Submitted:
07 October 2025
Posted:
09 October 2025
You are already at the latest version
Abstract
The emergence of multidrug-resistant (MDR) "superbugs" is a serious threat to world health because of the interdependence of environmental ecosystems, agriculture, and human medicine within the single health framework. Horizontal gene transfer (HGT), residue persistence, and excessive antibiotic use have accelerated the development of antimicrobial resistance (AMR) by 2050, it is predicted that AMR would be responsible for 8.22 million deaths. To evade conventional therapies, bacteria employ sophisticated resistance mechanisms such enzymatic inactivation (e.g., β-lactamases, carbapenemases), target alteration (e.g., PBP2a in MRSA), efflux pumps, biofilm formation, and reduced membrane permeability. In light of this dilemma, new strategies are needed to restore the efficacy of antibiotics and stop the spread of resistance. Advances in antibiotic adjuvants, like efflux pump blockers and β-lactamase inhibitors like avibactam, complement existing drugs to combat resistance. The arsenal against MDR pathogens is further diversified by phage therapy, CRISPR-Cas systems, anti-virulence inhibitors, combinatorial therapy and vaccinations. However, challenges persist, biofilm resilience, plasmid-borne anti-CRISPR defences, and ecological risks of gene-editing tools necessitate rigorous mitigation AI-driven diagnostics, metagenomics, and genomics together offer a revolutionary approach to AMR management and surveillance. To address the superbug epidemic & pandemic, the paper integrates the genetic, molecular, and ecological facets of AMR and highlights the vital need for international collaboration, sustainable practices, and One Health-aligned policies.
Keywords:
multidrug resistance
; superbugs
; antibiotic stewardship
; novel therapies
; infection control
Introduction:
The World Health Organization (WHO) revealed multidrug-resistant pathogens/superbugs as a significant global health threat. The Interagency Coordinating Group on antimicrobial resistance (AMR) has projected that by 2050, the mortality rate attributable to AMR may rise, with an estimated annual death toll of around 10 million individuals 1,2. Multidrug resistant pathogens are defined as bacteria, viruses, fungi, or parasites that have developed resistance to standard treatments and no longer respond due to factors such as misuse of antimicrobial drugs, consumption of contaminated food, poor sanitation practices, inadequate infection control measures, among others 3. Antibiotic resistance (ABR) dissemination involves multiple environmental reservoirs, including soil, water, hospitals, industrial sites, farm waste, and various polluted ecological niches 4,5. The movement or spread of pathogens carrying resistance genes is facilitated among humans, animals, and their environment 6–8. In this context, the collaboration between Chile and Sweden has advanced the One Health–One World initiative aimed at controlling ABR globally 9. One Health calls for integrated global action to safeguard health across all species and ecosystems which amplifies the risks of disease transmission, antimicrobial resistance, and ecological impact. The deep connection is shown in Figure 1. Factors contributing to the local and global dissemination of ABR include improper use of antibiotics in both animals and humans, polluted surroundings, and inadequate measures for controlling infections 4,10–12 . Thus, the One Health–One World effort is an important worldwide movement that unifies practice and policy across disciplines to prevent global dangers and enhance health outcomes. At every level, from local to global, cross-sector collaborations, standardized regulations, and robust governance are essential to its success.
The irrational use of antibiotics in humans, animals, communities, and their environments has led to the emergence of resistance reservoirs, where drug residues or resistance genes persist in the environment. The swift emergence of newly evolved resistance mechanisms, coupled with the limited effectiveness of drugs/treatments against recurrent microbial infections, results in prolonged illness, escalating healthcare expenses, deteriorating disease conditions, and heightened mortality risk. Over the past few decades, many infectious agents and parasites have developed significant resistance levels, leading to high morbidity and mortality rates; these are thus known as “superbugs” [13].
Typically, superbugs or multidrug resistance arise from natural processes, but the morbidity rate escalates in immunodeficient conditions such as diabetes, HIV infection and others. Several types of bacteria are quickly developing resistance to antibiotics listed by the WHO [1]. The interconnection of human, animal, and environmental health has intensified the spread of AMR through agricultural overuse, environmental pollution, and global travel. By 2050, AMR is forecasted to claim 10 million lives annually, driven by Gram-negative "superbugs" like carbapenem-resistant Enterobacteriaceae and methicillin-resistant Staphylococcus aureus (MRSA) [14]. These pathogens exploit diverse resistance strategies: enzymatic degradation of antibiotics (e.g., metallo-β-lactamases), efflux pumps (e.g., AcrAB-TolC), target mutations (e.g., altered DNA gyrase), and biofilm-mediated persistence. Among the different bacteria causing clinical infections viz, Klebsiella pneumoniae, Escherichia coli, Serratia marcescens, Pseudomonas aeruginosa, Staphylococcus aureus, Proteus mirabilis, and so on, are some of the most vicious ones emerging at an unprecedented rate with huge impact on public health [1].
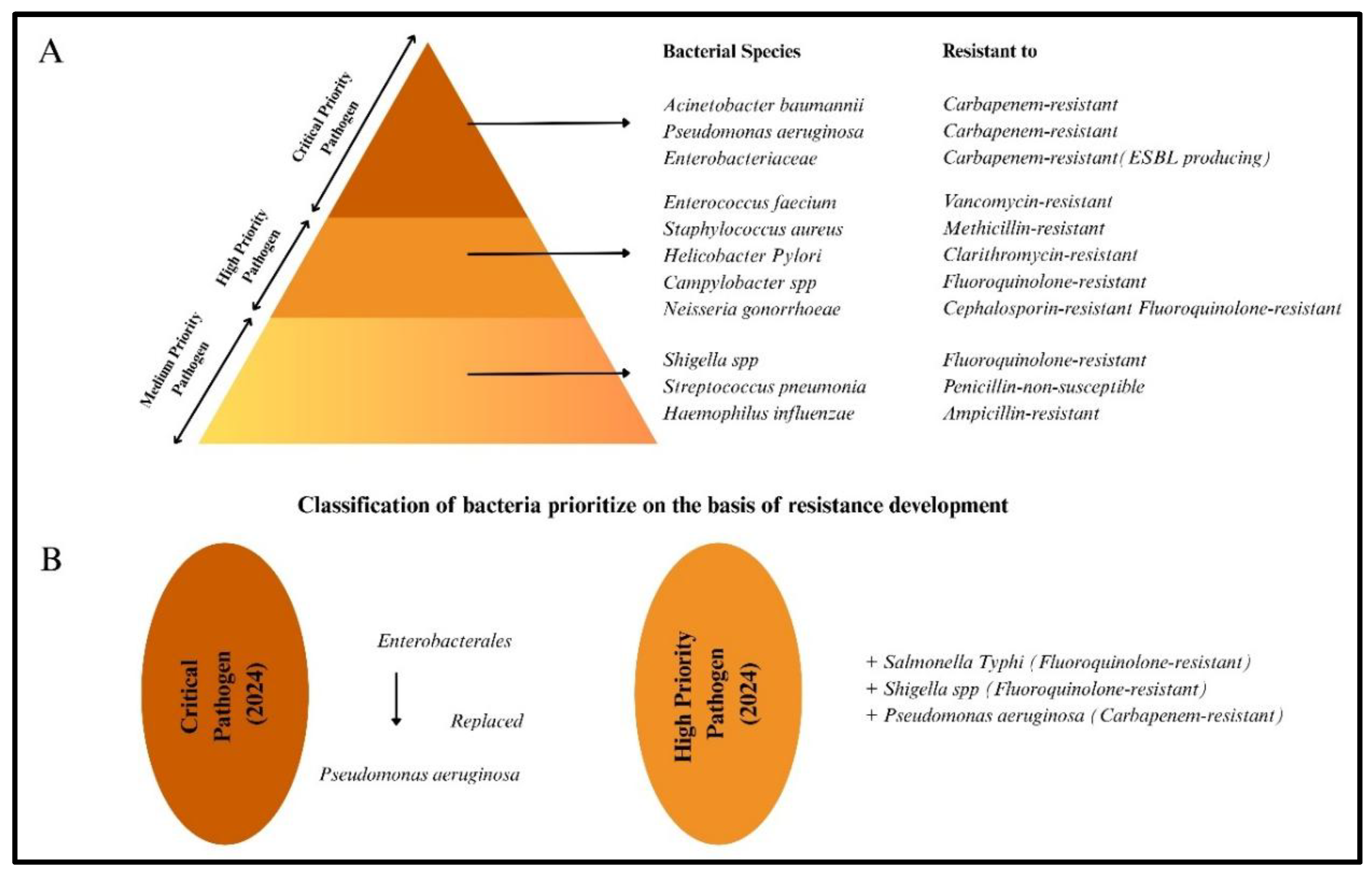
The agricultural sector, responsible for two-thirds of global antibiotic consumption, has been a crucible for resistance evolution. Prophylactic use in livestock, growth promoters, and inadequate hygiene practices have seeded resistance genes (blaCTX-M, mcr-1) into ecosystems, transmitted to humans via food chains and environmental pathways [15,16,17]. Several strategies are essential to enhance the efficacy of antibiotics and eliminate MDR. One of the most effective strategies used in clinical settings to combat infection is the use of an adjuvant in conjunction with antibiotics [18]. Simultaneously, CRISPR-Cas systems and phage therapy offers a targeted solution for multidrug-resistant (MDR) infections by specifically infecting and destroying bacterial strains without harming beneficial bacteria [19]. The CRISPR-Cas system has targeted various AMR genes, including blaOXA-232, blaNDM, blaCTX-M, ermB, vanA, mecA, fosA3, blaKPC, and mcr-1 [20], Some research has investigated the role of CRISPR in suppressing virulence genes such as enterotoxin genes, tsst1, and iutA in S. aureus and K.pneumoniae [20]. The priority of bacterial pathogen has classified by WHO and shown in Figure 1A [2].
Figure 2.
Priority of Bacteria Based on Antimicrobial Resistance: 2A. Critical priority pathogens include bacteria pose the most significant threat due to their high resistance levels and limited treatment options. High priority pathogens include bacteria exhibit increasing resistance and can cause severe infections. Medium priority pathogens – While they are not as resistant as those in the higher categories but still need urgent attention. 2B. The enterobacteria have displaced Pseudomonas aeruginosa as a critical priority, dark brown (Left side) and have been promoted to high priority categories according to the WHO 2024 report while new pathogens, some as depicted as light brown circle (Right side) have been added to high priority categories.
Figure 2.
Priority of Bacteria Based on Antimicrobial Resistance: 2A. Critical priority pathogens include bacteria pose the most significant threat due to their high resistance levels and limited treatment options. High priority pathogens include bacteria exhibit increasing resistance and can cause severe infections. Medium priority pathogens – While they are not as resistant as those in the higher categories but still need urgent attention. 2B. The enterobacteria have displaced Pseudomonas aeruginosa as a critical priority, dark brown (Left side) and have been promoted to high priority categories according to the WHO 2024 report while new pathogens, some as depicted as light brown circle (Right side) have been added to high priority categories.
Risk of Antibiotics and Antibiotics Resistance:
Antibiotics is frequently used in the primary treatment for bacterial infections, and their efficacy is being increasingly undermined by the growing problem of antimicrobial resistance. The choice of antibiotics for treatment management depends on the host and parasites, viz; Isoniazid and Rifampicin are antibiotics used to manage tuberculosis infections in humans while certain antibiotics, such as ionophores and flavophospholipol, are specifically employed in veterinary medicine. Meanwhile, various classes of antibiotics are regularly prescribed in both human medicine and veterinary practice [21,106]. In addition to their use in medicine and veterinary practices, antibiotics like streptomycin and tetracyclines are utilized in horticulture for disease prevention or treatment, including the management of bacterial infections such as fire blight in apple and pear orchards [22].
In human medicine, antibiotics are mainly prescribed to treat infections and, in some cases, as a preventive measure, such as post-surgical prophylaxis. However, their usage differs between companion animals and food-producing animals in veterinary settings. In pets, antibiotic prescriptions closely resemble those in human healthcare [23]. Conversely, in livestock production, antibiotics are employed not just for treating infections but also as feed additives and growth promoters. Unlike in animals, the application of prophylactic antibiotics in humans is less frequent and is primarily reserved for serious infectious diseases such as meningococcal infections. In such instances, antibiotics are given solely to individuals who have had prolonged contact with infected persons [24]. The use of antibiotics as growth promoters in food animals has been recognized as a significant factor in antimicrobial resistance, since extended sub-therapeutic exposure encourages the development and dissemination of resistant bacteria among animal populations. These microorganisms, which are resistant, can subsequently be passed on to humans through either direct exposure from the environment or through the food supply [23].
Recognizing these risks, the FAO, OIE, and WHO have recommended against using antibiotics for growth promotion as part of global efforts to combat AMR. Consequently, several countries, including those in Europe, Canada, and the United States, have developed and enforced regulations to restrict antibiotic use in food production [23]. The ten of most deadly pathogens showed resistance to common antibiotics has been enumerated in percentage and shown in Figure 3 A.
A superbug refers to a bacterial strain that has developed resistance to several antibiotics, complicating treatment with conventional medications. These organisms, which are resistant to multiple drugs, can lead to severe infections in both humans and animals, particularly within hospitals and healthcare environments. The emergence of superbugs can be attributed mainly to the excessive and inappropriate use of antibiotics, which is increasingly jeopardizing global public health. The resistance associated factors can be accumulated and acts as a reservoir as shown in Figure 4C. The basic strategies to mitigate ABR are recommended and illustrated in Figure 4D. The diagram emphasizes major sources and routes for the transmission and dissemination of antibiotic-resistant bacteria, including wastewater, livestock, aquaculture, healthcare, and industrial effluents. Certain genes, such as AmpC (AmpC β-lactamases), blaSHV (SHV-type β-lactamases), TrxA (thioredoxin), and TrxB (thioredoxin reductase), contribute to intrinsic resistance against antibiotics like β-lactams, aminoglycosides, rifampicin, triclosan, and fluoroquinolones [25]. On the other hand, acquired resistance occurs when bacteria develop new ways to defend themselves against antibiotics. This can happen through mechanisms such as preventing antibiotic penetration, actively pumping the drugs out (efflux), modifying drug targets, or breaking down antibiotics using enzymes Figure 4A [26]. Bacteria can develop and acquire antibiotic resistance not just through the horizontal transfer of resistance genes (via plasmids, transposons, and integrons) but also through mutations in their chromosomal DNA, as illustrated in Fig.4B. These spontaneous genetic mutations can occur naturally during replication or be triggered by environmental pressures, such as the presence of antibiotics. Bacteria that possess such mutations are more likely to thrive and reproduce when these mutations provide a survival advantage, thereby transmitting these resistant characteristics to their descendants. chromosomal mutations serve as a basic and adaptive method for bacteria to develop resistance. Although these genetic alterations spread at a slower rate than plasmid-mediated genes, they are frequently irreversible and can result in significant drug resistance, creating major obstacles for treatment and infection management [27,28,105].
The rise of multidrug-resistant (MDR) bacteria with acquired resistance is a growing global health concern. Some well-known antibiotic resistance genes (ARGs), such as blaTEM, blaCTX-M, blaKPC, blaNDM, blaVIM, and mcr-1, have been linked to the spread of MDR strains of E. coli and P. aeruginosa [29]. The mcr-1 gene, which confers resistance to colistin, is a significant example of how antibiotic resistance spreads through the One Health framework. It was first identified in raw meat samples from China and later found in E. coli isolates from both migratory birds and humans [30,31]. Pathogenic bacteria have gradually developed antimicrobial resistance (ABR) over time because of evolutionary processes shaped by various factors [11,32]. Key contributors to the global distribution of antimicrobial resistance (ABR) within a One Health framework include intensive food production systems, the globalization of food supply chains, international travel (which aids in the dissemination of resistance genes), climate change, population growth, and urbanization. The growing burden of ABR is closely associated with several factors, including the overuse of antibiotics in both humans and animals (such as livestock, pets, and aquaculture), the availability of antibiotics without prescriptions, increased international trade and travel, movements of migratory birds, displaced populations like refugees, environmental changes, poor sanitation and hygiene practices, and the introduction of unmetabolized antibiotics or their residues into ecosystems [10,11,33].These factors play a role in the genetic selection pressure on bacteria, resulting in both vertical and horizontal transmission of resistance genes among bacterial species. This mechanism encourages the dissemination of antimicrobial resistance genes (ARGs) in the environment and aids in the growth of multidrug-resistant (MDR) pathogens within communities [33]. The real volume of antibiotics used as medications is often accessible without a prescription or available over the counter. This unrestricted access plays a role in the emergence of antibiotic-resistant bacteria [34,35]. Nonetheless, study indicates that the lowering of endemic diseases and mortality rates in African nations has been greatly aided by the expanded availability of antibiotics, improved sanitation, and vaccination campaigns [36]. Plasmids, transposons, and integrons also aid in the dissemination of resistance genes. In order to promote co-selection under antibiotic pressure, the blaNDM-1 gene, which codes for the NDM-1 metallo-β-lactamase, is usually found on plasmids that carry other resistance determinants (bleMBL for bleomycin resistance) [37,38,39]. By capturing and expressing resistance cassettes through site-specific recombination, integrons in A. baumannii allow for quick adaptation to novel medications [40].
To address the growing threat of antimicrobial resistance (ABR), it is essential to reduce global antibiotic usage. Antibiotic use recommendations may require customization according to geographic factors and the incidence of infectious diseases. In the fields of agriculture and animal health, antibiotics are also commonly employed. It is estimated that around 66% of all antibiotics used worldwide are consumed in animal production. Nevertheless, numerous nations have enacted limitations on their application as growth promoters. It was estimated that 131,000 tons of antibiotics were used in global livestock production in 2013, with projections indicating this amount will increase to 200,000 tons by 2030 [41,42].
ESKAPE Pathogens Antibiotic Resistance and Mechanisms:
The six highly virulent and antibiotic-resistant bacterial pathogens that make up ESKAPE are the main contributors to antibiotic resistance worldwide. They do this by producing enzymes (ESBL, carbapenemases), altering their targets (PBP2a, Van genes), using efflux pumps, losing porin, and forming biofilms.
Currently the term “superbug” is usually applied to the ESKAPE pathogens, which are named for their ability to escape antibiotic effects [43,44]. The behaviour of pathogens against antibiotics is shown and listed in Figure 5. Their appearance reflects a trend of human mistakes and its excessive use. The World Health Organization issued a warning in 2014 that antimicrobial resistance could lead to a “post-antibiotic era”. By 2016, the United Nations had compared its global threat level to that of climate change in annual report. As of 2021, AMR was accountable for 1.14 million deaths annually, surpassing the death tolls of HIV/AIDS and malaria, with forecasts estimating that this figure could reach 10 million by 2050 [1,2].
Alternatives Strategies and Adjuvants for ABR Treatments:
In the treatment of MDR infections, a fundamental strategy is to combine antibiotics with agents that have a synergistic effect. Pairs of β-lactam and β-lactamase inhibitors, like ceftazidime-avibactam, get around resistance by shielding the antibiotic from being broken down by enzymes. Avibactam, which is a diazabicyclooctane (DBO) inhibitor, inactivates Class A (e.g., KPC), Class C (AmpC), and certain Class D (OXA-48) β-lactamases, thereby restoring the effectiveness of ceftazidime against carbapenem-resistant Enterobacteriaceae [45,46,103]. Likewise, meropenem-vaborbactam aims at K. pneumoniae carbapenemase, showing over 90% effectiveness in clinical trials [47,48]. In the case of Gram-negative “superbugs” such as A. baumannii, combinations of colistin and rifampicin are effective in disrupting biofilms and increasing bactericidal activity [49,50]. First-generation inhibitors like clavulanic acid and tazobactam are ineffective against MBLs. Novel agents like aspergillomarsamine A (AMA), a fungal-derived chelator, strip zinc ions from NDM-1, restoring meropenem’s activity [51]. Ebselen, an organoselenium compound, covalently modifies the active site of NDM-1, reducing carbapenem MICs by 4–35-fold in K. pneumoniae [52,53].
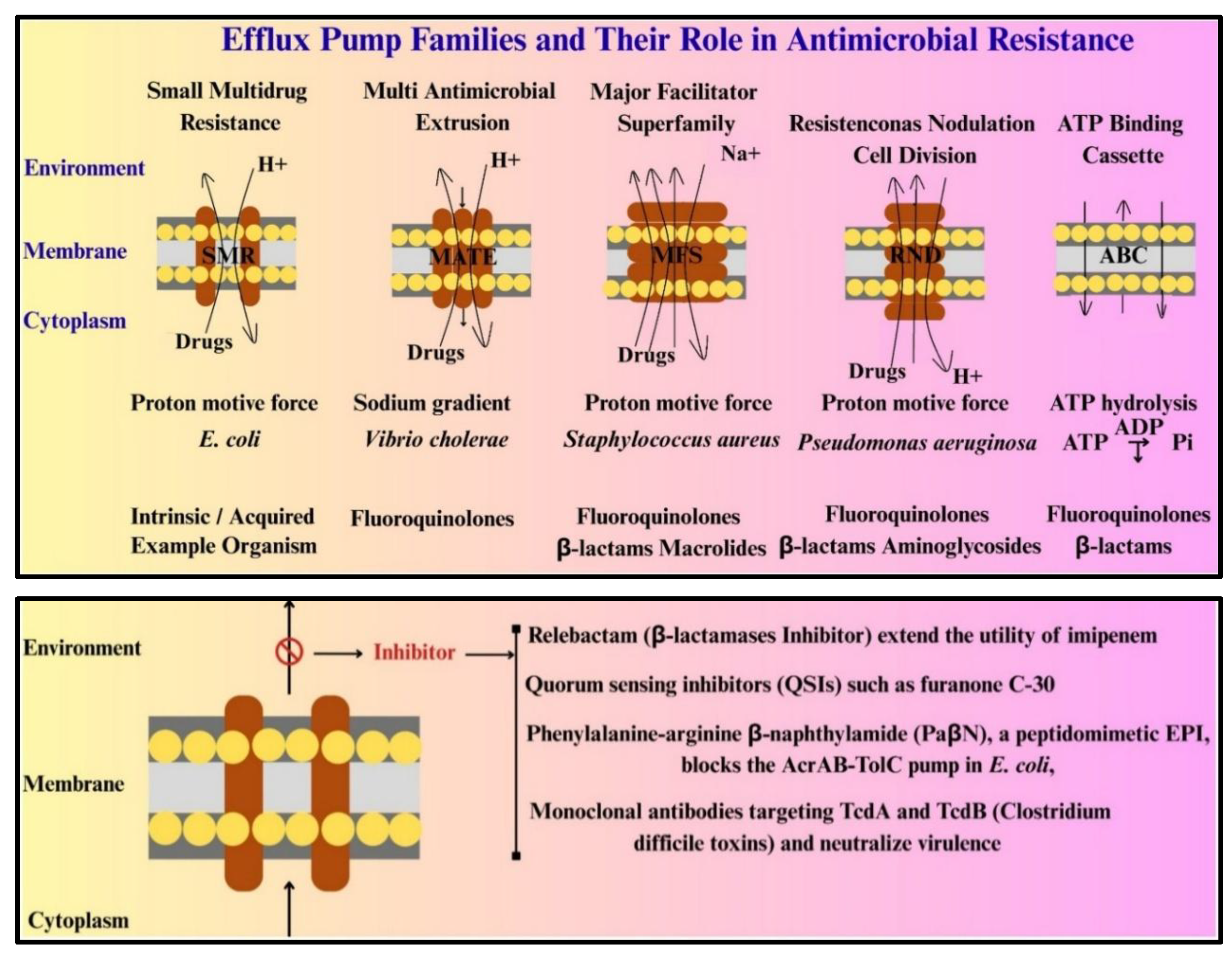
Five fundamental families of efflux pumps—SMR (Small Multidrug Resistance), MATE (Multi Antimicrobial Extrusion), MFS (Major Facilitator Superfamily), RND (Resistance Nodulation and cell Division), and ABC (ATP Binding Cassette) play a crucial role in both intrinsic and acquired resistance. Exemplified by AcrAB-TolC in E. coli, the RND family creates tripartite complexes that span the inner membrane, periplasm, and outer membrane. These pumps expel a variety of substrates, such as β-lactams, fluoroquinolones, and tetracyclines, utilizing proton motive force [43,44]. The overexpression of acrAB, which is controlled by transcriptional activators such as MarA and SoxS, has been associated with ciprofloxacin resistance in urinary tract infections [54,55]. Plasmid-borne pumps like OqxAB in Klebsiella pneumoniae make treatment even more challenging. The oqxAB operon, which is often bordered by IS26 transposons, disseminates quickly via horizontal gene transfer (HGT). However, natural compounds like reserpine and 5’-methoxyhydrocarpin inhibit NorA and MFS pumps in S. aureus, reversing resistance to ciprofloxacin and erythromycin [56,57,58]. Synthetic EPIs, such as MBX2319, target the substrate-binding pocket of AcrB (a component of AcrAB-TolC), enhancing levofloxacin activity against MDR Enterobacteriaceae. Despite progress, clinical translation remains hampered by toxicity; arylpiperazines, for instance, exhibit off-target serotonin receptor agonism [59,60].
Adjuvants are Non-Antibiotic Compounds That Potentiate Antimicrobial Activity:
| Class I (Anti-resistance agents) | These include efflux pump inhibitors (EPIs) and β-lactamase inhibitors. Phenylalanine-arginine β-naphthylamide (PaβN), a peptidomimetic EPI, blocks the AcrAB-TolC pump in E. coli, increasing intracellular concentrations of fluoroquinolones and tetracyclines [61]. For β-lactamases, next-generation inhibitors like relebactam (a Diazabicyclooctane (DBO derivative) extend the utility of imipenem against MBL-producing strains [62]. |
| Class II (Anti-virulence agents) | Quorum sensing inhibitors (QSIs) such as furanone C-30 disrupt P. aeruginosa biofilm formation by mimicking AHL signals, reducing pyocyanin and elastase production [63,64]. Monoclonal antibodies targeting Clostridium difficile toxins TcdA and TcdB neutralize virulence without affecting bacterial growth, offering a pathogen-specific approach [65]. |
Figure 6.
Efflux pump regulation in antimicrobial resistance bacteria; Antimicrobial resistance-related efflux pump families, displaying their energy sources, representative bacterial species, and related antibiotic classes. MATE (Multi Antimicrobial Extrusion) uses a sodium gradient; RND (Resistance-Nodulation-Division) uses a proton motive force across inner and outer membranes; ABC (ATP-Binding Cassette) uses ATP hydrolysis; and SMR (Small Multidrug Resistance) and MFS (Major Facilitator Superfamily) use a proton motive force. The examples of organisms and representative antibiotic resistances are illustrated above for every family. The antibacterial potential of drugs can be increased by employing various efflux inhibitors to alter the resistance property of microorganisms as shown in lower panel.
Figure 6.
Efflux pump regulation in antimicrobial resistance bacteria; Antimicrobial resistance-related efflux pump families, displaying their energy sources, representative bacterial species, and related antibiotic classes. MATE (Multi Antimicrobial Extrusion) uses a sodium gradient; RND (Resistance-Nodulation-Division) uses a proton motive force across inner and outer membranes; ABC (ATP-Binding Cassette) uses ATP hydrolysis; and SMR (Small Multidrug Resistance) and MFS (Major Facilitator Superfamily) use a proton motive force. The examples of organisms and representative antibiotic resistances are illustrated above for every family. The antibacterial potential of drugs can be increased by employing various efflux inhibitors to alter the resistance property of microorganisms as shown in lower panel.
Anti-Virulence and Gene Editing Strategies:
Salicylidene acylhydrazides inhibit type III secretion systems (T3SS) in Gram-negative bacterium, blocking effector protein delivery to host cells [66,67]. In P. aeruginosa, the anti-quorum sensing agent hamamelitannin disperses biofilms by interfering with peptidoglycan remodelling, sensitizing bacteria to vancomycin [68]. Gene editing and transgenic methods provide new ways to combat antibiotic-resistant bacteria. These techniques have been applied to control bacterial infections such as mastitis, which requires extensive antibiotic treatment. Engineered lysozyme and lysostaphin have been used as alternatives to antibiotics for this purpose. While these approaches are still undergoing regulatory evaluation, they hold promise as potential substitutes for antibiotics [69,70]. Whole genome sequencing (WGS)-based mutagenesis is also used to identify viral genes that affect replication, helping researchers find new gene-editing targets [71,72]. Genome-based tools like RNA sequencing (RNAseq) can identify bacterial regulators, virulence factors, and genes involved in infection. Instead of killing bacteria, anti-virulence (AV) inhibitors work by preventing the expression of genes responsible for bacterial infections. For example, virstatin is an AV inhibitor that reduces virulence in Vibrio cholerae by downregulating the ToxT regulator gene [73,74].
Vaccines and Antibiotic Resistance:
Vaccines are crucial in the fight against antibiotic resistance because they reduce the need for antibiotic use [75]. In addition to lowering the need for antibiotics, vaccinations also limit the emergence and spread of antibiotic resistance by lowering the prevalence of infectious illnesses. Effective vaccines can be developed to stop resistant bacterial infections, which have the potential to dramatically raise the illness burden in both high- and low-income countries. The possible obstacles to vaccine development that need to be overcome to forecast the potential impact of next-generation vaccines on AMR. The development of vaccines benefits from the use of sophisticated genomic methods like comparative and functional genomics. These tools aid in the identification of conserved pathogen components that can be tested as subunit vaccines and targeted by antimicrobials agents [76,77,78]. There are numerous studies that demonstrate the effectiveness of vaccines in preventing antibacterial resistance, such as the reduction of antibiotic-resistant S. pneumoniae infections in both adults and children by the Pneumococcal Conjugate Vaccine (PCV), the significant reduction of resistant Hib cases globally by the Haemophilus influenzae type b (Hib) Vaccine, the reduction of inappropriate antibiotic use for viral respiratory infections by the Influenza Vaccine, and the prevention of drug-resistant Salmonella typhi by the Typhoid conjugate vaccine (TCV) [79,80,81,82].
The virulence components that bacterial pathogens express, including polysaccharides, toxins, adhesins, and effector proteins, can be bound by monoclonal antibodies (mAbs). A bispecific mAb intended to eliminate a bacterial virulence factor and a combination of two mAbs that target the exopolysaccharide of Pseudomonas aeruginosa are among the most sophisticated examples [83]. The fact that mAbs normally only detect one target, but bacterial pathogenesis is typically multivariate, presents a significant hurdle when employing them to treat bacterial infections. However, recent developments in technology have overcome this constraint. To improve complement activation, for example, the Fc component can be engineered to create hexamers (HexaBody technology), as shown against Neisseria gonorrhoeae. Bispecific monoclonal antibodies have also been developed to target Pseudomonas aeruginosa, while potent monoclonal antibodies have been generated to neutralize Clostridium difficile toxins A and B [84]. In addition, bioconjugation has emerged as a promising strategy for addressing antimicrobial resistance and advancing vaccine development.
Bacteriophage Therapy:
One effective method of addressing antimicrobial resistance (AMR) is bacteriophage therapy. Viruses known as bacteriophages, selectively infect and lyse bacteria without endangering human cells or the good microbiota. The high specificity of phages, in contrast to traditional antibiotics, enables them to target resistant bacterial strains with the least amount of collateral damage [85]. Phages and modified phage proteins, such as endolysins and depolymerases, have gained attention as a means of combating bacterial infections due to the growing issue of antibiotic resistance [86,87,88]. Phages may be used in aquaculture, veterinary medicine, and food safety, including the management of E. coli O157:H7 in animals [89,90].
To overcome newly developed resistance mechanisms, they can also co-evolve with bacteria. Developments in synthetic biology and genetic engineering have significantly improved phage therapy by allowing the creation of tailored or cocktail formulations that increase potency and host range. Phage treatment is gradually being acknowledged as a potential supplement or substitute for antibiotics in the fight against AMR, despite the fact that there are still obstacles to overcome, including regulatory approval, large-scale manufacture, and stability assurance. Phage therapy has shown promise in the fight against bacterial pathogens that are resistant to antibiotics, such as Pseudomonas aeruginosa and Staphylococcus aureus [91,92]. To increase the effectiveness of treatment for infections brought on by P. aeruginosa and Clostridium difficile, researchers are looking into localized delivery techniques such oral administration into the gastrointestinal tract or direct inhalation into the lungs [93,94]. Phage formulations can be made using encapsulation or adsorption to increase stability and efficacy. Additionally, phages have the potential to be CRISPR–Cas systems delivery vehicles, allowing for precise genetic targeting of resistance determinants and bacterial pathogens [85].
Technological Advancements:
Antibiotic resistance is a growing global health issue, and advanced tools are needed to study its underlying causes. Metagenomics, a sequencing-based approach for analyzing microbial communities, is widely used to study antibiotic resistance. This technique helps researchers to understand how bacteria acquire ARGs through horizontal gene transfer (HGT) and track the spread of resistance in different environments [95]. Metagenomics also allows researchers to study microbial diversity by reconstructing complete genomes and detecting species abundance [96,97]. Since it does not require culturing bacteria, it is a practical tool for environmental research.
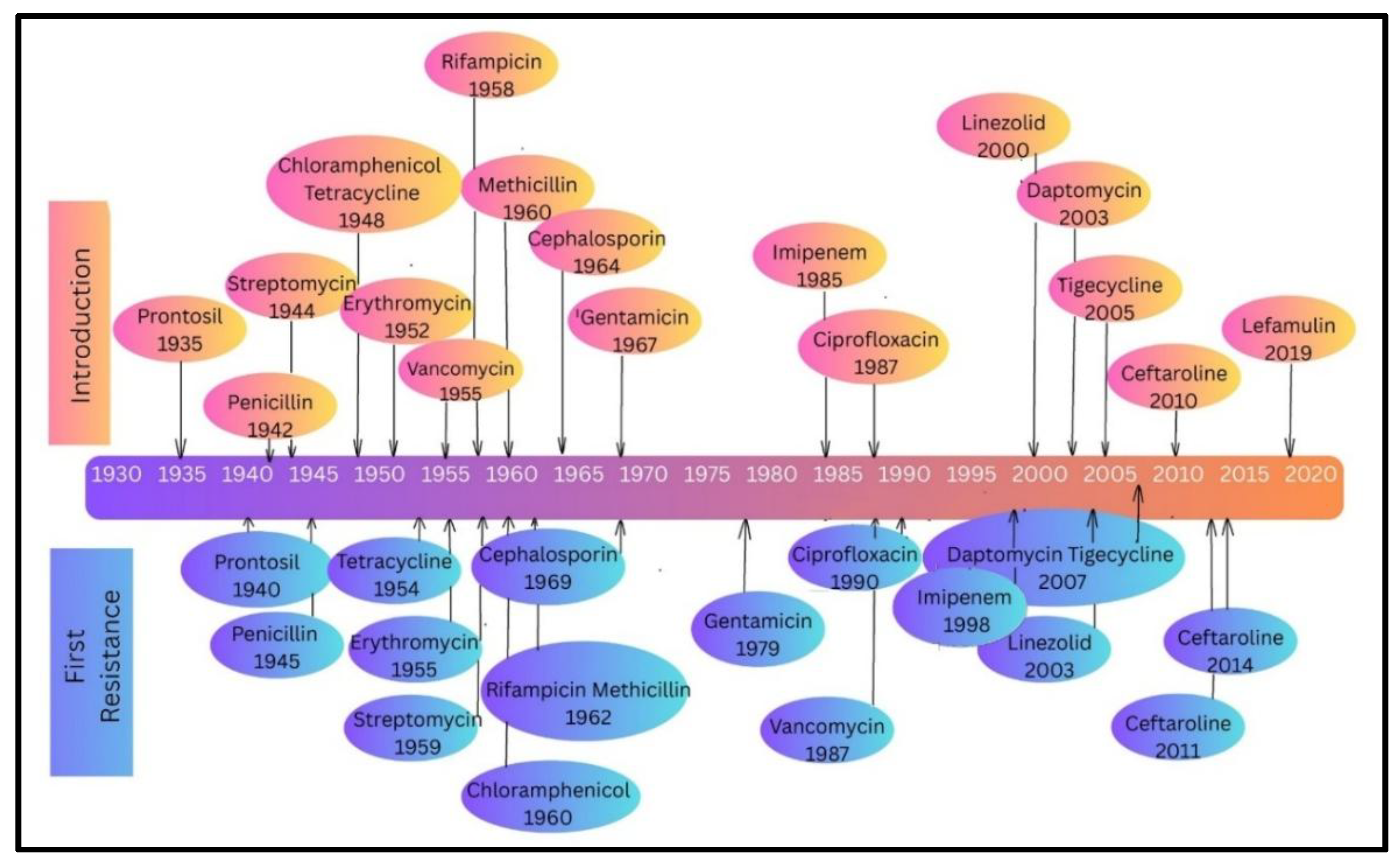
Figure 7.
History of antibiotics and its resistance: The timeline displays the year that bacterial resistance to antibiotics was first documented (blue-purple ovals, bottom) and the introduction of different antibiotics (pink-orange ovals, top). Every antibiotic has a vertical connection to the resistance year that corresponds to it. The gradient colour bar indicates when the specific antibiotic was first used in clinical settings and when resistance to that antibiotic was initially identified.
Figure 7.
History of antibiotics and its resistance: The timeline displays the year that bacterial resistance to antibiotics was first documented (blue-purple ovals, bottom) and the introduction of different antibiotics (pink-orange ovals, top). Every antibiotic has a vertical connection to the resistance year that corresponds to it. The gradient colour bar indicates when the specific antibiotic was first used in clinical settings and when resistance to that antibiotic was initially identified.
The metagenomics may not detect rare bacterial populations carrying ARGs, and its ability to identify bacterial variants with different resistance traits is limited. Combining metagenomics with functional genomics can help overcome these limitations [98]. Whole genome sequencing (WGS) and next-generation sequencing (NGS) have greatly improved research on antibiotic resistance across different domains of One Health. WGS is a powerful tool for studying antibiotic resistance and predicting bacterial resistance profiles. However, inconsistencies between WGS-based predictions and traditional culture-based testing need to be resolved. Identifying ARGs in different One Health sectors is crucial due to the complexity of microbial communities. Recent studies have highlighted the role of mobile genetic elements (MGEs) in spreading antibiotic resistance across various environments [99,100].
The rise of antibiotic-resistant "superbugs" represents one of the most pressing challenges in modern medicine. Traditional antibiotics, once hailed as miracle drugs, are increasingly rendered ineffective due to the rapid evolution of bacterial defence mechanisms. In this landscape, CRISPR-Cas systems have emerged as a revolutionary tool to combat antimicrobial resistance (AMR). This section explores the current state of CRISPR-Cas antimicrobial development, design principles, delivery mechanisms, and the ecological challenges that influence their efficacy.
Artificial Intelligence and Machine Learning Use in Surveillance of ABR:
The artificial intelligence (AI) and machine learning (ML), which provide quick, sophisticated, evidence-based solutions that improve decision-making by combining clinical information, microbiome insights, epidemiological and lab data in healthcare [101]. AI technologies are particularly promising in tackling complex global health concerns like antimicrobial resistance (AMR) because they can quickly analyse large amounts of data and produce logical, well-informed outputs more precisely [102]. Large-scale data management and interpretation capabilities make AI/ML systems ideal for addressing AMR-related issues around the world. They can predict which individuals are at risk of acquiring or already have antibiotic-resistant illnesses by analysing complex health records [103]. Moreover, by effectively screening large chemical compound libraries for potency against resistant infections, these methods aid in medication discovery, drug repurposing, and combination therapy [104]. By determining the transmission of AMR genes, evaluating risk factors, and suggesting novel diagnostic and treatment approaches, AI/ML systems also support public health. Artificial intelligence and machine learning (AI/ML) could reduce antimicrobial resistance (AMR) using a variety of learning strategies, as illustrated in Figure 8. These models can provide insight into combination therapy, the creation of novel antimicrobials, and the repurposing of current drugs by analysing their molecular structures.
Figure 8.
Two main tactics to fight antimicrobial resistance (AMR) are shown. (A)Conventional methods of controlling AMR using conventional medical and public health measures (B)New therapeutic techniques to fight antibiotic resistance.
Figure 8.
Two main tactics to fight antimicrobial resistance (AMR) are shown. (A)Conventional methods of controlling AMR using conventional medical and public health measures (B)New therapeutic techniques to fight antibiotic resistance.
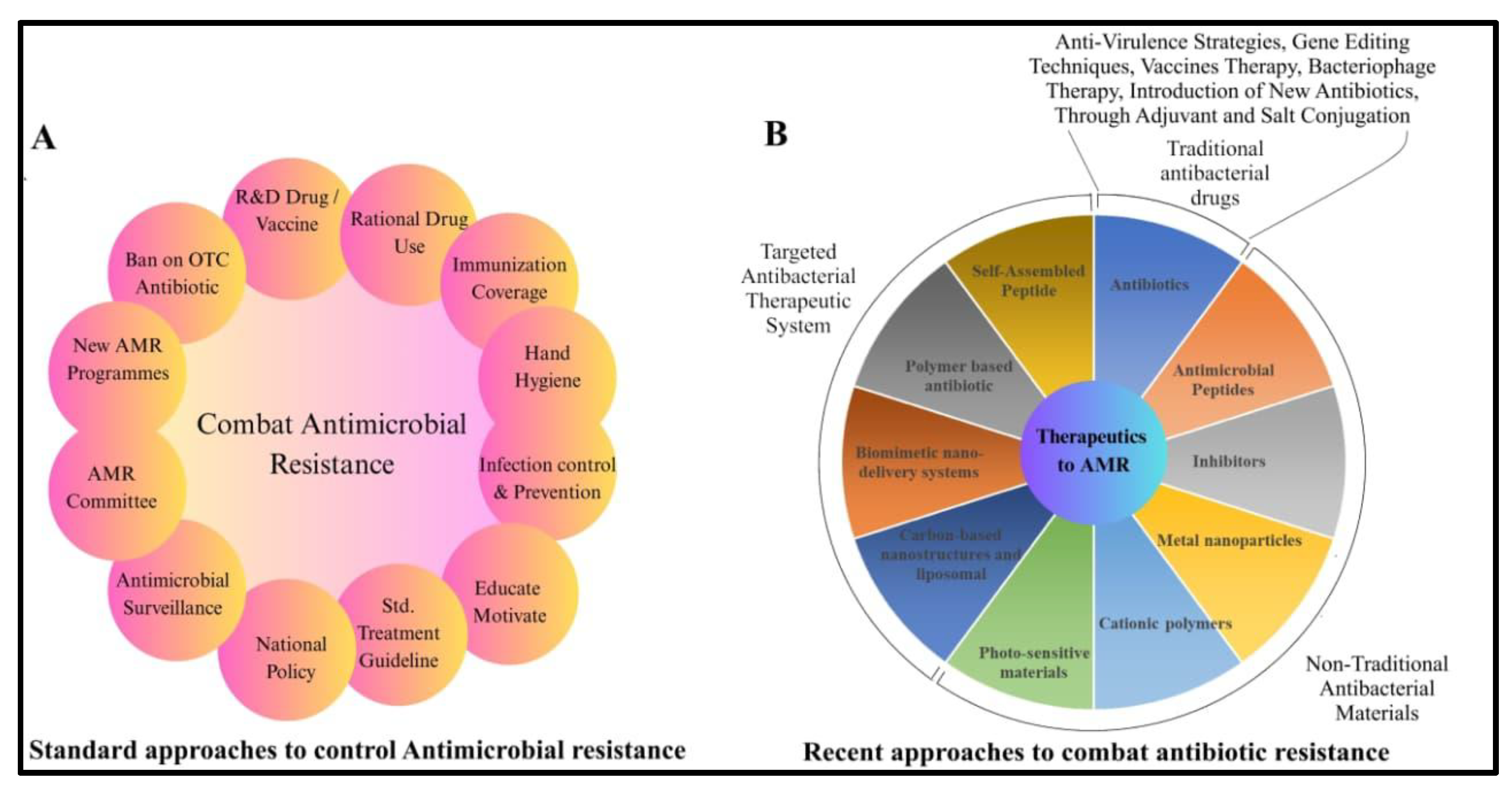
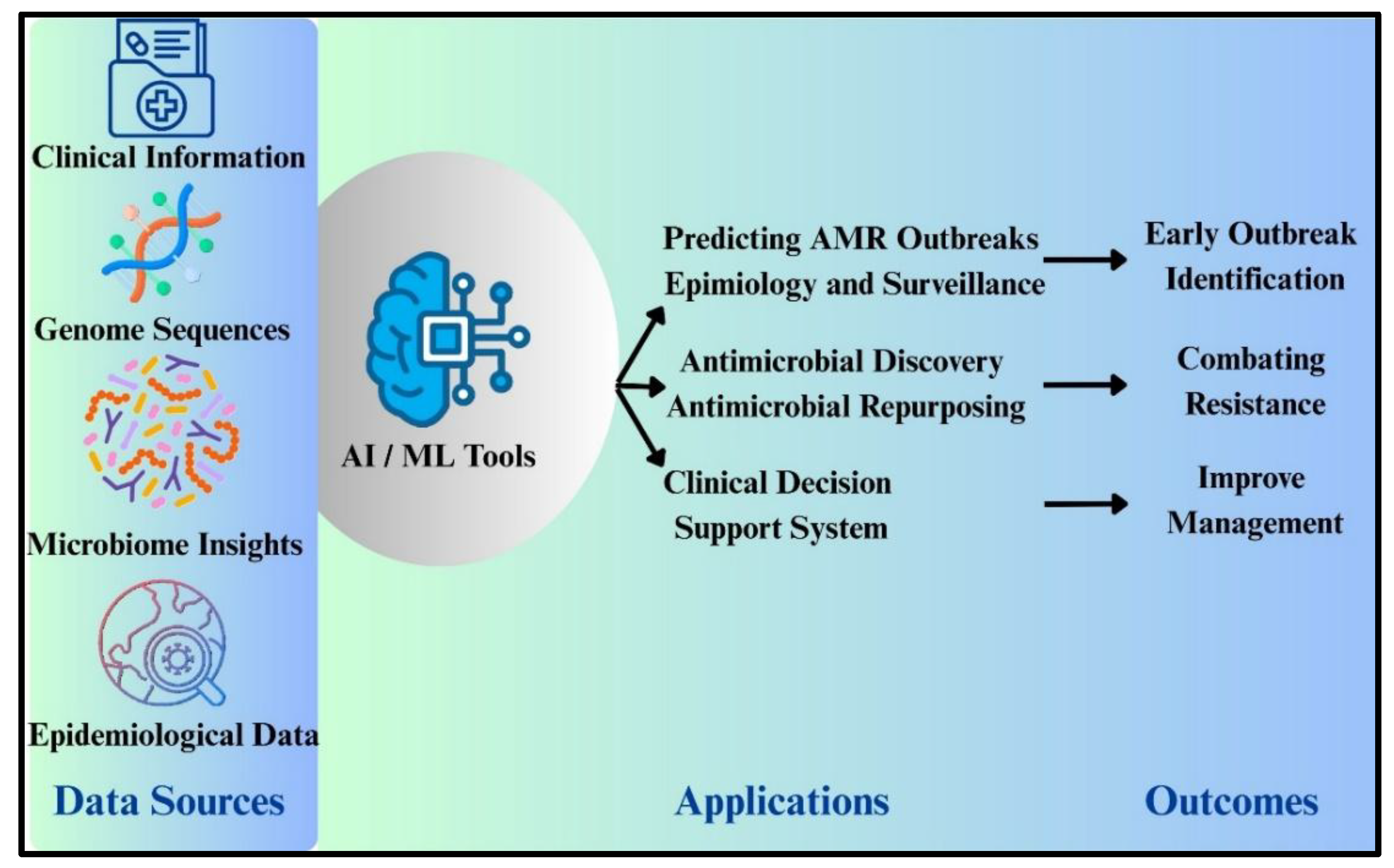
Figure 9.
Artificial intelligence and machine learning revolutionize healthcare by combining clinical and lab research as a data source to produce useful results like effectively managing resistance and identifying outbreaks, repurposing the drugs.
Figure 9.
Artificial intelligence and machine learning revolutionize healthcare by combining clinical and lab research as a data source to produce useful results like effectively managing resistance and identifying outbreaks, repurposing the drugs.
Conclusion and Future Prospective:
Multidrug-resistant (MDR) bacteria constitute a serious and rising global health concern, compromising the effectiveness of current antibiotics and endangering decades of medical progress. Increases in morbidity, mortality, and healthcare expenses are caused by MDR pathogens like K. pneumoniae, A. baumannii, and P. aeruginosa, which are fuelled by antibiotic abuse, poor infection management, and a lack of therapeutic innovation. Strengthened surveillance, careful use of antibiotics, and quick development of new treatments are all necessary to counter this menace. A coordinated One Health strategy that incorporates environmental, animal, and human health interventions is needed to combat MDR bacteria. Developing next-generation medications and alternative treatments, implementing quick diagnostics, improving worldwide resistance monitoring, implementing antimicrobial stewardship regulations, increasing public awareness, and lowering environmental antibiotic contamination are among the top priorities. To ensure that antibiotics continue to be effective for upcoming generations, strategic, cooperative work is necessary.
Author Contributions
Souvik and Tiyasa are co–first authors, each having made significant contributions to the review article presented in this paper. The study was designed, analyzed, and interpreted by DKS. All authors have reviewed the manuscript and agree to take responsibility for the integrity and accuracy of the work.
Data Sharing Statement
The datasets used and analysed during the current study available from the corresponding author on reasonable request.
Disclosure
All authors have no conflicts of interest regarding this work.
References
- Sati H, Carrara E, Savoldi A, et al. The WHO Bacterial Priority Pathogens List 2024: a prioritisation study to guide research, development, and public health strategies against antimicrobial resistance. Lancet Infect Dis. Published online September 1, 2025. [CrossRef]
- Organisation Mondiale de la Santé. WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Saudi Med J. 2017;(February).
- Ahmed SK, Hussein S, Qurbani K, et al. Antimicrobial resistance: Impacts, challenges, and future prospects. Journal of Medicine, Surgery, and Public Health. 2024;2. [CrossRef]
- Marti E, Variatza E, Balcazar JL. The role of aquatic ecosystems as reservoirs of antibiotic resistance. Trends Microbiol. 2014;22(1). [CrossRef]
- Huijbers PMC, Blaak H, De Jong MCM, Graat EAM, Vandenbroucke-Grauls CMJE, De Roda Husman AM. Role of the Environment in the Transmission of Antimicrobial Resistance to Humans: A Review. Environ Sci Technol. 2015;49(20). [CrossRef]
- Woolhouse MEJ, Ward MJ. Sources of antimicrobial resistance. Science (1979).American Association for the Advancement of Science. 2013;341(6153):1460-1461. [CrossRef]
- Holmes AH, Moore LSP, Sundsfjord A, et al. Understanding the mechanisms and drivers of antimicrobial resistance. The Lancet. 2016;387(10014). [CrossRef]
- Aslam B, Khurshid M, Arshad MI, et al. Antibiotic Resistance: One Health One World Outlook. Front Cell Infect Microbiol. 2021;11. [CrossRef]
- Cabrera-Pardo JR, Lood R, Udekwu K, et al. A One Health – One World initiative to control antibiotic resistance: A Chile - Sweden collaboration. One Health. 2019;8. [CrossRef]
- Sassi A, Basher NS, Kirat H, et al. The Role of the Environment (Water, Air, Soil) in the Emergence and Dissemination of Antimicrobial Resistance: A One Health Perspective. Antibiotics. 2025;14(8):764. [CrossRef]
- Pandey S, Doo H, Keum GB, et al. Antibiotic resistance in livestock, environment and humans: One Health perspective. J Anim Sci Technol. 2024;62(2). [CrossRef]
- Burow E, Käsbohrer A. Risk Factors for Antimicrobial Resistance in Escherichia coli in Pigs Receiving Oral Antimicrobial Treatment: A Systematic Review. Microbial Drug Resistance. 2017;23(2). [CrossRef]
- Naghavi M, Vollset SE, Ikuta KS, et al. Global burden of bacterial antimicrobial resistance 1990–2021: a systematic analysis with forecasts to 2050. The Lancet. 2024;404(10459):1199-1226. [CrossRef]
- Lee AS, De Lencastre H, Garau J, et al. Methicillin-resistant Staphylococcus aureus. Nat Rev Dis Primers. 2018;4. [CrossRef]
- Manyi-Loh C, Mamphweli S, Meyer E, Okoh A. Antibiotic use in agriculture and its consequential resistance in environmental sources: Potential public health implications. Molecules. 2018;23(4). [CrossRef]
- Li Y, Sun QL, Shen Y, et al. Rapid increase in prevalence of carbapenem-resistant enterobacteriaceae (CRE) and emergence of colistin resistance gene mcr-1 in CRE in a hospital in Henan, China. J Clin Microbiol. 2018;56(4). [CrossRef]
- Ramadan AA, Abdelaziz NA, Amin MA, Aziz RK. Novel blaCTX-M variants and genotype-phenotype correlations among clinical isolates of extended spectrum beta lactamase-producing Escherichia coli. Sci Rep. 2019;9(1). [CrossRef]
- Kumar V, Yasmeen N, Pandey A, et al. Antibiotic adjuvants: synergistic tool to combat multi-drug resistant pathogens. Front Cell Infect Microbiol. 2023;13. [CrossRef]
- Mafe AN, Büsselberg D. Phage Therapy in Managing Multidrug-Resistant (MDR) Infections in Cancer Therapy: Innovations, Complications, and Future Directions. Pharmaceutics.Multidisciplinary Digital Publishing Institute (MDPI). 2025;17(7). [CrossRef]
- Okesanya OJ, Ahmed MM, Ogaya JB, et al. Reinvigorating AMR resilience: leveraging CRISPR–Cas technology potentials to combat the 2024 WHO bacterial priority pathogens for enhanced global health security—a systematic review. Trop Med Health.BioMed Central Ltd. 2025;53(1). [CrossRef]
- Zenner D, Beer N, Harris RJ, Lipman MC, Stagg HR, Van Der Werf MJ. Treatment of latent tuberculosis infection: An updated network meta-analysis. Ann Intern Med.American College of Physicians. 2017;167(4):248-255. [CrossRef]
- Vidaver AK. Uses of antimicrobials in plant agriculture. Clinical Infectious Diseases. 2002;34(SUPPL. 3). [CrossRef]
- McEwen SA, Collignon PJ. Antimicrobial Resistance: a One Health Perspective. Microbiol Spectr. 2018;6(2). [CrossRef]
- Kasimanickam V, Kasimanickam M, Kasimanickam R. Antibiotics Use in Food Animal Production: Escalation of Antimicrobial Resistance: Where Are We Now in Combating AMR? Medical sciences. 2021;9(1). [CrossRef]
- Kakoullis L, Papachristodoulou E, Chra P, Panos G. Mechanisms of antibiotic resistance in important gram-positive and gram-negative pathogens and novel antibiotic solutions. Antibiotics. 2021;10(4). [CrossRef]
- Blair JMA, Webber MA, Baylay AJ, Ogbolu DO, Piddock LJV. Molecular mechanisms of antibiotic resistance. Nat Rev Microbiol. 2015;13(1). [CrossRef]
- Michaelis C, Grohmann E. Horizontal Gene Transfer of Antibiotic Resistance Genes in Biofilms. Antibiotics. 2023;12(2). [CrossRef]
- Rodríguez-Beltrán J, DelaFuente J, León-Sampedro R, MacLean RC, San Millán Á. Beyond horizontal gene transfer: the role of plasmids in bacterial evolution. Nat Rev Microbiol. 2021;19(6). [CrossRef]
- Aslam B, Wang W, Arshad MI, et al. Antibiotic resistance: a rundown of a global crisis. Infect Drug Resist. 2018;11. [CrossRef]
- Liu Z, Zhang TT, Yu J, et al. Excess Body Weight during Childhood and Adolescence Is Associated with the Risk of Multiple Sclerosis: A Meta-Analysis. Neuroepidemiology. 2016;47(2). [CrossRef]
- McFarland L V., Evans CT, Goldstein EJC. Strain-specificity and disease-specificity of probiotic efficacy: A systematic review and meta-analysis. Front Med (Lausanne). 2018;5(MAY). [CrossRef]
- Hernando-Amado S, Coque TM, Baquero F, Martínez JL. Defining and combating antibiotic resistance from One Health and Global Health perspectives. Nat Microbiol. 2019;4(9). [CrossRef]
- Cycoń M, Mrozik A, Piotrowska-Seget Z. Antibiotics in the soil environment—degradation and their impact on microbial activity and diversity. Front Microbiol. 2019;10(MAR). [CrossRef]
- Jim YK, Shakow A, Mate K, Vanderwarker C, Gupta R, Farmer P. Limited good and limited vision: Multidrug-resistant tuberculosis and global health policy. Soc Sci Med. 2005;61(4 SPEC. ISS.). [CrossRef]
- Auta A, Hadi MA, Oga E, et al. Global access to antibiotics without prescription in community pharmacies: A systematic review and meta-analysis. Journal of Infection. 2019;78(1). [CrossRef]
- Keenan JD, Bailey RL, West SK, et al. Azithromycin to Reduce Childhood Mortality in Sub-Saharan Africa. New England Journal of Medicine. 2018;378(17). [CrossRef]
- Yong D, Toleman MA, Giske CG, et al. Characterization of a new metallo-β-lactamase gene, bla NDM-1, and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob Agents Chemother. 2009;53(12). [CrossRef]
- Singh AR. Science, names giving and names calling: Change NDM-1 to PCM. Mens Sana Monogr. 2011;9(1). [CrossRef]
- Pollini S, Maradei S, Pecile P, et al. FIM-1, a new acquired metallo-β-lactamase from a Pseudomonas aeruginosa clinical isolate from Italy. Antimicrob Agents Chemother. 2013;57(1). [CrossRef]
- Bhat BA, Mir RA, Qadri H, et al. Integrons in the development of antimicrobial resistance: critical review and perspectives. Front Microbiol. 2023;14. [CrossRef]
- Aarestrup FM, Seyfarth AM, Emborg HD, Pedersen K, Hendriksen RS, Bager F. Effect of abolishment of the use of antimicrobial agents for growth promotion on occurrence of antimicrobial resistance in fecal enterococci from food animals in Denmark. Antimicrob Agents Chemother. 2001;45(7). [CrossRef]
- Van Boeckel TP, Brower C, Gilbert M, et al. Global trends in antimicrobial use in food animals. Proc Natl Acad Sci U S A. 2015;112(18). [CrossRef]
- Fernando DM, Kumar A. Resistance-Nodulation-Division multidrug efflux pumps in Gram-negative bacteria: Role in virulence. Antibiotics. 2013;2(1). [CrossRef]
- Sulavik MC, Houseweart C, Cramer C, et al. Antibiotic susceptibility profiles of Escherichia coli strains lacking multidrug efflux pump genes. Antimicrob Agents Chemother. 2001;45(4). [CrossRef]
- Buynak JD. β-Lactamase inhibitors: A review of the patent literature (2010 - 2013). Expert Opin Ther Pat. 2013;23(11). [CrossRef]
- Marshall S, Hujer AM, Rojas LJ, et al. Can ceftazidime-avibactam and aztreonam overcome β-lactam resistance conferred by metallo-β-lactamases in Enterobacteriaceae? Antimicrob Agents Chemother. 2017;61(4). [CrossRef]
- Marino A, Maraolo AE, Mazzitelli M, et al. Head-to-head: meropenem/vaborbactam versus ceftazidime/avibactam in ICUs patients with KPC-producing K. pneumoniae infections– results from a retrospective multicentre study. Infection. Published online 2025. [CrossRef]
- Guillot MF, Cerezuela MM, Ramírez P. New evidence in severe pneumonia: meropenemvaborbactam. Revista Espanola de Quimioterapia. 2022;35. [CrossRef]
- Petrosillo N, Chinello P, Proietti MF, et al. Combined colistin and rifampicin therapy for carbapenem-resistant Acinetobacter baumannii infections: Clinical outcome and adverse events [1]. Clinical Microbiology and Infection. 2005;11(8). [CrossRef]
- Lee HJ, Bergen PJ, Bulitta JB, et al. Synergistic activity of colistin and rifampin combination against multidrug-resistant acinetobacter baumannii in an in vitro pharmacokinetic/ pharmacodynamic model. Antimicrob Agents Chemother. 2013;57(8). [CrossRef]
- Sychantha D, Rotondo CM, Tehrani KHME, Martin NI, Wright GD. Aspergillomarasmine A inhibits metallo-β-lactamases by selectively sequestering Zn2+. Journal of Biological Chemistry. 2021;297(2). [CrossRef]
- Jin W Bin, Xu C, Cheung Q, et al. Bioisosteric investigation of ebselen: Synthesis and in vitro characterization of 1,2-benzisothiazol-3(2H)-one derivatives as potent New Delhi metallo-β-lactamase inhibitors. Bioorg Chem. 2020;100. [CrossRef]
- Lu Q, Cai Y, Xiang C, et al. Ebselen, a multi-target compound: Its effects on biological processes and diseases. Expert Rev Mol Med. 2021;23. [CrossRef]
- Albarri O, AlMatar M, Öcal MM, Köksal F. Overexpression of Efflux Pumps AcrAB and OqxAB Contributes to Ciprofloxacin Resistance in Clinical Isolates of K. pneumoniae. Curr Protein Pept Sci. 2022;23(5). [CrossRef]
- Weston N, Sharma P, Ricci V, Piddock LJV. Regulation of the AcrAB-TolC efflux pump in Enterobacteriaceae. Res Microbiol. 2018;169(7-8). [CrossRef]
- Bharatham N, Bhowmik P, Aoki M, et al. Structure and function relationship of OqxB efflux pump from Klebsiella pneumoniae. Nat Commun. 2021;12(1). [CrossRef]
- Schmitz FJ, Fluit AC, Lückefahr M, et al. The effect of reserpine, an inhibitor of multidrug efflux pumps, on the in-vitro activities of ciprofloxacin, sparfloxacin and moxifloxacin against clinical isolates of Staphylococcus aureus. Journal of Antimicrobial Chemotherapy. 1998;42(6). [CrossRef]
- Costa SS, Viveiros M, Amaral L, Couto I. Multidrug Efflux Pumps in Staphylococcus aureus: an Update. Open Microbiol J. 2013;7(1). [CrossRef]
- Vargiu A V., Ruggerone P, Opperman TJ, Nguyen ST, Nikaido H. Molecular mechanism of MBX2319 inhibition of Escherichia coli AcrB multidrug efflux pump and comparison with other inhibitors. Antimicrob Agents Chemother. 2014;58(10). [CrossRef]
- White M, Lenzi K, Dutcher LS, et al. 124. Impact of Levofloxacin MIC on Outcomes with Levofloxacin Step-down Therapy in Enterobacteriaceae Bloodstream Infections. Open Forum Infect Dis. 2019;6(Supplement_2). [CrossRef]
- Lamers RP, Cavallari JF, Burrows LL. The Efflux Inhibitor Phenylalanine-Arginine Beta-Naphthylamide (PAβN) Permeabilizes the Outer Membrane of Gram-Negative Bacteria. PLoS One. 2013;8(3). [CrossRef]
- Karaiskos I, Galani I, Daikos GL, Giamarellou H. Breaking Through Resistance: A Comparative Review of New Beta-Lactamase Inhibitors (Avibactam, Vaborbactam, Relebactam) Against Multidrug-Resistant Superbugs. Antibiotics.Multidisciplinary Digital Publishing Institute (MDPI). 2025;14(5). [CrossRef]
- Bové M, Bao X, Sass A, Crabbé A, Coenye T. The quorum-sensing inhibitor furanone C-30 rapidly loses its tobramycin-potentiating activity against Pseudomonas aeruginosa biofilms during experimental evolution. Antimicrob Agents Chemother. 2021;65(7). [CrossRef]
- Proctor CR, McCarron PA, Ternan NG. Furanone quorum-sensing inhibitors with potential as novel therapeutics against Pseudomonas aeruginosa. J Med Microbiol. 2020;69(2). [CrossRef]
- Kroh HK, Jensen JL, Wellnitz S, et al. Mouse monoclonal antibodies against Clostridioides difficile toxins TcdA and TcdB target diverse epitopes for neutralization. Freitag NE, ed. Infect Immun. Published online August 22, 2025. [CrossRef]
- Anantharajah A, Buyck JM, Sundin C, Tulkens PM, Mingeot-Leclercq MP, Van Bambeke F. Salicylidene acylhydrazides and hydroxyquinolines act as inhibitors of type three secretion systems in pseudomonas aeruginosa by distinct mechanisms. Antimicrob Agents Chemother. 2017;61(6). [CrossRef]
- Dahlgren MK, Zetterström CE, Gylfe Å, Linusson A, Elofsson M. Statistical molecular design of a focused salicylidene acylhydrazide library and multivariate QSAR of inhibition of type III secretion in the Gram-negative bacterium Yersinia. Bioorg Med Chem. 2010;18(7). [CrossRef]
- Brackman G, Breyne K, De Rycke R, et al. The Quorum Sensing Inhibitor Hamamelitannin Increases Antibiotic Susceptibility of Staphylococcus aureus Biofilms by Affecting Peptidoglycan Biosynthesis and eDNA Release. Sci Rep. 2016;6. [CrossRef]
- Zhao H, Brooks SA, Eszterhas S, et al. Globally deimmunized lysostaphin evades human immune surveillance and enables highly efficacious repeat dosing. Sci Adv. 2020;6(36). [CrossRef]
- Zha J, Li J, Su Z, Akimbekov N, Wu X. Lysostaphin: Engineering and Potentiation toward Better Applications. J Agric Food Chem. 2022;70(37). [CrossRef]
- Royer-Bertrand B, Rivolta C. Whole genome sequencing as a means to assess pathogenic mutations in medical genetics and cancer. Cellular and Molecular Life Sciences. 2015;72(8). [CrossRef]
- Beal MA, Glenn TC, Somers CM. Whole genome sequencing for quantifying germline mutation frequency in humans and model species: Cautious optimism. Mutat Res Rev Mutat Res. 2012;750(2). [CrossRef]
- Hung DT, Shakhnovich EA, Pierson E, Mekalanos JJ. Small-molecule inhibitor of Vibrio cholerae virulence and intestinal colonization. Science (1979). 2005;310(5748). [CrossRef]
- Woodbrey AK, Onyango EO, Kovacikova G, Kull FJ, Gribble GW. A Modified ToxT Inhibitor Reduces Vibrio cholerae Virulence in Vivo. Biochemistry. 2018;57(38). [CrossRef]
- Micoli F, Bagnoli F, Rappuoli R, Serruto D. The role of vaccines in combatting antimicrobial resistance. Nat Rev Microbiol. 2021;19(5). [CrossRef]
- Lipsitch M, Siber GR. How can vaccines contribute to solving the antimicrobial resistance problem? mBio. 2016;7(3). [CrossRef]
- Jansen KU, Anderson AS. The role of vaccines in fighting antimicrobial resistance (AMR). Hum Vaccin Immunother. 2018;14(9). [CrossRef]
- Hasso-Agopsowicz M, Sparrow E, Cameron AM, et al. The role of vaccines in reducing antimicrobial resistance: A review of potential impact of vaccines on AMR and insights across 16 vaccines and pathogens. Vaccine. 2024;42(19):S1-S8. [CrossRef]
- Gilsdorf JR. Hib Vaccines: Their Impact on Haemophilus influenzae Type b Disease. Journal of Infectious Diseases. 2021;224. [CrossRef]
- Sempere J, Llamosí M, López Ruiz B, et al. Effect of pneumococcal conjugate vaccines and SARS-CoV-2 on antimicrobial resistance and the emergence of Streptococcus pneumoniae serotypes with reduced susceptibility in Spain, 2004–20: a national surveillance study. Lancet Microbe. 2022;3(10). [CrossRef]
- van Heuvel L, Paget J, Dückers M, Caini S. The impact of influenza and pneumococcal vaccination on antibiotic use: an updated systematic review and meta-analysis. Antimicrob Resist Infect Control. 2023;12(1). [CrossRef]
- Nampota-Nkomba N, Carey ME, Jamka LP, Fecteau N, Neuzil KM. Using Typhoid Conjugate Vaccines to Prevent Disease, Promote Health Equity, and Counter Drug-Resistant Typhoid Fever. Open Forum Infect Dis. 2023;10. [CrossRef]
- DiGiandomenico A, Keller AE, Gao C, et al. A multifunctional bispecific antibody protects against Pseudomonas aeruginosa. Sci Transl Med. 2014;6(262). [CrossRef]
- Gulati S, Beurskens FJ, de Kreuk BJ, et al. Complement alone drives efficacy of a chimeric antigonococcal monoclonal antibody. PLoS Biol. 2019;17(6). [CrossRef]
- Lima R, Del Fiol FS, Balcão VM. Prospects for the use of new technologies to combat multidrug-resistant bacteria. Front Pharmacol. 2019;10(JUN). [CrossRef]
- ault P, Leclerc T, Jennes S, et al. Efficacy and tolerability of a cocktail of bacteriophages to treat burn wounds infected by Pseudomonas aeruginosa (PhagoBurn): a randomised, controlled, double-blind phase 1/2 trial. Lancet Infect Dis. 2019;19(1). [CrossRef]
- Aslam B, Arshad MI, Aslam MA, et al. Bacteriophage Proteome: Insights and Potentials of an Alternate to Antibiotics. Infect Dis Ther. 2021;10(3). [CrossRef]
- Patel DR, Bhartiya SK, Kumar R, Shukla VK, Nath G. Use of Customized Bacteriophages in the Treatment of Chronic Nonhealing Wounds: A Prospective Study. International Journal of Lower Extremity Wounds. 2021;20(1). [CrossRef]
- Sabouri S, Sepehrizadeh Z, Amirpour-Rostami S, Skurnik M. A minireview on the in vitro and in vivo experiments with anti-Escherichia coli O157:H7 phages as potential biocontrol and phage therapy agents. Int J Food Microbiol. 2017;243. [CrossRef]
- Almeida GMF, Mäkelä K, Laanto E, Pulkkinen J, Vielma J, Sundberg LR. The fate of bacteriophages in recirculating aquaculture systems (RAS)—towards developing phage therapy for RAS. Antibiotics. 2019;8(4). [CrossRef]
- Fischetti VA. Development of phage lysins as novel therapeutics: A historical perspective. Viruses. 2018;10(6). [CrossRef]
- Merabishvili M, Pirnay JP, Verbeken G, et al. Quality-controlled small-scale production of a well-defined bacteriophage cocktail for use in human clinical trials. PLoS One. 2009;4(3). [CrossRef]
- Sahota JS, Smith CM, Radhakrishnan P, et al. Bacteriophage Delivery by Nebulization and Efficacy Against Phenotypically Diverse Pseudomonas aeruginosa from Cystic Fibrosis Patients. J Aerosol Med Pulm Drug Deliv. 2015;28(5). [CrossRef]
- Nale JY, Spencer J, Hargreaves KR, Trzepiński P, Douce GR, Clokie MRJ. Bacteriophage combinations significantly reduce Clostridium difficile growth in vitro and proliferation in vivo. Antimicrob Agents Chemother. 2016;60(2). [CrossRef]
- de Abreu RC, Fernandes H, da Costa Martins PA, Sahoo S, Emanueli C, Ferreira L. Native and bioengineered extracellular vesicles for cardiovascular therapeutics. Nat Rev Cardiol. 2020;17(11). [CrossRef]
- Chen K, Pachter L. Bioinformatics for whole-genome shotgun sequencing of microbial communities. PLoS Comput Biol. 2005;1(2). [CrossRef]
- De R. Metagenomics: Aid to combat antimicrobial resistance in diarrhea. Gut Pathog. 2019;11(1). [CrossRef]
- Liu S, Moon CD, Zheng N, Huws S, Zhao S, Wang J. Opportunities and challenges of using metagenomic data to bring uncultured microbes into cultivation. Microbiome. 2022;10(1). [CrossRef]
- Khedkar S, Smyshlyaev G, Letunic I, et al. Landscape of mobile genetic elements and their antibiotic resistance cargo in prokaryotic genomes. Nucleic Acids Res. 2022;50(6). [CrossRef]
- Mackenzie JS, Jeggo M. The one health approach-why is it so important? Trop Med Infect Dis. 2019;4(2). [CrossRef]
- Macesic N, Polubriaginof F, Tatonetti NP. Machine learning: Novel bioinformatics approaches for combating antimicrobial resistance. Curr Opin Infect Dis. 2017;30(6). [CrossRef]
- Rajpurkar P, Chen E, Banerjee O, Topol EJ. AI in health and medicine. Nat Med. 2022;28(1). [CrossRef]
- Blechman SE, Wright ES. Applications of Machine Learning on Electronic Health Record Data to Combat Antibiotic Resistance. Journal of Infectious Diseases.Oxford University Press. 2024;230(5):1073-1082. [CrossRef]
- Vamathevan J, Clark D, Czodrowski P, et al. Applications of machine learning in drug discovery and development. Nat Rev Drug Discov. 2019;18(6). [CrossRef]
- Wachino, Jun-ichi. "Horizontal Gene Transfer Systems for Spread of Antibiotic Resistance in Gram-Negative Bacteria." Microbiology and Immunology. 2025; 69.7: 367-376.
- Pfaller, Michael A. "Flavophospholipol use in animals: positive implications for antimicrobial resistance based on its microbiologic properties." Diagnostic microbiology and infectious disease. 2006; 56.2: 115-121.
Figure 1.
Concept of One Health Approach: The One Health framework's roles for clinical medicine, disease surveillance, human, animal and environment health are represented by the circle differently in networks. The failure of single One Health framework component could compromise the overall effectiveness of policies. Therefore, the integration of environment, animal, and human health is necessary to attain holistic well-being and avert global health hazards like pandemics, antibiotic resistance, and diseases linked to climate change.
Figure 1.
Concept of One Health Approach: The One Health framework's roles for clinical medicine, disease surveillance, human, animal and environment health are represented by the circle differently in networks. The failure of single One Health framework component could compromise the overall effectiveness of policies. Therefore, the integration of environment, animal, and human health is necessary to attain holistic well-being and avert global health hazards like pandemics, antibiotic resistance, and diseases linked to climate change.
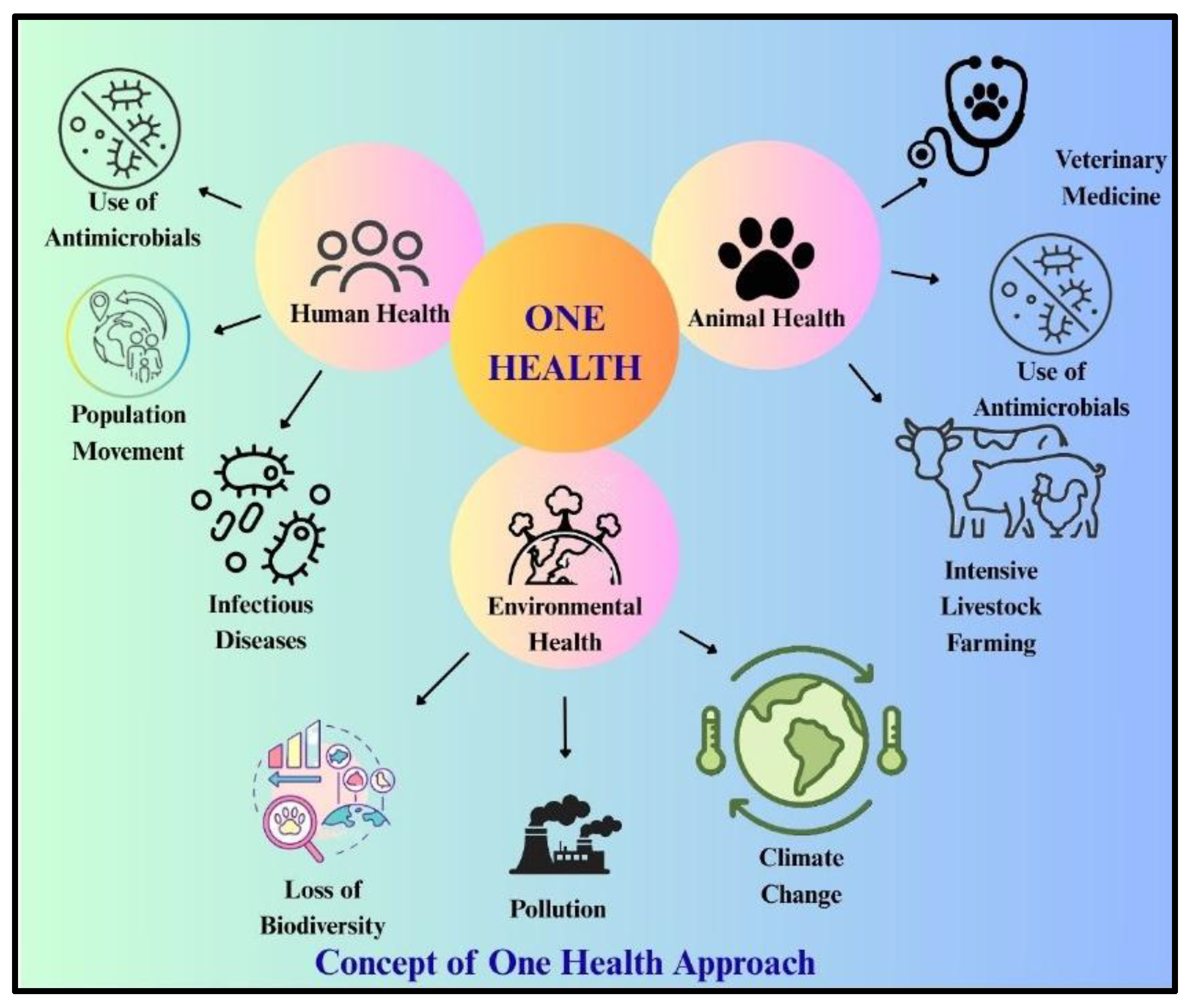
Figure 3.
The top ten most deadly pathogen-drug combos by AMR burden worldwide: A. The proportion of attributable deaths in 2021 for each pathogen-resistance combination are highlighted with the relative impact of each pairing globally. Staphylococcus aureus (Miticillin) showed dominant affects in compared to others. B. Deaths Attributable and Associated with AMR by Age Group (1990 vs. 2021). Neonatal (>1Year), Early childhood (≥ 9 years), Adolescents (≥19 years), Young Adults (≥29 years), Early Middle Age (≥39 years), Middle Age (≥49 years), Late Middle Age (≥ 59 years), Seniors / Elderly (≥ 69 years), Older Seniors (≥ 79 years), Very Old (≥ 89 years) and oldest age group (≤ 90 years) is correlated with antimicrobial resistance. Early childhood is the time with the highest rates of attributable and associated deaths. While the trends are partly reversed beyond the more senior age group, the accompanying death rate is lower in 2021 compared to 1990. In a similar vein, fewer deaths have been observed in 2021 than in 1990.
Figure 3.
The top ten most deadly pathogen-drug combos by AMR burden worldwide: A. The proportion of attributable deaths in 2021 for each pathogen-resistance combination are highlighted with the relative impact of each pairing globally. Staphylococcus aureus (Miticillin) showed dominant affects in compared to others. B. Deaths Attributable and Associated with AMR by Age Group (1990 vs. 2021). Neonatal (>1Year), Early childhood (≥ 9 years), Adolescents (≥19 years), Young Adults (≥29 years), Early Middle Age (≥39 years), Middle Age (≥49 years), Late Middle Age (≥ 59 years), Seniors / Elderly (≥ 69 years), Older Seniors (≥ 79 years), Very Old (≥ 89 years) and oldest age group (≤ 90 years) is correlated with antimicrobial resistance. Early childhood is the time with the highest rates of attributable and associated deaths. While the trends are partly reversed beyond the more senior age group, the accompanying death rate is lower in 2021 compared to 1990. In a similar vein, fewer deaths have been observed in 2021 than in 1990.
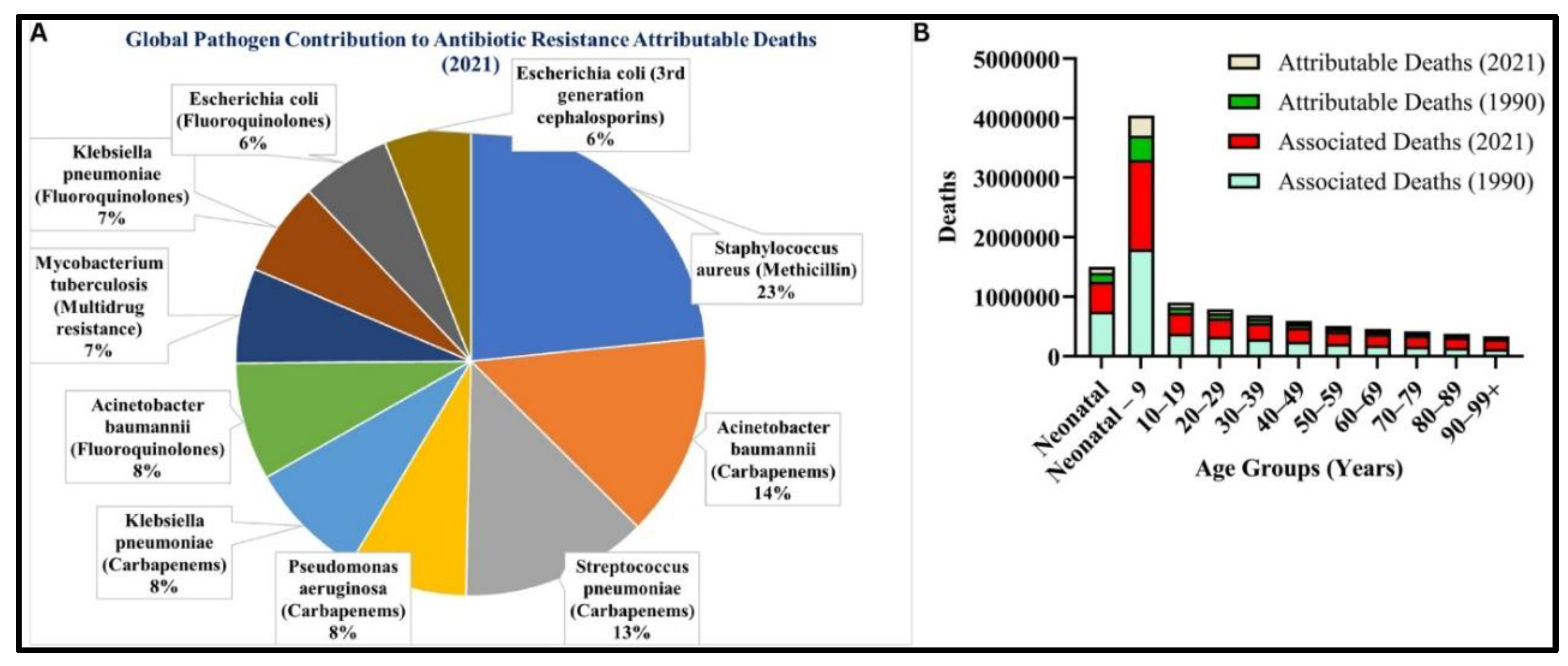
Figure 4.
Molecular mechanisms of bacteria resisting antibiotics: A. Illustration of antibiotic resistance, a single bacterial colony was isolated on a petri plate. The activation of pathogens' pathophysiological activity, which is controlled by the genetic makeup of bacteria, can result in the development of resistance traits in bacteria. Antibiotics (highlighted blue star) are broken down enzymatically by activating enzymes (light brown) insight the cells that initiate break down antibiotics (light green) before damage the bacterial cells. Activation of antibiotic modifying enzyme (dark brown) causes inactivation of antibiotics (light pink) a. antibiotics breakdown b. Antibiotic modifying enzymes chemically alter antibiotics to neutralize their effect c. Bacteria could modify the target molecules in their own cells that antibiotics normally attack, preventing antibiotics from binding and disrupting bacterial function d. The efflux pump is used by bacteria to expel antibiotics from their cells e. Reducing the permeability of cell walls prevent antibiotics from entering the cell. B. Bacteria acquire and evolve resistance: Chromosome mutations can be caused by duplication, insertion, deletion, and the acquisition of additional promoters. The horizontal gene transfer, or sexual reproduction, is one of the main causes of the development of resistance in bacteria. C. Antibiotics resistance Reservoirs: Mismanagement practices in the environment can lead to the development of resistance, and associated strains can be stored and function as reservoirs (like in a network), which is the main cause of resistance flow among the population and the failure of one health strategy. D. Basic Key Highlights for management of resistance in favour of one health policy.
Figure 4.
Molecular mechanisms of bacteria resisting antibiotics: A. Illustration of antibiotic resistance, a single bacterial colony was isolated on a petri plate. The activation of pathogens' pathophysiological activity, which is controlled by the genetic makeup of bacteria, can result in the development of resistance traits in bacteria. Antibiotics (highlighted blue star) are broken down enzymatically by activating enzymes (light brown) insight the cells that initiate break down antibiotics (light green) before damage the bacterial cells. Activation of antibiotic modifying enzyme (dark brown) causes inactivation of antibiotics (light pink) a. antibiotics breakdown b. Antibiotic modifying enzymes chemically alter antibiotics to neutralize their effect c. Bacteria could modify the target molecules in their own cells that antibiotics normally attack, preventing antibiotics from binding and disrupting bacterial function d. The efflux pump is used by bacteria to expel antibiotics from their cells e. Reducing the permeability of cell walls prevent antibiotics from entering the cell. B. Bacteria acquire and evolve resistance: Chromosome mutations can be caused by duplication, insertion, deletion, and the acquisition of additional promoters. The horizontal gene transfer, or sexual reproduction, is one of the main causes of the development of resistance in bacteria. C. Antibiotics resistance Reservoirs: Mismanagement practices in the environment can lead to the development of resistance, and associated strains can be stored and function as reservoirs (like in a network), which is the main cause of resistance flow among the population and the failure of one health strategy. D. Basic Key Highlights for management of resistance in favour of one health policy.
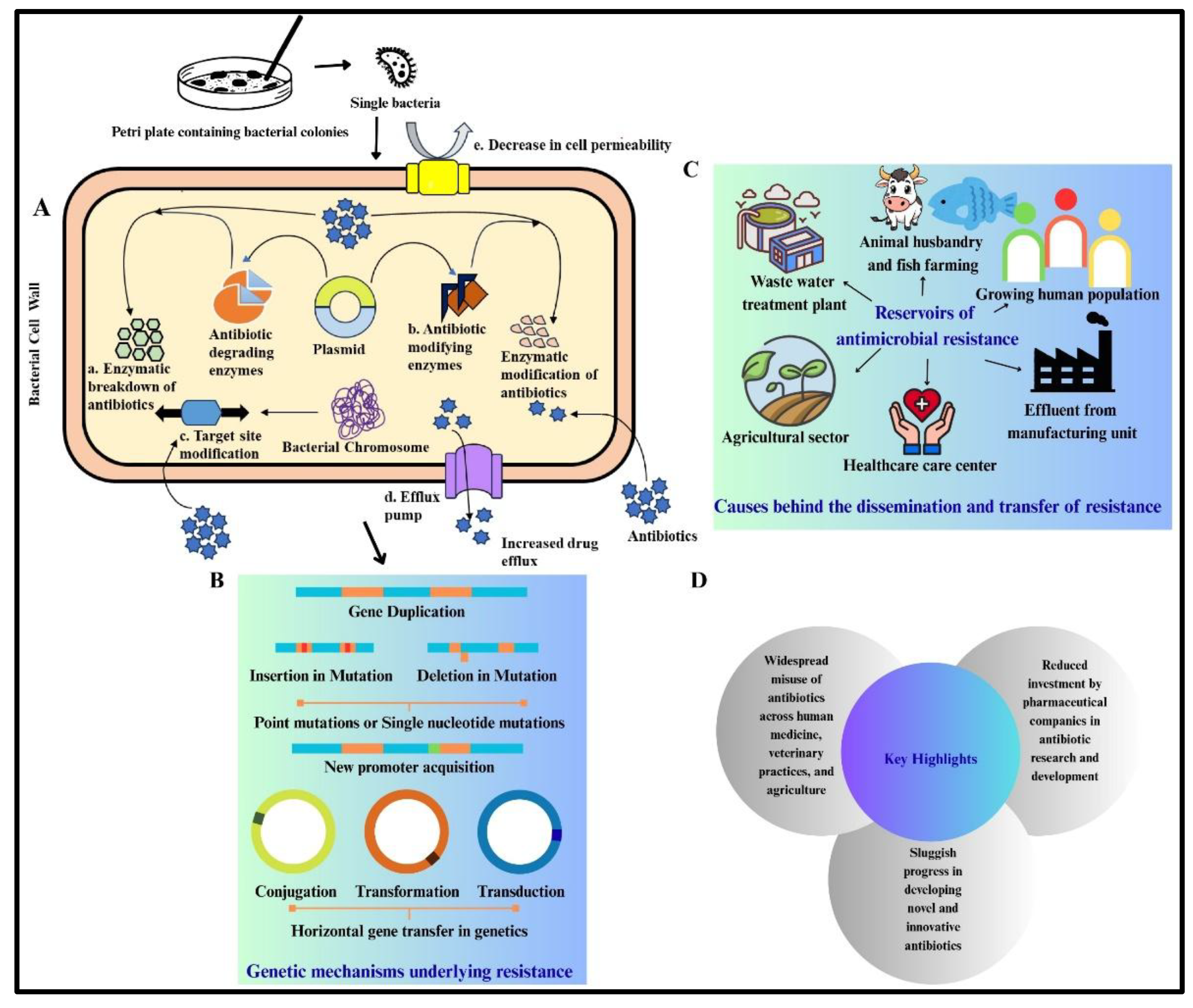
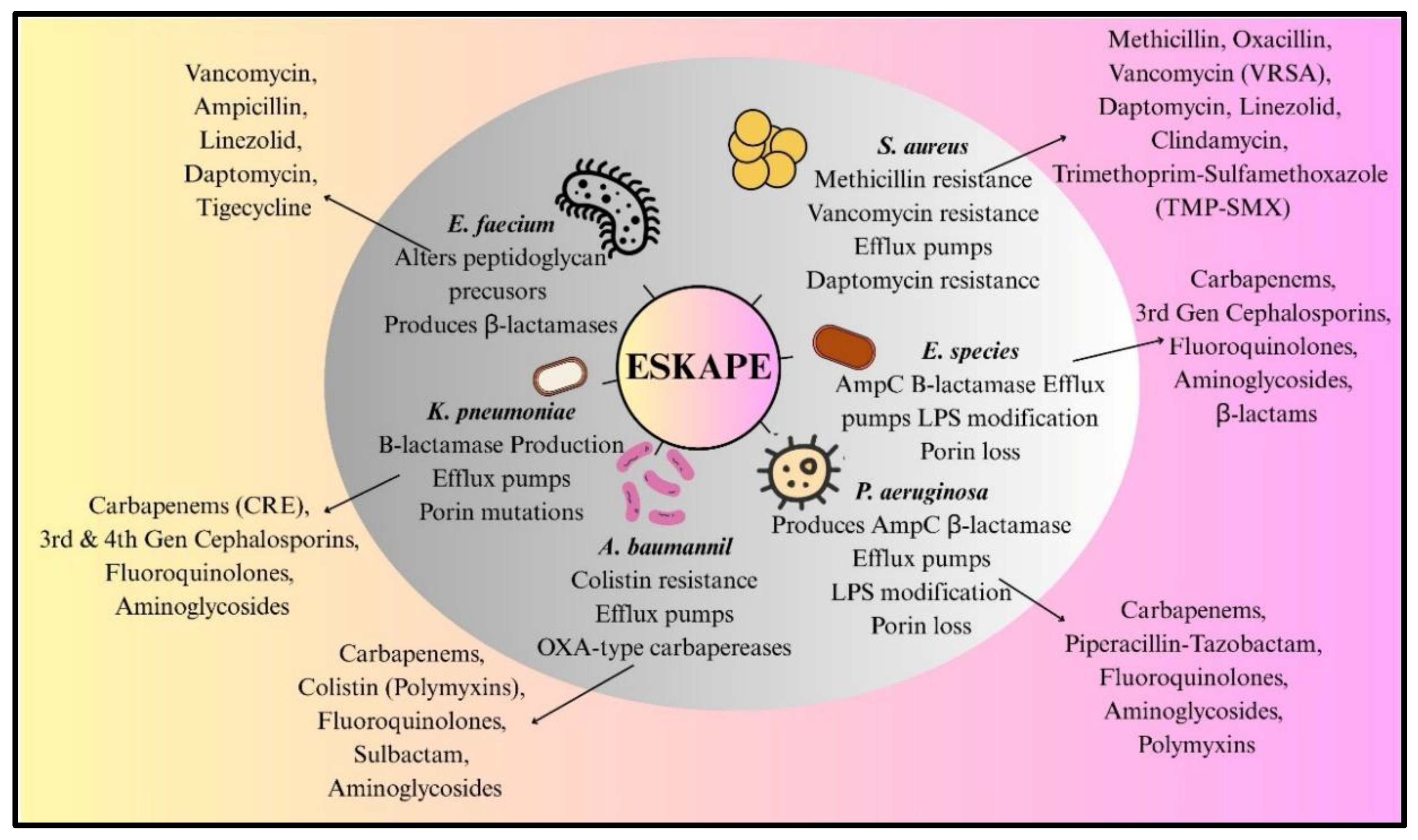
Figure 5.
ESKAPE pathogens response to antibiotics: ESKAPE pathogens (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp.) are highly resistant to antibiotics. They evade treatment through mechanisms like enzyme production, efflux pumps, target modification, reduced permeability, and biofilm formation, as illustrated above. The resistance of these species to antibiotics is also well documented in literature and illustrated above for specific bacterial species.
Figure 5.
ESKAPE pathogens response to antibiotics: ESKAPE pathogens (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp.) are highly resistant to antibiotics. They evade treatment through mechanisms like enzyme production, efflux pumps, target modification, reduced permeability, and biofilm formation, as illustrated above. The resistance of these species to antibiotics is also well documented in literature and illustrated above for specific bacterial species.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



