Submitted:
19 November 2025
Posted:
20 November 2025
Read the latest preprint version here
Abstract
Background: Aging brains are shaped by a persistent dialogue between declining neurogenesis and rising neuroinflammation. Neural stem cells progressively lose regenerative capacity, while microglia and astrocytes shift toward maladaptive states that erode synaptic plasticity and cognition. This convergence defines inflammaging, a slow yet relentless process that undermines resilience. However, the field remains hampered by critical gaps: incomplete mapping of microglial heterogeneity, poorly understood epigenetic scars from inflammasome signaling, lack of longitudinal data, unclear niche-specific immune mechanisms, and uncertain cross-species relevance. This review addresses these pressing barriers, aiming to transform fragmented insights into actionable strategies. Summary: I chart how neurogenesis and neuroinflammation operate in continuous dialogue, identify five major knowledge gaps, and evaluate strategies to reprogram this interaction. Approaches include longitudinal imaging, niche-focused immunomodulation, glial subtype reprogramming, brain-penetrant inflammasome inhibitors, and CRISPR-based epigenetic editing. Each strategy is mapped against translational potential, short-term feasibility, and long-term vision, with emphasis on how mechanistic precision can guide clinical innovation. Conclusion: Here I highlight that neurogenic potential is not entirely lost with age but may be preserved or restored by tuning immune and epigenetic environments. This review proposes a roadmap for reshaping the aging brain’s fate, offering mechanistically grounded strategies to delay cognitive decline. Beyond neurology, the work underscores a broader principle: by integrating cellular plasticity with immune modulation, science edges closer to re-engineering resilience across the lifespan.
Keywords:
neurogenesis
; neuroinflammation
; aging brain
; microglia
; hippocampus
; cognitive decline
; Alzheimer disease
; inflammasomes
; epigenetics
; translational research
1. Introduction
The brain remains a dynamic organ across the lifespan, continuously reshaped by the birth of new neurons in specialized niches such as the hippocampus [1,2]. Far from being a relic of development, adult neurogenesis enriches learning, memory, and emotional resilience, safeguarding adaptability in a changing environment [3,4]. Yet this plasticity is not inexhaustible. With aging, neurogenic output wanes, cognitive reserve diminishes, and vulnerability to neurodegeneration grows [5,6]. This tension between a system designed for renewal and its gradual attrition defines a central challenge for brain health, setting the stage for how neurogenesis and neuroinflammation intersect in the aging brain [7,8].
Aging is accompanied by a persistent, low-grade inflammatory state often termed inflammaging, a process distinct from acute infection yet equally influential in shaping brain health [9,10]. In this slow fire, microglia gradually lose their homeostatic balance, adopting pro-inflammatory phenotypes that release cytokines and chemokines [11,12]. Astrocytes amplify this tone, shifting toward reactive states that erode trophic support and disrupt neuronal networks [13,14]. Vascular changes weaken the blood-brain barrier (BBB), while peripheral immune signals infiltrate and reinforce local inflammation [15,16]. Together, these subtle but enduring perturbations accumulate over decades, progressively altering cellular behavior and circuit resilience [17,18,19].
Across the aging hippocampus, two intertwined trajectories emerge: a steady decline in neurogenesis and a progressive rise in neuroinflammation [20,21]. Diminished neural stem cell activity and impaired maturation of adult-born neurons reduce pattern separation, flexibility in memory strategies, and mood regulation [22,23]. At the same time, microglia shift toward a proinflammatory state, releasing cytokines such as interleukin-1 beta (IL-1β) and TNF while offering reduced trophic support, thereby altering the niche [22,24]. Rather than separate phenomena, these arcs converge into a bidirectional dialogue in which inflammation curtails neurogenesis, and neurogenic failure amplifies vulnerability to inflammatory stressors [8,25].
Microglia operate as finely tuned gatekeepers of the neurogenic niche, shaping whether new neurons thrive or fail [26,27]. In youthful contexts, they clear apoptotic cells, sculpt synapses with precision, and secrete trophic factors such as brain-derived neurotrophic factor (BDNF) and IGF-1 that sustain progenitor proliferation and survival [26,28]. Yet chronic inflammatory tone rewires their functions: cytokine release intensifies, complement-driven pruning accelerates, and phagocytic activity becomes biased toward eliminating viable cells [29,30]. This shift suppresses neurogenesis, disrupts circuit integration, and fosters vulnerability [27,31]. Crucially, youthful microglia are not merely less reactive; they are actively programmed toward pro-neurogenic states that are progressively lost with age [29,32].
Experimental models demonstrate that inflammatory insults sharply constrain adult hippocampal neurogenesis [33,34]. Acute lipopolysaccharide challenges, chronic peripheral inflammation, or autoimmune insults reduce progenitor proliferation, neuronal survival, and integration [35,36]. Conversely, targeted interventions ranging from pharmacological agents to trophic factors rescue aspects of neurogenesis by dampening microglial activation or restoring signaling cascades such as PI3K-Akt, ERK, or wingless-related integration site signaling pathway (Wnt)/β-catenin [37,38]. Yet evidence cautions against simple “anti-inflammation” strategies: both excessive suppression and prolonged activation can be deleterious [31,39]. Outcomes depend on timing, intensity, and niche context, underscoring the need for mechanistic precision in modulating pathways, microglial states, and local environments [8,40].
Despite mounting evidence, several gaps blunt causal inference and stall translation. First, microglial diversity is strikingly region-specific, varying across niches and brain regions in ways that defy simple categorization [41,42]. Second, inflammasome-driven epigenetic reprogramming can lock progenitors and microglia into low-neurogenic states, yet its dynamics remain elusive [43,44]. Third, longitudinal tracking of neuroimmune interactions across the lifespan is scarce, leaving temporal trajectories speculative [45,46]. Fourth, incomplete maps of vascular and T-cell signals within niches obscure critical modulators of microglial tone [47,48]. Finally, cross-species disconnects confound human relevance, undermining therapeutic extrapolation from animal models [49,50].
This review sets out a playbook for rewiring the neuroimmune dialogue by linking mechanistic insight to translational strategy. I map unresolved gaps to actionable approaches: longitudinal neuroimmune imaging to capture temporal causality, niche-focused immunomodulation to tune local signals, and glial subtype reprogramming to restore supportive states or even generate new neurons. I highlight brain-penetrant nod-like receptor protein 3 (NLRP3) inhibitors and nucleic acid therapeutics as near-term strategies to break maladaptive IL-1β loops, while CRISPR-based epigenetic editing represents a longer-horizon tool to reset maladaptive chromatin programs. Together, these advances reframe therapeutic feasibility in aging.
Preserving a youthful neurogenic niche holds the promise of sustaining cognitive reserve, delaying neurodegeneration, and enhancing resilience across the lifespan [51]. Mechanistic advances reveal that both lifestyle factors and molecular interventions can counter inflammaging, rejuvenate progenitors, and restore plasticity [52]. The challenge is moving from associations to actionable strategies that align biology with translation [53]. This review charts that trajectory by first detailing the intertwined biology of neurogenesis and neuroinflammation, then interrogating five critical gaps that obscure causality, and finally evaluating emerging strategies with human applicability in view, guiding readers from concept to clinic-ready hypotheses.
Figure 1.
Neurogenesis–neuroinflammation in the aging brain This figure will show a young vs. aged brain/neurogenic niche: in the young panel, active neural stem cells in the dentate gyrus (DG) with supportive microglia secreting brain-derived eutrophic factor (BDNF) and growth factors; in the aged panel, fewer newborn neurons and activated microglia (releasing cytokines, IL-1β) alongside infiltrating T cells. BDNF, brain-derived eutrophic factor; DG, dentate gyrus, IL-1β, interleukin-1 beta).
Figure 1.
Neurogenesis–neuroinflammation in the aging brain This figure will show a young vs. aged brain/neurogenic niche: in the young panel, active neural stem cells in the dentate gyrus (DG) with supportive microglia secreting brain-derived eutrophic factor (BDNF) and growth factors; in the aged panel, fewer newborn neurons and activated microglia (releasing cytokines, IL-1β) alongside infiltrating T cells. BDNF, brain-derived eutrophic factor; DG, dentate gyrus, IL-1β, interleukin-1 beta).
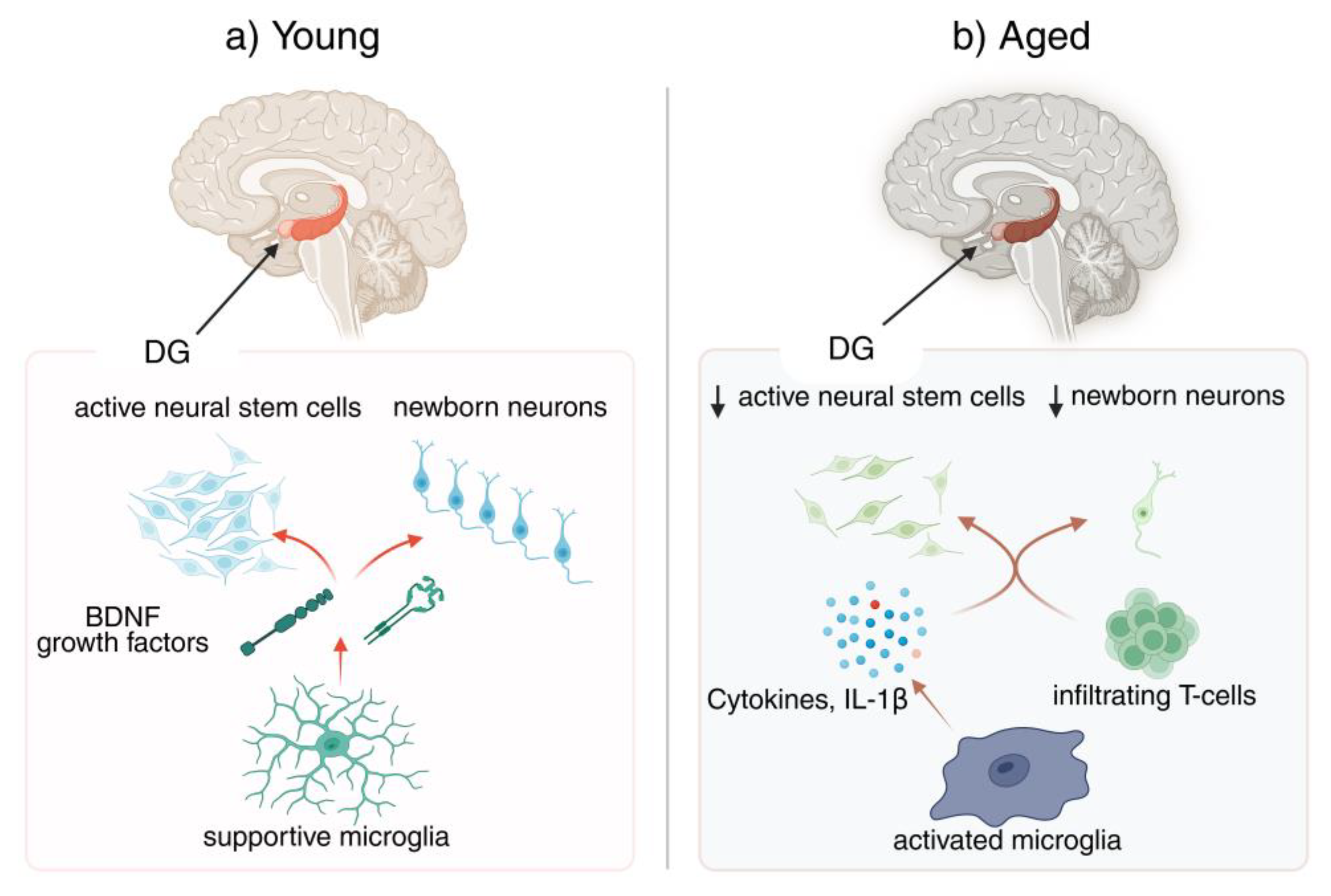
2. Neurogenesis and Neuroinflammation in the Aging Brain: An Overview
Adult mammalian brains retain a limited capacity for neurogenesis, confined mainly to the dentate gyrus (DG) of the hippocampus and the SVZ zone, where neural stem cells generate new neurons that integrate into existing circuits [54,55]. This process diminishes with age, as stem cells proliferate less and fewer neurons survive to maturity [22,56]. In parallel, microglia gradually adopt a primed, pro-inflammatory phenotype, releasing cytokines such as IL-1β and TNFα that impair progenitor proliferation and neuronal differentiation [57,58]. By contrast, anti-inflammatory and trophic factors like IL-4, IL-10, IGF-1, and BDNF promote neurogenesis [59,60]. The balance between these opposing signals shifts during “inflammaging,” when systemic immune mediators and infiltrating cells increasingly shape the neurogenic niche [57,61,62,63].
2.1. Adult Neurogenesis: Mechanisms and Age-Related Decline (~400 words)
Adult neurogenesis in the mammalian brain occurs primarily in two discrete regions, the subgranular zone of the hippocampus and the SVZ zone of the forebrain, where astrocyte-like neural stem cells sustain lifelong plasticity by generating new neurons and glia [64,65]. Within these niches, stem cells undergo sequential steps of proliferation, lineage commitment, and differentiation into intermediate progenitors that ultimately mature into functional granule neurons or glial cells [66,67]. Newly generated neurons progress through migration, synaptic integration, and circuit incorporation, thereby reshaping hippocampal and olfactory networks while maintaining a dynamic balance between neuronal and glial lineages [68,69].
The regulation of adult neurogenesis reflects a delicate interplay between intrinsic genetic programs and extrinsic environmental cues [60,70]. Transcription factors and epigenetic mechanisms orchestrate lineage progression, guiding neural stem cells from quiescence toward neuronal or glial differentiation [54,71]. Simultaneously, the neurogenic niche provides trophic support, vascular inputs, glial signaling, and neuronal activity that sustain proliferation and integration [72,73]. Acting as a dynamic coordinator, the niche integrates these signals to preserve stem cell function and ensure a balanced neurogenic output under physiological conditions [74,75].
Rodent studies consistently demonstrate that adult neurogenesis undergoes a steep decline with advancing age, marked by reduced progenitor proliferation and a shrinking contribution of new neurons to hippocampal circuits [76,77]. While neural stem cells persist, their output is curtailed by prolonged quiescence, asymmetric division, and intrinsic alterations such as diminished lamin B1 expression [78,79]. Equally decisive is the aging microenvironment: decreased trophic support, vascular dysfunction, and elevated TGF-β and inflammatory signaling constrain neurogenic potential [80,81]. These findings underscore that the decline is not due to progenitor loss but to niche deterioration, which restricts activation and differentiation despite preserved stem cell reservoirs [82,83,84].
Evidence from human postmortem and imaging studies indicates that hippocampal neurogenesis likely persists across adulthood, with several investigations detecting thousands of immature neurons in healthy individuals well into the eighth or even ninth decade [1,85]. Yet, other studies describe steep age-related reductions in proliferation and neurogenic markers, despite the continued presence of progenitor cells [22,86]. These conflicting findings are often attributed to methodological differences in tissue processing and marker detection [87,88]. The resulting uncertainty contrasts with rodent data, where neurogenic potential is retained but niche decline dominates, creating a translational dilemma that frames ongoing cross-species comparisons [86,89].
2.2. Neuroinflammation in Aging: Microglia and Beyond
The aging brain is characterized by a progressive remodeling of its immune landscape, where a state of low-grade but chronic neuroinflammation becomes a defining hallmark [90,91,92]. Central to this shift are microglia, the resident immune cells that gradually lose their homeostatic and reparative functions while adopting pro-inflammatory, neurotoxic phenotypes [11,12,93]. Hallmarks of aged microglia include altered transcriptomes, dystrophic morphology, impaired phagocytosis, and exaggerated cytokine release [11,94,95]. Yet microglia do not act alone; astrocytic immunosenescence and peripheral immune inputs further amplify inflammatory tone, contrasting sharply with the supportive environment of younger brains [96,97,98].
Aged microglia are marked by dystrophic morphology, diminished phagocytic capacity, and transcriptional reprogramming that favors pro-inflammatory gene expression over reparative functions [11,99,100]. This deterioration is compounded by microglial priming, a process in which prior immune or metabolic challenges leave cells in a state of innate immune memory, heightening their responsiveness to subsequent insults [101,102,103]. Primed microglia release exaggerated amounts of cytokines such as IL-1β, IL-6, and TNFα, impairing synaptic plasticity and accelerating neurodegeneration [17,102,104]. Even when replaced experimentally, aged microglia retain their hyperreactivity due to niche-driven cues, underscoring how priming locks biases the aging brain into a maladaptive inflammatory state [100,105].
Chronic activation of inflammatory pathways is a hallmark of microglial aging, with NF-κB signaling and NLRP3 inflammasome activity emerging as central drivers [106,107]. Their persistent activation sustains the release of IL-1β, IL-6, and TNFα, creating a hostile milieu that erodes neuronal survival and actively suppresses adult neurogenesis [108,109,110]. These cytokines impair progenitor proliferation, bias glial differentiation, and disrupt synaptic plasticity, gradually shifting the neurogenic niche from supportive to inhibitory [8,111,112]. Importantly, mitochondrial dysfunction and oxidative stress further amplify NF-κB and NLRP3 activity, locking the system into a cycle of chronic inflammation that undermines regenerative capacity in the aging brain [110,113,114,115,116].
With advancing age, the blood–brain barrier becomes increasingly permeable, weakening its selective function and permitting infiltration of peripheral immune cells [16,117]. Among these, CD8⁺ T cells accumulate in neurogenic niches of aged mice and humans, where they release interferon-γ and other cytokines that suppress neural stem cell proliferation and neuronal differentiation [118,119]. Aged microglia further facilitate this process by secreting chemokines and remodeling the niche microenvironment, creating a feed-forward inflammatory loop [47,120]. In sharp contrast, the young brain maintains a largely anti-inflammatory, pro-neurogenic milieu, highlighting how immune remodeling with age tilts the balance away from regeneration toward chronic dysfunction [31,120,121].
2.3. Microglia–Neural Stem Cell Crosstalk
Microglia have emerged as central orchestrators of adult neurogenesis, engaging in a continuous dialogue with neural stem and progenitor cells that shapes every stage of the process [28,122,123]. Far from passive sentinels, they actively sculpt the neurogenic niche by phagocytosing apoptotic newborn cells, thereby maintaining homeostasis and determining which neurons survive to maturity [123,124]. Microglia also refine synaptic connections of adult-born neurons through selective pruning, ensuring proper integration into existing circuits [27,28,125]. Beyond these structural roles, their secretome exerts powerful influence, releasing context-dependent cues that either promote proliferation and differentiation or restrict neurogenesis, highlighting their dual capacity as nurturers or inhibitors [123,126].
In the young brain, microglia frequently adopt phenotypes that nurture rather than hinder neurogenesis [127,128,129]. By secreting trophic factors such as BDNF, IGF-1, and TGF-β, they stimulate neural stem cell proliferation, guide differentiation, and promote survival of newborn neurons [26,129,130]. Environmental enrichment and physical activity further enhance this supportive role, shifting microglia toward anti-inflammatory states that amplify plasticity and circuit integration [26,131,132]. M2-polarized microglia in particular foster neuronal differentiation and synaptic maturation, underscoring their capacity to translate systemic and local signals into pro-neurogenic outcomes [133,134,135]. This trophic partnership highlights microglia as crucial allies in sustaining hippocampal resilience early in life [26,128,136].
Aging and chronic stress profoundly disrupt microglia–neural stem cell interactions by driving microglia toward pro-inflammatory, neurotoxic states [101,137]. In this maladaptive phenotype, microglia secrete elevated levels of IL-1β, TNFα, and IL-6, which suppress NSC proliferation, reduce BDNF availability, and block the maturation of newborn neurons [12,93,138]. At the same time, microglia lose their phagocytic balance, leading to excessive or aberrant pruning that compromises neuronal survival and synaptic plasticity [11,93,139]. Impaired autophagy and metabolic dysfunction exacerbate these changes, locking aged and stressed microglia into states that foster chronic inflammation and progressively undermine the regenerative potential of the neurogenic niche [93,94,140].
Microglia–NSC crosstalk is mediated by finely tuned molecular pathways, with the C-X3-C motif chemokine ligand 1 (CX3CL1)–C-X3-C motif chemokine receptor 1 (CX3CR1) axis emerging as a central regulator of microglial activation, synaptic integration, and neurogenic support [141,142]. This signaling maintains microglial quiescence, limits cytokine release, and facilitates proper maturation of adult-born neurons, while its disruption impairs dendritic spine formation and neurogenesis [142,143,144]. Other modulators, including cytokines, chemokines, and extracellular vesicles (EVs), complement this dialogue by shaping microglial states and their influence on progenitors [145,146,147]. With aging, these pathways shift from protective to maladaptive, fostering chronic inflammation and reduced neurogenic output, thereby highlighting their therapeutic relevance [141,148] (Table 1).
3. Critical Gaps in Current Knowledge
In this section, I outline five critical gaps that currently limit our understanding of how neuroinflammation shapes neurogenesis across the aging trajectory. These challenges range from fundamental biology to translational relevance. First, the field lacks a clear picture of region-specific microglial diversity in aging brains and how this heterogeneity impacts stem cell dynamics. Second, the role of inflammasome-driven epigenetic alterations in sustaining or amplifying neurogenic decline remains poorly defined. Third, the absence of longitudinal data obscures temporal causality between inflammation and neurogenesis. Fourth, the contribution of niche-specific immune mechanisms is still unresolved. Finally, cross-species disconnects complicate translation, underscoring the urgency for human-relevant models.
3.1. Gap 1 – Region-Specific Microglial Diversity in Aging
Microglia are increasingly recognized as a heterogeneous population whose identities vary across brain regions rather than fitting a uniform template [41]. Transcriptomic and single-cell profiling studies reveal distinct gene expression and morphological features in microglia from the cortex, hippocampus, cerebellum, and other regions, with some subsets tuned toward surveillance and others toward immune activation [150]. Aging accentuates these differences, reshaping transcriptional signatures in a region-dependent manner and amplifying selective vulnerabilities [41]. This diversity represents a critical yet underexplored determinant of brain aging and resilience [91].
Aging imprints distinct signatures on microglia across brain regions, revealing striking contrasts in phenotype and function [41]. In the hippocampus, transcriptomic analyses show upregulation of adhesion and motility genes, aligning with greater sensitivity to inflammatory and metabolic stress, while cerebellar microglia appear comparatively stable [41]. Experimental challenges further underscore this diversity: TNFα or systemic LPS elicit robust and prolonged activation in hippocampal microglia, yet only muted responses in other regions [151]. These findings highlight that microglial aging is not uniform [41]. What remains unresolved is how such region-specific shifts shape neuronal survival, plasticity, and ultimately cognitive aging [91].
Neurogenic regions such as the DG and SVZ zone rely on close interactions between neural stem cells and local microglia, yet whether these microglia display unique aging trajectories remains an unresolved question [21]. Evidence suggests that niche-resident microglia have specialized roles, from supporting neuroblast migration to modulating survival signals [152]. With age, however, these populations undergo positional remodeling and progressive activation that may create antineurogenic environments [120]. What is missing is systematic mapping of their transcriptional and functional states across the lifespan [21]. Without such resolution, it is difficult to disentangle whether neurogenesis declines mainly from local niche deterioration or reflects broader systemic shifts in microglial aging [153].
Resolving how microglial diversity shapes neurogenic decline carries profound implications for therapy [154]. If hippocampal microglia are particularly prone to adopting pro-inflammatory, anti-neurogenic profiles with aging, while SVZ microglia preserve more supportive functions, this could help explain selective vulnerabilities in cognition and neurodegeneration [155]. Such distinctions suggest that interventions need not silence microglia globally but instead target maladaptive phenotypes in specific regions [42]. High-resolution profiling of microglial states in neurogenic versus non-neurogenic regions will therefore be critical [156]. These insights could enable tailored strategies that restore hippocampal neurogenesis locally while preserving beneficial immune surveillance elsewhere [157].
3.2. Gap 2 – Inflammasome-Driven Epigenetic Alterations
Persistent activation of the NLRP3 inflammasome has emerged as a hallmark of brain aging, shaping a chronic inflammatory environment that disrupts neuronal and stem cell homeostasis [158]. Unlike acute responses, which are transient and protective, aged microglia remain locked in an overactivated state, driving continual secretion of IL-1β and interleukin-18 (IL-18) [159]. This sustained output fuels neuroinflammation, amplifies synaptic dysfunction, and accelerates neuronal loss [158]. Evidence from Alzheimer’s disease (AD) and other age-related contexts shows that NLRP3 activation is maintained by metabolic stressors and amyloid accumulation, marking it as a central instigator of the pro-inflammatory niche characteristic of the aged brain [160].
A growing body of evidence shows that aged microglia carry an epigenetic memory of past inflammatory encounters, leaving behind enduring “scars” that sustain maladaptive activity [161]. Hypomethylation of the IL-1β promoter, for instance, maintains excessive cytokine release in aging brains and drives persistent neuroinflammation [161]. Such chromatin-based reprogramming distinguishes transient immune responses from long-lasting dysfunction [161]. Neural stem cells in inflamed niches may undergo similar repressive modifications at pro-neurogenic loci, reducing their regenerative potential [162]. Parallels with hematopoietic stem cells, where chronic inflammasome signaling reshapes enhancer accessibility, underscore how inflammation imprints itself epigenetically to constrain stem cell function across tissues [163].
Whether inflammasome signaling directly reshapes the epigenome of neural stem cells in the DG or SVZ zone remains an unresolved question [164]. NSCs in these regions may acquire repressive chromatin marks that blunt their regenerative responses long after inflammatory cues dissipate, yet systematic evidence is lacking [164]. Do aged NSCs inherit such “epigenetic scars,” locking them into diminished neurogenic potential? [164] Current studies describe epigenetic regulation in adult NSCs and chromatin remodeling during aging, but they rarely examine inflammasome-driven mechanisms [71]. Without detailed chromatin and transcriptomic maps of inflamed niches, the link between persistent inflammation and neurogenic failure remains speculative [164].
If inflammasome activity imprints lasting epigenetic scars on neural stem cells, these changes could persistently dampen neurogenesis and lower the ceiling of regenerative responses, even after inflammatory cues subside. However, studies of ischemia and stroke in rodents and humans demonstrate that locally activated, injury-responsive stem cells with reprogrammed phenotypes can still be recruited within damaged areas and contribute to neurogenesis [278,279]. Thus, rather than an absolutely “locked” state, inflammasome-driven epigenetic memory may shift neural stem cells along a continuum of reduced but still reactivatable potential, with important implications for how we design interventions to reset the niche.
3.3. Gap 3 – Longitudinal Dynamics of Neuroimmune Interactions
Most studies examining the interplay between neuroinflammation and neurogenesis rely on static snapshots, typically contrasting young and old animals or measuring endpoints after an inflammatory insult [167]. While such designs capture broad differences, they cannot reconstruct dynamic trajectories or reveal causal order [168]. It remains unclear whether inflammatory changes precede neurogenic decline, arise in parallel, or follow as a secondary consequence [167]. Cross-sectional single-cell and epigenomic studies have enriched our understanding of cell states, yet they provide only frozen moments in time, leaving the temporal choreography of neuroimmune aging unresolved [169].
The absence of longitudinal tracking obscures causal interpretation of neuroimmune interactions in aging [170]. We still do not know precisely when microglial priming begins to meaningfully suppress neurogenesis, or whether short-lived insults such as infection, stress, or metabolic imbalance leave enduring dents in neuronal production [45]. Most evidence comes from cross-sectional or endpoint analyses, which show associations but not sequence [170]. Without continuous data, we cannot determine if chronic inflammation initiates, parallels, or simply follows the decline in neurogenic potential [170].
The timing of intervention may prove as critical as the intervention itself [171]. If inflammation-driven suppression of neurogenesis begins earlier than currently assumed, therapeutic windows may lie in mid-life rather than late life [171]. Evidence from stroke, ’s models, and systemic inflammation shows that acute inflammatory episodes often precede lasting neurogenic decline, and that early modulation of microglial or inflammasome activity can preserve regenerative capacity [38]. These findings underscore that therapies applied too late risk diminished efficacy, whereas timely intervention may sustain lifelong plasticity [172].
Closing the temporal gap will require methodological advances that move beyond static measures [173]. Longitudinal in vivo imaging of both neurogenesis and neuroinflammation, coupled with emerging PET tracers, offers one promising path [174]. Parallel development of peripheral and central biomarkers, alongside chronic experimental paradigms rather than acute LPS challenges, is equally critical [175]. Ultimately, integrated strategies that link molecular, cellular, and systems-level dynamics are needed to capture how neuroimmune interactions unfold across the lifespan [176].
Clarifying the temporal sequence between inflammation and neurogenesis is pivotal for understanding brain aging [21]. If chronic inflammation proves to be a driver, consequence, or both in neurogenic decline, this will fundamentally reshape strategies for preserving neural plasticity [177]. Untangling this interplay is therefore essential for precision approaches that safeguard neurogenic capacity across the lifespan and ultimately inform how we design therapies to maintain cognition and resilience in aging [178].
3.4. Gap 4 – Niche-Specific Immune Mechanisms
The subgranular zone (SGZ) of the hippocampus and the SVZ of the lateral ventricles form highly specialized neurogenic niches, distinct from the broader brain parenchyma [73]. These microenvironments bring together neural stem cells, progenitors, astrocytes, microglia, endothelial cells, and, in aging, even infiltrating immune cells [73]. Yet, despite advances in transcriptomic and proteomic profiling, we still lack a clear map of which immune and inflammatory signals within these niches directly regulate stem cell activity [179]. Equally unresolved is whether resident or infiltrating immune cells dominate in suppressing neurogenesis during aging [179].
Growing evidence implicates inflammatory cues as major inhibitors of neurogenesis within the SVZ and subgranular zones [179]. Microglial-derived cytokines such as IL-1β, TNFα, and IL-6 consistently emerge as candidates, while monocyte infiltration and CD8+ T cell activity in the SVZ have also been linked to reduced neurogenic potential [118]. Yet causality and relative contributions remain unresolved [180]. Systemic inflammation further complicates matters by altering blood–brain barrier integrity and selectively reshaping niche immune composition, but the permeability and vulnerability of these sites are still poorly defined [181].
Not all immune influences within the neurogenic niche are detrimental [122]. Signals such as TGF-β, IL-10, IGF-1, and CX3CL1 are increasingly recognized as protective factors that can sustain or even restore neurogenesis [182]. Yet whether these mediators act in a niche-specific manner and how their decline contributes to age-related collapse of neurogenic capacity remain unanswered questions [183]. It is also unclear whether immune checkpoints or anti-inflammatory feedback loops normally shield stem cells from inflammatory stress but fail with aging, leaving the niche vulnerable to irreversible dysfunction [183].
The idea of a “niche immunome” has emerged as a powerful framework to decode the immune and inflammatory signals that shape neurogenic niches [184]. Single-cell and spatial transcriptomic approaches now allow systematic profiling of the SGZ and SVZ across age, revealing immune pathways that bulk parenchymal studies cannot resolve [118]. Such resolution is crucial for distinguishing local immune regulation of neural stem cells from generalized brain inflammation, and for uncovering niche-specific vulnerabilities that may define regenerative potential in aging [185].
Bridging neuroimmunology with regenerative neuroscience requires moving beyond broad immunosuppression toward niche-specific interventions [186]. Without precise insight into the immune circuits of the SGZ and SVZ, therapies risk silencing protective signals while failing to restore neurogenesis [186]. Evidence from aging models shows that targeted modulation of microglia, T cells, or cytokine pathways can rejuvenate neurogenic capacity, underscoring the therapeutic promise of restoring local immune balance [100]. Defining these mechanisms positions the niche immunome as a critical frontier for precision interventions in brain aging [186].
3.5. Gap 5 – Translational and Cross-Species Disconnects
Most of what we know about the interplay between neuroinflammation and neurogenesis comes from rodent studies, yet rodents differ profoundly from humans in biology, lifespan, and environment [187]. Mechanisms that restore neurogenesis or cognition in mice often fail in clinical settings because molecular programs, immune responses, and even circadian rhythms diverge across species [95]. Human microglia show distinct transcriptional heterogeneity, and adult neurogenesis itself is limited and debated in humans compared with rodents [188]. This translational disconnect remains a central barrier, slowing progress from mechanistic insight to therapies that could counteract age-related cognitive decline [187].
Rodents display strikingly robust adult neurogenesis, both in the hippocampal DG and in the SVZ–olfactory bulb pathway, where thousands of new neurons are continually produced and integrated [189]. In contrast, humans lack meaningful SVZ-driven olfactory bulb neurogenesis, and hippocampal neurogenesis, though reported, is modest, controversial, and appears to decline with age [187]. Some studies suggest persistence across the lifespan, while others argue it is virtually absent in adulthood [89]. This lack of consensus complicates translational efforts, as strategies that reliably boost rodent neurogenesis—such as environmental enrichment, exercise, or pharmacological interventions—may have little impact in humans [190]. Without resolving this debate, applying pro-neurogenic therapies clinically remains fraught with uncertainty [191].
Human neuroinflammation diverges markedly from rodent models, complicating translational efforts [192]. Single-cell studies reveal that while core microglial programs are conserved, human microglia exhibit greater transcriptional heterogeneity, unique complement and phagocytic modules, and a baseline preactivated state not mirrored in rodents [188]. Moreover, human immune aging is shaped by lifelong infections, systemic comorbidities, and lifestyle exposures absent in laboratory animals, producing compounded inflammatory stress [94]. These differences raise concerns that interventions restoring neurogenesis in mice, such as exercise, cytokine modulation, or small molecules, may not yield comparable benefits in the aged human brain, underscoring the need for human-specific models and biomarkers [193].
Closing the translational gap requires models that capture the complexity of the human neuroimmune environment more faithfully than rodents [194]. Non-human primates offer closer physiology, yet complementary systems such as human brain organoids and induced pluripotent stem cell (iPSC)-derived microglia–NSC co-cultures now provide scalable and mechanistically precise tools [195]. These platforms permit interrogation of key pathways like NLRP3 or CX3CR1 in human-relevant contexts, while enabling controlled testing of immunomodulatory and pro-neurogenic therapies [194]. By combining primate studies with organoid and chip-based systems, researchers can generate clinically predictive insights that accelerate the translation of neuroimmune discoveries into interventions for aging-related decline [193].
Without human-specific insight, immunomodulatory therapies risk being blunt instruments, either failing to restore neurogenesis or disrupting essential immune functions [110]. Bridging species differences is therefore indispensable. By refining targets within human-relevant systems, interventions can be designed to preserve or even rejuvenate neurogenic capacity in aging, offering a path to meaningfully rewire the brain’s fate in clinical reality [196,197].
Figure 2.
Region-specific microglial diversity and aging. Region-specific microglial heterogeneity across the aging brain, illustrating how transcriptional and morphological diversification emerges in distinct neural niches. The figure contrasts microglial states in the hippocampus, cortex, and cerebellum, using either quantitative panels (e.g., bar plot or heatmap derived from single-cell RNA sequencing datasets comparing young versus aged microglia) or conceptual illustrations showing divergent activation profiles. Hippocampal microglia are depicted as more primed and pro-inflammatory with age, cortical microglia display mixed transitional phenotypes, while cerebellar microglia retain comparatively ramified and homeostatic features. This visualization highlights that microglial aging is non-uniform across regions, underscoring differential vulnerability of neurogenic pathways and supporting the rationale for spatially tailored therapeutic strategies. DAM, disease-associated microglia.
Figure 2.
Region-specific microglial diversity and aging. Region-specific microglial heterogeneity across the aging brain, illustrating how transcriptional and morphological diversification emerges in distinct neural niches. The figure contrasts microglial states in the hippocampus, cortex, and cerebellum, using either quantitative panels (e.g., bar plot or heatmap derived from single-cell RNA sequencing datasets comparing young versus aged microglia) or conceptual illustrations showing divergent activation profiles. Hippocampal microglia are depicted as more primed and pro-inflammatory with age, cortical microglia display mixed transitional phenotypes, while cerebellar microglia retain comparatively ramified and homeostatic features. This visualization highlights that microglial aging is non-uniform across regions, underscoring differential vulnerability of neurogenic pathways and supporting the rationale for spatially tailored therapeutic strategies. DAM, disease-associated microglia.
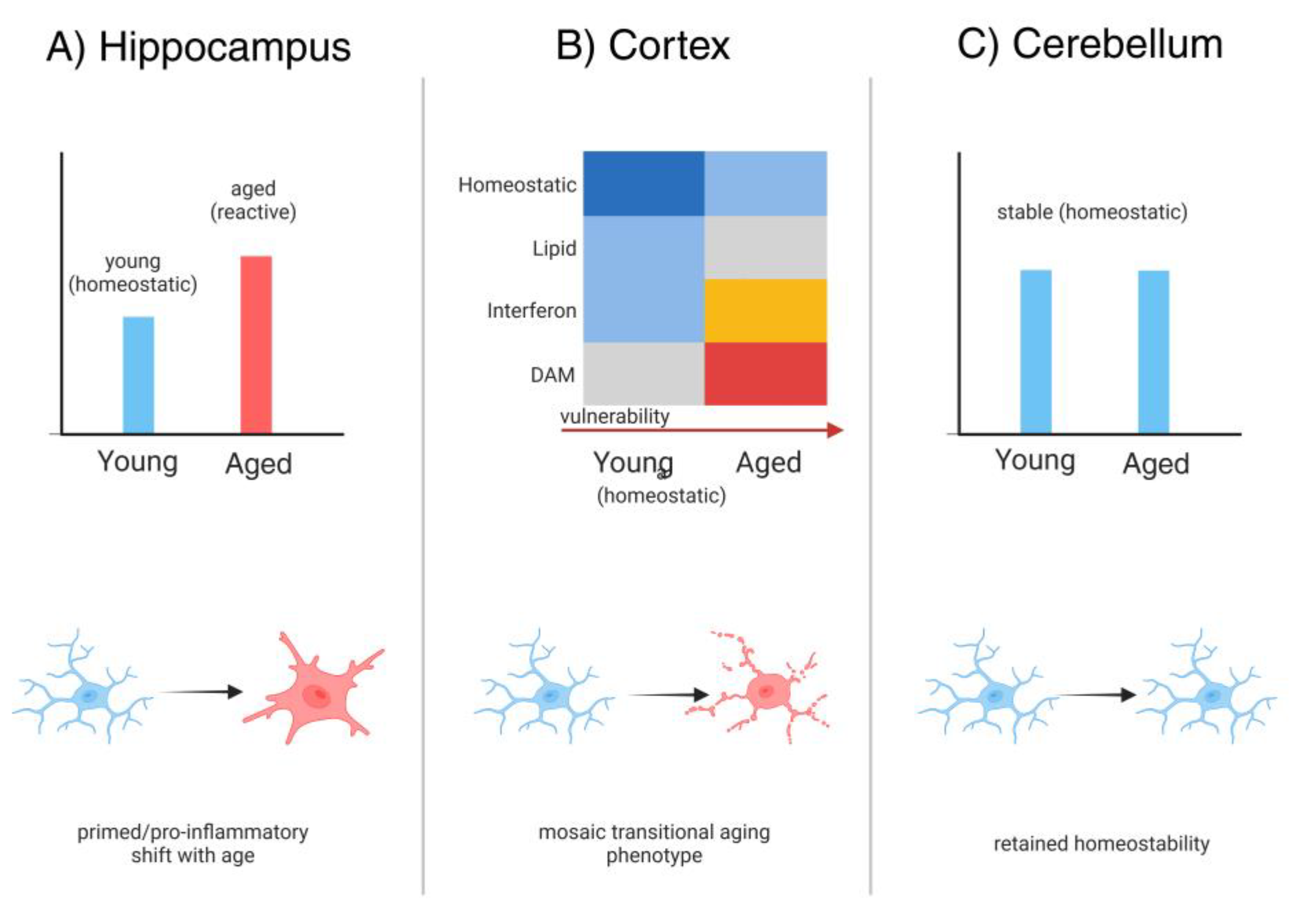

Table 2.
Five key knowledge gaps in neurogenesis–neuroinflammation. Five key knowledge gaps linking neurogenesis and neuroinflammation, summarizing unresolved questions, current unknowns, biological and clinical relevance, and candidate methodological strategies. Columns include: Gap, Description of Unknown, Why it Matters / Potential Consequences, and Suggested Approaches. Representative entries may include regional microglial specialization, age-dependent inflammatory plasticity, inflammasome–neurogenesis coupling, long-term effects of transient immune activation, and sex-specific neuroimmune interactions. This table provides a rapid reference to complement Section 3 and guide hypothesis formulation, experimental design, and translational priority setting.
Table 2.
Five key knowledge gaps in neurogenesis–neuroinflammation. Five key knowledge gaps linking neurogenesis and neuroinflammation, summarizing unresolved questions, current unknowns, biological and clinical relevance, and candidate methodological strategies. Columns include: Gap, Description of Unknown, Why it Matters / Potential Consequences, and Suggested Approaches. Representative entries may include regional microglial specialization, age-dependent inflammatory plasticity, inflammasome–neurogenesis coupling, long-term effects of transient immune activation, and sex-specific neuroimmune interactions. This table provides a rapid reference to complement Section 3 and guide hypothesis formulation, experimental design, and translational priority setting.
| Gap | Description of Unknown | Why it Matters / Consequences | Suggested Approaches |
|---|---|---|---|
| 1. Regional Microglial Diversity | Limited understanding of how microglial phenotypes differ across brain regions and influence neurogenesis | Regional vulnerabilities exist (hippocampus vs. olfactory bulb); lack of clarity hampers targeted interventions | Single-cell RNA-seq, region-specific lineage tracing, conditional microglial manipulation |
| 2. Inflammasome Dynamics in Aging | Unresolved timeline of NLRP3/other inflammasome activation in aged niches | Unclear when inflammasome priming becomes irreversible; timing critical for therapeutic window | Longitudinal transcriptomics, in vivo biosensors, inducible knockout models |
| 3. Crosstalk Between Peripheral and CNS Immunity | Mechanisms of how peripheral T cells and cytokines reshape neurogenic niches remain obscure | Infiltrating T cells alter NSC fate; missing mechanistic detail limits translation to systemic therapies | Fate-mapping of immune infiltration, parabiosis, targeted blockade of adhesion molecules |
| 4. Beneficial vs. Detrimental Microglial States | Poorly defined markers distinguishing pro-neurogenic vs. antineurogenic microglial states | Current therapies risk indiscriminate immunosuppression; need precision immunomodulation | Multi-omics integration (proteome, epigenome), machine-learning-based state classification, microglia-specific drug screens |
| 5. Non-coding RNA & Extracellular Vesicle Signaling | Roles of EV cargo (miRNAs, lncRNAs) in regulating neurogenesis under inflammation are underexplored | Missed therapeutic opportunities; EVs may carry both detrimental and reparative signals | High-resolution EV profiling, CRISPR-based RNA manipulation, engineered EV delivery systems |
CNS, Central Nervous System; EV, Extracellular Vesicle; lncRNA, Long Non-Coding RNA; miRNA, MicroRNA; NLRP3, NOD-Like Receptor Pyrin Domain-Containing Protein 3; NSC, Neural Stem Cell; RNA-seq, RNA Sequencing.
4. Strategies and Emerging Approaches to Bridge the Gaps
Having outlined the critical knowledge gaps, this section discusses five key strategies to address these gaps and modulate the neuroimmune dialogue for therapeutic benefit. Each subsection corresponds to a specific strategy highlighted in the abstract (longitudinal neuroimmune imaging, niche-focused immunomodulation, glial subtype reprogramming, brain-penetrant NLRP3 inhibition, and CRISPR-based epigenetic editing). For each approach, I will describe the concept, provide examples of current research or tools, and discuss how it can help fill one or more of the gaps identified in Section 3. I will also comment on the feasibility and timeline: which strategies are nearer-term vs longer-term, and how they could be implemented in animal models or clinically.
Figure 3.
Neuroimmune interventions – from mechanism to therapy. Conceptual overview of five neuroimmune intervention strategies designed to preserve or rejuvenate adult neurogenesis, presented as modular schematic panels corresponding to Section 4.1–4.5: “Imaging,” “Immunomodulation,” “Glial Reprogramming,” “NLRP3 Inhibition,” and “Epigenetic Editing.” Each mini-diagram visually maps the mechanistic target to its therapeutic goal, such as longitudinal neuroimmune monitoring (Imaging), targeted suppression or rerouting of maladaptive immune signals (Immunomodulation), phenotype conversion or functional tuning of glial cells (Glial Reprogramming), blockade of inflammasome-driven IL-1β signaling (NLRP3 Inhibition), and locus-specific chromatin regulation or transcriptional correction (Epigenetic Editing). Collectively, this schematic bridges mechanistic insights with actionable therapeutic trajectories, offering a visual roadmap for designing next-generation neuroimmune interventions. IL-1β, interleukin-1 beta; NLPR3, NLR family pyrin domain containing 3.
Figure 3.
Neuroimmune interventions – from mechanism to therapy. Conceptual overview of five neuroimmune intervention strategies designed to preserve or rejuvenate adult neurogenesis, presented as modular schematic panels corresponding to Section 4.1–4.5: “Imaging,” “Immunomodulation,” “Glial Reprogramming,” “NLRP3 Inhibition,” and “Epigenetic Editing.” Each mini-diagram visually maps the mechanistic target to its therapeutic goal, such as longitudinal neuroimmune monitoring (Imaging), targeted suppression or rerouting of maladaptive immune signals (Immunomodulation), phenotype conversion or functional tuning of glial cells (Glial Reprogramming), blockade of inflammasome-driven IL-1β signaling (NLRP3 Inhibition), and locus-specific chromatin regulation or transcriptional correction (Epigenetic Editing). Collectively, this schematic bridges mechanistic insights with actionable therapeutic trajectories, offering a visual roadmap for designing next-generation neuroimmune interventions. IL-1β, interleukin-1 beta; NLPR3, NLR family pyrin domain containing 3.
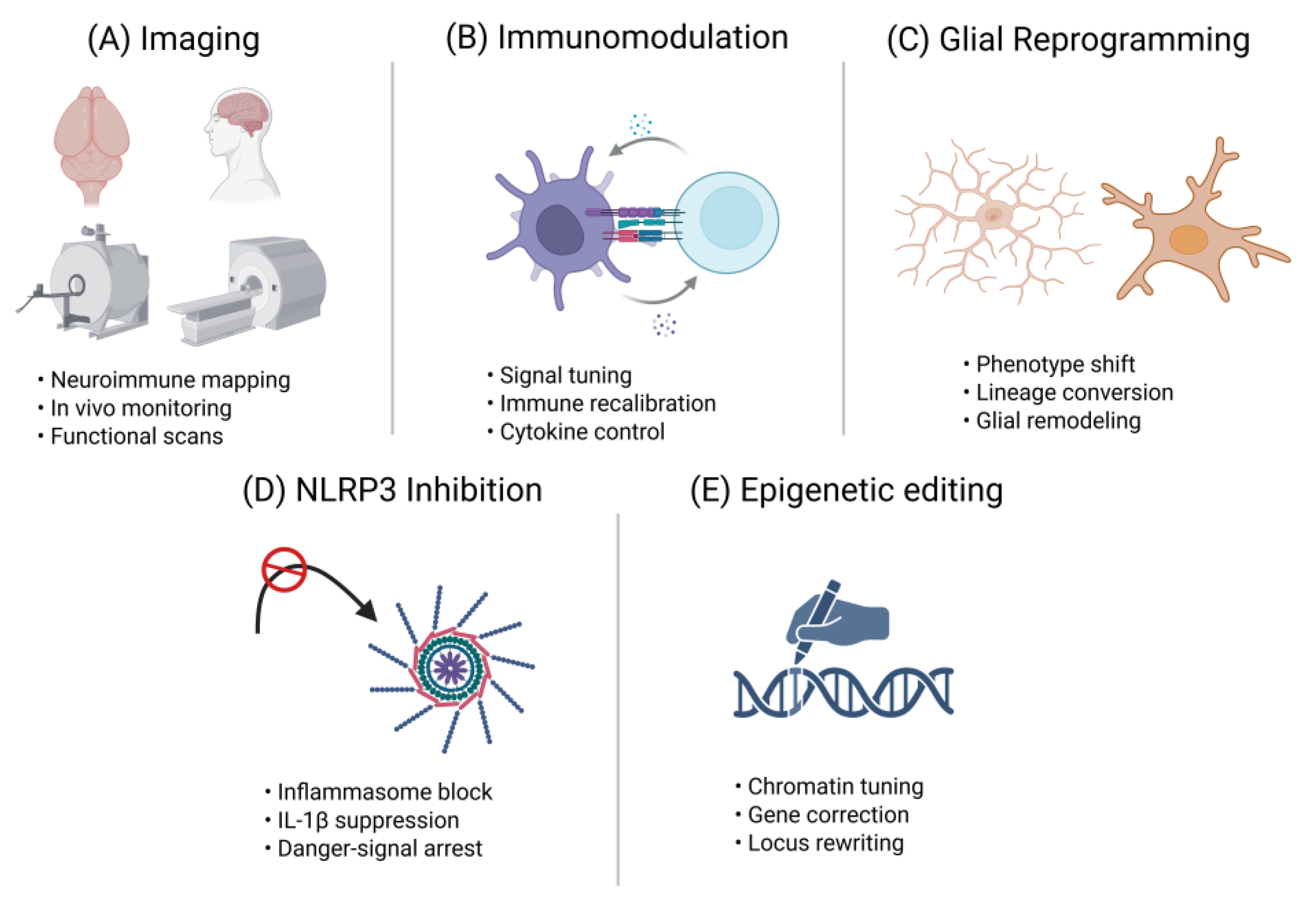
4.1. Longitudinal Neuroimmune Imaging
Longitudinal neuroimmune imaging is emerging as a transformative approach to address critical gaps in our understanding of brain aging [199]. It directly tackles Gap 3 by moving beyond cross-sectional “snapshot” methods, enabling dynamic monitoring of neurogenesis and neuroinflammation over time within the same subject [200]. Such temporal resolution can reveal whether surges in microglial activation precede, coincide with, or follow changes in neural stem cell activity [199,201]. Equally important, this strategy advances Gap 5 by providing non-invasive, cross-species applications through PET, magnetic resonance imaging (MRI), and two-photon microscopy [202]. By linking mechanistic insights from animal models to clinically relevant biomarkers in humans, it strengthens translational bridges [202,203].
Two-photon microscopy has revolutionized animal neuroimaging by enabling real-time, longitudinal observation of microglia–neuron interactions in the hippocampal neurogenic niche [204]. Using chronic cranial windows, researchers can track the same cells over weeks, revealing processes such as synaptic pruning, phagocytosis, and modulation of neural stem cell activity [204,205]. Fluorescent reporters and genetic labeling strategies further enhance cellular specificity, allowing precise mapping of immune–neural interactions [206]. These methods provide unmatched mechanistic insight into how inflammation and neurogenesis co-evolve, yet they remain invasive, limited to small animals, and restricted in imaging depth [207]. Despite these constraints, animal imaging offers critical proof-of-concept data that inspire translational strategies in humans [207,208].
Positron emission tomography and magnetic resonance imaging have become indispensable for longitudinal neuroimmune imaging, spanning both preclinical and clinical domains [209]. Translocator protein 18 kDa (TSPO)-PET is the most established approach for visualizing activated microglia, yet interpretation is hampered by low specificity, multicellular expression, and genetic polymorphisms that affect ligand binding [210]. To overcome these limitations, experimental PET tracers such as [^18F]FLT have been explored for labeling proliferating cells, with proof-of-concept studies showing that inhibiting tracer efflux enables detection of neurogenesis in vivo [209,211]. MRI provides a crucial complement, offering high-resolution structural measures such as hippocampal atrophy and functional connectivity readouts [212]. Together, PET and MRI form a translational bridge, linking cellular-level processes to human biomarkers and paving the way for therapeutic monitoring [213,214,215,216].
Next-generation imaging strategies are reshaping how we study neuroimmune dynamics. New PET tracers are being designed to distinguish between pro-inflammatory and anti-inflammatory microglial states, with promising targets such as P2X7R and P2Y12R offering phenotype-specific resolution [217,218]. Other tracers aim to directly label neurogenic processes or capture early astrocytic responses [217,219,220]. Hybrid modalities like PET/MR and dual-modal probes enhance spatial and molecular precision, while computational approaches including radiomics and machine learning refine signal interpretation and predict outcomes [221]. Integrating imaging readouts with peripheral biomarkers from blood or cerebrospinal fluid (CSF) promises a multimodal framework that could accelerate translation toward clinically actionable neuroimmune biomarkers [222].
The future of longitudinal neuroimmune imaging hinges on overcoming key technical and conceptual barriers [174]. New PET tracers are being developed to distinguish pro- and anti-inflammatory microglial phenotypes or to directly visualize neurogenic processes, promising greater specificity than TSPO-based tools [223]. Hybrid modalities such as PET/MRI enable integration of molecular and structural data, while computational approaches including radiomics and machine learning refine interpretation and enhance predictive power [224,225]. Pairing imaging with peripheral biomarkers from blood or CSF offers a multimodal strategy that could transform neuroimmune profiling, but achieving reliable, clinically translatable applications will require coordinated innovation and rigorous cross-species validation [174,226].
4.2. Niche-Focused Immunomodulation
Niche-focused immunomodulation refers to strategies that directly target the immune microenvironment of neurogenic regions such as the hippocampal DG and the SVZ zone, offering a sharp contrast to broad systemic immunosuppression [227]. These niches are not only central to sustaining neurogenesis but also uniquely accessible for precision therapies [227]. Localized approaches include intranasal delivery of cytokines that preferentially concentrate in ventricular areas, biomaterials or hydrogels engineered for sustained release of modulators, and blood–brain barrier-permeable compounds that accumulate within neurogenic zones [228]. Such strategies have shown that tailoring immune signals at the site of neural stem cell activity can stimulate neurogenesis while minimizing systemic risks [227]. By focusing interventions where they are most needed, niche-targeted approaches provide a rational and clinically appealing pathway to restore or preserve brain plasticity in aging and disease [227,229].
Aged and inflamed neurogenic niches often recruit CD8+ T cells and monocytes that secrete interferon-γ and other inhibitory factors, directly suppressing stem cell proliferation and neurogenesis [118]. Neutralizing these detrimental influences has emerged as a promising strategy to protect niche integrity [230]. Approaches include preventing immune cell entry with antibodies targeting adhesion molecules like CD44, or blunting their effects through cytokine neutralizers such as IL-8 blockade. Experimental work shows that anti-inflammatory agents, including indomethacin and minocycline, can preserve hippocampal neurogenesis during inflammatory insults, underscoring the therapeutic potential of immune blockade [231]. While systemic immunosuppression risks broad deficits, restricting these strategies to the niche could selectively alleviate inhibitory signaling without impairing host defenses [118]. Such focused modulation offers a rational pathway to counter age- and disease-associated immune pressures while preserving the regenerative capacity of the brain [118].
Enhancing pro-neurogenic immune signals represents a complementary strategy to blocking detrimental drivers, aiming instead to amplify reparative pathways within neurogenic niches [122]. Skewing microglia toward an “M2-like” phenotype through IL-4, IL-13, TGF-β mimetics, or nanomaterial-based modulators has shown proof-of-concept benefits, improving neurogenesis in models of aging, injury, and neurodegeneration [232]. Beyond immune skewing, engineered astrocytes or transplanted neural stem cells can be programmed to secrete protective cytokines and trophic factors, creating self-sustaining pro-regenerative feedback loops [233]. Biomaterials and hydrogels further extend these approaches by providing sustained, localized release of immune modulators within the hippocampus or SVZ [234]. Such strategies capitalize on the unique accessibility of neurogenic zones to reprogram niche immunity from within [122]. By strengthening protective cues locally, rather than relying on systemic administration, therapies may more effectively counter the age-related decline of neurogenesis while minimizing adverse immune suppression [122].
Implementing niche-focused immunomodulation requires overcoming significant technical hurdles but also opens remarkable translational opportunities [186]. Precision delivery systems such as focused ultrasound can transiently open the blood–brain barrier in hippocampal or SVZ regions, enabling local administration of gene vectors, cytokines, or antibodies with reduced systemic spillover [235]. Engineered stem cell grafts or EVs offer additional routes to sustain protective immunomodulation directly within the niche, while gene therapy vectors can be tailored for long-term expression of pro-neurogenic signals [236]. Challenges remain, including achieving delivery accuracy, sustaining therapeutic effects, and accounting for heterogeneity across neurogenic zones [186]. These considerations tie closely to Gap 1, highlighting regional microglial diversity, and Gap 5, emphasizing cross-species disconnects that complicate translation [186]. By integrating innovative tools with human-relevant models, niche-focused immunomodulation emerges as a therapeutic bridge, transforming mechanistic insights into precision strategies to rejuvenate neurogenesis in aging brains [186].
4.3. Glial Subtype Reprogramming
Glial subtype reprogramming represents a bold therapeutic paradigm in which resident glia are reshaped to foster neural repair and adult neurogenesis [237]. Two main strategies define this field. Phenotypic reprogramming focuses on restoring dysfunctional microglia or astrocytes to neuroprotective states, for example by inhibiting inflammatory cascades or promoting cross-talk that favors protective cytokine release [238]. Lineage reprogramming goes further, converting astrocytes into neurons or neural progenitors through transcription factors such as NeuroD1, DLX2, or Neurog2, or with small-molecule cocktails capable of inducing neuronal fates in vivo [239]. Together, these approaches seek to remodel the neurogenic niche, counteract age-related decline, and generate new avenues for brain rejuvenation rooted in cellular plasticity [240].
Phenotypic reprogramming of microglia has emerged as a compelling strategy to restore neurogenic potential within the hippocampal niche [241]. Central to this approach is the inhibition of NF-κB and related inflammatory cascades, which drive the release of cytokines that suppress neural stem cell proliferation [242]. Pharmacological interventions such as indole derivatives, natural compounds like mangiferin or costunolide, and small molecules targeting PI3K–Akt or Nrf2 signaling have successfully reduced pro-inflammatory activity while promoting M2-like polarization [108]. Similarly, exosome-based delivery of miR-124 or growth factors such as FGF1 rejuvenated microglial transcriptomes and enhanced neurogenesis [243]. Experimental evidence demonstrates that these interventions not only dampen pathological inflammation but also improve hippocampal neurogenic output, leading to cognitive and behavioral recovery in stress, injury, and neurodegenerative models [241].
Lineage reprogramming of astrocytes has revealed an extraordinary potential to regenerate neurons within damaged or aged brains [244]. Breakthrough studies demonstrate that transcription factors such as NeuroD1, DLX2, or Neurogenin-2 can directly convert reactive astrocytes into functional neurons in vivo, with newly generated cells integrating into local circuits and restoring behavioral function after injury or stroke [239]. Complementary approaches employ cocktails of small molecules to reprogram astrocytes into neurons or progenitor-like cells without viral vectors, offering a more clinically attractive route [245]. Both genetic and chemical strategies have produced proof-of-concept evidence that even reactive astrocytes in diseased or inflamed contexts can be redirected toward a neuronal fate [246]. These advances suggest that lineage reprogramming may one day augment or replace lost neurogenesis, transforming astrocytes into reservoirs of neuronal replacement [247].
Glial subtype reprogramming provides a bold strategy to tackle Gap 1, the challenge of regional microglial diversity, and Gap 2, the influence of inflammasome-driven epigenetic alterations [30]. By restoring homeostatic or neuroprotective microglial states, interventions counteract inhibitory cytokine cascades that suppress neural stem cells [248]. At the same time, lineage reprogramming of astrocytes into neurons or progenitors enlarges the neurogenic reservoir, directly compensating for age-related decline [240]. This dual action both mitigates maladaptive immune signaling and boosts neuronal output, reframing glia not as barriers but as therapeutic substrates for rejuvenating neurogenic niches [249].
Glial subtype reprogramming faces formidable but surmountable translational challenges [249]. Precision of delivery remains paramount, as viral vectors and gene editing tools pose risks of off-target effects, immune responses, and uncontrolled proliferation [250]. The heterogeneity of niches adds further complexity, demanding context-sensitive strategies rather than blanket interventions [251]. Innovative technologies such as CRISPR-based regulation, hydrogel-rationed delivery systems, and inducible gene circuits offer avenues for safer, more controlled reprogramming [252]. Ultimately, this approach represents a bold therapeutic frontier: the potential to “rewrite” the aging brain’s fate by generating neurons from glia and restoring supportive immune states within neurogenic niches [237].
4.4. Brain-Penetrant NLRP3 Inflammasome Inhibitors
The NLRP3 inflammasome has emerged as a central orchestrator of chronic neuroinflammation, with microglial activation driving sustained release of IL-1β that disrupts neural stem cell function and impairs neurogenesis [253]. While systemic inhibition of this pathway shows anti-inflammatory promise, the distinct challenge in brain disorders lies in achieving effective suppression within the central nervous system (CNS)[254]. Small-molecule inhibitors capable of crossing the blood–brain barrier, such as nlrp3 inflammasome inhibitor mcc950 (MCC950) and newer candidates like NT-0796 or ASP0965, represent a breakthrough class [254]. By directly targeting microglial inflammasome activity in situ, these compounds address age- and disease-related priming of neuroinflammation that perpetuates cognitive decline and accelerates neurogenic failure [255].
MCC950 has served as the prototypical NLRP3 inhibitor, demonstrating consistent ability to cross the blood–brain barrier and attenuate microglial activation across diverse models of stress, injury, and neurodegeneration [256]. By suppressing caspase-1 activation and IL-1β release, it preserves neural progenitor proliferation and mitigates cognitive decline in contexts such as AD, stroke, traumatic brain injury (TBI), and depression-like states [257]. Building on this foundation, newer derivatives such as NP3-253 and novel bicyclic scaffolds have been designed for improved CNS penetration, stability, and potency [258]. Preclinical studies with these compounds confirm that inflammasome inhibition protects hippocampal neurogenesis, underscoring the therapeutic promise of this mechanistically targeted approach [259].
New brain-penetrant NLRP3 inhibitors such as NT-0796 and BGE-102 are advancing into early clinical trials, marking a pivotal step in translating inflammasome biology into therapy [254]. These compounds demonstrate robust CNS exposure and have been shown to lower neuroinflammatory biomarkers in humans, offering a promising route to intervene in age-related cognitive decline and mild cognitive impairment [254,260]. By disrupting IL-1β–driven feedback loops, they directly address Gap 2, mitigating inflammasome-driven epigenetic alterations that lock microglia into pro-inflammatory states [261]. At the same time, their clinical development speaks to Gap 5, bridging preclinical insights with druggable, human-relevant strategies aimed at rejuvenating neurogenic niches [262].
Chronic NLRP3 activation in microglia not only drives IL-1β release but also imprints maladaptive epigenetic programs, including hypomethylation of inflammatory promoters that sustain reactivity. Such “trained” states perpetuate neurotoxic signaling, impair neurogenesis, and foster astrocytic dysfunction [263]. Inhibitors like MCC950 and next-generation brain-penetrant compounds can disrupt this loop, dampening acute cytokine production while gradually reprogramming microglial memory toward a less inflammatory phenotype [264]. This mechanistic depth extends their value beyond transient blockade, suggesting that inflammasome inhibition may restore a supportive niche by stabilizing microglial identity and relieving epigenetic brakes on neurogenesis [264].
While brain-penetrant NLRP3 inhibitors hold strong therapeutic promise, challenges remain in balancing efficacy with safety [265]. Risks include off-target immunosuppression, uncertain timing of intervention, and limited knowledge of long-term effects [265]. Refining specificity through next-generation scaffolds, selective inflammasome modulators, and combinatorial, biomarker-guided strategies could mitigate these concerns [266]. Ultimately, NLRP3 inhibition represents one of the most tangible near-term pharmacological routes to rejuvenating the neurogenic niche, translating mechanistic insights on inflammasome-driven pathology into clinically actionable therapies for aging and neurodegeneration [267].
4.5. CRISPR-Based Epigenetic Editing
CRISPR-based epigenetic editing harnesses catalytically inactive Cas9 (dCas9) fused to effector domains that alter chromatin or DNA methylation, enabling locus-specific regulation of gene expression without introducing double-strand breaks [268]. This distinguishes it from conventional genome editing by allowing reversible, non-mutagenic interventions [268]. For example, dCas9-DNMT3A or DNMT3A/3L fusions can deposit methylation at promoters to silence inflammatory genes, whereas dCas9-TET1 can induce targeted demethylation to reactivate silenced loci such as Oct4 or Fgf21 [269]. Additional configurations, including KRAB- or Ezh2-dCas9 fusions, deposit repressive histone marks, while SunTag-TET systems amplify demethylase recruitment for strong activation [270]. Together, these tools offer unprecedented precision in modulating immune and neurogenic pathways at the epigenetic level [268].
CRISPR-based epigenetic editing directly addresses Gap 2 by enabling the reversal of maladaptive methylation states in neural and immune cells shaped by aging or chronic inflammation [269]. For example, targeting dCas9-DNMT3A to the IL1β promoter in aged microglia could restore silencing through re-methylation, thereby reducing chronic inflammatory drive [271]. Conversely, dCas9-TET1 applied to neurogenic loci such as BDNF or Oct4 can relieve age-induced repression and reactivate transcription, reinstating neurogenic potential in stem cells [269]. Proof-of-concept studies with Yamanaka factors or partial reprogramming confirm that rejuvenating epigenetic marks restores neurogenesis and cognitive capacity in aged niches [240]. Unlike transient cytokine blockade, this strategy reprograms cellular memory itself, offering durable restoration of youthful transcriptional states and opening new avenues for neuroregenerative therapy [272].
Preclinical studies highlight CRISPR-based epigenetic editing as a versatile platform to reshape neuronal and immune gene expression without introducing DNA breaks [273]. In tauopathy models, dCas9-p300 activation of Gad1 restored synaptic inhibition and cognition, while targeted methylation of the APP promoter in Alzheimer’s mice reduced amyloid pathology and memory decline. CRISPRoff approaches have even created heritable transcriptional memory, demonstrating sustained regulation across divisions [273]. In immune cells, epigenetic reprogramming stabilized lineage-specific expression, underscoring durability [274]. Hypothetically, maintaining neurotrophin expression in aged neural stem cells or silencing astrocytic inflammatory mediators could rejuvenate neurogenic niches [275]. By enabling precise, durable, and programmable control of maladaptive states, CRISPR epigenetic editing directly addresses Gap 5, offering a forward-looking strategy to translate mechanistic insight into therapies for neurodegeneration and cognitive decline [273].
Translating CRISPR-based epigenetic editing into the brain faces formidable challenges, with delivery standing as the most immediate hurdle [276]. Viral vectors such as adeno-associated virus (AAV)s provide durable expression but risk insertional mutagenesis and immunogenicity, while nonviral platforms like nanoparticles, nanocapsules, and engineered peptide coatings promise safer, localized delivery yet remain under development [276]. Equally pressing is the need to ensure locus specificity, as off-target chromatin remodeling could introduce unpredictable, long-lasting effects [277]. Despite these risks, incremental advances in vector design and precision editing suggest that durable, brain-targeted interventions are attainable [276]. In the long term, CRISPR epigenetic editing may become a transformative therapeutic modality, capable of permanently resetting maladaptive cellular states, rejuvenating neural stem cell potential, and sustaining neurogenesis well into aging [278].
Table 3.
Emerging therapeutic strategies targeting the neuroimmune axis. Overview of emerging therapeutic strategies aimed at modulating the neuroimmune axis to protect or restore adult neurogenesis, outlining representative tools, intended biological effects, and current translational maturity. Columns include: Strategy, Examples / Tools, Goal / Effect, and Stage of Development. Representative entries may feature longitudinal neuroimmune imaging modalities (e.g., [^18F]FLT PET, TSPO-PET), brain-penetrant NLRP3 inflammasome inhibitors (e.g., MCC950, NT-0796) [279] , in vivo glial reprogramming vectors (e.g., AAV-NeuroD1), precision epigenetic gene editing platforms (e.g., CRISPR-dCas9), and niche-targeted immunomodulatory therapeutics. This table offers a rapid translational snapshot for investigators evaluating feasibility, clinical readiness, and mechanistic alignment across intervention classes.
Table 3.
Emerging therapeutic strategies targeting the neuroimmune axis. Overview of emerging therapeutic strategies aimed at modulating the neuroimmune axis to protect or restore adult neurogenesis, outlining representative tools, intended biological effects, and current translational maturity. Columns include: Strategy, Examples / Tools, Goal / Effect, and Stage of Development. Representative entries may feature longitudinal neuroimmune imaging modalities (e.g., [^18F]FLT PET, TSPO-PET), brain-penetrant NLRP3 inflammasome inhibitors (e.g., MCC950, NT-0796) [279] , in vivo glial reprogramming vectors (e.g., AAV-NeuroD1), precision epigenetic gene editing platforms (e.g., CRISPR-dCas9), and niche-targeted immunomodulatory therapeutics. This table offers a rapid translational snapshot for investigators evaluating feasibility, clinical readiness, and mechanistic alignment across intervention classes.
| Strategy | Examples / Tools | Goal / Effect | Stage of Development |
| Longitudinal Imaging | [^18F]FLT-PET for neurogenesis, TSPO-PET for microglial activation | Enables in vivo monitoring of neurogenesis and neuroinflammation across lifespan | Preclinical for neurogenesis tracers; TSPO-PET in human use |
| Brain-Penetrant NLRP3 Inhibitors | MCC950, NT-0796, BGE-102 | Reduce chronic IL-1β release, restore neurogenic potential | Preclinical to Phase 1 clinical trials |
| Glial Reprogramming | AAV-NeuroD1, SOX2-based astrocyte-to-neuron conversion | Replace lost neurons; rejuvenate circuits | Proof-of-concept in rodents |
| CRISPR Epigenetic Editing | CRISPR-dCas9 targeting IL-1β/NLRP3 loci; enhancer repression | Long-term silencing of pro-inflammatory genes without DNA cleavage | Lab-stage; in vitro and early in vivo |
| Niche Immunomodulation | Anti-IL-1β, anti-TNF, IL-6R antibodies; microglia-specific modulators | Dampens chronic inflammation in neurogenic niches | Several agents in AD, MCI, depression trials |
| Extracellular Vesicle (EV) Therapeutics | Engineered EVs carrying miRNAs, BDNF, or IGF-1 cargo | Deliver pro-neurogenic and anti-inflammatory signals | Preclinical; first-in-human safety studies emerging |
| Lifestyle & Activity-Based Interventions | Exercise, enriched environment, caloric modulation | Boost endogenous IGF-1/BDNF, reduce inflammatory priming | Multiple human cohort studies and ongoing clinical trials |
| Small-Molecule Neurotrophic Enhancers | TrkB agonists, phosphodiesterase inhibitors | Enhance BDNF signaling, promote synaptic/neurogenic resilience | Early-stage clinical testing, mixed outcomes |
| Microglial State Modulation | CSF1R inhibitors, TREM2 agonists | Shift microglia from pro-inflammatory to reparative states | Preclinical; TREM2 antibodies in Phase 2 AD trials |
| Combinatorial Approaches | NLRP3 inhibitor + exercise; anti-TNF + BDNF mimetics | Target multiple axes (inflammatory and trophic) simultaneously | Conceptual and early preclinical testing |
AAV, Adeno-Associated Virus; AD, Alzheimer’s Disease; BDNF, Brain-Derived Neurotrophic Factor; CSF1R, Colony-Stimulating Factor 1 Receptor; CRISPR, Clustered Regularly Interspaced Short Palindromic Repeats; dCas9, Deactivated CRISPR-Associated Protein 9; EV, Extracellular Vesicle; [^18F]FLT, Fluorothymidine Labeled With Fluorine-18; IGF-1, Insulin-Like Growth Factor-1; IL-1β, Interleukin-1 Beta; IL-6R, Interleukin-6 Receptor; MCI, Mild Cognitive Impairment; miRNA, MicroRNA; NLRP3, NOD-Like Receptor Pyrin Domain-Containing Protein 3; PET, Positron Emission Tomography; SOX2, SRY-Box Transcription Factor 2; TNF, Tumor Necrosis Factor; TrkB, Tropomyosin Receptor Kinase B; TSPO, Translocator Protein 18kDa.
5. Comparative Perspectives: Human vs. Animal Models
Understanding how rodent and human data align—or diverge—is essential for evaluating the translational relevance of neurogenesis and neuroinflammation research [89]. Animal models provide mechanistic precision, offering evidence for persistent but declining neurogenic activity and for microglial shifts that shape brain plasticity across the lifespan [190]. Human studies, however, reveal greater uncertainty, complicated by methodological variability and ethical constraints [280]. By contrasting these perspectives, we can identify both the strengths and limitations of each approach, setting the stage for a closer examination of adult hippocampal neurogenesis across species [89].
5.1. Adult Neurogenesis: Rodents vs. Humans
Adult rodent studies have firmly established that hippocampal neurogenesis is robust in youth and declines with age, yet it never disappears entirely [76]. Bromodeoxyuridine labeling and lineage tracing demonstrate that new granule cells continue to be generated in the DG, although proliferation rates drop dramatically with aging, from nearly three percent of granule cells in young adults to less than half a percent in old animals [281]. Even under stressors such as ischemia or stroke, aged rodents retain the capacity for injury-induced neurogenesis, with locally activated neural stem and progenitor cells emerging in peri-infarct regions, although their efficiency and neuronal differentiation are reduced [282]. Complementary observations in human stroke and ischemic pathology indicate that neurogenic progenitors and stem-like cells can also be mobilized within or adjacent to injured areas, suggesting that endogenous stem cell pools remain at least partly reprogrammable and capable of contributing to structural repair, even in the aged brain [279,281]. These findings confirm a persistent, though diminished, neurogenic reservoir across the lifespan [76].
In humans, evidence for adult hippocampal neurogenesis remains strikingly divided [283]. Boldrini and colleagues reported thousands of immature neurons persisting even in older adults, whereas Sorrells and collaborators argued that new neurons are virtually absent beyond childhood [284]. Much of this divergence stems from methodological factors: antigen preservation, fixation times, and tissue sampling critically determine whether markers like doublecortin (DCX) or polysialylated neural cell adhesion molecule (PSA-NCAM) are detectable [85]. Reviews emphasize that small differences in processing can yield opposite conclusions, making consensus elusive [88]. The debate continues, with most agreeing that technical rigor, standardized protocols, and multimodal approaches are essential to resolve this controversy [285].
5.2. Microglial States Across Species
Rodent studies have provided a detailed atlas of microglial aging, revealing consistent transcriptional and metabolic shifts that define an “inflammaging” signature [286]. Single-cell RNA sequencing across the mouse lifespan uncovers multiple microglial states, with aging marked by heightened chemokine expression and reprogramming of metabolic pathways [287]. A particularly striking feature is the emergence of lipid droplet–accumulating microglia, which display defective phagocytosis, exaggerated cytokine release, and altered lipid metabolism [11]. Proteomic and transcriptomic analyses further demonstrate reduced homeostatic signaling, increased glycolysis, and overlap with disease-associated microglia (DAM) [288]. Collectively, these findings establish aged rodent microglia as pro-inflammatory, metabolically reprogrammed, and primed for maladaptive responses to stress or injury [286].
Human microglia display both striking overlaps with rodents and distinct aging trajectories that underscore species divergence [289]. Transcriptomic studies reveal that while a conserved core program exists, humans show unique regulation of adhesion, cytoskeletal, and complement-related genes, alongside greater transcriptional heterogeneity with age [188]. Unlike rodents, aged human microglia often develop dystrophic morphologies and altered responses to neurodegeneration [95]. Yet shared features emerge: chronic systemic inflammation accelerates microglial aging across species, and both mice and humans exhibit increased T cell infiltration in the SVZ zone, reshaping the neurogenic niche [290]. These parallels and divergences highlight the importance of comparative perspectives for translational relevance.
5.3. Inflammatory Pathways and Neuroimmune Crosstalk
Rodent studies have revealed how inflammasome priming and glial crosstalk shape neurogenic outcomes in aging and disease [291]. Activation of the microglial NLRP3 inflammasome drives the conversion of astrocytes into a neurotoxic A1 state, suppressing neurogenesis and impairing cognition, while genetic deletion of Nlrp3 or treatment with inhibitors such as MCC950 restores function [292]. Similarly, interferon-gamma (IFN-γ)–primed microglia impair neural stem cell proliferation, an effect reversible by janus kinase/signal transducer and activator of transcription 1 (JAK/STAT1) blockade [138]. Tri-culture models confirm that microglia–astrocyte interactions amplify inflammatory cascades, while mitochondrial dysfunction further exaggerates NLRP3 activity [116]. These findings underscore how precisely manipulable rodent systems delineate pathways where inflammation curtails hippocampal neurogenesis.
In humans, evidence for inflammasome activation is largely indirect, derived from postmortem analyses, CSF biomarkers, and emerging imaging studies [293]. Elevated IL-1β, IL-18, and inflammasome proteins such as ASC and caspase-1 have been reported in neurodegenerative disease and TBI, often correlating with severity or outcome [294]. Immunohistochemistry shows co-localization of NLRP3 with glial markers in Alzheimer’s tissue, while iPSC-derived microglia link genetic risk factors to inflammasome priming [295]. Yet interpretation is complicated by timing, chronic disease progression, and comorbidities, making causal inference far less straightforward than in controlled rodent experiments [293].
5.4. Intervention Efficacy and Translational Readiness
Rodent studies provide strong evidence that lifestyle and experimental interventions can enhance neurogenesis and preserve cognition well into aging and disease [296,297]. Aerobic exercise reduces microglial inflammasome activity through irisin signaling, restoring hippocampal neurogenesis and memory in Parkinson’s models [264]. Environmental enrichment, with or without exercise, consistently improves learning and reduces anxiety-like behaviors, while also limiting aberrant neurogenesis after stroke [298]. Mechanistically, these effects arise from reduced inflammation, epigenetic reprogramming, and enhanced plasticity across hippocampal subregions [299]. Direct manipulations, such as BDNF overexpression or sodium lactate administration, reproduce the benefits of enrichment and exercise, underscoring their causal link to neurogenesis and cognitive resilience [300].
In humans, lifestyle interventions such as exercise, cognitive engagement, and diet consistently improve cognition and brain health, yet their link to neurogenesis remains indirect, inferred from changes in neuroplasticity and neurotrophic signaling rather than direct cellular evidence [301]. Clinical trials of non-steroidal anti-inflammatory drugs (NSAD) in AD have been disappointing, with large-scale meta-analyses showing no meaningful benefit and even highlighting adverse events [302]. This discrepancy with epidemiological associations underscores the complexity of timing and target specificity in human disease [303]. More selective approaches, particularly NLRP3 inflammasome inhibitors, represent an emerging avenue with stronger mechanistic rationale, but translation is still in its infancy, awaiting proof of efficacy and safety in controlled human trials [265].
5.5. Bridging the Gap: Models, Ethics, and Future Outlook (≈150–180 Words)
Rodent models permit invasive manipulations such as ablation, lineage tracing, and precise genetic editing, approaches that are fundamentally restricted in humans due to ethical and practical barriers [304]. Human studies instead rely on observational designs, neuroimaging, and pharmacological interventions, offering indirect but clinically relevant insights [305]. While nonhuman primates and organoid-based chimeras help bridge gaps, their use is also constrained by ethical scrutiny and feasibility [306,307]. Consequently, translational progress depends on integrating mechanistic detail from animal work with non-invasive, ethically sound human research strategies that refine, reduce, and replace invasive experimentation where possible [308].
Future progress will hinge on combining advanced imaging and circulating biomarkers with humanized models that integrate microglia into brain organoids [309]. Such systems allow dynamic visualization of neuron–glia crosstalk, capture human-specific inflammatory signatures, and provide a platform for testing therapeutic strategies [310]. Rodent models remain indispensable for dissecting mechanisms and enabling invasive manipulations that cannot be performed in humans [311]. Yet innovation must increasingly center on organoid-based and biomarker-driven approaches, ensuring translation captures the complexity of human neuroimmunity while retaining the mechanistic clarity offered by animal studies.
Figure 4.
Comparative Overview of Neurogenic Capacity in Rodents and Humans. Neurogenesis remains robust across the lifespan in rodents, accompanied by higher microglial density and phenotypic diversity, along with active subventricular zone (SVZ) neurogenic activity. In contrast, adult humans exhibit markedly reduced neurogenic output, diminished microglial diversity, and limited or absent SVZ-driven neuronal production. While rodents display an age-related tapering in neuronal survival, humans demonstrate minimal or near-absent levels. Preclinical interventions such as physical activity, anti-inflammatory modulation, imaging-guided strategies, targeted pharmacology, or gene-editing platforms show pronounced benefits in animal models, whereas human findings remain largely associative or under investigation. SVZ, subventricular zone.
Figure 4.
Comparative Overview of Neurogenic Capacity in Rodents and Humans. Neurogenesis remains robust across the lifespan in rodents, accompanied by higher microglial density and phenotypic diversity, along with active subventricular zone (SVZ) neurogenic activity. In contrast, adult humans exhibit markedly reduced neurogenic output, diminished microglial diversity, and limited or absent SVZ-driven neuronal production. While rodents display an age-related tapering in neuronal survival, humans demonstrate minimal or near-absent levels. Preclinical interventions such as physical activity, anti-inflammatory modulation, imaging-guided strategies, targeted pharmacology, or gene-editing platforms show pronounced benefits in animal models, whereas human findings remain largely associative or under investigation. SVZ, subventricular zone.
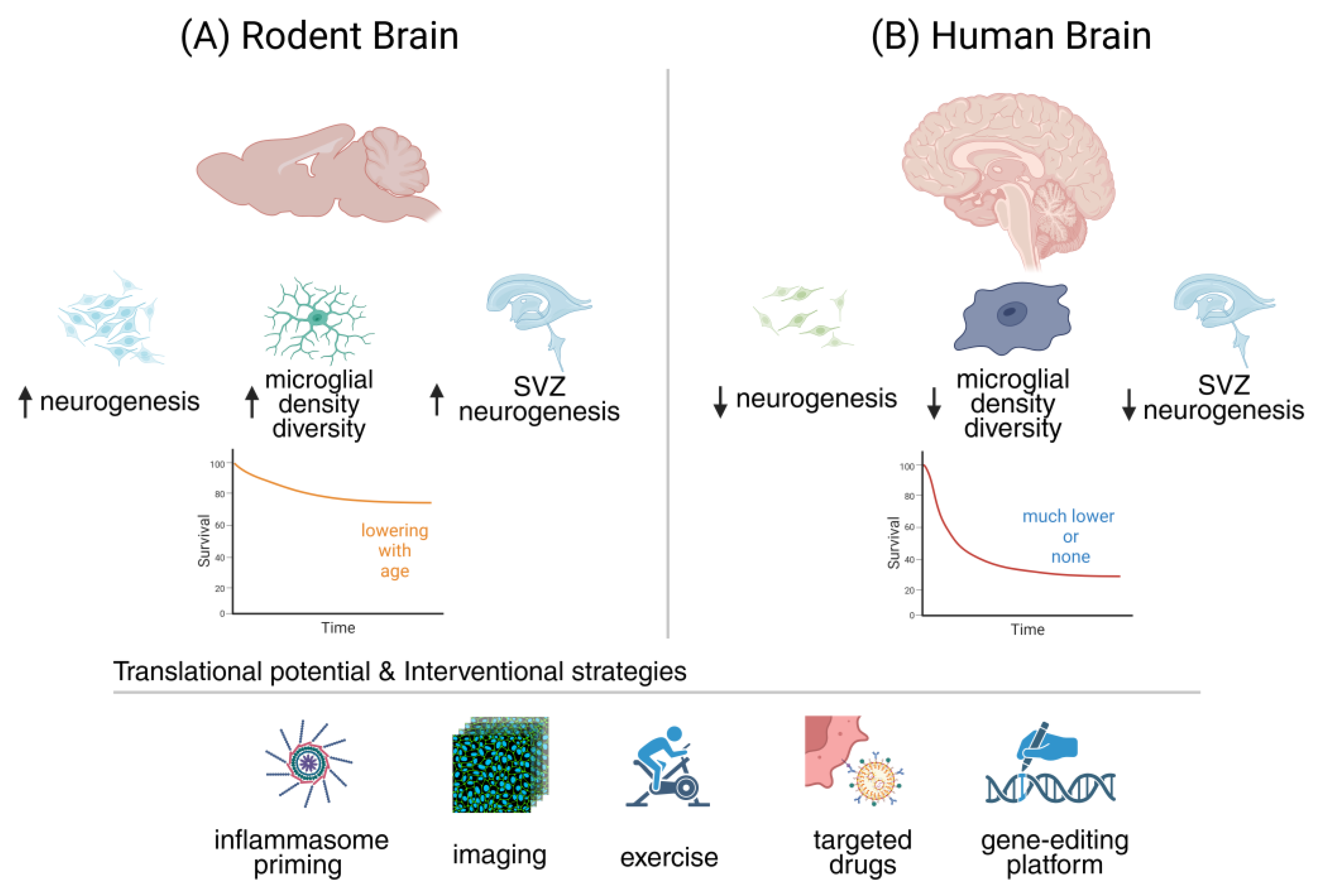
Table 4.
Comparative characteristics – neurogenesis and neuroinflammation in mice vs. humans. Side-by-side comparison of core biological, cellular, and aging-related features of adult hippocampal neurogenesis and neuroinflammatory remodeling in rodents versus humans. Columns include: Aspect, Rodents (Murine), and Humans. Representative comparison points may include baseline levels of adult neurogenesis, age-related rates of decline, microglial density and activation phenotypes across lifespan, peripheral immune cell infiltration into the central nervous system, and responsiveness to lifestyle-based interventions such as exercise or environmental enrichment. This table serves as a rapid translational reference, highlighting biological similarities, species-specific divergence, and areas where human data remain limited or method-dependent.
Table 4.
Comparative characteristics – neurogenesis and neuroinflammation in mice vs. humans. Side-by-side comparison of core biological, cellular, and aging-related features of adult hippocampal neurogenesis and neuroinflammatory remodeling in rodents versus humans. Columns include: Aspect, Rodents (Murine), and Humans. Representative comparison points may include baseline levels of adult neurogenesis, age-related rates of decline, microglial density and activation phenotypes across lifespan, peripheral immune cell infiltration into the central nervous system, and responsiveness to lifestyle-based interventions such as exercise or environmental enrichment. This table serves as a rapid translational reference, highlighting biological similarities, species-specific divergence, and areas where human data remain limited or method-dependent.
| Aspect | Rodents (Murine) | Humans |
| Adult hippocampal neurogenesis (baseline) | Thousands of new neurons per day in young adult hippocampus; robust measurable pools | Far fewer (hundreds/day in young adults by some estimates); highly variable depending on methodology |
| Age of significant decline in neurogenesis |
Detectable decline starting mid-life (12–18 months); still measurable in aged animals | Steep decline reported from middle age; ongoing debate whether residual neurogenesis persists in elderly |
| Microglial density and activation state in aging | Well-characterized shift to 'primed' phenotype with pro-inflammatory gene expression and reduced phagocytic resolution | Less comprehensive; aged human microglia show pro-inflammatory signatures, distinct subsets identified via single-cell transcriptomics |
| Peripheral immune cell involvement in CNS with age | Increased infiltration of T cells (especially CD8⁺) into hippocampus and SVZ with aging; enhances IFN-γ tone | Limited but growing evidence; T-cell presence in human hippocampus in aging and neurodegeneration; mechanisms less defined |
| Evidence for exercise or enrichment effects | Exercise and enriched environments robustly increase neurogenesis and improve cognition in mice | Human studies show hippocampal volume increases and cognitive benefits; direct evidence for neurogenesis boost is indirect (MRI, blood biomarkers) |
| Inflammasome/NLRP3 activation with age | Strong evidence for NLRP3-driven IL-1β increase in aged rodent hippocampus, reducing neurogenesis | Human post-mortem and transcriptomic studies support NLRP3 upregulation in aging brain; functional causality harder to confirm |
| Translational caveats | High plasticity, short lifespan, and controlled environments amplify experimental effects | Human variability, long lifespan, and heterogeneous exposures complicate translation; methodological debates on detecting neurogenesis |
CD8⁺, Cluster of Differentiation 8-Positive T Cell; CNS, Central Nervous System; IFN-γ, Interferon-Gamma; IL-1β, Interleukin-1 Beta; MRI, Magnetic Resonance Imaging; NLRP3, NOD-Like Receptor Pyrin Domain-Containing Protein 3; SVZ, Subventricular Zone.
6. Integrating Mechanisms with Therapeutics: Toward Rewiring the Aging Brain
The convergence of mechanistic insight with therapeutic innovation marks a critical frontier in efforts to reshape the course of brain aging [312,313]. Rather than viewing neurogenesis decline and neuroinflammation as inevitable hallmarks of senescence, emerging work reveals them as modifiable processes that can be recalibrated through precise interventions [21]. By aligning detailed knowledge of inflammasome signaling, microglial states, and epigenetic regulation with translational tools such as advanced imaging, targeted drugs, and gene-editing platforms, we begin to chart a roadmap for reprogramming resilience. This section considers how these once-disparate strategies may synergize, offering realistic short-term goals and bold long-term visions for delaying or even reversing cognitive decline [314,315].
6.1. Mechanistic Gaps as Opportunities (~400 words)
Gaps in translation should not be seen as obstacles but as navigational markers guiding innovation [316]. Comparative work between animal models and humans consistently reveals mismatches in pathology, timing, and response, yet these very mismatches highlight where new discoveries can be made [317]. Limitations in rodents have prompted the development of humanized organoids, large animal models, and network-based approaches that better capture human complexity [318]. In this way, every identified gap points to a therapeutic or conceptual opportunity, and the strategies that follow can be viewed as direct responses to these translational signposts [319].
Microglial heterogeneity (Gap 1) offers more than complexity; it provides a framework for designing region-specific and state-selective interventions [41]. Rodent studies have mapped diverse transcriptional states across age and pathology, while human transcriptomics confirm that these subtypes exist but follow distinct trajectories, guiding translational priorities [166,320]. Parallel to this, inflammasome-driven epigenetic regulation (Gap 2) emerges as a fertile ground for therapeutic innovation. NLRP3 inhibitors (Strategy 4.4) and CRISPR-based epigenetic editing (Strategy 4.5) exemplify approaches that can reset maladaptive inflammatory programs [265,321]. Thus, what appears as translational limitation is simultaneously the rationale for precision interventions that reprogram cellular states in aging brain niches [249].
Gap 3 highlights systemic influences as a critical axis where rodent and human data converge [322,323]. Parabiosis studies and controlled inflammatory challenges in mice demonstrate the causal power of circulating factors to accelerate or reverse brain aging, while in humans these influences are tracked indirectly through biomarkers, immune profiling, and neuroimaging [324]. Gap 4 reframes modeling constraints as an impetus for innovation, driving the creation of humanized organoids with microglia or vascular integration [325]. Together, these approaches shrink the translational distance, allowing rodent mechanistic insights to be anchored in human relevance and opening new opportunities for therapeutic discovery [326,327].
Gap 5 represents the pivot from mechanism to medicine, where insights from animal models begin to shape clinical opportunity [319,328]. Rodent studies demonstrate that targeting the NLRP3 inflammasome, boosting neurotrophic signaling, or harnessing epigenetic editing can restore neurogenesis and rescue cognition, inspiring translational efforts such as NLRP3 inhibitors now advancing toward human trials [329]. Lifestyle-based interventions likewise show convergent benefits, even if mechanisms differ across species [330]. Comparative perspectives remind us of limits but also highlight optimism: each gap becomes a guidepost, pointing directly to the therapeutic strategies most likely to rejuvenate neurogenesis and combat cognitive decline [331].
6.2. Translational Roadmap
Longitudinal imaging stands at the forefront of translational progress, offering a way to monitor neuroinflammation and neurogenesis non-invasively across time [174]. Advances in PET tracers, from second-generation TSPO ligands to newer targets such as COX and P2X7 receptors, promise higher specificity and functional insight [175]. Complementary multiparametric MRI approaches add spatial and physiological context, enabling integration with PET for richer biomarker panels [222]. Yet, robust clinical translation requires rigorous standardization, multicenter reproducibility, and careful correlation of imaging signals with cognitive outcomes [332]. Establishing validated tracers and harmonized protocols will turn imaging into a reliable bridge between mechanistic insight and therapeutic monitoring [333].
Rodent studies have shown that intranasal delivery of EVs, nanoparticles, or viral vectors can modulate microglial activation and promote neuroprotection, and the challenge now lies in scaling these strategies to non-human primates [334]. Advances in engineered AAVs and synthetic promoters already enable selective targeting of glial populations, while refined capsid variants reduce peripheral exposure and off-target effects [335]. Non-viral systems, such as EVs and lipid nanoparticles, add further flexibility and safety [336]. Critical milestones include demonstrating long-term safety, reproducibility, and circuit specificity, ensuring that reprogramming interventions translate effectively into clinically viable therapies [337,338].
NLRP3 inhibitors are moving from preclinical promise to clinical evaluation, with trials in AD now focusing on safety, dose optimization, and early cognitive outcomes as essential milestones [267]. Small molecules such as MCC950, OLT1177, and JC124, alongside emerging biologics, have consistently reduced neuroinflammation and improved cognition in rodent models, and several are progressing into human testing [259,329,339]. Their success will hinge on demonstrating blood–brain barrier penetration, tolerability, and biomarker validation [262,267]. Looking ahead, combinatorial strategies pairing inflammasome inhibition with lifestyle or behavioral interventions may enhance efficacy and broaden therapeutic relevance [262].
CRISPR-based epigenetic editing is advancing rapidly in preclinical research, offering the unprecedented possibility of rewriting maladaptive molecular memory at specific genomic loci [275]. Current efforts center on delivery strategies, with AAV vectors providing strong CNS transduction and nanoparticles emerging as safer, non-viral alternatives [340]. The key challenges are achieving locus specificity, ensuring durability without irreversibility, and minimizing immune or off-target effects [341]. These tools hold unique promise for modulating inflammatory or neurogenic pathways directly at the epigenetic level, yet clinical translation will depend on overcoming safety, delivery, and regulatory hurdles with rigorous precision [275].
The future of translation lies in tailoring interventions to individual neuroinflammatory and epigenetic landscapes, moving beyond one-size-fits-all approaches [342]. Stratifying patients through biomarker and pharmacogenomic profiling could guide the choice of pharmacological, behavioral, or gene-based strategies, while combinatorial therapies may unlock greater synergy than single modalities [343]. Lifestyle programs could complement NLRP3 inhibition or epigenetic editing, and regenerative tools may be personalized for niche restoration [344]. The roadmap remains incremental, yet each milestone brings us closer to clinically rejuvenating neurogenesis and sustaining cognition across aging and disease [186,203].
6.3. Ethical and Clinical Considerations (~400 Words)
Precision epigenetic editing holds transformative therapeutic potential, yet it brings significant safety challenges that cannot be overlooked [345]. CRISPR-based approaches risk unintended off-target modifications, unpredictable durability of changes, and immune responses triggered by delivery vectors such as AAV or nanoparticles [346]. In the central nervous system, even small errors may have lasting effects, raising concerns about circuit stability and tumorigenesis [347]. Long-term surveillance will be essential to detect delayed consequences of editing [348]. Preclinical pipelines must therefore prioritize reversible, temporally controlled systems and comprehensive off-target profiling before these strategies advance into first-in-human trials [349].
Glial reprogramming represents one of the most exciting frontiers in regenerative neuroscience, yet its clinical translation is shadowed by profound safety concerns [350]. Converting glia into neurons carries risks of aberrant network activity, seizure induction, or tumorigenesis if new cells fail to integrate correctly [237]. Proper synaptic incorporation and maintenance of circuit balance are therefore paramount, as incomplete or uncontrolled conversion could destabilize neural networks [351]. The promise of replacing lost neurons must be weighed against an ethical duty to protect vulnerable patients, ensuring that regenerative enthusiasm does not outpace rigorous safety and ethical oversight [249].
Immune modulation in the aging brain presents a profound double-edged challenge. While suppressing microglial overactivation or dampening inflammasome signaling may restore plasticity and cognitive resilience, excessive suppression risks compromising pathogen defense, elevating vulnerability to infection or even cancer [352]. Calibrating these interventions in elderly patients therefore requires a fine balance between rejuvenation and safety [352]. Stratification by immune competence, genetic background, and comorbidities will be critical for minimizing harm [353]. Ultimately, personalized immune interventions must progress cautiously, ensuring that therapeutic innovation aligns with the biological realities of aging immunity.
Preventive interventions in normal aging raise complex ethical dilemmas, particularly when the therapies under consideration carry high risks [354]. At what threshold does delaying cognitive decline justify invasive gene editing or immune modulation in individuals without disease? Autonomy and informed consent must remain central, yet both are challenged by uncertainty in predicting benefit [354]. Societal concerns also arise, from medicalizing normal aging to reinforcing inequities in access [354]. Defining clear thresholds of clinical risk versus potential gain is therefore essential before preventive neuroenhancement can be ethically endorsed.
Regulatory pathways in neurodegenerative interventions differ markedly, with small molecules such as NLRP3 inhibitors often advancing more rapidly than gene or cell-based therapies, which require complex oversight [265]. Designing trials that capture both safety and meaningful outcomes remains essential [355]. Beyond adverse events, endpoints must include cognition, neurogenesis biomarkers, and quality of life to demonstrate true clinical relevance [355]. Harmonized protocols, long-term monitoring, and adaptive trial designs will be critical [356]. Ultimately, ethical rigor and regulatory foresight provide the scaffolding for translating scientific breakthroughs into safe, responsible therapies [357].
7. Conclusions
Neurogenesis and neuroinflammation exist in continuous dialogue, shaping how the brain ages and responds to stress [12]. This review has highlighted that disrupting this dialogue accelerates decline, while recalibrating it can preserve or even restore cognitive resilience. Mapping key gaps and aligning them with targeted strategies provides a roadmap for intervention [2]. The central message is clear: the neurogenic potential of the aging brain is not lost but remains accessible if the immune environment is carefully tuned. Even in late life, glia and neural stem cells retain remarkable plasticity. Harnessing this latent capacity requires converging approaches, from modulating microglial states and inflammasome signaling to applying epigenetic editing, gene therapy, and novel imaging biomarkers [358,359]. These strategies are more than incremental advances; they represent a shift in how we conceptualize and attempt to reshape the brain’s fate during aging. The novelty and significance of these approaches lie in their integrative scope. By combining mechanistic insights with innovative technologies, we are beginning to see the contours of interventions that could delay neurodegeneration and protect cognition. The challenge now is translation: embedding safety, ethics, and rigorous trial design into every step. With continued interdisciplinary collaboration, the prospect of actively guiding the neuroimmune dialogue toward healthier brain aging is within reach [2,255].
Author Contributions
Conceptualization, M.T.; methodology, M.T.; software, M.T.; validation, M.T.; formal analysis, M.T.; investigation, M.T.; resources, M.T.; data curation, M.T.; writing—original draft preparation, M.T.; writing—review and editing, M.T..; visualization, M.T.; supervision, M.T.; project administration, M.T.; funding acquisition, M.T. All author has read and agreed to the published version of the manuscript.
Funding
This work was supported by the HUN-REN Hungarian Research Network.
Acknowledgments
The graphical abstract and figures were created using biorender.com.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AD | Alzheimer’s disease |
| AAV | adeno-associated virus |
| BBB | blood-brain barrier |
| BDNF | brain-derived neurotrophic factor |
| CNS | central nervous system |
| CRISPR | clustered regularly interspaced short palindromic repeats |
| CSF | cerebrospinal fluid |
| CX3CL1 | C-X3-C motif chemokine ligand 1 |
| CX3CR1 | C-X3-C motif chemokine receptor 1 |
| DAM | disease-associated microglia |
| DCX | doublecortin |
| DG | dentate gyrus |
| EVs | extracellular vesicles |
| IFN-γ | interferon-gamma |
| IL-1β | interleukin-1 beta |
| IL-18 | interleukin-18 |
| iPSC | induced pluripotent stem cell |
| JAK/STAT1 | janus kinase/signal transducer and activator of transcription 1 |
| MCC950 | nlrp3 inflammasome inhibitor mcc950 |
| MRI | magnetic resonance imaging |
| NLRP3 | nod-like receptor protein 3 |
| NSAID | non-steroidal anti-inflammatory drug |
| PET | positron emission tomography |
| PSA-NCAM | polysialylated neural cell adhesion molecule |
| RNA-seq | RNA sequencing |
| SVZ | subventricular zone |
| TBI | traumatic brain injury |
| TNF-α | tumor necrosis factor-alpha |
| TSPO | translocator protein 18 kDa |
| Wnt | wingless-related integration site signaling pathway |
References
- Boldrini, M.; Fulmore, C.A.; Tartt, A.N.; Simeon, L.R.; Pavlova, I.; Poposka, V.; Rosoklija, G.B.; Stankov, A.; Arango, V.; Dwork, A.J.; et al. Human Hippocampal Neurogenesis Persists throughout Aging. Cell Stem Cell 2018, 22, 589-599.e585. [CrossRef]
- Babcock, K.R.; Page, J.S.; Fallon, J.R.; Webb, A.E. Adult Hippocampal Neurogenesis in Aging and Alzheimer's Disease. Stem Cell Reports 2021, 16, 681-693. [CrossRef]
- Anacker, C.; Hen, R. Adult hippocampal neurogenesis and cognitive flexibility - linking memory and mood. Nat Rev Neurosci 2017, 18, 335-346. [CrossRef]
- de Lima, E.P.; Laurindo, L.F.; Catharin, V.C.S.; Direito, R.; Tanaka, M.; Jasmin Santos German, I.; Lamas, C.B.; Guiguer, E.L.; Araújo, A.C.; Fiorini, A.M.R.; et al. Polyphenols, Alkaloids, and Terpenoids Against Neurodegeneration: Evaluating the Neuroprotective Effects of Phytocompounds Through a Comprehensive Review of the Current Evidence. Metabolites 2025, 15. [CrossRef]
- Culig, L.; Chu, X.; Bohr, V.A. Neurogenesis in aging and age-related neurodegenerative diseases. Ageing Res Rev 2022, 78, 101636. [CrossRef]
- Toda, T.; Parylak, S.L.; Linker, S.B.; Gage, F.H. The role of adult hippocampal neurogenesis in brain health and disease. Mol Psychiatry 2019, 24, 67-87. [CrossRef]
- Valero, J.; Bernardino, L.; Cardoso, F.L.; Silva, A.P.; Fontes-Ribeiro, C.; Ambrósio, A.F.; Malva, J.O. Impact of Neuroinflammation on Hippocampal Neurogenesis: Relevance to Aging and Alzheimer's Disease. J Alzheimers Dis 2017, 60, S161-s168. [CrossRef]
- Amanollahi, M.; Jameie, M.; Heidari, A.; Rezaei, N. The Dialogue Between Neuroinflammation and Adult Neurogenesis: Mechanisms Involved and Alterations in Neurological Diseases. Mol Neurobiol 2023, 60, 923-959. [CrossRef]
- Mészáros, Á.; Molnár, K.; Nógrádi, B.; Hernádi, Z.; Nyúl-Tóth, Á.; Wilhelm, I.; Krizbai, I.A. Neurovascular Inflammaging in Health and Disease. Cells 2020, 9. [CrossRef]
- Jurcau, M.C.; Jurcau, A.; Cristian, A.; Hogea, V.O.; Diaconu, R.G.; Nunkoo, V.S. Inflammaging and Brain Aging. Int J Mol Sci 2024, 25. [CrossRef]
- Marschallinger, J.; Iram, T.; Zardeneta, M.; Lee, S.E.; Lehallier, B.; Haney, M.S.; Pluvinage, J.V.; Mathur, V.; Hahn, O.; Morgens, D.W.; et al. Lipid-droplet-accumulating microglia represent a dysfunctional and proinflammatory state in the aging brain. Nat Neurosci 2020, 23, 194-208. [CrossRef]
- Wendimu, M.Y.; Hooks, S.B. Microglia Phenotypes in Aging and Neurodegenerative Diseases. Cells 2022, 11. [CrossRef]
- Lawrence, J.M.; Schardien, K.; Wigdahl, B.; Nonnemacher, M.R. Roles of neuropathology-associated reactive astrocytes: a systematic review. Acta Neuropathol Commun 2023, 11, 42. [CrossRef]
- Clarke, L.E.; Liddelow, S.A.; Chakraborty, C.; Münch, A.E.; Heiman, M.; Barres, B.A. Normal aging induces A1-like astrocyte reactivity. Proc Natl Acad Sci U S A 2018, 115, E1896-e1905. [CrossRef]
- Propson, N.E.; Roy, E.R.; Litvinchuk, A.; Köhl, J.; Zheng, H. Endothelial C3a receptor mediates vascular inflammation and blood-brain barrier permeability during aging. J Clin Invest 2021, 131. [CrossRef]
- Elahy, M.; Jackaman, C.; Mamo, J.C.; Lam, V.; Dhaliwal, S.S.; Giles, C.; Nelson, D.; Takechi, R. Blood-brain barrier dysfunction developed during normal aging is associated with inflammation and loss of tight junctions but not with leukocyte recruitment. Immun Ageing 2015, 12, 2. [CrossRef]
- von Bernhardi, R.; Eugenín-von Bernhardi, L.; Eugenín, J. Microglial cell dysregulation in brain aging and neurodegeneration. Front Aging Neurosci 2015, 7, 124. [CrossRef]
- de Lima, E.P.; Tanaka, M.; Lamas, C.B.; Quesada, K.; Detregiachi, C.R.P.; Araújo, A.C.; Guiguer, E.L.; Catharin, V.; de Castro, M.V.M.; Junior, E.B.; et al. Vascular Impairment, Muscle Atrophy, and Cognitive Decline: Critical Age-Related Conditions. Biomedicines 2024, 12. [CrossRef]
- Nunes, Y.C.; Mendes, N.M.; Pereira de Lima, E.; Chehadi, A.C.; Lamas, C.B.; Haber, J.F.S.; Dos Santos Bueno, M.; Araújo, A.C.; Catharin, V.C.S.; Detregiachi, C.R.P.; et al. Curcumin: A Golden Approach to Healthy Aging: A Systematic Review of the Evidence. Nutrients 2024, 16. [CrossRef]
- Ramnauth, A.D.; Tippani, M.; Divecha, H.R.; Papariello, A.R.; Miller, R.A.; Nelson, E.D.; Thompson, J.R.; Pattie, E.A.; Kleinman, J.E.; Maynard, K.R.; et al. Spatiotemporal analysis of gene expression in the human dentate gyrus reveals age-associated changes in cellular maturation and neuroinflammation. Cell Rep 2025, 44, 115300. [CrossRef]
- Wu, Y.; Korobeynyk, V.I.; Zamboni, M.; Waern, F.; Cole, J.D.; Mundt, S.; Greter, M.; Frisén, J.; Llorens-Bobadilla, E.; Jessberger, S. Multimodal transcriptomics reveal neurogenic aging trajectories and age-related regional inflammation in the dentate gyrus. Nat Neurosci 2025, 28, 415-430. [CrossRef]
- Mathews, K.J.; Allen, K.M.; Boerrigter, D.; Ball, H.; Shannon Weickert, C.; Double, K.L. Evidence for reduced neurogenesis in the aging human hippocampus despite stable stem cell markers. Aging Cell 2017, 16, 1195-1199. [CrossRef]
- Bedrosian, T.A.; Houtman, J.; Eguiguren, J.S.; Ghassemzadeh, S.; Rund, N.; Novaresi, N.M.; Hu, L.; Parylak, S.L.; Denli, A.M.; Randolph-Moore, L.; et al. Lamin B1 decline underlies age-related loss of adult hippocampal neurogenesis. Embo j 2021, 40, e105819. [CrossRef]
- Ishijima, T.; Nakajima, K. Inflammatory cytokines TNFα, IL-1β, and IL-6 are induced in endotoxin- stimulated microglia through different signaling cascades. Sci Prog 2021, 104, 368504211054985. [CrossRef]
- Tanaka, M.; Battaglia, S. From Biomarkers to Behavior: Mapping the Neuroimmune Web of Pain, Mood, and Memory. Biomedicines 2025, 13. [CrossRef]
- Araki, T.; Ikegaya, Y.; Koyama, R. The effects of microglia- and astrocyte-derived factors on neurogenesis in health and disease. Eur J Neurosci 2021, 54, 5880-5901. [CrossRef]
- Ekdahl, C.T. Microglial activation - tuning and pruning adult neurogenesis. Front Pharmacol 2012, 3, 41. [CrossRef]
- Früholz, I.; Meyer-Luehmann, M. The intricate interplay between microglia and adult neurogenesis in Alzheimer's disease. Front Cell Neurosci 2024, 18, 1456253. [CrossRef]
- Al-Onaizi, M.; Al-Khalifah, A.; Qasem, D.; ElAli, A. Role of Microglia in Modulating Adult Neurogenesis in Health and Neurodegeneration. Int J Mol Sci 2020, 21. [CrossRef]
- Gao, C.; Jiang, J.; Tan, Y.; Chen, S. Microglia in neurodegenerative diseases: mechanism and potential therapeutic targets. Signal Transduct Target Ther 2023, 8, 359. [CrossRef]
- Sanchez-Molina, P.; Almolda, B.; Giménez-Llort, L.; González, B.; Castellano, B. Chronic IL-10 overproduction disrupts microglia-neuron dialogue similar to aging, resulting in impaired hippocampal neurogenesis and spatial memory. Brain Behav Immun 2022, 101, 231-245. [CrossRef]
- De Lucia, C.; Rinchon, A.; Olmos-Alonso, A.; Riecken, K.; Fehse, B.; Boche, D.; Perry, V.H.; Gomez-Nicola, D. Microglia regulate hippocampal neurogenesis during chronic neurodegeneration. Brain Behav Immun 2016, 55, 179-190. [CrossRef]
- Domínguez-Rivas, E.; Ávila-Muñoz, E.; Schwarzacher, S.W.; Zepeda, A. Adult hippocampal neurogenesis in the context of lipopolysaccharide-induced neuroinflammation: A molecular, cellular and behavioral review. Brain Behav Immun 2021, 97, 286-302. [CrossRef]
- Chesnokova, V.; Pechnick, R.N.; Wawrowsky, K. Chronic peripheral inflammation, hippocampal neurogenesis, and behavior. Brain Behav Immun 2016, 58, 1-8. [CrossRef]
- Rusznák, K.; Horváth Á, I.; Pohli-Tóth, K.; Futácsi, A.; Kemény, Á.; Kiss, G.; Helyes, Z.; Czéh, B. Experimental Arthritis Inhibits Adult Hippocampal Neurogenesis in Mice. Cells 2022, 11. [CrossRef]
- Zonis, S.; Pechnick, R.N.; Ljubimov, V.A.; Mahgerefteh, M.; Wawrowsky, K.; Michelsen, K.S.; Chesnokova, V. Chronic intestinal inflammation alters hippocampal neurogenesis. J Neuroinflammation 2015, 12, 65. [CrossRef]
- Liu, Q.; Zhang, J.; Xiao, C.; Su, D.; Li, L.; Yang, C.; Zhao, Z.; Jiang, W.; You, Z.; Zhou, T. Akebia saponin D protects hippocampal neurogenesis from microglia-mediated inflammation and ameliorates depressive-like behaviors and cognitive impairment in mice through the PI3K-Akt pathway. Front Pharmacol 2022, 13, 927419. [CrossRef]
- Ekdahl, C.T.; Claasen, J.H.; Bonde, S.; Kokaia, Z.; Lindvall, O. Inflammation is detrimental for neurogenesis in adult brain. Proc Natl Acad Sci U S A 2003, 100, 13632-13637. [CrossRef]
- Golia, M.T.; Poggini, S.; Alboni, S.; Garofalo, S.; Ciano Albanese, N.; Viglione, A.; Ajmone-Cat, M.A.; St-Pierre, A.; Brunello, N.; Limatola, C.; et al. Interplay between inflammation and neural plasticity: Both immune activation and suppression impair LTP and BDNF expression. Brain Behav Immun 2019, 81, 484-494. [CrossRef]
- Miguel-Hidalgo, J.J.; Pang, Y. Role of neuroinflammation in the establishment of the neurogenic microenvironment in brain diseases. Current Tissue Microenvironment Reports 2021, 2, 17-28.
- Grabert, K.; Michoel, T.; Karavolos, M.H.; Clohisey, S.; Baillie, J.K.; Stevens, M.P.; Freeman, T.C.; Summers, K.M.; McColl, B.W. Microglial brain region-dependent diversity and selective regional sensitivities to aging. Nat Neurosci 2016, 19, 504-516. [CrossRef]
- Tan, Y.L.; Yuan, Y.; Tian, L. Microglial regional heterogeneity and its role in the brain. Mol Psychiatry 2020, 25, 351-367. [CrossRef]
- Yu, H.; Chang, Q.; Sun, T.; He, X.; Wen, L.; An, J.; Feng, J.; Zhao, Y. Metabolic reprogramming and polarization of microglia in Parkinson's disease: Role of inflammasome and iron. Ageing Res Rev 2023, 90, 102032. [CrossRef]
- Petralla, S.; De Chirico, F.; Miti, A.; Tartagni, O.; Massenzio, F.; Poeta, E.; Virgili, M.; Zuccheri, G.; Monti, B. Epigenetics and Communication Mechanisms in Microglia Activation with a View on Technological Approaches. Biomolecules 2021, 11. [CrossRef]
- Li, X.; Li, Y.; Jin, Y.; Zhang, Y.; Wu, J.; Xu, Z.; Huang, Y.; Cai, L.; Gao, S.; Liu, T.; et al. Transcriptional and epigenetic decoding of the microglial aging process. Nat Aging 2023, 3, 1288-1311. [CrossRef]
- Wang, W.; Wang, M.; Yang, M.; Zeng, B.; Qiu, W.; Ma, Q.; Jing, X.; Zhang, Q.; Wang, B.; Yin, C.; et al. Transcriptome dynamics of hippocampal neurogenesis in macaques across the lifespan and aged humans. Cell Res 2022, 32, 729-743. [CrossRef]
- Zhang, X.; Wang, R.; Chen, H.; Jin, C.; Jin, Z.; Lu, J.; Xu, L.; Lu, Y.; Zhang, J.; Shi, L. Aged microglia promote peripheral T cell infiltration by reprogramming the microenvironment of neurogenic niches. Immun Ageing 2022, 19, 34. [CrossRef]
- Bisht, K.; Okojie, K.A.; Sharma, K.; Lentferink, D.H.; Sun, Y.Y.; Chen, H.R.; Uweru, J.O.; Amancherla, S.; Calcuttawala, Z.; Campos-Salazar, A.B.; et al. Capillary-associated microglia regulate vascular structure and function through PANX1-P2RY12 coupling in mice. Nat Commun 2021, 12, 5289. [CrossRef]
- Brubaker, D.K.; Lauffenburger, D.A. Translating preclinical models to humans. Science 2020, 367, 742-743. [CrossRef]
- Gault, N.; Szele, F.G. Immunohistochemical evidence for adult human neurogenesis in health and disease. WIREs Mech Dis 2021, 13, e1526. [CrossRef]
- Cutler, R.R.; Kokovay, E. Rejuvenating subventricular zone neurogenesis in the aging brain. Curr Opin Pharmacol 2020, 50, 1-8. [CrossRef]
- Ji, S.; Xiong, M.; Chen, H.; Liu, Y.; Zhou, L.; Hong, Y.; Wang, M.; Wang, C.; Fu, X.; Sun, X. Cellular rejuvenation: molecular mechanisms and potential therapeutic interventions for diseases. Signal Transduct Target Ther 2023, 8, 116. [CrossRef]
- Gillotin, S.; Sahni, V.; Lepko, T.; Hanspal, M.A.; Swartz, J.E.; Alexopoulou, Z.; Marshall, F.H. Targeting impaired adult hippocampal neurogenesis in ageing by leveraging intrinsic mechanisms regulating Neural Stem Cell activity. Ageing Res Rev 2021, 71, 101447. [CrossRef]
- Niklison-Chirou, M.V.; Agostini, M.; Amelio, I.; Melino, G. Regulation of Adult Neurogenesis in Mammalian Brain. Int J Mol Sci 2020, 21. [CrossRef]
- Abbott, L.C.; Nigussie, F. Adult neurogenesis in the mammalian dentate gyrus. Anat Histol Embryol 2020, 49, 3-16. [CrossRef]
- Apple, D.M.; Solano-Fonseca, R.; Kokovay, E. Neurogenesis in the aging brain. Biochem Pharmacol 2017, 141, 77-85. [CrossRef]
- Nicaise, A.M.; Willis, C.M.; Crocker, S.J.; Pluchino, S. Stem Cells of the Aging Brain. Front Aging Neurosci 2020, 12, 247. [CrossRef]
- Peng, H.; Whitney, N.; Wu, Y.; Tian, C.; Dou, H.; Zhou, Y.; Zheng, J. HIV-1-infected and/or immune-activated macrophage-secreted TNF-alpha affects human fetal cortical neural progenitor cell proliferation and differentiation. Glia 2008, 56, 903-916. [CrossRef]
- Vidal, P.M.; Lemmens, E.; Dooley, D.; Hendrix, S. The role of "anti-inflammatory" cytokines in axon regeneration. Cytokine Growth Factor Rev 2013, 24, 1-12. [CrossRef]
- Liang, Z.; Jin, N.; Guo, W. Neural stem cell heterogeneity in adult hippocampus. Cell Regen 2025, 14, 6. [CrossRef]
- Calabrese, V.; Santoro, A.; Monti, D.; Crupi, R.; Di Paola, R.; Latteri, S.; Cuzzocrea, S.; Zappia, M.; Giordano, J.; Calabrese, E.J.; et al. Aging and Parkinson's Disease: Inflammaging, neuroinflammation and biological remodeling as key factors in pathogenesis. Free Radic Biol Med 2018, 115, 80-91. [CrossRef]
- Tanaka, M.; Vécsei, L. From Microbial Switches to Metabolic Sensors: Rewiring the Gut-Brain Kynurenine Circuit. Biomedicines 2025, 13. [CrossRef]
- Barbalho, S.M.; Leme Boaro, B.; da Silva Camarinha Oliveira, J.; Patočka, J.; Barbalho Lamas, C.; Tanaka, M.; Laurindo, L.F. Molecular Mechanisms Underlying Neuroinflammation Intervention with Medicinal Plants: A Critical and Narrative Review of the Current Literature. Pharmaceuticals (Basel) 2025, 18. [CrossRef]
- Duan, X.; Kang, E.; Liu, C.Y.; Ming, G.L.; Song, H. Development of neural stem cell in the adult brain. Curr Opin Neurobiol 2008, 18, 108-115. [CrossRef]
- Obernier, K.; Alvarez-Buylla, A. Neural stem cells: origin, heterogeneity and regulation in the adult mammalian brain. Development 2019, 146. [CrossRef]
- Kempermann, G.; Song, H.; Gage, F.H. Neurogenesis in the Adult Hippocampus. Cold Spring Harb Perspect Biol 2015, 7, a018812. [CrossRef]
- Cope, E.C.; Gould, E. Adult Neurogenesis, Glia, and the Extracellular Matrix. Cell Stem Cell 2019, 24, 690-705. [CrossRef]
- Sakamoto, M.; Kageyama, R.; Imayoshi, I. The functional significance of newly born neurons integrated into olfactory bulb circuits. Front Neurosci 2014, 8, 121. [CrossRef]
- Deshpande, A.; Bergami, M.; Ghanem, A.; Conzelmann, K.K.; Lepier, A.; Götz, M.; Berninger, B. Retrograde monosynaptic tracing reveals the temporal evolution of inputs onto new neurons in the adult dentate gyrus and olfactory bulb. Proc Natl Acad Sci U S A 2013, 110, E1152-1161. [CrossRef]
- Faigle, R.; Song, H. Signaling mechanisms regulating adult neural stem cells and neurogenesis. Biochim Biophys Acta 2013, 1830, 2435-2448. [CrossRef]
- Shi, J.; Wang, Z.; Wang, Z.; Shao, G.; Li, X. Epigenetic regulation in adult neural stem cells. Front Cell Dev Biol 2024, 12, 1331074. [CrossRef]
- Horgusluoglu, E.; Nudelman, K.; Nho, K.; Saykin, A.J. Adult neurogenesis and neurodegenerative diseases: A systems biology perspective. Am J Med Genet B Neuropsychiatr Genet 2017, 174, 93-112. [CrossRef]
- Bátiz, L.F.; Castro, M.A.; Burgos, P.V.; Velásquez, Z.D.; Muñoz, R.I.; Lafourcade, C.A.; Troncoso-Escudero, P.; Wyneken, U. Exosomes as Novel Regulators of Adult Neurogenic Niches. Front Cell Neurosci 2015, 9, 501. [CrossRef]
- Li, Y.; Guo, W. Neural Stem Cell Niche and Adult Neurogenesis. Neuroscientist 2021, 27, 235-245. [CrossRef]
- Quaresima, S.; Istiaq, A.; Jono, H.; Cacci, E.; Ohta, K.; Lupo, G. Assessing the Role of Ependymal and Vascular Cells as Sources of Extracellular Cues Regulating the Mouse Ventricular-Subventricular Zone Neurogenic Niche. Front Cell Dev Biol 2022, 10, 845567. [CrossRef]
- Kuhn, H.G.; Dickinson-Anson, H.; Gage, F.H. Neurogenesis in the dentate gyrus of the adult rat: age-related decrease of neuronal progenitor proliferation. J Neurosci 1996, 16, 2027-2033. [CrossRef]
- Lee, S.W.; Clemenson, G.D.; Gage, F.H. New neurons in an aged brain. Behav Brain Res 2012, 227, 497-507. [CrossRef]
- Bin Imtiaz, M.K.; Jaeger, B.N.; Bottes, S.; Machado, R.A.C.; Vidmar, M.; Moore, D.L.; Jessberger, S. Declining lamin B1 expression mediates age-dependent decreases of hippocampal stem cell activity. Cell Stem Cell 2021, 28, 967-977.e968. [CrossRef]
- Bast, L.; Calzolari, F.; Strasser, M.K.; Hasenauer, J.; Theis, F.J.; Ninkovic, J.; Marr, C. Increasing Neural Stem Cell Division Asymmetry and Quiescence Are Predicted to Contribute to the Age-Related Decline in Neurogenesis. Cell Rep 2018, 25, 3231-3240.e3238. [CrossRef]
- Pineda, J.R.; Daynac, M.; Chicheportiche, A.; Cebrian-Silla, A.; Sii Felice, K.; Garcia-Verdugo, J.M.; Boussin, F.D.; Mouthon, M.A. Vascular-derived TGF-β increases in the stem cell niche and perturbs neurogenesis during aging and following irradiation in the adult mouse brain. EMBO Mol Med 2013, 5, 548-562. [CrossRef]
- Buckwalter, M.S.; Yamane, M.; Coleman, B.S.; Ormerod, B.K.; Chin, J.T.; Palmer, T.; Wyss-Coray, T. Chronically increased transforming growth factor-beta1 strongly inhibits hippocampal neurogenesis in aged mice. Am J Pathol 2006, 169, 154-164. [CrossRef]
- DeCarolis, N.A.; Kirby, E.D.; Wyss-Coray, T.; Palmer, T.D. The Role of the Microenvironmental Niche in Declining Stem-Cell Functions Associated with Biological Aging. Cold Spring Harb Perspect Med 2015, 5. [CrossRef]
- Tanaka, M.; Vécsei, L. A Decade of Dedication: Pioneering Perspectives on Neurological Diseases and Mental Illnesses. Biomedicines 2024, 12. [CrossRef]
- Jiménez Peinado, P.; Urbach, A. From Youthful Vigor to Aging Decline: Unravelling the Intrinsic and Extrinsic Determinants of Hippocampal Neural Stem Cell Aging. Cells 2023, 12. [CrossRef]
- Moreno-Jiménez, E.P.; Flor-García, M.; Terreros-Roncal, J.; Rábano, A.; Cafini, F.; Pallas-Bazarra, N.; Ávila, J.; Llorens-Martín, M. Adult hippocampal neurogenesis is abundant in neurologically healthy subjects and drops sharply in patients with Alzheimer's disease. Nat Med 2019, 25, 554-560. [CrossRef]
- Knoth, R.; Singec, I.; Ditter, M.; Pantazis, G.; Capetian, P.; Meyer, R.P.; Horvat, V.; Volk, B.; Kempermann, G. Murine features of neurogenesis in the human hippocampus across the lifespan from 0 to 100 years. PLoS One 2010, 5, e8809. [CrossRef]
- Flor-García, M.; Terreros-Roncal, J.; Moreno-Jiménez, E.P.; Ávila, J.; Rábano, A.; Llorens-Martín, M. Unraveling human adult hippocampal neurogenesis. Nat Protoc 2020, 15, 668-693. [CrossRef]
- Terreros-Roncal, J.; Flor-García, M.; Moreno-Jiménez, E.P.; Rodríguez-Moreno, C.B.; Márquez-Valadez, B.; Gallardo-Caballero, M.; Rábano, A.; Llorens-Martín, M. Methods to study adult hippocampal neurogenesis in humans and across the phylogeny. Hippocampus 2023, 33, 271-306. [CrossRef]
- Seki, T. Understanding the Real State of Human Adult Hippocampal Neurogenesis From Studies of Rodents and Non-human Primates. Front Neurosci 2020, 14, 839. [CrossRef]
- Gulen, M.F.; Samson, N.; Keller, A.; Schwabenland, M.; Liu, C.; Glück, S.; Thacker, V.V.; Favre, L.; Mangeat, B.; Kroese, L.J.; et al. cGAS-STING drives ageing-related inflammation and neurodegeneration. Nature 2023, 620, 374-380. [CrossRef]
- Costa, J.; Martins, S.; Ferreira, P.A.; Cardoso, A.M.S.; Guedes, J.R.; Peça, J.; Cardoso, A.L. The old guard: Age-related changes in microglia and their consequences. Mech Ageing Dev 2021, 197, 111512. [CrossRef]
- Koellhoffer, E.C.; McCullough, L.D.; Ritzel, R.M. Old Maids: Aging and Its Impact on Microglia Function. Int J Mol Sci 2017, 18. [CrossRef]
- Ana, B. Aged-Related Changes in Microglia and Neurodegenerative Diseases: Exploring the Connection. Biomedicines 2024, 12. [CrossRef]
- Škandík, M.; Friess, L.; Vázquez-Cabrera, G.; Keane, L.; Grabert, K.; Cruz De Los Santos, M.; Posada-Pérez, M.; Baleviciute, A.; Cheray, M.; Joseph, B. Age-associated microglial transcriptome leads to diminished immunogenicity and dysregulation of MCT4 and P2RY12/P2RY13 related functions. Cell Death Discov 2025, 11, 16. [CrossRef]
- Edler, M.K.; Mhatre-Winters, I.; Richardson, J.R. Microglia in Aging and Alzheimer's Disease: A Comparative Species Review. Cells 2021, 10. [CrossRef]
- O'Neil, S.M.; Hans, E.E.; Jiang, S.; Wangler, L.M.; Godbout, J.P. Astrocyte immunosenescence and deficits in interleukin 10 signaling in the aged brain disrupt the regulation of microglia following innate immune activation. Glia 2022, 70, 913-934. [CrossRef]
- Tamatta, R.; Pai, V.; Jaiswal, C.; Singh, I.; Singh, A.K. Neuroinflammaging and the Immune Landscape: The Role of Autophagy and Senescence in Aging Brain. Biogerontology 2025, 26, 52. [CrossRef]
- Lutshumba, J.; Nikolajczyk, B.S.; Bachstetter, A.D. Dysregulation of Systemic Immunity in Aging and Dementia. Front Cell Neurosci 2021, 15, 652111. [CrossRef]
- Norden, D.M.; Godbout, J.P. Review: microglia of the aged brain: primed to be activated and resistant to regulation. Neuropathol Appl Neurobiol 2013, 39, 19-34. [CrossRef]
- Elmore, M.R.P.; Hohsfield, L.A.; Kramár, E.A.; Soreq, L.; Lee, R.J.; Pham, S.T.; Najafi, A.R.; Spangenberg, E.E.; Wood, M.A.; West, B.L.; et al. Replacement of microglia in the aged brain reverses cognitive, synaptic, and neuronal deficits in mice. Aging Cell 2018, 17, e12832. [CrossRef]
- Niraula, A.; Sheridan, J.F.; Godbout, J.P. Microglia Priming with Aging and Stress. Neuropsychopharmacology 2017, 42, 318-333. [CrossRef]
- Norden, D.M.; Muccigrosso, M.M.; Godbout, J.P. Microglial priming and enhanced reactivity to secondary insult in aging, and traumatic CNS injury, and neurodegenerative disease. Neuropharmacology 2015, 96, 29-41. [CrossRef]
- Neher, J.J.; Cunningham, C. Priming Microglia for Innate Immune Memory in the Brain. Trends Immunol 2019, 40, 358-374. [CrossRef]
- Hoeijmakers, L.; Heinen, Y.; van Dam, A.M.; Lucassen, P.J.; Korosi, A. Microglial Priming and Alzheimer's Disease: A Possible Role for (Early) Immune Challenges and Epigenetics? Front Hum Neurosci 2016, 10, 398. [CrossRef]
- O'Neil, S.M.; Witcher, K.G.; McKim, D.B.; Godbout, J.P. Forced turnover of aged microglia induces an intermediate phenotype but does not rebalance CNS environmental cues driving priming to immune challenge. Acta Neuropathol Commun 2018, 6, 129. [CrossRef]
- Wahl, D.; Risen, S.J.; Osburn, S.C.; Emge, T.; Sharma, S.; Gilberto, V.S.; Chatterjee, A.; Nagpal, P.; Moreno, J.A.; LaRocca, T.J. Nanoligomers targeting NF-κB and NLRP3 reduce neuroinflammation and improve cognitive function with aging and tauopathy. J Neuroinflammation 2024, 21, 182. [CrossRef]
- Li, Y.; Xia, Y.; Yin, S.; Wan, F.; Hu, J.; Kou, L.; Sun, Y.; Wu, J.; Zhou, Q.; Huang, J.; et al. Targeting Microglial α-Synuclein/TLRs/NF-kappaB/NLRP3 Inflammasome Axis in Parkinson's Disease. Front Immunol 2021, 12, 719807. [CrossRef]
- Lei, L.Y.; Wang, R.C.; Pan, Y.L.; Yue, Z.G.; Zhou, R.; Xie, P.; Tang, Z.S. Mangiferin inhibited neuroinflammation through regulating microglial polarization and suppressing NF-κB, NLRP3 pathway. Chin J Nat Med 2021, 19, 112-119. [CrossRef]
- Wang, T.; Liu, Y.; Lu, Y.; Chi, L. NTN-1 attenuates amyloid-β-mediated microglial neuroinflammation and memory impairment via the NF-κB pathway and NLRP3 inflammasome in a rat model of Alzheimer's disease. Front Aging Neurosci 2025, 17, 1516399. [CrossRef]
- Fornari Laurindo, L.; Aparecido Dias, J.; Cressoni Araújo, A.; Torres Pomini, K.; Machado Galhardi, C.; Rucco Penteado Detregiachi, C.; Santos de Argollo Haber, L.; Donizeti Roque, D.; Dib Bechara, M.; Vialogo Marques de Castro, M.; et al. Immunological dimensions of neuroinflammation and microglial activation: exploring innovative immunomodulatory approaches to mitigate neuroinflammatory progression. Front Immunol 2023, 14, 1305933. [CrossRef]
- Borsini, A.; Zunszain, P.A.; Thuret, S.; Pariante, C.M. The role of inflammatory cytokines as key modulators of neurogenesis. Trends Neurosci 2015, 38, 145-157. [CrossRef]
- Kong, X.; Gong, Z.; Zhang, L.; Sun, X.; Ou, Z.; Xu, B.; Huang, J.; Long, D.; He, X.; Lin, X.; et al. JAK2/STAT3 signaling mediates IL-6-inhibited neurogenesis of neural stem cells through DNA demethylation/methylation. Brain Behav Immun 2019, 79, 159-173. [CrossRef]
- Salminen, A.; Ojala, J.; Kaarniranta, K.; Kauppinen, A. Mitochondrial dysfunction and oxidative stress activate inflammasomes: impact on the aging process and age-related diseases. Cell Mol Life Sci 2012, 69, 2999-3013. [CrossRef]
- Tanaka, M.; Szabó, Á.; Vécsei, L. Redefining Roles: A Paradigm Shift in Tryptophan-Kynurenine Metabolism for Innovative Clinical Applications. Int J Mol Sci 2024, 25. [CrossRef]
- Juhász, L.; Spisák, K.; Szolnoki, B.Z.; Nászai, A.; Szabó, Á.; Rutai, A.; Tallósy, S.P.; Szabó, A.; Toldi, J.; Tanaka, M.; et al. The Power Struggle: Kynurenine Pathway Enzyme Knockouts and Brain Mitochondrial Respiration. J Neurochem 2025, 169, e70075. [CrossRef]
- Sarkar, S.; Malovic, E.; Harishchandra, D.S.; Ghaisas, S.; Panicker, N.; Charli, A.; Palanisamy, B.N.; Rokad, D.; Jin, H.; Anantharam, V.; et al. Mitochondrial impairment in microglia amplifies NLRP3 inflammasome proinflammatory signaling in cell culture and animal models of Parkinson's disease. NPJ Parkinsons Dis 2017, 3, 30. [CrossRef]
- Hansen, C.E.; Vacondio, D.; van der Molen, L.; Jüttner, A.A.; Fung, W.K.; Karsten, M.; van Het Hof, B.; Fontijn, R.D.; Kooij, G.; Witte, M.E.; et al. Endothelial-Ercc1 DNA repair deficiency provokes blood-brain barrier dysfunction. Cell Death Dis 2025, 16, 1. [CrossRef]
- Dulken, B.W.; Buckley, M.T.; Navarro Negredo, P.; Saligrama, N.; Cayrol, R.; Leeman, D.S.; George, B.M.; Boutet, S.C.; Hebestreit, K.; Pluvinage, J.V.; et al. Single-cell analysis reveals T cell infiltration in old neurogenic niches. Nature 2019, 571, 205-210. [CrossRef]
- Moreno-Valladares, M.; Moreno-Cugnon, L.; Silva, T.M.; Garcés, J.P.; Saenz-Antoñanzas, A.; Álvarez-Satta, M.; Matheu, A. CD8(+) T cells are increased in the subventricular zone with physiological and pathological aging. Aging Cell 2020, 19, e13198. [CrossRef]
- Solano Fonseca, R.; Mahesula, S.; Apple, D.M.; Raghunathan, R.; Dugan, A.; Cardona, A.; O'Connor, J.; Kokovay, E. Neurogenic Niche Microglia Undergo Positional Remodeling and Progressive Activation Contributing to Age-Associated Reductions in Neurogenesis. Stem Cells Dev 2016, 25, 542-555. [CrossRef]
- Fonken, L.K.; Gaudet, A.D. Neuroimmunology of healthy brain aging. Curr Opin Neurobiol 2022, 77, 102649. [CrossRef]
- Chintamen, S.; Imessadouene, F.; Kernie, S.G. Immune Regulation of Adult Neurogenic Niches in Health and Disease. Front Cell Neurosci 2020, 14, 571071. [CrossRef]
- Diaz-Aparicio, I.; Paris, I.; Sierra-Torre, V.; Plaza-Zabala, A.; Rodríguez-Iglesias, N.; Márquez-Ropero, M.; Beccari, S.; Huguet, P.; Abiega, O.; Alberdi, E.; et al. Microglia Actively Remodel Adult Hippocampal Neurogenesis through the Phagocytosis Secretome. J Neurosci 2020, 40, 1453-1482. [CrossRef]
- Sierra, A.; Encinas, J.M.; Deudero, J.J.; Chancey, J.H.; Enikolopov, G.; Overstreet-Wadiche, L.S.; Tsirka, S.E.; Maletic-Savatic, M. Microglia shape adult hippocampal neurogenesis through apoptosis-coupled phagocytosis. Cell Stem Cell 2010, 7, 483-495. [CrossRef]
- Kurematsu, C.; Sawada, M.; Ohmuraya, M.; Tanaka, M.; Kuboyama, K.; Ogino, T.; Matsumoto, M.; Oishi, H.; Inada, H.; Ishido, Y.; et al. Synaptic pruning of murine adult-born neurons by microglia depends on phosphatidylserine. J Exp Med 2022, 219. [CrossRef]
- Jiang, X.; Yi, S.; Liu, Q.; Zhang, J. The secretome of microglia induced by IL-4 of IFN-γ differently regulate proliferation, differentiation and survival of adult neural stem/progenitor cell by targeting the PI3K-Akt pathway. Cytotechnology 2022, 74, 407-420. [CrossRef]
- Matsui, T.K.; Mori, E. Microglia support neural stem cell maintenance and growth. Biochem Biophys Res Commun 2018, 503, 1880-1884. [CrossRef]
- Wlodarczyk, A.; Holtman, I.R.; Krueger, M.; Yogev, N.; Bruttger, J.; Khorooshi, R.; Benmamar-Badel, A.; de Boer-Bergsma, J.J.; Martin, N.A.; Karram, K.; et al. A novel microglial subset plays a key role in myelinogenesis in developing brain. Embo j 2017, 36, 3292-3308. [CrossRef]
- Mallard, C.; Tremblay, M.E.; Vexler, Z.S. Microglia and Neonatal Brain Injury. Neuroscience 2019, 405, 68-76. [CrossRef]
- Harley, S.B.R.; Willis, E.F.; Shaikh, S.N.; Blackmore, D.G.; Sah, P.; Ruitenberg, M.J.; Bartlett, P.F.; Vukovic, J. Selective Ablation of BDNF from Microglia Reveals Novel Roles in Self-Renewal and Hippocampal Neurogenesis. J Neurosci 2021, 41, 4172-4186. [CrossRef]
- Xu, H.; Gelyana, E.; Rajsombath, M.; Yang, T.; Li, S.; Selkoe, D. Environmental Enrichment Potently Prevents Microglia-Mediated Neuroinflammation by Human Amyloid β-Protein Oligomers. J Neurosci 2016, 36, 9041-9056. [CrossRef]
- Mee-Inta, O.; Zhao, Z.W.; Kuo, Y.M. Physical Exercise Inhibits Inflammation and Microglial Activation. Cells 2019, 8. [CrossRef]
- Choi, J.Y.; Kim, J.Y.; Kim, J.Y.; Park, J.; Lee, W.T.; Lee, J.E. M2 Phenotype Microglia-derived Cytokine Stimulates Proliferation and Neuronal Differentiation of Endogenous Stem Cells in Ischemic Brain. Exp Neurobiol 2017, 26, 33-41. [CrossRef]
- Orihuela, R.; McPherson, C.A.; Harry, G.J. Microglial M1/M2 polarization and metabolic states. Br J Pharmacol 2016, 173, 649-665. [CrossRef]
- Vay, S.U.; Flitsch, L.J.; Rabenstein, M.; Rogall, R.; Blaschke, S.; Kleinhaus, J.; Reinert, N.; Bach, A.; Fink, G.R.; Schroeter, M.; et al. The plasticity of primary microglia and their multifaceted effects on endogenous neural stem cells in vitro and in vivo. J Neuroinflammation 2018, 15, 226. [CrossRef]
- Nelson, L.H.; Peketi, P.; Lenz, K.M. Microglia Regulate Cell Genesis in a Sex-dependent Manner in the Neonatal Hippocampus. Neuroscience 2021, 453, 237-255. [CrossRef]
- Jurgens, H.A.; Johnson, R.W. Dysregulated neuronal-microglial cross-talk during aging, stress and inflammation. Exp Neurol 2012, 233, 40-48. [CrossRef]
- Zhang, J.; He, H.; Qiao, Y.; Zhou, T.; He, H.; Yi, S.; Zhang, L.; Mo, L.; Li, Y.; Jiang, W.; et al. Priming of microglia with IFN-γ impairs adult hippocampal neurogenesis and leads to depression-like behaviors and cognitive defects. Glia 2020, 68, 2674-2692. [CrossRef]
- Carrier, M.; Šimončičová, E.; St-Pierre, M.K.; McKee, C.; Tremblay, M. Psychological Stress as a Risk Factor for Accelerated Cellular Aging and Cognitive Decline: The Involvement of Microglia-Neuron Crosstalk. Front Mol Neurosci 2021, 14, 749737. [CrossRef]
- Afridi, R.; Lee, W.H.; Suk, K. Microglia Gone Awry: Linking Immunometabolism to Neurodegeneration. Front Cell Neurosci 2020, 14, 246. [CrossRef]
- Mecca, C.; Giambanco, I.; Donato, R.; Arcuri, C. Microglia and Aging: The Role of the TREM2-DAP12 and CX3CL1-CX3CR1 Axes. Int J Mol Sci 2018, 19. [CrossRef]
- Pawelec, P.; Ziemka-Nalecz, M.; Sypecka, J.; Zalewska, T. The Impact of the CX3CL1/CX3CR1 Axis in Neurological Disorders. Cells 2020, 9. [CrossRef]
- Vukovic, J.; Colditz, M.J.; Blackmore, D.G.; Ruitenberg, M.J.; Bartlett, P.F. Microglia modulate hippocampal neural precursor activity in response to exercise and aging. J Neurosci 2012, 32, 6435-6443. [CrossRef]
- Bolós, M.; Perea, J.R.; Terreros-Roncal, J.; Pallas-Bazarra, N.; Jurado-Arjona, J.; Ávila, J.; Llorens-Martín, M. Absence of microglial CX3CR1 impairs the synaptic integration of adult-born hippocampal granule neurons. Brain Behav Immun 2018, 68, 76-89. [CrossRef]
- Morton, M.C.; Neckles, V.N.; Seluzicki, C.M.; Holmberg, J.C.; Feliciano, D.M. Neonatal Subventricular Zone Neural Stem Cells Release Extracellular Vesicles that Act as a Microglial Morphogen. Cell Rep 2018, 23, 78-89. [CrossRef]
- Chen, X.; Jiang, M.; Li, H.; Wang, Y.; Shen, H.; Li, X.; Zhang, Y.; Wu, J.; Yu, Z.; Chen, G. CX3CL1/CX3CR1 axis attenuates early brain injury via promoting the delivery of exosomal microRNA-124 from neuron to microglia after subarachnoid hemorrhage. J Neuroinflammation 2020, 17, 209. [CrossRef]
- Qian, H.D.; Song, X.Y.; He, G.W.; Peng, X.N.; Chen, Y.; Huang, P.; Zhang, J.; Lin, X.Y.; Gao, Q.; Zhu, S.M.; et al. Müller Glial-Derived Small Extracellular Vesicles Mitigate RGC Degeneration by Suppressing Microglial Activation via Cx3cl1-Cx3cr1 Signaling. Adv Healthc Mater 2025, 14, e2404306. [CrossRef]
- Fritze, J.; Muralidharan, C.; Stamp, E.; Ahlenius, H. Microglia undergo disease-associated transcriptional activation and CX3C motif chemokine receptor 1 expression regulates neurogenesis in the aged brain. Dev Neurobiol 2024, 84, 128-141. [CrossRef]
- Gemma, C.; Bachstetter, A.D.; Bickford, P.C. Neuron-Microglia Dialogue and Hippocampal Neurogenesis in the Aged Brain. Aging Dis 2010, 1, 232-244.
- Barko, K.; Shelton, M.; Xue, X.; Afriyie-Agyemang, Y.; Puig, S.; Freyberg, Z.; Tseng, G.C.; Logan, R.W.; Seney, M.L. Brain region- and sex-specific transcriptional profiles of microglia. Front Psychiatry 2022, 13, 945548. [CrossRef]
- Spencer, S.J.; Basri, B.; Sominsky, L.; Soch, A.; Ayala, M.T.; Reineck, P.; Gibson, B.C.; Barrientos, R.M. High-fat diet worsens the impact of aging on microglial function and morphology in a region-specific manner. Neurobiol Aging 2019, 74, 121-134. [CrossRef]
- Ribeiro Xavier, A.L.; Kress, B.T.; Goldman, S.A.; Lacerda de Menezes, J.R.; Nedergaard, M. A Distinct Population of Microglia Supports Adult Neurogenesis in the Subventricular Zone. J Neurosci 2015, 35, 11848-11861. [CrossRef]
- Smith, L.K.; White, C.W., 3rd; Villeda, S.A. The systemic environment: at the interface of aging and adult neurogenesis. Cell Tissue Res 2018, 371, 105-113. [CrossRef]
- Lana, D.; Magni, G.; Landucci, E.; Wenk, G.L.; Pellegrini-Giampietro, D.E.; Giovannini, M.G. Phenomic Microglia Diversity as a Druggable Target in the Hippocampus in Neurodegenerative Diseases. Int J Mol Sci 2023, 24. [CrossRef]
- Sato, K. Effects of Microglia on Neurogenesis. Glia 2015, 63, 1394-1405. [CrossRef]
- Chintamen, S.; Gaur, P.; Vo, N.; Bradshaw, E.M.; Menon, V.; Kernie, S.G. Distinct microglial transcriptomic signatures within the hippocampus. PLoS One 2024, 19, e0296280. [CrossRef]
- McKee, C.G.; Hoffos, M.; Vecchiarelli, H.A.; Tremblay, M. Microglia: A pharmacological target for the treatment of age-related cognitive decline and Alzheimer's disease. Front Pharmacol 2023, 14, 1125982. [CrossRef]
- McGroarty, J.; Salinas, S.; Evans, H.; Jimenez, B.; Tran, V.; Kadavakollu, S.; Vashist, A.; Atluri, V. Inflammasome-Mediated Neuroinflammation: A Key Driver in Alzheimer's Disease Pathogenesis. Biomolecules 2025, 15. [CrossRef]
- Xu, W.; Huang, Y.; Zhou, R. NLRP3 inflammasome in neuroinflammation and central nervous system diseases. Cell Mol Immunol 2025, 22, 341-355. [CrossRef]
- Khilazheva, E.D.; Mosiagina, A.I.; Panina, Y.A.; Belozor, O.S.; Komleva, Y.K. Impact of NLRP3 Depletion on Aging-Related Metaflammation, Cognitive Function, and Social Behavior in Mice. Int J Mol Sci 2023, 24. [CrossRef]
- Zhang, X.; Kracht, L.; Lerario, A.M.; Dubbelaar, M.L.; Brouwer, N.; Wesseling, E.M.; Boddeke, E.; Eggen, B.J.L.; Kooistra, S.M. Epigenetic regulation of innate immune memory in microglia. J Neuroinflammation 2022, 19, 111. [CrossRef]
- Kamei, N.; Day, K.; Guo, W.; Haus, D.L.; Nguyen, H.X.; Scarfone, V.M.; Booher, K.; Jia, X.Y.; Cummings, B.J.; Anderson, A.J. Injured inflammatory environment overrides the TET2 shaped epigenetic landscape of pluripotent stem cell derived human neural stem cells. Sci Rep 2024, 14, 25186. [CrossRef]
- Itokawa, N.; Oshima, M.; Koide, S.; Takayama, N.; Kuribayashi, W.; Nakajima-Takagi, Y.; Aoyama, K.; Yamazaki, S.; Yamaguchi, K.; Furukawa, Y.; et al. Epigenetic traits inscribed in chromatin accessibility in aged hematopoietic stem cells. Nat Commun 2022, 13, 2691. [CrossRef]
- Zocher, S.; Toda, T. Epigenetic aging in adult neurogenesis. Hippocampus 2023, 33, 347-359. [CrossRef]
- Das, N.D.; Chai, Y.G. Neuroinflammation on the Epigenetics. Neural Stem Cells: New Perspectives 2013, 381.
- Kodi, T.; Sankhe, R.; Gopinathan, A.; Nandakumar, K.; Kishore, A. New Insights on NLRP3 Inflammasome: Mechanisms of Activation, Inhibition, and Epigenetic Regulation. J Neuroimmune Pharmacol 2024, 19, 7. [CrossRef]
- Holleman, J.; Daniilidou, M.; Kåreholt, I.; Aspö, M.; Hagman, G.; Udeh-Momoh, C.T.; Spulber, G.; Kivipelto, M.; Solomon, A.; Matton, A.; et al. Diurnal cortisol, neuroinflammation, and neuroimaging visual rating scales in memory clinic patients. Brain Behav Immun 2024, 118, 499-509. [CrossRef]
- Dutta, P.; Quax, R.; Crielaard, L.; Badiali, L.; Sloot, P.M.A. Inferring temporal dynamics from cross-sectional data using Langevin dynamics. R Soc Open Sci 2021, 8, 211374. [CrossRef]
- Mroczek, M.; Desouky, A.; Sirry, W. Imaging Transcriptomics in Neurodegenerative Diseases. J Neuroimaging 2021, 31, 244-250. [CrossRef]
- Argiris, G.; Stern, Y.; Habeck, C. Quantifying Age-Related Changes in Brain and Behavior: A Longitudinal versus Cross-Sectional Approach. eNeuro 2021, 8. [CrossRef]
- Sochocka, M.; Diniz, B.S.; Leszek, J. Inflammatory Response in the CNS: Friend or Foe? Mol Neurobiol 2017, 54, 8071-8089. [CrossRef]
- Voloboueva, L.A.; Sun, X.; Xu, L.; Ouyang, Y.B.; Giffard, R.G. Distinct Effects of miR-210 Reduction on Neurogenesis: Increased Neuronal Survival of Inflammation But Reduced Proliferation Associated with Mitochondrial Enhancement. J Neurosci 2017, 37, 3072-3084. [CrossRef]
- Lee, N.; Choi, J.Y.; Ryu, Y.H. The development status of PET radiotracers for evaluating neuroinflammation. Nucl Med Mol Imaging 2024, 58, 160-176. [CrossRef]
- Jain, P.; Chaney, A.M.; Carlson, M.L.; Jackson, I.M.; Rao, A.; James, M.L. Neuroinflammation PET Imaging: Current Opinion and Future Directions. J Nucl Med 2020, 61, 1107-1112. [CrossRef]
- Narayanaswami, V.; Dahl, K.; Bernard-Gauthier, V.; Josephson, L.; Cumming, P.; Vasdev, N. Emerging PET Radiotracers and Targets for Imaging of Neuroinflammation in Neurodegenerative Diseases: Outlook Beyond TSPO. Mol Imaging 2018, 17, 1536012118792317. [CrossRef]
- Chen, Z.; Haider, A.; Chen, J.; Xiao, Z.; Gobbi, L.; Honer, M.; Grether, U.; Arnold, S.E.; Josephson, L.; Liang, S.H. The Repertoire of Small-Molecule PET Probes for Neuroinflammation Imaging: Challenges and Opportunities beyond TSPO. J Med Chem 2021, 64, 17656-17689. [CrossRef]
- Zhang, W.; Sun, H.S.; Wang, X.; Dumont, A.S.; Liu, Q. Cellular senescence, DNA damage, and neuroinflammation in the aging brain. Trends Neurosci 2024, 47, 461-474. [CrossRef]
- Jin, R.; Chan, A.K.Y.; Wu, J.; Lee, T.M.C. Relationships between Inflammation and Age-Related Neurocognitive Changes. Int J Mol Sci 2022, 23. [CrossRef]
- Gonzalez-Perez, O.; Gutierrez-Fernandez, F.; Lopez-Virgen, V.; Collas-Aguilar, J.; Quinones-Hinojosa, A.; Garcia-Verdugo, J.M. Immunological regulation of neurogenic niches in the adult brain. Neuroscience 2012, 226, 270-281. [CrossRef]
- Das, S.; Basu, A. Inflammation: a new candidate in modulating adult neurogenesis. J Neurosci Res 2008, 86, 1199-1208. [CrossRef]
- Galea, I. The blood-brain barrier in systemic infection and inflammation. Cell Mol Immunol 2021, 18, 2489-2501. [CrossRef]
- Wu, X.; Shen, Q.; Chang, H.; Li, J.; Xing, D. Promoted CD4(+) T cell-derived IFN-γ/IL-10 by photobiomodulation therapy modulates neurogenesis to ameliorate cognitive deficits in APP/PS1 and 3xTg-AD mice. J Neuroinflammation 2022, 19, 253. [CrossRef]
- Brunet, A.; Goodell, M.A.; Rando, T.A. Ageing and rejuvenation of tissue stem cells and their niches. Nat Rev Mol Cell Biol 2023, 24, 45-62. [CrossRef]
- Parkitny, L.; Maletic-Savatic, M. Glial PAMPering and DAMPening of Adult Hippocampal Neurogenesis. Brain Sci 2021, 11. [CrossRef]
- Allen, W.E.; Blosser, T.R.; Sullivan, Z.A.; Dulac, C.; Zhuang, X. Molecular and spatial signatures of mouse brain aging at single-cell resolution. Cell 2023, 186, 194-208.e118. [CrossRef]
- Velikic, G.; Maric, D.M.; Maric, D.L.; Supic, G.; Puletic, M.; Dulic, O.; Vojvodic, D. Harnessing the Stem Cell Niche in Regenerative Medicine: Innovative Avenue to Combat Neurodegenerative Diseases. Int J Mol Sci 2024, 25. [CrossRef]
- Duque, A.; Arellano, J.I.; Rakic, P. An assessment of the existence of adult neurogenesis in humans and value of its rodent models for neuropsychiatric diseases. Mol Psychiatry 2022, 27, 377-382. [CrossRef]
- Geirsdottir, L.; David, E.; Keren-Shaul, H.; Weiner, A.; Bohlen, S.C.; Neuber, J.; Balic, A.; Giladi, A.; Sheban, F.; Dutertre, C.A.; et al. Cross-Species Single-Cell Analysis Reveals Divergence of the Primate Microglia Program. Cell 2019, 179, 1609-1622.e1616. [CrossRef]
- Lim, D.A.; Alvarez-Buylla, A. The Adult Ventricular-Subventricular Zone (V-SVZ) and Olfactory Bulb (OB) Neurogenesis. Cold Spring Harb Perspect Biol 2016, 8. [CrossRef]
- Denoth-Lippuner, A.; Jessberger, S. Formation and integration of new neurons in the adult hippocampus. Nat Rev Neurosci 2021, 22, 223-236. [CrossRef]
- Tosoni, G.; Ayyildiz, D.; Bryois, J.; Macnair, W.; Fitzsimons, C.P.; Lucassen, P.J.; Salta, E. Mapping human adult hippocampal neurogenesis with single-cell transcriptomics: Reconciling controversy or fueling the debate? Neuron 2023, 111, 1714-1731.e1713. [CrossRef]
- Nutma, E.; Fancy, N.; Weinert, M.; Tsartsalis, S.; Marzin, M.C.; Muirhead, R.C.J.; Falk, I.; Breur, M.; de Bruin, J.; Hollaus, D.; et al. Translocator protein is a marker of activated microglia in rodent models but not human neurodegenerative diseases. Nat Commun 2023, 14, 5247. [CrossRef]
- Pediaditakis, I.; Kodella, K.R.; Manatakis, D.V.; Le, C.Y.; Barthakur, S.; Sorets, A.; Gravanis, A.; Ewart, L.; Rubin, L.L.; Manolakos, E.S.; et al. A microengineered Brain-Chip to model neuroinflammation in humans. iScience 2022, 25, 104813. [CrossRef]
- Tian, A.; Bhattacharya, A.; Muffat, J.; Li, Y. Expanding the neuroimmune research toolkit with in vivo brain organoid technologies. Dis Model Mech 2025, 18. [CrossRef]
- Balestri, W.; Sharma, R.; da Silva, V.A.; Bobotis, B.C.; Curle, A.J.; Kothakota, V.; Kalantarnia, F.; Hangad, M.V.; Hoorfar, M.; Jones, J.L.; et al. Modeling the neuroimmune system in Alzheimer's and Parkinson's diseases. J Neuroinflammation 2024, 21, 32. [CrossRef]
- Tanaka, M. Parkinson's Disease: Bridging Gaps, Building Biomarkers, and Reimagining Clinical Translation. Cells 2025, 14. [CrossRef]
- Tanaka, M.; Battaglia, S. Dualistic Dynamics in Neuropsychiatry: From Monoaminergic Modulators to Multiscale Biomarker Maps. Biomedicines 2025, 13. [CrossRef]
- De Schepper, S.; Crowley, G.; Hong, S. Understanding microglial diversity and implications for neuronal function in health and disease. Dev Neurobiol 2021, 81, 507-523. [CrossRef]
- Takkinen, J.S.; López-Picón, F.R.; Al Majidi, R.; Eskola, O.; Krzyczmonik, A.; Keller, T.; Löyttyniemi, E.; Solin, O.; Rinne, J.O.; Haaparanta-Solin, M. Brain energy metabolism and neuroinflammation in ageing APP/PS1-21 mice using longitudinal (18)F-FDG and (18)F-DPA-714 PET imaging. J Cereb Blood Flow Metab 2017, 37, 2870-2882. [CrossRef]
- Wu, Y.; Bottes, S.; Fisch, R.; Zehnder, C.; Cole, J.D.; Pilz, G.A.; Helmchen, F.; Simons, B.D.; Jessberger, S. Chronic in vivo imaging defines age-dependent alterations of neurogenesis in the mouse hippocampus. Nat Aging 2023, 3, 380-390. [CrossRef]
- Mathys, H.; Adaikkan, C.; Gao, F.; Young, J.Z.; Manet, E.; Hemberg, M.; De Jager, P.L.; Ransohoff, R.M.; Regev, A.; Tsai, L.H. Temporal Tracking of Microglia Activation in Neurodegeneration at Single-Cell Resolution. Cell Rep 2017, 21, 366-380. [CrossRef]
- Kreisl, W.C.; Kim, M.J.; Coughlin, J.M.; Henter, I.D.; Owen, D.R.; Innis, R.B. PET imaging of neuroinflammation in neurological disorders. Lancet Neurol 2020, 19, 940-950. [CrossRef]
- Tanaka, M. From Serendipity to Precision: Integrating AI, Multi-Omics, and Human-Specific Models for Personalized Neuropsychiatric Care. Biomedicines 2025, 13. [CrossRef]
- Kamei, R.; Urata, S.; Maruoka, H.; Okabe, S. In vivo Chronic Two-Photon Imaging of Microglia in the Mouse Hippocampus. J Vis Exp 2022. [CrossRef]
- Padmashri, R.; Tyner, K.; Dunaevsky, A. Implantation of a Cranial Window for Repeated In Vivo Imaging in Awake Mice. J Vis Exp 2021. [CrossRef]
- Xiong, H.; Tang, F.; Guo, Y.; Xu, R.; Lei, P. Neural circuit changes in neurological disorders: Evidence from in vivo two-photon imaging. Ageing Res Rev 2023, 87, 101933. [CrossRef]
- Ulivi, A.F.; Castello-Waldow, T.P.; Weston, G.; Yan, L.; Yasuda, R.; Chen, A.; Attardo, A. Longitudinal Two-Photon Imaging of Dorsal Hippocampal CA1 in Live Mice. J Vis Exp 2019. [CrossRef]
- Ren, W.; Ji, B.; Guan, Y.; Cao, L.; Ni, R. Recent Technical Advances in Accelerating the Clinical Translation of Small Animal Brain Imaging: Hybrid Imaging, Deep Learning, and Transcriptomics. Front Med (Lausanne) 2022, 9, 771982. [CrossRef]
- Guglielmetti, C.; Levi, J.; Huynh, T.L.; Tiret, B.; Blecha, J.; Tang, R.; VanBrocklin, H.; Chaumeil, M.M. Longitudinal Imaging of T Cells and Inflammatory Demyelination in a Preclinical Model of Multiple Sclerosis Using (18)F-FAraG PET and MRI. J Nucl Med 2022, 63, 140-146. [CrossRef]
- Best, L.; Ghadery, C.; Pavese, N.; Tai, Y.F.; Strafella, A.P. New and Old TSPO PET Radioligands for Imaging Brain Microglial Activation in Neurodegenerative Disease. Curr Neurol Neurosci Rep 2019, 19, 24. [CrossRef]
- Tamura, Y.; Takahashi, K.; Takata, K.; Eguchi, A.; Yamato, M.; Kume, S.; Nakano, M.; Watanabe, Y.; Kataoka, Y. Noninvasive Evaluation of Cellular Proliferative Activity in Brain Neurogenic Regions in Rats under Depression and Treatment by Enhanced [18F]FLT-PET Imaging. J Neurosci 2016, 36, 8123-8131. [CrossRef]
- Schwarz, C.G. Uses of Human MR and PET Imaging in Research of Neurodegenerative Brain Diseases. Neurotherapeutics 2021, 18, 661-672. [CrossRef]
- Mannheim, J.G.; Schmid, A.M.; Schwenck, J.; Katiyar, P.; Herfert, K.; Pichler, B.J.; Disselhorst, J.A. PET/MRI Hybrid Systems. Semin Nucl Med 2018, 48, 332-347. [CrossRef]
- Tanaka, M.; He, Z.; Han, S.; Battaglia, S. Editorial: Noninvasive brain stimulation: a promising approach to study and improve emotion regulation. Front Behav Neurosci 2025, 19, 1633936. [CrossRef]
- Tanaka, M.; Battaglia, S.; Liloia, D. Navigating Neurodegeneration: Integrating Biomarkers, Neuroinflammation, and Imaging in Parkinson's, Alzheimer's, and Motor Neuron Disorders. Biomedicines 2025, 13. [CrossRef]
- Valotto Neto, L.J.; Reverete de Araujo, M.; Moretti Junior, R.C.; Mendes Machado, N.; Joshi, R.K.; Dos Santos Buglio, D.; Barbalho Lamas, C.; Direito, R.; Fornari Laurindo, L.; Tanaka, M.; et al. Investigating the Neuroprotective and Cognitive-Enhancing Effects of Bacopa monnieri: A Systematic Review Focused on Inflammation, Oxidative Stress, Mitochondrial Dysfunction, and Apoptosis. Antioxidants (Basel) 2024, 13. [CrossRef]
- Chauveau, F.; Winkeler, A.; Chalon, S.; Boutin, H.; Becker, G. PET imaging of neuroinflammation: any credible alternatives to TSPO yet? Mol Psychiatry 2025, 30, 213-228. [CrossRef]
- Beaino, W.; Janssen, B.; Kooij, G.; van der Pol, S.M.A.; van Het Hof, B.; van Horssen, J.; Windhorst, A.D.; de Vries, H.E. Purinergic receptors P2Y12R and P2X7R: potential targets for PET imaging of microglia phenotypes in multiple sclerosis. J Neuroinflammation 2017, 14, 259. [CrossRef]
- Zhou, R.; Ji, B.; Kong, Y.; Qin, L.; Ren, W.; Guan, Y.; Ni, R. PET Imaging of Neuroinflammation in Alzheimer's Disease. Front Immunol 2021, 12, 739130. [CrossRef]
- Parker, C.A.; Nutt, D.J.; Tyacke, R.J. Imidazoline-I2 PET Tracers in Neuroimaging. Int J Mol Sci 2023, 24. [CrossRef]
- Zaharchuk, G. Next generation research applications for hybrid PET/MR and PET/CT imaging using deep learning. Eur J Nucl Med Mol Imaging 2019, 46, 2700-2707. [CrossRef]
- Aiello, M.; Cavaliere, C.; Fiorenza, D.; Duggento, A.; Passamonti, L.; Toschi, N. Neuroinflammation in Neurodegenerative Diseases: Current Multi-modal Imaging Studies and Future Opportunities for Hybrid PET/MRI. Neuroscience 2019, 403, 125-135. [CrossRef]
- Beaino, W.; Janssen, B.; Vugts, D.J.; de Vries, H.E.; Windhorst, A.D. Towards PET imaging of the dynamic phenotypes of microglia. Clin Exp Immunol 2021, 206, 282-300. [CrossRef]
- Chiu, F.Y.; Yen, Y. Imaging biomarkers for clinical applications in neuro-oncology: current status and future perspectives. Biomark Res 2023, 11, 35. [CrossRef]
- Tanaka, M. Special Issue "Translating Molecular Psychiatry: From Biomarkers to Personalized Therapies". Int J Mol Sci 2025, 26. [CrossRef]
- Fleischer, V.; Brummer, T.; Muthuraman, M.; Steffen, F.; Heldt, M.; Protopapa, M.; Schraad, M.; Gonzalez-Escamilla, G.; Groppa, S.; Bittner, S.; et al. Biomarker combinations from different modalities predict early disability accumulation in multiple sclerosis. Front Immunol 2025, 16, 1532660. [CrossRef]
- Vassal, M.; Martins, F.; Monteiro, B.; Tambaro, S.; Martinez-Murillo, R.; Rebelo, S. Emerging Pro-neurogenic Therapeutic Strategies for Neurodegenerative Diseases: A Review of Pre-clinical and Clinical Research. Mol Neurobiol 2025, 62, 46-76. [CrossRef]
- Praça, C.; Rai, A.; Santos, T.; Cristovão, A.C.; Pinho, S.L.; Cecchelli, R.; Dehouck, M.P.; Bernardino, L.; Ferreira, L.S. A nanoformulation for the preferential accumulation in adult neurogenic niches. J Control Release 2018, 284, 57-72. [CrossRef]
- Tanaka, M.; Tuka, B.; Vécsei, L. Navigating the Neurobiology of Migraine: From Pathways to Potential Therapies. Cells 2024, 13. [CrossRef]
- Li, X.; Wang, H.; Zhang, Q.; Sun, X.; Zhang, M.; Wang, G. Inhibition of adult hippocampal neurogenesis induced by postoperative CD8 + T-cell infiltration is associated with cognitive decline later following surgery in adult mice. J Neuroinflammation 2023, 20, 227. [CrossRef]
- Brocke, S.; Piercy, C.; Steinman, L.; Weissman, I.L.; Veromaa, T. Antibodies to CD44 and integrin alpha4, but not L-selectin, prevent central nervous system inflammation and experimental encephalomyelitis by blocking secondary leukocyte recruitment. Proc Natl Acad Sci U S A 1999, 96, 6896-6901. [CrossRef]
- Guo, J.; Tang, X.; Deng, P.; Hui, H.; Chen, B.; An, J.; Zhang, G.; Shi, K.; Wang, J.; He, Y.; et al. Interleukin-4 from curcumin-activated OECs emerges as a central modulator for increasing M2 polarization of microglia/macrophage in OEC anti-inflammatory activity for functional repair of spinal cord injury. Cell Commun Signal 2024, 22, 162. [CrossRef]
- Marchetti, B.; Tirolo, C.; L'Episcopo, F.; Caniglia, S.; Testa, N.; Smith, J.A.; Pluchino, S.; Serapide, M.F. Parkinson's disease, aging and adult neurogenesis: Wnt/β-catenin signalling as the key to unlock the mystery of endogenous brain repair. Aging Cell 2020, 19, e13101. [CrossRef]
- Zhu, R.; Zhu, X.; Zhu, Y.; Wang, Z.; He, X.; Wu, Z.; Xue, L.; Fan, W.; Huang, R.; Xu, Z.; et al. Immunomodulatory Layered Double Hydroxide Nanoparticles Enable Neurogenesis by Targeting Transforming Growth Factor-β Receptor 2. ACS Nano 2021, 15, 2812-2830. [CrossRef]
- Dubey, S.; Heinen, S.; Krantic, S.; McLaurin, J.; Branch, D.R.; Hynynen, K.; Aubert, I. Clinically approved IVIg delivered to the hippocampus with focused ultrasound promotes neurogenesis in a model of Alzheimer's disease. Proc Natl Acad Sci U S A 2020, 117, 32691-32700. [CrossRef]
- Bonetto, V.; Grilli, M. Neural stem cell-derived extracellular vesicles: mini players with key roles in neurogenesis, immunomodulation, neuroprotection and aging. Front Mol Biosci 2023, 10, 1187263. [CrossRef]
- Li, H.; Chen, G. In Vivo Reprogramming for CNS Repair: Regenerating Neurons from Endogenous Glial Cells. Neuron 2016, 91, 728-738. [CrossRef]
- Shinozaki, Y.; Shibata, K.; Yoshida, K.; Shigetomi, E.; Gachet, C.; Ikenaka, K.; Tanaka, K.F.; Koizumi, S. Transformation of Astrocytes to a Neuroprotective Phenotype by Microglia via P2Y(1) Receptor Downregulation. Cell Rep 2017, 19, 1151-1164. [CrossRef]
- Guo, Z.; Zhang, L.; Wu, Z.; Chen, Y.; Wang, F.; Chen, G. In vivo direct reprogramming of reactive glial cells into functional neurons after brain injury and in an Alzheimer's disease model. Cell Stem Cell 2014, 14, 188-202. [CrossRef]
- Xu, L.; Ramirez-Matias, J.; Hauptschein, M.; Sun, E.D.; Lunger, J.C.; Buckley, M.T.; Brunet, A. Restoration of neuronal progenitors by partial reprogramming in the aged neurogenic niche. Nat Aging 2024, 4, 546-567. [CrossRef]
- Zhang, J.; Rong, P.; Zhang, L.; He, H.; Zhou, T.; Fan, Y.; Mo, L.; Zhao, Q.; Han, Y.; Li, S.; et al. IL4-driven microglia modulate stress resilience through BDNF-dependent neurogenesis. Sci Adv 2021, 7. [CrossRef]
- Cai, B.; Seong, K.J.; Bae, S.W.; Kook, M.S.; Chun, C.; Lee, J.H.; Choi, W.S.; Jung, J.Y.; Kim, W.J. Water-Soluble Arginyl-Diosgenin Analog Attenuates Hippocampal Neurogenesis Impairment Through Blocking Microglial Activation Underlying NF-κB and JNK MAPK Signaling in Adult Mice Challenged by LPS. Mol Neurobiol 2019, 56, 6218-6238. [CrossRef]
- Yang, Y.; Ye, Y.; Kong, C.; Su, X.; Zhang, X.; Bai, W.; He, X. MiR-124 Enriched Exosomes Promoted the M2 Polarization of Microglia and Enhanced Hippocampus Neurogenesis After Traumatic Brain Injury by Inhibiting TLR4 Pathway. Neurochem Res 2019, 44, 811-828. [CrossRef]
- Qin, R.; Lai, X.; Xu, W.; Qin, Q.; Liang, X.; Xie, M.; Chen, L. The Mechanisms and Application Prospects of Astrocyte Reprogramming into Neurons in Central Nervous System Diseases. Curr Neuropharmacol 2025. [CrossRef]
- Zhang, L.; Yin, J.C.; Yeh, H.; Ma, N.X.; Lee, G.; Chen, X.A.; Wang, Y.; Lin, L.; Chen, L.; Jin, P.; et al. Small Molecules Efficiently Reprogram Human Astroglial Cells into Functional Neurons. Cell Stem Cell 2015, 17, 735-747. [CrossRef]
- Ma, Y.; Xie, H.; Du, X.; Wang, L.; Jin, X.; Zhang, Q.; Han, Y.; Sun, S.; Wang, L.; Li, X.; et al. In vivo chemical reprogramming of astrocytes into neurons. Cell Discov 2021, 7, 12. [CrossRef]
- Huang, L.; Lai, X.; Liang, X.; Chen, J.; Yang, Y.; Xu, W.; Qin, Q.; Qin, R.; Huang, X.; Xie, M.; et al. A promise for neuronal repair: reprogramming astrocytes into neurons in vivo. Biosci Rep 2024, 44. [CrossRef]
- Revuelta, M.; Urrutia, J.; Villarroel, A.; Casis, O. Microglia-Mediated Inflammation and Neural Stem Cell Differentiation in Alzheimer's Disease: Possible Therapeutic Role of K(V)1.3 Channel Blockade. Front Cell Neurosci 2022, 16, 868842. [CrossRef]
- Greșiță, A.; Hermann, D.M.; Boboc, I.K.S.; Doeppner, T.R.; Petcu, E.; Semida, G.F.; Popa-Wagner, A. Glial Cell Reprogramming in Ischemic Stroke: A Review of Recent Advancements and Translational Challenges. Transl Stroke Res 2025, 16, 1811-1835. [CrossRef]
- Feng, X.; Li, Z.; Liu, Y.; Chen, D.; Zhou, Z. CRISPR/Cas9 technology for advancements in cancer immunotherapy: from uncovering regulatory mechanisms to therapeutic applications. Exp Hematol Oncol 2024, 13, 102. [CrossRef]
- Tai, W.; Xu, X.M.; Zhang, C.L. Regeneration Through in vivo Cell Fate Reprogramming for Neural Repair. Front Cell Neurosci 2020, 14, 107. [CrossRef]
- Mahmoudi, N.; Wang, Y.; Moriarty, N.; Ahmed, N.Y.; Dehorter, N.; Lisowski, L.; Harvey, A.R.; Parish, C.L.; Williams, R.J.; Nisbet, D.R. Neuronal Replenishment via Hydrogel-Rationed Delivery of Reprogramming Factors. ACS Nano 2024, 18, 3597-3613. [CrossRef]
- Zhao, Z.; Wang, Y.; Zhou, R.; Li, Y.; Gao, Y.; Tu, D.; Wilson, B.; Song, S.; Feng, J.; Hong, J.S.; et al. A novel role of NLRP3-generated IL-1β in the acute-chronic transition of peripheral lipopolysaccharide-elicited neuroinflammation: implications for sepsis-associated neurodegeneration. J Neuroinflammation 2020, 17, 64. [CrossRef]
- Harrison, D.; Billinton, A.; Bock, M.G.; Doedens, J.R.; Gabel, C.A.; Holloway, M.K.; Porter, R.A.; Reader, V.; Scanlon, J.; Schooley, K.; et al. Discovery of Clinical Candidate NT-0796, a Brain-Penetrant and Highly Potent NLRP3 Inflammasome Inhibitor for Neuroinflammatory Disorders. J Med Chem 2023, 66, 14897-14911. [CrossRef]
- Ward, R.; Li, W.; Abdul, Y.; Jackson, L.; Dong, G.; Jamil, S.; Filosa, J.; Fagan, S.C.; Ergul, A. NLRP3 inflammasome inhibition with MCC950 improves diabetes-mediated cognitive impairment and vasoneuronal remodeling after ischemia. Pharmacol Res 2019, 142, 237-250. [CrossRef]
- Gordon, R.; Albornoz, E.A.; Christie, D.C.; Langley, M.R.; Kumar, V.; Mantovani, S.; Robertson, A.A.B.; Butler, M.S.; Rowe, D.B.; O'Neill, L.A.; et al. Inflammasome inhibition prevents α-synuclein pathology and dopaminergic neurodegeneration in mice. Sci Transl Med 2018, 10. [CrossRef]
- Dempsey, C.; Rubio Araiz, A.; Bryson, K.J.; Finucane, O.; Larkin, C.; Mills, E.L.; Robertson, A.A.B.; Cooper, M.A.; O'Neill, L.A.J.; Lynch, M.A. Inhibiting the NLRP3 inflammasome with MCC950 promotes non-phlogistic clearance of amyloid-β and cognitive function in APP/PS1 mice. Brain Behav Immun 2017, 61, 306-316. [CrossRef]
- Mackay, A.; Velcicky, J.; Gommermann, N.; Mattes, H.; Janser, P.; Wright, M.; Dubois, C.; Brenneisen, S.; Ilic, S.; Vangrevelinghe, E.; et al. Discovery of NP3-253, a Potent Brain Penetrant Inhibitor of the NLRP3 Inflammasome. J Med Chem 2024, 67, 20780-20798. [CrossRef]
- Kuwar, R.; Rolfe, A.; Di, L.; Blevins, H.; Xu, Y.; Sun, X.; Bloom, G.S.; Zhang, S.; Sun, D. A Novel Inhibitor Targeting NLRP3 Inflammasome Reduces Neuropathology and Improves Cognitive Function in Alzheimer's Disease Transgenic Mice. J Alzheimers Dis 2021, 82, 1769-1783. [CrossRef]
- Barbalho, S.M.; Laurindo, L.F.; de Oliveira Zanuso, B.; da Silva, R.M.S.; Gallerani Caglioni, L.; Nunes Junqueira de Moraes, V.B.F.; Fornari Laurindo, L.; Dogani Rodrigues, V.; da Silva Camarinha Oliveira, J.; Beluce, M.E.; et al. AdipoRon's Impact on Alzheimer's Disease-A Systematic Review and Meta-Analysis. Int J Mol Sci 2025, 26. [CrossRef]
- Xu, Y.; Yang, Y.; Chen, X.; Jiang, D.; Zhang, F.; Guo, Y.; Hu, B.; Xu, G.; Peng, S.; Wu, L.; et al. NLRP3 inflammasome in cognitive impairment and pharmacological properties of its inhibitors. Transl Neurodegener 2023, 12, 49. [CrossRef]
- McManus, R.M.; Latz, E. NLRP3 inflammasome signalling in Alzheimer's disease. Neuropharmacology 2024, 252, 109941. [CrossRef]
- Li, S.; Fang, Y.; Zhang, Y.; Song, M.; Zhang, X.; Ding, X.; Yao, H.; Chen, M.; Sun, Y.; Ding, J.; et al. Microglial NLRP3 inflammasome activates neurotoxic astrocytes in depression-like mice. Cell Rep 2022, 41, 111532. [CrossRef]
- Zhao, R.; Tian, X.; Xu, H.; Wang, Y.; Lin, J.; Wang, B. Aerobic Exercise Restores Hippocampal Neurogenesis and Cognitive Function by Decreasing Microglia Inflammasome Formation Through Irisin/NLRP3 Pathway. Aging Cell 2025, 24, e70061. [CrossRef]
- Vande Walle, L.; Lamkanfi, M. Drugging the NLRP3 inflammasome: from signalling mechanisms to therapeutic targets. Nat Rev Drug Discov 2024, 23, 43-66. [CrossRef]
- Mammoliti, O.; Carbajo, R.; Perez-Benito, L.; Yu, X.; Prieri, M.L.C.; Bontempi, L.; Embrechts, S.; Paesmans, I.; Bassi, M.; Bhattacharya, A.; et al. Discovery of Potent and Brain-Penetrant Bicyclic NLRP3 Inhibitors with Peripheral and Central In Vivo Activity. J Med Chem 2025, 68, 4848-4887. [CrossRef]
- Barczuk, J.; Siwecka, N.; Lusa, W.; Rozpędek-Kamińska, W.; Kucharska, E.; Majsterek, I. Targeting NLRP3-Mediated Neuroinflammation in Alzheimer's Disease Treatment. Int J Mol Sci 2022, 23. [CrossRef]
- Cai, R.; Lv, R.; Shi, X.; Yang, G.; Jin, J. CRISPR/dCas9 Tools: Epigenetic Mechanism and Application in Gene Transcriptional Regulation. Int J Mol Sci 2023, 24. [CrossRef]
- Liu, X.S.; Wu, H.; Ji, X.; Stelzer, Y.; Wu, X.; Czauderna, S.; Shu, J.; Dadon, D.; Young, R.A.; Jaenisch, R. Editing DNA Methylation in the Mammalian Genome. Cell 2016, 167, 233-247.e217. [CrossRef]
- O'Geen, H.; Ren, C.; Nicolet, C.M.; Perez, A.A.; Halmai, J.; Le, V.M.; Mackay, J.P.; Farnham, P.J.; Segal, D.J. dCas9-based epigenome editing suggests acquisition of histone methylation is not sufficient for target gene repression. Nucleic Acids Res 2017, 45, 9901-9916. [CrossRef]
- Vojta, A.; Dobrinić, P.; Tadić, V.; Bočkor, L.; Korać, P.; Julg, B.; Klasić, M.; Zoldoš, V. Repurposing the CRISPR-Cas9 system for targeted DNA methylation. Nucleic Acids Res 2016, 44, 5615-5628. [CrossRef]
- Lu, Y.; Brommer, B.; Tian, X.; Krishnan, A.; Meer, M.; Wang, C.; Vera, D.L.; Zeng, Q.; Yu, D.; Bonkowski, M.S.; et al. Reprogramming to recover youthful epigenetic information and restore vision. Nature 2020, 588, 124-129. [CrossRef]
- Nuñez, J.K.; Chen, J.; Pommier, G.C.; Cogan, J.Z.; Replogle, J.M.; Adriaens, C.; Ramadoss, G.N.; Shi, Q.; Hung, K.L.; Samelson, A.J.; et al. Genome-wide programmable transcriptional memory by CRISPR-based epigenome editing. Cell 2021, 184, 2503-2519.e2517. [CrossRef]
- Pattali, R.K.; Ornelas, I.J.; Nguyen, C.D.; Xu, D.; Divekar, N.S.; Nuñez, J.K. CRISPRoff epigenetic editing for programmable gene silencing in human cells without DNA breaks. bioRxiv 2024. [CrossRef]
- Yim, Y.Y.; Teague, C.D.; Nestler, E.J. In vivo locus-specific editing of the neuroepigenome. Nat Rev Neurosci 2020, 21, 471-484. [CrossRef]
- Xiong, K.; Wang, X.; Feng, C.; Zhang, K.; Chen, D.; Yang, S. Vectors in CRISPR Gene Editing for Neurological Disorders: Challenges and Opportunities. Adv Biol (Weinh) 2025, 9, e2400374. [CrossRef]
- Lee, H.; Rho, W.Y.; Kim, Y.H.; Chang, H.; Jun, B.H. CRISPR-Cas9 Gene Therapy: Non-Viral Delivery and Stimuli-Responsive Nanoformulations. Molecules 2025, 30. [CrossRef]
- Shang, J.; Song, F.; Zhang, Z.; Chen, D.; Yang, S. Application of novel CRISPR tools in brain therapy. Life Sci 2024, 352, 122855. [CrossRef]
- NodThera’s NLRP3 Inhibitor NT-0796 Reverses Neuroinflammation in Parkinson’s Disease Phase Ib/IIa Trial. Available online: https://www.nodthera.com/news/nodtheras-nlrp3-inhibitor-nt-0796-reverses-neuroinflammation-in-parkinsons-disease-phase-ib-iia-trial/#:~:text=NodThera%27s%20NLRP3%20Inhibitor%20NT,biomarkers%20in%20Parkinson%27s%20disease%20patients (accessed on 11.18).
- Moreno-Jiménez, E.P.; Terreros-Roncal, J.; Flor-García, M.; Rábano, A.; Llorens-Martín, M. Evidences for Adult Hippocampal Neurogenesis in Humans. J Neurosci 2021, 41, 2541-2553. [CrossRef]
- Arellano, J.I.; Rakic, P. Modelling adult neurogenesis in the aging rodent hippocampus: a midlife crisis. Front Neurosci 2024, 18, 1416460. [CrossRef]
- Darsalia, V.; Heldmann, U.; Lindvall, O.; Kokaia, Z. Stroke-induced neurogenesis in aged brain. Stroke 2005, 36, 1790-1795. [CrossRef]
- Simard, S.; Matosin, N.; Mechawar, N. Adult Hippocampal Neurogenesis in the Human Brain: Updates, Challenges, and Perspectives. Neuroscientist 2025, 31, 141-158. [CrossRef]
- Kuhn, H.G.; Toda, T.; Gage, F.H. Adult Hippocampal Neurogenesis: A Coming-of-Age Story. J Neurosci 2018, 38, 10401-10410. [CrossRef]
- Kempermann, G.; Gage, F.H.; Aigner, L.; Song, H.; Curtis, M.A.; Thuret, S.; Kuhn, H.G.; Jessberger, S.; Frankland, P.W.; Cameron, H.A.; et al. Human Adult Neurogenesis: Evidence and Remaining Questions. Cell Stem Cell 2018, 23, 25-30. [CrossRef]
- Holtman, I.R.; Raj, D.D.; Miller, J.A.; Schaafsma, W.; Yin, Z.; Brouwer, N.; Wes, P.D.; Möller, T.; Orre, M.; Kamphuis, W.; et al. Induction of a common microglia gene expression signature by aging and neurodegenerative conditions: a co-expression meta-analysis. Acta Neuropathol Commun 2015, 3, 31. [CrossRef]
- Hammond, T.R.; Dufort, C.; Dissing-Olesen, L.; Giera, S.; Young, A.; Wysoker, A.; Walker, A.J.; Gergits, F.; Segel, M.; Nemesh, J.; et al. Single-Cell RNA Sequencing of Microglia throughout the Mouse Lifespan and in the Injured Brain Reveals Complex Cell-State Changes. Immunity 2019, 50, 253-271.e256. [CrossRef]
- Flowers, A.; Bell-Temin, H.; Jalloh, A.; Stevens, S.M., Jr.; Bickford, P.C. Proteomic anaysis of aged microglia: shifts in transcription, bioenergetics, and nutrient response. J Neuroinflammation 2017, 14, 96. [CrossRef]
- Galatro, T.F.; Holtman, I.R.; Lerario, A.M.; Vainchtein, I.D.; Brouwer, N.; Sola, P.R.; Veras, M.M.; Pereira, T.F.; Leite, R.E.P.; Möller, T.; et al. Transcriptomic analysis of purified human cortical microglia reveals age-associated changes. Nat Neurosci 2017, 20, 1162-1171. [CrossRef]
- He, R.; Zhang, Q.; Wang, L.; Hu, Y.; Qiu, Y.; Liu, J.; You, D.; Cheng, J.; Cao, X. Exploring the feasibility of using mice as a substitute model for investigating microglia in aging and Alzheimer's disease though single cell analysis. PLoS One 2024, 19, e0311374. [CrossRef]
- Qi, C.; Yan, Y.; Cao, Q.; Zou, L.; Li, S.; Yang, Q.; Deng, Q.; Wu, B.; Song, B. Elucidating the mechanisms underlying astrocyte-microglia crosstalk in hippocampal neuroinflammation induced by acute diquat exposure. Environ Sci Pollut Res Int 2024, 31, 15746-15758. [CrossRef]
- Hou, B.; Zhang, Y.; Liang, P.; He, Y.; Peng, B.; Liu, W.; Han, S.; Yin, J.; He, X. Inhibition of the NLRP3-inflammasome prevents cognitive deficits in experimental autoimmune encephalomyelitis mice via the alteration of astrocyte phenotype. Cell Death Dis 2020, 11, 377. [CrossRef]
- Anderson, F.L.; Biggs, K.E.; Rankin, B.E.; Havrda, M.C. NLRP3 inflammasome in neurodegenerative disease. Transl Res 2023, 252, 21-33. [CrossRef]
- Adamczak, S.; Dale, G.; de Rivero Vaccari, J.P.; Bullock, M.R.; Dietrich, W.D.; Keane, R.W. Inflammasome proteins in cerebrospinal fluid of brain-injured patients as biomarkers of functional outcome: clinical article. J Neurosurg 2012, 117, 1119-1125. [CrossRef]
- Chou, V.; Pearse, R.V., 2nd; Aylward, A.J.; Ashour, N.; Taga, M.; Terzioglu, G.; Fujita, M.; Fancher, S.B.; Sigalov, A.; Benoit, C.R.; et al. INPP5D regulates inflammasome activation in human microglia. Nat Commun 2023, 14, 7552. [CrossRef]
- Ma, C.L.; Ma, X.T.; Wang, J.J.; Liu, H.; Chen, Y.F.; Yang, Y. Physical exercise induces hippocampal neurogenesis and prevents cognitive decline. Behav Brain Res 2017, 317, 332-339. [CrossRef]
- Figueiredo Godoy, A.C.; Frota, F.F.; Araújo, L.P.; Valenti, V.E.; Pereira, E.; Detregiachi, C.R.P.; Galhardi, C.M.; Caracio, F.C.; Haber, R.S.A.; Fornari Laurindo, L.; et al. Neuroinflammation and Natural Antidepressants: Balancing Fire with Flora. Biomedicines 2025, 13. [CrossRef]
- Mohd Sahini, S.N.; Mohd Nor Hazalin, N.A.; Srikumar, B.N.; Jayasingh Chellammal, H.S.; Surindar Singh, G.K. Environmental enrichment improves cognitive function, learning, memory and anxiety-related behaviours in rodent models of dementia: Implications for future study. Neurobiol Learn Mem 2024, 208, 107880. [CrossRef]
- Methi, A.; Islam, M.R.; Kaurani, L.; Sakib, M.S.; Krüger, D.M.; Pena, T.; Burkhardt, S.; Liebetanz, D.; Fischer, A. A Single-Cell Transcriptomic Analysis of the Mouse Hippocampus After Voluntary Exercise. Mol Neurobiol 2024, 61, 5628-5645. [CrossRef]
- Choi, S.H.; Bylykbashi, E.; Chatila, Z.K.; Lee, S.W.; Pulli, B.; Clemenson, G.D.; Kim, E.; Rompala, A.; Oram, M.K.; Asselin, C.; et al. Combined adult neurogenesis and BDNF mimic exercise effects on cognition in an Alzheimer's mouse model. Science 2018, 361. [CrossRef]
- Phillips, C. Lifestyle Modulators of Neuroplasticity: How Physical Activity, Mental Engagement, and Diet Promote Cognitive Health during Aging. Neural Plast 2017, 2017, 3589271. [CrossRef]
- Asthana, A.; Tripathi, S.; Agarwal, R. Role of Nonsteroidal Anti-Inflammatory Drugs as a Protective Factor in Alzheimer's Disease: A Systematic Review and Meta-Analysis. Neurol India 2024, 72, 1144-1151. [CrossRef]
- Rivers-Auty, J.; Mather, A.E.; Peters, R.; Lawrence, C.B.; Brough, D. Anti-inflammatories in Alzheimer's disease-potential therapy or spurious correlate? Brain Commun 2020, 2, fcaa109. [CrossRef]
- Cetin, A.; Komai, S.; Eliava, M.; Seeburg, P.H.; Osten, P. Stereotaxic gene delivery in the rodent brain. Nat Protoc 2006, 1, 3166-3173. [CrossRef]
- Kalincik, T.; Roos, I.; Sharmin, S. Observational studies of treatment effectiveness in neurology. Brain 2023, 146, 4799-4808. [CrossRef]
- Barnhart, A.J.; Dierickx, K. A Tale of Two Chimeras: Applying the Six Principles to Human Brain Organoid Xenotransplantation. Camb Q Healthc Ethics 2023, 1-17. [CrossRef]
- Erler, A. Human brain organoid transplantation: testing the foundations of animal research ethics. Neuroethics 2024, 17, 20.
- Neziri, S.; Köseoğlu, A.E.; Deniz Köseoğlu, G.; Özgültekin, B.; Özgentürk, N. Animal models in neuroscience with alternative approaches: Evolutionary, biomedical, and ethical perspectives. Animal Model Exp Med 2024, 7, 868-880. [CrossRef]
- Mrza, M.A.; He, J.; Wang, Y. Integration of iPSC-Derived Microglia into Brain Organoids for Neurological Research. Int J Mol Sci 2024, 25. [CrossRef]
- Schafer, S.T.; Mansour, A.A.; Schlachetzki, J.C.M.; Pena, M.; Ghassemzadeh, S.; Mitchell, L.; Mar, A.; Quang, D.; Stumpf, S.; Ortiz, I.S.; et al. An in vivo neuroimmune organoid model to study human microglia phenotypes. Cell 2023, 186, 2111-2126.e2120. [CrossRef]
- Ao, Z.; Cai, H.; Wu, Z.; Song, S.; Karahan, H.; Kim, B.; Lu, H.C.; Kim, J.; Mackie, K.; Guo, F. Tubular human brain organoids to model microglia-mediated neuroinflammation. Lab Chip 2021, 21, 2751-2762. [CrossRef]
- Pagotto, G.L.O.; Santos, L.; Osman, N.; Lamas, C.B.; Laurindo, L.F.; Pomini, K.T.; Guissoni, L.M.; Lima, E.P.; Goulart, R.A.; Catharin, V.; et al. Ginkgo biloba: A Leaf of Hope in the Fight against Alzheimer's Dementia: Clinical Trial Systematic Review. Antioxidants (Basel) 2024, 13. [CrossRef]
- Gonzales, M.M.; Garbarino, V.R.; Pollet, E.; Palavicini, J.P.; Kellogg, D.L., Jr.; Kraig, E.; Orr, M.E. Biological aging processes underlying cognitive decline and neurodegenerative disease. J Clin Invest 2022, 132. [CrossRef]
- Tanaka, M.; Vécsei, L. Revolutionizing our understanding of Parkinson's disease: Dr. Heinz Reichmann's pioneering research and future research direction. J Neural Transm (Vienna) 2024, 131, 1367-1387. [CrossRef]
- Rao, R.V.; Subramaniam, K.G.; Gregory, J.; Bredesen, A.L.; Coward, C.; Okada, S.; Kelly, L.; Bredesen, D.E. Rationale for a Multi-Factorial Approach for the Reversal of Cognitive Decline in Alzheimer's Disease and MCI: A Review. Int J Mol Sci 2023, 24. [CrossRef]
- Fekonja, L.S.; Forkel, S.J.; Aydogan, D.B.; Lioumis, P.; Cacciola, A.; Lucas, C.W.; Tournier, J.D.; Vergani, F.; Ritter, P.; Schenk, R.; et al. Translational network neuroscience: Nine roadblocks and possible solutions. Netw Neurosci 2025, 9, 352-370. [CrossRef]
- Barron, H.C.; Mars, R.B.; Dupret, D.; Lerch, J.P.; Sampaio-Baptista, C. Cross-species neuroscience: closing the explanatory gap. Philos Trans R Soc Lond B Biol Sci 2021, 376, 20190633. [CrossRef]
- Moreno-Gonzalez, I.; Garcia-Martin, J.; Marongiu, R. Editorial: Animal models of Alzheimer's disease and other dementias: past, present, and future. Front Aging Neurosci 2024, 16, 1539837. [CrossRef]
- Vitek, M.P.; Araujo, J.A.; Fossel, M.; Greenberg, B.D.; Howell, G.R.; Rizzo, S.J.S.; Seyfried, N.T.; Tenner, A.J.; Territo, P.R.; Windisch, M.; et al. Translational animal models for Alzheimer's disease: An Alzheimer's Association Business Consortium Think Tank. Alzheimers Dement (N Y) 2020, 6, e12114. [CrossRef]
- Sun, N.; Victor, M.B.; Park, Y.P.; Xiong, X.; Scannail, A.N.; Leary, N.; Prosper, S.; Viswanathan, S.; Luna, X.; Boix, C.A.; et al. Human microglial state dynamics in Alzheimer's disease progression. Cell 2023, 186, 4386-4403.e4329. [CrossRef]
- Szabó, Á.; Galla, Z.; Spekker, E.; Szűcs, M.; Martos, D.; Takeda, K.; Ozaki, K.; Inoue, H.; Yamamoto, S.; Toldi, J.; et al. Oxidative and Excitatory Neurotoxic Stresses in CRISPR/Cas9-Induced Kynurenine Aminotransferase Knockout Mice: A Novel Model for Despair-Based Depression and Post-Traumatic Stress Disorder. Front Biosci (Landmark Ed) 2025, 30, 25706. [CrossRef]
- Pluvinage, J.V.; Wyss-Coray, T. Systemic factors as mediators of brain homeostasis, ageing and neurodegeneration. Nat Rev Neurosci 2020, 21, 93-102. [CrossRef]
- Szabó, Á.; Galla, Z.; Spekker, E.; Martos, D.; Szűcs, M.; Fejes-Szabó, A.; Fehér, Á.; Takeda, K.; Ozaki, K.; Inoue, H.; et al. Behavioral Balance in Tryptophan Turmoil: Regional Metabolic Rewiring in Kynurenine Aminotransferase II Knockout Mice. Cells 2025, 14. [CrossRef]
- Higgins-Chen, A.T.; Thrush, K.L.; Levine, M.E. Aging biomarkers and the brain. Semin Cell Dev Biol 2021, 116, 180-193. [CrossRef]
- Sun, X.-Y.; Ju, X.-C.; Li, Y.; Zeng, P.-M.; Wu, J.; Zhou, Y.-Y.; Shen, L.-B.; Dong, J.; Chen, Y.-J.; Luo, Z.-G. Generation of vascularized brain organoids to study neurovascular interactions. elife 2022, 11, e76707.
- Martos, D.; Lőrinczi, B.; Szatmári, I.; Vécsei, L.; Tanaka, M. Decoupling Behavioral Domains via Kynurenic Acid Analog Optimization: Implications for Schizophrenia and Parkinson's Disease Therapeutics. Cells 2025, 14. [CrossRef]
- Lei, T.; Zhang, X.; Fu, G.; Luo, S.; Zhao, Z.; Deng, S.; Li, C.; Cui, Z.; Cao, J.; Chen, P.; et al. Advances in human cellular mechanistic understanding and drug discovery of brain organoids for neurodegenerative diseases. Ageing Res Rev 2024, 102, 102517. [CrossRef]
- Tanaka, M.; Battaglia, S.; Giménez-Llort, L.; Chen, C.; Hepsomali, P.; Avenanti, A.; Vécsei, L. Innovation at the Intersection: Emerging Translational Research in Neurology and Psychiatry. Cells 2024, 13. [CrossRef]
- Lonnemann, N.; Hosseini, S.; Marchetti, C.; Skouras, D.B.; Stefanoni, D.; D'Alessandro, A.; Dinarello, C.A.; Korte, M. The NLRP3 inflammasome inhibitor OLT1177 rescues cognitive impairment in a mouse model of Alzheimer's disease. Proc Natl Acad Sci U S A 2020, 117, 32145-32154. [CrossRef]
- Ngandu, T.; Lehtisalo, J.; Korkki, S.; Solomon, A.; Coley, N.; Antikainen, R.; Bäckman, L.; Hänninen, T.; Lindström, J.; Laatikainen, T.; et al. The effect of adherence on cognition in a multidomain lifestyle intervention (FINGER). Alzheimers Dement 2022, 18, 1325-1334. [CrossRef]
- Polis, B.; Samson, A.O. Addressing the Discrepancies Between Animal Models and Human Alzheimer's Disease Pathology: Implications for Translational Research. J Alzheimers Dis 2024, 98, 1199-1218. [CrossRef]
- Young, P.N.E.; Estarellas, M.; Coomans, E.; Srikrishna, M.; Beaumont, H.; Maass, A.; Venkataraman, A.V.; Lissaman, R.; Jiménez, D.; Betts, M.J.; et al. Imaging biomarkers in neurodegeneration: current and future practices. Alzheimers Res Ther 2020, 12, 49. [CrossRef]
- Wagatsuma, K.; Miwa, K.; Akamatsu, G.; Yamao, T.; Kamitaka, Y.; Sakurai, M.; Fujita, N.; Hanaoka, K.; Matsuda, H.; Ishii, K. Toward standardization of tau PET imaging corresponding to various tau PET tracers: a multicenter phantom study. Ann Nucl Med 2023, 37, 494-503. [CrossRef]
- Yang, L.; Han, B.; Zhang, Z.; Wang, S.; Bai, Y.; Zhang, Y.; Tang, Y.; Du, L.; Xu, L.; Wu, F.; et al. Extracellular Vesicle-Mediated Delivery of Circular RNA SCMH1 Promotes Functional Recovery in Rodent and Nonhuman Primate Ischemic Stroke Models. Circulation 2020, 142, 556-574. [CrossRef]
- Jüttner, J.; Szabo, A.; Gross-Scherf, B.; Morikawa, R.K.; Rompani, S.B.; Hantz, P.; Szikra, T.; Esposti, F.; Cowan, C.S.; Bharioke, A.; et al. Targeting neuronal and glial cell types with synthetic promoter AAVs in mice, non-human primates and humans. Nat Neurosci 2019, 22, 1345-1356. [CrossRef]
- Upadhya, R.; Madhu, L.N.; Attaluri, S.; Gitaí, D.L.G.; Pinson, M.R.; Kodali, M.; Shetty, G.; Zanirati, G.; Kumar, S.; Shuai, B.; et al. Extracellular vesicles from human iPSC-derived neural stem cells: miRNA and protein signatures, and anti-inflammatory and neurogenic properties. J Extracell Vesicles 2020, 9, 1809064. [CrossRef]
- Tanaka, M.; Szatmári, I.; Vécsei, L. Quinoline Quest: Kynurenic Acid Strategies for Next-Generation Therapeutics via Rational Drug Design. Pharmaceuticals (Basel) 2025, 18. [CrossRef]
- Giacomoni, J.; Åkerblom, M.; Habekost, M.; Fiorenzano, A.; Kajtez, J.; Davidsson, M.; Parmar, M.; Björklund, T. Identification and validation of novel engineered AAV capsid variants targeting human glia. Front Neurosci 2024, 18, 1435212. [CrossRef]
- Naeem, A.; Prakash, R.; Kumari, N.; Ali Khan, M.; Quaiyoom Khan, A.; Uddin, S.; Verma, S.; Ab Robertson, A.; Boltze, J.; Shadab Raza, S. MCC950 reduces autophagy and improves cognitive function by inhibiting NLRP3-dependent neuroinflammation in a rat model of Alzheimer's disease. Brain Behav Immun 2024, 116, 70-84. [CrossRef]
- Meneghini, V.; Peviani, M.; Luciani, M.; Zambonini, G.; Gritti, A. Delivery Platforms for CRISPR/Cas9 Genome Editing of Glial Cells in the Central Nervous System. Front Genome Ed 2021, 3, 644319. [CrossRef]
- Shi, L.; Li, S.; Zhu, R.; Lu, C.; Xu, X.; Li, C.; Huang, X.; Zhao, X.; Mao, F.; Li, K. CRISPRepi: a multi-omic atlas for CRISPR-based epigenome editing. Nucleic Acids Res 2025, 53, D901-d913. [CrossRef]
- Griñán-Ferré, C.; Bellver-Sanchis, A.; Guerrero, A.; Pallàs, M. Advancing personalized medicine in neurodegenerative diseases: The role of epigenetics and pharmacoepigenomics in pharmacotherapy. Pharmacol Res 2024, 205, 107247. [CrossRef]
- Ahmad, S.R.; Zeyaullah, M.; Khan, M.S.; AlShahrani, A.M.; Altijani, A.A.G.; Ali, H.; Dawria, A.; Mohieldin, A.; Alam, M.S.; Mohamed, A.O.A. Pharmacogenomics for neurodegenerative disorders - a focused review. Front Pharmacol 2024, 15, 1478964. [CrossRef]
- Deng, S.; Xie, H.; Xie, B. Cell-based regenerative and rejuvenation strategies for treating neurodegenerative diseases. Stem Cell Res Ther 2025, 16, 167. [CrossRef]
- Ueda, J.; Yamazaki, T.; Funakoshi, H. Toward the Development of Epigenome Editing-Based Therapeutics: Potentials and Challenges. Int J Mol Sci 2023, 24. [CrossRef]
- Khoshandam, M.; Soltaninejad, H.; Mousazadeh, M.; Hamidieh, A.A.; Hosseinkhani, S. Clinical applications of the CRISPR/Cas9 genome-editing system: Delivery options and challenges in precision medicine. Genes Dis 2024, 11, 268-282. [CrossRef]
- Pei, W.D.; Zhang, Y.; Yin, T.L.; Yu, Y. Epigenome editing by CRISPR/Cas9 in clinical settings: possibilities and challenges. Brief Funct Genomics 2020, 19, 215-228. [CrossRef]
- Tremblay, F.; Xiong, Q.; Shah, S.S.; Ko, C.W.; Kelly, K.; Morrison, M.S.; Giancarlo, C.; Ramirez, R.N.; Hildebrand, E.M.; Voytek, S.B.; et al. A potent epigenetic editor targeting human PCSK9 for durable reduction of low-density lipoprotein cholesterol levels. Nat Med 2025, 31, 1329-1338. [CrossRef]
- Gemberling, M.P.; Siklenka, K.; Rodriguez, E.; Tonn-Eisinger, K.R.; Barrera, A.; Liu, F.; Kantor, A.; Li, L.; Cigliola, V.; Hazlett, M.F.; et al. Transgenic mice for in vivo epigenome editing with CRISPR-based systems. Nat Methods 2021, 18, 965-974. [CrossRef]
- Wang, F.; Cheng, L.; Zhang, X. Reprogramming Glial Cells into Functional Neurons for Neuro-regeneration: Challenges and Promise. Neurosci Bull 2021, 37, 1625-1636. [CrossRef]
- Lentini, C.; d'Orange, M.; Marichal, N.; Trottmann, M.M.; Vignoles, R.; Foucault, L.; Verrier, C.; Massera, C.; Raineteau, O.; Conzelmann, K.K.; et al. Reprogramming reactive glia into interneurons reduces chronic seizure activity in a mouse model of mesial temporal lobe epilepsy. Cell Stem Cell 2021, 28, 2104-2121.e2110. [CrossRef]
- Matt, S.M.; Johnson, R.W. Neuro-immune dysfunction during brain aging: new insights in microglial cell regulation. Curr Opin Pharmacol 2016, 26, 96-101. [CrossRef]
- Filgueira, L.; Larionov, A.; Lannes, N. The Influence of Virus Infection on Microglia and Accelerated Brain Aging. Cells 2021, 10. [CrossRef]
- Flick, C.; Zamani, E.D.; Stahl, B.C.; Brem, A. The future of ICT for health and ageing: unveiling ethical and social issues through horizon scanning foresight. Technological Forecasting and Social Change 2020, 155, 119995.
- Hansson, O. Biomarkers for neurodegenerative diseases. Nat Med 2021, 27, 954-963. [CrossRef]
- Husain, M. Smarter adaptive platform clinical trials in neurology. Brain 2022, 145, 409-410. [CrossRef]
- Grill, J.D.; Karlawish, J. Implications of FDA Approval of a First Disease-Modifying Therapy for a Neurodegenerative Disease on the Design of Subsequent Clinical Trials. Neurology 2021, 97, 496-500. [CrossRef]
- Fumagalli, M.; Lombardi, M.; Gressens, P.; Verderio, C. How to reprogram microglia toward beneficial functions. Glia 2018, 66, 2531-2549. [CrossRef]
- Liang, T.; Zhang, Y.; Wu, S.; Chen, Q.; Wang, L. The Role of NLRP3 Inflammasome in Alzheimer's Disease and Potential Therapeutic Targets. Front Pharmacol 2022, 13, 845185. [CrossRef]
Table 1.
Molecular pathways linking inflammation and neurogenesis. Molecular pathways connecting inflammation and adult neurogenesis, summarizing key cytokines, chemokines, neurotrophic factors, and intracellular signaling hubs relevant to stem cell regulation in the adult brain. Each entry outlines the primary cellular source, directional impact on neural stem cell proliferation, differentiation, or survival, age-associated expression shifts, and potential therapeutic interventions or pharmacological modulators targeting the listed molecule or pathway [149] . Examples include IL-1β (microglial-derived negative regulator), TNFα (context-dependent neurogenic suppressor), CX3CL1/CX3CR1 (microglia–neuron communication axis), IGF-1 and BDNF (trophic pro-neurogenic drivers), as well as inflammasome nodes such as NLRP3. This reference table aims to support target discovery, translational prioritization, and mechanistic modeling across neuroinflammation-neuroplasticity research.
Table 1.
Molecular pathways linking inflammation and neurogenesis. Molecular pathways connecting inflammation and adult neurogenesis, summarizing key cytokines, chemokines, neurotrophic factors, and intracellular signaling hubs relevant to stem cell regulation in the adult brain. Each entry outlines the primary cellular source, directional impact on neural stem cell proliferation, differentiation, or survival, age-associated expression shifts, and potential therapeutic interventions or pharmacological modulators targeting the listed molecule or pathway [149] . Examples include IL-1β (microglial-derived negative regulator), TNFα (context-dependent neurogenic suppressor), CX3CL1/CX3CR1 (microglia–neuron communication axis), IGF-1 and BDNF (trophic pro-neurogenic drivers), as well as inflammasome nodes such as NLRP3. This reference table aims to support target discovery, translational prioritization, and mechanistic modeling across neuroinflammation-neuroplasticity research.
| Molecule/Pathway | Source / Cell Type | Effect on Neurogenesis | Relevance in Aging | Targeted by (if any) |
|---|---|---|---|---|
| IL-1β | Activated microglia | Inhibits NSC proliferation and newborn neuron survival; blocks maturation | Chronically elevated with NF-κB/NLRP3 activation; contributes to hostile niche | NLRP3 inhibitors (MCC950, NT-0796), anti-IL-1 drugs |
| TNF-α | Activated microglia | Suppresses progenitor proliferation and neuronal differentiation | Increased in microglial 'primed' states during inflammaging | TNF pathway blockers |
| IL-6 | Activated microglia/astrocytes | Reduces NSC proliferation; impairs plasticity | Elevated with chronic NF-κB/NLRP3 signaling | Anti-IL-6 agents (exploratory) |
| IFN-γ | Infiltrating CD8⁺ T cells; activated microglia | Suppresses NSC proliferation; antineurogenic bias | T-cell accumulation in aged niches; drives microglial priming | JAK/STAT inhibitors |
| NLRP3 inflammasome | Microglia | Sustains IL-1β/IL-18; locks antineurogenic programs | Persistently activated in aging; imprints epigenetic 'scars' | Brain-penetrant NLRP3 inhibitors |
| NF-κB | Microglia/astrocytes | Pro-inflammatory transcription; suppresses neurogenesis | Chronically active with oxidative stress; feeds cytokine loop | Pathway modulators (research) |
| Complement (C1q/C3) | Microglia/astrocytes | Accelerated pruning; survival loss of newborns | Heightened with chronic inflammatory tone | Complement inhibitors |
| CX3CL1–CX3CR1 | Neurons → microglia | Maintains microglial quiescence; supports maturation/integration | Protective tone wanes with age; disruption impairs neurogenesis | CX3CR1/CX3CL1 agonists |
| IGF-1 | Microglia, niche cells | Promotes NSC proliferation and survival | Declines with aging; part of youthful pro-neurogenic secretome | IGF-1 delivery/mimetics |
| BDNF / TrkB | Microglia, neurons | Enhances proliferation, maturation, survival; plasticity | Reduced availability under chronic inflammation | TrkB agonists; BDNF delivery |
| TGF-β | Microglia/astrocytes, niche | Context-dependent; supports homeostasis in youth | Elevated tonic signaling with age constrains neurogenesis | TGF-β tuning (local) |
| IL-10 | Microglia/astrocytes | Pro-neurogenic, supports integration | Protective signals decline with age | Cytokine augmentation |
| PI3K–Akt / ERK / Wnt–β-catenin | NSCs; microglia-modulated | Downstream pro-neurogenic cascades | Suppressed under inflammatory milieu | Small-molecule activators |
| CD8⁺ T-cell entry | Peripheral T cells | IFN-γ-mediated suppression of NSCs | Accumulate in aged SGZ/SVZ; feed-forward loop | Blockade of entry/adhesion |
| Extracellular vesicles (EVs) | Microglia/astrocytes/NSCs | Shift microglial phenotype; support neurogenesis | Therapeutic EVs restore youthful tone | EV-based miR/growth factor delivery |
Akt, Protein Kinase B (PKB); BDNF, Brain-Derived Neurotrophic Factor; C1q, Complement Component 1q; C3, Complement Component 3; CD8⁺, Cluster of Differentiation 8-Positive T Cell; CX3CL1, Chemokine (C-X3-C Motif) Ligand 1; CX3CR1, Chemokine (C-X3-C Motif) Receptor 1; ERK, Extracellular Signal-Regulated Kinase; EVs, Extracellular Vesicles; IFN-γ, Interferon-Gamma; IGF-1, Insulin-Like Growth Factor-1; IL-1β, Interleukin-1 Beta; IL-6, Interleukin-6; IL-10, Interleukin-10; JAK/STAT, Janus Kinase / Signal Transducer and Activator of Transcription; MCC950, NLRP3 Inflammasome Inhibitor (Small Molecule); NF-κB, Nuclear Factor Kappa-Light-Chain-Enhancer of Activated B Cells; NLRP3, NOD-Like Receptor Pyrin Domain-Containing Protein 3; NSC, Neural Stem Cell; NT-0796, Brain-Penetrant NLRP3 Inhibitor; PI3K, Phosphoinositide 3-Kinase; SGZ, Subgranular Zone; SVZ, Subventricular Zone; TGF-β, Transforming Growth Factor-Beta; TNF-α, Tumor Necrosis Factor-Alpha; TrkB, Tropomyosin Receptor Kinase B; Wnt, Wingless-Related Integration Site.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



