Submitted:
08 October 2025
Posted:
10 October 2025
You are already at the latest version
Abstract
Wheat (Triticum aestivum L.) is the most widely cultivated staple food crop globally. As a primary food source for 35-40% of the world's population, the stability of its yield is directly linked to global food security. However, extreme weather events triggered by climate change have led to wheat yield reduction, making it an urgent issue to enhance wheat's stress tolerance against drought and high temperatures. In this study, we successfully isolated and cloned an inositol oxidase gene from wheat. Further research revealed that high-temperature and drought stresses significantly increased the expression level of the TaMIOXA gene in wheat leaves. A batch of overexpressing lines was obtained via Agrobacterium-mediated transformation. Compared with the control group, wheat plants with molecularly modified TaMIOXA overexpression exhibited stronger resistance to high temperatures and drought, with their survival rate significantly increased by 10% to 40%. This confirms that overexpression of inositol oxidase significantly enhances wheat's tolerance to drought and high temperatures.
Keywords:
Wheat
; Inositol Oxidase
; Drought
; High Temperatures
; Stress Resistance
1. Introduction
Wheat (Triticum aestivum L.) is the most widely cultivated staple crop worldwide[1], spanning a broad latitudinal range from 60° N to 40° S and occupying 30.7 % of the global grain-cropping area[2]. It serves as a primary food source for 35–40 % of the world’s population[3], supplying approximately 20 % of humanity’s caloric and protein requirements[4]. Consequently, the stability of wheat yields is directly linked to global food security[5]. However, extreme weather events driven by climate change are threatening wheat production systems at an unprecedented rate [6]. Current studies indicate that for every 1 °C rise in global temperature, the environmental adaptability of existing wheat varieties declines by 8.7 %. High temperatures directly damage the photosynthetic apparatus of wheat leaves, disrupting their chloroplast structure [7] and reducing Rubisco activity by 55–70 %, which severely weakens carbon assimilation [8]. Moreover, stress-induced bursts of reactive oxygen species (ROS) compromise the cellular membrane system, trigger oxidative stress [9], and accelerate plant senescence [10]. In the water dimension, drought stress leads to abnormal stomatal opening and closing, markedly decreasing water use efficiency and thereby inhibiting nutrient uptake and material transport [11]. In addition, drought stress damages the cellular membrane system of crops [12], elevates the production of reactive oxygen species (ROS), and disrupts the balance between ROS generation and scavenging [13]. This leads to membrane lipid peroxidation and oxidative stress in plant cells, resulting in increased malondialdehyde (MDA) content and heightened cell-membrane permeability[14]. Moreover, drought stress inactivates membrane-associated proteins, enzymes, and other biomolecules, impairing the structure and function of the biofilm and, in severe cases, causing crop death[15].
Currently, research on plant resistance to abiotic stresses such as drought and high temperature has evolved into a multidimensional, multilevel systematic framework. Significant progress has been made in screening and functional analysis of related genes. For instance, TaSnRK2.10 enhances drought tolerance by phosphorylating TaERD15 and TaENO1[2], whereas TaSHN1 improves drought resistance by modulating leaf architecture without compromising yield[16]. In terms of hormonal and signaling regulation, progesterone contributes to the stability of photosystem II under heat stress[17]. Moreover, endoplasmic reticulum stress-related miRNAs are widely implicated in stress responses[18]. In rye and maize, exogenous application of 2,4-epibrassinolide and introduction of C4-type PEPC genes have been shown to markedly enhance photosynthetic performance and improve tolerance to high temperatures[6,19]. In addition, members of transcription-factor families such as MYB and SPL (e.g., TaMYB31 and TaSPL6) exert opposite regulatory effects on drought-response processes, with MYB acting as a positive regulator and SPL as a negative regulator[20,21]. Moreover, other crop genes—including maize ZmSCE1e, ZmERF21, and tobacco NtabDOG1L—enhance drought and heat tolerance through sumoylation modification and the regulation of hormone-signaling pathways and antioxidant mechanisms, respectively[22,23,24]. The down-regulation of the cytokinin receptor SlHK2 enhances tomato resistance to combined stresses[25], while the heat shock factor TaHsfA2d also participates in the response to phosphorus deficiency[26]. These studies systematically elucidate the complex regulatory network involving multiple species and genes in abiotic stress responses, and they provide a wealth of candidate genes and a solid theoretical foundation for the genetic improvement of wheat stress resistance.
Myo-inositol oxygenase (MIOX) is a pivotal metabolic enzyme that plays a crucial role in plant growth, development, and metabolism, particularly in response to abiotic stress[27]. As a key enzyme in the inositol metabolic pathway, MIOX regulates the cellular inositol pool, thereby influencing the synthesis of L-ascorbic acid (L-AsA) and the biosynthesis of cell-wall polysaccharides[28]. Moreover, MIOX significantly contributes to plant tolerance against various abiotic stresses, including drought, salinity, and low temperature. For example, overexpression of the MIOX gene in cotton enhances the plant’s drought and salt resistance[29], while its overexpression in Arabidopsis improves tolerance to a range of abiotic stresses[30]. Current studies have demonstrated that MIOX is associated with plant resistance across various species. Elevating the expression level of inositol oxidase enhances plant tolerance to abiotic stresses. Moreover, the expression pattern of the MIOX gene varies among different tissues and is induced by drought, salinity, and cold stress.
In view of the increasingly severe drought and high-temperature conditions that have affected Henan in recent years [31], we aimed to investigate the role of TaMIOX in wheat tolerance to these stresses. We selected Zhengmai 7698(郑麦7698), a principal cultivar in Henan, and carried out a 7-day drought treatment. Leaf samples from both the drought-stressed group and the well-watered control group were analyzed using quantitative reverse transcription PCR (RT qPCR). To broaden the genetic background, two additional wheat varieties—Yangmai 13(扬麦13) and Bainong 207(百农207)—were subsequently included. After 24 hours of exposure to high temperatures, the RT qPCR results demonstrated a marked upregulation of TaMIOX expression in wheat under both drought and heat stress. Here, total RNA was isolated from wheat leaves, and the myo-inositol oxygenase gene in the Triticum aestivum A-genome (TaMIOXA) coding sequence was amplified from reverse-transcribed cDNA. Finally, the pCAMBIA1302 TaMIOXA overexpression plasmid was constructed and introduced into three wheat genotypes. The resulting transgenic lines were then subjected to RT qPCR and leaf protein extraction, confirming that the pCAMBIA1302 TaMIOXA construct functions effectively in wheat. The transgenic lines overexpressing inositol oxidase and the corresponding control plants were subjected to simulated drought and high-temperature stress. The overexpression group displayed a markedly higher survival rate than the control group, demonstrating that elevated TaMIOX expression significantly enhances wheat tolerance to combined drought and heat stress.
2. Results
2.1. Gene Sequence Information of Inositol Oxidase
The coding sequence of TaMIOXA obtained from wheat was aligned with the reference sequence deposited in the public database (Figure S1). One nucleotide substitution was identified across all four wheat varieties. The phylogenetic tree was built using the retrieved CDS of TaMIOXA to elucidate its evolutionary relationship with other homologous genes (Figure 1).
2.2. Changes in Inositol Oxidase Transcription Level Under Drought and High-Temperature Stress
We first subjected the cultivar Zhengmai 7698 to the drought regimen and subsequently extracted total RNA from its leaves. Quantitative reverse transcription PCR (RT qPCR) analysis revealed that drought stress significantly upregulated the transcription of the inositol oxidase gene in Zhengmai 7698. Subsequently, two additional wheat cultivars—Yangmai 13 and Bainong 207—were subjected to 40 °C heat treatment for 24 h. Quantitative reverse transcription PCR (RT qPCR) analysis revealed that the abundant transcript of Ta MIOX was significantly upregulated in all three wheat varieties. This demonstrates that both drought and high-temperature stress markedly upregulated TaMIOX transcription in wheat leaves(Figure 2).
2.3. Construction of W
For the construction of the pCAMBIA1302-TaMIOXA vector plasmid map, RNA was isolated from wheat leaves, and the TaMIOXA coding sequence was amplified from reverse-transcribed cDNA. The pCAMBIA1302 TaMIOXA overexpression plasmid (Figure 3) was first assembled and subsequently introduced into Agrobacterium tumefaciens. Agrobacterium-mediated infection then yielded a batch of wheat plants transiently overexpressing TaMIOXA.
Next, total RNA was extracted from the leaves of three independent TaMIOXA overexpression wheat lines and subjected to RT-qPCR analysis. As shown in Figure 4, the expression level of TaMIOXA was significantly upregulated in transgenic lines compared to the wild-type control group.
Subsequently, leaf proteins from the three wheat genotypes were extracted and purified using a His tag affinity protocol. The purified proteins were then resolved using SDS PAGE (Figure 5). A distinct band corresponding to the expected molecular weight (~61.7 kDa) was observed in the supernatant of TaMIOX overexpression lines, whereas no detectable band appeared in the purified fractions from the control plants. These results confirm that the pCAMBIA1302 TaMIOXA construct is functionally expressed in wheat leaves.
2.4. Detection of Various Indicators of Overexpressing Strains
2.4.1. Phenotype of Drought and High-Temperature Treatment
After the initial characterization, the three wheat cultivars were grown under identical conditions for 73 days and then subjected to combined drought and high-temperature stress treatment for 7 days, followed by a 10-day recovery period with adequate irrigation (Figure 6). Survival rates were recorded and are summarized in Table 1. The inositol oxidase overexpression lines displayed markedly higher survival rates than their respective wild-type controls. Notably, the TaMIOXA overexpression line of Yangmai 13 achieved the highest survival rate (53.33 %), representing a 40 % increase relative to the control. Among the three cultivars, the TaMIOXA overexpression line of Zhengmai 7698 exhibited the most pronounced improvement in survival compared to its control group. The quantitative PCR analysis of the three wheat cultivars revealed that lines exhibiting high TaMIOXA expression displayed markedly enhanced tolerance to both drought and high-temperature stress. Among the tested varieties, the Zhengmai 7698 cultivar showed the most pronounced improvement in stress resistance.
2.4.2. Detection of Physiological Indexes
To further elucidate the role of TaMIOXA in the tolerance of wheat to high temperatures and drought, five plants were randomly selected from each treatment group after exposure to combined heat–drought stress. The leaves were then subjected to 3,3′-diaminobenzidine (DAB) staining, and the accumulation of hydrogen peroxide (H₂O₂) was examined microscopically (Figure 7). Through DAB staining observations [32], we found that the content of H2O2 in wheat leaves from the overexpression group was significantly lower than that of the control group after high-temperature and drought treatment.
The quantitative determination of H2O2 content before and after stress treatment corroborated the DAB-staining results. As shown in Figure 8, the H2O2 levels in all TaMIOXA-overexpression lines were significantly lower than the corresponding control plants. Specifically, the control plants Bainong 207, Yangmai 13, and Zhengmai 7698 exhibited H2O2 concentrations 1.04-, 1.19-, and 1.27-fold higher, respectively, than those of overexpression lines. Moreover, the relative accumulation of H2O2 (post-treatment versus pre-treatment) in the control groups was 1.42-fold, 1.02-fold, and 1.47-fold greater than that of the overexpression groups (Figure 8).
We also measured the catalase (CAT) activity of wheat before and after a 1-day drought and high-temperature treatment (Figure 9). As shown in Figure 9, the CAT activities of all TaMIOXA-overexpression lines were lower than those of the corresponding control plants after the stress. Specifically, the CAT activities in the control groups of Bainong 207, Yangmai 13, and Zhengmai 7698 were 1.13-, 1.08-, and 1.72-fold higher, respectively, than in the overexpression lines. Moreover, the increase in CAT activity from pre- to post-treatment was markedly greater in the control plants. The fold-increase in the control groups of Bainong 207, Yangmai 13, and Zhengmai 7698 was 1.58-, 1.11-, and 4.00-fold higher.
As illustrated in Figure 10, panel (a) displays the malondialdehyde (MDA) content in both control and TaMIOXA-overexpressing groups across three wheat varieties before and after combined drought and heat stress treatment. Panel (b) presents the cumulative MDA content values before and after treatment. Here, we found an interesting phenomenon, normal in the face of abiotic stress in plants: MDA content will gradually increase, accelerating cell death. However, we performed a one-day dry-heat treatment on three wheat varieties. It was found that compared with the control group, the MDA accumulation in the Yangmai 13 overexpression group decreased more. The MDA reduction in the control plants of Bainong 207 and Zhengmai 7698 was 1.67- and 1.83-fold greater, respectively, than that observed in the overexpression lines. This shows that the increase in MIOXA expression will reduce the content of MDA in wheat.
The detection results of glucuronic acid (GlcA) content in the control and TaMIOXA-overexpression groups (Figure 11) demonstrated that GlcA levels in all overexpression lines were lower than those in the corresponding controls. The reduction in GlcA in the control plants of Bainong 207, Yangmai 13, and Zhengmai 7698 was 1.09-, 1.05-, and 1.51-fold greater, respectively, than in the overexpression lines—an outcome that contradicts our initial hypothesis. Consequently, overexpression of TaMIOXA leads to a decrease in GlcA content.
In addition, the detection results of POD (peroxidase) and SOD (superoxide dismutase) showed that(Figure 12) the enzyme activity of POD is not significantly affected by TaMIOXA overexpression. The results of SOD enzyme activity were similar to those of CAT enzyme activity, and the increase in enzyme activity in the overexpression group was significantly lower than that in the control group. This indicates that the reaction substrate of TaMIOXA may partially overlap with the substrate of SOD, but the specific type of superoxide consumed requires further investigation.
3. Discussion
We isolated the TaMIOXA gene from four wheat cultivars—Chinese Spring, Zhengmai 7698, Yangmai 13, and Bainong 207. Compared with the reference sequence, all four isolates shared an identical nucleotide substitution that resulted in a single amino-acid change, converting tyrosine to cysteine at position 185 (Y185C). This mutation likely represents a novel natural variant that arose through regional or environmental selection. In addition, we designed distinct primers (Supplementary Material 2) for the PCR amplification of TaMIOXB (Triticum aestivum myo-inositol oxygenase B) and TaMIOXD (Triticum aestivum myo-inositol oxygenase D) based on the CDS sequences of TaMIOXA, TaMIOXB, and TaMIOXD. Despite numerous attempts, neither allele could be amplified from any developmental stage of the four wheat varieties.
In this study, we generated transient TaMIOXA-overexpressing lines in three wheat cultivars and subjected them to heat and drought stress. Our results demonstrate that TaMIOXA overexpression markedly enhances wheat tolerance to both drought and high temperatures; this effect is especially pronounced in Zhengmai 76978, a principal wheat variety cultivated in Henan Province. This demonstrates that up-regulating TaMIOXA expression offers a novel strategy for mitigating wheat yield losses caused by recent dry and hot wind events under increasingly extreme high-temperature and drought conditions. TaMIOXA can be used as a screening marker for new drought- and high-temperature-resistant wheat varieties. However, for the same overexpression strains, the RT-qPCR results obtained the lowest overexpression for Yangmai 13, which may be related to the breeding screening. The primary cultivation region of Yangmai 13 lies south of the Bainong 207 and Zhengmai 7698 zones, where temperatures are comparatively higher. Targeted breeding has endowed Yangmai 13 with enhanced tolerance to elevated temperatures, and its baseline expression of inositol oxidase is relatively high. This elevated expression may also explain why the control group of Yangmai 13 exhibited the greatest survival rate after 7-day combined drought and heat stress treatment. As such, inositol oxidase can serve as a novel screening marker for wheat varieties exhibiting high temperature and drought tolerance.
In addition, studies investigating which oxidant is consumed by plant inositol oxidase during its catalytic activity are scarce. The prevailing view is that plant inositol oxidase functions as an oxidase that utilizes molecular oxygen. Other radioisotope-labeling studies have demonstrated that, after 18O2 labeling, a portion of the oxygen atoms incorporated into the reaction product of glucuronic acid originates from molecular oxygen [33].
In this study, simultaneous monitoring of multiple wheat parameters during treatment allowed us to propose novel hypotheses concerning the oxidative substrates and stress-resistance mechanisms involved in the TaMIOXA reaction. After 1 day of treatment, the H2O2 levels in all overexpression groups were significantly lower than those in the control group. The cumulative amount of H2O2 in the overexpression groups was also comparatively reduced. On the contrary, the CAT enzyme activity of all overexpression wheat groups after drought and high-temperature treatment was lower than that of the control group. The level of CAT enzyme activity before and after treatment in the control group was significantly higher than that in the overexpression group. A comparison of cumulative H2O2 levels before and after wheat treatment indicates that the reduction in H2O2 accumulation observed in the overexpression lines is not attributable to catalase activity but is directly associated with the overexpression of TaMIOXA. TaMIOXA likely consumes H2O2 directly during the oxidation of inositol.
In this study, we employed the AlphaFold 3 online service to generate an ab initio model of TaMIOXA and subsequently performed molecular docking with AutoDock to systematically screen and investigate the oxidation substrates and their corresponding binding sites on TaMIOXA. The results are displayed in a Pymol map (Figure 13), and the corresponding hydrogen bond interaction network is marked. The mechanism of action and binding sites of inositol oxidase, inositol, and hydrogen peroxide were further predicted, as shown in Figure 13. The amino acids that have direct hydrogen bond interaction with inositol are serine (S103), aspartic acid (D104), histidine (H139), aspartic acid (D140), histidine (H212), histidine (H238), serine (S239), aspartic acid (D271), and lysine (K275). Arginine (R76), methionine (M208), aspartic acid (D214), and arginine (R236) all experience direct hydrogen bonding interactions with hydrogen peroxide.
We performed a one-day dry-heat treatment test on three wheat cultivars. Apart from the control group of Yangmai 13, which showed an increase in MDA accumulation, the MDA content decreased in all other wheat groups. The increase in TaMIOXA expression during the early phase of treatment could have enhanced ASA synthesis while consuming a portion of MDA. Yangmai 13, the principal cultivar grown in southern regions, exhibits enhanced tolerance to high-temperature stress. Consequently, the expression level of TaMIOXA in the control group of Yangmai 13 was low, and the consumption (depletion) of malondialdehyde (MDA) was reduced. Therefore, we infer that TaMIOXA enhances wheat tolerance to high temperatures and drought, both due to the scavenging of reactive oxygen species and the promotion of ASA synthesis and other related pathways.
Finally, we measured the glucuronic acid content in both the control and overexpression groups of the three wheat varieties. The results demonstrated that the overexpression lines had a significantly lower glucuronic acid concentration than the controls, with the most pronounced decrease observed in the Zhengmai 7698 cultivar. Studies have demonstrated that overexpression of the inositol oxidase (MIOX) gene in Arabidopsis thaliana [34], tomato (Solanum lycopersicum)[35], and cucumber (Cucumis sativus) [36] markedly enhances the content of ascorbic acid (AsA). Combining the phenotypic observations and indices of three wheat cultivars after dry-heat treatment, we hypothesize that, during the early stages of abiotic stress, the expression of inositol oxidase is upregulated. This enzyme scavenges accumulated reactive oxygen species (ROS) and produces glucuronic acid. Subsequently, a portion of glucuronic acid may be converted into ascorbic acid (ASA) via the ASA pathway, thereby reducing malondialdehyde (MDA) levels and enhancing the stress tolerance of wheat.
4. Materials and Methods
4.1. Isolation and Cloning of TaMIOXA
Total RNA was extracted from wheat leaves using the total RNA extraction kit TRIpure Reagent(Heruibio, Fujian, China)[37], and then, RNA was reverse-transcribed into cDNA using a HisScript RT premix kitHisScriptⅡQ RT superMix for qPCR (Vazyme, Nanjing, China) [38,39]. The primers were designed based on the nucleotide sequence of TaMIOXA. The cDNA was amplified by polymerase chain reaction (PCR) using 2 × Es Taq MasterMix (Dye). The PCR product was then recovered using agarose gel electrophoresis and purified with the HiPure Gel Pure DNA Mini Kit (Magen, Guangzhou, China). The recovered products were submitted for sequencing; the resulting reads were aligned using SnapeGene® 6.0.2 and subsequently translated into amino acid sequences [40].
4.2. Bioinformatics Analysis of TaMIOXA
The nucleotide sequence of TaMIOXA was translated into a protein sequence using SnapeGene ® 6.0.2 software. The protein structure of TaMIOXA was predicted using the AlphaFold Server (https://alphafoldserver.com/fold/5c40373bf5717a2e) provided by Google DeepMind. The platform is based on the AlphaFold3 model and can process various biomolecular inputs such as proteins and nucleic acids. The default parameters are used for prediction, and the custom template is not enabled. In order to perform molecular docking, this study obtained the three-dimensional structure files (format PDB) of hydrogen peroxide (H2O2) and superoxide anion (O2-) from the NCBI PubChem database (https://pubchem.ncbi.nlm.nih.gov/). The PubChem CIDs of the compounds were 784 and 5359597, respectively, and the structure of inositol was derived from ChemSpider (Inositol | C6H12O6).
The translated amino acid sequences were analyzed using BLAST on NCBI, and the sequences with significant homology were selected for download [41]. Subsequently, phylogenetic tree analysis was performed using MEGA 7.0 [42]. After that, we used Autodock4.26 to perform molecular docking on the protein prediction structure of TaMIOXA [43], and the results were displayed using Pymol [44].
4.3. Expression Analysis of the TaMIOXA Gene
According to the method described by Wang et al.[45], the expression of TaMIOXA was evaluated via RT-qPCR. Customized RT-qPCR primers (Table S1) were produced by targeting the conserved sequences of actin and VhMYB60 genes [46]. For the RT-qPCR process, we used the SYBR Green Realtime PCR premix from TaKaRa in Beijing, China [47] to ensure accurate and reliable results. The relative expression of TaMIOXA was determined using the 2-ΔΔCt method. The experiment was repeated using three techniques [48].
4.4. Preparation of Vector and Transformation of Wheat Plants
In this study, based on the coding-region sequence of the TaMIOXA gene and the sequence features of the pCAMBIA1302-GFP vector (Abcam, Hong Kong, China), NcoI and BglII were selected as restriction sites, and specific primers incorporating these sites were designed for homologous recombination experiments (Table S1). The pCAMBIA1302-GFP vector was linearized with NcoI and BglII Restriction Enzymes [49], and the two restriction sites together with the homologous arms were fused to the target gene via PCR. Finally, the overexpression vector was constructed by homologous recombination. Agrobacterium tumefaciens was ligated to the empty vector and the successfully ligated TaMIOXA-pCAMBIA1300-GFP expression vector. Subsequently, the constructed vector plasmid was transferred into Agrobacterium tumefaciens and transformed into wheat via Agrobacterium tumefaciens to obtain positive plants [50].
4.5. Plant Materials and Treatments
According to the planting conditions at different latitudes in the Huang-Huai wheat region, three wheat varieties were selected: Zhengmai 7698 (34°16′ N–36°22′ N), Yangmai 13 (29°–33° N), and Bainong 207 (31°23′ N–36°22′ N). From each variety, 150 seeds were chosen, primarily ovoid or elliptical, with intact morphology, a balanced seed-coat to endosperm ratio, and fine surface striations or hairs. After sterilization with hydrogen peroxide [51], the seeds were divided into two groups: a control group and a group subjected to Agrobacterium-mediated transformation. Following 2–3 days of germination, 50 healthy seeds from each treatment were transferred to cultivation pots, with 6–8 seeds per pot and 0.14 kg of dry soil per pot, which was then fully saturated with water. All treatment groups were maintained at 22 °C and 80 % ± 5 % relative humidity under a photoperiod of 16 h light/8 h dark and cultivated for 60–70 days. In each group, 30 healthy plants with complete roots, stems, and leaves were selected for stress exposure. After sampling, the samples were stored at –80 °C for further analysis.
4.6. Analysis of Physiological Indicators After Stress Overexpression of TaMIOXA in Wheat
Wheat plants in the overexpression groups (B207A, Y13A, Z7698A) and the control groups (B207CK, Y13CK, Z7698CK) were cultured in a light incubator for 10 weeks at 22 °C and 85 % relative humidity, under a 16 h light/8 h dark photoperiod. The overexpression and control groups were simultaneously subjected to drought and high-temperature stress for 7 days, with conditions of 40 °C and 50 % relative humidity for 14 h during the day, and 25 °C and 50 % relative humidity for 8 h at night. After drought and high-temperature stress, the plants were re-watered and allowed to recover for 10 days. During the stress and recovery periods, the growth condition of the wheat was recorded. Leaves were harvested, rapidly frozen in liquid nitrogen, and then stored at −80 °C for subsequent experiments. We quantified malondialdehyde (MDA) using the thiobarbituric acid (TBA) assay [6]. Hydrogen peroxide content was measured using the ammonium molybdate colorimetric method [52]; glucuronic acid was determined with high-performance liquid chromatography (HPLC); catalase (CAT) activity was assessed via UV spectrophotometry [10]; peroxidase (POD) activity was assessed using the guaiacol oxidation assay [53]; and superoxide dismutase (SOD) activity was assessed using the nitroblue tetrazolium (NBT) reduction assay [54], respectively.
5. Conclusions
In summary, we subjected wheat to drought and heat stress and observed that the transcription level of inositol oxidase significantly increased. To verify the role of inositol oxidase in wheat under abiotic stress, we generated three TaMIOXA-overexpressing wheat lines. After subjecting the plants to high-temperature and drought treatments, the results demonstrated that TaMIOXA overexpression markedly enhanced the tolerance of wheat to both heat and drought. The simultaneous detection of various physiological indices in wheat revealed that inositol oxidase enhances the plant’s stress resistance through the coordinated action of multiple pathways(Figure 14)
Under abiotic stress conditions, wheat plants accumulate reactive oxygen species (ROS), which upregulate the expression of inositol oxygenase. This enzyme consumes ROS (such as hydrogen peroxide) and myo-inositol to produce glucuronic acid [55]. The accumulated glucuronic acid is partially converted into cell wall components such as hemicellulose and pectin, while another portion serves as a precursor for ascorbic acid (ASA) synthesis [56]. This process activates the ASA biosynthetic pathway, ultimately leading to the production of ASA. AsA, together with superoxide dismutase (SOD), synergistically scavenges malondialdehyde (MDA) in plants [57]. This series of reactions maintains the plant’s physiological stability and improves its resistance to abiotic stress. Therefore, a synergistic interaction between the inositol metabolic pathway and other ASA biosynthetic pathways may modulate the ASA content in plants by influencing the expression or activity of related enzymes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Material. 1: Sequence alignment of reference TaMIOXA (TraesCS7A02G357800) and the PCR-amplified wheat sequence. Table S1: Primers used in this study.
Author Contributions
Conceptualization, Kun Cheng and Wenming Zheng; methodology, Kun Cheng; software, Sen Zhang and Shuaitao Huang; validation, Wenmei Zhou, Wenjing Qi ,Yimeng Liu and Zihan Zhang; formal analysis, Kun Cheng and Kunpu Zhang; investigation, Pengfei Shi; resources, Pengfei Shi and Yuhang Liu; data curation, Sen Zhang and Shuaitao Huang.; writing—original draft preparation, Sen Zhang; writing—review and editing, Kun Cheng and Wenming Zheng; project administration, Wenming Zheng; funding acquisition, Kun Cheng and Daowen Wang. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Sub-project of the National Key R&D Program of China, grant number 2021YFF1000203-12.
Data Availability Statement
The data are available in the Supplementary Materials.
Acknowledgments
We want to express our gratitude to the National Key R&D Program of China for funding this project.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| TaMIOXA | myo-inositol oxygenase gene in Triticum aestivum A-genome (Triticum urartu) |
| TaMIOXB | myo-inositol oxygenase gene in Triticum aestivum B-genome (Aegilops speltoides) |
| TaMIOXD | myo-inositol oxygenase gene in Triticum aestivum D-genome (Aegilops tauschii) |
| B207A | TaMIOXA overexpressing lines of wheat cultivar Bainong 207 |
| Y13207A | TaMIOXA overexpressing lines of wheat cultivar Yangmai 13 |
| Z7698A | TaMIOXA overexpressing lines of wheat cultivar Zhengmai 7698 |
| B207CK | control line of wheat cultivar Baonong 207 |
| Y13207CK | control line of wheat cultivar Yangmai 13 |
| Z7698CK | control line of wheat cultivar Zhengmai 7698 |
| CAT | catalase |
| SOD | superoxide dismutase |
| POD | peroxidase |
| GlcA | glucuronic acid |
| ASA | ascorbic acid |
| MDA | malondialdehyde |
References
- Wang, Y.; Yang, M.; Wei, S.; Qin, F.; Zhao, H.; Suo, B., Identification of Circular RNAs and Their Targets in Leaves of Triticum aestivum L. under Dehydration Stress. Front Plant Sci 2016, 7, 2024. [CrossRef]
- Zhang, Y.; Wang, J.; Li, Y.; Zhang, Z.; Yang, L.; Wang, M.; Zhang, Y.; Zhang, J.; Li, C.; Li, L.; Reynolds, M. P.; Jing, R.; Wang, C.; Mao, X., Wheat TaSnRK2.10 phosphorylates TaERD15 and TaENO1 and confers drought tolerance when overexpressed in rice. Plant Physiol 2023, 191, (2), 1344-1364. [CrossRef]
- Li, H.; Yu, M.; Zhu, X.; Nai, F.; Yang, R.; Wang, L.; Liu, Y.; Wei, Y.; Ma, X.; Yu, H.; Wang, X., TaGSr contributes to low-nitrogen tolerance by optimizing nitrogen uptake and assimilation in Arabidopsis. Environmental and Experimental Botany 2024, 219, 105657. [CrossRef]
- Reynolds, M. P.; Lewis, J. M.; Ammar, K.; Basnet, B. R.; Crespo-Herrera, L.; Crossa, J.; Dhugga, K. S.; Dreisigacker, S.; Juliana, P.; Karwat, H.; Kishii, M.; Krause, M. R.; Langridge, P.; Lashkari, A.; Mondal, S.; Payne, T.; Pequeno, D.; Pinto, F.; Sansaloni, C.; Schulthess, U.; Singh, R. P.; Sonder, K.; Sukumaran, S.; Xiong, W.; Braun, H. J., Harnessing translational research in wheat for climate resilience. J Exp Bot 2021, 72, (14), 5134-5157. [CrossRef]
- Rathor, P.; Rouleau, V.; Gorim, L. Y.; Chen, G.; Thilakarathna, M. S., Humalite enhances the growth, grain yield, and protein content of wheat by improving soil nitrogen availability and nutrient uptake. Journal of Plant Nutrition and Soil Science 2024, 187, (2), 247-259. [CrossRef]
- Zhao, G.; Xu, H.; Zhang, P.; Su, X.; Zhao, H., Effects of 2,4-epibrassinolide on photosynthesis and Rubisco activase gene expression in Triticum aestivum L. seedlings under a combination of drought and heat stress. Plant Growth Regulation 2017, 81, (3), 377-384. [CrossRef]
- Li, H.; Xu, H.; Zhang, P.; Gao, M.; Wang, D.; Zhao, H., High temperature effects on D1 protein turnover in three wheat varieties with different heat susceptibility. Plant Growth Regulation 2017, 81, (1), 1-9. [CrossRef]
- Perdomo, J. A.; Capó-Bauçà, S.; Carmo-Silva, E.; Galmés, J., Rubisco and Rubisco Activase Play an Important Role in the Biochemical Limitations of Photosynthesis in Rice, Wheat, and Maize under High Temperature and Water Deficit. Front Plant Sci 2017, 8, 490. [CrossRef]
- Liu, J.; Hasanuzzaman, M.; Wen, H.; Zhang, J.; Peng, T.; Sun, H.; Zhao, Q., High temperature and drought stress cause abscisic acid and reactive oxygen species accumulation and suppress seed germination growth in rice. Protoplasma 2019, 256, (5), 1217-1227. [CrossRef]
- Ding, H.; Ma, D.; Huang, X.; Hou, J.; Wang, C.; Xie, Y.; Wang, Y.; Qin, H.; Guo, T., Exogenous hydrogen sulfide alleviates salt stress by improving antioxidant defenses and the salt overly sensitive pathway in wheat seedlings. Acta Physiologiae Plantarum 2019, 41, (7), 123. [CrossRef]
- Guo, X.; Xin, Z.; Yang, T.; Ma, X.; Zhang, Y.; Wang, Z.; Ren, Y.; Lin, T., Metabolomics Response for Drought Stress Tolerance in Chinese Wheat Genotypes (Triticum aestivum). Plants (Basel) 2020, 9, (4). [CrossRef]
- Maseyk, K.; Lin, T.; Cochavi, A.; Schwartz, A.; Yakir, D., Quantification of leaf-scale light energy allocation and photoprotection processes in a Mediterranean pine forest under extensive seasonal drought. Tree Physiol 2019, 39, (10), 1767-1782. [CrossRef]
- Bouremani, N.; Cherif-Silini, H.; Silini, A.; Bouket, A. C.; Luptakova, L.; Alenezi, F. N.; Baranov, O.; Belbahri, L., Plant Growth-Promoting Rhizobacteria (PGPR): A Rampart against the Adverse Effects of Drought Stress. In Water, 2023; Vol. 15. [CrossRef]
- Zhang, L.; Yan, S.; Zhang, S.; Yan, P.; Wang, J.; Zhang, H., Glutathione, carbohydrate and other metabolites of Larix olgensis A. Henry reponse to polyethylene glycol-simulated drought stress. PLoS One 2021, 16, (11), e0253780. [CrossRef]
- Liu, F.; Zhao, Y.; Wang, X.; Wang, B.; Xiao, F.; He, K., Physiological response and drought resistance evaluation of Gleditsia sinensis seedlings under drought-rehydration state. Sci Rep 2023, 13, (1), 19963. [CrossRef]
- Bi, H.; Shi, J.; Kovalchuk, N.; Luang, S.; Bazanova, N.; Chirkova, L.; Zhang, D.; Shavrukov, Y.; Stepanenko, A.; Tricker, P.; Langridge, P.; Hrmova, M.; Lopato, S.; Borisjuk, N., Overexpression of the TaSHN1 transcription factor in bread wheat leads to leaf surface modifications, improved drought tolerance, and no yield penalty under controlled growth conditions. Plant Cell Environ 2018, 41, (11), 2549-2566. [CrossRef]
- Xue, R. L.; Wang, S. Q.; Xu, H. L.; Zhang, P. J.; Li, H.; Zhao, H. J., Progesterone increases photochemical efficiency of photosystem II in wheat under heat stress by facilitating D1 protein phosphorylation. Photosynthetica 2017, 55, (4), 664-670. [CrossRef]
- Chen, Y.; Yu, X., Endoplasmic reticulum stress-responsive microRNAs are involved in the regulation of abiotic stresses in wheat. Plant Cell Rep 2023, 42, (9), 1433-1452. [CrossRef]
- Qi, X.; Xu, W.; Zhang, J.; Guo, R.; Zhao, M.; Hu, L.; Wang, H.; Dong, H.; Li, Y., Physiological characteristics and metabolomics of transgenic wheat containing the maize C(4) phosphoenolpyruvate carboxylase (PEPC) gene under high temperature stress. Protoplasma 2017, 254, (2), 1017-1030. [CrossRef]
- Zhao, Y.; Cheng, X.; Liu, X.; Wu, H.; Bi, H.; Xu, H., The Wheat MYB Transcription Factor TaMYB(31) Is Involved in Drought Stress Responses in Arabidopsis. Front Plant Sci 2018, 9, 1426. [CrossRef]
- Zhao, Y.; He, J.; Liu, M.; Miao, J.; Ma, C.; Feng, Y.; Qian, J.; Li, H.; Bi, H.; Liu, W., The SPL transcription factor TaSPL6 negatively regulates drought stress response in wheat. Plant Physiol Biochem 2024, 206, 108264. [CrossRef]
- Dai, X.; Wang, Y.; Chen, Y.; Li, H.; Xu, S.; Yang, T.; Zhang, X.; Su, X.; Xia, Z., Overexpression of NtDOG1L-T Improves Heat Stress Tolerance by Modulation of Antioxidant Capability and Defense-, Heat-, and ABA-Related Gene Expression in Tobacco. Front Plant Sci 2020, 11, 568489. [CrossRef]
- Wang, H.; Wang, M.; Xia, Z., Overexpression of a maize SUMO conjugating enzyme gene (ZmSCE1e) increases Sumoylation levels and enhances salt and drought tolerance in transgenic tobacco. Plant Sci 2019, 281, 113-121. [CrossRef]
- Wang, Z.; Zhao, X.; Ren, Z.; Abou-Elwafa, S. F.; Pu, X.; Zhu, Y.; Dou, D.; Su, H.; Cheng, H.; Liu, Z.; Chen, Y.; Wang, E.; Shao, R.; Ku, L., ZmERF21 directly regulates hormone signaling and stress-responsive gene expression to influence drought tolerance in maize seedlings. Plant Cell Environ 2022, 45, (2), 312-328. [CrossRef]
- Mushtaq, N.; Wang, Y.; Fan, J.; Li, Y.; Ding, J., Down-Regulation of Cytokinin Receptor Gene SlHK2 Improves Plant Tolerance to Drought, Heat, and Combined Stresses in Tomato. Plants (Basel) 2022, 11, (2). [CrossRef]
- Zhao, Y.; Miao, J.; He, J.; Tian, X.; Gao, K.; Ma, C.; Tian, X.; Men, W.; Li, H.; Bi, H.; Liu, W., Wheat heat shock factor TaHsfA2d contributes to plant responses to phosphate deficiency. Plant Physiol Biochem 2022, 185, 178-187. [CrossRef]
- Alok, A.; Singh, S.; Kumar, P.; Bhati, K. K., Potential of engineering the myo-inositol oxidation pathway to increase stress resilience in plants. Mol Biol Rep 2022, 49, (8), 8025-8035. [CrossRef]
- Ivanov Kavkova, E.; Blöchl, C.; Tenhaken, R., The Myo-inositol pathway does not contribute to ascorbic acid synthesis. Plant Biol (Stuttg) 2019, 21 Suppl 1, (Suppl Suppl 1), 95-102. [CrossRef]
- Li, Z.; Liu, Z.; Wei, Y.; Liu, Y.; Xing, L.; Liu, M.; Li, P.; Lu, Q.; Peng, R., Genome-wide identification of the MIOX gene family and their expression profile in cotton development and response to abiotic stress. PLoS One 2021, 16, (7), e0254111. [CrossRef]
- Nepal, N.; Yactayo-Chang, J. P.; Medina-Jiménez, K.; Acosta-Gamboa, L. M.; González-Romero, M. E.; Arteaga-Vázquez, M. A.; Lorence, A., Mechanisms underlying the enhanced biomass and abiotic stress tolerance phenotype of an Arabidopsis MIOX over-expresser. Plant Direct 2019, 3, (9), e00165. [CrossRef]
- Flood and Drought Disaster Prevention and Control Division, D. o. W. R. o. H. P.; Li, L.; Junfeng, L.; Zhipeng, G.; Hao, S.; Di, W.; Rongrong, G., Henan Province: Precise measures to fully ensure the water supply demand for drought resistance, planting and seedling protection. China Flood & Drought Management 2024, 34, (8), 81-82.
- Carril, P.; da Silva, A. B.; Tenreiro, R.; Cruz, C., An Optimized in situ Quantification Method of Leaf H(2)O(2) Unveils Interaction Dynamics of Pathogenic and Beneficial Bacteria in Wheat. Front Plant Sci 2020, 11, 889. [CrossRef]
- Moskala, R.; Reddy, C. C.; Minard, R. D.; Hamilton, G. A., An oxygen-18 tracer investigation of the mechanism of myo-inositol oxygenase. Biochem Biophys Res Commun 1981, 99, (1), 107-13. [CrossRef]
- Lorence, A.; Chevone, B. I.; Mendes, P.; Nessler, C. L., myo-inositol oxygenase offers a possible entry point into plant ascorbate biosynthesis. Plant Physiol 2004, 134, (3), 1200-5. [CrossRef]
- Munir, S.; Mumtaz, M. A.; Ahiakpa, J. K.; Liu, G.; Chen, W.; Zhou, G.; Zheng, W.; Ye, Z.; Zhang, Y., Genome-wide analysis of Myo-inositol oxygenase gene family in tomato reveals their involvement in ascorbic acid accumulation. BMC Genomics 2020, 21, (1), 284. [CrossRef]
- Xue, Z. Main Pathway and Key Gene Contribution to the Increasing AsA Concentration in Cucumber Leaves under Nitrogen Deficiency. Chinese Academy of Agricultural Sciences, 2016.
- Tatineni, S.; Alexander, J.; Qu, F., Differential Synergistic Interactions Among Four Different Wheat-Infecting Viruses. Front Microbiol 2021, 12, 800318. [CrossRef]
- Wang, C.; Gan, J.; Mi, X., On four species of the genus Argiope Audouin, 1826 (Araneae, Araneidae) from China. Zookeys 2021, 1019, 15-34. [CrossRef]
- Yan, Y. L.; Zhang, C.; Hao, J.; Wang, X. L.; Ming, J.; Mi, L.; Na, J.; Hu, X.; Wang, Y., DPPA2/4 and SUMO E3 ligase PIAS4 opposingly regulate zygotic transcriptional program. PLoS Biol 2019, 17, (6), e3000324. [CrossRef]
- Wang, X.; Qin, Z.; Zhang, M.; Shang, B.; Li, Z.; Zhao, M.; Tang, Q.; Tang, Q.; Luo, J., Immunogenicity and protection of recombinant self-assembling ferritin-hemagglutinin nanoparticle influenza vaccine in mice. Clin Exp Vaccine Res 2025, 14, (1), 23-34. [CrossRef]
- Sayers, E. W.; Bolton, E. E.; Brister, J. R.; Canese, K.; Chan, J.; Comeau, D. C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; Madej, T.; Marchler-Bauer, A.; Lanczycki, C.; Lathrop, S.; Lu, Z.; Thibaud-Nissen, F.; Murphy, T.; Phan, L.; Skripchenko, Y.; Tse, T.; Wang, J.; Williams, R.; Trawick, B. W.; Pruitt, K. D.; Sherry, S. T., Database resources of the national center for biotechnology information. Nucleic Acids Res 2022, 50, (D1), D20-d26. [CrossRef]
- Tan, B.; Yan, L.; Li, H.; Lian, X.; Cheng, J.; Wang, W.; Zheng, X.; Wang, X.; Li, J.; Ye, X.; Zhang, L.; Li, Z.; Feng, J., Genome-wide identification of HSF family in peach and functional analysis of PpHSF5 involvement in root and aerial organ development. PeerJ 2021, 9, e10961. [CrossRef]
- Wu, Z.; Du, Y.; Li, Z.; Guo, R.; Li, Y.; Wei, J.; Yin, X.; Su, L., Soldier Caste-Specific Protein 1 Is Involved in Soldier Differentiation in Termite Reticulitermes aculabialis. Insects 2022, 13, (6). [CrossRef]
- Valdés-Tresanco, M. S.; Valdés-Tresanco, M. E.; Valiente, P. A.; Moreno, E., AMDock: a versatile graphical tool for assisting molecular docking with Autodock Vina and Autodock4. Biol Direct 2020, 15, (1), 12. [CrossRef]
- Wang, L.; Li, H.; Zhao, C.; Li, S.; Kong, L.; Wu, W.; Kong, W.; Liu, Y.; Wei, Y.; Zhu, J. K.; Zhang, H., The inhibition of protein translation mediated by AtGCN1 is essential for cold tolerance in Arabidopsis thaliana. Plant Cell Environ 2017, 40, (1), 56-68. [CrossRef]
- Galbiati, M.; Matus, J. T.; Francia, P.; Rusconi, F.; Cañón, P.; Medina, C.; Conti, L.; Cominelli, E.; Tonelli, C.; Arce-Johnson, P., The grapevine guard cell-related VvMYB60 transcription factor is involved in the regulation of stomatal activity and is differentially expressed in response to ABA and osmotic stress. BMC Plant Biol 2011, 11, 142. [CrossRef]
- Wang, Y.; Yang, K.; Bai, C.; Yin, D.; Li, G.; Qi, K.; Wang, G.; Li, Y., Development of a SYBR Green I real-time PCR for the detection of the orf virus. AMB Express 2017, 7, (1), 21. [CrossRef]
- Zhang, L.; Cui, X.; Yang, L.; Raziq, A.; Hao, S.; Zeng, W.; Huang, J.; Cao, Y.; Duan, Q., Nontransformation methods for studying signaling pathways and genes involved in Brassica rapa pollen-stigma interactions. Plant Physiol 2024, 196, (3), 1802-1812. [CrossRef]
- Zhu, X.; Rong, W.; Wang, K.; Guo, W.; Zhou, M.; Wu, J.; Ye, X.; Wei, X.; Zhang, Z., Overexpression of TaSTT3b-2B improves resistance to sharp eyespot and increases grain weight in wheat. Plant Biotechnol J 2022, 20, (4), 777-793. [CrossRef]
- Gao, Y.; Yang, J.; Duan, W.; Ma, X.; Qu, L.; Xu, Z.; Yang, Y.; Xu, J., NtRAV4 negatively regulates drought tolerance in Nicotiana tabacum by enhancing antioxidant capacity and defence system. Plant Cell Rep 2022, 41, (8), 1775-1788. [CrossRef]
- Davoudpour, Y.; Schmidt, M.; Calabrese, F.; Richnow, H. H.; Musat, N., High resolution microscopy to evaluate the efficiency of surface sterilization of Zea Mays seeds. PLoS One 2020, 15, (11), e0242247. [CrossRef]
- Ji, X.; Hou, H.; Wang, X.; Qiu, Y.; Ma, Y.; Wang, S.; Guo, S.; Huang, S.; Zhang, C., Effect of dietary Glycyrrhiza polysaccharides on growth performance, hepatic antioxidant capacity and anti-inflammatory capacity of broiler chickens. Res Vet Sci 2024, 167, 105114. [CrossRef]
- Wang, Y.; Li, X.; Liu, N.; Wei, S.; Wang, J.; Qin, F.; Suo, B., The iTRAQ-based chloroplast proteomic analysis of Triticum aestivum L. leaves subjected to drought stress and 5-aminolevulinic acid alleviation reveals several proteins involved in the protection of photosynthesis. BMC Plant Biol 2020, 20, (1), 96. [CrossRef]
- Su, X.; Fan, X.; Shao, R.; Guo, J.; Wang, Y.; Yang, J.; Yang, Q.; Guo, L., Physiological and iTRAQ-based proteomic analyses reveal that melatonin alleviates oxidative damage in maize leaves exposed to drought stress. Plant Physiol Biochem 2019, 142, 263-274. [CrossRef]
- Choudhury, F. K.; Rivero, R. M.; Blumwald, E.; Mittler, R., Reactive oxygen species, abiotic stress and stress combination. Plant J 2017, 90, (5), 856-867. [CrossRef]
- Eixelsberger, T.; Horvat, D.; Gutmann, A.; Weber, H.; Nidetzky, B., Isotope Probing of the UDP-Apiose/UDP-Xylose Synthase Reaction: Evidence of a Mechanism via a Coupled Oxidation and Aldol Cleavage. Angew Chem Int Ed Engl 2017, 56, (9), 2503-2507. [CrossRef]
- Ali, B.; Kumar, S.; Sui, X.; Niu, J.; Yang, J.; Zheng, M.; Tang, Y.; Li, H., Exogenous acetylsalicylic acid mitigates cold stress in common bean seedlings by enhancing antioxidant defense and photosynthetic efficiency. Front Plant Sci 2025, 16, 1589706. [CrossRef]
Figure 1.
A phylogenetic tree was generated using the wheat-derived TaMIOXA sequence.
Figure 2.
(a) Expression of TaMIOXA in Zhengmai 7698 after 5-day drought treatment. ( b) Expression levels of TaMIOXA in Bainong 207, Yangmai 13, and Zhengmai 7698 following 24-hour combined drought and high-temperature treatment for MIOXA overexpression lines.
Figure 2.
(a) Expression of TaMIOXA in Zhengmai 7698 after 5-day drought treatment. ( b) Expression levels of TaMIOXA in Bainong 207, Yangmai 13, and Zhengmai 7698 following 24-hour combined drought and high-temperature treatment for MIOXA overexpression lines.
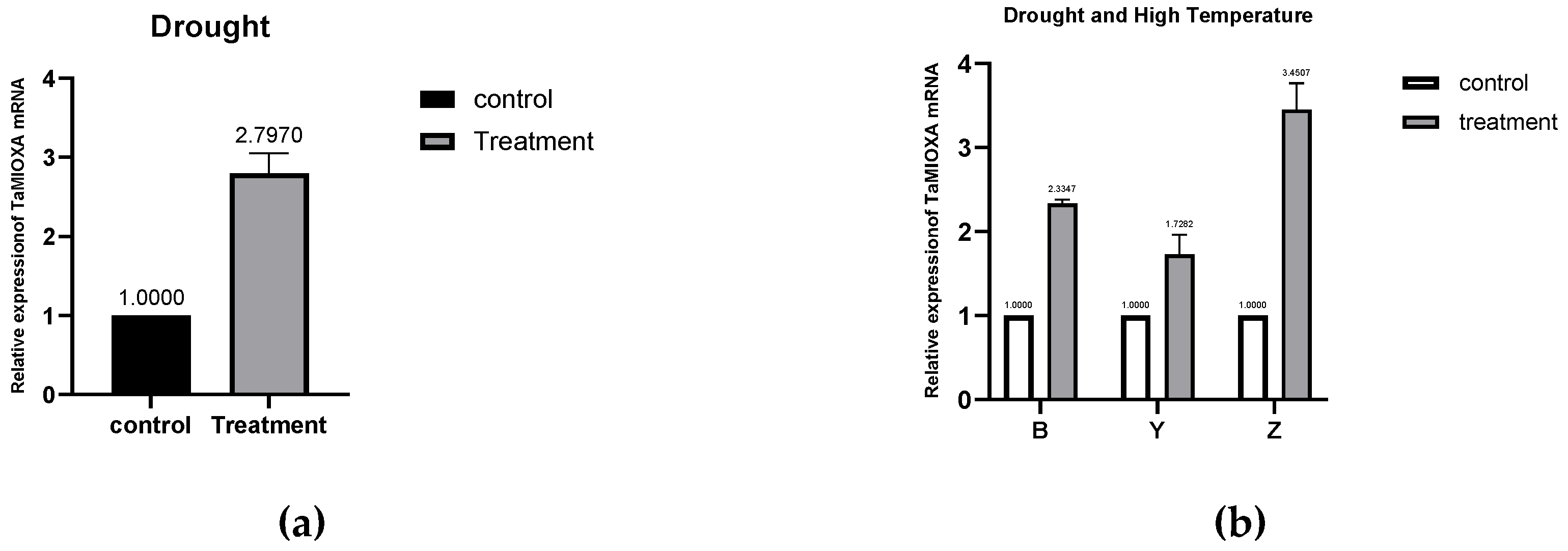
Figure 3.
pCAMBIA1302-TaMIOXA plasmid map.
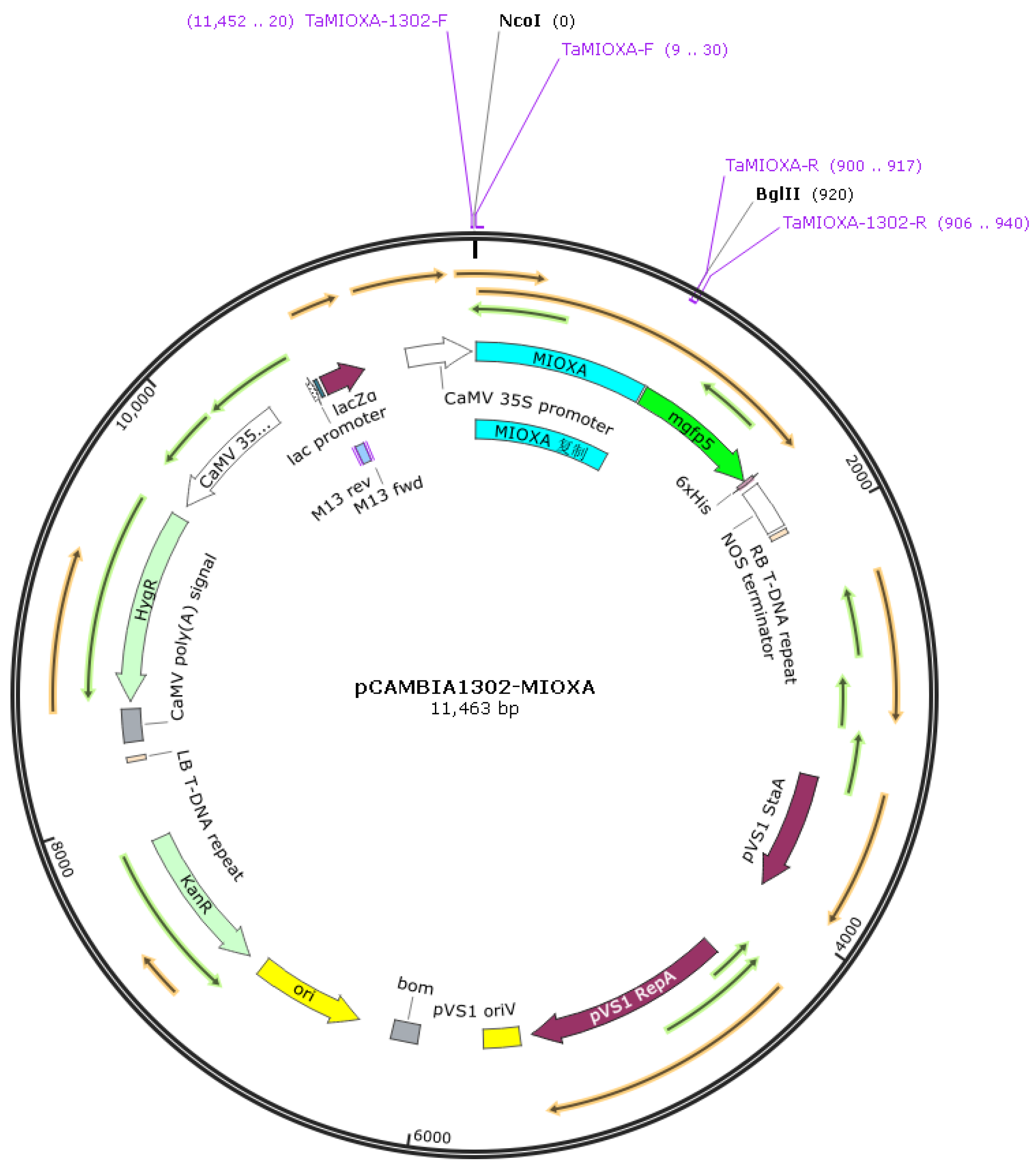
Figure 4.
(a) The expression level of TaMIOXA in the Bainong 207 overexpression strain. (b) The expression level of TaMIOXA in the Yangmai 13 overexpression strain. (c) The expression level of TaMIOXA in the Zhengmai 7698 overexpression strain.
Figure 4.
(a) The expression level of TaMIOXA in the Bainong 207 overexpression strain. (b) The expression level of TaMIOXA in the Yangmai 13 overexpression strain. (c) The expression level of TaMIOXA in the Zhengmai 7698 overexpression strain.
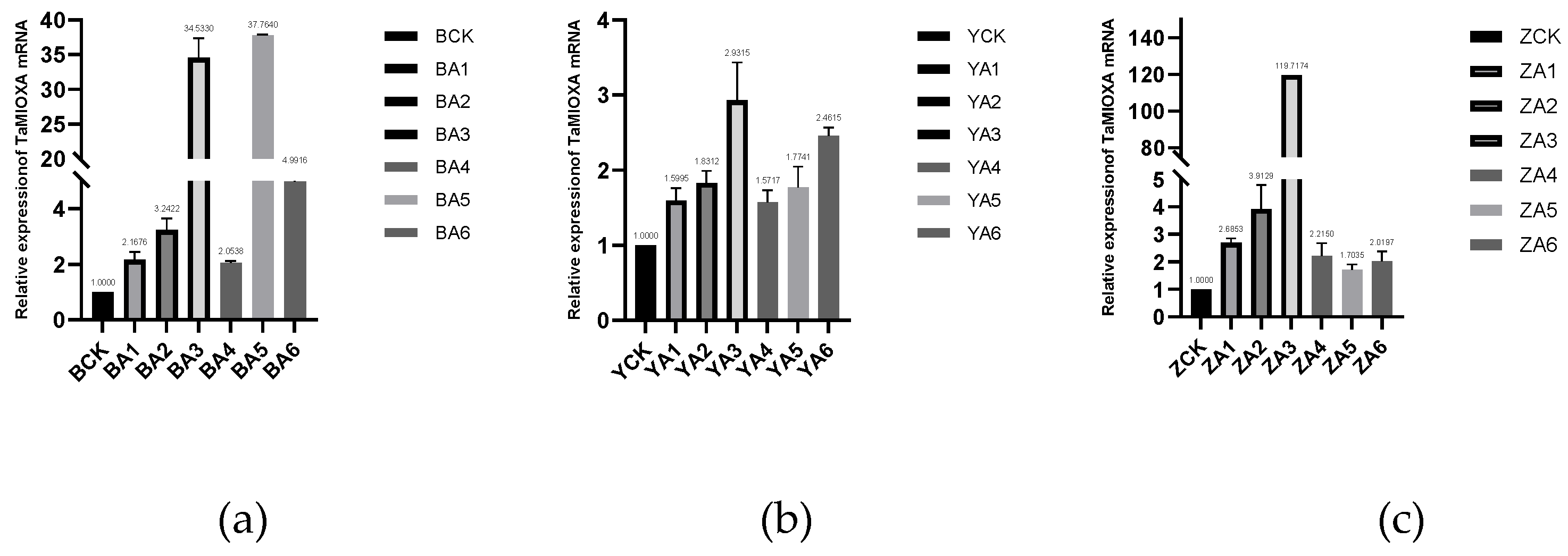
Figure 5.
SDS-PAGE electrophoresis results for the extraction of wheat leaf protein. (a) Bainong 207. (b) Yangmai 13. (c) Zhengmai 7698.
Figure 5.
SDS-PAGE electrophoresis results for the extraction of wheat leaf protein. (a) Bainong 207. (b) Yangmai 13. (c) Zhengmai 7698.

Figure 6.
Images of control and overexpression groups subjected to dry-heat treatment for 0 days and 7 days, followed by 10 days of recovery from rehydration. (a) Bainong 207. (b) Yangmai 13.(c)Zhengmai 7698. D0 (Day 0); D7 (Day 7); R10 (Recovery Day 10).
Figure 6.
Images of control and overexpression groups subjected to dry-heat treatment for 0 days and 7 days, followed by 10 days of recovery from rehydration. (a) Bainong 207. (b) Yangmai 13.(c)Zhengmai 7698. D0 (Day 0); D7 (Day 7); R10 (Recovery Day 10).

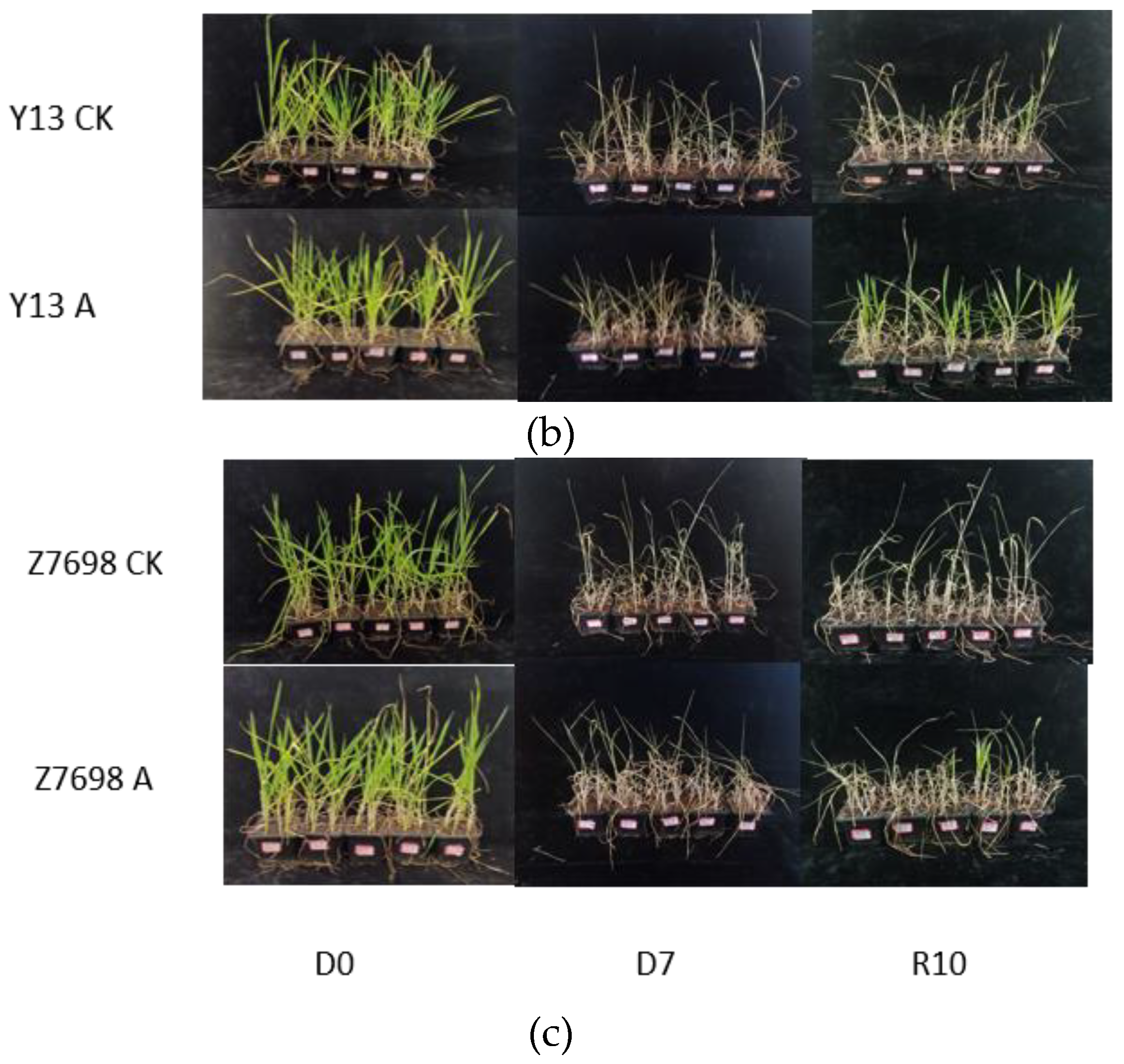
Figure 7.
The control and overexpression groups were subjected to 24 h dry-heat treatment, after which the leaves were examined using DAB staining. (a) Bainong 207. (b) Yangmai 13. (c) Zhengmai 7698.
Figure 7.
The control and overexpression groups were subjected to 24 h dry-heat treatment, after which the leaves were examined using DAB staining. (a) Bainong 207. (b) Yangmai 13. (c) Zhengmai 7698.

Figure 8.
The control and TaMIOXA-overexpression groups were subjected to 24 h dry-heat treatment, after which the above-ground tissues of the wheat plants were harvested for hydrogen peroxide (H₂O₂) quantification. (a) H₂O₂ content in the three wheat cultivars (Bainong 207, Yangmai 13, and Zhengmai 7698) for both control and overexpression lines, measured before and after the 24 h dry-heat stress test. (b) A cumulative change in H₂O₂ levels (post-treatment minus pre-treatment) for the same three cultivars, comparing control and overexpression groups after the 24 h dry-heat exposure test.
Figure 8.
The control and TaMIOXA-overexpression groups were subjected to 24 h dry-heat treatment, after which the above-ground tissues of the wheat plants were harvested for hydrogen peroxide (H₂O₂) quantification. (a) H₂O₂ content in the three wheat cultivars (Bainong 207, Yangmai 13, and Zhengmai 7698) for both control and overexpression lines, measured before and after the 24 h dry-heat stress test. (b) A cumulative change in H₂O₂ levels (post-treatment minus pre-treatment) for the same three cultivars, comparing control and overexpression groups after the 24 h dry-heat exposure test.
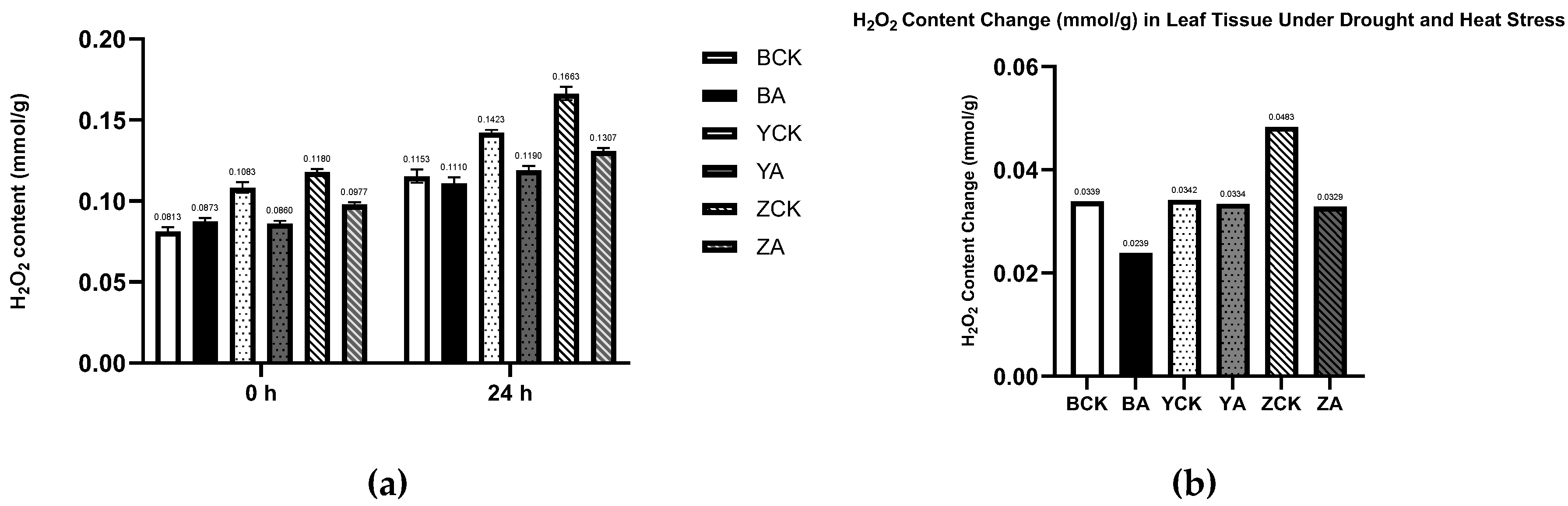
Figure 9.
The control and TaMIOXA-overexpression groups were subjected to a 24 h dry-heat treatment, after which the above-ground tissues of the wheat plants were harvested for catalase (CAT) activity measurement. (a) CAT activity in the three wheat cultivars (Bainong 207, Yangmai 13, and Zhengmai 7698) for both control and overexpression lines, measured before and after the 24 h dry-heat stress. (b) Cumulative changes in CAT activity (post-treatment minus pre-treatment) for the same three cultivars, comparing control and overexpression.
Figure 9.
The control and TaMIOXA-overexpression groups were subjected to a 24 h dry-heat treatment, after which the above-ground tissues of the wheat plants were harvested for catalase (CAT) activity measurement. (a) CAT activity in the three wheat cultivars (Bainong 207, Yangmai 13, and Zhengmai 7698) for both control and overexpression lines, measured before and after the 24 h dry-heat stress. (b) Cumulative changes in CAT activity (post-treatment minus pre-treatment) for the same three cultivars, comparing control and overexpression.
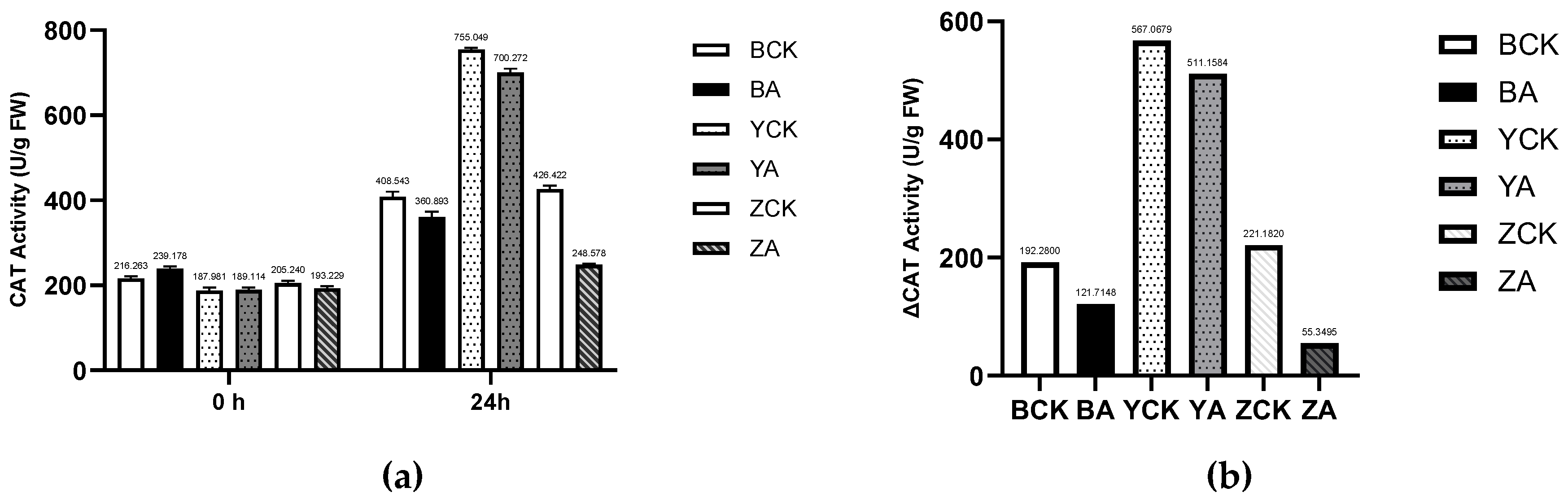
Figure 10.
The control and TaMIOXA-overexpression groups were subjected to 24 h dry-heat treatment, after which the above-ground tissues of the wheat plants were harvested for malondialdehyde (MDA) measurement. (a) MDA content in the three wheat cultivars (Bainong 207, Yangmai 13, and Zhengmai 7698) was measured for both control and overexpression lines, before and after the 24 h dry-heat stress test. (b) Net change in MDA levels (post-treatment minus pre-treatment) for the same three cultivars, comparing control and overexpression groups after 24 h of dry-heat exposure.
Figure 10.
The control and TaMIOXA-overexpression groups were subjected to 24 h dry-heat treatment, after which the above-ground tissues of the wheat plants were harvested for malondialdehyde (MDA) measurement. (a) MDA content in the three wheat cultivars (Bainong 207, Yangmai 13, and Zhengmai 7698) was measured for both control and overexpression lines, before and after the 24 h dry-heat stress test. (b) Net change in MDA levels (post-treatment minus pre-treatment) for the same three cultivars, comparing control and overexpression groups after 24 h of dry-heat exposure.
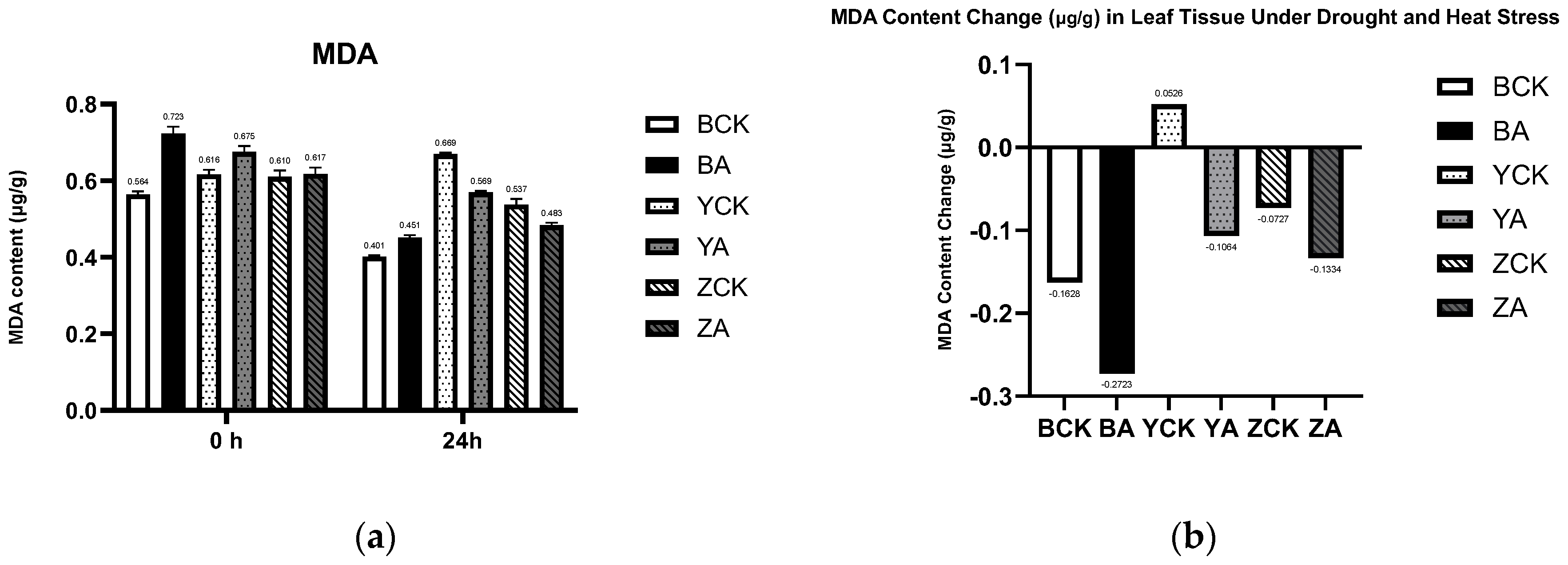
Figure 11.
The glucuronic acid (GlcA) content was measured in the above-ground tissues of wheat from both the control and TaMIOXA-overexpression groups.
Figure 11.
The glucuronic acid (GlcA) content was measured in the above-ground tissues of wheat from both the control and TaMIOXA-overexpression groups.

Figure 12.
Pl. The control and TaMIOXA-overexpression groups were subjected to 24 h dry-heat treatment, after which the above-ground tissues of the wheat plants were harvested for peroxidase (POD) and superoxide dismutase (SOD) activity assays. (a) POD activity in the three wheat cultivars (Bainong 207, Yangmai 13, and Zhengmai 7698) for both control and overexpression lines, measured before and after the 24 h dry-heat stress test. (b) The net change in POD activity (post-treatment minus pre-treatment) for the same three cultivars, comparing control and overexpression groups after 24 h dry-heat exposure. (c) SOD activity in the three wheat cultivars for control and overexpression lines, measured before and after the 24 h dry-heat treatment test. (d) The net change in SOD activity (post-treatment minus pre-treatment) for the three cultivars, comparing control and overexpression groups, after the 24 h dry-heat exposure test.
Figure 12.
Pl. The control and TaMIOXA-overexpression groups were subjected to 24 h dry-heat treatment, after which the above-ground tissues of the wheat plants were harvested for peroxidase (POD) and superoxide dismutase (SOD) activity assays. (a) POD activity in the three wheat cultivars (Bainong 207, Yangmai 13, and Zhengmai 7698) for both control and overexpression lines, measured before and after the 24 h dry-heat stress test. (b) The net change in POD activity (post-treatment minus pre-treatment) for the same three cultivars, comparing control and overexpression groups after 24 h dry-heat exposure. (c) SOD activity in the three wheat cultivars for control and overexpression lines, measured before and after the 24 h dry-heat treatment test. (d) The net change in SOD activity (post-treatment minus pre-treatment) for the three cultivars, comparing control and overexpression groups, after the 24 h dry-heat exposure test.
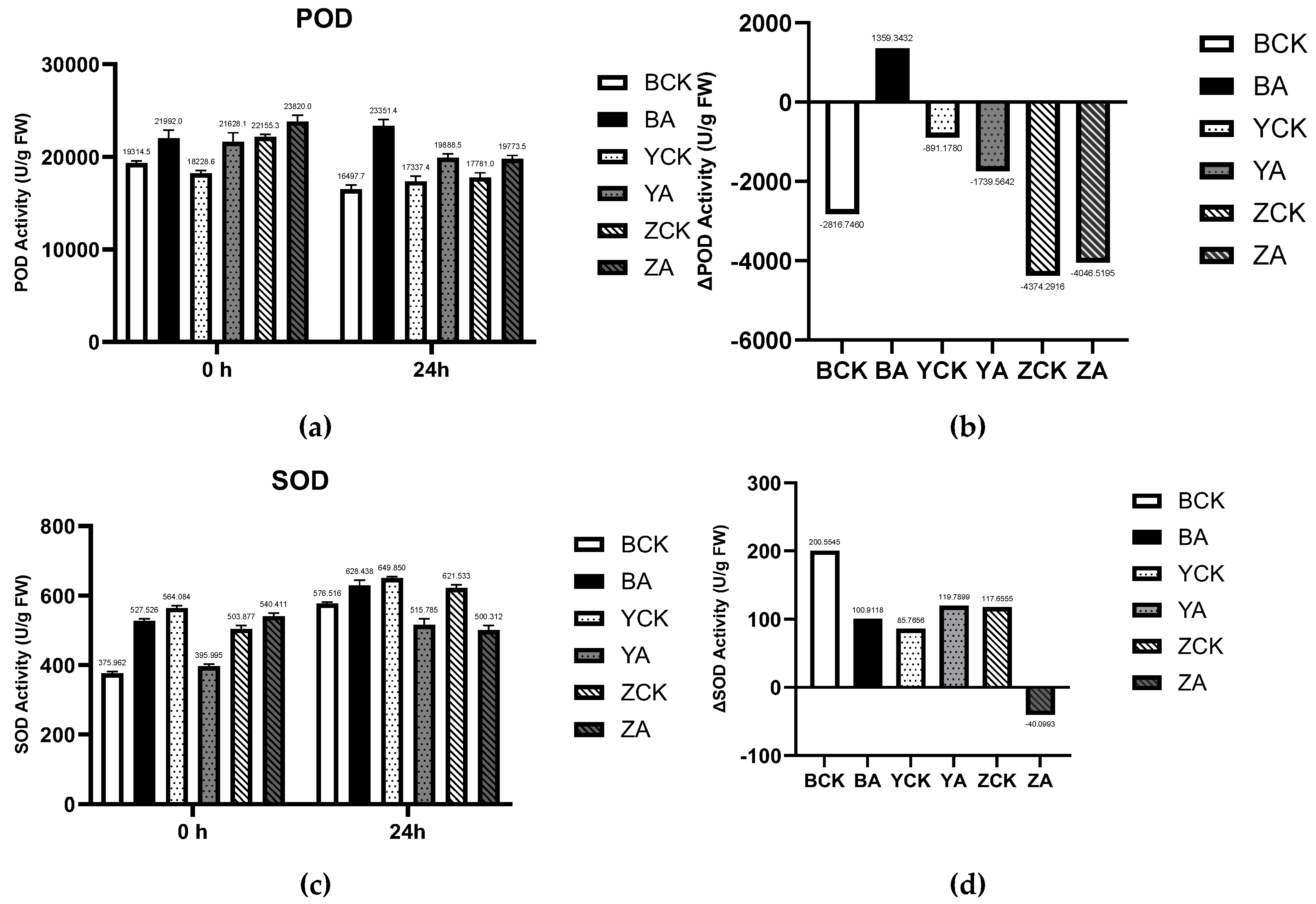
Figure 13.
Molecular docking results of the TaMIOXA protein. (a) Protein modeling results of inositol oxygenase generated by AlphaFold 3. (b) The hydrogen bonding network from the docking of inositol oxygenase with inositol. (c) The hydrogen bonding network from the docking of inositol oxygenase with hydrogen peroxide.
Figure 13.
Molecular docking results of the TaMIOXA protein. (a) Protein modeling results of inositol oxygenase generated by AlphaFold 3. (b) The hydrogen bonding network from the docking of inositol oxygenase with inositol. (c) The hydrogen bonding network from the docking of inositol oxygenase with hydrogen peroxide.

Figure 14.
Myo-inositol oxygenase pathway and hypothesized related pathways under drought and high-temperature stress.
Figure 14.
Myo-inositol oxygenase pathway and hypothesized related pathways under drought and high-temperature stress.
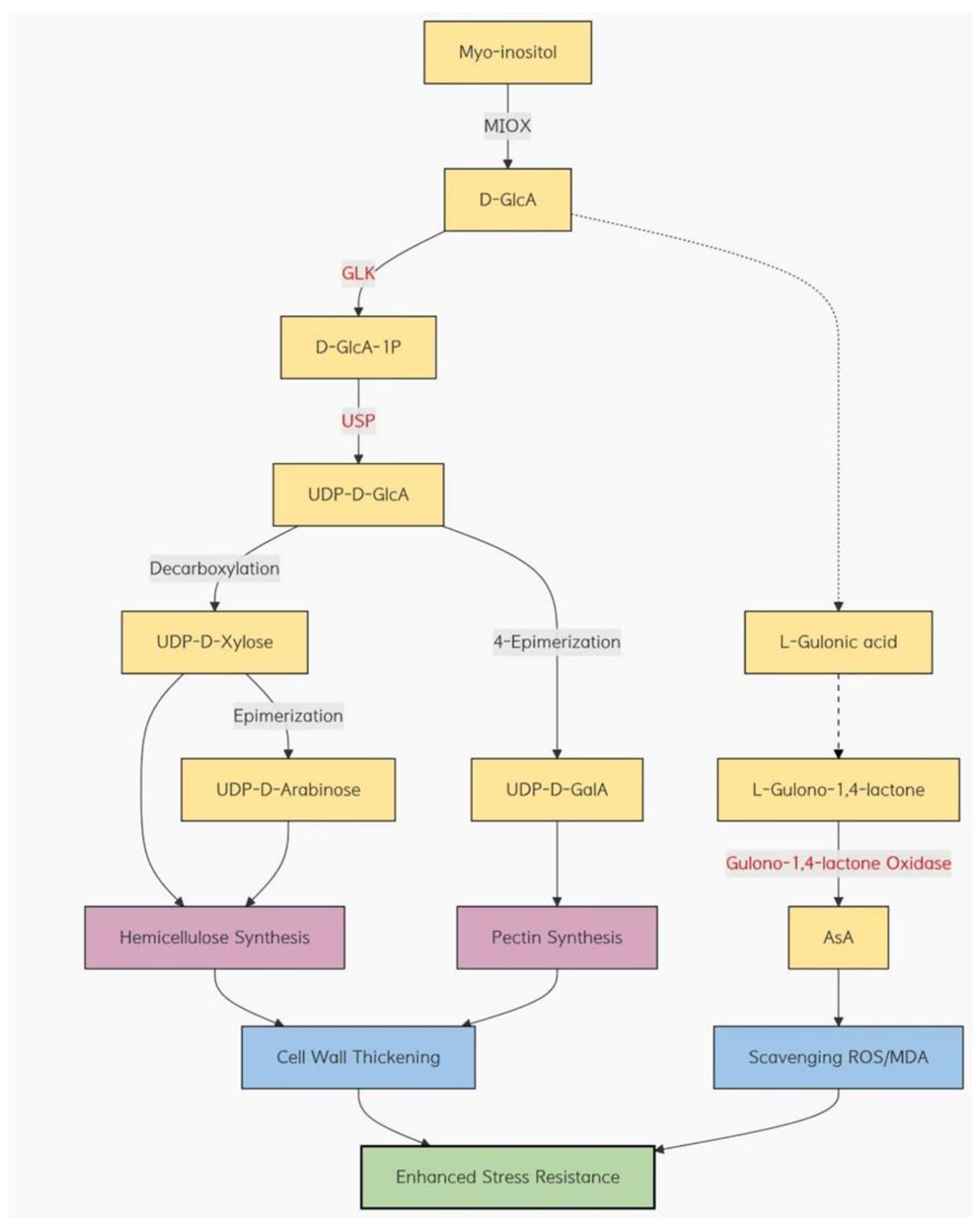

Table 1.
The recovery of wheat under stress conditions.
| Survival Count (Plants) | Initial Count (Plants) | Survival Rate (%) | ||
|---|---|---|---|---|
| CK | B207 CK | 2 | 30 | 6.67 |
| Y13 CK | 4 | 30 | 13.33 | |
| Z7698 CK | 0 | 30 | 0 | |
| OE-TaMIOX-A | B207 A | 5 | 30 | 16.67 |
| Y13 A | 16 | 30 | 53.33 | |
| Z7698 A | 12 | 30 | 40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



