
Submitted:
02 October 2025
Posted:
03 October 2025
You are already at the latest version
Abstract
Plant viruses are useful tools for quickly and easily producing recombinant proteins in plants. Compared to systems that use genetically modified plants, viral vectors are easier to work with and can produce recombinant proteins faster and in larger amounts. Recently, there has been growing interest in using plant viruses as vectors to make vaccines, either as whole proteins or as small parts displayed on the surface of modified viral particles. Several viruses, like tobacco mosaic virus, potato virus X, and cowpea mosaic virus, have been well developed for this purpose. Vaccines made using these viruses target various human and animal diseases and have often triggered immune responses and provided protection against infections. This review looks at the benefits of using plant virus vectors, the progress in developing different viral vector systems, and immune studies that support the idea of vaccines made from plant viruses.
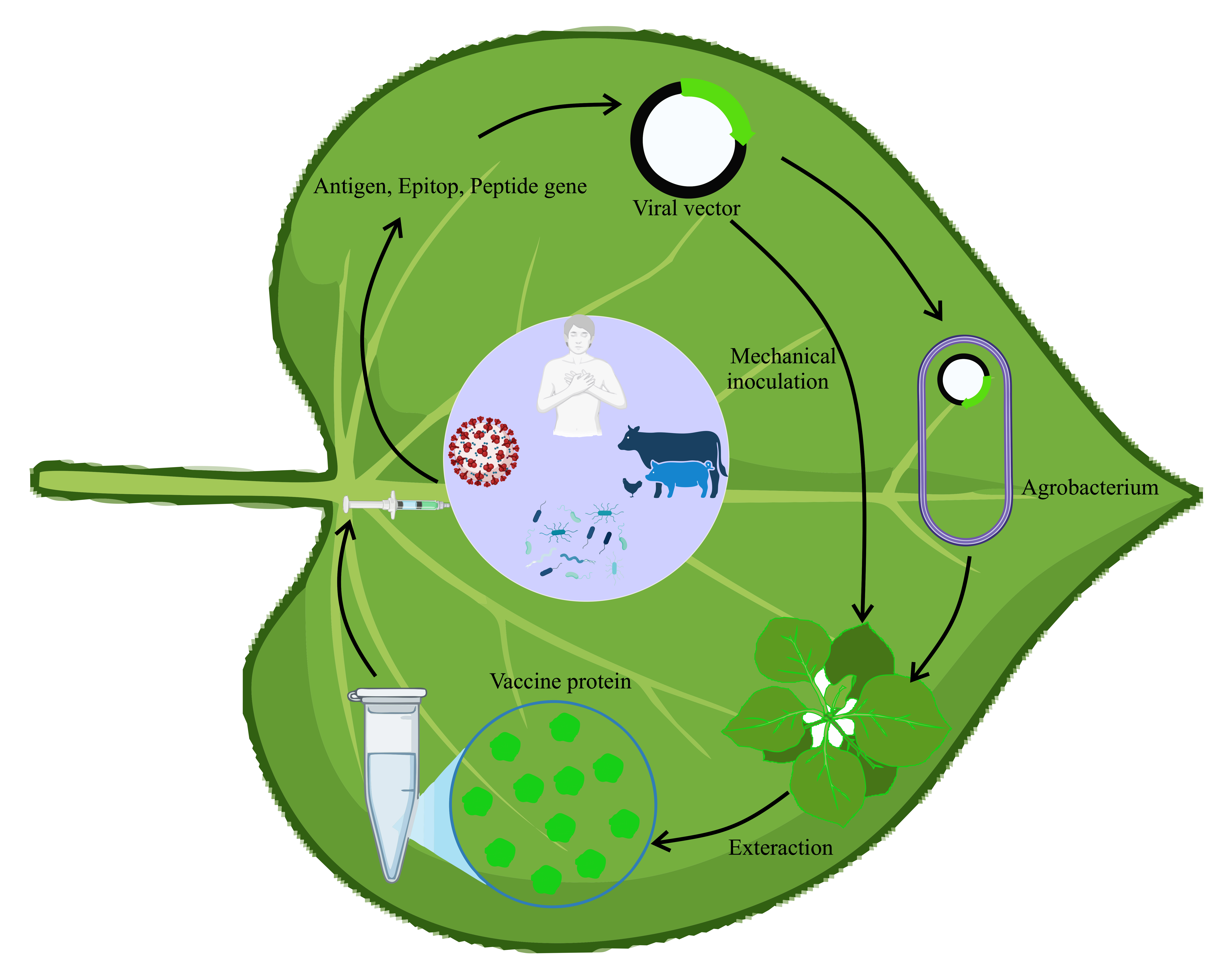
Keywords:
plant virus
; genetically modified plant
; potato virus X
; tobacco mosaic virus
; Alfalfa mosaic virus
1. Introduction
Vaccinology started in 1789 with Edward Jenner’s smallpox vaccine with pivotal disease prevention [1,2]. In the 19th century, Louis Pasteur contributed to the field by creating vaccines through pathogen attenuation [3]. Cell culture techniques developed by the mid-20th century led to the creation of inactivated and attenuated vaccines, which played a crucial role in eradicating outbreak diseases and saving millions of lives [2]. Before the late 20th century, conventional vaccines primarily consisted of inactivated or weakened pathogens, toxoids, proteins, or bacterial polysaccharides, prior to the emergence of molecular biology and significantly reduced the prevalence and severity of diseases[4]. Despite ongoing research for decades, effective vaccines for specific human pathogens are still lacking due to challenges such as high genetic variability, persistent or latent infections, and the difficulty in eliciting sterilizing immunity [4,5]. Furthermore, traditional vaccines face major limitations, such as safety risks from the reversion of live attenuated viruses, contamination with live organisms, and potential adverse effects in immunocompromised individuals[4]. Their production is also expensive, time-consuming, and requires high biosafety standards along with specialized laboratory facilities [4,6].
Recent advancements in genetic engineering, immunology, structural biology, and nanotechnology have led to the emergence of innovative vaccine platforms. These innovations encompass a range of cutting-edge technologies in the field of vaccines, including recombinant viral vectors, virus-like particles (VLPs), mRNA vaccines, synthetic DNA vaccines, and bacterial vector vaccines [4]. Such technologies provide significant advantages over traditional vaccines, facilitating rapid and efficient responses to new pathogens and pandemics. Recombinant plant viral vectors are highlighted as a promising platform among various technologies. The use of plant viruses in vaccine development can be rile on genome capability for encode foreign antigens, intrinsic adjuvant properties for eliciting innate immune responses, and safety. Furthermore, they can also be engineered for targeted antigen delivery to specific cells or tissues. Plant virus expression vectors enable rapid, large-scale gene expression without the need for extensive plant transformation, often achieving substantial levels within days. This production level can be scaled up by increasing the number of host plants. Moreover, the purification of vaccine proteins can be achieved using a few simple steps, or in some cases via only partial purification [7]. These properties make plant virus expression vectors versatile for use to address infectious diseases in the Third World today.
2. Plants as Biofactory in Vaccine Development
Vaccines can be produced in various host systems, including bacteria, yeast, insect cells, and mammalian cells [8]. Advancements in molecular biology in the 1980s facilitated the creation of subunit vaccines through the utilization of proteins sourced from pathogens. While mammalian cell systems are commonly used for vaccine production, vaccine production poses safety risks due to potential toxins and infectious agents, as microbial contamination can remain undetected post-purification, complicating quality control during large-scale manufacturing [9,10]. E. coli is deficient in key post-translational modifications, and while yeast and insect cells present some alternatives, their immunological disparities restrict their effectiveness.
The late 1980s, Plants have been suggested as a promising platform for recombinant protein production [11]. Today, producing biopharmaceuticals (drugs and vaccines) in plants using molecular biology techniques know with term of ‘molecular farming [12]. Plant-based platforms for vaccine production provide various benefits compared to conventional expression systems. “First, Plant-made vaccines are significantly cost-effective, with production costs potentially up to 1,000 times lower than mammalian cell systems and up to 50 times lower than E. coli bioreactors, based on protein yields[13]. Plants utilize sunlight, carbon dioxide, water, and nutrients in photosynthesis to produce materials and energy. This not only lowers production costs but also improves scalability, with a larger cultivation area directly increasing output [14]. Second, Plants are a safer option for human and veterinary applications than mammalian cell systems because they do not harbor animal pathogens like microbes or prions [15]. Third, Plant systems can perform eukaryotic post-translational modifications, like glycosylation and disulfide bond formation, which are essential for maintaining the structural integrity and functionality of vaccine proteins [14]. “Fourth, Plant-based platforms provide remarkable scalability, allowing quick production capacity expansion and significantly reducing manufacturing timelines, which is vital for addressing infectious disease outbreaks and public health emergencies [16]. Lastly, plant-made vaccines can be produced in various edible parts of the plant including fruits, leaves, seeds, tubers, and vegetables—and are referred to as ‘edible vaccines [12,14]. Thus, Plants have become a compelling medium for producing vaccines. The common strategies that available for vaccine production in plants are stable transformation of the plant nuclear genome, stable transformation of the chloroplast genome and viral transient expression.
3. Plant Viral Transient expression in Vaccine Development
Plant viruses have facilitated the introduction of foreign genes into plants since the early 1980s, with advances in molecular biology and plant virology enabling the development of enhanced expression systems. These Recent systems offer advantages such as rapid, high-level transgene expression and improved transgene containment, particularly in movement defective systems, by eliminating vertical and horizontal gene transfer [17]. Plant viruses exhibit structural flexibility and genetic diversity, enabling the design of viruses tailored for biotechnological applications. These viruses can enhance immune responses more effectively than conventional vaccines, highlighting their potential in vaccine development [18]. The ability of plant viruses to present multiple antigens leads to strong immune responses, proving valuable for next-generation vaccines with improved immunogenicity and efficacy [19,20]. pVLPs are not only significant in vaccinology but also serve as effective vehicles for drug delivery, capable of encapsulating therapeutic molecules and delivering them directly to target cells, which reduces off-target effects and improves therapeutic results [21,22]. Another method employs plant viruses as adjuvants in vaccine development, enhancing safety by not replicating in animal cells. In contrast, plant viral-based vaccines typically do not require additional adjuvants, simplifying their composition and formulation compared to other vaccines.
Plant viruses are effective in producing recombinant proteins, facilitating large-scale synthesis of heterologous proteins, especially costly ones such as vaccines and antibodies. Plant viruses act as cellular parasites, producing many viral proteins and genomes, while inhibiting Post-transcriptional gene silencing (PTGS) and protein degradation, thereby facilitating systemic infection in plants. Plant viral vectors offer benefits as an alternative to conventional transgenic systems, including faster expression, higher yield production, cost and time savings, and adaptability for high throughput and scalability.
4. Plant Viruses Vectors -Based Strategies in Vaccine Development
Technical advances in molecular biology, including cDNA generation from RNA templates, have broadened the search for expression systems in single-stranded RNA viruses, which are prevalent among plant viruses[23]. Additionally, the application of Agrobacterium tumefaciens has facilitated viral infection [14,24]. Generally, viral infection can start through mechanical inoculation of viral particles on leaves or by transfecting nucleic acids. The technique known as “agroinfection” employs Agrobacterium tumefaciens to directly and efficiently deliver cDNA to the plant cell nucleus. The cDNA constructs in the plant expression cassette produce an infectious, independently replicating nucleic acid following nuclear transcription and processing in host cells[14]. Agroinfection enables the utilization of viruses that, while not mechanically transmissible in nature, require a specific insect vector to commence the infection process. Viral vectors are categorized into gene substitution vectors, gene insertion vectors, modular vector systems, and peptide display vectors.

Table 1.
Examples of plant viruses expression vectors used in the development of vaccine.
| Plant viral vector | Pathogen or protein/epitope | References |
|---|---|---|
| Tobacco mosaic virus | SARS-CoV-2 spike protein, bovine herpes virus-gD protein, major birch pollen antigen plasmodium antigen | [25,26,27] |
| Physalis mottle virus (PhMV) | Influenza A virus (M2e) | [28] |
| Potato virus X | scFv | [29] |
| Zucchini yellow mosaic virus | Anti-HIV proteins | [30] |
| Cowpea mosaic virus | SARS-CoV-2 spike protein, Human papillomavirus 16 L1 protein bearing the M2e influenza epitope | [31,32] |
| Alfalfa mosaic virus | HIV | [33] |
| Pod pepper vein yellows virus (PoPeVYV), | hepatitis B surface antigen (HBsAg) | [34] |
4.1. Gene Insertion Vectors
Gene insertion vectors are modified viruses that include an extra open reading frame (ORF) designed for the target protein [17]. The target protein can be expressed through a fusion with the viral coat protein (CP) or independently without CP [14,17]. TMV and potato virus are notable vectors, both featuring single-stranded positive-strand RNA genomes [17]. Insertion of the CAT gene between the motor protein and CP genes enables TMV to replicate, form subgenomic RNAs, assemble accurately, and express the intended gene activity [35,36]. A viral expression vector derived from Potato virus X (PVX) was created, which incorporates a bacterial β-glucuronidase gene. The vector, pGC3, successfully infected Nicotiana clevelandii and N. tabacum, demonstrating GUS expression in the leaves [37]. A common vector is one derived from Cowpea mosaic virus (CPMV). A CPMV/S-2A-GFP construct, fused with jellyfish green fluorescent protein, was inoculated into cowpea, resulting in a 1% expression of total soluble protein, with systemic infection observed 10 days after inoculation [38]. However, Use of the amplified subgenomic promoter resulted in homologous recombination, causing vector instability and deletion of the foreign gene [36]. The first generation of viral vectors has revealed issues, notably the risk of complete viruses disseminating systemically in host plants, which raises biosafety and biosecurity concerns. Large exogenous inserts can impair virus assembly and the infection area, whereas a cleavage site may lead to a combination of free and fused proteins. Promoter duplication can result in homologous recombination of repeating sequences, leading to instability in constructs, despite significant attempts to address these challenges [14].
4.2. Gene Replacement Vectors
Gene substitution or replacement vectors allow for the replacement of an endogenous viral sequence with a gene of interest from a different source [17]. A gene replacement vector is created by substituting an endogenous viral sequence with a gene of interest (GOI). The viral genome does not include all elements required for protein expression, such as CP. CP is essential for the systemic movement of the majority of viruses. Advancements in technology, such as vacuum infiltration, facilitate the extensive distribution of viral vectors across entire plants. The removal of CP allows for the effective elimination of bio-contamination threats, as it is no longer necessary for systemic movement. Additionally, viral vectors can accommodate larger inserts when the viral CP is substituted with a gene of interest, offering advantages over traditional gene insertion vectors. Musiychuk et al. created pBID4, a launch vector designed to enhance the expression, stability, and immunogenicity of modified lichenase and its fusion products with proteins like plague, anthrax, and influenza [39]. For instance, two forms of antigen hemagglutinin (HA) from the influenza type A/Vietnam/04 (H5N1) virus were expressed: the globular domain fused with LicKM (LicKM-H5GD) and the stem domain fused with LicKM (LicKM-H5SD). Both proteins were extracted from Nicotiana benthamiana at 4-7 days post-infiltration and purified to over 80% purity using affinity chromatography. These plant-made vaccines demonstrated specific binding to antibodies in sheep sera raised against the HA of the influenza type A/Vietnam/04 virus [17,39]. Moreover, using a vector that contains the entire viral genome and gene of interest (GOI) can lead to a large plasmid size, which may diminish the efficiency and flexibility of genetic engineering or cellular expression systems.
4.3. Modular or Deconstructed Vectors
Modular or deconstructed vectors represent an advanced category of viral expression systems, developed from the insight that not all viral components are necessary or advantageous for expression vectors. This concept allows for the disassembly of viruses into distinct genomic elements that can function together during the infection process akin to wild-type multipartite genome viruses. Moreover, agroinfection provided the technical possibility to co-deliver multiple different components [17]. In the modular system, the viral genome is segmented into provectors or minimal vectors that include only essential viral elements to enhance protein expression, such as RNA-dependent RNA polymerase (RdRp) [40,41]. To create an integrated system, provectors must be introduced into a single cell through agroinfiltration and co-expressed to operate as a complete expression system. In modular systems, viral components are divided into separate sections and introduced into binary vectors within an Agrobacterium strain, which are then mixed and co-infiltrated into plant leaves. This strategy enables the reduction of the replicon size to accommodate transgene insertion, while other essential viral components for vector function are supplied during the infection process. One of the most widely used vectors currently is the deconstructed system based on TMV, developed by Icon Genetics in Halle, Germany, which was recently acquired by Bayer Innovation GmbH.
4.4. Peptide Display Vectors
In this strategy, the target gene is fused with viral CP gene to display on the surface of virus particles (CVPs) [17]. The foreign peptides can directly be fused at the N- or C terminus of CP or indirectly joined with CP via an extra sequence, e.g., a readthrough translational context [42] or an F2A sequence [43]. The gene fusion should not disrupted replication, RNA and protein production, self-assembly viral particles. Although these requirements sometimes n crestrict the size of inserts. Peptide display vectors have been actively used since 1986, and popular tool in plant-made vaccine production [44]. Subunit vaccines use small peptide or protein epitopes, but their limited size hinders recognition by antigen-presenting cells, resulting in weak or incomplete immune responses unless presented on carriers or with adjuvants [45]. Plant viruses by enlarging epitope size can enhance identification and immune responses [46,47]. Additionally, plant viruses are confined to their host-plant range and lack the ability to replicate or cause disease in animals, ensuring biosafety for vaccine and therapeutic applications. Lico et al. incorporated the nucleoprotein (NP) sequence of the influenza A virus to the N-terminus sequence of PVX, in order to generate NP-CVPs [48]. Foreign peptides fused with PVX CP were displayed on NP-CVPs, yielding ~1.1 mg/g in N. benthamiana. Immunized mice produced antigen-specific CD8+ T cells without adjuvants, indicating CVPs can activate cell-mediated immunity and serve as effective vaccine platforms.
5. Type of Plant Viral Vectors
5.1. Comovirus (Cowpea Mosaic Virus (CPMV))
CPMV-based expression vectors used as a peptide display system in the 1990s [49]. In this system, short antigenic peptides known as epitopes are expressed on the surface of CPMV. Cowpea CPMV features a highly ordered, three-dimensional icosahedral symmetry, allowing repetitive display of multiple epitope copies on its surface, which enhances immunogenicity and establishes it as an effective vaccine carrier platform [14,50]. CPMV achieves high yields in plants, producing 0.5 to 2 mg purified virus per gram of fresh leaf tissue. Its high titer, combined with nonpathogenicity in mammalian cells, makes CPMV an ideal epitope presentation platform for vaccines, balancing both biosafety and yield effectively [14,51]. There are three main sites for epitope insertion in CPMV coat proteins: the βB-βC loop of the small coat protein (predominant site), the βC’-βC’’ loop of the small coat protein, and the βE-αB loop of the large coat protein [14,52]. A variety of approaches, including gene substitution, gene insertion, deconstructed vectors and transgene combined virus vectors, have been used to modify CPMV as a gene expression system[14,53].
5.2. Tobamovirus (Tobacco Mosaic Virus (TMV))
Tobamovirus genus be long to family Virgaviridae, is rigid, monopartite, rod-shaped plant viruses with a positive-stranded RNA genome. N-terminus, C-terminus and a 60s loop are exposed on the viral surface and suitable to bioconjugations [54]. Furthermore, TMV’s stability, modifiability, and self-assembly make it ideal for peptide display [55TMV was the first plant virus engineered for molecular farming and was used to display a type 3 poliovirus epitope, showcasing its utility as a peptide display platform {Haynes, 1986 #56]. Turpen et al. used TMV to display Plasmodium epitopes, producing malarial subunit vaccines in tobacco plants [56]. The immunogenic epitopes such as G5-24 from the rabies virus (RV) glycoprotein, and 5B19 from murine hepatitis virus (MHV) S-glycoprotein, were successfully displayed on the surface of TMV [55]. Takamatsu et al. replaced TMV’s coat protein gene with a bacterial chloramphenicol acetyltransferase gene (CAT) gene [57].
The genome of TMV has been ‘deconstructed’ into two separate modules, as depicted in Figure 1. One module contains the portions of the genome required for replication, while the other contains cassettes designed for insertion of foreign genes. In this way, biopharmaceutical proteins can be expressed at high levels in a viral replicon system in plants
5.3. Potexvirus (Potato Virus X (PVX))
Potexvirus Vectors
Several potexviruses have been used to engineer expression vectors inclusive of Alternanthera mosaic virus (AltMV) (Beernink and Whitham, 2023; Lim et al., 2010), Foxtail mosaic virus (FoMV) (Bouton et al., 2018; Liu et al., 2016; Mei et al., 2016, 2019), cassava common mosaic virus (CsCMV) (Tuo et al., 2021), Cymbidium mo saic virus (CymMV) (Hsieh et al., 2013), pepino mosaic virus (PepMV) (Abrahamian et al., 2021; Sempere et al., 2011), narcissus mosaic virus (NMV) (Zhang et al., 2013), Zygocactus X virus (ZVX) (Koenig et al., 2006), Plantago asiatica mosaic virus (PlAMV) (Minato et al., 2014), potato virus X (PVX) [Salazar-González et al., 2015] and bamboo mosaic virus (BaMV) (Chen et al., 2017; Lin et al., 1996; Liou et al., 2014). Of these, PVX has been successfully used for generation of vaccine candidates against cancer and other diseases. The following section describes recently published findings on the potential applications of PVX-derived vectors as vaccine candidates.
Potato Virus X
Potato virus X (PVX) is is the type member of the genus Potexvirus and contains 1300 CP subunits encapsidating a single (+)-sense RNA having a cap structure at its 5 ‘end and a poly-A tail at its 3’ end. The genome harbors five ORFs [Salazar-González et al., 2015] that code for the RNA-dependent RNA polymerase essential for replication, three movement proteins called the triple gene block (TGB) as well as the CP gene necessary for PVX assembly, cell-to-cell infection through the plasmodesmata and for systemic movement.
PVX functions as an inimitable nano-scaffold for development of vaccines by genetic engineering of the virus to display on its surface immunogenic peptides that can elicit strong immune response. Scientists have genetically modified PVX to chemically conjugate or fuse foreign peptides to the PVX CP subunits to generate chimeric virus particles (CVPs). Such modified forms of the PVX particles can enable multivalent presentation of antigens, augmenting their capability to target B cells and induce robust immune reactions in animal models. The filamentous, flexible structure of PVX makes it an effectual nanocarrier for antigen presentation to the immune system. Plants infected with the engineered constructs of PVX can generate high yields of the CVPs capable of display of the desired antigens or peptides. The surface of the PVX CVPs are densely covered with several copies of the same antigenic peptide, inducing a strong immune response. Biodistribution analyses reveal that these PVX CVPs are well-tolerated, in addition to displaying broad biodistribution and having high bioavailability in animal models. PVX antigen carriers exhibit a tropism for B cells that are essential for antibody responses and immune memory. PVX provides a versatile platform enabling the display of various types of antigens and peptides for several vaccine applications.
The filamentous structure of the PVX viral particles enables them to carry large payloads that are favorable for biomedical imaging as this requires the formation of multi-functional scaffolds having high aspect ratio [Röder et al., 2019]. PVX is capable of superior homing into tumors and has better retention properties when compared to that of spherical nanoparticles. Since PVX is comprised of a protein-based nanoparticle, its inimitable functional characteristics combined with augmented biocompatibility makes it more appropriate for biomedical applications than that of synthetic nanomaterials. Furthermore, PVX nanoparticles have diminished toxicity in vivo in addition to better pharmacokinetic profiles.
PVX vectors enable high-level production of recombinant proteins, rapid transport and systemic infection within the plant. In plant systems, PVX has been employed as a full-length expression vector with capability to infect distal tissues. It has also been used as a deleted vector wherein genes important for systemic and local movement are deleted. Many approaches have been undertaken to generate PVX-derived transient expression vectors. These methodologies include: the usage of a duplicated sub-genomic promoter to execute exogenous expression of genes [Baulcombe et al., 1995], foreign protein expression wherein the foreign gene is directly fused to the truncated CP N-terminus [Marusic et al. 2001; Uhde et al. 2005], the antigen of interest and the CP expressed through a single transcription unit wherein the FMDV 2A autocatalytic peptide is inserted between both the ORFs [Santa Cruz et al., 1996] so that a ribosomal skip mechanism can release two independent peptides derived from a single transcription unit, and expression of a bicistronic mRNA harboring both the foreign gene as well as the CP gene divided by an internal ribosome binding site and driven by the sub-genomic promoter of the CP [Toth et al., 2001].
In a pioneering study, the pGR106 expression vector derived from PVX was used in the expression of the HPV L2 and E7 proteins in N. benthamiana [Čerˇovská et al. 2008]. This resulted in a yield of 2.8-4.3 mg of the proteins per 100 gm fresh leaves. In an effort to generate vaccine prototypes of HIV, chimeric virus particles were produced wherein the HIV type I gp41 ELDKWA epitope was fused to the PVX CP N-terminus. Immunogenicity investigations in mice models showed that intraperitonial or intranasal immunization of mice led to high expression levels of IgG and IgA antibodies specific to HIV even when these virus particles were administered in the absence of adjuvants [Marusic et al., 2001]. Čerˇovská et al. (2012) produced a fusion protein containing an epitope from the L2 minor capsid protein of HPV 16 and the PVX CP. This generated a yield of 170 mg of this protein for every kg of fresh leaf tissue. Test mice that were subcutaneously immunized with the protein showed the elicitation of antibodies against both the L2 epitope and the PVX CP. The Influenza virus M2 protein conserved trans-membrane domain was fused with the Hepatitis B core antigen (HBc) and expression of this fusion protein using a PVX vector resulted in a yield of 2% of the TSP. The purified form of this protein was found to be immunogenic in mice wherein the immune reaction was Th1 polarized [Ravin et al., 2012]. PVX expressing the HPV-16 E7 protein in N. benthamiana when administered along with the Quil A adjuvant was found to be highly immunogenic in mice by eliciting both cell-mediated and antibody responses [Franconi et al., 2002, 2006]. The IgG antibody isotypic profile revealed that both Th1 and Th2 immune responses occurred and 40% of the mice remained free of the HPV tumors following challenge with a tumor cell line expressing the E7 protein. Additionally, the tumors that did emerge in mice vaccinated with the engineered plant-expressed E7 were notably smaller in volume compared to those that emerged in untreated mice.
CVPs of PVX displaying the 2F5 linear epitope (2F5e) of the HIV-1 gp41 envelope protein are able to elicit the expression of epitope-specific IgG and IgA, upon intranasal or intraperitoneal administration in normal mice models without the necessity for co-administered adjuvants. Significantly, when human dendritic cells were pulsed with these PVX CVPs, it activated the 2F5e-specific primary antibody response in acute combined immunodeficient mice that were engrafted with the human lymphocytes [Lico et al., 2015].
PVX CVPs that display the Influenza virus MHC class I-restricted peptide have been engineered wherein these CVPs were found to be genetically stable and elicited epitope-specific CD8+ T cells without the necessity of adjuvant co-delivery [Lico et al., 2009). PVX CVPs are superior epitope carriers endowed with adjuvant properties likely because of its complex particulate structure in addition to the presence of the PVX viral RNA that may stimulate TLR-7 on the antigen presenting cells (Jobsri et al., 2015). Due to these inimitable features, PVX has the inherent capability to elicit whole immune responses. PVX CVPs have been tailored to display the Staphylococcus aureus fibronectin-binding protein D2 peptide (Brennan et al., 1999), two E2 glycoprotein peptides of the classical swine fever virus [Marconi et al., 2006] and the R9 peptide from the HCV hypervariable region I [Uhde-Holzem et al., 2010].
PVX particles that display a peptide derived from the HER2 extracellular domain have been constructed through conjugation to the Lys residue on the PVX CP surface via a two-step reaction by means of a a heterofunctional NHS-(poly)ethyleneglycole (PEG)4-maleimide linker (Shukla et al., 2014a). The antigen HER2 is typically expressed on breast cancer cells wherein it induces B cell activation and antibody production. These chemically derivatized particles of PVX upon testing in mice models were shown to be effectual in eliciting the overcoming of the HER2 immunological tolerance in HER2-positive human cancer cells. PVX nanoparticles labeled with A647 upon bioconjugation with a 12 amino acid peptide having affinity to the EGF receptor (EFGR) are capable of detecting and imaging carcinoma cell lines upregulating EGFR and show preference to partition to cancer cells rather than into macrophages [Chariou et al., 2015]. Moreover, PVX particles that were conjugated to the HER2 peptide specific to the Herceptin (Trastuzumab) monoclonal antibody showed capability to stimulate the HER2-positive cell line apoptosis [Esfandiari et al., 2015].
PVX VNPs functionalized with protein A fragments can be used in the display of whole antibody molecules as protein A binds to the IgG heavy chain Fc region [Wen and Steinmetz, 2016; Uhde-Holzem et al., 2015]. Such PVX particles could then be employed as a plug-and play mechanism to display a variety of antibodies with an orientational control level not accomplishable through chemical conjugation. The heterogeneity of primary Sjögren’s syndrome (pSjS) that is a chronic systemic autoimmune disorder often impedes diagnosis. Lipo peptide derived from lipocalin, a human autoantigen upon display on PVX VNPs served as a nanoparticle system specific to sera of pSjS patients with higher reactivity compared to the peptide alone [Tinazzi et al., 2015]. Deconstructed vectors derived from PVX have been variously used as expression systems. Efficient expression of the gene insert has been achieved by replacement of the TGB movement protein and CP genes [Komarova et al., 2006]. PTGS suppression enhanced protein yields in which transiently co-expressed TBSV P19 or TEV HC-Pro respectively resulted in 44% and 83% augmentation of gene expression [Larsen and Curtis, 2012]. Such a gene silencing suppression mechanism was also used to enhance the expression of a canine oral papillomavirus highly labile L1 vaccine protein from a PVX vector wherein this protein was fused to a chloroplast targeting peptide [Azhakanandam et al., 2007].
Pepino Mosaic Virus (PepMV)
The widespread PepMV, a Potexvirus is a propitious candidate for expression of recombinant proteins in plants due to its high-level accretion in its hosts as well as the absence of acute infection symptoms. PepMV undergoes distribution into most plant parts, establishing infection in flowers and roots. These properties render this virus as an attractive system for use as a tool to silence plant genes in various tissues.
Sempere et al., 2011 report the generation of a PepMV-based vector that has the ability to establish systemic foreign protein expression in N. benthamiana plants. Several strategies were used to enable stable expression of transgenes inclusive of substitution of the CP gene, CP subgenomic promoter duplication and a translational gene fusion. A stable PepMV vector was produced by the expression of the transgene as a fusion with the CP by the incorporation of the sequence that codes for the foot-and-mouth disease virus (FMDV) 2A catalytic peptide between the two genes. Through this, they achieved high recombinant protein expression, Incorporation of a translational enhancer into the PepMV-based vector enabled rapid foreign protein expression with protein levels as high as 4% of the total soluble protein [Abrahamian et al., 2021]. Such expression levels demonstrate that PepMV-based vectors are promising for the large-scale biosynthesis of pharmaceutical compounds.
5.4. Alfamovirus (Alfalfa Mosaic Virus (AlMV))
Alfalfa mosaic Virus-Based Vectors and Vaccine Development
Alfalfa mosaic virus is the type member of the genus Alfamovirus belonging to the family Bromoviridae with worldwide prevalence and broad host range. AMV spreads upon sowing of infected seeds or by aphid vectors [https://www.creative-biostructure.com/structural-research-of-alfalfa-mosaic-virus.html#:~:text=Alfalfa%20mosaic%20virus%20(AMV)%20of,%2C%20drug%20delivery%2C%20and%20nanotechnology]. AMV is a complex virus constituted by four distinct types of particles inclusive of three rod-shaped particles (30 to 57 nm) and one spherical particle. The rod-shaped particles have a hemispherical end containing pentagonal symmetry besides which there occurs a cylindrical portion having hexagonal extensions in different configurations. AMV harbors a single-stranded RNA genome constituted by RNA1, RNA2 and RNA3. Additionally, the subgenomic RNA4 is generated by transcription from the RNA3 negative-sense strand. Each of the genomic RNAs is encapsidated inside the rod-shaped particles in addition to RNA4. Each of RNA1 and RNA2 contains an ORF respectively coding for the P1 and P2 subunits of the viral replicase. RNA3 has 2 ORFs from which movement proteins as well as capsid proteins are expressed. The translation products of AMV-CP or RNA4 are necessary for viral RNA replication, packaging, transmittance and infection.
AMV has been engineered as VLPs towards vaccine development owing to its structural stability, immunogenic properties and capability to express foreign antigens upon genetic modification. VLPs of AMV can be generated in plants and employed in the deliverance of antigens for various diseases including cancers and infectious diseases through stimulation of specific T-cell and antibody responses. A major advantage of AMV is non-cytolytic effect wherein it does not kill cells directly, enabling efficient immune modulation within the tumor microenvironment and allowing for in situ vaccination.
Particles of AMV serve as platforms for the display of foreign antigens, stimulating the immune system to recognize the antigens and respond to them. AMV has been genetically engineered to express antigen targets inside host plant cells following which the resultant virus particles can be purified from plant tissue for use as vaccine components. AMV by itself naturally induces immune responses, thereby functioning as an inherent immunostimulant. AMV has been investigated for in situ vaccination wherein AMV nanoparticles are injected directly into the tumor to trigger the immune system to combat the cancer, converting the immunosuppressive tumor microenvironment into an immunostimulatory form, decelerating tumor growth and delaying metastasis.
AMV recruits immune cells and modulates cytokines that makes it a propitious candidate for use as therapeutic vaccines towards controlling several diseases. AMV particles show high stability, are easier to handle and have extended shelf life. Being a plant virus, it is non-pathogenic to humans which makes it a safer option compared to other vaccine platforms. The production of AMV in plants is economical and affords for rapid mass production, particularly in response to unprecedented increase in the demand for vaccines.
The Pfs25 protein expressed on the surfaces of Plasmodium falciparum gametes, ookinetes and zygotes has been considered as a prime target for the development of malaria transmission blocking vaccines. This Pfs25 malarial vaccine candidate has been engineered as a fusion with the AMV CP and expressed in plants [Chichester et al., 2018]. This vaccine candidate named Pfs25 VLP-FhCMB has been manufactured at pilot scale in N. benthamiana plants as per GMP guidelines. Phase I and first in-human study using healthy adult volunteers was performed in combination with the Alhydrogel® adjuvant wherein the plant-derived Pfs25-AMV VLP purified from the plants was injected into the subjects. At doses that were tested in this study, this vaccine formulation was found to elicit good antibody reactions at total protein doses higher than 30 mg and found to be safe with no incidence of vaccine-associated adverse events and toxicity. This proved that the vaccine had an acceptable tolerability and safety profile.
AMV can carry small antigenic peptides as fusions with its coat proteins and induce antigen-specific immune response. The Bacillus anthracis protective antigen domain-4 small loop 15 amino acid epitope (PA-D4s) was inserted immediately following the first 25 amino acids from the N-terminus of AMV CP [Brodzik et al., 2005]. This retained the genomic activation and association of the CP to viral RNAs. The resultant recombinant AMV particles were efficaciously expressed in tobacco plants, allowing for facile purification and immunological analyses in addition to exhibiting systemic proliferation and extended stability in the plants. Mice injected intraperitonially with the recombinant AMV particles expressing the PA-D4s epitope showed distinctive immune response wherein the immunized mice sera could recognize both the AMV CP as well as the Bacillus anthracis PA antigen.
Shahgolzari et al., 2021 showed that AMV upon administration as an in-situ vaccine elicited a variety of immune changes inside the tumor microenvironment leading to immune-enabled efficacy against the 4T1 murine triple-negative breast cancer model, which is highly aggressive, metastatic and recalcitrant to treatment with conventional immunotherapy. Mechanistic investigations showed that AMV ISV augments the production of inflammatory cytokines, immune effector cell infiltration and costimulatory molecules in addition to downregulating immune-suppressive molecules. In situ AMV vaccination switches the tumor microenvironment from its immunosuppressive mode to an immunostimulatory mode. AMV nanoparticles are neither cytolytic nor trigger apoptosis of the tumor cells in a direct manner but instead stimulate immune reactions against the tumor. Thus, AMV is naturally immunogenic, and this property can be used to develop AMV as an immunostimulatory agent for in situ vaccination into tumors. AMV ISV notably delays the growth of the tumors and tumor metastasis. The cytokines secreted by ISV with AMV provide direct evidence for its immunostimulatory properties reflecting on immune modulation mechanisms. AMV elicits more granulocytes inside the tumors and alters the tumor microenvironment through stimulating and recruiting immune-modulating cytokines leading to phenotypic changes within the immune cells.
A chimeric peptide containing the rabies virus nucleoprotein and glycoprotein determinants was fused to the AMV CP N-terminus and cloned into an AMV-based vector. When this fusion protein was transfected into spinach and N. benthamiana plants, it led to high level (0.4 mg/g FLW) accretion of the recombinant virus particles displaying the chimeric rabies virus epitopes [Yusibov et al., 2002]. Intraperitoneal administration of the vaccine formulation elicited robust systemic neutralizing antibody response in murine models and protected the mice against a lethal challenge of the rabies virus for at least 120 days. This proved the prolonged duration of the protective immune responses induced by plant-derived antigens and propounded that rabies virus epitope immunological memory was established by this immunogen. The chimeric peptide that was displayed on the recombinant AMV particles showed higher immunogenicity than the non-conjugated peptide formulation upon co-delivery with adjuvant in mouse models. Oral delivery of spinach leaves expressing these purified AMV particles induced strong mucosal IgA response and IgG / IgA serum immunity specific to rabies virus. Similar results were also achieved in humans demonstrating that plant-derived chimeric VLPs or cPVPs displaying rabies virus epitopes may afford a safer, economical and non-parenteral vaccine alternative for use in the developing world where rabies disease is more prevalent.
References
- Riedel, S. ; Edward Jenner and the history of smallpox and vaccination. Proc (Bayl Univ Med Cent) 2005, 18, 21–5. [Google Scholar]
- Koff, W.C.; et al. , Accelerating Next-Generation Vaccine Development for Global Disease Prevention. Science 2013, 340, 1232910. [Google Scholar]
- Rappuoli, R.; et al. , Vaccines, new opportunities for a new society. Proceedings of the National Academy of Sciences 2014, 111, 12288–12293. [Google Scholar]
- Henríquez, R. and I. Muñoz-Barroso, Viral vector- and virus-like particle-based vaccines against infectious diseases: A minireview. Heliyon 2024, 10, e34927. [Google Scholar] [PubMed]
- Wijesinghe, V.N.; et al. Current vaccine approaches and emerging strategies against herpes simplex virus (HSV). Expert review of vaccines 2021, 20, 1077–1096. [Google Scholar]
- Andrei, G. ; Vaccines and Antivirals: Grand Challenges and Great Opportunities. Frontiers in Virology 2021, Volume 1 - 2021.
- Hefferon, K.L. ; Plant virus expression vectors set the stage as production platforms for biopharmaceutical proteins. Virology 2012, 433, 1–6. [Google Scholar] [PubMed]
- Cid, R. and J. Bolívar, Platforms for Production of Protein-Based Vaccines: From Classical to Next-Generation Strategies. Biomolecules 2021, 11(8).
- Larrick, J.W. and D.W. Thomas, Producing proteins in transgenic plants and animals. Current opinion in biotechnology 2001, 12, 411–418. [Google Scholar] [PubMed]
- Tiwari, S.; et al. Plants as bioreactors for the production of vaccine antigens. Biotechnol Adv 2009, 27, 449–67. [Google Scholar] [CrossRef]
- Daniell, H. ; S.J. Streatfield, and E.P. Rybicki, Advances in molecular farming: key technologies, scaled up production and lead targets. Plant Biotechnol J 2015, 13, 1011–2. [Google Scholar]
- Shanmugaraj, B. ; I.B. CJ, and W. Phoolcharoen, Plant Molecular Farming: A Viable Platform for Recombinant Biopharmaceutical Production. Plants (Basel) 2020, 9. [Google Scholar]
- Huang, X.; et al. Escherichia coli-derived virus-like particles in vaccine development. npj Vaccines 2017, 2, 3. [Google Scholar] [CrossRef]
- Tran, H.H. ; Developing a plant virus-based expression system for the expression of vaccines against Porcine Reproductive and Respiratory Syndrome Virus. 2017, The University of Western Ontario (Canada).
- Mason, H.S.; et al. Edible plant vaccines: applications for prophylactic and therapeutic molecular medicine. Trends in molecular medicine 2002, 8, 324–329. [Google Scholar] [CrossRef]
- Lomonossoff, G.P. and M.-A. D’Aoust, Plant-produced biopharmaceuticals: a case of technical developments driving clinical deployment. Science 2016, 353, 1237–1240. [Google Scholar] [CrossRef] [PubMed]
- Lico, C. ; Q. Chen, and L. Santi, Viral vectors for production of recombinant proteins in plants. Journal of cellular physiology 2008, 216, 366–377. [Google Scholar] [CrossRef] [PubMed]
- Rybicki, E.P. ; Plant molecular farming of virus--like nanoparticles as vaccines and reagents. Wiley Interdisciplinary Reviews: Nanomedicine and Nanobiotechnology 2020, 12, e1587. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q. and H. Lai, Plant-derived virus-like particles as vaccines. Human vaccines & immunotherapeutics 2013, 9, 26–49. [Google Scholar]
- Scotti, N. and E.P. Rybicki, Virus-like particles produced in plants as potential vaccines. Expert review of vaccines 2013, 12, 211–224. [Google Scholar] [CrossRef]
- Nuñez-Rivera, A.; et al. Brome mosaic virus-like particles as siRNA nanocarriers for biomedical purposes. Beilstein Journal of Nanotechnology 2020, 11, 372–382. [Google Scholar] [CrossRef]
- Villanueva-Flores, F.; et al. A novel formulation of Asparaginase encapsulated into virus-like particles of brome mosaic virus: In vitro and in vivo evidence. Pharmaceutics 2023, 15, 2260. [Google Scholar] [CrossRef]
- Lico, C. ; Q. Chen, and L. Santi, Viral vectors for production of recombinant proteins in plants. J Cell Physiol 2008, 216, 366–77. [Google Scholar] [CrossRef]
- Grimsley, N.; et al. “Agroinfection,” an alternative route for viral infection of plants by using the Ti plasmid. Proc Natl Acad Sci U S A 1986, 83, 3282–6. [Google Scholar] [CrossRef]
- Phiri, K. and L. Grill, Development of a Candidate TMV Epitope Display Vaccine against SARS-CoV-2. Vaccines 2024, 12, 448. [Google Scholar] [CrossRef]
- Pérez Filgueira, D.M.; et al. Bovine herpes virus gD protein produced in plants using a recombinant tobacco mosaic virus (TMV) vector possesses authentic antigenicity. Vaccine 2003, 21, 4201–4209. [Google Scholar] [CrossRef]
- Wagner, B.; et al. Plant virus expression systems for transient production of recombinant allergens in Nicotiana benthamiana. Methods 2004, 32, 227–234. [Google Scholar] [CrossRef]
- Blokhina, E.A.; et al. Chimeric Virus-like Particles of Physalis Mottle Virus as Carriers of M2e Peptides of Influenza a Virus. Viruses 2024, 16, 1802. [Google Scholar] [CrossRef] [PubMed]
- Smolenska, L.; et al. Production of a functional single chain antibody attached to the surface of a plant virus. FEBS Letters 1998, 441, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Arazi, T.; et al. Production of Antiviral and Antitumor Proteins MAP30 and GAP31 in Cucurbits Using the Plant Virus Vector ZYMV-AGII. Biochemical and Biophysical Research Communications 2002, 292, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Rivera, O.A.; et al. Cowpea Mosaic Virus Nanoparticle Vaccine Candidates Displaying Peptide Epitopes Can Neutralize the Severe Acute Respiratory Syndrome Coronavirus. ACS Infectious Diseases 2021, 7, 3096–3110. [Google Scholar] [CrossRef] [PubMed]
- Matić, S.; et al. Efficient production of chimeric Human papillomavirus 16 L1 protein bearing the M2e influenza epitope in Nicotiana benthamiana plants. BMC Biotechnology 2011, 11, 106. [Google Scholar] [CrossRef]
- Yusibov, V.; et al. Antigens produced in plants by infection with chimeric plant viruses immunize against rabies virus and HIV-1. Proceedings of the National Academy of Sciences 1997, 94, 5784–5788. [Google Scholar] [CrossRef]
- Wang, L.; et al. Development of a pod pepper vein yellows virus-based expression vector for the production of heterologous protein or virus like particles in Nicotiana benthamiana. Virus Research 2025, 355, 199559. [Google Scholar] [CrossRef]
- Takamatsu, N.; et al. Expression of bacterial chloramphenicol acetyltransferase gene in tobacco plants mediated by TMV-RNA. Embo j 1987, 6, 307–11. [Google Scholar] [CrossRef]
- Dawson, W.O.; et al. A tobacco mosaic virus-hybrid expresses and loses an added gene. Virology 1989, 172, 285–92. [Google Scholar] [CrossRef]
- Chapman, S. ; T. Kavanagh, and D. Baulcombe, Potato virus X as a vector for gene expression in plants. Plant J 1992, 2, 549–57. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, K.; et al. Engineering cowpea mosaic virus RNA-2 into a vector to express heterologous proteins in plants. Virology 2000, 267, 159–73. [Google Scholar] [CrossRef] [PubMed]
- Musiychuk, K.; et al. A launch vector for the production of vaccine antigens in plants. Influenza and Other Respiratory Viruses 2007, 1, 19–25. [Google Scholar] [CrossRef]
- Giritch, A.; et al. Rapid high-yield expression of full-size IgG antibodies in plants coinfected with noncompeting viral vectors. Proceedings of the National Academy of Sciences 2006, 103, 14701–14706. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.S. and W.R. Curtis, RNA viral vectors for improved Agrobacterium-mediated transient expression of heterologous proteins in Nicotiana benthamiana cell suspensions and hairy roots. BMC Biotechnology 2012, 12, 21. [Google Scholar] [CrossRef]
- Sugiyama, Y.; et al. Systemic production of foreign peptides on the particle surface of tobacco mosaic virus. FEBS Letters 1995, 359, 247–250. [Google Scholar] [CrossRef]
- Uhde-Holzem, K.; et al. Immunogenic properties of chimeric potato virus X particles displaying the hepatitis C virus hypervariable region I peptide R9. Journal of Virological Methods 2010, 166, 12–20. [Google Scholar] [CrossRef]
- Haynes, J.R.; et al. Development of a Genetically–Engineered, Candidate Polio Vaccine Employing the Self–Assembling Properties of the Tobacco Mosaic Virus Coat Protein. Bio/Technology 1986, 4, 637–641. [Google Scholar] [CrossRef]
- Bachmann, M.F. and G.T. Jennings, Vaccine delivery: a matter of size, geometry, kinetics and molecular patterns. Nature Reviews Immunology 2010, 10, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Marconi, G.; et al. In planta production of two peptides of the Classical Swine Fever Virus (CSFV) E2 glycoprotein fused to the coat protein of potato virus X. BMC Biotechnology 2006, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- Marusic, C.; et al. Chimeric Plant Virus Particles as Immunogens for Inducing Murine and Human Immune Responses against Human Immunodeficiency Virus Type 1. Journal of Virology 2001, 75, 8434–8439. [Google Scholar] [CrossRef] [PubMed]
- Lico, C.; et al. Plant-produced potato virus X chimeric particles displaying an influenza virus-derived peptide activate specific CD8+ T cells in mice. Vaccine 2009, 27, 5069–5076. [Google Scholar] [CrossRef]
- Porta, C.; et al. Development of cowpea mosaic virus as a high-yielding system for the presentation of foreign peptides. Virology 1994, 202, 949–955. [Google Scholar] [CrossRef]
- Patel, B.K.; et al. Cowpea Mosaic Virus (CPMV)-Based Cancer Testis Antigen NY-ESO-1 Vaccine Elicits an Antigen-Specific Cytotoxic T Cell Response. ACS Appl Bio Mater 2020, 3, 4179–4187. [Google Scholar] [CrossRef]
- Beiss, V.; et al. Cowpea Mosaic Virus Outperforms Other Members of the Secoviridae as In Situ Vaccine for Cancer Immunotherapy. Mol Pharm 2022, 19, 1573–1585. [Google Scholar] [CrossRef]
- Langeveld, J.P.M.; et al. Inactivated recombinant plant virus protects dogs from a lethal challenge with canine parvovirus. Vaccine 2001, 19, 3661–3670. [Google Scholar] [CrossRef]
- Sainsbury, F.; et al. Expression of multiple proteins using full-length and deleted versions of cowpea mosaic virus RNA-2. Plant Biotechnology Journal 2008, 6, 82–92. [Google Scholar] [CrossRef]
- Namba, K. and G. Stubbs, Structure of Tobacco Mosaic Virus at 3.6 Å Resolution: Implications for Assembly. Science 1986, 231, 1401–1406. [Google Scholar] [CrossRef]
- Bendahmane, M.; et al. Display of epitopes on the surface of tobacco mosaic virus: impact of charge and isoelectric point of the epitope on virus-host interactions. J Mol Biol 1999, 290, 9–20. [Google Scholar] [CrossRef]
- Turpen, T.H.; et al. Malaria Epitopes Expressed on the surface of Recombinant Tobacco Mosaic Virus. Bio/Technology 1995, 13, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Takamatsu, N.; et al. Production of enkephalin in tobacco protoplasts using tobacco mosaic virus RNA vector. FEBS Letters 1990, 269, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.- S.; Vaira, A.M.; Domier, L.L.; Lee, S.C.; Kim, H.G. & Hammond, J. Efficiency of VIGS and gene expression in a novel bipar tite potexvirus vector delivery system as a function of strength of TGB1 silencing suppression. Virology 2010, 402, 149–163. [Google Scholar] [PubMed]
- Bouton, C.; King, R.C.; Chen, H.; Azhakanandam, K.; Bieri, S.; Hammond- Kosack, K.E.; et al. Foxtail mosaic virus: a viral vector for pro tein expression in cereals. Plant Physiology 2018, 177, 1352–1367. [Google Scholar] [CrossRef]
- Liu, N.; Xie, K.; Jia, Q.; Zhao, J.; Chen, T.; Li, H.; et al. Foxtail mosaic virus- induced gene silencing in monocot plants. Plant Physiology 2016, 171, 1801–1807. [Google Scholar] [CrossRef]
- Mei, Y.; Zhang, C.; Kernodle, B.M.; Hill, J.H. & Whitham, S. A. A foxtail mosaic virus vector for virus- induced gene silencing in maize. Plant Physiology 2016, 171, 760–772. [Google Scholar]
- Mei, Y.; Beernink, B.M.; Ellison, E.E.; Konečná, E.; Neelakandan, A.K.; Voytas, D.F.; et al. (2019) Protein expression and gene editing in monocots using foxtail mosaic virus vectors. Plant Direct, 3, e00181.
- Tuo, D.; Zhou, P.; Yan, P.; Cui, H.; Liu, Y.; Wang, H.; et al. A cassava common mosaic virus vector for virus- induced gene silencing in cassava. Plant Methods 2021, 17, 74. [Google Scholar] [CrossRef]
- Hsieh, M.-H.; Lu, H.-C.; Pan, Z.-J.; Yeh, H.-H.; Wang, S.-S.; Chen, W.-H.; et al. Optimizing virus- induced gene silencing efficiency with cymbidium mosaic virus in Phalaenopsis flower. Plant Science 2013, 201-202, 25–41. [Google Scholar] [CrossRef]
- Abrahamian, P.; Hammond, J.; Hammond, R.W. Development and optimization of a Pepino mosaic virus- based vector for rapid expression of heterologous proteins in plants. Applied Microbiology and Biotechnology 2021, 105, 627–645. [Google Scholar] [CrossRef]
- Sempere, R.N.; Gomez, P.; Truniger, V.; Aranda, M.A. Development of expression vectors based on Pepino mosaic virus. Plant Methods 2011, 7, 6. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L.; Hunter, D.; Voogd, C.; Joyce, N.; Davies, K. A narcissus mosaic viral vector system for protein expression and flavonoid production. Plant Methods 2013, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Koenig, R. ; Lesemann D-E.; Loss, S.; Engelmann, J.; Commandeur, U.; Deml, G. et al. Zygocactus virus X- based expression vectors and formation of rod- shaped virus- like particles in plants by the expressed coat proteins of beet necrotic yellow vein virus and soil- borne cereal mosaic virus. Journal of General Virology 2006, 87, 439–443. [Google Scholar] [PubMed]
- Minato, N.; Komatsu, K.; Okano, Y.; Maejima, K.; Ozeki, J.; Senshu, H.; et al. Efficient foreign gene expression in planta using a Plantago asiatica mosaic virus- based vector achieved by the strong RNA- silencing suppressor activity of TGBp1. Archives of Virology 2014, 159, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Chen T-H.; Hu C-C.; Liao J-T.; Lee Y-L.; Huang Y-W.; Lin N-S. et al. Production of Japanese encephalitis virus antigens in plants using bamboo mosaic virus- based vector. Frontiers in Microbiology 2017, 8, 788.
- Lin, N.S.; Lee, Y.S.; Lin, B.Y.; Lee, C.W.; Hsu, Y.H. The open read ing frame of bamboo mosaic potexvirus satellite RNA is not essen tial for its replication and can be replaced with a bacterial gene. Proceedings of the National Academy of Sciences of the United States of America 1996, 93, 3138–3142. [Google Scholar] [CrossRef]
- Liou M-R.; Huang Y-W.; Hu C-C.; Lin N-S.; Hsu Y-H. A dual gene- silencing vector system for monocot and dicot plants. Plant Biotechnology Journal 2014, 12, 330–343. [CrossRef]
- Salazar-González JA, Bañuelos-Hernández B, Rosales-Mendoza S. Current status of viral expression systems in plants and perspectives for oral vaccines development. Plant Mol Biol. 2015, 87, 203–17. [CrossRef]
- Baulcombe D, Chapman S, Santa Cruz S. Jellyfish green f luorescent protein as a reporter for virus infections. Plant J 1995, 7, 1045–1053. [CrossRef]
- Marusic C, Rizza P, Lattanzi L, Mancini C, Spada M, Belardelli F, Benvenuto E, Capone I Chimeric plant virus parti cles as immunogens for inducing murine and human immune responses against human immunodeficiency virus type 1. J Virol 2001, 75, 8434–8439. [CrossRef]
- Uhde K, Fischer R, Commandeur U Expression of multiple foreign epitopes presented as synthetic antigens on the surface of Potato virus X particles. Arch Virol 2005, 150, 327–340. [CrossRef]
- Santa Cruz S, Chapman S, Roberts AG, Roberts IM, Prior DAM, Oparka KJ. Assembly and movement of a plant virus car rying a green fluorescent protein overcoat. Proc Natl Acad Sci U S A 1996, 93, 6286–6290. [Google Scholar] [CrossRef]
- Toth RL, Chapman S, Carr F, Santa Cruz S. A novel strategy for the expression of foreign genes from plant virus vectors. FEBS Lett 2001, 489, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Čerˇovská N, Hoffmeisterová H, Pecenková T, Moravec T, Synková H, Plchová H, Velemínský J. Transient expression of HPV16 E7 peptide (aa 44–60) and HPV16 L2 peptide (aa 108–120) on chimeric potyvirus-like particles using Potato virus X-based vector. Protein Expr Purif 2008, 58, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Čerˇovská N, Hoffmeisterova H, Moravec T, Plchova H, Folwarc zna J, Synkova H, Ryslava H, Ludvikova V, Smahel M. Transient expression of human papillomavirus type 16 L2 epitope fused to N- and C-terminus of coat protein of Potato virus X in plants. J Biosci 2012, 37, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Ravin NV, Kotlyarov RY, Mardanova ES, Kuprianov VV, Migunov AI, Stepanova LA, Tsybalova LM, Kiselev OI, Skryabin KG. Plant-produced recombinant influenza vaccine based on virus-like HBc particles carrying an extracellular domain of M2 protein. Biochemistry (Mosc) 2012, 77, 33–40. [Google Scholar] [CrossRef]
- Franconi, R.; Di Bonito, P.; Dibello, F.; Accardi, L.; Muller, A.; Cirilli, A.; Simeone, P.; Dona, M.G.; Venuti, A.; and Giorgi, C. Plant derived-human papillomavirus 16 E7 oncoprotein induces immune response and specific tumour protection. Cancer Res. 2002, 62, 3654–3658. [Google Scholar]
- Franconi R, Massa S, Illiano E, et al. Exploiting the Plant Secretory Pathway to Improve the Anticancer Activity of a Plant-Derived HPV16 E7 Vaccine. International Journal of Immunopathology and Pharmacology. [CrossRef]
- Röder J, Dickmeis C and Commandeur U. Small, Smaller, Nano: New Applications for Potato Virus X in Nanotechnology. Front. Plant Sci. 2019, 10, 158. [Google Scholar] [CrossRef]
- Lico C, Benvenuto E and Baschieri S. The Two-Faced Potato Virus X: From Plant Pathogen to Smart Nanoparticle. Front. Plant Sci 2015, 6, 1009. [CrossRef]
- Lico, C.; Mancini, C.; Italiani, P.; Betti, C.; Boraschi, D.; Benvenuto, E.; et al. Plant-produced potato virus X chimeric particles displaying an influenza virus derived peptide activate specific CD8+ T cells in mice. Vaccine 2009, 27, 5069–5076. [Google Scholar] [CrossRef]
- Jobsri, J.; Allen, A.; Rajagopal, D.; Shipton, M.; Kanyuka, K.; Lomonossoff, G. P.; et al. Plant virus particles carrying tumour antigen activate TLR7 and Induce high levels of protective antibody. PLoS ONE 2015, 10, e0118096. [Google Scholar] [CrossRef]
- Brennan, F. R.; Jones, T. D.; Longstaff, M.; Chapman, S.; Bellaby, T.; Smith, H.; et al. Immunogenicity of peptides derived from a fibronectin-binding protein of S. aureus expressed ontwodifferentplant viruses. Vaccine 1999, 17, 1846–1857. [Google Scholar] [CrossRef]
- Marconi, G.; Albertini, E.; Barone, P.; De Marchis, F.; Lico, C.; Marusic, C.; et al. In planta production of two peptides of the Classical Swine Fever Virus (CSFV) E2 glycoprotein fused to the coat protein of potato virus X. BMC Biotechnol. 2006, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- Uhde-Holzem, K.; Schlösser, V.; Viazov, S.; Fischer, R.; and Commandeur, U. Immunogenic properties of chimeric potato virus X particles displaying the hepatitis C virus hypervariable region I peptide R9. J. Virol. Methods 2010, 166, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Wen, A.M.; Commandeur, U.; and Steinmetz, N.F. Presentation of HER2epitopes using a filamentous plant virus-based vaccination platform. J. Mater. Chem. B 2014, 2, 6249–6258. [Google Scholar] [CrossRef] [PubMed]
- Chariou, P. L.; Lee, K. L.; Wen, A. M.; Gulati, N. M.; Stewart, P. L.; and Steinmetz, N. F. Detection and imaging of aggressive cancer cells using an epidermal growth factor receptor (EGFR)-targeted filamentous plant virus-based nanoparticle. Bioconjug. Chem. 2015, 26, 262–269. [Google Scholar] [CrossRef]
- Esfandiari, N.; Arzanani, M. K.; Soleimani, M.; Kohi-Habibi, M.; and Svendsen, W. E. A new application of plant virus nanoparticles as drug delivery in breast cancer. Tumour. Biol. 2015. [CrossRef]
- Sempere RN, Gómez P, Truniger V, Aranda MA. Development of expression vectors based on pepino mosaic virus. Plant Methods. 2011, 7, 6. [Google Scholar] [CrossRef]
- Abrahamian, P.; Hammond, J. & Hammond, R. W. Development and optimization of a Pepino mosaic virus- based vector for rapid expression of heterologous proteins in plants. Applied Microbiology and Biotechnology 2021, 105, 627–645. [Google Scholar]
- Beernink BM, Whitham SA. Foxtail mosaic virus: A tool for gene function analysis in maize and other monocots. Mol Plant Pathol. 2023, 24, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Wen AM, Steinmetz NF. Design of virus-based nanomaterials for medicine, biotechnology, and energy. Chem Soc Rev. 2016, 45, 4074–126. [Google Scholar] [CrossRef] [PubMed]
- Uhde-Holzem K, McBurney M, Tiu BD, Advincula RC, Fischer R, Commandeur U, Steinmetz NF. Macromol. Biosci. 2015. [CrossRef]
- Tinazzi E, Merlin M, Bason C, Beri R, Zampieri R, Lico C, Bartoloni E, Puccetti A, Lunardi C, Pezzotti M, Avesani L. Front. Plant Sci. 2015, 6. [PubMed: 25657654].
- Komarova TV, Skulachev MV, Zvereva AS, Schwartz AM, Dorokhov YL, Atabekov JG. Biochemistry (Mosc). 2006, 71, 846–850.
- Larsen JS, Curtis WR. BMC Biotechnol. 2012, 12, 21.
- Azhakanandam K, Weissinger SM, Nicholson JS, Qu R, Weissinger AK. Plant Mol. Biol. 2007, 63, 393–404.
- Chichester JA, Green BJ, Jones RM, Shoji Y, Miura K, Long CA, Lee CK, Ockenhouse CF, Morin MJ, Streatfield SJ, Yusibov V. Safety and immunogenicity of a plant-produced Pfs25 virus-like particle as a transmission blocking vaccine against malaria: A Phase 1 dose-escalation study in healthy adults. Vaccine 2018, 36, 5865–5871. [Google Scholar] [CrossRef]
- R. Brodzik, K. Bandurska, D. Deka, M. Golovkin, H. Koprowski, Advances in alfalfa mosaic virus-mediated expression of anthrax antigen in planta, Biochemical and Biophysical Research Communications, Volume 338, Issue 2, 2005, Pages 717-722, ISSN 0006-291X,. [CrossRef]
- Shahgolzari M, Pazhouhandeh M, Milani M, Fiering S, Khosroushahi AY. Alfalfa mosaic virus nanoparticles-based in situ vaccination induces antitumor immune responses in breast cancer model. Nanomedicine (Lond). 2021, 16, 97–107. [Google Scholar] [CrossRef]
- Yusibov V, Hooper DC, Spitsin SV, Fleysh N, Kean RB, Mikheeva T, et al. Expression in plants and immu nogenicity of plant virus-based experimental rabies vaccine. Vaccine 2002, 20, 3155–64. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



