
Submitted:
18 October 2025
Posted:
20 October 2025
You are already at the latest version
Abstract
Urban parks serve a vital function in delivering essential ecosystem services, including biodiversity maintenance and the improvement of urban well-being. This study explores how tree stand characteristics impact the hemeroby, naturalness, and functional diversity of herbaceous cover in two parks in Dnipro, Ukraine. Utilizing structural equation modelling we identified four key gradients influencing herbaceous diversity: stand density/thinning, diversity/uniformity, cooling potential, and heating potential. Higher tree density and diversity lead to cooler temperatures and increased soil and air humidity, which in turn lessens hemeroby and promotes naturalness. Reduced light intensity under denser canopies bolsters the establishment of natural herbaceous communities. Hemeroby and naturalness also impact functional diversity, which can be divided into richness, evenness, and divergence. Environmental disturbances often reduce functional richness while increasing evenness and divergence; in contrast, greater naturalness is related to increased evenness but decreased richness. Our results highlight the importance of tree stand structure in shaping microclimate and functional diversity within urban parks. A thorough understanding of these interactions is crucial for developing park management strategies that enhance biodiversity and ecosystem stability. By merging ecological theory with urban planning, this study provides a framework for sustainable management of green spaces, ensuring lasting ecological and social benefits.
Keywords:
hemeroby
; naturalness
; urban green space
; functional traits
; structural equation modelling
1. Introduction
Urban parks provide numerous ecosystem services that enhance the environment and quality of life in urban areas [1]. They hold significant aesthetic and cultural value and contribute to the conservation of biodiversity [2]. Parks assist in alleviating the urban heat island effect, decreasing stormwater runoff, absorbing pollutants like gases and dust, and can also capture heavy metals [3]. One of the critical ecological functions of parks is the removal of greenhouse gases, mainly through the ability of trees to absorb carbon dioxide [4]. The influence of tree stands on other ecosystem elements, including microclimatic conditions, soil composition, and herbaceous cover, is crucial for the overall functioning of park stands [5]. Urban parks provide a variety of ecosystem services, including the purification of water and air, the reduction of wind and noise, carbon sequestration, microclimate regulation, habitat provision for wildlife, recreational opportunities and the establishment of conditions that promote the social and psychological well-being of city residents [6]. The quantity and quality of these ecosystem services depend on the structure of the park’s plantings. A diverse array of tree-planting strategies can be employed to optimize specific ecosystem services, such as air purification and mitigating urban heat island effects [7]. However, these strategies must also consider how a park’s stand structure influences the functioning of other park components, which can either facilitate or hinder the intended outcomes of park design.
The forest canopy significantly influences the environment by determining light availability, resource allocation, microclimate variability, and biotic interactions [8]. These interactions, in turn, shape the functional diversity of herbaceous plants [9]. The tree canopy in urban parks, comprising natural forests and artificial tree plantations, significantly influences environmental regimes [10]. The density and species composition of the tree canopy in urban parks significantly affect illumination levels [11]. A dense tree cover creates favorable conditions for herbaceous species that are adapted to shady environments [12], while a sparse tree cover fosters an environment that supports light-loving species, thereby enhancing plant community diversity [13,14]. The accumulation of leaves and other organic residues beneath the canopy forms a litter layer that significantly influences the composition and productivity of herbaceous plants [15]. Nutrient-rich tree litter promotes the growth of herbaceous species with high ecological demands for soil fertility [16], while dense tree litter layers can inhibit the germination of less competitive species [17]. Competition for resources between trees and herbaceous plants can restrict access to water and minerals for less resilient species [18], thereby altering the functional diversity of the herbaceous cover [19]. The microclimatic effects of the tree canopy regulate temperature and humidity [20], creating favourable conditions for moisture-loving and temperature-sensitive herbaceous species [21]. In urban parks with a more diverse tree composition, a mosaic of conditions is established that supports the growth of herbaceous species with varying ecological adaptations [22]. The predominance of homogeneous plantations or species with allelopathic effects can limit the functional diversity of herbaceous vegetation [23]. Furthermore, it is crucial to recognize the significant anthropogenic pressures that parks face, as reflected in the concepts of hemeroby and naturalness. Anthropogenic factors, such as recreational pressure and changes in the physical state and chemical composition of the soil and climate, interact with the influence of the tree canopy to affect the functional diversity of the herbaceous cover. The term ‘hemeroby’ is employed to indicate the degree of anthropogenic transformation of the ecosystem, whereas ‘naturalness’ is used to indicate the level of departure from the most natural state of the ecosystem [24]. Further research is needed to elucidate the impact of hemeroby and naturalness on the ecosystem functions of urban parks [25]. Urban parks possess the capacity to resist hemeroby, and their naturalness is a source of enhanced beneficial ecosystem services.
The diversity of herbaceous communities within parks plays a crucial role in ensuring the efficiency and sustainability of ecosystem functions [26], and holds significant aesthetic value in urban settings [27]. The varied perennial herbaceous communities found in urban environments can contribute to biodiversity conservation, water resource preservation, and the protection of culturally significant flora [28]. The presence and biodiversity of herbaceous cover in urban parks create opportunities for physical activity and recreation, which, in turn, provide health benefits that positively affect the immune function of visitors [29]. Parks and green spaces should be designed to provide adequate leisure facilities, as well as opportunities for physical activity. This can be accomplished by ensuring that sufficient herbaceous cover is available for individuals engaged in sedentary leisure activities [30]. The aesthetic quality of urban parks is enhanced by the presence of trees arranged in strips or groups, rather than by the uniform coverage of grass [31]. Limitations in solar radiation due to variations in tree density can diminish the recreational and aesthetic functions of park plantations and hinder the development of herbaceous cover [32]. Therefore, further investigations are necessary to assess the relationships between tree stands and herbaceous cover in parks, with the aim of understanding the role of these elements in providing ecosystem services.
The herbaceous cover observed in urban parks can be categorized into two main types: natural and adventive [33]. The proportions of these categories indicate the extent to which the plant communities deviate from their natural state. The presence of non-native plant species in urban parks results from the availability of unoccupied niches, as well as the relatively favourable soil and water conditions that support plant survival. Non-native plants can be found in various elements of urban parks, including lawns, flower beds, tree plantations, and hedges [34,35]. The proliferation of non-natives in urban parks is a natural phenomenon that occurs without the need for additional financial or labour resources. However, the ecological benefits of these plants should not be overlooked. They play crucial roles in mitigating urban heat, sequestering carbon, preventing erosion, retaining stormwater, and regulating noise levels [36]. Furthermore, weeds provide a habitat for wildlife [37]. The phenomenon of global climate change has led to a significant expansion of summer weather patterns, which has, in turn, facilitated the spread of non-native plant species in urban parks. This proliferation of non-native plants may also have beneficial effects on the enhancement of ecosystem services in a warming climate [38]. Biological traits influence the distribution of spontaneous plants within urban tree plantations [39]. The functional diversity of parks is responsive to the landscape matrix [40].
The functional traits of a species are defined as morphological, physiological, or phenological features that directly or indirectly influence an organism’s functional capacity, including its ability to grow, reproduce, and survive [41]. An examination of the functional traits of plants reveals their capacity to adapt to diverse environmental conditions, thereby elucidating the various biological strategies employed by species for dispersal and reproduction [42]. These functional traits serve as the foundation for calculating indices of functional diversity within communities [43]. The functional diversity of plant communities serves as an indicator of their capacity to adapt to environmental conditions and to perform essential ecosystem functions. The way of distribution of seeds and fruits or their morpho-physiological features illustrate the influence of various environmental factors on plant reproduction and the interconnectivity between plants and animals [44]. The observed changes in leaf morphology may indicate degradation processes induced by nutrient enrichment or salinity [45]. Environmental disturbances lead to abiotic filtration, resulting in the formation of communities dominated by species that have adapted to these perturbations and share similar characteristics [46]. The invasive nature of a plant can be assessed through various traits. Invasive species often grow more rapidly than native species. Additionally, these invasive plants frequently exhibit a larger specific leaf area, a longer flowering period, greater fecundity, and more efficient seed dispersal [47]. The functional traits of invasive plant species serve as proxies for evaluating their impacts on a range of ecosystem services [48]. Numerous metrics for measuring functional diversity have been developed, which can be categorized into three groups, each assessing a different aspect of functional diversity: functional richness, functional evenness, and functional divergence [49]. Although the theoretical foundation for indicators of functional diversity is well established, there is a lack of empirical evidence regarding their actual behaviour across ecological gradients. There is limited information on changes in the indices of functional diversity of plant communities along the hemeroby–naturalness gradient. The impact of tree stands in park plantations on the functional diversity of herbaceous cover in parks remains largely unknown. Addressing these issues is crucial for gaining a deeper understanding of the dynamics of diversity and the functions of urban parks as holistic ecosystems, as well as for justifying optimal park management strategies.
The aims of this study were to address the following questions: 1) How do the structural components of the stand influence the impact of tree plantations on microclimatic conditions in the park? 2) How do tree plantations affect the hemeroby and naturalness of the herbaceous cover in urban parks? 3) How do hemeroby and naturalness influence the variability in functional diversity of herbaceous cover parks?
2. Materials and Methods
2.1. Study Location
The study was conducted in two parks located in the city of Dnipro, Ukraine: Ivan Starov Square and Students’ Park, each covering an area of 11.2 hectares. Ivan Starov Square (48°27′N, 35°3′E) is situated in the historical centre of the city and has experienced a significant amount of recreational activity. In contrast, Students’ Park (48°25′N, 35°2′E) is located on the campus of Oles Honchar Dnipro National University, resulting in a considerably lower level of recreational use. The Walter–Lieth diagram (Walter & Lieth, 1960) was constructed using the climatol library on the basis of data from the Center for Environmental Data Analysis (https://data.ceda.ac.uk/). The mean annual precipitation for the period from 2018 to 2023 was 498 mm. The highest mean daily temperature recorded in August was 30.2 °C. The mean annual temperature is 11.1 °C (Figure S1).
The vegetation cover in Ivan Starov Square consists of two distinct layers: trees and herbaceous plants. The sample locations were distributed throughout the study area, and their exact coordinates were determined using GPS (Garmin eTrex, ± 5 m). At each sample site, the species composition and number of trees within a 5-meter radius were recorded, along with the number of herbaceous plants. The environmental characteristics within samples sites were also measured. In Ivan Starov Square, vegetation cover and environmental properties were measured at 150 sample sites arranged in a quasi-regular grid (Figure 1). The topography of Ivan Starov Square is relatively flat, with elevations ranging from 115 to 128 meters. Additionally, the park management practices are relatively uniform. The sample site density in Ivan Starov Square was one location per 748 m2. In contrast, the geomorphological characteristics of Students’ Park are highly diverse, with terrain elevations ranging from 131 to 165 meters. The park has also undergone partial reconstruction [1,50,51]. To accommodate the observed heterogeneity, the density of the sample sites was increased. In Students’ Park, vegetation cover and environmental measurements were taken at 230 sample sites arranged in a quasi-regular grid, resulting in a survey density of one sample site per 501 m2.
2.2. Herbaceous Layer Plants Recording
The presence of all species of vascular plants in the herbaceous layer of the 3 × 3 m relevé plot in the centre of each sample site was recorded during the study period, which spanned from July 7 to July 12, 2024. The projected species cover of the herbaceous plants is expressed as a percentage. Critical specimens were collected and identified using microscopy. Plant taxonomy was based on the Euro+Med Plantbase (http://ww2.bgbm.org/EuroPlusMed).
2.3. Measurement of Soil and Microclimatic Properties
The soil moisture content was measured using an MG–44 soil moisture meter (Ukraine), with readings taken at a depth of 5–7 cm. This device has a measurement resolution of 0.1% and an associated error margin of 1%. The soil temperature within the 7–10 cm layer was assessed using a digital thermometer (TC–3M, Ukraine). The air temperature and atmospheric humidity at a height of 1.5 m were recorded with a HUATO HE–173 temperature and humidity logger (China). Illuminance at the same height was measured using an RSE–174 lux meter (Germany). The electrical conductivity of the soil was measured in situ with an HI 76305 sensor (Hanna Instruments, Woonsocket, RI), which operates in conjunction with a portable HI 993310 tester. This instrument is designed to evaluate the total electrical conductivity of the soil, which is defined as the combined conductivity of air, water, and soil particles. The results from this device are presented in units of soil salt concentration, expressed in grams per litter. A comparison of the HI 76305 measurements with laboratory data allowed for the estimation of the unit conversion factor, which is 1 dS/m = 155 mg/l [52,53]. The data on the environmental properties were subjected to principal component analysis.
2.4. Canopy Structure and Gap Light Transmission Indices
The crown cover of the tree stand (hereafter referred to as “the Cover”) was evaluated through visual inspection, with the results expressed as a percentage. The canopy structure and gap light transmission indices were extracted from true-colour fisheye photographs using Gap Light Analyzer (GLA) software. The use of hemispherical smartphone photography allowed for a rapid assessment of the forest canopy and light regime. This method serves as a suitable alternative to traditional camera-based hemispherical photography, providing comparable results in a more efficient and cost-effective manner [54]. The following indices were evaluated: canopy openness, expressed as a percentage, indicates the proportion of the open sky visible beneath the forest canopy. This index is calculated exclusively from a hemispherical photograph and does not account for the influence of the surrounding topography. The Leaf Area Index (LAI) represents the effective leaf area integrated over zenith angles ranging from 0° to 60° [55]. Direct Transparency refers to the amount of direct solar radiation transmitted through the canopy, measured in moles per square meter per day (mol/m2/day). Diffuse Transparency denotes the amount of diffuse solar radiation transmitted by the canopy, also measured in moles per square meter per day (mol/m2/day). Total Transparency (Trans) is the sum of both direct and diffuse transparency, expressed in moles per square meter per day (mol/m2/day).
2.5. Plant Functional Traits
The grass height in each plot was measured on five occasions. The following morphological parameters were considered functional properties of plants (Table S1): leaf area (mm2), leaf area per leaf dry mass (mm2/mg), leaf dry mass per leaf fresh mass (mg/g), leaf nitrogen (N) content per leaf dry mass (mg/g), plant height (cm), seed mass (g), and phenological parameters, including the beginning of the flowering period (month), end of flowering (month), time of seed dispersal (month), and seed bank duration (years) [56]. Dispersal distance classes were taken from Lososová et al. [57]. Ellenberg-type ecological indicator values were employed to estimate the environmental requirements of plant species, specifically in terms of moisture (W), soil pH (R), temperature (T), light (L), and salinity (S) [58]. Ecological indicators offer numerous advantages, as they facilitate a comprehensive assessment of environmental conditions, integrate environmental parameters over extended periods, circumvent costly and time-consuming instrumental measurements, and reflect anthropogenic impacts. Similar to abiotic environmental factors, naturalness and degradation can be viewed as a continuum ranging from artificial (i.e., degraded) to ‘natural’ conditions. Species exhibit varying responses to anthropogenic degradation; some plants are typically found in predominantly natural or near-natural environments, while others can tolerate, benefit from, or even require anthropogenic influences. This variability allows for the assessment of a site’s naturalness or degradation based on its species composition. To this end, two types of indicator value systems have been developed: naturalness and hemeroby. Hemeroby measures the extent of ecosystem transformation due to anthropogenic pressure, while naturalness indicates the degree to which an ecosystem approximates the ideal least disturbed state [59]. The projected cover of herbaceous plant species was used as a weight in the calculation of the hemeroby assessment:
Plant life-form (climamorphes) are classified according to Raunkiaer [64]. Pollenochores are groups of plants characterised by their method of pollination; for spore-producing plants, this distinction is based on the type of gamete transfer. Diasporochores, also known as ecobiochores, are groups of plants that exhibit varying mechanisms for the dispersal of their diasporas [65]. Pollenochores and diasporochores are defined according to Tarasov [66]. The Shannon index (Tree Diversity) was calculated to assess the diversity of the tree stands.
2.6. Species and Functional Diversity Indices
The calculation of distances between pairs of species on the basis of functional traits for species collected in communities was conducted via the mFD package [67]. This study considered the following species and functional diversity indices: species richness (Sp_richn), Gini-Simpson (GSimpson), functional richness (FRic), functional evenness (FEve), functional divergence (FDiv), functional originality (FOri), functional specialization (FSpe), functional identity (FIde), functional dispersion (FDis), Rao’s quadratic entropy (FunRao), functional redundancy (FunRedundancy), functional mean pairwise distance (Fmpd), functional mean nearest neighbour distance (Fnnd). Species richness (Sp_richn) is the number of species present in a community [68]. The Gini-Simpson (GSimpson) index is a measure of species diversity [69]. GSimpson, FunRao, and FunRedundancy were calculated using the rao.diversity function from the SYNCSA package [70]. The response of the functional diversity indices to the gradient of environmental factors was elucidated using a quadratic model, which is suitable for describing a ‘hump-backed’ relationship. Additionally, a linear model and a model with one intercept were considered; however, neither of these were statistically significant. The optimal model was identified by utilizing a set of indices and employing the compare_performance function from the performance package [71].
2.7. Tree Community Ordination
A nonmetric multidimensional scaling (NMDS) analysis was conducted to evaluate tree species composition. We chose NMDS because of the presence of nonlinear gradients in species composition. This ordination method was implemented using the vegan package [72]. Various data transformations were tested, including untransformed, log-transformed, square root (sqrt)-transformed, column total-normalized, column maximum-normalized, frequency-transformed, normalised, range-transformed, Hellinger-transformed, chi-square-transformed, Wisconsin-transformed, and arcsine-transformed data. Additionally, we evaluated several distance metrics: Euclidean, Manhattan, Gower, Bray-Curtis, Kulczynski, Morisita, Horn–Morisita, Cao, Jaccard, Mountford, Raup-Crick, Canberra, Chao, Mahalanobis, Binomial, and Clark distances. The goal was to identify a combination of data transformation and distance metric to generate a distance matrix most strongly correlated with the environmental variables. Rank correlations were calculated using the rankindex function of the vegan package. The Gower distance combined with arcsine data transformation produced the best results and was subsequently used for the NMDS procedure. The NMDS spatial variability model was developed in ArcGIS 10.2.
2.8. Structural Equation Modelling
The impacts of urban park trees on the environment and herbaceous cover under anthropogenic pressure are complex. While not all factors and processes involved in these interactions can be measured directly, they are crucial for understanding the structure and dynamics of urban ecosystems. Structural equation modelling enables to test hypotheses regarding the causal relationships between factors and their effects, as well as to evaluate latent factors that cannot be directly measured but are significant. The method of partial least squares was employed for structural equation modelling [73]. The calculations were conducted using the seminr package [74].
3. Results
3.1. The Parameters of the Urban Park Tree Stand
The tree stands in the park plantations comprised 47 species or subspecies of woody plants (Table S1). The distribution of the number of trees per plot followed a Poisson distribution (λ = 2.4, χ2 = 2.7, P = 0.74). The tree stand density in Students’ Park was significantly greater than that in Ivan Starov Square (Mann–Whitney test, P < 0.001) (Table 1).
The Shannon tree diversity index was 0.9 ± 0.6, and this index did not differ between the parks (t = 0.41, P = 0.68). Visually assessed canopy cover was not significantly different between the parks (t = 0.08, P = 0.93). However, the canopy openness and transparency scores were greater for the stand in Ivan Starov Square than for the stand in Students’ Park. The leaf area index (LAI) was greater for the stand in Students’ Park than for the stand in Ivan Starov Square.
3.2. Nonmetric Multidimensional Scaling of Urban Park Tree Communities
The dependence of stress statistic derived after NMDS on the number of dimensions indicated that a four-dimensional solution was sufficient for a qualitative representation of the tree community space in a low-dimensional space (Figure S2). Fitting the stand parameters to the extracted dimensions enabled a meaningful interpretation of these dimensions. Dimension 1 indicated the variability of the stand structure, which was associated with the variable number of trees in the survey plot, the diversity of the stand, and the transparency of the stand canopy (Figure 2). Among the trees, Acer platanoides and Robinia pseudoacacia were indicators of high stand density, diversity, and low canopy transparency. The indicators of the contrasting conditions included Acer pseudoplatanus, Acer negundo, Ailanthus altissima, Aesculus hippocastanum and several other tree species. The parks exhibited similar patterns of spatial variability in dimension 1 (Figure 3).
Negative values for dimension 1 were observed in the central areas of the parks, whereas the peripheral regions displayed positive values for this dimension. The LAI was most sensitive to dimension 2, with Acer platanoides serving as the key indicator among the tree species. The spatial patterns suggested the presence of comparable structures in the tree canopy of the parks, resulting from the formative influence of Acer platanoides, which contributed to the increase in LAI values. The LAI served as an indicator for dimension 3, which exhibited an increase in LAI due to the proliferation of Acer negundo and Gleditsia triacanthos and a decrease in Acer platanoides and Ulmus pumila. In both parks, elevated values of dimension 3 were predominantly found in the peripheral areas. Dimension 4 indicated a trend of increasing stand diversity coupled with greater canopy openness, set against a backdrop of a decreasing LAI. The primary marker for this dimension was Acer platanoides. This dimension also showed a similar spatial pattern in both parks.
3.3. The Properties of the Environment and Herbaceous Cover
The principal component analysis of the variability in environmental and herbaceous cover properties revealed two principal components with eigenvalues greater than one (Table 2). Principal components 1 and 2 can be meaningfully interpreted as the gradients of hemeroby and naturalness, respectively. An increase in hemeroby was correlated with higher light intensity, soil and air temperatures, a decrease in atmospheric humidity, and a reduction in soil moisture and electrical conductivity. Furthermore, an increase in hemeroby was linked to greater functional diversity within plant communities and a decrease in the height of the herbaceous cover. Conversely, a decline in the naturalness of the herbaceous community was associated with increased light intensity, species richness, height, and projective cover of the herbaceous layer.
3.4. Influence of Tree Stands on Environmental Properties and Herbaceous Cover
The dimensions derived from the results of multidimensional scaling of the stand structure were correlated with the environmental properties and herbaceous cover. The positive values of NMDS 1 indicated conditions characterized by elevated soil and air temperatures, as well as increased levels of illumination (Table 2). Furthermore, the positive values of NMDS 1 suggested a greater degree of hemeroby and less natural conditions, which were associated with greater projective cover of herbaceous plants. Conversely, an increase in the naturalness of ecosystems was associated with higher levels of soil and air humidity. The correlation structure for NMDS 2 closely resembled that of NMDS 1, with the notable exception of the lack of a statistically significant correlation between NMDS 2 and the projective cover of herbaceous vegetation and naturalness. NMDS 3 was also responsive to hemeroby and naturalness, with its distinctive feature being its sensitivity to soil electrical conductivity and grass height. NMDS 4 illustrated the variability of projective cover and light in relation to the stand.
3.5. Functional Axes of Herbaceous Plants
The variability of plant traits in the herbaceous layer was characterized by five functional axes (Table 3). Functional axis 1 indicated consistent changes in plant properties related to their hemeroby and naturalness. Hemeroby plants exhibited elevated leaf nitrogen content, delayed onset and termination of the flowering period, increased dispersal potential, and heightened light requirements.
The plants with higher values for axis 1 were typically therophytes, whereas those with lower values were predominantly synzoochores (Figure 4). Additionally, functional axis 2 was associated with hemeroby and the naturalness of plants, highlighting the significant dispersal potential of hemeroby species and their relatively smaller seed sizes. The plants with higher values on functional axis 2 were typically hemicryptophytes, anemophilous plants, and ballistochores, whereas those with lower values were generally seedlings of phanerophyte or nanophanerophyte species. The results for functional axis 3 indicated that species with a greater degree of hemeroby exhibited a lower specific leaf area and a greater dry weight of leaves relative to their wet weight. Positive values on functional axis 1 corresponded to hemicryptophytes and entomophilous plants, whereas lower values were associated with seedlings of phanerophytes and synzoochores. The findings demonstrated that functional axes 4 and 5 were independent of hemeroby or naturalness. Functional axis 4 allowed for the differentiation of species on the basis of their sensitivity to nutrient content. Species exhibiting greater sensitivity to nutrient levels were also found to be sensitive to the soil moisture content, displaying greater height and larger leaf areas in terms of dry weight. Conversely, plants that were less sensitive to the soil nutrient content presented greater dry weights of leaves than wet weights and presented a greater tendency toward thermophily and a more continental habitat.
A higher value on functional axis 4 was indicative of ballistochores. Dimension 5 distinguished plants on the basis of their flowering date. The plants that began flowering later were taller, had a greater affinity for light, and had smaller leaf areas in terms of dry weight. Higher values on functional axis 5 were indicative of geophytes and hemicryptophytes, whereas lower values were associated with therophytes.
3.6. The Functional Diversity Indices of Herbaceous Plant Communities Along the Gradient of Hemeroby and Naturalness
The functional diversity indices exhibited distinct patterns of response along the gradients of hemeroby and naturalness (Figure 5 and Figure 6).
An increase in hemeroby was associated with a linear increase in FunRedundancy, Fide_PC1, Fide_PC2, Fide_PC5, and Fori, whereas Fnnd and Fspe showed a linear decrease. The responses of Fric, FunRao, Fide_PC3, Fide_PC4, Feve, and Fdiv to the hemeroby gradient were characterized by a ‘hump-backed’ pattern. In contrast, the responses of Fdis and GSimpson to the hemeroby gradient were irregular. The functional diversity indices Fdis, Fdiv, Fmpd, and Fnnd displayed a linear response pattern along the naturalness gradient, whereas the other indices exhibited a ‘hump-backed’ response. Notably, the explanatory power of the respective response models along the naturalness gradient is relatively low. The indices of functional diversity underwent cluster analysis, resulting in the identification of three clusters, which can be conditionally designated as follows (Figure S3): Functional Richness (Fric, Fdis, GSimpson, FunRao, FunRedundancy, Fide_PC3, Fide_PC5), Functional Evenness (Feve, Fori, Fide_PC1, Fide_PC2), and Functional Divergence (Fdiv, Fmpd, Fnnd, Fspe, Fide_PC4). These clusters are considered latent variables in the subsequent analysis.
3.7. Structural Equation Modelling of the Influence of Tree Stands on the Environmental Properties and Functional Diversity of Herbaceous Cover
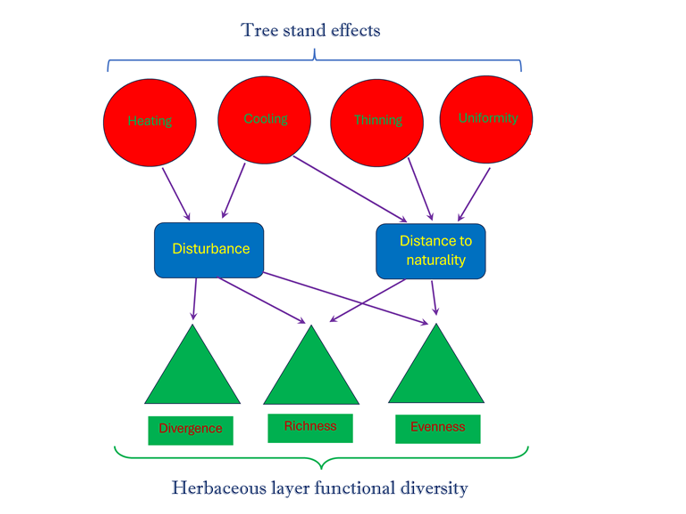
The results obtained in the previous stage of the study allowed for the proposal of the following structural model (Figure 7). The structural features of the stand, which influence the effects on the environment and herbaceous vegetation cover, were represented through four latent variables: Heating Potential, Cooling Potential, Density/Thinning of the stand, and Diversity/Uniformity of the stand’s species composition. NMDS1, ..., NMDS4, stand crown properties, stand density, and diversity indicators were considered as indicators of the latent variables. NMDS3 and Canopy Openness were identified as statistically significant indicators of Heating Potential. NMDS3, Canopy, and LAI were statistically significant indicators of Cooling Potential. NMDS1, Trans, and Tree Number were statistically significant indicators of Thinning Potential. NMDS4 and Tree Diversity were statistically significant indicators of Stand Uniformity.
The changes in the environmental properties were represented by two latent variables. The indicators of hemeroby and naturalness were meaningfully interpreted as Disturbance and Distance to Naturalness. The Disturbance increased with increasing air and soil temperatures, decreasing air and soil humidity, and decreasing soil electrical conductivity. Distance to Naturalness increased with increasing light availability, species richness, and grassland height. An increase in Heating Potential resulted in a rise in Disturbance, whereas an increase in Cooling Potential led to a decrease in Disturbance. Cooling Potential reduces Distance to Naturalness, whereas Thinning and Stand Uniformity contribute to a greater distance of communities from the state of naturalness.
The indices Fric, FunRedundance, and Fide_PC3 were identified as statistically significant indicators of functional richness. The results indicated that Fide_PC1, Fide_PC2, and Feve were statistically significant indicators of functional evenness. Fdiv and Fide_PC4 were recognized as statistically significant indicators of functional divergence. The model demonstrated that an increase in disturbance led to a decrease in both functional richness and functional divergence, while simultaneously resulting in an increase in functional evenness. Furthermore, an increase in the distance to naturality was associated with an increase in functional evenness and a decrease in functional richness. However, the effect of distance to naturality on functional divergence was not statistically significant.
4. Discussion
The formation of tree stands in city parks results from a combination of factors [75,76]. Park plantations can be established based on the remnants of the natural forest vegetation that previously existed at the site [77]. Trees in parks may be planted by humans, utilizing both native flora and non-native tree species for this purpose [78]. The existing tree stand can be influenced by park management and reconstruction efforts [1]. The propagation and spread of both native and non-native trees can occur spontaneously within a park as a result of biological processes [79]. These processes contribute to a complex mosaic in the structure of the park’s tree stand, which can be quantified [80]. A key factor in the variability of park plantations is the alteration in stand density. Tree stand density is an essential marker for understanding the structure of natural forests and urban parks alike [81]. The results indicate that stand density variability is associated with changes in stand diversity and crown openness. Alterations in stand density influence the distribution of environmental factors such as light, temperature, and humidity of soil and air [82]. The modification of environmental conditions caused by trees significantly influences on both the growth dynamics of woody plants [83], and the comfort levels experienced by visitors in urban parks [84]. The density and diversity of tree stands influence the physical and chemical properties of the soil [85], the carbon content and primary productivity of ecosystems [86], and the phytomass of herbaceous cover (Li et al., 2022). Furthermore, the density of a stand plays a crucial role in determining the amount of space available for plant growth within the community [88] and the natural resources accessible to individual trees [89]. Elevated stand density also intensifies competition for space and resources among individual trees, thereby inhibiting overall stand growth [90].
Quantitative alterations in stand density are correlated with variations in the diversity of woody plant communities. The influence of stand density on the functional characteristics of vegetation cover can be elucidated as a result of changes in water utilization and the effects of the stand on the distribution and infiltration of precipitation (Wang et al., 2021). These factors subsequently impact the intensity of soil erosion and the conditions of soil nutrients [92]. The most diverse park plantations within the studied areas are characterized by communities that include both the native species Acer platanoides and the introduced species Robinia pseudoacacia. This phenomenon has been identified as a thinning/increasing stand density trend.
The dynamics in question have a substantial impact on the transparency of the canopy of the stand and the degree to which the stand diverges from its hypothetical natural state. A higher-density Robinia plantation can capture greater amounts of light energy due to its elevated leaf area index (LAI) and canopy density, resulting in an increased photosynthetic capacity for carbon fixation [93]. The thinning of the tree stands in urban parks resulting from management interventions is evidenced by alterations in the condition of the herbaceous cover. Thinning may occur due to the removal of aged or diseased trees, the expansion of pedestrian pathways, or the establishment of children’s play areas or sports facilities within the park. Notably, a decline in the naturalness of the herbaceous cover may coincide with an increase in ecosystem dissimilarities, such as an increase in the proportion of allergenic plants within the plant community, which could adversely affect the health of park visitors.
The increase in the proportion of the native species Acer platanoides has contributed to an increase in the LAI, resulting in a cooling effect within the park environment. This phenomenon can be considered a significant ecosystem function, which, in addition to its environmental implications, fosters conditions conducive to reducing hemeroby and restoring herbaceous cover to a more natural state. These findings underscore the critical role of native flora in the development of parklands and align with existing research indicating that native plants fulfil multiple ecosystem functions in urban green spaces. Furthermore, they support urban biodiversity and provide ecosystem services more effectively than non-native plant species do, highlighting the necessity of prioritizing native species in urban horticultural practices [94]. Park management strategies should prioritize the preservation of natural vegetation pockets, as their ecological significance is substantial in terms of optimizing environmental conditions and enhancing the aesthetic appeal of the park. When restoring or establishing park plantings, native flora should be preferred. However, it is also important to consider that introduced species possess considerable potential for the development of park plantings, particularly in areas subjected to high anthropogenic pressures, characterized by elevated levels of hemeroby and reduced naturalness. Adventive species exhibit a high degree of resilience to adverse conditions, thereby facilitating the establishment of park plantations that can provide substantial ecosystem services in proximity to sources of environmental degradation. Consequently, the incorporation of resilient adventitious species is recommended for planting in areas adjacent to urban thoroughfares.
The openness of tree crown stands is a significant factor influencing the potential for surface heating within park ecosystems. The characteristics of the crown, including overall structure, biomass distribution, and spatial arrangement of crown elements, play crucial roles in the functioning of both forests (Li et al., 2022) and park ecosystems [95]. The crown layer of a forest or park determines the intensity of light interception and influences water and gas exchange within the ground layer [96]. Different tree species present in park plantations exhibit varying aesthetic appearances and crown characteristics. The transparency of the crown impacts the functional attributes of trees and their aesthetic perception, which can be quantified using the LAI [97]. The structure of the crown layer significantly influences the productivity of natural forest stands and urban parks [98,99]. The crown structure of individual trees serves as a reliable indicator of their vitality, reflecting the adaptive strategies employed by trees in response to various factors, including competition for growth, environmental conditions, and climate change [100]. The morphology of tree crowns in urban parks substantially affects both the volume of the crown and the ecosystem services rendered by the trees, such as the provision of shade area and density [101]. Nonmetric multidimensional scaling (NMDS3) analysis revealed that adventitious species, such as Acer negundo and Gleditsia triacanthos, can create favourable conditions that mitigate surface heating potential. An increase in surface heating potential is directly associated with elevated levels of hemeroby. Furthermore, the homogeneity of the stand contributed to a decline in the naturalness of the herbaceous cover. Therefore, establishing more diverse tree stands is essential for fostering conditions that promote a more natural state of herbaceous cover.
Hemeroby and naturalness are frequently perceived as opposing yet complementary concepts [59]. The findings suggest a symmetry breaking between hemeroby and naturalness, indicating a certain degree of independence between these phenomena. Hemeroby is influenced by processes associated with the thermal balance of the Earth’s surface, specifically the potential for heating and cooling. In contrast, naturalness is affected by alterations in the structure of the stand, including its density and diversity, as well as changes in the cooling potential. Urban parks stand to play a crucial role in mitigating the urban heat island effect. Research indicates that an urban park’s area and perimeter are positively correlated with the intensity of the park’s cooling effect [102]. The cooling effect is contingent not only upon the park’s area but also on its structural characteristics. Tree species’ composition significantly influences horizontal and vertical cooling, thereby affecting the dynamics of favourable microclimatic conditions [103]. Multilayered plant communities within urban parks are particularly effective in cooling the urban environment. Tree stand density, area, tree height, and solar radiation transmittance significantly influence temperature reduction and increase relative humidity [104,105]. The LAI is a widely utilized canopy indicator representing the one-sided leaf area relative to the crown projection area of trees and shrubs [106]. This metric facilitates the identification of numerous critical factors and processes within the urban forest ecosystem, including shading, cooling, regeneration, and the hydrological cycle [107,108]. The LAI indicates the potential for shading and cooling effects in environments influenced by urban parks [109]. Additionally, crown structure is a pivotal factor affecting the diversity of other layers within forest and park ecosystems [110].
Hemeroby and naturalness significantly influence functional diversity indices of the herbaceous layer of park plantations, which can be more accurately understood as the relationship whereby alterations in the hemeroby and naturalness of plant communities impact their functional characteristics. The herbaceous layer of park plantations plays a crucial role in ecosystem functions and serves as a reliable indicator of environmental properties and the extent of anthropogenic disturbance [111]. The findings align with the generalisations that functional diversity can be conceptualized in terms of functional richness, functional evenness, and functional divergence. For the corresponding latent variables, the nominal indices (Fric, Feve, and Fdiv) are statistically significant indicators. Notably, functional identity indices (Fide) typically exhibit the highest loadings on the respective latent variables. Functional identity (FIde) refers to the weighted average position of a species within an assemblage along each functional axis or the mean trait values for the assemblage along each axis of the studied functional space [112]. Fides are derived from functional axes that can be interpreted in relation to plant functional properties, thereby facilitating the connection between formal indices of functional diversity and functional properties. Consequently, the functional richness of a community is altered by changes in the aggregation of functional properties characterized by functional axes 3 and 5. An increase in functional richness is observed under conditions where a greater proportion of plants exhibit a larger leaf area relative to leaf dry mass, specifically hemicryptophytes, alongside a decrease in the proportion of therophytes. Therefore, an increase in hemeroby and a decrease in naturalness result in a reduction in functional richness, which is attributed to a decrease in the specific leaf area of cryptophytes and an increase in the specific leaf area of therophytes. This outcome is consistent with previous research indicating a correlation between the proportion of therophytes and the level of hemeroby [113,114]. The potential for enhancing the role of therophytes within the community may arise from a reduction in functional richness as a consequence of anthropogenic pressures.
The functional evenness (FEve) indices describe how the feature space is filled [43]. These indices of functional evenness are sensitive to the community’s potential for productivity, reliability, and vulnerability to invasion [115]. Changes in functional evenness are also reflected in functional axes 1 and 2. This suggests that an increase in functional evenness is associated with a reduction in leaf area and seed weight, a decrease in humidity and naturalness, an extension of flowering duration and seed bank longevity, alterations in light regimes, climate continuity, and an increase in hemeroby levels. Consequently, the increase in functional evenness serves as an indicator of the degree of hemeroby. Functional traits are markers for plant species that have adapted to anthropogenic pressures. These traits are closely aligned with environmental trends that correlate with conditions conducive to the proliferation of hemeroby. These trends include a reduction in soil and air moisture, an increase in soil and air temperatures, and an increase in light intensity. The reduction in leaf area serves as an adaptation to excessive light conditions. Reproductive traits play crucial roles in the survival of plant species facing anthropogenic pressures. It is highly probable that the expansion of hemeroby is accompanied by a decline in the overrepresentation of functional properties within the community, subsequently influencing the enhancement of functional evenness.
Functional divergence (FDiv) refers to the deviation of species abundance from the centre of gravity of the vertices that define the convex hull of the studied assemblage [116]. Functional divergence serves as an indicator of functional axis 4, which reflects the sensitivity of functional divergence to the proportion of plant species within a community that exhibits a high demand for soil nitrogen and moisture. It has been observed that functional divergence increases with a more significant proportion of plant species that require elevated nitrogen levels, are taller, and have larger specific leaf areas. These species are characterized by extreme traits relative to the overall community, and their prevalence tends to rise in response to heightened anthropogenic impacts. This observation aligns with findings that suggest that urban species share similar ecological traits, including a preference for drier, less acidic, and more fertile environments [117]. Hemeroby negatively correlated with the ratio of carbon to nitrogen concentrations in the soil [118], suggesting that species with relatively higher nitrogen requirements have ecological advantages in the context of anthropogenic environmental changes. The urban environment is characterized by elevated nitrogen availability [119]. Consequently, the ability to adapt to excess nitrogen is a critical factor for survival and success amid increasing hemeroby. These findings underscore the importance of selecting and planting nitrogen-tolerant plant species in urban forest management [120]. Notably, the species richness of native grasses and shrubs has been shown to decline significantly in response to heightened nitrogen levels, whereas the species richness of introduced species has remained unaffected; however, nitrogen does promote an increase in their coverage [121]. This contrasting influence of nitrogen content elucidates that the proliferation of nitrogen-loving plant species, in response to rising hemeroby, is accompanied by a functional restructuring of the community, which is evidenced by an increase in functional divergence.
The results obtained enable the formulation of practical guidelines for the management of urban parks. Open tree canopies or insufficient tree density can increase surface heating and create unfavourable microclimatic conditions. This can elevate temperatures and decrease soil moisture and humidity in recreational areas for park visitors. Dense tree canopies with a high leaf area index can provide shade and lower temperatures, contributing to a comfortable environment for visitors and enhancing the natural appearance of the herbaceous cover. This aspect is particularly significant in the context of global climate change. Using native tree species (e.g., Acer platanoides) helps to enhance the cooling effect, reduce hemeroby, and restore the naturalness of the herbaceous cover. Introduced species can be beneficial in areas with high levels of anthropogenic pressure, where resilience to stressful conditions is necessary. Reducing the density of plantations through thinning increases the potential for heating and promotes the growth of hemeroby, which can diminish the natural quality of the ecosystem. Multi-layered plantations with high species diversity provide a better cooling effect and improve microclimatic conditions. The cooling potential can be optimised by increasing planting density and selecting species with extensive leaf coverage. Creating zones with varying vegetation densities to balance heating and cooling potential offers opportunities to establish optimal conditions for visitors to relax in the park. The preservation and restoration of natural areas contribute to enhancing the natural quality and aesthetics of parks. These measures will ensure effective management of urban parks, thereby improving their ecosystem services and visitor comfort.
5. Conclusions
The influence of urban park tree canopies on their surrounding environments and herbaceous cover can be characterised through various gradients, including density/thinning, diversity/homogeneity of the stand, heating potential, and cooling potential. A notable consequence of increased hemeroby within an urban park is a rise in light levels, increases in air and soil temperatures, and reductions in electrical conductivity and soil moisture content. Furthermore, there has been a decrease in air humidity within the park. An increase in hemeroby correlates with an increase in heating potential, whereas a decrease in hemeroby is associated with an intensification of cooling potential within the park. Conversely, the cooling potential mitigates how much the grass layer deviates from its natural state. In contrast, the uniformity and thinning of the tree contribute to an increase in this deviation. Functional diversity indices can be categorised on the basis of their sensitivity to functional richness, evenness, and divergence. Functional identity indices aid in interpreting the variability observed in functional diversity indices concerning the functional properties of plants. Consequently, changes in functional richness depend on variations in the relative proportions of hemicryptophytes and therophytes. The functional evenness of a given ecosystem may fluctuate due to the plants’ preferences for light intensity and associated traits, including leaf area, seed weight, humidity preference, level of naturalness, flowering time, seed bank duration, climate continuity, and hemeroby level. A shift in the proportion of plants requiring nitrogen compounds and soil moisture leads to a change in functional divergence.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Supplement_1.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, O.K. and O.Z.; methodology, I.I.; software, K.H.; validation, O.L., O.K. and H.T.; formal analysis, O.Z.; investigation, H.T.; resources, O.K.; data curation, O.Z.; writing—original draft preparation, O.K.; writing—review and editing, O.Z.; visualization, H.T.; supervision, I.I.; project administration, K.H.; funding acquisition, O.Z.
Funding
This research received no external funding.
Data Availability Statement
Data on plant survey results and protocols of the calculations are available in open access, with the corresponding links provided in the main text of the article.
Acknowledgments
The authors gratefully acknowledge the students of Bogdan Khmelnitsky Melitopol State Pedagogical University and Oles Honchar Dnipro National University for their participation in the fieldwork, which substantially contributed to the completion of this project.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Molozhon, K.O.; Lisovets, O.I.; Kunakh, O.M.; Zhukov, O.V. The Structure of Beta-Diversity Explains Why the Relevance of Phytoindication Increases under the Influence of Park Reconstruction. Regul. Mech. Biosyst. 2023, 14, 634–651. [Google Scholar] [CrossRef]
- Myalkovsky, R.; Plahtiy, D.; Bezvikonnyi, P.; Horodyska, O.; Nebaba, K. Urban Parks as an Important Component of Environmental Infrastructure: Biodiversity Conservation and Recreational Opportunities. Ukr. J. For. Wood Sci. 2023, 14, 57–72. [Google Scholar] [CrossRef]
- Nowak, D.J.; Crane, D.E. Carbon Storage and Sequestration by Urban Trees in the USA. Environ. Pollut. 2002, 116, 381–389. [Google Scholar] [CrossRef]
- Lemanowicz, J.; Haddad, S.A.; Bartkowiak, A.; Lamparski, R.; Wojewódzki, P. The Role of an Urban Park’s Tree Stand in Shaping the Enzymatic Activity, Glomalin Content and Physicochemical Properties of Soil. Sci. Total Environ. 2020, 741, 140446. [Google Scholar] [CrossRef] [PubMed]
- Komlyk, Y.; Ponomarenko, O.; Zhukov, O. A Hemeroby Gradient Reveals the Structure of Bird Communities in Urban Parks. Biosyst. Divers. 2024, 32, 426–436. [Google Scholar] [CrossRef]
- Mexia, T.; Vieira, J.; Príncipe, A.; Anjos, A.; Silva, P.; Lopes, N.; Freitas, C.; Santos-Reis, M.; Correia, O.; Branquinho, C.; et al. Ecosystem Services: Urban Parks under a Magnifying Glass. Environ. Res. 2018, 160, 469–478. [Google Scholar] [CrossRef]
- Bodnaruk, E.W.; Kroll, C.N.; Yang, Y.; Hirabayashi, S.; Nowak, D.J.; Endreny, T.A. Where to Plant Urban Trees? A Spatially Explicit Methodology to Explore Ecosystem Service Tradeoffs. Landsc. Urban Plan. 2017, 157, 457–467. [Google Scholar] [CrossRef]
- Lenk, A.; Richter, R.; Kretz, L.; Wirth, C. Effects of Canopy Gaps on Microclimate, Soil Biological Activity and Their Relationship in a European Mixed Floodplain Forest. Sci. Total Environ. 2024, 941, 173572. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Chen, H.Y.H.; Hautier, Y.; Bao, D.; Xu, M.; Yang, B.; Zhao, Z.; Zhang, L.; Yan, E. Functionally Diverse Tree Stands Reduce Herbaceous Diversity and Productivity via Canopy Packing. Funct. Ecol. 2022, 36, 950–961. [Google Scholar] [CrossRef]
- Kunakh, O.M.; Lisovets, O.I.; Yorkina, N.V.; Zhukova, Y.O. Phytoindication Assessment of the Effect of Reconstruction on the Light Regime of an Urban Park. Biosyst. Divers. 2021, 29, 84–93. [Google Scholar] [CrossRef]
- Öztürk, M.; Ağırtaş, L. Canopy Parameters for Tree and Shrub Species Compositions in Differently Intervened Land Uses of an Urban Park Landscape. Build. Environ. 2021, 206, 108340. [Google Scholar] [CrossRef]
- Nieto-Lugilde, D.; Lenoir, J.; Abdulhak, S.; Aeschimann, D.; Dullinger, S.; Gégout, J.; Guisan, A.; Pauli, H.; Renaud, J.; Theurillat, J.; et al. Tree Cover at Fine and Coarse Spatial Grains Interacts with Shade Tolerance to Shape Plant Species Distributions across the Alps. Ecography (Cop.). 2015, 38, 578–589. [Google Scholar] [CrossRef]
- Ren, Y.; Guo, M.; Yin, F.; Zhang, M.-J.; Wei, J. Tree Cover Improved the Species Diversity of Understory Spontaneous Herbs in a Small City. Forests 2022, 13, 1310. [Google Scholar] [CrossRef]
- Kunakh, O.M.; Volkova, A.M.; Tutova, G.F.; Zhukov, O.V. Diversity of Diversity Indices: Which Diversity Measure Is Better? Biosyst. Divers. 2023, 31, 131–146. [Google Scholar] [CrossRef]
- Rawlik, M.; Kasprowicz, M.; Jagodziński, A.M.; Kaźmierowski, C.; Łukowiak, R.; Grzebisz, W. Canopy Tree Species Determine Herb Layer Biomass and Species Composition on a Reclaimed Mine Spoil Heap. Sci. Total Environ. 2018, 635, 1205–1214. [Google Scholar] [CrossRef]
- González, I.; Sixto, H.; Rodríguez-Soalleiro, R.; Cañellas, I.; Fuertes, A.; Oliveira, N. How Can Leaf-Litter from Different Species Growing in Short Rotation Coppice Contribute to the Soil Nutrient Pool? For. Ecol. Manage. 2022, 520, 120405. [Google Scholar] [CrossRef]
- Zhang, X.; Ni, X.; Heděnec, P.; Yue, K.; Wei, X.; Yang, J.; Wu, F. Litter Facilitates Plant Development but Restricts Seedling Establishment during Vegetation Regeneration. Funct. Ecol. 2022, 36, 3134–3147. [Google Scholar] [CrossRef]
- Nambiar, E.K.S.; Sands, R. Competition for Water and Nutrients in Forests. Can. J. For. Res. 1993, 23, 1955–1968. [Google Scholar] [CrossRef]
- Reczyńska, K.; Orczewska, A.; Yurchenko, V.; Wójcicka-Rosińska, A.; Świerkosz, K. Changes in Species and Functional Diversity of the Herb Layer of Riparian Forest despite Six Decades of Strict Protection. Forests 2022, 13, 747. [Google Scholar] [CrossRef]
- Meili, N.; Manoli, G.; Burlando, P.; Carmeliet, J.; Chow, W.T.L.; Coutts, A.M.; Roth, M.; Velasco, E.; Vivoni, E.R.; Fatichi, S. Tree Effects on Urban Microclimate: Diurnal, Seasonal, and Climatic Temperature Differences Explained by Separating Radiation, Evapotranspiration, and Roughness Effects. Urban For. Urban Green. 2021, 58, 126970. [Google Scholar] [CrossRef]
- Verheyen, K.; Gillerot, L.; Blondeel, H.; De Frenne, P.; De Pauw, K.; Depauw, L.; Lorer, E.; Sanczuk, P.; Schreel, J.; Vanneste, T.; et al. Forest Canopies as Nature-based Solutions to Mitigate Global Change Effects on People and Nature. J. Ecol. 2024, 112, 2451–2461. [Google Scholar] [CrossRef]
- Dylewski, Ł.; Banaszak-Cibicka, W.; Maćkowiak, Ł.; Dyderski, M.K. How Do Urbanization and Alien Species Affect the Plant Taxonomic, Functional, and Phylogenetic Diversity in Different Types of Urban Green Areas? Environ. Sci. Pollut. Res. 2023, 30, 92390–92403. [Google Scholar] [CrossRef]
- Xu, Y.; Chen, X.; Ding, L.; Kong, C.-H. Allelopathy and Allelochemicals in Grasslands and Forests. Forests 2023, 14, 562. [Google Scholar] [CrossRef]
- Fehrenbach, H.; Grahl, B.; Giegrich, J.; Busch, M. Hemeroby as an Impact Category Indicator for the Integration of Land Use into Life Cycle (Impact) Assessment. Int. J. Life Cycle Assess. 2015, 20, 1511–1527. [Google Scholar] [CrossRef]
- Tkachuk, R.; Nykytiuk, Y.; Komorna, О.; Kravchenko, O.; Zymaroieva, A. Ecological Groups of Birds of Zhytomyr Region (Ukraine) in Relation to Thermal Regime and Their Future Prospects in the Context of Global Climate Change. Biosyst. Divers. 2024, 32, 297–305. [Google Scholar] [CrossRef]
- Paudel, S.; States, S.L. Urban Green Spaces and Sustainability: Exploring the Ecosystem Services and Disservices of Grassy Lawns versus Floral Meadows. Urban For. Urban Green. 2023, 84, 127932. [Google Scholar] [CrossRef]
- Ignatieva, M.; Hedblom, M. An Alternative Urban Green Carpet. Science (80-. ). 2018, 362, 148–149. [Google Scholar] [CrossRef]
- Norton, B.A.; Bending, G.D.; Clark, R.; Corstanje, R.; Dunnett, N.; Evans, K.L.; Grafius, D.R.; Gravestock, E.; Grice, S.M.; Harris, J.A.; et al. Urban Meadows as an Alternative to Short Mown Grassland: Effects of Composition and Height on Biodiversity. Ecol. Appl. 2019, 29. [Google Scholar] [CrossRef]
- Lai, H.; Flies, E.J.; Weinstein, P.; Woodward, A. The Impact of Green Space and Biodiversity on Health. Front. Ecol. Environ. 2019, 17, 383–390. [Google Scholar] [CrossRef]
- Mazumdar, S.; Chong, S.; Jalaudin, B.; Wardle, K.; Merom, D. Green Grass in Urban Parks Are a Necessary Ingredient for Sedentary Recreation. J. Park Recreat. Admi. 2024. [Google Scholar] [CrossRef]
- Aboufazeli, S.; Jahani, A.; Farahpour, M. Aesthetic Quality Modeling of the Form of Natural Elements in the Environment of Urban Parks. Evol. Intell. 2024, 17, 327–338. [Google Scholar] [CrossRef]
- Dąbrowski, P. The Effect of Light Availability on Leaf Area Index, Biomass Production and Plant Species Composition of Park Grasslands in Warsaw. Plant Soil Environ. 2013, 59, 543–548. [Google Scholar] [CrossRef]
- Bayón, Á.; Godoy, O.; Maurel, N.; van Kleunen, M.; Vilà, M. Proportion of Non-Native Plants in Urban Parks Correlates with Climate, Socioeconomic Factors and Plant Traits. Urban For. Urban Green. 2021, 63, 127215. [Google Scholar] [CrossRef]
- Li, X.-P.; Fan, S.-X.; Hao, P.-Y.; Dong, L. Temporal Variations of Spontaneous Plants Colonizing in Different Type of Planted Vegetation-a Case of Beijing Olympic Forest Park. Urban For. Urban Green. 2019, 46, 126459. [Google Scholar] [CrossRef]
- Tkachuk, R.; Nykytiuk, Y.; Komorna, О.; Zymaroieva, A. Global Climate Change Promotes the Expansion of Rural and Synanthropic Bird Species: The Case of Zhytomyr Region (Ukraine). Biosyst. Divers. 2024, 32, 183–192. [Google Scholar] [CrossRef]
- Robinson, S.L.; Lundholm, J.T. Ecosystem Services Provided by Urban Spontaneous Vegetation. Urban Ecosyst. 2012, 15, 545–557. [Google Scholar] [CrossRef]
- Ilie, D.; Cosmulescu, S. Spontaneous Plant Diversity in Urban Contexts: A Review of Its Impact and Importance. Diversity 2023, 15, 277. [Google Scholar] [CrossRef]
- Tredici, P. Del Spontaneous Urban Vegetation: Reflections of Change in a Globalized World. Nat. Cult. 2010, 5, 299–315. [Google Scholar] [CrossRef]
- Omar, M.; Al Sayed, N.; Barré, K.; Halwani, J.; Machon, N. Drivers of the Distribution of Spontaneous Plant Communities and Species within Urban Tree Bases. Urban For. Urban Green. 2018, 35, 174–191. [Google Scholar] [CrossRef]
- Czortek, P.; Pielech, R. Surrounding Landscape Influences Functional Diversity of Plant Species in Urban Parks. Urban For. Urban Green. 2020, 47, 126525. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.-L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the Concept of Trait Be Functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Asanok, L.; Marod, D.; Duengkae, P.; Pranmongkol, U.; Kurokawa, H.; Aiba, M.; Katabuchi, M.; Nakashizuka, T. Relationships between Functional Traits and the Ability of Forest Tree Species to Reestablish in Secondary Forest and Enrichment Plantations in the Uplands of Northern Thailand. For. Ecol. Manage. 2013, 296, 9–23. [Google Scholar] [CrossRef]
- Schleuter, D.; Daufresne, M.; Massol, F.; Argillier, C. A User’s Guide to Functional Diversity Indices. Ecol. Monogr. 2010, 80, 469–484. [Google Scholar] [CrossRef]
- Galetti, M.; Guevara, R.; Côrtes, M.C.; Fadini, R.; Von Matter, S.; Leite, A.B.; Labecca, F.; Ribeiro, T.; Carvalho, C.S.; Collevatti, R.G.; et al. Functional Extinction of Dirds Drives Rapid Evolutionary Changes in Seed Size. Science (80-. ). 2013, 340, 1086–1090. [Google Scholar] [CrossRef]
- Wright, T.E.; Kasel, S.; Tausz, M.; Bennett, L.T. Leaf Traits of Eucalyptus Arenacea (Myrtaceae) as Indicators of Edge Effects in Temperate Woodlands of South-Eastern Australia. Aust. J. Bot. 2013, 61, 365. [Google Scholar] [CrossRef]
- Ding, Y.; Zang, R.; Letcher, S.G.; Liu, S.; He, F. Disturbance Regime Changes the Trait Distribution, Phylogenetic Structure and Community Assembly of Tropical Rain Forests. Oikos 2012, 121, 1263–1270. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M. Traits Associated with Invasiveness in Alien Plants: Where Do We Stand. In Biological Invasions. Ecological Studies, vol 193; Nentwig, W., Ed.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2008; pp. 97–125. [Google Scholar]
- Milanović, M.; Knapp, S.; Pyšek, P.; Kühn, I. Linking Traits of Invasive Plants with Ecosystem Services and Disservices. Ecosyst. Serv. 2020, 42, 101072. [Google Scholar] [CrossRef]
- Pakeman, R.J. Functional Diversity Indices Reveal the Impacts of Land Use Intensification on Plant Community Assembly. J. Ecol. 2011, 99, 1143–1151. [Google Scholar] [CrossRef]
- Zhukov, O.; Kunakh, O.; Yorkina, N.; Tutova, A. Response of Soil Macrofauna to Urban Park Reconstruction. Soil Ecol. Lett. 2023, 5, 220156. [Google Scholar] [CrossRef]
- Lisovets, O.; Khrystov, O.; Kunakh, O.; Zhukov, O. Application of Hemeroby and Naturalness Indicators for Monitoring the Aquatic Macrophyte Communities in Protected Areas. Biosyst. Divers. 2024, 32, 270–277. [Google Scholar] [CrossRef]
- Pennisi, B.V.; van Iersel, M. 3 Ways to Measure Medium EC. GMPro 2002, 22, 46–48. [Google Scholar]
- Yorkina, N.V.; Teluk, P.; Umerova, A.; Budakova, V.S.; Zhaley, O.A.; Ivanchenko, K.O.; Zhukov, O. V Assessment of the Recreational Transformation of the Grass Cover of Public Green Spaces. Agrology 2021, 4, 10–20. [Google Scholar] [CrossRef]
- Bianchi, S.; Cahalan, C.; Hale, S.; Gibbons, J.M. Rapid Assessment of Forest Canopy and Light Regime Using Smartphone Hemispherical Photography. Ecol. Evol. 2017, 7, 10556–10566. [Google Scholar] [CrossRef]
- Stenberg, P.; Linder, S.; Smolander, H.; Flower-Ellis, J. Performance of the LAI-2000 Plant Canopy Analyzer in Estimating Leaf Area Index of Some Scots Pine Stands. Tree Physiol. 1994, 14, 981–995. [Google Scholar] [CrossRef]
- Kattge, J.; Diaz, S.; Lavorel, S.; Prentice, I.; Leadley, P.; Bönisch, G.; Garnier, E.; Westoby, M.; Reich, P.; Wright, I.; et al. TRY - a Global Database of Plant Traits. Glob. Chang. Biol. 2011, 17, 2905–2935. [Google Scholar] [CrossRef]
- Lososová, Z.; Axmanová, I.; Chytrý, M.; Midolo, G.; Abdulhak, S.; Karger, D.N.; Renaud, J.; Van Es, J.; Vittoz, P.; Thuiller, W. Seed Dispersal Distance Classes and Dispersal Modes for the European Flora. Glob. Ecol. Biogeogr. 2023, 32, 1485–1494. [Google Scholar] [CrossRef]
- Dengler, J.; Jansen, F.; Chusova, O.; Hüllbusch, E.; Nobis, M.P.; Van Meerbeek, K.; Axmanová, I.; Bruun, H.H.; Chytrý, M.; Guarino, R.; et al. Ecological Indicator Values for Europe (EIVE) 1.0. Veg. Classif. Surv. 2023, 4, 7–29. [Google Scholar] [CrossRef]
- Erdős, L.; Bede-Fazekas, Á.; Bátori, Z.; Berg, C.; Kröel-Dulay, G.; Magnes, M.; Sengl, P.; Tölgyesi, C.; Török, P.; Zinnen, J. Species-Based Indicators to Assess Habitat Degradation: Comparing the Conceptual, Methodological, and Ecological Relationships between Hemeroby and Naturalness Values. Ecol. Indic. 2022, 136, 108707. [Google Scholar] [CrossRef]
- Frank, D.; Klotz, S. Biologisch-Ökologische Daten Zur Flora Der DDR; M.artin-Luther- Universität: Halle-Wittenberg, Halle (Saale), 1990. [Google Scholar]
- Goncharenko, I.V. Fitoindykaciya Antropogennogo Navantazhennya [Phytoindication of Anthropogenic Factor]; Serednyak T.K.: Dnipro (in Ukranian), 2017. [Google Scholar]
- Borhidi, A. Social Behaviour Types, the Naturalness and Relative Ecological Indicator Values of the Higher Plants in the Hungarian Flora. Acta Bot. Hung. 1995, 39, 97–181. [Google Scholar]
- Grime, J.P. Vegetation Classification by Reference to Strategies. Nature 1974, 250, 26–31. [Google Scholar] [CrossRef]
- Raunkiaer, C. Plant Life Forms; Clarendon Press: Oxford, 1937. [Google Scholar]
- Zhukov, O.; Lisovets, O.; Molozhon, K. Differential Ecomorphic Analysis of Urban Park Vegetation. IOP Conf. Ser. Earth Environ. Sci. 2023, 1254, 1–21. [Google Scholar] [CrossRef]
- Tarasov, V.V. Flora of Dnipropetrovsk and Zaporizhia Regions; Travleev, A.P., Ed.; Dnipropetrovsk: Lira (in Ukranian), 2012. [Google Scholar]
- Magneville, C.; Loiseau, N.; Albouy, C.; Casajus, N.; Claverie, T.; Escalas, A.; Leprieur, F.; Maire, E.; Mouillot, D.; Villéger, S. MFD: An R Package to Compute and Illustrate the Multiple Facets of Functional Diversity. Ecography (Cop.). 2022, 2022. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity. Curr. Biol. 2021, 31, R1174–R1177. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688–688. [Google Scholar] [CrossRef]
- Debastiani, V.J.; Pillar, V.D. SYNCSA—R Tool for Analysis of Metacommunities Based on Functional Traits and Phylogeny of the Community Components. Bioinformatics 2012, 28, 2067–2068. [Google Scholar] [CrossRef]
- Lüdecke, D.; Ben-Shachar, M.; Patil, I.; Waggoner, P.; Makowski, D. Performance: An R Package for Assessment, Comparison and Testing of Statistical Models. J. Open Source Softw. 2021, 6, 3139. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Community Ecology Package. R Package Version 2.5-2. 2018.
- Hair, J.F.; Hult, G.T.M.; Ringle, C.M.; Sarstedt, M.; Danks, N.P.; Ray, S. Partial Least Squares Structural Equation Modeling (PLS-SEM) Using R; C.lassroom Companion: Business; Springer International Publishing: Cham, 2021; ISBN 978-3-030-80518-0. [Google Scholar]
- Ray, S.; Danks, N.; Calero Valdez, A. SEMinR: Domain-Specific Language for Building, Estimating, and Visualizing Structural Equation Models in R. SSRN Electron. J. 2021. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, H. Tree Composition and Diversity in Relation to Urban Park History in Hong Kong, China. Urban For. Urban Green. 2022, 67, 127430. [Google Scholar] [CrossRef]
- Molozhon, K.O.; Lisovets, O.I.; Kunakh, O.M.; Zhukov, O.V. Increased Soil Penetration Resistance Drives Degrees of Hemeroby in Vegetation of Urban Parks. Biosyst. Divers. 2023, 31. [Google Scholar] [CrossRef]
- Shanahan, D.F.; Lin, B.B.; Gaston, K.J.; Bush, R.; Fuller, R.A. What Is the Role of Trees and Remnant Vegetation in Attracting People to Urban Parks? Landsc. Ecol. 2015, 30, 153–165. [Google Scholar] [CrossRef]
- Nielsen, A.B.; van den Bosch, M.; Maruthaveeran, S.; van den Bosch, C.K. Species Richness in Urban Parks and Its Drivers: A Review of Empirical Evidence. Urban Ecosyst. 2014, 17, 305–327. [Google Scholar] [CrossRef]
- Padayachee, A.L.; Irlich, U.M.; Faulkner, K.T.; Gaertner, M.; Procheş, Ş.; Wilson, J.R.U.; Rouget, M. How Do Invasive Species Travel to and through Urban Environments? Biol. Invasions 2017, 19, 3557–3570. [Google Scholar] [CrossRef]
- Planchuelo, G.; Kowarik, I.; von der Lippe, M. Endangered Plants in Novel Urban Ecosystems Are Filtered by Strategy Type and Dispersal Syndrome, Not by Spatial Dependence on Natural Remnants. Front. Ecol. Evol. 2020, 8. [Google Scholar] [CrossRef]
- Burkhart, H.E. Comparison of Maximum Size–Density Relationships Based on Alternate Stand Attributes for Predicting Tree Numbers and Stand Growth. For. Ecol. Manage. 2013, 289, 404–408. [Google Scholar] [CrossRef]
- Moreno, G.; Cubera, E. Impact of Stand Density on Water Status and Leaf Gas Exchange in Quercus Ilex. For. Ecol. Manage. 2008, 254, 74–84. [Google Scholar] [CrossRef]
- Martín-Benito, D.; Del Río, M.; Heinrich, I.; Helle, G.; Cañellas, I. Response of Climate-Growth Relationships and Water Use Efficiency to Thinning in a Pinus Nigra Afforestation. For. Ecol. Manage. 2010, 259, 967–975. [Google Scholar] [CrossRef]
- Talal, M.L.; Santelmann, M.V.; Tilt, J.H. Urban Park Visitor Preferences for Vegetation – An On-site Qualitative Research Study. PLANTS, PEOPLE, PLANET 2021, 3, 375–388. [Google Scholar] [CrossRef]
- Forrester, D.I.; Collopy, J.J.; Beadle, C.L.; Baker, T.G. Effect of Thinning, Pruning and Nitrogen Fertiliser Application on Light Interception and Light-Use Efficiency in a Young Eucalyptus Nitens Plantation. For. Ecol. Manage. 2013, 288, 21–30. [Google Scholar] [CrossRef]
- Cai, H.; Di, X.; Chang, S.X.; Jin, G. Stand Density and Species Richness Affect Carbon Storage and Net Primary Productivity in Early and Late Successional Temperate Forests Differently. Ecol. Res. 2016, 31, 525–533. [Google Scholar] [CrossRef]
- Li, Q.; Liu, Z.; Jin, G. Impacts of Stand Density on Tree Crown Structure and Biomass: A Global Meta-Analysis. Agric. For. Meteorol. 2022, 326, 109181. [Google Scholar] [CrossRef]
- Farooq, T.H.; Shakoor, A.; Rashid, M.H.U.; Zhang, S.; Wu, P.; Yan, W. Annual Growth Progression, Nutrient Transformation, and Carbon Storage in Tissues of Cunninghamia Lanceolata Monoculture in Relation to Soil Quality Indicators Influenced by Intraspecific Competition Intensity. J. Soil Sci. Plant Nutr. 2021, 21, 3146–3158. [Google Scholar] [CrossRef]
- Lochhead, K.D.; Comeau, P.G. Relationships between Forest Structure, Understorey Light and Regeneration in Complex Douglas-Fir Dominated Stands in South-Eastern British Columbia. For. Ecol. Manage. 2012, 284, 12–22. [Google Scholar] [CrossRef]
- Mäkinen, H.; Hein, S. Effect of Wide Spacing on Increment and Branch Properties of Young Norway Spruce. Eur. J. For. Res. 2006, 125, 239–248. [Google Scholar] [CrossRef]
- Wang, T.; Xu, Q.; Gao, D.; Zhang, B.; Zuo, H.; Jiang, J. Effects of Thinning and Understory Removal on the Soil Water-Holding Capacity in Pinus Massoniana Plantations. Sci. Rep. 2021, 11, 13029. [Google Scholar] [CrossRef]
- Razafindrabe, B.H.N.; He, B.; Inoue, S.; Ezaki, T.; Shaw, R. The Role of Forest Stand Density in Controlling Soil Erosion: Implications to Sediment-Related Disasters in Japan. Environ. Monit. Assess. 2010, 160, 337–354. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhou, J.; Zhou, H.; Zhao, Z. Photosynthetic Carbon Fixation Capacity of Black Locust in Rapid Response to Plantation Thinning on the Semiarid Loess Plateau in China. Pakistan J. Bot. 2019, 51. [Google Scholar] [CrossRef] [PubMed]
- Tartaglia, E.S.; Aronson, M.F.J. Plant Native: Comparing Biodiversity Benefits, Ecosystem Services Provisioning, and Plant Performance of Native and Non-Native Plants in Urban Horticulture. Urban Ecosyst. 2024, 27, 2587–2611. [Google Scholar] [CrossRef]
- Hofman, J.; Bartholomeus, H.; Janssen, S.; Calders, K.; Wuyts, K.; Van Wittenberghe, S.; Samson, R. Influence of Tree Crown Characteristics on the Local PM 10 Distribution inside an Urban Street Canyon in Antwerp (Belgium): A Model and Experimental Approach. Urban For. Urban Green. 2016, 20, 265–276. [Google Scholar] [CrossRef]
- Binkley, D.; Stape, J.L.; Ryan, M.G. Thinking about Efficiency of Resource Use in Forests. For. Ecol. Manage. 2004, 193, 5–16. [Google Scholar] [CrossRef]
- Vogt, J.; Gillner, S.; Hofmann, M.; Tharang, A.; Dettmann, S.; Gerstenberg, T.; Schmidt, C.; Gebauer, H.; Van de Riet, K.; Berger, U.; et al. Citree: A Database Supporting Tree Selection for Urban Areas in Temperate Climate. Landsc. Urban Plan. 2017, 157, 14–25. [Google Scholar] [CrossRef]
- Kim, J.; Kang, Y.; Kim, D.; Son, S.; Kim, E.J. Carbon Storage and Sequestration Analysis by Urban Park Grid Using I-Tree Eco and Drone-Based Modeling. Forests 2024, 15, 683. [Google Scholar] [CrossRef]
- Ponomarenko, O.; Komlyk, Y.; Tutova, H.; Zhukov, O. Landscape Diversity Mapping Allows Assessment of the Hemeroby of Bird Species in a Modern Industrial Metropolis. Biosyst. Divers. 2024, 32, 470–483. [Google Scholar] [CrossRef]
- Vanderwel, M.C.; Lopez, E.L.; Sprott, A.H.; Khayyatkhoshnevis, P.; Shovon, T.A. Using Aerial Canopy Data from UAVs to Measure the Effects of Neighbourhood Competition on Individual Tree Growth. For. Ecol. Manage. 2020, 461, 117949. [Google Scholar] [CrossRef]
- Franceschi, E.; Moser-Reischl, A.; Rahman, M.; Pauleit, S.; Pretzsch, H.; Rötzer, T. Crown Shapes of Urban Trees-Their Dependences on Tree Species, Tree Age and Local Environment, and Effects on Ecosystem Services. Forests 2022, 13, 748. [Google Scholar] [CrossRef]
- Yao, X.; Yu, K.; Zeng, X.; Lin, Y.; Ye, B.; Shen, X.; Liu, J. How Can Urban Parks Be Planned to Mitigate Urban Heat Island Effect in “Furnace Cities” ? An Accumulation Perspective. J. Clean. Prod. 2022, 330, 129852. [Google Scholar] [CrossRef]
- Chen, J.; Jin, S.; Du, P. Roles of Horizontal and Vertical Tree Canopy Structure in Mitigating Daytime and Nighttime Urban Heat Island Effects. Int. J. Appl. Earth Obs. Geoinf. 2020, 89, 102060. [Google Scholar] [CrossRef]
- Zhang, Z.; Lv, Y.; Pan, H. Cooling and Humidifying Effect of Plant Communities in Subtropical Urban Parks. Urban For. Urban Green. 2013, 12, 323–329. [Google Scholar] [CrossRef]
- Nykytiuk, Y.; Kravchenko, O.; Komorna, O.; Bambura, V.; Seredniak, D. Spatial and Temporal Variation of the Rainfall Erosivity Factor in Polissya and Forest-Steppe of Ukraine. Biosyst. Divers. 2024, 32. [Google Scholar] [CrossRef]
- Parker, G.G. Tamm Review: Leaf Area Index (LAI) Is Both a Determinant and a Consequence of Important Processes in Vegetation Canopies. For. Ecol. Manage. 2020, 477, 118496. [Google Scholar] [CrossRef]
- Zhang, H.; Yao, R.; Luo, Q.; Yang, Y. Estimating the Leaf Area Index of Urban Individual Trees Based on Actual Path Length. Build. Environ. 2023, 245, 110811. [Google Scholar] [CrossRef]
- Tutova, H.; Lisovets, О.; Kunakh, O.; Zhukov, O. The Future of the Kakhovka Reservoir after Ecocide: Afforestation and Ecosystem Service Recovery through Emergent Willow-Popular Communities. Stud. Biol. 2025, 19, 171–194. [Google Scholar] [CrossRef]
- Moser, A.; Rötzer, T.; Pauleit, S.; Pretzsch, H. Structure and Ecosystem Services of Small-Leaved Lime (Tilia Cordata Mill.) and Black Locust (Robinia Pseudoacacia L.) in Urban Environments. Urban For. Urban Green. 2015, 14, 1110–1121. [Google Scholar] [CrossRef]
- Onoszko, K.; Ruiz Gómez, F.J.; Lazzaro, L.; Lora González, Á.; González-Moreno, P. Diversity Patterns of Herbaceous Community in Environmental Gradients of Dehesa Ecosystems. Glob. Ecol. Conserv. 2024, 54, e03162. [Google Scholar] [CrossRef]
- Kunakh, O.M.; Ivanko, I.A.; Holoborodko, K.K.; Lisovets, O.I.; Volkova, A.M.; Zhukov, O.V. Urban Park Layers: Spatial Variation in Plant Community Structure. Biosyst. Divers. 2022, 30, 274–288. [Google Scholar] [CrossRef]
- Garnier, E.; Cortez, J.; Billès, G.; Navas, M.-L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; Aubry, D.; Bellmann, A.; et al. Plant Functional Markers Capture Ecosystem Properties during Secondary Succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- Kim, Y.-M.; Zerbe, S.; Kowarik, I. Human Impact on Flora and Habitats in Korean Rural Settlements | Vliv Lidské Činnosti Na Stanoviště Korejských Vesnic a Jejich Flóru. Preslia 2002, 74, 409–419. [Google Scholar]
- Kunakh, O.; Lisovets, O.; Podpriatova, N.; Zhukov, O. Plant Community Hemeroby Is a Reliable Indicator of the Dynamics of Reclamation of Lands Disturbed by Mining. Ekológia (Bratislava) 2024, 43, 43–53. [Google Scholar] [CrossRef]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional Richness, Functional Evenness and Functional Divergence: The Primary Components of Functional Diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New Multidimensional Functional Diversity Indices for a Multifaceted Framework in Functional Ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef]
- Hill, M.O.; Roy, D.B.; Thompson, K. Hemeroby, Urbanity and Ruderality: Bioindicators of Disturbance and Human Impact. J. Appl. Ecol. 2002, 39, 708–720. [Google Scholar] [CrossRef]
- Testi, A.; De Nicola, C.; Dowgiallo, G.; Fanelli, G. Correspondences between Plants and Soil/Environmental Factors in Beech Forests of Central Apennines: From Homogeneity to Complexity. Rend. LINCEI 2010, 21, 27–43. [Google Scholar] [CrossRef]
- Hu, C.-C.; Liu, X.-Y. Plant Nitrogen-Use Strategies and Their Responses to the Urban Elevation of Atmospheric Nitrogen Deposition in Southwestern China. Environ. Pollut. 2022, 311, 119969. [Google Scholar] [CrossRef]
- Xia, N.; Guo, Y.; Tian, Y.; Li, B.; Liu, X.; Vries, W. Ecological Effects of Nitrogen Deposition on Urban Forests: An Overview. Front. Agric. Sci. Eng. 2022, 9. [Google Scholar] [CrossRef]
- Valliere, J.M.; Bucciarelli, G.M.; Bytnerowicz, A.; Fenn, M.E.; Irvine, I.C.; Johnson, R.F.; Allen, E.B. Declines in Native Forb Richness of an Imperiled Plant Community across an Anthropogenic Nitrogen Deposition Gradient. Ecosphere 2020, 11. [Google Scholar] [CrossRef]
Figure 1.
Locations of environmental characteristics measurements and plant recording sites in Ivan Starov Square and Students Park (Dnipro, Ukraine).
Figure 1.
Locations of environmental characteristics measurements and plant recording sites in Ivan Starov Square and Students Park (Dnipro, Ukraine).

Figure 2.
The locations of the tree species in the space of dimensions derived from nonmetric multidimensional scaling, which were adjusted to reflect the changes characterizing the conditions of the park stand. Tree species exhibiting a distance greater than 0.15 from the origin are depicted. LAI represents the Leaf Area Index, Trans represents the transparency of the tree canopy, Tree_number represents the number of trees per recording plot, Tree_diversity represents the Shannon diversity index of the tree community per the recording plot, Canopy represents the closeness of the park canopy, determined via the visual method, and Canopy_Openness represents the openness of the park canopy, determined via the fisheye method.
Figure 2.
The locations of the tree species in the space of dimensions derived from nonmetric multidimensional scaling, which were adjusted to reflect the changes characterizing the conditions of the park stand. Tree species exhibiting a distance greater than 0.15 from the origin are depicted. LAI represents the Leaf Area Index, Trans represents the transparency of the tree canopy, Tree_number represents the number of trees per recording plot, Tree_diversity represents the Shannon diversity index of the tree community per the recording plot, Canopy represents the closeness of the park canopy, determined via the visual method, and Canopy_Openness represents the openness of the park canopy, determined via the fisheye method.
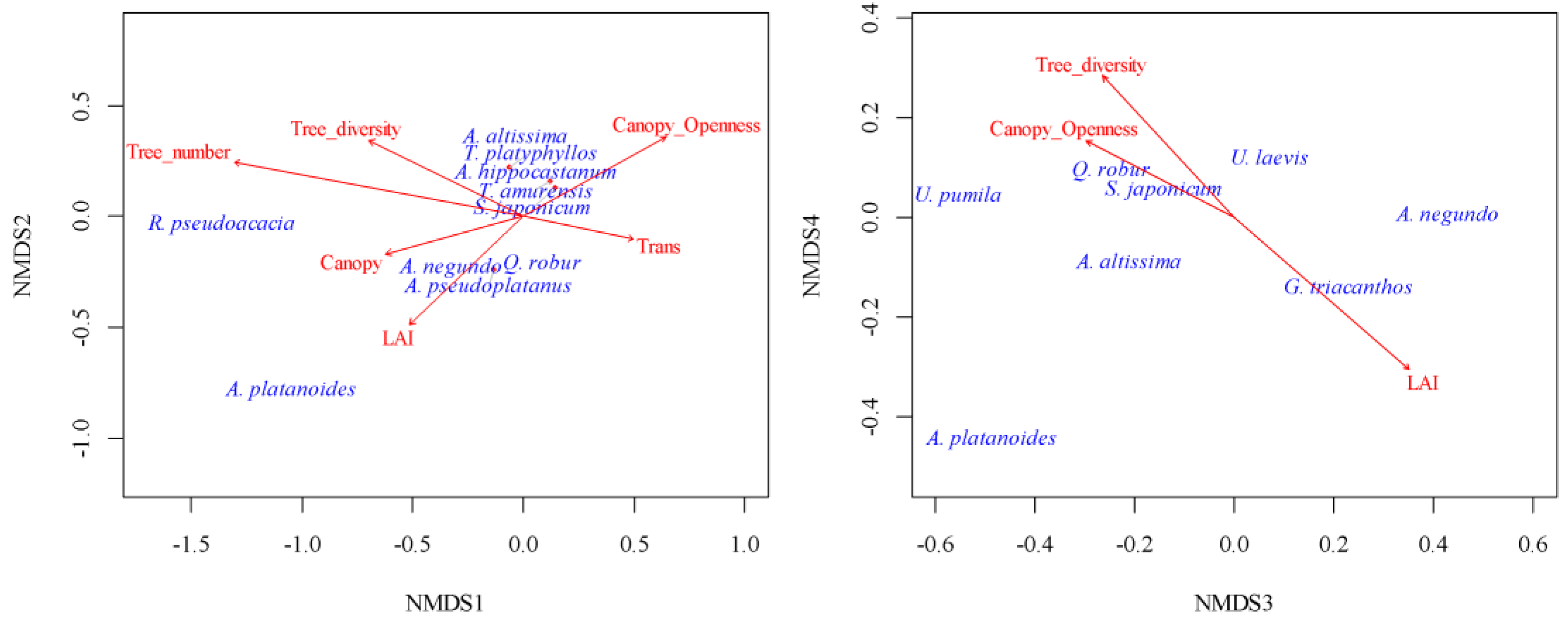
Figure 3.
Spatial variation in NMDS within the examined parks.
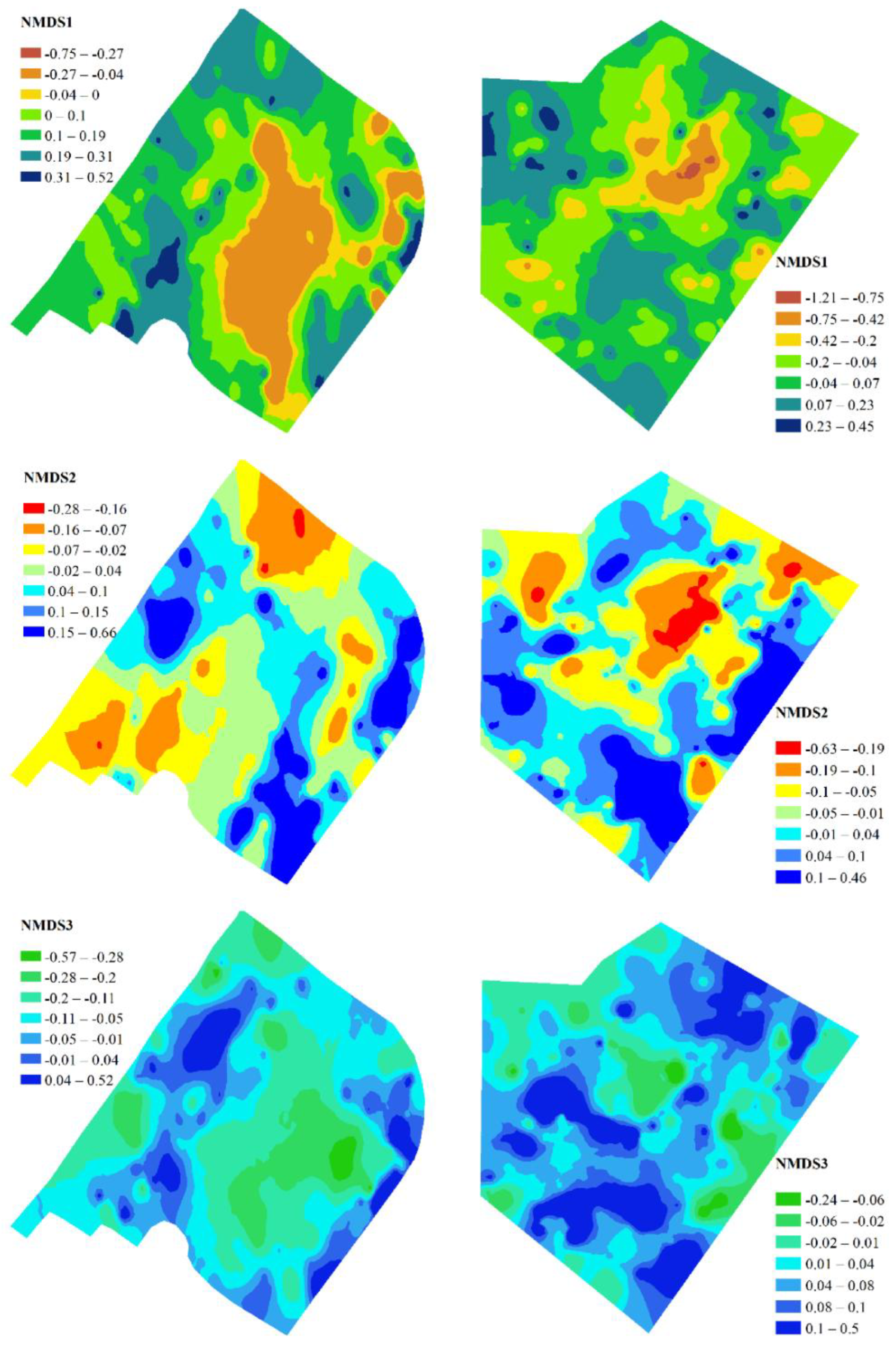
Figure 4.
Boxplot of the variability of functional axes PC1, ..., PC5 depending on climamorphs, pollenochores and diasporochores. Climamorph: G is a geophyte, HKr is a hemicryptophyte, nPh is a nanophanerophyte, Ph is a phanerophyte, and T is a therophyte. Pollenochor: Anph is an anemophilous plant, and Ent is an entomophilous plant. Diasporochor: Anch is an anemochor, Bal is a ballistochor, Bar is a barochor, Endz is an endozoochor, EpZ is an epizoochor, and Synz is a synzoochor.
Figure 4.
Boxplot of the variability of functional axes PC1, ..., PC5 depending on climamorphs, pollenochores and diasporochores. Climamorph: G is a geophyte, HKr is a hemicryptophyte, nPh is a nanophanerophyte, Ph is a phanerophyte, and T is a therophyte. Pollenochor: Anph is an anemophilous plant, and Ent is an entomophilous plant. Diasporochor: Anch is an anemochor, Bal is a ballistochor, Bar is a barochor, Endz is an endozoochor, EpZ is an epizoochor, and Synz is a synzoochor.
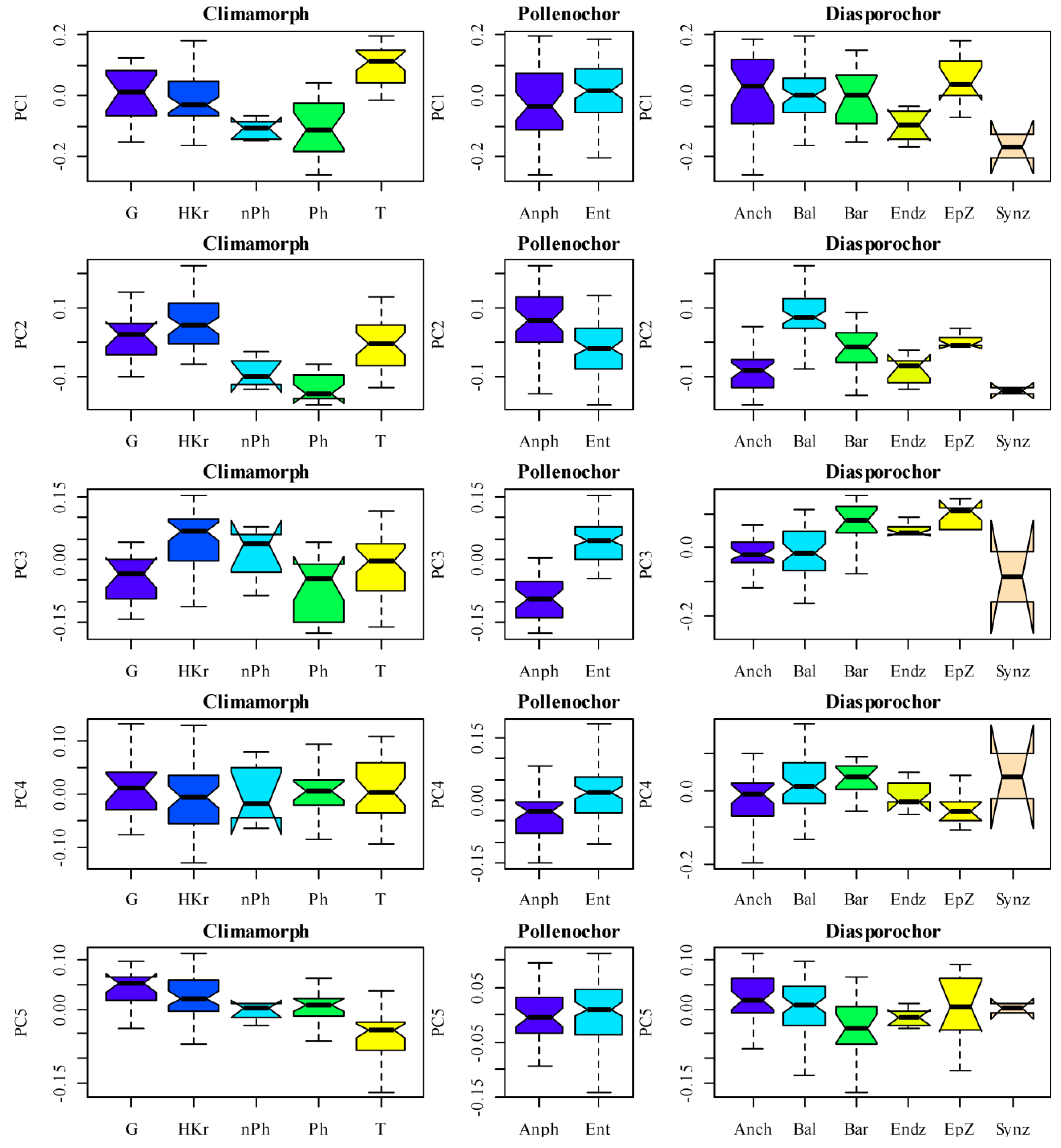
Figure 5.
The response of functional diversity indices along the gradient of the hemeroby factor (PC1) is illustrated in the scatter plot, where blue dots represent the functional diversity indices across the PC1 gradient. The green line denotes the polynomial approximation of the relationship when it is identified as the optimal model, whereas the red line similarly indicates the polynomial approximation for the best model. In cases where the optimal model is a constant, no line is presented. The adjusted R-squared (Radj2) reflects the proportion of variation in the functional diversity index that is accounted for by the respective model. The P-value is derived from the results of the Likelihood-Ratio Test (LRT), which compares the best model to a model that includes only a constant. The abscissa represents the PC1 scores, and the ordinate indicates the values of the functional diversity indices for the grass layer in urban parks: Fmpd is the functional mean pairwise distance, Fnnd is functional mean nearest neighbour distance, FEve is the functional evenness, FRic is the functional richness, FDiv is the functional divergence, FOri is the functional originality, FSpe is the functional specialization, Fide_PC1, … PC5 are the Functional identity indeces, GSimpson is the Gini-Simpson, FunRao is the Rao quadratic entropy, FunRedundancy is the functional redundancy.
Figure 5.
The response of functional diversity indices along the gradient of the hemeroby factor (PC1) is illustrated in the scatter plot, where blue dots represent the functional diversity indices across the PC1 gradient. The green line denotes the polynomial approximation of the relationship when it is identified as the optimal model, whereas the red line similarly indicates the polynomial approximation for the best model. In cases where the optimal model is a constant, no line is presented. The adjusted R-squared (Radj2) reflects the proportion of variation in the functional diversity index that is accounted for by the respective model. The P-value is derived from the results of the Likelihood-Ratio Test (LRT), which compares the best model to a model that includes only a constant. The abscissa represents the PC1 scores, and the ordinate indicates the values of the functional diversity indices for the grass layer in urban parks: Fmpd is the functional mean pairwise distance, Fnnd is functional mean nearest neighbour distance, FEve is the functional evenness, FRic is the functional richness, FDiv is the functional divergence, FOri is the functional originality, FSpe is the functional specialization, Fide_PC1, … PC5 are the Functional identity indeces, GSimpson is the Gini-Simpson, FunRao is the Rao quadratic entropy, FunRedundancy is the functional redundancy.
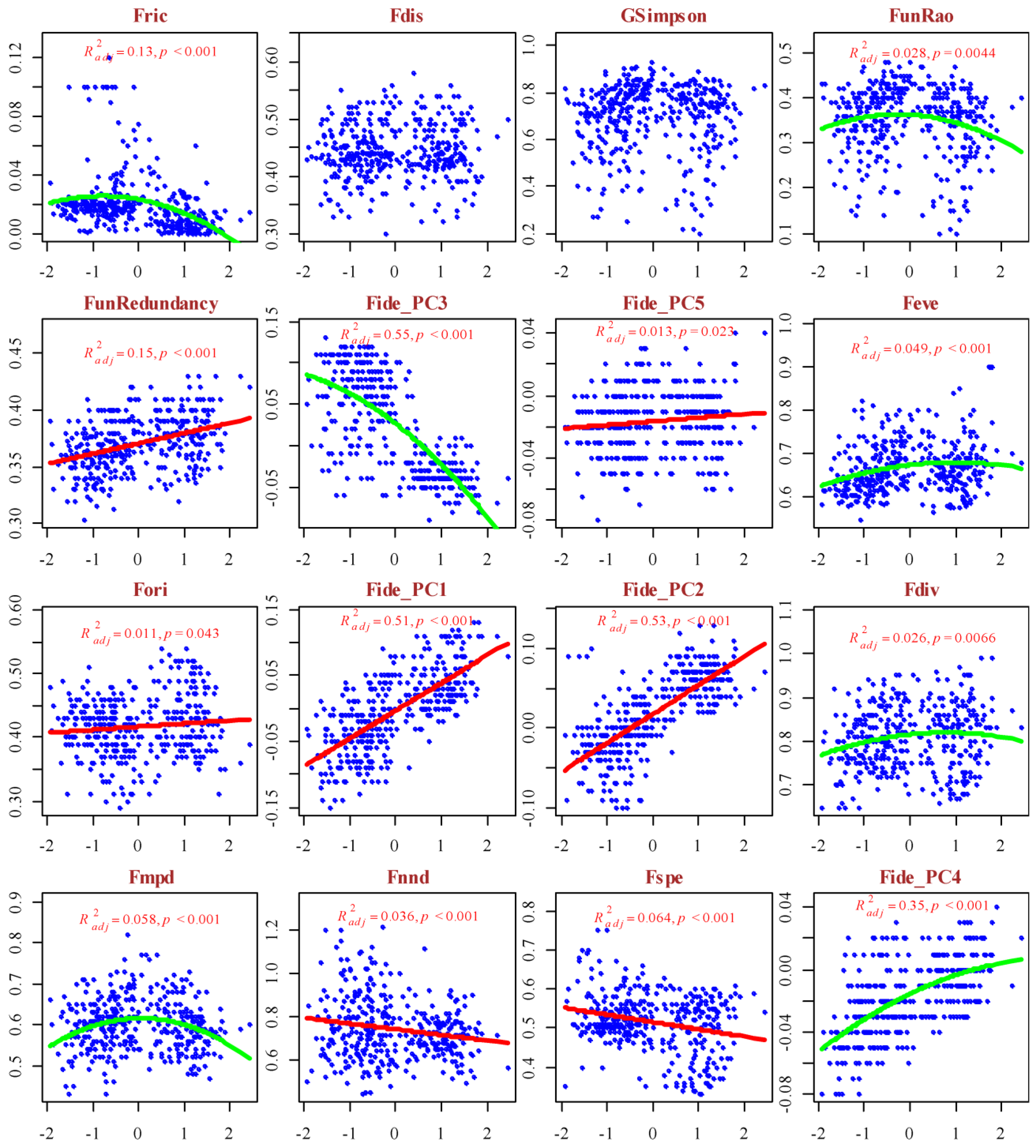
Figure 6.
The response of functional diversity indices along the gradient of the naturalness factor (PC2) is illustrated in the scatter plot, where blue dots represent the functional diversity indices across the PC1 gradient. The green line denotes the polynomial approximation of the relationship when it is identified as the optimal model, whereas the red line similarly indicates the polynomial approximation for the best model. In cases where the optimal model is a constant, no line is presented. The adjusted R-squared (Radj2) reflects the proportion of variation in the functional diversity index that is accounted for by the respective model. The P-value is derived from the results of the Likelihood-Ratio Test (LRT), which compares the best model to a model that includes only a constant. The abscissa represents the PC1 scores, and the ordinate indicates the values of the functional diversity indices for the grass layer in urban parks: Fmpd is the functional mean pairwise distance, Fnnd is functional mean nearest neighbour distance, FEve is the functional evenness, FRic is the functional richness, FDiv is the functional divergence, FOri is the functional originality, FSpe is the functional specialization, Fide_PC1, … PC5 are the Functional identity indeces, GSimpson is the Gini-Simpson, FunRao is the Rao quadratic entropy, FunRedundancy is the functional redundancy.
Figure 6.
The response of functional diversity indices along the gradient of the naturalness factor (PC2) is illustrated in the scatter plot, where blue dots represent the functional diversity indices across the PC1 gradient. The green line denotes the polynomial approximation of the relationship when it is identified as the optimal model, whereas the red line similarly indicates the polynomial approximation for the best model. In cases where the optimal model is a constant, no line is presented. The adjusted R-squared (Radj2) reflects the proportion of variation in the functional diversity index that is accounted for by the respective model. The P-value is derived from the results of the Likelihood-Ratio Test (LRT), which compares the best model to a model that includes only a constant. The abscissa represents the PC1 scores, and the ordinate indicates the values of the functional diversity indices for the grass layer in urban parks: Fmpd is the functional mean pairwise distance, Fnnd is functional mean nearest neighbour distance, FEve is the functional evenness, FRic is the functional richness, FDiv is the functional divergence, FOri is the functional originality, FSpe is the functional specialization, Fide_PC1, … PC5 are the Functional identity indeces, GSimpson is the Gini-Simpson, FunRao is the Rao quadratic entropy, FunRedundancy is the functional redundancy.
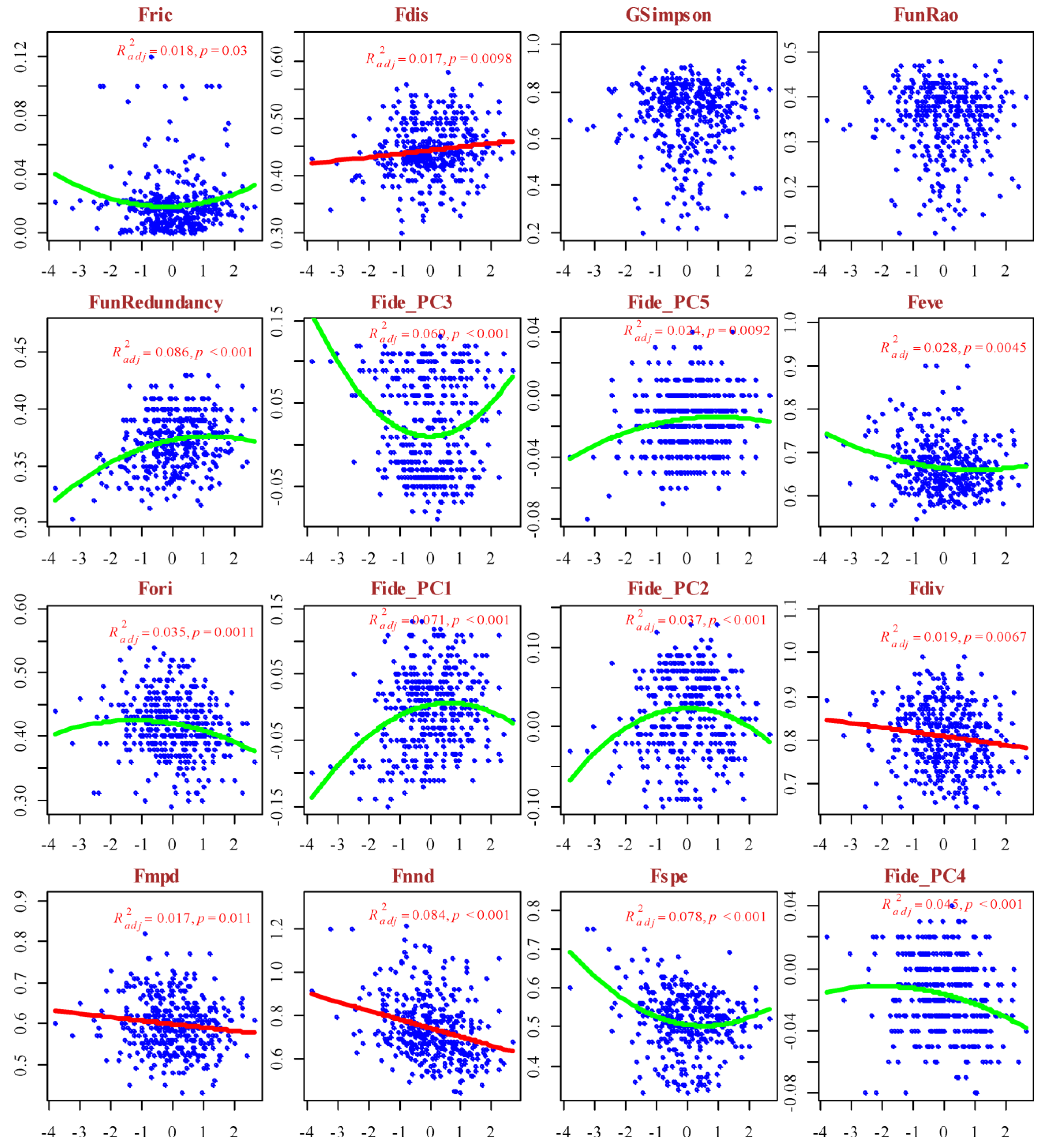
Figure 7.
Structural equation modelling of the effects of tree stands on environmental transformation, disturbance levels and functional properties of herbaceous layers in urban parks. The latent variables are depicted within hexahedra, whereas the manifest variables (indicators) are represented in quadrangles. Asterisks denote the statistical significance of the factor loadings: *** indicates P < 0.001, ** indicates P < 0.01, and * indicates P < 0.05. Fmpd is the functional mean pairwise distance, Fnnd is the functional mean nearest neighbour distance, FEve is the functional evenness, FRic is the functional richness, FDiv is the functional divergence, FOri is the functional originality, FSpe is the functional specialization, Fide_PC1, … PC5 are the Functional identity indices, GSimpson is the Gini-Simpson diversity, FunRao is the Rao quadratic entropy, FunRedundancy is the functional redundancy.
Figure 7.
Structural equation modelling of the effects of tree stands on environmental transformation, disturbance levels and functional properties of herbaceous layers in urban parks. The latent variables are depicted within hexahedra, whereas the manifest variables (indicators) are represented in quadrangles. Asterisks denote the statistical significance of the factor loadings: *** indicates P < 0.001, ** indicates P < 0.01, and * indicates P < 0.05. Fmpd is the functional mean pairwise distance, Fnnd is the functional mean nearest neighbour distance, FEve is the functional evenness, FRic is the functional richness, FDiv is the functional divergence, FOri is the functional originality, FSpe is the functional specialization, Fide_PC1, … PC5 are the Functional identity indices, GSimpson is the Gini-Simpson diversity, FunRao is the Rao quadratic entropy, FunRedundancy is the functional redundancy.
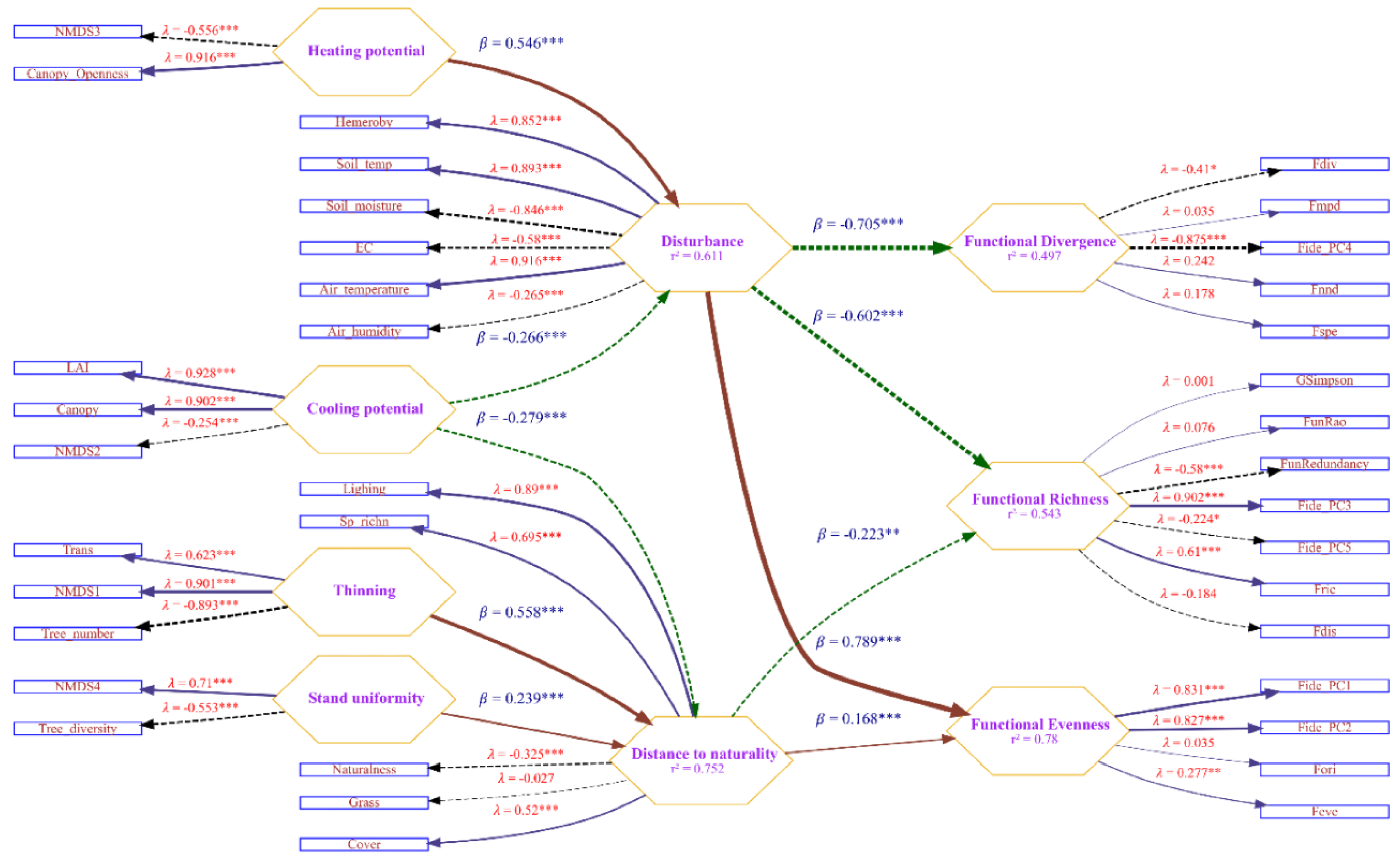

Table 1.
Parameters of the tree stands of the park plantations.
| Tree stand parameter | Park | Total (N = 380) |
Test | P-value | |
| Ivan Starov Square (N = 150) | Students’ Park (N = 230) |
||||
| Tree number per sample plot | 2.0±1.2 | 2.7±1.6 | 2.4±1.5 | Mann-Whitney | < 0.001 |
| Tree stand Shannon diversity | 0.9±0.6 | 0.9±0.6 | 0.9±0.6 | Student’s t-test | 0.68 |
| Cover, % | 51.7±19.9 | 51.5±26.0 | 51.6±23.7 | Student’s t-test | 0.93 |
| Canopy openness, % | 57.0±17.4 | 27.5±13.4 | 39.1±20.9 | Student’s t-test | < 0.001 |
| Leaf area index (LAI) | 0.9±0.6 | 2.1±0.8 | 1.6±0.9 | Student’s t-test | < 0.001 |
| Transparency (direct) | 5.1±3.9 | 4.4±2.9 | 4.7±3.3 | Student’s t-test | 0.044 |
| Transparency (diffuse) | 5.4±3.7 | 4.1±2.4 | 4.6±3.0 | Student’s t-test | < 0.001 |
| Transparency (total) | 10.5±7.3 | 8.5±4.9 | 9.3±6.1 | Student’s t-test | 0.002 |
Table 2.
Correlations among the soil, air, and herbaceous cover properties, as well as the dimensions, were derived from the results of nonmetric multidimensional scaling of the park tree stands. Only statistics that achieved significance at p < 0.05 are included.
Table 2.
Correlations among the soil, air, and herbaceous cover properties, as well as the dimensions, were derived from the results of nonmetric multidimensional scaling of the park tree stands. Only statistics that achieved significance at p < 0.05 are included.
| Variable | Factor loadings | Dimensions, derived from the results of nonmetric multidimensional scaling | ||||
| PC1, λ = 4.9 |
PC2, λ = 1.8 |
NMDS 1 | NMDS 2 | NMDS 3 | NMDS 4 | |
| Soil temperature (log-transformed) | 0.88 | – | 0.35 | 0.24 | –0.27 | – |
| Soil moisture (log-transformed) | –0.82 | – | –0.20 | –0.12 | 0.29 | – |
| Soil electric conductivity | –0.56 | – | – | – | 0.15 | – |
| Lighing (log-transformed) | 0.67 | 0.36 | 0.42 | 0.25 | –0.19 | 0.10 |
| Air temperature | 0.91 | – | 0.31 | 0.26 | –0.34 | – |
| Air humidity | –0.90 | – | –0.34 | –0.26 | 0.33 | – |
| Grass height | –0.46 | 0.66 | – | – | 0.23 | – |
| Herbaceous cover | – | 0.80 | 0.24 | – | – | 0.16 |
| Hemeroby | 0.84 | – | 0.27 | 0.21 | –0.38 | – |
| Naturalness | – | –0.40 | –0.15 | – | 0.11 | – |
| Herb species richness | 0.30 | 0.56 | 0.36 | – | – | 0.27 |
| PC1 | – | – | 0.36 | 0.22 | –0.35 | – |
| PC2 | – | – | 0.49 | – | – | 0.46 |
Table 3.
Relationships between all traits and all axes of the functional space. For continuous traits a linear model was computed, and the Pearson r was returned. For other types of traits, a Kruskal-Wallis test was computed, and the Kruskal-Wallis eta2 statistics were returned. Only those statistically significant at p < 0.05 are presented only.
Table 3.
Relationships between all traits and all axes of the functional space. For continuous traits a linear model was computed, and the Pearson r was returned. For other types of traits, a Kruskal-Wallis test was computed, and the Kruskal-Wallis eta2 statistics were returned. Only those statistically significant at p < 0.05 are presented only.
| Trait | Test | Functional axes | ||||
| PC1 | PC2 | PC3 | PC4 | PC5 | ||
| Leaf area | Linear Model | –0.32 | –0.58 | – | – | – |
| Leaf area per leaf dry mass | Linear Model | – | – | 0.49 | –0.29 | –0.40 |
| Leaf dry mass per leaf fresh mass | Linear Model | –0.58 | – | –0.34 | – | – |
| Leaf nitrogen (N) content per leaf dry mass | Linear Model | 0.47 | –0.21 | 0.28 | –0.49 | – |
| Plant height | Linear Model | – | – | – | –0.65 | 0.44 |
| Beginning of flowering period | Linear Model | 0.35 | – | – | – | 0.65 |
| End of flowering | Linear Model | 0.59 | 0.35 | 0.30 | – | – |
| Time of seed dispersal | Linear Model | 0.25 | – | – | – | 0.28 |
| Seed mass | Linear Model | –0.31 | –0.51 | – | – | – |
| Dispersal distance class | Linear Model | 0.58 | –0.57 | –0.18 | – | – |
| Seedbank duration | Linear Model | 0.76 | 0.23 | – | – | – |
| Light regime | Linear Model | 0.65 | 0.44 | – | – | 0.20 |
| Temperatures | Linear Model | – | –0.41 | –0.39 | 0.34 | – |
| Continentality of climate | Linear Model | 0.30 | 0.36 | – | 0.22 | – |
| Humidity | Linear Model | –0.41 | –0.23 | – | –0.63 | – |
| Acidity | Linear Model | – | – | – | – | – |
| Nutrients availability | Linear Model | – | –0.21 | 0.28 | –0.73 | – |
| Naturalness | Linear Model | –0.76 | –0.19 | 0.20 | – | – |
| Hemeroby | Linear Model | 0.84 | 0.21 | –0.24 | – | – |
| Climamorph | Kruskal-Wallis | 0.44 | 0.45 | 0.29 | – | 0.41 |
| Pollenochor | Kruskal-Wallis | – | 0.15 | 0.57 | 0.12 | – |
| Diasporochor | Kruskal-Wallis | 0.06 | 0.63 | 0.20 | 0.11 | 0.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



