Submitted:
02 October 2025
Posted:
06 October 2025
You are already at the latest version
Abstract
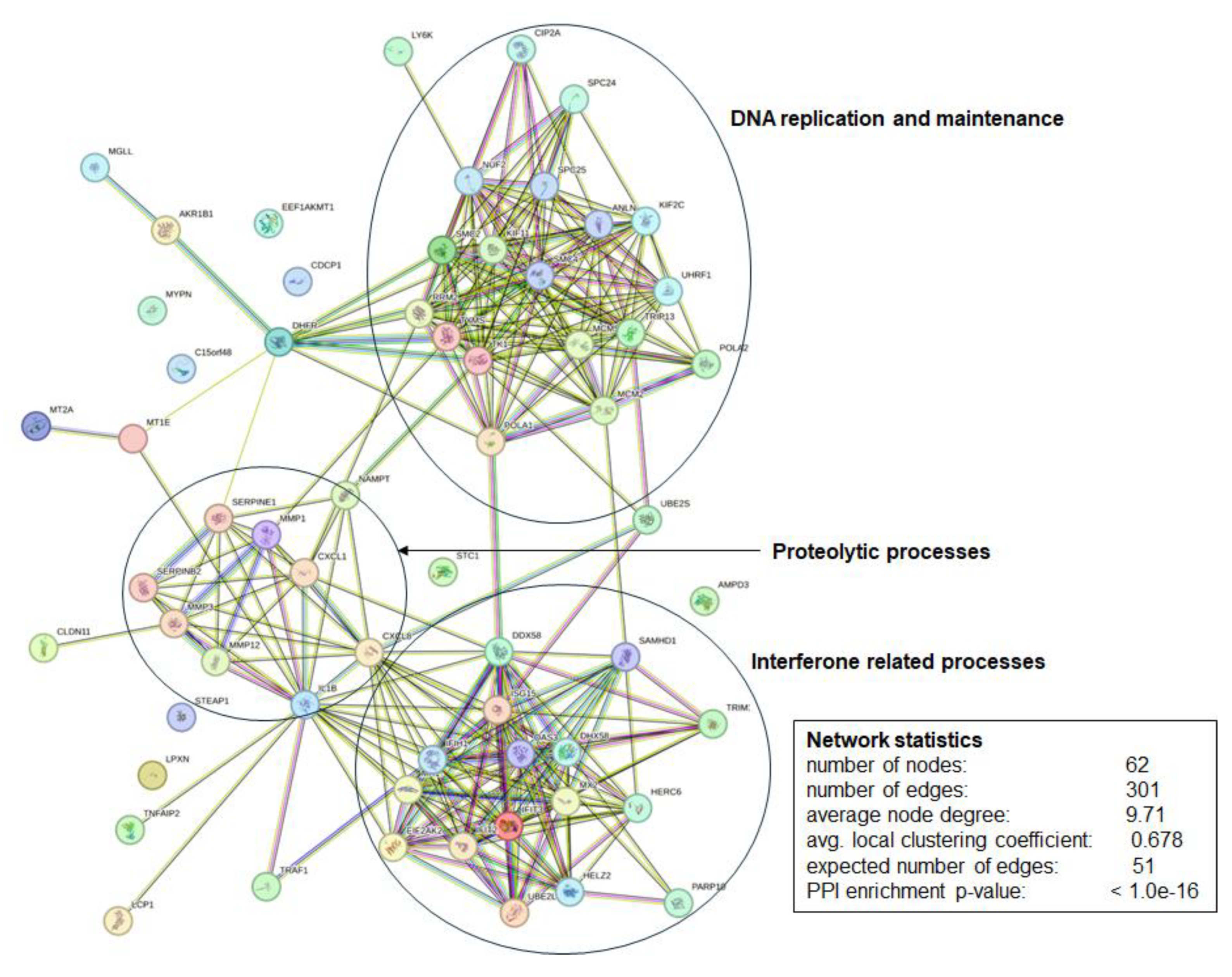
Toxoplasma gondii, the causative agent of toxoplasmosis widespread in animals and humans, is an intracellular apicomplexan protozoan parasite infecting a variety of host cells. Gene editing using CRISPR-Cas9 has become a standard tool to investigate the molecular genetics of this interaction. With respect to gene knock-out (KO) studies, the general paradigm implies that the gene of interest is expressed in the wildtype and that only the gene of interest is affected by the knock-out. Consequently, the observed phenotype depends on presence or absence of genes of interest. To challenge this paradigm, we knocked out two open reading frames (ORFs) constitutively expressed in T. gondii ShSp1 tachyzoites, but not essential, namely ORF 297720 encoding a trehalose-6-phosphatase homolog and ORF 319730 encoding a You2 C2C2 zinc finger homolog. We analyzed the proteomes of tachyzoites isolated at a late stage of infection, of intracellular tachyzoites and of host cells at an early stage of infection. The intended KO proteins were present in the T. gondii Sp1 wildtype but absent in the KO clones. Moreover, besides differentially expressed (DE) proteins specific to each KO, 17 DE proteins common to both KOs were identified in isolated and 39 in intracellular tachyzoites. Moreover, 76 common DE proteins were identified in host cells. Network and enrichment analyses showed that these proteins were functionally related to antiviral defense mechanisms. These results indicate that the KO of a gene of interest may not only affect the expression of other genes of the target organism, in our case T. gondii, but also the gene expression of its host cells. Therefore, phenotypes of KO strains may not be causally related to the KO of a given gene.
Keywords:
1. Introduction
2. Results
2.1. Molecular Genetic Characterization of the Knock-Outs
2.2. Growth Characteristics of the KO Clones
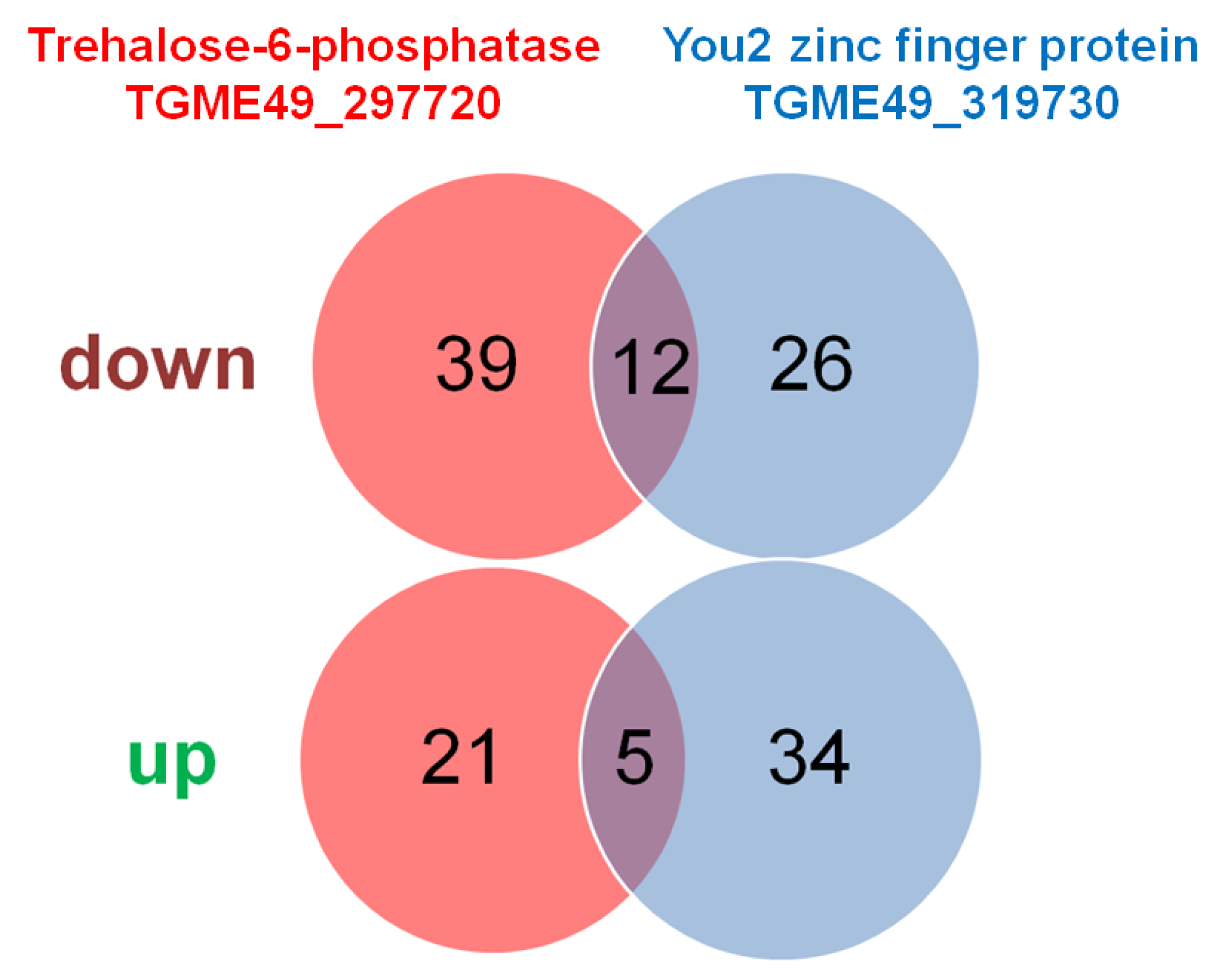
2.3. Differentially Expressed Proteins in Isolated Tachyzoites
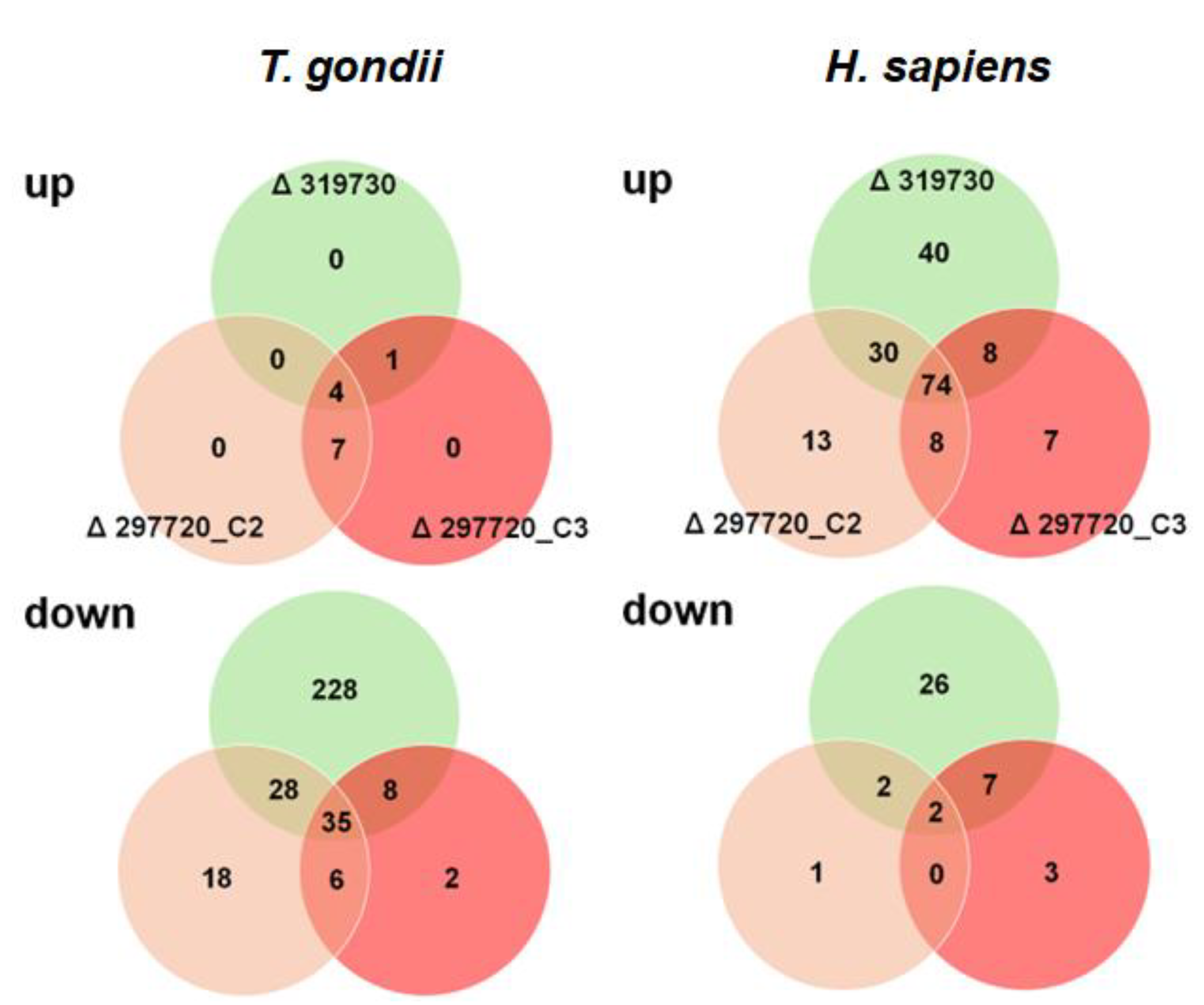
2.4. Differentially Expressed Proteins in Infected Host Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. In vitro Culture and Parasite Maintenance
4.3. Isolation of Parasites
4.4. Generation of the Knock-Out Strains
4.5. Growth Analysis
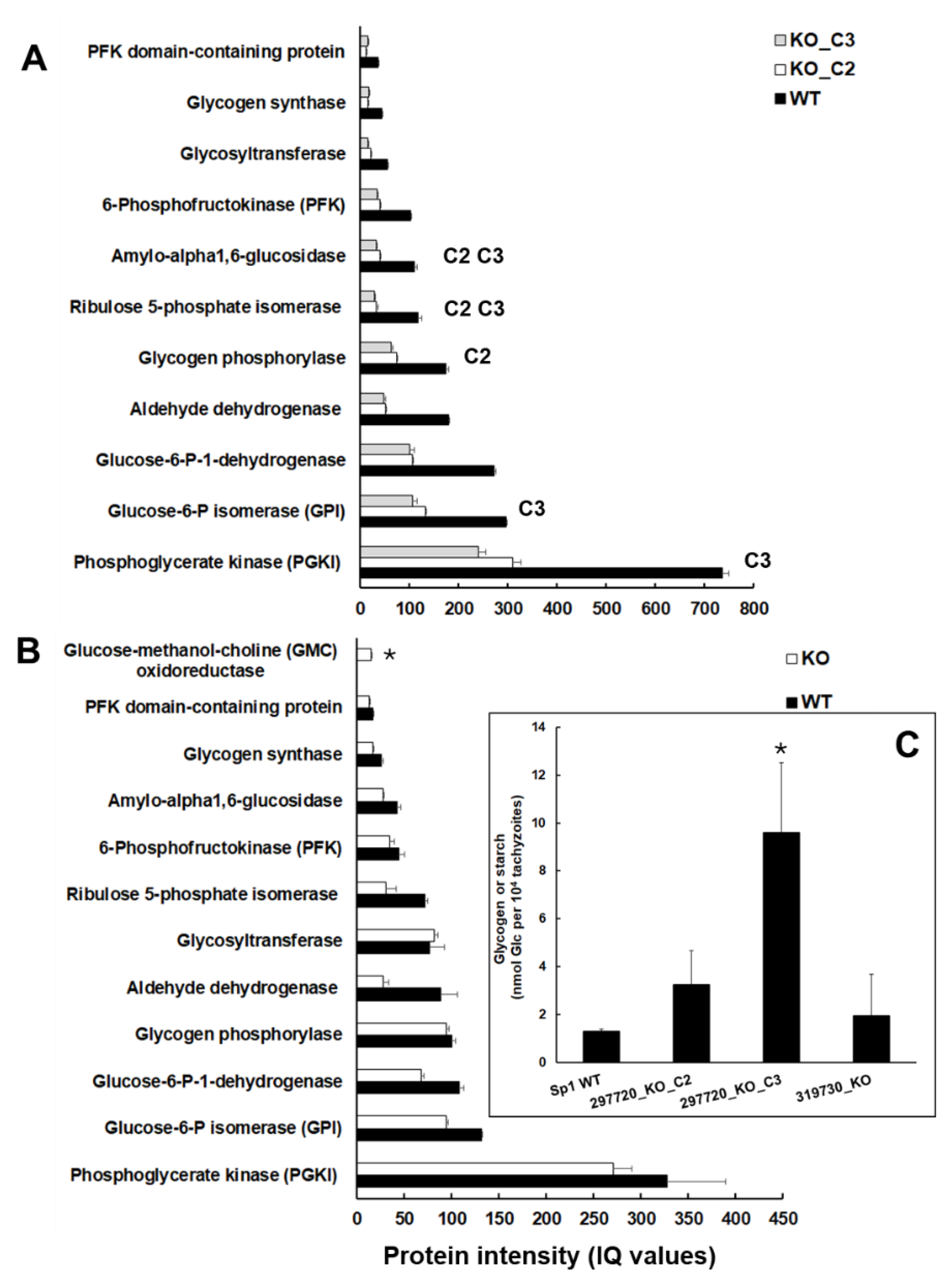
4.6. Quantification of Glycogen/Starch
4.7. Proteomic Analysis of Isolated Tachyzoites
4.8. Proteomic Analysis of Infected Host Cells
4.9. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukes, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.W.; Burki, F.; et al. Revisions to the classification, nomenclature, and diversity of eukaryotes. J Eukaryot Microbiol 2019, 66, 4–119. [Google Scholar] [CrossRef]
- Dubey, J.P. Outbreaks of clinical toxoplasmosis in humans: five decades of personal experience, perspectives and lessons learned. Parasit Vectors 2021, 14, 263. [Google Scholar] [CrossRef]
- Beltrame, A.; Kim, K.; Weiss, L.M. Human toxoplasmosis: current advances in the field. Curr Opin Infect Dis 2025. [Google Scholar] [CrossRef] [PubMed]
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Weiss, L.M. Toxoplasma gondii: the model apicomplexan. Int J Parasitol 2004, 34, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Sidik, S.M.; Hackett, C.G.; Tran, F.; Westwood, N.J.; Lourido, S. Efficient genome engineering of Toxoplasma gondii using CRISPR/Cas9. PLoS One 2014, 9, e100450. [Google Scholar] [CrossRef]
- Hildebrandt, F.; A, N.M.; Treeck, M. A CRISPR view on genetic screens in Toxoplasma gondii. Curr Opin Microbiol 2025, 83, 102577. [Google Scholar] [CrossRef]
- Sidik, S.M.; Huet, D.; Ganesan, S.M.; Huynh, M.H.; Wang, T.; Nasamu, A.S.; Thiru, P.; Saeij, J.P.; Carruthers, V.B.; Niles, J.C.; et al. A Genome-wide CRISPR screen in Toxoplasma Identifies essential apicomplexan genes. Cell 2016, 166, 1423–1435 e1412. [Google Scholar] [CrossRef]
- Müller, J.; Boubaker, G.; Müller, N.; Uldry, A.C.; Braga-Lagache, S.; Heller, M.; Hemphill, A. Investigating antiprotozoal chemotherapies with novel proteomic tools-chances and limitations: A critical review. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Hänggeli, K.P.A.; Hemphill, A.; Müller, N.; Heller, M.; Uldry, A.C.; Braga-Lagache, S.; Müller, J.; Boubaker, G. Comparative proteomic analysis of Toxoplasma gondii RH wild-type and four SRS29B (SAG1) knock-out clones reveals significant differences between individual strains. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Anghel, N.; Müller, J.; Serricchio, M.; Jelk, J.; Butikofer, P.; Boubaker, G.; Imhof, D.; Ramseier, J.; Desiatkina, O.; Paunescu, E.; et al. Cellular and molecular targets of nucleotide-tagged trithiolato-bridged arene ruthenium complexes in the protozoan parasites Toxoplasma gondii and Trypanosoma brucei. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Fritz, H.M.; Buchholz, K.R.; Chen, X.; Durbin-Johnson, B.; Rocke, D.M.; Conrad, P.A.; Boothroyd, J.C. Transcriptomic analysis of Toxoplasma development reveals many novel functions and structures specific to sporozoites and oocysts. PLoS One 2012, 7, e29998. [Google Scholar] [CrossRef] [PubMed]
- Mayoral, J.; Tomita, T.; Tu, V.; Aguilan, J.T.; Sidoli, S.; Weiss, L.M. Toxoplasma gondii PPM3C, a secreted protein phosphatase, affects parasitophorous vacuole effector export. PLoS Pathog 2020, 16, e1008771. [Google Scholar] [CrossRef]
- Antunes, A.V.; Shahinas, M.; Swale, C.; Farhat, D.C.; Ramakrishnan, C.; Bruley, C.; Cannella, D.; Robert, M.G.; Corrao, C.; Coute, Y.; et al. In vitro production of cat-restricted Toxoplasma pre-sexual stages. Nature 2024, 625, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Baptista, C.G.; Hosking, S.; Gas-Pascual, E.; Ciampossine, L.; Abel, S.; Hakimi, M.A.; Jeffers, V.; Le Roch, K.; West, C.M.; Blader, I.J. The Toxoplasma gondii F-Box protein L2 functions as a repressor of stage specific gene expression. PLoS Pathog 2024, 20, e1012269. [Google Scholar] [CrossRef]
- Kerbler, S.M.; Armijos-Jaramillo, V.; Lunn, J.E.; Vicente, R. The trehalose 6-phosphate phosphatase family in plants. Physiol Plant 2023, 175, e14096. [Google Scholar] [CrossRef] [PubMed]
- Beraki, T.; Hu, X.; Broncel, M.; Young, J.C.; O'Shaughnessy, W.J.; Borek, D.; Treeck, M.; Reese, M.L. Divergent kinase regulates membrane ultrastructure of the Toxoplasma parasitophorous vacuole. Proc Natl Acad Sci U S A 2019, 116, 6361–6370. [Google Scholar] [CrossRef]
- Gosztyla, M.L.; Zhan, L.; Olson, S.; Wei, X.; Naritomi, J.; Nguyen, G.; Street, L.; Goda, G.A.; Cavazos, F.F.; Schmok, J.C.; et al. Integrated multi-omics analysis of zinc-finger proteins uncovers roles in RNA regulation. Mol Cell 2024, 84, 3826–3842 e3828. [Google Scholar] [CrossRef]
- Rashidi, S.; Vieira, C.; Mansouri, R.; Ali-Hassanzadeh, M.; Ghani, E.; Karimazar, M.; Nguewa, P.; Manzano-Roman, R. Host cell proteins modulated upon Toxoplasma infection identified using proteomic approaches: a molecular rationale. Parasitol Res 2022, 121, 1853–1865. [Google Scholar] [CrossRef]
- Kemp, L.E.; Yamamoto, M.; Soldati-Favre, D. Subversion of host cellular functions by the apicomplexan parasites. FEMS Microbiol Rev 2013, 37, 607–631. [Google Scholar] [CrossRef]
- Bougdour, A.; Tardieux, I.; Hakimi, M.A. Toxoplasma exports dense granule proteins beyond the vacuole to the host cell nucleus and rewires the host genome expression. Cell Microbiol 2014, 16, 334–343. [Google Scholar] [CrossRef]
- Bougdour, A.; Durandau, E.; Brenier-Pinchart, M.P.; Ortet, P.; Barakat, M.; Kieffer, S.; Curt-Varesano, A.; Curt-Bertini, R.L.; Bastien, O.; Coute, Y.; et al. Host cell subversion by Toxoplasma GRA16, an exported dense granule protein that targets the host cell nucleus and alters gene expression. Cell Host Microbe 2013, 13, 489–500. [Google Scholar] [CrossRef]
- Rego-Costa, A.; Debarre, F.; Chevin, L.M. Chaos and the (un)predictability of evolution in a changing environment. Evolution 2018, 72, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Skinner, J.E.; Molnar, M.; Vybiral, T.; Mitra, M. Application of chaos theory to biology and medicine. Integr Physiol Behav Sci 1992, 27, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Toker, D.; Sommer, F.T.; D'Esposito, M. A simple method for detecting chaos in nature. Commun Biol 2020, 3, 11. [Google Scholar] [CrossRef]
- Barbieri, S.; Gotta, M. Order from chaos: cellular asymmetries explained with modelling. Trends Cell Biol 2024, 34, 122–135. [Google Scholar] [CrossRef]
- Winzer, P.; Anghel, N.; Imhof, D.; Balmer, V.; Ortega-Mora, L.M.; Ojo, K.K.; Van Voorhis, W.C.; Müller, J.; Hemphill, A. Neospora caninum: Structure and fate of multinucleated complexes induced by the bumped kinase inhibitor BKI-1294. Pathogens 2020, 9. [Google Scholar] [CrossRef]
- Winzer, P.; Müller, J.; Aguado-Martinez, A.; Rahman, M.; Balmer, V.; Manser, V.; Ortega-Mora, L.M.; Ojo, K.K.; Fan, E.; Maly, D.J.; et al. In vitro and In vivo effects of the bumped kinase inhibitor 1294 in the related cyst-forming apicomplexans Toxoplasma gondii and Neospora caninum. Antimicrob Agents Chemother 2015, 59, 6361–6374. [Google Scholar] [CrossRef]
- Mukhopadhyay, D.; Arranz-Solis, D.; Saeij, J.P.J. Toxoplasma GRA15 and GRA24 are important activators of the host innate immune response in the absence of TLR11. PLoS Pathog 2020, 16, e1008586. [Google Scholar] [CrossRef]
- Arranz-Solis, D.; Warschkau, D.; Fabian, B.T.; Seeber, F.; Saeij, J.P.J. Late embryogenesis abundant proteins contribute to the resistance of Toxoplasma gondii oocysts against environmental stresses. mBio 2023, 14, e0286822. [Google Scholar] [CrossRef]
- Hänggeli, K.P.A.; Hemphill, A.; Müller, N.; Schimanski, B.; Olias, P.; Müller, J.; Boubaker, G. Single- and duplex TaqMan-quantitative PCR for determining the copy numbers of integrated selection markers during site-specific mutagenesis in Toxoplasma gondii by CRISPR-Cas9. Plos One 2022, 17. [Google Scholar] [CrossRef]
- Vermathen, M.; Müller, J.; Furrer, J.; Müller, N.; Vermathen, P. 1H HR-MAS NMR spectroscopy to study the metabolome of the protozoan parasite Giardia lamblia. Talanta 2018, 188, 429–441. [Google Scholar] [CrossRef]
- Müller, J.; Boller, T.; Wiemken, A. Pools of non-structural carbohydrates in soybean root nodules during water stress. Physiologia Plantarum 1996, 98, 723–730. [Google Scholar] [CrossRef]
- Semeraro, M.; Boubaker, G.; Scaccaglia, M.; Müller, J.; Vigneswaran, A.; Hanggeli, K.P.A.; Amdouni, Y.; Kramer, L.H.; Vismarra, A.; Genchi, M.; et al. Transient adaptation of Toxoplasma gondii to exposure by thiosemicarbazone drugs that target ribosomal proteins is associated with the upregulated expression of tachyzoite transmembrane proteins and transporters. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Alvarez-Jarreta, J.; Amos, B.; Aurrecoechea, C.; Bah, S.; Barba, M.; Barreto, A.; Basenko, E.Y.; Belnap, R.; Blevins, A.; Bohme, U.; et al. VEuPathDB: the eukaryotic pathogen, vector and host bioinformatics resource center in 2023. Nucleic Acids Res 2024, 52, D808–D816. [Google Scholar] [CrossRef] [PubMed]
- UniProt, C. UniProt: a worldwide hub of protein knowledge. Nucleic Acids Res 2019, 47, D506–D515. [Google Scholar] [CrossRef]
- Pham, T.V.; Henneman, A.A.; Jimenez, C.R. iq: an R package to estimate relative protein abundances from ion quantification in DIA-MS-based proteomics. Bioinformatics 2020, 36, 2611–2613. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.C.; Gorenstein, M.V.; Li, G.Z.; Vissers, J.P.; Geromanos, S.J. Absolute quantification of proteins by LCMSE: a virtue of parallel MS acquisition. Mol Cell Proteomics 2006, 5, 144–156. [Google Scholar] [CrossRef]
- Huber, W.; von Heydebreck, A.; Sultmann, H.; Poustka, A.; Vingron, M. Variance stabilization applied to microarray data calibration and to the quantification of differential expression. Bioinformatics 2002, 18 Suppl 1, S96–104. [Google Scholar] [CrossRef]
- Silver, J.D.; Ritchie, M.E.; Smyth, G.K. Microarray background correction: maximum likelihood estimation for the normal-exponential convolution. Biostatistics 2009, 10, 352–363. [Google Scholar] [CrossRef]
- Kammers, K.; Cole, R.N.; Tiengwe, C.; Ruczinski, I. Detecting significant changes in protein abundance. EuPA Open Proteom 2015, 7, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate - a practical and powerful approach to multiple testing. Journal of the Royal Statistical Society Series B-Statistical Methodology 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Uldry, A.C.; Maciel-Dominguez, A.; Jornod, M.; Buchs, N.; Braga-Lagache, S.; Brodard, J.; Jankovic, J.; Bonadies, N.; Heller, M. Effect of sample transportation on the proteome of human circulating blood extracellular vesicles. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
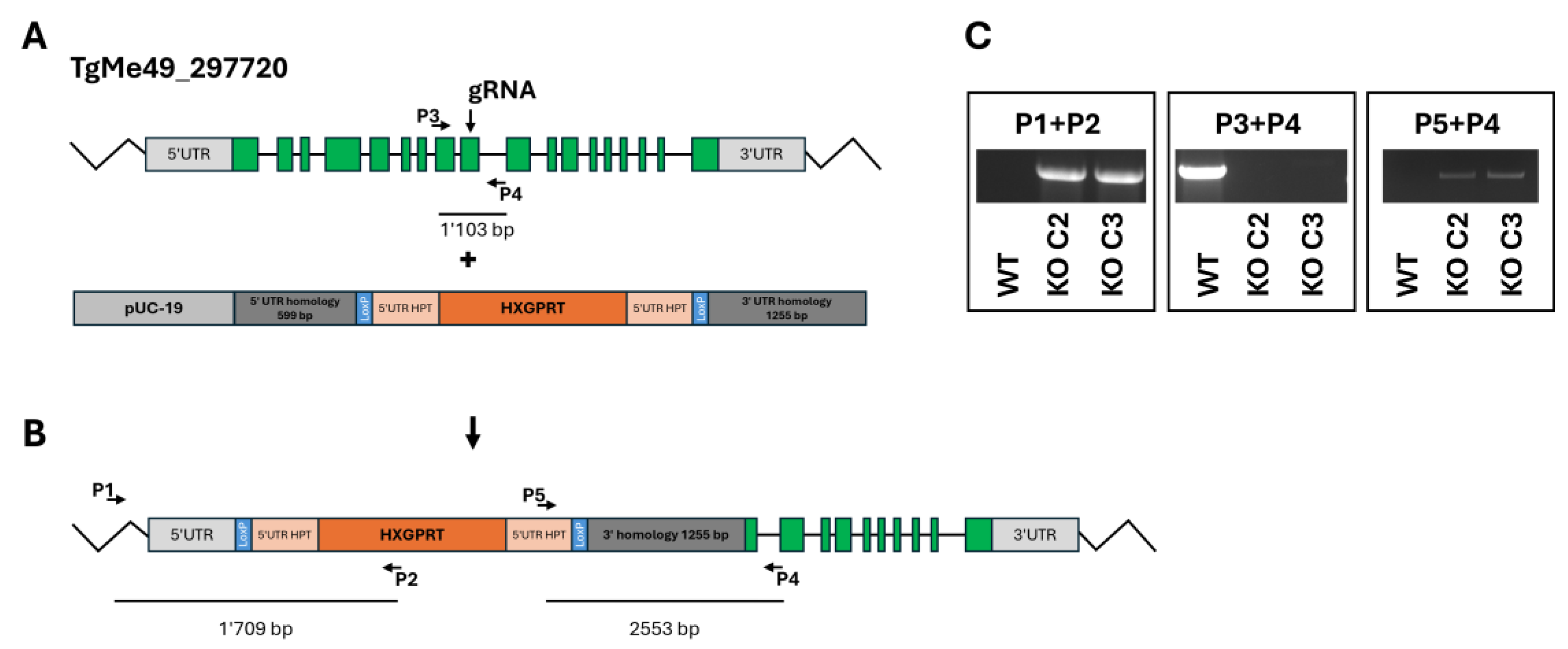
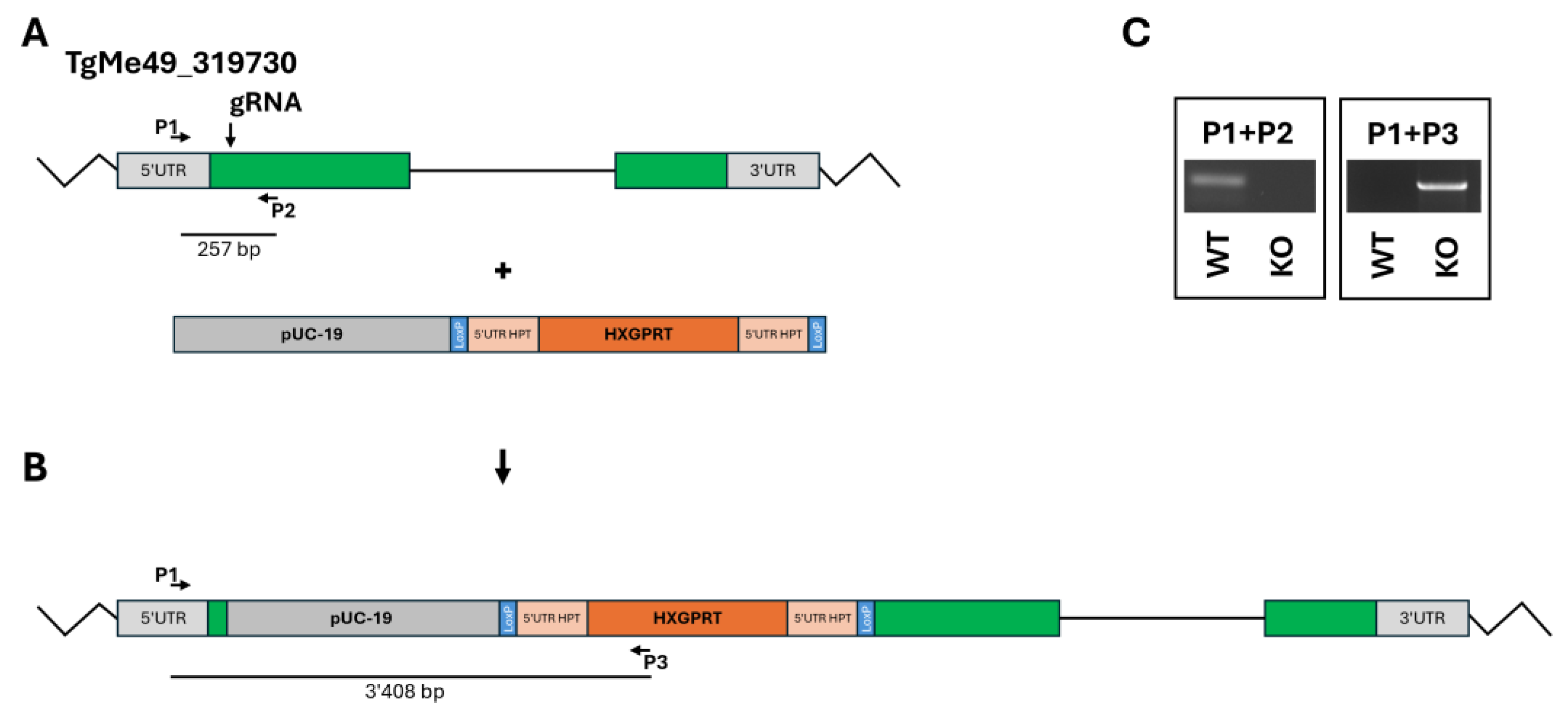
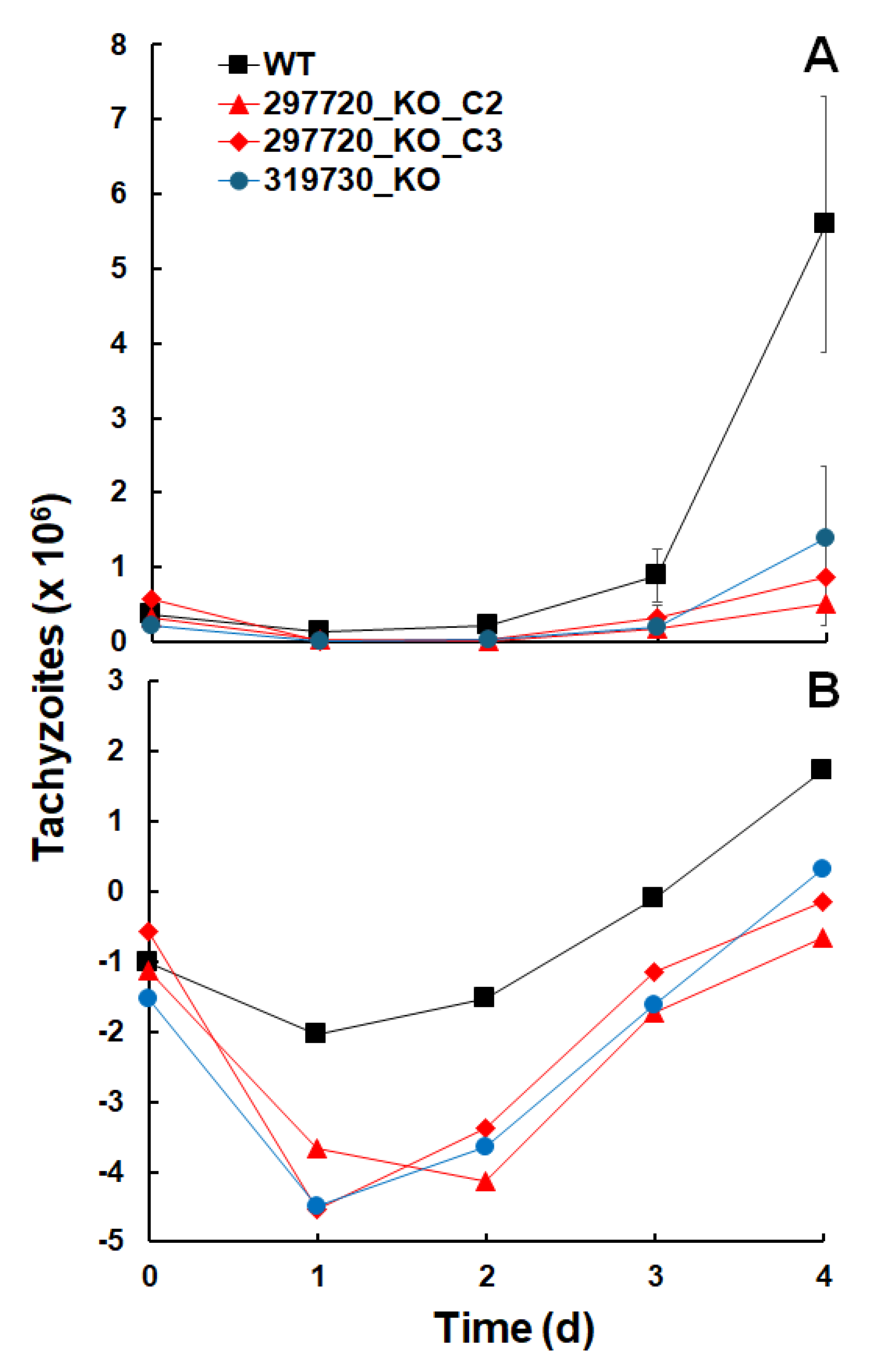
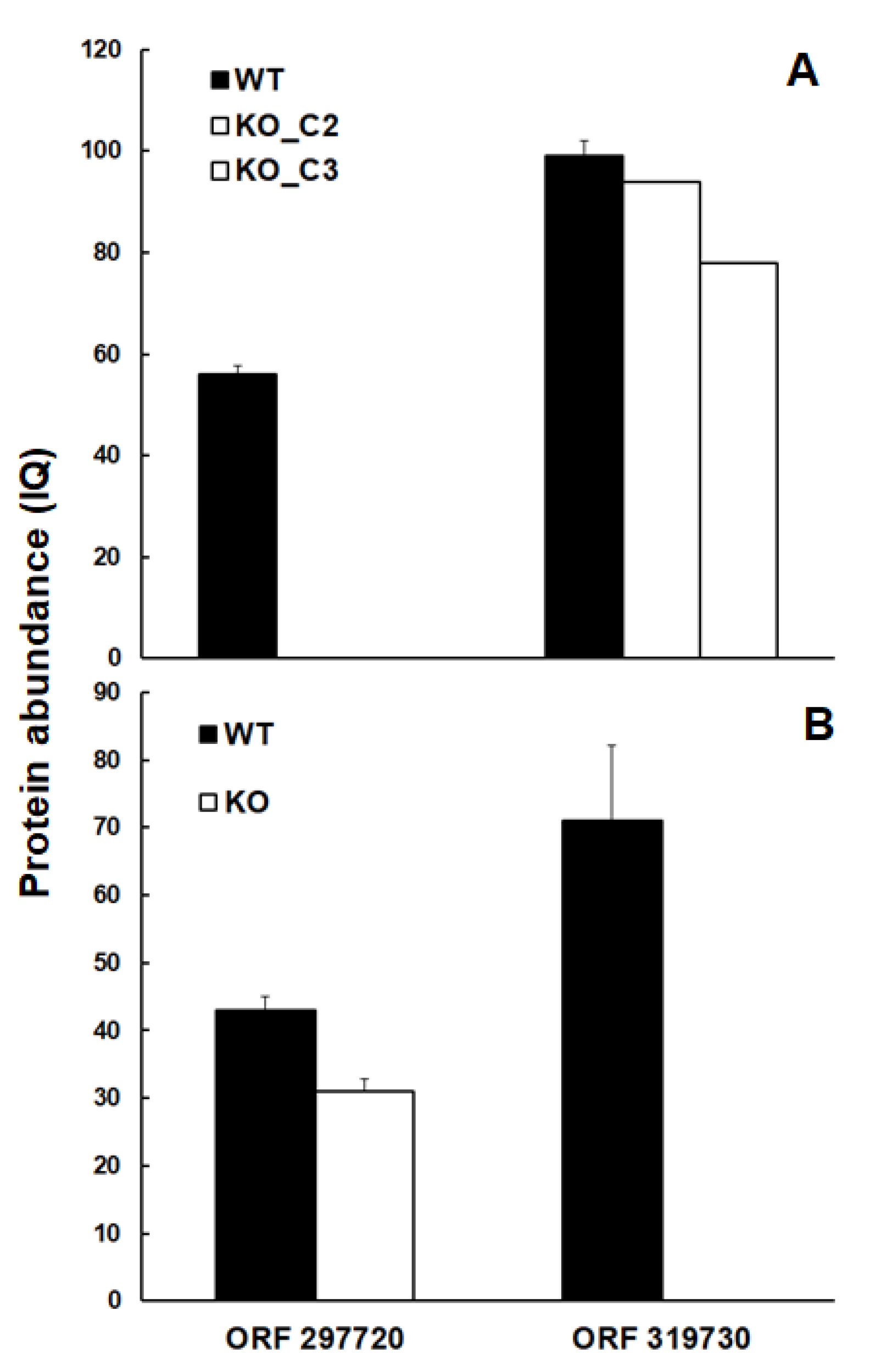
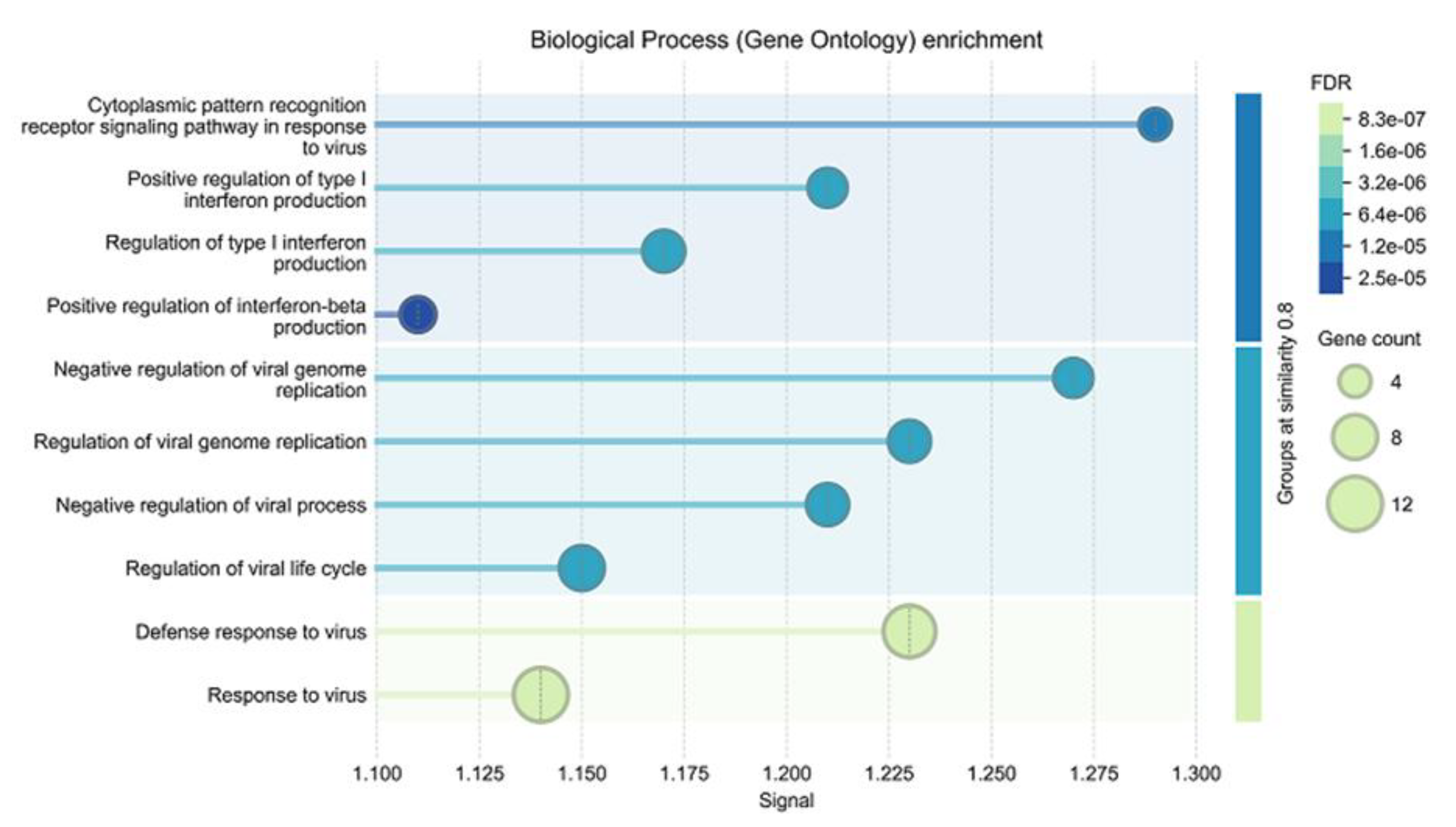

| Strain | Logarithmic growth rate (d-1) | Doubling time (d) |
|---|---|---|
| T. gondii Sp1 wildtype | 1.63 | 0.43 |
| 297720_KO_C2 | 1.73 | 0.40 |
| 297720_KO_C3 | 1.62 | 0.43 |
| 319730_KO | 1.99 | 0.35 |
| Parameter | TGME49_297720 KO vs WT | TGME49_319730 KO vs WT |
|---|---|---|
| Unique T. gondii peptides | 113213 | 60863 |
| Non-redundant T. gondii proteins | 5367 | 4185 |
| Datasets | Table S1 | Table S2 |
| Common downregulated proteins | Common upregulated proteins | ||
|---|---|---|---|
| GN | Description | GN | Description |
| TGME49_200250 | Microneme protein MIC17A | TGME49_224770 | SAG-related sequence SRS40D |
| TGME49_203720 | Vitamin k epoxide reductase family protein | TGME49_273980 | Hypothetical protein |
| TGME49_207210 | Hypothetical protein | TGME49_277230 | Hypothetical protein |
| TGME49_209985 | cAMP-dependent protein kinase | TGME49_292260 | SAG-related sequence SRS36B |
| TGME49_213480 | Hypothetical protein | TGME49_320250 | SAG-related sequence SRS15A |
| TGME49_243680 | Dihydrodipicolinate reductase | ||
| TGME49_280570 | SAG-related sequence SRS35A | ||
| TGME49_291040 | Lactate dehydrogenase LDH2 | ||
| TGME49_308093 | Rhoptry kinase family protein (incomplete catalytic triad) | ||
| TGME49_309930 | Melibiase subfamily protein GRA65 | ||
| TGME49_318880 | Hypothetical protein | ||
| TGME49_319090 | IgA-specific serine endopeptidase | ||
| Protein family or function | TGME49_297720 | TGME49_319730 | ||
|---|---|---|---|---|
| KO down | KO up | KO down | KO up | |
| Metabolism or transport | 8 | 0 | 5 | 6 |
| Signaling | 3 | 1 | 0 | 3 |
| Protein binding or modification | 4 | 1 | 3 | 0 |
| Nucleic acid binding or modification | 6 | 3 | 6 | 3 |
| BAG or MAG | 1 | 0 | 1 | 0 |
| GRA | 0 | 1 | 1 | 2 |
| MIC | 3 | 0 | 1 | 0 |
| ROP | 2 | 0 | 0 | 1 |
| SAG | 4 | 5 | 2 | 6 |
| Unknown | 8 | 10 | 7 | 13 |
| Sum | 39 | 21 | 26 | 34 |
| GN | Description | Metabolic step |
|---|---|---|
| TGME49_226910 | Amylo-alpha-1,6-glucosidase | Starch degradation |
| TGME49_231920 | Oxidoreductase, short chain dehydrogenase/reductase family protein | Unknown redox process |
| TGME49_238200 | Alpha/beta hydrolase fold domain-containing protein | Unknown hydrolytic process |
| TGME49_239310 | Ribulose 5-phosphate isomerase | Pentose phosphate shunt |
| TGME49_257750 | Homocysteine s-methyltransferase domain-containing protein | Methionine biosynthesis |
| TGME49_297720 | Trehalose-phosphatase (the intended KO) | Trehalose biosynthesis |
| TGME49_313050 | Oxidoreductase, short chain dehydrogenase/reductase family protein | Unknown redox process |
| TGME49_320630 | Phosphotransferase enzyme family protein | Activation of alcohol group |
| GN | Description | Metabolic step |
|---|---|---|
| Downregulated in KO | ||
| TGME49_215490 | Novel putative transporter NPT1 | Transmembrane transport |
| TGME49_222160 | Aldehyde dehydrogenase | Cytosolic redox process |
| TGME49_266640 | Acetyl-coenzyme A synthetase 2, putative | Acetyl-CoA biosynthesis |
| TGME49_268860 | Enolase 1 | Glycolysis in bradyzoites |
| TGME49_301210 | NAD(P) transhydrogenase subunit beta, putative | NADH-NADPH interconversion |
| Upregulated in KO | ||
| TGME49_219230 | AMP-binding enzyme domain-containing protein | Lipid metabolism |
| TGME49_227100 | Glutaredoxin 5 | Fe-S-cluster biosynthesis |
| TGME49_277240 | NTPase I | Nucleotide hydrolysis |
| TGME49_294640 | Ribonucleoside-diphosphate reductase large chain | dNTP biosynthesis |
| TGME49_313950 | Glucose-methanol-choline (GMC) oxidoreductase | Various redox processes |
| TGME49_321570 | beta-hydroxyacyl-acyl carrier protein dehydratase (FABZ) | Lipid metabolism |
| T. gondii | H. sapiens | |
|---|---|---|
| Unique peptides | 156329 | |
| Non-redundant proteins | 2178 | 7755 |
| Data bases | ToxoDB | Uniprot |
| GN | Description | KO strain |
|---|---|---|
| Downregulated in KO | ||
| TGME49_203720 | Vitamin k epoxide reductase family protein | All KO strains |
| TGME49_207210 | Hypothetical protein | All KO strains |
| TGME49_209985 | Rhoptry protein ROP42 | All KO strains |
| TGME49_280570 | SAG-related sequence SRS35A | All KO strains |
| TGME49_291040 | Lactate dehydrogenase LDH2 | All KO strains |
| TGME49_309930 | Dense granule protein GRA56 | All KO strains |
| TGME49_258230 | Rhoptry kinase family protein ROP20 | TGME49_297720 KO clones C2 and C3 |
| TGME49_297720 | Trehalose-6-P phosphatase (the intended KO) | TGME49_297720 KO clones C2 and C3 |
| TGME49_200250 | Microneme protein MIC17A | TGME49_319730 KO clone |
| TGME49_202020 | DnAK-TPR | TGME49_319730 KO clone |
| TGME49_207160 | SAG-related sequence SRS49D | TGME49_319730 KO clone |
| TGME49_207210 | Hypothetical protein | TGME49_319730 KO clone |
| TGME49_209755 | Cyst matrix protein MAG2 | TGME49_319730 KO clone |
| TGME49_216140 | Tetratricopeptide repeat-containing protein ANK1 | TGME49_319730 KO clone |
| TGME49_222160 | Aldehyde dehydrogenase | TGME49_319730 KO clone |
| TGME49_260600 | mRNA-binding protein PUF1 | TGME49_319730 KO clone |
| TGME49_264660 | SAG-related sequence SRS44 | TGME49_319730 KO clone |
| TGME49_308093 | rhoptry kinase family protein (incomplete catalytic triad) | TGME49_319730 KO clone |
| TGME49_319560 | Microneme protein MIC3 | TGME49_319730 KO clone |
| TGME49_319730 | You2 zinc finger protein (the intended KO) | TGME49_319730 KO clone |
| Upregulated in KO | ||
| TGME49_273980 | Dense granule protein GRA80 | All KO clones |
| TGME49_277230 | Dense granule protein GRA82 | All KO clones |
| TGME49_224760 | SAG-related sequence SRS40E | TGME49_297720 KO clones C2 and C3 |
| TGME49_239090 | SAG-related sequence SRS23 | TGME49_297720 KO clones C2 and C3 |
| TGME49_278370 | Toxoplasma gondii family A protein | TGME49_297720 KO clones C2 and C3 |
| TGME49_292260 | SAG-related sequence SRS36B | TGME49_297720 KO clones C2 and C3 |
| TGME49_322010 | Myosin-light-chain kinase | TGME49_319730 KO clone |
| GN | Description | Remarks |
|---|---|---|
| Downregulated in KO | ||
| TGME49_207930 | Phosphatidylethanolamine-binding protein | Dense granule protein, expression in bradyzoites and oocysts |
| TGME49_232780 | Basal complex component BCC1 | Phosphorylated, also present in oocysts |
| TGME49_258230 | Rhoptry kinase family protein ROP20 | DE also in isolated tachyzoites |
| TGME49_268860 | Enolase 1 | Bradyzoite marker |
| TGME49_297720 | Trehalose-phosphatase | Intended KO |
| TGME49_313780 | Hypothetical protein | Present in tachyzoite conoid proteome |
| Upregulated in KO | ||
| TGME49_224760 | SAG-related sequence SRS40E | DE also in isolated tachyzoites |
| TGME49_230180 | Dense granule protein GRA24 | Virulence marker |
| TGME49_239090 | SAG-related sequence SRS23 | DE also in isolated tachyzoites |
| TGME49_265870 | Pantoate-beta-alanine ligase | Nucleus located, phosphorylated |
| TGME49_278370 | Toxoplasma gondii family A protein | DE also in isolated tachyzoites |
| TGME49_292260 | SAG-related sequence SRS36B | DE also in isolated tachyzoites |
| TGME49_308950 | Histidine acid phosphatase superfamily protein | Dense granule protein |
| Name | Sequence |
|---|---|
| TGME49_297720 KO | |
| gRNA_TgME49_297720_fwd | AAGTTGAATCATCAACATGCCTGTGGG |
| gRNA_TgME49_297720_rev | AAAACCCACAGGCATGTTGATGATTCA |
| TgME49_297720_P1 | CCAGTGACGACGAGTGTTGA |
| TgME49_297720_P2 | CAACTCCTCGCCGAAGTAAG |
| TgME49_297720_P3 | TCTCGACTTGTCCATCCCCT |
| TgME49_297720_P4 | GAAGTGGGCGTTGTTTACCG |
| TgME49_297720_P5 | ACAGTCTCACCTCGCCTTGT |
| TGME49_319730 KO | |
| gRNA_TgME49_319730_fwd | AAGTTGCGGTACCCTTCGCTGGAGGAG |
| gRNA_TgME49_319730_rev | AAAACTCCTCCAGCGAAGGGTACCGCA |
| TgME49_319730_P1 | TTTTCACCTTCGGTCTCTCG |
| TgME49_319730_P2 | CGTTCGAGTGCTGTGTGTCT |
| TgME49_319730_P3 | CAACTCCTCGCCGAAGTAAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



