Submitted:
01 October 2025
Posted:
02 October 2025
You are already at the latest version
Abstract
Number processing confers significant adaptive advantages, enabling organisms to navigate their environment and maximize survival outcomes. Extensive evidence across species demonstrates that numerical cognition is a ubiquitous cognitive trait that enhances fitness by supporting critical behaviors such as foraging, hunting, and intergroup conflict. In this review, we examine the evolutionary and developmental roots of numerical cognition, focusing on its functional role in social contexts. First, we report research findings on the use of numerical information in various social behaviors among a wide range of animals. Then, we discuss how selective pressures arising from social complexity, like group-size, hierarchical structure, and cooperation, contribute to the refinement of numerical abilities during social interactions. Finally, we explore how early social deprivation during ontogeny may shape the development of numerical cognition. We present a novel and integrative perspective on the evolutionary and developmental link between numerical skills and social life.
Keywords:
numerical cognition
; number sense
; social intelligence hypothesis
; social cognition
1. Numerical Cognition in Animals
Since the beginning of the 20th century, animal numerical competences have been vastly documented, and in the last decades, the interest in cross-species numerical cognition has constantly increased [1,2,3]. The first inquiry into numerical abilities in animals was quite unfortunate, marked by the famous case of Clever Hans. Hans was a horse that was trained by Wilhelm von Osten in the early 1900s to solve complex arithmetic calculations by tapping the results with his hoof. Initially believed by most of the scientific community, it was later revealed that Hans was not actually solving mathematical tasks through counting. Instead, it was a classic example of the Observer effect: the horse was detecting and interpreting unintentional behavioral cues from its experimenter. Hans would continue tapping until he detected these cues, which indicated when he had reached the correct answer [4].
Only in the 1930s, a systematic investigation of numerical abilities in animals started. The pioneer was Otto Koehler, who studied numerical cognition in several birds such as jackdaw (Corvus monedula), budgerigars (Melopsittacus undulates), and pigeons (Columba livia) [5]. A few years later, Jacky Emmerton and colleagues (1997; 1998) consolidated Koehler’s findings and documented pigeons’ ability to discriminate relatively smaller quantities (1,2,3) from larger ones (4,5,6,7) [6,7]. As research in this area progressed, evidence of numerical abilities emerged across diverse species, including rats (Rattus norvegicus) [8], macaques (Macaca mulatta) [9], and domestic chicks (Gallus gallus) [10].
Investigation of numerical abilities in animals and newborn humans has revealed the presence of a Number sense, a system for numerosity estimation [11,12]. This system emerges in the first days of human life. Being pre-symbolic and pre-verbal, Number sense is considered as the foundations of more complex numerical reasoning and symbolic mathematic acquisition [13,14,15].
The phylogenetic ubiquity [16,17,18,19,20,21,22] and precocious availability [23,24,25,26,27,28] of the Number sense have led researchers to consider it as one of the core systems present at birth, which contribute to representing the most relevant environmental aspects such as objects, agents, spatial relationships, social partners, and numerosity [29,30,31]. Number sense involves two subsystems: the Object File System (OFS) and the Analogue Magnitude System (AMS). OFS supports the ability to rapidly and accurately process up to three/four elements, by identifying each object based on its peculiar features (e.g., color, shape), and opening a file for each object in the working memory [32]. Instead, AMS enables estimations of numerosity beyond four as well as other magnitudes such as length, duration, weight [33]. Remarkably, AMS follows the Weber’s law [34,35], according to which, the discriminability between two magnitudes becomes increasingly less accurate and slower as the ratio approaches 1: in fact the ratio effect reflects a combination of the numerical distance effect [34,36,37] and the numerical size effect [38].
This fundamental cognitive ability is rooted in specialized neural circuits that have been increasingly mapped across species [12,39], with number processing engaging different brain regions [40]. Although the location and mechanisms of numerical skills are best understood in the human and primate brain, neurophysiological evidence in vertebrates, such as mammals, birds and fishes, have demonstrated that the pallial regions of the endbrain are involved in number representations [41,42]. The discovery of number neurons in the brain of different animal species has shed light on the neurobiological basis of the Number sense [43,44,45,46,47,48]. Intriguingly, these neurons are localized in homologous structures across species: the nidopallium caudolaterale in birds and the prefrontal cortex in primates [49].
This shared neurobiological foundation underlies the Number sense, and it encompasses various cognitive abilities [50], which serve as precursors for the subsequent development of higher numerical skills such as quantity and proto-numerical discrimination, proportional reasoning, and ordinal processing [51,52].
Quantity discrimination is the ability to differentiate between two sets of items to determine which is "more" or "less" numerous [31]. Quantity discrimination has been documented in different species and taxa, including primates [53,54,55], mammals [56,57,58], birds [59,60,61,62], fish [63,64,65,66,67], amphibians [68,69,70], reptiles [71,72,73,74], and even invertebrates [75,76,77,78,79,80].
Foraging behavior provides a classic context for observing quantitative discrimination. The seminal study by Krebs and colleagues (1974) demonstrated that various species preferentially select numerically advantageous food sources, consistent with the predictions of the Optimal Foraging Theory [81]. According to it, animals maximize energetic gain by choosing food patches that provide the greatest rewards relative to the time and energy expended [81]. This would lead animals to choose patches with more abundant food items. However, it is important to note that foraging could be influenced by non-numerical factors, such as food size. Therefore, animals may sometimes prioritize larger items over more numerous ones. Indeed, changes in numerosity (the number of items in a set) often correlate with changes in continuous variables like volume, surface area, density, and contour length [82]. To determine if animals truly perceive discrete numerosity, researchers carefully controlled continuous variables in laboratory settings. The discovery that animals are capable of discriminating between sets based solely on numerosity, indicates proto-numerical rather than just quantity discrimination abilities [47,83,84,85,86].
Effective foraging in competitive environments critically depends on the ability to understand and use proportional reasoning. In a pivotal experiment, free-living mallards were offered different numbers of pieces of bread thrown at different rates and on opposite sides of a lake. Birds divided themselves between the resource patches as if they were simultaneously considering the amount of food and the number of animals feeding at each site [87]. The ability to use proportional reasoning is present across diverse taxa, including birds [88,89], fish [90], and primates [91,92,93].
Discriminating between two sets is not only used to maximize food intake, but it also provides other fitness advantages. Numerical abilities are equally important in reproductive strategies, such as parental care and mate selection. Numerical assessment in reproductive contexts extends across various species shaping behaviours and strategies. For instance, male mosquitofish (Gambusia holbrooki) demonstrate a preference for groups with more females, likely to increase their mating opportunities [94]. In fish populations, males often adjust their reproductive tactics, such as sperm allocation, based on the number of males and females [95]. Similarly, male mealworm beetles (Tenebrio molitor) modulate mate guarding duration based on the number of rivals encountered to mitigate sperm competition risk and optimize reproductive success [96,97]. Numerical abilities can also enhance offspring survival, as seen in American coots (Fulica americana), which reduce brood parasitism by keeping track of the number of eggs they lay and rejecting extras introduced by parasitic individuals [98].
This capacity to track items and process numerical information may have evolved particularly in social species such as primates, cetaceans, corvids, and parrots [60]. Indeed, within social environments, this capacity is likely to provide a key adaptive advantage during competition between groups and predation, as will be explored in the section Social Dynamics Require Numbers.
Another fundamental aspect of number processing is ordinality. It encompasses two distinct cognitive abilities: a) the processing of an item's position within a series, and b) the understanding of relative magnitude, where adding an item to a set makes it larger than the previous set but smaller than the following one [31].
The processing of an item's position within a series has been demonstrated in various animal species, including rats [22,99], domestic chicks [100,101], nutcrackers [102], zebrafish [103], and honeybees [104]. Such ability enables animals to better orient themselves and navigate their environments, counting landmarks as reference points [104].
Instead, the understanding of relative magnitude has so far been documented in macaques [16], and pigeons [105].
These findings suggest that non-human animals can process ordinal relationships abstractly and infer hierarchical numerical rules. The ability to organize stimuli according to their numerical value, creating a sequential and linear arrangement where each element precedes the one that follows, is a key cognitive skill underlying the process of transitive inference [106]. Transitive inference is a deductive process through which individuals infer a relationship between two non-adjacent items based on their known relationships with a third item (e.g., if A > B and B > C, then A > C). This process relies on an internal representation of the entire sequence that shares mechanisms with the number sense, as evidenced by Weber’s law [16,107].
Relating objects or individuals is a fundamental building block for establishing hierarchical relationships within a social group [108].
Indeed, enumerating conspecifics and tracking dominance relations between individuals relies on core numerical mechanisms. These mechanisms offer clear adaptive advantages including foraging, navigation, predator avoidance, hunting, and mating [109,110] (see Table 1). Are these numerical abilities merely useful for individual fitness or specifically shaped and required by the complexities of social living?
2. Social Dynamics Requires Numbers
Within a broad adaptive landscape, social life presents a particularly complex set of challenges that leverage and may have driven the refinement of numerical processing. Numerical cognition thus plays a key role in group dynamics and constitutes a foundation for ecological success in many social species. Here, three key numerical processes used in social dynamics will be examined (Error! Reference source not found.). First, the capacity to estimate the number of conspecifics in a group is vital for assessing competitive advantage, optimizing hunting efficiency, and minimizing predation risk. Second, representing social hierarchies is essential for minimizing aggressive encounters. Finally, collective decision-making often relies on numerical thresholds, which enable efficient and cohesive group choices. Numerical processing advantages and their possible evolutionary pressure in social contexts are summarized in Figure 1.
2.1. Counting the Group Members
A fundamental ability observed in most social species is the capacity to estimate the number of conspecifics in a group. For group-living species, the adaptive tuning of group size is critical, as it often affects fitness outcomes [137,138,139]. Social species frequently rely on proto-numerical discrimination to assess competitive advantage during intergroup conflicts, determining whether a rival group is larger or smaller, and playing a pivotal role in territory defense [111,140]. For example, in the seminal study of McComb and colleagues (1994), female lions (Panthera leo) were less likely to engage in approach behavior when confronted with the simulated roars of three unknown intruders compared to a single intruder. The lionesses’ behavior, either alone or in a group, was strongly influenced by the ratio of defenders to intruders, underscoring the importance of numerical odds in social conflict [111]. Similar numerical assessment during intergroup conflict was reported in wild spotted hyenas (Crocuta crocuta) [112], chimpanzees (Pan troglodytes) [113], black howler monkey (Alouatta pigra) [114], subdesert mesite (Monias benschi) [115] and in free-ranging dogs (Canis lupus familiaris) [116]. This widespread behavior aligns with Game Theory predictions, where assessing relative numbers and strength before engaging in conflict is essential to maximizing the payoff of engagement, as larger groups typically prevail [141].
This numerical skill is also particularly crucial for social carnivorans in making hunting decisions. Proto-numerical assessment allows them to select foraging patches with greater prey availability or less dangerous prey and coordinate cooperative hunting efforts [110]. Studies on carnivores have shown that numerical assessments are often prioritized over other factors during decision-making processes [140]. Specifically, during hunting, numerical discrimination informs decisions on which foraging patches to exploit or which prey to target based on size, choices critically dependent on the size of the hunting party itself [142].
Crucially, estimating group numbers confers advantages not only for predators in hunting contexts but also to prey in defensive ones. The formation of social groups is a common evolutionary response to predation risk, providing individuals with several adaptive benefits [143]. The mechanisms that enhance safety of individuals in groups include the confusion effect, which makes it harder for a predator to single out and attack an individual prey animal; the many-eyes effect, which increases the likelihood of detecting predators; and the dilution effect, wherein an individual’s risk of predation decreases as the number of group members increases [144,145,146].
Many fish utilize their ability to choose larger, safer social groups when navigating unfamiliar or potentially dangerous environments [109,147]. For example, guppies (Poecilia reticulata) and three-spined stickleback (Gasterosteus aculeatus) can distinguish between groups of different sizes, with guppies discriminating between 4 and 5 individuals [118] and sticklebacks between 6 and 7 [119]. Moreover, mosquitofish show ratio-dependent discrimination, successfully discriminating 4 vs. 8 and 4 vs. 10, but not 4 vs. 6 or 4 vs. 7 [63]. This proto-numerical ability enables prey species to reduce their predation risk by preferentially joining larger groups.
The same capability of domestic chicks to spontaneously approach the larger group of conspecifics [135,148,149] has also been used to test proto-arithmetic abilities, namely the capacity to operate on non-symbolic numerical representations through basic addition and subtraction [31]. For example, in one study, newborn chicks reared with five objects (social companions) were able to track and compare group sizes when the objects disappeared one by one behind screens (e.g., 1+1 vs. 1+1+1). This ability persisted even under more complex conditions involving both addition and subtraction (e.g., 1+2 vs. 4–2), suggesting a spontaneous capacity to form and manipulate mental representations of quantity [135].
These findings highlight how basic social dynamics benefit from numerical abilities, suggesting that proto-numerical discrimination is tightly linked to social demands. Across taxa, predation pressure and the challenges of group living have acted as strong selective forces shaping the evolution of numerical cognition as a tool for survival.
2.2. Ordering the Group
Animals living in social groups must not only count potential allies, rivals, or prey, but also navigate complex social hierarchies. To achieve this, they must recognize individuals, track social relationships, and assess their own status within the hierarchical order of the group. Critically, this allows individuals to minimize aggressive encounters with higher-ranking conspecifics, who typically pose greater physical threats. Such skills are essential for avoiding direct confrontations that could result in injury or even death [150].
Using playback experiments, Bergman and colleagues (2003) demonstrated that baboons not only recognize the rank orders of various group members but also understand which matrilineal kin group each individual belongs to. This indicates that primates organize social concepts hierarchically, and highlights their sophisticated cognitive abilities [108]. Hierarchy is not limited to primates. Ravens (Corvus corax), for instance, provide compelling evidence of hierarchical understanding [122]. Remarkably, ravens exhibit such responses not only for members of their own group but also for those of adjacent groups [150,151,152]. Similar findings have been observed in geese (Anser anser) [123], and domestic chickens (Gallus gallus) [124,125], suggesting that this ability is a fundamental cognitive building block in species with dominance hierarchies [153,154].
Another example comes from the study by Daisley and colleagues (2021) in which transitive inference reasoning was studied in relation to chicks’ social rank and sex. The results showed that females outperformed males in this task. Moreover, females’ performance decreased linearly from the lowest to highest ranks, while in males, middle-ranked individuals had the best performance [124]. These findings have been attributed to sex-specific social behaviors: female chickens typically form a rigid linear hierarchy, where low-ranking individuals closely monitor dominance relationships to avoid conflict. On the other hand, a few males control territories, with males of intermediate rank competing more for mating opportunities [155].
Crucially, invertebrates also exhibit transitive inference. Paper wasps (Polistes dominula and Polistes metricus) despite miniature brains, could organize color pairs into linear hierarchies and infer novel relationships. Notably, both species performed equally well despite P. dominula's live in more complex social groups compared to P. metricus' smaller groups [126]. This shared ability despite differing social complexity suggests that the cognitive processing of dominance hierarchies may be evolutionarily conserved, potentially linked to ancestral dominance behaviors rather than driven by current socio-ecological demands [126]. This contrasts sharply with honeybees (Apis mellifera), which fail transitive inference tests [156], likely reflecting the lack of ecological relevance of this cognitive ability in their non-hierarchical societies where social roles are fixed.
In conclusion, the ability to infer and predict social relationships is crucial for navigating the complex dynamics of group life, increasing the chances of survival by optimizing conflict resolution and reproductive success.
2.3. Group Decision
Group-living animals, beyond static assessment of companions, or the ranking order, rely on numbers for crucial dynamic and coordinated actions. Numerical processing has a key role in group decision making, which often relies on collective mechanisms. Among these, quorum sensing is a key process by which behavioral responses are triggered once a threshold number of individuals perform a key action [129]. To achieve this, individuals must correctly perceive and estimate the number of conspecifics engaged in the specific action. Crucially, such group-level responses often rely on relative proportions rather than absolute counts, consistent with Weber's Law, which states that sensitivity to differences between numerical sets depends on their ratio rather than absolute difference [129].
Compelling examples come from white-faced capuchin monkeys (Cebus capucinus), where adult individuals use specific vocalizations (trills) to initiate or redirect group movement. The group typically responds and moves in the direction indicated by the trilling individual only after a sufficient number of group members appear to support the initiative, either by position or vocal reply [130]. Similar evidence from wild olive baboons (Papio anubis) revealed that collective movement is governed by shared, majority-based decision rules [131]. The likelihood of a baboon joining a group increases in an S-shaped curve as one subgroup gains a numerical advantage over another. However, baboons are only willing to compromise and follow a different direction when there's minimal disagreement between subgroups about which way to go.
Similar mechanisms for group decision are present in other highly social species such as African wild dogs (Lycaon pictus) [132], and meerkats (Suricata suricatta) [133]. This process enables individuals to choose among mutually exclusive alternatives while maintaining group cohesion [127].
In social insects, quorum decisions are vital for processes such as selecting a new nest site. For instance, in the Japanese ant (Myrmecina nipponica), the quorum thresholds for nest selection increase with colony size [127]. Ants employ an analogue magnitude mechanism to assess group size, so their accuracy diminishes with increasing stimulus magnitude due to limitations consistent with Weber’s Law [127]. This ratio-dependent mechanism implies that ants’ numerical discrimination abilities decrease as the size of the group grows, affecting their capacity for optimal collective actions like resource allocation during colony fission or coordinating foraging efforts.
Honeybees (Apis mellifera) also demonstrate quorum sensing in their decision-making processes. For example, they initiate swarming when 10–15 scout bees gather at a single location [134]. During house-hunting, both bees and ants (Temnothorax rugatulus) rely on quorum processes to select among potential nest sites [128,134].
The studies of quorum sensing in various species highlight the benefits of numerical competence in collective decision making. Numerical discrimination and ordinal processing are fundamental cognitive abilities with profound socio-ecological significance. As research continues to uncover the cognitive mechanisms underlying these abilities, it becomes increasingly clear that numerical competence is a critical component of animal social behavior, shaping interactions and outcomes across a wide array of species.
3. Evolutionary Pressure
The diverse scenarios outlined above highlight how numerical abilities contribute directly to social behaviors. Indeed, environmental and social demands act as potent evolutionary pressures in shaping cognitive traits. Numerical cognition may have emerged repeatedly across evolution suggesting possible evolutionary convergence in phylogenetically distant taxa, highlighting its adaptive value [157]. In the present section, we expand this perspective by examining both ecological and social pressures that drive the evolution of cognition across species.
3.1. The Adaptive Value of Cognition
One of the most prominent hypotheses about selective pressure for cognitive development is the Ecological Intelligence Hypothesis (EIH) [158,159]. According to it, the drivers of cognitive evolution are ecological challenges, such as unpredictability of resource availability, habitat complexity, and predation risks [160,161,162,163,164]. Challenges such as parasitism [165], predation [166,167], habitat complexity [168], foraging behavior [169,170] may have been selective pressures for variation in cognitive abilities, such as numerical processing. For example, in the case of extractive foraging aimed at obtaining inaccessible, hidden or protected foods, primates may have evolved higher cognition and intelligent tool use [159,171]. Likewise, the seasonal nature of many primate diets, such as frugivores, requires cognitive mapping, which is a representative expression of an individual's knowledge about the spatial and environmental relations of geographic space [172]. Indeed, according to the EIH, memorizing the spatial and temporal availability of food could have been one of the pressures for the development of higher cognitive skills [172,173]. Analogous to observations in primates, compelling evidence exists for similar cognitive adaptations in birds. For example, scrub jays (Aphelocoma californica) and Clark's nutcrackers (Nucifraga columbiana) hide food to ensure survival during periods of food scarcity, demonstrating a complex capacity for memory and planning [174,175].
EIH suggests that ecological challenges were the drivers of larger relative brain size, since activities such as food acquisition require spatiotemporal skills and/or manipulative skills, as mainly documented in primates [176,177,178,179]. Notably, corvids, compared to other avian groups, possess larger relative brain sizes and are known for their use of extractive foraging techniques that resemble primates’ techniques, such as the use of sticks to catch insects hiding in crevices [180].
Crucially, in primates, specialized diets may directly select for numerical cognition. Black-handed spider monkeys (Ateles geoffroyi) are highly frugivorous primates like chimpanzees and orangutans. Their need to optimize foraging in patchy, ephemeral fruit distributions likely favors quantitative assessments of food patches [181]. Their excelling in complex food ratios discrimination (e.g., 4 vs. 5 items), aligns with the EIH predictions.
Notably, spider monkeys also share a key social trait with great apes: fission-fusion dynamics, suggesting that ecological pressures may synergize with social complexity to drive cognitive enhancements. Their proficiency in abstract numerosity tasks (e.g., discrimination of dot arrays on the basis of their numerosity when continues variables are controlled) further supports that numerical cognition is adaptive not only for ecological decisions but also for tracking fluctuating subgroup composition.
3.2. The Social Intelligence Hypothesis
Along with ecological challenges, social living is also a critical driver for cognitive traits. This is the central argument of the Social Intelligence Hypothesis (SIH) [182,183,184], which states that enhanced cognitive abilities emerge in group-living species, because social living imposes selective pressures that demand advanced cognition (e.g., cooperation and deception) [122,185].
Evidence from several taxa, including birds, fish, rodents, and primates, suggests that individuals who live in larger social groups exhibit better cognitive performance [186]. Existing literature illustrates how social factors can be the cause of intraspecific variation in cognitive performances, highlighting their relevance in influencing cognitive development, which in turns reflects on fitness [186].
Therefore, evidence suggests that an individual's cognitive development is influenced by their social surroundings.
This relationship becomes evident also on the neural level: A large body of research uses measures of brain size or brain regions (e.g., neocortex, amygdala, cerebrum, cerebellum, and telencephalon) as a measurement for cognitive abilities [186,187]. For example, the encephalization quotient (a measure of brain size relative to body mass) varies significantly among species [188] and has been associated with variation in cognitive performance [189]. This is also true for the neocortex. The neocortex is a complex brain structure that controls and regulates higher functions such as spatial reasoning, planning and decision-making. Studies from numerous taxa (primates, bats, carnivorous mammals and cetaceans) found that neocortex size is positively correlated with social group size [190,191,192]. Such a correlation is supposed to stem from the need to remember, track and manage relationships in larger groups [193].
However, other studies seem to contradict the positive correlation between brain and group size [194]. Indeed, the link between sociality and brain size strongly reported in primates is not as clear in other groups, such as ungulates [195], lemurs [196], birds [197,198,199], corvids [200]. The use of neuroanatomical measures as a proxy for cognition, is still a topic of debate given both the negative [201] as well as positive findings [202], and also the fact that different taxonomic groups show similar cognitive abilities despite very different brain structures [203].
Diverging from the use of brain size as a proxy for cognition, the study of MacLean and colleagues (2013) focused on the association between group size and cognition across primates [204]. Their findings indicate that group size was a predictor of social cognition, measured as the ability to understand a competitor's visual perspective. Instead, group size did not predict a non-social cognitive measure, i.e. inhibitory control in a food retrieval task. Another question that needs to be addressed when considering the SIH is which cognitive traits are expected to respond to evolutionary changes in social systems. To date we can identify two hypotheses, one in line with the domain-specific hypothesis [205] and the other with the domain-general hypothesis [206]. According to the domain-specific hypothesis, greater social complexity leads to selection for cognitive skills that are specific to social life [207]. Contrarily, based on the domain-general hypothesis, social complexity favors both social and non-social cognitive skills [208]. However, it is important to point out that the relationship between group size and cognition is neither universal nor unidirectional. A recent study tested spontaneous proto-numerical discrimination in wild Western Australian magpies (Gymnorhina tibicen dorsalis). The results showed that individuals from smaller groups exhibited enhanced discrimination accuracy compared to those from larger groups [62]. Two primary explanations account for these findings: a) smaller groups face higher intergroup conflict costs, selecting for better numerical assessment skills; b) fewer group members increase vigilance demands ("many-eyes effects") enhancing foraging-related discrimination [62]. This highlights how group size, along with task-specific demands and ecological context can influence cognitive abilities in different ways, necessitating nuanced interpretations of cognitive adaptations.
3.3. Refining the Proxies of Sociality
It can be argued that, group size is not ideal proxy for social complexity [209]. If it is not group size alone that determines a complex society, it could be necessary to consider the type and quality of relationships among individuals [210,211,212]. Indeed, a good proxy of social complexity could also include mating system, dominance style and breeding strategy [209]. One definition we could adopt is from Freeberg and colleagues (2012), who define complex social systems as “those in which individuals frequently interact in many different contexts with many different individuals, and often repeatedly interact with many of the same individuals over time” [213]. Multiple studies show how different species (primates and corvids) living in fission-fusion society, i.e., complex societies where there is frequently splitting and merging in subgroups of variable composition, outperform those that live in more stable groups in cognitive tests used to assess different aspects of impulse control and behavioral flexibility [214,215].
Even though neuroanatomical measures may not be a good proxy for cognition, it is still worth mentioning that a larger brain does not only characterize larger groups, but also groups with low within-group kinship. These groups are distinct by reproductive suppression of adult females, alloparental provisioning and rare reproductive division of labor between group members. Larger brains have also been found in groups where individuals need to make strategic decisions to manage conflicts, specifically Machiavellian strategies of social manipulation and deception; such as leading animals towards or away from places; deflecting aggression onto innocent third parties; concealing the glances that would reveal a hidden food, or concealing the food itself in the hand or under the body [173,205,216]. Indeed, the ‘Machiavellian intelligence’ or ‘social brain’ hypothesis, proposes that large brains are an adaptation to the social complexity produced by living in long-term social groups. Social tactics that seem sophisticated, such as deception and cooperation, may be built on quite simple cognitive mechanisms: the main capacity that is needed is efficient memory, to differentiate individuals and keep track of the webs of kinship, dominance, and mutual favor-trading that make up social life [217]. Actually, it has been suggested that the most challenging societies are those where individuals live in multi-generational units, members are able to recognize each other, there is cooperation and/or competition for resources and the majority of learning happens during social development [218].
Even though results in numerous species, such as spotted hyenas [219], are very promising and once again sustain the SIH, at the moment it is still necessary to further compare cognitive abilities among more and less complex social systems, to affirm that evolution of brain and behaviors have occurred in response to social complexity. For example, from the Reader and Laland’s (2002) comparative analysis of 445 observations of social learning [220], no correlation was found between social group size and social learning frequency. Currently, there is still little evidence regarding social learning being an adaptive specialization to particular environmental demands, beyond favoring behavioral flexibility in general [221].
On the other hand, the evolution towards larger brains could also have ecological explanations. Two prominent hypotheses in this domain are the Extractive Foraging Theory [159] and the Cognitive Mapping Hypothesis [158]. The former posits that the cognitive demands of accessing limited embedded foods through tool use and complex object manipulation drove brain expansion in primate ancestors. The latter proposes that the challenge of locating scattered food sources in rainforests led to enhanced spatial memory, mental mapping, and enlarged brains. These perspectives show how the discourse on the driving forces for brain evolution is still open to debate [222]. Overall, Reader and Laland study [220] support the idea that the EIH and SIH should not be considered alternatives to one another, since the sources that favored the evolution of a large primate executive brain were multiple [173,190,222].
Additionally, a recent meta-analysis of 103 studies across 17 taxonomic orders shows a correlation between sociality and cognition in various species, contexts, and measures, thus supporting the SIH as an evolutionary and developmental explanation for cognitive variations [187]. Nevertheless, authors have prompted recommendations for expanded research in underrepresented taxa, standardized cognitive and neuroanatomical tests, and collaborative programs to enhance understanding of cognitive evolution [187].
Whether sustaining the SIH, or the EIH, the driver of cognitive evolution could be considered the same in both cases, that is individuals gathering and processing information to mitigate uncertainty [223,224]. Indeed, treating social and ecological variables as independent could not be a proper approach, social species need to solve ecological problems in a social context, and social complexity itself may have evolved as a response to ecological factors [186,225].
Finally, the literature extensively explores the impact of social interaction on neurodevelopment and cognitive performance. While evolutionary pressures are significant, it is equally important to consider how sociality shapes cognitive development. This dual perspective raises a critical developmental question: How do insufficient or inappropriate social inputs affect brain development and cognitive function?
4. Effects of Early Social Deprivation
To understand the reason as to why early social experiences are so impactful and have long lasting consequences, it is necessary to understand and define some well-studied phenomena, such as the difference between sensitive and critical periods. Sensitive periods are a limited time window during development where there is a heightened potential for neural (re)organization in response to environmental inputs, whereas critical periods are a special class of sensitive periods where the neural system requires definitive inputs to organize and develop physiologically [226,227]. A well-known example of a sensitive period is song learning in songbirds, where young birds must be exposed to adult songs during a specific developmental window to learn their species-specific song patterns effectively. The timing of this sensitive period varies between species and can be influenced by both experience and hormones [228]. Instead, an example of critical period is filial imprinting in ducks and domestic chickens, where within a few days of hatching, these animals imprint on stimuli that they identify as the parent [229,230,231]. It is noteworthy that the phenomenon of filial imprinting has been extensively utilized in domestic chicks as a paradigm for investigating numerical cognition [135,148,232].
The experiences faced during limited periods in life can powerfully shape many aspects of perceptual, cognitive, and emotional capabilities, including the capacity to form strong social relationships and responses to stress due to changes in the hypothalamic-pituitary-adrenal axis [233]. For instance, Liu et al. (1997) demonstrated that maternal care in the early days of life significantly influences the development of the HPA axis in rats [233]. Specifically, the deprivation of maternal care leads to reduced expression of glucocorticoid receptors in the hippocampus, resulting in an exaggerated stress response in adulthood [233]. Notably, the experience must be of a particular kind, and it must occur within a certain period of time for the behavior to develop normally. Indeed, in the absence of species or age expected cognitive and social inputs, animals face a condition defined as deprivation [234]. In non-human animals social deprivation occurs as social isolation from their littermates and mothers [235] and maternal separation [236]; while in humans it manifests mainly as neglect, institutionalization, or living in impoverished environments [237,238].
Some researchers hypothesize that the lack of social complexity in an early environment may results in a neural structure designed to deal with low complexity environments [234]. This evidence would be explained by the phenomenon of synaptic pruning, a process in which unused or less active neural connections are eliminated. Synaptic pruning is a central force in the remodeling the brain’s neuronal connections throughout development in response to experience [239]. For example, animals raised in a deprived environment such as in an empty cage with no conspecifics, toys, and novel stimuli, exhibit reductions in the number of synapses per neuron [240], the density of cortical dendritic spines [241], the branching and length of dendrites [242,243], and cortical depth [244].
Critically, research reveals that early social isolation disrupts oligodendrocyte maturation and myelination in the prefrontal cortex through specific molecular pathways. Mice isolated during a critical postnatal period (during the fourth and fifth weeks of life) reduces expression of key genes, including neurogulin-1 (NRG1) and its receptor ErbB3. NRG1 is a signaling protein that plays a crucial role in neurodevelopment, particularly in the formation and maintenance of myelin sheaths. It acts by binding to and activating ErbB receptors, especially ErbB3, on oligodendrocytes (the cells responsible for producing myelin in the central nervous system). This NRG1-ErbB3 signaling pathway is essential for oligodendrocyte differentiation and the initiation of myelination. The reduction in NRG1 and ErbB3 expression leads to downregulation of myelin-related genes (e.g., MBP, MAG), causing permanent deficits in brain connectivity and social impairments [245].
According to other studies, the consequences of early social environments can manifest as long-term alterations of large-scale gene expression patterns in the brain [246,247]. For example, one study by Weaver and colleagues (2006) found that rats that received high levels of maternal care during the first week of life had increased expression of the glucocorticoid receptor gene in the hippocampus, compared to rats that received low levels of maternal care [247]. The glucocorticoid receptor gene is involved in the regulation of the hypothalamic–pituitary–adrenal axis, which is responsible for the body's stress response, making those who received high levels of maternal care more resilient to stress later in life.
Similarly, in rhesus monkeys, early postnatal deprivation of maternal care induces differences in gene expression in the amygdala. Sabatini and colleagues (2007) identified GUCY1A3 as a gene whose expression was significantly reduced in monkeys separated from their mothers at one week of age [246]. GUCY1A3 is involved in the production of the enzyme guanylate cyclase, which plays a role in various processes, including synaptic plasticity, learning, memory, and stress response.
Moreover, as a result of lack of complexity and absence of sensory, cognitive, and social stimulation, there are other mechanisms affected, such as epigenetic regulation [248]. Early social environments can lead to persistent alterations of brain gene expression [249,250]. For example, juvenile cooperatively breeding cichlids (Neolamprologus pulcher) raised in socially deprived conditions, specifically with sibling brood mates only, show less submission after being attacked by dominants compared to fish raised in a social group with parents, subordinates, and same-clutch siblings [251], and down-regulation of their Glucocorticoid Receptor 1 gene in the telencephalon compared to fish raised in a social group with parents, subordinates, and siblings from the same clutch [252].
Several studies have also demonstrated both the short- and long-term effects that early social experiences have on the physiological response to stress [253,254,255]. Rodents exposed to adverse rearing environments exhibit heightened reactivity and impaired ability of the hypothalamic-pituitary-adrenal axis to effectively terminate the stress response [256,257]. Caldji and colleagues (2000) assessed stress in rats by measuring corticosterone levels in response to maternal separation [256]. Corticosterone serves as a sensitive and readily quantifiable indicator of maternal deprivation stress in rat pups, reflecting hypothalamic-pituitary-adrenal axis activity and responding to both deprivation and interventions mimicking maternal care [258]. Indeed, extensive animal research has demonstrated that exposure to insufficient maternal care or maternal deprivation early in life results in protracted stress responses [259]. This phenomenon is observed across species; for example, in rhesus monkeys and Goeldi’s monkeys (Callimico goeldii), both Suomi (1999) and Dettling and colleagues (1998) measured stress by assessing cortisol, the primate equivalent of rodent corticosterone, levels in response to social separation and maternal separation, respectively [255,259].
Furthermore, early maternal deprivation affects other neurochemical pathways in both animals and humans [260]. An example is the dopaminergic system, a complex network of neurons involved in a variety of functions, including reward and motivation [261,262], movement and coordination [263,264], learning and memory [265,266], attention and focus [267], and emotion [268,269], where deprivation can cause a reduction of dopamine receptor binding in certain brain regions. The effects of isolation impair animals’ cognitive abilities. These include i) selective attention, that allows individuals to focus their attention on a particular task, object, or stimuli while ignoring other irrelevant or distracting information in the environment [270]; ii) inhibitory control, that involves the ability to suppress or inhibit automatic or prepotent responses, thoughts, or behaviors to achieve a specific goal or task [271]; iii) planning, which is the cognitive process of anticipating and organizing actions and behaviors to achieve a specific goal or objective; iv) working memory, that is responsible for temporarily holding and manipulating information for the purpose of completing complex cognitive tasks; v) problem solving, which is the cognitive process of identifying, analyzing, and resolving problems or obstacles that stand in the way of achieving a particular goal or objective [226].
The effects of early social deprivation have been demonstrated in numerous taxa such as fish [272], birds [273,274], rodents [275], primates [276], and humans [277] and the range of effects can extend from neuro-development, gene expression, physiology, cognition and even fitness. For example, mother-reared and peer-reared rhesus macaques acquired a higher social rank than young raised with limited access to conspecifics, suggesting that early social environment can give long-term effects that can affect even fitness [276].
Despite the effects of early social experience having been extensively studied in other cognitive domains [277], there remains a significant gap in understanding its specific role in the ontogenetic development of numerical cognition [109]. Regarding numerical cognition, its ontogeny remains understudied, particularly in understanding how social experience influences development. Few studies in animals were conducted on numerical cognition development. For example, the study by Sheardown and colleagues (2022) investigated proto-numerical discrimination in zebrafish (Danio rerio) across early ontogeny (21–33 days post-fertilization) [121]. The results show that fish’s ability in proto-numerical discrimination improves with age. While this study did not manipulate social experience, leaving open questions about its role, the study by Bisazza and colleagues (2010) on guppies (Poecilia reticulata) tested the impact of social experience by raising fish either in groups or in pairs [117]. At 20 days of age, guppies required social experience to discriminate large numerosity (4 vs. 8), whereas by 40 days, this ability emerged regardless of rearing conditions. Together, these studies suggest that while maturation drives basic numerical capacities, social experience may accelerate the refinement of number discrimination.
In line with this, recent findings suggest that social context can enhance numerical discrimination. Newborn chicks, when reared with individually distinctive objects (treated as social companions) were able to successfully discriminate between sets of 3 vs. 4 items, a comparison they typically fail when the object are all the same [136]. This improvement is thought to result from enhanced object individuation, which facilitates encoding in working memory via the Object File System. These findings suggest that social relevance and perceptual individuation can enhance non-symbolic numerical abilities by engaging working memory mechanisms dedicated to tracking individual objects [136].
Yet, how specific social experiences shape individual cognition remains underexplored, particularly in early developmental stages [186]. Within a social group, different group members experience different information-processing demands, depending on their pattern of agonistic and affiliative interactions, and the strength and number of their relationships [278]. Further evidence is needed to clarify this interaction across species and contexts.
5. Conclusions & Future Directions
This review highlights the crucial role of numerical abilities for a wide range of adaptive social behaviors in animals. Numerical cognition appears to be refined in response to the increasing demands of living in complex social environments. In support of the Social Intelligence Hypothesis, this perspective places numerical cognition not merely as a by-product of ecological challenges, but as a core adaptive function for navigating social environments [140]. Although cognitive abilities may have appeared several times in response to different ecological pressures, in social animals these abilities, once established, actively enhance social functioning—facilitating group cohesion, coordination, and survival. In this way, numerical cognition not only reflects but enables social complex behaviors.
Research on numerical cognition across various animal species has revealed the existence of common mechanisms for numerical processing [12,39], suggesting that similar evolutionary pressures have shaped the Number sense. The capacity to rapidly and accurately estimate numerosity and track small sets, together with skills such as proto-numerical discrimination, ordinal processing, proportional reasoning, and basic arithmetic directly contribute to social behaviors. Individuals with more refined numerical competences are likely to interact more effectively in social groups, which in turn can increase their fitness and promote the stability of group structures. As a result, numerical abilities may become widespread within populations where group living confers survival advantages, thus creating a positive evolutionary feedback loop between numerical cognition and sociality.
Empirical studies investigating how varying degrees of social enrichment relate to the development of numerical capacities could yield valuable insights. Future research may benefit from longitudinal approaches that assess numerical abilities across developmental stages, while combining behavioral measures with neuroimaging techniques to explore the neural correlates of experience-related differences.
A particularly relevant aspect concerns the distinction between early predispositions and acquired components of numerical cognition and the extent to which social interactions facilitate its development. Given the salience of numerical information in social contexts, it is plausible to hypothesize that the development of numerical abilities may be modulated by social environmental factors at an ontogenetic level. Furthermore, it can be postulated that the impairments observed in other cognitive domains due to social deprivation may also extend to numerical processing.
A deeper understanding of how the social environment modulates the development of numerical cognition will have significant implications. It will inform optimal rearing and environmental practices to promote healthy cognitive development, and shed light on neural mechanisms underlying the influence of early experience on the development of higher-order cognitive functions. Such research agenda promises to provide fundamental insights into the developmental pathways that underlie the extraordinary numerical competencies that have emerged through the selective pressures of social complexity.

Table 2.
Main pressures of the evolution of numerical cognition. The evolutionary pressure and the advantages of developing numerical processing are related to individual behavior and social behavior.
Table 2.
Main pressures of the evolution of numerical cognition. The evolutionary pressure and the advantages of developing numerical processing are related to individual behavior and social behavior.
| Pressure | Advantages of numerical processing | |
|---|---|---|
| Individual behavior | Resources availability | Discriminating food resources (feeding) |
| Mating opportunities | Discriminating number of male and female (reproduction) | |
| Habitat complexity | Counting landmark as reference point (navigation) | |
| Parasitism | Reject parasitized eggs (parental care) | |
| Social behavior | Collective foraging and hunting | Optimal distribution during foraging and numerical assessments during hunting strategies (feeding) |
| Territory defense | Use numerical assessment to establish advantages during intergroup conflict (aggression and defense) | |
| Predatory risk | Join larger groups for defense from predators (defense) | |
| Group organization | Ordering group member in hierarchy (navigate social system) | |
| Group cohesion | Initiate collective action (decision-making) |
Author Contributions
Conceptualization, M.M., A.F., and R.R; Writing – Original Draft Preparation, M.M.; Writing – Review & Editing, M.M., A.F., and R.R.; Supervision, R.R.; Funding Acquisition, R.R.
Funding
Italian Ministry of Education and Research through the Research Project of National Relevance (PRIN) – 2022 Prot. 202254RHRT, to R.R., and Italian Ministry of Education and Research through the Research Project of National Relevance (PRIN) – 2022 PNRR Prot. P2022TKY7B to R.R.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| OFS | Object File System |
| AMS | Analogue Magnitude System |
| EIH | Ecological Intelligence Hypothesis |
| SIH | Social Intelligence Hypothesis |
References
- Butterworth, B. Can Fish Count?: What Animals Reveal About Our Uniquely Mathematical Minds, New edition; Basic Books, 2022; ISBN 978-1-5416-2081-0. [Google Scholar]
- Rugani, R. Towards Numerical Cognition’s Origin: Insights from Day-Old Domestic Chicks. Philos Trans R Soc Lond B Biol Sci 2017, 373, 20160509. [Google Scholar] [CrossRef]
- Vallortigara, G. Comparative Cognition of Number and Space: The Case of Geometry and of the Mental Number Line. Philos Trans R Soc Lond B Biol Sci 2017, 373, 20170120. [Google Scholar] [CrossRef]
- Pfungst, O. Das Pferd des Herrn von Osten: Der kluge Hans. Ein Beitrag zur experimentellen Tier- und ...; J. A. Barth, 1907.
- Koehler, O. Vom Erlernen unbenannter Anzahlen bei Vögeln. Naturwissenschaften 1941, 29, 201–218. [Google Scholar] [CrossRef]
- Emmerton, J.; Lohmann, A.; Niemann, J. Pigeons’ Serial Ordering of Numerosity with Visual Arrays. Animal Learning & Behavior 1997, 25, 234–244. [Google Scholar] [CrossRef]
- Emmerton, J. Numerosity Differences and Effects of Stimulus Density on Pigeons’ Discrimination Performance. Animal Learning & Behavior 1998, 26, 243–256. [Google Scholar] [CrossRef]
- Meck, W.H.; Church, R.M. A Mode Control Model of Counting and Timing Processes. J Exp Psychol Anim Behav Process 1983, 9, 320–334. [Google Scholar] [CrossRef]
- Jordan, K.E.; Brannon, E.M. A Common Representational System Governed by Weber’s Law: Nonverbal Numerical Similarity Judgments in 6-Year-Olds and Rhesus Macaques. Journal of Experimental Child Psychology 2006, 95, 215–229. [Google Scholar] [CrossRef]
- Rugani, R.; Regolin, L.; Vallortigara, G. Discrimination of Small Numerosities in Young Chicks. Journal of Experimental Psychology: Animal Behavior Processes 2008, 34, 388–399. [Google Scholar] [CrossRef]
- Dehaene, S. The Number Sense : How the Mind Creates Mathematics; New York : Oxford University Press, 2011; ISBN 978-0-19-975387-1. [Google Scholar]
- Lorenzi, E.; Kobylkov, D.; Vallortigara, G. Is There an Innate Sense of Number in the Brain? Cereb Cortex 2025, 35, bhaf004. [Google Scholar] [CrossRef]
- Burr, D.C.; Anobile, G.; Arrighi, R. Psychophysical Evidence for the Number Sense. Philosophical Transactions of the Royal Society B: Biological Sciences 2018, 373, 20170045. [Google Scholar] [CrossRef] [PubMed]
- Halberda, J.; Mazzocco, M.M.M.; Feigenson, L. Individual Differences in Non-Verbal Number Acuity Correlate with Maths Achievement. Nature 2008, 455, 665–668. [Google Scholar] [CrossRef]
- Starr, A.; Libertus, M.E.; Brannon, E.M. Number Sense in Infancy Predicts Mathematical Abilities in Childhood. Proc Natl Acad Sci U S A 2013, 110, 18116–18120. [Google Scholar] [CrossRef] [PubMed]
- Brannon, E.M.; Terrace, H.S. Ordering of the Numerosities 1 to 9 by Monkeys. Science 1998, 282, 746–749. [Google Scholar] [CrossRef]
- Hauser, M.D.; Carey, S.; Hauser, L.B. Spontaneous Number Representation in Semi–Free–Ranging Rhesus Monkeys. Proceedings of the Royal Society of London. Series B: Biological Sciences 2000, 267, 829–833. [Google Scholar] [CrossRef]
- Rugani, R.; Vallortigara, G.; Priftis, K.; Regolin, L. Number-Space Mapping in the Newborn Chick Resembles Humans’ Mental Number Line. Science 2015, 347, 534–536. [Google Scholar] [CrossRef]
- Agrillo, C.; Piffer, L.; Bisazza, A.; Butterworth, B. Evidence for Two Numerical Systems That Are Similar in Humans and Guppies. PLOS ONE 2012, 7, e31923. [Google Scholar] [CrossRef]
- Howard, S.R.; Avarguès-Weber, A.; Garcia, J.E.; Greentree, A.D.; Dyer, A.G. Numerical Ordering of Zero in Honey Bees. Science 2018, 360, 1124–1126. [Google Scholar] [CrossRef] [PubMed]
- Kilian, A.; Yaman, S.; von Fersen, L.; Güntürkün, O. A Bottlenose Dolphin Discriminates Visual Stimuli Differing in Numerosity. Learning & Behavior 2003, 31, 133–142. [Google Scholar] [CrossRef]
- Davis, H.; Bradford, S.A. Counting Behavior by Rats in a Simulated Natural Environment. Ethology 1986, 73, 265–280. [Google Scholar] [CrossRef]
- Izard, V.; Sann, C.; Spelke, E.S.; Streri, A. Newborn Infants Perceive Abstract Numbers. Proceedings of the National Academy of Sciences 2009, 106, 10382–10385. [Google Scholar] [CrossRef] [PubMed]
- Wynn, K. Addition and Subtraction by Human Infants. Nature 1992, 358, 749–750. [Google Scholar] [CrossRef]
- Xu, F.; Spelke, E.S. Large Number Discrimination in 6-Month-Old Infants. Cognition 2000, 74, B1–B11. [Google Scholar] [CrossRef]
- Feigenson, L.; Carey, S.; Spelke, E. Infants’ Discrimination of Number vs. Continuous Extent. Cognitive Psychology 2002, 44, 33–66. [Google Scholar] [CrossRef]
- Lipton, J.S.; Spelke, E.S. Origins of Number Sense: Large-Number Discrimination in Human Infants. Psychol Sci 2003, 14, 396–401. [Google Scholar] [CrossRef]
- Xu, F. Numerosity Discrimination in Infants: Evidence for Two Systems of Representations. Cognition 2003, 89, B15–B25. [Google Scholar] [CrossRef] [PubMed]
- Kinzler, K.D.; Spelke, E.S. Core Systems in Human Cognition. Prog Brain Res 2007, 164, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Spelke, E.S. Core Knowledge. American Psychologist 2000, 55, 1233–1243. [Google Scholar] [CrossRef]
- Vallortigara, G.; Chiandetti, C.; Rugani, R.; Sovrano, V.A.; Regolin, L. Animal Cognition. Wiley Interdiscip Rev Cogn Sci 2010, 1, 882–893. [Google Scholar] [CrossRef]
- Trick, L.M.; Pylyshyn, Z.W. Why Are Small and Large Numbers Enumerated Differently? A Limited-Capacity Preattentive Stage in Vision. Psychol Rev 1994, 101, 80–102. [Google Scholar] [CrossRef]
- Feigenson, L.; Carey, S.; Hauser, M. The Representations Underlying Infants’ Choice of More: Object Files Versus Analog Magnitudes. Psychol Sci 2002, 13, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Gallistel, C.R.; Gelman, R. Preverbal and Verbal Counting and Computation. Cognition 1992, 44, 43–74. [Google Scholar] [CrossRef]
- Wynn, K. Psychological Foundations of Number: Numerical Competence in Human Infants. Trends in Cognitive Sciences 1998, 2, 296–303. [Google Scholar] [CrossRef]
- Feigenson, L.; Libertus, M.E.; Halberda, J. Links Between the Intuitive Sense of Number and Formal Mathematics Ability. Child Dev Perspect 2013, 7, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Piazza, M.; Facoetti, A.; Trussardi, A.N.; Berteletti, I.; Conte, S.; Lucangeli, D.; Dehaene, S.; Zorzi, M. Developmental Trajectory of Number Acuity Reveals a Severe Impairment in Developmental Dyscalculia. Cognition 2010, 116, 33–41. [Google Scholar] [CrossRef]
- Moyer, R.S.; Landauer, T.K. Time Required for Judgements of Numerical Inequality. Nature 1967, 215, 1519–1520. [Google Scholar] [CrossRef]
- Nieder, A. The Evolutionary History of Brains for Numbers. Trends in Cognitive Sciences 2021, 25, 608–621. [Google Scholar] [CrossRef]
- Peters, L.; De Smedt, B. Arithmetic in the Developing Brain: A Review of Brain Imaging Studies. Developmental Cognitive Neuroscience 2018, 30, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Vogel, S.E.; De Smedt, B. Developmental Brain Dynamics of Numerical and Arithmetic Abilities. npj Sci. Learn. 2021, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, E.; Perrino, M.; Vallortigara, G. Numerosities and Other Magnitudes in the Brains: A Comparative View. Front. Psychol. 2021, 12. [Google Scholar] [CrossRef]
- Ditz, H.M.; Nieder, A. Neurons Selective to the Number of Visual Items in the Corvid Songbird Endbrain. Proceedings of the National Academy of Sciences 2015, 112, 7827–7832. [Google Scholar] [CrossRef]
- Wagener, L.; Nieder, A. Categorical Representation of Abstract Spatial Magnitudes in the Executive Telencephalon of Crows. Current Biology 2023, 33, 2151–2162.e5. [Google Scholar] [CrossRef]
- Sawamura, H.; Shima, K.; Tanji, J. Numerical Representation for Action in the Parietal Cortex of the Monkey. Nature 2002, 415, 918–922. [Google Scholar] [CrossRef]
- Kobylkov, D.; Mayer, U.; Zanon, M.; Vallortigara, G. Number Neurons in the Nidopallium of Young Domestic Chicks. Proceedings of the National Academy of Sciences 2022, 119, e2201039119. [Google Scholar] [CrossRef]
- Lorenzi, E.; Perrino, M.; Messina, A.; Zanon, M.; Vallortigara, G. Innate Responses to Numerousness Reveal Neural Activation in Different Brain Regions in Newly-Hatched Visually Naïve Chicks. Heliyon 2024, 10. [Google Scholar] [CrossRef]
- Luu, P.; Nadtochiy, A.; Zanon, M.; Moreno, N.; Messina, A.; Petrazzini, M.E.M.; Torres Perez, J.V.; Keomanee-Dizon, K.; Jones, M.; Brennan, C.H.; et al. Neural Basis of Number Sense in Larval Zebrafish. bioRxiv 2024. [Google Scholar] [CrossRef] [PubMed]
- Ditz, H.M.; Nieder, A. Numerosity Representations in Crows Obey the Weber–Fechner Law. Proceedings of the Royal Society B: Biological Sciences 2016, 283, 20160083. [Google Scholar] [CrossRef] [PubMed]
- Szűcs, D.; Devine, A.; Soltesz, F.; Nobes, A.; Gabriel, F. Cognitive Components of a Mathematical Processing Network in 9-Year-Old Children. Developmental Science 2014, 17, 506–524. [Google Scholar] [CrossRef]
- Feigenson, L.; Dehaene, S.; Spelke, E. Core Systems of Number. Trends Cogn Sci 2004, 8, 307–314. [Google Scholar] [CrossRef]
- Nieder, A.; Dehaene, S. Representation of Number in the Brain. Annual Review of Neuroscience 2009, 32, 185–208. [Google Scholar] [CrossRef]
- Barnard, A.M.; Hughes, K.D.; Gerhardt, R.R.; DiVincenti, L.; Bovee, J.M.; Cantlon, J.F. Inherently Analog Quantity Representations in Olive Baboons (Papio Anubis). Front. Psychol. 2013, 4. [Google Scholar] [CrossRef]
- Beran, M.J.; Evans, T.A.; Leighty, K.A.; Harris, E.H.; Rice, D. Summation and Quantity Judgments of Sequentially Presented Sets by Capuchin Monkeys (Cebus Apella). American Journal of Primatology 2008, 70, 191–194. [Google Scholar] [CrossRef]
- Flombaum, J.I.; Junge, J.A.; Hauser, M.D. Rhesus Monkeys (Macaca Mulatta) Spontaneously Compute Addition Operations over Large Numbers. Cognition 2005, 97, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Kilian, A.; Yaman, S.; von Fersen, L.; Güntürkün, O. A Bottlenose Dolphin Discriminates Visual Stimuli Differing in Numerosity. Learning & Behavior 2003, 31, 133–142. [Google Scholar] [CrossRef]
- Panteleeva, S.; Reznikova, Z.; Vygonyailova, O. Quantity Judgments in the Context of Risk/Reward Decision Making in Striped Field Mice: First “Count,” Then Hunt. Front. Psychol. 2013, 4. [Google Scholar] [CrossRef]
- Perdue, B.M.; Talbot, C.F.; Stone, A.M.; Beran, M.J. Putting the Elephant Back in the Herd: Elephant Relative Quantity Judgments Match Those of Other Species. Anim Cogn 2012, 15, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Emmerton, J.; Renner, J.C. Scalar Effects in the Visual Discrimination of Numerosity by Pigeons. Learn Behav 2006, 34, 176–192. [Google Scholar] [CrossRef]
- Pepperberg, I.M. Cognitive and Communicative Abilities of Grey Parrots. Applied Animal Behaviour Science 2006, 100, 77–86. [Google Scholar] [CrossRef]
- Kelly, E.M. Counting on Your Friends: The Role of Social Environment on Quantity Discrimination. Behav Processes 2016, 128, 9–16. [Google Scholar] [CrossRef]
- Hunter, H.; Blackburn, G.; Ashton, B.J.; Ridley, A.R. Group Size Affects Spontaneous Quantity Discrimination Performance in Wild Western Australian Magpies (Gymnorhina Tibicen Dorsalis). Anim Cogn 2025, 28, 41. [Google Scholar] [CrossRef]
- Agrillo, C.; Dadda, M.; Serena, G.; Bisazza, A. Do Fish Count? Spontaneous Discrimination of Quantity in Female Mosquitofish. Anim Cogn 2008, 11, 495–503. [Google Scholar] [CrossRef]
- Agrillo, C.; Miletto Petrazzini, M.E.; Tagliapietra, C.; Bisazza, A. Inter-Specific Differences in Numerical Abilities Among Teleost Fish. Front. Psychol. 2012, 3. [Google Scholar] [CrossRef]
- Gómez-Laplaza, L.M.; Gerlai, R. Spontaneous Discrimination of Small Quantities: Shoaling Preferences in Angelfish (Pterophyllum Scalare). Anim Cogn 2011, 14, 565–574. [Google Scholar] [CrossRef]
- Potrich, D.; Sovrano, V.A.; Stancher, G.; Vallortigara, G. Quantity Discrimination by Zebrafish (Danio Rerio). Journal of Comparative Psychology 2015, 129, 388–393. [Google Scholar] [CrossRef]
- Stancher, G.; Sovrano, V.A.; Potrich, D.; Vallortigara, G. Discrimination of Small Quantities by Fish (Redtail Splitfin, Xenotoca Eiseni). Anim Cogn 2013, 16, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Krusche, P.; Uller, C.; Dicke, U. Quantity Discrimination in Salamanders. Journal of Experimental Biology 2010, 213, 1822–1828. [Google Scholar] [CrossRef]
- Stancher, G.; Rugani, R.; Regolin, L.; Vallortigara, G. Numerical Discrimination by Frogs (Bombina Orientalis). Anim Cogn 2015, 18, 219–229. [Google Scholar] [CrossRef]
- Uller, C.; Jaeger, R.; Guidry, G.; Martin, C. Salamanders (Plethodon Cinereus) Go for More: Rudiments of Number in an Amphibian. Anim Cogn 2003, 6, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Gazzola, A.; Vallortigara, G.; Pellitteri-Rosa, D. Continuous and Discrete Quantity Discrimination in Tortoises. Biology Letters 2018, 14, 20180649. [Google Scholar] [CrossRef]
- Miletto Petrazzini, M.E.; Fraccaroli, I.; Gariboldi, F.; Agrillo, C.; Bisazza, A.; Bertolucci, C.; Foà, A. Quantitative Abilities in a Reptile (Podarcis Sicula). Biology Letters 2017, 13, 20160899. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.-C.; Whiting, M.J.; Hsieh, M.-Y.; Shaner, P.-J.L.; Lin, S.-M. Superior Continuous Quantity Discrimination in a Freshwater Turtle. Frontiers in Zoology 2021, 18, 49. [Google Scholar] [CrossRef]
- Szabo, B.; Noble, D.W.A.; McCloghry, K.J.; Monteiro, M.E.S.; Whiting, M.J. Spontaneous Quantity Discrimination in a Family-Living Lizard. Behav Ecol 2021, 32, 686–694. [Google Scholar] [CrossRef]
- Giurfa, M. Honeybees Foraging for Numbers. J Comp Physiol A 2019, 205, 439–450. [Google Scholar] [CrossRef]
- Howard, S.R.; Avarguès-Weber, A.; Garcia, J.E.; Greentree, A.D.; Dyer, A.G. Numerical Cognition in Honeybees Enables Addition and Subtraction. Science Advances 2019, 5, eaav0961. [Google Scholar] [CrossRef]
- Pahl, M.; Si, A.; Zhang, S. Numerical Cognition in Bees and Other Insects. Front. Psychol. 2013, 4. [Google Scholar] [CrossRef]
- Rodríguez, R.L.; Briceño, R.D.; Briceño-Aguilar, E.; Höbel, G. Nephila Clavipes Spiders (Araneae: Nephilidae) Keep Track of Captured Prey Counts: Testing for a Sense of Numerosity in an Orb-Weaver. Anim Cogn 2015, 18, 307–314. [Google Scholar] [CrossRef]
- Tanner, C.J. Numerical Assessment Affects Aggression and Competitive Ability: A Team-Fighting Strategy for the Ant Formica Xerophila. Proceedings of the Royal Society B: Biological Sciences 2006, 273, 2737–2742. [Google Scholar] [CrossRef]
- Yang, T.-I.; Chiao, C.-C. Number Sense and State-Dependent Valuation in Cuttlefish. Proceedings of the Royal Society B: Biological Sciences 2016, 283, 20161379. [Google Scholar] [CrossRef]
- Krebs, J.R.; Ryan, J.C.; Charnov, E.L. Hunting by Expectation or Optimal Foraging? A Study of Patch Use by Chickadees. Animal Behaviour 1974, 22, 953–IN3. [Google Scholar] [CrossRef]
- Davis, H.; Pérusse, R. Numerical Competence in Animals: Definitional Issues, Current Evidence, and a New Research Agenda. Behavioral and Brain Sciences 1988, 11, 561–615. [Google Scholar] [CrossRef]
- Bogale, B.A.; Kamata, N.; Mioko, K.; Sugita, S. Quantity Discrimination in Jungle Crows, Corvus Macrorhynchos. Animal Behaviour 2011, 82, 635–641. [Google Scholar] [CrossRef]
- Swenson, L.C. One versus Two Discrimination by Whitenecked Ravens (Corvus Cryptoleucus) with Non-Number Dimensions Varied. Animal Behaviour 1970, 18, 454–460. [Google Scholar] [CrossRef]
- Potrich, D.; Zanon, M.; Vallortigara, G. Archerfish Number Discrimination. eLife 2022, 11, e74057. [Google Scholar] [CrossRef]
- Bortot, M.; Agrillo, C.; Avarguès-Weber, A.; Bisazza, A.; Miletto Petrazzini, M.E.; Giurfa, M. Honeybees Use Absolute Rather than Relative Numerosity in Number Discrimination. Biology Letters 2019, 15, 20190138. [Google Scholar] [CrossRef]
- Harper, D.G.C. Competitive Foraging in Mallards: “Ideal Free’ Ducks. Animal Behaviour 1982, 30, 575–584. [Google Scholar] [CrossRef]
- Rugani, R.; Vallortigara, G.; Regolin, L. The Use of Proportion by Young Domestic Chicks (Gallus Gallus). Anim Cogn 2015, 18, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Rugani, R.; McCrink, K.; de Hevia, M.-D.; Vallortigara, G.; Regolin, L. Ratio Abstraction over Discrete Magnitudes by Newly Hatched Domestic Chicks (Gallus Gallus). Sci Rep 2016, 6, 30114. [Google Scholar] [CrossRef]
- Godin, J.-G.J.; Keenleyside, M.H. Foraging on Patchily Distributed Prey by a Cichlid Fish (Teleostei, Cichlidae): A Test of the Ideal Free Distribution Theory. Animal Behaviour 1984, 32, 120–131. [Google Scholar] [CrossRef]
- Drucker, C.B.; Rossa, M.A.; Brannon, E.M. Comparison of Discrete Ratios by Rhesus Macaques (Macaca Mulatta). Anim Cogn 2016, 19, 75–89. [Google Scholar] [CrossRef]
- Rakoczy, H.; Clüver, A.; Saucke, L.; Stoffregen, N.; Gräbener, A.; Migura, J.; Call, J. Apes Are Intuitive Statisticians. Cognition 2014, 131, 60–68. [Google Scholar] [CrossRef]
- Rakoczy, H.; Clüver, A.; Saucke, L.; Stoffregen, N.; Gräbener, A.; Migura, J.; Call, J. Apes Are Intuitive Statisticians. Cognition 2014, 131, 60–68. [Google Scholar] [CrossRef]
- Agrillo, C.; Dadda, M.; Serena, G. Choice of Female Groups by Male Mosquitofish (Gambusia Holbrooki). Ethology 2008, 114, 479–488. [Google Scholar] [CrossRef]
- Pilastro, A.; Scaggiante, M.; Rasotto, M.B. Individual Adjustment of Sperm Expenditure Accords with Sperm Competition Theory. Proceedings of the National Academy of Sciences 2002, 99, 9913–9915. [Google Scholar] [CrossRef]
- Carazo, P.; Font, E.; Forteza-Behrendt, E.; Desfilis, E. Quantity Discrimination in Tenebrio Molitor: Evidence of Numerosity Discrimination in an Invertebrate? Anim Cogn 2009, 12, 463–470. [Google Scholar] [CrossRef]
- Carazo, P.; Fernández-Perea, R.; Font, E. Quantity Estimation Based on Numerical Cues in the Mealworm Beetle (Tenebrio Molitor). Front. Psychol. 2012, 3. [Google Scholar] [CrossRef]
- Lyon, B.E. Egg Recognition and Counting Reduce Costs of Avian Conspecific Brood Parasitism. Nature 2003, 422, 495–499. [Google Scholar] [CrossRef]
- Suzuki, K.; Kobayashi, T. Numerical Competence in Rats (Rattus Norvegicus): Davis and Bradford (1986) Extended. Journal of Comparative Psychology 2000, 114, 73–85. [Google Scholar] [CrossRef]
- Rugani, R.; Regolin, L.; Vallortigara, G. Rudimental Numerical Competence in 5-Day-Old Domestic Chicks (Gallus Gallus): Identification of Ordinal Position. Journal of Experimental Psychology: Animal Behavior Processes 2007, 33, 21–31. [Google Scholar] [CrossRef]
- Rugani, R.; Zhang, Y.; Scarsi, B.; Regolin, L. Hybro Chicks Outperform Ross308 in a Numerical-Ordinal Task. Cognitive and Behavioral Comparisons between 2 Broiler Strains of Newborn Domestic Chicks (Gallus Gallus). Poultry Science 2023, 102, 103148. [Google Scholar] [CrossRef] [PubMed]
- Rugani, R.; Kelly, D.M.; Szelest, I.; Regolin, L.; Vallortigara, G. Is It Only Humans That Count from Left to Right? Biology Letters 2010, 6, 290–292. [Google Scholar] [CrossRef] [PubMed]
- Potrich, D.; Rugani, R.; Sovrano, V.A.; Regolin, L.; Vallortigara, G. Use of Numerical and Spatial Information in Ordinal Counting by Zebrafish. Sci Rep 2019, 9, 18323. [Google Scholar] [CrossRef] [PubMed]
- Chittka, L.; Geiger, K. Can Honey Bees Count Landmarks? Animal Behaviour 1995, 49, 159–164. [Google Scholar] [CrossRef]
- Scarf, D.; Colombo, M. Knowledge of the Ordinal Position of List Items in Pigeons. Journal of Experimental Psychology: Animal Behavior Processes 2011, 37, 483–487. [Google Scholar] [CrossRef]
- Lyons, I.M.; Vogel, S.E.; Ansari, D. On the Ordinality of Numbers: A Review of Neural and Behavioral Studies. Prog Brain Res 2016, 227, 187–221. [Google Scholar] [CrossRef]
- Brunamonti, E.; Mione, V.; Bello, F.D.; Pani, P.; Genovesio, A.; Ferraina, S. Neuronal Modulation in the Prefrontal Cortex in a Transitive Inference Task: Evidence of Neuronal Correlates of Mental Schema Management. J. Neurosci. 2016, 36, 1223–1236. [Google Scholar] [CrossRef]
- Bergman, T.J.; Beehner, J.C.; Cheney, D.L.; Seyfarth, R.M. Hierarchical Classification by Rank and Kinship in Baboons. Science 2003, 302, 1234–1236. [Google Scholar] [CrossRef]
- Agrillo, C.; Bisazza, A. Understanding the Origin of Number Sense: A Review of Fish Studies. Philosophical Transactions of the Royal Society B: Biological Sciences 2018, 373, 20160511. [Google Scholar] [CrossRef]
- Nieder, A. The Adaptive Value of Numerical Competence. Trends in Ecology & Evolution 2020, 35, 605–617. [Google Scholar] [CrossRef]
- McComb, K.; Packer, C.; Pusey, A. Roaring and Numerical Assessment in Contests between Groups of Female Lions, Panthera Leo. Animal Behaviour 1994, 47, 379–387. [Google Scholar] [CrossRef]
- Benson-Amram, S.; Heinen, V.K.; Dryer, S.L.; Holekamp, K.E. Numerical Assessment and Individual Call Discrimination by Wild Spotted Hyaenas, Crocuta Crocuta. Animal Behaviour 2011, 82, 743–752. [Google Scholar] [CrossRef]
- Wilson, M.L.; Hauser, M.D.; Wrangham, R.W. Does Participation in Intergroup Conflict Depend on Numerical Assessment, Range Location, or Rank for Wild Chimpanzees? Animal Behaviour 2001, 61, 1203–1216. [Google Scholar] [CrossRef]
- Kitchen, D.M. Experimental Test of Female Black Howler Monkey (Alouatta Pigra) Responses to Loud Calls from Potentially Infanticidal Males: Effects of Numeric Odds, Vulnerable Offspring, and Companion Behavior. American Journal of Physical Anthropology 2006, 131, 73–83. [Google Scholar] [CrossRef]
- Seddon, N.; Tobias, J.A. Communal Singing in the Cooperatively Breeding Subdesert Mesite Monias Benschi: Evidence of Numerical Assessment? Journal of Avian Biology 2003, 34, 72–80. [Google Scholar] [CrossRef]
- Bonanni, R.; Natoli, E.; Cafazzo, S.; Valsecchi, P. Free-Ranging Dogs Assess the Quantity of Opponents in Intergroup Conflicts. Anim Cogn 2011, 14, 103–115. [Google Scholar] [CrossRef]
- Bisazza, A.; Piffer, L.; Serena, G.; Agrillo, C. Ontogeny of Numerical Abilities in Fish. PLoS One 2010, 5, e15516. [Google Scholar] [CrossRef]
- Lucon-Xiccato, T.; Dadda, M. Individual Guppies Differ in Quantity Discrimination Performance across Antipredator and Foraging Contexts. Behav Ecol Sociobiol 2016, 71, 13. [Google Scholar] [CrossRef]
- Mehlis, M.; Thünken, T.; Bakker, T.C.M.; Frommen, J.G. Quantification Acuity in Spontaneous Shoaling Decisions of Three-Spined Sticklebacks. Anim Cogn 2015, 18, 1125–1131. [Google Scholar] [CrossRef]
- Bshary, R.; Noë, R. Red Colobus and Diana Monkeys Provide Mutual Protection against Predators. Animal Behaviour 1997, 54, 1461–1474. [Google Scholar] [CrossRef] [PubMed]
- Sheardown, E.; Torres-Perez, J.V.; Anagianni, S.; Fraser, S.E.; Vallortigara, G.; Butterworth, B.; Miletto-Petrazzini, M.E.; Brennan, C.H. Characterizing Ontogeny of Quantity Discrimination in Zebrafish. Proc Biol Sci 2022, 289, 20212544. [Google Scholar] [CrossRef]
- Massen, J.J.M.; Pašukonis, A.; Schmidt, J.; Bugnyar, T. Ravens Notice Dominance Reversals among Conspecifics within and Outside Their Social Group. Nat Commun 2014, 5, 3679. [Google Scholar] [CrossRef] [PubMed]
- Weiß, B.M.; Kehmeier, S.; Schloegl, C. Transitive Inference in Free-Living Greylag Geese, Anser Anser. Animal Behaviour 2010, 79, 1277–1283. [Google Scholar] [CrossRef]
- Daisley, J.N.; Vallortigara, G.; Regolin, L. Low-Rank Gallus Gallus Domesticus Chicks Are Better at Transitive Inference Reasoning. Commun Biol 2021, 4, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Degrande, R.; Amichaud, O.; Piégu, B.; Cornilleau, F.; Jardat, P.; Ferreira, V.H.B.; Colson, V.; Lansade, L.; Calandreau, L. Transitive Reasoning in the Adult Domestic Hen in a Six-Term Series Task. Anim Cogn 2024, 27, 77. [Google Scholar] [CrossRef]
- Tibbetts, E.A.; Agudelo, J.; Pandit, S.; Riojas, J. Transitive Inference in Polistes Paper Wasps. Biology Letters 2019, 15, 20190015. [Google Scholar] [CrossRef]
- Cronin, A.L. Ratio-Dependent Quantity Discrimination in Quorum Sensing Ants. Anim Cogn 2014, 17, 1261–1268. [Google Scholar] [CrossRef]
- Doering, G.N.; Pratt, S.C. Queen Location and Nest Site Preference Influence Colony Reunification by the Ant Temnothorax Rugatulus. Insect. Soc. 2016, 63, 585–591. [Google Scholar] [CrossRef]
- Franks, N.R.; Stuttard, J.P.; Doran, C.; Esposito, J.C.; Master, M.C.; Sendova-Franks, A.B.; Masuda, N.; Britton, N.F. How Ants Use Quorum Sensing to Estimate the Average Quality of a Fluctuating Resource. Sci Rep 2015, 5, 11890. [Google Scholar] [CrossRef]
- Boinski, S. Vocal Coordination of Troop Movement among White-Faced Capuchin Monkeys, Cebus Capucinus. American Journal of Primatology 1993, 30, 85–100. [Google Scholar] [CrossRef]
- Strandburg-Peshkin, A.; Farine, D.R.; Couzin, I.D.; Crofoot, M.C. Shared Decision-Making Drives Collective Movement in Wild Baboons. Science 2015, 348, 1358–1361. [Google Scholar] [CrossRef]
- Walker, R.H.; King, A.J.; McNutt, J.W.; Jordan, N.R. Sneeze to Leave: African Wild Dogs (Lycaon Pictus) Use Variable Quorum Thresholds Facilitated by Sneezes in Collective Decisions. Proceedings of the Royal Society B: Biological Sciences 2017, 284, 20170347. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, C.A.H.; Sumpter, D.J.T.; Manser, M.B. Moving Calls: A Vocal Mechanism Underlying Quorum Decisions in Cohesive Groups. Proceedings of the Royal Society B: Biological Sciences 2010, 278, 1482–1488. [Google Scholar] [CrossRef]
- Visscher, P.K.; Seeley, T.D. Coordinating a Group Departure: Who Produces the Piping Signals on Honeybee Swarms? Behav Ecol Sociobiol 2007, 61, 1615–1621. [Google Scholar] [CrossRef]
- Rugani, R.; Fontanari, L.; Simoni, E.; Regolin, L.; Vallortigara, G. Arithmetic in Newborn Chicks. Proceedings of the Royal Society B: Biological Sciences 2009, 276, 2451–2460. [Google Scholar] [CrossRef]
- Rugani, R.; Loconsole, M.; Simion, F.; Regolin, L. Individually Distinctive Features Facilitate Numerical Discrimination of Sets of Objects in Domestic Chicks. Sci Rep 2020, 10, 16408. [Google Scholar] [CrossRef]
- Angulo, E.; Luque, G.M.; Gregory, S.D.; Wenzel, J.W.; Bessa-Gomes, C.; Berec, L.; Courchamp, F. Review: Allee Effects in Social Species. Journal of Animal Ecology 2018, 87, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Clutton-Brock Breeding Together: Kin Selection and Mutualism in Cooperative Vertebrates. Science 2002, 296, 69–72. [CrossRef]
- Cockburn, A. Evolution of Helping Behavior in Cooperatively Breeding Birds. Annual Review of Ecology and Systematics 1998, 29. [Google Scholar] [CrossRef]
- Benson-Amram, S.; Gilfillan, G.; McComb, K. Numerical Assessment in the Wild: Insights from Social Carnivores. Philosophical Transactions of the Royal Society B: Biological Sciences 2018, 373, 20160508. [Google Scholar] [CrossRef]
- Parker, G.A. Assessment Strategy and the Evolution of Fighting Behaviour. Journal of Theoretical Biology 1974, 47, 223–243. [Google Scholar] [CrossRef] [PubMed]
- Watts, D.P.; Mitani, J.C. Hunting Behavior of Chimpanzees at Ngogo, Kibale National Park, Uganda. International Journal of Primatology 2002, 23, 1–28. [Google Scholar] [CrossRef]
- Caro, T.M. Antipredator Defenses in Birds and Mammals; University of Chicago Press, 2005; ISBN 978-0-226-09436-6. [Google Scholar]
- Hamilton, W.D. Geometry for the Selfish Herd. Journal of Theoretical Biology 1971, 31, 295–311. [Google Scholar] [CrossRef]
- Kenward, R.E. Hawks and Doves: Factors Affecting Success and Selection in Goshawk Attacks on Woodpigeons. Journal of Animal Ecology 1978, 47, 449–460. [Google Scholar] [CrossRef]
- Krause, J.; Ruxton, G.D. Living in Groups; OUP Oxford, 2002; ISBN 978-0-19-850818-2. [Google Scholar]
- Hager, M.C.; Helfman, G.S. Safety in Numbers: Shoal Size Choice by Minnows under Predatory Threat. Behav Ecol Sociobiol 1991, 29, 271–276. [Google Scholar] [CrossRef]
- Rugani, R.; Regolin, L.; Vallortigara, G. Imprinted Numbers: Newborn Chicks’ Sensitivity to Number vs. Continuous Extent of Objects They Have Been Reared With. Developmental Science 2010, 13, 790–797. [Google Scholar] [CrossRef]
- Loconsole, M.; Regolin, L.; Rugani, R. Asymmetric Number–Space Association Leads to More Efficient Processing of Congruent Information in Domestic Chicks. Front. Behav. Neurosci. 2023, 17. [Google Scholar] [CrossRef]
- Bugnyar, T. Why Are Ravens Smart? Exploring the Social Intelligence Hypothesis. J Ornithol 2024, 165, 15–26. [Google Scholar] [CrossRef]
- Braun, A.; Bugnyar, T. Social Bonds and Rank Acquisition in Raven Nonbreeder Aggregations. Animal Behaviour 2012, 84, 1507–1515. [Google Scholar] [CrossRef]
- Lazareva, O.F.; Smirnova, A.A.; Bagozkaja, M.S.; Zorina, Z.A.; Rayevsky, V.V.; Wasserman, E.A. Transitive Responding in Hooded Crows Requires Linearly Ordered Stimuli. Journal of the Experimental Analysis of Behavior 2004, 82, 1–19. [Google Scholar] [CrossRef]
- Fernald, R.D. Cognitive Skills and the Evolution of Social Systems. Journal of Experimental Biology 2017, 220, 103–113. [Google Scholar] [CrossRef]
- MacLean, E.L.; Merritt, D.J.; Brannon, E.M. Social Complexity Predicts Transitive Reasoning in Prosimian Primates. Animal Behaviour 2008, 76, 479–486. [Google Scholar] [CrossRef]
- Chase, I.D. Behavioral Sequences During Dominance Hierarchy Formation in Chickens. Science 1982, 216, 439–440. [Google Scholar] [CrossRef] [PubMed]
- Benard, J.; Giurfa, M. A Test of Transitive Inferences in Free-Flying Honeybees: Unsuccessful Performance Due to Memory Constraints. Learn Mem 2004, 11, 328–336. [Google Scholar] [CrossRef]
- Regolin, L.; Loconsole, M.; Rosa-Salva, O.; Brosche, K.; Macchinizzi, M.; Felisatti, A.; Rugani, R. Numerical Cognition in Birds. Nat Rev Psychol 2025, 1–15. [Google Scholar] [CrossRef]
- Milton, K. Foraging Behaviour and the Evolution of Primate Intelligence. In Machiavellian intelligence: Social expertise and the evolution of intellect in monkeys, apes, and humans; Clarendon Press/Oxford University Press: New York, NY, US, 1988; ISBN 978-0-19-852179-2. [Google Scholar]
- Parker, S.T.; Gibson, K.R. Object Manipulation, Tool Use and Sensorimotor Intelligence as Feeding Adaptations in Cebus Monkeys and Great Apes. Journal of Human Evolution 1977, 6, 623–641. [Google Scholar] [CrossRef]
- Lefebvre, L.; Sol, D. Brains, Lifestyles and Cognition: Are There General Trends? Brain Behavior and Evolution 2008, 72, 135–144. [Google Scholar] [CrossRef]
- Morand-Ferron, J.; Cole, E.F.; Quinn, J.L. Studying the Evolutionary Ecology of Cognition in the Wild: A Review of Practical and Conceptual Challenges. Biological Reviews 2016, 91, 367–389. [Google Scholar] [CrossRef] [PubMed]
- Sayol, F.; Maspons, J.; Lapiedra, O.; Iwaniuk, A.N.; Székely, T.; Sol, D. Environmental Variation and the Evolution of Large Brains in Birds. Nat Commun 2016, 7, 13971. [Google Scholar] [CrossRef] [PubMed]
- Shumway, C.A. Habitat Complexity, Brain, and Behavior. Brain Behavior and Evolution 2008, 72, 123–134. [Google Scholar] [CrossRef]
- Soler, J.J.; Peralta-Sánchez, J.M.; Martín-Vivaldi, M.; Martín-Platero, A.M.; Flensted-Jensen, E.; Møller, A.P. Cognitive Skills and Bacterial Load: Comparative Evidence of Costs of Cognitive Proficiency in Birds. Naturwissenschaften 2012, 99, 111–122. [Google Scholar] [CrossRef]
- Dunn, J.C.; Cole, E.F.; Quinn, J.L. Personality and Parasites: Sex-Dependent Associations between Avian Malaria Infection and Multiple Behavioural Traits. Behav Ecol Sociobiol 2011, 65, 1459–1471. [Google Scholar] [CrossRef]
- Brown, C.; Braithwaite, V.A. Effects of Predation Pressure on the Cognitive Ability of the Poeciliid Brachyraphis Episcopi. Behavioral Ecology 2005, 16, 482–487. [Google Scholar] [CrossRef]
- Ferrari, M.C.O. Short-Term Environmental Variation in Predation Risk Leads to Differential Performance in Predation-Related Cognitive Function. Animal Behaviour 2014, 95, 9–14. [Google Scholar] [CrossRef]
- Tebbich, S.; Teschke, I. Coping with Uncertainty: Woodpecker Finches (Cactospiza Pallida) from an Unpredictable Habitat Are More Flexible than Birds from a Stable Habitat. PLOS ONE 2014, 9, e91718. [Google Scholar] [CrossRef] [PubMed]
- Mazza, V.; Jacob, J.; Dammhahn, M.; Zaccaroni, M.; Eccard, J.A. Individual Variation in Cognitive Style Reflects Foraging and Anti-Predator Strategies in a Small Mammal. Sci Rep 2019, 9, 10157. [Google Scholar] [CrossRef]
- Sonnenberg, B.R.; Branch, C.L.; Pitera, A.M.; Bridge, E.; Pravosudov, V.V. Natural Selection and Spatial Cognition in Wild Food-Caching Mountain Chickadees. Current Biology 2019, 29, 670–676.e3. [Google Scholar] [CrossRef]
- Melin, A.D.; Young, H.C.; Mosdossy, K.N.; Fedigan, L.M. Seasonality, Extractive Foraging and the Evolution of Primate Sensorimotor Intelligence. Journal of Human Evolution 2014, 71, 77–86. [Google Scholar] [CrossRef]
- Tomasello, M. Primate Cognition: Introduction to the Issue. Cognitive Science 2000, 24, 351–361. [Google Scholar] [CrossRef]
- Whiten, A.; Byrne, R.W. Machiavellian Intelligence II: Extensions and Evaluations; Cambridge University Press, 1997; ISBN 978-0-521-55949-2. [Google Scholar]
- Balda, R.P.; Kamil, A.C. Long-Term Spatial Memory in Clark’s Nutcracker, Nucifraga Columbiana. Animal Behaviour 1992, 44, 761–769. [Google Scholar] [CrossRef]
- Raby, C.R.; Alexis, D.M.; Dickinson, A.; Clayton, N.S. Planning for the Future by Western Scrub-Jays. Nature 2007, 445, 919–921. [Google Scholar] [CrossRef]
- Clutton-Brock, T.H.; Harvey, P.H. Primates, Brains and Ecology. Journal of Zoology 1980, 190, 309–323. [Google Scholar] [CrossRef]
- DeCasien, A.R.; Williams, S.A.; Higham, J.P. Primate Brain Size Is Predicted by Diet but Not Sociality. Nat Ecol Evol 2017, 1, 1–7. [Google Scholar] [CrossRef]
- Heldstab, S.A.; Kosonen, Z.K.; Koski, S.E.; Burkart, J.M.; van Schaik, C.P.; Isler, K. Manipulation Complexity in Primates Coevolved with Brain Size and Terrestriality. Sci Rep 2016, 6, 24528. [Google Scholar] [CrossRef] [PubMed]
- Powell, L.E.; Isler, K.; Barton, R.A. Re-Evaluating the Link between Brain Size and Behavioural Ecology in Primates. Proceedings of the Royal Society B: Biological Sciences 2017, 284, 20171765. [Google Scholar] [CrossRef]
- Kabadayi, C.; Taylor, L.A.; von Bayern, A.M.P.; Osvath, M. Ravens, New Caledonian Crows and Jackdaws Parallel Great Apes in Motor Self-Regulation despite Smaller Brains. Royal Society Open Science 2016, 3, 160104. [Google Scholar] [CrossRef] [PubMed]
- Bosshard, T.C.; Salazar, L.T.H.; Laska, M. Numerical Cognition in Black-Handed Spider Monkeys (Ateles Geoffroyi). Behavioural Processes 2022, 201, 104734. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, R.I.M. The Social Brain Hypothesis. Evolutionary Anthropology: Issues, News, and Reviews 1998, 6, 178–190. [Google Scholar] [CrossRef]
- Humphrey, N.K. The Social Function of Intellect. In Growing points in ethology; Cambridge U Press: Oxford, England, 1976. [Google Scholar]
- Jolly, A. Lemur Social Behavior and Primate Intelligence. Science 1966, 153, 501–506. [Google Scholar] [CrossRef]
- Brosnan, S.F.; Bshary, R. Cooperation and Deception: From Evolution to Mechanisms. Philosophical Transactions of the Royal Society B: Biological Sciences 2010, 365, 2593–2598. [Google Scholar] [CrossRef]
- Ashton, B.J.; Thornton, A.; Ridley, A.R. An Intraspecific Appraisal of the Social Intelligence Hypothesis. Philosophical Transactions of the Royal Society B: Biological Sciences 2018, 373, 20170288. [Google Scholar] [CrossRef]
- Speechley, E.M.; Ashton, B.J.; Foo, Y.Z.; Simmons, L.W.; Ridley, A.R. Meta-Analyses Reveal Support for the Social Intelligence Hypothesis. Biological Reviews 2024, 99, 1889–1908. [Google Scholar] [CrossRef]
- Mace, G.M.; Harvey, P.H.; Clutton-Brock, T.H. Brain Size and Ecology in Small Mammals. Journal of Zoology 1981, 193, 333–354. [Google Scholar] [CrossRef]
- Deaner, R.O.; Isler, K.; Burkart, J.; van Schaik, C. Overall Brain Size, and Not Encephalization Quotient, Best Predicts Cognitive Ability across Non-Human Primates. Brain Behavior and Evolution 2007, 70, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Barton, R.A. The Evolutionary Ecology of the Primate Brain. 1999.
- Connor, R.C.; Mann, J.; Tyack, P.L.; Whitehead, H. Social Evolution in Toothed Whales. Trends in Ecology & Evolution 1998, 13, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Joffe, T.H.; Dunbar, R.I.M. Visual and Socio–Cognitive Information Processing in Primate Brain Evolution. Proceedings of the Royal Society of London. Series B: Biological Sciences 1997, 264, 1303–1307. [Google Scholar] [CrossRef]
- Dunbar, R.I.M.; Shultz, S. Evolution in the Social Brain. Science 2007, 317, 1344–1347. [Google Scholar] [CrossRef]
- Holekamp, K.E. Questioning the Social Intelligence Hypothesis. Trends in Cognitive Sciences 2007, 11, 65–69. [Google Scholar] [CrossRef]
- Shultz, S.; Dunbar, R. i. m Both Social and Ecological Factors Predict Ungulate Brain Size. Proceedings of the Royal Society B: Biological Sciences 2005, 273, 207–215. [Google Scholar] [CrossRef]
- MacLean, E.L.; Barrickman, N.L.; Johnson, E.M.; Wall, C.E. Sociality, Ecology, and Relative Brain Size in Lemurs. Journal of Human Evolution 2009, 56, 471–478. [Google Scholar] [CrossRef]
- Beauchamp, G.; Fernández-Juricic, E. Is There a Relationship between Forebrain Size and Group Size in Birds? Evol Ecol Res 2004, 6, 833–842. [Google Scholar]
- Burish, M.J.; Kueh, H.Y.; Wang, S.S.-H. Brain Architecture and Social Complexity in Modern and Ancient Birds. Brain Behavior and Evolution 2004, 63, 107–124. [Google Scholar] [CrossRef]
- Iwaniuk, A.N.; Hurd, P.L. The Evolution of Cerebrotypes in Birds. Brain Behavior and Evolution 2005, 65, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Iwaniuk, A.N.; Arnold, K.E. Is Cooperative Breeding Associated With Bigger Brains? A Comparative Test in the Corvida (Passeriformes). Ethology 2004, 110, 203–220. [Google Scholar] [CrossRef]
- Healy, S.D.; Rowe, C. Costs and Benefits of Evolving a Larger Brain: Doubts over the Evidence That Large Brains Lead to Better Cognition. Animal Behaviour 2013, 86, e1–e3. [Google Scholar] [CrossRef]
- MacLean, E.L.; Hare, B.; Nunn, C.L.; Addessi, E.; Amici, F.; Anderson, R.C.; Aureli, F.; Baker, J.M.; Bania, A.E.; Barnard, A.M.; et al. The Evolution of Self-Control. Proceedings of the National Academy of Sciences 2014, 111, E2140–E2148. [Google Scholar] [CrossRef] [PubMed]
- Güntürkün, O.; Bugnyar, T. Cognition without Cortex. Trends Cogn Sci 2016, 20, 291–303. [Google Scholar] [CrossRef] [PubMed]
- MacLean, E.L.; Sandel, A.A.; Bray, J.; Oldenkamp, R.E.; Reddy, R.B.; Hare, B.A. Group Size Predicts Social but Not Nonsocial Cognition in Lemurs. PLOS ONE 2013, 8, e66359. [Google Scholar] [CrossRef]
- Whiten, A.; Byrne, R.W. Tactical Deception in Primates. Behavioral and Brain Sciences 1988, 11, 233–244. [Google Scholar] [CrossRef]
- Deaner, R.O.; van Schaik, C.P.; Johnson, V. Do Some Taxa Have Better Domain-General Cognition than Others? A Meta-Analysis of Nonhuman Primate Studies. Evol Psychol 2006, 4, 147470490600400114. [Google Scholar] [CrossRef]
- Gigerenzer, G. The Modularity of Social Intelligence. In Adaptive Thinking: Rationality in the Real World; Gigerenzer, G., Ed.; Oxford University Press, 2002; ISBN 978-0-19-515372-9. [Google Scholar]
- Reader, S.M.; Hager, Y.; Laland, K.N. The Evolution of Primate General and Cultural Intelligence. Philosophical Transactions of the Royal Society B: Biological Sciences 2011, 366, 1017–1027. [Google Scholar] [CrossRef]
- Kappeler, P.M. A Framework for Studying Social Complexity. Behav Ecol Sociobiol 2019, 73, 13. [Google Scholar] [CrossRef]
- Cords, M.; Aureli, F.; Silk, J.B.; Call, J. CHAPTER 9 Reconciliation and Relationship Qualities. In Natural Conflict Resolution; Aureli, F., Ed.; University of California Press, 2000; pp. 177–198. ISBN 978-0-520-92493-2. [Google Scholar]
- Emery, N.J.; Seed, A.M.; von Bayern, A.M.P.; Clayton, N.S. Cognitive Adaptations of Social Bonding in Birds. Philosophical Transactions of the Royal Society B: Biological Sciences 2007, 362, 489–505. [Google Scholar] [CrossRef] [PubMed]
- Shultz, S.; Dunbar, R.I.M. Social Bonds in Birds Are Associated with Brain Size and Contingent on the Correlated Evolution of Life-History and Increased Parental Investment. Biological Journal of the Linnean Society 2010, 100, 111–123. [Google Scholar] [CrossRef]
- Freeberg, T.M.; Dunbar, R.I.M.; Ord, T.J. Social Complexity as a Proximate and Ultimate Factor in Communicative Complexity. Philosophical Transactions of the Royal Society B: Biological Sciences 2012, 367, 1785–1801. [Google Scholar] [CrossRef]
- Amici, F.; Aureli, F.; Call, J. Fission-Fusion Dynamics, Behavioral Flexibility, and Inhibitory Control in Primates. Current Biology 2008, 18, 1415–1419. [Google Scholar] [CrossRef]
- Bond, A.B.; Kamil, A.C.; Balda, R.P. Serial Reversal Learning and the Evolution of Behavioral Flexibility in Three Species of North American Corvids (Gymnorhinus Cyanocephalus, Nucifraga Columbiana, Aphelocoma Californica). Journal of Comparative Psychology 2007, 121, 372–379. [Google Scholar] [CrossRef]
- Lukas, D.; Clutton-Brock, T. Social Complexity and Kinship in Animal Societies. Ecology Letters 2018, 21, 1129–1134. [Google Scholar] [CrossRef] [PubMed]
- Zuberbühler, K.; Byrne, R.W. Social Cognition. Current Biology 2006, 16, R786–R790. [Google Scholar] [CrossRef]
- References. In Animal Social Complexity: Intelligence, Culture, and Individualized Societies; de Waal, F.B.M., Tyack, P.L., Eds.; Harvard University Press, 2013; pp. 495–593 ISBN 978-0-674-41913-1.
- Holekamp, K.E.; Sakai, S.T.; Lundrigan, B.L. Social Intelligence in the Spotted Hyena (Crocuta Crocuta). Philosophical Transactions of the Royal Society B: Biological Sciences 2007, 362, 523–538. [Google Scholar] [CrossRef]
- Reader, S.M.; Laland, K.N. Social Intelligence, Innovation, and Enhanced Brain Size in Primates. Proceedings of the National Academy of Sciences 2002, 99, 4436–4441. [Google Scholar] [CrossRef]
- Lefebvre, L.; Giraldeau, L.-A. Is Social Learning an Adaptive Specialization. In Social learning in animals: The roots of culture; Academic Press: San Diego, CA, US, 1996; ISBN 978-0-12-273965-1. [Google Scholar]
- Deaner, R.O.; Nunn, C.L.; van Schaik, C.P. Comparative Tests of Primate Cognition: Different Scaling Methods Produce Different Results. Brain Behavior and Evolution 2000, 55, 44–52. [Google Scholar] [CrossRef]
- Dall, S.R.X.; Giraldeau, L.-A.; Olsson, O.; McNamara, J.M.; Stephens, D.W. Information and Its Use by Animals in Evolutionary Ecology. Trends in Ecology & Evolution 2005, 20, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, A.S.; Stephens, D.W. Reliability, Uncertainty, and Costs in the Evolution of Animal Learning. Current Opinion in Behavioral Sciences 2016, 12, 73–79. [Google Scholar] [CrossRef]
- Jetz, W.; Rubenstein, D.R. Environmental Uncertainty and the Global Biogeography of Cooperative Breeding in Birds. Current Biology 2011, 21, 72–78. [Google Scholar] [CrossRef]
- Brett, Z.H.; Humphreys, K.L.; Fleming, A.S.; Kraemer, G.W.; Drury, S.S. Using Cross-Species Comparisons and a Neurobiological Framework to Understand Early Social Deprivation Effects on Behavioral Development. Development and Psychopathology 2015, 27, 347–367. [Google Scholar] [CrossRef]
- Knudsen, E.I. Sensitive Periods in the Development of the Brain and Behavior. Journal of Cognitive Neuroscience 2004, 16, 1412–1425. [Google Scholar] [CrossRef]
- Brainard, M.S.; Doupe, A.J. What Songbirds Teach Us about Learning. Nature 2002, 417, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Bolhuis, J.J.; Honey, R.C. Imprinting, Learning and Development: From Behaviour to Brain and Back. Trends in Neurosciences 1998, 21, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Di Giorgio, E.; Loveland, J.L.; Mayer, U.; Rosa-Salva, O.; Versace, E.; Vallortigara, G. Filial Responses as Predisposed and Learned Preferences: Early Attachment in Chicks and Babies. Behavioural Brain Research 2017, 325, 90–104. [Google Scholar] [CrossRef]
- Ramsay, A.O.; Hess, E.H. A Laboratory Approach to the Study of Imprinting. The Wilson Bulletin 1954, 66, 196–206. [Google Scholar]
- Versace, E.; Vallortigara, G. Origins of Knowledge: Insights from Precocial Species. Front. Behav. Neurosci. 2015, 9. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Diorio, J.; Tannenbaum, B.; Caldji, C.; Francis, D.; Freedman, A.; Sharma, S.; Pearson, D.; Plotsky, P.M.; Meaney, M.J. Maternal Care, Hippocampal Glucocorticoid Receptors, and Hypothalamic-Pituitary-Adrenal Responses to Stress. Science 1997, 277, 1659–1662. [Google Scholar] [CrossRef]
- Sheridan, M.A.; McLaughlin, K.A. Dimensions of Early Experience and Neural Development: Deprivation and Threat. Trends in Cognitive Sciences 2014, 18, 580–585. [Google Scholar] [CrossRef]
- Hatch, A.; Wiberg, G.S.; Balazs, T.; Grice, H.C. Long-Term Isolation Stress in Rats. Science 1963, 142, 507–507. [Google Scholar] [CrossRef]
- Suomi, S.J. Early Determinants of Behaviour: Evidence from Primate Studies. British Medical Bulletin 1997, 53, 170–184. [Google Scholar] [CrossRef]
- Bowlby, J. Maternal Care and Mental Health. Bulletin of the World Health Organization 1951, 3, 355–533. [Google Scholar]
- Rutter, M.; Team, the E. and R.A. (ERA) study Developmental Catch-up, and Deficit, Following Adoption after Severe Global Early Privation. The Journal of Child Psychology and Psychiatry and Allied Disciplines 1998, 39, 465–476. [CrossRef]
- Petanjek, Z.; Judaš, M.; Šimić, G.; Rašin, M.R.; Uylings, H.B.M.; Rakic, P.; Kostović, I. Extraordinary Neoteny of Synaptic Spines in the Human Prefrontal Cortex. Proceedings of the National Academy of Sciences 2011, 108, 13281–13286. [Google Scholar] [CrossRef]
- Turner, A.M.; Greenough, W.T. Differential Rearing Effects on Rat Visual Cortex Synapses. I. Synaptic and Neuronal Density and Synapses per Neuron. Brain Research 1985, 329, 195–203. [Google Scholar] [CrossRef]
- Globus, A.; Rosenzweig, M.R.; Bennett, E.L.; Diamond, M.C. Effects of Differential Experience on Dendritic Spine Counts in Rat Cerebral Cortex. Journal of Comparative and Physiological Psychology 1973, 82, 175–181. [Google Scholar] [CrossRef]
- Greenough, W.T.; Volkmar, F.R. Pattern of Dendritic Branching in Occipital Cortex of Rats Reared in Complex Environments. Experimental Neurology 1973, 40, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Volkmar, F.R.; Greenough, W.T. Rearing Complexity Affects Branching of Dendrites in the Visual Cortex of the Rat. Science 1972, 176, 1445–1447. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.C.; Law, F.; Rhodes, H.; Lindner, B.; Rosenzweig, M.R.; Krech, D.; Bennett, E.L. Increases in Cortical Depth and Glia Numbers in Rats Subjected to Enriched Environment. Journal of Comparative Neurology 1966, 128, 117–125. [Google Scholar] [CrossRef]
- Makinodan, M.; Rosen, K.M.; Ito, S.; Corfas, G. A Critical Period for Social Experience–Dependent Oligodendrocyte Maturation and Myelination. Science 2012, 337, 1357–1360. [Google Scholar] [CrossRef]
- Sabatini, M.J.; Ebert, P.; Lewis, D.A.; Levitt, P.; Cameron, J.L.; Mirnics, K. Amygdala Gene Expression Correlates of Social Behavior in Monkeys Experiencing Maternal Separation. J. Neurosci. 2007, 27, 3295–3304. [Google Scholar] [CrossRef]
- Weaver, I.C.G.; Meaney, M.J.; Szyf, M. Maternal Care Effects on the Hippocampal Transcriptome and Anxiety-Mediated Behaviors in the Offspring That Are Reversible in Adulthood. Proceedings of the National Academy of Sciences 2006, 103, 3480–3485. [Google Scholar] [CrossRef]
- Baroncelli, L.; Scali, M.; Sansevero, G.; Olimpico, F.; Manno, I.; Costa, M.; Sale, A. Experience Affects Critical Period Plasticity in the Visual Cortex through an Epigenetic Regulation of Histone Post-Translational Modifications. J. Neurosci. 2016, 36, 3430–3440. [Google Scholar] [CrossRef]
- Champagne, F.A.; Curley, J.P. How Social Experiences Influence the Brain. Current Opinion in Neurobiology 2005, 15, 704–709. [Google Scholar] [CrossRef]
- Seckl, J.R. Glucocorticoids, Developmental ‘Programming’ and the Risk of Affective Dysfunction. In Progress in Brain Research; De Kloet, E.R., Oitzl, M.S., Vermetten, E., Eds.; Stress Hormones and Post Traumatic Stress Disorder Basic Studies and Clinical Perspectives; Elsevier, 2007; Vol. 167, pp. 17–34. [Google Scholar]
- Arnold, C.; Taborsky, B. Social Experience in Early Ontogeny Has Lasting Effects on Social Skills in Cooperatively Breeding Cichlids. Animal Behaviour 2010, 79, 621–630. [Google Scholar] [CrossRef]
- Nyman, C.; Fischer, S.; Aubin-Horth, N.; Taborsky, B. Effect of the Early Social Environment on Behavioural and Genomic Responses to a Social Challenge in a Cooperatively Breeding Vertebrate. Molecular Ecology 2017, 26, 3186–3203. [Google Scholar] [CrossRef]
- Fahlke, C.; Lorenz, J.G.; Long, J.; Champoux, M.; Suomi, S.J.; Higley, J.D. Rearing Experiences and Stress-Induced Plasma Cortisol as Early Risk Factors for Excessive Alcohol Consumption in Nonhuman Primates. Alcoholism: Clinical and Experimental Research 2000, 24, 644–650. [Google Scholar] [CrossRef]
- Francis, D.D.; Meaney, M.J. Maternal Care and the Development of Stress Responses. Current Opinion in Neurobiology 1999, 9, 128–134. [Google Scholar] [CrossRef]
- Suomi, S.J. Developmental Trajectories, Early Experiences, and Community Consequences: Lessons from Studies with Rhesus Monkeys. In Developmental health and the wealth of nations: Social, biological, and educational dynamics; The Guilford Press: New York, NY, US, 1999; ISBN 978-1-57230-454-3. [Google Scholar]
- Caldji, C.; Diorio, J.; Meaney, M.J. Variations in Maternal Care in Infancy Regulate the Development of Stress Reactivity. Biological Psychiatry 2000, 48, 1164–1174. [Google Scholar] [CrossRef]
- Ladd, C.O.; Huot, R.L.; Thrivikraman, K.V.; Nemeroff, C.B.; Plotsky, P.M. Long-Term Adaptations in Glucocorticoid Receptor and Mineralocorticoid Receptor Mrna and Negative Feedback on the Hypothalamo-Pituitary-Adrenal Axis Following Neonatal Maternal Separation. Biological Psychiatry 2004, 55, 367–375. [Google Scholar] [CrossRef]
- Oers, H.J.J. van; Kloet, E.R. de; Whelan, T.; Levine, S. Maternal Deprivation Effect on the Infant’s Neural Stress Markers Is Reversed by Tactile Stimulation and Feeding But Not by Suppressing Corticosterone. J. Neurosci. 1998, 18, 10171–10179. [Google Scholar] [CrossRef]
- Dettling, A.; Pryce, C.R.; Martin, R.D.; Döbeli, M. Physiological Responses to Parental Separation and a Strange Situation Are Related to Parental Care Received in Juvenile Goeldi’s Monkeys (Callimico Goeldii). Developmental Psychobiology 1998, 33, 21–31. [Google Scholar] [CrossRef]
- Hall, F. s.; Wilkinson, L. s.; Humby, T.; Robbins, T. w. Maternal Deprivation of Neonatal Rats Produces Enduring Changes in Dopamine Function. Synapse 1999, 32, 37–43. [Google Scholar] [CrossRef]
- Schultz, W. Neuronal Reward and Decision Signals: From Theories to Data. Physiol Rev 2015, 95, 853–951. [Google Scholar] [CrossRef] [PubMed]
- Wise, R.A. Dopamine, Learning and Motivation. Nat Rev Neurosci 2004, 5, 483–494. [Google Scholar] [CrossRef]
- Graybiel, A.M. The Basal Ganglia: Learning New Tricks and Loving It. Current Opinion in Neurobiology 2005, 15, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, T.; DeLong, M.R. Deep-Brain Stimulation for Basal Ganglia Disorders. Basal Ganglia 2011, 1, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Lisman, J.E.; Grace, A.A. The Hippocampal-VTA Loop: Controlling the Entry of Information into Long-Term Memory. Neuron 2005, 46, 703–713. [Google Scholar] [CrossRef]
- Shohamy, D.; Adcock, R.A. Dopamine and Adaptive Memory. Trends in Cognitive Sciences 2010, 14, 464–472. [Google Scholar] [CrossRef]
- Nieoullon, A. Dopamine and the Regulation of Cognition and Attention. Progress in Neurobiology 2002, 67, 53–83. [Google Scholar] [CrossRef]
- Nestler, E.J.; Carlezon, W.A. The Mesolimbic Dopamine Reward Circuit in Depression. Biological Psychiatry 2006, 59, 1151–1159. [Google Scholar] [CrossRef]
- Pizzagalli, D.A. Depression, Stress, and Anhedonia: Toward a Synthesis and Integrated Model. Annual Review of Clinical Psychology 2014, 10, 393–423. [Google Scholar] [CrossRef] [PubMed]
- Melzack, R.; Burns, S.K. Neurophysiological Effects of Early Sensory Restriction. Exp Neurol 1965, 13, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Gluck, J.P.; Sackett, G.P. Extinction Deficits in Socially Isolated Rhesus Monkeys (Macaca Mulatta). Developmental Psychology 1976, 12, 173–174. [Google Scholar] [CrossRef]
- Moretz, J.A.; Martins, E.P.; Robison, B.D. The Effects of Early and Adult Social Environment on Zebrafish (Danio Rerio) Behavior. Environ Biol Fish 2007, 80, 91–101. [Google Scholar] [CrossRef]
- Adkins-Regan, E.; Krakauer, A. Removal of Adult Males from the Rearing Environment Increases Preference for Same-Sex Partners in the Zebra Finch. Animal Behaviour 2000, 60, 47–53. [Google Scholar] [CrossRef]
- Bertin, A.; Richard-Yris, M.-A. Mothering during Early Development Influences Subsequent Emotional and Social Behaviour in Japanese Quail. Journal of Experimental Zoology Part A: Comparative Experimental Biology 2005, 303A, 792–801. [Google Scholar] [CrossRef]
- Bester-Meredith, J.K.; Marler, C.A. Social Experience During Development and Female Offspring Aggression in Peromyscus Mice. Ethology 2007, 113, 889–900. [Google Scholar] [CrossRef]
- Bastian, M.L.; Sponberg, A.C.; Sponberg, A.C.; Suomi, S.J.; Higley, J.D. Long-Term Effects of Infant Rearing Condition on the Acquisition of Dominance Rank in Juvenile and Adult Rhesus Macaques (Macaca Mulatta). Developmental Psychobiology 2003, 42, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Meaney, M.J.; Szyf, M. Maternal Care as a Model for Experience-Dependent Chromatin Plasticity? Trends in Neurosciences 2005, 28, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Barrett, L.; Henzi, S.P.; Lusseau, D. Taking Sociality Seriously: The Structure of Multi-Dimensional Social Networks as a Source of Information for Individuals. Philosophical Transactions of the Royal Society B: Biological Sciences 2012, 367, 2108–2118. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Numerical Processing in Social Contexts. Numerical information provides advantages during social behaviors such as inter-group conflict (assessing group size), navigating social hierarchy (ordering group members), and initiating collective action (e.g. estimating proportions of individuals involved in a specific behavior).
Figure 1.
Numerical Processing in Social Contexts. Numerical information provides advantages during social behaviors such as inter-group conflict (assessing group size), navigating social hierarchy (ordering group members), and initiating collective action (e.g. estimating proportions of individuals involved in a specific behavior).
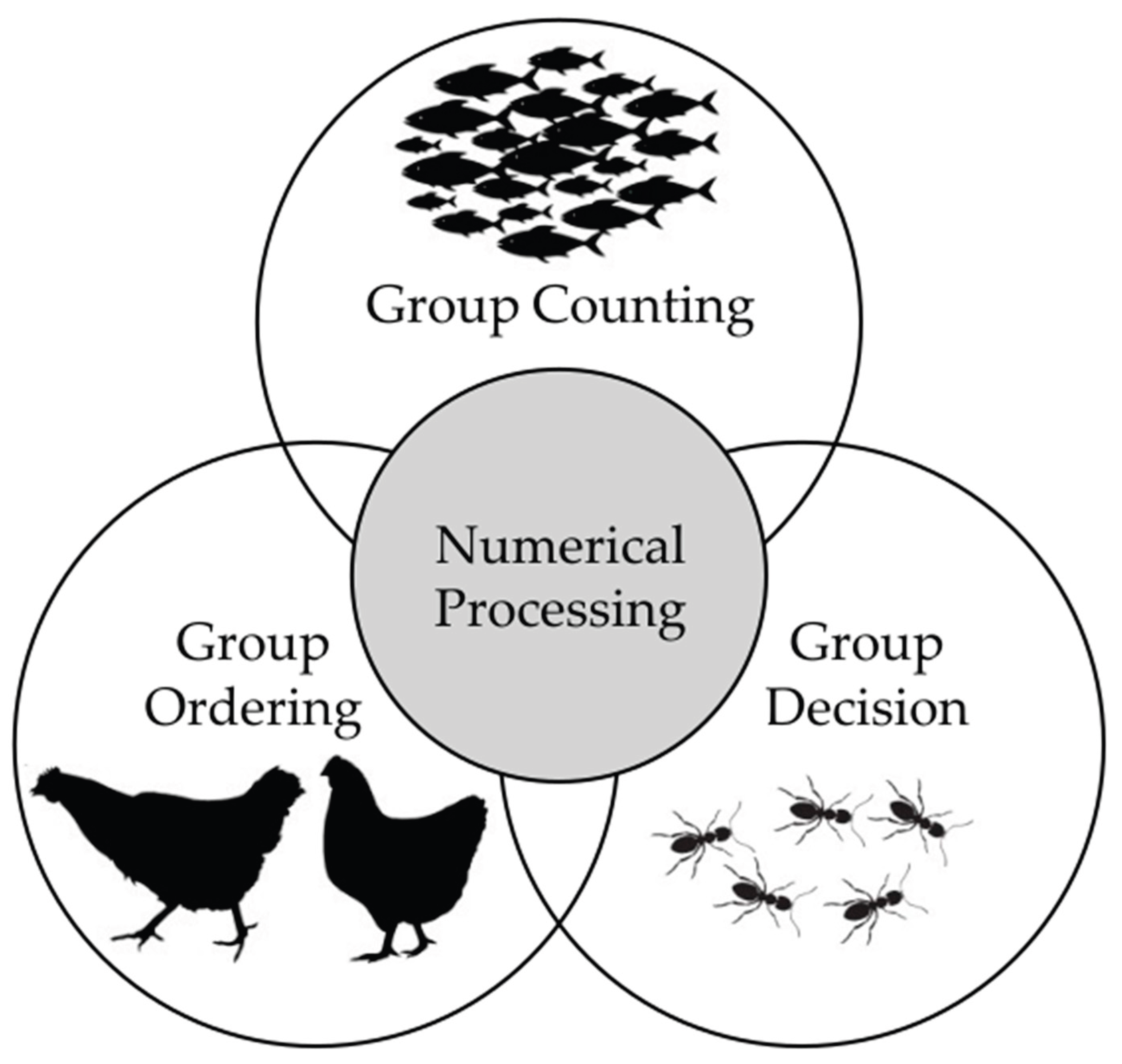
Table 1.
Social functions of numerical cognition across species. The table summarizes fundamental numerical abilities and their possible adaptive roles in social contexts across multiple taxa.
Table 1.
Social functions of numerical cognition across species. The table summarizes fundamental numerical abilities and their possible adaptive roles in social contexts across multiple taxa.
| Social function | Numerical abilities | Species |
|---|---|---|
| Assessment of groups size during conflicts | Proto-numerical Discrimination Proportional Reasoning |
Lions (Panthera leo) [111] Spotted hyenas (Crocuta crocuta) [112] Chimpanzees (Pan troglodytes) [113] Black howler monkey (Alouatta pigra) [114] Subdesert mesite (Monias benschi) [115] Free-ranging dogs (Canis lupus familiaris) [116] Western Australian magpies (Gymnorhina tibicen dorsalis) [62] |
| Protection from predators via group size | Proto-numerical Discrimination | Guppies (Poecilia reticulata) [117,118] Sticklebacks (Gasterosteus aculeatus) [119] Red colobus (Procolobus badius) [120] Diana monkey (Cercopithecus diana) [120] Zebrafish (Danio rerio) [121] |
| Navigation of dominance hierarchies | Ordinal Processing | Baboons (Papio anubis) [108] Ravens (Corvus corax) [122] Greylag geese (Anser anser) [123] Domestic chickens (Gallus gallus) [124,125] Paper wasps (Polistes dominula and Polistes metricus) [126] |
| Initiation of collective actions | Proto-numerical Discrimination Proportional Reasoning |
Ants (Myrmecina nipponica) [127,128,129] Capuchin monkeys (Cebus capucinus) [130] Baboons (Papio anubis) [131] African wild dogs (Lycaon pictus) [132] Meerkats (Suricata suricatta) [133] Honeybees (Apis mellifera) [134] |
| Balancing foraging trade-offs | Proportional Reasoning | Mallards (Anas platyrhynchos) [87] Cichlids (Astatotilapia burtoni) [90] |
| Tracking group members | Proto-Arithmetic | Domestic chicks (Gallus gallus): [135,136] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



