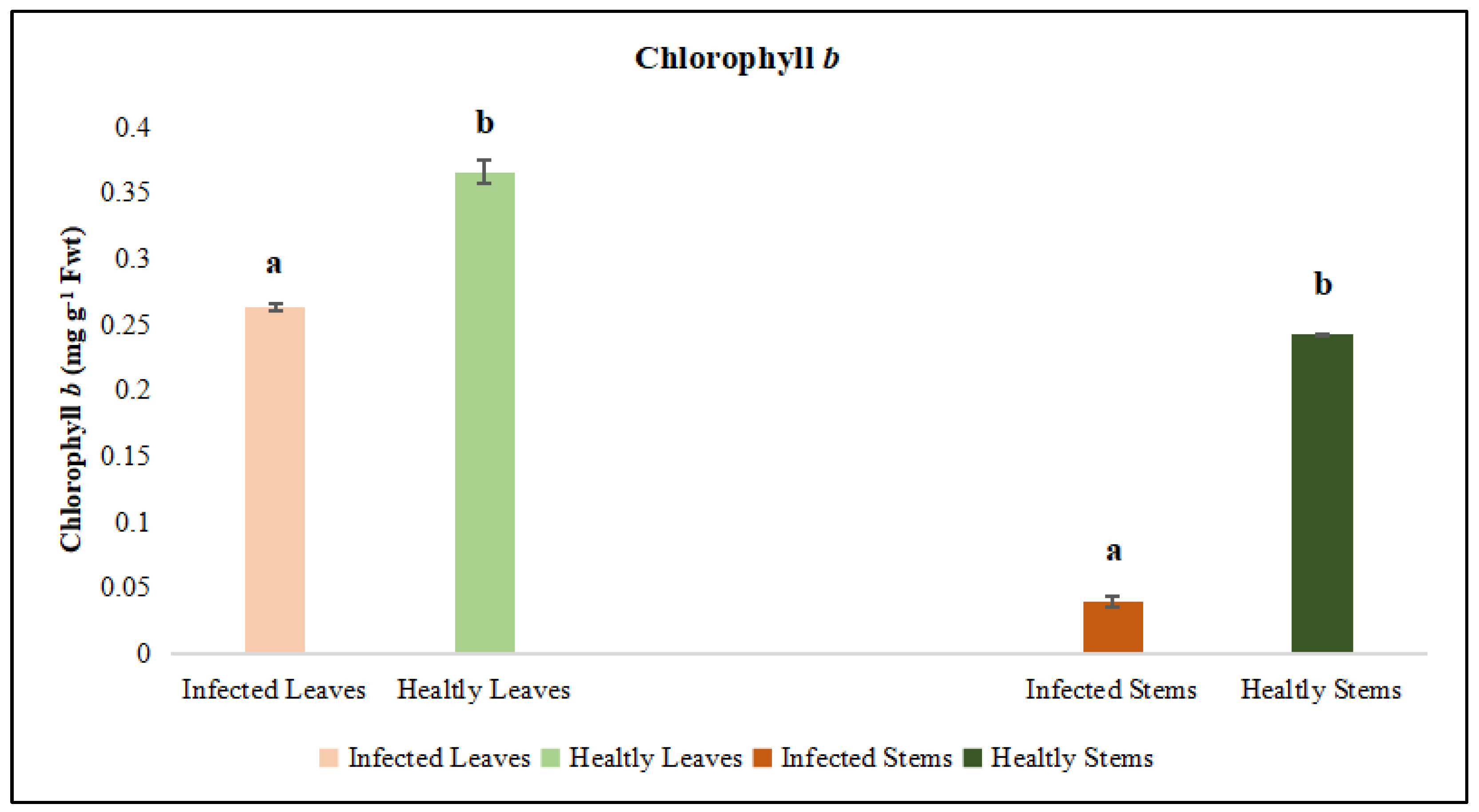
Figure 7.
Protein was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
Figure 7.
Protein was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
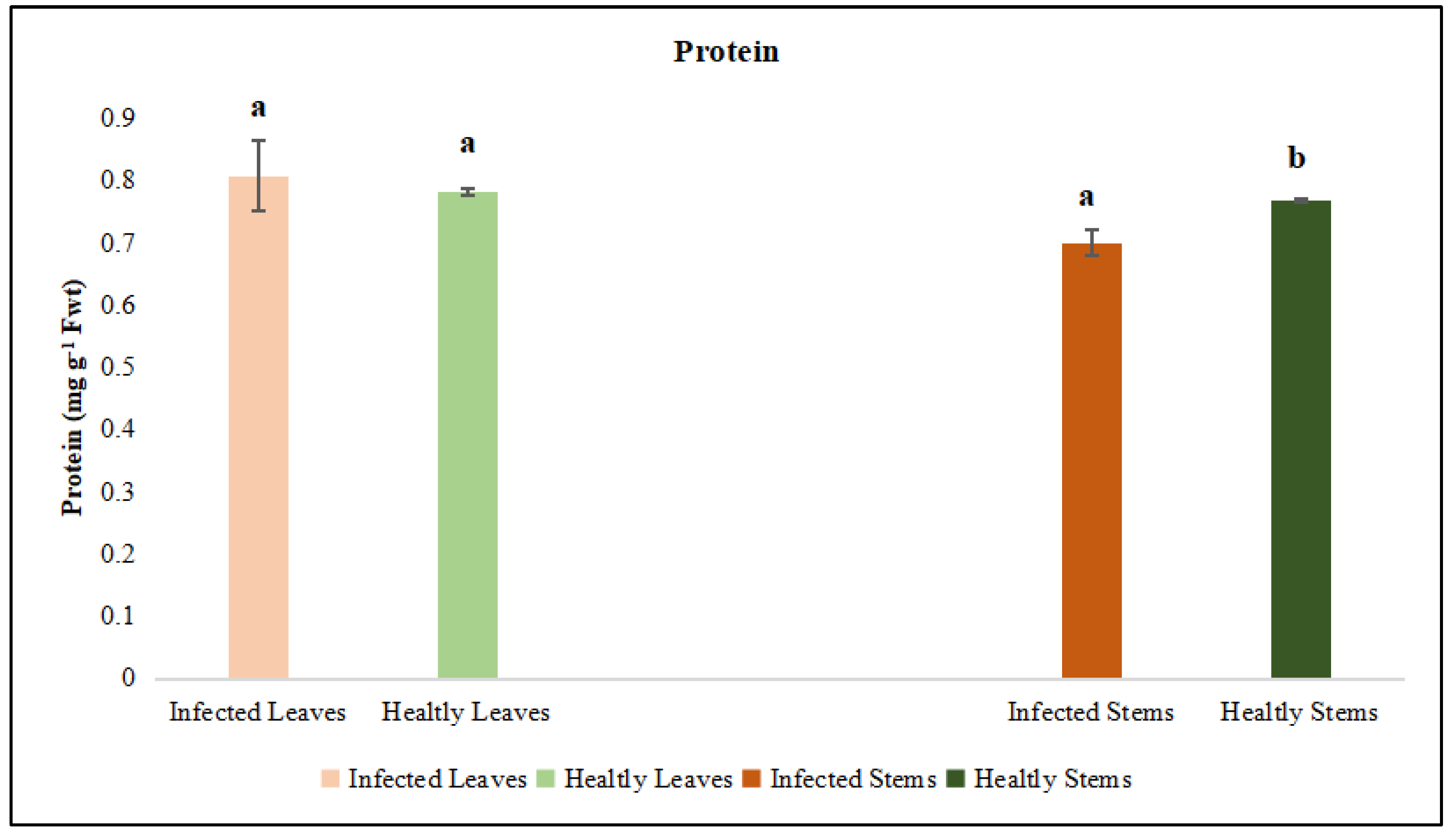
Figure 8.
Phenol was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
Figure 8.
Phenol was evaluated in the leaf and stem tissues of lucerne (Medicago sativa L. cv. Alsancak) infected with Cuscuta sp. Different letters above the bars indicate statistically significant differences according to the Least Significant Difference (LSD) test at P ≤ 0.05 (n = 3).
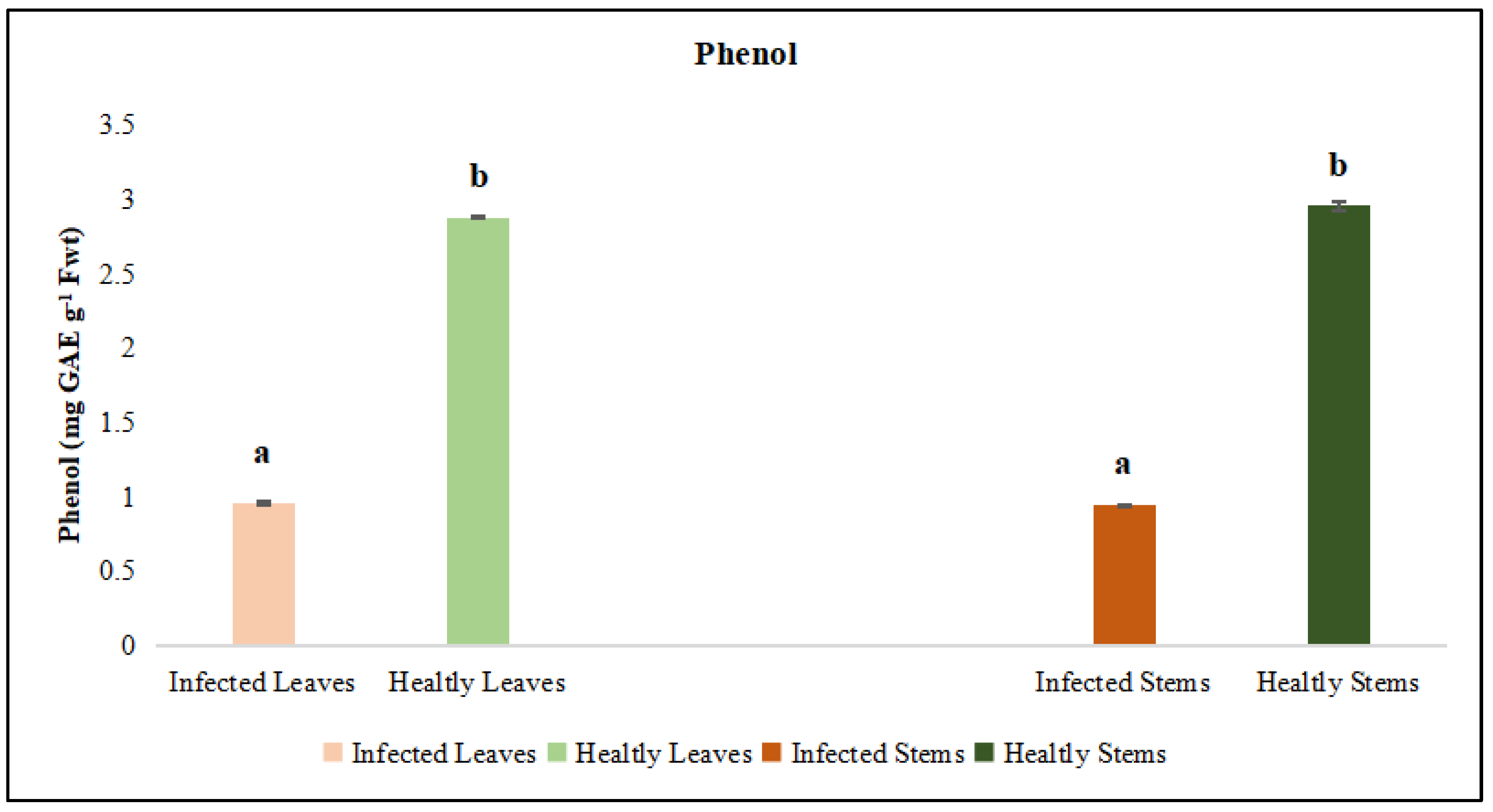
Figure 16.
DNA fragmentation in Medicago sativa L. cv. Alsancak followed by the infection of Cuscuta sp. plants. O’GeneRuler 100 bp DNA Ladder Plus was used to determine DNA fragmentation, 100 µg DNA was loaded for each well, and a size standard is shown on the left; a) uninfected Medicago sativa L. cv. Alsancak b) Medicago sativa L. cv. Alsancak infected with Cuscuta sp. c) Cuscuta sp.
Figure 16.
DNA fragmentation in Medicago sativa L. cv. Alsancak followed by the infection of Cuscuta sp. plants. O’GeneRuler 100 bp DNA Ladder Plus was used to determine DNA fragmentation, 100 µg DNA was loaded for each well, and a size standard is shown on the left; a) uninfected Medicago sativa L. cv. Alsancak b) Medicago sativa L. cv. Alsancak infected with Cuscuta sp. c) Cuscuta sp.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



