
Submitted:
06 October 2025
Posted:
10 October 2025
You are already at the latest version
Abstract
Quail farming requires balanced diets to ensure bird health and performance, and die-tary fiber and feed additives can influence immune and physiological responses. This study evaluated stimbiotic supplementation in diets with different fiber profiles on hematologi-cal and biochemical parameters of European quails (Coturnix coturnix coturnix). Six hundred quails, from 1 to 35 days of age, were assigned to 12 treatments with 5 replicates of 10 birds each. Diets included a corn-soy control, two basal diets with corn (low soluble fiber) or wheat (high soluble fiber), and three mixed diets combining corn and wheat in different proportions, each with or without 0.01% stimbiotic. Blood samples were collected at 14 and 35 days. In the early phase (1–14 days), stimbiotic in-creased hemoglobin and reduced total white blood cells, heterophils, and lymphocytes, indicating improved im-mune status, while fiber influenced liver enzyme activity and triglycerides. In the growth phase (15–35 days), stimbiotic maintained positive effects on hemoglobin and packed cell volume, and fiber increased eosinophils, modulating immune response without altering other parameters. In conclusion, fiber had a func-tional role, particularly during growth, and stimbiotic improved immunity in both phases, showing that both strategies enhance health and performance in European quails.
Keywords:
serum biochemistry
; hematology
; immunity
; metabolism
; nutrition
1. Introduction
Quail stand out as a poultry species of great economic importance due to the excellent quality of their meat, the high nutritional value of their eggs, and unique biological characteristics—such as early sexual maturity, high laying rate, and rapid growth—which make their farming a fast-return investment with low demand for space and feed [1]. The performance and health of these birds depend on balanced diets, especially during the initial (1–14 days) and growing (15–35 days) phases, when the gastrointestinal tract is still under development. In these phases, nutritional requirements differ from those of other poultry, with a higher demand for protein, lower for calcium, and higher for energy [2]. Such particularities, combined with genetic factors (breed and strain), sex, production purpose (meat or eggs), dietary energy density, nutrient bioavailability, and environmental and sanitary conditions, justify the need for specific nutritional recommendations for quail [3,4].
Corn and wheat are the main cereal grains used in poultry diets as energy sources due to their high starch content. However, these grains differ significantly in terms of the content and composition of non-starch polysaccharides (NSP). Wheat, for instance, presents a higher total NSP content, both in the soluble and insoluble fractions, compared to corn [3]. While the NSP profile of wheat is dominated by arabinoxylans, corn contains more cellulose and lower levels of soluble arabinoxylans [5].
Previously, dietary fiber was considered a diluting or antinutritional factor due to the presence of NSP [6,7]. However, in recent years, it has gained increasing attention due to its beneficial effects on gut health [8,9]. Since NSP are not digested by poultry enzymes, they are fermented by the gut microbiota, resulting in the production of short-chain fatty acids that favor the growth of beneficial bacteria [9,10,11,12,13,14,15].
In this context, stimbiotic emerges as a promising technological alternative. Composed of xylanase and xylo-oligosaccharides, it acts by signaling the microbiome to accelerate fiber fermentation, thereby promoting improvements in gut health and the immune system [16,17,18]. It is a dual-action additive—both energetic and prebiotic—capable of enhancing the fermentation of fibers already present in ingredients, although it is used at very low doses to directly impact the production of short-chain fatty acids.
The analysis of hematological and biochemical parameters is fundamental for assessing animal health. They serve as a mirror of the physiological state, providing essential information on organ function, the animal’s ability to adapt to nutritional and environmental challenges, and are crucial for identifying metabolic imbalances and various pathologies [19,20].
Recent studies have shown that stimbiotic supplementation exerts positive effects on gut health, productive performance, and the microbiota of broilers [9,14,15,21,22]. However, no studies have evaluated these effects in meat quail, nor have investigations explored their impacts on hematological and biochemical parameters, particularly during the initial and growing phases. This gap is critical, as supplementation and the definition of different fiber profiles directly influence the health, welfare, and physiological responses of the birds.
Therefore, the objective of this study was to evaluate the effect of different fiber profiles and stimbiotic supplementation on hematology and biochemical parameters in European quail, considering the 1–14 day and 15–35 day phases, and to identify possible metabolic responses and interactions between diet and additive.
2. Materials and Methods
2.1. Experimental Site and Ethics Committee
The experiment was conducted at the Poultry Experimental Module of the Center for Agricultural Sciences, at the Federal University of Paraíba – Campus II. A total of 600 one-day-old European quail (Coturnix coturnix coturnix) with an average weight of 10.17 ± 0.02 g were used. The experiment was submitted to the Animal Use Committee of the Federal University of Paraíba (CEUA-UFPB) under protocol No. 4639170425.
2.2. Housing
The treatments were arranged in a 6 × 2 factorial design, with 6 fiber profiles and 2 levels of stimbiotic. The birds were housed in galvanized wire cages measuring 100 × 33.3 × 20 cm (length × width × height), equipped with trough-type feeders and nipple drinkers, appropriate for the birds’ developmental stage. Feeders were supplied with the experimental diets twice daily, at 7:00 a.m. and 4:00 p.m. Throughout the experimental period, the birds had ad libitum access to feed and water.
2.3. Animals, Experimental Design and Experimental Diets
A total of 720 one-day-old European quail (Coturnix coturnix coturnix), with an average initial body weight of 10.17 ± 0.02 g, were used and distributed in a completely randomized design (CRD), with 12 treatments and 6 replicates of 10 birds each. The treatments were structured to allow comparison between different soluble fiber profiles in the diet. Initially, a control diet based on corn and soybean meal was used, representing the conventional standard employed in quail nutrition. Subsequently, two distinct basal diets were included: one composed exclusively of corn, characterized by a low soluble fiber content (L100), and another formulated solely with wheat, an ingredient recognized for its higher soluble fiber content (H100). To evaluate intermediate effects, three mixed diets were formulated by progressively combining wheat and corn in the proportions of L75:H25, L50:H50, and L25:H75, respectively. All diets were formulated to meet the recommendations of Silva and Costa [23] (Table 1 and Table 2). The chemical composition of the ingredients used in the formulation of the experimental diets was determined through Near Infrared Spectroscopy (NIRS). The six diets with different fiber profiles were administered with or without the addition of the stimbiotic (Signis, β-1,4-endo-xylanase, and xylo-oligosaccharides, AB Vista, Marlborough, UK) supplemented at 100g/ton of feed, totaling 12 dietary treatments (Figure 1A).
2.4. Data Collection
The experiment was conducted in two phases: initial (1 to 14 days) and growth (15 to 35 days). Hematological and biochemical analyses were evaluated in both phases. On days 14 and 35 of life, eight birds from each treatment were selected for blood collection, chosen based on the average body weight of the respective treatments. Prior to collection, the birds were fasted for 6 hours, and blood samples were obtained via jugular puncture using a 13 × 0.4 mm needle in syringes containing EDTA. The samples were individually stored in test tubes for hematological analyses. For leukocyte counting and differential analysis, blood smears were prepared immediately after collection from each bird (Figure 1B).
2.5. Collection of Biological Material and Laboratory Analyses
For the evaluation of biochemical parameters, blood samples were collected in plain tubes, which, after resting for 30 minutes, were centrifuged at 3,500 rpm for 1 minute to obtain the serum. The serum was then stored in Eppendorf tubes and frozen for subsequent analysis.
Biochemical analyses, as well as the staining and examination of blood smears, were performed at the Poultry Laboratory of the Federal University of Paraíba. For biochemical determinations, commercial kits (Labtest Diagnóstica S.A.®) were used to measure AST, ALT, GGT, proteins, triglycerides, and cholesterol, applied on the SINNOWA automatic biochemical analyzer (SX-260). Blood smears were stained using two routine stains, Dift-Quick and May-Grünwald Giemsa, which allowed differential leukocyte counting and the analysis of possible morphological alterations.
Immediately after collection, blood smears were prepared, air-dried, and subsequently stained with May-Grünwald-Giemsa. Packed cell volume (PCV) was determined using the microhematocrit method [24], and hemoglobin concentration was measured according to Weiss and Wardrop [25]. Hematimetric indices following Wintrobe’s method, including mean corpuscular volume (MCV) and mean corpuscular hemoglobin concentration (MCHC), were calculated using mathematical equations [26]. Total erythrocyte, leukocyte, and thrombocyte counts were performed manually in a Neubauer chamber with blood diluted in 0.01% toluidine blue at a ratio of 1:200 [27]. Differential leukocyte counts, morphological evaluation of blood cells, and hemoparasite screening were performed through examination of the blood smear under a light microscope at 1000× magnification. The heterophil-to-lymphocyte ratio (H:L) was determined according to Onbasilar and Aksoy [28]. All analyses were carried out at the Veterinary Hospital of the Center for Agricultural Sciences, Federal University of Paraíba.
2.6. Statistical Analysis
The data were subjected to Analysis of Variance (ANOVA) using R software version 4.2.0 [29] to determi-ne the effects of different fiber profiles (6 levels) and stimbiotic levels (2 levels), as well as their interactions, on the measured variables. For variables showing significant differences (P < 0.05), means were compared using Tukey’s test.
The variables were analyzed according to the following mathematical model:
Yijk=μ+αi+βj+(αβ)ij+ϵijk
Where:
- Yijk = response variable
- μ = overall mean
- αi = effect of the i-th fiber profile (i = 1 to 6)
- βj = effect of the j-th stimbiotic level (j = 1 to 2)
- (αβ)ij = interaction effect between the i-th fiber profile and the j-th stimbiotic level
- ϵijk = random error term associated with each observation, assumed to be normally distributed with mean zero and constant variance
3. Results
3.1. Hematological Parameters
Regarding the hematological parameters, it can be observed in Table 3 that during the initial phase (from the 1st to the 14th day of life), no significant interaction between fiber and stimbiotic was found for any of the variables analyzed. The parameters packed cell volume, erythrocyte count, mean corpuscular volume (MCV), mean corpuscular hemoglobin concentration (MCHC), and in the white blood cell series (Figure 2): eosinophils, basophils, monocytes, and heterophil/lymphocyte ratio showed no statistically significant differences as a function of fiber or stimbiotic. On the other hand, hemoglobin, total leukocyte count, heterophils, and lymphocytes were affected by the levels of stimbiotic inclusion in the diet. Hemoglobin was higher in animals that received stimbiotic, while total leukocyte count, heterophils, and lymphocytes were lower.
Platelet parameters, in turn, showed significant differences due to fiber inclusion. It was observed that the higher the fiber content, the lower the platelet counts, whereas in diets with lower fiber content, platelet counts were higher.
Table 4 presents the hematological values for the growth phase (15 and 35 days of age), and it can be observed that there was also no significant interaction between fiber and stimbiotic for any of the hematological variables. However, stimbiotic supplementation significantly increased packed cell volume and hemoglobin, while decreasing total leukocyte count and lymphocytes. Regarding fiber, only eosinophils showed a significant variation: birds on the control diet had the lowest eosinophil counts, whereas those on the diet with the highest fiber content (H100) exhibited the highest values.
3.2. Biochemical Parameters
In the biochemical analysis of the initial phase (Table 5), a significant interaction between stimbiotic and fiber was observed for alanine aminotransferase (ALT), as detailed in Figure 3. Evaluation of ALT levels showed significant differences among treatments. Birds in the control group receiving the stimbiotic exhibited the highest ALT concentration, 19.75 IU/L, which was statistically higher than most other treatments. In fiber profiles with stimbiotic, ALT values ranged from 11.38 IU/L (L75H25) to 16.75 IU/L (H100), with no significant differences among them. Treatments without stimbiotic showed lower and more homogeneous ALT levels, ranging from 9.50 IU/L (control) to 13.50 IU/L (L25H75), with no statistically significant differences between fiber profiles, except for the L25H75 treatment, which presented an intermediate value.
The variables albumin, aspartate aminotransferase (AST), cholesterol, gamma-glutamyl transferase (GGT), and total proteins did not show significant differences. However, triglycerides were significantly affected by both stimbiotic supplementation and fiber content. Triglyceride levels were higher with stimbiotic inclusion, and regarding fiber, the higher the fiber content, the lower the triglyceride values. In the growth phase, none of the evaluated biochemical variables showed statistical significance (Table 6).
4. Discussion
Packed cell volume (PCV), also known as hematocrit, represents the fraction of blood volume occupied by erythrocytes and is one of the main hematological indicators used to assess tissue oxygenation and hydration status in birds. In the growth phase (15 to 35 days) of the present experiment, this variable showed a statistically significant increase in the groups supplemented with the stimbiotic. This response may be related to greater efficiency in the absorption of nutrients essential for erythropoiesis, promoted by the intestinal modulation induced by the additive. According to Wang et al. [30] the use of stimbiotics in broiler diets significantly improves intestinal morphology, promoting increased villus height, higher villus-to-crypt ratio, and stimulation of genes related to epithelial barrier integrity. Such changes enhance the absorption of minerals like iron, phosphorus, and magnesium, whose bioavailability is crucial for proper hematopoietic function. Furthermore, as highlighted by Martinez et al. [31], stimbiotics contribute to creating a more stable and functional intestinal environment, optimizing nutrient digestibility. Therefore, the observed increase in PCV may reflect a positive physiological response to supplementation, demonstrating the indirect hematological potential of these additives through improved nutritional status.
Regarding hemoglobin, a significant increase was observed in both experimental phases in the groups supplemented with stimbiotic, suggesting a possible relationship with increased iron availability, favored by enhanced nutrient absorption provided by this functional additive. Hemoglobin is a hematological parameter directly influenced by iron bioavailability, which is essential for erythrocyte formation. According to Thrall et al. [32], physiological hemoglobin values in quail range from 4.0 to 5.2 g/L, and the data obtained in the present study were above this range, that is, higher than the normal reference values.
Among the variables that remained unchanged in both phases were hematimetric indices, mean corpuscular volume (MCV), mean corpuscular hemoglobin concentration (MCHC), eosinophils, basophils, monocytes, platelet counts, and the heterophil-to-lymphocyte ratio (H/L). In the initial phase, the quail were undergoing hematopoietic and immunological development, which may have limited the manifestation of perceptible physiological effects from stimbiotic supplementation. Furthermore, Garber and Parra [33] report that stimbiotics act indirectly in modulating systemic immunity, mainly by promoting an efferent intestinal environment where there is reduced activation of the immune axis in birds raised under good sanitary conditions. Thus, the absence of significant changes in some hematological variables, as observed in the present study, may reflect not the inefficacy of the stimbiotic, but rather the maintenance of a physiological state of immunological balance, without the need for substantial hematopoietic recruitment.
Hematimetric indices (MCV and MCHC), which assess erythrocyte morphological characteristics and mean hemoglobin content, did not show significant variations in either phase. This pattern indicates that stimbiotic supplementation, under the conditions studied, did not interfere with erythropoiesis or oxygen transport, maintaining physiological levels of red blood cell mass.
Total platelet counts, a parameter related to hemostatic function, also remained unchanged with additive inclusion, both in the initial and growth phases. This suggests that stimbiotic supplementation did not interfere with primary coagulation mechanisms or with physiological platelet activation in European quail (Coturnix coturnix coturnix).
In Table 3, regarding the initial phase (from 1 to 14 days), it is observed that fibers caused a significant variation in the number of platelets, with higher fiber content in the diet resulting in lower platelet counts. Platelets function to maintain blood hemostasis by releasing thromboplastin and other factors [34]. When associated with a higher-fiber diet, a lower cell count is observed, possibly related to improved intestinal health, favored by nutrient absorption in an intestinal environment enriched with beneficial bacteria stimulated by the presence of fiber. Paul et al. [35] reported that platelets constitutively express transcripts for both pro- and anti-inflammatory cytokines, indicating that elevated platelet levels in low-fiber diets may be related to an inflammatory response triggered by the diet composition.
Regarding leukocyte counts, variations were observed in both phases, with a consistent pattern of increased indices in the groups not supplemented with the stimbiotic. This response may be associated with greater exposure to physiological and environmental stress, since in birds, changes in total leukocyte numbers are commonly related to management, transport, subclinical infections, and adverse environmental conditions [32]. Thus, the data suggest that animals supplemented with the stimbiotic experienced lower metabolic stress, possibly due to improved intestinal integrity and nutrient absorption.
Lymphocytes showed significant variation in both experimental phases, with a higher proportion of these cells observed in the groups that did not receive a diet supplemented with the stimbiotic. Lymphocyte counts in birds can be influenced by various factors, including physiological, pathological, nutritional, and environmental aspects, since these cells constitute the main leukocyte population in the peripheral blood of birds [36] and play an essential role in the adaptive immune response. When comparing the results obtained in the two experimental phases, it is observed that lymphocyte counts were lower in animals supplemented with the stimbiotic. This reduction may be associated with a better immunological status and a more effective response to pathological challenges, highlighting the potential modulatory effect of the stimbiotic on the immune system.
Heterophile counts in birds can vary due to multiple physiological, pathological, and environmental factors and are considered an important indicator of the immunological and health status of birds. Significant changes were observed, with animals receiving the stimbiotic during the initial phase showing lower values than those that did not. As noted by Markowiak and Śliżewska [37], the gastrointestinal tract plays a significant immunological role and constitutes the most important barrier protecting the host against toxins, pathogens, and their effects, particularly inflammation. The presence of a beneficial microbiota promoted by the stimbiotic may reduce pathogen colonization, decrease the need for exaggerated inflammatory responses, and consequently stabilize or lower heterophile counts under healthy conditions.
Regarding leukocytes, the mean values of eosinophils, basophils, and monocytes also did not differ between the experimental groups in the two phases analyzed. The H/L ratio, traditionally used as a biomarker of stress in birds [38], also showed no significant difference between treatments. Considering that higher values of this ratio indicate a greater level of stress in birds, the results suggest that the stimbiotic, at the administered dosages and period, did not exert a significant modulatory effect on the stress response, at least not enough to alter this ratio between treatments.
When evaluating biochemical parameters, it is important to consider that they represent essential indicators of animal physiology, providing information about organ function, the capacity to adapt to nutritional, physiological, and environmental challenges, and helping to identify metabolic imbalances and pathologies [20]. According to [32], studies in birds focused on assessing the hepatic system are still relatively limited, with a scarcity of data regarding both reference values for these parameters and the sensitivity and specificity of the enzymes involved.
When analyzing Table 5, regarding the initial phase, it is observed that the only variable with a significant change was triglycerides. Their concentrations can vary according to sex, diet, and hormonal factors [20], considering the susceptibility of birds to elevated cortisol, especially when exposed to stress caused by captive rearing systems and grouping. Rezende et al. [39] reported triglyceride levels in broiler chickens between 128.9 and 140.2 mg/dL, while Evans et al. [40] and Silva et al. [41] reported values ranging from 136 to 166 mg/dL. The values obtained in this study were lower than these parameters, possibly due to differences in species and age.
In the initial phase (1–14 days), the inclusion of different fiber profiles in the diet significantly influenced serum triglyceride concentrations, while the stimbiotic showed a significant effect only on this variable. High-fiber diets, particularly those based on whole wheat (100H) or predominantly wheat (75H:25L, 50H:50L), resulted in lower triglyceride values compared to the control diet and the 100% corn formulation (100L). This reduction can be attributed to the higher proportion of soluble fiber present in wheat, which increases chyme viscosity, delays gastric emptying, and reduces lipid absorption, as well as possibly interfering with micellization and fatty acid absorption in the small intestine [42,43].
The hypolipidemic effect of soluble fiber may also be related to increased sequestration and excretion of bile acids, forcing the liver to use cholesterol for the synthesis of new bile salts, thereby indirectly modulating plasma lipid metabolism [42].
The use of the stimbiotic resulted in higher mean serum triglyceride values compared to diets without the additive, regardless of the fiber profile. This increase may be associated with the modulatory action of the stimbiotic on the intestinal microbiota, promoting the fermentation of undigested carbohydrates and increasing the production of short-chain fatty acids, particularly acetate, which acts as a precursor for hepatic lipogenesis in birds [44].
Although no significant interaction was observed for triglycerides, the main effects indicate that both fiber composition and the presence of the stimbiotic independently influenced the modulation of this variable [45]. The other biochemical parameters (albumin, AST, GGT, total protein, and cholesterol) did not show statistically relevant differences, except for ALT, which exhibited a significant interaction, suggesting the sensitivity of this enzyme to the specific combination of fiber profile and additive.
These findings reinforce the modulatory role of dietary fiber—particularly wheat—on lipid metabolism in young quails, possibly associated with the immaturity of the gastrointestinal tract at this stage, making the animals more responsive to changes in diet composition [43].
A direct relationship was also observed between a high-fiber diet and increased eosinophil counts in birds during the initial phase. This finding contrasts with low-fiber diets, which showed lower cell counts. Research by Cīrule et al. [46] suggests that this variation is related to stress, as eosinophil numbers tend to decrease under stressful conditions. The underlying mechanism is that fibers, when broken down into xylo-oligosaccharides, promote microbial fermentation and the production of volatile fatty acids. As Parra et al. [16] describe, these acids optimize digestion and nutrient absorption. This improvement in the digestive process reduces stress in the animals, which in turn explains the higher eosinophil levels observed in high-fiber diets.
When evaluating the interaction in the ALT variable, it is essential to consider that dietary fiber plays a vital role in the metabolic and digestive health of birds. In the case of the European quails studied, fiber sources such as wheat and corn have characteristics that directly influence ALT levels. Interpreting the increase in serum ALT concentration is challenging, as it may result from damage to various tissues [47] or from physiological variations, such as aging, which can elevate its values [32].
Wheat fiber, predominantly composed of soluble and insoluble fractions, has a higher fermentation capacity in the intestine, which can result in the production of volatile fatty acids beneficial to the intestinal mucosa and overall metabolism. According to Faria et al. [20], birds with severe liver damage may present normal or even reduced ALT values, indicating that the activity of this enzyme in the hepatic tissue of some species is diminished. Most bird species exhibit serum ALT values ranging from 19 to 50 IU/L [48,49].
Previous studies have shown that supplementation with stimbiotics improves productive performance, feed efficiency, intestinal microbiota composition, and gut health in broiler chickens [9,14,15,21,22]. Although the present study did not directly evaluate these parameters, the hematological and biochemical results indicate an improvement in the immunological status of quails, evidenced by reductions in leukocytes, lymphocytes, and heterophils, as well as increases in packed cell volume and hemoglobin. These findings suggest that the stimbiotic likely promoted effects similar to those reported in previous studies, favoring a healthier intestinal environment, optimizing nutrient absorption, and enhancing physiological and immunological balance in the birds (Figure 4).
Thus, the findings of the present study demonstrate that supplementation with a stimbiotic and variations in fiber profiles independently modulated hematological and biochemical parameters in broiler quails. Stimbiotic inclusion promoted increases in packed cell volume and hemoglobin, as well as reductions in leukocytes, lymphocytes, and heterophils, indicating lower physiological stress and improved immunological balance. On the other hand, different fiber profiles independently influenced certain variables, such as platelets and triglycerides, highlighting their role in intestinal and lipid metabolism. Although productive performance and gut health were not directly evaluated, the results suggest that the stimbiotic likely produced effects similar to those reported in broiler chickens, favoring nutrient absorption, physiological balance, and maintenance of a healthy immunological status in the birds.
5. Conclusions
The use of the stimbiotic is recommended in both the initial and growth phases, showing a positive impact on the birds' immune system. High-fiber diets, on the other hand, provide greater benefits during the growth phase.
Author Contributions
Conceptualization. L.M.C., A.V.d.L., A.A.F.d.S., R.d.S.S., J.K.G.C., X.R., M.R.d.L., F.G.P.C., R.R.G.; Methodology. L.M.C., A.V.d.L., E.G.d.S., M.G.d.S.B., R.d.S.S., J.K.G.C., X.R., M.R.d.L., F.G.P.C., R.R.G.; Software, L.M.C., A.V.d.L., E.G.d.S., N.L.R., R.d.S.S., M.G.d.S.B., F.G.P.C., R.R.G.; Formal analysis, L.M.C., A.A.F.d.S., M.G.d.S.B., F.G.P.C., N.L.R., R.R.G.; Writing—review and editing, L.M.C., A.V.d.L., E.G.d.S., R.d.S.S., M.G.d.S.B., N.L.R., M.R.d.L., F.G.P.C., R.R.G. All authors have read and agreed to the published version of the manuscript.
Funding
The Funding for this research was performed by National Council for Scientific and Technological Development (CNPq) Announcement nº 09/2023—Research Productivity Grants, project no. 313033/2023-4.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Ethics Committee of Federal University of Paraiba (protocol code 4639170425).
Informed Consent Statement
Not applicable.
Data Availability Statement
Dataset available on request from the authors.
Acknowledgments
The authors would like to thank the Center for Agricultural Sciences (CCA) of the Federal University of Paraíba (UFPB), the National Council for Scientific and Technological Development (CNPq), and the Coordination for the Improvement of Higher Education Personnel (CAPES). Special thanks are given to AB Vista for providing research materials used for the experiments.
Conflicts of Interest
Xavière Rousseau is an employee of AB Vista, the company that supplied the phytase (Quantum Blue) and stimbiotic (Signis) used in this study. This co-author was not involved in the preparation of the manuscript in any way that could bias the presentation of the company’s products. All authors declare that the research was conducted independently and that no commercial or financial relationships existed that could be construed as a potential conflict of interest.
References
- Cullere, M.; Woods, M.J.; Van Emmenes, L.; Pieterse, E.; Homman, L.C.; Zotte, A.D. Hermetia illucens Larvae Reared on Diferent Substrates in Broiler Quail Diets: E_ect on Physicochemical and Sensory Quality of the Quail Meat. Animals. [CrossRef]
- Silva, J.H.V.; Jordão Filho, j.; Costa, F.G.P. ; Lacerda. P.B; Vargas, D.G.V; Lima, M.R. Exigências nutricionais de codornas. Revista Brasileira de Saúde e Produção Animal. [CrossRef]
- Rostagno, H.S; Albino, L.F.T; Calderano, A.A.; Hannas, M.I.; Sakomura, N.K.; Costa, F.G.P; Rocha, G.C.; Saraiva, A.; Abreu, M. L. T. Tabelas Brasileiras para Aves e Suínos: composição de alimentos e exigências nutricionais. 5th edn. (UFV: Viçosa. Brazil). 2024.
- Costa, F.G.P; Lima, A.V; Souza, P.E.L; Rodrigues, A.B; Nascimento, C.H; Cavalcante, D.T; Lima, M.R; Kaneko, IN. Eficácia de diferentes fontes de metionina em dietas de codornas europeias. Observatorio de la Economía Latinoamericana. [CrossRef]
- Annison, G. Relationship between the levels of soluble nonstarch polysaccharides and the apparent metabolizable energy of wheats assayed in broiler chickens. J. Agric. Food Chem 1991, 1252–1256. [Google Scholar] [CrossRef]
- Jha, R. , Fouhse, J.M., Tiwari, U.P., Li, L., Willing, B.P. Dietary fiber and intestinal health of monogastric animals. Front. Vet. Sci 2019. 6. 48. [CrossRef]
- Mateos, G.; Fondevila, G.; C´amara, L. The importance of the fibre fraction of the feed in non-ruminant diets. In The Value of Fibre: Engaging the Second Brain for Animal Nutrition, 1st ed.; González-Ortiz, G., Bedford, M.R., Bach Knudsen, K.E., Courtin, C.M., Classen, H.L., Eds.; Wageningen Academic Publishers: Wageningen, Netherlands, 2019; pp. 61–83. [Google Scholar] [CrossRef]
- Jha, R. , Berrocoso, J. Dietary fiber utilization and its effects on physiological functions and gut health of swine. Animal 2015, 1441–1452. [Google Scholar] [CrossRef] [PubMed]
- kasireddy, B.; Lourenco, J.; Gonzalez-ortiz, G.; Bedford, M.R.; Olukosi, O.A. Growth performance, nutrient utilization, gut integrity, short-chain fatty acids, and gene expression in Eimeria-challenged broilers receiving stimbiotics and wheat bran as an additional fiber source. Poultry Science 2025, 4877. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.B. Challenges with nonfiber carbohydrate methods. Journal of Animal Science 2003, 3226–3232. [Google Scholar] [CrossRef] [PubMed]
- Bedford, M. , Cowieson, A.J. Exogenous enzymes and their effects on intestinal microbiology. Anim. Feed Sci Technol. [CrossRef]
- Macari, M.; Maiorka, A. Fisiologia das aves comerciais. (PROL Editora Gráfica: Jaboticabal. Brazil). 2017. 806p.
- Nguyen, H. , Bedford, M., Wu, S-B., Morgan, N.K. Soluble non-starch polysaccharide modulates broiler gastrointestinal tract environment. Poult. Sci, 2021; 100, 101183. [Google Scholar] [CrossRef]
- Jonchère, V.; Bussière, F.I.; Zemb, O.; Khaksar, V.; Cornaille, L.; Gambier, E.; Derouin-Tochon, F.; Hervo, F.; Schouler, C.; Rousseau, X.; Bedford, M.; Mignon, S.V.; Duclos, M.J.; Guabiraba, R.; Narcy, A. Differentiated short- and long-term impacts of a starter stimbiotic supplementation on gut health in broilers fed wheat and rye-based diets at homeostasis and under Eimeria tenella challenge. Animal Nutrition. [CrossRef]
- Toghyani, M.; Kim, E.; Macelline, S.P.; González-Ortiz, G.; Barekatain, R.; Liu, S.Y. Xylanase and stimbiotic supplementation improve broilers performance and nutrient digestibility across both wheat-barley and corn-based diets. Poultry Science 2025, 104, 105224. [Google Scholar] [CrossRef] [PubMed]
- Parra, D.; Greenwood, W.; Santos, T.T. A new perspective on maximising the potential of dietary fibre, xylanase and gut health. ABVista 2020, 26-27, 2020. [Google Scholar]
- Davies, C.; Gonzalez-Ortiz, G.; Rinttila, T.; Apajalahti, J.; Alyassin, M.; Bedford, M.R. Stimbiotic supplementation and xylose-rich carbohydrates modulate broiler's capacity to ferment fibre. Front Microbiol. [CrossRef]
- Veluri, S.; Gonzalez-Ortiz, G.; Bedford, M.R.; Olukosi, O.A. Interactive effects of a stimbiotic supplementation and wheat bran inclusion in corn- or wheat-based diets on growth performance, ileal digestibility, and expression of nutrient transporters of broilers chickens. Poult Sci. 2024, 103, 103178. [Google Scholar] [CrossRef] [PubMed]
- Rajman, M.; Juráni, M.; Lamosová, D.; Macajová, M.; Sedlacková, M.; Kostál, L.; Vyboh, P. The effects of feed restriction on plasma biochemistry in growing meat type chickens (Gallus gallus). Comparative Biochemistry and Physiology Part A: Molecular e Integrative Physiolog. [CrossRef]
- Faria, P.P.D.; Cruz, L.C.F.; Sampaio, S.A.; Borges, K.F.; Minafra, C.S. Análises bioquímicas para frango de corte – revisão. Revista Eletrônica NutriTime 2021, 18, 06. [Google Scholar]
- Lee, J.H.; Lee, B.; Rousseau, X.; Gomes, G.A.; Oh, H.J.; Kim, Y.J.; Chang, S.Y.; An, J.W.; Go, Y.B.; Song, D.C.; Cho, H.A.; Cho, J.H. Stimbiotic supplementation modulated intestinal inflammatory response and improved broilers performance in an experimentally induced necrotic enteritis infection model. Journal of Animal Science and Biotechnology. [CrossRef]
- Ren, Y.; Tian, Y.; Hou, M.; Zhao, Y.; Li, J.; Aftab, U.; Rousseau, X.; Jiang, R.; Kang, X.; Tian, Y.; Gong, Y. Evaluation of stimbiotic on growth performance and intestinal development of broilers fed corn- or wheat-based diets. Poultry Science 2023, 102, 103094. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.H.V.; Costa, F.G.P. Tabelas para codornas japonesas e europeias. 2th edn. (FUNEP: Jabuticabal. Brazil). 2009. 110p.
- Jain N., C. Essentials of Veterinary Hematology. 1st. (Wiley: Pennsylvania. EUA). 1993.
- Weiss D., J. , Wardrop K. Schalm's Veterinary Hematology. 6nd ed. (WileyBlackwell: Nova Jersey. EUA). 2010. 1206p.
- Kerr M., G. . Exames Laboratoriais em Medicina Veterinária: bioquímica clínica e hematologia. 2nd ed. (Roca: São Paulo. Brazil) 2003. 465p.
- Schalm O., W. , Jain N. C., Carroll E. J. Scham’s Veterinary Hematology. 4nd ed. (Lea & Febiger: Philadelphia. EUA). 1986. 1221p.
- Onbaşılar, E.E.; Aksoy, F.T. Stress parameters and immune response of layers under different cage floor and density conditions. Livest. prod. Sci. 2005, 95, 255–263. [Google Scholar] [CrossRef]
- R Core Team. R: a language and environment for statistical computing (ver. 4.2.0).” (R Foundation for Statistical Compu-ting: Vienna. Austria). 2022. https://www.R-project.org/.
- Wang, H.; et al. Stimbiotic improves intestinal morphology and tight junction integrity in broilers fed corn- or wheat-based diets. Animals. [CrossRef]
- Martinez, I.; Brito, A.B.; Costa, M.A.; Garcia, A. Fermentação de polissacarídeos não amiláceos e sua aplicação na nutrição de aves. aviNews Brasil 2024. [Google Scholar]
- Thrall, M.A. ; Weiser. G.; Allison, R.,; Campbell, T.W. Hematologia e Bioquímica Clínica Veterinária. 2. ed. (Roca: São Paulo. Brazil). 2014.
- Garber, A.; PARRA, D. Estimbióticos: um elemento essencial das estratégias de resiliência nutricional. aviNews Brasil 2021. [Google Scholar]
- Mitchell, E.B.; Johns, J. Avian hematology and Related Disorders. Vet Clin Exot Anim. [CrossRef]
- Paul, M.S.; Paolucci, S.; Barjesteh, N. ; Wood. R.D.; Schat, R.A.; Sharif, S. Characterization of Chicken Thrombocyte Responses to Toll-Like Receptor Ligands. Plos One. [CrossRef]
- Claver, J. A.; Quaglia, A. I. E. Comparative Morphology, Development, and Function of Blood Cells in Nonmammalian Vertebrates. Journal of Exotic Pet Medicine. [CrossRef]
- Markowiak, P.; Śliżewska, K. The role of probiotics, prebiotics and synbiotics in animal nutrition. Gut Pathogens. [CrossRef]
- Gross, W.B.; Siegel, H.S. Evaluation of the heterophil/lymphocyte ratio as a measure of stress in chickens. Avian Diseases 1983, 27, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Rezende, M.S.; Silva, P.L.; Guimarães, E.C.; Lellis, C.G.; Mundim, A.V. Variações fisiológicas, influência da idade e sexo no perfil bioquímico sanguíneo de aves da linhagem pesada de frango de corte na fase de recria. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, 2019. [Google Scholar] [CrossRef]
- Evans, A.J.; Bannister, D.W.; Whitehead, C.C.; Siller, W.G.; Wight, P.A.L. Changes in plasma lipid and glucose levels during the onset of fatty liver and kidney syndrome in chicks. Res. Vet. Sci. [CrossRef]
- Silva, P.R.L.; Freitas Neto, O.C.; Larurentiz, A.C.; Junqueira, O.M.; Fagliari, J.J. Blood serum components and serum protein test of Hybro-PG broilers of different ages. Braz. J. Poult. Sci, 2007. [Google Scholar] [CrossRef]
- Choct, M. Managing gut health through nutrition. British Poultry Science. [CrossRef]
- Svihus, B. Function of the digestive system. Journal of Applied Poultry Research. [CrossRef]
- Yue, Y.; Luasiri, P.; Li, J. , Laosam, P.; Sangsawad, P. Research advancements on the diversity and host interactions of the chicken gut microbiome. Frontiers in Veterinary Science. [CrossRef]
- Chesson, A. Non-starch polysaccharide-degrading enzymes in poultry diets: Influence of ingredients on the selection of activities. World's Poultry Science Journal. [CrossRef]
- Cirule, D.; Krama, T.; Vrublevska, J.; Rantala, M.J; Krams, I. A rapid effect of handling on counts of white blood cells in a wintering passerine bird: a more practical measure of stress? Journal of Ornithology. [CrossRef]
- Grunkemeyer, V.L. Advanced diagnostic approaches and current management of avian hepatic disorders. Veterinary Clinics: Exotic Animal Practice. [CrossRef]
- Lumeij, J.T. Avian clinical biochemistry. In Clinical biochemistry. In Clinical biochemistry of domestic animals, 5th ed.; Kaneko, J.J., Harvey, J.W., Bruss, M.L., Eds.; Academic Press: San Diego, USA, 1997; pp. 885–905. [Google Scholar] [CrossRef]
- Campbell, T. W. Veterinary hematology and clinical chemistry. 2. ed. (Wiley-Blackwell: Iowa, USA). 2012. 772p.
Figure 1.
Formation of the experimental treatments from three basal diets (control, high, and low fiber), with and without stimbiotic, resulting in 12 experimental treatments (A), and a schematic representation of blood collection and analyses (B).
Figure 1.
Formation of the experimental treatments from three basal diets (control, high, and low fiber), with and without stimbiotic, resulting in 12 experimental treatments (A), and a schematic representation of blood collection and analyses (B).
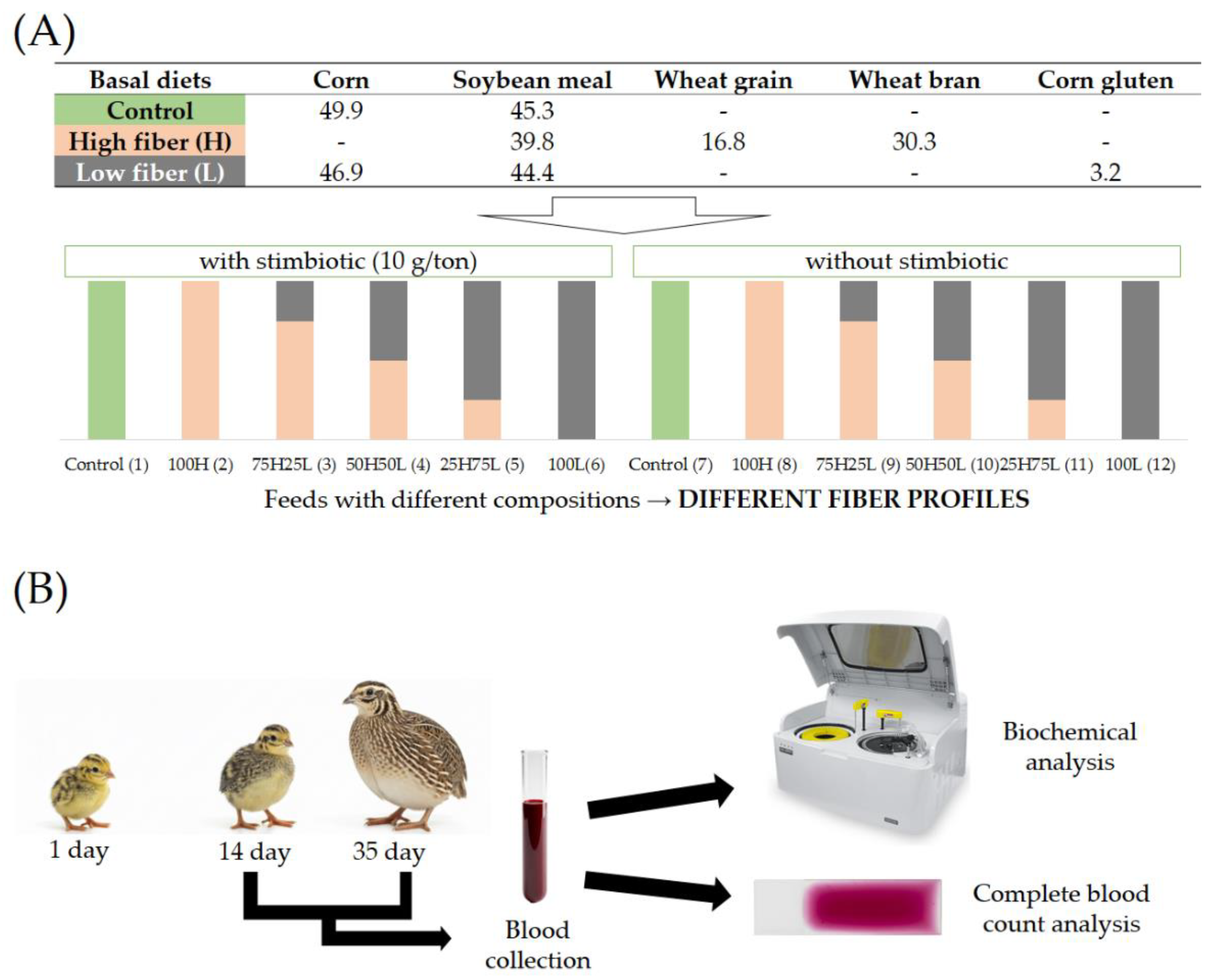
Figure 2.
Photomicrographs of blood cells from European quails (Coturnix coturnix coturnix). A – Erythrocytes (open arrow), heterophils (thick arrow), thrombocytes (short arrows), and lymphocytes (arrowhead); B – Erythrocytes (open arrow), lymphocytes (arrowhead), thrombocytes (short arrows), and monocytes (asterisk); C – Erythrocytes (open arrow), thrombocytes (short arrows), and lymphocytes (arrowhead); D – Erythrocytes (open arrows) and eosinophils (arrow); E – Eosinophils (arrow), heterophils (thick arrow), and lymphocyte (arrowhead); F – Basophils (thin arrow). Staining: May-Grünwald-Giemsa. Scale bars: A–F, 10 µm.
Figure 2.
Photomicrographs of blood cells from European quails (Coturnix coturnix coturnix). A – Erythrocytes (open arrow), heterophils (thick arrow), thrombocytes (short arrows), and lymphocytes (arrowhead); B – Erythrocytes (open arrow), lymphocytes (arrowhead), thrombocytes (short arrows), and monocytes (asterisk); C – Erythrocytes (open arrow), thrombocytes (short arrows), and lymphocytes (arrowhead); D – Erythrocytes (open arrows) and eosinophils (arrow); E – Eosinophils (arrow), heterophils (thick arrow), and lymphocyte (arrowhead); F – Basophils (thin arrow). Staining: May-Grünwald-Giemsa. Scale bars: A–F, 10 µm.
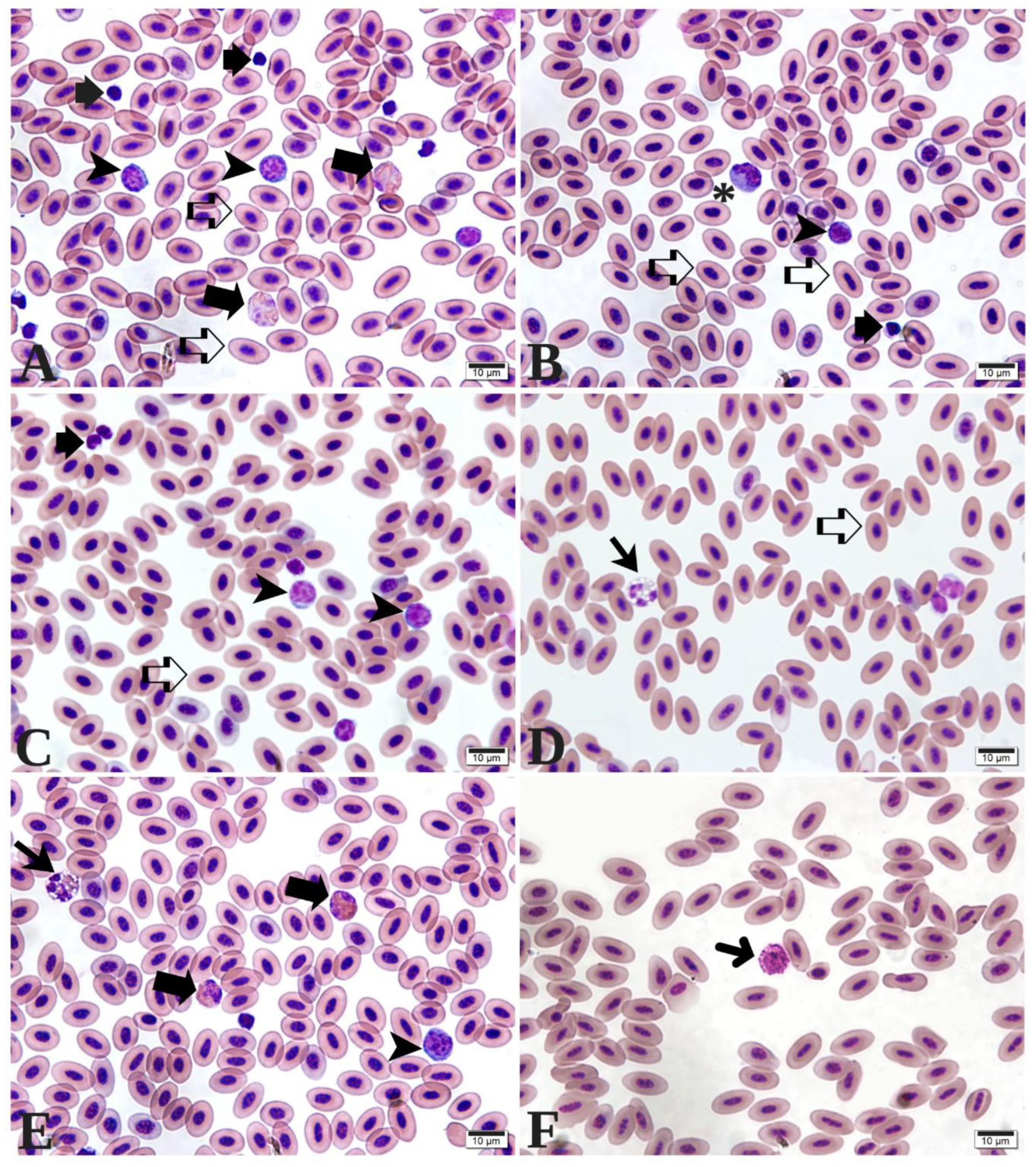
Figure 3.
Photomicrographs of blood cells from European quails (Coturnix coturnix coturnix). A – Erythrocytes (open arrow), heterophils (thick arrow), thrombocytes (short arrows), and lymphocytes (arrowhead); B – Erythrocytes (open arrow), lymphocytes (arrowhead), thrombocytes (short arrows), and monocytes (asterisk); C – Erythrocytes (open arrow), thrombocytes (short arrows), and lymphocytes (arrowhead); D – Erythrocytes (open arrows) and eosinophils (arrow); E – Eosinophils (arrow), heterophils (thick arrow), and lymphocyte (arrowhead); F – Basophils (thin arrow). Staining: May-Grünwald-Giemsa. Scale bars: A–F, 10 µm.
Figure 3.
Photomicrographs of blood cells from European quails (Coturnix coturnix coturnix). A – Erythrocytes (open arrow), heterophils (thick arrow), thrombocytes (short arrows), and lymphocytes (arrowhead); B – Erythrocytes (open arrow), lymphocytes (arrowhead), thrombocytes (short arrows), and monocytes (asterisk); C – Erythrocytes (open arrow), thrombocytes (short arrows), and lymphocytes (arrowhead); D – Erythrocytes (open arrows) and eosinophils (arrow); E – Eosinophils (arrow), heterophils (thick arrow), and lymphocyte (arrowhead); F – Basophils (thin arrow). Staining: May-Grünwald-Giemsa. Scale bars: A–F, 10 µm.
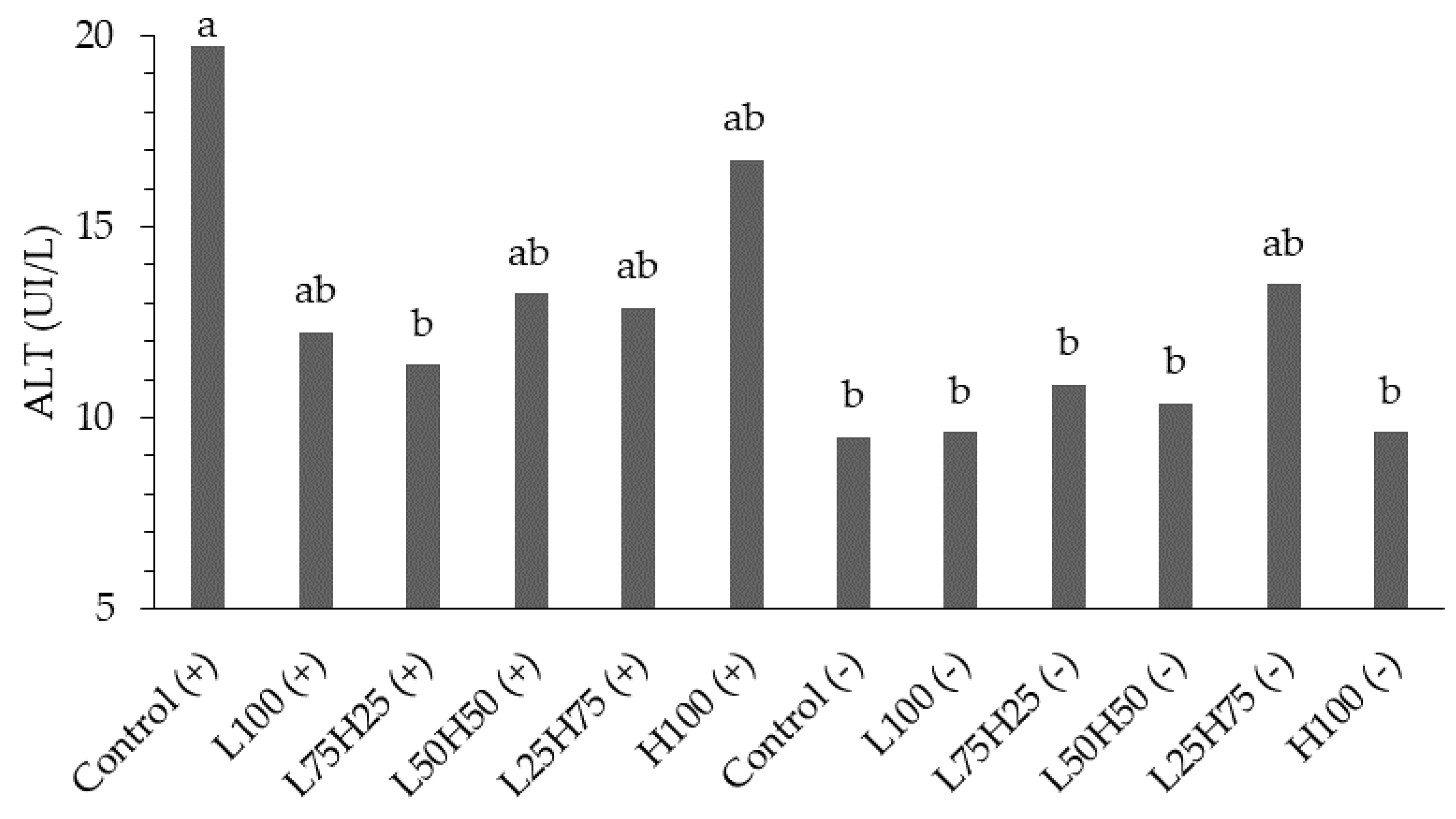
Figure 4.
Schemes of stimbiotic effects in quails, showing positive impacts on microbiota, gut health, immunity, and performance.
Figure 4.
Schemes of stimbiotic effects in quails, showing positive impacts on microbiota, gut health, immunity, and performance.

Table 1.
Composition and nutritional values of diets with different fiber profiles, with or without stimbiotic, for European quails in the starter phase (1–14 days).
Table 1.
Composition and nutritional values of diets with different fiber profiles, with or without stimbiotic, for European quails in the starter phase (1–14 days).
| Ingredients, % | With Stimbiotic | Without Stimbiotic | ||||||||||
| Control | 100H | 75H25L | 50H50L | 25H75L | 100L | Control | 100H | 75H25L | 50H50L | 25H75L | 100L | |
| Corn | 49.885 | 0.000 | 11.723 | 23.446 | 35.169 | 46.892 | 49.885 | 0.000 | 11.723 | 23.446 | 35.169 | 46.892 |
| Soybean meal | 45.331 | 39.758 | 40.913 | 42.068 | 43.223 | 44.378 | 45.331 | 39.758 | 40.913 | 42.068 | 43.223 | 44.378 |
| Whole wheat | 0.000 | 16.799 | 12.599 | 8.399 | 4.200 | 0.000 | 0.000 | 16.799 | 12.599 | 8.399 | 4.200 | 0.000 |
| Wheat bran | 0.000 | 30.337 | 22.753 | 15.169 | 7.584 | 0.000 | 0.000 | 30.337 | 22.753 | 15.169 | 7.584 | 0.000 |
| Corn gluten | 0.000 | 0.000 | 0.804 | 1.607 | 2.411 | 3.215 | 0.000 | 0.000 | 0.804 | 1.607 | 2.411 | 3.215 |
| Soybean oil | 1.544 | 10.100 | 8.145 | 6.190 | 4.235 | 2.280 | 1.544 | 10.100 | 8.145 | 6.190 | 4.235 | 2.280 |
| Dicalcium phosphate | 1.053 | 0.384 | 0.544 | 0.704 | 0.864 | 1.023 | 1.053 | 0.384 | 0.544 | 0.704 | 0.864 | 1.023 |
| Limestone | 1.110 | 1.484 | 1.395 | 1.306 | 1.218 | 1.129 | 1.110 | 1.484 | 1.395 | 1.306 | 1.218 | 1.129 |
| Salt | 0.393 | 0.389 | 0.386 | 0.384 | 0.382 | 0.379 | 0.393 | 0.389 | 0.386 | 0.384 | 0.382 | 0.379 |
| L-Lysine | 0.098 | 0.153 | 0.143 | 0.134 | 0.125 | 0.116 | 0.098 | 0.153 | 0.143 | 0.134 | 0.125 | 0.116 |
| DL-Methionine | 0.373 | 0.385 | 0.382 | 0.379 | 0.376 | 0.373 | 0.373 | 0.385 | 0.382 | 0.379 | 0.376 | 0.373 |
| L-Threonine | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| Mineral premix1 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 |
| Vitamin premix2 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 |
| Quantum Blue3 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 |
| Signis4 | 0.010 | 0.010 | 0.010 | 0.010 | 0.010 | 0.010 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| Inert | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.010 | 0.010 | 0.010 | 0.010 | 0.010 | 0.010 |
| Total | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 |
| Calculated nutrients | ||||||||||||
| CP, % | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 |
| ME, kcal/kg | 2900 | 2900 | 2900 | 2900 | 2900 | 2900 | 2900 | 2900 | 2900 | 2900 | 2900 | 2900 |
| CF, % | 2.808 | 4.855 | 4.386 | 3.916 | 3.446 | 2.977 | 2.808 | 4.855 | 4.386 | 3.916 | 3.446 | 2.977 |
| NDF, % | 13.049 | 19.383 | 17.960 | 16.538 | 15.115 | 13.693 | 13.049 | 19.383 | 17.960 | 16.538 | 15.115 | 13.693 |
| MM, % | 3.333 | 4.052 | 3.895 | 3.738 | 3.582 | 3.425 | 3.333 | 4.052 | 3.895 | 3.738 | 3.582 | 3.425 |
| Starch, % | 34.519 | 21.041 | 24.075 | 27.110 | 30.144 | 33.178 | 34.519 | 21.041 | 24.075 | 27.110 | 30.144 | 33.178 |
| Total NSP, % | 10.269 | 15.100 | 14.031 | 12.961 | 11.892 | 10.822 | 10.269 | 15.100 | 14.031 | 12.961 | 11.892 | 10.822 |
| Insoluble NSP, % | 8.365 | 12.181 | 11.354 | 10.527 | 9.699 | 8.872 | 8.365 | 12.181 | 11.354 | 10.527 | 9.699 | 8.872 |
| Soluble NSP, % | 1.904 | 2.919 | 2.677 | 2.434 | 2.192 | 1.950 | 1.904 | 2.919 | 2.677 | 2.434 | 2.192 | 1.950 |
| Total AX, % | 3.405 | 6.749 | 5.993 | 5.238 | 4.483 | 3.727 | 3.405 | 6.749 | 5.993 | 5.238 | 4.483 | 3.727 |
| Soluble AX, % | 0.327 | 0.900 | 0.762 | 0.625 | 0.487 | 0.350 | 0.327 | 0.900 | 0.762 | 0.625 | 0.487 | 0.350 |
| Insoluble AX, % | 3.078 | 5.847 | 5.230 | 4.612 | 3.995 | 3.377 | 3.078 | 5.847 | 5.230 | 4.612 | 3.995 | 3.377 |
| EE, % | 4.731 | 12.495 | 10.726 | 8.957 | 7.189 | 5.420 | 4.731 | 12.495 | 10.726 | 8.957 | 7.189 | 5.420 |
| Calcium, % | 0.850 | 0.850 | 0.850 | 0.850 | 0.850 | 0.850 | 0.850 | 0.850 | 0.850 | 0.850 | 0.850 | 0.850 |
| Available phosphorus, % | 0.320 | 0.320 | 0.320 | 0.320 | 0.320 | 0.320 | 0.320 | 0.320 | 0.320 | 0.320 | 0.320 | 0.320 |
| Sodium, % | 0.170 | 0.170 | 0.170 | 0.170 | 0.170 | 0.170 | 0.170 | 0.170 | 0.170 | 0.170 | 0.170 | 0.170 |
| Chlorine, % | 0.302 | 0.286 | 0.289 | 0.292 | 0.295 | 0.297 | 0.302 | 0.286 | 0.289 | 0.292 | 0.295 | 0.297 |
| Potassium, % | 0.989 | 1.127 | 1.094 | 1.060 | 1.027 | 0.994 | 0.989 | 1.127 | 1.094 | 1.060 | 1.027 | 0.994 |
| Digestible amino acids | ||||||||||||
| Lysine, % | 1.370 | 1.370 | 1.370 | 1.370 | 1.370 | 1.370 | 1.370 | 1.370 | 1.370 | 1.370 | 1.370 | 1.370 |
| Methionine, % | 0.703 | 0.692 | 0.695 | 0.697 | 0.699 | 0.702 | 0.703 | 0.692 | 0.695 | 0.697 | 0.699 | 0.702 |
| Methionine + Cysteine, % | 1.040 | 1.040 | 1.040 | 1.040 | 1.040 | 1.040 | 1.040 | 1.040 | 1.040 | 1.040 | 1.040 | 1.040 |
| Threonine, % | 0.856 | 0.787 | 0.803 | 0.819 | 0.835 | 0.852 | 0.856 | 0.787 | 0.803 | 0.819 | 0.835 | 0.852 |
| Tryptophan, % | 0.297 | 0.306 | 0.303 | 0.300 | 0.296 | 0.293 | 0.297 | 0.306 | 0.303 | 0.300 | 0.296 | 0.293 |
| Valine, % | 1.057 | 1.020 | 1.029 | 1.038 | 1.048 | 1.057 | 1.057 | 1.020 | 1.029 | 1.038 | 1.048 | 1.057 |
¹ Copper 10,000mg; Iodine 160mg; Manganese 14,000mg; Selenium 108mg and Zinc 14,000mg; 2 Mineral and vitamin premix: Levels per kg of product: Vit. at 2,090,000 IU; Vit. E 7,600mg; Vit D3 332,500 IU; Vit k3 950mg; Nicotinic Acid 8,500mg; Vit B1 475mg; Vit B12 3,800 mg; Vit. B2 1,900mg; Folic Acid 237.5mg; Biotin 38mb; Choline 72,000mg; Pantothenic Acid 3,800mg; 3 Phytase equivalent to the supplemental dose of 500 FTU/kg of feed; 4 Stimbiotic: β-1,4-endo-xylanase and xylo-oligosaccharides, AB Vista, Marlborough, UK, providing 16,000 BXU/kg; L100 = 100% high fiber – whole wheat and wheat bran; L75H25 = 75% high fiber + 25% low fiber; L50H50 = 50% high fiber + 50% low fiber; L25H75 = 25% high fiber + 75% low fiber; H100 = 100% low fiber – corn germ; CP = Crude Protein; ME = Metabolizable Energy; CF = Crude Fiber; NDF = Neutral Detergent Fiber; MM = Mineral Matter; NSP = Non-Starch Polysaccharides; AX = Arabinoxylans; EE = Ether Extract.
Table 2.
Composition and nutritional values of diets with different fiber profiles, with or without stimbiotic, for European quails in the growth phase (15–35 days).
Table 2.
Composition and nutritional values of diets with different fiber profiles, with or without stimbiotic, for European quails in the growth phase (15–35 days).
| Ingredients, % | With Stimbiotic | Without Stimbiotic | ||||||||||
| Control | 100H | 75H25L | 50H50L | 25H75L | 100L | Control | 100H | 75H25L | 50H50L | 25H75L | 100L | |
| Corn | 60.491 | 0.000 | 14.245 | 28.490 | 42.735 | 56.980 | 60.491 | 0.000 | 14.245 | 28.490 | 42.735 | 56.980 |
| Soybean meal | 34.809 | 28.153 | 29.773 | 31.393 | 33.012 | 34.632 | 34.809 | 28.153 | 29.773 | 31.393 | 33.012 | 34.632 |
| Whole wheat | 0.000 | 36.122 | 27.091 | 18.061 | 9.030 | 0.000 | 0.000 | 36.122 | 27.091 | 18.061 | 9.030 | 0.000 |
| Wheat bran | 0.000 | 3.761 | 2.821 | 1.880 | 0.940 | 0.000 | 0.000 | 3.761 | 2.821 | 1.880 | 0.940 | 0.000 |
| Corn gluten | 0.000 | 19.834 | 15.613 | 11.392 | 7.171 | 2.950 | 0.000 | 19.834 | 15.613 | 11.392 | 7.171 | 2.950 |
| Soybean oil | 2.055 | 9.628 | 7.929 | 6.231 | 4.532 | 2.833 | 2.055 | 9.628 | 7.929 | 6.231 | 4.532 | 2.833 |
| Dicalcium phosphate | 0.868 | 0.534 | 0.609 | 0.685 | 0.760 | 0.835 | 0.868 | 0.534 | 0.609 | 0.685 | 0.760 | 0.835 |
| Limestone | 0.937 | 1.114 | 1.074 | 1.033 | 0.993 | 0.953 | 0.937 | 1.114 | 1.074 | 1.033 | 0.993 | 0.953 |
| Salt | 0.345 | 0.263 | 0.280 | 0.298 | 0.315 | 0.333 | 0.345 | 0.263 | 0.280 | 0.298 | 0.315 | 0.333 |
| L-Lysine | 0.000 | 0.092 | 0.069 | 0.046 | 0.023 | 0.000 | 0.000 | 0.092 | 0.069 | 0.046 | 0.023 | 0.000 |
| DL-Methionine | 0.223 | 0.187 | 0.194 | 0.202 | 0.210 | 0.217 | 0.223 | 0.187 | 0.194 | 0.202 | 0.210 | 0.217 |
| L-Threonine | 0.059 | 0.100 | 0.088 | 0.077 | 0.065 | 0.054 | 0.059 | 0.100 | 0.088 | 0.077 | 0.065 | 0.054 |
| Mineral premix1 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 |
| Vitamin premix2 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 | 0.050 |
| Quantum Blue3 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 |
| Signis4 | 0.010 | 0.010 | 0.010 | 0.010 | 0.010 | 0.010 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| Inert | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.010 | 0.010 | 0.010 | 0.010 | 0.010 | 0.010 |
| Total | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 | 100.000 |
| Calculated nutrients | ||||||||||||
| CP, % | 22 | 22 | 22 | 22 | 22 | 22 | 22 | 22 | 22 | 22 | 22 | 22 |
| ME, kcal/kg | 3050 | 3050 | 3050 | 3050 | 3050 | 3050 | 3050 | 3050 | 3050 | 3050 | 3050 | 3050 |
| CF, % | 2.540 | 4.020 | 3.693 | 3.366 | 3.039 | 2.712 | 2.540 | 4.020 | 3.693 | 3.366 | 3.039 | 2.712 |
| NDF, % | 13.084 | 16.736 | 15.968 | 15.200 | 14.432 | 13.664 | 13.084 | 16.736 | 15.968 | 15.200 | 14.432 | 13.664 |
| MM, % | 2.805 | 3.611 | 3.439 | 3.268 | 3.096 | 2.925 | 2.805 | 3.611 | 3.439 | 3.268 | 3.096 | 2.925 |
| Starch, % | 40.580 | 27.205 | 30.131 | 33.056 | 35.982 | 38.908 | 40.580 | 27.205 | 30.131 | 33.056 | 35.982 | 38.908 |
| Total NSP, % | 9.017 | 14.000 | 12.901 | 11.803 | 10.704 | 9.605 | 9.017 | 14.000 | 12.901 | 11.803 | 10.704 | 9.605 |
| Insoluble NSP, % | 7.555 | 11.641 | 10.748 | 9.856 | 8.964 | 8.071 | 7.555 | 11.641 | 10.748 | 9.856 | 8.964 | 8.071 |
| Soluble NSP, % | 1.462 | 2.359 | 2.153 | 1.946 | 1.740 | 1.534 | 1.462 | 2.359 | 2.153 | 1.946 | 1.740 | 1.534 |
| Total AX, % | 3.322 | 6.380 | 5.691 | 5.001 | 4.312 | 3.622 | 3.322 | 6.380 | 5.691 | 5.001 | 4.312 | 3.622 |
| Soluble AX, % | 0.275 | 0.800 | 0.675 | 0.550 | 0.425 | 0.300 | 0.275 | 0.800 | 0.675 | 0.550 | 0.425 | 0.300 |
| Insoluble AX, % | 3.047 | 5.577 | 5.013 | 4.450 | 3.886 | 3.322 | 3.047 | 5.577 | 5.013 | 4.450 | 3.886 | 3.322 |
| EE, % | 5.344 | 11.705 | 10.296 | 8.887 | 7.478 | 6.070 | 5.344 | 11.705 | 10.296 | 8.887 | 7.478 | 6.070 |
| Calcium, % | 0.700 | 0.700 | 0.700 | 0.700 | 0.700 | 0.700 | 0.700 | 0.700 | 0.700 | 0.700 | 0.700 | 0.700 |
| Available phosphorus, % | 0.270 | 0.270 | 0.270 | 0.270 | 0.270 | 0.270 | 0.270 | 0.270 | 0.270 | 0.270 | 0.270 | 0.270 |
| Sodium, % | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 | 0.150 |
| Chlorine, % | 0.278 | 0.244 | 0.251 | 0.258 | 0.266 | 0.273 | 0.278 | 0.244 | 0.251 | 0.258 | 0.266 | 0.273 |
| Potassium, % | 0.831 | 0.892 | 0.880 | 0.868 | 0.857 | 0.845 | 0.831 | 0.892 | 0.880 | 0.868 | 0.857 | 0.845 |
| Digestible amino acids | ||||||||||||
| Lysine, % | 1.020 | 1.020 | 1.020 | 1.020 | 1.020 | 1.020 | 1.020 | 1.020 | 1.020 | 1.020 | 1.020 | 1.020 |
| Methionine, % | 0.510 | 0.471 | 0.479 | 0.488 | 0.497 | 0.506 | 0.510 | 0.471 | 0.479 | 0.488 | 0.497 | 0.506 |
| Methionine + Cysteine, % | 0.800 | 0.800 | 0.800 | 0.800 | 0.800 | 0.800 | 0.800 | 0.800 | 0.800 | 0.800 | 0.800 | 0.800 |
| Threonine, % | 0.780 | 0.780 | 0.780 | 0.780 | 0.780 | 0.780 | 0.780 | 0.780 | 0.780 | 0.780 | 0.780 | 0.780 |
| Tryptophan, % | 0.242 | 0.236 | 0.237 | 0.239 | 0.240 | 0.241 | 0.242 | 0.236 | 0.237 | 0.239 | 0.240 | 0.241 |
| Valine, % | 0.880 | 0.920 | 0.913 | 0.906 | 0.899 | 0.892 | 0.880 | 0.920 | 0.913 | 0.906 | 0.899 | 0.892 |
¹ Copper 10,000mg; Iodine 160mg; Manganese 14,000mg; Selenium 108mg and Zinc 14,000mg; 2 Mineral and vitamin premix: Levels per kg of product: Vit. at 2,090,000 IU; Vit. E 7,600mg; Vit D3 332,500 IU; Vit k3 950mg; Nicotinic Acid 8,500mg; Vit B1 475mg; Vit B12 3,800 mg; Vit. B2 1,900mg; Folic Acid 237.5mg; Biotin 38mb; Choline 72,000mg; Pantothenic Acid 3,800mg; 3 Phytase equivalent to the supplemental dose of 500 FTU/kg of feed; 4 Stimbiotic: β-1,4-endo-xylanase and xylo-oligosaccharides, AB Vista, Marlborough, UK, providing 16,000 BXU/kg; L100 = 100% high fiber – whole wheat and wheat bran; L75H25 = 75% high fiber + 25% low fiber; L50H50 = 50% high fiber + 50% low fiber; L25H75 = 25% high fiber + 75% low fiber; H100 = 100% low fiber – corn germ; CP = Crude Protein; ME = Metabolizable Energy; CF = Crude Fiber; NDF = Neutral Detergent Fiber; MM = Mineral Matter; NSP = Non-Starch Polysaccharides; AX = Arabinoxylans; EE = Ether Extract.
Table 3.
Hemogram of European quails (Coturnix coturnix coturnix) at 14 days of age, fed diets with different fiber profiles, with or without stimbiotic supplementation during the period from 1 to 14 days.
Table 3.
Hemogram of European quails (Coturnix coturnix coturnix) at 14 days of age, fed diets with different fiber profiles, with or without stimbiotic supplementation during the period from 1 to 14 days.
| Variables | Stimbiotic | Fiber | SEM | P-Value | ||||||||
| - | + | Control | 100H | 75H25L | 50H50L | 25H75L | 100L | S | F | S*F | ||
| Packed cell volume, L/L | 0.341 | 0.343 | 0.351 | 0.344 | 0.334 | 0.337 | 0.342 | 0.343 | 2.76 | 0.6586 | 0.6129 | 0.8699 |
| Erythrocyte count, ×10¹²/L | 2.49 | 2.39 | 2.45 | 2.41 | 2.29 | 2.44 | 2.46 | 2.60 | 0.39 | 0.2353 | 0.3883 | 0.7497 |
| Hemoglobin, g/dL | 6.61b | 7.35a | 7.16 | 7.02 | 6.86 | 6.87 | 6.91 | 7.08 | 0.75 | <0.001 | 0.8249 | 0.2448 |
| MCV, fL | 139 | 145.95 | 145.37 | 145.71 | 151.14 | 140.45 | 142.01 | 132.11 | 20.30 | 0.1555 | 0.2193 | 0.6958 |
| MCHC, g/dL | 19.41 | 21.04 | 20.44 | 20.34 | 20.52 | 20.24 | 20.22 | 20.63 | 1.37 | 0.0564 | 0.9600 | 0.0560 |
| Total platelets, ×10⁹/L | 39.26 | 35.96 | 45.87a | 31.69b | 34.22b | 35.07b | 40.37ab | 36.25b | 10.78 | 0.0848 | 0.0062 | 0.3595 |
| Total leukocyte count, ×10⁹/L | 8.81a | 6.70b | 8.07 | 7.49 | 6.12 | 7.42 | 8.91 | 8.44 | 3.69 | 0.0065 | 0.3626 | 0.1383 |
| Monocyte, ×10⁹/L | 0.02 | 0.02 | 0.04 | 0.01 | 0.01 | 0.01 | 0.02 | 0.02 | 0.05 | 0.3533 | 0.4431 | 0.4219 |
| Lymphocyte, ×10⁹/L | 5.85a | 4.79b | 5.25 | 5.07 | 4.68 | 5.11 | 6.45 | 5.31 | 2.48 | 0.0392 | 0.4652 | 0.0797 |
| Eosinophil, ×10⁹/L | 0.28 | 0.28 | 0.38 | 0.17 | 0.15 | 0.21 | 0.21 | 0.31 | 0.26 | 0.1043 | 0.1500 | 0.4244 |
| Heterophile, ×10⁹/L | 2.49a | 1.58b | 2.22 | 2.04 | 1.13 | 1.93 | 2.11 | 2.71 | 1.80 | 0.0162 | 0.2735 | 0.5212 |
| Basophil, ×10⁹/L | 0.19 | 0.17 | 0.18 | 0.19 | 0.15 | 0.14 | 0.12 | 0.08 | 0.13 | 0.0509 | 0.1763 | 0.3053 |
| H/L | 0.43 | 0.33 | 0.42 | 0.40 | 0.24 | 0.38 | 0.33 | 0.51 | 0.73 | 0.5346 | 0.3642 | 0.7329 |
MCV = mean corpuscular volume; MCHC = mean corpuscular hemoglobin concentration; H/L = heterophile/lymphocyte ratio. The symbol (+) indicates supplementation with the stimbiotic and (–) indicates no supplementation. Fiber profiles: Control (corn- and soybean meal-based diet); L100 (100% high fiber – whole wheat and wheat bran); L75H25 (75% high fiber + 25% low fiber); L50H50 (50% high fiber + 50% low fiber); L25H75 (25% high fiber + 75% low fiber); H100 (100% low fiber – corn germ). Different letters in the same row indicate significant differences according to Tukey’s test (p < 0.05). SEM = standard error of the mean. E = stimbiotic; F = fiber; E×F = stimbiotic × fiber interaction.
Table 4.
Hemogram of European quails (Coturnix coturnix coturnix) at 35 days of age, fed diets with different fiber profiles, with or without stimbiotic supplementation during the period from 1 to 35 days.
Table 4.
Hemogram of European quails (Coturnix coturnix coturnix) at 35 days of age, fed diets with different fiber profiles, with or without stimbiotic supplementation during the period from 1 to 35 days.
| Variables | Stimbiotic | Fiber | SEM | P-Value | ||||||||
| - | + | Control | 100H | 75H25L | 50H50L | 25H75L | 100L | S | F | S*F | ||
| Packed cell volume, L/L | 0.403b | 0.428a | 0.423 | 0.427 | 0.404 | 0.401 | 0.415 | 0.427 | 5.15 | 0.0205 | 0.6528 | 0.7959 |
| Erythrocyte count, ×10¹²/L | 3.54 | 3.76 | 4.41 | 3.20 | 3.23 | 3.13 | 4.56 | 3.43 | 2.94 | 0.7117 | 0.5789 | 0.2184 |
| Hemoglobin, g/dL | 9.18b | 9.89a | 9.88 | 9.69 | 8.99 | 9.64 | 9.60 | 9.45 | 1.33 | 0.0117 | 0.5370 | 0.1136 |
| MCV, fL | 128.11 | 127.78 | 127.42 | 132.79 | 127.41 | 129.64 | 122.61 | 127.71 | 24.82 | 0.9482 | 0.9270 | 0.2774 |
| MCHC, g/dL | 22.33 | 22.66 | 21.89 | 23.04 | 22.31 | 24.07 | 21.67 | 21.97 | 3.82 | 0.6817 | 0.4941 | 0.1258 |
| Total platelets, ×10⁹/L | 35.98 | 32.79 | 37.69 | 38.50 | 33.66 | 27.70 | 33.57 | 35.16 | 12.37 | 0.2022 | 0.1832 | 0.8435 |
| Total leukocyte count, ×10⁹/L | 9.09a | 7.71b | 7.03 | 8.90 | 8.25 | 9.72 | 9.13 | 7.38 | 3.11 | 0.0386 | 0.1256 | 0.0803 |
| Monocyte, ×10⁹/L | 0.16 | 0.14 | 0.35 | 0.03 | 0.02 | 0.06 | 0.41 | 0.03 | 0.80 | 0.9282 | 0.5604 | 0.2698 |
| Lymphocyte, ×10⁹/L | 6.66a | 5.77b | 5.56 | 6.02 | 6.58 | 7.17 | 6.11 | 5.82 | 2.10 | 0.0437 | 0.3156 | 0.1298 |
| Eosinophil, ×10⁹/L | 0.48 | 0.46 | 0.30b | 0.78a | 0.34b | 0.57ab | 0.48ab | 0.37b | 0.38 | 0.8338 | 0.0055 | 0.1298 |
| Heterophile, ×10⁹/L | 1.74 | 1.36 | 1.04 | 1.76 | 1.27 | 1.82 | 2.43 | 1.05 | 1.76 | 0.2950 | 0.2155 | 0.2246 |
| Basophil, ×10⁹/L | 0.11 | 0.11 | 0.13 | 0.11 | 0.04 | 0.09 | 0.18 | 0.11 | 0.20 | 0.9000 | 0.5426 | 0.5995 |
| H/L | 0.26 | 0.24 | 0.19 | 0.29 | 0.19 | 0.25 | 0.40 | 0.18 | 0.84 | 0.6100 | 0.5350 | 0.9460 |
MCV = mean corpuscular volume; MCHC = mean corpuscular hemoglobin concentration; H/L = heterophile/lymphocyte ratio. The symbol (+) indicates supplementation with the stimbiotic and (–) indicates no supplementation. Fiber profiles: Control (corn- and soybean meal-based diet); L100 (100% high fiber – whole wheat and wheat bran); L75H25 (75% high fiber + 25% low fiber); L50H50 (50% high fiber + 50% low fiber); L25H75 (25% high fiber + 75% low fiber); H100 (100% low fiber – corn germ). Different letters in the same row indicate significant differences according to Tukey’s test (p < 0.05). SEM = standard error of the mean. E = stimbiotic; F = fiber; E×F = stimbiotic × fiber interaction.
Table 5.
Biochemical parameters of European quails (Coturnix coturnix coturnix) at 14 days of age, fed diets with different fiber profiles, with or without stimbiotic supplementation during the period from 1 to 14 days.
Table 5.
Biochemical parameters of European quails (Coturnix coturnix coturnix) at 14 days of age, fed diets with different fiber profiles, with or without stimbiotic supplementation during the period from 1 to 14 days.
| Variables | Stimbiotic | Fiber | SEM | P-Value | ||||||||
| - | + | Control | 100H | 75H25L | 50H50L | 25H75L | 100L | S | F | S*F | ||
| Albumin, g/dL | 1.14 | 1.1 | 1.17 | 1.01 | 1.16 | 1.13 | 1.14 | 1.13 | 0.15 | 0.2573 | 0.0652 | 0.636 |
| ALT, UI/L | 10.58 | 14.38 | 14.6 | 13.19 | 13.19 | 11.81 | 11.13 | 10.94 | 4.52 | 0.5768 | 0.1662 | 0.0083 |
| AST, UI/L | 218.71 | 214.42 | 230.08 | 224.25 | 218.81 | 215.88 | 215.69 | 194.5 | 58.63 | 0.7208 | 0.6337 | 0.4337 |
| GGT, UI/L | 1.29 | 1.27 | 1.25 | 1.29 | 1.71 | 1.46 | 1.15 | 0.86 | 1.19 | 0.9018 | 0.4749 | 0.3318 |
| Total protein, g/dL | 2.94 | 2.88 | 2.98 | 2.76 | 2.96 | 2.95 | 2.91 | 2.94 | 0.34 | 0.3248 | 0.5286 | 0.2842 |
| Cholesterol, mg/dL | 149.45 | 140.68 | 136.08 | 142.74 | 157.26 | 152.46 | 146.29 | 138.56 | 29.22 | 0.105 | 0.2962 | 0.6456 |
| Triglycerides, mg/dL | 77.51b | 115.58a | 144.86a | 73.36b | 72.82b | 93.84b | 104.84ab | 90.07b | 46.91 | 0.0001 | 0.0003 | 0.3197 |
ALT = alanine aminotransferase; AST = aspartate aminotransferase; GGT = gamma-glutamyl transferase. The symbol (+) indicates supplementation with the stimbiotic and (–) indicates no supplementation. Fiber profiles: Control (corn- and soybean meal-based diet); L100 (100% high fiber – whole wheat and wheat bran); L75H25 (75% high fiber + 25% low fiber); L50H50 (50% high fiber + 50% low fiber); L25H75 (25% high fiber + 75% low fiber); H100 (100% low fiber – corn germ). Different letters in the same row indicate significant differences according to Tukey’s test (p < 0.05). SEM = standard error of the mean. E = stimbiotic; F = fiber; E×F = stimbiotic × fiber interaction.
Table 6.
Biochemical parameters of European quails (Coturnix coturnix coturnix) at 35 days of age, fed diets with different fiber profiles, with or without stimbiotic supplementation during the period from 1 to 35 days.
Table 6.
Biochemical parameters of European quails (Coturnix coturnix coturnix) at 35 days of age, fed diets with different fiber profiles, with or without stimbiotic supplementation during the period from 1 to 35 days.
| Variables | Stimbiotic | Fiber | SEM | P-Value | ||||||||
| - | + | Control | 100H | 75H25L | 50H50L | 25H75L | 100L | S | F | S*F | ||
| Albumin, g/dL | 5.61 | 4.94 | 13.89 | 1.24 | 1.36 | 1.40 | 12.65 | 1.31 | 26.91 | 0.9048 | 0.5368 | 0.2791 |
| ALT, UI/L | 12.72 | 12.625 | 11.74 | 9.84 | 13.75 | 16.50 | 12.41 | 11.88 | 12.99 | 0.9393 | 0.7968 | 0.4774 |
| AST, UI/L | 198.69 | 212.96 | 206.18 | 204.79 | 201.88 | 199.44 | 199.86 | 222.13 | 52.29 | 0.1868 | 0.3386 | 0.0774 |
| GGT, UI/L | 37.72 | 89.94 | 15.10 | 36.53 | 45.07 | 137.76 | 94.89 | 53.89 | 139.21 | 0.0726 | 0.1520 | 0.9620 |
| Total protein, g/dL | 3.35 | 3.63 | 3.35 | 3.20 | 3.50 | 3.70 | 3.88 | 3.30 | 0.84 | 0.1013 | 0.1995 | 0.3563 |
| Cholesterol, mg/dL | 186.54 | 195.79 | 199.75 | 169.79 | 182.72 | 208.11 | 188.26 | 196.86 | 63.10 | 0.4795 | 0.6011 | 0.4710 |
| Triglycerides, mg/dL | 285.94 | 216.99 | 190.46 | 244.43 | 294.29 | 158.16 | 324.71 | 300.87 | 244.26 | 0.1749 | 0.3409 | 0.2480 |
ALT = alanine aminotransferase; AST = aspartate aminotransferase; GGT = gamma-glutamyl transferase. The symbol (+) indicates supplementation with the stimbiotic and (–) indicates no supplementation. Fiber profiles: Control (corn- and soybean meal-based diet); L100 (100% high fiber – whole wheat and wheat bran); L75H25 (75% high fiber + 25% low fiber); L50H50 (50% high fiber + 50% low fiber); L25H75 (25% high fiber + 75% low fiber); H100 (100% low fiber – corn germ). Different letters in the same row indicate significant differences according to Tukey’s test (p < 0.05). SEM = standard error of the mean. E = stimbiotic; F = fiber; E×F = stimbiotic × fiber interaction.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



