Submitted:
02 October 2025
Posted:
03 October 2025
You are already at the latest version
Abstract
The therapeutic clotting factor VIII (FVIII) is known for its particular immunogenicity, with nearly 30% of the hemophilic patients developing neutralizing antibodies against the infused protein. The root cause of this immunogenicity is still not well understood, but intrinsic factors, such as FVIII by-products, have been linked to the immunological response elicited. Bioengineering of the FVIII molecule has been improving its recombinant (rhFVIII) production in many aspects, mainly enhancing its expression and stability. Assessment of immunogenicity for novel recombinant isoforms is crucial for further development and scaling-up processes, particularly due to the unpredictable antigenic properties and their impact on neutralizing antibody formation. In the present study, we describe a bioengineered human recombinant FVIII (rhFVIII-H6A), which induces lower immunogenicity in a murine model of hemophilia A. The rhFVIII-H6A product is characterized by a B domain-deleted heavy chain (HCh), with the C-terminal of the B domain fused to the light chain (BΔ-LCh). Compared to plasma-derived FVIII (pdFVIII) and rhFVIII reference products, treating hemophilic mice with rhFVIII-H6A induced lower levels of anti-FVIII antibody formation, including those with inhibitory neutralizing activity, while no difference was observed in the functional activity of rhFVIII-H6A in reverting the in vivo hemophilia phenotype. In addition, our results indicate that deleting the major part of the B domain from the HCh might lower the immunogenicity of novel rhFVIII products.
Keywords:
factor VIII
; procoagulant factor
; immunogenicity
; bioengineering
; recombinant
1. Introduction
Factor VIII (FVIII) replacement therapy employs either FVIII concentrates from blood plasma or, alternatively, recombinant human FVIII, which is the standard of care for managing hemophilia A, a genetic bleeding disorder caused by plasmatic deficiency of this procoagulant factor. Recombinant human FVIII (rhFVIII), especially plasma/albumin-free third-generation rhFVIII products, represents a safer alternative to plasma-derived products due to a reduced risk of transmission of blood-borne pathogens. High costs are associated with rhFVIII production, since the expression yields are approximately 2 to 3 orders of magnitude lower than those achieved with recombinant proteins of similar sizes, such as the recombinant human Factor V [1]. In addition, FVIII is known for its particular immunogenicity, with 25-35% of the hemophilic patients developing neutralizing alloantibodies against the infused FVIII [2,3]. Alloantibodies formation directly interferes with the safety and efficacy of therapeutic products, affecting their toxicity, pharmacokinetics and pharmacodynamics parameters. The risk of alloantibodies development and the severity of the clinical effects vary among therapeutic proteins, examples including insulin, growth hormone, granulocyte-macrophage colony-stimulating factor (GM-CSF), erythropoietin and interferons [4]. FVIII alloantibodies neutralize the FVIII functional activity by affecting the pharmacokinetic parameters, completely inhibiting drug activity. The alloantibodies to therapeutic factor human VIII in hemophilia A represent the major complication associated with FVIII replacement therapy [5,6]. Accordingly, therapeutic bioengineered molecules have been subjected to strict regulatory demands, among which immunogenicity studies, and these molecules have been strategically assessed during their different product development stages. Genetically modified animal models have been useful for the evaluation of the immunogenic potential of several biopharmaceuticals in pre-clinical trials [7]. Hemophilia A mice, produced by targeted disruption of the fviii gene, have been used as a model system to study the immunogenicity of human FVIII [8]. After serial intravenous injections of human FVIII using a dose per body mass similar to that of humans, hemophilia A mice develop inhibitors [9]. The inhibitor response in hemophilia A mice includes antibodies that recognize the immune-dominant Arg484-Ile508 A2 epitope, which is recognized by human hemophilia A patients [10,11], although the extent to which other B cell epitopes in the murine model and human hemophilia are similar is still unknown. Overall, hemophilia A mice constitute a powerful model system to study rhFVIII immunogenicity and dissect the eventual immune responses. In the present study, we highlight the assessment of immunogenicity during the development of a B-domain deleted (BDD) recombinant FVIII product.
2. Results
2.1. Production of the Bioengineered Factor VIII (rhFVIII-H6A) Presenting an Artificial Light Chain (LCh) Fused to Part of the B Domain
The recombinant human factor VIII (rhFVIII-H6A) was produced in our laboratory by employing the CHO-DG44-derived H6A cell clone, as previously described by Demasi, 2016 [13]. The H6A FVIII variant was described initially by Yonemura and colleagues [15] and contained a B-domain deleted HCh, and an artificial light chain (BΔ-LCh, described by Chen, 1999 [14]) fused to part of the B domain. The rhFVIII-H6A product was purified from media conditioned for 24 to 48 hours using anion exchange and heparin pseudo-affinity chromatographic steps, followed by an ultrafiltration/diafiltration step for formulation, employing a membrane with a 100 kDa cut-off and a formulation solution similar to rhFVIII (Kogenate® FS). rhFVIII-H6A recovery was monitored throughout the purification procedure using the chromogenic FVIII activity assay and SDS-PAGE, followed by Coomassie blue staining and LCh Western blot. Overexpression of the rhFVIII-H6A product is associated with the production of two main protein bands, namely: one doublet of 80kDa, corresponding to the fully proteolytically processed LCh isoforms starting at Glu1649 and Asp1658, and a 90kDa protein band, corresponding to proteolytically unprocessed B-LCh primary translation product starting at Asp1563 [11]. In contrast, the LCh electrophoretic profile of pdFVIII (Fahndi®) and of the full-length rhFVIII (Kogenate® FS) is characterized by the detection of only one 80kDa protein band. The rhFVIII-H6A-specific activity was assessed throughout the purification and formulation steps. The final formulated rhFVIII-H6A preparation was obtained after five batches and independent cycles of purification. The specific activity was compared to that of both reference products employed in this work, namely: pdFVIII and rhFVIII. The purification protocol yielded an rhFVIII-H6A preparation displaying an intermediate specific activity value (135 IU/mg) when compared to pdFVIII (10 IU/mg) and rhFVIII (4,500 IU/mg) (Figure 1).
2.2. Low Immunogenicity of rhFVIII-H6A in a Murine Model of Hemophilia A
We first evaluated the immunogenicity of the rhFVIII-H6A isoform in comparison with highly purified full-length human rhFVIII (~4,500 IU/mg). Since our rhFVIII-H6A preparation displayed a specific activity of approximately 130 IU/mg, indicating that only approximately 3% of the protein content is represented by FVIIII, we also included the pdFVIII commercial preparation, which has a specific activity of approximately 10 IU/mL, with only 0.2% of the protein content being represented by FVIII. The immunization protocol involved treatment by retro-orbital injection of six- to eight-week-old fviii knockout mice once a week with 50 IU/Kg of pdFVIII, rhFVIII, rhFVIII-H6A or vehicle (N=5 per group) for four weeks. The 50 IU/kg dose was elected because it corresponds to the dose regimen employed in the prophylactic treatment of hemophilic patients (Bayer Healthcare LLC, 2021). Five weeks after beginning this treatment, plasma samples from the treated groups were evaluated for reactivity against rhFVIII (Figure 2). Total anti-rhFVIII antibody titers in the plasma samples were assessed ex vivo by ELISA, employing wells coated with rhFVIII. Figure 2A shows that rhFVIII and pdFVIII preparations induced significantly higher values of anti-rhFVIII IgG titers (1,000±0 and 8,400±9,990, respectively), than those observed in PBS-treated (Sham) mice (60±22) (p<0.05 for rhFVIII, and p<0.01 for pdFVIII). Titers of total anti-rhFVIII antibodies found in the plasma of mice treated with rhFVIII-H6A were markedly lower (250±229) than those observed with rhFVIII and pdFVIII. Although the rhFVIII-H6A group presented a markedly reduced total average anti-rhFVIII titers when compared to pdFVIII and rhFVIII (4 and 34 times lower, respectively), this difference was only statistically significant when compared to the pdFVIII reference product (p<0.05). Additionally, the average titer of the rhFVIII-H6A group is higher than that of the control group, but it was also not possible to establish statistical significance. These results suggest that only a small fraction of the immunogenicity elicited by FVIII can be attributed to its minimal inactive pro-FVIII heterodimer isoform. Figure 2B shows that all animals (N=5 for each group tested) treated with either rhFVIII or pdFVIII presented a positive (NBU/mL>0.5) Nijmegen-Bethesda titer of rhFVIII inhibitors, displaying a mean of 28±17 NBU/mL and 13±7 NBU/mL, respectively. Remarkably, only one animal (N=5) treated with rhFVIII-H6A presented a positive Nijmegen-Bethesda titer (2.5NBU/mL). The group of animals treated with rhFVIII-H6A displayed a mean of 0.5±1.3BU/mL, a value that is markedly (56 and 26 times) lower than that observed for rhFVIII and pdFVIII, although not statistically significant when compared to the latter group (Figure 2B). Together, these results indicate a trend for markedly decreased anti-FVIII immune response elicited by the rhFVIII-H6A isoform, suggesting that this isoform constitutes a less immunogenic FVIII configuration.
2.3. In Vivo Functional Activity Evaluation of rhFVIII-H6A
The in vivo hemostatic activity of FVIII was assessed using a murine model of severe hemophilia A through a tail bleeding hemostatic assay, based on blood clotting [14]. We evaluated the proportion of animals presenting a full hemostatic response to an intermediate dose (50IU/kg) of FVIII within the 120min of observation. Hemophilic mice retro-orbitally injected with a 50IU/kg dose of FVIII present a partial hemostatic response, which is associated with approximately 60-70% probability to present a full hemostatic response within 120-min. Compared to the reference products pdFVIII (Fahndi®) and rhFVIII (Kogenate® FS), the rhFVIII-H6A product presented a similar in vivo functional hemostatic activity (Figure 3).
3. Discussion
Inhibitory anti-FVIII antibody formation renders treatment of hemophilia A patients ineffective, representing the most severe complication, resulting in the need for new therapies with bypassing agents and cumbersome and expensive immune tolerance induction protocols [18,19]. Several factors related to the production and purification of therapeutic proteins impact their immunogenic potential, i.e., amino acid sequence and the structure of the molecule; post-translational modifications, such as glycosylation profile and proteolytic processing; the presence of impurities; the physical state of the molecules and the method utilized for viral inactivation [19,20,21]. The root cause of FVIII immunogenicity is unclear, including potential contributions of some dissimilarities among rhFVIII products within their bioengineered chemical structure and post-translational modification patterns. Here, we evaluated the immunogenicity of FVIII products with different characteristics, including structure (full-length vs. BDD heterodimer), presence of vWF (von Willebrand factor, contained in pdFVIII), and purity level. Most rhFVIII products explore the deletion or truncation of the B-domain, being produced using non-human cell lines. The contribution of structural domains to the immunogenicity of the BDD rhFVIII is unclear, so the minimal inactive pro-FVIII heterodimer rhFVIII-H6A was bioengineered by deleting the B domain from the HCh [14], with part of it fused to the light chain (BΔ-LCh) [15]. Most anti-FVIII inhibitory antibodies recognize A2, C1, and C2 FVIII domains [23,24,25,26,27], while anti-B domain antibodies demonstrate low inhibitory activity against FVIII [28]. As other moroctocog alfa products (Refacto AF® and Xyntha®), our recombinant product of rhFVIII-H6A presents only five putative glycosylation sites compared to the 25 sites present in the canonical FVIII and is produced using the CHO cell line. It is not clear whether this differential glycosylation pattern could contribute to the differential immunogenicity potential of B-deleted rFVIII. Still, it is one structural aspect that could be further investigated since extensive evidence of the involvement of N-glycans in proteolytic degradation, aggregation, denaturation, pharmacodynamics, pharmacokinetics, and particularly immunogenicity of therapeutic proteins is available [29]. Exploring two hemophilia A experimental models, Lai et al. reported that the N-linked glycosylated rhFVIII produced in BHK cells (Kogenate® FS) was more immunogenic than that produced in CHO cells (Xyntha®) and that incomplete occupancy of N-linked sites led to the formation of IgM- and IgG-FVIII immune complexes [30]. Moreover, Arthur et al. reported that non-human glycans can regulate anti–FVIII antibody formation in mice, and that BHK-derived rhFVIII possesses higher levels of non-human glycans compared with CHO-derived rhFVIII [31]. The immunogenicity of artificially generated FVIII aggregates in animal models indicates that size, quantity, nature, and structure are essential in initiating unwanted immune responses toward native FVIII [32]. However, a comparative analysis of six commercially available rhFVIII products, including BDD or truncated, revealed no association between molecular structure and sub-visible particles or high-molecular-weight protein species [33,34]. Molecular aggregation studies of three rhFVIII products indicated more variable amounts of aggregates after reconstitution of ReFacto AF® and Kogenate® than of Advate®[35]. Protein aggregates have been linked to increased immune responses to the monomeric form of the protein [36,37,38]. Regarding rhFVIII, it has been considered that aggregated forms do not enhance the immunogenicity, but act as a distinct antigen [39].
Another potential factor associated with FVIII immunogenicity is that pre-formed complexes containing vWF increase the half-life and decrease the FVIII immunogenicity in animal studies [40]. One possible mechanism would be that the degradation of FVIII-vWF complexes, carried out by spleen macrophages, prevents the uptake of FVIII by antigen-presenting cells [41,42,43,44]. In accordance, vWF blocks FVIII binding to the macrophage mannose receptor (CD206) in a dose-dependent manner [45], and an in vitro activation of FVIII-specific CD4+T cells by dendritic cells is abrogated upon removal of the N-linked glycosylation site [46]. However, the notion that FVIII products containing VWF may be less immunogenic is controversial. Clinical evidence of inhibitors in hemophilia A indicates pdFVIII products with intact VWF may be less immunogenic [47], resulting in a lower incidence of inhibitors than rhFVIII [48,49]. Our data (Figure 2B) indicate a tendency for lower immunogenicity of pdFVIII compared to rFVIII, suggesting a potential effect of vWF in suppressing the immunogenic response toward FVIII. The physiological context associated with activation of the blood coagulation cascade, leading to thrombin production and creating a pro-inflammatory context per se could also act as a trigger, generating signals that could stimulate an immune response to FVIII [50,51,52,53]. It has been postulated that the immunogenicity elicited by FVIII could be related, at least in part, to its function in potentiating the blood coagulation intrinsic pathway during the hemostatic response, thereby generating the inflammatory response context required to induce an immunological response to FVIII. According to this hypothesis, a combination of FVIII presence, which in hemophilic patients represents a protein for which tolerance may not have been completely induced, and a co-stimulation signal triggered by thrombin’s action, would induce the formation of inhibitory antibodies to FVIII. Skupsky and colleagues [54] (2009) studied the immunogenicity of human heat-denatured FVIII in hemophilic mice and found that the denatured form was less immunogenic than the native form. In addition, these authors observed a decrease in lymphocyte response and in antibody formation against FVIII in hemophilic mice treated with the anticoagulant warfarin or the Hirudin 4 thrombin inhibitor. These data could suggest a role for thrombin formation through the pro-coagulant activity of FVIII, inducing co-stimulation of FVIII inhibitor formation. Interestingly, complexed or not to vWF, rhFVIII was shown not to present co-stimulatory signals for human dendritic cells [55].
However, Meeks and colleagues (2012) [56] suggested that the primary determinant of immunogenicity to factor VIII in a murine model is independent of its pro-coagulant function. In agreement, Gangadharan et al. reported that thrombin generation or associated coagulation processes do not modulate the anti-FVIII antibody response. Instead, these authors reported that warfarin anticoagulant treatment failed to reduce immunogenicity [57]. To enhance the study of anti-FVIII specific responses, FVIII was recently engineered by incorporating the model antigen ovalbumin to study CD4 T cell responses [58]. We investigated the in vivo hemostatic activity of the purified and formulated rhFVIII-H6A compared to the pdFVIII and rhFVIII reference products, observing similar hemostatic activity among these different sources of FVIII. At this point, we could not determine which factors could contribute to rhFVIII-H6A displaying unexpectedly low immunogenicity in a murine model of hemophilia A. The fact that the rhFVIII-H6A preparations were partially purified could be a confounding factor. However, it should also be mentioned that pdFVIII products contain many other proteins as well, which may influence their FVIII immunogenicity. Importantly and with some relevance to the context of decreasing the immunogenicity of rFVIII for better treatment of Hemophilia A patients, we demonstrated that the treatment with the bioengineered rhFVIII-H6A (compared to plasma-derived and recombinant reference products) corrected the hemophilic phenotype in vivo, inducing very low levels of antibody formation and especially low FVIII neutralizing activity. This work would contribute to addressing the important unmet needs of current prophylaxis for Hemophilia A, particularly in the research of factor FVIII products with lower immunogenicity. Overcoming the limitations resulting from the immunogenicity, the development of better and safer therapy products for the treatment of hemophilia A patients could be a new alternative.
4. Materials and Methods
4.1. Culture of CHO-DG44 Cell Clones Overexpressing rhFVIII
CHO-DG44 cells [12] were co-transfected with pIQID-HC and pIQID-LC dhfr-bicistronic expression plasmids [13] using the Lipofectamine 2000 reagent (Invitrogen) to selection of a cell clone named CHO-DG44-H6A, as described by Demasi, 2016 [13]: the amplified HC cDNA encodes the FVIII contained 19-aa signal peptide and A1 and A2 domains (Ala1 to Arg740) [14]; the cDNA sequence encoding the derivative of FVIII LC (B-LC), described by Yonemura et al. [15] encodes the 19-aa FVIII signal peptide, the B-domain C-terminal portion (Asp1563 to Arg1648), and the entire LC (domains A3, C1, and C2) (Glu1649 to Tyr2332). The representative structure of FVIII is shown as Supplementary material.
The human recombinant FVIII-producing CHO-DG44-H6A cells were expanded in HAM’s F12 medium (Invitrogen) supplemented with 10% heat-inactivated fetal bovine serum (FBS) (Invitrogen) supplemented with 100µg/mL ampicillin, 100µg/mL streptomycin, and 1.2g/L sodium bicarbonate, in a humidified atmosphere of 2%CO2 in air, at 37 °C.
4.2. Purification of rhFVIII-H6A
Purification of rhFVIII-H6A was carried out from medium conditioned by the H6A clone for 24-48 hours, upon cell culturing as a monolayer using α-MEM supplemented with 7% FBS, in three steps, namely: two chromatographic steps and one ultrafiltration. The chromatographic steps were performed using the Akta Purifier UPC 100 system, controlled by the Unicorn 5.3 software (General Electric Healthcare). Anion-exchange chromatography was carried out using prepacked 5mL Hitrap Q-Sepharose Fast Flow columns (GE Healthcare). Column equilibration was achieved with A1 buffer (5mM CaCl2), and unbound proteins were washed out using 3.8% of the A2 buffer (5 mM CaCl2; 2M NaCl) in the A1 buffer. Stepwise elution was carried out using three different NaCl concentrations, namely: 5CV of 26%, 5CV of 50%, and 5CV of 100% from the A2 buffer. Pseudo-affinity chromatography using prepacked 1 mL columns Hitrap Heparin HP (GE Healthcare) was employed. Equilibration was carried out with B1 buffer (150mM NaCl; 5mM CaCl2), and washing of unbound proteins was carried out sequentially with 1% and 24% of the B2 buffer (150mM NaCl; 500mM CaCl2) in the B1 buffer. The System was rewashed with 18CV of 24% B2 buffer. Next, elution was performed using 100% B2 buffer. Samples were then 8-fold concentrated twice through ultrafiltration (1,500xg at 4 °C) using Amicon Ultra-4 100kDa (Millipore Corporation, Carrigtwohill, CO, Ireland) with one step of dilution (1:7) in formulation solution (1,3% Sucrose; 25mg/mL Glycine; 23mM Histidine; 36mEq/L NaCl; 80μg/mL Polysorbate 80; 20mM Imidazole; 0,06µg/mL CuCl2; pH7,3). Samples were sterilized using 0.22µm membranes and snap-frozen. Representative chromatograms and typical recovery results are provided in the Supplementary material.
4.3. Quantification of Total Proteins
Total protein was quantified using the colorimetric assay based on bicinchoninic acid, namely: the BCA Protein Assay commercial kit (Lot CI48587) from Pierce (ThermoFisher Scientific, Rockford, Illinois, USA), following the manufacturer’s instructions.
4.4. FVIII Chromogenic Assay
FVIII activity was quantified employing a chromogenic activity assay (Coatest SP4 FVIII, Chromogenix, Italy) using the acetic acid-stopped endpoint method according to the manufacturer’s instructions. All samples were diluted to yield values within the linear range of the assay.
4.5. Western Blot Analysis
Samples collected from all purification steps were fractionated in 8% SDS-polyacrylamide gel electrophoresis. The resolved proteins were blotted onto nitrocellulose membranes (Bio-Rad, Hercules, CA), which were blocked overnight at 4 °C with 5% nonfat milk in TBS-T buffer containing 0.05% Tween 20. The blots were probed in TBS-T/milk with the anti-LCh (Chemicon Mab-038, Merck-Millipore, MA, USA) (dilution 1:750) and then developed using (HRP)-conjugated secondary antibodies (Invitrogen) and ECL Plus chemiluminescent detection reagent (GE HealthCare, NJ, USA), according to the manufacturer’s instructions.
4.6. Reference Products
The plasma-derived factor VIII Fanhdi® (Lot IBVC0XEXG1, Grifols Institute, Parets del Valles, BA, Spain) was donated by the Brazilian Program of Coagulopathies from The Ministry of Health. The recombinant factor VIII Kogenate® FS (Lot 27N2JP2, Bayer AG, Leverkusen, NRW, Germany) was donated by the Secretary of Health from the Federal District of Brazil.
4.7. Animals
FVIII knockout mice (B6;129S4-F8tm1Kaz/J strain), generated through targeted exon-6 disruption [8], were purchased from The Jackson Laboratory (Bar Harbor, ME, USA). FVIII wild-type mice (129J/Sv strain) were purchased from the Animal Facility of the University of São Paulo Biomedical Sciences Institute (ICB/USP, São Paulo, SP, Brazil). All mice procedures were approved by the Ethics Committee on Animal Care and Use of the Chemistry Institute of the University of São Paulo (Protocol number 20/2012), being in accordance with the Brazilian National Council for Control of Animal Experimentation (CONCEA).
4.8. Functional Activity of FVIII
A quantitative tail bleeding hemostatic assay was carried out as previously described [16]. Hemophilic mice were anesthetized and treated with FVIII by retro-orbital injection. Ten minutes after FVIII administration, the tip of the mice’s tails was transected at 1mm diameter using a ring template, and the time for coagulation was quantified. Bleeding was recorded every 15-min by blotting the blood accumulated on the mouse tail tip during the 15-min observational interval onto a 3 mm Whatmann cellulose filter paper.
4.9. Induction of Immunological Response to Factor VIII
Induction of immunological response was assessed using the murine model of severe hemophilia A. Mice were divided into four groups of five animals each (N=5) for treatment with vehicle (formulation solution) or 50IU/kg of rhFVIII-H6A or reference products rhFVIII (Kogenate® FS) and pdFVIII (Fanhdi®). Mice received four weekly administrations, and five weeks from the beginning of the treatment, 180µL of blood samples were collected directly into tubes containing 20µL of 130mM sodium citrate by puncturing the submandibular vein using an 18G needle. Samples were centrifuged for 5 min at 2,000xg, and plasma samples were immediately frozen for further analysis.
4.10. Dosage of Total Anti-Factor VIII Antibodies
Titers of anti-FVIII immunoglobulins G (IgG) were measured in duplicate for the plasma of hemophilic mice treated with rhFVIII-H6A, rhFVIII (Kogenate® FS), pdFVIII (Fanhdi®), or vehicle by enzymatic immunoassay (ELISA). Ant-FVIII titers were assessed in 96-well Nunc MaxiSorp coated with rhFVIII. Briefly, the coating was carried out with rhFVIII in 50µL of carbonate buffer pH 9.4 for 16h at 4 °C, followed by washing with 0.1% Tween 20 in phosphate-buffered saline solution (PBSA). Non-specific binding sites were blocked with 200µL of 2% (w/v) bovine serum albumin (BSA) in PBSA at room temperature and humid atmosphere for 2h. The wells were washed three times using 0.1% Tween 20 and incubated for 2h at 37 °C with 100µL samples of plasma from hemophilic mice, which were serially diluted in blocking solution. Analysis was carried out with plasma dilution curves from animals treated with rhFVIII-H6A 1:25, 1:50, 1:100, 1:500, 1:1,000; Kogenate® FS 1:100, 1:500, 1:1000, 1:5,000, 1:10,000, 1:25,000; Fanhdi® 1:1,000, 1:5,000, 1:10,000, 1:25,000, 1:50,000, 1:75,000; and vehicle 1:25, 1:50, 1:100, 1:500, 1:1000. The wells were washed three times using 0.1% Tween 20 and then incubated with anti-mouse secondary antibody IgG from Invitrogen (Life Technologies, Burlingame, CA, USA) conjugated to peroxidase and diluted 1:2,000 in blocking solution, at room temperature and humid atmosphere for 1h and 30min. The wells were then washed three times with 0.1% Tween 20, and reactions were revealed using the TMB ELISA kit solution (Thermo Fisher Scientific, Rockford, Illinois, USA), according to the manufacturer’s instructions. Absorbance was measured at 405nm using Molecular Devices SpectraMax M2 (LLC Molecular Devices, Sunnyvale, CA, USA). Antibody titers were estimated based on the highest dilution presenting a positive signal, defined as the absorbance value greater than 0.05, representing the mean of the negative control plus the standard deviation value multiplied by three.
4.11. Dosage of Total Inhibitory Factor VIII Antibodies
Titers of inhibitors were detected in the plasma of hemophilic mice treated with vehicle rhFVIII-H6A, rhFVIII (Kogenate® FS), or pdFVIII (Fanhdi®) based on the Nijmegen-modified Bethesda assay [17]. Plasma samples from the animals previously treated with rhFVIII-H6A, Kogenate® FS, Fanhdi®, or vehicle were first incubated at 58 °C for 90min to inactivate any residual FVIII activity, followed by centrifugation at 4,000xg for 2min to remove any debris. Plasma samples were serially diluted in commercial human FVIII-deficient plasma (Affinity Biologicals Inc, Ancaster, Ontario, Canada). For the group of animals treated with rhH6A, samples were diluted 1:5 and 1:25; for Kogenate® FS at 1:10 to 1:50; for Fanhdi® at 1:20, 1:30, 1:100, and 1:150; and for the vehicle, which was defined as ‘plasma-test’ at 1:4 and 1:20. Triplicate samples containing only FVIII-deficient plasma were used as ‘plasma-control’ samples. ‘Plasma-test’ and ‘plasma-control’ samples were incubated for 2h at 37 °C with an equal volume (45µL) of 2IU/mL of FVIII (Fanhdi®) in FVIII-deficient plasma buffered with 100mM Imidazole at pH 7.4. After the incubation period, the FVIII activity was evaluated in ‘plasma-test’ and ‘plasma-control’ samples by the in vitro chromogenic assay. The FVIII activity obtained in the ‘plasma-test’ samples was expressed relative to that obtained in the ‘control plasma’ samples and then referred to as ‘remaining FVIII activity (%). Quantification of inhibitors was performed using linear regression on a semi-logarithmic scale, assuming that 100% of the remaining FVIII activity corresponded to 0.0 NBU/mL and that 50% of the remaining activity corresponded to 1.0 NBU/mL. The remaining activity values were only estimated between 25% and 75% from the FVIII activity of control plasma samples, since only this interval would linearly correlate to the presence of FVIII inhibitors.
4.12. Statistical Analysis
The different levels of total and inhibitory anti-factor VIII antibodies formation among the plasma of hemophilic animals treated with rhFVIII-H6A, rhFVIII (Kogenate® FS), pdFVIII (Fanhdi®), or vehicle (Sham) were compared through nonparametric one-way analysis of variance Kruskal–Wallis test, followed by Dunn’s multiple comparison test using GraphPad 5.0 (GraphPad Software, CA, USA). Statistical significance was set at p<0.05. Data are expressed as the mean±SD representing independent animals.
Author Contributions
ESM and MAAD designed and executed the experiments, discussed the results and wrote the manuscript; TLD executed ELISA assays and reviewed the manuscript; MCS discussed the results and reviewed the manuscript.
Data Availability Statement
Purification data may be found in a data supplement available in the online version of this article. For original data, please email the corresponding author.
Acknowledgements
This study was funded by the Brazilian Higher Education Personnel Improvement Coordination (CAPES) grant number 88881.068070/2014-01, the National Council for Scientific and Technological Development (CNPq) to INCT-REGENERA grant number 465656/2014-5, and the São Paulo State Research Foundation – FAPESP, Thematic grant Number 2016/05311-2. ESM held a pre-doctoral fellowship from CNPq. Donations of reference products by the Brazilian Program of Coagulopathies and by The Ministry of Health and the Secretary of Health from the Federal District of Brazil.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Pipe, S.W.; Morris, J.A.; Shah, J.K.R. Differential interaction of coagulation factor VIII and factor V with protein chaperones calnexin and calreticulin. J. Biol. Chem. 1998, 273, 8537–8544. [Google Scholar] [CrossRef]
- Ljung, R.; Auerswald, G.; Benson, G.; et al. Inhibitors in haemophilia A and B: Management of bleeds, inhibitor eradication and strategies for difficult-to-treat patients. Eur J Haematol 2019, 102, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Scharrer, I.; Bray, G.L.; Neutzling, O. Incidence of inhibitors in haemophilia A patients - a review of recent studies of recombinant and plasma-derived factor VIII concentrates. Haemophilia 1999, 5, 145–154. [Google Scholar] [CrossRef]
- Schellekens, H.; Casadevall, N. Immunogenicity of recombinant human proteins: causes and consequences. J. Neurol. 2004, 251, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Ebbers, H.C.; Crow, S. a, Vulto, A.G., Schellekens, H. Interchangeability, immunogenicity and biosimilars. Nat Biotechnol. 2012, 30, 1186–1190. [Google Scholar] [CrossRef]
- Koren, E.; Smith, H.W.; Shores, E.; et al. Recommendations on risk-based strategies for detection and characterization of antibodies against biotechnology products. J. Immunol. Methods. 2008, 333, 1–9. [Google Scholar] [CrossRef]
- Barbosa, M.D.F.S. Immunogenicity of biotherapeutics in the context of developing biosimilars and biobetters. Drug Discov. Today. 2011, 16, 345–353. [Google Scholar] [CrossRef]
- Bi, L.; Lawler, A.M.; Antonarakis, S.E.; High, K.A.; Gearhart, J.D.; Kazazian, H.H. Targeted disruption of the mouse factor VIII gene produces a model of haemophilia A. Nat. Genet. 1995, 10, 119–121. [Google Scholar] [CrossRef]
- Healey, J.F.; Parker, E.T.; Barrow, R.T.; Langley, T.J.; Church, W.R.; Lollar, P. The Comparative Immunogenicity of Human and Porcine Factor VIII in Hemophilia A Mice. Thromb Haemost. 2009, 102, 35. [Google Scholar] [CrossRef]
- Healey, J.F.; Lubin, I.M.; Nakai, H.; Saenko, E.L.; Hoyer, L.W.; Scandella, D.; Lollar, P. Residues 484-508 contain a major determinant of the inhibitory epitope in the A2 domain of human factor VIII. J Biol Chem. 1995, 270, 14505–14509. [Google Scholar] [CrossRef]
- Parker, E.T.; Healey, J.F.; Barrow, R.T.; Craddock, H.N.; Lollar, P. Reduction of the inhibitory antibody response to human factor VIII in hemophilia A mice by mutagenesis of the A2 domain B-cell epitope. Blood. 2004, 104, 704–710. [Google Scholar] [CrossRef]
- Urlaub, G.; Mitchell, P.J.; Kas, E.; Chasin, L.A.; Funanage, V.L.; Myoda, T.T.; Hamlin, J. Effect of gamma rays at the dihydrofolate reductase locus: deletions and inversions. Somat Cell Mol Genet 1986, 12, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Demasi, M.A.; de, S. Molina, E.; Bowman-Colin, C.; Lojudice, F.H.; Muras, A.; Sogayar, M.C. Enhanced Proteolytic Processing of Recombinant Human Coagulation Factor VIII B-Domain Variants by Recombinant Furins. Mol Biotechnol 2016, 58, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Fang, X.D.; Zhu, J.; et al. The gene expression of coagulation factor VIII in mammalian cell lines. Thrombosis Research 1999, 95, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Yonemura, H.; Sugawara, K.; Nakashima, K.; et al. Efficient production of recombinant human factor VIII by co-expression of the heavy and light chains. Protein Eng. 1993, 6, 669–674. [Google Scholar] [CrossRef]
- Molina, E.S.; Fujita, A.; Sogayar, M.C.; Demasi, M.A. A quantitative and humane tail bleeding assay for efficacy evaluation of antihaemophilic factors in haemophilia A mice. Haemophilia 2014, 20, e392–e398. [Google Scholar] [CrossRef]
- Verbruggen, B.; van Heerde, W.L.; Laros-van Gorkom, B. a P.. Improvements in factor VIII inhibitor detection: From Bethesda to Nijmegen. Semin. Thromb. Hemost. 2009, 35, 752–759. [Google Scholar] [CrossRef]
- Messori, A. Inhibitors in Hemophilia A: A Pharmacoeconomic Perspective. Semin Thromb Hemost 2018, 44, 561–567. [Google Scholar] [CrossRef]
- Scott, D.W.; Pratt, K.P. Factor VIII: Perspectives on Immunogenicity and Tolerogenic Strategies. Front Immunol 2020, 10, 3078. [Google Scholar] [CrossRef]
- European Medicines Agency. Guideline on the clinical investigation of recombinant and plasma-derived factor VIII products. EMA/CHMP/BPWP/144533/2009 rev. 2. Committee for Medicinal Products for Human Use (CHMP). London. 26 July 2018. https://www.ema.europa.eu/en/documents/scientific-guideline/guideline-clinical-investigation-recombinant-and-human-plasma-derived-factor-viii-products-revision-2_en.pdf.
- Hermeling, S.; Crommelin, D.J.A.; Schellekens, H.; Jiskoot, W. Structure-immunogenicity relationships of therapeutic proteins. Pharm. Res. 2004, 21, 897–903. [Google Scholar] [CrossRef]
- Singh, S.K.; Cousens, L.P.; Alvarez, D.; Mahajan, P.B. Determinants of immunogenic response to protein therapeutics. Biologicals 2012, 40, 364–368. [Google Scholar] [CrossRef]
- Meeks, S.L.; Healey, J.F.; Parker, E.T.; Barrow, R.T.; Lollar, P. Antihuman factor VIII C2 domain antibodies in hemophilia A mice recognize a functionally complex continuous spectrum of epitopes dominated by inhibitors of factor VIII activation. Blood 2007, 110, 4234–4242. [Google Scholar] [CrossRef]
- Pratt, K.P.; Qian, J.; Ellaban; et al. Immunodominant T-cell epitopes in the factor VIII C2 domain are located within an inhibitory antibody binding site. Thromb. Haemost. 2004, 92, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Scandella, D.H.; Nakai, H.; Felch, M.; Mondorf, W.; Scharrer, I.; Hoyer, L.W.; Saenko, E.L. In hemophilia A and autoantibody inhibitor patients: the factor VIII A2 domain and light chain are most immunogenic. Thromb. Res. 2001, 101, 377–385. [Google Scholar] [CrossRef]
- Wroblewska, A.; van Haren, S.D.; Herczenik; et al. Modification of an exposed loop in the C1 domain reduces immune responses to factor VIII in hemophilia A mice. Blood 2012, 119, 5294–5300. [Google Scholar] [CrossRef] [PubMed]
- Wuerth, M.E.; Cragerud, R.K.; Clint Spiegel, P. Structure of the Human Factor VIII C2 Domain in Complex with the 3E6 Inhibitory Antibody. Scientific Reports 2015, 5:1 5, 1–11. [Google Scholar] [CrossRef]
- Lavigne-Lissalde, G.; Lacroix-Desmazes, S.; Wootla, B.; et al. Molecular characterization of human B domain-specific anti-factor VIII monoclonal antibodies generated in transgenic mice. Thromb. Haemost. 2007, 98, 138–147. [Google Scholar] [PubMed]
- Zhou, Q.; Qiu, H. The Mechanistic Impact of N-Glycosylation on Stability, Pharmacokinetics, and Immunogenicity of Therapeutic Proteins. J Pharm Sci 2019, 108, 1366–1377. [Google Scholar] [CrossRef]
- Lai, J.D.; Swystun, L.L.; Cartier, D.; et al. N-linked glycosylation modulates the immunogenicity of recombinant human factor VIII in hemophilia A mice. Haematologica 2018, 103, 1925–1936. [Google Scholar] [CrossRef]
- Arthur, C.M.; Zerra, P.E.; Shin, S.; et al. Nonhuman glycans can regulate anti-factor VIII antibody formation in mice. Blood 2022, 139, 1312–1317. [Google Scholar] [CrossRef]
- Pisal, D.S.; Kosloski, M.P.; Middaugh, C.R.; Bankert, R.B.; Balu-Iyer, S. Native-like aggregates of factor VIII are immunogenic in von Willebrand factor deficient and hemophilia a mice. J. Pharm. Sci. 2012, 101, 2055–2065. [Google Scholar] [CrossRef]
- Anzengruber, J.; Lubich, C.; Prenninger, T.; et al. Comparative analysis of marketed factor VIII products: recombinant products are not alike vis-a-vis soluble protein aggregates and subvisible particles. J Thromb Haemost 2018, 16, 1176–1181. [Google Scholar] [CrossRef] [PubMed]
- Baunsgaard, D.; Nielsen, A.D.; Nielsen; et al. A comparative analysis of heterogeneity in commercially available recombinant factor VIII products. Haemophilia 2018, 24, 880–887. [Google Scholar] [CrossRef] [PubMed]
- Cristofaro, R.; de Sacco, M.; Lancellotti, S.; Berruti, F.; Garagiola, I.; Valsecchi, C.; Basso, M.; Stasio, E.d.; Peyvandi, F. Molecular Aggregation of Marketed Recombinant FVIII Products: Biochemical Evidence and Functional Effects. TH Open 2019, 3, e123. [Google Scholar] [CrossRef]
- Lundahl, M.L.E.; Fogli, S.; Colavita, P.E.; Scanlan, E.M. Aggregation of protein therapeutics enhances their immunogenicity: causes and mitigation strategies. RSC Chem Biol 2021, 2, 1004–1020. [Google Scholar] [CrossRef] [PubMed]
- Ratanji, K.D.; Derrick, J.P.; Dearman, R.J.; Kimber, I. Immunogenicity of therapeutic proteins: influence of aggregation. J Immunotoxicol 2014, 11, 99–109. [Google Scholar] [CrossRef]
- Rosenberg, A.S. Effects of protein aggregates: an immunologic perspective. AAPS J 2006, 8. [Google Scholar] [CrossRef]
- Purohit, V.S.; Middaugh, C.R.; Balasubramanian, S. Influence of aggregation on immunogenicity of recombinant human factor VIII in hemophilia A mice. J. Pharm. Sci. 2006, 95, 358–371. [Google Scholar] [CrossRef]
- Delignat, S.; Repessé; Y; Navarrete; et al. Immunoprotective effect of von Willebrand factor towards therapeutic factor VIII in experimental haemophilia A. Haemophilia 2012, 18, 248–254. [Google Scholar] [CrossRef]
- Dasgupta, S.; Repessé; Y; Bayry, J. ; et al. VWF protects FVIII from endocytosis by dendritic cells and subsequent presentation to immune effectors. Blood 2007, 109, 610–612. [Google Scholar] [CrossRef]
- Kaveri, S.v.; Dasgupta, S.; Andre, S.; et al. Factor VIII inhibitors: role of von Willebrand factor on the uptake of factor VIII by dendritic cells. Haemophilia 2007, 13, 61–64. [Google Scholar] [CrossRef]
- Lacroix-Desmazes, S.; Repessé; Y; Kaveri, S. v.; Dasgupta, S. The role of VWF in the immunogenicity of FVIII. Thromb Res 2008, 122, S3–6. [Google Scholar] [CrossRef] [PubMed]
- Sorvillo, N.; Hartholt, R.B.; Bloem, E.; et al. von Willebrand factor binds to the surface of dendritic cells and modulates peptide presentation of factor VIII. Haematologica 2016, 101, 309. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Navarrete, A.M.; Bayry, J.; et al. A role for exposed mannosylations in presentation of human therapeutic self-proteins to CD4+ T lymphocytes. Proc Natl Acad Sci U S A 2007, 104, 8965–8970. [Google Scholar] [CrossRef] [PubMed]
- Delignat, S.; Rayes, J.; Dasgupta, S.; et al. Removal of Mannose-Ending Glycan at Asn2118 Abrogates FVIII Presentation by Human Monocyte-Derived Dendritic Cells. Front Immunol 2020, 11. [Google Scholar] [CrossRef]
- Ettingshausen, C.E. , Kreuz, W. Recombinant vs. plasma-derived products, especially those with intact VWF, regarding inhibitor development. Haemophilia 2006, 12, 102–106. [Google Scholar] [CrossRef]
- Calvez, T.; Chambost, H.; D’Oiron, R.; et al. Analyses of the FranceCoag cohort support differences in immunogenicity among one plasma-derived and two recombinant factor VIII brands in boys with severe hemophilia A. Haematologica 2018, 103, 179–189. [Google Scholar] [CrossRef]
- Peyvandi, F.; Mannucci, P.M.; Garagiola, I.; et al. A Randomized Trial of Factor VIII and Neutralizing Antibodies in Hemophilia A. New England Journal of Medicine 2016, 374, 2054–2064. [Google Scholar] [CrossRef]
- Eaton, D.; Rodriguez, H.; Vehar, G.A. Proteolytic processing of human factor VIII. Correlation of specific cleavages by thrombin, factor Xa, and activated protein C with activation and inactivation of factor VIII coagulant activity. Biochemistry 1986, 25, 505–512. [Google Scholar] [CrossRef]
- Esmon, C.T. Interactions between the innate immune and blood coagulation systems. Trends. Immunol. 2004, 25, 536–542. [Google Scholar] [CrossRef]
- Walker, F.J.; Fay, P.J. Regulation of blood coagulation by the protein C system. FASEB J 1992, 6, 2561–2567. [Google Scholar] [CrossRef]
- Wilhelm, A.R.; Parsons, N.A.; Samelson-Jones, B.J.; et al. Activated protein C has a regulatory role in factor VIII function. Blood 2021, 137, 2532–2543. [Google Scholar] [CrossRef]
- Skupsky, J.; Zhang, A.; Su, Y.; Scott, D.W. A role for thrombin in the initiation of the immune response to therapeutic factor VIII. Blood 2009, 114, 4741–4748. [Google Scholar] [CrossRef]
- Pfistershammer, K.; Stöckl, J.; Siekmann, J.; Turecek, P.L.; Schwarz, H.P.; Reipert, B.M. Recombinant factor VIII and factor VIII-von Willebrand factor complex do not present danger signals for human dendritic cells. Thromb. Haemost. 2006, 96, 309–316. [Google Scholar] [CrossRef]
- Meeks, S.L.; Cox, C.L.; Healey; et al. A major determinant of the immunogenicity of factor VIII in a murine model is independent of its procoagulant function. Blood 2012, 120, 2512–2520. [Google Scholar] [CrossRef]
- Gangadharan, B.; Delignat, S.; Ollivier; et al. Role of coagulation-associated processes on factor VIII immunogenicity in a mouse model of severe hemophilia A. J Thromb Haemost 2014, 12, 2065–2069. [Google Scholar] [CrossRef]
- Zerra, P.E.; Parker, E.T.; Baldwin, W.H.; et al. Engineering a Therapeutic Protein to Enhance the Study of Anti-Drug Immunity. Biomedicines 2022, 10, 1724. [Google Scholar] [CrossRef] [PubMed]
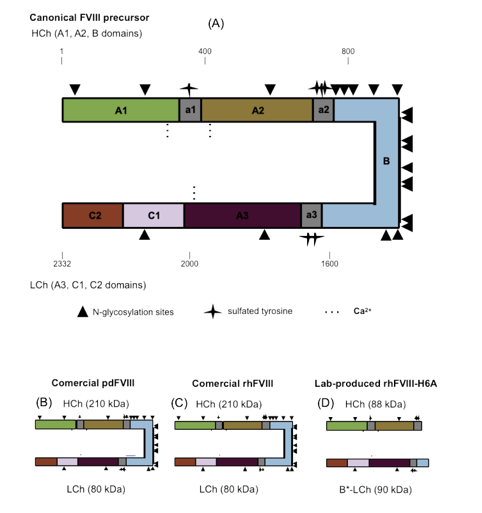
Figure 1.
Purity, structural integrity analysis, and specific activity of the recombinant Factor VIII produced in our laboratory (rhFVIII-H6A) compared to reference commercial products. (A) Factor VIII light chain (LCh) Western blot analysis of the pool of samples derived from media conditioned by the H6A cell clone (well 2) (0.04 IU); after the anion exchange chromatography step (well 3) (0.07 IU); after the affinity chromatography step (well 4) (0.13 IU); retained fractions after the 100 kDa ultrafiltration step (well 5) (0.49 IU); rhFVIII-H6A final formulation (well 6) (0.5 IU); recombinant product Kogenate® FS (well 7) (0.5 IU); and plasma derived Fanhdi® (well 9) (0.5 IU). Molecular weight ladder was applied in well number 1 and water in well number 8. (B) Evaluation of the protein mixture complexity was carried out for the same samples described in (A) using 8% denaturing polyacrylamide gel electrophoresis, followed by staining with Coomassie blue G-250 colloidal using 2 IU of formulated rhH6A (well 6); 2 IU of rhFVIII Kogenate® FS (well 7) and 1 IU pdFVIII Fanhdi® (well 9). (C) Factor VIII specific activity obtained for formulated rhFVIII-H6A. Specific activity was quantified based on chromogenic in vitro Factor VIII activity assay and protein quantification using colorimetric bicinchoninic acid assay.
Figure 1.
Purity, structural integrity analysis, and specific activity of the recombinant Factor VIII produced in our laboratory (rhFVIII-H6A) compared to reference commercial products. (A) Factor VIII light chain (LCh) Western blot analysis of the pool of samples derived from media conditioned by the H6A cell clone (well 2) (0.04 IU); after the anion exchange chromatography step (well 3) (0.07 IU); after the affinity chromatography step (well 4) (0.13 IU); retained fractions after the 100 kDa ultrafiltration step (well 5) (0.49 IU); rhFVIII-H6A final formulation (well 6) (0.5 IU); recombinant product Kogenate® FS (well 7) (0.5 IU); and plasma derived Fanhdi® (well 9) (0.5 IU). Molecular weight ladder was applied in well number 1 and water in well number 8. (B) Evaluation of the protein mixture complexity was carried out for the same samples described in (A) using 8% denaturing polyacrylamide gel electrophoresis, followed by staining with Coomassie blue G-250 colloidal using 2 IU of formulated rhH6A (well 6); 2 IU of rhFVIII Kogenate® FS (well 7) and 1 IU pdFVIII Fanhdi® (well 9). (C) Factor VIII specific activity obtained for formulated rhFVIII-H6A. Specific activity was quantified based on chromogenic in vitro Factor VIII activity assay and protein quantification using colorimetric bicinchoninic acid assay.
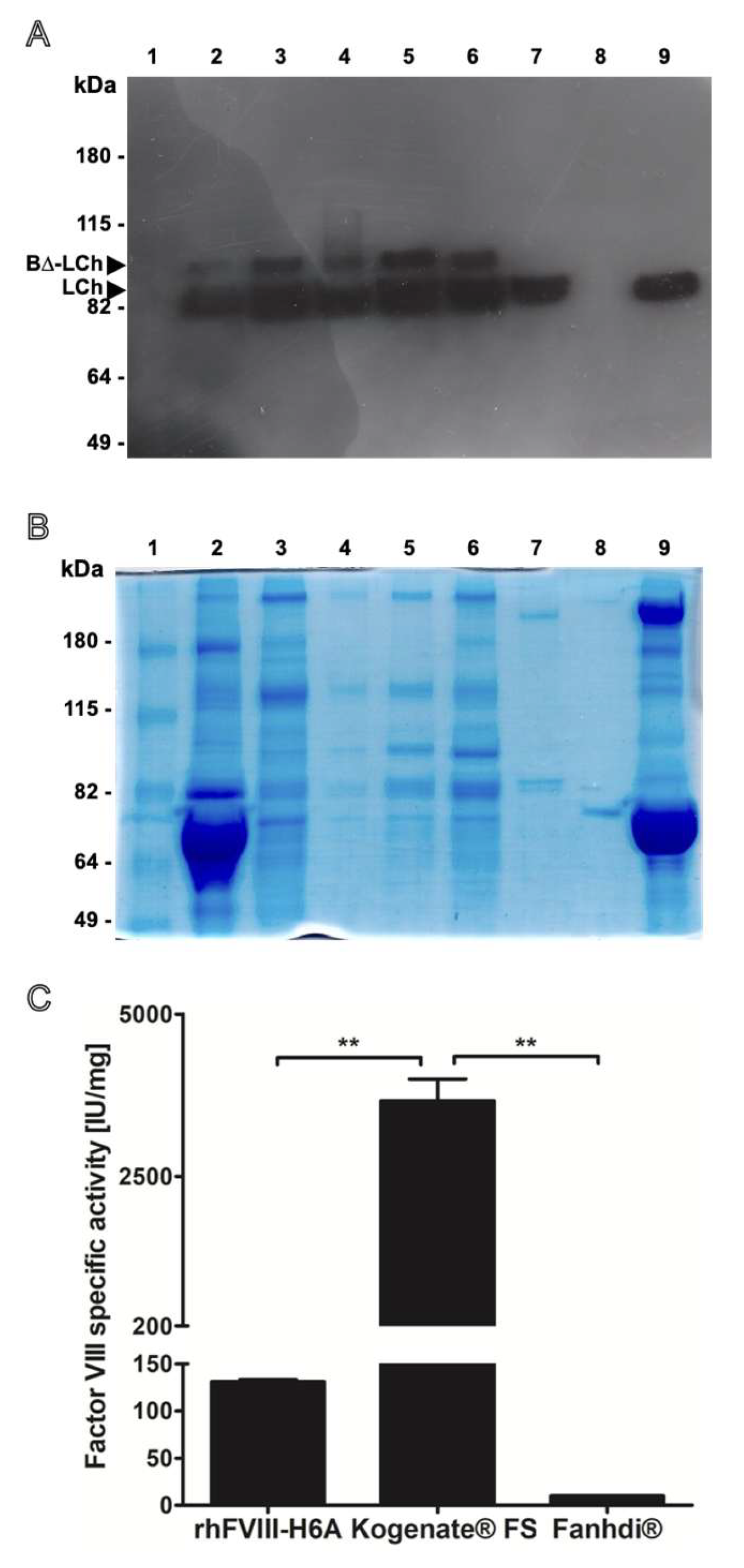
Figure 2.
Alloantibodies quantification in plasma of hemophilic mice after weekly treatments with the bioengineered Factor VIII (rhFVIII-H6A) compared to reference products. The animal model of severe hemophilia A was treated with four weekly administrations of vehicle, named Sham, or 50 IU/Kg of rhFVIII-H6A produced in our laboratory, commercial rhFVIII (Kogenate® FS), or commercial pdFVIII (Fanhdi®). Five weeks after the beginning of the treatment, anti-factor VIII antibodies were quantified in the plasma samples (N=5 per group). (A) Scatter plot of total anti-factor VIII antibodies, which were quantified by immunoenzymatic assay. (B) Scatter plot of anti-factor VIII neutralizing antibodies, which were quantified using the Nijmegen-Bethesda assay. One-way non-parametric analysis of variance followed by multiple comparison test (* 0.01<P<0.05, ** 0.001<P <0.01).
Figure 2.
Alloantibodies quantification in plasma of hemophilic mice after weekly treatments with the bioengineered Factor VIII (rhFVIII-H6A) compared to reference products. The animal model of severe hemophilia A was treated with four weekly administrations of vehicle, named Sham, or 50 IU/Kg of rhFVIII-H6A produced in our laboratory, commercial rhFVIII (Kogenate® FS), or commercial pdFVIII (Fanhdi®). Five weeks after the beginning of the treatment, anti-factor VIII antibodies were quantified in the plasma samples (N=5 per group). (A) Scatter plot of total anti-factor VIII antibodies, which were quantified by immunoenzymatic assay. (B) Scatter plot of anti-factor VIII neutralizing antibodies, which were quantified using the Nijmegen-Bethesda assay. One-way non-parametric analysis of variance followed by multiple comparison test (* 0.01<P<0.05, ** 0.001<P <0.01).
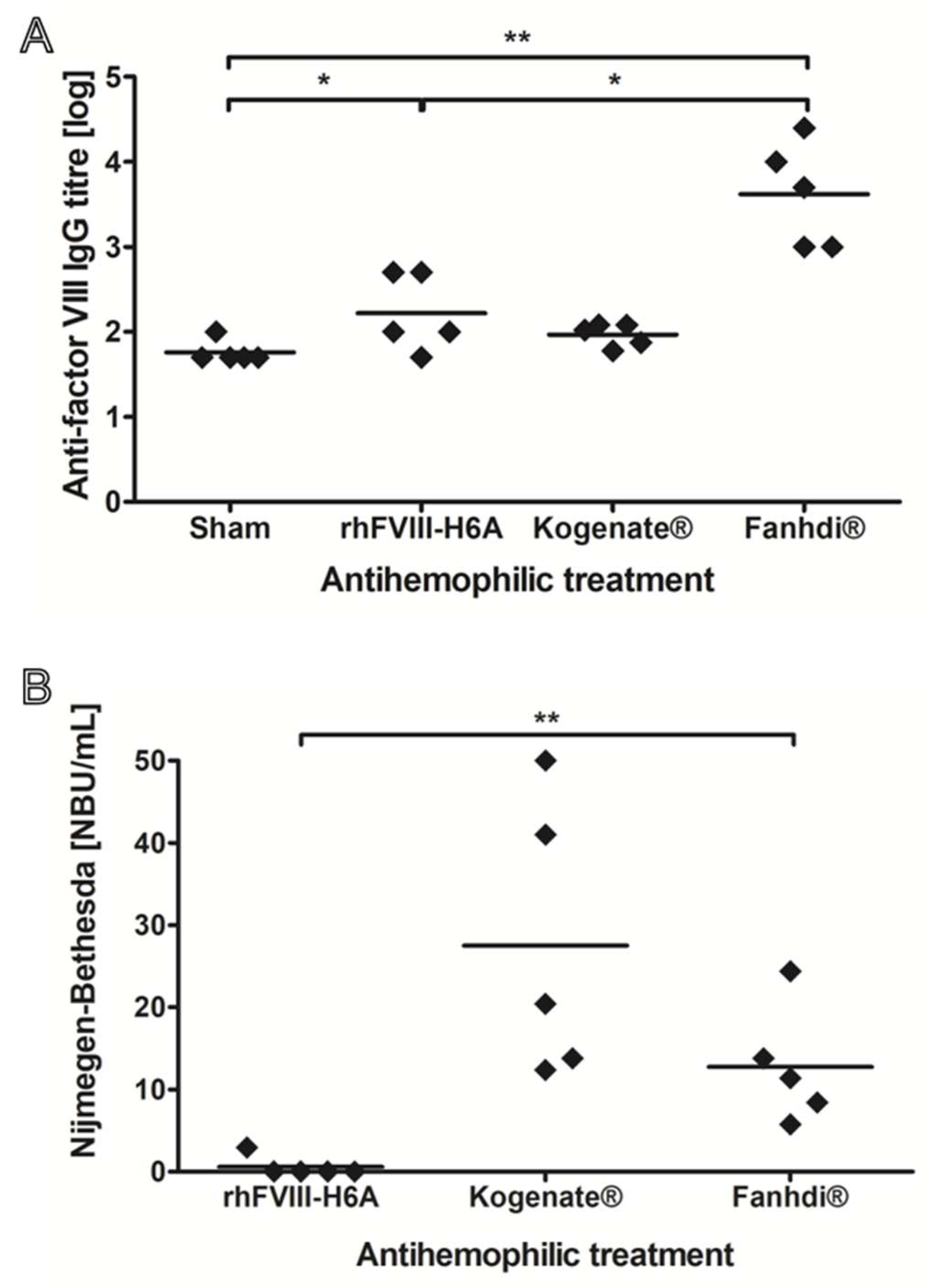
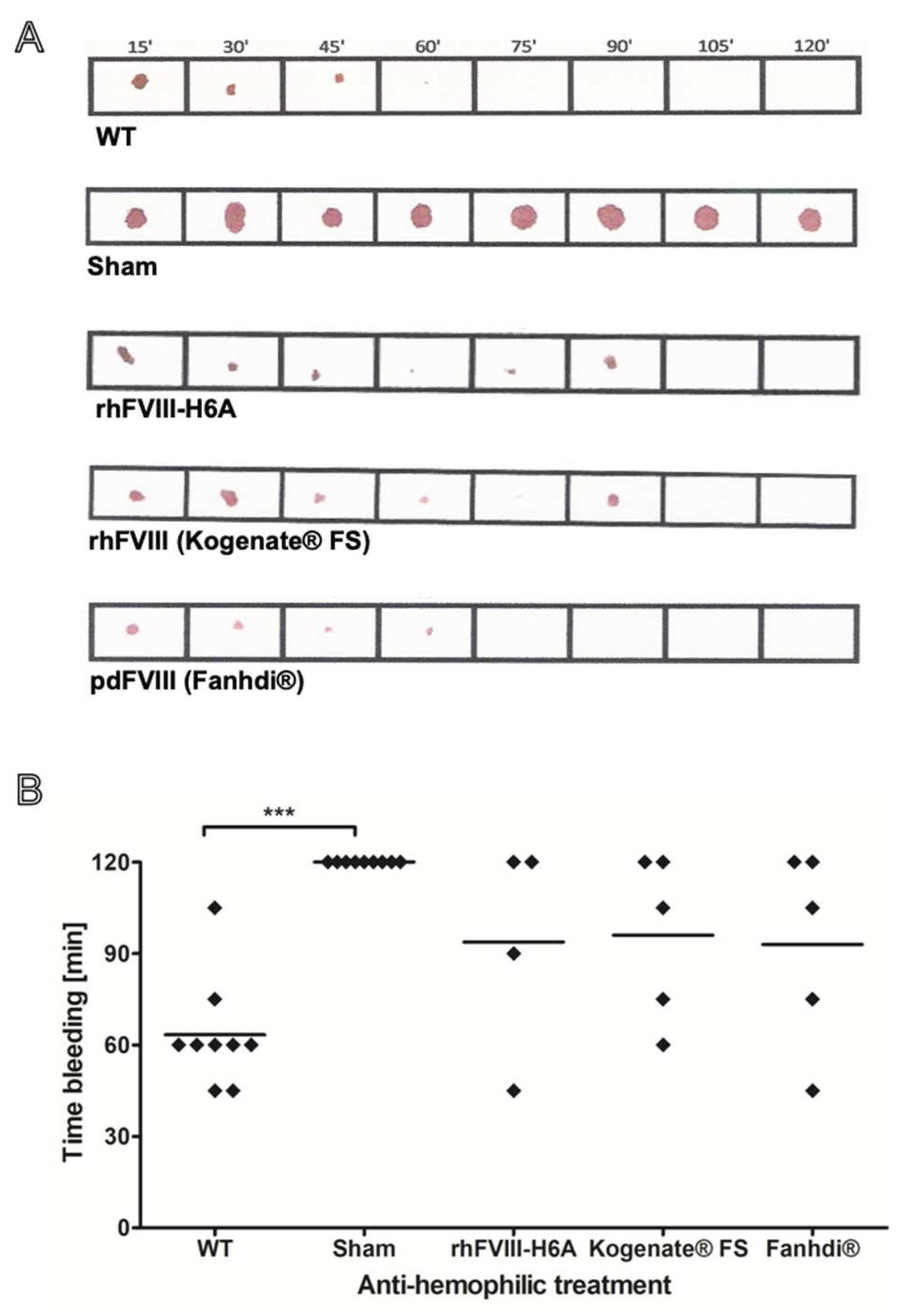
Figure 3.
Functional in vivo activity of the bioengineered factor VIII produced in our laboratory (rhFVIII-H6A) compared to reference products. The anti-hemophilic effect was assessed by inducing a hemorrhagic event in Factor VIII knockout mice previously treated with factor VIII. The reversion of severe hemophilia A phenotype was estimated based on an assay involving tail bleeding interruption, indicating stable clot formation. The same background wild-type (WT) mice were used as a positive control of physiologic clot formation. WT animals were treated with vehicle (N=9) and hemophilic animals were treated with vehicle, named Sham (N=9), or 50 IU/Kg of rhFVIII-H6A (N=4), commercial rhFVIII (Kogenate® FS) (N=5), or commercial pdFVIII (Fanhdi®) (N=5). After 10 min of retro-orbital injections, a surgical blade was used to transect the animal’s tail at a level of 1 mm in diameter. The bleeding time was recorded on filter paper every 15 min until reaching 120 min, when the experiment was interrupted by tail cauterization of the Sham group. (A) Representative bleeding records obtained on paper filters for WT, rhH6A, and reference products. (B) Scatter plotting of bleeding time values. One-way analysis of variance followed by multiple comparison test (*** P<0.001).
Figure 3.
Functional in vivo activity of the bioengineered factor VIII produced in our laboratory (rhFVIII-H6A) compared to reference products. The anti-hemophilic effect was assessed by inducing a hemorrhagic event in Factor VIII knockout mice previously treated with factor VIII. The reversion of severe hemophilia A phenotype was estimated based on an assay involving tail bleeding interruption, indicating stable clot formation. The same background wild-type (WT) mice were used as a positive control of physiologic clot formation. WT animals were treated with vehicle (N=9) and hemophilic animals were treated with vehicle, named Sham (N=9), or 50 IU/Kg of rhFVIII-H6A (N=4), commercial rhFVIII (Kogenate® FS) (N=5), or commercial pdFVIII (Fanhdi®) (N=5). After 10 min of retro-orbital injections, a surgical blade was used to transect the animal’s tail at a level of 1 mm in diameter. The bleeding time was recorded on filter paper every 15 min until reaching 120 min, when the experiment was interrupted by tail cauterization of the Sham group. (A) Representative bleeding records obtained on paper filters for WT, rhH6A, and reference products. (B) Scatter plotting of bleeding time values. One-way analysis of variance followed by multiple comparison test (*** P<0.001).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



