
Submitted:
09 October 2025
Posted:
09 October 2025
You are already at the latest version
Abstract
Triple-negative breast cancer (TNBC) has the poorest prognosis among all breast cancer subtypes, largely due to the lack of targeted therapies and its resistance to both chemotherapy and immunotherapy. A deeper understanding of TNBC biology is therefore critical for identifying therapeutic targets. Molecular subtyping of TNBC, first introduced over a decade ago, has significantly advanced our knowledge of the disease’s biology. However, tumor heterogeneity remains a major factor contributing to poor clinical outcomes and treatment resistance. The integration of multi-omics technologies including genomic, transcriptomic, and proteomic analyses, offers a powerful approach to further dissect tumor heterogeneity and accelerate the discovery of new biomarkers and therapeutic targets. This review aims to highlight the potential utility and evolving role of multiomics (-omics) in improving our understanding of TNBC biology - particularly tumor heterogeneity - ultimately facilitating the development of novel therapies and actionable strategies to treat the disease.
Keywords:
triple-negative breast cancer
; multiomics
; tumor heterogeneity
; biomarkers
; therapeutic targets
1. Introduction
Despite advancements in diagnosis and treatment, breast cancer has an incidence of over 2 million worldwide and remains the leading cause of cancer mortality for women [1,2]. Triple-negative breast cancer (TNBC) refers to a heterogenous group of breast cancers that lack expression of ER, PR, and HER2. Despite recent advances with immunotherapy (IO) and antibody-drug conjugates (ADCs), TNBC remains the most aggressive subtype, characterized by a high risk of recurrence and a short overall survival in the metastatic setting [3,4]. As a result, there continues to be a need to identify biomarkers for diagnosis, prognosis, and treatment of these aggressive breast cancers.
Multiomics (also referred to as -omics) represent a potential avenue for interrogation of these novel gene targets. They are multimodal techniques that identify biological molecules (i.e., mRNA in the case of transcriptomics [5]) to gain a comprehensive understanding of biology and disease pathology. They generate tremendous amounts of data, which in the hands of skilled bioinformaticians can be used to comprehensively profile heterogeneous biologic samples. These techniques can be used to identify novel genes of interest from a background of 100s if not 1000s of genes, ultimately helping researchers work more efficiently to seek answers.
The purpose of this review is to explore current applications of -omics analyses and examine their potential role in the identification of novel biomarkers, with the goal of guiding the future of research on TNBC.
2. Multiomic Approaches to Cancer Research
Multiomics technologies can be used to identify mRNA (transcriptomics), DNA (genomics), protein (proteomics), metabolites (metabolomics), and lipids (lipidomics). Transcriptomic approaches explore expression levels of different genes on a sample-wide (bulk RNA-seq) or single-cell (scRNA-seq) level. Recent advancements have allowed these principles to be applied directly onto histologic sections to assess gene expression with spatial context (spatial transcriptomics). Table 1 contains an overview of some of these different -omics approaches, along with their strengths and weaknesses. Integrated approaches (e.g., single-cell RNA-sequencing combined with spatial transcriptomics) optimize each approach’s strengths to identify clinically relevant trends.
Multiomic techniques have been used to better understand the tumor biology of different cancers, from melanomas to gliomas and adenocarcinomas [12,13,14,15]. They have helped to better understand mechanisms behind response to checkpoint inhibitors [16] as well as assess why some tumors are less responsive to immunotherapy [17]. They can also be used to monitor tumorigenic signaling between malignant and non-malignant cells [18]. In breast cancer specifically, they have been used to identify transcriptionally divergent subtypes of breast cancers [19], and to analyze samples of HER2+ tumor in conjunction with histology [20]. Further, attempts have been made at using -omics to annotate HER2 status on histology [21] as well as to delineate the expression profiles of invasive ductal carcinoma and ductal carcinoma in-situ [22]. Recently, they have been used for the identification of novel biomarkers for breast cancer diagnosis and treatment [23].
3. Multiomic Approaches in Triple Negative Breast Cancer
Breast cancer has been classically divided into intrinsic subtypes based on expression of ER, PR, and HER2 in the PAM50 assay. These types include luminal A and B (both expressing hormone receptors, with Luminal B expressing high Ki-67), HER2-enriched, and basal-like (which is primarily composed of TNBCs). These molecular subtypes of breast cancer have clinical implications on risk and survival [24,25].
Triple negative breast cancer accounts for approximately 15% of breast cancers that lack expression of ER, PR, and HER2 [26]. They tend to be larger, higher in tumor grade, and are more aggressive than other breast cancer types [3]. Because of their lack of hormone receptors or HER2 expression, no targeted therapies currently exist. Currently treatment of TNBC relies on chemotherapy [26]. Only around 30% of TNBC cases respond to neoadjuvant chemotherapy [27], and even with successful chemotherapy, patients have higher risk of relapse and death after treatment [4]. Immunotherapies have demonstrated some effect, but some tumors are immune-cold and do not respond [28,29]. Although TNBC tumors are generally similar in their poor prognostic outcomes, there exists great heterogeneity between tumors – pathologically, mutationally, and transcriptionally [27]. Thus, there is a need to identify 1) prognostic indicators to stratify risk for TNBC cancers and 2) markers to target TNBC (and its different specific subtypes).
3.1. Triple Negative Breast Cancer Molecular Subtypes Based on Gene Set Enrichment Analysis
Due to lack of non-cytotoxic therapies, concerted efforts have been made to classify TNBCs molecularly for possible identification of novel treatment targets. Six primary subtypes, collectively known as TNBCtype, were initially identified based on gene set enrichment analyses (GSEA), comparing relative expression of different canonical pathways from microarray data [26,30,31,32]. These initial subtypes are summarized in Table 2. Interestingly, while TNBCs overall do not express ER or PR, the LAR subtype was enriched in pathways associated with hormone response. Since initial stratification, these TNBCtypes have been refined to four current subtypes (TNBCtype-4), with laser microdissection of histology revealing the original immunomodulatory (IM) and mesenchymal stem-like (MSL) groups to be non-tumor associated cell lines.
The molecular TNBCtype-4 classifications have been shown to be clinically relevant [33], notably as predictors of complete pathologic response rates to neoadjuvant chemotherapy, with basal-like 1 (BL1) having the best response and basal-like 2 (BL2) and luminal androgen receptor (LAR) the worst [31,34]. Further, multiomic investigations into these subtypes have revealed genes of interest that represent potential avenues of treatment for their associated subtypes.

Table 3.
TNBCtype-4 Targets.
| Subtype | Gene Findings | Citation |
| BL1 | Upregulated DNA/RNA synthesis, cell division, and nuclear export | [35] |
| BL2 | Upregulated extracellular matrix, collagen, cell junction, and cell membrane components | [35] |
| M | Lowly express PD-L1, making immunotherapy less effective | [36] |
| LAR | PRC-2, enhances chemotherapy response Genetic dependency on CCND1 GPX4, can be inhibited to cause ferroptosis Activating mutation in PIK3CA |
[33,34,36,37] |
The classification of these molecular subtypes of TNBC based on GSEA has been a landmark finding, however further studies reveal that TNBC complexity persists even past these subtypes. For example, while both “basal-like”, the BL1 and BL2 subtypes have notable differences that have been identified with further study. The BL2 subtype is enriched in genes that contribute to cancer invasion and motility [35], which could help explain why the BL2 subtype of TNBCs have worse observed responses to treatment.
MicroRNAs, which are single stranded, non-coding RNA that can post-transcriptionally modify mRNA and regulate cancer phenotype, represent an additional layer of complexity with these subtypes [38]. When compared with BL2, BL1 cancers have been demonstrated to upregulated the miR-17-92 cluster of microRNA [35], which are known to suppress E2F1 and PTEN, both of which are growth control proteins [39,40,41,42].
Even further, the LAR subtype of TNBC has been shown to exist on a spectrum. Distinct LAR subgroups (“sub-subtypes”) have been identified, with one being more biologically similar to mesenchymal TNBC and another being similar to basal TNBC [43]. Additionally, outside of these TNBCsubtype-4 analyses, comparisons of different TNBC groups have identified other markers that may help elucidate the mechanisms behind TNBC’s tumor heterogeneity.
3.2. Differentially Expressed Gene Analysis of Triple Negative Breast Cancer
In order to better understand mechanisms behind TNBC growth and tumorigenesis, research has been conducted comparing different phenotypes of TNBC to find differentially expressed genes (DEGs). DEG analysis is a method of identifying potential genes of interest when comparing different populations of study [44]. Out of 100s if not 1000s of genes, it identifies genes of interest that are over-expressed or under-expressed in one population versus another. Comparisons can be inter-sample (between separate biological samples, i.e., comparing expression profiles between cancers with different TNM staging) or intra-sample (within the same sample, i.e., comparing different cell populations within the same tumor) [45].
The fundamental idea behind DEG analysis is that differences in gene expression ultimately contribute to differences in observed phenotype – whether that be disease aggressiveness or treatment response. Identification of these DEGs can be a vital step in understanding how different disease pathologies come to be and can be helpful when exploring pheonmenon that are incredibly complex and driven by a variety of factors. In the case of triple negative breast cancer, DEG analysis can be performed with -omics techniques to investigate genes which underly differences in prognosis and treatment response.
3.2.1. Immune-Related
The cancer-immunity cycle, where the innate immune system catalyzes the adaptive response to tumor antigens, defines the body’s response to cancers [46]. Mechanisms to evade this system [47] allow cancer cells to proliferate and for tumors to become malignant. Yet, even when the immune system is unable prevent cancer development, the advent of immunotherapies, like checkpoint inhibitors and monoclonal antibodies, have proven effective at harnessing the immune system’s natural capabilities to treat cancer [48]. For example in high-risk, early stage TNBC, the immune checkpoint inhibitor pembrolizumab has achieved significant improvements in pathologic complete response[49,50]. In advanced metastatic disease with positive PD-L1 expression, pembrolizumab has also demonstrated significant effects on survival[51] and has become standard of care. Despite these advancements in treatment, TNBCs are heterogeneous, and a subset have molecular profiles that in some way impact their ability to respond to immunotherapy. -Omics techniques utilizing differential analysis have been used to identify key players in this chain, from immune response to immune therapy.
To evaluate the spectrum of immune reactions to TNBC development, differential gene expression analysis can be used to compare TNBC samples of different severities. These analyses help identify novel markers for downstream analysis. For example, macrophage inhibitory factor (MIF) is secreted by cancer cells and has been thought to drive immune evasion [52,53,54]. In Chen et al., RNA-sequencing across TNBC samples of varying severity revealed differential expression of MIF, with higher expression of this gene correlating with higher T stage. Furthermore, through the integration of other -omics approaches, Chen et al. were able to implicate a possible mechanism underlying this finding. Single-cell RNA-sequencing analysis, which can be used to infer cell-to-cell interactions, identified epithelial breast cancer cells which secrete MIF, potentially mediating the polarization of macrophages into the M2 phenotype. Spatial transcriptomics corrobated this result, with epithelial cell markers and MIF expression found to be expressed in concurrent regions [55].
Even with the adoption of novel immunotherapies, TNBC has proven to be a formidable foe, with many cases of TNBC being non-responsive to immune checkpoint inhibitors [28,29]. Much like with tumor severity, differential analysis of -omics data can be performed to better understand the genes that are involved in this phenomenon. In Liu et al., analysis of single-cell sequencing data from both immune checkpoint responders and non-responders, revealed SIGLEC9 (in myeloid cells) and TNFSF9 (in dendritic cells) to be expressed highly in non-responders compared to responders [56]. Interestingly, -omics techniques have similarly implicated SIGLEC9-expressing macrophages in other cancers that fail to respond to anti-PD-1 therapy [57], as well as TNFSF9 in metastatic progression [58], altogether suggesting that these two genes somehow contribute to immune-cold responses to immunotherapy in TNBC. On the other hand, investigations of immunotherapy response have also identified genes that could be important predictors of successful response to immune checkpoint inhibitors. In Zhang et al.’s comparison of single-cell RNA-sequencing from responders and non-responders, CXCL13 expressed in T cells was found to be enriched in responders prior to treatment. Additionally, for responders who received combination chemo- and immunotherapy, the population of CXCL13-expressing T-cells significantly expanded from baseline. This pattern was not mimicked in patients who responded to monotherapy paclitaxel, who interestingly exhibited decreased levels of CXCL13+ T cells after treatment. These observations in single-cell sequencing suggest that not only is CXCL13 significant for TNBC response to immunotherapy – the addition of paclitaxel with these treatments may actually inhibit the expansion of CXCL13+ cells and dampen the effect of immune checkpoint blockers [59].
Differential analysis of gene expression and protein translation can be further applied to the intrinsic subtypes of breast cancer to explore immune therapy response. In Zhu et al., differential analysis of proteomics comparing all intrinsic breast cancer subtypes, revealed CD73, a protein which inhibits immune responses, to be enriched in TNBC. To explain this finding, the authors implicated OTUD4, which stabilizes CD73 [60] in TNBC cells, causing its eventual accumulation and downstream immunosuppressive effects. With additional spatial trancriptomic analysis, OTUD4 and CD73 were found to be positively correlated in malignant regions across samples of TNBC [61]. Here, -omics techniques allowed the authors to rapidly profile hundreds of proteins to identify a potential protein for further investigation. Armed with this knowledge, Zhu et al. investigated possible treatment modalities targeting the OTUD4/CD73 axis, by developing a pharmacologic inhibitor of OTUD4, ST80, which restored T-cell function and increased the efficacy of immune checkpoint therapy [61].
3.2.2. Epithelial Cells
The principles of differential gene expression have also been applied to the epithelial cells that make up TNBC tumors. Through single-cell RNA-sequencing, Kim et al. sought to establish markers that separate TNBC epithelial cells from epithelial cells from other breast cancers. Using this method, the authors identifed 12 DEGs which were highly expressed in TNBC epithelial cells compared to normal breast tissue, ER+, and HER2+ breast cancer [62]. These included multiple genes that are involved in epithelial to mesenchymal transition: vimentin (VIM) and caldesmon (CALD1).
In epithelial cell cancers (carcinomas) the epithelial to mesenchymal transition has been heavily implicated in cancer progression and metastasis. During this process, epithelial cells shed their epithelial characteristics in favor of upregulating mesenchymal cell markers. Under typical conditions, this transition contributes to wound healing, however in carcinomas, it increases cellular motility and allows epithelial cell cancers to become invasive [63]. Further, genes involved in this process have been associated with drug-resistance [64], underlying the importance of studying these genes through techniques like -omics.
VIM, an intermediate filament protein found in mesenchymal cells [65], is required for the epithelial to mesenchymal transition and has been shown to promote TNBC invasion [66,67]. Silencing of the estrogen receptor leads to the development of the vimentin cytoskeleton [68], which is believed to be one of the driving factors behind the aggressiveness of TNBC. Further, pre-clinical experiments assessing the effect of vimentin knockdown on tumorgenicity have exhibited decreased metastatic progression, suggesting the possibility for VIM not only as a marker of TNBC, but also for future therapeutic investigation as well [67].
CALD1 encodes caldesmon, an actin-binding protein that regulates cell motility via actomyosin systems in both muscle and non-muscle cells [69,70]. Caldesmon has previously been implicated in other carcinomas, including colorectal cancer [69,71]. In breast cancer cells, silencing of the estrogen receptor led to CALD1 upregulation [68]. Further studies of CALD1 in breast cancer have suggested that CALD1 could be a marker for tamoxifen-resistance and cancer recurrence [72].
In sum, applying differential gene expression analysis to data from -omics techniques has identified many markers of TNBC – some associated with better or worse treatment response, cancer severity, or prognosis. These markers are shown in Table 4.
3.3. Additional Applications of -Omics in TNBC
Other -omic techniques, like whole genome sequencing or exome sequencing can be used to identify mutants and copy number variants that occur in TNBC. These techniques can be used to better understand how the mutation development contributes to TNBC development and response to treatment.
Kim et al. used exome sequencing to study the development of chemoresistance in treatment-naïve TNBC cases. They demonstrated broad categories of mutant responses to neoadjuvant chemotherapy: clonal persistance (where mutant frequencies were altered but mutants remained after therapy) and clonal extinction (where mutant clones were eradicated post-treatment). Further, Kim et al. performed single-nuclei RNA sequencing to identify phenotypic differences between chemoresistant and chemoresponsive tumors. They identified genes from several processes, including extracellular matrix degradation, angiogenesis, epithelial to mesenchymal transition, AKT1 signaling via mTOR, and hypoxia, that were upregulated in chemoresistant TNBC after neaodjuvant chemotherapy [73]. In this case, -omics were used to better understand the different phenotypic responses that TNBC can have to therapy.
4. Discussion
Because each of the multiomic approaches have their own strengths and weaknesses, the most compelling of research studies integrate multiple approaches in their investigations of triple negative breast cancer. Studies like Chen et al. [55] and Kim et al. [73] draw upon multiple -omics techniques to provide the most thorough analysis of TNBCs. Such approaches can be used to profile 100s if not 1000s of genes in order to identify potential novel biomarkers of TNBC. These isolated biomarkers can then be further validated preclinically using in vitro cell lines and animal models. Finally, the most promising of these genes can finally be evaluated clinically, to explore potential new avenues of TNBC classification, risk-stratification, and treatment. This proposed pipeline of -omics to bedside investigation is illustrated in Figure 1.
While the downstream effects of -omics implementation in TNBC research have yet to be fully realized, these techniques have already advanced our understanding of TNBC. For instance, the discovery of the TNBCtype-4 subtypes of triple negative breast cancer has led to investigations of new targeted therapies, exploring individualized therapies against genes enriched in each of the molecular subtypes of TNBC [74]. The knowledge that basal-like TNBCs upregulate growth factor genes has led to investigations into the efficacy of targeting epidermal growth factor receptor (EGFR) [75,76], vascular endothelial growth factor receptor (VEGFR) [77,78], and fibroblast growth factor receptor (FGFR) [33,79]. Furthermore, the identification of the LAR subtype has driven research into androgen receptor targeting [33] as well as into the effects of PI3K inhibitors [74], which have been shown to make cancers more susceptible to DNA-damaging agents [80].
The TNBCtype-4 subtypes of triple negative breast cancer have also demonstrated some potential for stratifying responses to chemotherapy regimens. For instance, the p53 family of tumor suppressors has been shown to be associated with the basal-like classes of TNBC, suggesting its status as a potential predictive marker for response to paclitaxel/cisplatin with everolimus [74].
Further applications of -omics techniques using differential gene analysis have identified key genes that distinguish TNBC from other types of breast cancer. These analyses can also identify how different subtypes (and even sub-subtypes) of TNBC differ from each other, to help us understand the heterogeneity that underlies all the different phenotypes of TNBC that are observed. However, because of this complexity, true understanding of TNBC remains elusive. Even within the TNBCsubtype-4 classifications, sub-subtypes exist. Additional markers that have been discovered independently of the TNBCsubtype-4 classifications are further evidence of the deep heterogeneity of TNBCs. It is unlikely that a “one size fits all” or even a “one size fits most” is possible with TNBC – as treatment responses have been demonstrated to be multifactorial, and not wholly dependent on any single known marker, an idea that has been exemplified with programed death ligand 1 (PD-L1). The advent of immunotherapies has been an exciting advancement in oncology, as immunotherapies like atezolizumab and pembrolizumab can be used in a targeted fashion against PD-L1/PD-1. In the case of TNBC however, PD-L1 status is a poor predictor of tumor response to atezolizumab with carboplatin. Although the drug directly targets PD-L1, certain subtypes of TNBC can respond to immunotherapy regardless of their PD-L1 status, suggesting that this one gene is not capable of stratifying TNBC into responders and non-responders [81].
It is clear that triple negative breast cancers have vast complexity. In spite of the nature of the disease, -omics techniques have incredible potential to tackle this challenge. In particular, spatially resolved -omics technologies enable researchers to understand TNBC tumor cells in the context of their microenvironment, providing insights into cell-to-cell interactions, immune infiltration, and stromal contributions to tumor behavior. Advancements and further applications of -omics, like the advent of sub-cellular resolution for spatial transcriptomics with the Xenium platform [82] or the profiling of non-coding sequences like microRNA [83] or long non-coding mRNA [84] make the field even more exciting. While cost remains a concern for multiomics studies (the techniques themselves are often costly to run even without considering labor costs), as with any new and emerging technology, the expenses will continue to decline as more innovations are made to improve these techniques. Finally, as -omics methods and analysis improve, it is our hope that they can be further integrated into our studies of TNBC.
5. Conclusions
Triple negative breast cancer remains one of the most complex and poorly understood cancers to afflict humanity. It has significant epidemiological consequences, and because of its aggressiveness and lack of expression of targetable ER, PR, or HER2, it continues to be poor prognostically. As such, there is a need to continue exploring novel biomarkers for classification and treatment of TNBC. Multiomics (or -omics) are multimodal techniques that are used to identify, profile, and understand biological molecules, such as DNA, RNA, or proteins. These molecules define biology and drive the mechanisms behind survival as well as disease, including TNBC.
In the hands of skilled bioinformaticians, multiomics can be used to tackle the complexity of TNBC, in a way that was not previously possible. These techniques can be used to explore 100s if not 1000s of genes between multiple experimental conditions, to better understand TNBC broadly and potentially identify novel biomarkers for downstream experimentation. As these technologies continue to advance, they should be further integrated as tools used to study TNBC, just as mouse models or clinical trials have been.
Author Contributions
Conceptualization, Y.P.; writing—original draft preparation, C.J.; writing—review and editing, Y.P.; visualization, C.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wilkinson, L.; Gathani, T. Understanding breast cancer as a global health concern. Br J Radiol 2022, 95, 20211033. [Google Scholar] [CrossRef]
- Ginsburg, O.; Bray, F.; Coleman, M.P.; Vanderpuye, V.; Eniu, A.; Kotha, S.R.; Sarker, M.; Huong, T.T.; Allemani, C.; Dvaladze, A.; et al. The global burden of women’s cancers: A grand challenge in global health. The Lancet 2017, 389, 847–860. [Google Scholar] [CrossRef]
- Carey, L.A.; Dees, E.C.; Sawyer, L.; Gatti, L.; Moore, D.T.; Collichio, F.; Ollila, D.W.; Sartor, C.I.; Graham, M.L.; Perou, C.M. The triple negative paradox: Primary tumor chemosensitivity of breast cancer subtypes. Clin Cancer Res 2007, 13, 2329–2334. [Google Scholar] [CrossRef] [PubMed]
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-negative breast cancer: Clinical features and patterns of recurrence. Clin Cancer Res 2007, 13, 4429–4434. [Google Scholar] [CrossRef] [PubMed]
- Nassar, S.F.; Raddassi, K.; Wu, T. Single-Cell Multiomics Analysis for Drug Discovery. Metabolites 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Papalexi, E.; Satija, R. Single-cell RNA sequencing to explore immune cell heterogeneity. Nat Rev Immunol 2018, 18, 35–45. [Google Scholar] [CrossRef]
- Ziegenhain, C.; Vieth, B.; Parekh, S.; Reinius, B.; Guillaumet-Adkins, A.; Smets, M.; Leonhardt, H.; Heyn, H.; Hellmann, I.; Enard, W. Comparative Analysis of Single-Cell RNA Sequencing Methods. Mol Cell 2017, 65, 631–643.e634. [Google Scholar] [CrossRef]
- Zheng, B.; Fang, L. Spatially resolved transcriptomics provide a new method for cancer research. J Exp Clin Cancer Res 2022, 41, 179. [Google Scholar] [CrossRef]
- Xiao, X.; Juan, C.; Drennon, T.; Uytingco, C.R.; Vishlaghi, N.; Sokolowskei, D.; Xu, L.; Levi, B.; Sammarco, M.C.; Tower, R.J. Spatial transcriptomic interrogation of the murine bone marrow signaling landscape. Bone Research 2023, 11, 59. [Google Scholar] [CrossRef]
- Viode, A.; van Zalm, P.; Smolen, K.K.; Fatou, B.; Stevenson, D.; Jha, M.; Levy, O.; Steen, J.; Steen, H. A simple, time- and cost-effective, high-throughput depletion strategy for deep plasma proteomics. Sci Adv 2023, 9, eadf9717. [Google Scholar] [CrossRef]
- Nurchis, M.C.; Radio, F.C.; Salmasi, L.; Heidar Alizadeh, A.; Raspolini, G.M.; Altamura, G.; Tartaglia, M.; Dallapiccola, B.; Pizzo, E.; Gianino, M.M.; et al. Cost-Effectiveness of Whole-Genome vs Whole-Exome Sequencing Among Children With Suspected Genetic Disorders. JAMA Netw Open 2024, 7, e2353514. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, D.; Peng, M.; Tang, L.; Ouyang, J.; Xiong, F.; Guo, C.; Tang, Y.; Zhou, Y.; Liao, Q.; et al. Single-cell RNA sequencing in cancer research. J Exp Clin Cancer Res 2021, 40, 81. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.X.; Taramelli, R.; Pedrini, E.; Knijnenburg, T.; Huang, S. Extracting Intercellular Signaling Network of Cancer Tissues using Ligand-Receptor Expression Patterns from Whole-tumor and Single-cell Transcriptomes. Scientific Reports 2017, 7, 8815. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Tao, Y.; Chen, G.; Shi, T. Systematic expression analysis of ligand-receptor pairs reveals important cell-to-cell interactions inside glioma. Cell Communication and Signaling 2019, 17, 48. [Google Scholar] [CrossRef]
- Hosein, A.N.; Huang, H.; Wang, Z.; Parmar, K.; Du, W.; Huang, J.; Maitra, A.; Olson, E.; Verma, U.; Brekken, R.A. Cellular heterogeneity during mouse pancreatic ductal adenocarcinoma progression at single-cell resolution. JCI Insight 2019, 4. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, W.; Jankovic, V.; Golubov, J.; Poon, P.; Oswald, E.M.; Gurer, C.; Wei, J.; Ramos, I.; Wu, Q.; et al. Combination cancer immunotherapy targeting PD-1 and GITR can rescue CD8(+) T cell dysfunction and maintain memory phenotype. Sci Immunol 2018, 3. [Google Scholar] [CrossRef]
- Li, B.; Severson, E.; Pignon, J.-C.; Zhao, H.; Li, T.; Novak, J.; Jiang, P.; Shen, H.; Aster, J.C.; Rodig, S.; et al. Comprehensive analyses of tumor immunity: Implications for cancer immunotherapy. Genome Biology 2016, 17, 174. [Google Scholar] [CrossRef]
- Calbo, J.; van Montfort, E.; Proost, N.; van Drunen, E.; Beverloo, H.B.; Meuwissen, R.; Berns, A. A Functional Role for Tumor Cell Heterogeneity in a Mouse Model of Small Cell Lung Cancer. Cancer Cell 2011, 19, 244–256. [Google Scholar] [CrossRef]
- Wu, S.Z.; Al-Eryani, G.; Roden, D.L.; Junankar, S.; Harvey, K.; Andersson, A.; Thennavan, A.; Wang, C.; Torpy, J.R.; Bartonicek, N.; et al. A single-cell and spatially resolved atlas of human breast cancers. Nat Genet 2021, 53, 1334–1347. [Google Scholar] [CrossRef]
- Vickovic, S.; Eraslan, G.; Salmén, F.; Klughammer, J.; Stenbeck, L.; Schapiro, D.; Äijö, T.; Bonneau, R.; Bergenstråhle, L.; Navarro, J.F.; et al. High-definition spatial transcriptomics for in situ tissue profiling. Nat Methods 2019, 16, 987–990. [Google Scholar] [CrossRef]
- Andersson, A.; Larsson, L.; Stenbeck, L.; Salmén, F.; Ehinger, A.; Wu, S.; Al-Eryani, G.; Roden, D.; Swarbrick, A.; Borg, Å. Spatial deconvolution of HER2-positive breast tumors reveals novel intercellular relationships. bioRxiv 2020. [Google Scholar] [CrossRef]
- Yoosuf, N.; Navarro, J.F.; Salmén, F.; Ståhl, P.L.; Daub, C.O. Identification and transfer of spatial transcriptomics signatures for cancer diagnosis. Breast Cancer Res 2020, 22, 6. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Sahoo, S.; Brien, R.; Jung, S.; Humphries, B.; Lee, W.; Cheng, Y.H.; Zhang, Z.; Luker, K.E.; Wicha, M.S.; et al. Single-cell RNA-sequencing of migratory breast cancer cells: Discovering genes associated with cancer metastasis. Analyst 2019, 144, 7296–7309. [Google Scholar] [CrossRef] [PubMed]
- Prat, A.; Pineda, E.; Adamo, B.; Galván, P.; Fernández, A.; Gaba, L.; Díez, M.; Viladot, M.; Arance, A.; Muñoz, M. Clinical implications of the intrinsic molecular subtypes of breast cancer. Breast 2015, 24 (Suppl. 2), S26–S35. [Google Scholar] [CrossRef]
- Parker, J.S.; Mullins, M.; Cheang, M.C.U.; Leung, S.; Voduc, D.; Vickery, T.; Davies, S.; Fauron, C.; He, X.; Hu, Z.; et al. Supervised Risk Predictor of Breast Cancer Based on Intrinsic Subtypes. J Clin Oncol 2023, 41, 4192–4199. [Google Scholar] [CrossRef]
- Abramson, V.G.; Lehmann, B.D.; Ballinger, T.J.; Pietenpol, J.A. Subtyping of triple-negative breast cancer: Implications for therapy. Cancer 2015, 121, 8–16. [Google Scholar] [CrossRef]
- Lehmann, B.D.; Pietenpol, J.A. Clinical implications of molecular heterogeneity in triple negative breast cancer. Breast 2015, 24 (Suppl. 2), S36–S40. [Google Scholar] [CrossRef]
- Keenan, T.E.; Tolaney, S.M. Role of Immunotherapy in Triple-Negative Breast Cancer. J Natl Compr Canc Netw 2020, 18, 479–489. [Google Scholar] [CrossRef]
- Steiner, M.; Tan, A.R. The evolving role of immune checkpoint inhibitors in the treatment of triple-negative breast cancer. Clin Adv Hematol Oncol 2021, 19, 305–315. [Google Scholar]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J Clin Invest 2011, 121, 2750–2767. [Google Scholar] [CrossRef]
- Masuda, H.; Baggerly, K.A.; Wang, Y.; Zhang, Y.; Gonzalez-Angulo, A.M.; Meric-Bernstam, F.; Valero, V.; Lehmann, B.D.; Pietenpol, J.A.; Hortobagyi, G.N.; et al. Differential response to neoadjuvant chemotherapy among 7 triple-negative breast cancer molecular subtypes. Clin Cancer Res 2013, 19, 5533–5540. [Google Scholar] [CrossRef]
- Lehmann, B.D.; Jovanović, B.; Chen, X.; Estrada, M.V.; Johnson, K.N.; Shyr, Y.; Moses, H.L.; Sanders, M.E.; Pietenpol, J.A. Refinement of Triple-Negative Breast Cancer Molecular Subtypes: Implications for Neoadjuvant Chemotherapy Selection. PLoS ONE 2016, 11, e0157368. [Google Scholar] [CrossRef]
- Lehmann, B.D.; Pietenpol, J.A. Identification and use of biomarkers in treatment strategies for triple-negative breast cancer subtypes. J Pathol 2014, 232, 142–150. [Google Scholar] [CrossRef]
- Zhang, W.; Li, E.; Wang, L.; Lehmann, B.D.; Chen, X.S. Transcriptome Meta-Analysis of Triple-Negative Breast Cancer Response to Neoadjuvant Chemotherapy. Cancers 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Kalecky, K.; Modisette, R.; Pena, S.; Cho, Y.R.; Taube, J. Integrative analysis of breast cancer profiles in TCGA by TNBC subgrouping reveals novel microRNA-specific clusters, including miR-17-92a, distinguishing basal-like 1 and basal-like 2 TNBC subtypes. BMC Cancer 2020, 20, 141. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, B.D.; Colaprico, A.; Silva, T.C.; Chen, J.; An, H.; Ban, Y.; Huang, H.; Wang, L.; James, J.L.; Balko, J.M.; et al. Multi-omics analysis identifies therapeutic vulnerabilities in triple-negative breast cancer subtypes. Nat Commun 2021, 12, 6276. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Xiao, Y.; Ding, J.H.; Jin, X.; Ma, D.; Li, D.Q.; Shi, J.X.; Huang, W.; Wang, Y.P.; Jiang, Y.Z.; et al. Ferroptosis heterogeneity in triple-negative breast cancer reveals an innovative immunotherapy combination strategy. Cell Metab 2023, 35, 84–100.e108. [Google Scholar] [CrossRef]
- Hussen, B.M.; Hidayat, H.J.; Salihi, A.; Sabir, D.K.; Taheri, M.; Ghafouri-Fard, S. MicroRNA: A signature for cancer progression. Biomed Pharmacother 2021, 138, 111528. [Google Scholar] [CrossRef]
- Knoll, S.; Emmrich, S.; Pützer, B.M. The E2F1-miRNA cancer progression network. Adv Exp Med Biol 2013, 774, 135–147. [Google Scholar] [CrossRef]
- O’Donnell, K.A.; Wentzel, E.A.; Zeller, K.I.; Dang, C.V.; Mendell, J.T. c-Myc-regulated microRNAs modulate E2F1 expression. Nature 2005, 435, 839–843. [Google Scholar] [CrossRef]
- Chawra, H.S.; Agarwal, M.; Mishra, A.; Chandel, S.S.; Singh, R.P.; Dubey, G.; Kukreti, N.; Singh, M. MicroRNA-21’s role in PTEN suppression and PI3K/AKT activation: Implications for cancer biology. Pathol Res Pract 2024, 254, 155091. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Shi, G.; Zhang, Q.; Wu, Q.; Li, B.; Zhang, Z. MicroRNA-20b promotes cell growth of breast cancer cells partly via targeting phosphatase and tensin homologue (PTEN). Cell Biosci 2014, 4, 62. [Google Scholar] [CrossRef] [PubMed]
- Jovanovic, B.; Church, S.E.; Gorman, K.M.; North, K.; Richardson, E.T.; DiLullo, M.; Attaya, V.; Kasparian, J.; Mohammed-Abreu, A.; Kirkner, G.; et al. Integrative multi-omic profiling of triple-negative breast cancer for identifying suitable therapies. Clinical Cancer Research 2024. [Google Scholar] [CrossRef] [PubMed]
- Khozyainova, A.A.; Valyaeva, A.A.; Arbatsky, M.S.; Isaev, S.V.; Iamshchikov, P.S.; Volchkov, E.V.; Sabirov, M.S.; Zainullina, V.R.; Chechekhin, V.I.; Vorobev, R.S.; et al. Complex Analysis of Single-Cell RNA Sequencing Data. Biochemistry (Mosc) 2023, 88, 231–252. [Google Scholar] [CrossRef]
- Potter, S.S. Single-cell RNA sequencing for the study of development, physiology and disease. Nat Rev Nephrol 2018, 14, 479–492. [Google Scholar] [CrossRef]
- Demaria, O.; Cornen, S.; Daëron, M.; Morel, Y.; Medzhitov, R.; Vivier, E. Harnessing innate immunity in cancer therapy. Nature 2019, 574, 45–56. [Google Scholar] [CrossRef]
- Vinay, D.S.; Ryan, E.P.; Pawelec, G.; Talib, W.H.; Stagg, J.; Elkord, E.; Lichtor, T.; Decker, W.K.; Whelan, R.L.; Kumara, H.; et al. Immune evasion in cancer: Mechanistic basis and therapeutic strategies. Semin Cancer Biol 2015, 35, S185–S198. [Google Scholar] [CrossRef]
- Kumar, A.R.; Devan, A.R.; Nair, B.; Vinod, B.S.; Nath, L.R. Harnessing the immune system against cancer: Current immunotherapy approaches and therapeutic targets. Mol Biol Rep 2021, 48, 8075–8095. [Google Scholar] [CrossRef]
- Pusztai, L.; Denkert, C.; O’Shaughnessy, J.; Cortes, J.; Dent, R.; McArthur, H.; Kümmel, S.; Bergh, J.; Park, Y.H.; Hui, R.; et al. Event-free survival by residual cancer burden with pembrolizumab in early-stage TNBC: Exploratory analysis from KEYNOTE-522. Ann Oncol 2024, 35, 429–436. [Google Scholar] [CrossRef]
- Schmid, P.; Cortes, J.; Pusztai, L.; McArthur, H.; Kümmel, S.; Bergh, J.; Denkert, C.; Park, Y.H.; Hui, R.; Harbeck, N.; et al. Pembrolizumab for Early Triple-Negative Breast Cancer. N Engl J Med 2020, 382, 810–821. [Google Scholar] [CrossRef]
- Cortes, J.; Rugo, H.S.; Cescon, D.W.; Im, S.A.; Yusof, M.M.; Gallardo, C.; Lipatov, O.; Barrios, C.H.; Perez-Garcia, J.; Iwata, H.; et al. Pembrolizumab plus Chemotherapy in Advanced Triple-Negative Breast Cancer. N Engl J Med 2022, 387, 217–226. [Google Scholar] [CrossRef]
- Yan, L.; Wu, M.; Wang, T.; Yuan, H.; Zhang, X.; Zhang, H.; Li, T.; Pandey, V.; Han, X.; Lobie, P.E.; et al. Breast Cancer Stem Cells Secrete MIF to Mediate Tumor Metabolic Reprogramming That Drives Immune Evasion. Cancer Res 2024, 84, 1270–1285. [Google Scholar] [CrossRef]
- Balogh, K.N.; Templeton, D.J.; Cross, J.V. Macrophage Migration Inhibitory Factor protects cancer cells from immunogenic cell death and impairs anti-tumor immune responses. PLoS ONE 2018, 13, e0197702. [Google Scholar] [CrossRef] [PubMed]
- Kang, I.; Bucala, R. The immunobiology of MIF: Function, genetics and prospects for precision medicine. Nat Rev Rheumatol 2019, 15, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Liu, H.; Hong, B.; Xiao, Y.; Qian, Y. MIF as a potential diagnostic and prognostic biomarker for triple-negative breast cancer that correlates with the polarization of M2 macrophages. Faseb j 2024, 38, e23696. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.W.; Lin, J.; Hou, R.; Cai, Z.; Gong, Y.; He, P.A.; Yang, J. Single-cell RNA-seq reveals the metabolic status of immune cells response to immunotherapy in triple-negative breast cancer. Comput Biol Med 2024, 169, 107926. [Google Scholar] [CrossRef]
- Mei, Y.; Wang, X.; Zhang, J.; Liu, D.; He, J.; Huang, C.; Liao, J.; Wang, Y.; Feng, Y.; Li, H.; et al. Siglec-9 acts as an immune-checkpoint molecule on macrophages in glioblastoma, restricting T-cell priming and immunotherapy response. Nat Cancer 2023, 4, 1273–1291. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Y.; Yang, Y.; Liu, F.; Jiang, Z.; Jiang, Z. TNFSF9 promotes metastasis of pancreatic cancer by regulating M2 polarization of macrophages through Src/FAK/p-Akt/IL-1β signaling. Int Immunopharmacol 2022, 102, 108429. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.; Mo, H.; Hu, X.; Gao, R.; Zhao, Y.; Liu, B.; Niu, L.; Sun, X.; Yu, X.; et al. Single-cell analyses reveal key immune cell subsets associated with response to PD-L1 blockade in triple-negative breast cancer. Cancer Cell 2021, 39, 1578–1593.e1578. [Google Scholar] [CrossRef]
- Ma, X.; Wan, R.; Wen, Y.; Liu, T.; Song, Y.; Zhu, Y. Deubiquitinating enzyme OTUD4 regulates metastasis in triple-negative breast cancer by stabilizing Snail1. Exp Cell Res 2024, 434, 113864. [Google Scholar] [CrossRef]
- Zhu, Y.; Banerjee, A.; Xie, P.; Ivanov, A.A.; Uddin, A.; Jiao, Q.; Chi, J.J.; Zeng, L.; Lee, J.Y.; Xue, Y.; et al. Pharmacological suppression of the OTUD4/CD73 proteolytic axis revives antitumor immunity against immune-suppressive breast cancers. J Clin Invest 2024, 134. [Google Scholar] [CrossRef]
- Kim, M.; Yang, W.; Hong, D.; Won, H.S.; Yoon, S. A Retrospective View of the Triple-Negative Breast Cancer Microenvironment: Novel Markers, Interactions, and Mechanisms of Tumor-Associated Components Using Public Single-Cell RNA-Seq Datasets. Cancers 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Shim, J.S. Targeting Epithelial-Mesenchymal Transition (EMT) to Overcome Drug Resistance in Cancer. Molecules 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Fang, Z.; Ma, J. Regulatory mechanisms and clinical significance of vimentin in breast cancer. Biomed Pharmacother 2021, 133, 111068. [Google Scholar] [CrossRef] [PubMed]
- Grasset, E.M.; Dunworth, M.; Sharma, G.; Loth, M.; Tandurella, J.; Cimino-Mathews, A.; Gentz, M.; Bracht, S.; Haynes, M.; Fertig, E.J.; et al. Triple-negative breast cancer metastasis involves complex epithelial-mesenchymal transition dynamics and requires vimentin. Sci Transl Med 2022, 14, eabn7571. [Google Scholar] [CrossRef]
- Winter, M.; Meignan, S.; Völkel, P.; Angrand, P.O.; Chopin, V.; Bidan, N.; Toillon, R.A.; Adriaenssens, E.; Lagadec, C.; Le Bourhis, X. Vimentin Promotes the Aggressiveness of Triple Negative Breast Cancer Cells Surviving Chemotherapeutic Treatment. Cells 2021, 10. [Google Scholar] [CrossRef]
- Al Saleh, S.; Al Mulla, F.; Luqmani, Y.A. Estrogen receptor silencing induces epithelial to mesenchymal transition in human breast cancer cells. PLoS ONE 2011, 6, e20610. [Google Scholar] [CrossRef]
- Alnuaimi, A.R.; Nair, V.A.; Malhab, L.J.B.; Abu-Gharbieh, E.; Ranade, A.V.; Pintus, G.; Hamad, M.; Busch, H.; Kirfel, J.; Hamoudi, R.; et al. Emerging role of caldesmon in cancer: A potential biomarker for colorectal cancer and other cancers. World J Gastrointest Oncol 2022, 14, 1637–1653. [Google Scholar] [CrossRef]
- Sobue, K.; Sellers, J.R. Caldesmon, a novel regulatory protein in smooth muscle and nonmuscle actomyosin systems. J Biol Chem 1991, 266, 12115–12118. [Google Scholar] [CrossRef]
- Ma, W.Q.; Miao, M.C.; Ding, P.A.; Tan, B.B.; Liu, W.B.; Guo, S.; Er, L.M.; Zhang, Z.D.; Zhao, Q. CALD1 facilitates epithelial-mesenchymal transition progression in gastric cancer cells by modulating the PI3K-Akt pathway. World J Gastrointest Oncol 2024, 16, 1029–1045. [Google Scholar] [CrossRef]
- De Marchi, T.; Timmermans, A.M.; Smid, M.; Look, M.P.; Stingl, C.; Opdam, M.; Linn, S.C.; Sweep, F.C.; Span, P.N.; Kliffen, M.; et al. Annexin-A1 and caldesmon are associated with resistance to tamoxifen in estrogen receptor positive recurrent breast cancer. Oncotarget 2016, 7, 3098–3110. [Google Scholar] [CrossRef]
- Kim, C.; Gao, R.; Sei, E.; Brandt, R.; Hartman, J.; Hatschek, T.; Crosetto, N.; Foukakis, T.; Navin, N.E. Chemoresistance Evolution in Triple-Negative Breast Cancer Delineated by Single-Cell Sequencing. Cell 2018, 173, 879–893.e813. [Google Scholar] [CrossRef]
- Mayer, I.A.; Abramson, V.G.; Lehmann, B.D.; Pietenpol, J.A. New strategies for triple-negative breast cancer--deciphering the heterogeneity. Clin Cancer Res 2014, 20, 782–790. [Google Scholar] [CrossRef] [PubMed]
- Baselga, J.; Gómez, P.; Greil, R.; Braga, S.; Climent, M.A.; Wardley, A.M.; Kaufman, B.; Stemmer, S.M.; Pêgo, A.; Chan, A.; et al. Randomized phase II study of the anti-epidermal growth factor receptor monoclonal antibody cetuximab with cisplatin versus cisplatin alone in patients with metastatic triple-negative breast cancer. J Clin Oncol 2013, 31, 2586–2592. [Google Scholar] [CrossRef] [PubMed]
- Carey, L.A.; Rugo, H.S.; Marcom, P.K.; Mayer, E.L.; Esteva, F.J.; Ma, C.X.; Liu, M.C.; Storniolo, A.M.; Rimawi, M.F.; Forero-Torres, A.; et al. TBCRC 001: Randomized phase II study of cetuximab in combination with carboplatin in stage IV triple-negative breast cancer. J Clin Oncol 2012, 30, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Burstein, H.J.; Elias, A.D.; Rugo, H.S.; Cobleigh, M.A.; Wolff, A.C.; Eisenberg, P.D.; Lehman, M.; Adams, B.J.; Bello, C.L.; DePrimo, S.E.; et al. Phase II study of sunitinib malate, an oral multitargeted tyrosine kinase inhibitor, in patients with metastatic breast cancer previously treated with an anthracycline and a taxane. J Clin Oncol 2008, 26, 1810–1816. [Google Scholar] [CrossRef]
- von Minckwitz, G.; Eidtmann, H.; Rezai, M.; Fasching, P.A.; Tesch, H.; Eggemann, H.; Schrader, I.; Kittel, K.; Hanusch, C.; Kreienberg, R.; et al. Neoadjuvant chemotherapy and bevacizumab for HER2-negative breast cancer. N Engl J Med 2012, 366, 299–309. [Google Scholar] [CrossRef]
- Turner, N.; Lambros, M.B.; Horlings, H.M.; Pearson, A.; Sharpe, R.; Natrajan, R.; Geyer, F.C.; van Kouwenhove, M.; Kreike, B.; Mackay, A.; et al. Integrative molecular profiling of triple negative breast cancers identifies amplicon drivers and potential therapeutic targets. Oncogene 2010, 29, 2013–2023. [Google Scholar] [CrossRef]
- Ibrahim, Y.H.; García-García, C.; Serra, V.; He, L.; Torres-Lockhart, K.; Prat, A.; Anton, P.; Cozar, P.; Guzmán, M.; Grueso, J.; et al. PI3K inhibition impairs BRCA1/2 expression and sensitizes BRCA-proficient triple-negative breast cancer to PARP inhibition. Cancer Discov 2012, 2, 1036–1047. [Google Scholar] [CrossRef]
- Lehmann, B.D.; Abramson, V.G.; Dees, E.C.; Shah, P.D.; Ballinger, T.J.; Isaacs, C.; Santa-Maria, C.A.; An, H.; Gonzalez-Ericsson, P.I.; Sanders, M.E.; et al. Atezolizumab in Combination With Carboplatin and Survival Outcomes in Patients With Metastatic Triple-Negative Breast Cancer: The TBCRC 043 Phase 2 Randomized Clinical Trial. JAMA Oncol 2024, 10, 193–201. [Google Scholar] [CrossRef]
- Marco Salas, S.; Kuemmerle, L.B.; Mattsson-Langseth, C.; Tismeyer, S.; Avenel, C.; Hu, T.; Rehman, H.; Grillo, M.; Czarnewski, P.; Helgadottir, S.; et al. Optimizing Xenium In Situ data utility by quality assessment and best-practice analysis workflows. Nat Methods 2025, 22, 813–823. [Google Scholar] [CrossRef]
- Chen, C.; Li, Y.; Wei, W.; Lu, Y.; Zou, B.; Zhang, L.; Shan, J.; Zhu, Y.; Wang, S.; Wu, H.; et al. A precise microdissection strategy enabled spatial heterogeneity analysis on the targeted region of formalin-fixed paraffin-embedded tissues. Talanta 2024, 278, 126501. [Google Scholar] [CrossRef]
- Pinkney, H.R.; Black, M.A.; Diermeier, S.D. Single-Cell RNA-Seq Reveals Heterogeneous lncRNA Expression in Xenografted Triple-Negative Breast Cancer Cells. Biology 2021, 10. [Google Scholar] [CrossRef]
Figure 1.
Proposed Pipeline Using -Omics Techniques to Identify Biomarkers in TNBC.
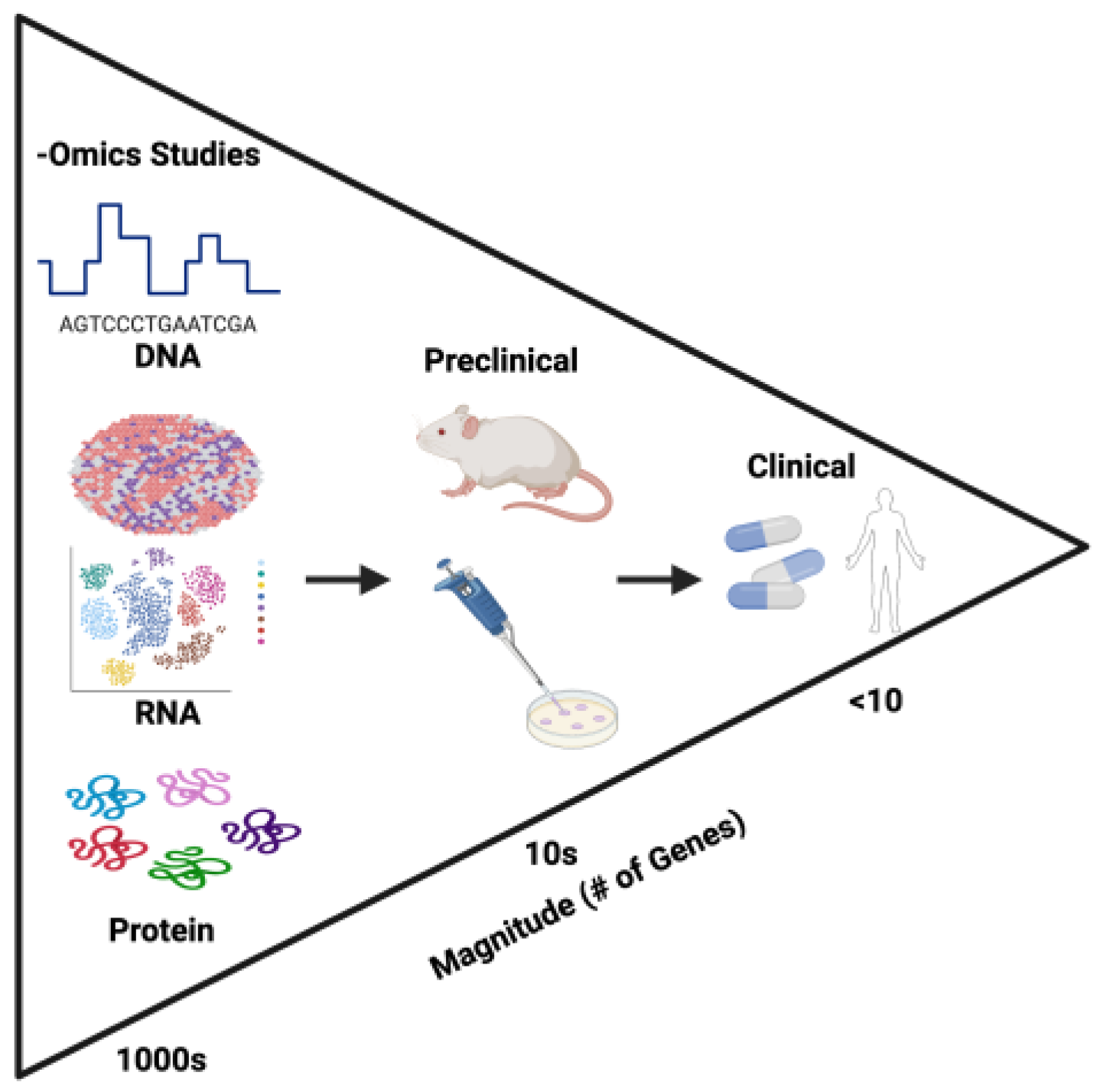
Table 1.
Overview of Multiomic Approaches.
| Technique | Description | Strength | Weakness |
| Single-cell transcriptomics (scRNA-seq) | Cellular level analysis of mRNA expression | High resolution. Identify cell populations in heterogeneous samples [6] | Cost [7]. Lacks spatial context. |
| Spatial transcriptomics | Analysis of mRNA from sample on slide | Trends with spatial context. Entire transcriptome probe sets available. Promising future as more advanced products are released | Cost [8]. Low resolution. Relatively low transcript capture in certain tissue types (e.g., mineralized tissue [9]) |
| Microarray gene expression analysis | Collection of mRNAs using array of known probes | Targeted studies | Can only identify expression of known probes. |
| Bulk RNA-seq | Analysis of mRNA from whole sample | More sensitive than microarray and doesn’t require probes. | Primarily for broad sample-wide trends |
| Proteomics | Analysis of pro-tein structure and function | Clinically relevant protein identification. Cost-effective methods [10] | Complexity of protein structure. Difficulty for studying post-translational modifications. |
| Genomics | Analysis of DNA sequencing | Whole-genome sequencing becoming more readily available. Identification of mutations. Exome sequencing can identify copy number variations. | Cost [11]. May not reflect transcribed/translated protein. Variants detected of may be of varying significance. |
Table 2.
Original TNBCtype Classes.
| Subtype | Pathways | |
| Basal-like | Basal-like 1 (BL1) | Proliferative gene pathways (cell cycle, DNA replication), usually associated with high Ki-67 |
| Basal-like 2 (BL2) | Growth factor genes | |
| Immunomodulatory (IM)* | Immune cell signaling | |
| Mesenchymal | Mesenchymal-like (M) | Cell motility, cell differentiation, WNT, ALK, Extracellular matrix |
| Mesenchymal stem-like (MSL)** | Growth factor and epithelial-to-mesenchymal transition | |
| Luminal | Luminal androgen receptor (LAR) | Androgen/estrogen metabolism, Steroid biosynthesis, Porphyrin metabolism |
Table 4.
Potential Markers of TNBC Identified by -Omics Techniques.
| Markers | Techniques | Function | Citations |
| MIF | RNA-seq, scRNA-seq, Spatial transcriptomics | Regulates glucocorticoid immunosuppression mediating cell survival | [52,53,54,55] |
| CXCL13 | scRNA-seq | Expressed in T cells to induce proinflammatory signaling in macrophages | [59] |
| CD73/OTUD4 | Proteomics and Spatial transcriptomics | CD73 stabilizes OTUD4, causing accumulation and immunosuppression | [60,61] |
| VIM | scRNA-seq | Intermediate filament protein found in mesenchymal cells. Drives epithelial to mesenchymal transition | [65,66,67,68] |
| CALD1 | scRNA-seq | Actin-binding protein involved in cell motility. Drives epithelial to mesenchymal transition. | [68,69,70] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



