
Submitted:
04 October 2025
Posted:
06 October 2025
You are already at the latest version
Abstract
Ghrelin, the endogenous ligand of the growth hormone secretagogue receptor 1a (GHSR1a), has expanded from a “hunger hormone” to a pleiotropic regulator of whole-body physiology. This review synthesizes current advances spanning ghrelin biogenesis, signaling, and systems biology. Physiologically, preproghrelin processing and O-acylation by ghrelin O-acyltransferase (GOAT) generate acyl-ghrelin, the high-potency GHSR1a agonist; des-acyl ghrelin predominates in circu-lation and exerts context-dependent, GHSR1a-independent or low-potency effects, while truncated “mini-ghrelins” can act as com-petitive antagonists. Emergence of synthetic ligands, agonists, antago-nists and reverse-agonists provided tools necessary to decipher of GHSR1a activity. Recent cryo-EM structures of GHSR1a with peptide and small-molecule ligands reveal a bipartite binding pocket and pro-vide a framework for biased signaling, constitutive activity, and receptor partner selectivity. Beyond regulation of feeding and growth hormone release, ghrelin modulates glucose homeostasis, gastric secretion and motility, cardiovascular tone, bone remodeling, renal hemodynamics, and innate immunity. Ghrelin broadly dampens pro-inflammatory re-sponses and promotes reparative macrophage phenotypes. In the emerging field of ghrelin activity in central nervous system, ghrelin in-fluences neuroprotection, stress reactivity, sleep architecture, and is implicated in depression, Alzheimer’s disease, and substance abuse disorders. Practical and transitional aspects are also highlighted: ap-proaches for ghrelin stabilization; recent GHSR1a agonists/antagonists and inverse agonists findings; LEAP-2–based strategies; and emerging GOAT inhibitors. Together, structural insights and pathway selectivity position the ghrelin system as a druggable axis for the management of inflammatory diseases, neuropsychiatric and addiction conditions and for obesity treatment in post–GLP-1 receptor agonist era.
Keywords:
ghrelin
; GHSR1a
; G protein-coupled receptor
; GOAT
1. Introduction
In 1999, Kojima M. et al. utilized a screening method to associate orphan G protein-coupled receptors (GPCRs) with unknown ligands. Their method detected an increase in intracellular calcium levels induced by agonists, leading to the purification of a 28-amino-acid peptide from the gut. This peptide was identified as the natural ligand for the growth hormone secretagogue receptor 1a (GHSR1a) and was named “ghrelin”, derived from the Proto-Indo-European root “ghre”, signifying “grow” [1]. Since then, ghrelin has been discovered in many different organs, where it is involved in a wide range of processes and activities.
Comprehensive RT-PCR studies revealed that ghrelin mRNA is present in various tissues. The highest levels were observed in the stomach, whereas lower levels were detected in the liver, lung, kidney, and skeletal muscle [2]. In turn, at the protein level, ghrelin was detected in the stomach, small intestine, brain, cerebellum, pituitary, lung, skeletal muscle, pancreas, salivary glands, adrenal gland, ovary, and testis, with concentrations ranging from 0.05 to 1.43 ng/mg of homogenate protein. The lung and brain showed the highest levels. Protein was not found in the heart, liver, and kidney, highlighting a potential discrepancy between gene and protein expression [2]. Recent studies have shown that ghrelin is also expressed in the oral cavity since it was detected in saliva, gingival crevicular fluid (GCF), salivary glands, oral epithelial cells, and oral squamous cell carcinoma lesions. Its concentration is lowest in serum, while in GCF it is ~500-fold higher than in saliva [3,4].
The level of ghrelin-like immunoreactivity in plasma among healthy individuals, as determined by a specialized radioimmunoassay (RIA), stood at 117 ± 37 fmol/mL [5]. Ghrelin levels in the serum rise with age irrespective of sex. They fluctuate considerably over the course of the day, peaking during sleep [6]. Unlike other gut hormones, ghrelin levels in the plasma rise during fasting, and drop after eating [7]. Chronic high-calorie diets and obesity tend to lower plasma ghrelin levels in humans [7,8]. In rodents, long-term high-fat diets result in obesity and reduced stomach ghrelin production and secretion, simultaneously increasing the number of ghrelin-secreting cells [9]. However, the exact impact of increased adiposity on ghrelin production is unclear [10].
Ghrelin is reported to be the orexigenic peptide mainly secreted by the X/A-like cells, a group of unique endocrine cells in the gastrointestinal tract [11]. Norepinephrine mediates the increase in ghrelin levels before a meal, whereas the decrease in ghrelin after a meal is regulated by glucose and insulin, with insulin contributing additively [12,13]. In addition to reduced stomach production of ghrelin in obesity, decreased systemic levels have been observed in conditions such as male hypogonadism, untreated hyperthyroidism, polycystic ovary syndrome, and H. pylori-induced gastritis [14,15,16,17]. Conversely, elevated ghrelin levels are observed in conditions such as anorexia nervosa, in lean individuals, Prader-Willi syndrome, and following the eradication of H. pylori [17].
Two other research groups employed ghrelin-secreting cell lines from transgenic mice and primary cell cultures from mice and rats to demonstrate the mechanism underlying the regulation of ghrelin secretion [18,19]. The data demonstrate that a number of factors, including glucose, glucagon, dopamine, insulin, oxytocin, somatostatin, and long-chain fatty acids, directly act on the ghrelin-producing cells, thereby controlling ghrelin release [12,18,19,20]. Recently, significant progress in understanding of the ghrelin system was made, reflected by the determination of the ghrelin-bound GHSR1a structure, functional description of the receptor’s biased signaling and ongoing progress in structural and functional analysis of the ghrelin O-acyltransferase (GOAT). Together with the increased understanding of ghrelin’s role in immune and central nervous systems and an emerging role of ghrelin in dopamine-related signaling and addiction, ghrelin is again found at the forefront of research. This review aims to provide a comprehensive, modern update on ghrelin biology.
2. Processing and Maturation of Ghrelin
To carry out its diverse functions, ghrelin must undergo a series of maturation steps. Ghrelin prepropeptide gene (GHRL) (Figure 1) consists of 6 exons and 4 introns, where mature preproghrelin mRNA is translated into preproghrelin, a 117-amino-acid product [21,22]. One noteworthy feature of this construct is the presence of obestatin, a putative proteolytic fragment with activities opposite to those of ghrelin [22,23]. While ghrelin has an appetite-stimulating impact, obestatin, which is made up of 23 amino acids, is mainly linked to enhancing feelings of fullness [23]. Rats given obestatin had lower food intake, inhibited jejunal contractions, and gained less weight [24]. Human studies, however, could not demonstrate any connection between obestatin and anorexia in general, weight loss, or other dietary issues [25].
To release mature ghrelin, the 117-amino-acid precursor undergoes systematic proteolytic processing within the endoplasmic reticulum (ER) to form the 28-amino-acid ghrelin. The process begins with signal peptidase-mediated removal of the signal peptide at Arg23, essential for translocation to the ER [26]. In stomach X/A-like cells, where processing is best described, ghrelin undergoes O-octanoylation at Ser3, catalyzed by ER-localized enzyme ghrelin O-acyltransferase (GOAT), and is subsequently cleaved by the prohormone convertase PC1/3 at Arg51 to yield the active 28-amino-acid acylated form of ghrelin (AG) [26,27]. The GOAT enzyme was initially identified to be expressed in the gut and ghrelin O-octanoylation is essential for its binding to GHSR1a [27,28]. Recently, however, GOAT was detected in urine and blood of prostate cancer (PC) patients and, in this limited studies, its levels were reported to outperform PSA in predicting aggressive disease [29,30].
Some portion of preproghrelin undergoes C-terminal trimming by a carboxypeptidase-B-like enzyme, resulting in the 27-amino-acid sequence of ghrelin, lacking the C-terminal Arg28 [31]. This type of processing, commonly seen in other peptide hormones (e.g., endorphins [32], cholecystokinin [33]), often involves cleavage at, or near basic residues (Arg or Lys) [32,33]. The 27-residue isoform can also arise from alternative splicing. A splice variant, prepro-des-Gln14-ghrelin (116 residues), produces des-Gln14-ghrelin, a second endogenous GHSR ligand. However, des-Gln1144-ghrelin appears to be less abundant in humans compared to rodents, where the ratio of ghrelin to des-Gln14-ghrelin precursors varies by species (e.g., ~5:1 in rat, ~6:5 in mouse) [34].
Several studies have demonstrated the existence of truncated, bioactive ghrelin isoforms, collectively termed “mini-ghrelins”, that retain the essential N-terminal octanoyl modification and can modulate ghrelin receptor signaling [35,36,37]. Nearly a decade ago, Satou et al. identified activated protein C (APC), a serine protease found in bovine plasma, as a novel ghrelin endopeptidase. APC selectively cleaves human octanoylated ghrelin between Arg15 and Lys16, generating ghrelin(1–15) as the predominant cleavage product. In vivo, the administration of ProTac, a snake venom–derived pharmacological activator of APC, significantly enhanced this cleavage in mice [35].
Two additional, shorter isoforms of mini-ghrelin have also been identified. Ghrelin(1–14) likely arises via C-terminal trimming of ghrelin(1–15), possibly mediated by plasma carboxypeptidases that remove terminal basic residues. The shortest characterized variant, ghrelin(1–11), is generated by cleavage at the Arg11-Val12 bond in human ghrelin. This specific cleavage does not occur in rodents, where the corresponding residues are Lys-Ala, highlighting differences in protease recognition and substrate specificity [37].
Naturally occurring splice variants of the ghrelin gene (GHRL) have been identified across vertebrate species. Comparative genomic analysis of 77 species revealed that exon 2, which encodes part of the ghrelin peptide, is a symmetrical exon, allowing it to be skipped without altering the downstream reading frame. Exon 2 skipping produces a truncated preproghrelin transcript that encodes a 13-amino-acid peptide, followed immediately by the obestatin coding region. This exon 2-deleted isoform retains the overall structural organization of the preprohormone and has been detected in species such as mice and sheep, indicating evolutionary conservation and supporting its potential functional relevance [36].
Despite the preserved ability of mini-ghrelins to bind GHSR1a and inhibit calcium channel activity in vitro, similar to full-length ghrelin, mini-ghrelins do not elicit canonical ghrelin responses in vivo, such as food intake stimulation or hypothalamic c-Fos activation. Instead, these peptides act as competitive antagonists, inhibiting the binding and orexigenic effects of exogenous ghrelin [37].
The amino acid sequences of mammalian ghrelin exhibit significant conservation, especially in the N-termini, where the ten amino acids display complete identity. The structural likeness and the ongoing necessity for acyl-modification of the third residue indicate the pivotal importance of this N-terminal segment in the peptide’s function. Also, it’s noteworthy that rat and human ghrelin differ by only two amino acid residues and both show comparable potency in activation of the human GHSR1a [38,39].
3. The GOAT—A Single Enzyme for the Single Substrate
GOAT was independently discovered by two labs in 2008 [27,28]. It is a member of membrane-bound-O-acyltransferase (MBOAT) family and is an integral membrane protein with 11 transmembrane α-helices. The GOAT structure remains uncharacterized, due to inherent difficulty in structural characterization of integral membrane proteins. Structures of other enzymes from the same family; DltB, a bacterial alanyltransferase solved by X-ray diffraction [40] and human sterol O-acyltransferase solved by cryo-EM [41] provide some insight into the enzyme mechanism and inhibitor design strategies for GOAT. In recent years, a topology-guided, computational model of GOAT structure was developed by James L. Hougland’s group [41] and facilitated identification of the unique internal channel in GOAT structure, enabling modification of the ghrelin present in the ER lumen with cytoplasmic octanoyl-CoA. GOAT forms an ellipsoidal cone embedded in the ER membrane, with a narrow end facing the lumen and the wide side directed toward the cytoplasm. Octanoyl-CoA is recruited from the cytoplasmic side and is not transported across the membrane, but rather the catalytic machinery of GOAT attaches the fatty acid chain to the lumen-delivered ghrelin within the channel. Based on the absolute conservation and its critical position within the channel, His338 is proposed as the catalytic residue of GOAT.
It has been proposed that the unexpected biological activity of des-acyl ghrelin (DAG) may, at least in part, be mediated by re-acylation. Indeed, this emerging hypothesis was raised by several reports indicating that adipogenic activity of des-acyl ghrelin is mediated by the presence of GHSR1a in rats and suggested extracellular activity of GOAT enzyme [42]. Similarly, the presence and GOAT activity on exogenous des-acyl ghrelin were reported in mouse hippocampus [43]. This is consistent with earlier reports, which indicated that acyl-ghrelin is subjected to unidirectional transport through the blood–brain barrier (BBB) in mice. The mouse acyl-ghrelin transport was effective only in brain-to-blood direction and negligible in the opposing blood-to-brain influx, while des-acyl mouse ghrelin was subjected to passive, membrane diffusion-based transport. As acyl-ghrelin is readily detectable in mouse brain, these findings indirectly suggested the presence of the local, extracellular GOAT activity [44]. Indeed, it was demonstrated recently that GOAT is expressed on the surface of LNCaP and 22Rv1 prostate cancer cell lines and is able to bind and modify exogenous des-acyl ghrelin [45]. The current model of the potential extracellular GOAT activity postulates that GOAT is present within the cell membrane, where it binds and modifies the exogenous des-acyl ghrelin, which is then recognized by the adjacent GHSR1a, enabling signal transduction into the target cell.
Des-acyl ghrelin (DAG), the predominant form of ghrelin in the bloodstream, comprises approximately 90% of total circulating ghrelin [46]. While ghrelin acylation is crucial for its complete activity through GHSR1a [47], research indicates that DAG, despite exhibiting weak yet complete agonism, shows low potency in displacing ligand binding at GHSR1a [38]. Significant differences in potency have been observed: DAG has an EC50 ranging from 1.6 to 2.4 µM, compared to ghrelin’s EC50 of 2–2.6 nM, spanning three orders of magnitude [48]. Another study found an even more significant disparity, with ghrelin having an EC50 of 1–2 nM compared to over 3.5 µM for DAG, again indicating a ~1033-fold lower potency of DAG than acyl-ghrelin at GHSR1a [39]. Circulating levels of ghrelin and DAG range from 0.1 to 0.5 nmol/L, though assays detecting bound peptides show higher levels (3–4 nmol/L). Therefore, DAG levels in the bloodstream are much lower than the concentration needed to activate GHSR1a [49].
Nevertheless, by activating the survival-promoting extracellular signal-regulated kinase 1/2 (ERK1/2) and PI3K/Akt signaling pathways, DAG has been demonstrated to support adipogenesis and have an anti-apoptotic impact on cardiomyocytes [42,50]. DAG was also reported to mediate anxiety-like behavior [51], and shift adipose tissue residual macrophages to M2 [52] in mice. In addition, DAG has been reported to reduce alcohol intake in rats in a dopamine-dependent manner [53]. In contrast to DAG’s function, AG appears important in controlling autophagy, a cellular process that breaks down proteins and organelles [54,55,56]. Des-acyl ghrelin also induces food intake via orexin neurons, independent of GHSR1a in mice and rats [57], although contrasting data were also reported [58]. Acylated ghrelin is swiftly cleared from plasma with a half-life of 9–13 minutes, whereas total ghrelin (including DAG) persists for 27–34 minutes [59]. Early reports indicated that ratio of active ghrelin to total ghrelin was approximately 1:20, as measured in healthy human sera by polyclonal rabbit antibodies raised against N-terminal fragment of ghrelin (active) and C-terminal fragment (total), respectively [60]. Similar findings were reported in rat serum, where early reports indicated 1:5 ratio of active to total ghrelin [61,62]. The presence of des-acyl ghrelin was attributed to serum-mediated degradation of the active molecule, not to the production and release of the des-acyl molecule. Indeed, after 240 minutes of acyl ghrelin incubation with human serum, nearly 50% of the peptide was converted to des-acyl form and no further processing, including proteolysis, was observed [63]. Some other reports hinted at the possibility of even faster processing of ghrelin in human serum [64,65]. More pronounced effect was observed in rat serum, as already after 30 minutes, ~60% of ghrelin-derived peptide was converted to des-acyl form [63]. This activity was attributed to butyrylcholinesterase and possibly other esterases in human serum, whereas in rat serum, only carboxylesterase was involved [63,64,66]. Proteolytic degradation of ghrelin was observed not in serum, but in stomach, liver and kidney tissue homogenates, which led to the generation of the biologically inactive fragments, indicating that proteases may be involved in ghrelin processing locally, but rather not systemically [63]. In parallel, other activities were identified, including description of Acyl-Protein Thioesterase 1/Lysophospholipase (APT1) as ghrelin deacylating enzyme in rat stomach and in fetal bovine serum [67,68]. Unexpectedly, α2-macroglobulin hydrolase activity was identified in rat serum by active-site labeling approach with ghrelin-derived activity-based probe (ABP), potentially accounting for up to 50% of ghrelin deacylase activity [69]. Further, activated protein C (APC) in bovine plasma was described as ghrelin-cleaving protease, leading to the release of biologically active ghrelin1-15 octanoylated peptide [35].
These findings led to the investigations on the appropriate conditions of the optimal collection and storage of samples for the subsequent ghrelin detection. Indeed, some level of protection was observed after the addition of PMSF to serum samples [63] or after the addition of EDTA and aprotinin during sample collection [70]. Also, the acidification of the serum samples with HCl to pH 3-4 led to the significant protection of ghrelin, an observation consistent with the inhibition of serum esterases by low pH [70]; however, increased ghrelin deacylation was observed in the samples containing ≥100 mM HCl [65]. Further research indicated 4-(2-aminoethyl)benzenesulfonyl fluoride hydrochloride (AEBSF) as effective compound in the active ghrelin protection in the human serum samples [65]. In recent years alkyl fluorophosphonate inhibitors were proposed as the optimal compounds stabilizing ghrelin octanoylation in biological samples. This is exemplified by active ghrelin protection in the presence of methoxy arachidonyl fluorophosphonate (MAFP) in biological samples, including cell lysates and rat blood, a treatment superior to all previously described [71]. Indeed, addition of MAFP during the collection of the rat blood samples resulted in the unprecedented acyl-ghrelin protection and led to the detection of unexpectedly high active ghrelin levels [71]. It is worth noting that significant levels of ghrelin deacylase APT1 are released from the mouse-derived RAW264.7 macrophage cell line upon LPS stimulation and similar enzymes were identified in fetal bovine serum (FBS) [68], indicating the potential need for ghrelin stabilization even in cell culture-based assays. Indeed, it has been suggested that inherent instability of ghrelin in serum may lead to the significant underestimation of active/total ghrelin ratios, especially in the earlier reports, where no protection procedures were employed [71].
4. Ghrelin: Functionality Profile
Ghrelin has emerged as a pivotal factor in numerous physiological functions (Figure 2). The functional scope encompasses the growth hormone (GH) secretion; stimulation of the appetite and food intake; maintenance of glucose homeostasis; modulation of gastric secretion and gastrointestinal motility; improvement of the gut barrier function and circulation, and exerting immunomodulatory effects by downregulation of pro-inflammatory and upregulation of anti-inflammatory cytokines [1,72,73]. In addition, ghrelin enhances neuroprotection, contributes to cardiovascular functions such as lowering blood pressure and enhancing coronary blood flow, influences sleep/wake rhythms, increases expression of anti-apoptotic BCL-2 in lung cells, enhances renal blood flow and promotes the glomerular filtration in the kidney, and stimulates osteoblast proliferation and bone formation [74,75,76,77,78,79].
4.1. Ghrelin Serves as a “Hunger Hormone” Regulating Food Intake and Obesity
Ghrelin earned its nickname as the “hunger hormone” because it stimulates appetite by sending signals to the brain to indicate that it’s time to eat, thereby increasing food intake and promoting fat storage [80]. Ghrelin levels in serum increase before meals and decrease afterward [7,13]. Ghrelin plays a role in short-term food intake regulation and long-term body weight control by reducing fat utilization [81]. This effect on feeding is facilitated through GHSR1a, evidenced by the absence of its orexigenic impact in knockout mice [81].
By regulating appetite ghrelin acts as a key regulator of energy homeostasis integrating signals from peripheral nutritional status to the central nervous system. Specifically, ghrelin promotes increased food consumption and fat storage while potentially slowing overall metabolic rate and reducing the body’s capacity to burn fat, which helps conserve energy during periods of fasting or low nutrient availability [82]. In terms of energy balance, ghrelin contributes to efficient metabolic adaptations by modulating energy expenditure; for instance, it can induce changes that favor energy conservation, such as altering thermogenesis or substrate utilization in tissues like adipose and muscle. It also stimulates the release of growth hormone from the pituitary gland, which indirectly supports metabolic processes like protein synthesis, lipolysis, and glucose regulation [83]. Regarding glucose metabolism, ghrelin influences insulin secretion and sensitivity, often promoting a state that maintains blood sugar levels during fasting, though chronic elevations may contribute to insulin resistance in certain contexts. Additionally, ghrelin exerts control over lipid metabolism by regulating central and peripheral pathways, including the promotion of lipogenesis in the liver and adipose tissue, which aids in energy storage [84]. Emerging research suggests it may also play a protective role in preventing excessive obesity and insulin resistance during growth phases, such as in catch-up growth scenarios, by balancing energy metabolism with anabolic processes [85]. However, studies on ghrelin knockout models indicate that while it is important, its role in appetite and metabolism may be somewhat redundant, as animals lacking ghrelin do not always show significant reductions in food intake or metabolic disruptions [86]. Overall, ghrelin’s metabolic effects are context-dependent, varying with factors like nutritional status, circadian rhythms, and interactions with other hormones such as leptin.
4.2. Ghrelin is a Ligand for the Growth Hormone Secretagogue Receptor 1a (GHSR1a)
Ghrelin conveys its biological activity by binding to homo- and heterodimers of the growth hormone secretagogue receptor 1a (GHSR1a) (AG) and to yet-to-be-identified receptors (DAG) [1,57]. GHSR1a exhibits widespread distribution across tissues and organs, including the lung (especially in alveolar macrophages), kidney, heart, liver, intestine, and adipose tissue [87,88]. Notable locations also include the ventromedial and arcuate nuclei of the hypothalamus that govern feeding and body weight homeostasis [89]. Furthermore, a variety of immune cells, including monocytes, dendritic cells, B and T cells, as well as neutrophils express GHSR1a on their surface [88,90].
Numerous studies indicate that an acyl group on Ser3 is essential for ghrelin’s biological activity through GHSR1a. The position of the octanoylated Ser is fundamental: moving the acyl group to Ser2 retains partial activity, while C8:0 at Ser6 or Ser18 leads to the activity reduction. The fatty acid chain length also impacts the receptor recognition, as maximal activity is retained by C10:0 Ser3, C12:0 Ser3, and C16:0 Ser3 ghrelin variants, but is decreased in the case of the shorter C4:0 Ser3 or C2:0 Ser3 fatty acid modifications [39,91]. While substituting Ser3 with Trp3 preserves ghrelin activity, replacing it with aliphatic amino acids (Val, Leu, Ile) reduces it. The ester bond at Ser3 can be substituted with thioester or ether without affecting activity, highlighting the flexibility in chemical modifications [39]. The N-terminal positive charge and Phe4 are essential for activity and GHSR1a recognition. The minimal active fragment required for activation of GHSR1a is the N-terminal pentapeptide, including C8:0 Ser3 [38]. Amidation of the C-terminus enhances, whereas N-acylation diminishes activity [91,92]. Both acyl and des-acyl ghrelin exhibit a short α-helix conformation when bound to lipids [93]. The minimal core sequence necessary for GHSR1a activation covers the N-terminal 1GSS(Octanoyl)FL5 sequence with indispensable octanoylated Ser3 in the middle (Figure 1).
5. GHSR1a Signaling Pathways
GHSR1a belongs to the A family of G-protein coupled receptors (GPCRs), characterized by 7-transmembrane (7–TM) regions. Tucked away in cell membranes, these receptors are selective for a wide range of ligands, from sophisticated proteins to small molecule ligands like adrenaline. Classically GHSR1a transduces the signal via GHSR1a-Gαq–dependent pathway (Figure 3A), which leads to the increase in intracellular Ca2+, mediated by phospholipase C (PLC) activation and release of inositol 1,4,5-trisphosphate (IP3) and downstream activation of protein kinase C (PKC), Ca2+/calmodulindependent protein kinase-IIa (CamKII), and 5′ AMP-activated protein kinase (AMPK) [94,95,96]. In another G protein-related pathway( Figure 3C), GHSR1a –Gαi/o activates phosphoinositide 3-kinases (PI3K) to induce the activation of protein kinase A (PKA), PKCε, and serine/threonine protein kinase (AKT) [97,98]. Alternatively, G-protein independent signaling is based on the signal relay via GSHR1a-β-arrestin complex (Figure 3B), leading to the increase in receptor internalization, accompanied by extracellular signal-regulated kinases (ERK1/2) and AKT activation [99,100]. It was reported that recruitment of Gαq/11 is required for the regulation of food intake [101], while Gi2 is necessary for the regulation of insulin release [102].
In recent years, significant progress has been made, and structures of GHSR1a with several ligands have been determined. Recognition of ghrelin and the antagonist Compound 21 involves the binding pocket, which bifurcates into two cavities separated by a salt bridge formed between TM3 E1243.33 and TM6 R2836.55, with cavity I created between helices TM6 and TM7 of the receptor dedicated to the binding of the ghrelin N-termini and cavity II created between helices TM4 and TM5 recognizing the octanoylated side chain of Ser3 [103,104]. Further, the recent structure of GHSR1a with anamorelin, its small-molecule agonist, currently investigated for the cancer-related cachexia and anorexia treatment [105,106] confirmed this mode of binding [107]. Notably, the study combined cryo-EM structural analysis and in-cell functional assays to elucidate biased signaling of the GHSR1a. The repertoire of G-protein families recruited depends on ligand concentration and time; different ligands (agonists/antagonists/reverse agonists) stabilize distinct receptor conformations, producing a characteristic G-protein/β-arrestin fingerprint [107,108]. These findings provide the structural explanation for the GHSR1a biased signaling and indicate that selected ligand combinations might be chosen for the personalized treatment even in the case of SNP variations in the receptor.
As progress in the clinical development of drugs targeting GHSR1a is relatively slow, new data on the pharmacology of GHSR1a ligands are needed to design new, more selective ligands with predictable biological activity. The pharmacology of GHSR1a is highly complex, involving G protein-dependent and -independent signaling pathways and high constitutive activity. The functional selectivity and signaling bias of many GHSR1a-specific ligands is still not well described. Activity of several peptides was investigated in aspect of constitutive signaling, ligand-directed downstream GHSR1a signaling, functional selectivity, and signaling bias [109]. Biased ligand binding to GHSRs simultaneously stabilizes the receptor in a conformation that is able to selectively activate specific signaling pathways (Figure 3). Ghrelin can stabilize GHSRs in specific conformations that favor G-protein activation (Gαq/Gαi/o) or β-arrestin recruitment [110]. Interestingly, the activity of peptides (inverse agonist KwFwLL and agonist AwFwLL) depends on key residues in transmembrane helices III and IV. Such mutations change the efficacy of KwFwLL from full inverse agonism in the WT receptor to partial agonism in the mutated receptor. In contrast, ghrelin stabilizes a different conformation that is able to recruit Gαq, Gαi/o, and β-arrestin but not Gαs. These findings are in line with knowledge that the GHSR conformation induced by ghrelin in the presence of β-arrestin differs from ghrelin-induced conformation in the presence of Gαq [111]. Indeed, GHSR exhibits distinct conformations when activated by different ligands. There is a growing collection of agonists and antagonists for GHSR demonstrating a spectrum of activities. Ramirez et al. examined a group of biologically active substances including: ghrelin, Ibutamoren (MK-0677), L692,585, and [D-Lys3]-growth hormone–releasing peptide-6 (DLys3-GHRP-6), JMV2959, and [D-Arg1,D-Phe5,D-Trp7, 9,Leu11]-substance P (SP-analog) [112]. DLys3-GHRP-6 behaved as a partial antagonist with a strong bias toward GHSR1a–β-arrestin signaling, whereas JMV2959 acted as a full unbiased GHSR1a antagonist. Moreover, the SP-analog at high concentrations behaved as an inverse agonist increasing G-protein–dependent signaling, whereas, at low concentrations, the SP-analog attenuated β-arrestin–dependent signaling (Table 1).
Liver-expressed antimicrobial peptide 2 (LEAP-2) was first described as an antimicrobial peptide expressed in the liver [113]. Subsequent investigation revealed that LEAP-2 is an endogenous antagonist of GHSR1a blocking the action of ghrelin, by competing at the ghrelin-binding site [114]. LEAP-2 increases whereas ghrelin decreases in obesity, and LEAP-2 is implicated in obesity-related disorders including polycystic ovary syndrome and non-alcoholic fatty liver disease [115]. Recent studies have revealed that ghrelin attenuates obesity-induced myocardial injury through various signaling axes [116,117]. It has been shown that knockdown of LEAP-2 relieved hyperlipidemia, inflammation, and myocardial injury in obese mice by polarizing macrophages toward the M2 phenotype [118]. LEAP-2 is being investigated as a potential therapeutic target for obesity and related metabolic diseases, as it can impact food intake and body weight [119]. Recent studies have focused on developing LEAP-2 analogs with potential clinical application [120]. Despite complicated landscape of ghrelin signaling pathways, the understanding of the receptor structure and biased signaling is progressing. As of now only anamorelin, a small-molecule synthetic agonist of GHSR1a has found a limited clinical use in cancer patients cachexia treatment. Fo the summary on the progress in the clinical trials of ghrelin receptor ligands, please refer to the recent review by Bukhari [121].
Figure 3.
Signaling pathways associated with the ghrelin GHS-R1a receptor. Upon activation by ghrelin, GHSR1a engages Gαq/11 (A), Gαi/o (C) or β-arrestin (B) signaling. Each G-protein/β-arrestin complex is linked with unique intracellular activation pathway and specific physiological effects. p-: phosphorylated; AMPK: adenosine-monophosphate-activated protein kinase; DiAcGly: diacylglycerol; GTP: guanosine triphosphate; IP3: inositol 1,4,5-trisphosphate; PKC: Protein Kinase C; CamKII: Ca2+/calmodulindependent protein kinase-IIa; PLC: phospholipase C; PI3K: phosphoinositide 3-kinase; PKA: protein kinase A; AKT: serine/threonine protein kinase; ERK: extracellular signal-regulated kinases. Created with BioRender.com.
Figure 3.
Signaling pathways associated with the ghrelin GHS-R1a receptor. Upon activation by ghrelin, GHSR1a engages Gαq/11 (A), Gαi/o (C) or β-arrestin (B) signaling. Each G-protein/β-arrestin complex is linked with unique intracellular activation pathway and specific physiological effects. p-: phosphorylated; AMPK: adenosine-monophosphate-activated protein kinase; DiAcGly: diacylglycerol; GTP: guanosine triphosphate; IP3: inositol 1,4,5-trisphosphate; PKC: Protein Kinase C; CamKII: Ca2+/calmodulindependent protein kinase-IIa; PLC: phospholipase C; PI3K: phosphoinositide 3-kinase; PKA: protein kinase A; AKT: serine/threonine protein kinase; ERK: extracellular signal-regulated kinases. Created with BioRender.com.
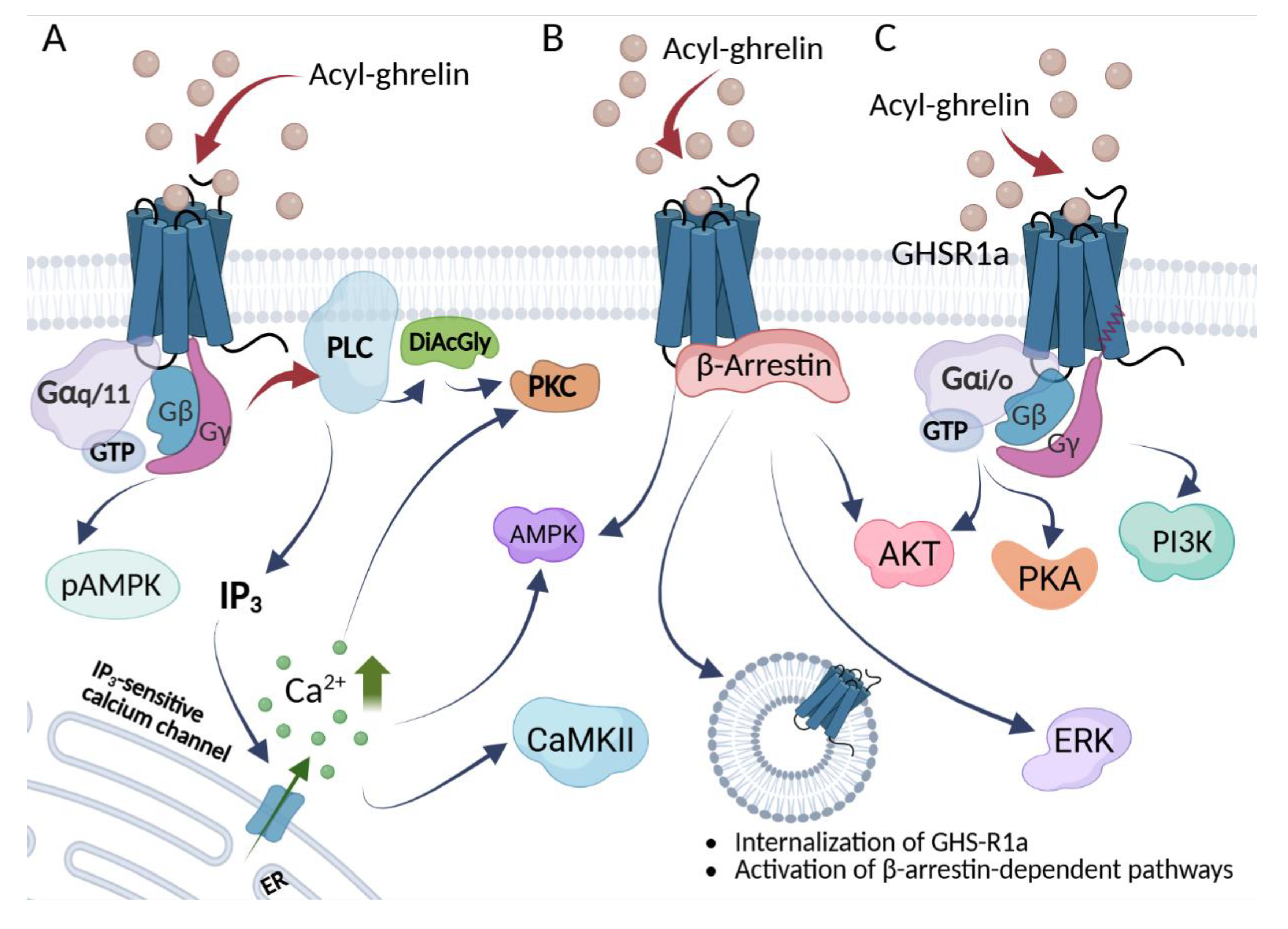

Table 1.
Selected ligands of the human growth hormone secretagogue receptor (GHSR1a).
| Ligand | Type | Activity | Signaling Pathways | Selected refs | |||
| Ca2+ mobilization | β-arrestin | GHSR1a intern. |
ERK Phosph. |
||||
| Ghrelin (human, acylated) | Endogenous peptide | Full agonist (canonical) | + | + | + | + | [1,112] |
| Des--acyl ghrelin (DAG) | Endogenous peptide (des--acyl) | Weak/low--potency agonist in vitro; often functionally GHSR1a--independent in vivo | [38] | ||||
| Mini-ghrelins (1–15, 1–14, 1–11) | Endogenous peptide fragments | Competitive antagonists | [37,38] | ||||
| LEAP--2 | Endogenous peptide/protein | Competitive antagonist | [114] | ||||
| Anamorelin | Small--molecule | Potent agonist | [107,122] | ||||
| Ibutamoren (MK--677) | Small--molecule | Potent, selective, orally active agonist | + | + | + | + | [112,123,124] |
| + | + | + | + | ||||
| L--692,585 | Small--molecule | Agonist | [112,125,126] | ||||
| JMV2959 | Small--molecule | Unbiased antagonist; bias-inverse agonist | +/- | Basal - | 0 | [112,127] | |
| Compound 21 (C21) | Small--molecule | Neutral antagonist | [103] | ||||
| PF-5190457 | Small--molecule | Orally active inverse agonist | [128,129] | ||||
| Basal - | 0 | ||||||
| [D--Lys3]--GHRP--6 | Peptide analog | Preferentially β-arrestin pathway blocker; bias-inverse agonist | [112,130,131] | ||||
| Substance P analog (D--Arg11,D--Phe55,D--Trp77,99, Leu1111--SP) |
Peptide analog | Inverse agonist at higher concentrations; attenuates β--arrestin at low concentrations | Basal - | Basal - | [112,132] | ||
| KwFwLL | Peptidomimetic | Inverse agonist | [133] | ||||
| AwFwLL | Peptidomimetic | Agonist | [133] | ||||
GHSR1a intern.: internalization of the receptor; ERK Phosph.: Extracellular signal-regulated kinase phosphorylation; Activity symbols: Basal -: reduction in the constitutive (basal) receptor activity; +: increase in signal; 0: no effect; +/- weak increase in signal.
6. Ghrelin as an Anti-Inflammatory Agent
Ghrelin is recognized for its broad anti-inflammatory effects across multiple physiological systems. In the central nervous system, it reduces neuronal damage following subarachnoid hemorrhage in rats and mitigates neuroinflammation in Alzheimer’s disease models [134]. Ghrelin’s immunoregulatory role is exemplified by its suppression of pro-inflammatory cytokine production. Notably, in aged septic rats, both ghrelin and GH attenuate immunosuppression via vagus nerve–dependent inhibition of transforming growth factor-beta (TGF-β) production (105).
In the gastrointestinal tract, ghrelin demonstrates therapeutic potential in relieving colitis, while also exerting beneficial effects in metabolic disorders such as type 2 diabetes [135,136]. Beyond these roles, ghrelin influences skeletal muscle, cardiovascular, and respiratory function [137,138].
In a murine model of elastase-induced emphysema, ghrelin treatment attenuated pulmonary inflammation, promoted macrophage polarization toward the M2 phenotype, reduced collagen deposition, and increased elastic fiber content, collectively contributing to improved alveolar architecture and lung function. Additionally, ghrelin improved cardiovascular dysfunction and increased both lean and total body mass [138].
Consistently, ghrelin has been shown to reduce inflammation and disease severity in numerous conditions, including sepsis, inflammatory bowel disease, arthritis, pancreatitis, obesity, autoimmune encephalomyelitis, and diabetic nephropathy [134,139,140,141]. In turn, this underscores the evident link between ghrelin and chronic diseases . Studies from many laboratories collectively provide robust evidence that ghrelin exerts anti-inflammatory effects in human monocytes and macrophages by reducing pro-inflammatory cytokine production, inhibiting inflammatory signaling pathways, promoting anti-inflammatory macrophage polarization, and decreasing adhesion and oxidative stress [142,143,144,145].
Ghrelin hinders the generation of pro-inflammatory cytokines from monocytes, T-cells, and macrophages [142]. It also hampers leptin-triggered expression of pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-, while boosting the production of anti-inflammatory cytokines like TGF-β and IL-10 [73,143]. HMGB1 (High Mobility Group Box 1) is a nuclear protein produced by the HMGB1 gene which stabilizes chromosome structure and regulates gene expression [146] and when translocated to the cytoplasm induces autophagy [147]. In immune cells, HMGB1 is directed for secretion [148] and acts as an alarmin, recognized by RAGE and TLR-family receptors [149]. It is secreted by diverse immune cells, including monocytes, macrophages, and dendritic cells, prompting the release of pro-inflammatory cytokines like TNF-, IL-1, IL-6, and IL-8, thus intensifying inflammation [150,151]. Ghrelin treatment impedes HMGB1 translocation from the nucleus to the cytoplasm, reducing its secretion and thereby dampening inflammation [142]. The delayed release of ghrelin by macrophages when stimulated by substances like LPS, IL-1, and TNF-may act as a regulatory mechanism to avert excessive inflammation [152].
Remarkably, GHSR deletion mutants (Ghsrmutant) of macrophages and microglia reduce their inflammatory responses to fructose, indicating that GHSR mediates fructose-induced inflammation. Furthermore, GHSR regulates fructose transport and metabolism, and mediates fructose-induced inflammatory activation through CREB–AKT–NF-κB and p38 MAPK signaling pathways [153]. These findings reveal that the nutrient-sensing receptor GHSR plays a crucial role in fructose-mediated inflammatory activation. Interestingly, at the molecular level, GHSR metabolically programs macrophage polarization through PKA–CREB–IRS2–AKT2 signaling pathway [154], an axis which is not characteristic for other cell types. Insulin signaling is a master regulatory pathway of metabolism, and is involved in macrophage polarization [155]. Insulin receptor deficiency in macrophages protects against inflammation [156,157,158], and IRS2 deletion promotes IL-4-induced M2 macrophage polarization [159]. Thus, nutrient-sensing ghrelin signaling is linked to immune regulation and meta-inflammation.
Obesity-related chronic inflammation, or meta-inflammation, has been linked to a wide range of metabolic dysfunctions such as insulin resistance [160,161], fatty acid dysregulation [162,163] and non-alcoholic fatty liver disease (NAFLD) [164,165]. Macrophages are among the first responders of inflammation in response to metabolic dysregulation [166]. Meta-inflammation through cytokines and other inflammatory mediators promotes macrophage recruitment and activation in tissues, leading to deleterious effects in, for example, adipose tissue and liver [167,168,169]. Macrophage diversity is described by the outdated M1/M2 model, created more than 20 years ago, which distinguishes between M1 or “classically” activated macrophages; and M2 or “alternatively” activated macrophages, based on the effect of in vitro macrophage stimulation with type 1 or type 2 cytokines [170]. In the more recent version, the “M1-like” phenotype is usually described as pro-inflammatory and is induced by Toll-like receptor (TLR) ligands and type 1 cytokines, namely IFN-γ and TNF-α; ‘M2-like’ macrophages, having anti-inflammatory properties, are activated by IL-4 or IL-13 and produce TGF-β. This nomenclature, although oversimplified, is widely used today [171,172]. Significantly, a considerable heterogeneity in morphology, function and expression of cell surface markers has been observed in resident macrophages from different organs [173]; moreover, co-expression of M1 as well as M2 genes has been described in subgroups of macrophages from most tumor types [174]. Therefore, it is now believed that macrophages form a continuous spectrum of phenotypes during polarization [175,176]. Single-cell RNA sequencing technology (scRNA-seq) confirms the multidimensional diversity of macrophage immunophenotype and function [177]. Ghrelin is known to stimulate appetite and promote obesity/insulin resistance via the G protein-coupled receptor (GPCR) [47,178,179,180,181]. GHSR1a expression is very high in macrophages and monocytes [142,182]. In mouse peritoneal macrophages, expression reaches 60% relative to the hypothalamus, the tissue with the highest expression [182]. Global ablation of GHSR1a promotes anti-inflammatory change in peritoneal macrophages and adipose tissue macrophages in aging mice [182]. Global GHSR1a ablation has also been shown to alleviate adipose tissue inflammation and non-alcoholic steatohepatitis (NASH) induced by high fructose corn syrup (HFCS) [183]. Adipose tissue macrophages of such mice have reduced expression of pro-inflammatory markers (M1), including monocyte chemoattractant protein-1 (MCP-1), TNF-α and inducible nitric oxide synthase (iNOS), while the anti-inflammatory markers arginase-1 (Arg-1) and macrophage galactose-type lectin-1 (Mgl-1) were increased [184]. In vitro studies have also shown that antagonist or knock-down (siRNA) of GHSR1a decreases the expression of proinflammatory cytokine genes in the LPS-stimulated macrophage cell line RAW264.7 [182,183]. These observations suggest that GHSR1a has a cell-autonomous effect in macrophages, and that GHSR1a probably plays a key role in macrophage programming. In myeloid-specific GHSR1a knockout mice tissue analysis showed significantly decreased monocyte/macrophage infiltration, pro-inflammatory activation, and lipid accumulation. Ex vivo, GHSR1a-deficient macrophages were protected against palmitic acid (PA)- or LPS-induced pro-inflammatory polarization, showing reduced glycolysis, increased fatty acid oxidation, and decreased NF-κB nuclear translocation [154]. As a G protein-coupled receptor, GHSR has a high ligand-independent constitutive activity of ~50% of its maximal activity [132,185,186]. Thus, the effect of GHSR1a in macrophages may be mediated by both ligand-dependent and independent (constitutive) mechanisms. Global deletion of GHSR1a does not alter serum ghrelin levels compared to wild-type mice under both fed and fasted conditions [179]. It is likely that the effect of GHSR1a in macrophages is mediated by both ghrelin-dependent and -independent mechanisms. Although evidence in primary human macrophages remains limited, it is speculated that even without endogenous ghrelin, GHSR1a may modulate macrophages by altering downstream signaling pathways. Most evidence comes from animal models (e.g., murine macrophages) or indirect studies in human cell lines (e.g., THP-1 macrophages), which may not reflect primary human macrophage behavior. There is no direct evidence in human macrophages suggesting that GHSR1a’s constitutive activity promotes pro-inflammatory responses. Constitutive activity alone does not appear to drive pathways like NF-κB activation or pro-inflammatory cytokine release in human macrophages [187,188] [187]. Thus, we speculate that the potential inflammatory outcome of GHSR1a’s constitutive activity may depend on the macrophage’s activation state (e.g., resting, M1, or M2) and the microenvironment. For instance, in resting human macrophages, constitutive activity likely maintains an anti-inflammatory tone, while in highly inflamed states, its impact might be overshadowed by dominant pro-inflammatory signals (e.g., LPS-induced TLR4 activation).
7. The Neuroimmune Connection—Ghrelin’s Role in the Nervous System
The hypothalamus is a small brain region located just below the thalamus, responsible for central regulation of appetite and energy homeostasis. Structurally, the hypothalamus consists of several distinct nuclei: the arcuate nucleus (ARC), paraventricular nucleus (PVN), lateral hypothalamic area (LHA), ventromedial nucleus (VMN), and dorsomedial nucleus (DMN). The ARC lies adjacent to the median eminence, a circumventricular organ characterized by fenestrated capillaries and an atypical blood–brain barrier (BBB). This proximity provides ARC neurons with relatively direct access to circulating hormones and nutrients without crossing a typical BBB. Owing to this unique anatomical feature, the ARC is considered a principal hypothalamic site for sensing peripheral metabolic signals. Two major neuronal populations are present in the ARC: (i) neurons co-expressing orexigenic (appetite-stimulating) neuropeptides, including neuropeptide Y (NPY) and agouti-related peptide (AgRP); and (ii) neurons expressing anorexigenic (appetite-limiting) neuropeptides, including proopiomelanocortin (POMC) and cocaine- and amphetamine-regulated transcript (CART). These populations serve as first-order integrators through which peripheral metabolic signals affecting appetite are relayed [189].
As in other organs, the function of ghrelin in the central nervous system (CNS) is mediated by activation of the GHSR1a. Its expression has been identified in the pituitary gland, ARC, VMN, PVN and, to some extent, in the DMN [190,191,192,193]. Ghrelin activity in hypothalamic neurons is part of the brain’s fatty-acid–sensing mechanism. Binding of ghrelin to GHSR1a activates PLC–IP3/DiAcGly–PKC signaling and increases intracellular calcium levels. This, in turn, activates calcium/calmodulin-dependent protein kinase 2 (CamKII), which phosphorylates AMP-activated protein kinase (AMPK) and promotes formation of a stable AMPK/CaMKK2 complex. This complex increases phosphorylation of acetyl-CoA carboxylase (ACC), lowering malonyl-CoA and thereby disinhibiting carnitine palmitoyltransferase-1 (CPT1A and CPT1C) [194,195,196,197]. CPT1A shuttles acylcarnitines, allowing long-chain fatty acids to cross the mitochondrial membrane for subsequent β-oxidation [198,199]. Enhanced fatty-acid metabolism can elevate mitochondrial reactive oxygen species (ROS) and upregulate the free-radical-scavenging uncoupling protein-2 (UCP2), which contributes to activation of NPY/AgRP neurons and stimulation of ghrelin-induced food intake [200]. The CPT1C isoform increases ceramide levels and upregulates NPY and AgRP while downregulating POMC expression, thereby increasing NPY/AgRP and decreasing POMC activity in the ARC, effectively stimulating appetite. In addition, ghrelin modulates NPY-related neurons in the PVN and ARC, increasing GABAergic suppression of POMC neurons, which further promotes appetite [201,202].
Beyond its classical roles in appetite regulation and pituitary signaling, ghrelin exhibits potent anti-inflammatory and neuroprotective properties [143]. Preclinical studies demonstrate that exogenous ghrelin maintains the blood–brain barrier (BBB) integrity, reduces oxidative damage, and limits neuronal apoptosis after traumatic brain injury (TBI) and ischemia–reperfusion injury (IRI) [203,204,205,206]. These effects are linked to modulation of fibroblast growth factor signaling [207] and activation of the PI3K/Akt pathway, which promotes neuronal survival via regulation of GSK-3β and Bcl-2 [208,209,210].
Ghrelin has been implicated in several mental disorders, including anorexia nervosa and bulimia—via appetite regulation—as well as depression, anxiety, and schizophrenia [211,212]. The ghrelin gene polymorphism Leu72Met has been associated with depression, but not with panic disorder, in patients [213].
Depression is a multifactorial disorder with genetic, neuroendocrine, immune, and metabolic influences [214,215]. Beyond monoamine deficiency [216], hypothalamus-pituitary-adrenal (HPA) axis dysregulation [217], excitatory/inhibitory imbalance [218], and neuroinflammation [219], growing evidence implicates ghrelin in mood regulation.
Ghrelin may influence depression in part through normalization of the hypothalamic–pituitary–adrenal (HPA) axis, which is dysregulated—often hyperactive—in patients with major depressive symptoms [220,221]. It has been reported that ghrelin administration increases cortisol and growth hormone (GH), with only a weak trend toward mood improvement in male patients [222], and that ghrelin can alleviate anxiety after short-term stress by activating the HPA axis [223]. In line with these findings, rat studies using ghrelin O-acyltransferase (GOAT) inhibitors—thus reducing active (acyl-)ghrelin—showed dampened HPA activity, suggesting that active ghrelin modulates HPA tone and may affect depressive phenotypes [224]. In patients, ghrelin administration alters sleep architecture, increasing non-REM and reducing REM sleep [222,225]. In the same vein are the recent results on the efficacy of the novel butyrylcholinesterase inhibitor (BChEI) in the Flinders Sensitive Line rat model of major depressive disorder. Application of BChEI improved behavioral, cognitive and reward-related parameters of treated animals in a GHSR1a-dependent manner, likely by increased protection of acyl-ghrelin from butyrylcholinesterase in serum [226].
Ghrelin also enhances dopaminergic signaling, and dysregulation of the dopamine system is linked to depressive symptoms [227]. Specifically, ghrelin activates ventral tegmental area (VTA) dopamine neurons, a mechanism also associated with appetite stimulation [228,229]. In mice, depressive-like behavior was reduced by ghrelin, an effect attributed to GHSR1a-dependent increases in dopamine levels and promotion of dopaminergic neuronal responses and synapse formation [230,231]; these effects were at least partially reversed by administration of a ghrelin receptor antagonist [232]. Preclinical evidence suggests that ghrelin inhibits serotonin synthesis via nitric oxide (NO)-dependent suppression of tryptophan hydroxylase [233], a mechanism that may promote depressive phenotypes. Conversely, ghrelin enhances dopaminergic and orexin signaling [234,235], supporting reward processing, stress resilience, and emotional regulation. Recently it was demonstrated that ghrelin-independent, constitutive activity of GHSR1a is required for the reversal (inhibition to excitation) of dopamine D2 receptor signaling in the spinal defecation center [236].
Stress elevates circulating ghrelin, modulating HPA axis activity and glucocorticoid release [237,238], though associations with cortisol in clinical studies remain inconsistent [239,240]. At the neuronal level, ghrelin stimulates hippocampal neurogenesis and synaptic plasticity through PI3K/Akt–MAPK pathways, upregulating brain-derived neurotrophic factor (BDNF) [241,242]. Ligand-free (apo) GHSR1a can heteromerize with dopamine receptor D1 (DRD1), shaping reward-related plasticity [243,244].
Beyond its potential as a therapeutic target, ghrelin has been proposed as a biomarker to monitor antidepressant treatment response [245] and to aid in differential diagnosis between depression and bipolar disorder [246,247].
Disruption of ghrelin signaling is increasingly implicated in Alzheimer’s disease (AD). While total plasma ghrelin is often unchanged [248,249], some studies report elevated circulating levels in AD and prodromal AD, inversely correlating with cognitive performance [249,250]. This paradox may reflect reduced local ghrelin availability, as supported by decreased temporal lobe ghrelin mRNA [251]. Central to ghrelin signaling disruption in AD are alterations in ghrelin receptor (GHSR) expression and regulation. GHSR mRNA is reduced in leukocytes and temporal lobes [250,251], yet hippocampal protein levels are increased, where Aβ directly binds and inhibits GHSR, impairing signaling [252]. GHSR1b, a splice variant that may suppress GHSR function, is also elevated in AD brains [251,253]. Additional modulation may involve liver-expressed antimicrobial peptide 2 (LEAP-2), which could exacerbate ghrelin resistance due to its association with obesity and metabolic syndrome, both recognized AD risk factors [254,255]. In recent years, ghrelin analogs were indeed demonstrated to have neuroprotective properties in vitro and in the AD triple transgenic mouse model in vivo [256,257]. Ghrelin activation may also be altered through ghrelin O-acyltransferase (GOAT). Increased MBOAT4 (GOAT) mRNA has been observed in leukocytes of AD patients [250], though its functional significance outside the gut remains unclear. Evidence for circulating “free” GOAT [30] raises additional questions about peripheral ghrelin activation in AD. Importantly, GOAT activity requires coenzyme A–activated fatty acids, especially medium-chain fatty acids (MCFAs), and dysregulated fatty acid metabolism in AD may further modulate ghrelin activation [258].
Ghrelin signaling plays a central role in alcohol use disorder (AUD) [259]. In the brain, ghrelin affects reward-related regions such as the amygdala [260], Edinger–Westphal nucleus [261], laterodorsal tegmental area, and lateral hypothalamus [262], modulating dopaminergic activity and reinforcing alcohol intake. Notably, chronic alcohol exposure upregulates GHSR expression within these structures [263], while ghrelin enhances alcohol-associated cues and neurochemical responses [264]. Preclinical studies show that ghrelin promotes alcohol intake, reward, and relapse primarily by stimulating mesolimbic dopaminergic VTA–nucleus accumbens (VTA–NAc) circuit [265]. Conversely, genetic deletion or pharmacological inhibition of GHSR (via antagonists or inverse agonists) consistently attenuates alcohol consumption, reduces motivational drive, and relapse-like behaviors across animal models [266,267]. Endogenous ghrelin dynamics appear context-dependent; acute alcohol administration reduces circulating ghrelin in male rats [268], whereas chronic exposure may lead to elevated levels [259]. Additional modulators of the ghrelin pathway, such as des-acyl ghrelin (DAG) and LEAP-2, may further influence addiction-related behaviors, though their roles remain less defined [269].
Beyond alcohol, ghrelin signaling broadly modulates the reinforcing properties of other addictive substances. In rodents, ghrelin enhances locomotor activity, reward, and conditioned place preference (CPP) associated with psychostimulants (cocaine, amphetamines, nicotine), opioids (morphine, fentanyl, oxycodone), and cannabinoids [270,271,272,273,274,275]. Conversely, pharmacological inhibition of GHSR attenuates drug-induced dopaminergic transmission, CPP, self-administration, and reinstatement across these substance classes [267]. Complementary human studies indicate associations between GHSR gene polymorphisms and substance dependence, as well as positive correlations between circulating ghrelin levels and craving intensity [129].
8. Conclusions
Ghrelin, an extraordinary molecule with its unique processing system remains a focus of the ongoing research and it is now known that its role extends far beyond its metabolic functions, exerting neuroprotective, anti-inflammatory, and neuromodulatory effects. Disrupted ghrelin signaling contributes to Alzheimer’s disease, addiction, and depression, where it regulates neuronal survival, reward pathways, and mood. As ghrelin-GHSR1a structure was recently solved and provided structural foundations for long-reported biased signaling, the precise pharmaceutical targeting of the ghrelin system may finally become feasible in the near future. Together with the functional GOAT model, the development of novel GOAT inhibitors and better understanding of the ghrelin post-translational processing, these findings position ghrelin as a promising therapeutic target not only in the obesity treatment in the post–GLP-1 receptor agonist era, but also in the management of inflammatory diseases, neuropsychiatric disorders and addiction-related therapy.
Author Contributions
HP writing—original draft preparation and reviewing, figure preparation; KG: writing—original draft preparation and reviewing; TK writing—original draft preparation, reviewing and editing, figure preparation, conceptualization, supervision, funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by SONATA Bis project granted by National Centre of Science, Poland (UMO-2016/22/E/NZ5/00332) (TK).
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
All figures were created in BioRender. Kantyka, T. (2025) https://BioRender.com/0inw92w; https://.
BioRender.com/6s2oynh
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| GHSR1a | growth hormone secretagogue receptor 1a |
| GOAT | ghrelin O-acyltransferase |
| LEAP-2 | Liver-expressed antimicrobial peptide 2 |
| GPCR | G protein-coupled receptor |
| GCF | gingival crevicular fluid |
| RIA | radioimmunoassay |
| CoA | Coenzyme A |
| ER | endoplasmic reticulum |
| PC1/3 | prohormone convertase 1/3 |
| PC | prostate cancer |
| APC | activated protein |
| MBOAT | membrane-bound-O-acyltransferase |
| AG | acyl ghrelin |
| DAG | des-acyl ghrelin |
| ERK1/2 | extracellular signal-regulated kinase ½ |
| PI3K | Phosphoinositide 3-kinase |
| AKT | Protein kinase B; serine/threonine protein kinase |
| APT1 | Acyl-Protein Thioesterase 1/Lysophospholipase |
| PMSF | phenylmethylsulfonyl fluoride |
| EDTA | Ethylenediaminetetraacetic acid |
| AEBSF | 4-(2-aminoethyl)benzenesulfonyl fluoride hydrochloride |
| MAFP | methoxy arachidonyl fluorophosphonate |
| FBS | fetal bovine serum |
| GH | growth hormone |
| PLC | phospholipase C |
| IP3 | inositol 1,4,5-trisphosphate |
| PKC | protein kinase C |
| DiAcGly | Diacylglycerol |
| CamKII | Ca2+/calmodulindependent protein kinase-IIa |
| AMPK | 5′ AMP-activated protein kinase |
| PKA | protein kinase A |
| PKCε | protein kinase Cε |
| TM | transmembrane helix |
| SNP | single-nucleotide polymorphism |
| DLys3-GHRP-6; D-Lys3 | [D-Lys3]-growth hormone–releasing peptide-6 |
| SP-analog | [D-Arg1,D-Phe5,D-Trp7, 9,Leu11]-substance P |
| HMGB1 | High Mobility Group Box 1 |
| NAFLD | non-alcoholic fatty liver disease |
| TLR | Toll-like receptor |
| scRNA-seq | Single-cell RNA sequencing |
| NASH | non-alcoholic steatohepatitis |
| MCP-1 | monocyte chemoattractant protein-1 |
| iNOS | inducible nitric oxide synthase |
| Arg-1 | arginase-1 |
| Mgl-1 | macrophage galactose-type lectin-1 |
| PA | palmitic acid |
| ARC | arcuate nucleus |
| PVN | paraventricular nucleus |
| LHA | lateral hypothalamic area |
| VMN | ventromedial nucleus |
| DMN | dorsomedial nucleus |
| BBB | blood–brain barrier |
| NPY | neuropeptide Y |
| AgRP | agouti-related peptide |
| POMC | proopiomelanocortin |
| CART | cocaine- and amphetamine-regulated transcript |
| CNS | central nervous system |
| ACC | acetyl-CoA carboxylase |
| CPT1 | carnitine palmitoyltransferase-1 |
| ROS | reactive oxygen species |
| UCP2 | uncoupling protein-2 |
| TBI | traumatic brain injury |
| IRI | ischemia–reperfusion injury |
| HPA | hypothalamus-pituitary-adrenal |
| BChEI | butyrylcholinesterase inhibitor |
| VTA | ventral tegmental area |
| NO | nitric oxide |
| BDNF | brain-derived neurotrophic factor |
| DRD1 | dopamine receptor D1 |
| AD | Alzheimer’s disease |
| GHSR1b | growth hormone secretagogue receptor 1b |
| AUD | alcohol use disorder |
| NAc | nucleus accumbens |
| CPP | conditioned place preference |
| GLP-1 | Glucagon-like peptide-1 |
References
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 1999, 402, 656–660. [CrossRef]
- Ghelardoni, S.; Carnicelli, V.; Frascarelli, S.; Ronca-Testoni, S.; Zucchi, R. Ghrelin tissue distribution: comparison between gene and protein expression. J Endocrinol Invest 2006, 29, 115–121. [CrossRef]
- Groschl, M.; Topf, H.G.; Bohlender, J.; Zenk, J.; Klussmann, S.; Dotsch, J.; Rascher, W.; Rauh, M. Identification of ghrelin in human saliva: production by the salivary glands and potential role in proliferation of oral keratinocytes. Clin Chem 2005, 51, 997–1006. [CrossRef]
- Ohta, K.; Laborde, N.J.; Kajiya, M.; Shin, J.; Zhu, T.; Thondukolam, A.K.; Min, C.; Kamata, N.; Karimbux, N.Y.; Stashenko, P.; et al. Expression and possible immune-regulatory function of ghrelin in oral epithelium. J Dent Res 2011, 90, 1286–1292. [CrossRef]
- Hosoda, H.; Kangawa, K. Standard sample collections for blood ghrelin measurements. Methods Enzymol 2012, 514, 113–126. [CrossRef]
- Drazen, D.L.; Vahl, T.P.; D’Alessio, D.A.; Seeley, R.J.; Woods, S.C. Effects of a fixed meal pattern on ghrelin secretion: evidence for a learned response independent of nutrient status. Endocrinology 2006, 147, 23–30. [CrossRef]
- Cummings, D.E.; Purnell, J.Q.; Frayo, R.S.; Schmidova, K.; Wisse, B.E.; Weigle, D.S. A preprandial rise in plasma ghrelin levels suggests a role in meal initiation in humans. Diabetes 2001, 50, 1714–1719. [CrossRef]
- Lee, H.M.; Wang, G.; Englander, E.W.; Kojima, M.; Greeley, G.H., Jr. Ghrelin, a new gastrointestinal endocrine peptide that stimulates insulin secretion: enteric distribution, ontogeny, influence of endocrine, and dietary manipulations. Endocrinology 2002, 143, 185–190. [CrossRef]
- Widmayer, P.; Goldschmid, H.; Henkel, H.; Kuper, M.; Konigsrainer, A.; Breer, H. High fat feeding affects the number of GPR120 cells and enteroendocrine cells in the mouse stomach. Front Physiol 2015, 6, 53. [CrossRef]
- Qi, X.; Reed, J.T.; Wang, G.; Han, S.; Englander, E.W.; Greeley, G.H., Jr. Ghrelin secretion is not reduced by increased fat mass during diet-induced obesity. Am J Physiol Regul Integr Comp Physiol 2008, 295, R429–435. [CrossRef]
- Konturek, P.C.; Brzozowski, T.; Pajdo, R.; Nikiforuk, A.; Kwiecien, S.; Harsch, I.; Drozdowicz, D.; Hahn, E.G.; Konturek, S.J. Ghrelin-a new gastroprotective factor in gastric mucosa. J Physiol Pharmacol 2004, 55, 325–336.
- Zhao, T.J.; Sakata, I.; Li, R.L.; Liang, G.; Richardson, J.A.; Brown, M.S.; Goldstein, J.L.; Zigman, J.M. Ghrelin secretion stimulated by beta1-adrenergic receptors in cultured ghrelinoma cells and in fasted mice. Proc Natl Acad Sci U S A 2010, 107, 15868–15873. [CrossRef]
- Williams, D.L.; Cummings, D.E.; Grill, H.J.; Kaplan, J.M. Meal-related ghrelin suppression requires postgastric feedback. Endocrinology 2003, 144, 2765–2767. [CrossRef]
- Pagotto, U.; Gambineri, A.; Vicennati, V.; Heiman, M.L.; Tschop, M.; Pasquali, R. Plasma ghrelin, obesity, and the polycystic ovary syndrome: correlation with insulin resistance and androgen levels. J Clin Endocrinol Metab 2002, 87, 5625–5629. [CrossRef]
- Riis, A.L.; Hansen, T.K.; Moller, N.; Weeke, J.; Jorgensen, J.O. Hyperthyroidism is associated with suppressed circulating ghrelin levels. J Clin Endocrinol Metab 2003, 88, 853–857. [CrossRef]
- Pagotto, U.; Gambineri, A.; Pelusi, C.; Genghini, S.; Cacciari, M.; Otto, B.; Castaneda, T.; Tschop, M.; Pasquali, R. Testosterone replacement therapy restores normal ghrelin in hypogonadal men. J Clin Endocrinol Metab 2003, 88, 4139–4143. [CrossRef]
- Lee, E.S.; Yoon, Y.S.; Park, C.Y.; Kim, H.S.; Um, T.H.; Baik, H.W.; Jang, E.J.; Lee, S.; Park, H.S.; Oh, S.W. Eradication of Helicobacter pylori increases ghrelin mRNA expression in the gastric mucosa. J Korean Med Sci 2010, 25, 265–271. [CrossRef]
- Gagnon, J.; Anini, Y. Insulin and norepinephrine regulate ghrelin secretion from a rat primary stomach cell culture. Endocrinology 2012, 153, 3646–3656. [CrossRef]
- Sakata, I.; Park, W.M.; Walker, A.K.; Piper, P.K.; Chuang, J.C.; Osborne-Lawrence, S.; Zigman, J.M. Glucose-mediated control of ghrelin release from primary cultures of gastric mucosal cells. Am J Physiol Endocrinol Metab 2012, 302, E1300–1310. [CrossRef]
- Iwakura, H.; Li, Y.; Ariyasu, H.; Hosoda, H.; Kanamoto, N.; Bando, M.; Yamada, G.; Hosoda, K.; Nakao, K.; Kangawa, K.; et al. Establishment of a novel ghrelin-producing cell line. Endocrinology 2010, 151, 2940–2945. [CrossRef]
- Seim, I.; Collet, C.; Herington, A.C.; Chopin, L.K. Revised genomic structure of the human ghrelin gene and identification of novel exons, alternative splice variants and natural antisense transcripts. BMC Genomics 2007, 8, 298. [CrossRef]
- Yanagi, S.; Sato, T.; Kangawa, K.; Nakazato, M. The Homeostatic Force of Ghrelin. Cell Metab 2018, 27, 786–804. [CrossRef]
- Zhang, J.V.; Ren, P.G.; Avsian-Kretchmer, O.; Luo, C.W.; Rauch, R.; Klein, C.; Hsueh, A.J. Obestatin, a peptide encoded by the ghrelin gene, opposes ghrelin’s effects on food intake. Science 2005, 310, 996–999. [CrossRef]
- Hassouna, R.; Zizzari, P.; Tolle, V. The ghrelin/obestatin balance in the physiological and pathological control of growth hormone secretion, body composition and food intake. J Neuroendocrinol 2010, 22, 793–804. [CrossRef]
- Mora, M.; Granada, M.L.; Roca, M.; Palomera, E.; Puig, R.; Serra-Prat, M.; Puig-Domingo, M. Obestatin does not modify weight and nutritional behaviour but is associated with metabolic syndrome in old women. Clin Endocrinol (Oxf) 2013, 78, 882–890. [CrossRef]
- Zhu, X.; Cao, Y.; Voogd, K.; Steiner, D.F. On the processing of proghrelin to ghrelin. J Biol Chem 2006, 281, 38867–38870. [CrossRef]
- Yang, J.; Brown, M.S.; Liang, G.; Grishin, N.V.; Goldstein, J.L. Identification of the acyltransferase that octanoylates ghrelin, an appetite-stimulating peptide hormone. Cell 2008, 132, 387–396. [CrossRef]
- Gutierrez, J.A.; Solenberg, P.J.; Perkins, D.R.; Willency, J.A.; Knierman, M.D.; Jin, Z.; Witcher, D.R.; Luo, S.; Onyia, J.E.; Hale, J.E. Ghrelin octanoylation mediated by an orphan lipid transferase. Proc Natl Acad Sci U S A 2008, 105, 6320–6325. [CrossRef]
- Hormaechea-Agulla, D.; Gomez-Gomez, E.; Ibanez-Costa, A.; Carrasco-Valiente, J.; Rivero-Cortes, E.; F, L.L.; Pedraza-Arevalo, S.; Valero-Rosa, J.; Sanchez-Sanchez, R.; Ortega-Salas, R.; et al. Ghrelin O-acyltransferase (GOAT) enzyme is overexpressed in prostate cancer, and its levels are associated with patient’s metabolic status: Potential value as a non-invasive biomarker. Cancer Lett 2016, 383, 125–134. [CrossRef]
- Gomez-Gomez, E.; Jimenez-Vacas, J.M.; Carrasco-Valiente, J.; Herrero-Aguayo, V.; Blanca-Pedregosa, A.M.; Leon-Gonzalez, A.J.; Valero-Rosa, J.; Fernandez-Rueda, J.L.; Gonzalez-Serrano, T.; Lopez-Miranda, J.; et al. Plasma ghrelin O-acyltransferase (GOAT) enzyme levels: A novel non-invasive diagnosis tool for patients with significant prostate cancer. J Cell Mol Med 2018, 22, 5688–5697. [CrossRef]
- Delporte, C. Structure and physiological actions of ghrelin. Scientifica (Cairo) 2013, 2013, 518909. [CrossRef]
- Kakidani, H.; Furutani, Y.; Takahashi, H.; Noda, M.; Morimoto, Y.; Hirose, T.; Asai, M.; Inayama, S.; Nakanishi, S.; Numa, S. Cloning and sequence analysis of cDNA for porcine beta-neo-endorphin/dynorphin precursor. Nature 1982, 298, 245–249. [CrossRef]
- Nakanishi, S.; Inoue, A.; Kita, T.; Nakamura, M.; Chang, A.C.; Cohen, S.N.; Numa, S. Nucleotide sequence of cloned cDNA for bovine corticotropin-beta-lipotropin precursor. Nature 1979, 278, 423–427. [CrossRef]
- Tanaka, M.; Hayashida, Y.; Iguchi, T.; Nakao, N.; Nakai, N.; Nakashima, K. Organization of the mouse ghrelin gene and promoter: occurrence of a short noncoding first exon. Endocrinology 2001, 142, 3697–3700. [CrossRef]
- Satou, M.; Nishi, Y.; Hishinuma, A.; Hosoda, H.; Kangawa, K.; Sugimoto, H. Identification of activated protein C as a ghrelin endopeptidase in bovine plasma. J Endocrinol 2015, 224, 61–73. [CrossRef]
- Seim, I.; Jeffery, P.L.; Thomas, P.B.; Walpole, C.M.; Maugham, M.; Fung, J.N.; Yap, P.Y.; O’Keeffe, A.J.; Lai, J.; Whiteside, E.J.; et al. Multi-species sequence comparison reveals conservation of ghrelin gene-derived splice variants encoding a truncated ghrelin peptide. Endocrine 2016, 52, 609–617. [CrossRef]
- Fernandez, G.; Fittipaldi, A.; Lufrano, D.; Mustafa, E.R.; Castrogiovanni, D.; Barrile, F.; De Francesco, P.N.; Tolosa, M.J.; Rodriguez, S.S.; Lalonde, T.; et al. Mini-ghrelins: Functional Characterization of N-terminal Peptides Derived From Ghrelin Proteolysis in Male Samples. Endocrinology 2025, 166. [CrossRef]
- Bednarek, M.A.; Feighner, S.D.; Pong, S.S.; McKee, K.K.; Hreniuk, D.L.; Silva, M.V.; Warren, V.A.; Howard, A.D.; Van Der Ploeg, L.H.; Heck, J.V. Structure-function studies on the new growth hormone-releasing peptide, ghrelin: minimal sequence of ghrelin necessary for activation of growth hormone secretagogue receptor 1a. J Med Chem 2000, 43, 4370–4376. [CrossRef]
- Matsumoto, M.; Hosoda, H.; Kitajima, Y.; Morozumi, N.; Minamitake, Y.; Tanaka, S.; Matsuo, H.; Kojima, M.; Hayashi, Y.; Kangawa, K. Structure-activity relationship of ghrelin: pharmacological study of ghrelin peptides. Biochem Biophys Res Commun 2001, 287, 142–146. [CrossRef]
- Ma, D.; Wang, Z.; Merrikh, C.N.; Lang, K.S.; Lu, P.; Li, X.; Merrikh, H.; Rao, Z.; Xu, W. Crystal structure of a membrane-bound O-acyltransferase. Nature 2018, 562, 286–290. [CrossRef]
- Campana, M.B.; Irudayanathan, F.J.; Davis, T.R.; McGovern-Gooch, K.R.; Loftus, R.; Ashkar, M.; Escoffery, N.; Navarro, M.; Sieburg, M.A.; Nangia, S.; et al. The ghrelin O-acyltransferase structure reveals a catalytic channel for transmembrane hormone acylation. J Biol Chem 2019, 294, 14166–14174. [CrossRef]
- Hopkins, A.L.; Nelson, T.A.; Guschina, I.A.; Parsons, L.C.; Lewis, C.L.; Brown, R.C.; Christian, H.C.; Davies, J.S.; Wells, T. Unacylated ghrelin promotes adipogenesis in rodent bone marrow via ghrelin O-acyl transferase and GHS-R(1a) activity: evidence for target cell-induced acylation. Sci Rep 2017, 7, 45541. [CrossRef]
- Murtuza, M.I.; Isokawa, M. Endogenous ghrelin-O-acyltransferase (GOAT) acylates local ghrelin in the hippocampus. J Neurochem 2018, 144, 58–67. [CrossRef]
- Banks, W.A.; Tschop, M.; Robinson, S.M.; Heiman, M.L. Extent and direction of ghrelin transport across the blood-brain barrier is determined by its unique primary structure. J Pharmacol Exp Ther 2002, 302, 822–827. [CrossRef]
- Campana, M.B.; Davis, T.R.; Novak, S.X.; Cleverdon, E.R.; Bates, M.; Krishnan, N.; Curtis, E.R.; Childs, M.D.; Pierce, M.R.; Morales-Rodriguez, Y.; et al. Cellular Uptake of a Fluorescent Ligand Reveals Ghrelin O-Acyltransferase Interacts with Extracellular Peptides and Exhibits Unexpected Localization for a Secretory Pathway Enzyme. ACS Chem Biol 2023, 18, 1880–1890. [CrossRef]
- Li, K.; Li, J.; Zheng, J.; Qin, S. Reactive Astrocytes in Neurodegenerative Diseases. Aging Dis 2019, 10, 664–675. [CrossRef]
- Kojima, M.; Kangawa, K. Ghrelin: structure and function. Physiol Rev 2005, 85, 495–522. [CrossRef]
- Gauna, C.; van de Zande, B.; van Kerkwijk, A.; Themmen, A.P.; van der Lely, A.J.; Delhanty, P.J. Unacylated ghrelin is not a functional antagonist but a full agonist of the type 1a growth hormone secretagogue receptor (GHS-R). Mol Cell Endocrinol 2007, 274, 30–34. [CrossRef]
- Lambert, E.; Lambert, G.; Ika-Sari, C.; Dawood, T.; Lee, K.; Chopra, R.; Straznicky, N.; Eikelis, N.; Drew, S.; Tilbrook, A.; et al. Ghrelin modulates sympathetic nervous system activity and stress response in lean and overweight men. Hypertension 2011, 58, 43–50. [CrossRef]
- Pei, X.M.; Yung, B.Y.; Yip, S.P.; Ying, M.; Benzie, I.F.; Siu, P.M. Desacyl ghrelin prevents doxorubicin-induced myocardial fibrosis and apoptosis via the GHSR-independent pathway. Am J Physiol Endocrinol Metab 2014, 306, E311–323. [CrossRef]
- Mahbod, P.; Smith, E.P.; Fitzgerald, M.E.; Morano, R.L.; Packard, B.A.; Ghosal, S.; Scheimann, J.R.; Perez-Tilve, D.; Herman, J.P.; Tong, J. Desacyl Ghrelin Decreases Anxiety-like Behavior in Male Mice. Endocrinology 2018, 159, 388–399. [CrossRef]
- Yuan, F.; Zhang, Q.; Dong, H.; Xiang, X.; Zhang, W.; Zhang, Y.; Li, Y. Effects of Des-acyl Ghrelin on Insulin Sensitivity and Macrophage Polarization in Adipose Tissue. J Transl Int Med 2021, 9, 84–97. [CrossRef]
- Witley, S.; Edvardsson, C.E.; Aranas, C.; Tufvesson-Alm, M.; Stalberga, D.; Green, H.; Vestlund, J.; Jerlhag, E. Des-acyl ghrelin reduces alcohol intake and alcohol-induced reward in rodents. Transl Psychiatry 2024, 14, 277. [CrossRef]
- Rodriguez, A.; Gomez-Ambrosi, J.; Catalan, V.; Rotellar, F.; Valenti, V.; Silva, C.; Mugueta, C.; Pulido, M.R.; Vazquez, R.; Salvador, J.; et al. The ghrelin O-acyltransferase-ghrelin system reduces TNF-alpha-induced apoptosis and autophagy in human visceral adipocytes. Diabetologia 2012, 55, 3038–3050. [CrossRef]
- Slupecka, M.; Wolinski, J.; Pierzynowski, S.G. The effects of enteral ghrelin administration on the remodeling of the small intestinal mucosa in neonatal piglets. Regul Pept 2012, 174, 38–45. [CrossRef]
- Bonfili, L.; Cuccioloni, M.; Cecarini, V.; Mozzicafreddo, M.; Palermo, F.A.; Cocci, P.; Angeletti, M.; Eleuteri, A.M. Ghrelin induces apoptosis in colon adenocarcinoma cells via proteasome inhibition and autophagy induction. Apoptosis 2013, 18, 1188–1200. [CrossRef]
- Toshinai, K.; Yamaguchi, H.; Sun, Y.; Smith, R.G.; Yamanaka, A.; Sakurai, T.; Date, Y.; Mondal, M.S.; Shimbara, T.; Kawagoe, T.; et al. Des-acyl ghrelin induces food intake by a mechanism independent of the growth hormone secretagogue receptor. Endocrinology 2006, 147, 2306–2314. [CrossRef]
- Asakawa, A.; Inui, A.; Fujimiya, M.; Sakamaki, R.; Shinfuku, N.; Ueta, Y.; Meguid, M.M.; Kasuga, M. Stomach regulates energy balance via acylated ghrelin and desacyl ghrelin. Gut 2005, 54, 18–24. [CrossRef]
- Tong, J.; Dave, N.; Mugundu, G.M.; Davis, H.W.; Gaylinn, B.D.; Thorner, M.O.; Tschop, M.H.; D’Alessio, D.; Desai, P.B. The pharmacokinetics of acyl, des-acyl, and total ghrelin in healthy human subjects. Eur J Endocrinol 2013, 168, 821–828. [CrossRef]
- Yoshimoto, A.; Mori, K.; Sugawara, A.; Mukoyama, M.; Yahata, K.; Suganami, T.; Takaya, K.; Hosoda, H.; Kojima, M.; Kangawa, K.; et al. Plasma ghrelin and desacyl ghrelin concentrations in renal failure. J Am Soc Nephrol 2002, 13, 2748–2752. [CrossRef]
- Hosoda, H.; Kojima, M.; Matsuo, H.; Kangawa, K. Ghrelin and des-acyl ghrelin: two major forms of rat ghrelin peptide in gastrointestinal tissue. Biochem Biophys Res Commun 2000, 279, 909–913. [CrossRef]
- Murakami, N.; Hayashida, T.; Kuroiwa, T.; Nakahara, K.; Ida, T.; Mondal, M.S.; Nakazato, M.; Kojima, M.; Kangawa, K. Role for central ghrelin in food intake and secretion profile of stomach ghrelin in rats. J Endocrinol 2002, 174, 283–288. [CrossRef]
- De Vriese, C.; Gregoire, F.; Lema-Kisoka, R.; Waelbroeck, M.; Robberecht, P.; Delporte, C. Ghrelin degradation by serum and tissue homogenates: identification of the cleavage sites. Endocrinology 2004, 145, 4997–5005. [CrossRef]
- Schopfer, L.M.; Lockridge, O.; Brimijoin, S. Pure human butyrylcholinesterase hydrolyzes octanoyl ghrelin to desacyl ghrelin. Gen Comp Endocrinol 2015, 224, 61–68. [CrossRef]
- Blatnik, M.; Soderstrom, C.I. A practical guide for the stabilization of acylghrelin in human blood collections. Clin Endocrinol (Oxf) 2011, 74, 325–331. [CrossRef]
- Chen, V.P.; Gao, Y.; Geng, L.; Parks, R.J.; Pang, Y.P.; Brimijoin, S. Plasma butyrylcholinesterase regulates ghrelin to control aggression. Proc Natl Acad Sci U S A 2015, 112, 2251–2256. [CrossRef]
- Shanado, Y.; Kometani, M.; Uchiyama, H.; Koizumi, S.; Teno, N. Lysophospholipase I identified as a ghrelin deacylation enzyme in rat stomach. Biochem Biophys Res Commun 2004, 325, 1487–1494. [CrossRef]
- Satou, M.; Nishi, Y.; Yoh, J.; Hattori, Y.; Sugimoto, H. Identification and characterization of acyl-protein thioesterase 1/lysophospholipase I as a ghrelin deacylation/lysophospholipid hydrolyzing enzyme in fetal bovine serum and conditioned medium. Endocrinology 2010, 151, 4765–4775. [CrossRef]
- Eubanks, L.M.; Stowe, G.N.; De Lamo Marin, S.; Mayorov, A.V.; Hixon, M.S.; Janda, K.D. Identification of alpha2 macroglobulin as a major serum ghrelin esterase. Angew Chem Int Ed Engl 2011, 50, 10699–10702. [CrossRef]
- Hosoda, H.; Doi, K.; Nagaya, N.; Okumura, H.; Nakagawa, E.; Enomoto, M.; Ono, F.; Kangawa, K. Optimum collection and storage conditions for ghrelin measurements: octanoyl modification of ghrelin is rapidly hydrolyzed to desacyl ghrelin in blood samples. Clin Chem 2004, 50, 1077–1080. [CrossRef]
- McGovern-Gooch, K.R.; Rodrigues, T.; Darling, J.E.; Sieburg, M.A.; Abizaid, A.; Hougland, J.L. Ghrelin Octanoylation Is Completely Stabilized in Biological Samples by Alkyl Fluorophosphonates. Endocrinology 2016, 157, 4330–4338. [CrossRef]
- Sangiao-Alvarellos, S.; Cordido, F. Effect of ghrelin on glucose-insulin homeostasis: therapeutic implications. Int J Pept 2010, 2010. [CrossRef]
- Wu, R.; Dong, W.; Cui, X.; Zhou, M.; Simms, H.H.; Ravikumar, T.S.; Wang, P. Ghrelin down-regulates proinflammatory cytokines in sepsis through activation of the vagus nerve. Ann Surg 2007, 245, 480–486. [CrossRef]
- Moon, M.; Kim, H.G.; Hwang, L.; Seo, J.H.; Kim, S.; Hwang, S.; Kim, S.; Lee, D.; Chung, H.; Oh, M.S.; et al. Neuroprotective effect of ghrelin in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mouse model of Parkinson’s disease by blocking microglial activation. Neurotox Res 2009, 15, 332–347. [CrossRef]
- Zhang, G.; Yin, X.; Qi, Y.; Pendyala, L.; Chen, J.; Hou, D.; Tang, C. Ghrelin and cardiovascular diseases. Curr Cardiol Rev 2010, 6, 62–70. [CrossRef]
- Szentirmai, E.; Kapas, L.; Sun, Y.; Smith, R.G.; Krueger, J.M. Spontaneous sleep and homeostatic sleep regulation in ghrelin knockout mice. Am J Physiol Regul Integr Comp Physiol 2007, 293, R510–517. [CrossRef]
- Li, B.; Zeng, M.; He, W.; Huang, X.; Luo, L.; Zhang, H.; Deng, D.Y. Ghrelin protects alveolar macrophages against lipopolysaccharide-induced apoptosis through growth hormone secretagogue receptor 1a-dependent c-Jun N-terminal kinase and Wnt/beta-catenin signaling and suppresses lung inflammation. Endocrinology 2015, 156, 203–217. [CrossRef]
- Gubina, N.V.; Kupnovytska, I.H.; Mishchuk, V.H.; Markiv, H.D. Ghrelin Levels and Decreased Kidney Function in Patients with Early Stages of Chronic Kidney Disease Against the Background of Obesity. J Med Life 2020, 13, 530–535. [CrossRef]
- Fukushima, N.; Hanada, R.; Teranishi, H.; Fukue, Y.; Tachibana, T.; Ishikawa, H.; Takeda, S.; Takeuchi, Y.; Fukumoto, S.; Kangawa, K.; et al. Ghrelin directly regulates bone formation. J Bone Miner Res 2005, 20, 790–798. [CrossRef]
- Wu, J.T.; Kral, J.G. Ghrelin: integrative neuroendocrine peptide in health and disease. Ann Surg 2004, 239, 464–474. [CrossRef]
- Cummings, D.E. Ghrelin and the short- and long-term regulation of appetite and body weight. Physiol Behav 2006, 89, 71–84. [CrossRef]
- Sovetkina, A.; Nadir, R.; Fung, J.N.M.; Nadjarpour, A.; Beddoe, B. The Physiological Role of Ghrelin in the Regulation of Energy and Glucose Homeostasis. Cureus 2020, 12, e7941. [CrossRef]
- Pradhan, G.; Samson, S.L.; Sun, Y. Ghrelin: much more than a hunger hormone. Curr Opin Clin Nutr Metab Care 2013, 16, 619–624. [CrossRef]
- Varela, L.; Vazquez, M.J.; Cordido, F.; Nogueiras, R.; Vidal-Puig, A.; Dieguez, C.; Lopez, M. Ghrelin and lipid metabolism: key partners in energy balance. J Mol Endocrinol 2011, 46, R43–63. [CrossRef]
- Zhang, L.; Liu, J.; Gao, D.; Li, D. Role of ghrelin in promoting catch-up growth and maintaining metabolic homeostasis in small-for-gestational-age infants. Front Pediatr 2024, 12, 1395571. [CrossRef]
- Wortley, K.E.; Anderson, K.D.; Garcia, K.; Murray, J.D.; Malinova, L.; Liu, R.; Moncrieffe, M.; Thabet, K.; Cox, H.J.; Yancopoulos, G.D.; et al. Genetic deletion of ghrelin does not decrease food intake but influences metabolic fuel preference. Proc Natl Acad Sci U S A 2004, 101, 8227–8232. [CrossRef]
- Venables, G.; Hunne, B.; Bron, R.; Cho, H.J.; Brock, J.A.; Furness, J.B. Ghrelin receptors are expressed by distal tubules of the mouse kidney. Cell Tissue Res 2011, 346, 135–139. [CrossRef]
- Davenport, A.P.; Bonner, T.I.; Foord, S.M.; Harmar, A.J.; Neubig, R.R.; Pin, J.P.; Spedding, M.; Kojima, M.; Kangawa, K. International Union of Pharmacology. LVI. Ghrelin receptor nomenclature, distribution, and function. Pharmacol Rev 2005, 57, 541–546. [CrossRef]
- Cowley, M.A.; Smith, R.G.; Diano, S.; Tschop, M.; Pronchuk, N.; Grove, K.L.; Strasburger, C.J.; Bidlingmaier, M.; Esterman, M.; Heiman, M.L.; et al. The distribution and mechanism of action of ghrelin in the CNS demonstrates a novel hypothalamic circuit regulating energy homeostasis. Neuron 2003, 37, 649–661. [CrossRef]
- Hattori, N.; Saito, T.; Yagyu, T.; Jiang, B.H.; Kitagawa, K.; Inagaki, C. GH, GH receptor, GH secretagogue receptor, and ghrelin expression in human T cells, B cells, and neutrophils. J Clin Endocrinol Metab 2001, 86, 4284–4291. [CrossRef]
- Van Craenenbroeck, M.; Gregoire, F.; De Neef, P.; Robberecht, P.; Perret, J. Ala-scan of ghrelin (1-14): interaction with the recombinant human ghrelin receptor. Peptides 2004, 25, 959–965. [CrossRef]
- Matsumoto, M.; Kitajima, Y.; Iwanami, T.; Hayashi, Y.; Tanaka, S.; Minamitake, Y.; Hosoda, H.; Kojima, M.; Matsuo, H.; Kangawa, K. Structural similarity of ghrelin derivatives to peptidyl growth hormone secretagogues. Biochem Biophys Res Commun 2001, 284, 655–659. [CrossRef]
- De Ricco, R.; Valensin, D.; Gaggelli, E.; Valensin, G. Conformation propensities of des-acyl-ghrelin as probed by CD and NMR. Peptides 2013, 43, 62–67. [CrossRef]
- Lee, J.H.; Patel, K.; Tae, H.J.; Lustig, A.; Kim, J.W.; Mattson, M.P.; Taub, D.D. Ghrelin augments murine T-cell proliferation by activation of the phosphatidylinositol-3-kinase, extracellular signal-regulated kinase and protein kinase C signaling pathways. FEBS Lett 2014, 588, 4708–4719. [CrossRef]
- Chen, J.H.; Huang, S.M.; Chen, C.C.; Tsai, C.F.; Yeh, W.L.; Chou, S.J.; Hsieh, W.T.; Lu, D.Y. Ghrelin induces cell migration through GHS-R, CaMKII, AMPK, and NF-kappaB signaling pathway in glioma cells. J Cell Biochem 2011, 112, 2931–2941. [CrossRef]
- Bayliss, J.A.; Lemus, M.B.; Stark, R.; Santos, V.V.; Thompson, A.; Rees, D.J.; Galic, S.; Elsworth, J.D.; Kemp, B.E.; Davies, J.S.; et al. Ghrelin-AMPK Signaling Mediates the Neuroprotective Effects of Calorie Restriction in Parkinson’s Disease. J Neurosci 2016, 36, 3049–3063. [CrossRef]
- Cavalier, M.; Crouzin, N.; Ben Sedrine, A.; de Jesus Ferreira, M.C.; Guiramand, J.; Cohen-Solal, C.; Fehrentz, J.A.; Martinez, J.; Barbanel, G.; Vignes, M. Involvement of PKA and ERK pathways in ghrelin-induced long-lasting potentiation of excitatory synaptic transmission in the CA1 area of rat hippocampus. Eur J Neurosci 2015, 42, 2568–2576. [CrossRef]
- Mousseaux, D.; Le Gallic, L.; Ryan, J.; Oiry, C.; Gagne, D.; Fehrentz, J.A.; Galleyrand, J.C.; Martinez, J. Regulation of ERK1/2 activity by ghrelin-activated growth hormone secretagogue receptor 1A involves a PLC/PKCvarepsilon pathway. Br J Pharmacol 2006, 148, 350–365. [CrossRef]
- Santos-Zas, I.; Lodeiro, M.; Gurriaran-Rodriguez, U.; Bouzo-Lorenzo, M.; Mosteiro, C.S.; Casanueva, F.F.; Casabiell, X.; Pazos, Y.; Camina, J.P. beta-Arrestin signal complex plays a critical role in adipose differentiation. Int J Biochem Cell Biol 2013, 45, 1281–1292. [CrossRef]
- Bouzo-Lorenzo, M.; Santo-Zas, I.; Lodeiro, M.; Nogueiras, R.; Casanueva, F.F.; Castro, M.; Pazos, Y.; Tobin, A.B.; Butcher, A.J.; Camina, J.P. Distinct phosphorylation sites on the ghrelin receptor, GHSR1a, establish a code that determines the functions of ss-arrestins. Sci Rep 2016, 6, 22495. [CrossRef]
- Mende, F.; Hundahl, C.; Plouffe, B.; Skov, L.J.; Sivertsen, B.; Madsen, A.N.; Luckmann, M.; Diep, T.A.; Offermanns, S.; Frimurer, T.M.; et al. Translating biased signaling in the ghrelin receptor system into differential in vivo functions. Proc Natl Acad Sci U S A 2018, 115, E10255–E10264. [CrossRef]
- Dezaki, K.; Kakei, M.; Yada, T. Ghrelin uses Galphai2 and activates voltage-dependent K+ channels to attenuate glucose-induced Ca2+ signaling and insulin release in islet beta-cells: novel signal transduction of ghrelin. Diabetes 2007, 56, 2319–2327. [CrossRef]
- Shiimura, Y.; Horita, S.; Hamamoto, A.; Asada, H.; Hirata, K.; Tanaka, M.; Mori, K.; Uemura, T.; Kobayashi, T.; Iwata, S.; et al. Structure of an antagonist-bound ghrelin receptor reveals possible ghrelin recognition mode. Nat Commun 2020, 11, 4160. [CrossRef]
- Wang, Y.; Guo, S.; Zhuang, Y.; Yun, Y.; Xu, P.; He, X.; Guo, J.; Yin, W.; Xu, H.E.; Xie, X.; et al. Molecular recognition of an acyl-peptide hormone and activation of ghrelin receptor. Nat Commun 2021, 12, 5064. [CrossRef]
- Fujii, H.; Yamada, Y.; Iihara, H.; Kobayashi, R.; Suzuki, A. Anamorelin in the Management of Cancer Cachexia: Clinical Efficacy, Challenges, and Future Directions. Anticancer Res 2025, 45, 865–881. [CrossRef]
- Yamaguchi, A.; Kamada, H.; Semba, S.; Kato, N.; Teraoka, Y.; Mizumoto, T.; Tamaru, Y.; Hatakeyama, T.; Kouno, H.; Shibata, Y.; et al. Real-world effectiveness of anamorelin in patients with unresectable and relapse pancreatic cancer: a prospective observational study. J Gastrointest Oncol 2025, 16, 1268–1279. [CrossRef]
- Shiimura, Y.; Im, D.; Tany, R.; Asada, H.; Kise, R.; Kurumiya, E.; Wakasugi-Masuho, H.; Yasuda, S.; Matsui, K.; Kishikawa, J.I.; et al. The structure and function of the ghrelin receptor coding for drug actions. Nat Struct Mol Biol 2025, 32, 531–542. [CrossRef]
- Masuho, I.; Ostrovskaya, O.; Kramer, G.M.; Jones, C.D.; Xie, K.; Martemyanov, K.A. Distinct profiles of functional discrimination among G proteins determine the actions of G protein-coupled receptors. Sci Signal 2015, 8, ra123. [CrossRef]
- Hedegaard, M.A.; Holst, B. The Complex Signaling Pathways of the Ghrelin Receptor. Endocrinology 2020, 161. [CrossRef]
- Mary, S.; Damian, M.; Louet, M.; Floquet, N.; Fehrentz, J.A.; Marie, J.; Martinez, J.; Baneres, J.L. Ligands and signaling proteins govern the conformational landscape explored by a G protein-coupled receptor. Proc Natl Acad Sci U S A 2012, 109, 8304–8309. [CrossRef]
- Damian, M.; Marie, J.; Leyris, J.P.; Fehrentz, J.A.; Verdie, P.; Martinez, J.; Baneres, J.L.; Mary, S. High constitutive activity is an intrinsic feature of ghrelin receptor protein: a study with a functional monomeric GHS-R1a receptor reconstituted in lipid discs. J Biol Chem 2012, 287, 3630–3641. [CrossRef]
- Ramirez, V.T.; van Oeffelen, W.; Torres-Fuentes, C.; Chruscicka, B.; Druelle, C.; Golubeva, A.V.; van de Wouw, M.; Dinan, T.G.; Cryan, J.F.; Schellekens, H. Differential functional selectivity and downstream signaling bias of ghrelin receptor antagonists and inverse agonists. FASEB J 2019, 33, 518–531. [CrossRef]
- Krause, A.; Sillard, R.; Kleemeier, B.; Kluver, E.; Maronde, E.; Conejo-Garcia, J.R.; Forssmann, W.G.; Schulz-Knappe, P.; Nehls, M.C.; Wattler, F.; et al. Isolation and biochemical characterization of LEAP-2, a novel blood peptide expressed in the liver. Protein Sci 2003, 12, 143–152. [CrossRef]
- Ge, X.; Yang, H.; Bednarek, M.A.; Galon-Tilleman, H.; Chen, P.; Chen, M.; Lichtman, J.S.; Wang, Y.; Dalmas, O.; Yin, Y.; et al. LEAP2 Is an Endogenous Antagonist of the Ghrelin Receptor. Cell Metab 2018, 27, 461–469 e466. [CrossRef]
- Lu, X.; Huang, L.; Huang, Z.; Feng, D.; Clark, R.J.; Chen, C. LEAP-2: An Emerging Endogenous Ghrelin Receptor Antagonist in the Pathophysiology of Obesity. Front Endocrinol (Lausanne) 2021, 12, 717544. [CrossRef]
- Lang, Y.Y.; Xu, X.Y.; Liu, Y.L.; Ye, C.F.; Hu, N.; Yao, Q.; Cheng, W.S.; Cheng, Z.G.; Liu, Y. Ghrelin Relieves Obesity-Induced Myocardial Injury by Regulating the Epigenetic Suppression of miR-196b Mediated by lncRNA HOTAIR. Obes Facts 2022, 15, 540–549. [CrossRef]
- Liu, Y.; Xu, X.Y.; Shen, Y.; Ye, C.F.; Hu, N.; Yao, Q.; Lv, X.Z.; Long, S.L.; Ren, C.; Lang, Y.Y.; et al. Ghrelin protects against obesity-induced myocardial injury by regulating the lncRNA H19/miR-29a/IGF-1 signalling axis. Exp Mol Pathol 2020, 114, 104405. [CrossRef]
- Lang, Y.; Liu, Y.; Ye, C.; Tang, X.; Cheng, Z.; Xie, L.; Feng, L.; Liu, Y. Loss of LEAP-2 alleviates obesity-induced myocardial injury by regulating macrophage polarization. Exp Cell Res 2023, 430, 113702. [CrossRef]
- Liu, D.; Li, S. LEAP2: Next game-changer of pharmacotherapy for overweight and obesity? Cell Rep Med 2022, 3, 100612. [CrossRef]
- Hola, L.; Zelezna, B.; Karnosova, A.; Kunes, J.; Fehrentz, J.A.; Denoyelle, S.; Cantel, S.; Blechova, M.; Sykora, D.; Myskova, A.; et al. A Novel Truncated Liver Enriched Antimicrobial Peptide-2 Palmitoylated at its N-Terminal Antagonizes Effects of Ghrelin. J Pharmacol Exp Ther 2022, 383, 129–136. [CrossRef]
- Bukhari, S.N.A. An insight into the multifunctional role of ghrelin and structure activity relationship studies of ghrelin receptor ligands with clinical trials. Eur J Med Chem 2022, 235, 114308. [CrossRef]
- Garcia, J.M.; Polvino, W.J. Pharmacodynamic hormonal effects of anamorelin, a novel oral ghrelin mimetic and growth hormone secretagogue in healthy volunteers. Growth Horm IGF Res 2009, 19, 267–273. [CrossRef]
- Patchett, A.A.; Nargund, R.P.; Tata, J.R.; Chen, M.H.; Barakat, K.J.; Johnston, D.B.; Cheng, K.; Chan, W.W.; Butler, B.; Hickey, G.; et al. Design and biological activities of L-163,191 (MK-0677): a potent, orally active growth hormone secretagogue. Proc Natl Acad Sci U S A 1995, 92, 7001–7005. [CrossRef]
- Copinschi, G.; Van Onderbergen, A.; L’Hermite-Baleriaux, M.; Mendel, C.M.; Caufriez, A.; Leproult, R.; Bolognese, J.A.; De Smet, M.; Thorner, M.O.; Van Cauter, E. Effects of a 7-day treatment with a novel, orally active, growth hormone (GH) secretagogue, MK-677, on 24-hour GH profiles, insulin-like growth factor I, and adrenocortical function in normal young men. J Clin Endocrinol Metab 1996, 81, 2776–2782. [CrossRef]
- Jacks, T.; Hickey, G.; Judith, F.; Taylor, J.; Chen, H.; Krupa, D.; Feeney, W.; Schoen, W.; Ok, D.; Fisher, M.; et al. Effects of acute and repeated intravenous administration of L-692,585, a novel non-peptidyl growth hormone secretagogue, on plasma growth hormone, IGF-1, ACTH, cortisol, prolactin, insulin, and thyroxine levels in beagles. J Endocrinol 1994, 143, 399–406. [CrossRef]
- Smith, R.G.; Cheng, K.; Schoen, W.R.; Pong, S.S.; Hickey, G.; Jacks, T.; Butler, B.; Chan, W.W.; Chaung, L.Y.; Judith, F.; et al. A nonpeptidyl growth hormone secretagogue. Science 1993, 260, 1640–1643. [CrossRef]
- Moulin, A.; Demange, L.; Berge, G.; Gagne, D.; Ryan, J.; Mousseaux, D.; Heitz, A.; Perrissoud, D.; Locatelli, V.; Torsello, A.; et al. Toward potent ghrelin receptor ligands based on trisubstituted 1,2,4-triazole structure. 2. Synthesis and pharmacological in vitro and in vivo evaluations. J Med Chem 2007, 50, 5790–5806. [CrossRef]
- Bhattacharya, S.K.; Andrews, K.; Beveridge, R.; Cameron, K.O.; Chen, C.; Dunn, M.; Fernando, D.; Gao, H.; Hepworth, D.; Jackson, V.M.; et al. Discovery of PF-5190457, a Potent, Selective, and Orally Bioavailable Ghrelin Receptor Inverse Agonist Clinical Candidate. ACS Med Chem Lett 2014, 5, 474–479. [CrossRef]
- Lee, M.R.; Tapocik, J.D.; Ghareeb, M.; Schwandt, M.L.; Dias, A.A.; Le, A.N.; Cobbina, E.; Farinelli, L.A.; Bouhlal, S.; Farokhnia, M.; et al. The novel ghrelin receptor inverse agonist PF-5190457 administered with alcohol: preclinical safety experiments and a phase 1b human laboratory study. Mol Psychiatry 2020, 25, 461–475. [CrossRef]
- Cheng, K.; Chan, W.W.; Barreto, A., Jr.; Convey, E.M.; Smith, R.G. The synergistic effects of His-D-Trp-Ala-Trp-D-Phe-Lys-NH2 on growth hormone (GH)-releasing factor-stimulated GH release and intracellular adenosine 3′,5′-monophosphate accumulation in rat primary pituitary cell culture. Endocrinology 1989, 124, 2791–2798. [CrossRef]
- Traebert, M.; Riediger, T.; Whitebread, S.; Scharrer, E.; Schmid, H.A. Ghrelin acts on leptin-responsive neurones in the rat arcuate nucleus. J Neuroendocrinol 2002, 14, 580–586. [CrossRef]
- Holst, B.; Cygankiewicz, A.; Jensen, T.H.; Ankersen, M.; Schwartz, T.W. High constitutive signaling of the ghrelin receptor--identification of a potent inverse agonist. Mol Endocrinol 2003, 17, 2201–2210. [CrossRef]
- Holst, B.; Mokrosinski, J.; Lang, M.; Brandt, E.; Nygaard, R.; Frimurer, T.M.; Beck-Sickinger, A.G.; Schwartz, T.W. Identification of an efficacy switch region in the ghrelin receptor responsible for interchange between agonism and inverse agonism. J Biol Chem 2007, 282, 15799–15811. [CrossRef]
- Dhurandhar, E.J.; Allison, D.B.; van Groen, T.; Kadish, I. Hunger in the absence of caloric restriction improves cognition and attenuates Alzheimer’s disease pathology in a mouse model. PLoS One 2013, 8, e60437. [CrossRef]
- Bansal, V.; Ryu, S.Y.; Blow, C.; Costantini, T.; Loomis, W.; Eliceiri, B.; Baird, A.; Wolf, P.; Coimbra, R. The hormone ghrelin prevents traumatic brain injury induced intestinal dysfunction. J Neurotrauma 2010, 27, 2255–2260. [CrossRef]
- Delhanty, P.J.; Huisman, M.; Baldeon-Rojas, L.Y.; van den Berge, I.; Grefhorst, A.; Abribat, T.; Leenen, P.J.; Themmen, A.P.; van der Lely, A.J. Des-acyl ghrelin analogs prevent high-fat-diet-induced dysregulation of glucose homeostasis. FASEB J 2013, 27, 1690–1700. [CrossRef]
- Wu, C.S.; Wei, Q.; Wang, H.; Kim, D.M.; Balderas, M.; Wu, G.; Lawler, J.; Safe, S.; Guo, S.; Devaraj, S.; et al. Protective Effects of Ghrelin on Fasting-Induced Muscle Atrophy in Aging Mice. J Gerontol A Biol Sci Med Sci 2020, 75, 621–630. [CrossRef]
- Rocha, N.N.; de Oliveira, M.V.; Braga, C.L.; Guimaraes, G.; Maia, L.A.; Padilha, G.A.; Silva, J.D.; Takiya, C.M.; Capelozzi, V.L.; Silva, P.L.; et al. Ghrelin therapy improves lung and cardiovascular function in experimental emphysema. Respir Res 2017, 18, 185. [CrossRef]
- Chorny, A.; Anderson, P.; Gonzalez-Rey, E.; Delgado, M. Ghrelin protects against experimental sepsis by inhibiting high-mobility group box 1 release and by killing bacteria. J Immunol 2008, 180, 8369–8377. [CrossRef]
- Makki, K.; Froguel, P.; Wolowczuk, I. Adipose tissue in obesity-related inflammation and insulin resistance: cells, cytokines, and chemokines. ISRN Inflamm 2013, 2013, 139239. [CrossRef]
- Granado, M.; Priego, T.; Martin, A.I.; Villanua, M.A.; Lopez-Calderon, A. Anti-inflammatory effect of the ghrelin agonist growth hormone-releasing peptide-2 (GHRP-2) in arthritic rats. Am J Physiol Endocrinol Metab 2005, 288, E486–492. [CrossRef]
- Dixit, V.D.; Schaffer, E.M.; Pyle, R.S.; Collins, G.D.; Sakthivel, S.K.; Palaniappan, R.; Lillard, J.W., Jr.; Taub, D.D. Ghrelin inhibits leptin- and activation-induced proinflammatory cytokine expression by human monocytes and T cells. J Clin Invest 2004, 114, 57–66. [CrossRef]
- Waseem, T.; Duxbury, M.; Ito, H.; Ashley, S.W.; Robinson, M.K. Exogenous ghrelin modulates release of pro-inflammatory and anti-inflammatory cytokines in LPS-stimulated macrophages through distinct signaling pathways. Surgery 2008, 143, 334–342. [CrossRef]
- Kizaki, T.; Maegawa, T.; Sakurai, T.; Ogasawara, J.E.; Ookawara, T.; Oh-ishi, S.; Izawa, T.; Haga, S.; Ohno, H. Voluntary exercise attenuates obesity-associated inflammation through ghrelin expressed in macrophages. Biochem Biophys Res Commun 2011, 413, 454–459. [CrossRef]
- Gasmi, A.; Noor, S.; Menzel, A.; Dosa, A.; Pivina, L.; Bjorklund, G. Obesity and Insulin Resistance: Associations with Chronic Inflammation, Genetic and Epigenetic Factors. Curr Med Chem 2021, 28, 800–826. [CrossRef]
- Bustin, M. Regulation of DNA-dependent activities by the functional motifs of the high-mobility-group chromosomal proteins. Mol Cell Biol 1999, 19, 5237–5246. [CrossRef]
- Tang, D.; Kang, R.; Livesey, K.M.; Cheh, C.W.; Farkas, A.; Loughran, P.; Hoppe, G.; Bianchi, M.E.; Tracey, K.J.; Zeh, H.J., 3rd; et al. Endogenous HMGB1 regulates autophagy. J Cell Biol 2010, 190, 881–892. [CrossRef]
- Bonaldi, T.; Talamo, F.; Scaffidi, P.; Ferrera, D.; Porto, A.; Bachi, A.; Rubartelli, A.; Agresti, A.; Bianchi, M.E. Monocytic cells hyperacetylate chromatin protein HMGB1 to redirect it towards secretion. EMBO J 2003, 22, 5551–5560. [CrossRef]
- Klune, J.R.; Dhupar, R.; Cardinal, J.; Billiar, T.R.; Tsung, A. HMGB1: endogenous danger signaling. Mol Med 2008, 14, 476–484. [CrossRef]
- Wang, H.; Vishnubhakat, J.M.; Bloom, O.; Zhang, M.; Ombrellino, M.; Sama, A.; Tracey, K.J. Proinflammatory cytokines (tumor necrosis factor and interleukin 1) stimulate release of high mobility group protein-1 by pituicytes. Surgery 1999, 126, 389–392.
- Andersson, U.; Wang, H.; Palmblad, K.; Aveberger, A.C.; Bloom, O.; Erlandsson-Harris, H.; Janson, A.; Kokkola, R.; Zhang, M.; Yang, H.; et al. High mobility group 1 protein (HMG-1) stimulates proinflammatory cytokine synthesis in human monocytes. J Exp Med 2000, 192, 565–570. [CrossRef]
- Wang, H.; Bloom, O.; Zhang, M.; Vishnubhakat, J.M.; Ombrellino, M.; Che, J.; Frazier, A.; Yang, H.; Ivanova, S.; Borovikova, L.; et al. HMG-1 as a late mediator of endotoxin lethality in mice. Science 1999, 285, 248–251. [CrossRef]
- Shen, Z.; Liu, Z.; Wang, H.; Landrock, D.; Noh, J.Y.; Zang, Q.S.; Lee, C.H.; Farnell, Y.Z.; Chen, Z.; Sun, Y. Fructose induces inflammatory activation in macrophages and microglia through the nutrient-sensing ghrelin receptor. FASEB J 2025, 39, e70412. [CrossRef]
- Kim, D.M.; Lee, J.H.; Pan, Q.; Han, H.W.; Shen, Z.; Eshghjoo, S.; Wu, C.S.; Yang, W.; Noh, J.Y.; Threadgill, D.W.; et al. Nutrient-sensing growth hormone secretagogue receptor in macrophage programming and meta-inflammation. Mol Metab 2024, 79, 101852. [CrossRef]
- Yu, T.; Gao, M.; Yang, P.; Liu, D.; Wang, D.; Song, F.; Zhang, X.; Liu, Y. Insulin promotes macrophage phenotype transition through PI3K/Akt and PPAR-gamma signaling during diabetic wound healing. J Cell Physiol 2019, 234, 4217–4231. [CrossRef]
- Mauer, J.; Chaurasia, B.; Plum, L.; Quast, T.; Hampel, B.; Bluher, M.; Kolanus, W.; Kahn, C.R.; Bruning, J.C. Myeloid cell-restricted insulin receptor deficiency protects against obesity-induced inflammation and systemic insulin resistance. PLoS Genet 2010, 6, e1000938. [CrossRef]
- Baumgartl, J.; Baudler, S.; Scherner, M.; Babaev, V.; Makowski, L.; Suttles, J.; McDuffie, M.; Tobe, K.; Kadowaki, T.; Fazio, S.; et al. Myeloid lineage cell-restricted insulin resistance protects apolipoproteinE-deficient mice against atherosclerosis. Cell Metab 2006, 3, 247–256. [CrossRef]
- Knuever, J.; Willenborg, S.; Ding, X.; Akyuz, M.D.; Partridge, L.; Niessen, C.M.; Bruning, J.C.; Eming, S.A. Myeloid Cell-Restricted Insulin/IGF-1 Receptor Deficiency Protects against Skin Inflammation. J Immunol 2015, 195, 5296–5308. [CrossRef]
- Rached, M.T.; Millership, S.J.; Pedroni, S.M.A.; Choudhury, A.I.; Costa, A.S.H.; Hardy, D.G.; Glegola, J.A.; Irvine, E.E.; Selman, C.; Woodberry, M.C.; et al. Deletion of myeloid IRS2 enhances adipose tissue sympathetic nerve function and limits obesity. Mol Metab 2019, 20, 38–50. [CrossRef]
- Uysal, K.T.; Wiesbrock, S.M.; Marino, M.W.; Hotamisligil, G.S. Protection from obesity-induced insulin resistance in mice lacking TNF-alpha function. Nature 1997, 389, 610–614. [CrossRef]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J Clin Invest 2003, 112, 1821–1830. [CrossRef]
- McGarry, J.D. Banting lecture 2001: dysregulation of fatty acid metabolism in the etiology of type 2 diabetes. Diabetes 2002, 51, 7–18. [CrossRef]
- Sachithanandan, N.; Graham, K.L.; Galic, S.; Honeyman, J.E.; Fynch, S.L.; Hewitt, K.A.; Steinberg, G.R.; Kay, T.W. Macrophage deletion of SOCS1 increases sensitivity to LPS and palmitic acid and results in systemic inflammation and hepatic insulin resistance. Diabetes 2011, 60, 2023–2031. [CrossRef]
- Czaja, A.J.; Manns, M.P. Advances in the diagnosis, pathogenesis, and management of autoimmune hepatitis. Gastroenterology 2010, 139, 58–72 e54. [CrossRef]
- Weber, A.; Boege, Y.; Reisinger, F.; Heikenwalder, M. Chronic liver inflammation and hepatocellular carcinoma: persistence matters. Swiss Med Wkly 2011, 141, w13197. [CrossRef]
- Bhargava, P.; Lee, C.H. Role and function of macrophages in the metabolic syndrome. Biochem J 2012, 442, 253–262. [CrossRef]
- Wynn, T.A.; Barron, L. Macrophages: master regulators of inflammation and fibrosis. Semin Liver Dis 2010, 30, 245–257. [CrossRef]
- Li, C.; Xu, M.M.; Wang, K.; Adler, A.J.; Vella, A.T.; Zhou, B. Macrophage polarization and meta-inflammation. Transl Res 2018, 191, 29–44. [CrossRef]
- Hertzel, A.V.; Yong, J.; Chen, X.; Bernlohr, D.A. Immune Modulation of Adipocyte Mitochondrial Metabolism. Endocrinology 2022, 163. [CrossRef]
- Mills, C.D.; Kincaid, K.; Alt, J.M.; Heilman, M.J.; Hill, A.M. M-1/M-2 macrophages and the Th1/Th2 paradigm. J Immunol 2000, 164, 6166–6173. [CrossRef]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: time for reassessment. F1000Prime Rep 2014, 6, 13. [CrossRef]
- Nahrendorf, M.; Swirski, F.K. Abandoning M1/M2 for a Network Model of Macrophage Function. Circ Res 2016, 119, 414–417. [CrossRef]
- Bleriot, C.; Chakarov, S.; Ginhoux, F. Determinants of Resident Tissue Macrophage Identity and Function. Immunity 2020, 52, 957–970. [CrossRef]
- Mulder, K.; Patel, A.A.; Kong, W.T.; Piot, C.; Halitzki, E.; Dunsmore, G.; Khalilnezhad, S.; Irac, S.E.; Dubuisson, A.; Chevrier, M.; et al. Cross-tissue single-cell landscape of human monocytes and macrophages in health and disease. Immunity 2021, 54, 1883–1900 e1885. [CrossRef]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat Rev Immunol 2008, 8, 958–969. [CrossRef]
- Guilliams, M.; Thierry, G.R.; Bonnardel, J.; Bajenoff, M. Establishment and Maintenance of the Macrophage Niche. Immunity 2020, 52, 434–451. [CrossRef]
- Ma, R.Y.; Black, A.; Qian, B.Z. Macrophage diversity in cancer revisited in the era of single-cell omics. Trends Immunol 2022, 43, 546–563. [CrossRef]
- Tschop, M.; Lahner, H.; Feldmeier, H.; Grasberger, H.; Morrison, K.M.; Janssen, O.E.; Attanasio, A.F.; Strasburger, C.J. Effects of growth hormone replacement therapy on levels of cortisol and cortisol-binding globulin in hypopituitary adults. Eur J Endocrinol 2000, 143, 769–773. [CrossRef]
- Sun, Y.; Wang, P.; Zheng, H.; Smith, R.G. Ghrelin stimulation of growth hormone release and appetite is mediated through the growth hormone secretagogue receptor. Proc Natl Acad Sci U S A 2004, 101, 4679–4684. [CrossRef]
- Lin, L.; Nuotio-Antar, A.M.; Ma, X.; Liu, F.; Fiorotto, M.L.; Sun, Y. Ghrelin receptor regulates appetite and satiety during aging in mice by regulating meal frequency and portion size but not total food intake. J Nutr 2014, 144, 1349–1355. [CrossRef]
- Lin, L.; Saha, P.K.; Ma, X.; Henshaw, I.O.; Shao, L.; Chang, B.H.; Buras, E.D.; Tong, Q.; Chan, L.; McGuinness, O.P.; et al. Ablation of ghrelin receptor reduces adiposity and improves insulin sensitivity during aging by regulating fat metabolism in white and brown adipose tissues. Aging Cell 2011, 10, 996–1010. [CrossRef]
- Lin, L.; Lee, J.H.; Buras, E.D.; Yu, K.; Wang, R.; Smith, C.W.; Wu, H.; Sheikh-Hamad, D.; Sun, Y. Ghrelin receptor regulates adipose tissue inflammation in aging. Aging (Albany NY) 2016, 8, 178–191. [CrossRef]
- Ma, X.; Lin, L.; Yue, J.; Pradhan, G.; Qin, G.; Minze, L.J.; Wu, H.; Sheikh-Hamad, D.; Smith, C.W.; Sun, Y. Ghrelin receptor regulates HFCS-induced adipose inflammation and insulin resistance. Nutr Diabetes 2013, 3, e99. [CrossRef]
- Yuan, F.; Ma, J.; Xiang, X.; Lan, H.; Xu, Y.; Zhao, J.; Li, Y.; Zhang, W. Improvement of Adipose Macrophage Polarization in High Fat Diet-Induced Obese GHSR Knockout Mice. Biomed Res Int 2018, 2018, 4924325. [CrossRef]
- Holst, B.; Holliday, N.D.; Bach, A.; Elling, C.E.; Cox, H.M.; Schwartz, T.W. Common structural basis for constitutive activity of the ghrelin receptor family. J Biol Chem 2004, 279, 53806–53817. [CrossRef]
- Petersen, P.S.; Woldbye, D.P.; Madsen, A.N.; Egerod, K.L.; Jin, C.; Lang, M.; Rasmussen, M.; Beck-Sickinger, A.G.; Holst, B. In vivo characterization of high Basal signaling from the ghrelin receptor. Endocrinology 2009, 150, 4920–4930. [CrossRef]
- Tanida, R.; Tsubouchi, H.; Yanagi, S.; Saito, Y.; Toshinai, K.; Miyazaki, T.; Takamura, T.; Nakazato, M. GHS-R1a deficiency mitigates lipopolysaccharide-induced lung injury in mice via the downregulation of macrophage activity. Biochem Biophys Res Commun 2022, 589, 260–266. [CrossRef]
- Noh, J.Y.; Herrera, M.; Patil, B.S.; Tan, X.D.; Wright, G.A.; Sun, Y. The expression and function of growth hormone secretagogue receptor in immune cells: A current perspective. Exp Biol Med (Maywood) 2022, 247, 2184–2191. [CrossRef]
- Yu, J.H.; Kim, M.S. Molecular mechanisms of appetite regulation. Diabetes Metab J 2012, 36, 391–398. [CrossRef]
- Guan, X.M.; Yu, H.; Palyha, O.C.; McKee, K.K.; Feighner, S.D.; Sirinathsinghji, D.J.; Smith, R.G.; Van der Ploeg, L.H.; Howard, A.D. Distribution of mRNA encoding the growth hormone secretagogue receptor in brain and peripheral tissues. Brain Res Mol Brain Res 1997, 48, 23–29. [CrossRef]
- Schellekens, H.; Dinan, T.G.; Cryan, J.F. Lean mean fat reducing “ghrelin” machine: hypothalamic ghrelin and ghrelin receptors as therapeutic targets in obesity. Neuropharmacology 2010, 58, 2–16. [CrossRef]
- Lim, C.T.; Kola, B.; Korbonits, M. The ghrelin/GOAT/GHS-R system and energy metabolism. Rev Endocr Metab Disord 2011, 12, 173–186. [CrossRef]
- Gnanapavan, S.; Kola, B.; Bustin, S.A.; Morris, D.G.; McGee, P.; Fairclough, P.; Bhattacharya, S.; Carpenter, R.; Grossman, A.B.; Korbonits, M. The tissue distribution of the mRNA of ghrelin and subtypes of its receptor, GHS-R, in humans. J Clin Endocrinol Metab 2002, 87, 2988. [CrossRef]
- Jiao, Z.T.; Luo, Q. Molecular Mechanisms and Health Benefits of Ghrelin: A Narrative Review. Nutrients 2022, 14. [CrossRef]
- Lam, T.K.; Schwartz, G.J.; Rossetti, L. Hypothalamic sensing of fatty acids. Nat Neurosci 2005, 8, 579–584. [CrossRef]
- He, W.; Lam, T.K.; Obici, S.; Rossetti, L. Molecular disruption of hypothalamic nutrient sensing induces obesity. Nat Neurosci 2006, 9, 227–233. [CrossRef]
- Obici, S.; Feng, Z.; Arduini, A.; Conti, R.; Rossetti, L. Inhibition of hypothalamic carnitine palmitoyltransferase-1 decreases food intake and glucose production. Nat Med 2003, 9, 756–761. [CrossRef]
- Lee, K.; Kerner, J.; Hoppel, C.L. Mitochondrial carnitine palmitoyltransferase 1a (CPT1a) is part of an outer membrane fatty acid transfer complex. J Biol Chem 2011, 286, 25655–25662. [CrossRef]
- Schlaepfer, I.R.; Joshi, M. CPT1A-mediated Fat Oxidation, Mechanisms, and Therapeutic Potential. Endocrinology 2020, 161. [CrossRef]
- Andrews, Z.B.; Liu, Z.W.; Walllingford, N.; Erion, D.M.; Borok, E.; Friedman, J.M.; Tschop, M.H.; Shanabrough, M.; Cline, G.; Shulman, G.I.; et al. UCP2 mediates ghrelin’s action on NPY/AgRP neurons by lowering free radicals. Nature 2008, 454, 846–851. [CrossRef]
- Chen, H.Y.; Trumbauer, M.E.; Chen, A.S.; Weingarth, D.T.; Adams, J.R.; Frazier, E.G.; Shen, Z.; Marsh, D.J.; Feighner, S.D.; Guan, X.M.; et al. Orexigenic action of peripheral ghrelin is mediated by neuropeptide Y and agouti-related protein. Endocrinology 2004, 145, 2607–2612. [CrossRef]
- Horvath, T.L.; Naftolin, F.; Kalra, S.P.; Leranth, C. Neuropeptide-Y innervation of beta-endorphin-containing cells in the rat mediobasal hypothalamus: a light and electron microscopic double immunostaining analysis. Endocrinology 1992, 131, 2461–2467. [CrossRef]
- Lopez, N.E.; Krzyzaniak, M.J.; Blow, C.; Putnam, J.; Ortiz-Pomales, Y.; Hageny, A.M.; Eliceiri, B.; Coimbra, R.; Bansal, V. Ghrelin prevents disruption of the blood-brain barrier after traumatic brain injury. J Neurotrauma 2012, 29, 385–393. [CrossRef]
- Raghay, K.; Akki, R.; Bensaid, D.; Errami, M. Ghrelin as an anti-inflammatory and protective agent in ischemia/reperfusion injury. Peptides 2020, 124, 170226. [CrossRef]
- Ku, J.M.; Taher, M.; Chin, K.Y.; Barsby, T.; Austin, V.; Wong, C.H.; Andrews, Z.B.; Spencer, S.J.; Miller, A.A. Protective actions of des-acylated ghrelin on brain injury and blood-brain barrier disruption after stroke in mice. Clin Sci (Lond) 2016, 130, 1545–1558. [CrossRef]
- Miao, Y.; Xia, Q.; Hou, Z.; Zheng, Y.; Pan, H.; Zhu, S. Ghrelin protects cortical neuron against focal ischemia/reperfusion in rats. Biochem Biophys Res Commun 2007, 359, 795–800. [CrossRef]
- Shao, X.; Hu, Q.; Chen, S.; Wang, Q.; Xu, P.; Jiang, X. Ghrelin Ameliorates Traumatic Brain Injury by Down-Regulating bFGF and FGF-BP. Front Neurosci 2018, 12, 445. [CrossRef]
- Sun, N.; Wang, H.; Ma, L.; Lei, P.; Zhang, Q. Ghrelin attenuates brain injury in septic mice via PI3K/Akt signaling activation. Brain Res Bull 2016, 124, 278–285. [CrossRef]
- Chen, L.; Wei, X.; Hou, Y.; Liu, X.; Li, S.; Sun, B.; Liu, X.; Liu, H. Tetramethylpyrazine analogue CXC195 protects against cerebral ischemia/reperfusion-induced apoptosis through PI3K/Akt/GSK3beta pathway in rats. Neurochem Int 2014, 66, 27–32. [CrossRef]
- Hong, Y.; Shao, A.; Wang, J.; Chen, S.; Wu, H.; McBride, D.W.; Wu, Q.; Sun, X.; Zhang, J. Neuroprotective effect of hydrogen-rich saline against neurologic damage and apoptosis in early brain injury following subarachnoid hemorrhage: possible role of the Akt/GSK3beta signaling pathway. PLoS One 2014, 9, e96212. [CrossRef]
- Mao, Q.; Wang, J.; Yang, Z.; Ding, R.; Lv, S.; Ji, X. The Pathologic Roles and Therapeutic Implications of Ghrelin/GHSR System in Mental Disorders. Depress Anxiety 2024, 2024, 5537319. [CrossRef]
- Lutter, M.; Sakata, I.; Osborne-Lawrence, S.; Rovinsky, S.A.; Anderson, J.G.; Jung, S.; Birnbaum, S.; Yanagisawa, M.; Elmquist, J.K.; Nestler, E.J.; et al. The orexigenic hormone ghrelin defends against depressive symptoms of chronic stress. Nat Neurosci 2008, 11, 752–753. [CrossRef]
- Nakashima, K.; Akiyoshi, J.; Hatano, K.; Hanada, H.; Tanaka, Y.; Tsuru, J.; Matsushita, H.; Kodama, K.; Isogawa, K. Ghrelin gene polymorphism is associated with depression, but not panic disorder. Psychiatr Genet 2008, 18, 257. [CrossRef]
- Yohn, C.N.; Gergues, M.M.; Samuels, B.A. The role of 5-HT receptors in depression. Mol Brain 2017, 10, 28. [CrossRef]
- Samuels, B.A.; Leonardo, E.D.; Gadient, R.; Williams, A.; Zhou, J.; David, D.J.; Gardier, A.M.; Wong, E.H.; Hen, R. Modeling treatment-resistant depression. Neuropharmacology 2011, 61, 408–413. [CrossRef]
- Shao, X.; Zhu, G. Associations Among Monoamine Neurotransmitter Pathways, Personality Traits, and Major Depressive Disorder. Front Psychiatry 2020, 11, 381. [CrossRef]
- Keller, J.; Gomez, R.; Williams, G.; Lembke, A.; Lazzeroni, L.; Murphy, G.M., Jr.; Schatzberg, A.F. HPA axis in major depression: cortisol, clinical symptomatology and genetic variation predict cognition. Mol Psychiatry 2017, 22, 527–536. [CrossRef]
- Duman, R.S.; Sanacora, G.; Krystal, J.H. Altered Connectivity in Depression: GABA and Glutamate Neurotransmitter Deficits and Reversal by Novel Treatments. Neuron 2019, 102, 75–90. [CrossRef]
- Tian, L.; Ma, L.; Kaarela, T.; Li, Z. Neuroimmune crosstalk in the central nervous system and its significance for neurological diseases. J Neuroinflammation 2012, 9, 155. [CrossRef]
- Vreeburg, S.A.; Hoogendijk, W.J.; van Pelt, J.; Derijk, R.H.; Verhagen, J.C.; van Dyck, R.; Smit, J.H.; Zitman, F.G.; Penninx, B.W. Major depressive disorder and hypothalamic-pituitary-adrenal axis activity: results from a large cohort study. Arch Gen Psychiatry 2009, 66, 617–626. [CrossRef]
- Vreeburg, S.A.; Zitman, F.G.; van Pelt, J.; Derijk, R.H.; Verhagen, J.C.; van Dyck, R.; Hoogendijk, W.J.; Smit, J.H.; Penninx, B.W. Salivary cortisol levels in persons with and without different anxiety disorders. Psychosom Med 2010, 72, 340–347. [CrossRef]
- Kluge, M.; Schussler, P.; Dresler, M.; Schmidt, D.; Yassouridis, A.; Uhr, M.; Steiger, A. Effects of ghrelin on psychopathology, sleep and secretion of cortisol and growth hormone in patients with major depression. J Psychiatr Res 2011, 45, 421–426. [CrossRef]
- Spencer, S.J.; Xu, L.; Clarke, M.A.; Lemus, M.; Reichenbach, A.; Geenen, B.; Kozicz, T.; Andrews, Z.B. Ghrelin regulates the hypothalamic-pituitary-adrenal axis and restricts anxiety after acute stress. Biol Psychiatry 2012, 72, 457–465. [CrossRef]
- Rucinski, M.; Ziolkowska, A.; Szyszka, M.; Hochol, A.; Malendowicz, L.K. Evidence suggesting that ghrelin O-acyl transferase inhibitor acts at the hypothalamus to inhibit hypothalamo-pituitary-adrenocortical axis function in the rat. Peptides 2012, 35, 149–159. [CrossRef]
- Kluge, M.; Gazea, M.; Schussler, P.; Genzel, L.; Dresler, M.; Kleyer, S.; Uhr, M.; Yassouridis, A.; Steiger, A. Ghrelin increases slow wave sleep and stage 2 sleep and decreases stage 1 sleep and REM sleep in elderly men but does not affect sleep in elderly women. Psychoneuroendocrinology 2010, 35, 297–304. [CrossRef]
- Olivier, N.; Harvey, B.H.; Gobec, S.; Shahid, M.; Kosak, U.; Zakelj, S.; Brink, C.B. A novel butyrylcholinesterase inhibitor induces antidepressant, pro-cognitive, and anti-anhedonic effects in Flinders Sensitive Line rats: The role of the ghrelin-dopamine cascade. Biomed Pharmacother 2025, 187, 118093. [CrossRef]
- Belujon, P.; Grace, A.A. Dopamine System Dysregulation in Major Depressive Disorders. Int J Neuropsychopharmacol 2017, 20, 1036–1046. [CrossRef]
- Mifune, H.; Tajiri, Y.; Sakai, Y.; Kawahara, Y.; Hara, K.; Sato, T.; Nishi, Y.; Nishi, A.; Mitsuzono, R.; Kakuma, T.; et al. Voluntary exercise is motivated by ghrelin, possibly related to the central reward circuit. J Endocrinol 2020, 244, 123–132. [CrossRef]
- Abizaid, A. Ghrelin and dopamine: new insights on the peripheral regulation of appetite. J Neuroendocrinol 2009, 21, 787–793. [CrossRef]
- Abizaid, A.; Liu, Z.W.; Andrews, Z.B.; Shanabrough, M.; Borok, E.; Elsworth, J.D.; Roth, R.H.; Sleeman, M.W.; Picciotto, M.R.; Tschop, M.H.; et al. Ghrelin modulates the activity and synaptic input organization of midbrain dopamine neurons while promoting appetite. J Clin Invest 2006, 116, 3229–3239. [CrossRef]
- Jerlhag, E.; Egecioglu, E.; Dickson, S.L.; Andersson, M.; Svensson, L.; Engel, J.A. Ghrelin stimulates locomotor activity and accumbal dopamine-overflow via central cholinergic systems in mice: implications for its involvement in brain reward. Addict Biol 2006, 11, 45–54. [CrossRef]
- Edvardsson, C.E.; Vestlund, J.; Jerlhag, E. A ghrelin receptor antagonist reduces the ability of ghrelin, alcohol or amphetamine to induce a dopamine release in the ventral tegmental area and in nucleus accumbens shell in rats. Eur J Pharmacol 2021, 899, 174039. [CrossRef]
- Ghersi, M.S.; Casas, S.M.; Escudero, C.; Carlini, V.P.; Buteler, F.; Cabrera, R.J.; Schioth, H.B.; de Barioglio, S.R. Ghrelin inhibited serotonin release from hippocampal slices. Peptides 2011, 32, 2367–2371. [CrossRef]
- Cerit, H.; Christensen, K.; Moondra, P.; Klibanski, A.; Goldstein, J.M.; Holsen, L.M. Divergent associations between ghrelin and neural responsivity to palatable food in hyperphagic and hypophagic depression. J Affect Disord 2019, 242, 29–38. [CrossRef]
- Mequinion, M.; Foldi, C.J.; Andrews, Z.B. The Ghrelin-AgRP Neuron Nexus in Anorexia Nervosa: Implications for Metabolic and Behavioral Adaptations. Front Nutr 2019, 6, 190. [CrossRef]
- Dehkhoda, F.; Ringuet, M.T.; Whitfield, E.A.; Mutunduwe, K.; Whelan, F.; Nowell, C.J.; Misganaw, D.; Xu, Z.; Piper, N.B.C.; Clark, R.J.; et al. Constitutive ghrelin receptor activity enables reversal of dopamine D2 receptor signaling. Mol Cell 2025, 85, 2246–2260 e2210. [CrossRef]
- Furness, J.B.; Hunne, B.; Matsuda, N.; Yin, L.; Russo, D.; Kato, I.; Fujimiya, M.; Patterson, M.; McLeod, J.; Andrews, Z.B.; et al. Investigation of the presence of ghrelin in the central nervous system of the rat and mouse. Neuroscience 2011, 193, 1–9. [CrossRef]
- Jensen, M.; Ratner, C.; Rudenko, O.; Christiansen, S.H.; Skov, L.J.; Hundahl, C.; Woldbye, D.P.; Holst, B. Anxiolytic-Like Effects of Increased Ghrelin Receptor Signaling in the Amygdala. Int J Neuropsychopharmacol 2016, 19. [CrossRef]
- Espelund, U.; Hansen, T.K.; Hojlund, K.; Beck-Nielsen, H.; Clausen, J.T.; Hansen, B.S.; Orskov, H.; Jorgensen, J.O.; Frystyk, J. Fasting unmasks a strong inverse association between ghrelin and cortisol in serum: studies in obese and normal-weight subjects. J Clin Endocrinol Metab 2005, 90, 741–746. [CrossRef]
- Schuessler, P.; Uhr, M.; Ising, M.; Schmid, D.; Weikel, J.; Steiger, A. Nocturnal ghrelin levels--relationship to sleep EEG, the levels of growth hormone, ACTH and cortisol--and gender differences. J Sleep Res 2005, 14, 329–336. [CrossRef]
- Buntwal, L.; Sassi, M.; Morgan, A.H.; Andrews, Z.B.; Davies, J.S. Ghrelin-Mediated Hippocampal Neurogenesis: Implications for Health and Disease. Trends Endocrinol Metab 2019, 30, 844–859. [CrossRef]
- Numakawa, T.; Odaka, H.; Adachi, N. Actions of Brain-Derived Neurotrophic Factor and Glucocorticoid Stress in Neurogenesis. Int J Mol Sci 2017, 18. [CrossRef]
- Price, M.L.; Ley, C.D.; Gorvin, C.M. The emerging role of heterodimerisation and interacting proteins in ghrelin receptor function. J Endocrinol 2021, 252, R23–R39. [CrossRef]
- Kern, A.; Mavrikaki, M.; Ullrich, C.; Albarran-Zeckler, R.; Brantley, A.F.; Smith, R.G. Hippocampal Dopamine/DRD1 Signaling Dependent on the Ghrelin Receptor. Cell 2015, 163, 1176–1190. [CrossRef]
- Ricken, R.; Bopp, S.; Schlattmann, P.; Himmerich, H.; Bschor, T.; Richter, C.; Elstner, S.; Stamm, T.J.; Schulz-Ratei, B.; Lingesleben, A.; et al. Ghrelin Serum Concentrations Are Associated with Treatment Response During Lithium Augmentation of Antidepressants. Int J Neuropsychopharmacol 2017, 20, 692–697. [CrossRef]
- Huang, K.L.; Chen, M.H.; Hsu, J.W.; Tsai, S.J.; Bai, Y.M. Using classification and regression tree modeling to investigate appetite hormones and proinflammatory cytokines as biomarkers to differentiate bipolar I depression from major depressive disorder. CNS Spectr 2021, 1–7. [CrossRef]
- Abdel Aziz, K.; Al-Mugaddam, F.; Sugathan, S.; Saseedharan, P.; Jouini, T.; Elamin, M.E.; Moselhy, H.; Aly El-Gabry, D.; Arnone, D.; Karam, S.M. Decreased acylated and total ghrelin levels in bipolar disorder patients recovering from a manic episode. BMC Psychiatry 2022, 22, 209. [CrossRef]
- Theodoropoulou, A.; Metallinos, I.C.; Psyrogiannis, A.; Vagenakis, G.A.; Kyriazopoulou, V. Ghrelin and leptin secretion in patients with moderate Alzheimer’s disease. J Nutr Health Aging 2012, 16, 472–477. [CrossRef]
- Cao, X.; Zhu, M.; He, Y.; Chu, W.; Du, Y.; Du, H. Increased Serum Acylated Ghrelin Levels in Patients with Mild Cognitive Impairment. J Alzheimers Dis 2018, 61, 545–552. [CrossRef]
- Yoshino, Y.; Funahashi, Y.; Nakata, S.; Ozaki, Y.; Yamazaki, K.; Yoshida, T.; Mori, T.; Mori, Y.; Ochi, S.; Iga, J.I.; et al. Ghrelin cascade changes in the peripheral blood of Japanese patients with Alzheimer’s disease. J Psychiatr Res 2018, 107, 79–85. [CrossRef]
- Gahete, M.D.; Rubio, A.; Cordoba-Chacon, J.; Gracia-Navarro, F.; Kineman, R.D.; Avila, J.; Luque, R.M.; Castano, J.P. Expression of the ghrelin and neurotensin systems is altered in the temporal lobe of Alzheimer’s disease patients. J Alzheimers Dis 2010, 22, 819–828. [CrossRef]
- Tian, J.; Guo, L.; Sui, S.; Driskill, C.; Phensy, A.; Wang, Q.; Gauba, E.; Zigman, J.M.; Swerdlow, R.H.; Kroener, S.; et al. Disrupted hippocampal growth hormone secretagogue receptor 1alpha interaction with dopamine receptor D1 plays a role in Alzheimer’s disease. Sci Transl Med 2019, 11. [CrossRef]
- Smith, R.G.; Van der Ploeg, L.H.; Howard, A.D.; Feighner, S.D.; Cheng, K.; Hickey, G.J.; Wyvratt, M.J., Jr.; Fisher, M.H.; Nargund, R.P.; Patchett, A.A. Peptidomimetic regulation of growth hormone secretion. Endocr Rev 1997, 18, 621–645. [CrossRef]
- M’Kadmi, C.; Cabral, A.; Barrile, F.; Giribaldi, J.; Cantel, S.; Damian, M.; Mary, S.; Denoyelle, S.; Dutertre, S.; Peraldi-Roux, S.; et al. N-Terminal Liver-Expressed Antimicrobial Peptide 2 (LEAP2) Region Exhibits Inverse Agonist Activity toward the Ghrelin Receptor. J Med Chem 2019, 62, 965–973. [CrossRef]
- Ebrahimpour, S.; Zakeri, M.; Esmaeili, A. Crosstalk between obesity, diabetes, and alzheimer’s disease: Introducing quercetin as an effective triple herbal medicine. Ageing Res Rev 2020, 62, 101095. [CrossRef]
- Popelova, A.; Kakonova, A.; Hruba, L.; Kunes, J.; Maletinska, L.; Zelezna, B. Potential neuroprotective and anti-apoptotic properties of a long-lasting stable analog of ghrelin: an in vitro study using SH-SY5Y cells. Physiol Res 2018, 67, 339–346. [CrossRef]
- Mengr, A.; Smotkova, Z.; Pacesova, A.; Zelezna, B.; Kunes, J.; Maletinska, L. Reduction of Neuroinflammation as a Common Mechanism of Action of Anorexigenic and Orexigenic Peptide Analogues in the Triple Transgenic Mouse Model of Alzheimer s Disease. J Neuroimmune Pharmacol 2025, 20, 18. [CrossRef]
- Zhao, T.J.; Liang, G.; Li, R.L.; Xie, X.; Sleeman, M.W.; Murphy, A.J.; Valenzuela, D.M.; Yancopoulos, G.D.; Goldstein, J.L.; Brown, M.S. Ghrelin O-acyltransferase (GOAT) is essential for growth hormone-mediated survival of calorie-restricted mice. Proc Natl Acad Sci U S A 2010, 107, 7467–7472. [CrossRef]
- Rasineni, K.; Thomes, P.G.; Kubik, J.L.; Harris, E.N.; Kharbanda, K.K.; Casey, C.A. Chronic alcohol exposure alters circulating insulin and ghrelin levels: role of ghrelin in hepatic steatosis. Am J Physiol Gastrointest Liver Physiol 2019, 316, G453–G461. [CrossRef]
- Landgren, S.; Engel, J.A.; Hyytia, P.; Zetterberg, H.; Blennow, K.; Jerlhag, E. Expression of the gene encoding the ghrelin receptor in rats selected for differential alcohol preference. Behav Brain Res 2011, 221, 182–188. [CrossRef]
- Ryabinin, A.E.; Cocking, D.L.; Kaur, S. Inhibition of VTA neurons activates the centrally projecting Edinger-Westphal nucleus: evidence of a stress-reward link? J Chem Neuroanat 2013, 54, 57–61. [CrossRef]
- Larsson, A.; Engel, J.A. Neurochemical and behavioral studies on ethanol and nicotine interactions. Neurosci Biobehav Rev 2004, 27, 713–720. [CrossRef]
- Suchankova, P.; Steensland, P.; Fredriksson, I.; Engel, J.A.; Jerlhag, E. Ghrelin receptor (GHS-R1A) antagonism suppresses both alcohol consumption and the alcohol deprivation effect in rats following long-term voluntary alcohol consumption. PLoS One 2013, 8, e71284. [CrossRef]
- Jerlhag, E.; Landgren, S.; Egecioglu, E.; Dickson, S.L.; Engel, J.A. The alcohol-induced locomotor stimulation and accumbal dopamine release is suppressed in ghrelin knockout mice. Alcohol 2011, 45, 341–347. [CrossRef]
- Loonen, A.J.; Ivanova, S.A. Circuits Regulating Pleasure and Happiness: The Evolution of the Amygdalar-Hippocampal-Habenular Connectivity in Vertebrates. Front Neurosci 2016, 10, 539. [CrossRef]
- Jerlhag, E.; Egecioglu, E.; Landgren, S.; Salome, N.; Heilig, M.; Moechars, D.; Datta, R.; Perrissoud, D.; Dickson, S.L.; Engel, J.A. Requirement of central ghrelin signaling for alcohol reward. Proc Natl Acad Sci U S A 2009, 106, 11318–11323. [CrossRef]
- Zallar, L.J.; Beurmann, S.; Tunstall, B.J.; Fraser, C.M.; Koob, G.F.; Vendruscolo, L.F.; Leggio, L. Ghrelin receptor deletion reduces binge-like alcohol drinking in rats. J Neuroendocrinol 2019, 31, e12663. [CrossRef]
- Yoshimoto, K.; Nagao, M.; Watanabe, Y.; Yamaguchi, T.; Ueda, S.; Kitamura, Y.; Nishimura, K.; Inden, M.; Marunaka, Y.; Hattori, H.; et al. Enhanced alcohol-drinking behavior associated with active ghrelinergic and serotoninergic neurons in the lateral hypothalamus and amygdala. Pharmacol Biochem Behav 2017, 153, 1–11. [CrossRef]
- Mustafa, E.R.; Cordisco Gonzalez, S.; Damian, M.; Cantel, S.; Denoyelle, S.; Wagner, R.; Schioth, H.B.; Fehrentz, J.A.; Baneres, J.L.; Perello, M.; et al. LEAP2 Impairs the Capability of the Growth Hormone Secretagogue Receptor to Regulate the Dopamine 2 Receptor Signaling. Front Pharmacol 2021, 12, 712437. [CrossRef]
- Jerlhag, E.; Egecioglu, E.; Dickson, S.L.; Engel, J.A. Ghrelin receptor antagonism attenuates cocaine- and amphetamine-induced locomotor stimulation, accumbal dopamine release, and conditioned place preference. Psychopharmacology (Berl) 2010, 211, 415–422. [CrossRef]
- Palotai, M.; Bagosi, Z.; Jaszberenyi, M.; Csabafi, K.; Dochnal, R.; Manczinger, M.; Telegdy, G.; Szabo, G. Ghrelin amplifies the nicotine-induced dopamine release in the rat striatum. Neurochem Int 2013, 63, 239–243. [CrossRef]
- Jerabek, P.; Havlickova, T.; Puskina, N.; Charalambous, C.; Lapka, M.; Kacer, P.; Sustkova-Fiserova, M. Ghrelin receptor antagonism of morphine-induced conditioned place preference and behavioral and accumbens dopaminergic sensitization in rats. Neurochem Int 2017, 110, 101–113. [CrossRef]
- Sustkova-Fiserova, M.; Puskina, N.; Havlickova, T.; Lapka, M.; Syslova, K.; Pohorala, V.; Charalambous, C. Ghrelin receptor antagonism of fentanyl-induced conditioned place preference, intravenous self-administration, and dopamine release in the nucleus accumbens in rats. Addict Biol 2020, 25, e12845. [CrossRef]
- You, Z.B.; Galaj, E.; Alen, F.; Wang, B.; Bi, G.H.; Moore, A.R.; Buck, T.; Crissman, M.; Pari, S.; Xi, Z.X.; et al. Involvement of the ghrelin system in the maintenance and reinstatement of cocaine-motivated behaviors: a role of adrenergic action at peripheral beta1 receptors. Neuropsychopharmacology 2022, 47, 1449–1460. [CrossRef]
- Charalambous, C.; Havlickova, T.; Lapka, M.; Puskina, N.; Slamberova, R.; Kuchar, M.; Sustkova-Fiserova, M. Cannabinoid-Induced Conditioned Place Preference, Intravenous Self-Administration, and Behavioral Stimulation Influenced by Ghrelin Receptor Antagonism in Rats. Int J Mol Sci 2021, 22. [CrossRef]
Figure 1.
Intracellular processing of ghrelin. The human GHRL gene resides on the short arm of chromosome 3p25-26 and consists of six exons. The ghrelin gene undergoes transcription and splicing to generate mature mRNA. This mRNA is translated into pre-proghrelin, which then undergoes translation to yield a 117 amino acid peptide. Within the ER, this precursor undergoes a systematic cleavage process to produce the mature 28 amino acid form of ghrelin. Before secretion, unmature non-acylated ghrelin undergoes acylation by ghrelin-O-acyltransferase (GOAT) in the presence of CoA as an acyl donor, facilitating its binding to the growth hormone secretagogue receptor 1a (GHSR1a). The highlighted box indicates the minimal active n-terminal pentapeptide core of the ghrelin sequence necessary for GHSR1a activation. Created with BioRender.com.
Figure 1.
Intracellular processing of ghrelin. The human GHRL gene resides on the short arm of chromosome 3p25-26 and consists of six exons. The ghrelin gene undergoes transcription and splicing to generate mature mRNA. This mRNA is translated into pre-proghrelin, which then undergoes translation to yield a 117 amino acid peptide. Within the ER, this precursor undergoes a systematic cleavage process to produce the mature 28 amino acid form of ghrelin. Before secretion, unmature non-acylated ghrelin undergoes acylation by ghrelin-O-acyltransferase (GOAT) in the presence of CoA as an acyl donor, facilitating its binding to the growth hormone secretagogue receptor 1a (GHSR1a). The highlighted box indicates the minimal active n-terminal pentapeptide core of the ghrelin sequence necessary for GHSR1a activation. Created with BioRender.com.
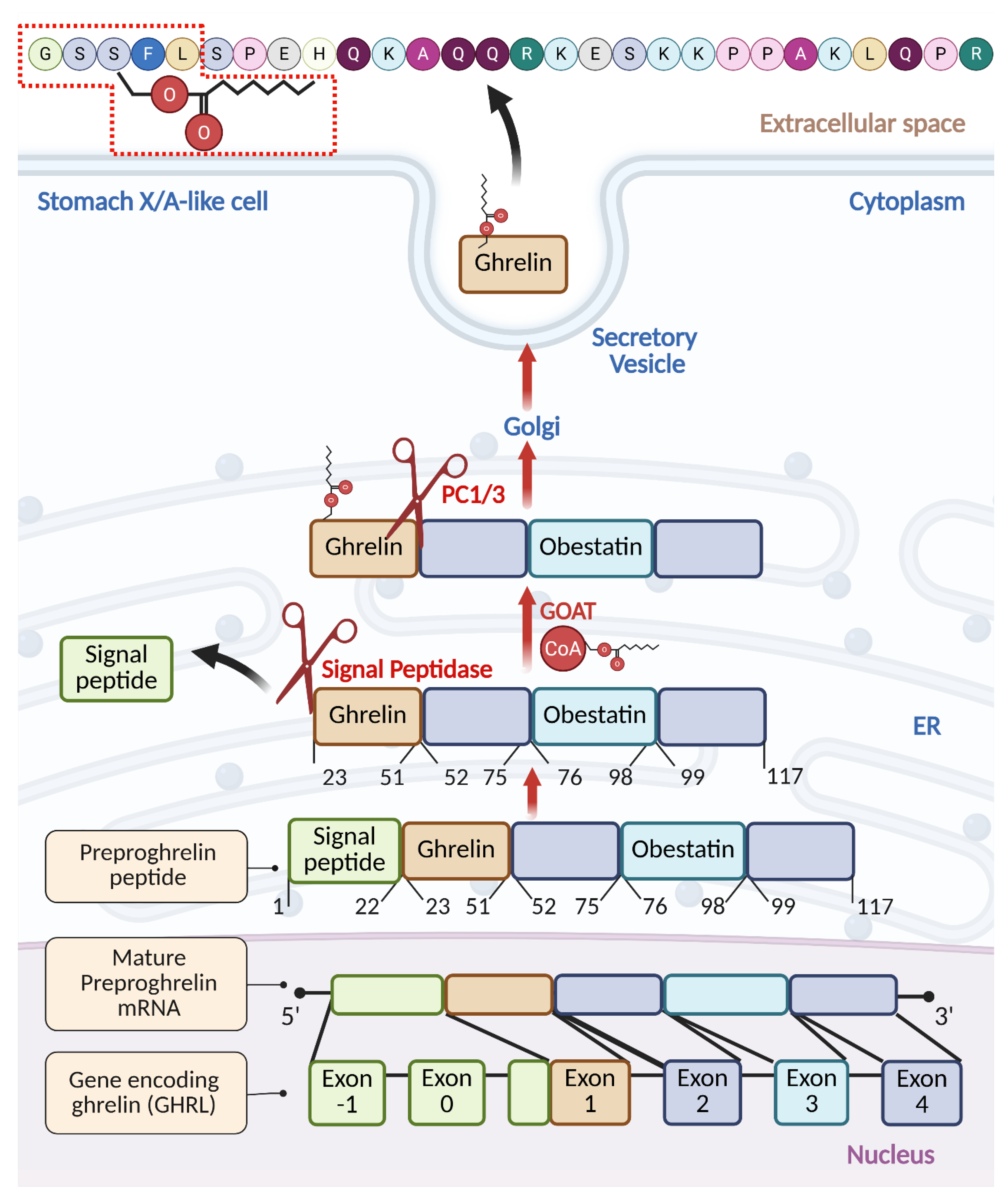
Figure 2.
The functionality profile of ghrelin. Schematic illustration of the hormonal actions of ghrelin in different target organs and tissues. The red arrows indicate the decrease, while blue arrows show the increase. Created with BioRender.com.
Figure 2.
The functionality profile of ghrelin. Schematic illustration of the hormonal actions of ghrelin in different target organs and tissues. The red arrows indicate the decrease, while blue arrows show the increase. Created with BioRender.com.
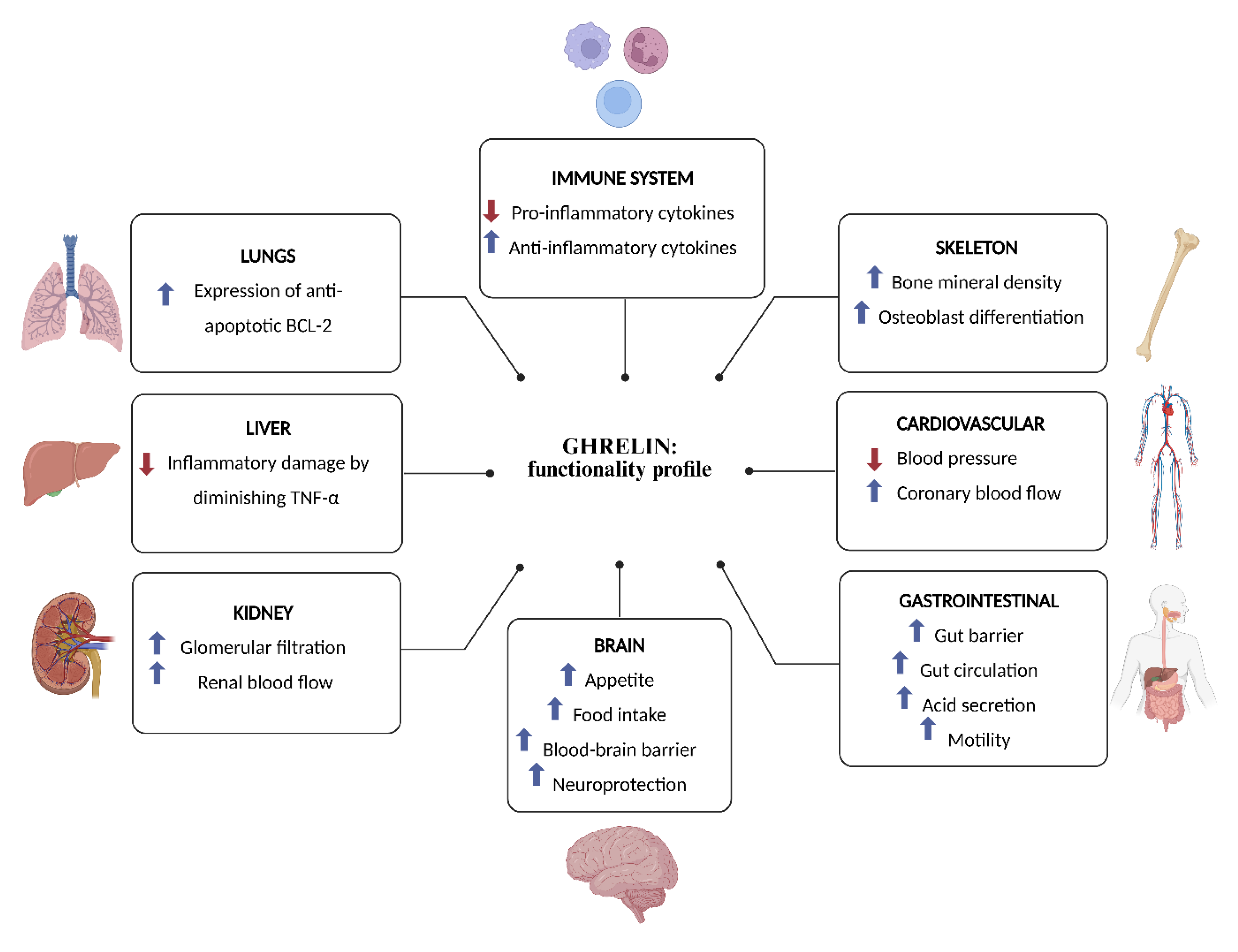
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



