
Submitted:
22 October 2025
Posted:
23 October 2025
You are already at the latest version
Abstract
This study aimed to determine the association between the content of flavonoids and an-14 tiviral effect of plant extracts against the dengue virus (DENV). Extracts from medicinal plants cul-15 tivated in Colombia were prepared by ultrasonic-assisted solvent extraction (UAE) and supercritical 16 fluid extraction (SFE). UHPLC/ESI-Q-Orbitrap-MS analysis identified forty-six flavonoids. Fourteen 17 extracts were tested for their ability to reduce the cytopathic effect (CPE) induced by DENV in Vero 18 cells. UAE extracts of Scutellaria coccinea, Scutellaria incarnata, and Lippia alba contained higher 19 amounts of flavonoid glycosides (from 97.0% to 87.9%) rather than aglycones, and effectively re-20 duced CPE- DENV (IC50: 3.0 to 65 μg/mL; SI: 0.4 to 71). UAE and SFE extracts from L. origanoides 21 with higher content of aglycones (41.7% to 93.4%) than glycosides (0.0 to 58.3%) did not reduce the 22 CPE- DENV. Cluster and one-way ANOVA analyses indicated that strong antiviral effect is associ-23 ated with increased levels of flavone glycosides in the extract. Docking analyses (AutoDock Vina) 24 revealed that flavonoid glycosides exhibited better binding affinity to target proteins (E, Gas6-Axl, 25 clathrin, and dynamin) than aglycones. This study establishes a scientific basis for using extracts 26 rich in flavonoid glycosides, particularly flavones, as starting points for developing plant-based 27 therapies to treat dengue.

Keywords:
dengue virus
; dengue treatment
; plant extracts
; flavonoids
1. Introduction
Dengue virus (DENV) causes a spectrum of disease ranging from asymptomatic infection to dengue fever and severe dengue shock syndrome [1]. According to the Pan American Health Organization, dengue fever increased significantly in the Americas in 2024 rising by 232% compared to 2023 and 421% compared to the average of the previous five years [2]. There are currently no specific antiviral treatments for dengue, so therapy is primarily supportive [3]. Although two vaccines have been approved in some countries, their use is constrained due to safety concerns and their limited effectiveness against all DENV serotypes [4]. Dengue is a complex disease, making the discovery of effective therapies challenging. Increasing evidence suggests that herbal medicines have the potential as sources for clinical dengue treatments [5]. Administration of such medicines after virus exposure has shown to reduce the risk of severe dengue [5,6]. Thus, analyzing plant extracts has become an important strategy to discover and develop alternative therapies for dengue [5,7].
Plant-derived extracts prepared from a variety of species cultivated in different countries have demonstrated antiviral properties against human viruses, including DENV [5,6,7]. Their antiviral effect varies according to the plant species, chemotype, and phenological stages. The chemical composition of plant extracts depends on both the species and the extraction technique used which, in turn, affects their antiviral activity [7,8,9]. Diverse secondary metabolites, especially phenolic compounds such as flavonoids, have shown the potential to inhibit a variety of human pathogenic viruses, including influenza H1N1, HIV, and Zika [7,8,9]. Further research is necessary to fully elucidate chemical profiles that explain the anti-DENV activity of plant extracts. Studies demonstrate that plant-derived flavonoids have antiviral properties and can inhibit DENV by interfering with viral replication, entry, or specific viral proteins [10,11].
The genus Scutellaria (Lamiaceae) includes about 400 species widely distributed across America, Europe and Asia [12]. S. baicalensis is recognized as a promising natural source of compounds for treating viral diseases [13]. The genus Lippia (Verbenaceae) comprises 150 species, primarily found in the Americas, tropical Africa and India [14]. Lippia alba (Mill.) N.E. Brown was the first plant approved by the French Medicines Agency for inclusion in the French Pharmacopoeia [15]. L. origanoides Kunth was included in the Formulário de Fitoterápicos da Farmacopéia Brasileira due to its strong antibacterial properties [16]. L. alba and L. origanoides are traditionally used to treat viral infections such as influenza, measles and gastroenteritis [17,18]. Extracts prepared from S. baicalensis and L. origanoides have demonstrated inhibitory effect on the replication of DENV and other enveloped viruses [19,20]. We have investigated biological activities of both Lippia species and Scutellaria species cultivated in Colombia, and their extract chemical composition have been documented [20,21,22,23,24].
Dengue has been identified as a significant and persistent public health concern in Colombia, which is among one of the most affected countries in the Americas. From 2012 to 2020, annual incidence rate ranged from 90.7 to 476.2 per 100,000 population, with the most recent outbreak reported in 2019 (465.9 per 100,000 population) [25]. Traditional Colombian herbal remedies are widely used to alleviate dengue symptoms, although few plants have been systematically studied for their therapeutic potential [26]. Research on the anti-DENV activity of plant extracts contributes to discovering alternative therapies to prevent severe dengue. In this study, we evaluated the in vitro anti-DENV activity of extracts from Scutellaria species and L. alba and L. origanoides and investigated the influence of their flavonoid content. In silico analyses were also performed to hypothesize how flavonoid in the extracts might interfere with DENV entry into Vero cells.
2. Results
2.1. Cytotoxicity
Fourteen extracts from Scutellaria, L. alba, and L. origanoides were selected for analysis, including eight prepared in this study and six in previous studies [20,21,22]. The characteristics of the extracts are summarized in Table 1, categorized by plant species, chemotype, and extraction technique. Eleven extracts were prepared using ultrasound-assisted extraction (UAE) at different temperatures (50 °C and 47 °C) and extraction time (60, 23, and 15 minutes). Three extracts from L. origanoides were prepared using supercritical fluid extraction (SFE).
Cytotoxicity of the extracts was assessed in Vero cells uninfected with DENV using crystal violet assays. Assays were carried out at corresponding time points as the antiviral assay to determine non-cytotoxic concentrations for the cell. All extracts were non-toxic at 100 µg/mL, the maximum concentration used for antiviral assays (Figure 1A). Additional assays following the standard protocol determined the 50% cytotoxicity concentration (CC50). Extracts from Scutellaria and L. alba, along with two extracts from L. origanoides (LopSE1 and LocSE2) reduced cell viability with CC50 values ranging from 23 µg/mL to 213 µg/mL (Figure 1B). The extract from S. ventenatii + incarnata (SivSE) and one extract from S. incarnata (SiSE1) exhibited the highest cytotoxicity (CC50: 23 and 50 µg/mL). The remaining six L. origanoides extracts showed no cytotoxicity at the maximum concentration tested, suggesting CC50 values greater than 250 µg/mL.
2.2. Antiviral Effect
Virus-induced cytopathic effect (CPE) is a surrogate measure of virus replication in vitro; therefore, lower DENV-induced CPE indicates stronger antiviral activity [27]. The extracts were tested at six nontoxic concentrations to evaluate their effect on CPE caused by DENV-1 and DENV-2 after treatment during virus adsorption to Vero cells. The presence of DENV in untreated cells was confirmed by detection of non-structural protein 1 (NS1 = 75 ± 1.97 PanBio units) using an ELISA kit. Antiviral effects were classified as: (i) strong, greater than 50% reduction in CPE of both virus serotypes with IC50 values lower than 55 µg/mL; (ii) weak, IC50 ≤ 55 µg/mL for only one serotype or IC50 between 55 and 100 µg/ml for one or both serotypes; and (iii) inactive, no significant CPE reduction in either serotype. Selectivity index (SI) was included in the analysis; extract with IC50 ≤ 55 µg/mL and SI ≥ 3.0 was considered as promising anti-DENV sample. The extracts exhibited variable anti-DENV effect and selectivity (Figure 2 and Table 2).
Extracts from S. coccinea (ScSE) and S. incarnata (SiSE1) exhibited strong anti-DENV effect. ScSE showed IC50 values < 53 µg/mL and SI values > 4.0, indicating promising anti-DENV sample. SiSE1 showed IC50 values < 40 µg/mL, but SI values < 3.0, suggesting reduced antiviral promise. The other S. incarnata extract (SiSE2) was inactive, significant reduction of the virus-induced CPE of both serotypes was not detected. The S. ventenatti + incarnata (SviSE) extract exhibited weak antiviral effect, reduced DENV-2 at IC50 of 65 µg/mL (SI of 0.4), but not DENV-1. Extracts from L. alba exhibited weak antiviral effect. LaiSE (citral chemotype) reduced the replication of both virus serotypes, but at IC50 of 81 µg/mL against DENV-1. LacSE (carvone chemotype) showed antiviral promise against DENV-2 (IC50 of 19 µg/mL and SI of 3.9), but was inactive against DENV-1. All eight extracts from L. origanoides were inactive against both virus serotypes.
2.3. UHPLC/ESI-Q-Orbitrap-MS Analysis of the Extracts Prepared in This Study
The analysis, based on retention times (Rt) and fragmentation patterns, identified only compounds belonging to the flavonoid class, including glycosides (n = 19), aglycones (n = 19) and methylated (n = 3) (Table 3 and Table 4).
Extracts from Scutellaria (UAE, 60 min at 50 °C) had higher concentrations of flavonoid glycosides than aglycones. Nine flavonoids were detected in S. incarnata (SiES2), with glycosides accounting for 64.7% (55-85 mg/g) of the total content. The predominant flavonoids were dihydrobaicalein-glucuronide, scutellarin, baicalin, baicalein, and wogonin. The flavonoid content of the S. ventenatti + S. incarnata (SviSE) extract was similar to that of SiES2, except for the absence of dihydrobaicalein-glucuronide and dihydrobaicalein. Flavonoid glycosides accounted for 55% (32/58 mg/g) of the SviSE content.
L. alba extracts (UAE, 60 min at 50 °C) had high flavonoid glycosides content, exceeding levels reported in extracts of the same species cultivated in Brazil [32], which may be attributed to variations in extraction methodology and cultivation practice of the plant. In the LaiSE (citral chemotype) extract were identified eight flavonoid glycosides accounting for 90.7% (16.6/18.3 mg/g) of total content. Tricin-7-diglucuronide, chrysoeriol-7-diglucuronide, chrysoeriol-7-glucuronide, tricin-glucuronide, and apigenin-7-glucuronide were predominant. Seven flavonoids aglycones were also identified, five from which were below detection limits. The LacSE (carvonal chemotype) extract exhibited a comparable profile, yet with higher amounts of flavonoid glycosides (97.1%, 20.7 / 21.3 mg/g).
L. origanoides extracts had higher flavonoid aglycone content than the other test extracts. Notable differences were observed in extracts from the carvacrol chemotype. LocSE1 (UAE, 23 min at 47 °C) contained twenty-four flavonoids, 82.3% (11.2/13.6 mg/g) of which were aglycones. Eriodictyol, naringenin, luteolin, quercetin, and cirsimaritin were predominant. The LocSE2 extract (UAE, 60 min at 50 °C) contained twelve flavonoids in higher amounts (259 mg/g vs. 13.6 mg/g), with aglycones accounting for 41.7%. The predominant flavonoids were eriodictyol, eriodictyol-7-glucoside, and luteolin-7-glucoside. The thymol chemotype LotSE (UAE, 23 min at 47 °C) extract contained twenty-three flavonoids. Aglycones accounted for 73.1% (42/57.4 mg/g), with eriodictyol, quercetin, and naringenin as the predominant. The phellandrene chemotype LopSE2 (UAE, 60 min at 50 °C) extract contained fourteen flavonoids, with aglycones accounting for 61.5% (228/371 mg/g) with eriodictyol, pinocembrin, and galangin as the predominant.
2.4. Relationship Between Anti-DENV Effect and the Flavonoid Content
Fourteen extracts listed in Table 1 were included in the analysis. The chemical compositions of six extracts that were analyzed in previous studies are shown in Table S1. The relationship between antiviral effect and the flavonoid content was investigated to test the hypothesis that extracts exhibiting anti-DENV effect (strong or weak) possess distinct chemical profiles compared to inactive extracts. Table 5 compares flavonoid classes identified in the fourteen extracts.
Extracts with the strongest anti-DENV effects (ScSE and SiSE1) contained high amounts of flavonoids (278 mg/g and 493 mg/g, respectively). Of these flavonoids, 83.4% and 86.1%, respectively, corresponded to flavone glycosides, with baicalin, dihydrobaicalein-glucuronide, and scutellarin being the predominant flavones. ScSE and SiSE1 also had small amounts of flavone aglycones (10.3% and 5%, respectively) and non-flavonoid compounds (6.4% and 10.5%), including verbascoside and umbelliferone-hexoside-pentoside (Table S1). In contrast, extracts with weak anti-DENV effects (SviSE, LacSE, and LaiSE) had significantly lower amounts of flavonoid (from 18.3 mg/g to 57.9 mg/g), though most of these were flavone glycosides (56.5%, 97%, and 90.7%, respectively). Among the extracts with anti-DENV effect, SviSE contained a significantly higher concentration (44.5%) of flavonoid aglycones, all of them belonging to the flavone class. The S. incarnata SiSE2 extract, lacking anti-DENV effect, had a chemical profile comparable to SviSE, except for the lack of dihydrobaicalein.
Extracts from L. origanoides lacking anti-DENV effect had the lowest concentrations of flavone glycosides (0 to 23.9%) but the highest of aglycones (61.5% to 98.1%), especially flavanone and flavanol + flavanonol, which were absent in extracts showing anti-DENV effects. Furthermore, UAE extracts (LopSE1, LopSe2, LocSE1, and LocSE) exhibited higher levels (17% to 34.4% vs. 0 to 4.6%) of flavonoid glycosides distinct to flavones, while SFE extracts lacked of flavonoid glycosides and contained small amounts of methylated flavonoids. The UAE-LopSE1 extract (phellandrene chemotype) had higher levels of flavonoid glycosides (77.4%) than aglycones (22.2%) compared to other L. origanoides extracts.
A Kohonen’s self-organized map algorithm, using percentages of flavonoid classes as input, grouped the fourteen extracts into two clusters (Figure 3A). Cluster 1 included five anti-DENV active extracts (Scutellaria and L. alba) along with the inactive S. incarnata extract (SiSE2). These extracts shared the highest flavone glycosides content and lowest flavonoid aglycones. Cluster 2 grouped seven inactive L. origanoides extracts marked by elevated flavonoid aglycones levels. The LopSE1 extract from L. origanoides was not included in cluster 2, as would have been expected, due to its distinct chemical profile. Figure 3B and 3C show that CPE values for DENV-1 and DENV-2 were significantly lower in cluster 1 compared to cluster 2 (Wilcoxon test, p < 0.01). This suggests a plausible relationship between the anti-DENV activity and a flavonoid profile characterized by a higher content of flavonoid glycosides, especially flavones, relative to flavonoid aglycones.
2.5. Molecular Interactions Between Flavonoids and DENV and Vero Cell Proteins
DENV infection of Vero cells is initiated by the interaction between its envelope (E) protein and specific cell surface receptors, including Axl receptor tyrosine kinase [34]. The virus enters cells via a non-classical endocytic pathway mediated by the clathrin and dynamin proteins [35,36,37]. To explore the role of flavonoids in the anti-DENV effect, an AutoDock Vina software was used to dock forty-six flavonoids found in fourteen extracts to the DENV-E, Gas6-Axl complex, clathrin, and dynamin. Binding energy values of ≤-7.55 kcal/mol indicate binding affinity between ligand and target. Figure 4 shows the relative concentrations of flavonoids in the extracts and their binding affinities to proteins. As expected, the analysis revealed varied binding affinities to targets. Generally, glycosides exhibited higher binding affinities (energy ranging from -10.22 to -8.01 kcal/mol) compared to aglycones (-8.23 to -7.90 kcal/mol), a difference that can be influenced by the presence of sugar. Most flavonoid glycosides exhibiting the lowest binding energies were the most abundant in extracts with anti-DENV effect.
The DENV-E protein structure consists of three domains [33], DI and DII domains form the DI/DII hinge region containing a conserved fusion loop. Conformational changes in this region lead to the fusion of the viral and cell endosomal membranes [37]. Nineteen glycosides (-9.16 to -7.72 kcal/mol) and fourteen aglycones (-8.06 to -7.55 kcal/mol) were predicted to bind to E. As for flavonoids found in extracts with antiviral effect, all flavone glycosides exhibited affinity (-8.78 to -7.72 kcal/mol) for E, except schaftoside. The lowest binding energies were exhibited by chrysoeriol-7-diglucuronide, chrysoeriol-7-glucuronide, luteolin-7-glucuronide, apigenin-7-glucuronide, and tricin-7-diglucuronide, all of which are predominant in L. alba extracts. All flavonoid aglycones, except cirsimaritin, exhibited affinity for E; the predominant baicalein, dihydrobaicalein, and apigenin showed the lowest energies (-7.96 to -7.84 kcal/mol). As for flavonoids present in extracts lacking antiviral effect (L. origanoides), 50% glycosides non-flavones (-8.08 to -7.73 kcal/mol) and 50% aglycones (-8.05 to -7.73 kcal/mol) exhibited affinity for E. Eriodyctiol-7-glucoside, eriodyctiol, and pinocembrin had the lowest binding energies. Flavonoids formed hydrogen bonds with amino acid residues at three consensus binding sites (Figure 5). Most glycosides were accommodated within the DII domain closer to the dimer interface (A/B), with the remaining glycosides located within the interface DIII/DI domains closer to the fusion loop. Most aglycones were accommodated within the detergent β-octyl glucoside (βOG) site in the hinge region, with the remaining aglycones located within the DII (A/B) interface.
DENV can bind to the Axl receptor via the Gas6 protein, which facilitates virus entry by bridging the viral envelope's phosphatidylserine to Axl [34]. Gas6 has two domains (Lg1 and Lg2) that interact with Axl domains (Ig1 and Ig2). Sixteen flavonoid glycosides were predicted to bind to Gas6 (-8.66 to -7.56 kcal/mol), whereas no aglycones (-7.52 to -6.83 kcal/mol) exhibited binding affinity. All flavone glycosides found in anti-DENV extracts exhibited affinity (-8.69 to -7.56 kcal/mol) for Gas6, except for schaftoside. The lowest binding energies were exhibited by chrysoeriol-7-diglucuronide, tricin-7-diglucuronide, luteolin-7-glucuronide, and luteolin-7-glucoside. In contrast, only two (eriodyctiol-7-glucoside and eriodyctiol-rhamnoside) from six glycosides non-flavones found in inactive extracts were predicted to bind to Gas6 (-7.77 kcal/mol and -7.76 kcal/mol, respectively). Flavonoid glycosides formed hydrogen bonds with amino acid residues at five consensus binding sites (Figure 6). Most flavonoids were located within the interface of Gas6-Lg2/Lg1 domains in which each of the monomers. The remaining flavonoids were accommodated within the Gas6-Lg1 domain, and the interface of the Gas6-Lg1/Axl domains within each of the monomers.
Dynamin possess a GTPase domain that binds to and hydrolyzes guanosine-5'-triphosphate (GTP), causing a conformational change crucial for its function in DENV and cell membranes fusion [39]. Small molecules targeting GTPase can interfere with virus internalization [39]. All twenty-three flavonoid glycosides (-9.96 to -7.68 kcal/mol), except quercetin-3-glucoside, were predicted to bind to GTPase. The glycosides with the lowest binding energies were chrysoeriol-7-diglucuronide, baicalin, isoliquiritin, apigenin-7-glucoside, and chrysoeriol-7-glucuronide. These flavonoids are found in anti-DENV extracts but not in inactive L. origanoides extracts. Seventeen aglycones exhibited binding affinity for GTPase (-8.26 to -7.73 kcal/mol). Baicalein, dihydrobaicalein, luteolin, and scutellarein had the lowest binding energies. The flavonoids formed hydrogen bonds with amino acid residues of five consensus binding sites (Figure 7). Most glycosides were accommodated at the G4 loop extending to the switch1 loop (G4-switch1 loops), while others were accommodated within the P-loop and a few at the switch1 or switch2, closer to the P-loop (switch1-P-loops and switch2-P-loops). Most aglycones were accommodated at the switch2-P-loops and G4-switch1-loops.
Clathrin is a transport protein whose N-terminal domain (CTD) has various peptide-binding sites, including the PWDLW motif (W-box) [38]. CTD has been proposed as a target for antiviral agents that hinder viral entry into the cell [40]. All of the flavonoid glycosides were predicted (-10.01 to -8.08 kcal/mol) to bind to CTD, except for quercetin-3-glucoside. The lowest binding energies were observed for chrysoeriol-7-diglucuronide, triicin-7-diglucuronide, apigenin-7-glucoside, baicalin, and scutellarin. All flavonoid aglycones except wogonin exhibited binding affinity for CTD (-8.22 to -7.75 kcal/mol). Luteolin, eriodictyol, hesperetin, nepetin, and baicalein had the lowest binding energies. Both glycosides and aglycones formed hydrogen bonds with amino acid residues in the W-box motif (Figure 8).
3. Discussion
This study analyzed fourteen plant extracts from Scutellaria and Lippia species cultivated in Colombia that were prepared under varying experimental conditions. Chemical analysis indicated that flavonoids constituted over 100% of their content. UAE-extracts exhibited significant differences in their chemical profiles. Retention of flavonoid glycosides was greater with shorter extraction times (5 or 23 minutes) than with longer times (15 or 60 minutes). This trend was evident in comparisons between SiSE1 vs SiSE2 as well as LopSE1 vs LopSE2 extracts. The reduction in flavonoid glycosides by prolonged extraction is likely due to excessive sonication that facilitate thermal, oxidative, and mechanical degradation, which particularly affect thermolabile glycosides and phenolic compounds [41,42]. Time-dependent variation in the extraction process critically influences the chemical profiles, and ultimately, the abundance of bioactive compounds [41]. SFE is a highly efficient technique for extracting aglycone-rich fractions with limited recovery of water-soluble or glycosylated phytochemicals [43]. The SFE extracts of L. origanoides contained high levels of flavonoid aglycones and lacked glycosides, reflecting the principle that nonpolar CO₂ extraction selectively enriches lipophilic compounds while excluding polar glycosides [43]. Chemical profiles of the extracts analyzed in this study differ from those reported for extracts of the same plant [19,28,29,30,32]. These variations can be explained by differences in growth stage and plant part, cultivation practice, and preparation technique.
The potential of crude plant extracts to inhibit DENV replication in vitro has been widely documented [5,6,7]. However, relationships between their chemical composition and anti-DENV effect remain to be elucidated. This study integrates UHPLC/ESI-Q-Orbitrap-MS data with CPE-DENV reduction data showing that higher contents of flavonoid glycosides, predominantly flavones, correlate with greater inhibitory effect on DENV replication in Vero cells. Moreover, increased aglycones contain is associated with lack of antiviral action, as evidenced by the extracts from L. origanoides. The distinction between S. inarnata extracts is notable: SiSE1 showed strong antiviral effect, whereas SiSE2 had none. SiSE2 contained lower flavonoid glycoside content (64.8% vs. 88.6%) and sevenfold higher aglycones (35.2% vs. 5.0%). In a previous study, we compared the anti-DENV effects of LopSE1 (77% glycosides) and LopSFE (no glycosides) extracts from L. origanoids. After treatment during DENV-1 adsorption on human hepatic cells, the level of viral NS1 protein was significantly reduced by LopSE1, but not by LopSFE. The flavonoid content is a determining factor in the antiviral action of plant extracts on virus others than DENV [8,44]. Glycosylation may enhance flavonoid interactions with virus particles or cell surface receptors by increasing polarity or specific binding conformations relative to aglycones [44]. A study revealed that baicalin exhibited higher anti-DENV activity than baicalein in virus-infected cells [45]. Other study revealed that myricetin and its glycosides exhibited higher antiviral activity against HIV-1 virus compared to their aglycone counterparts [46].
While mechanisms underlying plant extract antiviral activity are yet to be fully elucidated, flavonoids are widely accepted as primary agents [10,11]. Certain flavonoids can interfere with viral adsorption to host cells through the attachment and subsequent inhibition of proteins and molecules involved in the virus endocytosis [46,47]. The antiviral assay in this study evaluated the potential of the extracts to interfere with virus adsorption to cells. Reduction of DENV-CPE by Scutellaria and L. alba extracts suggests their flavonoids may block virus-cell membrane interactions. In silico analysis support this. We selected the envelope E protein of DENV, which plays an important role in the process that allows the virus to enter the cell [33]. Flavonoid glycosides from these extracts exhibited the strongest affinity for amino acids of the DII domain that interacts with receptors to promote viral entry into cells [33]. In addition, flavonoid aglycones bound to the βOG biding site, which has been established as a target for developing antivirals for dengue [48]. Numerous flavonoids have been identified as ligands for the DENV-E protein [11,49].
DENV uses a variety of cell receptors to gain entry into Vero cells [35,36,37]. The Gas6-Axl complex was selected for the in silico analysis. Flavonoid glycosides from anti-DENV active extracts showed good binding affinities to Gas6, suggesting potential biological interference with this receptor, whereas none of the flavonoid aglycones bound effectively. Flavonoid glycosides may have prevented DENV virions from adhering to Vero cells by blocking the PtdSer-Gas6 binding step. Disrupting the PtdSer-Gas6-Axl complex has been proposed as a potential therapeutic approach to inhibit DENV replication [36]. Flavonoids may also have entered the cells and affected the expression of Gas6 and Axl. Luteolin can downregulate the expression of these proteins in human cells [50].
Dynamin-dependent and clathrin-mediated endocytosis pathways are involved in DENV entry into Vero cells [33,34,35]. Targeting these pathways has been proposed as a strategy for developing antivirals [40,51]. The N-terminal domain (CTD) of clathrin and the GTPase domain of dynamin were selected for in silico analysis. Once again, flavonoid glycosides present in extracts with anti-DENV effect exhibited the best binding affinities with CTD and GTPase. Baicalein and scutellarin, abundant flavonoid aglycones in Scutellaria extracts, had the best binding affinity to both proteins. The flavonoids may have destabilized the W-box domain of clathrin-CTD and reduced the capacity of dynamin to exchange GDP for GTP. This, in turn, may have affected the functionality of both proteins, consequently impacting the endocytosis process of DENV in Vero cells. Also, it is plausible that the flavonoid treatment altered the fluidity of both the viral envelope and the cell membrane, thereby affecting their fusion process during virus endocytosis into the cell. The capacity of flavonoids to affect cell membrane fluidity (raft-breaking effect) has been documented [52].
4. Materials and Methods
4.1. Reagents
HPLC-grade acetonitrile, HPLC-grade formic acid (FA), isopropanol (98%), ammonium formate (AF, ≥99%), LC/MS-grade methanol, and potassium persulfate (≥98%) were obtained from Merck (Darmstadt, Germany). Standard substances (e.g., eriodictyol-7-glucoside, salvigenin, galangin, scutellarin, apigenin-7-glucuronide) were purchased from Sigma-Aldrich (St. Louis, MO, USA), Chemfaces (Wuhan,China) and Phytolab GmbH (Vestenbergsgreuth, Bavaria, Germany). MO, USA). Type I water was obtained from a Millipore Direct-QTM purification system. Eagle’s Minimum Essential Medium (MEM), fetal bovine serum (FBS), phosphate buffered saline (PBS), and antibiotics were purchased from Gibco (Grand Island, NY).
4.2. Viruses and Cell
Dengue virus type 1 (DENV-1) strain US/Hawaii/1944 and Dengue virus type-2 (DENV-2) New Guinea C strain (NGC) were used. Viruses were propagated titrated in BHK-21 cells as in a previous study [53]. African green monkey kidney (Vero) cell line (ATCC® CCL-81™) was cultured in Eagle’s Minimum Essential Medium (EMEM) containing 10% fetal bovine serum (FBS, Gibco, NY, U.S.A.) at 37 °C in the presence of 5% CO2.
4.3. Extracts
Plant material was provided by the Colombian government through Contract No. 270 for Access to Genetic Resources and Derived Products between the Ministry of Environment and Sustainable Development and the Industrial University of Santander. The plants used in this study were grown in the experimental plots at the Agroindustrial Pilot Complex of the National Center for Agroindustrialization of Aromatic and Medicinal Tropical Vegetables (CENIVAM) in the Industrial University of Santander (Bucaramanga, Colombia). The plants were dried in the dark, and leaves and stems were crushed and homogenized. Voucher specimens were deposited in the Herbarium of the Industrial University of Santander. Fourteen extracts were included in the analysis of their anti-DENV effect. Eight extracts were prepared in this study using the UAE technique following the protocol used in previous studies [20,22]. Briefly, dried leaves and stems (100 g) were crushed and the material was mixed with ethanol:water (20 mL; 70:30), the mixtures were subjected to ultrasonication (35 kHz, Elmasonic S15H, Singen, Germany) and filtered (Wattman No. 1 paper), and the residue was extracted again with ethanol:water (10 mL). Temperature of 50 oC for 60 min was used. The extracts were vacuum roto-evaporated in a Heidolph apparatus, dried in a VirTis AdVantage Plus tray lyophilizer, and stored at 4 ◦C in the absence of light. Six extracts were prepared in previous studies using UAE [20,22] or SFE techniques [21]. Each extract (1x105 µg/mL) was dissolved in 1% dimethyl sulfoxide (DMSO) and stored at -20 ◦C before analysis.
4.4. UHPLC/ESI-Q-Orbitrap-MS Analysis
UHPLC-Q-Orbitrap-MS/MS analysis was used for identifying and characterizing the chemical components of the eight extracts prepared in this study, in accordance with the protocol used in a previous study [22]. An Ultimate DionexTM 3000 UHPLC (Thermo Fisher Scientific, Bremen, Germany) connected to an Orbitrap™ mass detector Exactive Plus, Bremen, Germany), with with heated electrospray ionization (HESI-II; Thermo Fisher Scientific). operated in positive-ion acquisition mode (350 oC). Chromatographic separation was performed on 50 mm L × 2.1 mm I.D. Zorbax Eclipse XDB C18 column, 1.8 µm particle size (Sigma-Aldrich, St. Louis, MO, USA). The mobile phase was a mixture of 0.2% formic acid-water (A) and 0.2% formic acid-acetonitrile (B) at a flow rate of 0.3 mL/min. The initial gradient condition was A-100%, which linearly changed to B-100% in 8 min, held for 4 min, returned to A-100% in 1 min, followed by 3 min of equilibration. The injection volume was 2 µL. Mass spectrometry experiments were accomplished on a Q-Exactive Plus Orbitrap mass spectrometer (Thermo Scientific, Bremen, Germany) connected to a the UHPLC and equipped with an electrospray ionization (ESI) source. Samples were analyzed in negative ion modes. The parameters were as follows: mass range, m/z 80 to 1000; electrospray ionization temperature, 350 °C; capillary temperature, 320 ◦C; capillary voltage, 3.5 kV; higher-energy-collisional dissociation cell (HCD), 10- 40 eV range. Data were analyzed using Thermo Scientific™ Dionex™ Chromatography Data System (CDS) software, version 7.2 and Thermo Xcalibur 3.1 software (Thermo Scientific, CA, USA). Extract metabolites were identified by comparing their retention times (tR), exact ion masses, isotopic ratios and fragmentation patterns with those of standard substances and databases [31,54]. All samples were run in triplicates and the data were presented as the mean value ± standard deviation (SD) of mg per g of freeze-dried extract.
4.5. Crystal Violet Assays
The assay evaluated the effect of the fourteen extracts on the viability of non-DEN-infected Vero cells. Two different protocols, each including seven extract concentrations (3.12 to 250 µg/mL), were followed. Protocol 1: cell monolayers were treated with extract for 1.5 h, the extract was removed by washing with PBS, fresh medium MEM with 2% fetal bovine serum was added, and the plates were incubated at 37 ºC; 5% CO2 for five days. Culture supernatants were discarded and crystal violet solution (0.05%) was added followed by methanol, and then the plates were then analyzed using an ELISA plate reader at a wavelength of 570 nm. Untreated cells and cells treated with DMSO were run in parallel as control negative and positive, respectively. The higher concentration of extract that did not reduce cell viability respect to untreated control was calculated by regression analysis (GraphPad Software, San Diego, CA, USA). Protocol 2: cell monolayers were incubated in culture medium containing extract for 72 h, the extract was then removed by washing, and the cell viability was measured as in Protocol 1. Untreated cells and cells treated with DMSO were included as controls. The CC50 values were calculated by regression analysis as in previous studies [20,53].
4.6. Cytopathic Effect (CPE)-Based Antiviral Assay
Extracts were analyzed in the CPE-based antiviral assay according to the protocol used in a previous study [53]. DENV (1 PFU/cell) was adsorbed on Vero cells grown in 96-weel plate for 1.5 h at 37 ◦C; 5% CO2 in the presence of plant extract at non-cytotoxic concentrations (3.12 to 100 µg/ml); then, the cells were washed with PBS and fresh media without extract was added to allow virus replication. After five days of incubation at 37 ºC, the culture supernatant was discarded and cell viability measured using the crystal violet assay. Untreated DENV-infected cells and virus-infected / SDS-treated cells were included as negative and positive controls, respectively. DENV CPE (%) was calculated as follows: [(OD570 of infected treated cells / OD570 of uninfected untreated cells) x 100]. DENV CPE reduction was calculated as follows: DENV CPE reduction (%) = [(OD570 of infected treated cells - OD570 of negative control) / (OD570 of positive control - OD570 of negative control) × 100]. The concentration of extract that inhibited DENV-CPE by 50% (IC50) was calculated by nonlinear regression followed by the construction of a concentration-response curve (GraphPad Prism software version 8.0, San Diego, CA, U.S.A.). Selectivity index (SI = CC50 / IC50) values were calculated. Each extract was analyzed in triplicate in three independent assays.
4.7. Molecular Docking Analysis
Three-dimensional structures of target proteins were downloaded from the Protein Data Bank. DENV-2 E protein (PDB ID: 10AN), Gas6-Axl receptor (PDB ID: 2C5D), clathrin (CDT domain, PDB ID: 2XZG), and dynamin (GTPase domain, PDB ID: 2X2E). Structures of flavonoids were retrieved from the PubChem (https://pubchem.ncbi.nlm.nih.gov/ database (accessed on September 2024). The preparation of the target and ligands and molecular docking analyses were carried out using AutoDock Vina (Version 1.5.6, La Jolla, CA, USA), as described in previous studies [53,54]. The optimized protein structure was saved in the PDBQT file format. Default parameters were used, and the search exhaustiveness parameter was set to 100. Twenty five docked conformations were generated for each ligand using docking simulations. Three simulations were performed for each ligand–protein pair using seeds 6, 12, and 18. The ligand–protein interactions was displayed using the Discovery Studio Visualizer v21.1.0.20298 software.
4.8. Statistical Analysis
The relationship between antiviral activity and the flavonoid content was analyzed using two independent parameters: the chemical composition, represented as the absolute percentage (%) of different flavonoid classes (Table 5), and the antiviral effect, DENV-CPE (%) as a unidimensional variable ranging from 0 to 100. The fourteen extracts were clustered based on to their content of flavonoids and other compounds, using an unsupervised self-organized Kohonen Map [55]. One-way ANOVA and a Tukey-Kramer post hoc tests compared DENV-CPE (%) values between clusters, adopting a significance level of 0.05. The analysis was performed using the Matlab® R2021b software.
5. Conclusions
This study reports for the first time the correlation between the antiviral efficacy of extracts from medicinal plants cultivated in Colombia and their flavonoid content. Integration of antiviral assays, UHPLC analysis, and molecular docking analysis revealed that the content in flavonoid glycosides is a key determinant of the extracts’ anti-DENV effect. Extracts rich in flavonoid glycosides, particularly flavones, demonstrated the greatest antiviral efficacy. In contrast, extracts with lower levels of glycosides or lacking them did not show antiviral activity. Docking analysis suggests that the flavonoids present in the extracts could block the interaction between DENV particles and the cell membrane via different mechanisms. Based on IC50 and SI values from the CPE-based antiviral assay, the UAE-extract from S. coccinea can be classified as a prospective anti-DENV sample. This extract contains high concentrations of flavonoid glycosides with potential to inhibit DENV, and therefore could serve as a starting point for research on herbal medications for dengue treatment. Considerable evidence demonstrates the potential of Scutellaria species as an antiviral agent against various pathogenic viruses, including DENV.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Amount (mg/g) of compounds in extracts reported in previous studies. Table S2: Docking score (kcal/mol) flavonoid ligands of DENV-2 (E) and Vero cell (Cl, clathrin; DY, dynamin; GA, Gas6-Axl complex) proteins.
Author Contributions
Conceptualization, R.E.O.; methodology S.A.V.; E.Q.-R.; software and data curation, S.C.-O.; E.E.S.; writing—original draft preparation, R.E.O.; S.A.V; writing—review and editing, E.E.S. All authors have read and agreed to the published version of the manuscript.
Funding
The authors thank funding from the Ministry of Science, Technology and Innovation, the Ministry of Education, the Ministry of Industry, Commerce and Tourism, and ICETEX, Programme Ecosistema Científico-Colombia Científica, from the Francisco José de Caldas Fund; Grant RCFP44842-212-2018.
Institutional Review Board Statement
The Ministry of Environment and Sustainable Development. of Colombia supported the Universidad Industrial de Santander through the permit to genetic resources and derivatives access for bioprospecting (Contract No 270). The project RC-FP44842-212- 2018 was approved by the Scientific Research Ethical Committee (Record No. 15-2017, File No. 4110) from Universidad Industrial de Santander. The experiments and the chemical management were done according to national law (Resolution No 008430-1993) from the Ministry of Health of Colombia and the Institutional Manual of Integrated Management and Processes (PGOR-PGGA.05).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wilder-Smith, A.; Ooi, E.E.; Horstick, O.; Wills, B. Dengue. The Lancet. 2019, 393, 350–363. [Google Scholar] [CrossRef]
- Pan American Health Organization (PAHO). Dengue epidemiological situation in the region of the Americas, epidemiological week 35, 2025. PAHO: Washington, D.C. 2025. Available online: https://www.paho.org/en/documents/dengue-epidemiological-situation-region-americas-epidemiological-week-35-2025 (accessed on 24 September 2025).
- Obi, J.O.; Gutiérrez-Barbosa, H.; Chua, J.V.; Deredge, D.J. Current trends and limitations in dengue antiviral research. Trop Med Infect Dis. 2021, 30, 180. [Google Scholar] [CrossRef]
- Palanichamy Kala, M.; St John, A.L.; Rathore, A.P.S. Dengue: update on clinically relevant therapeutic strategies and vaccines. Curr Treat Options Infect Dis. 2023, 15, 27–52. [Google Scholar] [CrossRef] [PubMed]
- Rehman, B.; Ahmed, A.; Khan, S.; et al. Exploring plant-based dengue therapeutics: from laboratory to clinic. Trop Dis Travel Med Vaccines. 2024, 10, 23. [Google Scholar] [CrossRef]
- Rajapakse, S.; de Silva, N.L.; Weeratunga, P.; Rodrigo, C.; Sigera, C.; Fernando, S.D. Carica papaya extract in dengue: a systematic review and meta-analysis. BMC Complement Altern Med. 2019, 11, 265. [Google Scholar] [CrossRef]
- Adeosun, W.B.; Loots, D.T. Medicinal Plants against Viral Infections: A Review of Metabolomics Evidence for the Antiviral Properties and Potentials in Plant Sources. Viruses. 2024, 31, 218. [Google Scholar] [CrossRef]
- Zhao, L.; Qian, S.; Wang, X.; Si, T.; Xu, J.; Wang, Z.; Sun, Q.; Yang, Y.; Rong, R. UPLC-Q-Exactive/MS based analysis explore the correlation between components variations and anti-influenza virus effect of four quantified extracts of Chaihu Guizhi decoction. J Ethnopharmacol. 2024, 30, 319, 117318. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yuan, X.; Li, L.; Zhang, T.; Wang, B. Comparison of different methods for extraction of Cinnamomi ramulus: yield, chemical composition and in vitro antiviral activities, Nat. Prod. Res. 2017, 24, 2909–2913. [Google Scholar] [CrossRef] [PubMed]
- Badshah, S.L.; Faisal, S.; Muhammad, A.; Poulson, B.G.; Emwas, A.H.; Jaremko, M. Antiviral activities of flavonoids. Biomed Pharmacother. 2021, 140, 111596. [Google Scholar] [CrossRef]
- Loaiza-Cano, V.; Monsalve-Escudero, L.M.; Filho, C.D.S.M.B.; Martinez-Gutierrez, M.; Sousa, D.P. Antiviral role of phenolic compounds against dengue virus: A Review. Biomolecules. 2020, 24, 11. [Google Scholar] [CrossRef]
- Shen, J.; Li, P.; Liu, S.; Liu, Q.; Li, Y.; Sun, Y.; He, C.; Xiao, P. Traditional uses, ten-years research progress on phytochemistry and pharmacology, and clinical studies of the genus Scutellaria. J Ethnopharmacol. 2021, 30, 265, 113198. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, M.; Wang, M.; Lu, Y.; Wang, X.; Chen, X.; Yang, X.; Guo, H.; He, R.; Luo, Z. Scutellaria baicalensis: a promising natural source of antiviral compounds for the treatment of viral diseases. Chin J Nat Med. 2023, 21, 563–575. [Google Scholar] [CrossRef]
- Atkins, S. Verbenaceae: the families and genera of fowering plants. Berlin: Springer; 2004, 7, 449–68. [Google Scholar]
- Pascual, M.E.; Slowing, K.; Carretero, E.; Mata, D.S.; Villar, A. Lippia: traditional uses, chemistry and pharmacology: a review. J Ethnopharmacol. 2001, 76, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Hennebelle, T.; Sahpaz, S.; Joseph, H.; Bailleul, F. Ethnopharmacology of Lippia alba. J Ethnopharmacol. 2008, 5, 116, 211–22. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.R.; Leitão, G.G.; Fernandes, P.D.; Leitã, S.G. Ethnopharmacological studies of Lippia origanoides. Revista Brasileira de Farmacognosia. 2014, 24, 206–214. [Google Scholar] [CrossRef]
- Mamun-Or-Rashid, A. N. M.; Sen, M. K.; Jamal, M. A. H. M.; Nasrin, S. A comprehensive ethnopharmacological review on Lippia alba M. Int. J Biomed. Mater. Res. 2013, 1, 14–20. [Google Scholar]
- Zandi, K.; Lim, T.H.; Rahim, N.A.; Shu, M.H.; Teoh, B.T.; Sam, S.S.; Danlami, M.B.; Tan, K.K.; Abubakar, S. Extract of Scutellaria baicalensis inhibits dengue virus replication. BMC Complement Altern Med. 2013, 29, 13, 91. [Google Scholar] [CrossRef]
- Quintero-Rueda, E.; Velandia, S.A.; Ocazionez, R.E.; Stashenko, E.E.; Rondón-Villarreal, P. Comparative anti-dengue activities of ethanolic and supercritical extracts of Lippia origanoides Kunth: in-vitro and in-silico analyses. Recs Nat Prod. 2023, 1–18. [Google Scholar] [CrossRef]
- Arias, J.; Mejía, J.; Córdoba, Y.; Martínez, J.R.; Stashenko, E.E.; del Valle, J.M. Optimization of flavonoids extraction from Lippia graveolens and Lippia origanoides chemotypes with ethanol-modified supercritical CO2 after steam distillation, Ind. Crops. and Prod. 2020, 146, 112170. [Google Scholar] [CrossRef]
- Porras, S.M.; Saavedra, R.A.; Sierra, L.J.; González, R.T.; Martínez, J.R.; Stashenko, E.E. Chemical characterization and determination of the antioxidant properties of phenolic compounds in three Scutellaria sp. plants grown in Colombia. Molecules. 2023, 14, 28, 3474. [Google Scholar] [CrossRef]
- Quintero, W.L.; Moreno, E.M.; Pinto, S.M.L.; Sanabria, S.M.; Stashenko, E.; García, L.T. Immunomodulatory, trypanocide, and antioxidant properties of essential oil fractions of Lippia alba (Verbenaceae). BMC Complement Med Ther. 2021, 2, 21, 187. [Google Scholar] [CrossRef]
- Stashenko, E.E.; Martínez, J.R.; Cala, M.P.; Durán, D.C.; Caballero, D. Chromatographic and mass spectrometric characterization of essential oils and extracts from Lippia (Verbenaceae) aromatic plants. J Sep Sci. 2013, 36, 192–202. [Google Scholar] [CrossRef]
- Rodríguez-Morales, A.J.; López-Medina, E.; Arboleda, I.; Cardona-Ospina, J.A.; Castellanos, J.E.; Faccini-Martínez, Á.A.; Gallagher, E.; Hanley, R.; Lopez, P.; Mattar, S.; Pérez, C.E.; Kastner, R.; Reynales, H.; Rosso, F.; Shen, J.; Villamil-Gómez, W.E.; Fuquen, M. The epidemiological impact of dengue in Colombia: a systematic review. Am J Trop Med Hyg. 2024, 29, 112, 182–188. [Google Scholar] [CrossRef]
- Pájaro-González, Y.; Oliveros-Díaz, A.F.; Cabrera-Barraza, J.; CerraDominguez, J.; Díaz-Castillo, F. Chapter 1- A review of medicinal plants used as antimicrobials in Colombia. In: Chassagne, F. (Ed.). Medicinal Plants as Anti-Infectives. Academic Press. 2022; pp. 3–57.
- Smee, D.F.; Hurst, B.L.; Evans, W.J.; Clyde, N.; Wright, S.; Peterson, C.; Jung, K.H.; Day, C.W. Evaluation of cell viability dyes in antiviral assays with RNA viruses that exhibit different cytopathogenic properties. J Virol Methods. 2017, 246, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.F.; Almeida, M.P.; Leite, M.F.; Schwaiger, S.; Stuppner, H.; Halabalaki, M.; Amaral, J.G.; David, J.M. Seasonal variation in the chemical composition of two chemotypes of Lippia alba. Food Chem. 2019, 273, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.Z.; Mukhopadhyay, S.; Robbins, R.J.; Harnly, J.M. Identification and quantification of flavonoids of Mexican oregano (Lippia graveolens) by LC-DAD-ESI/MS analysis. J. Food Compos. Anal. 2007, 20, 361–369. [Google Scholar] [CrossRef]
- Parolin-Trindade, G.; Perez-Pinheiro, G.; Silva, A.A.R.; Porcari, A.M.; Sawaya, A.C.H.F. Sources of variation of the chemical composition of Lippia origanoides Kunth (Verbenaceae). Nat Prod Res. 2025, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- The NIST Mass Spectrometry Data Center. The NIST Mass Spectral Search Program. Standard Reference Data Program of the National Institute of Standards and Technology, U.S.A.; National Institute of Standards and Technology: Gaithersburg, MD, USA, 2017. [Google Scholar]
- Timóteoa, P.; Kariotia, A.; Leitãob, S.G.; Vincieri, F.F.; Bilia, A.R. HPLC/DAD/ESI-MS Analysis of non-volatile constituents of three Brazilian chemotypes of Lippia alba (Mill.) N. E. Brown. Nat Prod Com. 2008, 3, 12, 2017–2020. [Google Scholar] [CrossRef]
- Zhang, X.; Jia, R.; Shen, H.; Wang, M.; Yin, Z.; Cheng, A. Structures and functions of the envelope glycoprotein in flavivirus infections. Viruses. 2017, 13, 9, 338. [Google Scholar] [CrossRef]
- Meertens, L.; Carnec, X.; Lecoin, M.P.; Ramdasi, R.; Guivel-Benhassine, F.; Lew, E.; Lemke, G.; Schwartz, O.; Amara, A. The TIM and TAM families of phosphatidylserine receptors mediate dengue virus entry. Cell Host Microbe. 2012, 12, 544–57. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Oliveira, C.; Freire, J.M.; Conceição, T.M.; Higa, L.M.; Castanho, M.A.; Da Poian, A.T. Receptors and routes of dengue virus entry into the host cells. FEMS Microbiol Rev. 2015, 39, 155–170. [Google Scholar] [CrossRef]
- Piccini, L.E.; Castilla, V.; Damonte, E.B. Dengue-3 Virus Entry into Vero Cells: Role of Clathrin-Mediated Endocytosis in the Outcome of Infection. PLoS One. 2015, 10, e0140824. [Google Scholar] [CrossRef]
- Acosta, E.G.; Castilla, V.; Damonte, E.B. Alternative infectious entry pathways for dengue virus serotypes into mammalian cells. Cell Microbiol. 2009, 11, 1533–1549. [Google Scholar] [CrossRef]
- Kirchhausen, T.; Owen, D.; Harrison, S.C. Molecular structure, function, and dynamics of clathrin-mediated membrane traffic. Cold Spring Harb Perspect Biol. 2014, 6, a016725. [Google Scholar] [CrossRef]
- Harper, C. B.; Popoff, M. R.; McCluskey, A.; Robinson, P. J.; Meunier, F. A. Targeting membrane trafficking in infection prophylaxis: dynamin inhibitors. Trends in Cell Biology. 2013, 23(2), 90–101. [Google Scholar] [CrossRef]
- Ho, M.R.; Tsai, T.T.; Chen, C.L.; et al. Blockade of dengue virus infection and viral cytotoxicity in neuronal cells in vitro and in vivo by targeting endocytic pathways. Sci Rep. 2017, 7, 6910. [Google Scholar] [CrossRef]
- Shen, L.; Pang, S.; Zhong, M.; Sun, Y.; Qayum, A.; Liu, Y.; Rashid, A.; Xu, B.; Liang, Q.; Ma, H.; Ren, X. A comprehensive review of ultrasonic assisted extraction (UAE) for bioactive components: Principles, advantages, equipment, and combined technologies. Ultrason Sonochem. 2023, 101, 106646. [Google Scholar] [CrossRef]
- Hou, M.; Hu, W.; Wang, A.; Xiu, Z.; Shi, Y.; Hao, K.; Sun, X.; Cao, D.; Lu, R.; Sun, J. Ultrasound-assisted extraction of total flavonoids from Pteris cretica L.: Process Optimization, HPLC Analysis, and Evaluation of Antioxidant Activity. Antioxidants (Basel). 2019, 24, 8, 425. [Google Scholar] [CrossRef] [PubMed]
- Uwineza, P.A.; Waśkiewicz, A. Recent advances in supercritical fluid extraction of natural bioactive compounds from natural plant materials. Molecules. 2020, 25, 3847. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J. Dietary flavonoid aglycones and their glycosides: Which show better biological significance? Crit Rev Food Sci Nutr. 2017, 57, 1874–1905. [Google Scholar] [CrossRef]
- Moghaddam, E.; Teoh, BT.; Sam, SS.; Lani, R.; Hassandarvish, P.; Chik, Z.; Yueh, A.; Abubakar, S.; Zandi, K. Baicalin, a metabolite of baicalein with antiviral activity against dengue virus. Sci Rep. 2014, 4, 5452. [Google Scholar] [CrossRef]
- Ortega, J.T.; Suárez, A.I.; Serrano, M.L.; et al. The role of the glycosyl moiety of myricetin derivatives in anti-HIV-1 activity in vitro. AIDS Res Ther. 2017, 14, 57. [Google Scholar] [CrossRef] [PubMed]
- Pourfarzad, F.; Gheibi, N.; Namdar, P.; Gholamzadeh-Khoei, S.; Gheibi, N. Flavonoids antiviral effects: focusing on entry inhibition of influenza and coronavirus. J Inflamm Dis. 2024, 28, e153347. [Google Scholar]
- Naresh, P.; Selvaraj, A.; Shyam Sundar, P.; Murugesan, S.; Sathianarayananan, S.; Namboori P. K, K.; Jubie, S. Targeting a conserved pocket (n-octyl-β-D–glucoside) on the dengue virus envelope protein by small bioactive molecule inhibitors. J. Biomol. Struct. Dyn 2022, 40, 4866–4878. [Google Scholar] [CrossRef] [PubMed]
- Waiba, A.; Phunyal, A.; Lamichhane, TR.; Ghimire, MP.; Nyaupane, H.; Phuyal, A.; et al. Computational insights into flavonoids inhibition of dengue virus envelope protein: ADMET profiling, molecular docking, dynamics, PCA, and end-state free energy calculations. PLoS One. 2025, 20, e0327862. [Google Scholar] [CrossRef]
- Li, X.; Chen, M.; Lei, X.; Huang, M.; Ye, W.; Zhang, R.; Zhang, D. Luteolin inhibits angiogenesis by blocking Gas6/Axl signaling pathway. Int J Oncol. 2017, 51, 2, 677–685. [Google Scholar] [CrossRef]
- Alkafaas, S.S.; Abdallah, A.M.; Ghosh, S.; Loutfy, S.A.; Elkafas, S.S.; Abdel-Fattah, N.F.; Hessien, M. Insight into the role of clathrin-mediated endocytosis inhibitors in SARS-CoV-2 infection. Rev Med Virol. 2023, 33, e2403. [Google Scholar] [CrossRef]
- Kapral-Piotrowska, J.; Strawa, J.W.; Jakimiuk, K.; Wiater, A.; Tomczyk, M.; Gruszecki, WI.; Pawlikowska-Pawlęga, B. Investigation of the membrane localization and interaction of selected flavonoids by NMR and FTIR Spectroscopy. Int J Mol Sci. 2023, 24, 15275. [Google Scholar] [CrossRef] [PubMed]
- Silva-Trujillo, L.; Quintero-Rueda, E.; Stashenko, E.E.; Conde-Ocazionez, S.; Rondón-Villarreal, P.; Ocazionez, R.E. Essential Oils from Colombian Plants: Antiviral Potential against Dengue Virus Based on Chemical Composition, In Vitro and In Silico Analyses. Molecules. 2022, 27, 6844. [Google Scholar] [CrossRef]
- Parra-Acevedo, V.; Ocazionez, R.E.; Stashenko, E.E.; Silva-Trujillo, L.; Rondón-Villarreal, P. Comparative virucidal activities of essential oils and alcohol-based solutions against enveloped virus surrogates: in vitro and in silico analyses. Molecules 2023, 28, 4156. [Google Scholar] [CrossRef] [PubMed]
- Kohonen, T.; Somervuo, P. How to make large self-organizing maps for nonvectorial data. Neural Networks 2002, 15, 945–952. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Cytotoxicity of the extracts in the crystal violet assay. A. Vero cells were treated with extract for 1.5 h, after which the extract was removed. Cell viability was measured five days after treatment. B. Cells were cultured in medium containing the extract for 72 h, after which the viability was measured. S. coccinea (ScSE), S. incarnata (SiSE1 and SiSE2) and S. ventenatti + incarnata (SviSE); L. alba carvone (LacSE) and citral (LaiSE) chemotypes; and L. origanoides phellandrene (LopSE1, LopSE2, LopSFE), carvacrol (LocSE1, LocSE2, LocSFE) and thymol (LotSE, LotSFE) chemotypes. Dimethylsulfoxide (DMSO) was used as the positive control. Data are presented as mean ± SD of six measurements from three independent analyses.
Figure 1.
Cytotoxicity of the extracts in the crystal violet assay. A. Vero cells were treated with extract for 1.5 h, after which the extract was removed. Cell viability was measured five days after treatment. B. Cells were cultured in medium containing the extract for 72 h, after which the viability was measured. S. coccinea (ScSE), S. incarnata (SiSE1 and SiSE2) and S. ventenatti + incarnata (SviSE); L. alba carvone (LacSE) and citral (LaiSE) chemotypes; and L. origanoides phellandrene (LopSE1, LopSE2, LopSFE), carvacrol (LocSE1, LocSE2, LocSFE) and thymol (LotSE, LotSFE) chemotypes. Dimethylsulfoxide (DMSO) was used as the positive control. Data are presented as mean ± SD of six measurements from three independent analyses.
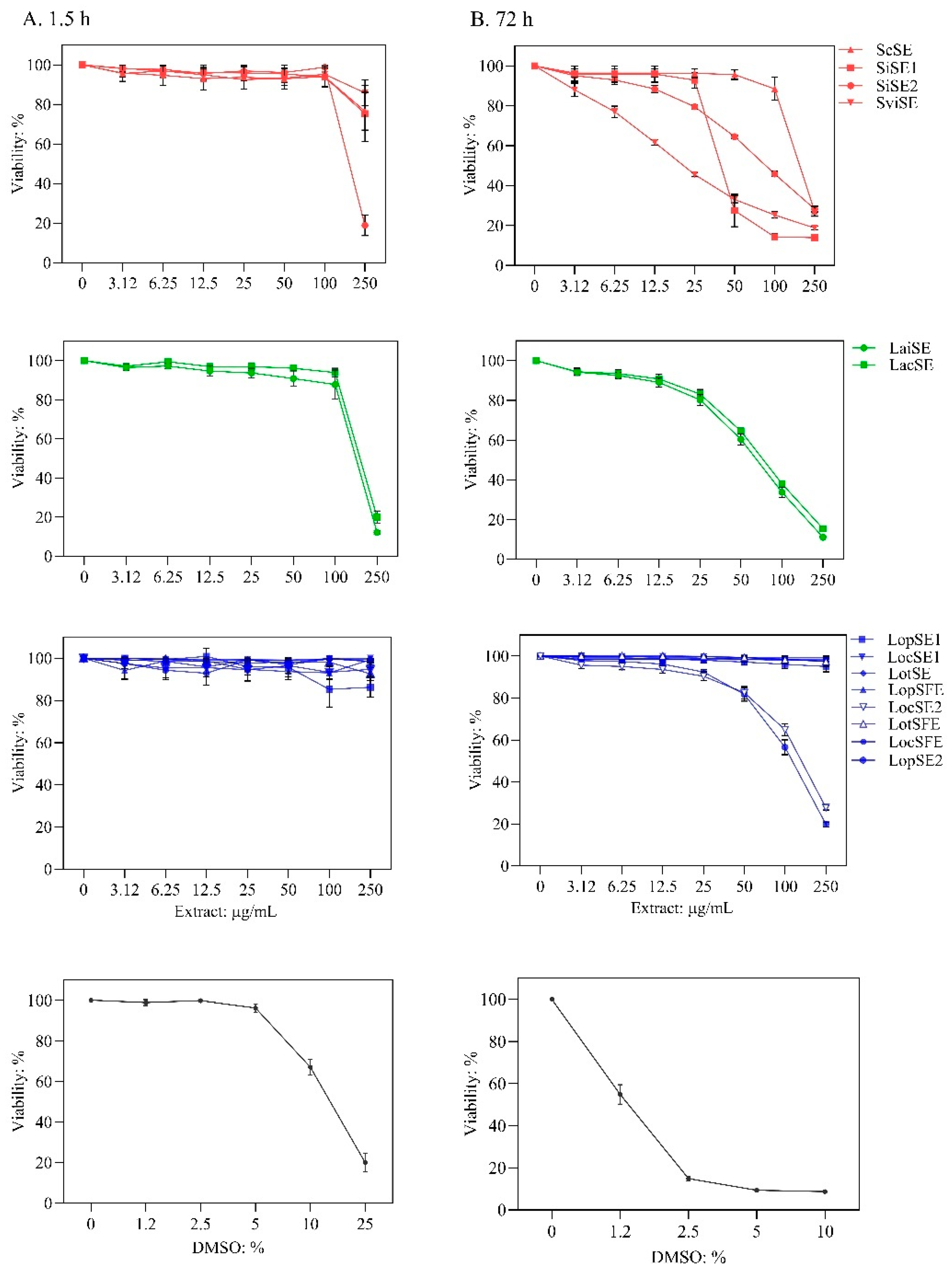
Figure 2.
Antiviral effect of the extracts against dengue virus (DENV-1 and DENV-2) in the cytopathic effect (CPE)-based assay. Extracts were added during virus adsorption to Vero cells. Quantification of crystal violet staining was performed to indirectly measure virus-induced CPE. DENV- CPE %: [(OD570 of virus-infected and extract-treated cells / OD570 of non-infected non-treated cells]. Scutellaria species (red), L. alba (green)and L. origanoides (blue) extracts. Sodium dodecyl sulfate (SDS) is an antiviral agent. Data are presented as mean ± SD of six measurements from three independent analyses.
Figure 2.
Antiviral effect of the extracts against dengue virus (DENV-1 and DENV-2) in the cytopathic effect (CPE)-based assay. Extracts were added during virus adsorption to Vero cells. Quantification of crystal violet staining was performed to indirectly measure virus-induced CPE. DENV- CPE %: [(OD570 of virus-infected and extract-treated cells / OD570 of non-infected non-treated cells]. Scutellaria species (red), L. alba (green)and L. origanoides (blue) extracts. Sodium dodecyl sulfate (SDS) is an antiviral agent. Data are presented as mean ± SD of six measurements from three independent analyses.
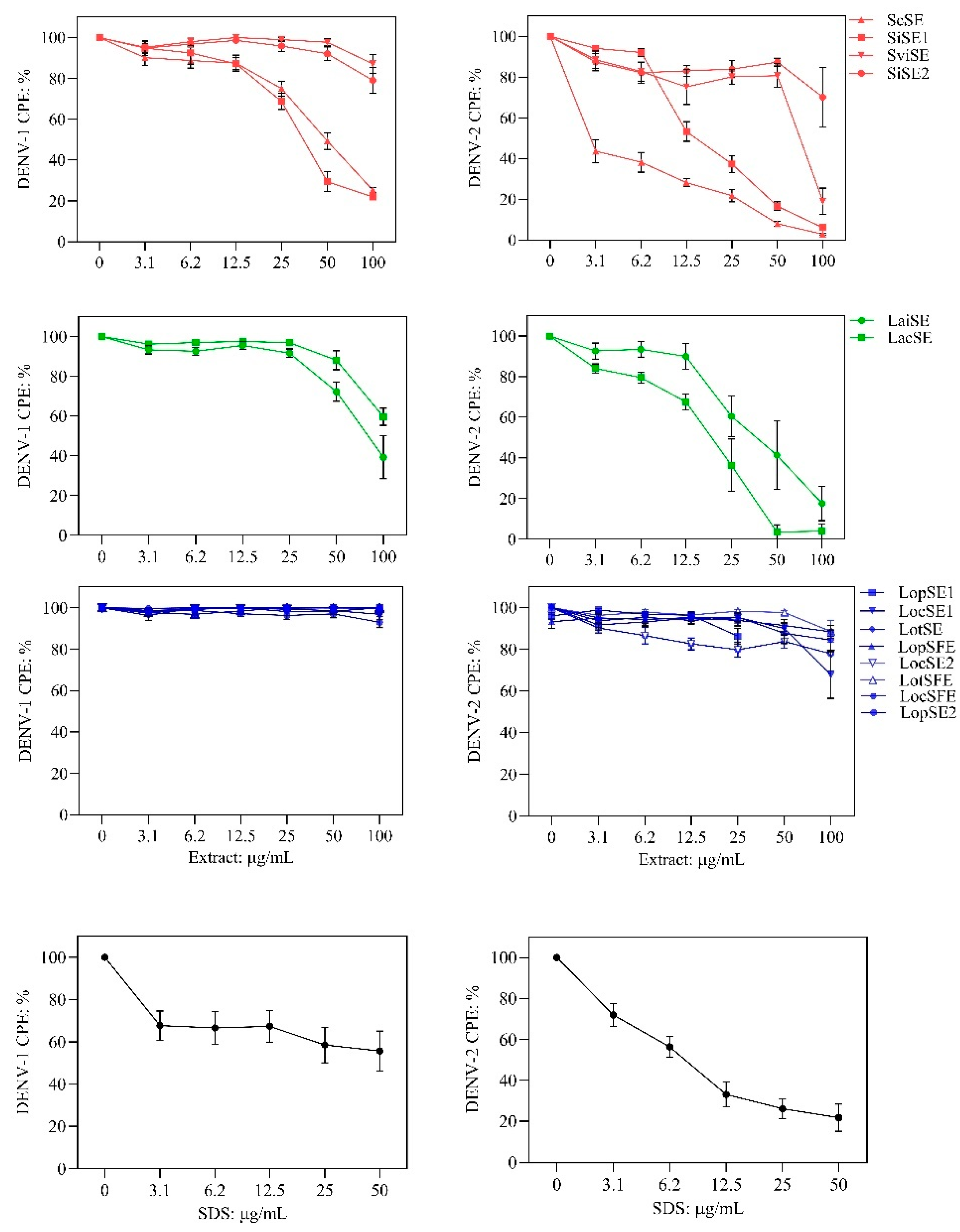
Figure 3.
Relationship between flavonoid profiles and anti-DENV effects of the extracts. A. Heatmap showing the content of flavonoid classes and revealing two clusters obtained by an unsupervised self-organizing map. B and C. Comparison of antiviral activity (CPE, cytopathic effect percentage) of clusters against DENV-1 (* Wilcoxon test Z = -2.969; p = 0.003) and DENV-2 (* Wilcoxon test Z = -2.785; p = 0.0053).
Figure 3.
Relationship between flavonoid profiles and anti-DENV effects of the extracts. A. Heatmap showing the content of flavonoid classes and revealing two clusters obtained by an unsupervised self-organizing map. B and C. Comparison of antiviral activity (CPE, cytopathic effect percentage) of clusters against DENV-1 (* Wilcoxon test Z = -2.969; p = 0.003) and DENV-2 (* Wilcoxon test Z = -2.785; p = 0.0053).
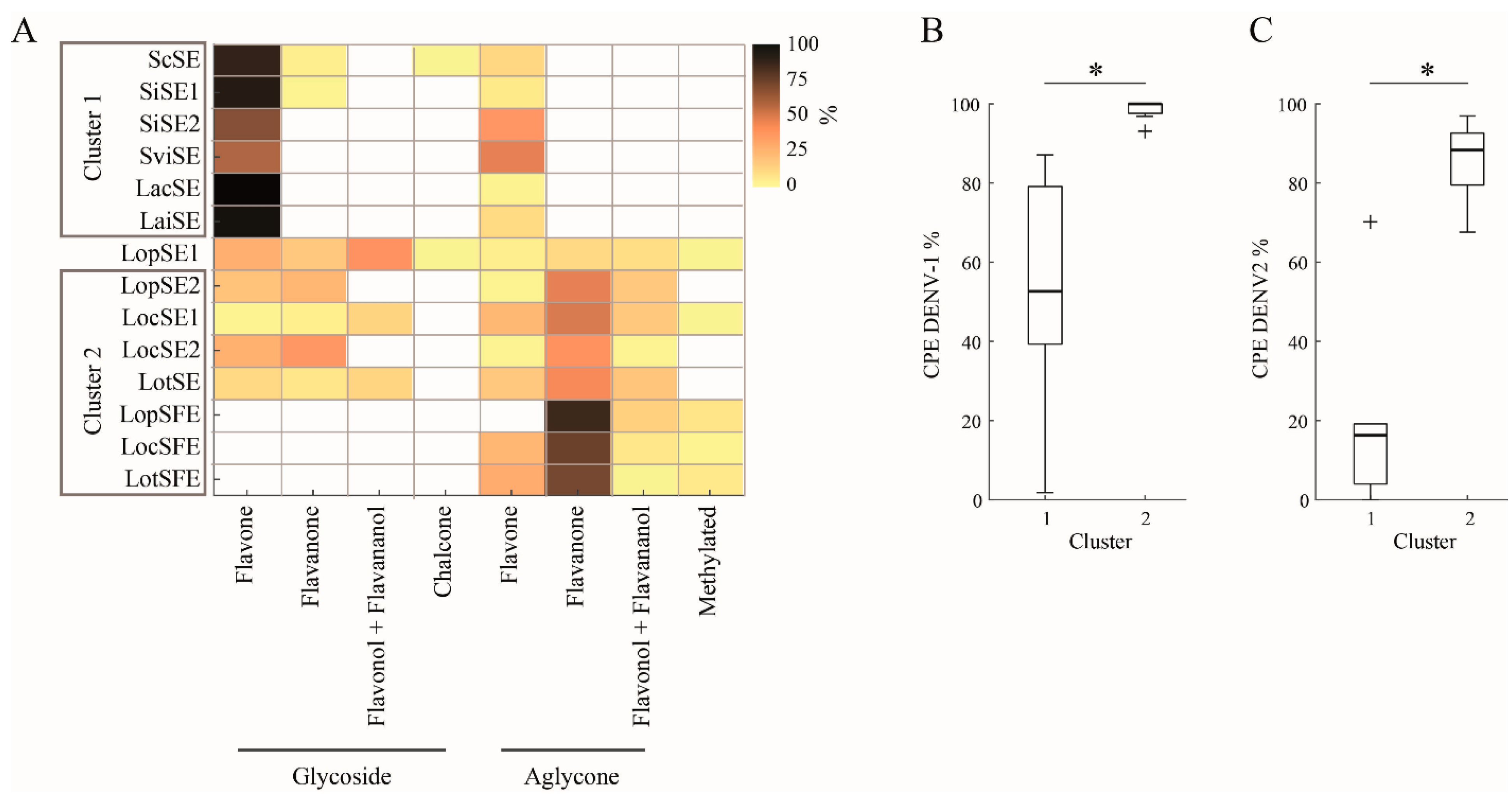
Figure 4.
Heatmaps showing the abundance of each flavonoid and its binding affinity for target proteins. A. Relative concentration (mg/g) of flavonoid in the extract (Table 4): 1, ScSE; 2, SiSE1; 3, SiSE2; 4, SviSE; 5, LacSE; 6, LaiSE; 7, LopSE1; 8, LopSE2; 9, LocSE1; 10, LocSE2; 11, LotSE; 11, LopSFE; 12, LocSFE; and 13, LotSFE. Scutellaria (1,2 and 4) and L. alba (6 and 7) exhibited anti-DENV effect, while the others extracts were inactive. * Low detection limit. B. Binding affinities (kcal/mol) of flavonoids for target proteins: E, DENV-2 E envelope; GX, Gas6-Axl receptor; Cl clathrin N-terminal domain (2XZG); DY, dynamin GTPase domain.
Figure 4.
Heatmaps showing the abundance of each flavonoid and its binding affinity for target proteins. A. Relative concentration (mg/g) of flavonoid in the extract (Table 4): 1, ScSE; 2, SiSE1; 3, SiSE2; 4, SviSE; 5, LacSE; 6, LaiSE; 7, LopSE1; 8, LopSE2; 9, LocSE1; 10, LocSE2; 11, LotSE; 11, LopSFE; 12, LocSFE; and 13, LotSFE. Scutellaria (1,2 and 4) and L. alba (6 and 7) exhibited anti-DENV effect, while the others extracts were inactive. * Low detection limit. B. Binding affinities (kcal/mol) of flavonoids for target proteins: E, DENV-2 E envelope; GX, Gas6-Axl receptor; Cl clathrin N-terminal domain (2XZG); DY, dynamin GTPase domain.

Figure 5.
Contact preferences at the DENV-E protein (PDB ID: 1OAN) of representative flavonoids. Norwogonin (1) within the DI/DII hinge region around the βOG pocket; chrysoeriol-7-diglucoronide (2) within the interface of DII domains; and baicalin (3) within the DIII/DI interface. Binding free energies (kcal/mol) are shown.
Figure 5.
Contact preferences at the DENV-E protein (PDB ID: 1OAN) of representative flavonoids. Norwogonin (1) within the DI/DII hinge region around the βOG pocket; chrysoeriol-7-diglucoronide (2) within the interface of DII domains; and baicalin (3) within the DIII/DI interface. Binding free energies (kcal/mol) are shown.
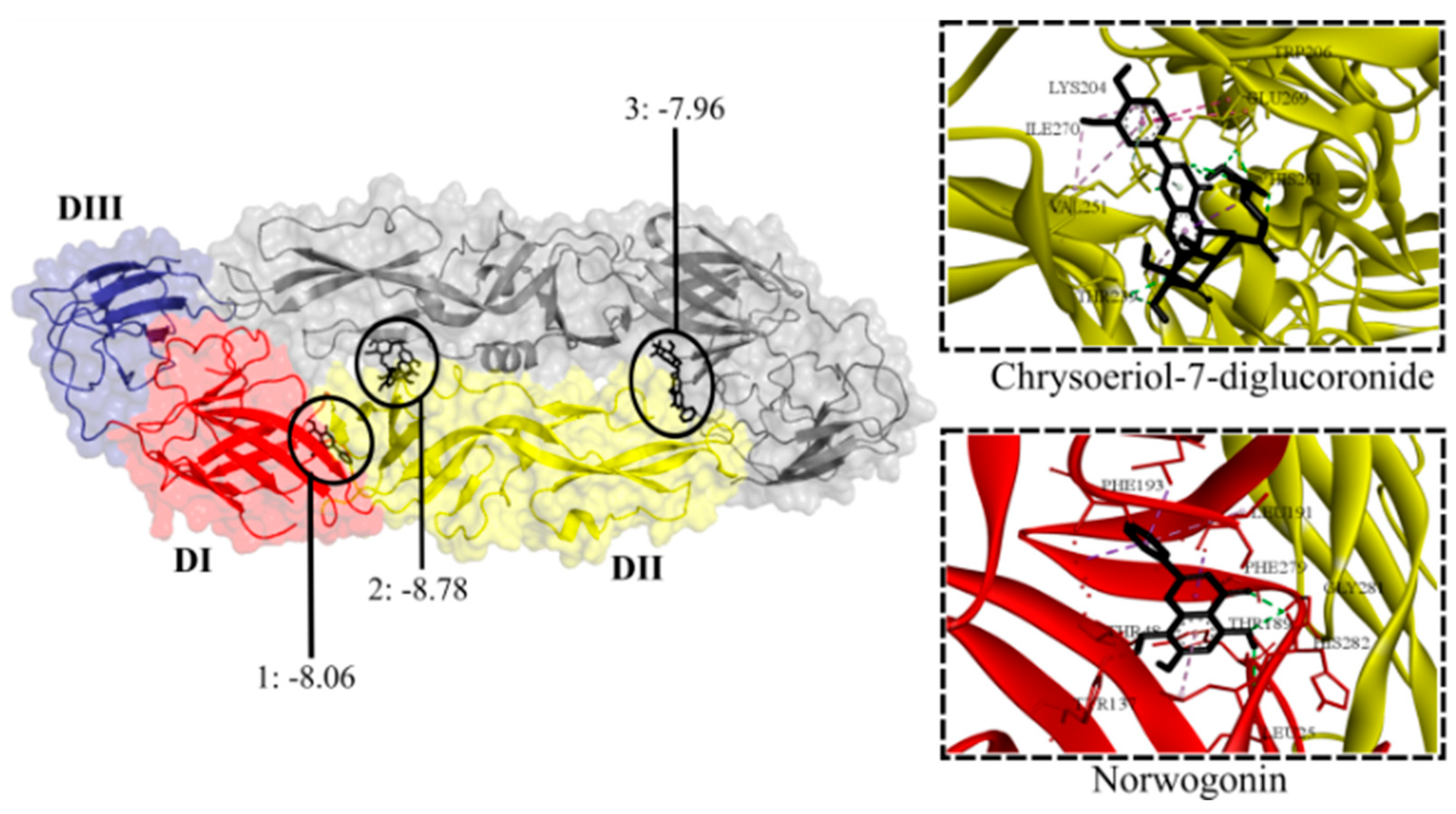
Figure 6.
Contact preferences at the Gas6-Axl complex (PDB ID: 2C5D) of representative flavonoids. Isocarthamidin-glucuronide (1) and chrysoeriol-7-glucuronide (2) within Gas6-Lg2/Lg1 domains; chrysoeriol-7-diglucuronide (3) within Gas6-Lg1 domain; tricin-7-diglucuronoside (4) within Gas6-Lg1/Axl-Ig2 domains; and eriodyctiol-rhamnoside (5) within Gas6-Lg1/Axl-Ig1 domains. Binding free energies (kcal/mol) are shown.
Figure 6.
Contact preferences at the Gas6-Axl complex (PDB ID: 2C5D) of representative flavonoids. Isocarthamidin-glucuronide (1) and chrysoeriol-7-glucuronide (2) within Gas6-Lg2/Lg1 domains; chrysoeriol-7-diglucuronide (3) within Gas6-Lg1 domain; tricin-7-diglucuronoside (4) within Gas6-Lg1/Axl-Ig2 domains; and eriodyctiol-rhamnoside (5) within Gas6-Lg1/Axl-Ig1 domains. Binding free energies (kcal/mol) are shown.
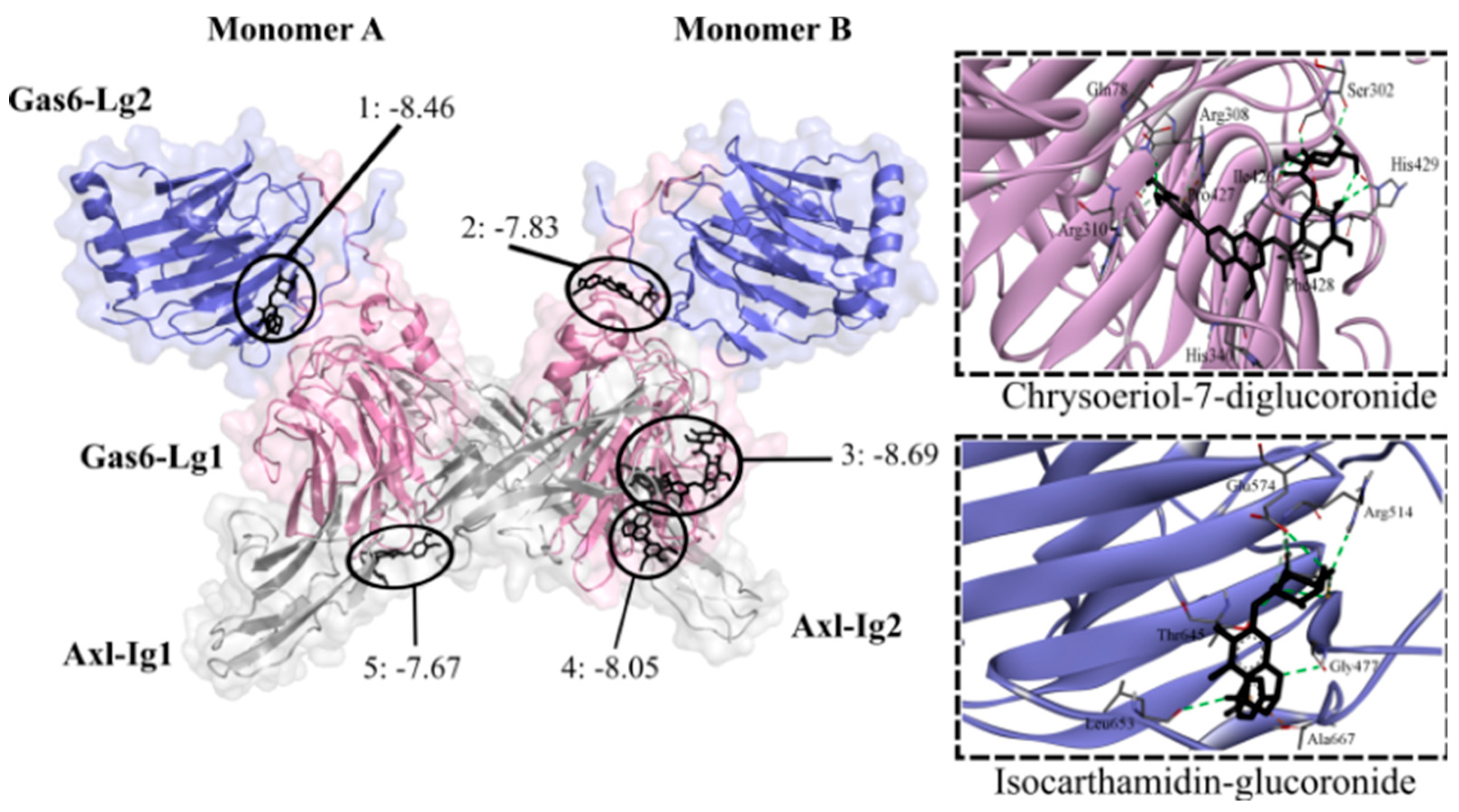
Figure 7.
Contact preferences at the GTPase domain (PDB ID: 2X2E) of representative flavonoids. Chrysoeriol-7-diglucuronide (1) within the G4-switch1-loops; apigenin-7-glucoside (2) closer to the switch1-loop; luteolin-7-glucuronide (3) within switch1-P-loops; dihydrobaicalein-glucuronide (4) closer to the P-loop; isocarthamidin-glucuronide (5) within the switch2-P-loops; and tricin-glucuronide (6) within the switch1-loop. Binding free energies (kcal/mol) are shown.
Figure 7.
Contact preferences at the GTPase domain (PDB ID: 2X2E) of representative flavonoids. Chrysoeriol-7-diglucuronide (1) within the G4-switch1-loops; apigenin-7-glucoside (2) closer to the switch1-loop; luteolin-7-glucuronide (3) within switch1-P-loops; dihydrobaicalein-glucuronide (4) closer to the P-loop; isocarthamidin-glucuronide (5) within the switch2-P-loops; and tricin-glucuronide (6) within the switch1-loop. Binding free energies (kcal/mol) are shown.
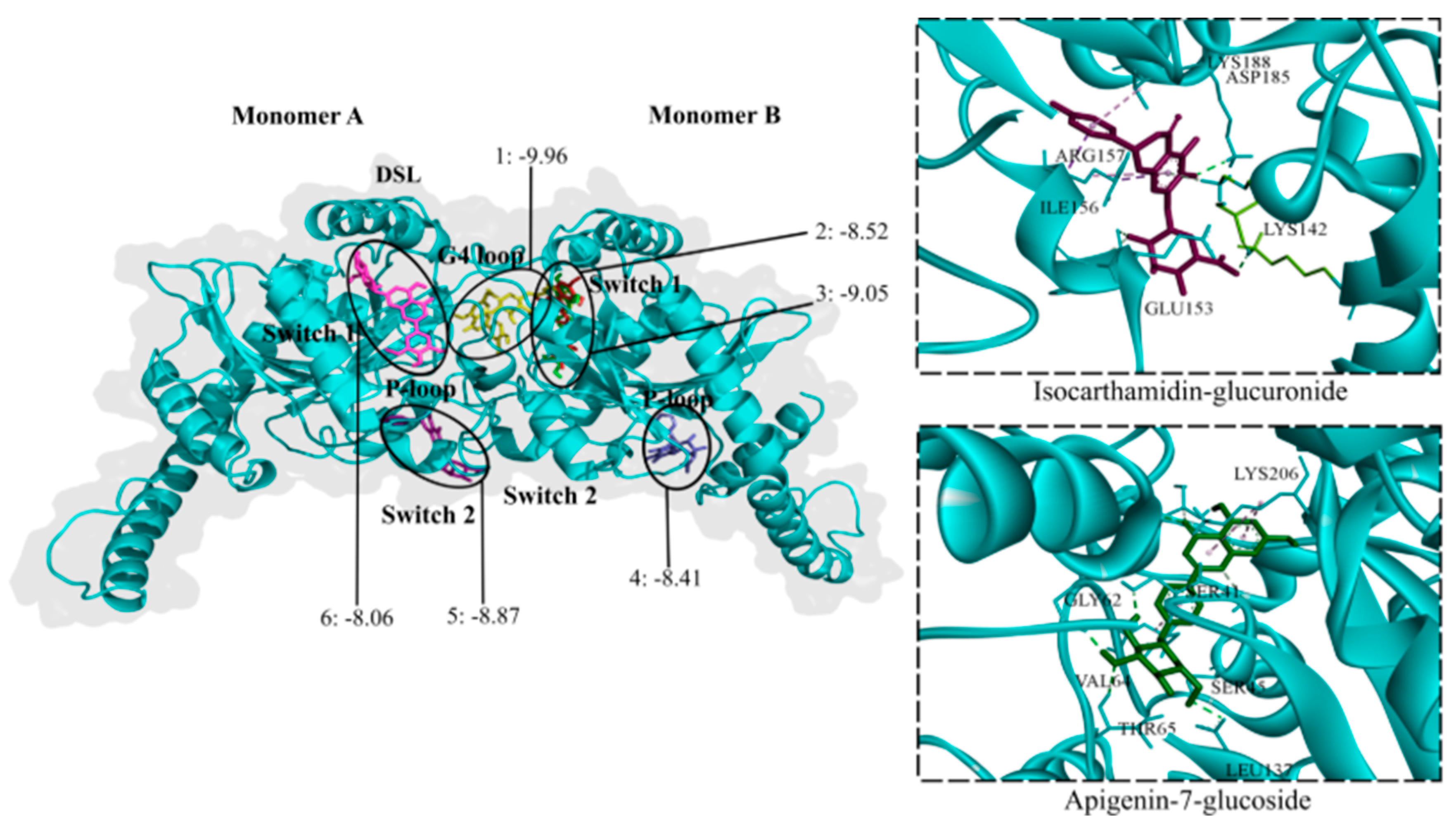
Figure 8.
Contact preferences at the CTD-clathrin domain (PDB ID: 2XZG) of representative flavonoids. Chrysoeriol-7-diglucuronide (1), scutellarin (2), dihydrobaicalein-glucuronide (3), and wogonoside (4) within the W-box motif. Binding free energies (kcal/mol) are shown.
Figure 8.
Contact preferences at the CTD-clathrin domain (PDB ID: 2XZG) of representative flavonoids. Chrysoeriol-7-diglucuronide (1), scutellarin (2), dihydrobaicalein-glucuronide (3), and wogonoside (4) within the W-box motif. Binding free energies (kcal/mol) are shown.
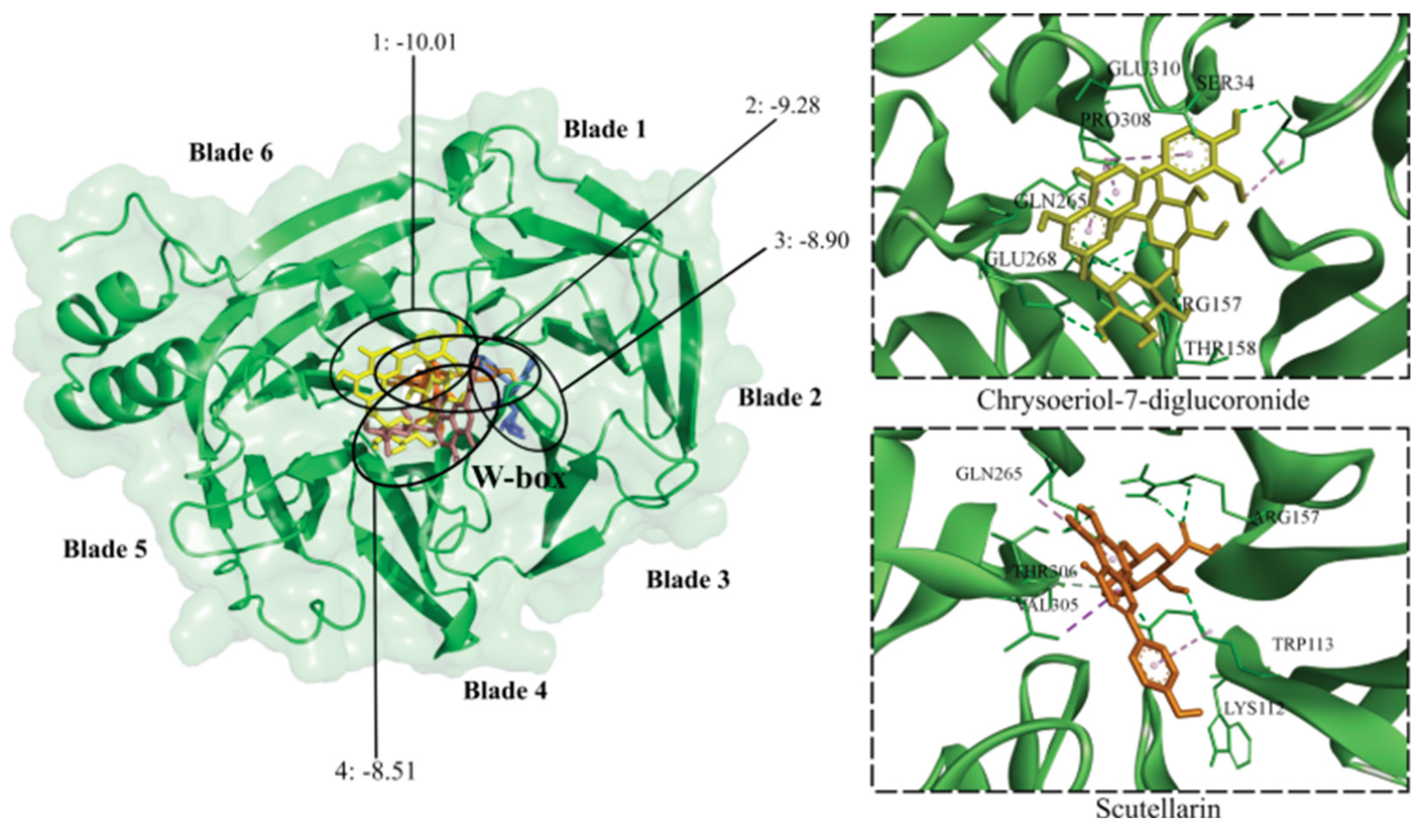

Table 1.
Extracts include in the study.
| Plant | Voucher | Extract code | Extraction | |
|---|---|---|---|---|
| Technique | Conditions | |||
| Scutellaria coccineaKunth | UIS219784 | ScSE | UAE | 15 min; 50 oC |
| Scutellaria incarnata Vent | UIS219783 | SiSE1 | UAE | 5 min; 50 oC |
| SiSE2 | UAE | 60 min; 50 oC | ||
| Scutellaria ventenatii + incarnata (hybrid) | UIS219785 | SviSE | UAE | 60 min; 50 oC |
| Lippia alba (Mill) N.E. Brown | UIS22031a | LacSE | UAE | 60 min; 50 oC |
| UIS22002b | LaiSE | UAE | 60 min; 50 oC | |
| Lippia origanoides Kunth | UIS22035c | LopSE1 | UAE | 23 min; 47 oC |
| LopSE2 | UAE | 60 min; 50 oC | ||
| LopSFE | SFE | 96 min; 307 bar | ||
| UIS22034d | LocSE1 | UAE | 23 min; 47 oC | |
| LocSE2 | UAE | 60 min; 50 oC | ||
| LocSFE | SFE | 96 min; 307 bar | ||
| UIS19799e | LotSE | UAE | 23 min; 47 oC | |
| LotSFE | SFE | 96 min; 307 bar | ||
Lippia chemotypes are based on the major components of their essential oils. a Carvone, b citral, c phellandrene, d carvacrol, and e thymol. UAE, ultrasound assisted solvent extraction. SFE, supercritical fluid extraction. The following extracts were prepared in previous studies: ScSE and SiSE1 [22]; LopSE1 [20]; and LopSFE, LocSFE and LotSFE [21].
Table 2.
Antiviral effect of the extracts on DENV-infected Vero cells.
| Extract Code |
CC50: µg/mL |
DENV-1 | DENV-2 | Effect | |||||
| CPE % |
IC50: µg/mL |
SI |
CPE: % |
IC50: µg/mL |
SI |
||||
| ScSEa | 213 ± 1.1 | 25 ± 1.2 | 52 ± 1.3 | 4.1 | 2.9 ± 0.45 | 3 ± 1.3 | 71 | Strong | |
| SiSE1a | 50 ± 1.2 | 22 ± 1.4 | 39 ± 1.2 | 1.2 | 6.1 ± 0.97 | 17 ± 1.2 | 2.9 | Strong | |
| SiSE2a | 91 ± 1.05 | 79 ± 6.5 | - | - | 70 ± 14.7 | - | - | Inactive | |
| SviSEa | 23 ± 2.0 | 87 ± 4.7 | - | - | 19 ± 6.5 | 65 ± 1.2 | 0.4 | Weak | |
| LaiSEb | 65 ± 1.1 | 39 ± 10.8 | 81 ± 1.1 | 0.8 | 17 ± 8.4 | 38 ± 1.2 | 1.7 | Weak | |
| LacSEb | 75 ± 1.0 | 60 ± 4.3 | - | - | 4 ± 3.7 | 19 ± 1.1 | 3.9 | Weak | |
| LopSE1c | >250.0 | 100 | - | - | 86 ± 3.7 | - | - | Inactive | |
| LopSE2c | 117 ± 1.0 | 93 ± 2.7 | - | - | 77 ± 8.4 | - | - | Inactive | |
| LopSFEc | >250.0 | 100 | - | - | 94 ± 3.2 | - | - | Inactive | |
| LocSE1c | >250.0 | 100 | - | - | 67 ± 11.2 | - | - | Inactive | |
| LocSE2c | 143 ± 1.1 | 100 | - | - | 88 ± 3.2 | - | - | Inactive | |
| LocSFEc | >250.0 | 96 ± 2.5 | - | - | 96 ± 2.0 | - | - | Inactive | |
| LotSEc | >250.0 | 100 | - | - | 84 ± 4.9 | - | - | Inactive | |
| LotSFEc | >250.0 | 99 ± 0.2 | - | - | 88 ± 5.2 | - | - | Inactive | |
a Scutellaria, b L. alba, and c L. origanoides extracts. CC50 >250 µg/mL was adopted for extracts that did not exhibit a 50% reduction in cell viability (Figure 1B). CPE %: percentage of virus-induced cytopathic effect after treatment with 100 µg/mL relative to the untreated control (100% CPE). SI, selectivity index (CC50 / IC50).
Table 3.
Flavonoids in the extracts prepared in the study identified by UHPLC/ESI-Q-Orbitrap-MS.
| Name | Formula | Exp. Masses [M+H]+ |
Δ ppm |
HCD, eV | Product ions | References | ||
|---|---|---|---|---|---|---|---|---|
| Product ions Fragment type |
Formula | (m/z) | ||||||
| Apigenin-7-glucoside | C2120O10 | 433.11287 | 0.11 | 30 | [(M+H)−C6H10O5]+ [(M+H)−C6H10O5−H2O]+ [(M+H)−C6H10O5−CO]+ [(M+H)−C6H10O5−C8H6O]+ [(M+H)−C6H10O5−C8H6O3]+ |
C15H11O5 C15H9O4 C14H11O4 C7H5O4 C7H5O2 |
271.06012 253.04997 243.06558 153.01848 121.02852 |
[19,22] |
| Apigenin-7-glucuronide | C21H18O11 | 447.09056 | 0.54 | 20 | [(M+H)−C6H8O6]+ [(M+H)−C6H8O6−C8H6]+ |
C15H11O5 C7H5O5 |
271.06033 169.01256 |
[19,22] |
| Baicalin | C21H18O11 | 447.09056 | 0.33 | 20 | [(M+H)−C6H8O6]+ [(M+H)−C6H8O6−C8H6]+ |
C15H11O5 C7H5O5 |
271.06033 169.01256 |
[19,22] |
| Chrysoeriol-7-diglucuronide | C28H28O18 | 653.13484 | 0.50 | 20 | [(M+H)−C6H8O6]+ [(M+H)−2C6H8O6]+ |
C22H21O12 C16H13O6 |
477.10275 301.07066 |
[28,31] |
| Chrysoeriol-7-glucuronide | C22H20O12 | 477.10275 | 0.20 | 20 | [(M+H)−C6H8O6]+ | C16H13O6 | 301.07066 | [28,31] |
| Dihydrobaicalein-glucuronide | C21H20O11 | 449.10602 | 0.44 | 10 | [(M+H−)C6H8O6]+ [(M+H)−C6H8O6−C8H8]+ [(M+H)−C6H8O6−C6H6O4]+ |
C15H13O5 C7H5O5 C9H7O |
273.07538 169.01341 131.04932 |
[22] |
| Eriodictyol-7-glucoside | C21H22O11 | 451.12327 | 0.48 | 10 | [(M+H)−C6H10O5]+ [(M+H)−C6H10O5−H2O]+ [(M+H)−C6H10O5−C8H8O2]+ |
C15H13O6 C15H11O5 C7H5O4 |
289.07062 271.05969 153.01810 |
[29,30] |
| Luteolin-7-glucuronide | C21H18O12 | 463.08710 | 1.57 | 30 | [(M+H)−C6H7O6]+ [(M+H)−C6H7O6−H2O]+ |
C21H17O12 C20 13CH17O12 C21H17O11 18O |
461,07211 462,07553 463,07739 |
[29,30] |
| Eriodictyol-rhamnoside | C21H22O10 | 435.12845 | 0.28 | 30 | [(M+H)−C6H10O4]+ [(M+H)−C6H10O4−H2O]+ [(M+H) −C6H10O4−H2O−C6H4O2]+ [(M+H)−C6H10O4−C8H8O2]+ |
C15H13O6 C15H11O5 C9H7O3 C7H5O4 |
289.07086 271.05937 163.03883 153.01846 |
[29] |
| Galangin- glucoside | C21H20O10 | 433.11287 | 0.11 | 20 | [(M+H)−C6H10O5]+ [(M+H)−C6H10O5−C2H2O]+ [(M+H)−C6H10O5−C6H4O2]+ [(M+H)−C6H10O5−C8H6O]+ |
C15H11O5 C13H9O4 C9H7O3 C7H5O4 |
271.06012 229.04935 163.03894 153.01825 |
[20,29] |
| Luteolin-7-glucoside | C21H20O11 | 449.10791 | 1.07 | 20 | [(M+H)−C6H10O5]+ [(M+H)−C6H10O5−C2H2O]+ [(M+H)−C6H10O5−C6H4O2]+ [(M+H)−C6H10O5−C8H6O2]+ |
C15H11O6 C13H9O5 C9H7O4 C7H5O4 |
287.05515 245.04386 179.03411 153.01846 |
[29,30] |
| Luteolin-rutinoside | C27H30O15 | 595.16524 | 0.86 | 10 | [(M+H)−C6H10O4]+ [(M+H)−C6H10O4−C6H10O5]+ [(M+H)−C6H10O4−C6H10O5−C8H6O2]+ [(M+H)−C6H10O4−C6H10O5−C8H6O3]+ |
C21H21O11 C15H11O6 C7H5O4 C7H5O3 |
449.10864 287.05530 153.01832 137.02325 |
[29,30] |
| Phloridzin | C21H24O10 | 437.14386 | 0.82 | 10 | [(M+H)−C6H10O5]+ [(M+H)−C6H10O5−H2O]+ [(M+H)−C6H10O5−C7H6O]+ |
C15H15O5 C15H13O4 C8H9O4 |
275.09131 257.08081 169.04961 |
[20,31] |
| Quercetin-3-glucoside | C21H20O12 | 465.10287 | 0.35 | 10 | [(M+H)−C6H10O5]+ [(M+H)−C6H10O5−H2O]+ [(M+H)−C6H10O5−C8H6O3]+ |
C15H11O7 C15H9O6 C7H5O4 |
303.04971 285.03970 153.01811 |
[20] |
| Scutellarin | C21H18O12 | 463.08762 | 0.42 | 10 | [(M+H)−C6H8O6]+ [(M+H)−C6H8O6−C6H4O3]+ |
C15H11O6 C9H7O3 |
287.05537 163.03885 |
[22] |
| Taxifolin-glucoside | C21H22O12 | 467.11829 | 0.24 | 20 | [(M+H)−C6H10O5]+ [(M+H)−C6H10O5−H2O]+ [(M+H)−C6H10O5−C8H8O3]+ |
C15H13O7 C15H11O6 C7H5O4 |
305.06561 287.05496 153.01832 |
[20] |
| Tricin-7-diglucuronoside | C29H30O19 | 683.14540 | 0.28 | 30 | [(M+H)−C6H8O6]+ [(M+H)−2C6H8O6]+ |
C23H23O13 C17H15O7 |
507.11331 331.08122 |
[31,32] |
| Tricin-glucuronide | C23H22O13 | 507.11331 | 1.03 | 20 | [(M+H)−C6H8O6]+ | C17H15O7 | 331.08063 | [31,32] |
| Wogonoside | C22H20O11 | 461.17773 | 0.47 | 20 | [(M+H)−C6H8O6]+ [(M+H)−C6H8O6−CH3]+• |
C16H13O5 C15H10O5 |
285.07535 270.05164 |
[22] |
| Acacetin | C16H12O5 | 285.07541 | 1.19 | 50 | [(M+H)−CH3]+• [(M+H)−CH3−CO]+• [(M+H)−C9H8O]+ [(M+H)−C8H6O4]+ |
C15H10O5 C14H10O4 C7H5O4 C8H7O |
270.05273 242.05754 153.01842 119.049228 |
[30,31] |
| Apigenin | C15H10O5 | 271.06042 | 0.82 | 40 | [(M+H)−H2O]+ [(M+H)−CO]+ [(M+H)−C8H6O]+ |
C15H9O4 C14H11O4 C7H5O4 |
253.04997 243.06558 153.01848 |
[21,22] |
| Baicalein | C15H10O5 | 271.05969 | 1.50 | 60 | [(M+H)−H2O]+ [(M+H)−H2O−CO]+ [(M+H)−C8H6]+ [(M+H)−C8H6−CO]+ [(M+H)−C8H6−CO−H2O]+ [(M+H)−C7H4O5]+ |
C15H9O4 C14H9O3 C7H5O5 C6H5O4 C6H3O3 C8H7 |
253.04919 225.05435 169.01305 141.01811 123.00773 103.04451 |
[19,22] |
| Chrysoeriol | C16H12O6 | 301.07036 | 0.99 | 30 | [(M+H)−CH3]+• [(M+H)−CH3−CO]+• [(M+H)−C9H8O2]+ [(M+H)−C7H4O4]+ [(M+H)−C8H6O5]+ |
C15H10O6 C14H10O5 C7H5O4 C9H9O2 C8H7O |
286.04718 258.05222 153.01855 149.05977 119.04939 |
[20,21] |
| Cirsimaritin | C17H14O6 | 315.08605 | 0.83 | 30 | [(M+H)−CH3]+• [(M+H)−2CH3]+• [(M+H)−CH3−H2O]+• [(M+H)−CH3−H2O−CO]+• [(M+H)−CH3−H2O−2CO]+• [(M+H)−C8H6O]+ |
C16H12O6 C15H9O6 C16H10O5 C15H10O4 C14H10O3 C9H9O5 |
300.06265 285.03961 282.05215 254.05727 226.06232 197.04445 |
[20,21] |
| Dihydrobaicalein | C15H12O5 | 273.07617 | 0.41 | 40 | [(M+H)−H2O]+ [(M+H)−C2H2O]+ [(M+H)−C8H8]+ [(M+H)−C6H6O4]+ |
C15H11O4 C13H11O4 C7H5O5 C9H7O |
255.06465 231.06482 169.01292 131.04906 |
[22] |
| Eriodictyol | C15H12O6 | 289.07105 | 0.57 | 20 | [(M+H)−H2O]+ [(M+H)−H2O−CO]+ [(M+H)−C6H6O2]+ [(M+H)−H2O−C6H4O2]+ [(M+H)−C8H8O2]+ |
C15H11O5 C14H11O4 C9H7O4 C9H7O3 C7H5O4 |
271.05969 243.05995 179.03374 163.03882 153.01810 |
[20,21] |
| Galangin | C15H10O5 | 271.06051 | 0.49 | 30 | [(M+H)−H2O]+ [(M+H)−C2H2O]+ [(M+H)−C6H4O2]+ [(M+H)−C8H6O]+ [(M+H)−C6H6O3]+ |
C15H9O4 C13H9O4 C9H7O3 C7H5O4 C9H5O2 |
253.04956 229.04935 163.03894 153.01825 145.02840 |
[20,21] |
| Hesperetin | C16H14O6 | 303.08620 | 0.35 | 20 | [(M+H)−H2O]+ [(M+H)−C2H2O]+ [(M+H)−C7H8O2]+ [(M+H)−C9H10O2]+ [(M+H)−C7H4O4]+ |
C16H13O5 C14H13O5 C9H7O4 C7H5O4 C9H11O2 |
285.07529 261.07568 179.03389 153.01828 151.07553 |
[20,21] |
| Luteolin | C15H10O6 | 287.05556 | 0.92 | 30 | [(M+H)−C2H2O]+ [(M+H)−C6H4O2]+ [(M+H)−C8H6O2]+ [(M+H)−C8H6O3]+ |
C13H9O5 C9H7O4 C7H5O4 C7H5O3 |
245.04393 179.03409 153.01822 137.02327 |
[21,24] |
| Nepetin | C16H12O7 | 317.06557 | 0.19 | 20 | [(M+H)−H2O]+ [(M+H)−CH2O]+ |
C16H11O6 C15H11O6 |
299.05501 287.05500 |
[30,31] |
| Naringenin | C15H12O5 | 273.07629 | 0.28 | 20 | [(M+H)−H2O]+ [(M+H)−C2H2O]+ [(M+H)−C8H8O]+ [(M+H)−C6H6O]+ [(M+H)−3C2H2O]+ |
C15H11O4 C13H11O4 C7H5O4 C9H7O4 C9H7O2 |
255.06464 231.06487 153.01810 179.03371 147.04393 |
[20,21] |
| Pinocembrin | C15H12O4 | 257.08132 | 0.20 | 20 | [(M+H)−H2O]+ [(M+H)−C2H2O]+ [(M+H)−C6H6]+ [(M+H)−C8H8]+ |
C15H11O3 C13H11O3 C9H7O4 C7H5O4 |
239.06998 215.07010 179.03415 153.01816 |
[21,24] |
| Quercetin | C15H10O7 | 303.05047 | 0.12 | 20 | [(M+H)−H2O]+ [(M+H)−2CO]+ [(M+H)−H2O−2CO]+ [(M+H)−C8H6O3]+ |
C15H9O6 C13H11O5 C13H9O4 C7H5O4 |
285.03787 247.06012 229.04951 153.01836 |
[20,21] |
| Sakuranetin | C16H14O5 | 287.09132 | 0.28 | 20 | (M+H)−C2H2O]+ [(M+H)−C6H6O]+ [(M+H)−C8H8O]+ [(M+H)−C7H8O3]+ |
C14H13O4 C10H9O4 C8H7O4 C9H7O2 |
245.08090 193.04971 167.03386 147.04402 |
[20,21] |
| Salvigenin | C18H16O6 | 329.10144 | 0.52 | 40 | [(M+H)−CH3]+∙ [(M+H)−CH3−H2O]+∙ [(M+H)−CH3−H2O−CO]+∙ |
C17H14O6 C17H12O5 C16H12O4 |
314.07794 296.06750 268.07200 |
[20,21] |
| Taxifolin | C15H12O7 | 305.06548 | 0.30 | 30 | [(M+H)−H2O]+ [(M+H)−H2O−CO]+ [(M+H)−H2O−2CO]+ [(M+H)−H2O−2CO−H2O]+ [(M+H)−C8H8O3]+ |
C15H11O6 C14H11O5 C13H11O4 C13H9O3 C7H5O4 |
287.05487 259.06003 231.06512 213.05463 153.01826 |
[20,21] |
| Scutellarein | C15H10O6 | 287.05444 | 0.57 | 60 | [(M+H)−H2O]+ [(M+H)−H2O−CO]+ [(M+H)−C8H6O]+ [(M+H)−C8H6O−CO]+ [(M+H)−C8H6O−CO−H2O]+ [(M+H)−C7H4O5]+ |
C15H9O5 C14H9O4 C7H5O5 C6H5O4 C6H3O3 C8H7O |
269.04416 241.04912 169.01280 141.01810 123.00771 119.04921 |
[22] |
| Wogonin | C16H12O5 | 285.07575 | 0.49 | 50 | [(M+H)−CH3]+• [(M+H)−CH3−CO]+• |
C15H10O5 C14H10O4 |
270.05176 242.05701 |
[22] |
| Methyl-apigenin | C16H12O5 | 285.07687 | 0.37 | 30 | [(M+H)−CH3]+• [(M+H)−CH3−CO]+• [(M+H)−C9H8O]+ |
C15H10O5 C14H10O4 C7H5O4 |
270.05219 242.05734 153.01848 |
[20,21] |
| Methyl-galangin | C16H12O5 | 285.07563 | 0.42 | 30 | [(M+H)−CH3]+• [(M+H)−CH3−CO]+• [(M+H)−CH3−CO−HCO]+• [(M+H)−C9H8O]+ |
C15H10O5 C14H10O4 C13H9O3 C7H5O4 |
270.05219 242.05734 213.05461 153.01830 |
[21] |
| Trimethyl-tricetin | C18H16O7 | 345.09659 | 0.80 | 30 | [(M+H)−CH3]+• [(M+H)−2CH3]+ [(M+H)−CH3−H2O]+• [(M+H)−CH3−H2O−CO]+• [(M+H)−C11H12O3]+ |
C17H14O7 C16H11O7 C17H12O6 C16H12O5 C7H5O4 |
330.07339 315.05063 312.06262 284.06769 153.01817 |
[20,31] |
Tentative identification based on comparison with [M+] or [M + H]+ ions reported in previous studies and the literature for Scutellaria spp. [19,22], L. origanoides [20,21,29,30] and L. alba [24,32]. Tentative identification based on comparison with molecule fragmentation pattern in mass spectra and on databases [31].
Table 4.
Amounts (mg/g) of flavonoids in extracts prepared in the study.
| Name | Scutellaria | L. alba | L. origanoides | |||||
|---|---|---|---|---|---|---|---|---|
| SiSE2 | SviSE | LacSE | LaiSE | LopSE2 | LocSE1 | LocSE2 | LotSE | |
| Flavonoid glycosides | ||||||||
| Apigenin-7-glucoside | <LOD | - | <LOD | <LOD | <LOD | 0.04 ± 0.01 | <LOD | 0.62 ± 0.2 |
| Apigenin-7-glucuronidea | - | - | 1.5 ± 0.09 | 2.3 ± 0.19 | 14 ± 1.00 | - | 27 ± 1.40 | - |
| Baicalin | 15.9 ± 0.02 | 16 ± 0.02 | - | - | - | - | - | - |
| Chrysoeriol-7-glucuronide | - | - | 3.8 ± 0.05 | 3.5 ± 0.01 | - | - | - | - |
| Chrysoeriol-7-diglucuronide | - | - | 4.9 ± 0.07 | 2.5 ± 0.03 | - | - | - | - |
| Dihydrobaicalein-glucuronidea | 23 ± 0.006 | <LOD | - | - | - | - | - | - |
| Eriodictyol-7-glucoside | - | - | - | - | 80 ± 2.00 | 0.36 ± 0.03 | 89 ± 1.30 | 3.2 ± 0.3 |
| Eriodictyol-rhamnoside | - | - | - | - | - | 0.13 ± 0.01 | - | 0.32 ± 0.03 |
| Galangin-glucoside | - | - | - | - | - | <LOD | - | - |
| Luteolin-7-glucoside | - | - | 0.6 ± 0.11 | 0.68 ± 0.04 | 49 ± 1.00 | 0.25 ± 0.02 | 35 ± 0.10 | 5.01 ± 0.3 |
| Luteolin-7-glucuronide | - | - | 0.8 ± 0.19 | 1.1 ± 0.33 | - | - | - | - |
| Luteolin-rutinoside | - | - | - | - | - | 0.02 ± 0.01 | - | 0.10 ± 0.01 |
| Quercetin-3-glucoside | - | - | - | - | <LOD | 1.5 ± 0.3 | <LOD | 6.2 ± 0.8 |
| Scutellarin | 11.4 ± 0.03 | 11.4 ± 0.03 | - | - | - | - | - | - |
| Taxifolin-glucoside | - | - | - | - | - | <LOD | - | <LOD |
| Tricin-glucuronide | - | - | 3.7 ± 0.12 | 2.8 ± 0.03 | - | - | - | - |
| Tricin-7-diglucuronoside | - | - | 5.5 ± 0.13 | 3.8 ± 0.04 | - | - | - | - |
| Wogonoside | 4.7 ± 0.03 | 4.7 ± 0.03 | - | - | - | - | - | - |
| Phloridzin | - | - | - | - | - | 0.01 ± 0.01 | - | 0.05 ± 0.04 |
|
Flavonoid aglycones |
||||||||
| Acacetin | - | - | <LOD | <LOD | - | - | ||
| Apigenin | - | - | 0.34 ± 0.06 | 1.2 ± 0.39 | - | 0.13 ± 0.01 | ||
| Baicalein | 10.1 ± 0.04 | 10 ± 0.04 | - | - | - | - | - | - |
| Chrysoeriol | - | - | - | - | - | 0.70 ± 0.04 | ||
| Cirsimaritin | - | - | 0.31 ± 0.02 | 0.40 ± 0.14 | - | 0.9 ± 0.04 | 4.3 ± 0.10 | 0.38 ± 0.01 |
| Baicalein | 10.1 ± 0.04 | 10 ± 0.04 | - | - | - | - | - | - |
| Chrysoeriol | - | - | - | - | - | 0.70 ± 0.04 | - | 2.3 ± 0.1 |
| Cirsimaritin | - | - | 0.31 ± 0.02 | 0.40 ± 0.14 | - | 0.9 ± 0.04 | - | 2.9 ± 0.2 |
| Dihydrobaicaleinc | 4.13 ± 0.007 | <LOD | - | - | - | - | ||
| Eriodictyol | - | - | - | - | 89 ± 1.00 | 4.5 ± 0.03 | ||
| Galangin | - | - | - | - | 50 ± 2.00 | <LOD | ||
| Hesperetin | - | - | - | - | - | 0.17 ± 0.01 | ||
| Luteolin | - | - | <LOD | <LOD | 7.3 ± 0.20 | 1.12 ± 0.05 | ||
| Naringenin | - | - | <LOD | <LOD | 6.0 ± 0.20 | 1.38 ± 0.05 | ||
| Nepetin | - | - | <LOD | <LOD | - | - | ||
| Pinocembrin | - | - | - | - | 71 ± 2.00 | 0.03 ± 0.01 | ||
| Quercetin | - | - | - | - | <LOD | 1.5 ± 0.1 | ||
| Sakuranetin | - | - | - | - | <LOD | 0.32 ± 0.02 | ||
| Salvigenin | - | - | <LOD | <LOD | - | - | ||
| Scutellarein | 0.2 ± 0.2 | 0.2 ± 0.20 | - | - | - | - | ||
| Taxifolin | - | - | - | - | 4.8 ± 0.10 | 0.47 ± 0.02 | ||
| Wogonin | 15.4 ± 0.03 | 15 ± 0.03 | - | - | - | - | ||
| Methylated flavonoids | ||||||||
| Methylapigenin | - | - | - | - | - | 0.03±0.01 | - | |
| Methylgalangin | - | - | - | - | <LOD | <LOD | - | |
| Trimethyltricetin | - | - | - | - | - | 0.03±0.01 | - | |
All extracts were prepared using the UAE technique (Table 1). Extracts fom S. incarnata (SiSE2), S. ventenatti + S. incarnata (SviSE), L. alba carvone (LacSE) and citral (LaiSE) chemotypes; and L. origanoides phellandrene (LopSE2), carvacrol (LocSE1 and LocSE2) and thymol (LotSE) chemotypes. a Quantification in baicalin; b quantification in wogonoside; c quantification in baicalein equivalents; d quantification in 1,3-dicafeoylquinic acid equivalents. LOD: limit of detection. Wogonoside (LOD = 0.05 mg/g), eriodyctiol-7-glucoside (LOD = 0.04 mg/g), baicalin (LOD = 0.80 mg/g), and scopoletin (LOD = 0.05 mg/g).
Table 5.
Antiviral effect and relative concentrations of flavonoid classes of the extracts.
| Extract |
Antiviral effect |
Total (mg/g) |
Glycoside (%) | Aglycone (%) |
Methylated (%) |
||||||||
| A | B | C | D | A | B | C | |||||||
| ScSE | Strong | 277.86 | 83.4 | 4.1 | 0 | 0.56 | 10.3 | 0 | 0 | 0 | |||
| SiSE1 | Strong | 492.98 | 86.1 | 2.5 | 0 | 0 | 5.0 | 0 | 0 | 0 | |||
| SviSE | Weak | 57.92 | 55.5 | 0 | 0 | 0 | 44.5 | 0 | 0 | 0 | |||
| LacSE | Weak | 21.37 | 97.0 | 0 | 0 | 0 | 3.0 | 0 | 0 | 0 | |||
| LaiSE | Weak | 18.34 | 90.7 | 0 | 0 | 0 | 9.3 | 0 | 0 | 0 | |||
| SiSE2 | Inactive | 84.92 | 64.8 | 0 | 0 | 0 | 35.2 | 0 | 0 | 0 | |||
| LopSE1 | Inactive | 27.58 | 24.7 | 15 | 37 | 0.9 | 3.9 | 10.1 | 8.0 | 0.4 | |||
| LopSE2 | Inactive | 371.10 | 17 | 21.6 | 0 | 0 | 2.0 | 44.7 | 14.8 | 0 | |||
| LocSE1 | Inactive | 13.59 | 2.3 | 3.6 | 11.0 | 0.07 | 21.0 | 47.1 | 14.5 | 0.4 | |||
| LocSE2 | Inactive | 259.00 | 23.9 | 34.4 | 0 | 0 | 1.7 | 38.2 | 1.8 | 0 | |||
| LotSE | Inactive | 57.44 | 10 | 6.1 | 10.8 | 0.1 | 15.1 | 41.4 | 16.2 | 0.2 | |||
| LopSFE | Inactive | 62.96 | 0 | 0 | 0 | 0 | 0.2 | 81.4 | 11.8 | 6.5 | |||
| LocSFE | Inactive | 19.85 | 0 | 0 | 0 | 0 | 21.4 | 70.7 | 6.0 | 1.9 | |||
| LotSFE | Inactive | 25.90 | 0 | 0 | 0 | 0 | 25.8 | 68.0 | 0.8 | 5.4 | |||
Table 6.
Binding mode predicted for the predominant flavonoids in extracts from Scutellaria species and L. alba to targets. Protein (PDB ID): Cl, clathrin N-terminal domain (2XZG); DENV-2 E, envelope (1OAN); DY, dynamin GTPase domain (2X2E); GX: Gas6-Axl receptor (2C5D).
Table 6.
Binding mode predicted for the predominant flavonoids in extracts from Scutellaria species and L. alba to targets. Protein (PDB ID): Cl, clathrin N-terminal domain (2XZG); DENV-2 E, envelope (1OAN); DY, dynamin GTPase domain (2X2E); GX: Gas6-Axl receptor (2C5D).
| Name/PubChem CID/formula | Target: site | Amino acids interacting through hydrogen bonds | kcal/mol | |||
|---|---|---|---|---|---|---|
| Chrysoeriol-7-diglucoronide 44258206  |
Cl: W-box | Ser34, Thr158, Gln265, Glu268, Arg157. | -10.01 ± 0.27 | |||
| DY: G4-switch1 | Ser41, Gly60, Lys113, Ala177, Asp180, Leu181, Lys206, Thr214, Asn236, Gln239. | -9.96 ± 0.44 | ||||
| E: DII (A/B) | Trp206, Thr239, His261, Thr265, Glu269, Ile270. | -8.78 ± 0.37 | ||||
| GX: Gas6-Lg1 | Gln78, Ser304, Gly307, Arg308, Leu309, Gln341, Ile426, Phe428, His429. | -8.69 ± 0.18 | ||||
| Baicalin 64982  |
DY: G4-switch1 | Ser41, Gly60, Ser61, Ala177, Asp180, Lys206, Thr214. | -9.67 ± 0.17 | |||
| Cl: W-box | Glu33, Lys112, Asn155, Arg157, The158, Met264, Gln265, Thr306 | -9.33 ± 0.49 | ||||
| GX: Gas6-Lg1/Lg2 | Met644, Thr645, Asp654, Leu655, Ala667. | -7.96 ± 0.18 | ||||
| E: DII/DI | Arg2, Glu44, Ile46, Asp98. | -7.96 ± 0.22 | ||||
| Tricin-7-diglucuronoside 131752191  |
Cl: W-box | Lys112, Arg157, Ser200, Phe201, Met264, Gln265, Ile266, Glu268, Thr306, Gly314. | -9.45 ± 0.17 | |||
| E: DII (A/B) | Leu65, Asp203, Lys204, Ala205, Val252, His261, Glu269. | -8.29 ± 0.21 | ||||
| DY: P-loop | Gln33, Arg67, Arg107, Gln117, Asn126, Asp130, Leu131, Lys166. | -7.88 ± 0.14 | ||||
| GX: Gas6-Lg1/Axl-Ig2 | Gln78, Ser302, Gly303, Arg308, Leu309, Gln341, Ile426, Phe428, His429. | -8.05 ± 0.20 | ||||
| Scutellarin 185617 
|
Cl: W-box | Trp111, Lys112, Trp113, Arg157, Thr158, Met264, Gln265, Thr306. | -9.28 ± 0.39 | |||
| DY: G4-switch1 | Ser61, Asp180, Leu181, Lys206, Thr214. | -8.31 ± 0.16 | ||||
| E: DII (A/B) | Asp203, Lys204, Ala205, Thr262, Leu264, Thr265, Glu269. | -8.13 ± 0.32 | ||||
| GX: Gas6-Lg1/Lg2 | Glu331, Asn438, Arg476, Gly477, Asp654, Glu657, Ala667. | -7.80 ± 0.17 | ||||
| Apigenin-7-glucoronide 5319484 
|
Cl: W-box | Lys112, Thr158, Gln265, Thr306 | -9.17 ± 0.27 | |||
| DY: G4-switch1 | Ser41, Ala42, Gly43, Lys44, Ser45, Ser46, Gly60, Val64, Thr65, Gln239. | -8.70 ± 0.16 | ||||
| E: DII (A/B) | Asp203, Lys204, Leu264, Thr262, Glu269. | -8.22 ± 0.17 | ||||
| GX: Gas6-Lg1/Lg2 | Phe328, Asn438, Arg476, Gly477, Asp654, Glu657, Ala667, His668. |
-7.82 ± 0.17 | ||||
| Luteolin-7-glucuronide 5280601  |
Cl: W-box | Glu33, Ser70, The109, Lys112, Ile153 | -9.10 ± 0.26 | |||
| DY: G4-switch1 | Ser41, Ala42, Gly43, Lys44, Ser46, Arg59, Gly60, Val64, Thr65, Lys206, Leu207, Asn236. | -8.52 ± 0.18 | ||||
| E: DII (A/B) | Lys204, Ala205, Thr265, Ala267, Glu269 | -8.41 ± 0.15 | ||||
| GX: Gas6-Lg2 | Gly477, Arg514, Asp654, Ala667. | -8.25 ± 0.21 | ||||
| Tricin-glucuronide 101939793 
|
Cl: W-box | Lys112, Arg157, Ser200, Phe201, Met264, Gln265, Ile266, Glu268, Thr306, Gly314. | -9.45 ± 0.17 | |||
| GX: Gas6-Lg1/Lg2 | Gln78, Ser302, Gly303, Arg308, Leu309, Gln341, Ile426, Phe428, His429. | -8.05 ± 0.20 | ||||
| E: DII (A/B) | Leu65, Asp203, Lys204, Ala205, Val252, His261, Glu269, Trp206. | -8.29 ± 0.20 | ||||
| DY: P-loop | Gln117, Asn126, Asp130, Leu131, Gln33, Arg67, Arg107, Lys166. | -7.88 ± 0.14 | ||||
| Dihydrobaicalein-glucuronide 14135324  |
Cl: W-box | Ser28, Ser70, Thr109, Gln152, Ile153. | -8.90 ± 0.27 | |||
| DY: P-loop | Gln17, Asn26, Asn121. | -8.41 ± 0.20 | ||||
| GX: Gas6-Lg2 | Gln78, Arg308. | -7.99 ± 0.20 | ||||
| E: DI/DII | Gly102, Asn103, His149, Gly152, Asn153, Asp154, Thr155. | -7.72 ± 0.34 | ||||
| Isocarthamidin-glucuronide 101274424  |
DY: switch2-P | Lys142, Glu153, Lys188. | -8.87 ± 0.26 | |||
| Cl: W-box | Trp113, Arg157, Met264, Gln265, Glu268, Thr30. | -8.78 ± 0.35 | ||||
| GX: Gas6-Lg1/Lg2 | Arg476, Gly477, Ser478, Arg514, Thr645. | -8.46 ± 0.18 | ||||
| E: DI/DII | Asn103, Val151, Asp154, Thr155, His244, Lys246. | -7.97 ± 0.24 | ||||
| Chrysoeriol-7-glucuronide 14630700 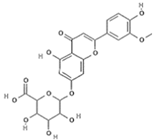 |
DY: G4-switch1 | Ser41, Val64, Thr65, Ser179, Asp180, Lys206, Thr214, Asp215. | -8.85 ± 0.43 | |||
| Cl: W-box | Met32, Asn155, Gln265, Thr306, Glu301. | -8.85 ± 0.27 | ||||
| E: DII (A/B) | Lys204, Trp206, Thr262, Leu264, Thr265, Glu269. | -8.37 ± 0.16 | ||||
| GX: Gas6-Lg1/Lg2 | Lys290, Arg467, Ser663, Asp664. | -7.83 ± 0.16 | ||||
| Luteolin-7-glucoside 5280637 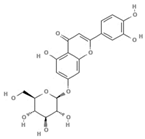
|
Cl: W-box | Ser70, Thr109, Lys112, Ile153. | -8.54 ± 0.26 | |||
| GX: Gas6-Lg2 | Ser478, Arg514, Cys643, Asp654, His668. | -8.18 ± 0.19 | ||||
| DY: G4-switch1 | Gly60, Ala177, Lys206, Leu209, Asp211, Asp236. | -8.00 ± 0.24 | ||||
| E: DII (A/B) | Lys204, Ala205, Thr262, Thr265. | -7.98 ± 0.17 | ||||
| Wogonoside 3084961  |
Cl: W-box | Arg157, Thr158, Ala160, Gln162, Phe201, Gln203, Glu268. | -8.51 ± 0.20 | |||
| E: DII (A/B) | Trp206, Gln256, Gly258, Leu264, Thr265, Glu269. | -7.94 ± 0.24 | ||||
| DY: G4-switch1 | Arg67, His85, Arg107, Asn121. | -7.69 ± 0.19 | ||||
| GX: Gas6-Lg1/Lg2 | Gln78, Leu309, Arg310, Gln341. | -7.69 ± 0.12 | ||||
| Baicalein 5281605 
|
DY: switch2-P | Thr141, Glu153, Arg157, Asp185. | -8.26 ± 0.36 | |||
| Cl: W-box | Asn155, Arg157, Val262, Met264, Gln265, Thr306. | -8.08 ± 0.47 | ||||
| E: βOG pocket | His27, Thr48, Gly281, His282. | -7.92 ± 0.76 | ||||
| Dihydrobaicalein 9816931 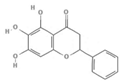
|
DY: switch2-P | Thr141, Glu153, Arg157, Asp185. | -8.25 ± 0.40 | |||
| Cl: W-box | Ser70, Lys83, Trp111. | 7.97 ± 0.45 | ||||
| E: βOG pocket | Leu25, His27, Thr48, Gly281, His282. | -7.84 ± 0.80 | ||||
| Wogonin 5281703 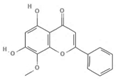 |
E: βOG pocket | Thr48, Tyr137, Gly281, His281. | -7.66 ± 0.80 | |||
Both apigenin-7-glucoronide and luteolin-7-glucoside were found in extracts from L. origanoides.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



