Submitted:
02 October 2025
Posted:
03 October 2025
You are already at the latest version
Abstract
The zona pellucida (ZP) is a glycoprotein-rich extracellular matrix essential for fertiliza-tion, early embryonic development, and implantation. Beyond its core functions, the ZP undergoes dynamic remodeling during oocyte maturation, involving regulated synthesis, assembly and conformational changes. Oviductal glycoprotein 1 (OVGP1; oviductin) in-tegrates into the ZP, modulating pore size, glycan composition, and structural homogene-ity to establish a species-specific barrier that prevents polyspermy and fine-tunes sperm–oocyte interactions. Exploiting ZP remodeling in assisted reproductive technologies offers new opportunities to enhance fertilization efficiency, embryo quality, and implantation success, including via assisted hatching or modulating ZP properties to overcome fertility limitations.
Keywords:
assisted reproductive techniques
; fertilization
; hatching
; maturation
; ovastacin
; oviductin
; sperm binding
; zona pellucida glycoproteins
1. Introduction
The zona pellucida (ZP) is a glycoprotein-rich extracellular matrix surrounding the mammalian oocyte, essential for oogenesis, fertilisation, and early embryonic development [1]. It is mainly composed of four glycoproteins: ZP1, ZP2, ZP3, and ZP4, whose structural integrity depends on non-covalent interactions. Each glycoprotein contains a linker region and a ZP module with two conserved subdomains, ZP-N and ZP-C, that regulate activity [2]. Its three-dimensional structure resembles an Ig-like fold [3]. The ZP is vital for species-specific fertilization as this barrier mediates sperm-oocyte binding. Previous research has examined the ability of sperm from distant mammalian orders (Carnivora, Primates, and Rodentia) to penetrate bovine oocytes by examining the role of bovine oviductal fluid (OF) and species-specific oviductal glycoprotein (OVGP1 or oviductin) from bovine, murine, or human sources in modulating the species-specificity of bovine and murine oocytes. Sperm from all the species were found to penetrate intact bovine ovarian oocytes to form hybrid embryos. However, contact with OF or bovine, murine, or human OVGP1, conferred the ZP species-specificity, allowing only the penetration of the corresponding sperm regardless of the ZP’s origin [4]. Glycosilation and microstructural analyses revealed that OVGP1 covers the pores present in the ZP and that OVGP1 glycosylation determines sperm specificity. This suggests specific fertilization capacity is acquired in the oviduct through the ZP’s incorporation of specific oviductin. Together, these findings highlight the importance of the ZP in oocyte maturation processes, fertilization, and embryo development, underscoring the need for further investigations to enhance current assisted reproductive techniques (ARTs).
2. Zona Pellucida Composition and Its Evolutionary Origin
The ZP is an extracellular matrix that surrounds all mammalian oocytes, eggs, and early embryos and plays vital roles during oogenesis, fertilization, and preimplantation development. The ZP is composed of three or four glycosylated proteins, ZP1–4, that are synthesized, processed, secreted, and assembled into long, cross-linked fibrils by growing oocytes. ZP proteins have an immunoglobulin-like three-dimensional structure and a ZP domain that consists of two subdomains, ZP-N and ZP-C, with ZP-N of ZP2 and ZP3 required for fibril assembly [2]. A ZP2–ZP3 dimer is located periodically along ZP fibrils that are cross-linked by ZP1, a protein with a proline-rich N terminus. Fibrils in the inner and outer regions of the ZP are oriented perpendicular and parallel to the oolemma, respectively, giving the ZP a multilayered appearance. Upon fertilization of eggs, modification of ZP2 and ZP3 results in changes in the ZP’s physical and biological properties that have important consequences. Certain structural features of ZP proteins suggest that they may be amyloid-like proteins. Following the fertilization of an egg by a single sperm, the egg coat or ZP hardens, and polyspermy is irreversibly blocked. These events are associated with the cleavage of the N-terminal region (NTR) of glycoprotein ZP2, a major subunit of ZP filaments. ZP2 processing is thought to inactivate sperm binding to the ZP, but its molecular consequences and connection with ZP hardening are unknown. Biochemical and structural studies show that cleavage of ZP2 triggers its oligomerization. Moreover, the structure of a native vertebrate egg coat filament, combined with AlphaFold predictions of human ZP polymers, reveals that two protofilaments consisting of type I (ZP3) and type II (ZP1/ZP2/ZP4) components interlock into a left-handed double helix from which the NTRs of type II subunits protrude [5]. Together, these data suggest that oligomerization of cleaved ZP2 NTRs extensively cross-links ZP filaments, rigidifying the egg coat and making it physically impenetrable to sperm.
ZP composition differs among species: in mice, it consists of ZP1, ZP2 (ZPA), and ZP3 (ZPC), while in cows, pigs, and dogs it is formed by ZP2 (ZPA), ZP4 (ZPB), and ZP3 (ZPC) [6]. Humans, rats, horses, cats, monkeys, rabbits, and hamsters express all four proteins [7,8] (Table 1; Figure 1). ZP thickness also varies, being ~16 μm in cows, 18 μm in pigs, 12 μm in humans, and 8 μm in mice [9,10,11], with pore sizes ranging from 50 to 182 nm [12]. Despite high conservation, the ZP exhibits structural variability contributing to species-specific fertilization. In many mammals, it initially acts as a non-specific barrier until interacting with oviductal fluid (OF), particularly OVGP1. Nevertheless, its conserved roles include sperm binding and selection, induction of the acrosome reaction, prevention of polyspermy, and protection of the oocyte and embryo [4]. These functions are both mechanical and immunological. The ZP also regulates embryo hatching and implantation, after which it is shed.
Structurally, the ZP is a heterogeneous matrix. ZP2 and ZP3 polymerize through their ZP-N subdomains, while ZP1 acts as a molecular crosslinker via its proline-rich N-terminal extension, stabilizing and conferring flexibility to the network [5,14]. These glycoproteins are secreted mainly by the oocyte, though granulosa cells contribute in some species, including humans [11,15]. They assemble into a fibrous network that provides elasticity, birefringence, and porosity, organized into asymmetrical outer, intermediate, and inner layers visible under polarized light microscopy [16]. In the mature ZP, filaments form two intertwined protofilaments in a left-handed helix: type I subunits (ZP3) and type II components (ZP1, ZP2, ZP4), replacing the earlier parallel–perpendicular model (Figure 2). After fertilization, N-terminal cleavage of ZP2 initiates oligomerization and cross-linking, stiffening the ZP and establishing a permanent polyspermy block in most mammals [5,17]. However, in lagomorphs such as rabbits, polyspermy prevention occurs at the oolemma, with multiple spermatozoa present in the perivitelline space [18,19].
The murine model has been central to functional studies of ZP proteins. ZP1-deficient female mice remain fertile but with reduced reproductive success, whereas ZP2- or ZP3-null mice are infertile and produce eggs without a ZP [21]. These mice also fail to establish gap junctions with follicular cells, impairing oocyte-follicle communication and resulting in infertility due to absent growing oocytes and multilayered follicles [22], although fertilization can be rescued by ICSI. Comparable effects have been reported in humans: mutations in ZP genes cause abnormal or absent ZPs, impaired oogenesis, and infertility [23,24,25,26,27,28,29]. For instance, hZP4 variants are associated with thin or irregular ZPs, suggesting defective assembly [30]. The ZP presence is conserved across vertebrates, with homologous envelopes in fish, amphibians, and reptiles that share structural and functional features, particularly in sperm-egg interactions. Phylogenetic analyses show that ZP genes underwent duplication and divergence, giving rise to the distinct ZP1–ZP4 proteins in mammals as species-specific adaptations for fertilization and polyspermy prevention. As noted above, while humans retain all four glycoproteins, mice lack ZP4 and cows, dogs, and pigs lack ZP1 [15,31]. Missing genes often persist as pseudogenes, with functional compensation by others. Conservation of ZP1–ZP3 ranges between 60–98% [32]. Sequence comparisons with human ZP proteins classify species into four groups: Group I (trout, 33%), Group II (frog, chicken, possum, 43–51%), Group III (mouse, rat, cow, pig, dog, 64–69%), and Group IV (macaques, chimpanzees, 93–99%), with >40% identity generally indicating functional conservation [33].
Gene subfamilies include ZPA, ZPB, ZPC, and ZPX [34], but other analyses identify six ancestral subfamilies: ZPA/ZP2, ZPB/ZP4, ZPC/ZP3, ZP1, ZPD, and ZPAX [35]. ZPD is restricted to some bird and amphibian lineages, while in monotremes like platypus, eight subfamilies exist (ZP1, ZP2, ZP3a, ZP3b, ZP3-2, ZP4, ZPAX, ZPY) [36]. In marsupials, ZP3 diversified into three subfamilies (ZP3-1a, ZP3-1b, ZP3-1c) [15]. These likely arose from duplications in a common ancestor of marsupials and placentals, with subsequent simplification in placental mammals, where most subfamilies were lost and only one functional ZP3 was retained, possibly reflecting adaptations to internal fertilization.
Traditionally, ZP3 has been considered the primary sperm receptor, mediating binding of capacitated sperm, with ZP2 acting as a secondary receptor for reacted sperm [37]. However, recent work shows that ZP2 also binds unreacted sperm through its N-terminal domain, triggering conformational changes that block polyspermy [20]. In humans, hZP2 alone can induce sperm binding and acrosome reaction, while hZP1 also contributes. Current proteomic and transcriptomic approaches are expanding knowledge of ZP proteins [15]. Most placental mammals, including humans, hamsters, rabbits, and cats, lack ZPY, ZPAX, and two ZP3 subfamilies, retaining only four [38,39,40,41]. Some species, like humans, chimpanzees, and cows, retain ZPAX pseudogenes, while others, including dogs, cows, and rodents, show pseudogenization of ZP1 or ZP4 [15,35,40].
3. Zona Pellucida Maturation
3.1. Zona Pellucida Formation During Ovarian Development
During follicular maturation, granulosa cells differentiate and acquire the ability to synthesize specific ZP glycoproteins. The ovarian microenvironment, enriched in signaling molecules and growth factors, strongly influences granulosa cell function and ZP formation. This dual origin highlights the interplay between oocytes and somatic cells in ZP formation, which is essential for fertilization and for early development. As noted, ZP thickness varies among species, for instance, ~6 µm in mice and ~20 µm in humans [21]. ZP formation begins early in folliculogenesis, when proteins secreted by the oocyte assemble into a fibrillar matrix that thickens as the oocyte matures [11,42]. While ZP maturation is mainly driven by the oocyte, some studies suggest cooperation with cumulus cells [11,15]. Communication between the oocyte and cumulus cells through gap junctions across the ZP has been documented in several species, including humans, monkeys, cows, pigs, dogs, and rabbits [43,44,45]. This bidirectional signaling ensures coordinated development of both cumulus cells and ZP. During oocyte maturation, cumulus expansion occurs in parallel with ZP thickening and increased birefringence (Table 2).
Oocyte maturation involves structural ZP changes synchronized with nuclear and cytoplasmic events, ensuring meiotic resumption, fertilization, and embryonic development. In bovine oocytes, cumulus–oocyte junction pores narrow during maturation [47]. In humans, immature ZPs appear smooth and non-porous, whereas mature ZPs become rougher and reticulated [48]. In pigs, the immature ZP is porous and loose, but becomes denser and more mesh-like upon maturation [48]. By contrast, equine ZPs remain compact with small pores regardless of maturity, underscoring interspecies differences. In pigs, incomplete ZP maturation during IVF likely contributes to high polyspermy rates, a defect partially corrected by OF exposure [49]. In cattle, both pore number and size correlate with sperm binding and penetration efficiency, serving as indicators of oocyte quality [50]. At ovulation, the ZP undergoes dynamic remodeling of its glycoprotein composition and architecture [51]. These changes, mediated by molecular signals and cellular interactions, involve rearrangements in glycoprotein distribution that influence both ZP structure and its role in sperm–egg recognition (Figure 3).
3.2. Synthesis and Polymerization of ZP Proteins
During folliculogenesis, oocytes secrete ZP glycoproteins together with a granulosa cell contribution in a coordinated process. ZP subunits co-polymerize into a two-protofilament filament: ZP3 provides the scaffold, ZP1 cross-links filaments via disulfide bonds, and ZP2, once cleaved after fertilization, oligomerizes and contributes to matrix hardening to block polyspermy [20]. ZP4, when present, supports fibril stabilization. ZP formation is regulated by hormonal signals, particularly FSH and estrogen, which control ZP gene expression. Altered expression or mutations in these genes can cause infertility due to defective ZP assembly [27].
3.3. Fertilization and ZP Modifications
Fertilization is the fusion of a haploid oocyte and a haploid sperm to form a diploid zygote, involving a series of cellular and biochemical events: the sperm acrosome reaction, recognition, binding, and penetration of the ZP, and fusion with the oolemma [52], with the ZP not directly involved in this last step. First, sperm undergo the acrosome reaction, in which the plasma membrane fuses with the outer acrosomal membrane, releasing hydrolytic enzymes and exposing previously hidden surface ligands [53]. Both acrosome-intact (AI) and acrosome-reacted (AR) sperm can bind the ZP [54], but only AR sperm reach the perivitelline space and fuse with the oolemma, making this reaction essential. Although the ZP was historically seen as the main trigger, only ~5% of sperm reaching the oocyte in mice are AI [55].
Next, sperm must penetrate the ZP, recognizing it in a species-specific manner. The identity of the ZP molecule acting as the sperm receptor remains debated [11,21,52]. Studies with purified human ZP proteins suggest ZP1, ZP3, and ZP4 bind mainly to AI sperm, while ZP2 prefers AR sperm [11]. In transgenic mice, human sperm bind oocytes expressing ZP2 but not ZP1, ZP3, or ZP4, suggesting a key role for ZP2 [11]. Proteolytic cleavage of ZP2’s N-terminal reduces sperm binding [56], and early domain-swapping studies supported ZP2 as the primary receptor [57]. However, mice expressing ZP2 lacking the N1 domain maintain binding capacity but show poor penetration and severe subfertility [20], casting doubt on ZP2’s receptor role. Conversely, ZP3, particularly its C-terminal region or associated oligosaccharides, may act as the sperm receptor [52]. Gene-deletion studies remain inconclusive, as ZP2 and ZP3 are both required for ZP formation in mice [58]. How sperm traverse the ZP network remains unclear [52].
After fertilization, the ZP is modified to block polyspermy. The cortical reaction releases enzymes that cleave ZP2, primarily via ovastacin and crosslink ZP3 through transglutaminase 2, increasing ZP rigidity [59]. This “ZP hardening” prevents further sperm penetration and reinforces structural integrity [60]. While ZP3 has been historically thought to trigger the acrosome reaction, most sperm initiate it before contacting the ZP, during transit through the oviduct or cumulus [61,62]. ZP3 primarily maintains ZP integrity, whereas ZP2 mediates post-acrosome-reaction binding, enabling sperm to reach the oolemma [57]. In bovines, our group showed that in vitro matured (IVM) oocytes, although genomically and cytoplasmically competent, often have porous ZPs. Exposure to OF or in vivo conditions induces conformational changes, incorporating OVGP1 and producing a more homogeneous, less porous structure, which may help prevent polyspermy. While ZP thinning is often attributed to blastomere divisions, proteolytic activity in the female tract also remodels the ZP to facilitate embryo hatching, influenced by asynchronous divisions and maternal factors [4,63]. These structural and functional changes are essential for hatching and implantation.
3.4. Journey Through the Oviduct
In animals with internal fertilization, the oviduct plays a central role by providing a specialized environment that supports gamete transport, final maturation, fertilization, and early embryonic development. Connecting the ovaries to the uterus, the oviduct comprises four anatomical regions: the infundibulum, ampulla, isthmus, and uterine-tubal junction. During ovulation, metaphase II oocytes surrounded by expanded cumulus cells are released into the infundibulum [13]. As they transit to the ampulla, oocytes undergo physiological modifications, notably within the ZP, the glycoprotein-rich extracellular matrix surrounding the oocyte [64].
Several studies emphasize the role of the ampullary region in facilitating both fertilization and ZP maturation, a process commonly termed ZP hardening. This modification occurs in two sequential phases: the first within the dominant follicle, and the second during the oocyte’s passage through the ampulla [13,65,66]. Coy et al. [49] introduced the concept of pre-fertilization ZP hardening, distinct from the post-fertilization cortical reaction. Oviduct-mediated ZP hardening is conserved across ungulates and complements oocyte-derived responses. In vitro co-culture with ampullary epithelial cells enhances ZP hardening, reduces polyspermy, and improves fertilization. Conditioned media from these cells, particularly when derived from direct interactions with cumulus-oocyte complexes, further support ZP hardening and early embryonic development [67,68].
Oviductal secretory cells produce high molecular weight glycoproteins, termed oviduct-specific glycoproteins (OGPs), which bind the ZP and perivitelline space during oocyte transit, with some persisting on embryos until implantation. Porcine IVF studies show that OVGP1 and peri-ovulatory OF promote ZP hardening and monospermy [64]. Cross-species studies using bovine OF reveal additional factors, including heat shock proteins (HSPs), protein disulfide isomerases (PDIs), and heparin-like glycosaminoglycans (GAGs), which collectively enhance ZP resistance to enzymatic digestion and sperm penetration. These findings illustrate that the oviduct provides a hormonally regulated, species-specific microenvironment actively shaping ZP maturation and fertilization competence [49].
Recent studies show that OVGP1 also remodels ZP architecture during oviductal transit. The C-terminal region of OVGP1 penetrates the ZP, creating a porous, net-like structure characteristic of fertilization-competent oocytes, increasing resistance to proteolytic digestion and promoting monospermy. Truncated OVGP1 lacking this domain fails to remodel the ZP effectively [63]. OVGP1 is also endocytosed by the oocyte, suggesting roles in oocyte–ZP communication and in early development. Immunolocalization studies detect OVGP1 in the microvilli of zygotes, the perivitelline space, and early embryo multivesicular bodies [69].
Although OVGP1 is critical in some species, knockout and pseudogene studies indicate it is not essential for fertilization in all species [70,71]. Its consistent in vivo and in vitro expression, including in bovine biomimetic systems [72], underscores its physiological relevance, where functional. This highlights the species-specific and redundant nature of oviductal regulation of ZP maturation [73]. In addition to soluble glycoproteins, extracellular vesicles (EVs) from oviductal epithelial cells carry proteins, lipids, and RNAs that traverse the ZP and deliver bioactive cargo, including OVGP1, into the ooplasm [74,75]. EVs from cumulus cells similarly penetrate the ZP during in vitro maturation, indicating a conserved intercellular communication mechanism [76]. These findings reinforce the concept of the ZP as a dynamic barrier facilitating selective molecular exchange during oviductal maturation and fertilization. The oviduct also supports early embryo development and transport, guiding fertilized oocytes to the uterine cavity. It provides growth factor-rich secretions, protects embryos from oxidative stress, and mediates the first embryo–maternal interactions [13,60,73,74]. Transport likely involves ciliary beating, muscular contractions, and fluid flow [77].
During the oocyte-to-embryo transition, the ZP acts as a protective barrier against mechanical stress and enzymatic degradation, maintaining structural integrity until blastocyst hatching [21]. ZP elasticity may serve as an indicator of embryo quality [78]. In vivo-derived porcine embryos exhibit more digestion-resistant ZP than in vitro counterparts, highlighting the role of oviduct-specific modifications [79]. Knockouts of ZP1 in mice or ZP4 in rabbits cause structural fragility, impaired transport, and developmental arrest, underscoring the ZP’s mechanical role in embryo survival [58,80]. Finally, the ZP facilitates smooth embryo transport by maintaining spherical shape and reducing adhesion to the oviductal epithelium [77,81]. Together, these observations highlight the dynamic crosstalk between the oviduct, oocyte, and early embryo, emphasizing the importance of replicating this microenvironment in ARTs.
3.5. Role in Oocyte Fertilization, Embryo Protection, and Embryo Implantation
During fertilization, the fusion of two highly differentiated haploid gametes is mediated by the ZP, which orchestrates three critical processes: species-specific sperm recognition, sperm binding to the oocyte, and prevention of polyspermy. Fusion of the sperm membrane with the oolemma triggers a cascade of structural and biochemical changes, including ZP hardening, which prevents polyspermy and the formation of nonviable polyploid embryos. One such mechanism is the cortical reaction, where cortical granules release their contents post-fertilization, causing the ZP matrix to lose its capacity to bind additional sperm [1,82].
A key component of this process is the cleavage of ZP2 glycoprotein. This cleavage is mediated by ovastacin, a cortical granule metalloendoprotease encoded by Astl, which targets the N-terminal domain of ZP2, and proteolytically degrades it, thereby preventing further sperm interaction. Functional evidence comes from Astl knockout mouse models, in which ZP2 remains uncleaved after fertilization. Consequently, sperm continue to bind the ZP of fertilized eggs, confirming that ovastacin-mediated ZP2 cleavage is essential for establishing a definitive block to polyspermy [83]. Supporting this, Maddirevula et al. [84] reported that adult women homozygous for a non-functional variant of ovastacin exhibit markedly reduced fertility.
Nishio et al. [20] further demonstrated that, following fertilization, ovastacin cleaves the N-terminal region of ZP2, triggering its oligomerization. This rearrangement results in extensive cross-linking between ZP filaments, reinforcing the structural integrity of the egg coat. The ZP is composed of a left-handed double-helical architecture formed by interwoven type I and type II protofilaments. Upon cleavage, liberated ZP2 fragments self-assemble, stabilizing the matrix, promoting filament compaction, and preventing polyspermy. While fetuin-B is acknowledged as an endogenous inhibitor of ovastacin [20], Von Wiegen et al. [85] showed that disruption of the ovastacin–fetuin-B balance alters ZP2 cleavage timing, causing premature zona hardening and reduced fertilization efficiency. Together, these findings underscore the critical importance of precise regulation of ZP2 proteolysis for ZP remodeling and reproductive success.
Transgenic mice with defective ZP2 cleavage (ZP2^mut) show that sperm continues to bind to the zona after fertilization, although monospermy is maintained. This suggests that ZP2 cleavage is essential for preventing post-fertilization sperm binding but not strictly required to block polyspermy [57]. ZP3 also undergoes rapid post-fertilization modifications, likely induced by cortical granule enzymes, which render it unrecognizable to sperm. Additionally, ZP3 experiences selective covalent crosslinking mediated by transglutaminase 2 (TGM2), a calcium-dependent enzyme secreted from the oocyte cortex via a SNARE-regulated pathway. TGM2 induces isopeptide bonds between glutamine and lysine residues within ZP3, reinforcing ZP structure, increasing matrix rigidity, and limiting further sperm penetration. Biochemical and functional assays in murine and porcine models identify ZP3 as the sole zona substrate of TGM2. Pharmacological inhibition or genetic ablation of TGM2 increases polyspermic fertilization, evidenced by elevated rates of polypronuclear zygotes and decreased zona resistance to proteolytic degradation. Conversely, recombinant TGM2 supplementation restores ZP integrity and reduces polyspermy in a concentration-dependent manner.
These post-fertilization biochemical alterations in ZP2 and ZP3, together with modifications of the ZP matrix and oolemma [20,21,56], collectively establish a robust block to polyspermy. Mouse models have been instrumental in elucidating ZP glycoprotein functions. ZP3 acts not only as a structural component but also as a signaling molecule. In ZP3-deficient mice, no two-cell embryos were recovered after mating, indicating fertilization or early development failure [58]. Similarly, embryos with enzymatically removed ZPs cannot progress through the oviduct and highlighting the ZP’s crucial structural role in embryo protection and transport.
Beyond fertilization, the ZP continues to play a vital role until implantation. It forms a protective shell around the preimplantation embryo, shielding it mechanically and preventing premature interactions with the maternal reproductive tract. In rabbits, embryos lacking ZP4 failed to develop to the blastocyst stage [80]. The intact ZP guides the embryo from the oviduct to the uterus, preventing ectopic implantation by separating the embryo from the oviductal epithelium [86]. Its glycoprotein-rich composition protects the embryo from mechanical stress and enzymatic degradation [87,88], and it may also exert immunomodulatory effects, shielding the embryo from maternal immune recognition [89]. As the embryo reaches the blastocyst stage, the ZP undergoes thinning and degradation during the hatching process, which is essential for successful implantation. Hatching is influenced by three main factors: mechanical expansion of the blastocyst, enzymatic ZP degradation by hydrolases secreted by trophectoderm cells, and proteases from uterine fluids that weaken the ZP [90].
3.6. Specificity of Sperm Binding
Successful mammalian fertilization involves a tightly coordinated sequence: sperm capacitation, acrosome reaction, species-specific sperm-egg recognition, penetration of the ZP, and membrane fusion with the oocyte. Species-specific gamete recognition ensures reproductive success by promoting genetic compatibility and maintaining species integrity. Despite its fundamental role, the molecular basis of sperm-oocyte interaction, particularly the species-specific features, remains partially understood. Evidence indicates that species specificity operates at multiple levels, from initial sperm-egg attraction to gamete membrane fusion [19].
Interspecies differences in binding affinities help prevent cross-species fertilization, as seen with reduced interaction between human JUNO and mouse IZUMO1 [91]. While the IZUMO1–JUNO interaction contributes to species specificity, ZP glycoproteins directly prevent heterologous fertilization. Genetic studies reveal that sperm-ZP binding is primarily mediated by the N-terminal domain of ZP2. Targeted mutagenesis of canonical O-glycosylation sites in mouse ZP3 (Ser^332 and Ser^334) does not impair fertilization, suggesting these carbohydrate moieties are dispensable in rodents [92].
In contrast, in bovine and porcine species, sialylated glycans are crucial for sperm binding and penetration. Terminal sialic acids, particularly α2-3-linked Neu5Ac and Neu5Gc, mediate this interaction, and their enzymatic removal significantly reduces sperm adhesion and fertilization [4]. These findings highlight a dual role for sialic acids in bovine ZP, supporting both nonspecific binding and species-specific recognition, modulated by the sialylation of ZP and OVGP1.
Species-specific fertilization barriers arise from differences in ZP architecture and glycosylation. Human ZP contains sialyl-Lewis^x epitopes absent in murine ZP, while many non-human mammals incorporate Neu5Gc, absent in humans due to CMAH inactivation [93]. These differences provide glycan-level cues for taxon-specific binding, although heterologous fertilization can still occur experimentally [4], suggesting that both ZP structure and glycosylation jointly modulate species specificity.
Extrinsic modulation by the oviductal environment further shapes ZP function. OF induces compositional and structural ZP changes that enhance oocyte maturation and sperm recognition [13,49]. OVGP1, an oviductal glycoprotein, is critical for species-specific fertilization. Only homologous OVGP1 enables sperm binding and penetration, while heterologous OVGP1 or absence of OVGP1 diminishes fertilization efficiency across multiple species, including hamsters, cows, pigs, and humans [4]. OVGP1 penetrates the ZP via its conserved C-terminal region, modifying glycosylation and coating ZP pores with sialic acid-rich oligosaccharides recognized in a species-specific manner. Scanning electron microscopy confirms uniform ZP coating only with homologous OVGP1, whereas heterologous protein or absence of OVGP1 results in incomplete coverage [4,94]. Thus, OVGP1 not only enhances fertilization efficiency but also reinforces species-specific sperm recognition, highlighting a cooperative mechanism in which intrinsic ZP properties and extrinsic oviductal factors jointly determine homologous fertilization.
3.7. Assisted Hatching: Rationale, Technique, and Clinical Outcomes
Zona pellucida thickness in early-stage embryos is strongly associated with implantation success. Comparative analyses have shown that embryos surrounded by zonae pellucidae with inadequate biomechanical properties exhibit reduced implantation potential, thereby reinforcing the concept that the zona pellucida fulfils a dual function in mediating gamete interaction and supporting subsequent embryonic development [95]. Implantation rates have also been correlated with ZP-related events such as birefringence and spontaneous hatching [96].
During in vitro culture, human embryos typically show gradual ZP thinning, whereas zygotes that fail to cleave demonstrate minimal change in ZP thickness [95,97]. Abnormalities in ZP structure or blastocyst hatching are linked to implantation failure and early pregnancy loss [90]. After embryo transfer, roughly 30% of embryos fail to implant, with failure rates rising to nearly 70% for blastocysts that do not undergo spontaneous hatching [96,98]. These observations underscore the critical importance of hatching for successful implantation and pregnancy [99]. Indeed, thawed blastocysts that re-expand or reach the full hatching stage are significantly more likely to result in pregnancy, underscoring the prognostic value of these morphological dynamics [100,101,102]. In vitro-cultured blastocysts often exhibit increased ZP hardening compared to their in vivo counterparts, impairing hatching and subsequent implantation [103]. In this context, assisted hatching has proven effective in enhancing implantation rates in both mouse [104] and human embryos [105].
Hatching is regulated by both physical and molecular factors. Physical factors include blastocyst expansion and ZP rupture, whereas molecular regulation involves multiple genes. Human hatched blastocysts show higher mRNA expression of CTSV, GATA3, and CGB compared to non-hatched counterparts [106]. Furthermore, dynamic expression of the Notch gene family suggests that Notch signaling plays a crucial role in mouse hatching progression, and disruptions in this pathway can impair the process, highlighting its importance in early embryonic development [107].
4. Clinical Implications and Future Perspectives
The ZP is a complex glycoprotein matrix whose absence or structural defects, whether due to abnormalities in glycoprotein composition or in receptors mediating critical events such as sperm recognition, acrosome reaction, or uterine implantation, represent a major cause of infertility in both animal models and humans [23,24,27,28,29]. Just as assisted hatching represents a major advancement in ARTs by facilitating ZP rupture and improving implantation [105], emerging research is exploring novel strategies targeting ZP structure and function to treat infertility and develop non-hormonal contraceptives.
Mutations in genes such as hZP4 can lead to an abnormally thin, discontinuous, and structurally deficient ZP, severely impairing sperm–oocyte interaction and reducing fertilization rates [30]. In a rabbit model, CRISPR/Cas9-mediated inactivation of ZP4 produced a thinner, unstable ZP and complete infertility, highlighting the essential structural role of ZP4 [80]. Partial weakening of the ZP preserved blastocyst development and editing efficiency, whereas complete removal increased mutation rates but compromised embryo viability. These findings illustrate the delicate balance between genome editing efficiency and embryonic health, emphasizing the importance of modulating ZP conditions during experimental manipulations.
Pathogenic variants in ZP1, ZP2, and ZP3 have been identified in women with genuine empty follicle syndrome (GEFS), a rare disorder in which oocytes degenerate despite normal ovarian stimulation [108,109]. These mutations disrupt ZP protein secretion and assembly, leading to infertility. CRISPR-based models provide valuable platforms to study such mutations, enhancing both diagnostics and the development of targeted therapies. Interestingly, not all oocyte-specific genes are essential for fertility: knockout of Oosp1, Oosp2, and Oosp3 in mice did not impair female fertility, highlighting the importance of in vivo functional screens to distinguish essential from dispensable reproductive genes [110]. Overall, CRISPR/Cas9 technologies are proving invaluable not only for precise gene editing but also for functional characterization of reproductive genes, informing which molecular pathways are genuinely critical for female reproductive success. The clinical translation of these approaches requires careful ethical consideration regarding safety, equitable access, and broader societal implications.
Our group recently demonstrated that OVGP1 can structurally modify the ZP to prevent polyspermy and cross-species sperm entry [4]. Specifically, bovine ZPs devoid of cytoplasm and unexposed to OF were penetrable by heterologous sperm, including human, while exposure to OVGP1 restored species-specificity. This approach suggests potential use of bovine ZPs for sperm selection in ICSI cycles, allowing identification of high-quality sperm based on acrosome reaction, DNA integrity, and blastocyst developmental potential.
Detailed understanding of ZP structure and function also enables the development of non-hormonal contraceptives. ZP-based vaccines have been widely explored to induce immunological infertility. For example, a monoclonal antibody (IE-3) or its scFv fragment targeting a defined N-terminal region of ZP2 has been shown to effectively block sperm binding and fertilization in mice without inducing ovarian damage or impairing follicular maturation [111].This peptide-based approach was effective in mouse models, and transgenic assays demonstrated that the human homolog competitively inhibited human sperm binding to humanized mouse oocytes, supporting its potential as a species-specific, non-hormonal contraceptive [112]. Similarly, immunization with recombinant ZP3 proteins has been reported to reduce fertility in animal models, although this effect is largely associated with ovarian follicle loss and apoptosis triggered by oxidative stress [113]. In dogs, native or recombinant ZP glycoproteins effectively inhibited fertility, representing a promising strategy for ethical wildlife population management [82]. Furthermore, targeting sperm proteins that interact with the ZP opens additional avenues for reversible, non-hormonal contraceptive strategies by competitive inhibition of sperm adhesion and penetration [31].
In conclusion, manipulation of the ZP represents a promising frontier in reproductive medicine, with potential applications in personalized infertility treatment, improved IVF outcomes, and the development of next-generation non-hormonal contraceptives.
Author Contributions
Original draft preparation, writing, figure and table preparation and revising, D.F., A.G; figure and table. Rest of the authors revised and edited this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants PID2021-122507OB-I00 awarded to A. Gutierrez-Adán, by Doctorados Industriales 2022 fellowship of the Comunidad de Madrid (IND2022/BIO-23646) awarded to D. de la Fuente and by AFRODITA from the European Union’s Horizon Europe programme under the MSCA Doctoral Network call, Grant Agreement No. 101120126 awarded to M. Prestianni.
Acknowledgments
We thank all colleagues and collaborators who contributed to this work.
Conflicts of Interest
Author Daniel de la Fuente was employed by the Fiv Center Clinic, 28023, Madrid, Spain. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AI | Acrosome-intact |
| AR | Acrosome-reacted |
| ARTs | Assisted Reproductive Techniques |
| EVs | Extracellular vesicles |
| GAGs | Heparin-like glycosaminoglycans |
| GEFs | Genuine empty follicle syndrome |
| HSPs | Heat shock proteins |
| IVM | in vitro matured |
| OF | Oviductal fluid |
| OVGP1 | Oviductal glycoprotein 1 |
| NTR | N-terminal region |
| TGM2 | Transglutaminase 2 |
| ZP | Zona pellucida |
References
- Wassarman, P.M. Zona Pellucida Glycoproteins. J Biol Chem. 2008, 283, 24285–24289. [Google Scholar] [CrossRef]
- Bokhove, M.; Jovine, L. Structure of Zona Pellucida Module Proteins. In Current Topics in Developmental Biology [Internet]; Elsevier, 2018; pp. 413–442, [cited 2025 Sept 9]; Available from: https://linkinghub.elsevier.com/retrieve/pii/S0070215318300395.
- Clark, G.F. The molecular basis of mouse sperm–zona pellucida binding: a still unresolved issue in developmental biology. REPRODUCTION 2011, 142, 377–381. [Google Scholar] [CrossRef]
- De La Fuente, D.; Maroto, M.; Cajas, Y.N.; Canon-Beltran, K.; Fernandez-Gonzalez, R.; Munoz-Maceda, A.; et al. Oviductin sets the species-specificity of the mammalian zona pellucida. eLife 2025, 13, RP101338. [Google Scholar] [CrossRef] [PubMed]
- Nishio, S.; Emori, C.; Wiseman, B.; Fahrenkamp, D.; Dioguardi, E.; Zamora-Caballero, S.; et al. ZP2 cleavage blocks polyspermy by modulating the architecture of the egg coat. Cell 2024, 187, 1440–1459.e24. [Google Scholar] [CrossRef]
- Goudet, G.; Mugnier, S.; Callebaut, I.; Monget, P. Phylogenetic Analysis and Identification of Pseudogenes Reveal a Progressive Loss of Zona Pellucida Genes During Evolution of Vertebrates1. Biol Reprod. 2008, 78, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Rico, M.J.; Jiménez-Movilla, M.; Llop, E.; Pérez-Oliva, A.B.; Ballesta, J.; Gutiérrez-Gallego, R.; et al. Hamster Zona Pellucida Is Formed by Four Glycoproteins: ZP1, ZP2, ZP3, and ZP4. J Proteome Res. 2009, 8, 926–941. [Google Scholar] [CrossRef] [PubMed]
- Stetson, I.; Avilés, M.; Moros, C.; García-Vázquez, F.A.; Gimeno, L.; Torrecillas, A.; et al. Four glycoproteins are expressed in the cat zona pellucida. Theriogenology 2015, 83, 1162–1173. [Google Scholar] [CrossRef]
- Tpfer-Petersen, E.; Ekhlasi-Hundrieser, M.; Tsolova, M. Glycobiology of fertilization in the pig. Int J Dev Biol. 2008, 52, 717–736. [Google Scholar] [CrossRef]
- Wiesak, T.; Wasielak, M.; Złotkowska, A.; Milewski, R. Effect of vitrification on the zona pellucida hardening and follistatin and cathepsin B genes expression and developmental competence of in vitro matured bovine oocytes. Cryobiology 2017, 76, 18–23. [Google Scholar] [CrossRef]
- Gupta, S.K. Human Zona Pellucida Glycoproteins: Binding Characteristics With Human Spermatozoa and Induction of Acrosome Reaction. Front Cell Dev Biol. 2021, 9, 619868. [Google Scholar] [CrossRef]
- Vanroose, G.; Nauwynck, H.; Soom, A.V.; Ysebaert, M.T.; Charlier, G.; Oostveldt, P.V.; et al. Structural Aspects of the Zona Pellucida of In Vitro-Produced Bovine Embryos: A Scanning Electron and Confocal Laser Scanning Microscopic Study1. Biol Reprod. 2000, 62, 463–469. [Google Scholar] [CrossRef]
- Avilés, M.; Gutiérrez-Adán, A.; Coy, P. Oviductal secretions: will they be key factors for the future ARTs? MHR Basic Sci Reprod Med. 2010, 16, 896–906. [Google Scholar] [CrossRef]
- Litscher, E.S.; Wassarman, P.M. Zona Pellucida Proteins, Fibrils, and Matrix. Annu Rev Biochem. 2020, 89, 695–715. [Google Scholar] [CrossRef]
- Moros-Nicolás, C.; Chevret, P.; Jiménez-Movilla, M.; Algarra, B.; Cots-Rodríguez, P.; González-Brusi, L.; et al. New Insights into the Mammalian Egg Zona Pellucida. Int J Mol Sci. 2021, 22, 3276. [Google Scholar] [CrossRef] [PubMed]
- Michelmann, H.; Rath, D.; Töpfer-Petersen, E.; Schwartz, P. Structural and Functional Events on the Porcine Zona Pellucida During Maturation, Fertilization and Embryonic Development: a Scanning Electron Microscopy Analysis. Reprod Domest Anim. 2007, 42, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Okumura, H.; Mizuno, A.; Iwamoto, E.; Sakuma, R.; Nishio, S.; Nishijima Kichi et, a.l. New insights into the role of microheterogeneity of ZP3 during structural maturation of the avian equivalent of mammalian zona pellucida. Mishra B, editor. PLOS ONE. 2023, 18, e0283087. [Google Scholar] [CrossRef]
- Gardner, A.J.; Evans, J.P. Mammalian membrane block to polyspermy: new insights into how mammalian eggs prevent fertilisation by multiple sperm. Reprod Fertil Dev. 2006, 18, 53. [Google Scholar] [CrossRef] [PubMed]
- Bhakta, H.H.; Refai, F.H.; Avella, M.A. The molecular mechanisms mediating mammalian fertilization. Development. 2019, 146, dev176966. [Google Scholar] [CrossRef]
- Nishio, S.; Emori, C.; Wiseman, B.; Fahrenkamp, D.; Dioguardi, E.; Zamora-Caballero, S.; et al. ZP2 cleavage blocks polyspermy by modulating the architecture of the egg coat. Cell. 2024, 187, 1440–1459.e24. [Google Scholar] [CrossRef]
- Wassarman, P.M.; Litscher, E.S. Female fertility and the mammalian egg’s zona pellucida. Histol Histopathol. 2024, 39, 1273–1284. [Google Scholar]
- Li, R.; Albertini, D.F. The road to maturation: somatic cell interaction and self-organization of the mammalian oocyte. Nat Rev Mol Cell Biol. 2013, 14, 141–152. [Google Scholar] [CrossRef]
- Dai, C.; Hu, L.; Gong, F.; Tan, Y.; Cai, S.; Zhang, S.; et al. ZP2 pathogenic variants cause in vitro fertilization failure and female infertility. Genet Med. 2019, 21, 431–440. [Google Scholar] [CrossRef]
- Sun, L.; Fang, X.; Chen, Z.; Zhang, H.; Zhang, Z.; Zhou, P.; et al. Compound heterozygous ZP1 mutations cause empty follicle syndrome in infertile sisters. Hum Mutat. 2019, 40, 2001–2006. [Google Scholar] [CrossRef]
- Liu, M.; Shen, Y.; Zhang, X.; Wang, X.; Li, D.; Wang, Y. Novel biallelic loss-of-function variants in ZP1 identified in an infertile female with empty follicle syndrome. J Assist Reprod Genet. 2020, 37, 2151–2157. [Google Scholar] [CrossRef]
- Zhang, Z.; Shangguan, T.; Li, Y.Y.; He, W. Infertility due to Lack of Zona Pellucida Caused by a Compound Heterozygous Mutation in ZP1 Gene. Reprod Dev Med. 2018, 2, 183–186. [Google Scholar] [CrossRef]
- Cao, Q.; Zhao, C.; Zhang, X.; Zhang, H.; Lu, Q.; Wang, C.; et al. Heterozygous mutations in ZP1 and ZP3 cause formation disorder of ZP and female infertility in human. J Cell Mol Med. 2020, 24, 8557–8566. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, K.; Dioguardi, E.; Nishio, S.; Villa, A.; Han, L.; Matsuda, T.; et al. Molecular basis of egg coat cross-linking sheds light on ZP1-associated female infertility. Nat Commun. 2019, 10, 3086. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zeng, Y.; Chen, H.; Zhou, Z.; Fu, J.; Sang, Q.; et al. A novel homozygous variant in ZP2 causes abnormal zona pellucida formation and female infertility. J Assist Reprod Genet. 2021, 38, 1239–1245. [Google Scholar] [CrossRef]
- Wei, X.; Li, Y.; Liu, Q.; Liu, W.; Yan, X.; Zhu, X.; et al. Mutations in ZP4 are associated with abnormal zona pellucida and female infertility. J Clin Pathol. 2022, 75, 201–204. [Google Scholar] [CrossRef]
- Tumova, L.; Zigo, M.; Sutovsky, P.; Sedmikova, M.; Postlerova, P. Ligands and Receptors Involved in the Sperm-Zona Pellucida Interactions in Mammals. Cells. 2021, 10, 133. [Google Scholar] [CrossRef]
- Litscher, E.S.; Wassarman, P.M. Zona Pellucida Proteins, Fibrils, and Matrix. Annu Rev Biochem. 2020, 89, 695–715. [Google Scholar] [CrossRef] [PubMed]
- Chothia, C.; Gough, J.; Vogel, C.; Teichmann, S.A. Evolution of the Protein Repertoire. Science. 2003, 300, 1701–1703. [Google Scholar] [CrossRef]
- Spargo, S.C.; Hope, R.M. Evolution and Nomenclature of the Zona Pellucida Gene Family. Biol Reprod. 2003, 68, 358–362. [Google Scholar] [CrossRef]
- Goudet, G.; Mugnier, S.; Callebaut, I.; Monget, P. Phylogenetic Analysis and Identification of Pseudogenes Reveal a Progressive Loss of Zona Pellucida Genes During Evolution of Vertebrates1. Biol Reprod. 2008, 78, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.M.; Tian, H.F.; Hu, Q.M.; Meng, Y.; Xiao, H.B. Evolution and multiple origins of zona pellucida genes in vertebrates. Biol Open. 2018, 7, bio036137. [Google Scholar] [CrossRef]
- Bleil, J.D.; Greve, J.M.; Wassarman, P.M. Identification of a secondary sperm receptor in the mouse egg zona pellucida: Role in maintenance of binding of acrosome-reacted sperm to eggs. Dev Biol. 1988, 128, 376–385. [Google Scholar] [CrossRef]
- Lefievre, L. Four zona pellucida glycoproteins are expressed in the human. Hum Reprod. 2004, 19, 1580–1586. [Google Scholar] [CrossRef]
- Izquierdo-Rico, M.J.; Jiménez-Movilla, M.; Llop, E.; Pérez-Oliva, A.B.; Ballesta, J.; Gutiérrez-Gallego, R.; et al. Hamster Zona Pellucida Is Formed by Four Glycoproteins: ZP1, ZP2, ZP3, and ZP4. J Proteome Res. 2009, 8, 926–941. [Google Scholar] [CrossRef]
- Stetson, I.; Izquierdo-Rico, M.J.; Moros, C.; Chevret, P.; Lorenzo, P.L.; Ballesta, J.; et al. Rabbit zona pellucida composition: A molecular, proteomic and phylogenetic approach. J Proteomics. 2012, 75, 5920–5935. [Google Scholar] [CrossRef]
- Stetson, I.; Avilés, M.; Moros, C.; García-Vázquez, F.A.; Gimeno, L.; Torrecillas, A.; et al. Four glycoproteins are expressed in the cat zona pellucida. Theriogenology. 2015, 83, 1162–1173. [Google Scholar] [CrossRef] [PubMed]
- Wassarman, P.M.; Litscher, E.S. The Mouse Egg’s Zona Pellucida. In Current Topics in Developmental Biology [Internet]; Elsevier, 2025; pp. 331–356, [cited 2025 Sept 10]; Available from: https://linkinghub.elsevier.com/retrieve/pii/S0070215318300036.
- Bleil, J.D.; Wassarman, P.M. Galactose at the nonreducing terminus of O-linked oligosaccharides of mouse egg zona pellucida glycoprotein ZP3 is essential for the glycoprotein’s sperm receptor activity.
- Dunbar, B.; Avery, S.; Lee, V.; Prasad, S.; Schwahn, D.; Schwoebel, E.; et al. The mammalian zona pellucida: its biochemistry, immunochemistry, molecular biology, and developmental expression. Reprod Fertil Dev. 1994, 6(, 331. [Google Scholar] [CrossRef]
- Bousquet, D.; Léveillé, M.C.; Roberts, K.D.; Chapdelaine, A.; Bleau, G. The cellular origin of the zona pellucida antigen in the human and hamster. J Exp Zool. 1981, 215, 215–218. [Google Scholar] [CrossRef]
- Pelletier, C.; Keefe, D.L.; Trimarchi, J.R. Noninvasive polarized light microscopy quantitatively distinguishes the multilaminar structure of the zona pellucida of living human eggs and embryos. Fertil Steril. 2004, 81, 850–856. [Google Scholar] [CrossRef]
- Suzuki, H.; Yang, X.; Foote, R.H. Surface alterations of the bovine oocyte and its investments during and after maturation and fertilization in vitro. Mol Reprod Dev. 1994, 38, 421–430. [Google Scholar] [CrossRef]
- Familiari, G.; Nottola, S.A.; Macchiarelli, G.; Micara, G.; Aragona, C.; Motta, P.M. Human zona pellucida during in vitro fertilization: An ultrastructural study using saponin, ruthenium red, and osmium-thiocarbohydrazide. Mol Reprod Dev. 1992, 32, 51–61. [Google Scholar] [CrossRef]
- Coy, P.; Cánovas, S.; Mondéjar, I.; Saavedra, M.D.; Romar, R.; Grullón, L.; et al. Oviduct-specific glycoprotein and heparin modulate sperm–zona pellucida interaction during fertilization and contribute to the control of polyspermy. Proc Natl Acad Sci. 2008, 105, 15809–15814. [Google Scholar] [CrossRef]
- Travnickova, I.; Hulinska, P.; Sladek, Z.; Skowronski, M.T.; Machatkova, M. Changes of the zona pellucida patterns during oocyte maturation, fertilization and embryo development in mammals: mini-review. Med J Cell Biol. 2022, 10, 23–28. [Google Scholar] [CrossRef]
- Oikawa, T.; Yanagimachi, R.; Nicolson, G.L. SPECIES DIFFERENCES IN THE LECTIN-BINDING SITES ON THE ZONA PELLUCIDA OF RODENT EGGS. Reproduction. 1975, 43, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, E.; Wright, G.J. Find and fuse: Unsolved mysteries in sperm–egg recognition. PLOS Biol. 2020, 18, e3000953. [Google Scholar] [CrossRef] [PubMed]
- Hirohashi, N.; Yanagimachi, R. Sperm acrosome reaction: its site and role in fertilization†. Biol Reprod. 2018, 99, 127–133. [Google Scholar] [CrossRef]
- Wassarman, P.M.; Litscher, E.S. Zona Pellucida Genes and Proteins: Essential Players in Mammalian Oogenesis and Fertility. Genes. 2021, 12, 1266. [Google Scholar] [CrossRef] [PubMed]
- La Spina, F.A.; Puga Molina, L.C.; Romarowski, A.; Vitale, A.M.; Falzone, T.L.; Krapf, D.; et al. Mouse sperm begin to undergo acrosomal exocytosis in the upper isthmus of the oviduct. Dev Biol. 2016, 411, 172–182. [Google Scholar] [CrossRef]
- Kang, I.; Koo, M.; Yoon, H.; Park, B.S.; Jun, J.H.; Lee, J. Ovastacin: An oolemma protein that cleaves the zona pellucida to prevent polyspermy. Clin Exp Reprod Med. 2023, 50, 154–159. [Google Scholar] [CrossRef]
- Avella, M.A.; Baibakov, B.; Dean, J. A single domain of the ZP2 zona pellucida protein mediates gamete recognition in mice and humans. J Cell Biol. 2014, 205, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Rankin, T.; Familari, M.; Lee, E.; Ginsberg, A.; Dwyer, N.; Blanchette-Mackie, J.; et al. Mice homozygous for an insertional mutation in the Zp3 gene lack a zona pellucida and are infertile. Development. 1996, 122, 2903–2910. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Lu, Y.; Miao, Y.; Dai, X.; Zhang, Y.; Xiong, B. Transglutaminase 2 crosslinks zona pellucida glycoprotein 3 to prevent polyspermy. Cell Death Differ. 2022, 29, 1466–1473. [Google Scholar] [CrossRef]
- Coy, P.; García-Vázquez, F.A.; Visconti, P.E.; Avilés, M. Roles of the oviduct in mammalian fertilization. REPRODUCTION. 2012, 144, 649–660. [Google Scholar] [CrossRef]
- Jin, M.; Fujiwara, E.; Kakiuchi, Y.; Okabe, M.; Satouh, Y.; Baba, S.A.; et al. Most fertilizing mouse spermatozoa begin their acrosome reaction before contact with the zona pellucida during in vitro fertilization. Proc Natl Acad Sci. 2011, 108, 4892–4896. [Google Scholar] [CrossRef]
- Muro, Y.; Buffone, M.G.; Okabe, M.; Gerton, G.L. Function of the Acrosomal Matrix: Zona Pellucida 3 Receptor (ZP3R/sp56) Is Not Essential for Mouse Fertilization1. Biol Reprod [Internet] 2012, 86. [cited 2025 Sept 10]. Available from: https://academic.oup.com/biolreprod/article-lookup/doi/10.1095/biolreprod.111.095877. [CrossRef]
- Algarra, B.; Maillo, V.; Avilés, M.; Gutiérrez-Adán, A.; Rizos, D.; Jiménez-Movilla, M. Effects of recombinant OVGP1 protein on in vitro bovine embryo development. J Reprod Dev. 2018, 64, 433–443. [Google Scholar] [CrossRef]
- Mondéjar, I.; Avilés, M.; Coy, P. The human is an exception to the evolutionarily-conserved phenomenon of pre-fertilization zona pellucida resistance to proteolysis induced by oviductal fluid. Hum Reprod. 2013, 28, 718–728. [Google Scholar] [CrossRef] [PubMed]
- Avilés, M.; El-Mestrah, M.; Jaber, L.; Castells, M.T.; Ballesta, J.; Kan, F.W.K. Cytochemical demonstration of modification of carbohydrates in the mouse zona pellucida during folliculogenesis. Histochem Cell Biol. 2000, 113, 207–219. [Google Scholar] [CrossRef]
- Avilés, M.; Okinaga, T.; Shur, B.D.; Ballesta, J. Differential expression of glycoside residues in the mammalian zona pellucida. Mol Reprod Dev. 2000, 57, 296–308. [Google Scholar] [CrossRef]
- Fang, X.; Bang, S.; Tanga, B.; Seo, C.; Zhou, D.; Seong, G.; et al. Oviduct epithelial cell-derived extracellular vesicles promote the developmental competence of IVF porcine embryos. Mol Med Rep. 2023, 27, 122. [Google Scholar] [CrossRef]
- Miyashita, N.; Akagi, S.; Somfai, T.; Hirao, Y. Serum-free spontaneously immortalized bovine oviduct epithelial cell conditioned medium promotes the early development of bovine in vitro fertilized embryos. J Reprod Dev. 2024, 70, 42–48. [Google Scholar] [CrossRef]
- Kan, F.W.K.; Roux, E.; Bleau, G. Immunolocalization of Oviductin in Endocytic Compartments in the Blastomeres of Developing Embryos in the Golden Hamster1. Biol Reprod. 1993, 48, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Balastegui-Alarcón, M.; Moros-Nicolás, C.; Ballesta, J.; Izquierdo-Rico, M.J.; Chevret, P.; Avilés, M. Molecular Evolution of the Ovgp1 Gene in the Subfamily Murinae. Animals. 2024, 15, 55. [Google Scholar] [CrossRef]
- Moros-Nicolás, C.; Fouchécourt, S.; Goudet, G.; Monget, P. Genes Encoding Mammalian Oviductal Proteins Involved in Fertilization are Subjected to Gene Death and Positive Selection. J Mol Evol. 2018, 86, 655–667. [Google Scholar] [CrossRef]
- Simintiras, C.A.; Fröhlich, T.; Sathyapalan, T.; Arnold, G.J.; Ulbrich, S.E.; Leese, H.J.; et al. Modelling aspects of oviduct fluid formation in vitro. Reproduction. 2017, 153, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Cerezales, S.; Ramos-Ibeas, P.; Acuña, O.S.; Avilés, M.; Coy, P.; Rizos, D.; et al. The oviduct: from sperm selection to the epigenetic landscape of the embryo†. Biol Reprod. 2018, 98, 262–276. [Google Scholar] [CrossRef]
- Almiñana, C.; Corbin, E.; Tsikis, G.; Alcântara-Neto, A.S.; Labas, V.; Reynaud, K.; et al. Oviduct extracellular vesicles protein content and their role during oviduct–embryo cross-talk. Reproduction. 2017, 154, 253–268. [Google Scholar] [CrossRef]
- Alcântara-Neto, A.S.; Schmaltz, L.; Caldas, E.; Blache, M.C.; Mermillod, P.; Almiñana, C. Porcine oviductal extracellular vesicles interact with gametes and regulate sperm motility and survival. Theriogenology 2020, 155, 240–255. [Google Scholar] [CrossRef]
- Fiorentino, G.; Merico, V.; Zanoni, M.; Comincini, S.; Sproviero, D.; Garofalo, M.; et al. Extracellular vesicles secreted by cumulus cells contain microRNAs that are potential regulatory factors of mouse oocyte developmental competence. Mol Hum Reprod. 2024, 30, gaae019. [Google Scholar] [CrossRef]
- Yuan, S.; Wang, Z.; Peng, H.; Ward, S.M.; Hennig, G.W.; Zheng, H.; et al. Oviductal motile cilia are essential for oocyte pickup but dispensable for sperm and embryo transport. Proc Natl Acad Sci. 2021, 118, e2102940118. [Google Scholar] [CrossRef]
- Murayama, Y.; Yoshida, K.; Takahashi, H.; Mizuno, J.; Akaishi, K.; Inui, H. Softening of the mouse zona pellucida during oocyte maturation. 2013 35th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC) [Internet], Osaka; IEEE, 2013; pp. 6834–6837, [cited 2025 Sept 10]. Available from: http://ieeexplore.ieee.org/document/6611127/.
- Kolbe, T.; Holtz, W. Differences in proteinase digestibility of the zona pellucida of in vivo and in vitro derived porcine oocytes and embryos. Theriogenology. 2005, 63, 1695–1705. [Google Scholar] [CrossRef] [PubMed]
- Lamas-Toranzo, I.; Fonseca Balvís, N.; Querejeta-Fernández, A.; Izquierdo-Rico, M.J.; González-Brusi, L.; Lorenzo, P.L.; et al. ZP4 confers structural properties to the zona pellucida essential for embryo development. eLife. 2019, 8, e48904. [Google Scholar] [CrossRef] [PubMed]
- Scully, D.M.; Xia, T.; Musina, G.R.; McCown, M.A.; Umezu, K.; Kircher, B.K.; et al. Region-specific roles of oviductal motile cilia in oocyte/embryo transport and fertility. Biol Reprod. 2025, 112, 651–662. [Google Scholar] [CrossRef]
- Gupta, S.K.; Srinivasan, V.A.; Suman, P.; Rajan, S.; Nagendrakumar, S.B.; Gupta, N.; et al. Contraceptive Vaccines Based on the Zona Pellucida Glycoproteins for Dogs and Other Wildlife Population Management. Am J Reprod Immunol. 2011, 66, 51–62. [Google Scholar] [CrossRef]
- Burkart, A.D.; Xiong, B.; Baibakov, B.; Jiménez-Movilla, M.; Dean, J. Ovastacin, a cortical granule protease, cleaves ZP2 in the zona pellucida to prevent polyspermy. J Cell Biol. 2012, 197, 37–44. [Google Scholar] [CrossRef]
- Maddirevula, S.; Coskun, S.; Al-Qahtani, M.; Aboyousef, O.; Alhassan, S.; Aldeery, M.; et al. ASTL is mutated in female infertility. Hum Genet. 2022, 141, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Von Wiegen, N.; Behl, C.; Körschgen, H. Crossing the barrier or how regulation of ovastacin controls fertilization and translates into clinical phenotypes. iScience. 2025, 28, 112976. [Google Scholar] [CrossRef] [PubMed]
- Yanagimachi, R. Mysteries and unsolved problems of mammalian fertilization and related topics. Biol Reprod. 2022, 106, 644–675. [Google Scholar] [CrossRef]
- Kuske, M.; Floehr, J.; Yiallouros, I.; Michna, T.; Jahnen-Dechent, W.; Tenzer, S.; et al. Limited proteolysis by acrosin affects sperm-binding and mechanical resilience of the mouse zona pellucida. Mol Hum Reprod. 2021, 27, gaab022. [Google Scholar] [CrossRef]
- Le, M.T.; Nguyen, T.V.; Dang, H.N.T.; Nguyen, Q.H.V. Formation of Blastocysts From Zona Pellucida–Free Oocytes: A Case Report on a Modified Technique in In Vitro Fertilization. Singh M, editor. Case Rep Obstet Gynecol. 2025, 2025, 5247242. [Google Scholar] [CrossRef]
- Clark, G.F. The role of carbohydrate recognition during human sperm-egg binding. Hum Reprod. 2013, 28, 566–577. [Google Scholar] [CrossRef]
- Ma, M.; Zhang, L.; Liu, Z.; Teng, Y.; Li, M.; Peng, X.; et al. Effect of blastocyst development on hatching and embryo implantation. Theriogenology. 2024, 21, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Satouh, Y.; Nishimasu, H.; Kurabayashi, A.; Morita, J.; Fujihara, Y.; et al. Structural and functional insights into IZUMO1 recognition by JUNO in mammalian fertilization. Nat Commun. 2016, 7, 12198. [Google Scholar] [CrossRef]
- Swann, C.A.; Cooper, S.J.B.; Breed, W.G. Molecular evolution of the carboxy terminal region of the zona pellucida 3 glycoprotein in murine rodents. Reproduction. 2007, 133, 697–708. [Google Scholar] [CrossRef]
- Pang, P.C.; Chiu, P.C.N.; Lee, C.L.; Chang, L.Y.; Panico, M.; Morris, H.R.; et al. Human Sperm Binding Is Mediated by the Sialyl-Lewisx Oligosaccharide on the Zona Pellucida. Science. 2011, 333, 1761–1764. [Google Scholar] [CrossRef]
- Algarra, B.; Han, L.; Soriano-Úbeda, C.; Avilés, M.; Coy, P.; Jovine, L.; et al. The C-terminal region of OVGP1 remodels the zona pellucida and modifies fertility parameters. Sci Rep. 2016, 6, 32556. [Google Scholar] [CrossRef] [PubMed]
- Priel, E.; Priel, T.; Szaingurten-Solodkin, I.; Wainstock, T.; Perets, Y.; Zeadna, A.; et al. Zona pellucida shear modulus, a possible novel non-invasive method to assist in embryo selection during in-vitro fertilization treatment. Sci Rep. 2020, 10, 14066. [Google Scholar] [CrossRef]
- Lu, X.; Liu, Y.; Cao, X.; Liu, S.Y.; Dong, X. Laser-assisted hatching and clinical outcomes in frozen-thawed cleavage-embryo transfers of patients with previous repeated failure. Lasers Med Sci. 2019, 34, 1137–1145. [Google Scholar] [CrossRef]
- Priel, E.; Mittelman, B.; Efraim, L.; Priel, T.; Szaingurten-Solodkin, I.; Har-Vardi, I. Hyperelastic models for the human zona pellucida and their implications on shear modulus estimation in the clinical practice. Sci Rep. 2024, 14, 31411. [Google Scholar] [CrossRef]
- Vani, V.; Vasan, S.S.; Adiga, S.K.; Varsha, S.R.; Seshagiri, P.B. Molecular regulators of human blastocyst development and hatching: Their significance in implantation and pregnancy outcome. Am J Reprod Immunol. 2023, 89, e13635. [Google Scholar] [PubMed]
- Guo, J.; Lu, W.F.; Liang, S.; Choi, J.W.; Kim, N.H.; Cui, X.S. Peroxisome proliferator-activated receptor δ improves porcine blastocyst hatching via the regulation of fatty acid oxidation. Theriogenology 2017, 90, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.H.; Lee, J.M.; Kim, S.K.; Youm, H.W.; Jee, B.C. Associations of post-warming embryo or blastocyst development with clinical pregnancy in vitrified embryo or blastocyst transfer cycles. Clin Exp Reprod Med. 2020, 47, 140–146. [Google Scholar] [CrossRef]
- Michailov, Y.; Friedler, S.; Saar-Ryss, B. Methods to improve frozen-thawed blastocyst transfer outcomes- the IVF laboratory perspective. J IVF-Worldw [Internet] 2025, 1. [cited 2025 Sept 10]. Available from: https://jivfww.scholasticahq.com/article/87541-methods-to-improve-frozen-thawed-blastocyst-transfer-outcomes-the-ivf-laboratory-perspective. [CrossRef]
- Yang, W.; Wang, Q.; Zhang, B.; Leung, R.K.K.; Deng, K.; Geng, S.; et al. Association between hatching status and pregnancy outcomes in single blastocyst transfers: a retrospective cohort analysis. J Assist Reprod Genet. 2025, 42, 1707–1715. [Google Scholar] [CrossRef]
- Mahdavinezhad, F.; Kazemi, P.; Fathalizadeh, P.; Sarmadi, F.; Hashemi, E.; Hajarian, H.; et al. In vitro versus In vivo: Development-, Apoptosis-, and Implantation-Related Gene Expression in Mouse Blastocyst. Iran J Biotechnol. 2019, 17, 90–97. [Google Scholar] [CrossRef]
- Hao, X.; Zhao, Y.T.; Ding, K.; Xue, F.R.; Wang, X.Y.; Yang, Q.; et al. Assisted Hatching Treatment of Piezo-Mediated Small Hole on Zona Pellucida in Morula Stage Embryos Improves Embryo Implantation and Litter Size in Mice. Front Cell Dev Biol. 2021, 9, 746104. [Google Scholar] [CrossRef]
- Liu, Y.; Jones, C.; Coward, K. The Current Practice of Assisted Hatching for Embryos in Fertility Centres: a General Survey. Reprod Sci. 2022, 29, 2664–2673. [Google Scholar] [CrossRef]
- Syrkasheva, A.G.; Dolgushina, N.V.; Romanov AYu Burmenskaya, O.V.; Makarova, N.P.; Ibragimova, E.O.; et al. Cell and genetic predictors of human blastocyst hatching success in assisted reproduction. Zygote. 2017, 25, 631–636. [Google Scholar] [CrossRef]
- Batista, M.R.; Diniz, P.; Torres, A.; Murta, D.; Lopes-da-Costa, L.; Silva, E. Notch signaling in mouse blastocyst development and hatching. BMC Dev Biol. 2020, 20, 9. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Chen, T.; Liu, Y.; Hou, Z.; Wu, K.; Cao, Y.; et al. The critical role of ZP genes in female infertility characterized by empty follicle syndrome and oocyte degeneration. Fertil Steril. 2021, 115, 1259–1269. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, Y.; Yuan, G.; Yang, J.; Liu, X.; Chen, S.; et al. A compound heterozygous mutation in ZP1 and two novel heterozygous cis mutations in ZP3 causes infertility in women presenting with empty follicle syndrome. J Ovarian Res. 2025, 18, 86. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, F.; Kodani, M.; Emori, C.; Kiyozumi, D.; Mori, M.; Fujihara, Y.; et al. CRISPR/Cas9-Mediated Genome Editing Reveals Oosp Family Genes are Dispensable for Female Fertility in Mice. Cells 2020, 9, 821. [Google Scholar] [CrossRef]
- Dioguardi, E.; Stsiapanava, A.; Fahrenkamp, E.; Han, L.; De Sanctis, D.; Inzunza, J.; et al. Structural basis of ZP2-targeted female nonhormonal contraception. Proc Natl Acad Sci. 2025, 122, e2426057122. [Google Scholar] [CrossRef]
- Avella, M.A.; Baibakov, B.A.; Jimenez-Movilla, M.; Sadusky, A.B.; Dean, J. ZP2 peptide beads select human sperm in vitro, decoy mouse sperm in vivo, and provide reversible contraception. Sci Transl Med [Internet] 2016, 8. [cited 2025 Sept 10]. Available from: https://www.science.org/doi/10.1126/scitranslmed.aad9946.
- Zhang, B.; Qu, G.; Nan, Y.; Zhou, E.M. Ovarian Oxidative Stress Induced Follicle Depletion After Zona Pellucida 3 Vaccination Is Associated With Subfertility in BALB/c Mice. Front Vet Sci. 2022, 9, 814827. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the regions (A, B, C, D, and E) of the oviductal glycoprotein 1 (OVGP1) in different mammalian species. Figure modified from [13].
Figure 1.
Schematic representation of the regions (A, B, C, D, and E) of the oviductal glycoprotein 1 (OVGP1) in different mammalian species. Figure modified from [13].
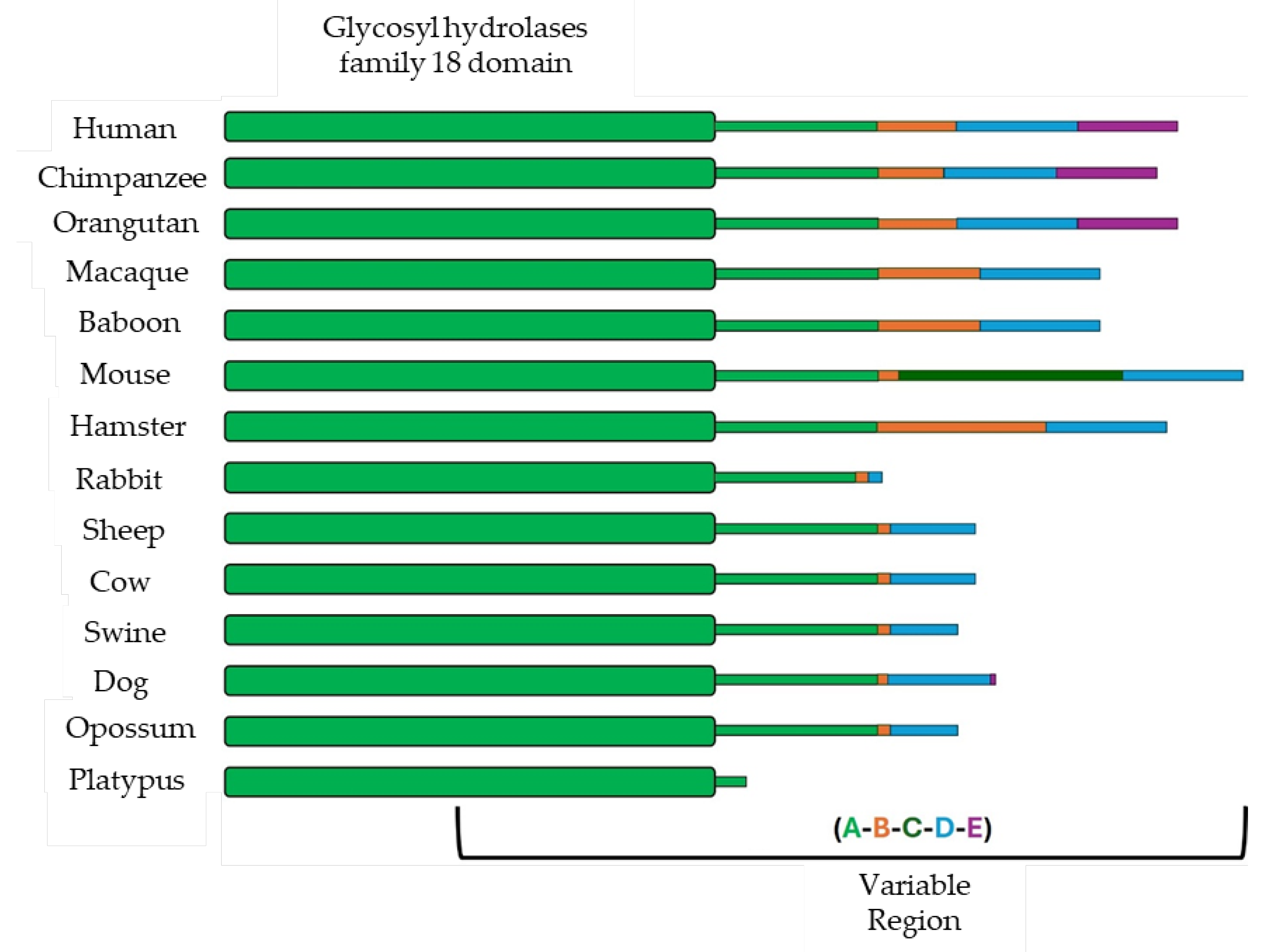
Figure 2.
Molecular architecture of zona pellucida (ZP) proteins, ZP composition in different eutherian mammals and protofilaments and filaments organization. Schematic representation of the four ZP glycoproteins (ZP1, ZP2, ZP3, and ZP4) highlighting the ZP module (red box) and showing the differences in their N-terminal region (a). ZP composition in representative mammalian groups: Mus musculus, with ZP4 pseudogenized; most eutherian mammals (humans, the genus Mus, rat, rabbit and hamster), expressing all 4 ZP proteins, and dog, cow, dolphin, fox, and sow, lacking ZP1 (b). Assembly of type I protofilaments (composed of ZP3 only) and type II protofilaments (formed by variable combinations of ZP1, ZP2, and/or ZP4) into filaments that constitute the ZP matrix (c). Figure modified from [20].
Figure 2.
Molecular architecture of zona pellucida (ZP) proteins, ZP composition in different eutherian mammals and protofilaments and filaments organization. Schematic representation of the four ZP glycoproteins (ZP1, ZP2, ZP3, and ZP4) highlighting the ZP module (red box) and showing the differences in their N-terminal region (a). ZP composition in representative mammalian groups: Mus musculus, with ZP4 pseudogenized; most eutherian mammals (humans, the genus Mus, rat, rabbit and hamster), expressing all 4 ZP proteins, and dog, cow, dolphin, fox, and sow, lacking ZP1 (b). Assembly of type I protofilaments (composed of ZP3 only) and type II protofilaments (formed by variable combinations of ZP1, ZP2, and/or ZP4) into filaments that constitute the ZP matrix (c). Figure modified from [20].
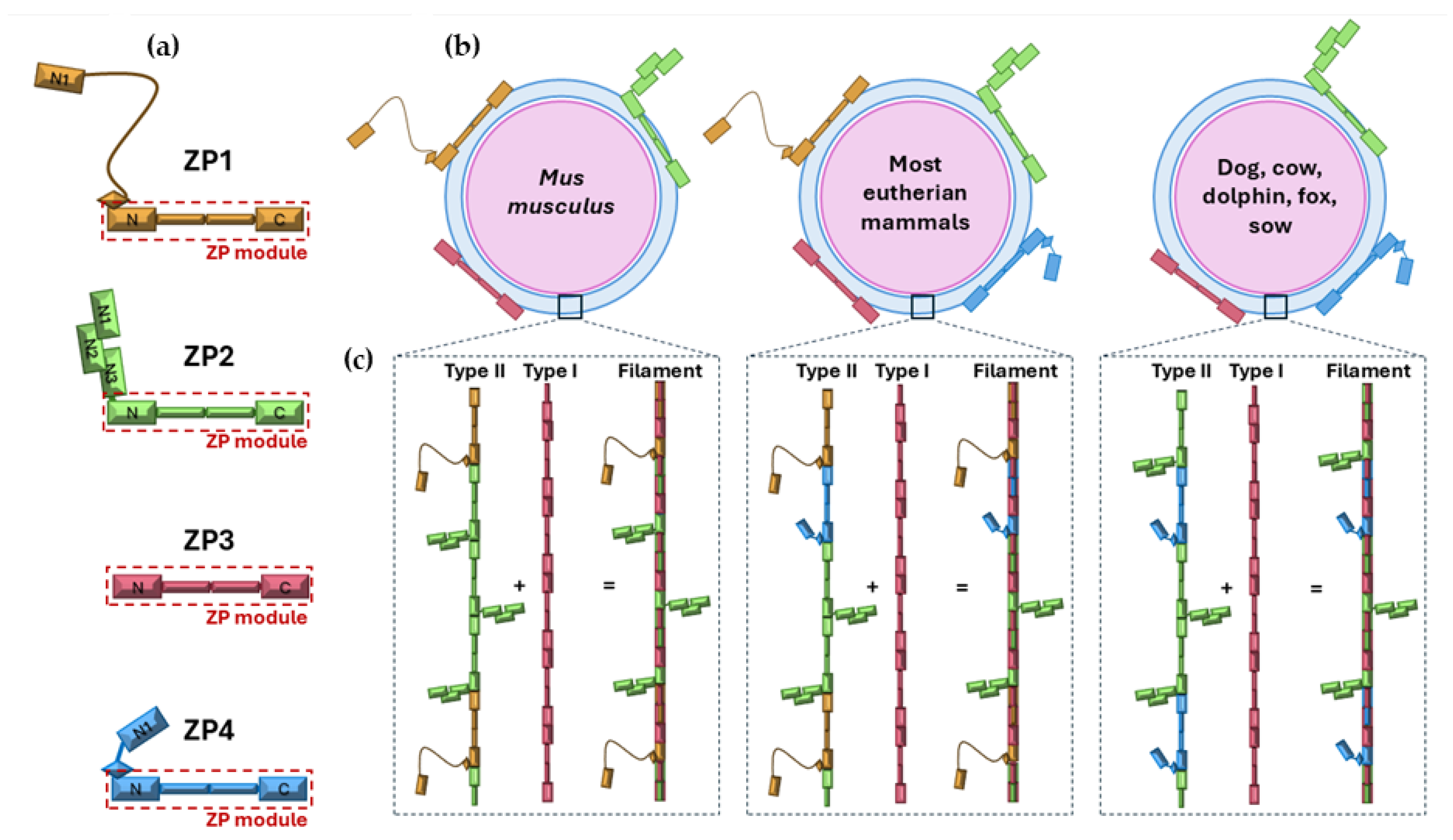
Figure 3.
(Adapted from [4]).Bovine ZP changes during oocyte maturation. Bovine in vitro matured (IVM) oocytes. High magnification reveals the ultrastructural characteristics of the ZP’s pores on the oocytes without bOVGP1 (a,b) and with bOVGP1 (c,d). Scale bars = 10 μm.
Figure 3.
(Adapted from [4]).Bovine ZP changes during oocyte maturation. Bovine in vitro matured (IVM) oocytes. High magnification reveals the ultrastructural characteristics of the ZP’s pores on the oocytes without bOVGP1 (a,b) and with bOVGP1 (c,d). Scale bars = 10 μm.

Table 1.
Proteins present in the zona pellucida (ZP) of different mammalian groups. In eutherian mammals, ZP3-1c is usually referred to as ZP3.
Table 1.
Proteins present in the zona pellucida (ZP) of different mammalian groups. In eutherian mammals, ZP3-1c is usually referred to as ZP3.
| Group of mammals | ZP1 | ZP2 | ZP3-1a | ZP3-1b | ZP3-1c | ZP3-2 | ZP4 | ZPAX | ZPY |
|---|---|---|---|---|---|---|---|---|---|
| Mus musculus | ✓ | ✓ | ✓ | ||||||
| Most eutherian mammals | ✓ | ✓ | ✓ | ✓ | |||||
| Dog, cow, sow, dolphin, fox | ✓ | ✓ | ✓ | ||||||
| American marsupials | ✓ | ✓ | ✓ | ✓ | |||||
| Australian marsupials | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||
| Monotremes | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ |
Table 2.
(Adapted from [46]). Average and variance values of ZP thickness and retardance measured in individual human oocytes and embryos. Values are presented as mean ± SD (one-tailed t-test; P < 0.05). a Statistically significant difference between immature and mature oocytes. b Statistically significant decrease between mature oocytes and embryos. c Statistically significant increase between mature oocytes and embryos.
Table 2.
(Adapted from [46]). Average and variance values of ZP thickness and retardance measured in individual human oocytes and embryos. Values are presented as mean ± SD (one-tailed t-test; P < 0.05). a Statistically significant difference between immature and mature oocytes. b Statistically significant decrease between mature oocytes and embryos. c Statistically significant increase between mature oocytes and embryos.
| Immature Oocytes | Mature Oocytes | Day 3 Embryos | |
|---|---|---|---|
| Layer 1 | |||
| Thickness (µm) | 10.5 ± 2.3 | 9.8 ± 2.1ᵇ | 7.9 ± 1.9 |
| Retardance (nm) | 3.26 ± 1.27 | 2.84 ± 1.07 | 3.00 ± 0.86 |
| Layer 2 | |||
| Thickness (µm) | 3.4 ± 0.6 | 3.7 ± 0.9 | 3.6 ± 1.2 |
| Retardance (nm) | 0.21 ± 0.07ᵃ | 0.24 ± 0.05ᶜ | 0.33 ± 0.29 |
| Layer 3 | |||
| Thickness (µm) | 6.5 ± 2.1 | 6.1 ± 1.7ᵇ | 3.7 ± 1.4 |
| Retardance (nm) | 0.91 ± 0.24 | 0.95 ± 0.23ᵇ | 0.78 ± 0.27 |
| Total Zona | |||
| Thickness (µm) | 20.4 ± 2.4 | 19.5 ± 2.2ᵇ | 15.2 ± 2.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



